Изобретение относится к биотехнологии, к генной инженерии, микробиологической и медицинской промышленности и представляет собой дрожжевой штамм - продуцент интерлейкина-2 человека и способ его получения.
Интерлейкин-2 человека относится к семейству цитокинов, сигнальных молекул иммунной системы, которые участвуют в формировании и регуляции защитных реакций организма. В рамках иммунной системы цитокины осуществляют взаимосвязь между неспецифическими защитными реакциями и специфическим иммунитетом. На уровне организма цитокины обеспечивают связь между иммунной, нервной, эндокринной и кроветворной системами (Симбирцев А.С. Цитокины и воспаление, 2002, т.1, с.9-16).
Благодаря использованию методов генной инженерии и биотехнологии многие цитокины получены в виде рекомбинантных препаратов, некоторые из них применяются в клинической практике в виде лекарственных средств (Кетлинский С.А. и др. Эндогенные иммуномодуляторы. СПб., 1992). Использование рекомбинантных цитокинов восполняет дефицит эндогенных молекул, повышает эффективность иммунотерапии за счет адекватной и целенаправленной медикаментозной коррекции иммунной дисфункции. В настоящее время иммунотерапия рекомбинантными цитокинами является одним из наиболее перспективных и постоянно расширяющихся направлений иммунофармакологии.
Основная функция интерлейкина-2 состоит в обеспечении клеточной составляющей адаптивного иммунитета. Интерлейкин-2 является фактором роста и дифференцировки Т-лимфоцитов, NK-клеток, обеспечивает дифференцировку Т-киллеров и способствует проявлению функциональной активности Т-хелперов. Интерлейкин-2 усиливает синтез иммуноглобулинов предварительно активированными В-лимфоцитами. Он непосредственно воздействует на моноциты, экспрессирующие рецептор интерлейкина-2. Широкий спектр биологических активностей интерлейкина-2 делает его ключевым компонентом иммунной системы (Taniguchi Т. and Minani Y., Cell, 1993, v.73, p.5-8; Jeal W. and Goa K.L., BioDrugs, 1997, v.7, p.285-317; Rodriquez Z.M. et al. Infection, 1996, v.24, p.115-120; Ярилин А.А. Основы иммунологии. М., 1999).
Наиболее перспективным подходом для получения цитокинов в значительных количествах является использование микроорганизмов в качестве продуцентов этих белков. Применение непатогенных микроорганизмов (дрожжей), не содержащих токсических и пирогенных факторов, в качестве продуцентов белков человека позволяет использовать рекомбинантные белки в клинической практике. Созданный ранее в лаборатории биохимической генетики БиНИИ СПбГУ штамм дрожжей Saccharomyces cerevisiae ВКПМ Y-791 - продуцент интерлейкина-2 человека синтезировал 30-50 мг биологически активного рекомбинантного белка на литр культуральной среды (Мясников А.Н. и др. Патент SU № 1770359, приоритет от 24 марта 1988). Лекарственная форма рекомбинантного дрожжевого интерлейкина-2 человека Ронколейкин успешно используется для лечения септических состояний, ряда онкологических и инфекционных заболеваний (Ронколейкин® - рекомбинантный интерлейкин-2 человека. Под ред. Козлова В.К. СПб., 2001). Перспектива широкого применения рекомбинантного дрожжевого интерлейкина-2 человека в клинической практике ставит задачу повышения уровня продукции этого белка в клетках дрожжей.
Задачей, на решение которой направлены предлагаемые изобретения, является создание штамма дрожжей Saccharomyces cerevisiae - продуцента рекомбинантного интерлейкина-2 человека и разработки способа его получения.
Для решения поставленной задачи создан штамма дрожжей Saccharomyces cerevisiae - продуцент рекомбинантного интерлейкина-2 человека и разработан способ получения штамма дрожжей Saccharomyces cerevisiae - продуцента рекомбинантного интерлейкина-2.
