Изобретение относится в основном к получению и использованию иммуноглобулиновых полипептидов, которые тормозят опосредованную PDGF пролиферацию клеток, воспроизводящих бета-рецептор фактора роста, произведенного из тромбоцитов (PDGF), человеческого типа.
Предпосылки изобретения
Фактор роста, полученный из тромбоцитов (PDGF), является сильным пролиферативным агентов в клетках мезенхимального происхождения (Antoniades, H. N. и др. (1979) Proc. Natl. Acad. Sci. USA 76: 1809-13; Bowen-Pope, D.F. и Ross, R. (1982) J. Bion. Chem. 257:5161-71; Heldin, C-H. и др. (1983) J. Biol. Chem. 258:10054-59). PDGF (ММ 30 кДа) представляет собой дисульфидсвязанный димер, состоящий из 2 гомологичных цепей, обозначенных А или В (Johnson, A. и др. (1982) Biochim. Biophys. Res. Commun. 104:66-74). Цепи могут объединяться с цепями того же или другого типа, что дает три изоформы АА, ВВ или АВ (Heldin, С-Н. и др. (1986) Nature 319:511-514). Митоген PDGF был впервые идентифицирован (Antoniades, H.N. (1979) Proc. Natl. Acad. Sci. USA 76:1809-13; Raines, E.W. и Ross, R. (1982) J. Biol. Chem. 257:5154-5160) и очищен от человеческих тромбоцитов (Raines, op. cit.), хотя последующий поиск показал, что несколько типов клеток, включающих васкулярные эндотелиальные клетки, васкулярные клетки гладкой мускулатуры, макрофаги и даже фибробласты синтезируют PDGF (Ross, R. и др. (1986) Cell 46:155-189).
Клеточная полиферация, вызванная всеми изоформами PDGF, осуществляется через посредство лигандной связи с рецептором PDGF (Heldin, C-H. (1983) op. cit., Ek, B. и др (1982) Nature 295:419-420; Glenn, K. и др. (1982) J. Biol. Chem. 257:5172-76; Frackelton, A. R. и др. (1984) J. Biol. Chem. 259:7909-15; Williams, L. T. и др. (1984) J. Biol. Chem. 259:5287-94). Рецептор PDGF (ММ 180 кДа) принадлежит к семейству тирозинкиназы и состоит из двух подтипов рецептора, обозначенных тип А (или тип альфа) (Matsui, T. и др. (1989) Science 243:800-804 и Claesson - Welsh, L. (1989) Proc. Natl. Acad. Sci. USA 86: 4917-4921) или тип В (или тип бета) (Yarden. Y. и др. (1986) Nature 323: 226-232 и Escobedo, J.A. и др. (1988) Science 240:1532-1534).
Сильное сродство связи PDGF с рецептором сопровождается димеризацией рецептора (Bishayee, S. и др. (1989) J. Biol. Chem. 264:11699-11705, Heldin, C-H. и др. (1989) J. Biol. Chem. 264:8905-8912) и аутофосфорилированием (Frackelton и др. op.cit.) и приводит к сложной серии внутриклеточных сигнальных событий, заканчивающихся в конечном итоге синтезом ДНК. Были клонированы гены бета-рецептора и альфа-рецептора человеческого и мышиного PDGF (Matsui и др. op. cit., Claesson-Welsh и др. op. cit., Yarden и др. op. cit. и Escobedo и др. op.cit.). Здесь рецепторы PDGF - тип А и тип альфа используют взаимозаменяемым образом, как тип В и тип бета.
Две изоформы рецепторов можно различить по их достаточно отличной специфичности связывания лиганда. Бета-рецептор PDGF связывает только В-цепь (изоформы ВВ и АВ), в то время как альфа-рецептор PDGF может связывать все формы PDGF (изоформы, содержащие А и/или В цепь (Matsui и др. op. cit., Claesson-Welsh и др. op. cit. и Seifert, R.A. и др. (1989) J. Biol. Chem. 264: 8771-8778). Рецептор PDGF проявляет высокую степень структурной гомологии к рецептору фактора стимуляции колонии макрофагов (Coussens, L. и др. (1986) Nature 320:277-280) и с-кит протоонкогенному продукту (Yarden и др. op. cit. ).
Рецепторы PDGF характеризуются внешнеклеточным доменом, который можно разграничить на пять Ig - подобных доменов (Домены 1-5) на основании их структуры, богатой β-пластом. Эти Ig повторы приблизительно в 100 аминокислот каждый имеют правильно распределенные цистеиновые остатки (за исключением четвертого повтора). Рецептор имеет один трансмембранный домен и цитоплазматический домен тирозин киназы (Williiams, L.T. и др. (1989) Science 243:1564-1570).
PDGF играет важную роль в нормальных физиологических процессах, таких как восстановление ткани и эмбриогенез (Ross, R. и др. op.cit.). Однако сейчас изучения предполагают, что этот сильный митоген участвует в патологических пролиферативных нарушениях и в развитии некоторых карцином (Ross, R. и др. op. cit.). Экспрессия цепи А PDGF и бета-рецептора PDGF была обнаружена в человеческих атеросклеротических бляшках гибридизацией in situ (Wilcox, J.N. и др. (1988) J. Clin. Invest. 82:1134-1143). Недавно появилось сообщение, Ferns и др. (1991) Scince 253:1129-1132), что поликлональное антитело к PDGF значительно снизило образование интимных повреждений в деэндотелиализированных сонных артериях атимических крыс. Предполагается, что PDGF замешан в патологии пролиферативных заболеваний в клетках мезенхимного происхождения (Nister, M. и др. (1984) Proc. Natl. Acad. Sci. USA 81:926-930, и Nister, M. и др. (1987) Cancer Res. 47:4953-4961). Golden и др. установили, что информация А цепи увеличивалась в участках интимальной гиперплазии у павиана для васкулярных трансплантатов ((1990) J. Vasc. Surg. 11: 580-585). PDGF также хемотаксичен для гладкой мускулатуры (Westermark, B. и др. (1990) Proc. Natl. Acad. Sci. USA 87:128-132), и PDGF тромбоцитов может быть причинным агентом для миграции и пролиферации клеток гладкой мускулатуры в сонной артерии крысы с балонным катетером, что приводит к значительному стенозу.
Изучение других факторов роста и их рецепторов продвинулось с изобретением антител против рецепторов. Например, антитела, которые опознают рецептор фактора эпидермального роста, как доказано, являются мощными инструментами в оценке механизма активации рецептора (Spaargaren, M. и др. (1991) J. Biol. Chem. 266:1733-1739). Антитела против рецепторов для интерлейкина-2 (IL-2) тормозят интернализацию IL-2 и таким образом тормозят последующую индукцию пролиферации ответных клеток (Dupre, V. и др. (1991) J. Biol. Chem. 1497-1501). Таким же образом моноклональное антитело против рецептора фактора эпидермального роста (EGF) тормозит стимулируемый эстрогеном рост клеточной линии МСF-7 аденокарциномы молочной железы человека (Eppstein, D.A. (1989) J. Cell. Physiol. 141:420-430). Такие антитела могут быть очень важны в лечении заболеваний, связанных с фактором роста.
Несколько групп имеют изолированные антитела против рецепторов PDGF, но эти антитела ограничены в своем применении (см., например, Kawahara, R.S. и др. (1987) Biochem, Biophys. Res. Commun. 147:839-845). Дополнительные моноклональные антитела против внеклеточного PDGF-связующего домена рецептора PDGF были получены из матки свиньи (Ronnerstrand. L. и Terracio, L. (1988) J. Biol. Chem. 263:10429-10435), но эти антитела не тормозят связывания PDGF с меткой 125I с фибробластами человека. О многочисленных антителах против рецептора PDGF, которые не тормозят активности PDGF, также были публикации Kanakaraj, P.S. и др. (1991) Biochemistry 30:1761-1767; Claesson-Welsh и др. (1989) J. Biol. Chem. 264:1742-1747; Seifert, R.A. и др. (1989) J. Biol. Chem. 264: 8771-8778; Kumjian, D.A. и др. (1989) Proc. Natl. Acad. Sci. USA 86:8232-8236; Bishayee и др. (1988) Mol. Cell. Biol. 8:3696-3702; Hart, C.E. 262: 10780-10785 и др. (1987) J. Biol. Chem. 262:10780-85; Escobedo. J.A. и др. (1988) J. Biol. Chem. 263:1482-1487; Daniel T.O. и др. (1987). J. Biol. Chem 262:9778-9784; Keating, M. T. и Williams L.T. (1987) J. Biol. Chem 262: 7932-7037; Kaslauskas, A. и J.A. Copper (1990), EMBO J. 9:3279-3286.
Таким образом, существует необходимость найти иммуноглобулин и другие агенты, способные специфическим образом тормозить активацию человеческого рецептора и/или пролиферацию клеток, воспроизводящих бета-рецептор PDGF человеческого типа. Такие агенты были бы полезны в картировании различных функциональных доменов рецептора и в разделении роли PDGF и его рецепторов в нормальных процессах и процессах заболевания.
Кроме того, такие агенты имели бы терапевтическую ценность в лечении пролиферативных заболеваний с участием PDGF, а также заболеваний, вовлекающих хемотаксис и миграцию с участием PDGF. Такие заболевания включают:
а) рестеноз, включая коронарный рестеноз после ангиопластики, атерэктомии или других инвазивных методов удаления бляшек, и почечный рестеноз или рестеноз периферических артерий после тех же процедур;
в) сосудистые пролиферативные явления и фиброз, связанный с другими формами острых повреждений, таких как легочный фиброз, связанный с синдромом респираторной патологии, почечный фиброз, связанный с нефритом, коронарный стеноз, связанный с болезнью Кавасаке, и сосудистые сужения, связанные с другими артериитами, такими как синдром Такаясу;
с) предотвращение сужений в венозных трансплантатах;
d) предотвращение сужений вследствие ускоренной миграции клеток гладкой мускулатуры и пролиферации в трансплантированных органах;
е) другие фиброзные процессы, такие как миофиброз; и
f) торможение пролиферации опухолевых клеток, которая происходит с участием PDGF.
Настоящее изобретение решает эти и другие задачи.
Содержание изобретения
Настоящее изобретение касается иммуноглобулиновых полипептидов, которые специфически связываются с бета-рецептором фактора роста, полученным из тромбоцитов человеческого типа (β PDGF-R), и связывание полипептида иммуноглобулина имеет следующие последствия (одно или несколько):
i) торможение PDGF ВВ связывания с рецептором;
ii) торможение индуцированного PDGF фосфорилирования β PDGF-R;
iii) торможение индуцированной PDGF димеризации β PDGF-R;
iv) торможение вызванного PDGF митогенеза клеток, воспроизводящих человеческий β PDGF-R; и
v) хемотаксиса и миграции клеток, воспроизводящих β PDGF-R, вызванных PDGF.
Предпочтительным вариантом изобретения является моноклональное антитело, такое как моноклональное антитело 2А1E2, которое представляет собой изотип IgG1.
Изолированные нуклеиновые кислоты, имеющие последовательность, практически идентичную кислотам, кодирующим для всего или части иммуноглобулинового полипептида, обладающего описанными свойствами, также включены в изобретение. Клеточная линия, транфицерованная, трансформированная или зараженная этими нуклеиновыми кислотами - другой вариант изобретения, так же, как способ получения иммуноглобулинового полипептида или его фрагментов новообразованием клеточной линии, содержащей заявленные нуклеиновые кислоты, и собиранием иммуноглобулиновых полипептидов или фрагментов.
