Изобретение относится к биохимии, в частности к применению известных бромпроизводных бензотетразин-1,3-диоксида общей формулы I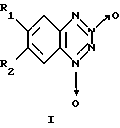
где, если R1=Н, то R2=Вr, или, если R1=Вr, то R2=Н,
в качестве необратимых ингибиторов Н,К-аденозинтрифосфатазы (Н,К-АТРазы).
Н, К-Аденозинтрифосфатаза (ЕС. 3.6.1.3, калий-протонтранспортирующая АТФ-гидролаза) является ферментом, который обеспечивает секрецию соляной кислоты в желудке позвоночных посредством АТФ-зависимого обмена внутриклеточных протонов на внеклеточные ионы калия. Увеличение активности данного фермента в результате патологических изменений в желудке или двенадцатиперстной кишке сопровождается повышением секреции соляной кислоты, что является одной из главных причин возникновения гастроэнтерологических заболеваний (например, язвенной болезни).
Известен ряд производных 2-сульфинилбензимидазола общей формулы II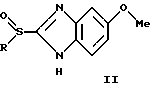
где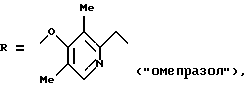
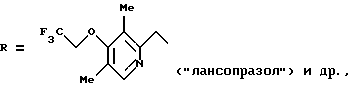
и других, входящих в качестве активных субстанций в фармпрепараты, применяемые в настоящее время для лечения язвы желудка и двенадцатиперстной кишки (Справочник Видаль, ЗАО "Астра Фарм-Сервис", М., 1999 г., с. Б-467, Б-342). Механизм фармакологического действия данных соединений основан на их молекулярной перегруппировке, протекающей в кислых условиях с образованием реакционноспособного сульфенимидного интермедиата, который химически взаимодействуете SH-группами Н,К-АТРазы, что приводит к необратимому ингибированию активности фермента (Р. Lindberg, P. Nordberg et al. "The mechanism of action of the gastric acid secretion inhibitor omeprazole". J. Med. Chem., 1986, v. 29, p. 1327-1329).
Необходимость проведения преинкубации в сильнокислой среде с последующей нейтрализацией значительным количеством буфера затрудняет применение вышеуказанных соединений в биохимических экспериментах.
Запатентованы замещенные пирроло[2,3-d]пиридазин-5,6-диоксиды общей формулы III
где A, R1-R5, X имеют указанные в патенте значения,
в качестве средств для лечения язвы желудка, вызванной Helicobacter pylori (Европейский патент 742218, СО7 D 487/04, oп. 1996 г.). Биохимические свойства вышеуказанных диоксидов не изучены.
Известны производные 1,2,5-оксадиазоло[3,4-d] пиридазин-5,6-диоксида общей формулы IV
где n=0 или 1,
в качестве специфических регуляторов активности нуклеотидзависимых ферментов (патент РФ 2130490, C12N 9/88, 9/99, 1997 г.)
Соединение вышеуказанной общей формулы IV, где n=0, не оказывало значительного влияния на активность Н,К-АТРазы, а его структурный аналог, где n=1, являлся донором оксида азота и вызывал активацию фермента.
Известны производные хиноксалин-1,4-диоксида общей формулы V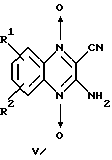
где R1 и R2 (независимо) = Н, Cl, F, CF3, СН3О и др.,
обладающие цитотоксическим действием и активирующиеся в условиях гипоксии (патент Великобритании N 2297089, CO7D 241/52, A61K 31/495, oп. 1996 г.). Влияние данных соединений на активность Н,К-АТРазы не изучено.
Известны производные 1,2,4-бензотриазин-1,4-диоксида общей формулы VI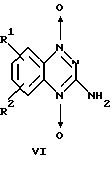
где R1=R2=H (тирапазамин) или R1 и R2 (независимо) = Н, СН3О, С1 и др.,
обладающие цитотоксическим действием и активирующиеся в условиях гипоксии (Е. М. Zeman, М. A. Baker et al. "Structure-activity relationships for benzotriazine di-N-oxides". Int., J. Radiat. Oncol. Biol. Phys., 1989, v. 16, p. 977-981). Влияние данных соединений на активность Н,К-АТРазы не изучено.