Штамм 1-60-Д578 получают из штамма 60-Д578, который трансформируют плазмидой pADPI с целью нарушения гена РЕР4. Для получения штамма 60-Д578 скрещивают штаммы 33-Д373 (MAT α pheA10 ade2-144,717 his7-l lys9-A21 ura3-52 trp1-289 leu2-3,112) и 5-Д502 (MATa arg6 pho3) Петергофской генетической коллекции (Андрианова В.М., Инге-Вечтомов С.Г. Каталог (указатель) Петергофской генетической коллекции дрожжей Saccharomyces cerevisiae. Л., 1988). В споровом потомстве отбирают штамм 60-Д578, содержащий мутации ade2-144,717, leu2-3,112, lys9-A21, arg6, pho3.
Для целенаправленного нарушения гена РЕР4 в штамме 60-Д578 конструируют плазмиду pADP1.
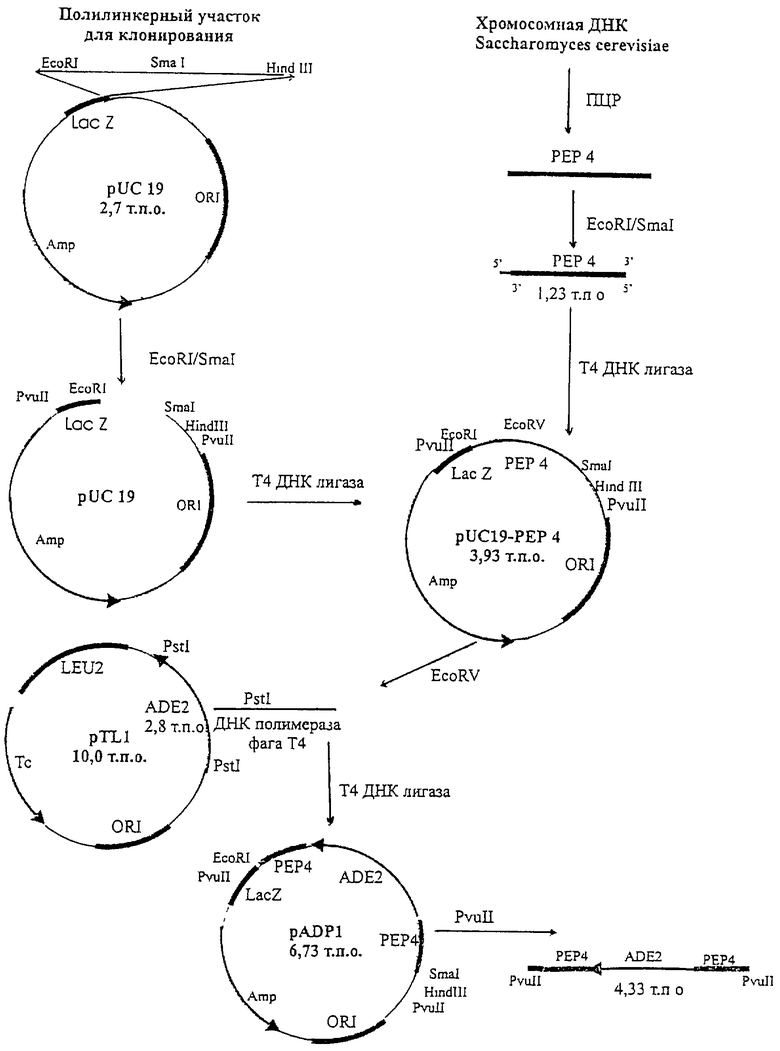
На чертеже представлена схема конструирования плазмиды pADP1.
На первом этапе синтезируют ген РЕР4 с помощью ПЦР, используя в качестве позитивного праймера последовательность 5'-ggaattcatgttcagcttga-3', содержащую сайт рестрикции для EcoRI, и в качестве негативного праймера последовательность 5'-tcccccgggtttagctcaaa-3', содержащую сайт рестрикции для SmaI, в качестве матрицы используют хромосомную ДНК дрожжей Saccharomyces cerevisiae.
