ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к использованию фармацевтической композиции, содержащей пептид, подавляющий аппетит, или фракцию, содержащую пептид, подавляющий аппетит, а также способ достижения регуляции аппетита при помощи этого пептида.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Глюкагон продуцируется панкреатической А-клеткой и выделяется в ответ на низкие уровни глюкозы в крови. Основным местом его действия является печень, где он стимулирует продуцирование глюкозы. Таким образом, он является основным гормоном, противодействующим инсулину в гомеостазе глюкозы крови (Unger, R. H. and L. Orci (1990). Glucagon, in: Diabetes Mellitus, 4th ed., New York, Elsevier, pp. 104-120).
Глюкагон процессируется из более крупного предшественника ограниченным протеолизом. Молекулярное клонирование гена глюкагона показало, что проглюкагоновый предшественник содержал не только глюкагон, но также два дополнительных подобных глюкагону пептида, названных GLP-1 и GLP-2. GLP-1 и GLP-2 кодируются различными оксонами, предполагающими различные биологические активности. Позднее было показано, что проглюкагоновый предшественник подвергался дифференциальному процессингу в трех различных тканях, известных для продуцирования проглюкагона: в панкреатической А-клетке, кишечной L-клетке и в центральной нервной системе (ЦНС). Глюкагон, таким образом, селективно вырезается из предшественника в островковой А-клетке, тогда как GLP-1 и GLP-2 селективно выделяются из кишечной L-клетки и ЦНС [обзор в (Unger, R. H. and L. Orci (1990). Glucagon. in: Diabetes Mellitus, 4th ed., New York, Elsevier, pp. 104-120).
Были идентифицированы специальные рецепторы GLP-1 (Thorens, В. (1992) Proc. Natl. Acad. Sci. USA 89: 8641-8645), которые несомненно отличаются от рецептора глюкагона (L.J. Jelinek, et al. (1993) Science 259: 1614-1616), и они имеют различное распределение в тканях (R. V. Campos, et al. (1994) Endocrinology 134: 2156-2164). GLP-1 высвобождается из L-клетки после приема пищи и функционирует в качестве гормона инкретина (т.е. он потенцирует индуцированное глюкозой выделение инсулина из панкреатической В-клетки). Рецептор GLP-1, таким образом, экспрессируется при высоких уровнях на поверхности островковых В-клеток (К. Moens, et al. (1996) Diabetes 45:257-261).
Была показана индукция кишечной эпителиальной пролиферации посредством GLP-2 (Drucker, D.J. et al. (1996) Proc. Natl. Acad. Sci. USA 93: 7911-7916) и было описано лечение кишечно-желудочных болезней путем роста клеток в среде GLP-2 (Drucker, D. J, and Keneford, J.R., WO 96/32414). До сих пор не было сообщения о рецепторе GLP-2.
Пептиды, происходящие из проглюкагона, и пищевое поведение
Нами ранее сообщался источник и установление трансплантируемых анорексических глюкагоном (О. D. Madsen, et al. (1993), Endocrinology 133: 2022-2030), а также гипогликемических инсулином в крысах (О.D. Madsen, et al. (1988) Proc. Natl. Acad. Sci. USA 85: 6652-6656). Такие опухоли можно получить из обычного клонального источника плюрипотентных MSL-клеток (О.D. Madsen, et al. (1986) J. Cell Biol. 103:2025-2034) и отражают процесс созревания для островковых А-клеток и В-клеток соответственно (О.D. Madsen, et al. (1993) Endocrinology 133:2022-2030).
Связанная с глюкагономой анорексия является очень тяжелым недугом: она имеет острое начало и приводит после нескольких дней к полному прекращению приема пищи. Эта серьезность анорексии совсем не сравнима с другими экспериментальными опухолями в грызунах и предполагает продуцирование глюкагономой очень сильнодействующего фактора насыщения, который действует посредством периферического пути введения. Ранее было показано, что анорексические глюкагономы проявляли нефизиологический процессинг, приводящий к образованию как глюкагона, так и GLP-1 (О.D. Madsen, et al. (1993) Endocrinology 133: 2022-2030). Более того, вариант неанорексической глюкагономы был не способен процессировать этот предшественник (О. D. Madsen, et al. (1995) Scand. J. Clin. Lab. Invest. 55, suppl 220:27-36). Потеря веса также упоминается в качестве компонента синдрома глюкагономы у человека (J.J. Holst (1985) Glucagon-producing tumors, in: Hormone-producing tumors of the gastrointestinal tract. New York, Churchill Livingstone, pp. 57-84), несмотря на высокую степень изменчивости ее у разных пациентов (S.J. Bhathena et al. (1981). Glucagonoma and glucagonoma syndrome, in: Glucagon. Physiology, pathophysiology and morphology of the pancreatic A-cells. New York, Elsevier, 413-438).
Глюкагон
Было показано, что глюкагон участвует в регуляции величины спонтанного приема пищи у крыс, но общее действие его минимально, и оказывает действие он через вагусные соединения с печенью (N. Geary, et al. (1993) Am. J. Physiol. 264; R116-R122). Это действие наблюдают только посредством печеночной инфузии глюкагона через воротную вену, тогда как внутрибрюшинное введение фармакологических доз не обнаруживает действия его на прием пищи у голодных крыс (О.D. Madsen, et al. (1993) Endocrinology 133:2022-2030).
GLP-1
Недавно сообщалось о центральной роли GLP-1 в регуляции кормления (М.D. Turton, et al. (1996) Nature 379: 69-72). Интрацеребровентрикулярное (ICV) введение GLP-1 ингибировало кормление у голодных крыс. И снова периферическое введение GLP-1 не оказывало влияния на пищевое поведение (М.D. Turton, et al. (1996) Nature 379: 69-72; O.D. Madsen, et al. (1993) Endocrinology 133: 2022-2030), предполагая, что опухоль, вызванная GLP-1, не может значительно содействовать наблюдаемой анорексии.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Было обнаружено, что GLP-2 оказывает сильное действие на ингибирование потребления пищи при введении периферически.
Предполагается, что GLP-2, обычно выделяемый вместе с GLP-1 из кишечных L-клеток, играет свою собственную отчетливую роль в качестве периферического фактора насыщения.
