Изобретение касается димеров "клеверных" пептидов, способов получения димеров "клеверных" пептидов, фармацевтических составов, включающих димеры "клеверных пептидов", и их применения для лечения желудочно-кишечных расстройств.
"Клеверные" пептиды образуют семейство пептидов, обнаруживаемых в основном в желудочно-кишечном тракте. "Клеверные" пептиды млекопитающих содержат один или более характерных доменов в форме клеверного листа [1], каждый из которых собран из аминокислотной последовательности, состоящей из 38 или 39 аминокислотных остатков, в которой 6 остатков полуцистина соединены в сочетании 1-5, 2-4 и 3-6, образуя таким образом характерную структуру в виде клеверного листа.
Известные к настоящему времени "клеверные" пептиды млекопитающих содержат или один, или два домена в форме клеверного листа (см. обзоры [3-5]), в то время как для лягушки Xenopus laevis описаны пептиды и белки, содержащие один [6], два [7], четыре [8] или шесть [9] доменов в форме клеверного листа. "Клеверными" пептидами млекопитающих, содержащими один домен, являются связанный с раком молочной железы pS2-пептид, известный к настоящему времени для человека [10, 11] и мыши [12], и кишечный "клеверный" фактор (ITF) человека [13, 14] и крысы [15, 16]. Для человека [17], свиньи [18] и мыши [17] описан спазмолитический полипептид (SP), содержащий два домена в форме клеверного листа. У людей все три "клеверных" пептида - hpS2, hITF и hSP - экспрессируются в норме в желудочно-кишечном тракте: hSP и hpS2 - в эпителиальном слое слизистой оболочки желудка [17, 19], a hITF - в эпителиальном слое слизистой оболочки тонкой и толстой кишки [13].
Физиологическая роль "клеверных" пептидов не очень хорошо выяснена. Сообщалось об усилении экспрессии "клеверных" пептидов в желудочно-кишечном тракте для ряда состояний, связанных с повреждением слизистой оболочки, таких как воспаление кишечника [20-22], язва желудка и двенадцатиперстной кишки [20, 23, 24]. На основании этих данных была предложена репаративная функция "клеверных" пептидов (например, в [22]). Недавно авторами работ [25-27] было получено доказательство ускорения восстановления поврежденной эпителиальной слизистой оболочки в результате действия "клеверных" пептидов. Механизм, с помощью которого "клеверные" пептиды способствуют выполнению их репаративной функции, может заключаться в образовании сшивок между муциновыми гликопротеинами с формированием вязкоупругого гелевого слоя, устойчивого к действию ферментов пищеварения [3, 28].
Клонирование однодоменного "клеверного" фактора кишечника крысы и человека и его применение для лечения повреждений желудочно-кишечного тракта описаны [29].
Обнаружена возможность получения димеров "клеверных" факторов, имеющих только один домен в форме клеверного листа и обладающих интересными фармакологическими свойствами.
В соответствии с этим настоящее изобретение касается "клеверного" пептида, содержащего единственный домен в форме клеверного листа и существующего в виде димера.
Как отмечено выше, предполагают, что "клеверные" пептиды способствуют заживлению пептических язв и других повреждений, относящихся к слизистой оболочке, благодаря стабилизации слизистого слоя кишечного тракта. Механизм подобной стабилизации в настоящее время не известен. Тем не менее рентгено-структурный анализ (ср. [28]) панкреатического спазмолитического полипептида (PSP) свиньи, образующего два домена в форме клеверного листа, показал, что большинство имеющихся в наличии аминокислотных остатков принимают участие в формировании щели шириной 8-10 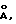 обнаруживаемой в каждом таком домене. Предварительные эксперименты показали, что эта щель способна вмещать часть олигосахаридной цепи, например углевода, связанного с муциновым гликопротеином. Если это так, то PSP с двумя подобными щелями могут сшивать молекулы муцинов друг с другом, помогая им образовывать защитный гель, покрывающий эпителий слизистой оболочки. Сейчас неизвестно, образуют ли in vivo димеры для осуществления схожей функции однодоменные "клеверные" пептиды (такие, как IFT и pS2) или у них другой механизм действия. Тем не менее в настоящее время считается, что димеры таких "клеверных" пептидов действительно могут сшивать молекулы муцинов, являясь поэтому активной формой этих пептидов.
обнаруживаемой в каждом таком домене. Предварительные эксперименты показали, что эта щель способна вмещать часть олигосахаридной цепи, например углевода, связанного с муциновым гликопротеином. Если это так, то PSP с двумя подобными щелями могут сшивать молекулы муцинов друг с другом, помогая им образовывать защитный гель, покрывающий эпителий слизистой оболочки. Сейчас неизвестно, образуют ли in vivo димеры для осуществления схожей функции однодоменные "клеверные" пептиды (такие, как IFT и pS2) или у них другой механизм действия. Тем не менее в настоящее время считается, что димеры таких "клеверных" пептидов действительно могут сшивать молекулы муцинов, являясь поэтому активной формой этих пептидов.
Другой аспект настоящего изобретения касается способа получения димера "клеверного" пептида, содержащего единичный домен в форме клеверного листа, способ, включающий культивирование соответствующих клеток-хозяев, трансформированных ДНК с последовательностью, кодирующей "клеверный" пептид, содержащий один домен в форме клеверного листа, в условиях, позволяющих наработку пептида, и выделение из культуры образующегося димера "клеверного" пептида.
Следующий аспект настоящего изобретения касается фармацевтических составов, включающих димер "клеверного" пептида, содержащий единственный домен в форме клеверного листа, совместно с фармацевтически приемлемым разбавителем или носителем.
Еще один аспект настоящего изобретения касается применения димера "клеверного" пептида, содержащего один домен в форме клеверного листа, в качестве лекарства и использования димера "клеверного" пептида, содержащего один домен в форме клеверного листа, для получения лекарства с целью профилактики и лечения желудочно-кишечных расстройств.
Димером "клеверного" фактора может быть, в частности, димер кишечного "клеверного" фактора (ITF) или пептида, связанного с раком молочной железы (pS2).
В частности, "клеверным" фактором является человеческий ITF, последовательность аминокислот мономера которого выглядит следующим образом:
ZEYVGLSANQCAVPAKDRVDCGYPHVTPKECNNRGCCFDSRIPGVPWCFKPLQEAECTF, где Z представляет собой остаток Glu, Gln или pyrGlu, или его гомолог, способный димеризоваться и проявляющий схожую активность;
или человеческий pS2, последовательность аминокислот мономера которого выглядит следующим образом:
ZAQTETCTVAPRERQNCGFPGVTPSQCANKGCCFDDTVRGVPWCFYPNTIDVPPEEECEF,
где Z представляет собой остаток Glu, Gln или pyrGlu, или его гомолог, способный димеризоваться и проявляющий схожую активность.
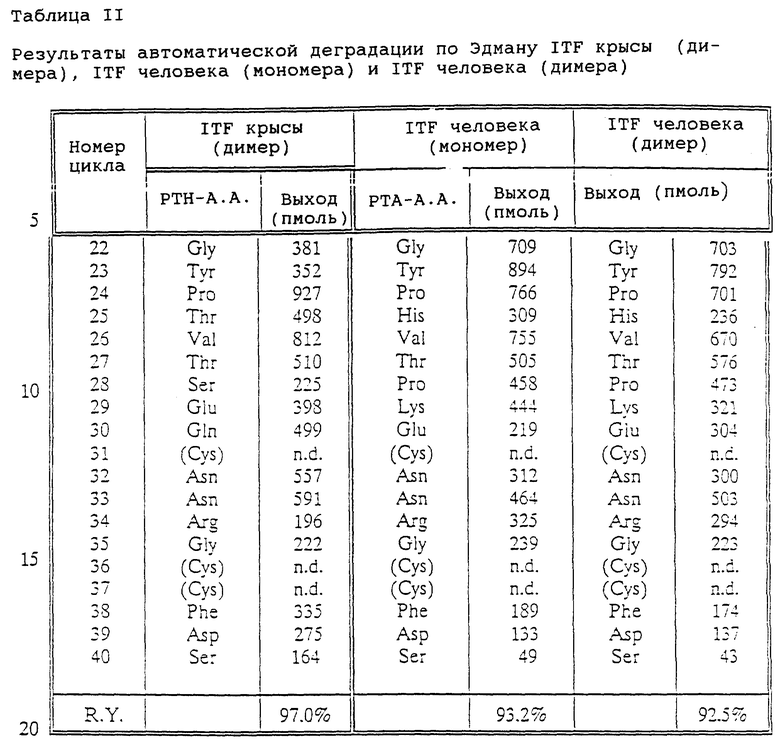
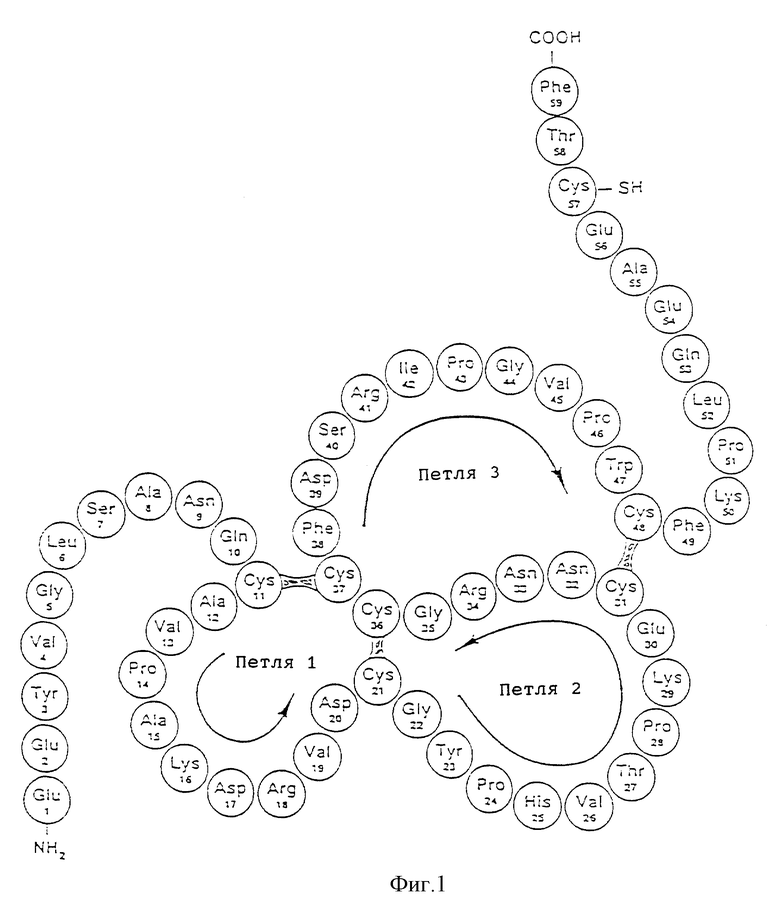
Гомологи ITF и pS2 охватывают пептиды с тем же самым характером расположения остатков цистеина и дисульфидных связей (фиг. 1) и определенной степенью гомологии (в смысле либо идентичности аминокислотных остатков в соответствующих положениях, либо наличия консервативных замен) в петлях 1, 2 и 3. Гомология аминокислотной последовательности в области петель может варьировать в интервале от 1 до 10 аминокислотных остатков, а число аминокислотных остатков в каждой петле (кроме остатков цистеина) - от 7 до 12, предпочтительно - от 9 до 10.