Синтезированный ген РЕР4 (1,23 т.п.о.) выделяют при помощи электрофореза в 0,7% агарозном геле, обрабатывают рестриктазами EcoRI и SmaI и лигируют с плазмидой pUC19 (2,7 т.п.о.) (Roberts R.J. Nucl. Acids Res., 1984, v.12, suppl. p.167-204), предварительно обработанной этими же рестриктазами. Полученной лигазной смесью трансформируют клетки штамма DH5α Escherichia coli (F'/endAl hsdR17 (r
Плазмиду pUC19-PEP4 обрабатывают рестриктазой EcoRV и лигируют с фрагментом ДНК, содержащим ген ADE2. Этот фрагмент получают при рестрикции плазмиды pTL1 (Останин К.В. и др. Биополимеры и клетка, 1988, т.4, с.272-279) рестриктазой Pst1, разделения фрагментов ДНК с помощью электрофореза в 0,7% агарозном геле, выделения фрагмента размером 2,80 т.п.о., затупления липких концов с помощью ДНК полимеразой I фага Т4. После лигирования полученной смесью трансформируют клетки штамма DH5α Escherichia coli (F'/endAl hsdR17 (r
Для получения штамма-продуцента интерлейкина-2 человека в штамм 1-60-Д578 вводят плазмиду pJDB(MSIL) (Мясников А.Н., Смирнов М.Н., Авот А.Я. и др. Патент РФ SU 1770359, БИ № 39, 1992) и отбирают трансформанты на среде без аденина и лейцина.
Экспрессия гена интерлейкина-2 человека в составе плазмиды pJDB(MSIL) находится под контролем дрожжевого промотора РНO5, содержащим области, обеспечивающие активацию транскрипции при отсутствии неорганического фосфата в культуральной среде, а также область инициации транскрипции. Промотор гена РНO5 относится к числу наиболее сильных дрожжевых промоторов. Уровень экспрессии генов, находящихся под контролем РНO5 промотора, эффективно регулируется экзогенным неорганическим фосфатом. Плазмида pJDB(MSIL) получена на основе челночного бактериально-дрожжевого вектора pJDB207, который является одним из наиболее стабильных и высококопийных векторов дрожжей. В его состав входит ген LEU2 дрожжей, являющийся селективным маркером для отбора трансформантов.
Штамм-реципиент 1-60-Д578 несет мутацию в гене LEU2, что позволяет отбирать трансформантов, несущих плазмиду pJDB(MSIL). Кроме того, он содержит нарушенный ген РЕР4. Мутация в этом гене приводит к отсутствию активности протеаз А и В, а также карбоксипептидазы Y в клетках дрожжей, что сопровождается повышением стабильности гетерологичных рекомбинантных белков (Hisch H.H. et al. In: Walton E.F., Yarranton G.T., Eds., Molecular and Cell Biology of Yeast, 1989, p.134-200). В отличие от штамма-прототипа ВКПМ Y-791, штамм 1-60-Д578 не несет мутации в гене РНO80. Мутация в этом гене сопровождается конститутивной экспрессией гена интерлейкина-2 человека независимо от концентрации неорганического фосфата. Синтез гетерологичного белка в течение всего периода культивирования штамма-продуцента является метаболической нагрузкой для клетки дрожжей и часто приводит к митотической нестабильности рекомбинантной плазмиды (Miyanohara A. et al., Proc. Natl. Acad. Sci., 1983, v.80, p.1-5).
Штамм дрожжей Saccharomyces cerevisiae 1-60-Д578 (MSIL) характеризуется следующими признаками.
Морфологические признаки.
Клетки округлой, слегка овальной формы, размером 5-10 мкм, часть клеток имеет на поверхности почки или соединена с дочерними клетками.
Культуральные признаки.
Клетки хорошо растут на полных органических средах:
ПЕП - 2% пептона, 2% глюкозы,
ПЕПФО - 2% пептона, 2% глюкозы, 0,1% однозамещенного фосфата калия,
YEPD - 2% пептона, 2% глюкозы, 1% дрожжевого экстракта.
Кроме того, клетки хорошо растут на минеральной среде SC: 0,67% Yeast Nitrogen Base (“Difco”, США), 2% глюкозы, 30 мг/л лизина, 30 мг/л аргинина, а также на других синтетических средах для дрожжей, содержащих 30 мг/л лизина, 30 мг/л аргинина.
При росте на твердых средах клетки образуют гладкие, круглые колонии с матовой поверхностью, светло-кремового цвета, край ровный.
При росте в жидких средах образуют интенсивную ровную суспензию. Культура имеет характерный запах дрожжей.
Физиолого-биохимические признаки.
Клетки растут в пределах от 4 до 37°. Оптимальной температурой выращивания является 30°. При росте в аэробных условиях клетки значительно закисляют среду. Оптимум рН для роста составляет 3,5-5,5.
