Изобретение является частичным продолжением заявки No 08/327690, зарегистрированной 24 октября 1994 года, которая является частичным продолжением PCT/US 94/05779, зарегистрированной 23 мая 1994 года и частичным продолжением заявки No 08/065725, зарегистрированной 21 мая 1993 года, которая является частичным продолжением PCT/US92/01636 и заявки No 07/662920, зарегистрированной 28 февраля 1991 года с передачей прав заявки No 08/164666, каждая из которых специально включена здесь во всей своей полноте в виде ссылок.
Область изобретения
Настоящее изобретение относится к белкам, используемым в качестве антикоагулянтов. Более подробно, настоящее изобретение относится к модифицированным формам Фактора VII, ингибирующим свертывание крови и фактор ткани.
Предпосылки к созданию изобретения
Свертывание крови представляет собой процесс, включающий сложное взаимодействие различных компонентов крови или факторов, который в конечном счете приводит к образованию фибринового сгустка. Обычно компоненты крови, которые участвуют в том, что определяется как коагуляционный "каскад", являются проэнзимами или зимогенами - энзиматически неактивными белками, которые превращаются в протеолитические ферменты при действии активатора, активированного фактора свертывающей системы крови. Факторы свертывания, которые подвергаются такому превращению, обычно относятся к "активным факторам" и обозначаются добавлением прописного индекса "а" (например, Фактор VIIa).
Активированный Фактор Х ("Ха") необходим для превращения протромбина в тромбин, который затем на заключительной стадии образования фибринового сгустка превращает фибриноген в фибрин. Существуют две системы или два метаболических пути, способствующих активации Фактора X. "Внутренний метаболический путь" охватывает те реакции, которые приводят к образованию тромбина посредством утилизации факторов, присутствующих только в плазме. Серия опосредованных протеазой реакций в итоге генерирует Фактор IXa, который в сочетании с Фактором VIIIa расщепляет Фактор Х в Ха. Идентичный протеолиз во "внешнем метаболическом пути" осуществляется Фактором VIIa и его кофактором, фактором ткани. Фактор ткани представляет собой связанный с мембраной белок и обычно не циркулирует в плазме. Однако при разрыве сосуда он может образовывать комплекс с Фактором VIIa для катализа активации Фактора Х или Фактора IX в присутствии иона Са++ и фосфолипида (Nemerson and Gentry, Biochem. 25: 4020-4033 (1986)). Несмотря на то, что относительное значение двух метаболических путей коагуляции в гемостазе остается неясным, в последние годы было найдено, что Фактор VII и фактор ткани играют основную роль в регуляции свертывания крови.
Фактор VII является маркерным гликопротеином плазмы, который циркулирует в крови в виде одноцепочечного зимогена. Зимоген каталитически неактивен (Williams et al. , J. Biol. Chem. 264:7536-7543 (1989); Rao et al., Proc. Natl. Acad. Sci. USA. 85:6687-6691 (1988)). Одноцепочечный Фактор VII может быть превращен in vitro в двухцепочечный Фактор VIIa Фактором Ха, Фактором ХIIа, Фактором IXa или тромбином. Фактор Ха считается основным физиологическим активатором Фактора VII. Подобно некоторым другим белкам плазмы, включенным в гемостаз, Фактору VII для активности, связанной с гамма-карбоксилированием многочисленных остатков глутаминовой кислоты, сосредоточенных на амино-конце белка, необходим витамин К. Эти гамма-карбоксилированные глутаминовые кислоты необходимы для взаимодействия Фактора VII с фосфолипидами в присутствии ионов металла.