Иммуноглобулиновые полипептиды изобретения имеют диагностическое, а также терапевтическое применение. Например, другой аспект изобретения включает методы лечения человека с заболеванием, связанным с пролиферацией клеток гладкой мускулатуры с вовлечением PDGF, назначением пациенту терапевтически эффективной дозы иммуноглобулинового полипептида настоящего изобретения.
Краткое описание чертежей
Фиг.1 Конструкция внеклеточного домена рекомбинантного человеческого бета-рецептора. Мутагенез делеции бета-рецептора полной длины был проведен, как описано в способах. РΔI относится к 5 внеклеточному домену бета-рецептора PDGF (аминокислоты I-499), который был получен делецией от кДНК бета-рецептора типа PDGF, кодоны для остатков аминокислот 500-1074 с использованием олигонуклеотида GTG TGA GGA ACG GGA AAT TCA TCG ACG GAC ATC CCC CGAC (последовательноать ID :1). РΔ2 относится к 4 внеклеточному домену рецептора PDGF (ак 1-384), который был получен делецией кодонов доя остатков аминокислот 385-1074 от кДНК с использованием олигонуклеотида GGA AGG TCG ATG TCT AGT TAA TCG ACG GAC ATC ССС CGAC (послед. ID 2). Предполагаемые Ig домены указаны следующим образом: D1 (ак 1-91), D2 (ак 92-181), D3 (ак 182-282), D4 (ак 283-384) и D5 (ак 385-499). Детерминанта пептида поликлональных антисывороток 1-3-5 указана выше.
Фиг. 2 А. Вестерн-блот выделенного внеклеточного бета-рецептора pΔ1-5 PDGF, 5 домен. Восстановленный (ряды 1-3) и невосстановленный (ряды 4-6) выделенный внеклеточный домен бета-рецептора PDGF (РΔ1-5, 5 μг/ряд) был подвергнут электрофорезу на 7% геле Лемли с последующим вестерн-переносом, как описано в способах. Нитроцеллюлоза была блокирована в физрастворе с фосфатным буфером (РBS), содержащем 2% молока, нарезалась на полоски и инкубировалась всю ночь при 4oС 60 μг/мл либо моноклональным антителом (МАb) 2А1Е2 (ряды 1 и 4), другим моноклональным антителом (1С7D5) бета-рецептора PDGF (ряды 2 и 5) либо неспецифичным моноклональным антителом (ряды 3 и 6). Нитроцеллюлозные полоски промывались РBS, содержащим 0,5% молока и 0,1% T Ween 20, инкубировались белком А с пометкой 125I 2 часа при комнатной температуре и подвергались рентгену. Стрелка указывает позицию рΔ1-5.
В. Иммуноосаждение выделенного внеклеточного бета-рецептора PDGF рΔ2-7 4 домена (2,6 μг). Выделенный бета-рецептор 4 домена рΔ2-7 (2,6 μг) инкубировался либо МАb 2А1Е2 (ряд 2,5 μг), IC7D5 (ряд 3,5 μг), либо неспецифичным МАb (ряд 4,5 μг) в конечном объеме в 500 μл в 1.Р. буфере 3 часа при 4oС. Сефароза СL4В белка А:К каждой трубке (60 μл 50% суспензии) была добавлена сефароза CL4B белка G (1:1) и инкубация продолжалась 2 часа при 4oС. Бусины отгонялись, промывались в 5Х в буфере 1.Р. и подвергались электрофорезу на 10% геле Лэммли. Гель переносился на нитроцеллюлозу и блокировался в РВS, содержащем 0,5% молока. Блот инкубировался с разбавлением 1:100 кроличьего поликлонального антитела бета-рецептора антиPDGF (1-3-5) в РВS, содержащем 0,5% молока. После инкубации в течение ночи блот промывался, инкубировался 125I-белком А, промывался и подвергался рентгену. Ряд 1 показывает стандарт РΔ2-7 без иммуноосаждения. Стрелка указывает позицию РΔ2-7.
С. Иммуноосаждение выделенного внеклеточного бета-рецептора PDGF МАb 2А1Е2. Внеклеточный бета-рецептор человеческого PDGF (рΔ1-5, 5 μг) был подвергнут иммуноосаждению 5 μг либо неспецифического МАb (ряд 2), МАb (ряд 2), МАb 2А1Е2 (ряд 3), либо 1C7D5 (ряд 4), как описано для А. Образцы обрабатывались и блот инкубировался кроличьим поликлональным бета-рецептором 1-3-5 антиPDGF (разбавление 1:100) и 125I-белком А, как описано для В. Ряд 1 содержит 5 μг стандартного рΔ1-5.
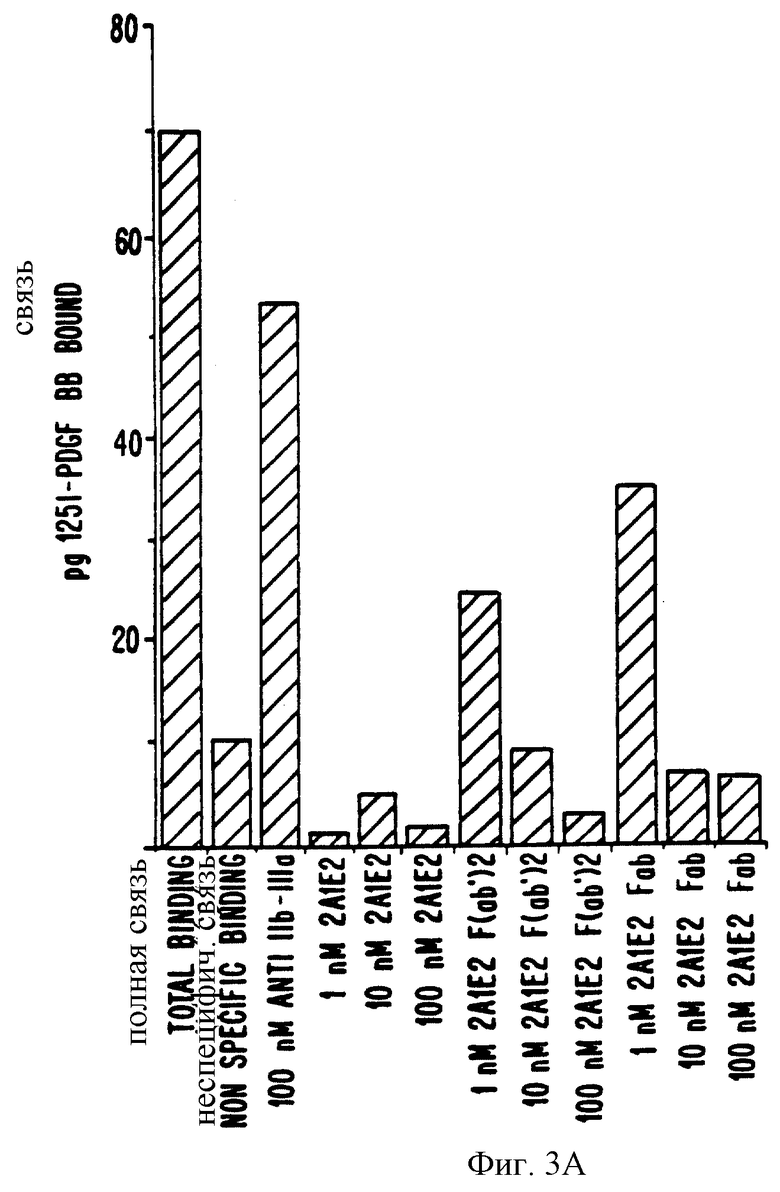
Фиг.3. Торможение 125I-PDGF BB связывания с клетками HR5.
А) Клетки HR5 инкубировались разными концентрациями MAb 2А1Е2 или контрольными MAbs/200 нМ анти-IIb/IIIa или 20 нМ 4С5С8), как описано в способах. Затем клетки инкубировались 125I-PDGF ВВ и связанная радиоактивность определялась, как описано. Неспецифическое связывание определяется как количество 125I-PDGF ВВ, связанного в присутствии 100-кратного избытка немеченного PDGF ВВ.
B) НR5 клетки инкубировались разными концентрациями MAb 2А1Е2, MAb 2А1Е2-F (ав' 2), или MAb 2А1Е2-Fab полной длины и 100 нМ анти-IIв/IIIа с полной длиной, как описано в способах. Измерялось общее связывание 125I-PDGF BB с клетками в присутствии и отсутствии MAbs и их производных. Количество 125I-PDGF BB, связанное в присутствии с 100-кратного избытка немеченного PDGF BB представляет неспецифическое связывание.
Фиг. 4. Торможение фосфорилирования клеток НR5 MAb 2А1Е2. Сливающиеся монослои клеток HR5 в 6-лунковых чашках предварительно инкубировались с дублированием различными MAbs с последующей инкубацией лигандом PDGFВВ, как описано в способах. Клетки растворялись и эквивалент одной 6-лунковой чашки подвергался электрофорезу на 7% геле Лэммли и переносился на нитроцеллюлозу. Вестерн-блот блокировался и затем инкубировался MAb антифосфотирозина. Затем блот инкубировался 125I-белком А и авторадиографировался. Ряды 3-7 представляют лунки, которые были ранее инкубированы либо 0,13 нМ MAb 2А1Е2 (ряд 3), 1,3 нМ MAb 2А1Е2 (ряд 4), 13,3 нМ MAb 2А1Е2 (ряд 5), 0,13 μМ MAb 2А1Е2 (ряд 6) или 0,53 μМ MAb 2А1Е2 (ряд 7), а потом 100 нг/мл PDGFВВ. Ряд 2 показывает степень фосфорилирования в присутствии 100 нг/мл PDGF ВВ, когда клетки вначале предварительно инкубировались 0,53 μМ неспецифичных МАb, и ряд 8 показывает фосфорилирование, вызванное PDGF ВВ, когда клетки предварительно инкубировались 0,53 μМ МАb 4С5С8 PDGF бета-рецептора. Стрелка указывает позицию человеческого PDGF бета-рецептора в полную длину.