В качестве источника углерода клетки могут использовать многие простые соединения, такие как глюкоза, сахароза, глицерин.
В качестве источника азота при условии добавки лизина и аргинина клетки используют минеральные соли в аммонийной форме, аминокислоты, мочевину.
Клетки способны к аэробному и анаэробному росту.
Существенными признаками штамма является ауксотрофность по лизину и аргинину и отсутствие потребности в лейцине и аденине.
Для получения штамма 1-60-Д578 с нарушенным геном РЕР4 используют плазмиду pADP1.
ПРИМЕР ОСУЩЕСТВЛЕНИЯ СПОСОБА
На первом этапе синтезируют ген РЕР4 с помощью ПЦР, используя в качестве матрицы хромосомную ДНК дрожжей Saccharomyces cerevisiae (см. чертеж). К 0,1 мкг хромосомной ДНК, растворенной в 5 мкл буфера ТЕ, добавляют 1 мкл 0.5 М NaOH и нагревают при 85° в течение 3 мин. Затем пробу быстро переносят в лед, добавляют 1 мкл 0.5 М НСl и далее используют в ПЦР.
В качестве позитивного праймера использовали последовательность 5'-ggaattcatgttcagcttga-3', содержащая сайт рестрикции для EcoRI, и в качестве негативного праймера - последовательность 5'-tcccccgggtttagctcaaa-3', содержащую сайт рестрикции для SmaI. Проба для ПЦР содержит 5 мкл матрицы, 30 рМ каждого праймера (по 2 мкл), 10 мкл 10-кратного раствора дезоксинуклеотидтрифосфатов (дНТФ), содержащего 1.25 мМ каждого дНТФ (дАТФ, дТТФ, дГТФ, дЦТФ), 10 мкл 10-кратного буфера для ПЦР (100 mM КСl, 100 тМ (NH4)2SO4, 200 тМ Трис-HCl, рН 8,8, 20 mM MgSO4, 1% тритон Х-100). В пробу добавляется дистиллированная H2O до конечного объема 100 мкл.
Далее пробу прогревают 5 мин при 95°С, охлаждают, добавляют 2.5 ед. (2.5 мкл) Вент-ДНК-полимеразы (“BioLabs”) и проводят 50 циклов ПЦР в следующих условиях: 1 мин при 95° (плавление цепей ДНК), 1 мин при 56° (отжиг праймеров), 1 мин при 72° (полимеразная реакция). После окончания ПЦР пробу инкубируют при 72° 5 мин.
Для доказательства того, что в ходе ПЦР синтезируется ген РЕР4, проводят электрофорез в 0,7% агарозном геле в буфере ТВЕ (0,1М трис-боратный буфер, рН 8,3, содержащий 1 мМ ЭДТА). Реакционную смесь вносят в лунки агарозного геля и проводят разделение фрагментов ДНК в течение 1-2 ч. По окончании разделения вырезают полоску геля, содержащую фрагмент ДНК размером 1,23 т.п.о., соответствующий гену РЕР4 Saccharomyces cerevisiae. Выделение ДНК из агарозного геля проводят по методике, разработанной фирмой QIAGEN. Полоску геля с фрагментом ДНК помещают в пробирку и добавляют раствор QX1 (300 мкл на 100 мг геля). Пробу нагревают до 50°, добавляют реактив QIAEX (10 мкл на 5 мкг ДНК) и инкубируют при 50° в течение 10 мин, периодически перемешивая. Далее центрифугируют 30 с при 15000 об/мин, супернатант отбрасывают, осадок дважды экстрагируют растворами QX2 и QX3, удаляют супернатант центрифугированием при 15000 об/мин в течение 30 с. Осадок высушивают на воздухе, растворяют в 20 мкл буфера ТЕ, центрифугируют 30 с при 15000 об/мин, супернатант переносят в новую пробирку.
Гидролиз фрагмента ДНК, содержащего синтезированный ген РЕР4 Saccharomyces cerevisiae, рестриктазами EcoRI и SmaI проводят в 10 мМ трис-хлоридном буфере (рН 7,5), содержащем 50 мМ хлористого натрия, 10 мМ хлористого магния и 1 мМ дитиотреитол. К 5 мкг плазмидной ДНК в объеме 20 мкл добавляют по 5 ед. каждой рестриктазы, после чего пробу инкубируют в течение 25 ч при 37°. Далее проводят лигирование с плазмидой pUC19, предварительно обработанной этими же рестриктазами.