Превращение зимогенного Фактора VII в активированную двухцепочечную молекулу происходит при расщеплении внутренней пептидной связи, расположенной примерно в середине молекулы. В человеческом Факторе VII сайт активационного расщепления находится при Arg152-lle153 (Hagen et al., Proc. Natl. Acad. Sci. USA. 83:24122416 (1986); Thim et al., Biochem. 27:7785-7793 (1988), обе работы включены здесь в виде ссылок). Бычий Фактор VII активируется расщеплением по аналогичной связи Arg152-Ile153 (Takeya et аl., J. Biol. Chem. 263: 14868-14877 (1988)). В присутствии фактора ткани, фосфолипидов и ионов кальция двухцепочечный Фактор VIIa ограниченным протеолизом быстро активирует Фактор Х или Фактор IX.
Часто бывает необходимым селективно блокировать коагуляционный каскад у пациента. Антикоагулянты, такие как гепарин, кумарин, производные кумарина, индандиона или другие агенты, могут быть использованы, например, при почечном диализе или в лечении тромбоза глубоких вен, коагулопатии потребления (DIC) и множества других медицинских нарушений. Например, обработка гепарином или экстракорпоральная обработка цитрат-ионом (US Patent 4500309) могут быть использованы при диализе, чтобы предотвратить коагуляцию в ходе лечения. Гепарин используется также для предотвращения тромбозов глубоких вен у пациентов, подвергающихся хирургической операции.
Однако лечение гепарином и другими антикоагулянтами может иметь нежелательные побочные эффекты. Все известные антикоагулянты, как правило, действуют по всему организму, а не специфично на сгусток. Например, гепарин может вызвать тяжелое кровотечение. Более того, имея время полувыведения примерно 80 минут, гепарин быстро исчезает из крови, тем самым вызывая необходимость частого введения. Из-за того что гепарин действует как кофактор антитромбина III (ATIII), а АТIII быстро убывает при лечении DIC, бывает очень трудно поддерживать надлежащую дозировку гепарина, что вызывает необходимость продолжительного мониторинга ATIII и уровней гепарина. Гепарин также неэффективен, если снижение ATIII экстремально. Кроме того, продолжительное использование гепарина может усилить агрегацию тромбоцитов и понизить их количество и явиться одной из причин развития остеопороза. Производные индандиона также могут иметь побочные токсические эффекты.
Было найдено, что, кроме антикоагулянтов, кратко описанных выше, некоторые природные белки обладают антикоагулянтной активностью. Например, Reutelingsperger (US Patent No 4736018) выделил антикоагулянтные белки из бычьей аорты и сосудов человеческой пупочной вены. Maki и др. (US Patent No 4732891) открыли антикоагулянтные белки в человеческой плаценте. Кроме того, в качестве терапевтического антикоагулянта был предложен ATIII (Schipper et al. , Lancet 1 (8069):854-856 (1978); Jordan, US Patent No 4386025; Bock et al., US Patent No 4517294).
Пролиферация клеток гладкой мышцы (SMCs) сосудистой стенки является важным событием при возникновении сосудистых поражений при атеросклерозе, после восстановления сосудов или в ответ на другие сосудистые повреждения. Например, лечение атеросклероза часто включает в себя очищение закупоренных сосудов ангиопластикой, эндартерэктомией или редукционной атерэктомией, или имплантацией шунтов - хирургические процедуры, при которых атеросклеротические бляшки спрессовываются или удаляются посредством катетеризации (ангиопластика), удаляются с артериальной стенки посредством рассечения (эндартерэктомия) или шунтируются природными или синтетическими имплантатами. Эти процедуры удаляют сосудистый эндотелий, нарушают находящуюся под ним внутреннюю оболочку (интиму) и приводят к гибели медиальных SMCs. Повреждение сопровождается пролиферацией медиальных SMCs и миграцией их во внутреннюю оболочку, что характерным образом проявляется в первые недели и вплоть до шести месяцев после повреждения и прекращается, когда происходит восстановление поверхностного эндотелиального слоя. У людей эти повреждения составляют примерно 20% клеток и 80% внеклеточного матрикса.