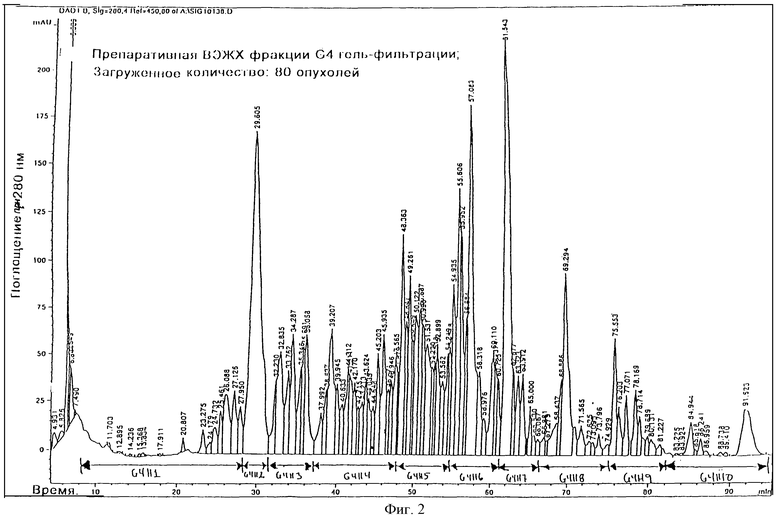
В соответствии с этим настоящее изобретение относится к использованию фармацевтической композиции, содержащей вместе с фармацевтически приемлемой средой или наполнителем фракцию ВЭЖХ экстракта глюкагономной опухоли, полученную экстракцией подкисленным этанолом, гель-фильтрацией и препаративной ВЭЖХ, причем эта фракция показана как фракция G4H9 на фиг.2 и содержит подобный глюкагону пептид 2 (GLP-2) в качестве основного компонента (т.е. более чем 40%) или содержащий любой один компонент этой фракции или комбинацию двух или более компонентов этой фракции.
В другом аспекте данное изобретение относится к использованию фармацевтической композиции, содержащей подобный глюкагону пептид-2 (GLP-2) или его вариант или гомолог, для профилактики или лечения болезней или нарушений, связанных с нарушенной регуляцией аппетита.
В другом аспекте данное изобретение относится к использованию фармацевтической композиции, содержащей пептид со следующей аминокислотной последовательностью
X1HX2DGSFSDEMNTX3LDX4LAX5X6DFINWLX7X8ТКIТDX9,
где Х1 представляет NH2, DFPEEVAIVEELGRR, DFPEEVTIVEELGRR, DFPEEVNIVEELRRR или его фрагмент,
X2 представляет Аlа или Gly,
X3 представляет Ilе или Val,
X4 представляет Asn, Ser или His,
X5 представляет Ala или Thr,
X6 представляет Arg или Lys,
X7 представляет Ilе или Leu,
X8 представляет Gln или His, или
X9 представляет ОН, Lys, Arg, Arg-Lys, Lys-Arg, Arg-Arg или Lys-Lys, для профилактики или лечения болезней или нарушений, связанных с нарушенной регуляцией аппетита.
Еще в одном аспекте это изобретение относится к способу лечения болезней или нарушений, связанных с нарушенной регуляцией аппетита, причем этот способ предусматривает введение индивидууму, нуждающемуся в таком лечении, количества пептида, как указано здесь, достаточного для подавления аппетита или вызывания насыщения у этого индивидуума.
Еще в одном аспекте это изобретение относится к использованию пептида, указываемого здесь, для изготовления лекарственного средства для профилактики или лечения болезней или нарушений, связанных с нарушенной регуляцией аппетита.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Предполагается, что в настоящем описании термин "пептид" включает зрелый пептид GLP-2 или его предшественник, а также его функциональный фрагмент, который по существу имеет активность этого пептида с полной длиной. Кроме того, термин "пептид" предназначается для включения гомологов этого пептида. Такие гомологи содержат аминокислотную последовательность, проявляющую степень идентичности по меньшей мере 50%, такую как по меньшей мере 75%, и более конкретно, по меньшей мере 90% идентичности с аминокислотной последовательностью GLP-2. Степень идентичности можно определить общепринятыми способами, см., например, Altshul et al., Bull. Math. Bio. 48:603-616, 1986, and Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA 89: 10915-10919, 1992.
Гомологи данного пептида могут иметь одну или несколько аминокислотных замен, делеций или инсерций. Эти изменения могут быть незначительными, т.е. являются консервативными аминокислотными заменами, которые не оказывают существенного влияния на складывание или активность пептида, небольшими делециями, типично от одной до пяти аминокислот, небольшими амино- или карбоксил-концевыми удлинениями, такими как амино-концевой метиониновый остаток, небольшой линкерный пептид, содержащий вплоть до 15 остатков, или небольшой вставкой, которая облегчает очистку, такой как полигистидиновый тракт, антигенный эпитоп или связывающий домен. Вообще см. Ford et al.. Protein Expression and Purification 2: 95-107, 1991. Примеры консервативных замен находятся в пределах группы основных аминокислот (аминокислот основного характера) (таких как аргинин, лизин, гистидин), кислотных аминокислот (таких как глутаминовая кислота и аспарагиновая кислота), полярных аминокислот (таких как глутамин и аспарагин), гидрофобных аминокислот (таких как лейцин, изолейцин, валин), ароматических аминокислот (таких как фенилаланин, триптофан, тирозин) и небольших аминокислот (таких как глицин, аланин, серин, треонин, метионин).
Гомолог может быть аллельным вариантом, т.е. альтернативной формой гена, который возникает посредством мутации, или модифицированным пептидом, кодированным мутированным геном, но имеющим по существу ту же самую активность, что и нативный пептид GLP-2. Следовательно, мутации могут быть молчащими (без изменения в кодированном пептиде) или могут кодировать пептиды, имеющие модифицированную аминокислотную последовательность.
Гомологом настоящего пептида может быть также видовой гомолог, т.е. пептид с аналогичной активностью, полученный из другого вида. Примерами видовых гомологов пептида GLP-2 являются человечий, бычий, крысиный, хомячковый, свиной GLP-2 и GLP-2 морских свинок.
В предпочтительном осуществлении настоящего изобретения пептид GLP-2 является пептидом, у которого Х1 представляет NH2, X2 представляет Аlа, X3 представляет Ilе, X4 представляет Аsn, X5 представляет Аlа, X6 представляет Аrg, X7 представляет Ilе, X8 представляет Gln или X9 представляет ОН. В частности, пептид имеет следующую аминокислотную последовательность.
HADGSFSDEMNTILDNLAARDFINWLIQTKI
ТD (GLP-2 человека), или
HADGSFSDEMNTILDNLATRDFINWLIQTKI
ТD (крысиный GLP-2), или
HADGSFSDEMNTVLDNLATRDFINWLLHTKIТD (свиной GLP-2).
Гомолог пептида можно выделить путем получения геномной или кДНК-библиотеки клетки нужного вида и скрининга на ДНК-последовательность, кодирующую весь или часть гомолога, с использованием синтетических олигонуклеотидных зондов в соответствии со стандартными способами, например, как описано Sambrook et al. , Molecular Cloning: A Laboratory Manual, 2nd. Ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1989, или при помощи полимеразной цепной реакции (ПЦР) с использованием специфических праймеров, как описано Sambrook et al., см. выше.