Известны нитропроизводные бензотетразин-1,3-диоксида общей формулы VII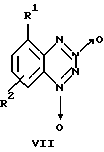
где R1=H и R2=NO2 или R1=NO2 и R2=H или NO2,
в качестве активаторов растворимой формы гуанилатциклазы (патент РФ 2123526, C12N 9/99, 9/88, 1997 г.). Влияние данных соединений на активность Н,К-АТРазы не изучено.
Известны бромпроизводные бензотетразин-1,3-диоксида вышеуказанной общей формулы I, где R1, R2 и R3 имеют соответствующие значения, в качестве продуктов химического синтеза (О.Ю. Смирнов, Дис. канд. хим. наук, ИОХ РАН им. Н. Д. Зелинского, М., 1995 г.).
Наиболее близким к бромпроизводным бензотетразин-1,3-диоксида общей формулы I по действию и структуре является 5,5'-дитиобис(2-нитробензойная) кислота (ДТНБ) формулы VIII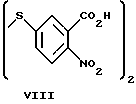
являющаяся модификатором SH-групп и необратимо ингибирующая Н,К-АТРазу (J. J. Schrijen, W. А. Н. М. Van Groningen-Luyben et al. "Studies on (K++H+)-ATPase. II. Role of sulrhydryl groups in its reaction mechanism". Biochim. Biophys. Acta, 1981, v. 640, p. 473-486 - прототип).
Недостатком данного соединения является неселективность его действия по отношению к другим ферментам (в частности, по отношению к Mg2+-АТРазе, Са2+ -АТРазе), а также сравнительно невысокая эффективность ингибирования.
Целью настоящего изобретения является выявление нового ингибитора Н,К-АТРазы с повышенной ингибирующей способностью и более селективным действием.
Поставленная цель достигается применением известных бромпроизводных бензотетразин-1,3-диоксида вышеуказанной общей формулы I, где R1 и R2 имеют приведенные значения, в качестве необратимых ингибиторов Н,К-АТРазы.
Соединения общей формулы I были получены известным способом - обработкой азотным ангидридом соответствующих бромпроизводных 2-(трет-6утип-NNO-азокси)анилина (О.Ю Смирнов, Дис. канд. хим. наук, ИОХ РАН им Н. Д. Зелинского, М., 1995 г. ) в среде органического растворителя.
Описание данного изобретения содержит 5 примеров, 3 таблицы и 6 рисунков, иллюстрирующих экспериментальный материал.
Пример 1. Ингибирующее действие бромпроизводных бензотетразин-1,3-диоксида (Вr-БТДО) общей формулы I на Н,К-АТРазу.
Препарат фракции микросом, обогащенный Н,К-АТРазой, получали из слизистой оболочки желудка кролика известным способом. Концентрацию белка измеряли способом Лоури в присутствии 1%-ного раствора дезоксихолата натрия. В качестве стандарта использовали бычий сывороточный альбумин.
Для измерения активности Н,К-АТРазы использовалась система сопряженных реакций:
1. АТР+Н2О-->ADP+Н2PO4 -+Н+ (АТРаза),
2. ADP+ФЕП-->АТР+пируват (пируваткиназа, ПК);
3. пируват+NADH-->лактат+NAD+ (лактатдегидрогеназа, ЛДГ).
Реакцию регистрировали по изменению оптической плотности раствора при 340 нм, которое происходит в результате уменьшения концентрации NADH в системе.
Среда определения активности содержала 0,3 М сахарозу, 5 мМ PIPES, 10 мМ Трис, рН 7,4, 3 мМ MgCl2, 1,5 мМ ФЕП, 3 мМ АТР, 0,2 мМ NADH, ЛДГ с активностью 10 единиц на пробу и ПК с активностью 15 единиц на пробу. Фракция микросом из слизистой оболочки желудка не обладала активностью NADH-редуктазы. Реакцию начинали добавлением микросом (10-15 мкг белка в пробу объемом 1 мл). Для измерения активности Н,К-АТРазы в пробу добавляли 1 мМ азида натрия, 10 мМ КС1, 5 мкМ СССР и 5 мкМ валиномицина. Для измерения активности Mg-АТРазы в пробу добавляли 0,1 мМ SCH 28080 (ингибитора Н,К-АТРазы).