Примерно у 30% пациентов и более, подвергшихся ангиопластике, эндартерэктомии или шунтированию, тромбоз и/или пролиферация SMCs во внутреннюю оболочку вызывает повторное слипание сосуда и в результате этого несостоятельность восстановительной хирургии. Такое закрытие сосуда после хирургического вмешательства известно как рестеноз.
Все еще существует необходимость в создании улучшенных составов с антикоагулянтной активностью, которые могут быть введены в сравнительно низких дозах и не дают нежелательных побочных эффектов, присущих традиционным антикоагулянтам. Настоящее изобретение реализует эту необходимость, обеспечивая антикоагулянты, которые действуют специфически в сайтах повреждения и дополнительно обеспечивают другие родственные преимущества.
Сущность
Приготовлены новые составы, включающие модифицированный Фактор VII, обладающие антикоагулянтными свойствами. Составы с модифицированным Фактором VII ингибируют также фактор ткани. Последовательность Фактора VII имеет, по крайней мере, одну аминокислотную модификацию, которая выбрана таким образом, что существенно снижает способность активированного Фактора VII катализировать активацию плазматических Факторов Х или XI и вследствие этого возможность ингибировать свертывающую активность. Новый Фактор VII имеет активный центр, модифицированный заменой, по крайней мере, одной аминокислоты и в своей модифицированной форме способен связывать фактор ткани. Для составов модифицированного Фактора VII типичной является в значительной степени чистая форма.
Составы изобретения особенно полезны для лечения больных, когда превращены в фармацевтические составы, которые могут быть даны пациентам, страдающим от различных болезней, чтобы лечить связанные с коагуляцией состояния. Такие молекулы модифицированного Фактора VII, способные связывать фактор ткани, но имеющие значительно сниженную способность катализировать активацию других факторов в свертывающем каскаде, могут обладать более длительным периодом полувыведения из плазмы и вследствие этого соответственно большей продолжительностью периода противосвертывающей активности по сравнению с другими анти коагулянтами.
Среди медицинских показаний для составов изобретения есть те, которые обычно лечат антикоагулянтами, такие как, например, тромбоз глубоких вен, легочная эмболия, удар, коагулопатия потребления (DIC), отложения фибрина в легких и почках, связанные с грам-отрицательной эндотоксемией, и инфаркт миокарда. Составы могут быть использованы для подавления сосудистого рестеноза, возникающего после механического повреждения сосуда, такого как повреждение, вызванное баллонной ангиопластикой, эндартерэктомией, редукционной атерэктомией, установкой стента, лазерной терапией или иссечением распадающейся ткани, или возникающего дополнительно к сосудистым имплантатам, стентам, шунтам или трансплантатам органов. Составы могут быть использованы для подавления отложения тромбоцитов и связанных с этим нарушений. Метод подавления коагуляции, сосудистого рестеноза или отложения тромбоцитов состоит во введении пациенту состава, включающего в себя модифицированный Фактор VII, имеющий, по крайней мере, одну замененную аминокислоту в каталитической триаде Ser344, Asp242 и His193 в количестве, достаточном для эффективного подавления коагуляции, сосудистого рестеноза или отложения тромбоцитов. Метод также находит применение в лечении острой закупорки коронарной артерии у больного и состоит во введении модифицированного Фактора VII, который включает в себя DEGR-Фактор VII в сочетании с активатором тканевого плазминогена (tРА) или стрептокиназой и может ускорять tPA-индуцированный тромболиз.
Характерно, что фармацевтические составы для введения людям будут состоять из белка модифицированного Фактора VII и фармацевтически приемлемых носителей и буферов.
В приведенных воплощениях человеческого и бычьего Фактора VII остаток Ser344 в активном центре модифицирован, заменен на Gly, Met, Thr или, предпочтительнее, на А1а. Такая замена может быть сделана отдельно или в комбинации с заменой(нами) в других сайтах каталитической триады, которая включает Asp242 и His193.
