Изобретение относится к области биологии, в частности молекулярной биологии и генетической инженерии, и может быть использовано для медико-биологических исследований и биотехнологического производства рекомбинантных белков путем их секреции клетками бактерий.
Известно, что для секреции экспрессируемых белков с использованием метилотрофных дрожжей Pichia pastoris широко используется вектор pPIC9 (Scorer, С. А., Clare, J.J., McCombie, W.R., Romanos, M.A. and Sreekrishna, K. (1994) Rapid selection using G418 of high copy number transformants of Pichia pastoris for high-level foreign gene expression. Biotechnology 12:181-184). В этом векторе для секреции полипептидов применяется лидерная последовательность препропептида альфа-фактора спаривания S. cerevisiae. Этот вектор был успешно применен для экспрессии гликопротеина Olea e 1 из оливкового дерева Olea europaea, являющегося аэроаллергеном. Данная система позволяет получать от 60 до 300 мг гликозилированного белка с правильным фолдингом на 1 л культуры (Huecas, S., Villalba, M., Gonzalez, E., Martinez-Ruiz, A., Rodriguez, R. (1999) Production and detailed characterization of biologically active olive pollen allergen Ole e 1 secreted by the yeast Pichia pastoris. Eur. J. Biochem. 261: 539-546), а также для секреции ряда других полипептидов эукариотической природы. Кроме того, лидерная последовательность препропептида альфа-фактора спаривания S. cerevisiae используется также в векторах для секреции гетерологичных белков в других штаммах дрожжей, Kluyveromyces lactis (Hsieh, H.P. and Da Silva, N.A. (1998) Partial-pKDl plasmids provide enhanced structural stability for heterologous protein production in K. lactis. Appl. Microbiol. Biotechnol. 49:411-416), Schizosaccaramyces pombe (Okada, H., Sekiya, Т., Yokoyama, K., Tohda, H., Kumagai, H. And Morikawa, Y. (1998) Efficient secretion of Trichoderma reesei cellobiohydrolase II in Schizosaccaramyces pombe. Appl. Microbiol. Biotechnol. 49: 301-308) и S. cerevisiae (Chung, B. H. and Park, K.S. (1998) Simple reducing proteolysis during secretory production of human hormone in Saccharomyces cerevisiae. Biotechnol Bioeng. 57: 245-249). Для секреции белков клетками эубактерий используется лидерный пептид белка внешней мембраны ОmpА (серия экспрессионных векторов pIN-III) (Ghraeb, J. , Kimura, H. , Takahara, M., Hsiung, H., Masui, Y., Inouye, M. (1984) Secretion cloning vectors in Escherichia coli. EMBO J. 3: 2437-2442). Он позволяет секрецию лишь в периплазматическое пространство, а не непосредственно в среду культивирования. Для секреции гибридных белков грамположительными бактериями применяется лидерный пептид пенициллиназы из алкалофильной бациллы, который используется вместе с полноразмерным геном пенициллиназы (Kato, С., Horikoshi, К. (1990) Polypeptide secretion expression vector, microorganism transformed with the same vector and production of polypeptide by the same microorganism. Patent: JP 1990100685-A).
Недостатками указанных способов является невозможность прямой секреции целевого продукта (например, белка) в культуральную жидкость с помощью грамотрицательных бактерий и невозможность применения описанных способов секреции к клеткам цианобактерий.
Задачей изобретения является разработка нового способа секреции белков клетками бактерий, включая цианобактерии, непосредственно в среду культивирования, что позволит упростить процесс последующей очистки целевого продукта и повысит его качество.
Указанная задача решается за счет применения новых химических веществ - лидерных последовательностей и сконструированных на их основе векторов, содержащих лидерные последовательности с последующей генетической трансформацией и культивированием бактерий-продуцентов, включая трансформированных фотоавтотрофных цианобактерий.
В векторах для трансформации клеток бактерий использованы молекулы нуклеиновой кислоты (Формула п.1 и 2), кодирующие лидерные пептиды (Формула п. 6 и 7, в результате чего получены штаммы бактерий-продуцентов, секретирующие целевой полипептид в среду культивирования. Суть изобретения состоит в том, что для получения штаммов бактерий-продуцентов используются векторы, содержащие оригинальные молекулы нуклеиновых кислот, кодирующие лидерные последовательности, которые обеспечивают секрецию белков непосредственно в среду культивирования. Полученные бактерии-продуценты, включая штаммы цианобактерий, секретируют целевой полипептид в среду культивирования, что обеспечивает его эффективное выделение и очистку.