Гомологи ITF и pS2 могут содержать одну или более аминокислотных замен, делеций или вставок. Преимущественная природа этих изменений такова, что они не влияют в значительной мере на сворачиваемость и активность белка. Размер небольших делеций обычно составляет от 1 до 3 аминокислотных остатков в области петель и от 1 до 10 аминокислотных остатков в N- и C-концевых областях; возможны одиночные амино- и карбоксиконцевые добавки, такие как аминоконцевой остаток метионина, небольшой линкерный пептид до 10 аминокислотных остатков или небольшая добавка, облегчающая очистку, такая как полигистидиновый тракт, антигенный эпитоп или связывающий домен (общие принципы см. в [30] ). Примерами консервативных замен могут быть замены в рамках группы основных аминокислот (например, аргинина, лизина, гистидина), кислых аминокислот (например, глутаминовой и аспарагиновой аминокислот), полярных аминокислот (например, глутамина и аспарагина), гидрофобных аминокислот (например, лейцина, изолейцина, валина), ароматических аминокислот (например, фенилаланина, триптофана, тирозина) и аминокислот с небольшим размером бокового радикала (например, глицина, аланина, серина, треонина, метионина).
Для специалистов в данной области должно быть очевидным, что подобные замены могут быть проведены вне областей, отвечающих за функцию молекулы, в результате чего получается по-прежнему активный полипептид. Существенные для активности настоящих "клеверных" пептидов аминокислоты могут быть идентифицированы в соответствии с известными специалистам методиками, такими как сайт-специфический и аланин-сканирующий мутагенез [31]. В последнем случае мутации вводят в каждый из аминокислотных остатков молекулы и получаемые мутантные молекулы проверяют на биологическую активность (например, на заживление слизистой оболочки, защиту слизистой оболочки, заживление язвы желудка) с целью идентификации тех аминокислотных остатков, которые отвечают за активность молекулы.
Возможен аллельный вариант гомолога, т.е. альтернативная форма гена, которая получается в результате мутации, или измененный пептид, кодируемый мутантным геном, но обладающий по существу такой же активностью как настоящий пептид. Следовательно, мутации могут быть "молчащие" (никаких изменений в кодируемом пептиде) или могут кодировать пептид с измененной аминокислотной последовательностью.
Гомологом настоящего "клеверного" пептида кроме этого может быть видовой гомолог, т.е. полипептид со схожей активностью, происходящий из другого вида, например из мыши, крысы, кролика, коровы, свиньи или лягушки.
В предпочтительном воплощении димер "клеверного" пептида по изобретению имеет значение молекулярного веса приблизительно 13000. Димер состоит из двух мономерных "клеверных" пептидов, связанных дисульфидной связью между двумя остатками цистеина в положении 57 для ITF-подобных мономеров или в положении 58 для рS2-подобных мономеров.
Предпочтительным является получение димера "клеверного" пептида с помощью технологии рекомбинантной ДНК. С этой целью последовательность ДНК, кодирующая "клеверный" пептид, может быть выделена посредством создания библиотеки геномной или кДНК и поиска в соответствии со стандартными методиками с помощью гибридизации с синтетическими олигонуклеотидными зондами [32] последовательности ДНК, кодирующей пептид целиком или частично. Предпочтительной в рамках настоящего изобретения является кодирующая пептид последовательность ДНК человеческого происхождения, т.е. из библиотеки геномной ДНК или кДНК человека.
Последовательность ДНК, кодирующую "клеверный" пептид, также можно получить путем химического синтеза с помощью традиционных стандартных методов, например фосфоамида(и)тного метода, описанного в [33], или метода, изложенного в [34]. В соответствии с фосфоамида(и)тным методом олигонуклеотиды синтезируются, например, в автоматическом ДНК-синтезаторе, затем очищаются, отжигаются, лигируются и клонируются в подходящих векторах.
Последовательность ДНК также может быть получена с помощью полимеразной цепной реакции с применением специфических праймеров, как, например, описано в [35, 36] или упомянутой выше работе [32].
Последовательность ДНК, кодирующую "клеверный" пептид, обычно вводят в рекомбинантный вектор, представляющий собой любой из векторов, пригодных для проведения процедур с рекомбинантной ДНК, и выбор вектора часто зависит от природы клетки-хозяина, в которую он должен быть введен. Так, данный вектор может быть автономно реплицирующимся вектором, например существуя вне хромосомы и реплицируясь независимо от хромосомальной репликации, т.е. быть плазмидой. С другой стороны, вектором может быть один из тех векторов, которые, попадая в клетку-хозяина, интегрируются в геном клетки-хозяина и реплицируются совместно с хромосомой(ами), в которую они интегрированы.
Предпочтительно, чтобы вектор являлся экспрессируемым вектором, в котором последовательность ДНК, кодирующая "клеверный" пептид, оперативно соединена с дополнительными участками, необходимыми для транскрипции ДНК. В общем случае экспрессируемый вектор происходит из плазмидной или вирусной ДНК или содержит элементы обоих. Термин "операбельно соединена" означает, что участки организованы таким образом, что функционируют во взаимодействии друг с другом для выполнения предназначенных целей, т.е. транскрипция начинается на промоторе и проходит на всем участке ДНК, кодирующем полипептид.
Промотором может быть любая последовательность ДНК, демонстрирующая транскрипционную активность в выбранной клетке-хозяине, происходящая из генов, кодирующих белки как гомологичные, так и гетерологичные клетке-хозяину.
Примерами подходящих для проведения транскрипции ДНК, кодирующей "клеверные" пептиды, в клетках млекопитающих промоторов являются промотор SV40 [37] , промотор МТ-1 (промотор гена металлотионеина) [38] или главный поздний промотор аденовируса 2.
Примером соответствующего промотора для использования в клетках насекомых является полиэдриновый промотор [39, 40], P10-промотор [41], промотор основного белка вируса полиэдроза Autoqrapha californica [42] , промотор "быстрого" раннего гена 1 бакуловируса [43, 44] или промотор замедленного раннего гена бакуловируса 39К [43, 44].
Соответствующими промоторами для использования в дрожжевых клетках-хозяевах являются промоторы дрожжевых гликолитических генов [45, 46], или генов алкогольдегидрогеназы [47], или TRI1- [48], или ADH2-4с-промоторы [49] .
Примерами подходящих для использования в нитчатых грибах как клетках-хозяевах промоторов являются, например, ADH3-промотор [50] или tpiA-промотор. В качестве примеров других полезных промоторов можно привести промоторы генов ТАКА-амилазы из A. oryzae, аспарагиновой протеиназы из Rhizomucor miehei, нейтральной α-амилазы из A. niger, кислотостабильной α-амилазы из A. niger, глюкоамилазы (gluA) из A. niger или A. awamori, липазы из Rhizomucor miehei, щелочной протеазы из A. oryzae, триозофосфатизомеразы из A. oryzae или ацетамидазы из A. nidulans. Предпочтительными являются промоторы генов ТАКА-амилазы и gluA. Соответствующие промоторы указаны, например, в [51] и [52].
Последовательность ДНК, кодирующая "клеверный" пептид, кроме того, при необходимости может быть оперативно связана с подходящим терминатором, таким как терминатор гена гормона роста человека [53], или, когда в качестве клеток-хозяев используются грибы, терминаторы TPI1 [46] или ADH3 [50]. Вектор также может включать такие элементы, как полиаденилатные сигналы (например, из Elb-области SV40 или аденовируса 5), последовательности, усиливающие транскрипцию (например, энхансер SV40), и последовательности, усиливающие трансляцию (такие, например, как кодирующие РНКы аденовируса VA).