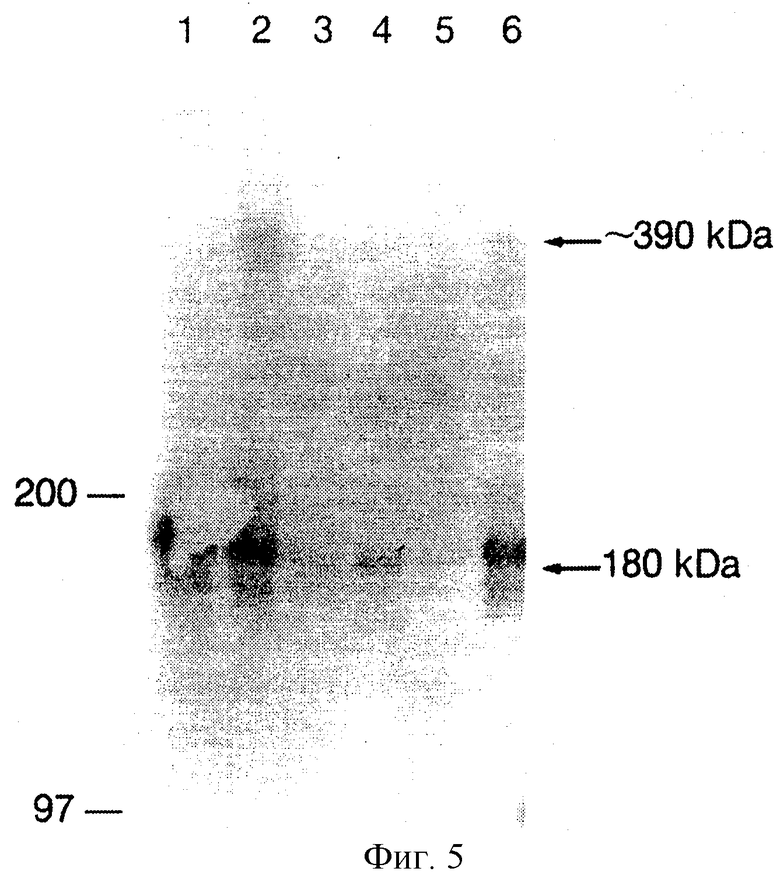
Фиг. 5. Торможение димеризации PDGF BB рецептора PDGF с помощью MAb 2А1E2, HR5 клетки инкубировались либо 0,13 нМ MAb 2А1Е2 (ряд 3), 0,13 μМ MAb 2А1Е2 (ряд 4), либо 1,3 μМ MAb 2А1Е2 (ряд 5), потом 100 нг/мл PDGF ВВ, и поперечная сшивка проводилась, как описано в способах. Ряд 6 представляет действие 0,1 μМ анти-IIb/IIIа МАb на димеризацию. Ряд 1 показывает относительное количество димера в отсутствие добавленного кросс-линкера и ряд 2 показывает количество димеризированного PDGF рецептора в отсутствие антитела.
Фиг. 6. Торможение митогенеза в клетках AG01423B MAb 2А1Е2. Клетки культивировались вместе и инкубировались, как описано в способах, различными концентрациями, либо MAb 2А1Е2 (окружности), либо нетормозящим контрольным MAb 4С5С8 бета-рецептора PDGF (затемненные квадраты) в присутствии 50 нг/мл PDGFBВ. Включение 3Н-тимидина измерялось, как описано.
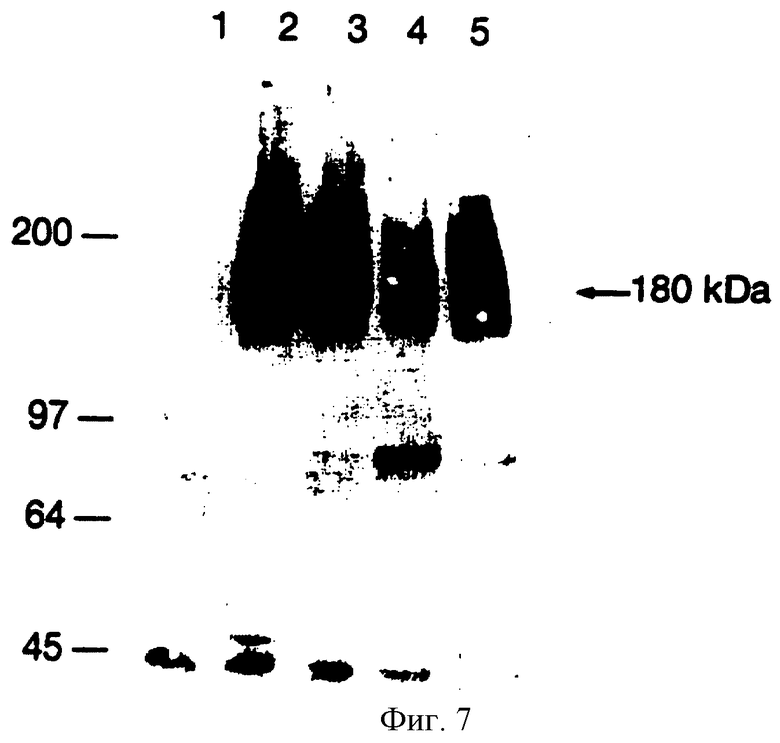
Фиг. 7. Торможение фосфолирования с помощью MAb 2А1Е2 в клетках гладкой мускулатуры павиана. Клетки гладкой мускулатуры павиана инкубировались PDGFBB (ряд 1) либо инкубировались 100 нг/мл PDGF BB в отсутствие МAb (ряд 2) либо в присутствии 2 нМ МАb 2А1Е2 (ряд 3), 200 нМ МАb 2А1Е2 (ряд 4) или 20 нМ МАb 2А1Е2 (ряд 5). Вызванное лигандом фосфорилирование определялось вестерн-анализом, как описано в способах.
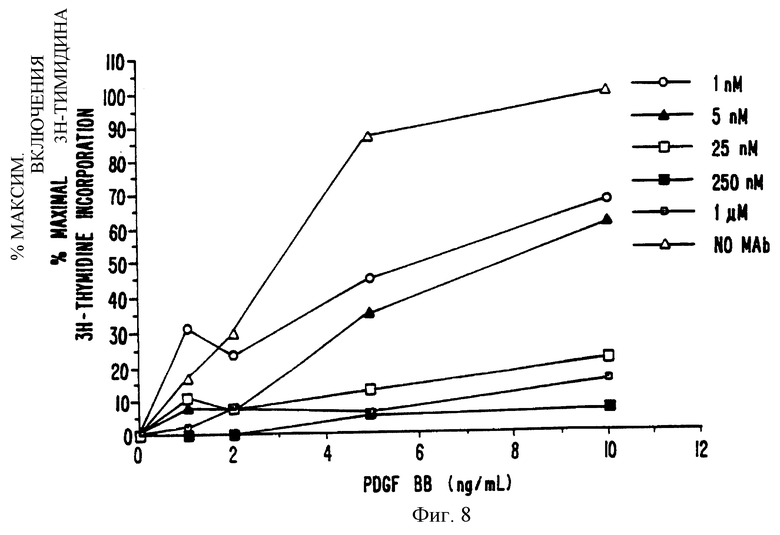
Фиг. 8. Торможение митогенеза посредством МАb 2А1Е2 в клетках гладкой мускулатуры павиана. Сливающиеся клетки гладкой мускулатуры павиана инкубировались 1 нМ, 5 нМ, 25 нМ, 250 нМ или 1 μМ МАb 2А1Е2 в присутствии различных концентраций PDGF ВВ. Включение 3Н-тимидина измерялось, как описано в способах. Данные выражены как процент максимального включенного 3Н-тимидина (приблизительно 30-50000 имп/м при насыщающей концентрации лиганда (10 нг/мл) в отсутствие МАb. Включение 3Н-тимидина в отсутствие PDGF ВВ составляет 5-8000 имп/м. График представляет в среднем 4 отдельных эксперимента.
Описание предпочтительных вариантов
Настоящее изобретение представляет собой иммуноглобулиновый полипептид, который специфически связывается с бета-рецептором PDGF человеческого типа. Антитело способно тормозить вызванный PDGF митогенез клеток, которые производят PDGF бета-рецептор человеческого типа на поверхности клеток. Изобретение полезно в диагностических применениях, а также в лечении заболеваний, вовлекающих пролиферацию с участием PDGF, миграцию и хемотаксис клеток с бета-рецептором PDGF человеческого типа.
Определения
а) Белки
Термины "пептид", "полипептид" или "белок" используются здесь взаимозаменяемым образом. Термин "значительная идентичность" со ссылкой на полипептиды указывает, что данный полипептид или белок по меньшей мере на 30% идентичен всему или части природного белка, обычно по меньшей мере идентичен на 70% и предпочтительно по меньшей мере на 95%.
Используемые здесь термины "изолированный, "фактически чистый" и "фактически гомогенный" используются взаимозаменяемым образом и описывают белок, который был отделен от компонентов, обычно сопровождающих его. Как правило, мономерный белок фактически чист, если по меньшей мере 60-75% образца демонстрируют единый полипептидный каркас. Малые варианты или химические модификации обычно имеют одинаковую полипептидную последовательность. Фактически очищенный белок, как правило, включает свыше 85-90% образца белка, более обычно - около 95%, и предпочтительно свыше 99% чистоты. Чистота белка или гомогенность может быть указана рядом способов, известных в области техники, такими как электрофорез на полиакриламидном геле образца белка с последующей визуализацией единой полипептидной полосы на полиакриламидном геле при окрашивании. Для некоторых целей потребуется высокое разрешение и для очистки используются ВЭЖХ или другие подобные средства.
Полипептид фактически свободен от естественных природных компонентов, когда он отделен от примесей, которые сопровождают его в его естественном соотношении. Таким образом, полипептид, который химически синтезирован или синтезирован в клеточной системе, отличной от клетки, от которой он в природе берет начало, будет фактически свободен от своих сопровождающих его в природе компонентов.
Белки могут быть очищены до значительной гомогенности стандартной технологией, хорошо известной в области техники, включающей селективное осаждение такими веществами, как сульфат аммония, колоночную хроматографию, методы иммуноочистки и другие. См. например. R. Scopes. Protein Purification: - Principles and Practice, Spriger - Verlag: New York (1982).
в) Нуклеиновые кислоты
Нуклеиновые кислоты могут быть ДНК или РНК. Относительно нуклеиновых кислот термин "значительная идентичность" указывает, что последовательности двух нуклеиновых кислот или их указанные части при оптимальном выравнивании и сравнении идентичны соответствующим нуклеотдиным вставкам или делециям по меньшей мере в 80% нуклеотидов, как правило, 90-95%, а предпочтительно 98-99,5% нуклеотидов.
Значительная идентичность последовательности нуклеиновой кислоты существует, если сегмент нуклеиновой кислоты гибридизируется при селективных условиях гибридизации с комплементом другой цепи нуклеиновой кислоты.
"Фактически комплементарная" также означает, что одна нуклеиновая кислота идентична другой или гибридизируется селективно с другой нуклеиновой кислотой. Как правило, селективная гибридизация происходит, если на фрагменте по меньшей мере в 14-25 нуклеотидов идентичность составляет по меньшей мере 55%, лучше около 75% и еще лучше около 90%. См. M. Kanehisa, Nucleic Acids Res. 12:203 (1984).
Точные условия гибридизации обычно включают солевые концентрации менее чем около 1M, чаще менее 500 мМ и предпочтительно менее 200 мМ. Температурные условия, как правило, будут выше 22oС, около 30oС, а предпочтительно более 37oС. Так как другие факторы могут существенно влиять на точность гибридизации, включая композицию основы и размер комплементарных цепей, присутствие органических растворителей и степень несоответствия основы, комбинация параметров более важна, чем абсолютная мера какого-либо одного из них.
"Изолированная" или "практически чистая" в отношении к нуклеиновым кислотам предполагает те кислоты, которые были очищены от других клеточных компонентов или других примесей, например других клеточных нуклеиновых кислот или белков стандартной технологией, включающей обработку щелочь/додецилсульфат натрия (SDS), центрирование в градиенте CsCl, колоночную хроматографию и другие хорошо известные в технике. См. F. Ausubel и др., изд. Current Protocols in Molecular Biology, Green Publishing and Wiley Interscience, New York (1987).
Нуклеиновая кислота "операбельно связана", если она вступает в функциональную взаимосвязь с другой последовательностью нуклеиновой кислоты. Например, промотор или усилитель операбельно связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности. Обычно операбельно связанный означает, что связанные последовательности нуклеиновой кислоты смежны, и если необходимо соединить кодирующие области двух белков, они смежны и находятся в рамке считывания.