Плазмиду pUC19 (2,7 т.п.о.) выделяют из клеток бактерий Escherichia coli, которые выращивают в течение ночи в 1 л питательной среды LB (1% пептона, 0,5% дрожжевого экстракта, 1% хлористого натрия), содержащей ампициллин в концентрации 50 мг/л. Клетки собирают центрифугированием при 5000 об/мин в течение 10 мин при 4°, суспендируют в 20 мл 25 мМ трис-хлоридного буфера (рН 8,0), содержащего 10 мМ ЭДТА и 50 мМ глюкозы, добавляют 30 мг лизоцима и инкубируют 10 мин при комнатной температуре. Далее добавляют 40 мл 0,2 М гидроокиси натрия, содержащей 1% додецилсульфата натрия, осторожно перемешивают и инкубируют в течение 10 мин при 4°. Раствор нейтрализуют добавлением 30 мл 3 М ацетата натрия (рН5,0) и выдерживают в течение 10 мин при 4°. После этого центрифугируют при 14000 об/мин в течение 40 мин при 4°. К супернатанту добавляют 0,6 объема изопропилового спирта, выдерживают 20 мин при комнатной температуре и центрифугируют при 14000 об/мин в течение 20 мин при 20°. Полученный осадок промывают 70% этиловым спиртом, высушивают в вакууме и растворяют в 4 мл дистиллированной воды. Далее добавляют 4,2 г хлористого цезия и 0,36 мл раствора бромистого этидия (10 мг/мл). Полученный раствор выдерживают в течение 1 ч при 4°, затем центрифугируют при 15000 об/мин в течение 15 мин. Супернатант центрифугируют при 70000 об/мин в течение 16 ч в центрифуге TL100 (Beckman). После центрифугирования отбирают полосу плазмидной ДНК (нижнюю из двух флюоресцирующих в ультрафиолетовом свете полос), дважды экстрагируют бромистый этидий равным объемом изоамилового спирта, разбавляют в два раза дистиллированной водой и осаждают плазмидную ДНК двумя объемами этилового спирта и 1/15 объема 3 М ацетата натрия (рН 5,0). Осадок собирают центрифугированием при 10000 об/мин в течение 10 мин, промывают 70% этиловым спиртом и растворяют в 0,5-1 мл буфера ТЕ (10 мМ трис-хлоридный буфер, рН 8,0, содержащий 1 мМ ЭДТА). Концентрацию плазмидной ДНК определяют по поглощению раствора при длине волны 260 нм. Чистоту препарата контролируют при помощи электрофореза в 0,7% агарозном геле в буфере ТВЕ.
Гидролиз плазмиды pUC19 рестриктазами EcoRI и SmaI осуществляют в условиях, описанных для расщепления синтезированного при помощи ПЦР гена РЕР4 Saccharomyces cerevisiae. Линеаризованную плазмидную ДНК выделяют по описанной выше методике фирмы QIAGEN, исключая стадию нагревания.
Для получения плазмиды pUC19-PEP4 проводят лигирование плазмиды pUC19, гидролизованной рестриктазами EcoRI и SmaI, и EcoRI/SmaI фрагмента гена PEP 4 Saccharomyces cerevisiae, синтезированного при помощи ПЦР. Для этого смешивают 0,5 мкг ДНК вектора и 0,1 мкг ДНК встройки в 10 мкл 70 мМ трис-хлоридного буфера (рН 7,6), содержащего 5 мМ дитиотреитола, 5 мМ хлористого магния, 1 мМ АТФ, добавляют 10 ед. ДНК-лигазы фага Т4 и инкубируют при 14° в течение ночи.