Рекомбинантный вектор также может включать последовательность ДНК, дающую возможность вектору реплицироваться в рассматриваемой клетке-хозяине. Примером подобной последовательности (когда клеткой-хозяином является клетка млекопитающего) является ориджин репликации SV40.
Когда клеткой-хозяином является дрожжевая клетка, соответствующими последовательностями, дающими возможность вектору реплицироваться, являются гены репликации REP 1-3 и ориджин репликации дрожжевой плазмиды 2μ.
Вектор также может содержать селективный маркер, например ген, продукт которого дополняет дефект клетки-хозяина, такой как ген дегидрофолатредуктазы (DHFR) или TPI-ген Schizosaccharomyces pombe (описанный в [54]), либо ген, придающий устойчивость к лекарствам, например ампициллину, канамицину, тетрациклину, хлорамфениколу, неомицину, гигромицину или метотрексату. Для нитчатых грибов селективные маркеры включают amdS, pyrG, argB, niaD или sC.
Для того чтобы направить "клеверный" пептид, описанный в настоящем изобретении, по секреторному пути клетки-хозяина, в рекомбинантный вектор может быть введена секреторная сигнальная последовательность (известная также как лидерная последовательность, предпропоследовательность или предпоследовательность). Секреторная сигнальная последовательность присоединяется к последовательности ДНК, кодирующей "клеверный" пептид, в корректной рамке считывания. Обычно сигнальная последовательность располагается с 5'-конца последовательности ДНК, кодирующей пептид. В качестве секреторной сигнальной последовательности может применяться сигнальная последовательность, в норме ассоциированная с данным пептидом, либо последовательность гена, кодирующего другой секреторный белок.
Для секреции из дрожжевых клеток секреторная сигнальная последовательность может кодировать любой сигнальный пептид, обеспечивающий эффективное направление экспрессированного "клеверного" пептида в секреторный путь клетки. Сигнальным пептидом может быть от природы имеющийся сигнальный пептид (либо его функциональная часть) или синтетический пептид. Было установлено, что соответствующими сигнальными пептидами являются сигнальный пептид α-фактора [54], сигнальный пептид амилазы слюны мыши [55], модифицированный сигнальный пептид карбоксипептидазы [56] , сигнальный пептид BAR1 дрожжей [57] или сигнальный пептид дрожжевой аспарагиновой протеазы 3 (YAP3) [57].
Кроме этого, для эффективной секреции в дрожжах между сигнальной последовательностью и последовательностью ДНК, кодирующей "клеверный" пептид, может быть введена последовательность, кодирующая лидерный пептид. Функция лидерного пептида заключается в том, что он позволяет экспрессированному пептиду направляться из эндоплазматического ретикулума в комплекс Гольджи и далее в секреторные визикулы для секреции в культуральную среду (т.е. направляет экспорт "клеверного" пептида через клеточную стенку или по меньшей мере через клеточную мембрану в периплазматическое пространство дрожжевой клетки). В качестве лидерного пептида может выступать лидерный пептид дрожжевого α-фактора (его использование описано, например, в [54, 59-63]). С другой стороны, лидерный пептид может быть искусственным, иными словами не природным. Синтетический лидерный пептид может, например, быть сконструирован как описано в [64] или [65].
С целью использования в нитчатых грибах сигнальный пептид можно удобно получать из гена, кодирующего амилазу или глюкоамилазу из Aspergillus sp., гена, кодирующего липазу или протеазу из Rhizomucor miehei или липазу из Humicola lanuginosa. Предпочтительным является сигнальный пептид гена, кодирующего ТАКА-амилазу из A. oryzae, нейтральную α-амилазу из A. niger, кислотостабильную α-амилазу из A. niger или глюкоамилазу из А. niger. Соответствующие сигнальные пептиды описаны в [51] и [66].
Для использования в клетках насекомых такой сигнальный пептид может быть получен на основе гена насекомого (ср. [67]), как, например, сигнальный пептид предшественника адипокинетического гормона чешуекрылого Manduca sexta (ср. [68]).
Методики, применяемые для лигирования последовательностей ДНК, кодирующих "клеверный" пептид, промотор и, возможно, терминатор и/или секреторную сигнальную последовательность, соответственно, и введения их в подходящие векторы, содержащие информацию, необходимую для репликации, хорошо известны специалистам в данной области (ср., например, [32]).
Клетка-хозяин, в которую вводят последовательность ДНК, кодирующую "клеверный" пептид, может представлять собой любую клетку, способную продуцировать данный пептид в димерной форме, включая клетки дрожжей, грибов и высших эукариотов.
Примерами соответствующих клеточных линий млекопитающих являются клеточные линии COS (АТСС CRL 1650), ВНК (АТСС CRL 1632, АТСС CCL 10), CHL (АТСС CCL 39) или СНО (АТСС CCL 61). Способы трансфекции клеток млекопитающих и экспрессии введенных в них последовательностей ДНК описаны в работах [69-75] .
Примеры соответствующих дрожжевых клеток включают клетки Saccharomyces spp. или Schizosaccharomyces spp., в особенности штаммы Saccharomyces cerevisiae или Saccharomyces kluyveri. Способы трансформации дрожжевых клеток гетерологичной ДНК и продуцирования в них гетерологичных полипептидов описаны, например, в [38, 54, 76-78], каждый из которых, таким образом, приведен в ссылках. Трансформированные клетки отбирают по фенотипу, определяемому по селективному маркеру, обычно по устойчивости к лекарственному препарату или способности расти в отсутствии определенного компонента питательной среды, например лейцина. Предпочтительным для использования в дрожжах является вектор РОТ1, описанный в [76]. Последовательности ДНК, кодирующей "клеверный" пептид, может предшествовать сигнальная последовательность, а возможно и лидерная последовательность, как, например, описано выше. Другими примерами соответствующих дрожжевых клеток являются штаммы Kluyveromyces, такие как К. lactis, Hansenula, например H. polymorpha, или Pichiа, например P.pastoris (cp. [79, 80]).
Примерами других грибных клеток являются клетки нитчатых грибов, например Aspergillus spp., Neurospora spp., Fusarium spp. или Trichoderma spp., в особенности A. oryzae, A. nidulans или A. niger. Применение Aspergillus spp. для экспрессии белков описано, например, в [51, 81, 82]. Трансформация F. oxysporum может, например, быть проведена, как описано в [83], a Trichoderma spp. - как в [84].
Когда в качестве клетки-хозяина используется клетка нитчатого гриба, она может быть трансформирована сконструированной по настоящему изобретению ДНК посредством интеграции последней в хромосому клетки-хозяина для получения рекомбинантной клетки-хозяина. Как правило, считается, что подобная интеграция имеет то преимущество, что введенная последовательность ДНК будет более стабильно поддерживаться в клетке. Интеграция сконструированных последовательностей ДНК в хромосому клетки-хозяина может быть осуществлена в соответствии с общепринятыми способами, например посредством гомологичной или гетерологичной рекомбинации.
Трансформация клеток насекомых и продукция в них гетерологичных полипептидов может быть осуществлена, как описано в патентах [39, 42-44, 85], включенных здесь в список литературы. В качестве клеточных линий насекомых, используемых как клетки-хозяева, подходит клеточная линия Lepidoptera, например клетки Spodoptera frugiperda или клетки Trichoplusia ni (ср. [86]). Для культивирования подходят условия, описанные, например, в [87] или [88] либо в любой из вышеупомянутых ссылок.
Подвергнутые трансформации или трансфекции описанные выше клетки-хозяева далее культивируют в соответствующей питательной среде в условиях, допускающих экспрессию "клеверного" пептида, после чего образовавшийся пептид полностью или частично может быть выделен из культуры в димерной форме. В качестве среды культивирования можно использовать любую общепринятую среду, пригодную для роста клеток-хозяев, например минимальную или комплексную среду, содержащую соответствующие добавки. Подходящие среды являются коммерчески доступными или их можно приготовить в соответствии с опубликованными рецептами (например, в соответствии с каталогами American Type Culture Collection). Продуцируемый клетками "клеверный" пептид далее может быть выделен из культуральной среды с помощью общепринятых методик, включающих отделение клеток-хозяев от среды посредством центрифугирования или фильтрования, осаждение белковых компонентов супернатанта или фильтрата с помощью соли, например сульфата аммония, очистку с использованием разнообразных хроматографических методик, например ионообменной хроматографии, гель-фильтрации, аффинной хроматографии или им подобных в зависимости от типа рассматриваемого полипептида.
Димер "клеверного" пептида можно вводить в фармацевтический состав по изобретению согласно любому из устоявшихся методов приготовления фармацевтических составов, например, как описано в [89]. Такой состав может быть изготовлен в пригодной для систематических инъекций или инфузий форме и таким образом может включать стерильную воду или изотонический раствор соли либо глюкозы. Составы можно стерилизовать с помощью стандартных методик стерилизации, хорошо известных специалистам в данной области. Полученные водные растворы можно сразу упаковать для использования или профильтровать в асептических условиях и высушить лиофильно; лиофилизованный препарат перед введением смешивают со стерильным водным раствором. Фармацевтический состав может содержать фармацевтически приемлемые добавочные вещества, необходимые для приближения к физиологическим условиям, такие как забуферивающие агенты, регулирующие тонус вещества и им подобные, например ацетат натрия, лактат натрия, хлорид натрия, хлорид калия, хлорид кальция и т.д.