Настоящее изобретение относится также к композиции, содержащей вариант пептида GLP-2. Этот вариант является вариантом, в котором один или несколько аминокислотных остатков были заменены на другие аминокислотные остатки. В конкретном предпочтительном осуществлении Аlа был замещен на Gly в положении 2 зрелого пептида. Ожидается, что этот вариант будет обладать более долгим временем полужизни в плазме, чем нативный пептид и имеет преимущество, поскольку доза, требуемая для получения адекватного подавляющего аппетит или вызывающего насыщение эффекта, будет обычно меньше.
Пептид GLP-2 или его гомолог или вариант, как указано выше, можно получить техникой рекомбинантных ДНК в соответствии с методиками, хорошо упрочившимися в этой области.
Более конкретно, ДНК-последовательность, кодирующую пептид GLP-2, можно выделить или синтезировать на основе опубликованной ДНК-последовательности препроглюкагона человека (сравни J.W. White et al., Nucleic Acids Res. 14, 1986, pp. 4719-4730; G.I. Bell et al. Nature 304, 1983, pp. 368-371), например, полученной посредством получения геномной или кДНК-библиотеки из подходящей ткани и скрининга ДНК последовательностей, кодирующих весь или часть пептида GLP-2 путем гибридизации с использованием синтетических олигонуклеотидных зондов в соответствии со стандартными методиками (cp. Sambrook et al. , см. выше). Для настоящей цели предпочтительна кодирующая пептид GLP-2 ДНК-последовательность человеческого происхождения.
ДНК-конструкцию, кодирующую пептид GLP-2, можно также получить синтетически принятыми стандартными способами, например способом фосфоамидита, описанным Beaucage and Caruthers, Tetrahedron Letters 22 (1981), 1859-1869, или способом, описанным Matters et al., EMBO Journal 3 (1984), 801-805. В соответствии со способом фосфоамидита, олигонуклеотиды синтезируют, например, в автоматическом ДНК-синтезаторе, очищают, ренатурируют, лигируют и клонируют в подходящих векторах.
Кроме того, ДНК-конструкция может быть смешанной синтетической и геномной, смешанной синтетической и кДНК или смешанной геномной и кДНК, полученной лигированием фрагментов синтетической, геномной или кДНК (что подходит), причем эти фрагменты соответствуют различным частям полной ДНК-конструкции, в соответствии со стандартными техниками.
ДНК-конструкцию можно также получить полимеразной цепной реацией с использованием специфических праймеров, например, как описано в патенте США 4683202 или Saiki et al., Science 239 (1988), 487-491, или Sambrook et al., см. выше.
В предпочтительном к настоящему времени осуществлении ДНК-конструкция содержит ДНК-последовательность, показанную на фиг.3 публикации G.I. Bell et al. , Nature 304, 1983, pp. 368-371, а также последовательности нуклеиновых кислот, кодирующие GLP-2 человека, но которые отличаются от ДНК-последовательности, показанной на фиг.3 публикации Bell et al, см. выше, вследствие вырожденности генетического кода. ДНК-конструкция дополнительно включает последовательности нуклеиновых кислот, которые гибридизуют с молекулой нуклеиновой кислоты (либо геномной, синтетической или кДНК, либо РНК), кодирующей GLP-2 человека в очень строгих условиях (т.е. предварительным пропитыванием в 5хSSC и предварительной гибридизацией в течение 1 ч приблизительно при 40oС в растворе 20% формамида, 5х раствора Denhardt, 50 мМ фосфата натрия, рН 6,8, и 50 мкг денатурированной, обработанной ультразвуком ДНК бычьего тимуса с последующей гибридизацией в том же самом растворе, дополненном 100 мкМ АТФ, в течение 18 ч приблизительно при 40oС, с последующим промыванием в 0,4хSSC при температуре около 45oС). Она может быть, например, ДНК-последовательностью, кодирующей GLP-2 из других видов, например крысиного, бычьего, хомячкового, свиного GLP-2 или GLP-2 морской свинки.
Для экспрессии GLP-2 ДНК-конструкцию, кодирующую пептид GLP-2, встраивают в подходящий рекомбинантный вектор. Он может быть любым вектором, который может быть подходящим образом подвергнут процедурам, производимым с рекомбинантными ДНК, и выбор вектора будет часто зависеть от клетки-хозяина, в которую он должен быть введен. Таким образом, вектор может быть автономно реплицирующимся вектором, т.е. вектором, который существует как экстрахромосомная единица, репликация которой не зависит от хромосомной репликации, т.е. плазмидой. Альтернативно, вектор может быть вектором, который при введении в клетку-хозяина интегрируется в геном этой клетки-хозяина и реплицируется вместе с хромосомой(ами), в которую он был интегрирован.
Вектор предпочтительно является экспрессирующим вектором, в котором ДНК-последовательность, кодирующая пептид GLP-2, надлежащим образом связана с дополнительными сегментами, необходимыми для транскрипции ДНК. Вообще экспрессирующий вектор происходит из плазмиды или вирусной ДНК, или он может содержать элементы того и другого. Термин "надлежащим образом связанный" означает, что сегменты расположены так, что они функционируют в соответствии с их нужными целями, например транскрипция инициируется промотором и продолжается по ДНК-последовательности, кодирующей этот пептид.
Промотором может быть любая ДНК-последовательность, которая проявляет транскрипционную активность в выбранной клетке-хозяине, и он может происходить из генов, кодирующих белки, либо гомологичные, либо гетерологичные клетке-хозяину.
Примерами подходящих промоторов для направления транскрипции ДНК, кодирующей пептид GLP-2 в клетках млекопитающих, являются промотор SV40 (Subramani et al., Mol. Cell Biol., 1 (1981), 854-864), промотор МТ-1 (ген металлотионеина) (Palmiter et al. , Science 222 (1983), 809-814) главный поздний промотор аденовируса 2.
Примером подходящего промотора для использования в клетках насекомых является промотор полиэдрина (патент США 4745051; Vasuvedan et al., FEBS Lett. 311 (1992) 7-11); промотор Р10 (J.M. Viak et al., J.Gen. Virology 69, 1988, pp. 765-776), промотор основного белка вируса полиэдроза Autographa californica (Европейский патент 397485), промотор немедленно-раннего гена 1 бакуловируса (патент США 5155037; патент США 5162222) или промотор задержанно-раннего гена 39К бакуловируса (патент США 5155037; патент США 5162222).
Примеры подходящих промоторов для использования в дрожжевых клетках-хозяевах включают промоторы из гликолитических генов дрожжей (Hitzeman et al. , J. Biol. Chem., 255 (1980), 12073-12080; Alber and Kawasaki, J. Mol. Appl. Gen. 1 (1982), 419-434) или генов алкогольдегидрогеназы (Young et al., in Genetic Engineering of Microorganisms for Chemicals (Hollaender et al., eds. ), Plenum Press, New York, 1982) или промоторы ТPI1 (патент США 4599311), или ADH2-4c (Russel et al., Nature 304 (1983), 652-654).