В другом аспекте изобретение связано с полинуклеотидной молекулой, состоящей из двух оперативно связанных областей кодирующих последовательностей, которые кодируют соответственно пре-пропептид и gla-домен витамин К-зависимого плазматического белка и лишенный gla-домена белок Фактора VII, при экспрессии упомянутый полинуклеотид кодирует молекулу Фактора VII, которая не активирует в достаточной степени плазматические Факторы Х или IX, но способна связывать фактор ткани. Молекула модифицированного Фактора VII, экспрессированная этим полинуклеотидом, является биологически активным антикоагулянтом, т. е. способна подавлять коагуляционный каскад и, следовательно, образование отложения фибрина или сгустка. С целью экспрессии модифицированного Фактора VII молекула полинуклеотида трансфицируется в линии клеток млекопитающих, такие как, например, ВНК, ВНК 570 или ВНК 293.
Краткое описание чертежей.
Фиг. 1 иллюстрирует конструкцию экспрессионного вектора с последовательностью ДНК для Фактора VII с модификацией Ser344, на Ala. Использованы символы 0-1, последовательность единицы картирования 0-1 аденовируса 5; Е, ген-энхансер (усилитель) вируса SV40; MLP, основной поздний промотор; SS, набор сайтов сплайсинга; и рА, сигнал полиаденилирования от SV40 в поздней ориентации.
Фиг. 2 показывает влияние струйного введения DEGR-Фактора VIIa на образование тромба (отложение тромбоцитов) на эндартерэктомизированной аорте бабуина по сравнению с обработанными физиологическим раствором контролями. Артерии измеряли через 60 минут. На данной модели острого сосудистого повреждения примата DEGR-Фактор VIIa заметно подавлял развитие обогащенных тромбоцитами тромбов.
Фиг.3 показывает результаты, полученные при инкубации клеток гладкой мышцы бабуина с возрастающими концентрациями или FVIIa (незакрашенные квадраты), или DEGR-FVIIa в присутствии постоянного количества FVIIa (5 нМ) (закрашенные квадраты). Уровень активации FX был затем определен с использованием хромогенного субстрата S-2222. Данные представлены в виде амидолитической активности, а именно как процент от активности, генерированной в присутствии только 5 нМ FVIIa.
Фиг.4 изображает размер площади внутренней оболочки после эндартерэктомии сонной артерии бабуина и обработки с DEGR-Фактором VIIa в течение 7 или 30 дней по сравнению с контрольными животными.
Фиг. 5 показывает отношение площади внутренней оболочки к сумме площадей внутренней и медиальной оболочки бедренной артерии бабуина после баллонного повреждения и обработки DEGR-Фактором VIIa, контрольная группа включала 5 сосудов, при 7-дневной обработке обследованы 11 сосудов, при 30-дневной обработке обследованы 2 сосуда (n - число обследованных сосудов).
Описание специфических воплощений.
Новый модифицированный Фактор VII, имеющий антикоагулянтную активность, обеспечивается настоящим изобретением. Модифицированный Фактор VII может быть в форме зимогена (т.е. одноцепочечной молекулы) или может быть расщеплен в активационном сайте. Таким образом, понятие "модифицированный Фактор VII" включает в себя модифицированный Фактор VII и молекулы модифицированного Фактора VIIa, которые связывают фактор ткани и подавляют активацию Фактора IX в IXa и Фактора Х в Ха. Составы модифицированного Фактора VII пригодны для введения различным млекопитающим, особенно человеку, с целью ингибирования коагуляционного каскада. Модифицированный Фактор VII может быть введен пациенту в сочетании с другими антикоагулянтами или вместо них.
Фактор VII играет важную роль в коагуляционном каскаде, особенно важным является включение его во внешний метаболический путь. Присутствуя в циркулирующей плазме в виде неактивного одноцепочечного проферментного белка и подвергнувшись активации, Фактор VIIa в сочетании с фактором ткани и ионами кальция активирует Фактор Х в Ха и Фактор IX в IХа с возможным образованием фибринового сгустка.