Фармацевтический состав по настоящему изобретению также может быть адаптирован для назального, трансдермального и ректального введения. Фармацевтически приемлемый наполнитель или разбавитель, используемый в составе, может представлять собой любой традиционный твердый наполнитель. Примерами твердых наполнителей являются лактоза, терра альба, сахароза, тальк, желатин, агар, пектин, аравийская камедь, стеарат магния и стеариновая кислота. Подобным же образом наполнитель или разбавитель может включать любой известный специалистам способствующий выведению материал, такой как глицерилмоностеарат или глицерилдистеарат, один либо в смеси с парафином. Содержание твердого наполнителя будет широко варьировать, однако обычно будет находиться в интервале от 25 мг до 1 г.
Концентрация "клеверного" пептида в таком составе может изменяться в широких пределах, например от приблизительно 5% до приблизительно 100% по весу. Предпочтительная концентрация лежит в области 50-100% по весу. Стандартная доза для такого состава обычно может содержать от приблизительно 1 мг до приблизительно 200 мг, предпочтительно от приблизительно 25 мг до приблизительно 75 мг, в частности приблизительно 50 мг пептида.
Согласно указанному выше считается, что представляемый в данном изобретении димер "клеверного" пептида является активной формой указанного пептида. В связи с этим предполагается, что он будет полезным в случае применения его с целью предупреждения или лечения заболеваний желудочно-кишечного тракта. Более конкретно предполагается использование его для лечения желудочных или пептических язв, воспалительных заболеваний кишечника, болезни Крона или повреждений кишечного тракта, вызванных радиационной терапией, бактериальными либо другими инфекциями и т.д. Доза полипептида, вводимого пациенту, будет изменяться в зависимости от типа и тяжести состояния лечащегося, но обычно лежит в интервале 0,1-1,0 мг/кг веса тела.
Дальнейшие детали настоящего изобретения описываются в примере со ссылками на прилагаемые чертежи, где
Фиг. 1 - предполагаемая структура кишечного "клеверного" фактора (ITF) человека. Первичная аминокислотная последовательность взята из работы [14], а дисульфидные связи расставлены исходя из гомологии с PSP и pS2 [2].
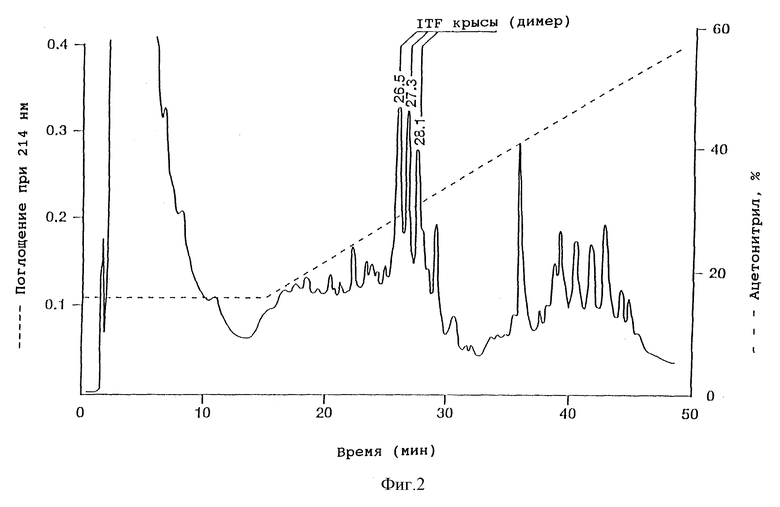
Фиг. 2 - обращенно-фазная ВЭЖХ супернатанта из дрожжевого штамма HW756, экспрессирующего ITF крысы, на колонке Vydac 214ТР54.
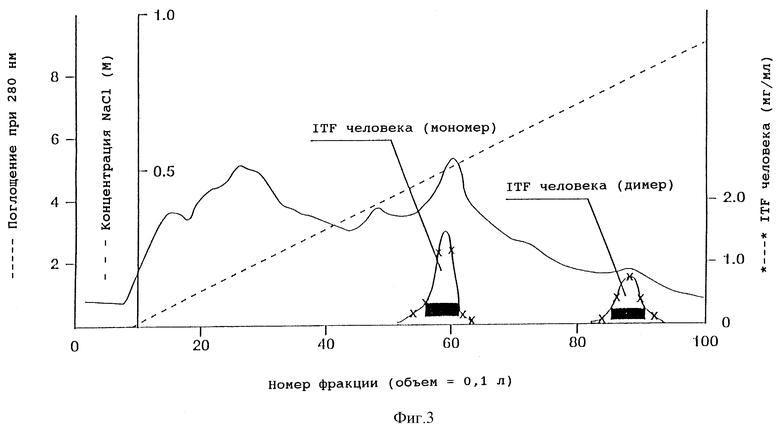
Фиг. 3 - ионообменная хроматография частично очищенного ITF человека на колонке с Fast Flow SP-Сефарозой. Количество мономера и димера ITF человека определяли с помощью аналитической ВЭЖХ. Окрашенные прямоугольники указывают фракции, собранные для дальнейшей очистки мономерной и димерной форм. Пунктирная линия соответствует концентрации NaCl в элюирующем растворителе.
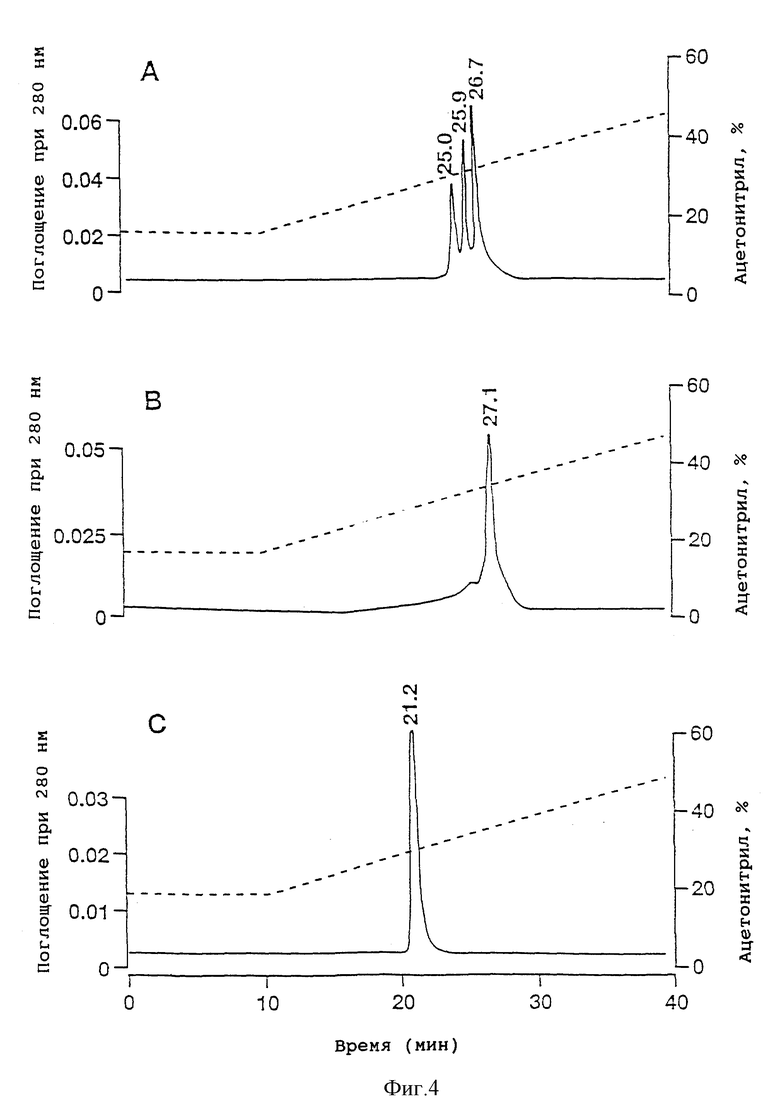
Фиг. 4 - обращенно-фазная ВЭЖХ очищенных димера ITF крысы (А), мономера ITF человека (В) и димера ITF человека (С) на колонке Vydac 214ТР54 C4. Пунктирная линия показывает концентрацию ацетонитрила в элюирующем растворителе.
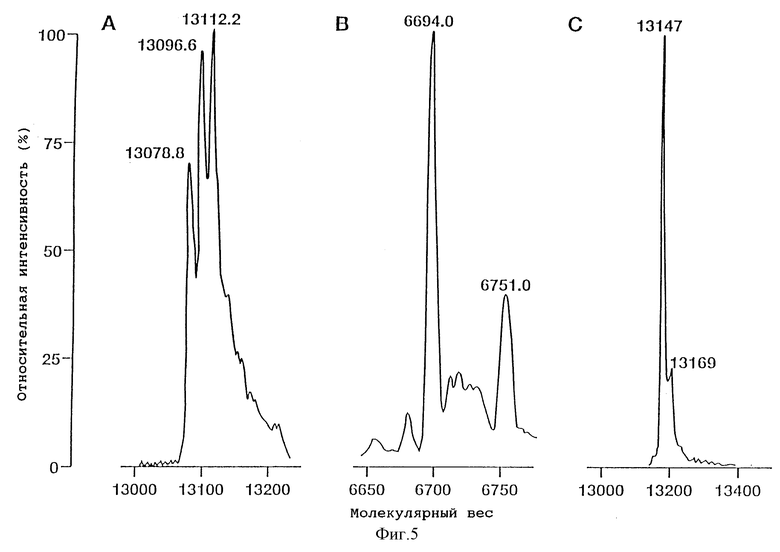
Фиг. 5 - масс-спектры очищенных ITF крысы (димера) (А), ITF человека (мономера) (В) и ITF человека (димера) (С).