Измерения проводили на спектрофотометре "Hitachi 200-20" при 37oС. Длина оптического пути составляла 1 см.
Для изучения влияния бромпроизводных бензотетразин-1,3-диоксида на активность Н, К-АТРазы суспензию микросом, содержащую 100 или 150 мкг белка, разводили средой выделения так, чтобы содержание белка в среде преинкубации составляло 10-15 мкг в 50 мкл и оставляли в течение 10 мин при комнатной температуре. Затем добавляли ингибитор и преинкубировали в течение 0,25-45 минут, через определенные промежутки времени отбирая аликвоты по 50 мкл, которые вносили в кюветы для измерения активности. Для измерения действия ингибиторов на Mg-АТРазу в кюветы для измерения активности добавляли 0,1 мМ SCH 28080 (ингибитора Н,К-АТРазы).
Анализ ингибирования (представление процесса ингибирования в виде суммы двух экспонент или одной экспоненты) и расчет констант скоростей ингибирования псевдопервого порядка проводили с использованием компьютерной программы Biochemical Utilites 3.0.
Отсутствие АТРазных активностей, не принадлежащих Н,К-АТРазе (Na,K-АТРазы, Mg-АТРазы, Са-АТРазы), в среде измерения контролировалось добавлением специфических ингибиторов (1 мМ уабаина, олигомицина, 1 мМ ЭДТА соответственно).
Фракцию микросом из слизистой оболочки желудка кролика преинкубировали с избытком изучаемого соединения (молярное соотношение ингибитор/ Н,К-АТРаза обычно превышало 100/1) при комнатной температуре в течение различных промежутков времени, а затем из смеси, содержащей комплекс фермент-ингибитор, отбирали аликвоты объемом 30-50 мкл и переносили их в среду для регистрации активности объемом 1 мл. В результате раствор, содержащий комплекс фермент-ингибитор, разводили в 20-30 раз. После разведения ингибирование Н,К-АТРазы не устранялось. Активность фермента после перенесения его в среду инкубации была линейна во времени в течение (по крайней мере) 15 минут, что свидетельствует об отсутствии медленной диссоциации комплекса фермент-ингибитор. Кроме того, ингибирующее действие бромпроизводных БТДО на активность Н,К-АТРазы развивалось во времени. Полученные данные показывают, что соединения общей формулы I являются необратимыми ингибиторами Н,К-АТРазы.
В описанных условиях бромпроизводные БТДО ингибировали К-активируемую, чувствительную к SCH-28080 АТРазную активность фракции микросом из слизистой оболочки желудка. Mg-АТРазная активность данной фракции, нечувствительная к SCH 28080, также как и азид-чувствительная АТРазная активность, данными соединениями не ингибировались. Таким образом, в описанных условиях бромпроизводные БТДО слективно ингибировали Н,К-АТРазу фракции микросом из слизистой оболочки желудка, не влияя на другие АТРазы данной фракции.
На рис. 1 представлена зависимость активности Н,К-АТРазы от времени преинкубации с различными концентрациями 7-Вr-БТДО (30, 60 и 100 мкМ) при различном молярном избытке ингибитора (50/1, 100/1 и 167/1). Расчеты молярного соотношения фермент/ингибитор проводили исходя из того, что процентное содержание Н, К-АТРазы в исследуемом препарате составляло около 30% от общего количества белка фракции микросом.
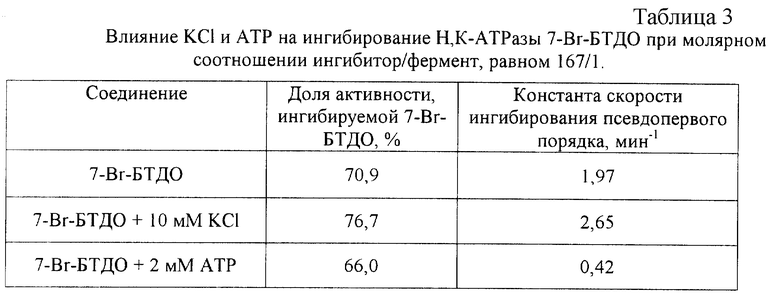
Как показано на рис.2, константы скорости ингибирования псевдопервого порядка, рассчитанные для ингибирования Н,К-АТРазы 7-Вr-БТДО, линейно зависели от концентрации ингибитора. Таким образом, константы скорости ингибирования второго порядка для всех трех соотношений белка и ингибитора были близки и составляли 1,75-2,07 M-1•мин-1 (табл.1).