Натсоящее изобретение обеспечивает возможность подавлять эту последовательность событий в коагуляционном каскаде, предотвращая или иным образом ингибируя активацию Факторов Х и IX с помощью Фактора VIIa. Белок Фактора VII этого изобретения имеет каталитический центр, который модифицирован таким образом, чтобы снизить каталитическую активность Фактора VIIa, но сохранить способность связывать фактор ткани. Молекула модифицированного Фактора VII конкурирует с нативным Фактором VII и/или VIIa за связвание с фактором ткани. В результате подавляется активация Факторов Х и IX,
В одном аспекте настоящего изобретения каталитическая активность Фактора VIIa подавляется химической дериватизацией каталитического центра, или триады. Дериватизация может быть осуществлена взаимодействием Фактора VII с необратимым ингибитором, таким как фосфорорганическое соединение, сульфонилхлорид, пептидсодержащий галометилкетон или азапептид, или, например, ацилированием. Предпочтительные пептидсодержащие галометилкетоны включают РРАСК (D-Phe-Pro-Arg-хлорметилкетон; см. US Patent No 4318904, включенный здесь в виде ссылки), D-Phe-Phe-Arg- и Phe-Phe-Arg-хлорметилкетон; и DEGRck (Dansyl-Glu-Gly-Arg-хлорметилкетон).
В другом аспекте каталитическая активность Фактора VIIa может быть также подавлена замещением, введением или удалением аминокислот. В представленных воплощениях замены аминокислот производятся в аминокислотной последовательности каталитической триады Фактора VII, определяемой здесь как область, содержащая аминокислоты, которые способствуют активности каталитического сайта Фактора VIIa. Замещения, введения или удаления в каталитической триаде обычно производят по аминокислоте, которая входит в каталитически и сайт или находится по соседству с ней. В белках человеческого и бычьего Фактора VII аминокислотами, образующими каталитическую "триаду", являются Ser344, Asp242 и His193 (нижний цифровой индекс указывает положение в аминокислотной последовательности). Каталитические сайты Фактора VII других видов млекопитающих могут быть определены с использованием доступной техники, включая помимо других выделение белка и анализ аминокислотной последовательности. Каталитические сайты могут быть также определены удлинением последовательности в присутствии других серинпротеаз, особенно химотрипсина, активный центр которого расшифрован ранее (Sigler et al., J. Mol. Biol. 35:143-164 (1968), включено здесь в виде ссылки), устанавливая с помощью упомянутого удлинения аналогичные аминокислотные остатки активного центра.
Замещения, введения или удаления аминокислот производятся таким образом, чтобы предотвратить или иным способом подавить активацию Факторов Х и/или Х Фактором VIIa. Модифицированный таким способом Фактор VII должен, однако, сохранять способность конкурировать с аутентичным Фактором VII и/или Фактором VIIa за связывание с фактором ткани в коагуляционном каскаде. Такая конкуренция легко определяется посредством, например, анализа свертываемости крови, как здесь описано, или анализом конкурентного связывания с использованием, например, клеточной линии, имеющей поверхностно-связанный фактор ткани, такой как клетки человеческой карциномы мочевого пузыря линии J82 (Sakai et al., J. Biol. Chem. 264:9980-9988 (1989), включено здесь в виде ссылки).