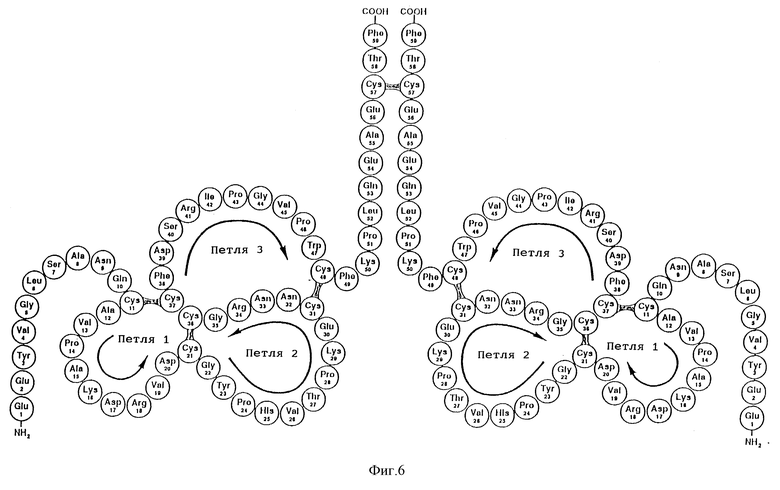
Фиг. 6 - структура димерной формы ITF человека.
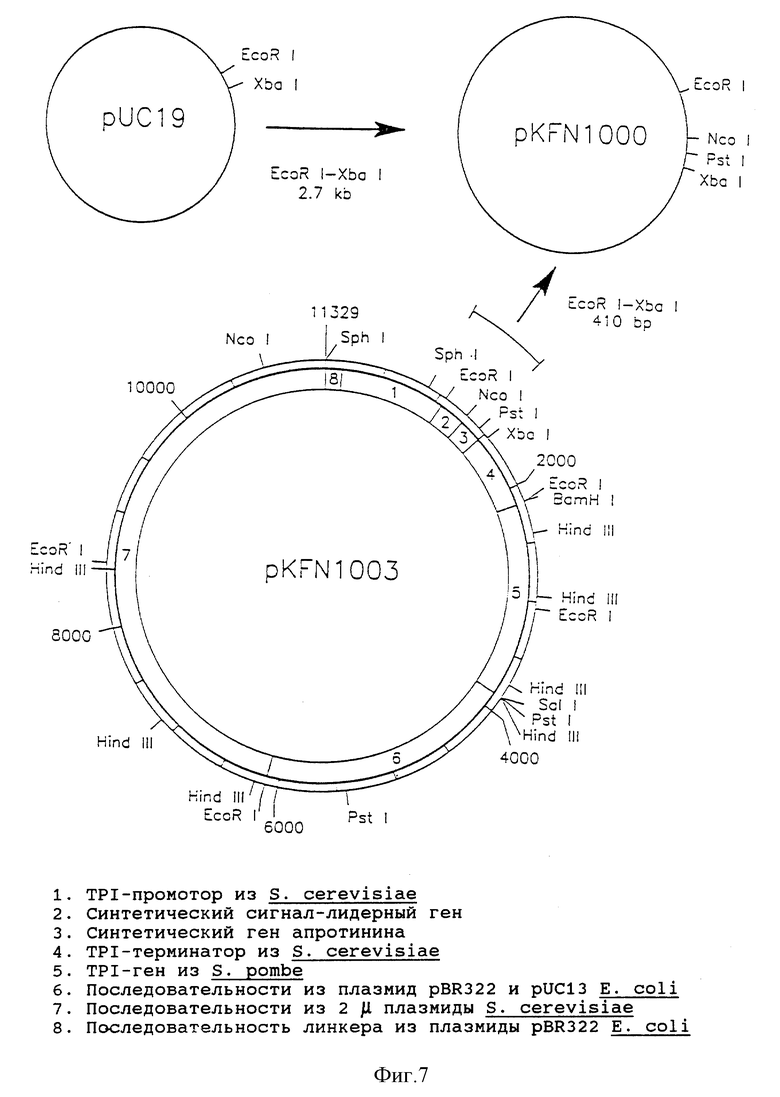
Фиг. 7 - рестрикционная карта плазмиды KFN 1003.
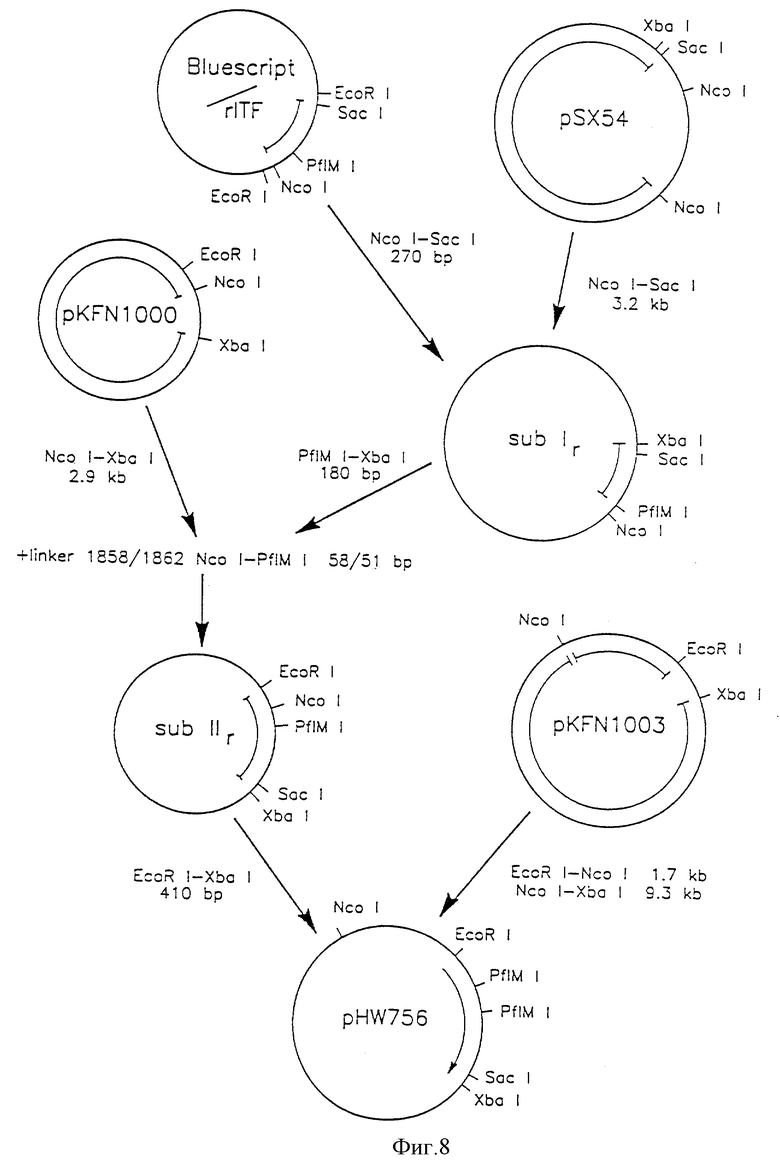
Фиг. 8 - конструкция плазмиды pHW756.
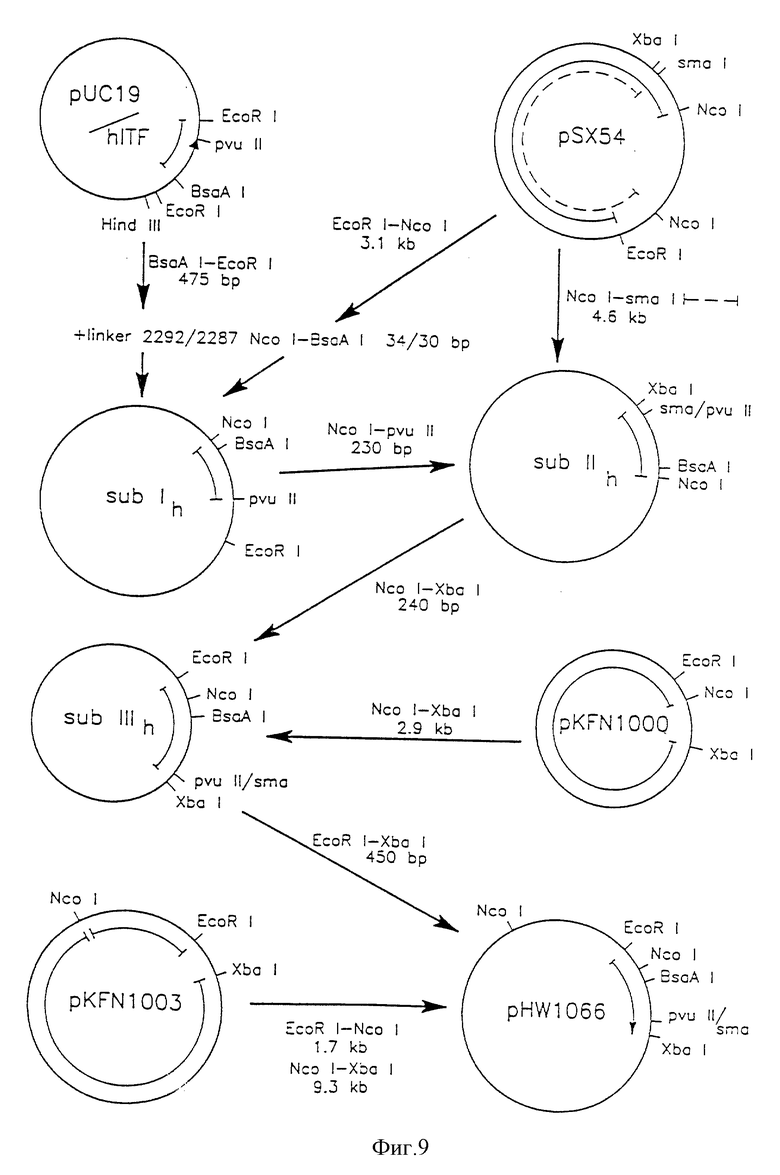
Фиг. 9 - конструкция плазмиды pHW1066.