Известно, что молекула Н, К-АТРазы содержит несколько высокоактивных SH-групп, которые подвергались селективной модификации с помощью известного аналога - ДТНВ. Кинетика ингибирования фермента под действием как бромпроизводных БТДО, так и ДТНБ описывалась моноэкспоненциальной кривой. Согласно данным прототипа при концентрации ДТНБ, равной 100 мкМ, константа скорости ингибирования Н, К-АТРазы псевдопервого порядка имела значение 0,044 мин-1, т. е. 7-Вr-БТДО как необратимый ингибитор данного фермента - в 45 раз более эффективен по сравнению с аналогичными свойствами известного аналога - ДТНБ. АТР защищал Н,К-АТРазу от ингибирования, вызванного как ДТНБ, так и Вr-БТДО.
На основании вышеприведенных данных можно сделать вывод о том, что 7-Вr-БДТО необратимо модифицирует существенные для активности аминокислотные остатки Н,К-АТРазы (по-видимому, SH-группу(ы)), расположенные на АТР-связывающем участке фермента.
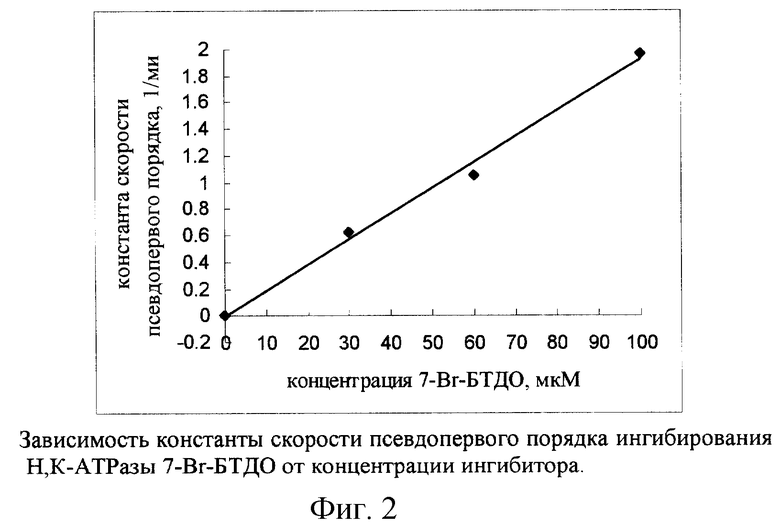
На рис.3 представлена зависимость активности Н,К-АТРазы от времени преинкубации с 6-Вr-БТДО при молярном соотношении ингибитор/фермент 167/1. Кривая, характеризующая данную зависимость, наилучшим образом описывается одной экспонентой (табл. 2). Константа скорости псевдопервого порядка для ингибирования Н, К-АТРазы под действием 6-Вr-БТДО несколько меньше (~ в 1,2 раза), чем под действием 7-Вr-БТДО (табл. 2). Тем не менее, 6-Вr-БТДО как необратимый ингибитор данного фермента ~ в 36 раз более эффективен по сравнению с аналогичными свойствами известного аналога - ДТНБ.
Пример 2. Влияние КС1 и АТР на процесс ингибирования Н,К-АТРазы бромпроизводными бензотетразин-1,3-диоксида общей формулы I.