Примерами подходящих промоторов для использования в клетках-хозяевах нитчатых грибов являются, например, промотор ADH3 (McKnight et al.. The EMBO J. 4 (1985), 2093-2099) или промотор tpiA. Примерами других пригодных промоторов являются промоторы, полученные из гена, кодирующего ТАКА-амилазу А. oryzae, аспарагиновую протеиназу Rhizomucor miehei, нейтральную α-амилазу A. niger, устойчивую к действию кислоты, α-амилазу А. niger или глюкоамилазу А niger или A. awamori (gluA), липазу Rhizomucor miehei, щелочную протеазу А. oryzae, триозофосфатизомеразу A. oryzae, или ацетамидазу А. nidylans. Предпочтительны промоторы ТАКА-амилазы и gluA.
Примеры подходящих промоторов для использования в бактериальных клетках-хозяевах включают промотор гена мальтогенной амилазы Bacillus stearothermophilus, гена альфа-амилазы Bacillus lichenifor mis, гена BAN-амилазы Bacillus amylol iquefaciens, гена щелочной протеазы Bacillus subtilis или гена ксилодазы Bacillus pumilus или промоторы фага Lambda PR или РL или промоторы lac, trp или tac E.coli.
ДНК-последовательность, кодирующая пептид GLP-2, может быть также, если нужно, надлежащим образом соединена с подходящим терминатором, таким как терминатор гормона роста человека (Palmiter et al., op. cit.) или (для грибковых хозяев) терминаторы TPI1 (Alber and Kawasaki, op. cit.) или ADH3 (McKnight et al., op. cit.). Вектор может дополнительно содержать элементы, такие как сигналы полиаденилирования (например из SV40 или области 5 Elb аденовируса), последовательности транскрипционного энхансера (например, энхансера SV40) и последовательности трансляционного энхансера (например, последовательности, кодирующие РНК аденовируса VA).
Рекомбинантный вектор может, кроме того, содержать ДНК-последовательность, дающую возможность вектору реплицироваться в нужной клетке-хозяине. Примером такой последовательности (когда клеткой-хозяином является клетка млекопитающего) является начало репликации SV40.
Когда клеткой-хозяином является дрожжевая клетка, подходящими последовательностями, дающими возможность вектору реплицироваться, являются гены репликации REP 1-3 и начало репликации дрожжевой плазмиды 2μ.
Вектор может содержать также селектируемый маркер, например ген, продукт которого комплементирует дефект в клетке-хозяине, такой как ген, кодирующий дигидрофолатредуктазу (DHFR) или ген TPI Schizosaccharomyces pombe (описаны Р. R. Russel, Gene 40, 1985, pp. 125-130), или ген, который несет устойчивость к лекарственному средству, например ампициллину, канамицину, тетрациклину, хлорамфениколу, неомицину, гигромицину или метотрексату. Для нитчатых грибов селектируемые маркеры включают amdS, pyrG, argB, niaD, sC.
Для направления пептида GLP-2 в секреторный путь клеток-хозяев секреторная сигнальная последовательность (известна также как лидерная последовательность, препропоследовательность или препоследовательность) может быть обеспечена в рекомбинантном векторе. Секреторную сигнальную последовательность соединяют с ДНК-последовательностью, кодирующей этот пептид в правильной рамке считывания. Секреторные сигнальные последовательности обычно расположены у 5'-конца ДНК-последовательности, кодирующей пептид. Секреторная сигнальная последовательность может быть последовательностью, обычно ассоциированной с пептидом, или может происходить из гена, кодирующего другой секретируемый белок.
Для секреции из дрожжевых клеток секреторная сигнальная последовательность может кодировать любой сигнальный пептид, который обеспечивает эффективное направление экспрессированного пептида в секреторный путь клетки. Сигнальный пептид может быть существующим в природе сигнальным пептидом или его функциональной частью или может быть синтетическим пептидом. Обнаружено, что подходящие сигнальные пептиды являются сигнальным пептидом α-фактора (ср. Патент США 4870008), сигнальным пептидом мышиной амилазы слюны (ср. О. Hagenbuchle et al., Nature 289, 1981, pp. 643-646), модифицированным сигнальным пептидом карбоксипептидазы (ср. L.A. Valls et al., Cell 48, 1987, pp. 887-897), дрожжевым сигнальным пептидом BAR1 (ср. WO/02670) или сигнальным пептидом дрожжевой аспарагиновой протеазы 3 (YAP3) (ср. М. Egel-Mitani et al., Yeast 6, 1990, pp. 127-137).
Для эффективной секреции в дрожжах последовательность, кодирующую лидерный пептид, можно также встроить перед сигнальной последовательностью и перед ДНК-последовательностью, кодирующей пептид GLP-2. Функцией лидерного пептида является позволение экспрессированному пептиду направляться из эндоплазматического ретикулума к аппарату Гольджи и далее к секреторной везикуле для секреции в культуральную среду (т.е. экспорт пептида через клеточную стенку или по меньшей мере через клеточную мембрану в периплазматический участок дрожжевой клетки). Лидерный пептид может быть лидером α-фактора дрожжей (использование которого описывается, например, в патенте США 4546082, Европейском патенте 16201, Европейском патенте 123294, Европейском патенте 123544 и Европейском патенте 163529). Альтернативно, лидерный пептид может быть синтетическим лидерным пептидом, который является, например, лидерным пептидом, не обнаруженном в природе. Синтетические лидерные пептиды можно, например, конструировать, как описано в WO 89/02463 или WO 92/11378.
Для использования в нитчатых грибах сигнальный пептид можно подходящим образом получить из гена, кодирующего амилазу или гликоамилазу Aspergillus sp., гена, кодирующего липазу или протеазу Rhizomucor miehei, липазу Hemicola lanuginosa. Сигнальный пептид предпочтительно получают из гена, кодирующего амилазу А. oryzae ТАКА, нейтральную α-амилазу A. niger, устойчивую к действию кислоты амилазу А. niger и глюкоамилазу А. niger.
Для использования в клетках насекомых сигнальный пептид можно подходящим образом получить из гена насекомого (ср. WO 90/05783), такой как сигнальный пептид предшественника адипокинетического гормона чешуекрылого Manduca sexta (ср. Патент США 5023328).
Методики, использованные для лигирования ДНК-последовательностей, кодирующих пептид GLP-2, промотор и необязательно терминатор и/или секреторную сигнальную последовательность соответственно и встраивания их в подходящие векторы, содержащие информацию, необходимую для репликации, хорошо известны специалистам в данной области (ср., например, Sambrook et al., op. cit.).