Полученной лигазной смесью трансформируют клетки штамма DH5α Escherichia coli (F’/endAl hsdR17 (r
Из выросших отдельных клонов трансформантов выделяют плазмидную ДНК pUC19-PEP4 (3,93 т.п.о.) при помощи методики, использованной для получения плазмиды pUC19, за исключением того что клетки Escherichia coli выращивают в 10 мл LB, и, соответственно, объемы всех растворов уменьшают в 100 раз. Кроме того, вместо стадии центрифугирования в градиенте плотности хлористого цезия проводят обработку ДНК панкреатической РНКазой. Для этого нуклеиновые кислоты, осажденные изопропиловым спиртом, растворяют в 100 мкл буфера ТЕ, добавляют 10 мкл раствора РНКазы (1мг/мл) и инкубируют 30 мин при 37°.
Далее плазмиду pUC19-PEP4 обрабатывают рестриктазой EcoRV в условиях, описанных для расщепления синтезированного при помощи ПЦР гена РЕР4 Saccharomyces cerevisiae, линеаризованную плазмидную ДНК выделяют по описанной выше методике фирмы QIAGEN, исключая стадию нагревания, и лигируют с фрагментом ДНК, содержащим ген ADE2 Saccharomyces cerevisiae. Этот фрагмент получают при рестрикции плазмиды pTL1 рестриктазой Pst1. Выделение плазмиды pTL1 и гидролиз ее рестриктазой Pst1 проводят в условиях, описанных выше для плазмиды pUC19. После гидролиза плазмиды pTL1 рестриктазой Pst1 проводят разделение фрагментов ДНК с помощью электрофореза в 0,7% агарозном геле и выделяют фрагмент 2,8 т.п.о. по описанной выше методике фирмы QIAGEN.
Полученный фрагмент обрабатывают ДНК полимеразой I фага Т4 в 50 мМ трис-хлоридном буфере (рН 7,8), содержащем 10мМ хлористого магния, 10 мМ дитиотреитола, 1 мМ АТФ, 25 мкг/мл бычьего сывороточного альбумина. К 5 мкг ДНК в объеме 20 мкл добавляют 5 ед. ДНК полимеразой I фага Т4, после чего пробу инкубируют в течение 8 ч при 16°С. и лигируют с плазмидой pUC19-PEP4, обработанной рестриктазой EcoRV. Лигазной смесью трансформируют клетки штамма DH5α Escherichia coli (F'/endAl hsdR17 (r
Полученным линейным фрагментом плазмиды pADP1 размером 4,33 т.п.о. трансформируют клетки штамма 60-Д578 для получения штамма 1-60-Д578 с нарушенным геном РЕР4.
Для получения штамма 60-Д578 скрещивают штаммы 33-Д373 (MAT α pheAl0 ade2-144,717 his7-1 lys9-A21 urа3-52 trpl-289 leu2-3,112) и 5-Д502 (МАТа arg6 pho3) Петергофской генетической коллекции. Полученный диплоид растет на минеральной среде без добавления аминокислот. Диплоид переносят на среду с ацетатом натрия (1% ацетат натрия, 0,5% хлористый калий) и через 4-5 дней спорулирующую культуру используют для изоляции гаплоидных спор. Спорулирующую культуру обрабатывают медицинским эфиром в течение 5 мин и высевают на полную среду YEPD. Выросшие на среде YEPD споры пересевают на полную среду и через 2 суток методом отпечатков переносят на селективные среды, определяют фенотип сегрегантов и отбирают штамм 60-Д578, содержащий мутации ade2-144,717, leu2-3,112, lys9-A21, arg6, pho3.
Для целенаправленного нарушения гена РЕР4 штамм 60-Д578 трансформируют линейным фрагментом плазмиды pADP1 размером 4,33 т.п.о., который содержит ген РЕР4 со встроенным геном ADE2. Клетки дрожжей штамма 60-Д578 выращивают в 100 мл среды YEPD до достижения культурой оптической плотности, соответствующей 2-4 ед. поглощения при длине волны 600 нм. Клетки дважды промывают стерильной водой, после чего суспендируют в 0,3 мл 100 мМ раствора ацетата лития и инкубируют при 30° в течение 30 мин. К 50 мкл полученной суспензии клеток добавляют 0,1-1 мкг плазмидной ДНК, 50 мкг ДНК спермы лосося, предварительно денатурированной нагреванием (10 мин при 100°), и 0,3 мл раствора 100 мМ ацетата лития, содержащего 40% полиэтиленгликоля 4000. Далее пробу инкубируют 30 мин при 30° и 20 мин при 42°, помещают на 15 с в ледяную баню и центрифугируют 10 с при 10000 об/мин. Клетки суспендируют в 1 мл стерильной воды и высевают на твердую среду SC, содержащую содержащих 30 мг/л лизина, 30 мг/л аргинина, 60 мг/л лейцина. Трансформанты отбирают по способности расти на среде, не содержащей аденина. Клоны трансформантов вырастают через 4-6 суток.