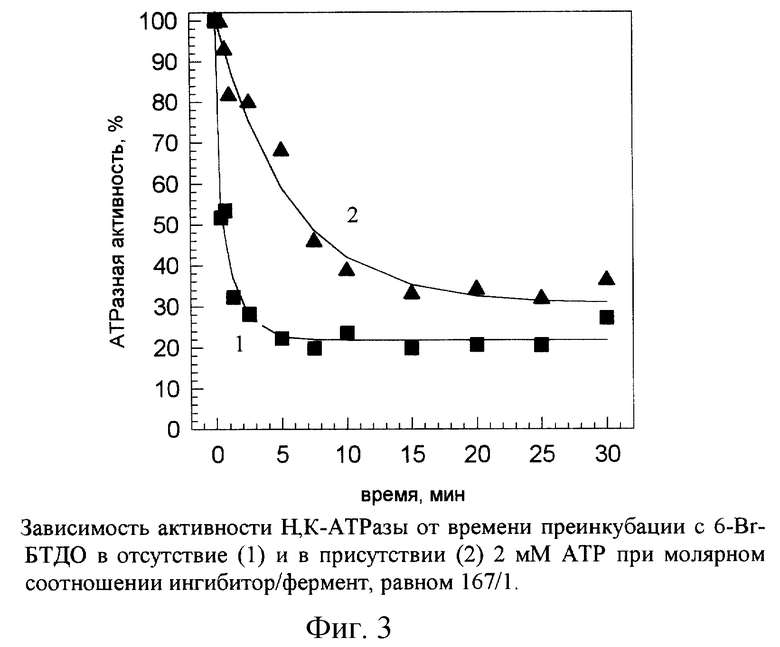
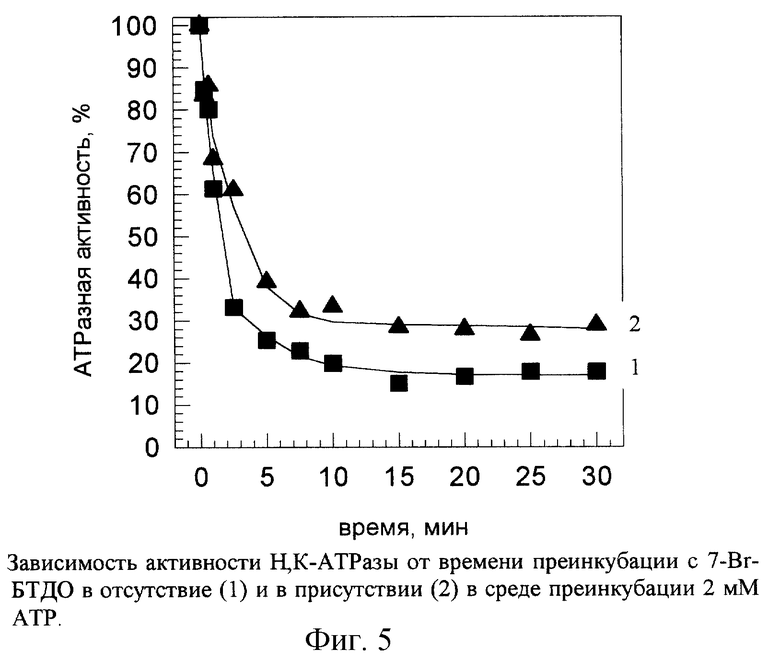
Ионы К+, переносимые через мембрану, принято рассматривать как субстрат или как активатор Н, К-АТРазы. Добавление в среду преинкубации 10 мМ КС1 ускоряло ингибирование данного фермента под действием 7-Вr-БТДО (рис.4, табл.3).
АТР является субстратом Н,К-АТРазы. Присутствие 2 мМ АТР в среде преинкубации, не содержащей КС1, приводило к тому, что скорость ингибирования активности фермента под действием 7-Вr-БТДО снижалась (рис.5): константа скорости ингибирования псевдопервого порядка в присутствии 2 мМ АТР уменьшалась почти в 5 раз (табл. 3). Таким образом, связывание АТР с Н,К-АТРазой существенно снижало способность определенных аминокислотных остатков фермента подвергаться химической модификации под действием 7-Вr-БТДО, что можно рассматривать как "защитный" эффект АТР.
При добавлении в среду преинкубации 2 мМ АТР скорость ингибирования Н, К-АТРазы под действием 6-Вr-БТДО уменьшалась, оставаясь при этом однофазным процессом (рис.3). Константа скорости ингибирования снижалась с 1,6 мин-1 до 0,2 мин-1, доля неингибируемой активности возрастала с 18 до 30%.