ДНК-последовательность, кодирующая пептид GLP-2, введенная в клетку-хозяина, может быть гомологичной или гетерологичной к нужному хозяину. Если она гомологична к клетке-хозяину, т.е. продуцируется клеткой-хозяином в природе, она обычно будет надлежащим образом соединена с другой промоторной последовательностью, или, если это возможно, с иной, чем в ее природном окружении секреторной сигнальной последовательностью и/или терминаторной последовательностью. Термин "гомологичный" предназначается для включения кДНК-последовательности, кодирующей нужный полипептид, нативный для организма-хозяина. Термин "гетерологичный" предназначается для включения ДНК-последовательности, не экспрессируемой клеткой-хозяином в природе. Таким образом, ДНК-последовательность может быть из другого организма или может быть синтетической последовательностью.
Клеткой-хозяином, в которую вводят ДНК-конструкцию или рекомбинантный вектор этого изобретения, может быть любая клетка, которая способна продуцировать данный пептид, она включает клетки бактерий, дрожжей, грибов и высшие эукариотические клетки.
Примерами бактериальных клеток-хозяев, которые при культивировании способны продуцировать пептид GLP-2, являются грамположительные бактерии такие как штаммы Bacillus, например штаммы В. subtilis, В. licheniformis, В. lentus, В. brevis, В. stearothermophilus, В. alkalophilus, В. amyloliquefaciens, В. coagulans, В. circulans, В. lautus, В. megatherium или В. thuringiensis, или штаммы Streptomyces, такие как S.lividans или S.murinus, или грамотрицательные бактерии, такие как Echerichia coli. Трансформацию бактерий можно проводить путем трансформации протопласта или использования компетентных клеток способом, известным самим по себе (ср. Sambrook et al., см. выше).
При экспрессии данного пептида в бактерии, такой как Е. coli, пептид может удерживаться в цитоплазме обычно в виде нерастворимых гранул (известных как тельца включения) или может направляться к периплазматическому пространству последовательностью бактериальной секреции. В первом случае клетки лизируют и гранулы выделяют и денатурируют, после чего пептид снова "снова укладывают" путем разбавления денатурирующим агентом. В последнем случае пептид можно выделить из периплазматического пространства разрывом клеток, например обработкой ультразвуком или осмотическим шоком, для высвобождения содержимого периплазматического пространства и выделения пептида.
Примерами подходящих клеточных линий млекопитающих являются клеточные линии COS (ATCC CRL 1650), ВНК (АТСС CRL 1632, АТСС CCL 10), CHL (ATCC CCL39) или СНО (АТСС CCL 61). Способы трансфекции клеток млекопитающих и экспрессии ДНК-последовательностей, введенных в клетки, описываются, например, в публикациях Kaufman and Sharp, J. Mol. Biol. 159 (1982), 601-621; Southern and Berg, J. Mol. Appl. Genet. 1 (1982), 327-341; Loyter et al., Proc. Natl. Acad. Sci. USA 79 (1982), 422-426; Wigler et al., Cell 14 (1978), 725; Corsaro and Pearson, Somatic Cell Genetics 7 (1981), 603, Graham and van der Eb, Virology 52 (1973), 456; and Neumann et al., EMBO J. 1 (1982), 841-845.
Примеры подходящих дрожжевых клеток включают клетки Saccharomyces spp. или Schizosaccharomyces spp., особенно штаммы Saccharomyces cerevisiae или Saccharomyces kluyveri. Способы трансформации дрожжевых клеток гетерологичной ДНК и продуцирования из них генерологичных полипептидов описываются, например, в патенте США 4599311, патенте США 4931373, патентах США 4870008, 5037743 и патенте США 4845975, все из которых, таким образом, вводятся в качестве ссылки. Трансформированные клетки отбирают определением фенотипа посредством отбираемого маркера, обычной устойчивости к лекарственному средству или способности расти в отсутствие определенного питательного вещества, например лейцина. Предпочтительным вектором для использования в дрожжах является вектор РОТ1, описанный в патенте США 4931373. ДНК-последовательности, кодирующей пептид GLP-2, может предшествовать сигнальная последовательность и необязательно лидерная последовательность, например, как описано выше. Следующими примерами подходящих дрожжевых клеток являются штаммы Kluyveromyces, такие как К. lactis, Hansenula, например, Н. polymorpha, или Pichia, например, Р. pastoris (ср. Gleeson et al., J. Gen. Microbiol. 132, 1986, pp. 3459-3465; патент США 4882279).
Примерами других грибковых клеток являются клетки нитчатых грибов, например Aspergillus spp., Neurospora spp., Fusarium spp. или Trichoderma spp., особенно штаммы А. oryzae, A. nidulans или A. niger. Использование Aspergillus spp. для экспрессии белков описывается, например, в Европейском патенте 272277 и Европейском патенте 230023. Трансформацию F. oxysporum можно, например, проводить, как описано Malardier et al., 1989, Gene 78: 147-156.
Когда в качестве клетки-хозяина используют нитчатые грибы, ее можно трансформировать подходящим образом ДНК-конструкцией, кодирующей пептид GLP-2, путем интегрирования ДНК-конструкции в хромосому хозяина для получения рекомбинантной клетки-хозяина. Обычно считается, что эта интеграция выгодна, когда более вероятно, что ДНК-последовательность стабильно сохраняется в клетке. Интеграцию ДНК-конструкции в хромосому хозяина можно проводить по обычным способам, например гомологичной или гетерологичной рекомбинацией.
Трансформацию клеток насекомых и продуцирование гетерологичных полипептидов в них можно проводить, как описано в патенте США 4745051; патенте США 4879236; патентах США 5155037; 5162222; Европейском патенте 397485, все из которых вводятся здесь ссылкой. Клеточными линиями насекомых, используемыми в качестве хозяина, приемлемо могут быть клеточные линии Lepidoptera, такие как клетки Spodoptera frugiperda или клетки Trichoplusia ni (ср. Патент США 5077214). Подходящие условия культивирования могут быть такие, как описаны, например, в WO 89/01029 или WO 89/01028, или любой из вышеуказанных ссылок.
Трансформированную или трансфикцированную клетку-хозяина, описанную выше, затем культивируют в подходящей питательной среде в условиях, позволяющих экспрессию пептида GLP-2, после чего полученный пептид GLP-2 выделяют из культуры.
Средой, используемой для культивирования клеток, может быть любая общепринятая среда, пригодная для роста клеток-хозяев, такая как минимальная или комплексная среда, содержащая подходящие добавки. Пригодны подходящие среды из коммерческих поставок, или их можно получить в соответствии с опубликованными рецептами (например, в каталоге American Type Culture Collection). Пептид GLP-2, продуцированный клетками, можно затем выделить из культивированной среды общепринятыми методиками, включая отделение клеток-хозяев от среды центрифугированием или фильтрацией, осаждение белковых компонентов супернатанта или фильтрата при помощи соли, например сульфата аммония, очистку различными хроматографическими методиками, например ионообменной хроматографией, гель-фильтрационной хроматографией, афинной хроматографией или тому подобное.