Изобретение имеет "изобретательский шаг", так как впервые предлагает способ секреции белков в бактериях с использованием молекул нуклеиновых кислот, кодирующих лидерные последовательности секретируемых белков цианобактерии, для получения полипептидов как в гетеротрофных клетках Е.coli, так и в фотоавтотрофных клетках Synechocystis. Применение этих секреторных лидерных последовательностей обеспечивает эффективную секрецию целевого белка в среду культивирования на уровне 50-75% от всего синтезированного в клетках целевого белка.
Изобретение иллюстрируется следующим примером.
Получение последовательности, конструирование вектора и синтез целевого белка.
Пример является одним из вариантов осуществления изобретения, служит для иллюстрации и не ограничивает область возможного использования объектов, изложенных в формуле изобретения, для синтеза и секреции в культуральную среду других целевых продуктов.
Краткое описание фигур.
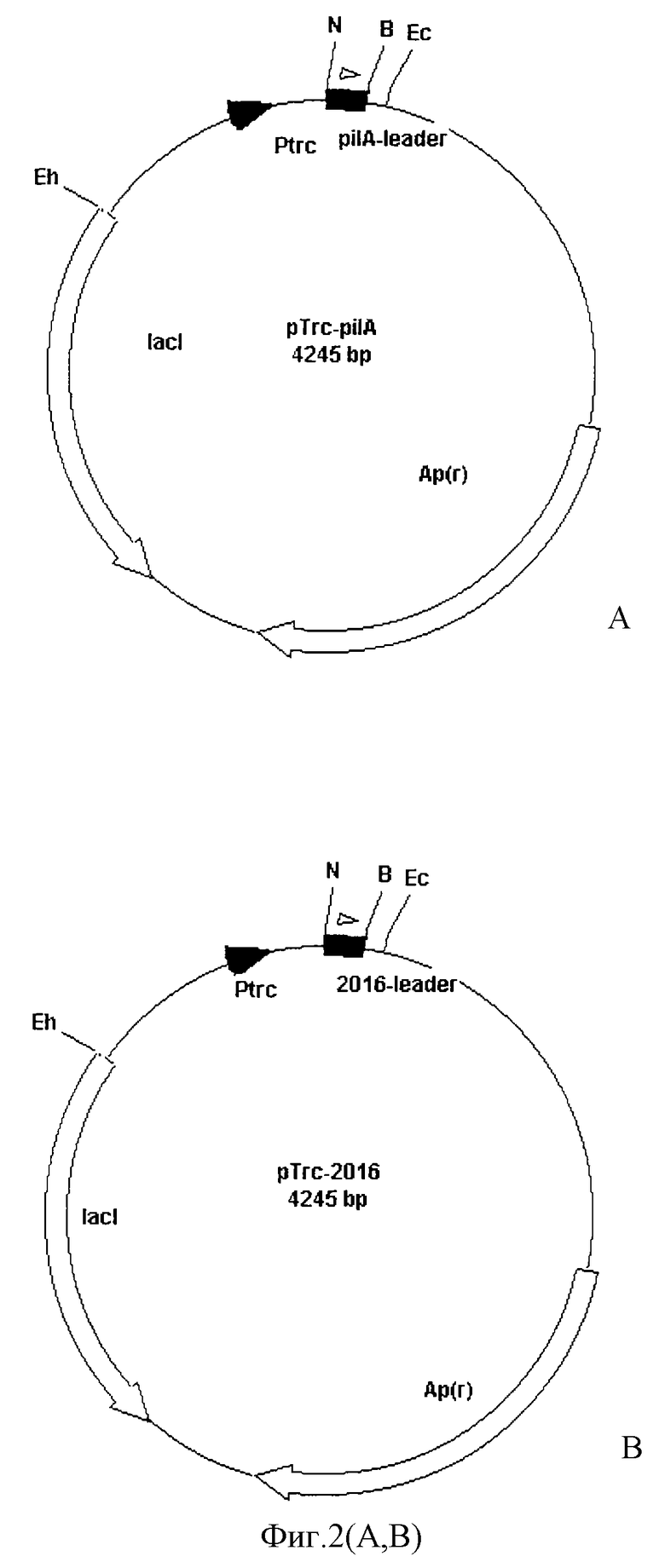
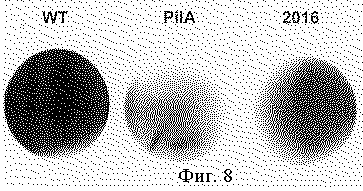
Фиг. 1 - молекулы нуклеиновых кислот, кодирующие лидерные последовательности для секреции полипептидов в культуральную среду. Фиг.2 (А, В) - схема векторов для экспрессии и секреции полипептидов в среду культивирования, содержащие в качестве действующей части молекулы нуклеиновых кислот, представленных на фиг.1. Фиг.3 (А, В) - схемы векторов для трансформации клеток бактерий, имеющие молекулы нуклеиновых кислот для секреции, промотора, целевого полипептида, гена устойчивости к антибиотику и участки нуклеиновых кислот для интеграции векторов в геном цианобактерии. Фиг.4 - тест на активность целевого полипептида в клетках Escherichia coli, трансформированных векторами, которые экспрессируют целевой полипептид. Фиг.5 - тест на активность целевого полипептида в среде культивирования клеток Escherichia coli, трансформированных векторами, которые экспрессируют целевой полипептид. Фиг. 6 - функциональный тест в ПААГ на активность целевого полипептида, секретируемого в среду культивирования клеток Escherichia coli. Фиг.7 - тест на активность целевого полипептида в клетках Synechocystis, трансформированных векторами, которые экспрессируют целевой полипептид. Фиг.8 - тест на активность целевого полипептида в среде культивирования клеток Synechocystis, трансформированных векторами, которые экспрессируют целевой полипептид. Фиг. 9 - функциональный тест в ПААГ на активность целевого полипептида в среде культивирования и в клетках Synechocystis, трансформированных векторами, которые экспрессируют целевой полипептид.
При определении белков, секретируемых цианобактерией Synechocystis в среду культивирования, обнаружили, что один из белков составляет до 70% от суммы секретируемых белков. Определение N-концевой последовательности белка обнаружило последовательность FTLIELLVVVIIIGVLAAIALP, в которой первый фенилаланин был метилирован. На основании этих данных был идентифицирован полноразмерный ген, получивший название РilА, и его возможный гомолог, белок, названный 2016, и определены нуклеотидные последовательности, кодирующие лидерные пептиды