Технология манипуляции с нуклеиновой кислотой такая, как субклонирование последовательностей нуклеиновой кислоты, кодирующих полипептиды в векторы экспрессии, метящие зонды, гибридизации ДНК и прочие, описаны, например, в Sambrook и др. (1989) Molecular Cloning: a Laboratory Manual (2 изд. том 1-3, Cold Spring Harbor Laboratory или Ausubel и др. изд. (1987) op. cit.).
"Векторы экспрессии", "клонирующие векторы" или "векторы" - часто плазмиды или другие молекулы нуклеиновой кислоты, которые способны к репликации в выбранной клетке хозяина. Векторы экспрессии могут воспроизводиться автономно или при вставке в геном клетки хозяина способами, хорошо известными в этой области. Векторы, которые воспроизводятся автономно, имеют начало репликации или автономную реплицирующую последовательность (ARS), которая функциональна в выбранной клетке (ах) хозяина. Часто представляется желательным, чтобы вектор мог использоваться более, чем в одной клетке хозяина, например в Е. соli, для клонирования и построения и в клетке млекопитающего для экспрессии.
Линии клетки млекопитающего часто используются в качестве клеток хозяина для экспрессии полипептидов, полученных от эукариотов. Размножение клеток млекопитающего в культуре per se хорошо известно. См. Tissue Culture, Academic Press, Kruse and Patterson изд. (1973). Линии клетки хозяина могут также включать такие организмы, как бактерии (например, Е.соli или В.Subtilis), дрожжи, волокнистые грибки, клетки растений или клетки насекомых среди других клеток.
"Трансформация" означает введение векторов, содержащих нужные нуклеиновые кислоты, непосредственно в клетки хозяина хорошо известными способами. Способы преобразования, которые различны в зависимости от типа клетки хозяина, включают электропорацию, трансфекцию с использованием хлорида кальция, рубидий хлорид кальций фосфата, ДЭАЭ-декстрана или других веществ, бомбардировку микропулями; липофекцию; заражение (если вектор является инфицирующим агентом) и другие методы. См. Sambrook и др. (1989) op.cit. и Ausubel и др. (изд.) (1987) op.cit. Ссылка на клетки, в которые были введены описанные выше нуклеиновые кислоты, включает также потомство таких клеток.
с) Антитела
Используемый здесь "иммуноглобулиновый полипептид" относится к молекулам, которые обладают специфической иммунореактивной активностью. Антитела представляют собой обычно тетрамеры иммуноглобулиновых полипептидов. Здесь термин "антитело" относится к белку, состоящему из одного или нескольких полипептидов, закодированных генами иммуноглобулина. Гены иммуноглобулина включают гены, кодирующие легкие цепи, которые могут быть типа лямбда или каппа, и гены, кодирующие тяжелые цепи. Типы тяжелых цепей - это альфа, гамма, дельта, эпсилон и мю. Карбоксиконцевые части тяжелых и легких цепей иммуноглобулина являются постоянными областями, в то время как аминоконцевые части кодируются бесчисленными генами вариабельной области иммуноглобулина. Вариабельные области иммуноглобулина - это части, которые обеспечивают специфичность распознавания антигена. В частности, специфичность заключается в областях определения комлементарности (CDR's), известных также как гипервариабельные области иммуноглобулина. Иммуноглобулин может существовать во множестве форм, включающих, например, Fv, Fab и F/аb/2, а также в одиночных цепях (например, Huston и др. Proc. Nat. Acad. Sci. USA 85: 5879-5883 (1988) и Bird и др. Science 242:423-426 (1988) (см. Hood и др. "Immunology", Benjamin, N.Y., 2-е изд. (1984) и Hunkapiller and Hood, Nature 323:15-16 (1986). Также могут использоваться антитела одиночной цепи, в которых гены комбинируются для тяжелой цепи и легкой цепи в единую кодирующую последовательность.
"Моноклоналольные антитела" можно получить различными технологиями, с которыми знакомы специалисты. Вкратце, клетки селезенки от животного, иммунизированного нужным антигеном, иммортализируются обычно слиянием с клеткой миеломы (см. Kohler and Milstein, Eur. J. Immunol. 6:511-519 (1976). Альтернативные методы иммортализации включают преобразование вирусов Эпштейна-Барра, онкогенезом или ретровирусами или другими методами, известными в данной области. Колонии, выращенные от одиночных иммортализированных клеток, просеиваются на получение антител нужной специфичности и сродства для антигена и культура моноклональных антител, полученных такими клетками, может быть усилена различной технологией, включающей введение в перитонеальную полость позвоночного хозяина.
Моноспецифичные иммуноглобулины можно также получить рекомбинантной технологией в прокариотных или эукариотных клетках хозяина.
"Химерные" антитела кодируются генами иммуноглобулина, которые были сконструированы так, что гены легкой и тяжелой цепи состоят из сегментов гена иммуноглобулина, принадлежащих разным видам. Например, вариабельные сегменты (V) генов от мышиного моноклонального антитела можно соединить с человеческими постоянными (С) сегментами. Такое химерное антитело, вероятно, будет менее антигенно к человеку, чем антитела с мышиными постоянными (неизмененными) областями, а также мышиными вариабельными областями.
Здесь термин химерное антитело также относится к антителу, которое включает иммуноглобулин, имеющий каркас типа человеческого и в котором любая неизменная область имеет идентичность последовательности полипептида с неизменной областью человеческого иммуноглобулина, так называемого "гуманизированного" иммуноглобулина по меньшей мере порядка 85-90%, а предпочтительно 95% (см., например, Публикацию РСТ WO 90/07861). Следовательно, все части такого "гуманизированного" иммуноглобулина за исключением возможно областей определения комплементарности (СDR's) фактически идентичны соответствующим частям одной или нескольких природных человеческих последовательностей иммуноглобулина.
Термин "область каркаса" здесь относится к тем частям вариабельных областей легкой и тяжелой цепи иммуноглобулина, которые относительно консервированы (т.е., кроме CDR's) среди различных иммуноглобулинов в одном виде, как определено Каbat и др. (1987): Sequences of Proteins of Immunologic Interest, 4-е изд. UP Dept. Health and Human Services ). Здесь "близкая к человеческой областькаркаса" означает область каркаса, которая в каждой существующей цепи включает по меньшей мере около 70 или больше остатков аминокислот, обычно 75-85 или больше остатков, идентичных остаткам в человеческом иммуноглобулине.
Последовательности ДНК человеческой неизменной области можно изолировать в соответствии с известными процедурами от множества человеческих клеток, но предпочтительно от иммортализированных В-клеток. Вариабельные области или CDRs для получения химерных иммуноглобулинов настоящего изобретения можно таким же образом получить из моноклональных антител, способных связываться с бета-рецептором PDGF человеческого типа, и они будут продуцироваться в любом подходящем источнике млекопитающих, включая мышей, крыс, кроликов или других позвноночных, способных продуцировать антитела хорошо известными способами. Подходящие исходные клетки для последовательностей ДНК и клетки хозяина для экспрессии и секреции иммуноглобулина можно получить из ряда источников, таких как Американская Коллекция типов культур ("Каталог клеточных линий и гибридом", пятое издание (1985) Rockville, Maryland, USA).
Кроме химерного и "гуманизированного" иммуноглобулинов, описанных здесь, другие фактически идентичные модифицированные иммуноглобулины можно легко построить и получить с использованием различных технологий рекомбинантных ДНК, хорошо известных специалистам. В общем модификацию генов легко провести с помощью многих известных технологий, таких как сайт-направленный мутагенез (см. Gillman и Smith, Grene 8:81-97 (1979) и S. Roberts и др., Nature 328: 731-734 (1987)).
В другом случае можно продуцировать фрагменты полипептида, включающие только часть первичной структуры иммуноглобулина. Например, может потребоваться получить фрагменты иммуноглобулинового полипептида, которые обладают одной или несколькими активностями иммуноглобулина (дополнение к, или другие, чем узнавание антигена, например, фиксацией комплемента).
Гены иммуноглобулина полностью или частично можно также комбинировать с функциональными областями из других генов (например, ферментами) или с другими молекулами, такими как токсины или метки, чтобы получить белки слияния (например "иммунотоксины"), обладающие новыми свойствами. В этих случаях слияния генов два компонента присутствуют внутри той же самой цепи полипептида. В другом случае иммуноглобулин или его фрагмент можно химически связать с токсином или меткой любой из множества хорошо известных химических процедур. Например, если метка или цитотоксический агент является белком и второй компонент - интактный иммуноглобулин, связь можно провести посредством гетеробифункциональных перекрестных сшивателей, например SPDP, карбодиимида, глютаральдегида и других.
Подходящие метки включают, например, радионуклеиды, ферменты, субстраты, кофакторы, ингибиторы, флюоресцеры, хемилюминесцеры, магнитные частицы. См., например, патенты по использованию таких меток: патенты США 3817837; 3850752; 3939350, 996345, 4277437, 4275149 и 4366241.
Иммунотоксины, включающие молекулы с одиночной цепью, могут также продуцироваться рекомбинантными средствами. Получение различных иммунотоксинов хорошо известно в области и способы можно найти, например, в "Monoclonal Antibody-Toxinconjugates: Aiming the Magic Bullet", Thorpe и др. Monoclonal Antibodies in Clinical Medicine, Acad. Press., стр. 168-190 (1982); Е. Vitetta, Science (1987) 238: 1098-1104 и G. Winter и C. Milstein, Nature (1991) 349:293-299.
Для использования в иммунотоксинах пригодно множество цитотоксических агентов. Цитотогксические агенты могут включать радионуклиды, такие как йод-131, иттрий-90, рений-188 и висмут-212; ряд химиотерапевтических лекарств, таких как виндезин, метотрексат, адриамицин и цисплатин; и цитотоксические белки, такие как рибосомные тормозящие белки, как антивирусный белок фитолакки, Pseudomonas экзотоксин А, рицин, дифтерийный токсин, цепь рицина А и т.п. или агент активный на поверхности клетки, такой как ферменты фосфолипазы (например, фосфолипаза С) (См. "Химерные токсины". Olsnes and Pihl, Pharmac. Ther, 15:355-381(1981) и "Моноклональные антитела для обнаружения и лечения рака", изд. Baldwin and Byers, стр. 159-179, 224-266, Academic Press (1985).
Описание изобретения
Иммуноглобулиновые полипептиды настоящего изобретения найдут применение в лечении, диагностике и других областях. Различные технические приемы, используемые в этих областях, обсуждаются, например, в Harlow и Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor, New York (1988) (включена сюда как ссылка для всех целей) и включают: иммунизацию животных для продуцирования иммуноглобулинов; получение моноколональных антител; мечение иммуноглобулинов для использования в качестве зондов; очистка иммуносродства; и иммунопробы.
Примером иммуноглобулинового полипептида настоящего изобретения является моноклональное антитело 2А1E2, описанное ниже, которое связывается специфически с рецептором человеческого PDGF типа бета. Моноклональное антитело 2А1E2, которое представляет собой изотип IgG1, было депонировано в Американской коллекции типовых культур, 12301 Park hown Drive, Rockvillе, Maryland 20231 (АТСС НВ10938) до даты подачи этой заявки.
Иммуноглобулиновые полипептиды антиPDGF рецептора настоящего изобретения можно получить иммунизацией животного очищенным или частично очищенным внеклеточным доменом рецептора человеческого PDGF бета-типа. Иммунизируемые животные могут быть одним любым из множества видов, которые способны иммунологически опознавать эпитопы, характерные для внеклеточного домена бета-рецептора человеческого PDGF, такие как лошадиные, мышиные и пр.