В фармацевтической композиции данного изобретения пептид GLP-2 может быть изготовлен любым из принятых способов изготовления фармацевтических композиций, например, как описано в Remington's Pharmaceutical Sciences, 1985. Композиция может быть в форме, подходящей для системной инъекции или инфузии, и может быть, как таковая, изготовлена с подходящим жидким наполнителем, таким как стерильная вода, или изотонический солевой раствор, или раствор глюкозы. Композиции можно стерилизовать общепринятыми методиками стерилизации, которые хорошо известны в данной области. Получаемые водные растворы можно упаковывать для использования или фильтровать в асептических условиях и лиофилизовать, причем лиофилизованный препарат смешивают со стерильным водным раствором перед введением. Композиция может содержать фармацевтически приемлемые вспомогательные вещества, которые требуются для создания близких физиологических условий, такие как буферные агенты, агенты, устанавливающие тоничность, и тому подобное, например ацетат натрия, лактат натрия, хлорид натрия, хлорид калия, хлорид кальция и так далее.
Фармацевтическая композиция настоящего изобретения может быть также приспособлена для назального, чрескожного, легочного или ректального введения. Фармацевтически приемлемым носителем или разбавителем, применяемым в композиции, может быть любой общепринятый твердый носитель. Примерами твердых носителей является лактоза, сульфат кальция, сахароза, тальк, желатин, агар, пектин, акация, стеарат магния и стеариновая кислота. Подобным же образом носитель или разбавитель может включать любой пролонгирующий высвобождение материал, известный в данной области, такой как глицерилмоностеарат или глицерилдистеарат, отдельно или в смеси с воском.
Может быть особенно выгодным получение композиции по данному изобретению в виде готовой препаративной формы с пролонгированным действием. Как таковую композицию можно изготовить в виде микрокапсул или микрочастиц, содержащих GLP-2, капсулированный подходящим фармацевтически приемлемым биоразрушаемым полимером или диспергированный в этом полимере, таком как полимолочная кислота, полигликолевая кислота или сополимер молочной кислоты и гликолевой кислоты.
Для назального введения препарат может содержать пептид GLP-2, растворенный или суспендированный в жидком носителе, особенно в водном носителе, для аэрозольного применения. Носитель может содержать добавки, такие как солюбилизирующие агенты, например пропиленгликоль, поверхностно-активные вещества, усилители абсорбции, такие как лецитин (фосфатидилхолин), или циклодекстрин, или консерванты, такие как парабены.
Обычно композиции настоящего изобретения приготовляют в виде унифицированной лекарственной формы, содержащей 0,5-5,00 мг данного пептида вместе с фармацевтически приемлемым носителем на унифицированную дозу.
Предполагается, что пептид GLP-2 благоприятен для использования подавления аппетита и обеспечении чувства насыщения, например, для профилактики или лечения болезней или нарушений, связанных с регуляцией нарушенного аппетита. Примерами таких болезней или нарушений является ожирение и диабет типа II. Доза пептида GLP-2, вводимого пациенту, будет варьировать в зависимости от типа и тяжести состояния, которое нужно лечить, но обычно она находится в диапазоне от около 10 мкг/кг до около 5 мг/кг массы тела.
В фармацевтической композиции этого изобретения пептид GLP-2 можно комбинировать с другим средством, подавляющим аппетит или вызывающим насыщение. Примером такого средства является GLP-1, который, как было показано, оказывает некоторое действие на подавление аппетита (см. M.D. Turton et al., Nature 379, 4 Januare 1996, pp. 69-72).
Далее предполагается, что пептид GLP-2 в подходящей меченой форме, например радиомеченый GLP-2, можно использовать для идентификации рецептора для GLP-2 в исследованиях по связыванию с использованием ткани(ей), которые, как ожидается, экспрессируют рецептор GLP-2, например ткани гипоталамуса. После локализации связыванием белком GLP-2 рецептор можно клонировать путем экспрессивного клонирования, т. е. путем приготовления кДНК-библиотеки нужной ткани, клонирования кДНК в подходящие векторы и введения векторов в подходящую клетку для осуществления экспрессии кДНК, после чего клон, экспрессирующий рецептор, идентифицируют путем связывания с GLP-2. Клеточную линию, стабильно экспрессирующую рецептор, можно затем использовать в анализе скринирования на агонисты GLP-2 (т.е. соединения, действующие на рецептор, чтобы вызвать насыщение или подавить аппетит) или антагонисты GLP-2 (т.е. соединения, которые создают антагонизм действию GLP-2 на рецептор, например для использования при лечении злокачественной анорексии или нервной анорексии).
Изобретение далее иллюстрируется следующими примерами, которые никоим образом не предназначены для ограничения объема заявляемого изобретения.
Пример 1
Экстракция опухолевой ткани подкисленным этанолом
Аноректические опухоли вызывали у крыс, как описано ранее (Madsen, O.D., et al. (1993) Endocrinology 133, 2022-2030). Пятьдесят аноректических опухолей 12C3AN (MSL-G-AN) (при -80oС), соответствующих 50,07 г сырой ткани, гомогенизировали при 4oС с 700 мл подкисленного этанола (96% этанол/0,7 М НСl, 3/1, об./об.). Гомогенизацию проводили в течение 5 мин в предварительно охлажденном (4oС) Waring Commercial-измельчителе на 2 литра при максимальной скорости. После гомогенизации смесь перемешивали при 4oС в течение 16 часов. Смесь центрифугировали при 9000 об/мин при 4oС в течение 1 часа. Объем супернатанта снижали до 20% при помощи вакуумного роторного испарителя. Во время этой операции, в которой основная часть этанола удаляется, образуется некоторое количество осадка. Этот осадок удаляли центрифугированием при 4oС в течение одного часа при 20000 об/мин. Супернатант, который все еще содержал некоторое количество подобного липиду материала, фильтровали и вводили к колонку LiChroprep RP-18 (Merck) (2,5х10 см), уравновешенную 0,1% трифторуксусной кислотой (ТФУ), при скорости потока 2 мл/мин. Колонку промывали 100 мл 0,1% ТФУ при скорости потока 4 мл/мин. Связанный материал элюировали 400 мл 0,1% ТФУ, содержащей 70% (об./об.) ацетонитрила. Ацетонитрил удаляли при помощи вакуумного роторного испарителя и получаемую смесь лиофилизовали. После лиофилизации материал растворяли в 50 мл воды и рН устанавливали до 5,3 с использованием 425 мкл 1н. NaOH. Дальнейшее титрование смеси до рН 6,0 приводило к образованию осадка. При обратном титровании до рН 5,3 этот осадок снова растворялся. Поэтому рН оставляли у значения 5,3 и смесь лиофилизовали.