этих белков. На основании этих данных были синтезированы синтетические олигонуклеотиды, соответствующие этим пептидам, которые были использованы для конструирования векторов для секреции белков в клетках Е. coli и Synechocystis. Нуклеотидная последовательность, кодирующая лидерный пептид PilA, состоит из 68 нуклеотидов и кодирует олигопептид из 23 аминокислот. Нуклеотидная последовательность, кодирующая лидерный пептид 2016, состоит из 59 нуклеотидов и кодирует олигопептид из 20 аминокислот. Предлагаемые конструкции содержат сильный конститутивный промотор Ptrc и синтетические лидерные последовательности для секреции.
Синтезированы молекулы нуклеиновых кислот, кодирующие лидерные последовательности для секреции полипептидов в культуральную среду и характеризующиеся формулой, представленной на фиг.1. Был проведен попарный отжиг комплиментарных синтетических олигонуклеотидов, представленных на фиг.1 с получением молекул двуцепочечных нуклеиновых кислот, имеющих с одной стороны выступающий конец для лигирования с сайтом, генерируемым эндонуклеазой рестрикции Nco I, а с другой эндонуклеазой рестрикции Bam HI. Полученные молекулы нуклеиновых кислот были лигированы в аналогичные сайты вектора рТrc99А с получением гибридных векторов, характеризующихся наличием промотора и последовательностей нуклеиновых кислот, обеспечивающих секрецию (фиг.2). Затем фрагменты векторов были вырезаны из полученных конструкций с помощью ферментов Ehe I и Bam HI, очищены с помощью электрофореза в агарозном геле и выделены из геля в чистой форме. Эти фрагменты были лигированы в сайты Hind III (конец предварительно регенерирован до тупого с помощью фрагмента Кленова ДНК-полимеразы I) и Bam HI плазмиды pLicB-Km, несущей ген licВ, кодирующий лихеназу из термофильной бактерии (конец предварительно регенерирован до тупого с помощью фрагмента Кленова ДНК-полимеразы I). Лигирование фрагментов было осуществлено таким образом, что рамка считывания лидерных пептидов совпадала с рамкой считывания гена licВ, в результате чего были получены векторы, характеризующиеся наличием промотора, молекул нуклеиновых кислот, обеспечивающих секрецию (фиг.1), гена целевого репортерного полипептида лихеназы licВ и гена устойчивости к антибиотику канамицину. При этом лидерпые пептиды и лихеназа образовали гибридные белки с одной рамкой считывания (фиг.2). Далее, фрагменты векторов, содержащие промотор, молекулы нуклеиновых кислот, обеспечивающие секрецию (фиг.1), ген целевого репортерного полипептида лихеназы и ген устойчивости к антибиотику канамицину, были выделены с использованием ферментов Ecl 137 I и Sma I и клонированы в сайт Ecl 137 I вектора p6803-platform, который содержал нейтральный фрагмент геномной ДНК цианобактерии Synechocystis, необходимый для интеграции в геном этой цианобактерии (фиг. 3). В результате были сконструированы векторы р6803-licB/рilА и р6803-licB/2016 для секреции белков клетками Escherichia coli и Synechocystis (фиг.3). При этом в клетках Е. coli векторы поддерживали автономную репликацию, а в клетках цианобактерии они были интегрированы в геном.