Пример 3. Зависимость скорости ингибирования Н,К-АТРазы под действием 7-бромбензотетразин-1,3-диоксида от рН среды.
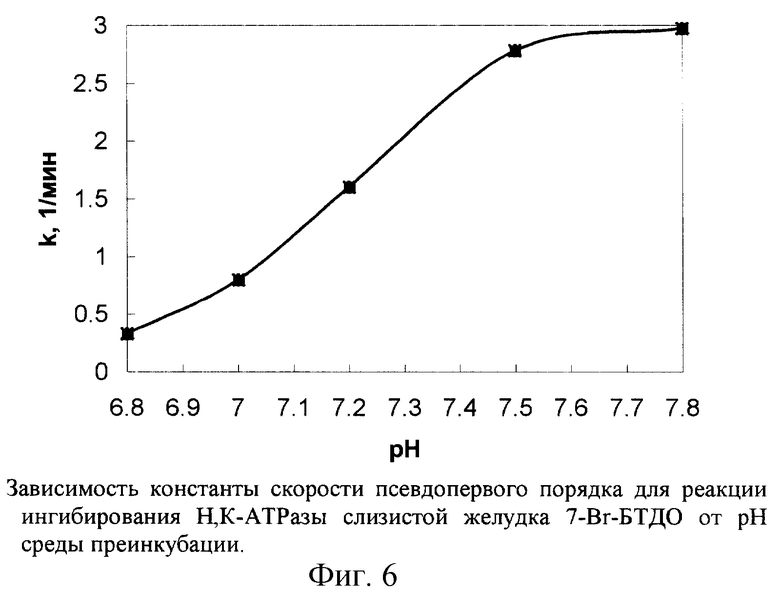
Изучение зависимости необратимого ингибирования фермента от рН среды позволяет с определенной точностью установить константу ионизации модифицируемой реакционноспособной группы белка, что может дать информацию о типе данного аминокислотного остатка.
На рис.6 представлена зависимость константы скорости ингибирования псевдопервого порядка для Н,К-АТРазы под действием 7-Вr-БТДО от рН среды. Данная зависимость имеет вид одной сигмоиды, что характерно для ионизации одного класса модифицируемых групп. Константа ионизации (рКа) аминокислотной группы фермента, участвующей в исследуемой реакции, имеет значение ~7,2, присущее остаткам цистеина или гистидина. Поскольку при близких значениях рН 7-Вr-БТДО практически не реагирует с имидазолом и имидазольной группой гистидина, ингибирование Н,К-АТРазы происходит в результате химического взаимодействия 7-Вr-БТДО, по-видимому, с остатком цистеина.
Пример 4. Изучение влияния 7-бромбензотетразин-1,3-диоксида на Са-АТРазу саркоплазматического ретикулума скелетных мышц кролика.
Мембранный препарат саркоплазматического ретикулума, содержащий 70% Са-АТРазы, получали из скелетных мышц кролика известным способом. Концентрацию белка измеряли по способу Лоури в присутствии 1%-ного раствора дезоксихолата натрия. В качестве стандарта использовали бычий сывороточный альбумин.
Измерение активности Са-АТРазы проводили тем же способом (с использованием системы сопряженных реакций), как и измерение активности Н,К-АТРазы (см. пример 1). Активность Са-АТРазы составляла 6,2 мкмоль/мин•мг белка. В присутствии 0,1 M 7-Вr-БТДО активность фермента не изменялась.
Пример 5. Изучение способности генерировать оксид азота бромпроизводными бензотетразин-1,3-диоксида общей формулы I.
Как указывалось ранее, некоторые гетероциклические N, N'-диоксиды (например, общей формулы IV) являются донорами NO. Известно также, что ингибирующее действие NO-генерирующих соединений на различные ферменты в аэробных условиях может быть опосредовано нитрозилированием функционально важных тиольных групп. Для того, чтобы оценить возможный вклад S-нитрозилирования в наблюдаемый ингибирующий эффект, была изучена способность бромпроизводных БТДО генерировать оксид азота.