Общий выход лиофилизованного материала из 50 опухолей был 359 мг сухого порошка.
Пример 2
Первая стадия очистки: гель-фильтрация на сефадексе G-75
Лиофилизованный материал (278 мг) из экстракта подкисленным этанолом, соответствующий 38 отдельным опухолям, снова растворяли в 20 мл 1 М НАс и вводили в колонку сефадекса G-75 (5х50см). Колонку уравновешивали и элюировали 1 М НAc при скорости потока 55 мл/ч и собирали фракции, соответствующие 10 мл элюата. Для каждой фракции регистрировали поглощение у 280 нм. Хроматограмма гель-фильтрации приводится на фиг.1. Отдельные фракции объединяли в следующие 5 основных фракций (пулы): G1 (фр.30-39), G2 (фр.40-45), G3 (фр. 46-66), G4 (фр. 67-91) и G5 (фр.92-118) и подвергали биоанализу после лиофилизации.
Пример 3
Вторая стадия очистки: Препаративная ВЭЖХ объединенной фракции (пула) G4
Некоторые из подавляющих аппетит пулов гель-фильтрации проявляли такую активность, которая должна присутствовать в пуле G4, этот пул далее фракционировали препаративной ВЭЖХ. Лиофилизованный материал G4 (соответствующий 80 опухолям) снова растворяли в 15 мл 0,1% ТФУ и при помощи насоса вводили в колонку Vydac 214ТР1022 С4 (2,2х25 см), уравновешенную в 0,1% ТФУ. Колонку промывали 20 мл 0,1% ТФУ, затем 100 мл МеСN/Н2O/ТФУ (10,0:89,9:0,1, об./об. /об. ). Материал элюировали при 25oС и скорости потока 4 мл/мин с линейным градиентом, образованным из MeCN/Н2О/ТФУ (10,0: 79,9:0,1, об./об./об.) и MeCN/Н2О/ТФУ (65,0: 34,9:0,1, об./об./об.) в течение 110 мин. УФ-поглощение проверяли у 214 нм и 280 нм. Хроматограмма ВЭЖХ (проверяли у 280 нм) приводится на фиг. 2. Получали фракции, соответствующие 10 основным пулам, как показано на фиг.2. Объем снижали приблизительно до 25% при помощи вакуумного роторного испарителя и фракции лиофилизовали и испытывали в биоанализе.
Подавляющую аппетит активность обнаружили во фракции G4H9 (пример 6), и пептиды этой фракции анализировали путем анализа аминокислотной последовательности и масс-спектрометрического анализа (пример 4).
Пример 4
Химическое исследование пептидов во фракции G4H9
Анализ аминокислотной последовательности проводили автоматизированным расщеплением по Эдману с использованием газового секвенатора Applied Biosystems Model 477, по существу так, как описано изготовителем. Масс-спектрометрический анализ проводили с использованием системы ЖХ/МС/МС API III (Sciex, Thornhill, Ont., Canada). Тройной квадрупольный инструмент имеет диапазон массы к заряду (m/z) 2400 и снабжен пневматически действующим электрораспылительным (называется также ионораспылительным) интерфейсом (Bruins, A. P. , Covey, T. R., and Henion, J.D.(1987) Anal. Chem. 59, 2642-2646 and Covey T. R. , Bonner, R. F., Shushan, B.I., and Henion, J.D. (1988) Rapid Commun. Mass Spectrom. 2, 249-256). Введение пробы проводили насосом с инфузионным шприцем (Sage Instruments, Cambridge, MA) через капилляр (75 мм внутр. диам.) со скоростью течения жидкости 0,5-1 мл/мин. Шкала m/z инструмента была калибрована монозаряженными ионами аммониевого аддукта поли(пропиленгликолей) (PPGs) при единичном разрешении. Точность измерений масс выше чем 0,02%.
Фракция G4H9
Было обнаружено, что доминирующий пептид в этой фракции имеет следующую аминокислотную последовательность:
HADGSFSDEMNTILDNLATRDFINWLIQTKIТD
Молекулярная масса, определенная масс-спектрометрией, была: 3796.
Этот пептид был идентичен крысиному GLP-2 (1-33). Были также обнаружены небольшие количества следующих двух пептидов:
DFPEEVAIAEELGRRHADGSFSDEMNTILDN LATRDFINWLIQTKITD и
HDEFERHAEGTFTSDVSSYLEGQAAKEFIAWLVКGR
Эти пептиды идентичны с крысиным GLP-2, который удлинен с N-конца спейсерным пептидом 2 и крысиным GLP-1 (1-36 амид) соответственно.
Пример 5
Способ испытания для измерения подавления аппетита у мышей
Мыши были лишены их нормального корма в течение двух дней и получали свободный доступ к 20% раствору глюкозы на третий день голодания. После 2-дневного периода голодания мышей инъецировали внутрибрюшинно 0,5 мл раствора, содержащего испытуемое вещество. Через тридцать минут после инъекции мышей раздельно помещали в одну из восьми испытательных камер на 15 см2 с сетчатой дверцей из нержавеющей стали и стеклянной трубкой для питья, которая выступала в камеру. Трубку для питья соединяли с резервуаром, содержащим 20% раствор глюкозы, и внутренняя часть трубки для питья содержала электрод, дающий возможность обнаруживать контакты при питье с раствором путем измерения течения слабого (незаметного) электрического тока через мышей при помощи электронного аппарата, соединенного с электродом трубки для питья и сетчатой дверцей из нержавеющей стали. Потребление раствора глюкозы измеряли в течение периода 10 минут путем электронной регистрации общего количества контакта с раствором сахарозы во время сеанса испытания. Степень подавления аппетита, вызванного данным испытуемым веществом, определяли статистическим сравнением продолжительности потребления сахарозы контрольными (обработаны носителем) мышами с продолжительностью потребления у мышей, обработанных испытуемым веществом. Степень подавления аппетита у обработанной группы мышей выражали как процент от ответной реакции контрольных групп.
Пример 6
Испытание на подавление аппетита у мышей фракцией, содержащей GLP-2
Мышей испытывали на подавление аппетита (см. пример 5) после обработки испытуемым веществом. Испытуемое вещество состояло из экстрактов опухоли аноректической глюкагономы, полученной в соответствии с примером 3 (фракция G4 гель-фильтрования) или в соответствии с примером 4 (фракция G4H9 ВЭЖХ), растворенных в забуференном фосфатном солевом растворе. Испытуемый раствор, содержащий лиофилизованный материал из фракции G4 гель-фильтрации, соответствующий 3,3 опухолям, подавлял потребление сахарозы на 72%. Из 10 субфракций ВЭЖХ фракции гель-фильтрации G4 (см. пример 4 и фиг.2) только GLP-2 содержащая фракция, G4P9, дала статистически значительное подавление аппетита, подавляя потребление сахарозы на 49%, когда давали лиофилизованный материал, соответствующий 5,3 опухоли.