Нарушение гена РЕР4 проверяют по снижению активности репрессибельной щелочной фосфатазы на среде ПЕП. Факт интеграции PEP4::ADE2 в хромосому доказывают, при помощи ПЦР, используя праймеры к гену РЕР4. В качестве матрицы используют хромосомную ДНК трансформантов.
Затем полученным штаммом 1-60-Д578 с нарушенным геном РЕР4 создают штамм - продуцент интерлейкина-2 человека.
Для получения штамма дрожжей Saccharomyces cerevisiae 1-60-Д578 (MSIL) - продуцента интерлейкина-2 человека, клетки дрожжей штамма 1-60-Д578 трансформируют плазмидой pJDB(MSIL). Клетки дрожжей выращивают в 100 мл среды YEPD до достижения культурой оптической плотности, соответствующей 2-4 ед. поглощения при длине волны 600 нм. Клетки дважды промывают стерильной водой, после чего суспендируют в 0,3 мл 100 мМ раствора ацетата лития и инкубируют при 30° в течение 30 мин. К 50 мкл полученной суспензии клеток добавляют 0,1-1 мкг плазмидной ДНК, 50 мкг ДНК спермы лосося, предварительно денатурированной нагреванием (10 мин при 100°), и 0,3 мл раствора 100 мМ ацетата лития, содержащего 40% полиэтиленгликоля 4000. Далее пробу инкубируют 30 мин при 30° и 20 мин при 42°, помещают на 15 с в ледяную баню и центрифугируют 10 с при 10000 об/мин. Клетки суспендируют в 1 мл стерильной воды и высевают на твердую среду SC, содержащую 30 мг/л лизина, 30 мг/л аргинина. Клоны трансформантов вырастают через 4-6 суток.
Для анализа продукции интерлейкина-2 человека клетками трансформантов их выращивают в 50 мл жидкой среды ПЕП до стационарной фазы роста. Клетки собирают центрифугированием при 3000 об/мин в течение 10 мин, промывают водой, суспендируют в 1 мл 50 мМ трис-хлоридного буфера (рН 7,5), содержащего 1 мМ ФМСФ (фенилметилсульфонилфторид), добавляют 1 г стеклянных бус (0,5 мм) и разрушают в дезинтеграторе “Braun” в течение 1 мин. Полученный гомогенат центрифугируют при 10000 об/мин в течение 10 мин, осадок промывают 10-кратным объемом того же буфера и суспендируют в 1 мл 50 мМ натрийфосфатного буфера (рН 7,0), содержащего 2% додецилсульфата натрия и 5% 2-меркаптоэтанола, и инкубируют 5 мин в кипящей водяной бане. По окончании инкубации пробу центрифугируют 15 мин при 12000 об/мин и в супернатанте определяют содержание интерлейкина-2 человека при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия и последующей гибридизации со специфическими антителами к интерлейкину-2 человека. Разделение белков проводят в 15% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ трис, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8). Параллельно проводят разделение белков контрольного штамма 1-60-Д578, выращенного и вскрытого в идентичных условиях. В качестве стандартов молекулярной массы используют карбоангидразу (31,0 кДа), ингибитор трипсина (21,5 кДа), миоглобин (16,9 кДа), лизоцим (14,4 кДа). По окончании электрофореза белки ренатурируют, выдерживая гели 15 мин в 10 мМ трис-хлоридном буфере (рН 7,5), содержащем 4М мочевину, 20 мМ ЭДТА, и переносят на нитроцеллюлозную мембрану в 25 мМ трис-192 мМ глициновом буфере (рН 8,3), содержащем 20% метилового спирта, при 30-40 В, в течение 1,5 ч. Далее мембрану выдерживают в буфере TBST (10 мМ трис-хлоридный буфер (рН 8,0), 150 мМ хлористого натрия, 0,05% твин-20, 1% бычьего сывороточного альбумина) в течение 2 ч при 37°. Затем помещают мембрану в тот же буфер, содержащий