Пример 1
МАТЕРИАЛЫ И МЕТОДЫ
Клонирование ITF крысы (rITF) и ITF человека (hITF)
Клонирование ITF крысы и человека выполняли, как описано в [15, 16, 29] (rITF) и [13, 14] (hITF).
Конструирование плазмид, экспрессирующих rITF и hITF
Согласно схемам, приведенным на фиг.7 - 9, были сконструированы экспрессирующие плазмиды pHW756 для секреции ITF крысы и pHW1066 для секреции ITF человека. Экспрессирующий дрожжевой вектор рКFN1003 (описан в [90]) представляет собой производную плазмиды СРОТ [91]. Он содержит в качестве селективного маркера TPI-ген Schizosaccharomyces pombe (РОТ) [53], промотор три-озофосфатизомеразы S. cerevisiae (ТРI) и терминатор, контролирующий экспрессию [46].
Ген ITF крысы первоначально клонировали в Bluescript II KS(-) (Stratagene), из которого его размножали в соответствии с фиг. 8. Вспомогательный вектор pSX54, обеспечивающий полезные для клонирования сайты, составлен из pUC18 и pDN1050 [92]. Синтетический ДНК-линкер Ncol-PflM1 имеет следующую последовательность нуклеотидов:
1858: 5'-CATGGCTGAAAGATTGGAAAAGAGACAAGAGTTCGTTGGTTTGTCTCCATCCC AATGT-3' 58 н.п.
1862: 5'-TTGGGATGGAGACAAACCAACGAACTCTTGTCTCTTTTCCAATCTTTCAGC - 3' 51 н. п.
Линкер кодирует 8 C-концевых аминокислот лидерной последовательности, как описано в [93], с несколькими изменениями в выборе кодонов, а N-концевая часть гена ITF крысы имеет следующую последовательность: QEFVGLSPSQC. Сигнальная и лидерная аминокислотные последовательности описаны там же:
MKAVFLVLSLIGFCWAQPVTGDESSVEIPEESLIIAENTTLANVAMAERLEKR.
Ген человеческого ITF клонировали в PUC19 и размножали в соответствии с фиг. 9. Синтетический ДНК-линкер Ncol-BsaAl имеет следующую последовательность:
2292: 5'-CATGGCTGAAAGATTGGAAAAGAGAGAAGAATAC-3' 34 н.п.
2287: 5'-GTATTCTTCTCTCTTTTCCAATCTTTCAGC-3' 30 н.п.
Линкер кодирует 8 C-концевых аминокислот лидерной последовательности, как описано для конструкции rITF, а тремя N-концевыми аминокислотами гена hITF являются EEY. Сигнальная и лидерная последовательности такие же, как и ранее.
Экспрессирующими плазмидами трансформировали штамм S.cerevisiae MT 663 (E2-7B X E11-36 a/α , δtpi/δtpi, pep 4-3/pep 4-3) с селекцией по росту на глюкозе в качестве единственного источника углерода.
Дрожжевые трансформанты, экспрессирующие ITF крысы и человека, были названы HW756 и HW1066 соответственно.
Ферментации
Описанные выше трансформанты культивировали в течение 72 ч при 30oC в дрожжевой пептон-декстрозной (YPD) среде [94], дополненной дрожжевым экстрактом (60 г/л). По окончании ферментаций были достигнуты значения оптических плотностей при 660 нм, равные 153 и 232 для HW756 (rITF) и HW1066 (hITF) соответственно. По окончании ферментации pH подводили до 2,5 с помощью 1 M фосфорной кислоты и дрожжевые клетки удаляли центрифугированием при 3000 об/мин в течение 15 мин.
Очистка рекомбинантного rITF
Концентрацию rITF в дрожжевой ферментационной питательной среде и фракциях, полученных во время очистки, измеряли с помощью аналитической ВЭЖХ. Аликвоты (обычно 50-200 мкл) наносили на C4 ВЭЖХ-колонку (0,46 x 25 см) Vydac 214TP54 для обращенно-фазной хроматографии, уравновешенную 0,1% (v/v) TFA в 15% (v/v) ацетонитриле, при 30oC и скорости потока 1,5 мл/мин. После 10 мин изократического элюирования концентрацию ацетонитрила в элюирующем растворителе увеличивали в течение 40 мин до 55%. Измеряли поглощение при 214 нм. Были обнаружены три пика, элюирующиеся с временами удерживания 26,5 мин, 27,3 мин и 28,2 мин (фиг. 2), представляющие собой димерные формы rITF. Количество пептидов определяли, используя калибровочный hSP-стандарт [93].
Уровень экспрессии рекомбинантного ITF крысы в данной дрожжевой системе составлял 113 мг/л.
С помощью центрифугирования из ферментера вместимостью 10 л было выделено 8,7 л ферментационного бульона. Для понижения проводимости супернатант был разбавлен 14,8 л дистиллированной воды. Образец наносили на колонку (5 x 42 см) с SP-Сефарозой Fast Flow (Pharmacia) при скорости потока 600 мл/ч. Перед использованием колонку уравновешивали 50 мМ формиатным буфером pH 3,7. ITF крысы элюировали с колонки 50 мМ муравьиной кислотой pH 3,7, содержащей 50 мМ NaCl. В процессе хроматографии при скорости потока 600 мл/ч отбирали фракции объемом по 100 мл, которые анализировали на содержание rITF. Фракции с предыдущей стадии, содержащие rITF, объединяли (2,3 л) и наносили на колонку (5 х 10 см) с Amberchrome (G-71). Перед использованием колонку уравновешивали 10 мМ аммоний-ацетатным буфером pH 4,8 при скорости потока 0,5 л/ч. После нанесения колонку промывали 0,5 л уравновешивающего буфера и элюировали 10 мМ аммоний-ацетатным буфером pH 4,8, содержащим 60% (v/v) этанола, при скорости потока 0,1 л/ч. Отбирали фракции, содержащие rITF, объемом 10 мл. Концентрацию этанола в объединенных фракциях повышали от 60% (v/v) до 87% (v/v) добавлением 2 объемов этанола (99,9%, v/v) и, охлаждая полученную смесь до температуры минус 25oC в течение 16 ч, осаждали rITF.
Осадок собирали центрифугированием в течение 1 ч при 10000 g и -25oC и перерастворяли при комнатной температуре в 130 мл 20 мМ муравьиной кислоты pH 3,0. Образец наносили на колонку (5 x 20 см) с SP-Сефарозой Fast Flow (Pharmacia) при скорости потока 50 мл/ч. Перед использованием колонку уравновешивали 20 мМ муравьиной кислотой pH 3,0. Пептиды элюировали с колонки с помощью линейного градиента, составленного из 1,5 л 50 мМ муравьиной кислоты pH 3,0 и 1,5 л муравьиной кислоты pH 3,0, содержащей 0,5 М NaCl. В процессе хроматографии при скорости потока 80 мл/ч отбирали фракции (10 мл), в которых измеряли оптическую плотность при длине волны 280 нм. В отобранных фракциях измеряли содержание rITF, после чего фракции, содержащие rITF, объединяли. Дальнейшую очистку ITF крысы проводили с помощью препаративной ВЭЖХ. Объединенные фракции (900 мл) наносили на препаративную С4 ВЭЖХ-колонку (2,2 x 25 см) Vydac 214TP1022, уравновешенную 0,1% (v/v) TFA. Пептиды элюировали при 25oC и скорости потока 5 мл/мин с помощью линейного градиента (540 мл), составленного из MeCN/H2O/TFA (10:89,9:0,1 v/v/v) и MeCN/H2O/TFA (65:34,9: 0,1 v/v/v). Регистрировали поглощение в ультрафиолетовой области при длине волны 280 нм, отбирали фракции объемом 10 мл и анализировали их на содержание rITF. Содержащие rITF фракции объединяли и полученный раствор уменьшали в объеме до 30% центрифугированием под вакуумом. Окончательное выделение rITF осуществляли лиофилизацией. Общее количество rITF, полученное из 8,7 л ферментационной среды, составило 236 мг, что соответствует общему 24% выходу по всем этапам очистки.
Очистка рекомбинантного hITF
Концентрацию hITF в дрожжевой ферментационной питательной среде и фракциях, полученных во время очистки, измеряли с помощью системы ВЭЖХ, идентичной описанной для rITF. Были обнаружены два пика, элюирующиеся с временами удерживания 21,2 мин и 27,1 мин (фиг. 2), представляющие собой по данным масс-спектрометрии и анализу последовательностей димерную и мономерную формы hITF. Уровень экспрессии рекомбинантного ITF человека в данной дрожжевой системе составил 90 мг/л.