Для определения оксида азота использовали известный способ, основанный на реакции оксида азота с кислородом воздуха в водной среде с образованием нитрита, количество которого измеряли по интенсивности окрашивания пробы продуктом реакции азосочетания с помощью спектрофотометра.
Проба конечным объемом 1 мл содержала 50 мМ калий-фосфатный буфер (рН 7,4), 0,5 мМ цистеин или глутатион, изучаемое соединение в концентрации 0,1 мМ и 0,2% диметилсульфоксид (ДМСО). В качестве отрицательного контроля использовали водный раствор ДМСО в концентрации 0,2%, а в качестве положительного контроля 0,1 мМ нитрит натрия, содержащий 0,2% ДМСО. Пробы инкубировали 60 мин при 37oС и добавляли последовательно 100 мкл 3 М ацетата натрия, 400 мкл 0,92% раствора сульфаниловой кислоты в 30% уксусной кислоте и 400 мкл 0,05% N-нафтилэтилендиамина. Пробы инкубировали 10 мин и измеряли оптическую плотность при длине волны 554 нм на спектрофотометре.
В вышеуказанных условиях при рН 7,4 не наблюдалось образование заметного количества нитрита (<< 0,01 моль нитрита/моль исходного соединения) в отсутствие тиолов В присутствии тиолов соединения общей формулы I также не генерировали нитрит. Таким образом, бромпроизводные бензотетразин-1,3-диоксида не являются спонтанными или тиолзависимыми донорами оксида азота.
Как следует из примера 1, соединения настоящего изобретения оказывают более выраженное ингибирующее действие на Н,К-АТРазу слизистой оболочки желудка кролика, чем эффект прототипа. Пример 2 показывает, что АТР снижает константу скорости ингибирования Н,К-АТРазы под действием бромпроизводных БТДО, тогда как ионы К+ ее несколько увеличивают. Из примера 3 следует, что, согласно характеру зависимости процесса ингибирования от рН среды инкубации и реакционной способности бромпроизводных бензотетразин-1,3-диоксида, модифицируются SH-группа (группы) фермента. Пример 4 показывает селективность ингибируюшего действия соединений общей формулы I на Н,К-АТРазу. Таким образом, применение указанных бромпроизводных бензотетразин-1,3-диоксида расширяет ассортимент специфических необратимых ингибиторов Н,К-АТРазы.
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕОБРАТИМЫЙ ИНГИБИТОР Ca-АДЕНОЗИНТРИФОСФАТАЗЫ САРКОПЛАЗМАТИЧЕСКОГО РЕТИКУЛУМА | 2001 |
|
RU2194756C1 |
| ИНГИБИТОР АГРЕГАЦИИ ТРОМБОЦИТОВ, ОБЛАДАЮЩИЙ АНТИМЕТАСТАТИЧЕСКИМ ДЕЙСТВИЕМ | 2001 |
|