Пример 7
Испытание на подавление аппетита у крыс синтетическим GLP-2
Мышей испытывали на подавление аппетита, как описано в примере 5, после обработки испытуемым веществом, состоящим из синтетического свиного GLP-2, растворенного в забуференном фосфатом солевом растворе. Свиной GLP-2 имеет следующую аминокислотную последовательность;
HADGSFSDEMNTVLDNLATRDFINWLLHTKIТD.
Внутрибрюшинная инъекция испытуемого раствора, содержащего 50 микрограмм синтетического свиного GLP-2, подавляла потребление сахарозы на 38%.
Пример 8
Способ испытания для измерения подавления аппетита у мышей
Способ такой же, как в примере 5, но вместо 20% сахарозы используют раствор детского молока (Complan®). Испытуемое вещество растворяют в наполнителе, состоящем из забуференного фосфатом солевого раствора с 1% альбумина. Испытуемые соединения, растворенные в наполнителе, инъецировали либо внутривенно (IV) в объеме 100 микролитров, либо интрацеребровентрикулярно (ICV) в объеме 10 микролитров.
Пример 9
Испытание на подавление аппетита у мышей синтетическим GLP-2
Мышей испытывали на подавление аппетита, как описано в примере 8, после обработки испытуемым веществом, состоящим из синтетического GLP-2 человека. GLP-2 человека имеет следующую аминокислотную последовательность:
HADGSFSDEMNTILDNLAARDFINWLIQTKIТD.
IV инъекция испытуемого раствора, содержащего 3 микрограмм синтетического GLP-2 человека, подавляла потребление молока на 24%, тогда как ICV инъекция 3 микрограмм и 10 микрограмм синтетического GLP-2 человека, подавляла потребление молока на 32% и 35% соответственно.
| название | год | авторы | номер документа |
|---|---|---|---|
| СТАБИЛИЗИРОВАННАЯ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ПЕПТИД | 2004 |
|
RU2421236C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОЛОГИЧНОГО БЕЛКА | 1994 |
|
RU2143495C1 |
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКОВ В ТРАНСФОРМИРОВАННЫХ ДРОЖЖЕВЫХ КЛЕТКАХ | 1999 |
|
RU2238323C2 |
| ДИМЕР "КЛЕВЕРНОГО" ПЕПТИДА | 1995 |
|
RU2162857C2 |
| ИЗОЛИРОВАННАЯ МОЛЕКУЛА ДНК, КОДИРУЮЩАЯ РЕЦЕПТОР ГЛЮКАГОНА, ДНК-КОНСТРУКЦИЯ, ЛИНИЯ КЛЕТОК, СПОСОБ ПОЛУЧЕНИЯ РЕЦЕПТОРА ГЛЮКАГОНА, ИЗОЛИРОВАННЫЙ ПЕПТИД РЕЦЕПТОРА ГЛЮКАГОНА, ИЗОЛИРОВАННОЕ АНТИТЕЛО, ЗОНД, СПОСОБ ОБНАРУЖЕНИЯ ПРИСУТСТВИЯ АНТАГОНИСТОВ ГЛЮКАГОНА | 1993 |
|
RU2184779C2 |
| ТОЛЕРОГЕННАЯ ДНК-ВАКЦИНА | 2017 |
|
RU2752608C2 |
| ПОЛИПЕПТИД ФАКТОРА КОАГУЛЯЦИИ VII ЧЕЛОВЕКА, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2002 |
|
RU2325401C2 |
| ВАРИАНТЫ ИЛ-21 | 2006 |
|
RU2412199C2 |
| ПРЕДШЕСТВЕННИК ИНСУЛИНА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 2000 |
|
RU2283846C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ ПОЛИПЕПТИДА В ДРОЖЖАХ | 1995 |
|
RU2167939C2 |
Изобретение относится к медицине. Предложена фармацевтическая композиция и способ, подавляющие аппетит, содержащие подавляющий аппетит пептид формулы (1). Изобретение позволяет более эффективно воздействовать на ингибирование потребления пищи при введении периферически. 4 с. и 13 з.п., 2 ил.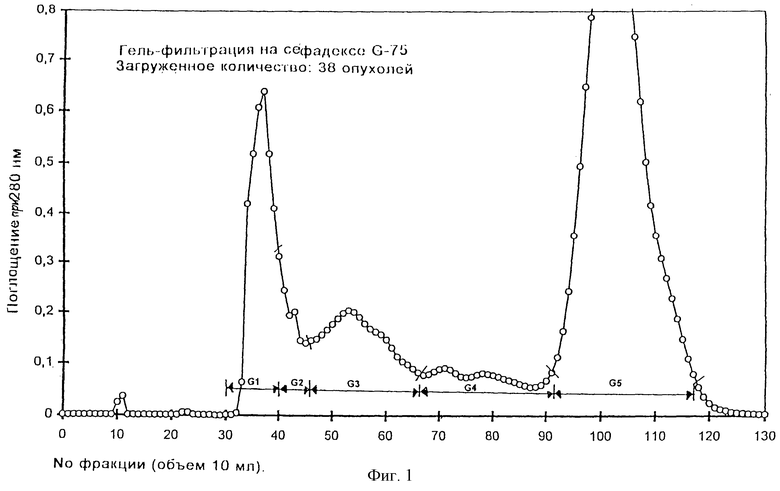
Х1HX2DGSFSDEMNTX3LDX4LAX5X6DFINWLX7X8TKITDX9
где Х1 представляет собой NH2, DFPEEVAIVEELGRR, DFPEEVTIVEELGRR, DFPEEVNIVEELRRR или их фрагмент;
Х2 представляет собой Ala или Gly;
Х3 представляет собой Ile или Val;
Х4 представляет собой Asn, Ser или His;
Х5 представляет собой Ala или Thr;
Х6 представляет собой Arg или Lys;
Х7 представляет собой Ile или Leu;
Х8 представляет собой Gln или His;
Х9 представляет собой ОН, Lys, Arg, Arg-Lys, Lys-Arg, Arg-Arg или Lys-Lys,
вместе с фармацевтически приемлемым наполнителем или разбавителем.
HADGSFSDEMNTILDNLAARDFINWLIQTKITD,
HADGSFSDEMNTILDNLATRDFINWLIQTKITD или
HADGSFSDEMNTVLDNLATRDFINWLLHTKITD.
| TURTON M.D | |||
| et al, Nature, 1996, 379, 69-72 | |||
| Способ лечения алиментарно-конституционального ожирения | 1988 |
|
SU1697823A1 |
| US 4632925 А, 30.12.1986. | |||