Моноклональные антитела изобретения можно получить иммортализацией последовательностей нуклеиновой кислоты, которые кодируют иммуноглобулиновые полипептиды или их части, которые связываются специфически с антигенными детерминантами, характерными для внеклеточного домена бета-рецептора человеческого PDGF. Процесс иммортализации можно проводить техникой слияния гибридомы, вирусным переобразованием лимфоцитов, производящих антитело, технической рекомбинантной ДНК или техникой, которая объединяет слияние клеток, вирусное преобразование и/или методики рекомбинантной ДНК.
В соответствии с одним аспектом изобретения клетки, продуцирующие моноклональные антитела антиPDGF рецептора, иммортализируются, использованием, например, техники преобразования вируса Эпштейна-Барра (EBV). Например, В лимфоциты, полученные из периферической крови, костного мозга, лимфатических узлов, небных миндалин и пр. пациентов, особенно иммунизированные рецептором PDGF или его частью, иммортализируются с использованием ЕВV в соответствии с методами, которые описаны в патенте США 4464465, Chan. и др. J. Immunol, 136:106 (1986).
Моноклональные антитела рецептора человеческого антиPDGF можно также получить другими способами, например использованием комбинации ЕВV или другой техники вирусной трансформации гибридомного слияния. Например, гибридомы можно создать слиянием стимулированных В клеток, полученных от индивида, иммунизированного рецептором PDGF или его частью, с партнером гетерогибридного слияния мышь/человек, разновидность которого была описана. См., например, патент США 4624921 и James и Bell, J. Immunol. Methods 100:5-40 (1987). Партнер слияния мышь/человек можно создать слиянем человеческих лимфоцитов, стимулированных или преобразованных ЕВV, с готовыми линиями мышиной миеломы, такими как NS-1 или РЗNS-1, в присутствии, например, полиэтиленгликоля. Гибрид должен соответствующе насыщаться лекарством, что можно сделать, культивируя гибрид в повышающихся концентрациях нужного лекарства, такого как 6-тиогуанин, уабаин или неомицин.
Гибридомы или лимфобластоидные клетки, которые выделяют нужное антитело, можно идентифицировать скринингом культуральных супернатантов на антитело, которое связывается с бета-рецептором PDGF. Предпочтительнее использовать скрининг для обнаружения тех антител, которые тормозят, например, митогенез с участием PDGF. Клетки, которые обладают нужной активностью, клонируются и субклонируются в соответствии с традиционной техникой и отслеживаются до тех пор, пока не идентифицируются стабильные иммортализированные линии, продуцирующие нужное моноклональное антитело рецептора антиPDGF. Под моноклональным антителом подразумевается антитело, продуцируемое клональной иммортализированной клеточной линией отдельно от клеток, производящих антитела с разной антигенсвязующей специфичностью. Таким образом, такие моноклональные антитела продуцируются изолированными от других моноклоналольных антител и соответственно в фактически чистой форме (относительно других антител) и в концентрации, обычно большей, чем обычно бывает в сыворотке из видов животных, которая служит в качестве источника В клетки.
В другом случае можно изолировать последовательности ДНК, которые кодируют иммуноглобулиновый полипептид рецептора человеческого антиPDGF или его часть, которая специфически связана с внеклеточным доменом рецептора PDGF, скринингом библиотеки ДНК от человеческих В клеток в соответствии с общим протоколом, как описано у Huse и др. Science 246:1275-1281 (1989), и затем клонированием и амплифицированием последовательностей, которые кодируют антитела рецептора антиPDGF (или связующий фрагмент) нужной специфичности.
Затем иммуноглобулины можно продуцировать введением вектора экспрессии, содержащего соответствующий ген иммуноглобулина или его часть, в подходящую клетку хозяина. Затем клеточная линия хозяина поддерживается в условиях, приемлемых для высокого уровня экспрессии нуклеотидных последовательностей иммуноглобулина, и при желании можно провести сбор и очистку легких цепей, тяжелых цепей, димеров легкой/тяжелой цепи или интактных антител, связывание фрагментов или получение других форм иммуноглоубина.
Подходящие клетки хозяина включают микроорганизмы, но тканевая клеточная культура млекопитающего или насекомого может быть предпочтительна для продуцирования моноклонального антитела настоящего изобретения (см. E. Winnacker, "От генов к клонам" VCH Publishers, N.Y., N.Y. (1987). В данной области был разработан ряд подходящих клеточных линий хозяина, способных выделять интактные иммуноглобулины, и они включают клеточную линию яичника китайского хомяка (СНО), но желательно использовать преобразованные В клетки или гибридомы.
После экспрессии целые антитела, их димеры, отдельные легкие и тяжелые цепи или другие формы иммуноглобулина настоящего изобретения можно очистить в соответствии со стандартными процедурами, включающими осаждение сульфатом аммония, аффинные колонки, колоночной хроматографией, гелевым электрофорезом и пр. (см., например, R. Scopes, Protein Purification, Springer-Verlag, N.Y. (1982). Для фармацевтических применений предпочитаются фактически чистые иммуноглобулины с гомогенностью 90-95%, лучше 98-99% или выше. После частичной очистки или нужной гомогенности полипептиды можно использовать терапевтически (включая экстракорпорально) или в разработке и проведении анализов, иммунофлюоресцентных окрашиваний и других процедурах (см. в общем Immunоlogical Methods, том 1 и 2, изд. Lefkovits and Pernis, Academic Press, New York (1979 и 1981).
Иммуноглобулиновые полипептиды, полученные по настоящему изобретению, могут представлять изотип IgG, IgM, IgA или IgD, а также быть любым из подклассов, как, например, IgG1, IgG2, IgG3 или IgG4. Используя технику рекомбинантной ДНК, можно легко провести "смену класса" изолированных иммуноглобулиновых полипептидов. В этом способе гены, кодирующие неизменные области, которые определяют изотип нужной молекулы иммуноглобулина, заменяются генами, кодирующими желаемый изотип или подкласс, как описано в европейской патентной публикации ЕР 314161, включенной сюда ссылкой. Иммуноглобулины с измененным классом можно также изолировать отбором клеток, которые перенесли спонтанное изменение с использованием известных способов селекции.
Назначение людям иммуноглобулиновых полипептидов, которые фактически нечеловеческие, может прояснить ответные реакции анти-антитело. Таким образом можно получить иммуноглобулиновые полипептиды антиPDGF рецептора настоящего изобретения, которые практически человеческие. Под термином "практически человеческий" понимается антитело или его связующий фрагмент, состоящий из последовательностей аминокислоты, которые по меньшей мере на 50% по происхождению человеческие, лучше на 70-80%, а еще лучше на 95-99% человеческие, предназначенные для повторных назначений в течение продолжительного периода, как в случае лечения нарушений клеточной пролиферации с участием PDGF. Здесь термин человеческое антитело охватывает антитела полностью человеческого происхождения, а также те, которые практически человеческие, если в тексте не указано иначе.
Поскольку изобретение моноклональных антител человеческого антиPDGF рецептора может быть трудным с помощью обычной техники иммортализации, вначале можно получить нечеловеческие антитела, а затем перенести через технику рекомбинантной ДНК антигенсвязующие области нечеловеческих антител, например, Fаb, области определения комплементарности (CDRs) или гипервариабельные области, на человеческие контактные области (Fc) или области каркаса, чтобы получить практически человеческие молекулы. Такие методы общеизвестны в области и описаны, например, в патенте США 4816397, публикации РСТ WO 90/07861 и ЕР публикациях 173494 и 239400.
Полученные химерные антитела или химерные иммуноглобулиновые полипептиды, которые специфически связываются с бета-рецептором PDGF человеческого типа и таким образом тормозят связывание PDGF с рецептором, также входят в объем настоящего изобретения. Типичным терапевтическим химерным антителом будет гибридный белок, состоящий из вариабельного (V) или антигенсвязующего домена от мышиного иммуноглобулина, специфичного для антигенной детерминанты бета-рецептора PDGF человеческого типа, и константантного (С) или эффективного домена от человеческого иммуноглобулина, хотя и для вариабельного и контактного доменов можно использовать домены от других видов млекопитающих. Здесь термин "химерное антитело" также относится к антителам, закодированным генами иммуноглобулина, в которых только области, определяющие комплементарность (СВR's), переносятся из иммуноглобулина, который специфически опознает антигенные детерминанты, остальная часть гена иммуноглобулина получена от человеческого (или другого млекопитающего) гена иммуноглобулина. Этот тип химерного антитела именуется "гуманизированный" (в случае использования гена человеческого иммуноглобулина) антителом.
Гипервариабельные области вариабельных доменов иммуноглобулиновых полипептидов антиPDGF рецептора являют собой другой аспект изобретения. Гипервариабельные области или CDRs вкупе с каркасными областями (теми частями вариабельных областей легкой и тяжелой цепи иммуноглобулина, которые относительно консервативны среди различных иммуноглобулинов в одном виде) позволяют иммуноглобулиновым полипептидам антиPDGF рецептора опознавать и таким образом связывать PDGF бета-рецептор человеческого типа. Гипервариабельные области можно клонировать и секвенировать. После идентификации эти области, которые наделены специфической способностью опознавания рецептора PDGF, можно клонировать
в вектор для эксперссии в хозяине как части другой молекулы иммуноглобулина или как белка слияния, например молекула носителя, которая функционирует на усилиение иммуногенности клонированного идиотопа.
Иммуноглобулиновые полипептиды антиPDGF рецептора изобретения будут обычно использовать интактно или как иммуногенные фрагменты, такие как фрагменты Fv, Fаb или F(аb')2. Фрагменты можно получить от антител традиционной техникой, как, например, протеолическим перевариванием антитела с использованим пепсина или папаина или техникой рекомбинантной ДНК, в которой ген или его часть, кодирующая желаемый фрагмент, клонируется или синтезируется и экспрессируется в ряде хозяев.
Специалистам понятно, что "антиидиотипические" антитела можно получить использованием специфического иммуноглобулина в качестве иммуногена в соответствии со стандартной технологией. Например, заражение или иммунизация полипептидом PDGF рецептора или его фрагмента индуцирует нейтрализующий иммуноглобулин, который на своем комбинирующем сайте вариабельного Fаb региона имеет изображение полипептида PDGF рецептора, которое уникально для этого конкретного иммуноглобулина, т. е. идиотип. Иммунизация таким антиPDGF-R иммуноглобулином индуцирует антиидиотипическое антитело, которое имеет конформацию на своем комбинирующем сайте, которая имитирует структуру первоначального PDGF-R антигена. Поэтому эти антиидиотипные антитела можно использовать вместо PDGF-R антигена для лечения заболеваний, опосредованных PDGF (см., например, Nisonoff (1991) J. Immunol. 147:2429-2438).
Иммуноглобулиновые полипептиды антиPDGF рецептора изобретения находят применение в терапевтических и диагностических методах и композициях. В лечебных целях иммуноглобулиновые полипептиды антиPDGF рецептора используются в качестве растворимого лиганда для бета-рецептора PDGF человеческого типа, маскируя рецептор или иначе тормозя молекулы PDGF в их связывании с рецептором, что приводит в итоге к торможению желательной миграции клеток и пролиферации.