Аминокислоты, которые образуют каталитический сайт Фактора VII, такие как Ser344, Asp242 и His193, человеческого и бычьего Фактора VII, могут быть или заменены, или удалены. В рамках настоящего изобретения предпочтительнее изменить только одну аминокислоту, сводя таким образом к минимуму вероятность усиления антигенности молекулы или подавления ее способности связывать фактор ткани, однако могут быть сделаны изменения двух или более аминокислот (замены, добавления или удаления), а также могут проводиться сочетания замены(н), добавления(ий) и удаления(ий). В избранном воплощении для человеческого и бычьего Фактора VII предпочтительнее замена Ser344 на Ala, но могут быть заменены Gly, Met, Thr или другие аминокислоты. Предпочтительно заменить Asp на Glu и His на Lys или Arg. В общем замена подбирается таким образом, чтобы как можно меньше разрушать третичную структуру белка. Модель Дайхоффа и др. (в Атласе структуры белка 1978. Natl. Biomed. Res. Found., Вашингтон, Колумбия), включенная здесь в виде ссылки, может быть использована в качестве руководства по выбору других аминокислотных заместителей. Можно внести изменение, как описано выше, в каталитический сайт соответствующей последовательности Фактора VII человека, быка или другого вида и тестировать полученный белок на подавление каталитической активности и возникающую антикоагулянтную активность, как здесь описано. Для модифицированного Фактора VII каталитическая активность будет существенно снижена, обычно она составляет менее 5% от каталитической активности соответствующих образцов Фактора VII дикого типа, преимущественно менее 1%.
Белки настоящего изобретения могут быть получены с использованием техники рекомбинантной ДНК. В общем клонированная последовательность ДНК Фактора VII дикого типа модифицируется, чтобы кодировать желаемый белок. Эта модифицированная последовательность затем встраивается в экспрессионный вектор, который в свою очередь трансформируется или вносится в клетки хозяина. Клетки высших эукариот, особенно культивированные клетки млекопитающих, являются предпочтительными в качестве клеток хозяина. Для человеческого Фактора VII известны полные нуклеотидная и аминокислотная последовательности. См. US Patent No 4784950 (который включен здесь в виде ссылки), где описаны клонирование и экспрессия человеческого рекомбинантного Фактора VII. Последовательность бычьего Фактора VII описана в работе Takeya et al., J. Biol. Chem. 263:14868-14872 (1988), которая включена в виде ссылки.
Изменения аминокислотной последовательности могут быть осуществлены различными методами. Для модификации последовательности ДНК используют сайт-специфичный мутагенез. Методы сайт-специфичного мутагенеза хорошо известны и описаны, например, Золлером и Смитом (ДНК 3:479-488 (1984)). Таким образом, используя нуклеотидные и аминокислотные последовательности Фактора VII, можно осуществить изменения по выбору.
Фактор VII, модифицированный в соответствии с настоящим изобретением, включает в себя те белки, у которых амино-концевая область (gla-домен) заменена gla-доменом одного из витамин К-зависимых плазматических белков Фактора IX, Фактора X, протромбина, белка С, белка S или белка Z. gla-Домены витамин К-зависимых плазматических белков характеризуются наличием остатков гамма-карбоксиглутаминовой кислоты и обычно имеют протяженность от примерно 30 до 40 аминокислот с С-концами, подобными положениям на границах экзон-интрон соответствующих генов. Методы получения Фактора VII с гетерологичным gla-доменом раскрыты в US Patent No 4784950, включенном здесь в виде ссылки.
Последовательности ДНК для использования в рамках настоящего изобретения будут типично кодировать препропептид на аминоконце белка Фактора VII для установления надлежащего посттрансляционного процессинга (например, гамма-карбоксилирование остатков глутаминовой кислоты) и секреции из клетки-хозяина. Препропептид может быть таким, как в Факторе VII, или другим витамин К-зависимым плазматическим белком, таким, как Фактор IX, Фактор X, протромбин, белок С или белок S. Как признают специалисты, могут быть осуществлены дополнительные модификации аминокислотной последовательности модифицированного Фактора VII, которые не ослабляют существенно способность белка действовать в качестве антикоагулянта. Например, Фактор VII, модифицированный по каталитической триаде, может быть также модифицирован и по сайту активационного расщепления с целью ингибировать превращение зимогенного Фактора VII в его активированную двухцепочечную форму, как описано в US Patent 5288629, включенном здесь в виде ссылки.