С помощью центрифугирования из ферментера вместимостью 10 л было выделено 8,0 л ферментационного бульона. Образец диализовали против 3 смен (каждая длительностью 24 ч) по 40 л 10 мМ муравьиной кислоты pH 2,5. Образец наносили (0,25 л/ч) на колонку (5 x 40 см) с SP-Сефарозой Fast Flow (Pharmacia). Колонку промывали 5 л 20 мМ муравьиной кислоты, pH 2,5 и 5 л муравьиной кислоты pH 2,5, содержащей 1 М NaCl. Отбирали фракции объемом по 100 мл и анализировали их на содержание hITF (фиг. 3). С колонки были элюированы две формы hITF, одна из которых представляла мономерную форму (элюируется при 0,5 М NaCl), а другая - димер hITF (элюируется при 0,78 М NaCI). Фракции, соответствующие каждой из форм, объединяли по отдельности.
Каждую из двух объединенных фракций делили на три равные части (объемом примерно по 700 мл) и наносили на колонку C4 Vydac 214TP1022 (2,2 x 25 см), уравновешенную 0,1% (v/v) TFA. Пептиды элюировали при 25oС и скорости потока 4 мл/мин с помощью линейного градиента (540 мл), составленного из MeCN/H2O/TFA (10: 89,9: 0,1 v/v/v) и MeCN/H2O/TFA (65:34,9:0,1 v/v/v). Поглощение в ультрафиолетовой области регистрировали при длине волны 280 нм, отбирали фракции объемом 10 мл и анализировали их на содержание hITF.
Фракции, содержащие hITF (мономер) и hITF (димер) соответственно, объединяли и осаждали пептиды, добавляя этанол до концентрации 90% (v/v) и выдерживая смесь в течение 72 ч при -25oC. Осадок собирали центрифугированием и лиофилизовали. Общее количество, полученное из 8,0 л ферментационной среды, составило 256 мг для мономера hITF и 133 мг для димера hITF, что соответствует общему выходу по всем этапам очистки в 50% и 65% для мономерной и димерной форм соответственно.
Характеристика рекомбинантных rITF и hITF
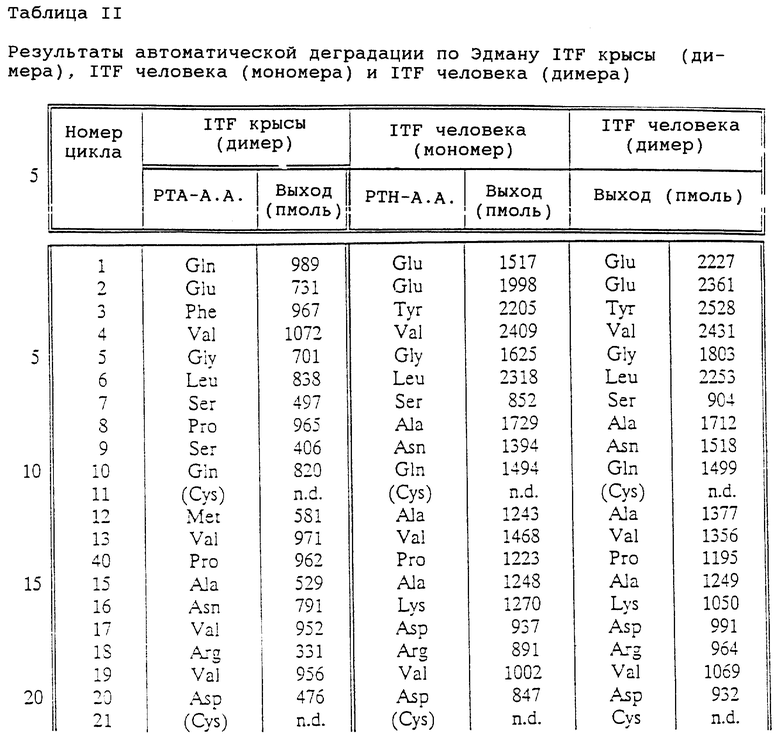
После гидролиза в 6 М HCl в вакуумированных запаянных ампулах в течение 24, 48 и 96 ч образцы (50 мкг) были проанализированы на автоматическом аминокислотном анализаторе фирмы Beckman (Model 121 MB). Количество остатков полуцистина определяли в виде S-β-(4-пиридилэтил)производного после восстановления дисульфидных связей трибутилфосфином [95] с последующим взаимодействием с 4-винилпиридином [96] . Гидролиз образцов, обработанных 4-винилпиридином, проводили в 4 М метансульфоновой кислоте или 3 М меркаптоэтансульфоновой кислоте при 110oC в течение 24 ч, как описано ранее. Анализ аминокислотной последовательности проводили с помощью автоматической процедуры деградации по Эдману, используя газо-фазовый секвенатор Model 470А фирмы Applied Biosystems [97].
Масс-спектрометрический анализ осуществляли на системе API III LC/MS/MS (Sciex, Thornhill, Ont., Canada). Тройной квадрупольный прибор, позволяющий определить значение отношения массы к заряду (m/z) в интервале до 2400, соединен с интерфейсом для пневмоэлектроспрея (называемого также ион-спреем) [98, 99] . Введение образца осуществляли с помощью шприцевого инфузионного насоса (Sage Instruments, Cambridge, MA) через вставной капилляр (внутр. д. 75 мкм) при скорости подачи жидкости 0,5-1 мкл/мин. Шкала m/z прибора была откалибрована с помощью однозарядных ионов аммониевого аддукта полипропиленгликолей (PPG) при единичном разрешении. Точность измерения масс обычно превосходит 0,02%.
На фиг. 4 показаны результаты аналитической ВЭЖХ для очищенных rITF (фиг. 4A) и hITF (фиг. 4B и 4C). Рекомбинантный rITF содержит смесь трех близких пептидов, для разделения которых не было предпринято никаких попыток. В результате масс-спектрометрического анализа были установлены три основных значения молекулярных весов - 13112,2; 13096,6 и 13078,8 (фиг. 5A). Расчетное значение молекулярного веса для мономерной формы ITF крысы, в которой Cys-57 имеет свободную -SH группу, составляет 6558,3. Расчетное значение молекулярного веса для димерной формы ITF крысы (например, с S-S мостиком между двумя Cys-57) составляет 13114,6. Из значений молекулярных весов, найденных для рекомбинантного ITF крысы, ясно, что все три пептида представляют собой димерную форму rITF. Для других "клеверных" пептидов, в которых N-концевым аминокислотным остатком является Gin, например PSP [17, 100], известно, что данный остаток имеет тенденцию к циклизации с образованием пирролидонкарбоновой кислоты (pyrGlu). Для ITF крысы, имеющего предсказанную N-концевую последовательность Gln-Glu-Phe-Val-Gly, разумно предположить, что N-концевой остаток Gln также способен циклизоваться с образованием pyrGlu. Подобное превращение будет приводить к уменьшению значения молекулярного веса ITF крысы (димер) на 17 (один pyrGlu) или 34 (два pyrGlu) единиц массы соответственно. Наблюдаемые значения молекулярных весов в 13096,6 и 13078,8 (фиг.5A) соответствуют димерной форме ITF крысы, в которой один и соответственно два N-концевых остатка Gln зациклизованы. Расчетные значения молекулярных весов для этих форм составляют 13097,6 и 13080,6, что находится в хорошем соответствии с экспериментально полученными величинами. Таким образом, из данных ВЭЖХ (фиг.4A) и масс-спектрометрического анализа (фиг. 5A) вытекает, что рекомбинантный ITF крысы включает 3 различные димерные формы: одну, содержащую 2 N-концевых Gln; одну - с 1 N-концевым Gln и 1 N-концевым pyrGlu; и одну - с 2 N-концевыми pyrGlu. В Таблице I приведен аминокислотный состав ITF крысы, находящийся в хорошем соответствии с ожидаемыми величинами.
фиг. 4B и 4C демонстрируют чистоту hITF (мономера) и hITF (димера) соответственно по данным аналитической ВЭЖХ. Димерная форма (фиг. 4C) выглядит относительно чистой, в то время как для мономерной (фиг. 4B) создается видимость загрязнения материалом, элюирующимся перед пептидом. Однако при рехроматографии материала основного пика получается аналогичный вид хроматограммы (данные не приводятся). По-видимому, это в большей степени указывает на нетипичное поведение hITF (мономера) при обращенно-фазной хроматографии, нежели на наличие примесей. Похожее поведение ранее мы наблюдали для высокоочищенного PSP свиньи, а также оно отмечалось для рекомбинантного hSP высокой степени очистки [93].
С помощью масс-спектрометрического анализа основного пика hITF (мономера) было установлено значение молекулярного веса, равное 6694,0 (фиг. 5B). Значение молекулярного веса, рассчитанное на основании аминокислотной последовательности (фиг. 1), составляет 6574,4 в предположении, что Cys-57 находится в -SH форме. Данные секвенирования (Табл. II) показывают наличие ожидаемой N-концевой последовательности: Glu-Glu-Tyr-Val-Gly-. Данные аминокислотного анализа (Табл. I) показывают ожидаемые величины за исключением наличия 7,3 (8) остатков цистеина. Наличие дополнительного остатка цистеина, соединенного с Cys-57, может увеличивать значение молекулярного веса до 6694,7, что очень близко к величине, найденной с помощью масс-спектрометрии (6694,0). Ввиду этого можно полагать, что в hITF (мономере) остаток Cys-57 связан дисульфидной связью с дополнительным остатком цистеина. Минорный пик (фиг. 5B) в масс-спектре может представлять собой другую производную Cys-57 или может быть обусловлен наличием в препарате примеси.