В фармавтических композициях иммуноглобулиновые полипептиды антиPDGF рецептора по изобретению, как описано здесь, назначаются индивиду, страдающему нарушением клеточной пролиферации, вызванной PDGF. В терапевтических применениях композиции назначаются пациенту в количестве, достаточном, чтобы блокировать клеточные рецепторы и таким образом пролиферацию и ее симптомы и/или осложнения. Количество, необходимое для этого, определяется как "терапевтически эффективная доза". Количества, эффективные для такого использования, будут зависеть от характера композиции, способа назначения, стадии и серьезности заболевания, веса и общего состояния здоровья пациента, а также от профессионального опыта лечащего врача, но обычно составляют от 0,01 до 100,0 мг/кг антитела в день с наиболее часто используемой дозировкой в 0,1 - 10,0 мг/кг антитела в день. Нужно помнить, что иммуноглобулиновые полипептиды антиPDGF рецептора и полученные из них пептидные композиции можно использовать в серьезных состояних заболевания, то есть в ситуациях угрозы жизни или потенциальной угрозы жизни. В таких случаях возможно с учетом мнения лечащего врача назначить значительно превышающие обычные дозы количества этих композиций. Так, при таких обстоятельствах наиболее предпочтительны моноклональные антитела человеческого антиPDGF рецептора или фактически человеческие антитела антиPDGF рецептора по изобретению.
Однократное или многократные назначения композиций можно проводить, учитывая уровень дозировки и схему, выбранную лечащим врачом. В любом случае фармацевтические препараты обеспечат достаточное количество полипептида антиPDGF рецептора, чтобы эффективно лечить пациента. Назначение должно начинаться при первых показаниях нежелательного проявления клеточной пролиферации или вскоре после диагностики и продолжаться до значительного снятия симптомов или в течение какого-то времени после этого. В точно установленных случаях заболевания потребуются нагружающие дозы с последующими поддерживающими дозами.
Фармацевтические композиции для терапевтического лечения предназначены для парентерального, местного, орального или локального применения. Предпочтительно композиции назначаются парентерально, например, внутривенно, подкожно, внутридермально или внутримышчено. Изобретение предлагает композиции для парентерального назначения, которые включают раствор иммуноглобулинового полипептида, растворенного или суспендированного в приемлемом носителе, предпочтительно водном. Можно использовать много водных носителей, например воду, забуференную воду, 0,4% солевой раствор, 0,3% глицин, гуалуроновую кислоту и другие. Эти композиции можно стерилизировать традиционной хорошо известной техникой стерилизации или стерильно фильтровать. Полученные водные растворы можно упаковывать для использования или лиофилизировать, лиофилизированный препарат соединяется со стерильным раствором перед введением. Композиции могут содержать фармацевтически приемлемые дополнительные вещества, необходимые для приближения к физиологическим условиям, такие как агенты регулирования рН и буферные агенты, агенты регулирования тоничности, смачивающие агенты и прочие, например, ацетат натрия, лактат натрия, хлорид натрия, хлорид калия, хлорид кальция, сорбитан монолаурат, триэтаноламин олеат и т.п.
Концентрация полипептидов иммуноглобулина антиPDGF рецептора в фармацевтических препаратах может значительно меняться, например от менее 1% до обычного уровня в 10-15% и до 50% или выше по весу и будет выбираться в основном объемом жидкости, вязкостью и пр. в соответствии с конкретным выбранным способом назначения.
Так, типичную фармацевтическую композицию для внутривенного вливания можно приготовить из 250 мл стерильного раствора Рингера и 100 мг иммуноглобулинового полипептида антиPDGF рецептора. Конкретные способы приготовления парентерально назначаемых композиций известны специалистам и описаны в подробностях в Remington's Pharmaceutical Science, 17 изд., Mack Publishing Company, Eаston, PA (1985).
Полипептиды и их фрагменты можно назначить через липосомы. Иммуноглобулиновые полипептиды антиPDGF рецептора могут служить для нацеливания липосом на конкретные ткани или клетки, воспроизводящие бета-рецептор человеческого PDGF. Липосомы включают эмульсии, пены, мицеллы, нерастворимые монослои, жидкие кристаллы, фосфолипидные дисперсии, пластинчатые слои и прочее. В этих препаратах нужный полипептид иммуноглобулина или его фрагмент включается как часть липосомы один или вкупе с молекулой, которая, например, токсична для целевых клеток. Липосомную суспензию, содержащую иммуноглобулиновый полипептид, можно назначить внутривенно, локально и т.п. в дозе, которая различна inter alia в зависимости от способа назначения, назначаемого пептида и степени заболевания.
Для твердых композиций полипептидов изобретения можно использовать традиционные нетоксичные твердые носители, которые включают, например, фармацевтические виды маннитола, лактозы, крахмала, стеарата магния, сахарина натрия, талька, целлюлозу, глюкозу, сахарозу, карбонат магния и пр. Для орального назначения фармацевтически приемлемая нетоксичная композиция готовится включением любого из обычно используемых наполнителей, таких как указанные выше носители, и 10-95% активного ингредиента, т.е. одного или нескольких иммуноглобулиновых полипептидов антиPDGF рецептора, а чаще 25-75%.
Для диагностических целей полипептиды изобретения могут быть мечеными или немечеными. Метка - это вещество, которое обеспечивает сигнал обнаружения любым из многих хорошо известных способов и указанных в литературе. Можно непосредственно поместить сами полипептиды иммуноглобулина. Или же намеченные антитела, включенные в изобретение, можно использовать в комбинации с другими антителами (вторыми антителами), которые помечены, и это опознают полипептиды антиPDGF рецептора настоящего изобретения. Например, помеченные антитела, специфичные для неизменных областей полипептидов антиPDGF рецептора, можно использовать для обнаружения полипептида, связанного с образцом.
Используется множество меток, таких как радионуклиды, фтор, ферменты, субстраты ферментов, кофакторы фермента, ингибиторы фермента, лиганды (в частности, гаптены) и т.п. Специалистам известны многие типы иммуноанализов.
Иммуноглобулиновые полипептиды антиPDGF рецептора и их фрагменты по настоящему изобретению можно использовать в различных иммуноанализах для обнаружения рецептора PDGF в физиологических образцах. Такие методы иммуноанализа могут включать жидкофазные иммуноанализы и вестерн-блот анализ, методы конкурентного и неконкуретнтного связывания с белком, твердофазный иммуноферментный анализ (ELISA) и другие общеиспользуемые и широко описанные в научной и патентной литературе, а также многие применяемые коммерчески.
Такие иммуноглобулины и пептиды можно использовать в технике иммуногистохимического окрашивания хорошо известными способами.
Следующий пример предлагается с целью иллюстрации, но не ограничения.
Пример
Материалы
PDGF ВВ был куплен у Amgen. Моноклональное антитело антифосфотирозина (МАb) РУ20 было приобретено у ICI. DМЕМ (модифицированная по способу Дульбекко среда Игла), RPM1 1640, F12, телячья сыворотка, раствор пенициллина-стрептомицина, С418-неомицин, 200мМ глютамина, IM HEPES (N-2-гидроксиэтилпиперазин-N'-2-этансульфоновая кислота), пируват натрия (11 мг/л) и физраствор с фосфатным буфером (РВS) приобретены от GIBCO. Фетальная телячья сыворотка (FCS) и подтиповой набор моноклонального антитела получены от Hyclone. Трис-фосфат натрия, борат натрия, уксусная кислота, пирофосфат натрия, фторид натрия, дитиотрейтол (ДТТ), этилендиамин тетрауксусная кислота - ЭДТК (EDTA), ЕGTA, натрий-додецил сульфат (SDS), натрий ортованадат, хлорид натрия (NaCl), лимонная кислота, фенилметилсульфонилфторид (РМSF), альбумин бычьей сыворотки (ВSA), Тритон Х100, ТWeen 20,2,2' Азино-бис(3-этилбензтиазолин)-6-сульфокислота (АВТS) и перекись водорода были получены от Sigmа. Козья антимышиная пероксидаза и гипоксантин-тиамин (НТ) получены от Boehringer Mannheim. Желатин, неокрашенные маркеры молекулярной массы белка и нитроцеллюлоза - от BioRad. Окрашенные маркеры высокомолекулярного белка - от BRL. Сефароза CL4B белка А, сефароза CL4B белка С, иммуночистый связующий буфер и наборы препаратов, F(аb')2 и Fab были доставлены от Pierce. Упакованные колонны PD10 - от Pharmacia. Маркеры молекулярной массы 125I-белка А и 14С были от Amersham. Реагент 125I-дийодо-Болтон-Хантера - от New England Nuclear. Метанол - от Burdick-Jackson. Тканевая культура поставлялась Costar. Клетки AG01523В были получены от АТСС. J.A. Escobedo (UCSF) любезно предоставили НР5 клетки. Первичные клетки гладкой мышцы плечевой артерии павиана были предоставлены J. Anderson и S. Hanson (Университет Эмори).
Способы
Клеточная культура. Клетки NIH ЗТЗ выдерживались обычно в ДМЕМ, содержащей 10% фетальной телячьей сыворотки. IX пенициллина-стрептомицина, 2 мМ глютамина и пируват натрия (0,11 мг/мл). СНО клетки, экспрессирующие внеклеточный домен (рΔ1-5) или бета-рецептор PDGF с полной длиной (НР 5), культивировались в RPM1, содержащем 10% FCS, IX пенициллина-стрептомицина, 2мМ глютамина, пируват натрия (0,11 мг/мл) и С418 (200 μг/мл). Клетки фибробласта крайней плоти человека (АС01523В) культивировались в DМЕМ, содержащей 10% FCS, IX пенициллина-стрептомицина, 2 мМ глютамина и пируват натрия (0,11 мг/мл). Моноклональные клетки гибридомы содержались в 50:50 DМЕМ:RPM1, содержащем 20% FCS, IX пенициллина-стрептомицина, 2 мМ глютамина, пируват натрия (0,1 мг/мл), IX НТ и 10% среду, когдиционированную макрофагами.
Конструирование клеточных линий, экспрессирующих усеченный бета-рецептор человеческого PDGF. Усечение кДНК бета-рецептора человеческого PDGF проводилось олинонуклеотид-направленным делеционным мутагенезом. Олигонуклеотид-направленный мутагенез ин витро проводился в соответствии с модифицированным методом Кункеля и др. (1987) Meth. Ensуmol. 154:367-382). Первоначально фрагмент кДНК ЕсоRI-Hind III в 3,9 т.п.н. всей кодирующей области бета-рецептора человеческого PDGF (остатки 32-499) был субклонирован в сайты ЕсоRI и Hind III m13 mp18, генерирующие вектор mp18PR (Maniatis и др. (1982) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Lab.N.Y.
Олигонуклеотиды были предназначены, чтобы делетировать части кДНК бета-рецептора PDGF, которые кодируют аминокислоты 499-1074 (PRΔ1: GTG TGA GGA ACG GGA AAT TCA TCG AAG GAC ATC CCC CGAC) или аминокислоты 384-1074 (PRΔ2: GGA AGG TCG ATG TCT AGT TAA TCG AAG GAC ATC CCC CGAC) (фиг.1). Стоп-кодон был введен после остатка 499 (PRΔ1) или остатка 384 (PRΔ2). Проверка субклонирования проводилась анализом переваривания рестрикционного фермента и секвенсированием терминации дидеокси-цепи (Sanger и др. (1977) Proc. Natl. Acad. Sci. USA 74:5463-5467, которая включена сюда ссылкой).