разведенные в 1000 раз мышиные моноклональные антитела к интерлейкину-2 человека (Boehringer), и инкубируют 2 ч при 37°. Далее трижды промывают мембрану буфером TBST и инкубируют 1 ч при 37° с разбавленным в 7000 раз конъюгатом видоспецифических антител к иммуноглобулинам мыши и пероксидазы хрена (Протеиновый контур, Санкт-Петербург). После отмывки мембраны буфером PBST (58 мМ двузамещенного фосфата натрия, 17 мМ однозамещенного фосфата натрия, 68 мМ хлористого натрия, 0,1% твин-20) добавляют раствор субстратов для пероксидазы: 0,02% DАВ(3,'3-диаминобензидин тетрагидрохлорид), 0,006% перекись водорода в 10 мМ трис-хлоридном буфере, рН 7,5. Параллельно окрашивают гели 0,15% раствором кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и отмывают в 10% уксусной кислоте. При сравнении спектра белков у штамма 1-60-Д578 (MSIL) обнаруживают появление дополнительной белковой полосы, дающей положительную реакцию с антителами к интерлейкину-2 человека, с молекулярной массой около 15,4 кДа, что соответствует молекулярной массе интерлейкина-2. Уровень синтеза интерлейкина 2 определяют, сравнивая интенсивность окрашивания полосы рекомбинантного белка с полосой стандартного интерлейкина-2 человека.
Согласно полученным данным, клетки дрожжей штамма 1-60-Д578 (MSIL) синтезируют около 60-65 мг интерлейкина-2 на литр культуры дрожжей.
Рекомбинантный интерлейкин-2 человека, синтезированный в клетках дрожжей, обладает биологической активностью, обеспечивая пролиферацию интерлейкин-2-зависимых Т-лимфоцитов мыши линии CTLL-2 (Gearing A., Thorpe R.J. Immunol. Meth., 1988, v.114, p.3-9). Активность рекомбинантного интерлейкина-2 составляет не менее 10 млн ед./л.
Суммируя вышесказанное, можно заключить, что преимуществом созданного штамма дрожжей Saccharomyces cerevisiae 1-60-Д578 (MSIL) по сравнению с прототипом ВКПМ Y-791 является более высокий уровень продукции интерлейкина-2 человека и большая стабильность рекомбинантного белка, что позволяет повысить выход целевого продукта.
Штамм дрожжей Saccharomyces cerevisiae 1-60-Д578 (MSIL) - продуцент интерлейкина-2 человека депонирован во Всероссийской коллекции промышленных микроорганизмов под номером ВКПМ Y-3079.
Изобретение относится к области генной инженерии и биотехнологии и может быть использовано в микробиологической и медицинской промышленности. Путем скрещивания штаммов 33-Д373 (МАТ α pheA10 ade2-144,717 his7-1 lys9-A21 ura3-52 trp1-289 leu2-3,112) и 5-Д502 (МАТа arg6 pho3) получают штамм 60-Д578, который затем трансформируют полученным из плазмиды pADP1 линейным фрагментом гена РЕР4, инактивированного встройкой гена ADE2. Использование предложенного штамма дрожжей Saccharomyces cerevisiae 1-60-Д578 обеспечивает высокий уровень продукции интерлейкина-2 человека. 2 с.п. ф-лы, 1 ил.
| RU 1830945 A1, 10.08.99 | |||
| Рекомбинантная плазмидная ДНК pJDB (MSIL), обеспечивающая синтез интерлейкина-2 человека в клетках дрожжей SасснаRомUсеS ceReUISIaL, способ ее получения и штамм дрожжей SасснаRомYсеS ceReUISIaL - продуцент интерлейкина-2 человека | 1988 |
|
SU1770359A1 |
| Способ прошивки трубной заготовки на трубопрокатном стане | 1961 |
|
SU142268A1 |
| СТИМУЛЯТОР РОСТА И ПРОДУКТИВНОСТИ ГУСЕНИЦ ТУТОВОГО ШЕЛКОПРЯДА | 0 |
|
SU152358A1 |
| WINGFIELS JM, DECKINSON JR Appl | |||
| Microbiol | |||
| Biotechnol., 36(6), 1992. | |||