Клетки бактерий были трансформированы полученными векторами. Анализируемые клоны Е. coli высевались уколом на чашки со стандартной средой LB, содержащей ампициллин (Ар) и канамицин (Km) в конечных концентрациях 100 и 50 мкг мл-1 соответственно, и выращивались в течение ночи при 37oС. Чашки заливали верхним агаром (0,7% агарозы в 50 мМ Трис-НСl рН8), содержащим 0,05% лихенана (Sigma, США). После застывания агара чашки инкубировали в течение 2 ч при 65oС. Окраску лихенана проводили 0,5%-ным водным раствором Конго красного в течение 10 мин при комнатной температуре и периодическом перемешивании. После окрашивания чашки отмывали 1М раствором NaCl три раза по 5 мин при комнатной температуре. Клетки трансформантов, несущие ген licВ, фланкированный лидерными последовательностями РilА и 2016 и находящийся под контролем сильного промотора, синтезировали лихеназу (фиг.4), в то время как клетки трансформантов, в которых ген licВ находится без промотора, не продуцировали лихеназу. Секреция лихеназы в культуральную среду тестировалась тестом на чашках. Клетки отделяли от культуральной среды двукратным центрифугированием с последующей фильтрацией через нейлоновые фильтры, не пропускающие клетки бактерий. Полученную среду смешивали с равным объемом 3%-ной агаризованной среды LB, разливали на чашки Петри и проводили тест на лихеназную активность, как описано выше (фиг.5). В клетках, трансформированных конструкциями, содержащими лидерные последовательности, наблюдали лихеназную активность в среде культивирования. В клетках, несущих конструкции без лидерных последовательностей, лихеназная активность не обнаруживалась (фиг.5).
Белки из культуральных сред концентрировали осаждением сульфатом аммония и разделяли гель-электрофорезом в денатурирующих условиях одновременно в 12%-ных ПААГ и в 12%-ных ПААГ, содержащих лихенан в концентрации 0,2%. Контрольные гели, не содержащие лихенан, окрашивали раствором Кумасси R-250 для визуализации всех белков, присутствовавших в культуральной жидкости. Гели с лихенаном окрашивали Конго красным для проявления активности лихеназы. В препарате, соответствующем контрольной плазмиде p6803-licB/control, несущей ген licВ без лидерной последовательности, не наблюдалось четко выраженных полос, представленных белком, соответствующим LicB (фиг.6,А). В препаратах, соответствующих плазмидам p6803-licB/pilA и р6803-licВ/2016 присутствуют две полосы в районе 32-36 кДа. Эти белки представляют собой две формы лихеназы - полноразмерную и процессированную внутриклеточной пептидазой Е. coli со стороны карбоксильного конца. Тест на лихеназную активность демонстрирует присутствие лихеназы в препаратах белков, полученных из сред культивирования клеток, трансформированных плазмидами р6803-licB/рilА и р6803-licB/2016, содержащими лидерные последовательности, и полное отсутствие лихеназной активности в контрольном препарате, соответствующем плазмиде p6803-licB/control, не содержащей лидерной последовательности. Экспрессия и секреция контрольного белка, лихеназы, в клетках Synechocystis, тестировалась таким же способом, как описано выше для клеток Е. coli. Клетки Synechocystis были трансформированы плазмидами p6803-licB/pilA и р6803-licВ/2016 с получением трансформантов, в которых экспрессионные кассеты были встроены в геном цианобактерии. В клетках трансформантов licВ/pilA и licВ/2016 наблюдалась эффективная экспрессия лихеназы (фиг.7). В обоих случаях наблюдали секрецию лихеназы в среду культивирования, как в тестах на чашках (фиг.8), так и в тестах с использованием разделения секретируемых белков с помощью ПААГ (фиг.9).