Модифицированные полипептиды бета-рецептора PDGF были субклонированы в ЕсоRI и Hind III сайты вектора экспрессии PBJI (Lin, A. и др. (1990) Science 249: 677-679, которая включена сюда ссылкой) и контрасфецированы вектором рSV2Neo (Southern P.J. и Berg (1982) J. Mol. Appl. Gien. 1:327-341, которая включена сюда ссылкой) в соотношении 1:10 в СНО-К1 клетки способом поглощения реагента липофектина (Felgner, P.L. и др. (1987) Proc. Natl. Acad. Sei. USA 84:7413-7417, которая включена сюда как ссылка). Трансфецированные клетки отбирались на устойчивость к С418-неомицину и индивидуальные клоны изолировались и сортировались при равной плотности клеток на высокий уровень экспрессии внеклеточного бета-рецептора в свободной от сыворотки среде. Экспрессия модифицированных внеклеточных белков бета-рецептора PDGF определялась анализом вестерн-блота с использованием кроличьей поликлональной сыворотки (Ав 1-3-5, см. фиг.1) против синтетического пептида, основанного на остатках 51-68 рецептора человеческого PDGF (SVLILTNLTGLDTGEYFC посл. 3). Для последующего получения белка использовались рекомбинантные клоны рΔ1-5 (экспрессирующие внеклеточный рецептор PDGF в полную длину ) и рΔ2-7 (экспрессирующие домены 1-4 внеклеточного рецептора PDGF).
Получение Маb против бета-рецептора человеческого PDGF. Антитела были получения, как описано Harlow и Lane (1988, Antibodies: A Lab. Manual. Cold Spring. Harbor Lab. N.Y.). Мыши иммунизировались частично очищенным внеклеточным доменом бета-рецептора PDGF (рΔ1-5), используя 50-100 μг (иммунизацию). Титр антитела в иммунизированных мышах определялся анализом ELISA следующим образом. Микротитровальные планшеты за 96 лунок Immunolon II покрывались на всю ночь при 4oС частично очищенным (10-15%) внеклеточным доменом бета-рецептора PDGF (200-300 мг/лунку). Остальные манипуляции проводились при комнатной температуре. Лунки блокировались 0,05 М Трис, рН 7, содержащим 100 мМ NaCl и 0,5% желатина в течение 1 часа. Планшеты инкубировались 2 часа разными разбавлениями мышиной сыворотки, промывались 5Х водным буфером (0,05 М Трис, рН 7, 100 мМ NaCl и 0,3% желатина) и инкубировались козьей антимышиной пероксидазой (1:100 в водном буфере) в течение 1 часа. Планшеты промывались, как описано ранее, и доводились 2,2'-азино-бис/3-этилбензтиазолин/6-сульфокислотой (АВТS, 1 мг/мл) в 0,1 М лимонной кислоты, 0,1 М двуосновного фосфата натрия, рН 4 с содержанием перекиси водорода (окончательная концентрация 0,003%). Определялась абсорбентность на 650 нм, и величины сравнивались с величинами, полученными с белком, очищенным до такой же степени из кондиционированной среды из СHO клеток, трансфицированных только вектором рВJ.
Мышами, которые проявляли высокую реактивность, пришлось пожертвовать и изолировать селезенки. Спленоциты были удалены и слиты с клетками РЗХ миеломы, как описано Harlow и Lane. Гибридомы сортировались с использованием того же анализа ELISA, и положительные гибридомы клонировались и повторно сортировались. Положительные моноклональные клетки культивировались и асциты готовились в Ваlв /с мыши, как описано Harlow и Lane (ор. cit.). Среда тканевой культуры использовалась для субтицирования МАbs с применением субтипирующего набора от Hyclone, как указано производителем.
Очистка антител. Антитела очищались на протеин А-сефароза СL4В следующим образом. Жидкость асцитов разбавлялась 1:5 в связующем буфере Immunopure (Pierce) и хроматографировалась на колонке протеина А-сефароза CL4В, уравновешенной в том же буфере. Проток собирался и колонка промывалась следующим буфером (Immunopure) 10 объемов колонны. Связанные IgGs элюмировались 0,1 М глицина, рН 2,8 и собирались в трубки, содержащие 2 М Трис, рН 11 (40 μл/мл) в качестве нейтрализующего агента. Пиковые фракции белка обнаруживались измерением абсорбции при 280 нм, группировались и диализировались в PBS (2 смены в 4 л каждая).
Получение протеолитических фрагментов МАb 2А1Е2 F(аb')2 и Fаb. F(аb')2. Фрагменты МАв 2А1Е2 были получены из интактного МАb с использованием набора F(аb')2 Ammunopure (Pierce) по инструкции производителя. Моноклональное антитело 2А1Е2 инкубировалось иммобилизированным пепсином 4 часа при 37oС при рН 4,2, и нерасщепленные фрагменты IgG и Fc отделялись от фрагментов F(аb')2 с использованием сефарозы СL4В белка А. Также были получены Fаb фрагменты с применением комплекта для получения Fаb Pierce Immunopure. IgG икубировался иммобилизированным папаином 5 часов при 37oС при рН 7, и непереваренные IgG и Fc фрагменты удалялись белком А. Образцы анрализировались SDS-PAGE после ферментного переваривания, и ELISA бета-рецептора PDGF определял потерю, при наличии таковой, опознавания антигена.
Анализ на иммунопреципитат. Иммунопреципитаты проводились модификацией процедуры Кесслера (1981, Meth. Ensymol. 73: 442-471). При использовании очищенного рΔ1-5 или рΔ2-7 белок инкубировался с МАb либо в течение 2 часов при комнатной температуре, либо в течение 12 часов при 4oС в иммунопреципитатноv (IP) буфере (40 мМ Трис, рН 8, 100 мМ NaCl, 10 мМ ЭДТК, 1 мМ ЕGТА и 1% Triton Х100). Если иммунопреципитату подвергался рецептор полной длины, клетки, экспрессирующие бета-рецептор PDGF, растворялись в IP буфере, содержащем 1 μМ ортованадата натрия и 1 мМ РМSF, и инкубировалась МАb на 8-12 часов при 4oС. Затем образцы инкубировались 50% суспензией смеси 1:1 белка А-сефарозы CL4В и белка С-сефарозы CL4В (50-100 μл/образец). После 1-2 часов при 4oС смола промывалась 3 циклами центрифугирования и повторного суспендирования в IP буфере, и наконец, смола кипятилась в растворяющем буфере образца Лэммли (50-100 μл/образец) (1970. Nature 227: 680-685, которая включена сюда ссылкой). Образцы подвергались SDS (додецилсульфат натрия) - PAGE (электрофорез в полиакриламидном геле) на 7 или 10%-ном геле Лэммли и затем переносились на нитроцеллюлозу. Вестерн- блот блокировался в блокирующем буфере (0,05 М Трис, рН 8, содержащем 0,5 NaCl и 4% ВSA) и инкубировался с первичным антителом 12 часов при 4oС. Нитроцеллюлоза промывалась блокирующим буфером и инкубировалась 125I-белком А (0,4 mCi/мл) на 1-2 часа при комнатной температуре и подвергалась радиоавтографии.
Радиойодирование PDGF ВВ. PDGF йодировался модификацией процедуры Болтон-Хантера, описанной Duan и др. (1991, J. Biol. Chem. 266:413-418). Вкратце, 125I-дийодореагент Болтона-Хантера (1 mCi) высушивался в азоте. Затем PDGF ВВ (2,5 г) добавлялся к 125I-дийодореагенту Болтон-Хантера в 10 μл 0,1 М бората натрия (рН 8,5) на 15 минут при 4oС. Реакционная смесь гасилась 500 μл 0,1 М бората натрия, 0,2 М глицина, рН 8,5, на 10 минут при 4oС. Этот материал подвергался гель-фильтрации на колонке РD 10, предварительно уравновешенной 0,3 М уксусной кислоты, содержащей 1 мг/мл BSA. Пиковые радиомеченные фракции белка обнаруживались использованием гамма-счетчика. Как правило, специфическая активность йодированного PDGF ВВ составляла 50000 имульсов/нг.
Связывание 125I-PDGF ВВ с интактными клетками НR5. Клетки HR5 собирались РВS, содержащим 2 мМ ЭДТК в течение 20 минут при 37oС. Промытые НR5 клетки (1х106/100 л) инкубировались в трех вариантах с различными концентрациями МАb (или F(аb')2 или Fаb фрагментов) в РВS, содержащем 0,5% BSA на 30 минут при комнатной температуре. HR5 клетки инкубировались 125I-PDGF ВВ (прибл. 1 нг/трубку) в отсутствие (полное связывание) или присутствии (неспецифическое связывание) 100-кратного избытка немеченных PDGF ВВ и белка-носителя (обедненная тромбоцитами плазма, 50 μл) 40 минут при комнатной температуре. Конечный объем инкубации составит 500 μл. Инкубационная смесь (400 μл) расслаивалась на Ficoll-paque (700 μл) и центрифугировалась. Всплывающий слой удалялся и в гранулах клеток определялась радиоактивность.
Анализ на фосфорилирование. НR5 клетки выращивались в 6-лунковых чашах и первичные клетки культивировались в 100 мм чашках. Клетки дважды промывались холодной свободной от сыворотки средой DМЕМ и инкубировались на льду 10 минут. Клетки предварительно инкубировались в двойном экземпляре Маb 2А1E2 30-45 минут на льду на вращаюащемся шейкере и затем к лункам добавлялся лиганд PDGF ВВ и инкубация продолжалась 1,5-2 часа. Клетки дважды промывались холодным РВS и растворялись либо в буфере Lysis (100 мМ Трис, рН 8,30 мМ пирофосфата натрия, 50 мМ фторида натрия, 5 мМ EGTA, 1% SDS, 100 мМ DTT), либо в IP буфере, оба содержали 1 мМ РМSF (фенилметилсульфонилфторид) и 1 μМ ортованадата натрия. Образцы обрабатывались далее до электрофореза.
Митогенез в человеческих клетках AG01523В фибробласта крайней плоти и клетки гладкой мускулатуры павиана. Человеческие клетки АG01523В фибробласта крейней плоти и клетки гладкой мускулатуры павиана выращивались вместе в 96-лунковых чашках. Клетки гладкой мускулатуры павиана приводились в состояние покоя инкубацией в течение ночи средой DМЕМ, содержащей 0,5% телячьей сыворотки. Затем клетки ингибировались в тройном экземпляре различными концентрациями МАb 2А1E2 в присутствии PDGF ВВ 18 часов при 37oС, а затем 5 часов при 37oС 2 μСi/лунку 3Н-тимидина. Контрольные первичные клетки гладкой мускулатур павиана инкубировались параллельно неспецифическим МАb и контрольные клетки AG01523В инкубировались нетормозящими МАb бета-рецептора антиPDGF (4С5С8). Затем лунки промывались ледяной 5% ТСА (трихлоруксусной кислотой) 2х250 μл) и растворялись 0,25 N NaOH (2х100 μл). Растворенные образцы переносились в сцинтилляционные ампулы и определялась радиоактивность.