RU2192857C1 |
| ПРОИЗВОДНЫЕ АНТРА[2,1-d]ИЗОТИАЗОЛ-3,6,11-ТРИОНА | 2002 |
|
RU2213744C1 |
| СПОСОБ ИНГИБИРОВАНИЯ КАСПАЗ | 2002 |
|
RU2233326C1 |
| ПРОИЗВОДНОЕ ПИРИДОПИРАЗИНДИОНА И ЕГО ПРИМЕНЕНИЕ В КАЧЕСТВЕ ПРОТИВОЯЗВЕННОГО СРЕДСТВА | 2010 |
|
RU2465274C2 |
| СПОСОБ ПРЯМОГО ИНГИБИРОВАНИЯ КАСПАЗ | 2002 |
|
RU2232809C1 |
| СПОСОБ ИНГИБИРОВАНИЯ КОРРОЗИИ И КОМПОЗИЦИИ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2001 |
|
RU2198245C2 |
| ПРОИЗВОДНЫЕ ТЕТРАФУРАЗАНО[3,4-b:3',4'-f:3",4"-j:3'",4'"-N][1,4,5,8,9,12,13,16]ОКТААЗАБ ИЦИКЛО[14.2.2]ЭЙКОЗА-4,8,12-ТРИЕНА И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2000 |
|
RU2167161C1 |
| АКТИВАТОР РАСТВОРИМОЙ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ | 1997 |
|
RU2123526C1 |
| ПРОИЗВОДНЫЕ (1,2,3-ТРИАЗОЛИЛ)-1,2,5-ОКСАДИАЗОЛА, ПОТЕНЦИРУЮЩИЕ NO-ЗАВИСИМУЮ АКТИВАЦИЮ РАСТВОРИМОЙ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ | 2000 |
|
RU2158265C1 |


Изобретение относится к биохимии, в частности к применению известных бромпроизводных бензотетразин-1,3-диоксида общей формулы I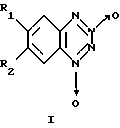
где R1 и R2 различны и означают водород или Вr,
в качестве необратимых ингибиторов Н, К-аденозинтрифосфатазы (Н,К-АТРазы). Новый ингибитор Н,К-АТРазы обладает повышенной ингибирующей способностью и более селективным действием. Бромпроизводные бензотетразин-1,3-диоксида (Вr-БТДО) необратимо селективно ингибируют Н,К-АТРазную активность фракции микросом из слизистой оболочки желудка. 6- и 7-Вr-БТДО как специфические необратимые ингибиторы Н,К-АТРазы более эффективны по сравнению с аналогичным действием 5,5'-дитиобис(2-нитробензойной)кислоты. 6 ил., 3 табл.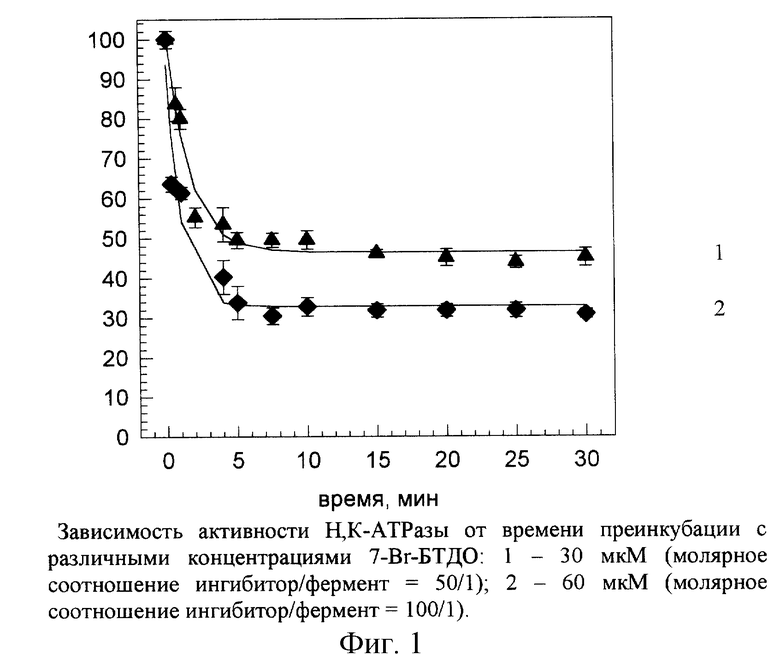
Применение бромпроизводных бензотетразин-1,3-диоксида общей формулы I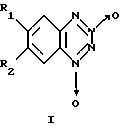
где, если R1= Н, то R2= Вr, или, если R1= Вr, то R2= Н,
в качестве необратимых ингибиторов Н, К-аденозинтрифосфатазы.
| СМИРНОВ О.Ю | |||
| Диссертация канд | |||
| хим | |||
| наук, ИОХ РАН им | |||
| Н.Д | |||
| Зелинского | |||
| - М., 1995 г | |||
| Устройство для автоматической расцепки железнодорожного подвижного состава | 1978 |
|
SU742218A1 |
| СПЕЦИФИЧЕСКИЙ РЕГУЛЯТОР АКТИВНОСТИ НУКЛЕОТИД-ЗАВИСИМЫХ ФЕРМЕНТОВ | 1997 |
|
RU2130490C1 |
| АКТИВАТОР РАСТВОРИМОЙ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ | 1997 |
|
RU2123526C1 |