Идентификация заявленных молекул нуклеиновых кислот, кодирующих лидерные последовательности для секреции полипептидов (Формула п.1 и 2) была осуществлена с помощью секвенирования векторов (Формула п.3-5) с использованием специфических праймеров для плазмиды рТrc99А. Секвенирование подтвердило идентичность структуры молекул нуклеиновых кислот (фиг.1). Идентификация заявленных олигопептидов (Формула п. 6 и 7) осуществлена путем сравнения аминоконцевых аминокислотных последовательностей белков, синтезированных в клетках, и процессированных белков, обнаруженных в культуральной жидкости. Секвенирование аминокислотных последовательностей подтвердило их идентичность последовательностям, представленным на фиг.1.
В результате заявляемого изобретения был разработан новый способ секреции белков в культуральную среду клетками бактерий, с использованием молекул нуклеиновых кислот, кодирующих лидерные последовательности секретируемых белков цианобактерии, для получения полипептидов как в гетеротрофных клетках Е. coli, так и в фотоавтотрофных клетках Synechocystis. Применение этих секреторных лидерных последовательностей обеспечивает эффективную секрецию целевого белка в среду культивирования на уровне 50-75% от всего синтезированного в клетках целевого белка.






Изобретение относится к молекулярной биологии и генетической инженерии и может быть использовано для медико-биологических исследований и биотехнологического производства рекомбинантных белков путем их секреции клетками бактерий. Молекула нуклеиновой кислоты (НК), кодирующая лидерную последовательность для секреции полипептидов в культуральную среду, имеет установленную нуклеотидную последовательность. Последовательность приведена в формуле изобретения. Вектор получен на основе плазмиды р6803-platform и содержит в качестве действующей части молекулу НК, кодирующую лидерную последовательность для секреции полипептидов в культуральную среду. Вектор может содержать фрагмент геномной ДНК цианобактерий Synechocystis для интеграции в геном этой цианобактерии. Представлены олигопептиды размером 20 и 23 аминокислоты. Аминокислотные последовательности приведены в формуле изобретения. Клетку бактерий трансформируют вектором и культивируют. Секреция целевого белка происходит непосредственно в среду. Используют бактерии E.coli или цианобактерии. Изобретение позволяет упростить процесс очистки целевого продукта и повысить его качество. 7 с. и 7 з.п. ф-лы, 9 ил.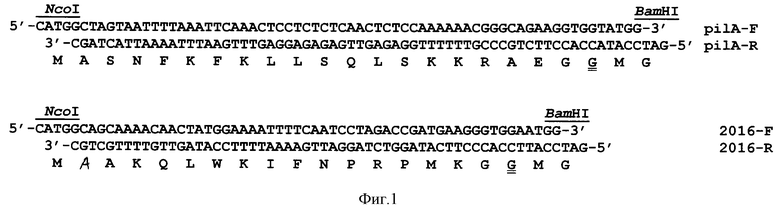
CCATGGCTAGTAATTTTAAATTCAAACTCCTCTCTCAACTCTCCAAAAAAСGGGCAGAAGGTGGTATGGATCC,
кодирующая лидерную последовательность для секреции полипептидов в культуральную среду.
CCATGGCAGCAAAACAACTATGGAAAATTTTCAATCCTAGACCGATGAAGGGTGGAATGGATCC,
кодирующая лидерную последовательность для секреции полипептидов в культуральную среду.
MASNFKFKLLSQLSKKRAEGGMG,
являющийся лидерным пептидом для секреции белков.
MАAKQLWKIFNPRPMKGGMG,
являющийся лидерным пептидом для секреции белков.
| ЕЗЕПЧУК Ю.В | |||
| Биомолекулярные основы патогенности бактерий | |||
| - М.: Наука, 1977, с.106-137 | |||
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, БЕЛОК И СПОСОБ ПОЛУЧЕНИЯ БЕЛКА | 1991 |
|
RU2103364C1 |
| ЗАПОРНОЕ УСТРОЙСТВО ДЛЯ ТРУБОПРОВОДОВ | 1995 |
|
RU2100685C1 |