Анализ на димеризацию. Слитые HR5 клетки культивировались, как описано выше в 100 мм чашках. Клетки промывались дважды в холодном РВS и инкубировались разными концентрациями МАb 2А1E2 в РВS, содержащем ВSA (1,5 мг/мл) и 25мМ НЕРЕS 1 час при 4oС. К клеткам добавлялся PDGF ВВ и инкубация продолжалась 2 часа при 4oС. Клетки промывались дважды в холодном РВS и инкубировались 30 минут при 4oС кросс-линкером BS3 (0,75 мг/планшет) в РВS, содержащем 25 мМ НЕРЕS. Реакция заканчивалась разбавлением в гасящем буфере (0,025 М Трис, рН 7,4, содержащем 150 мМ NaCl; 10 мл/планшет. Клетки экстрагировались 20 минут при 4oС в IP буфере, содержащем 1 мМ РМSF и 100 μМ ортованадата натрия (0,5 мл/планшет). Лизаты клеток иммуноосаждались всю ночь при 4oС с использованием античеловеческого бета-рецептора Аb (АВ88, разбавление 1: 500). Затем к каждому образцу добавлялось CL4В белка А (60μл 50% суспензии). После 1 часа при 4oС бусины промывались последовательно РВS, содержащим 0,5% NP40, 0,5 М хлорида лития с 0,5% NP40, 0,5 М хлорида лития и, наконец, водой. Образцы растворялись солюбилизирующим буфером Лэммли и подвергались электрофорезу на SDS-полиакриламидном геле с градиентом 3-8% с последующим вестерн-переносом на нитроцеллюлозу. Вестерн-блоты инкубировались или с антифосфотирозинными МАb (1:1000), или с Аb88 (1:500) всю ночь при 4oС, а затем 125I-белком А (0,15 μCi/мл) 2 часа при комнатной температуре. Затем вестерн-блоты подвергались радиоавтографии.
Результаты
Свойства МАb 2А1E2, МАb 2А1E2-моноклональное антитело IgG1, вестерн-анализ показывает, что МАb 2А1E2 опознает невосстановленный бета-рецептор человеческого PDGF (фиг.2, часть А, ряд 4), но не опознает восстановленный белок (фиг. 2, часть А, ряд 1). МАb 2А1E2 иммуноосаждает из раствора внеклеточный рецептор полной длины (рΔ1-5); остатки 1-499) (фиг.2, часть С, ряд 3). Однако, если очищенный рΔ2-7 (без домена 5; остатки 1-384) является антигеном, реактивности нет (фиг.2, часть В, ряд 2).
Дозозависимое ингибирование связывания 125I-PDGF ВВ с клетками НR5 посредством МАb 2А1E2. Если НR5 клетки (СНО-К клетки, которые экспрессируют бета-рецептор человеческого PDGF) инкубированы вначале с МАb 2А1E2, а потом 125I-PDGF ВВ, значительное торможение (48,1%) наблюдается даже при такой низкой концентрации, как 0,1 нМ МАb, в сравнении с клетками, необработанными МАb 2А1E2. Если использовать 1 нМ или большую концентрацию МАb 2А1E2, достигается 100% торможение связывания лиганда с бета-рецептором PDGF полной длины на клетках (фиг. 3, часть А). Если 200 нМ другого нетормозящего МАв (4С5С8) использовать в предварительной инкубации с бета-рецептором PDGF, степень торможения составляет только 36,1%. Это сравнимо с действием 200 нМ нерелевантного МАb (анти-IIb/IIIa) (19,18% торможения). Степень связывания 125I-PDGF ВВ в присутствии 1 нМ МАb 2A1E2 эквивалентна степени связывания лиганда, наблюдаемого в присутствии 1500-кратного избытка немеченного PDGF ВВ (фиг. 3, часть А), или степени связывания лиганда с нетрансфецированными СНО клетками, которые не экспрессируют человеческий рецептор PDGF (данные не показаны).
Чтобы определить, происходит ли наблюдаемое торможение с МАb 2A1E2 благодаря стерическому несоответствию, мы приготовили F(аb1)2 и Fаb фрагменты МАb. Эти протеолитические фрагменты антитела все не опознавали бета-рецептор PDGF в ELISA (данные не показаны), хотя их активность несколько снизилась. Когда их использовали в анализе связывания радиомеченного лиганда, мы выявили, что фрагменты антитела все же тормозили связывание 125I-PDGF ВВ с бета-рецептором PDGF полной длины на НR5 клетках с зависимостью от концентрации (фиг. 3, часть В). Однако полное торможение наблюдалось с 10 нМ МАb 2А1Е2 F(аb1)2 или Fаb фрагментами, а 1 нМ интактного антитела полностью тормозил связывание. Связывание лиганда в присутствии 1 нМ F(аb1)2 фрагментов тормозило на 64,5% и связывание в присутствии 1 нМ Fаb фрагментов тормозилось на 50%.
Торможение фосфорилирования посредством МАb 2А1Е2. Как показано на фиг. 4, МАb 2А1Е2 специфически тормозило вызванное PDGF фосфорилирование в НR5 клетках в зависимости от концентрации с торможением приблизительно в 50% при концентрации 1,3 нМ (ряд 4) и 100% при 13,3 нМ МАb 2А1Е2 (ряд 5). Контрольное нерелевантное МАb не имело действия (ряд 3) и МАb 4С5С8, которое также культивировалось против бета-рецептора человеческого PDGF, но опознавало другой эпитоп, не имело действия на фосфорилирование, вызванное лигандом (ряд 8).
Действие МАb 2А1E2 на вызванную PDGF ВВ димеризацию бета-рецептора человеческого PDGF. Лечение НR5 клеток PDGF ВВ приводит к фосфорилированию с участием лиганда (фиг. 5, белок в 180 кДа в рядах 1-6) и димеризации (фиг.5, белок в 390 кДа в рядах 2 и 6) бета-рецептора PDGF. Если НR5 клетки вначале предварительно инкубировать МАb 2A1E2, а затем PDGF ВВ, димеризация тормозится на всех тестуемых концентрациях (фиг.5, ряды 3, 4 и 5). Эта данные указывают, что связывание МАb 2А1Е2 на рецепторе исключает связывание лиганда и димеризацию рецептора.
Торможение митогенеза МАb 2А1Е2. Как показано на фиг.6, МАb 2А1Е2 тормозит вызванный PDGF ВВ митогенез в клетках АG01523В фибробласта человеческой крайней плоти с зависимостью от концентрации с максимальным торможением (69,55%) при концентрации в 1,3 μМ. Когда использовалось нетормозящее МАb, 4С5С8, мы не обнаружили значительного торможения митогенеза. Повышение концентрации МАb 2А1Е2 не улучшило степень торможения в основном из-за того, что эти клетки также экспрессируют альфа-рецептор PDGF (данные не показаны), который не связывается или не тормозится 2А1Е2.
Мы также определили действие различных концентраций МАb 2А1Е2 на первичные клетки гладкой мускулатуры из артерии павиана. Фосфорилирование рецептора PDGF с участием PDGF ВВ в артериальных клетках гладкой мускулатуры тормозилось 200 нМ и 20 нМ МАb 2А1Е2 (фиг.7, ряды 4 и 5 соответственно).
Как видно из фиг. 8, 1 нМ МАb 2А1Е2 тормозит внедрение 3Н-тимидина в присутствии 1-2 нг/мл PDGF ВВ а 90%, и 25 нМ МАb тормозит митогенез на 80% при концентрациях лиганда от 1 до 10 нг/мл. Концентрации МАb 2А1Е2 более 250 нМ тормозят митогенез на 90% при всех тестуемых концентрациях лиганда. При использовании нерелевантных МАb не наблюдается значительного действия на митогенез в клетках гладкой мускулатуры павиана.
В общем, мы раскрыли моноклональное антитело, МАb 2А1Е2, которое высоко специфично для бета-рецептора человеческого PDGF. МАb 2А1Е2 тормозит связывание PDGF с бета-рецептором PDGF человеческого типа при наномолярных концентрациях и, таким образом, тормозит активацию рецептора, как указано, торможением фосфорилирования и димеризации с участием лиганда. Антитело тормозит митогенез при микромолярных концентрациях. Протеолитические фрагменты МАb сохраняют функцию торможения, как определено торможением связывания 125I-PDGF ВВ (фиг.3А). Наблюдается специфическое и значительное торможение вызванного лигандом аутофосфорилирования бета-рецептора PDGF (фиг.4) при концентрациях, таких низких, как 1,3 нМ МАb 2А1Е2. Следовательно, мы выявили полное торможение митогенеза в НR5 клетках, вызванного PDGF (данные не показаны) при 0,1 М МАb 2А1Е2, и в клетках человеческого фибробласта крайней плоти (АG01523В) 70% торможение достигается при концентрации в 1 μМ МАb 2А1Е2.
МАb 2А1Е2 было также тестировано на перекрестную реактивность с клетками гладкой мускулатуры из плечевой артерии павиана. Фосфорилирование (фиг.7) и митогенез (фиг.8), вызванные PDGF ВВ тормозились до 80% 20-25 нМ МАb 2А1Е2. Моноколональное антитело 2А1Е2 не реагирует перекрестно с рецепторами PDGF от собаки, крысы, мыши или свиньи, а также не реагирует перекрестным образом с альфа-рецептором человеческого PDGF (данные не показаны).
Предлагаемое здесь описание является иллюстративным, но не ограничивающим. Специалистам после ознакомления с описанием будут очевидны многие варианты. Поэтому объем изобретения определяется не описанием, а ссылкой на прилагаемую формулу изобретения с полным объемом эквивалентов, на которые дает право формула.





Изобретение относится к иммунобиотехнологии. Очищенный иммуноглобулиновый полипептид получен с использованием очищенного или частично очищенного внеклеточного домена рецептора человеческого PDGF β-типа. Полипептид специфически связывается с рецептором фактора роста, происходящего из клеток, экспрессирующих рецептор человеческого β-PDGF-R, и не связывается специфически с α-PDGF-R человека. Полипептид представляет собой моноклональное антитело 2А1Е2 и может быть связан с определенной меткой. Фрагмент упомянутого полипептида имеет аминокислотную последовательность, идентичную последовательности области, определяющей его комплементарность. Полипептид или его фрагмент входит в состав композиции для диагностики и лечения нарушений клеточной пролиферации, вызванной PDGF. Полипептид или его фрагмент получают в процессе культивирования экспрессирующей полипептид клеточной линии, преимущественно гибридомы АТСС НВ 109038. 5 с. и 7 з.п.ф-лы, 11 ил.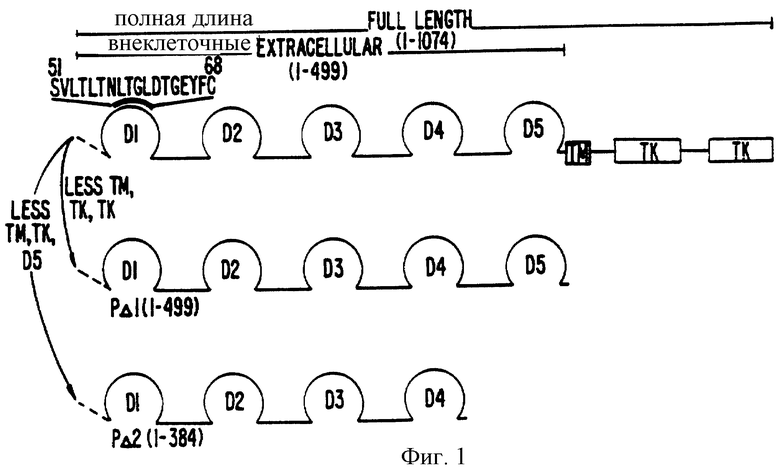
| KAWAHARA R.S | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Biochem | |||
| Biophys | |||
| Res | |||
| Commun., 15.09.1987, v | |||
| Раздвижной паровозный золотник со скользящими по его скалке поршнями и упорными для них шайбами | 1922 |
|
SU147A1 |