Расчетное значение молекулярного веса hITF (димера), в котором два мономера соединены дисульфидной связью между двумя остатками Cys-57, составляет 13146,8. Это значение находится в хорошем соответствии с величиной, определенной с помощью масс-спектрометрии (13147, фиг. 5C). Другой пик (13169) вероятно представляет собой Na+ аддукт hITF (димера). Результаты анализа аминокислотной последовательности (Табл. II) и состава (Табл. I) также находятся в хорошем соответствии с ожидаемыми величинами.
Пример 2
Оценка биологического действия димерного ITF с использованием in vitro модели эпителиального барьера (Heather Kindon et al., Trefoil Peptide Protection of Intestinal Epithelial Barrier Function; Gastroenterology 1995; 109 : 516-523)
ITF человека или крысы в димерной форме и/или гликопротеин добавляли к Transwell монослоям клеточной линии T84, полученной из раковой опухоли прямой кишки человека.
Интактный монослой пропускал менее 4% инертного маркера [3Н]маннита за 4 часа. В результате его обработки токсическими лецитин фитогемагглютинином (1 мг/мл), олеиновой кислотой (8 ммоль/л) и таурохлорной кислотой (12 ммоль/л) или токсином А из Clostridium difficile (0,7 мкг/мл) барьерная функция ослабевала и обработанный таким образом монослой пропускал 36%, 62% и 45% [3Н] маннита соответственно.
Добавление ITF человека (1-5 мкг/мкл) приводило к ослаблению повреждения целостности монослоя на величину вплоть до 52%. Защита усиливалась (до 95%) при одновременном присутствии муцинового гликопротеина прямой кишки человека.
Аналогичный эффект наблюдался, когда добавляли ITF крысы сам по себе или в присутствии муцинового гликопротеина прямой кишки человека.
Способность этих веществ обеспечивать некоторую защиту, когда они добавлены сами по себе, указывает на то, что защитные свойства присущи каждому из них и не являются результатом взаимодействия двух компонентов.
Концентрация как ITF, так и муцинового гликопротеина, использованная в этом исследовании, эквивалентна той, которая по оценкам имеет место на поверхности желудочно-кишечного тракта, следовательно, действие на такой модели эпителиального монослоя сравнимо с физиологическим, и можно сделать вывод, что ITF как человека, так и крысы, в сочетании с муциновыми гликопротеинами защищает слизистую оболочку желудочно-кишечного тракта от множества повреждений.
Пример 3
Влияние экзогенного ITF на экспериментальное повреждение желудка у крыс in vivo (Mark W. Babyatsky, Gastroenterology 1996, 110; 489-497)
Повреждение желудка индуцировали либо внутрижелудочным введением абсолютного этанола (1,0 мл), либо подкожным введением индометацина (20 мг/кг). В различных дозах (0,5, 1, 5 и 10 мг, каждая доза растворена в 1,0 мл дистиллированной воды) вводили димерный ITF крысы в различные моменты времени как до, так и после повреждения. В качестве контроля использовали носитель или бычий сывороточный альбумин. Затем крыс умерщвляли и исследовали повреждение желудка (макроскопически и гистологически). В результате введения ITF через рот наблюдался значительный защитный эффект по отношению к повреждениям, вызываемым как этанолом, так и индаметацином (P < 0,005).
Пример 4
Получение рекомбинантного pS2 человека
Рекомбинантный pS2 человека в мономерной и димерной форме были получены в Escherichia coli с использованием рекомбинантной плазмиды для экспресии pEZZ18, кодирующей либо серин (для мономера), либо цистеин (для димера) в положении 58 этого белка (Chadwick MP, May FEB. Westley BR. Production and comparison of mature single domain "trefoil" peptides pNR-2/pS2 Cys58 and pNR- 2/pS2 Ser58. Biochem. J. 1995, 308; 1001-1007). Эта система экспрессии продуцирует составной белок, содержащий два искусственных домена: полученный из Staphylococcus aureus белок А и белок pS2. Рекомбинантные составные белки очищали с помощью аффинной хроматографии на lgG сефарозе и высвобождали белок pS2 путем расщепления фактором Ха. Затем мономер pS2 Ser58 очищали анионобменной хроматографией. Белок pS2 Cys58 обрабатывали цистеином, очищали ионобменной хроматографией, и димер очищали гель-фильтрацией. Молекулярные массы мономера и димера, измеренные с помощью масс-спектрометрии с лазерной десорбцией, составили 6645 и 13378 Да и хорошо согласуются с теоретическими 6674 и 13340 Да.
Пример 5
Оценка биологического действия димерного или мономерного pS2 человека с использованием in vitro и in vivo модели ранения и восстановления (Tania Marchbank et al., Dimerisation of human pS2..., Journal of pathology, vol. 185; 153-158 (1998))
Модель in vitro
Скорость миграции клеток к лидерному краю поврежденного монослоя клеточной линии НТ29 прямой кишки человека увеличивалась при добавлении димерного или мономерного pS2 (0,65-325 мкг/мл). Димерный pS2 оказывал более сильное действие, чем мономерный, во всех испытанных дозах.
Модель in vivo
Действие димерного и мономерного pS2 человека (в мономерном Cys58 заменен на Ser58) сравнивали на модели заживления раны у крыс. Димерный pS2 в дозе 25 и 50 мкг/кг в час показал 50%-ное и 70%-ное восстановление повреждения желудка, индуцированного индометацином. Мономерный pS2 в той же дозе был значительно менее эффективным в восстановлении повреждения (примерно в половину).
Согласно вышесказанному, можно сделать вывод, что pS2 восстанавливает повреждение желудка и имеет защитное и заживляющее действие, причем димерный pS2 более эффективен, чем мономерный.
Пример 6
Получение димера pS2 крысы
Тотальная РНК была экстрагирована из различных тканей крыс, в частности из желудка и кишечника. С помощью обратной транскриптазы и ПЦР с различными праймерами была получена однонитевая кДНК.
С использованием полученной кДНК и плазмиды для экспрессии было осуществлено клонирование и экспрессия pS2 в E.coli, как описано в примере 4. Клонирование и экспрессия pS2 крысы в дрожжах осуществлялась так же, как описано для ITF человека и крысы в примере 1.
Пример 7
В связи с тем, что "клеверные" пептиды не разрушаются в пищеварительном тракте, одним из путей их введения является пероральный. Эти пептиды могут быть введены, например, в форме таблетки, капсулы или пилюли или суспендированы в растворе, таком как сироп, который проглатывает пациент. В альтернативном случае раствор, содержащий указанный пептид, может быть введен в виде желудочного промывания или в форме суппозитория. Фармацевтические композиции, содержащие соединение по изобретению, могут быть приготовлены по традиционным технологиям, например как описано в Remington: The Science and Practise of Pharmacy, 19th Ed., 1995.
Типичная таблетка, которую готовят по традиционной технологии, имеет следующую пропись, мг;
Сердцевина:
Активное соединение (ITF димер человека) - 10
Коллоидный диоксид кремния (Aerosil) - 1,5
Микрокристаллическая целлюлоза (Avicel) - 70
Модифицированная целлюлозная смола (Ac-Di-Sol) - 7,5
Стеарат магния - Ad.
Оболочка:
HPMC - 9
Mywacett 9-40 Т - 0,9
Сначала готовят сердцевинный материал таблеток, а затем его покрывают оболочкой. Mywacett представляет собой ацилированный моноглицерид, используемый для пленочного покрытия.







Изобретение касается димеров "клеверных" пептидов. "Клеверный" пептид содержит единственный домен в форме клеверного листа и представляет собой ITF человека или ITF крысы или pS2 человека или pS2 крысы. Пептид находится в форме димера, в котором два идентичных мономера соединены дисульфидной связью между двумя остатками цистеина. Молекулярный вес пептида около 13000. Культивируют подходящую клетку-хозяина, трансформированную ДНК, кодирующей ITF человека или крысы или pS2 человека или крысы и отделяют димерную форму от мономерной. Фармацевтическая композиция включает "клеверный" пептид и фармацевтически приемлемый разбавитель. Изобретение позволяет на основе димерных форм "клеверных" пептидов получать лекарства, используемые для заживления повреждений слизистой оболочки желудочно-кишечного тракта, например пептической язвы. 11 с. и 4 з.п. ф-лы, 9 ил., 2 табл.
| WO 9214837 А1, 03.09.1992 | |||
| Thim, R | |||
| Устройство двукратного усилителя с катодными лампами | 1920 |
|
SU55A1 |
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
| Poulsom R., Wright N.A | |||
| Am.J | |||
| Physiol | |||
| САННЫЙ ВЕЛОСИПЕД С ВЕДУЩИМ КОЛЕСОМ, СНАБЖЕННЫМ ШИПАМИ | 1921 |
|
SU265A1 |
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
| Hoffmann W., and Hauser F | |||
| Trends Biochem | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
| RU 94023808 А1, 27.06.1996 | |||
| RU 94031167 А1, 20.04.1994 | |||
| ПРОИЗВОДНЫЕ ПЕНТАПЕПТИДОВ | 1991 |
|
RU2010799C1 |