Экспрессионные векторы для использования в осуществлении настоящего изобретения будут включать в себя промотор, способный направлять транскрипцию клонированного гена или кДНК. Предпочтительными промоторами для использования в культуре клеток млекопитающих являются вирусные и клеточные промоторы. Вирусные включают промотор от SV40 (Subramani et al., Mol. Cell Bio). 1:854-864 (1981)) и промотор от CMV (цитомегаловируса) (Boshart et al. , Cell 41:521-530 (1985)). Особо предпочтительным вирусным промотором является основной поздний промотор аденовируса 2 (Kaufman and Sharp. Mol. Cell Biol. 2: 1304-1319 (1982)). Клеточные промоторы включают мышиный каппа-промотор (Bergman et al. , Proc. Natl. Acad. Scl. USA 81:7041-7045 (1983)) и мышиный VH промотор (Loh et al., Cell 33:85-93 (1983)). Особо предпочтительным клеточным промотором является мышиный металлотионеиновый промотор I (Palmiter et al., Science 222:809-814 (1983)). Экспрессионные векторы могут также содержать набор сайтов сплайсинга РНК, расположенных по ходу транскрипции от промотора и против хода транскрипции от сайта включения для последовательности Фактора VII. Предпочтительные сайты сплайсинга РНК могут быть получены из аденовирусов и/или генов иммуноглобуллина. В экспрессионном векторе содержится также сигнал полиаденилирования, расположенный по ходу транскрипции от сайта включения. Особо предпочтительные сигналы полиаденилирования включают ранний или поздний сигнал от SV40 (Kaufman and Sharp, там же), сигнал полиаденилирования из области Е1Ь аденовируса 5, терминатор гена человеческого гормона роста (De Noto er al. Nucl. Acids Res. 9:3719-3730 (1981)), сигнал полиаденилирования гена человеческого Фактора VII или гена бычьего Фактора VII. Экспрессионные векторы могут также включать некодирующую вирусную лидерную последовательность, такую как у аденовируса 2, расположенную между промотором и сайтами сплайсинга РНК; энхансерные последовательности, такие как энхансер от SV40.
Клонированные последовательности ДНК вводятся в культуру клеток млекопитающих, например, опосредованной фосфатом кальция трансфекцией (Wigler et al. , Cell 14:725-732 (1978); Corsaro and Pearson, Somatic Cell Genetics 7: 603-616, 1981; Graham and Van der Eb, Virology 52d:456-467 (1973)) или электропорацией (Neumann et al., EMBO J. 1:841-845 (1982)). Для идентификации и отбора клеток, экспрессирующих экзогенную ДНК, в клетки наряду с основным геном или кДНК обычно вводится ген, который определяет селектируемый фенотип (селектируемый маркер). Предпочтительные селектируемые маркеры включают гены, которые определяют устойчивость к таким препаратам, как неомицин, гигромицин и метотрексат. Селектируемый маркер может быть амплифицируемым селектируемым маркером. Предпочтительным амплифицируемым селективным маркером является последовательность дигидрофолатредуктазы (DHFR). Селектируемые маркеры рассмотрены Тилли (Технология клетки млекопитающих. Butterworth Publishers, Стонхем, Массачусетс; включено здесь в виде ссылки). Выбор селектируемого маркера доступен для обычного специалиста.
Селектируемые маркеры могут быть введены в клетку в отдельной плазмиде в одно и то же время с основным геном или могут быть введены в одной с ним плазмиде. При расположении в одной и той же плазмиде селектируемый маркер и основной ген могут контролироваться различными промоторами или одним и тем же промотором, последняя схема дает дицистронную единицу генетического кода. Конструкции этого типа известны специалистам (например, Levinson and Simonsen, US Patent 4713339). Полезным может быть также прибавление дополнительной ДНК, известной как "носитель ДНК", к смеси, которая вводится в клетку.
После введения ДНК в клетки они растут в подходящей ростовой среде, обычно Т-2 дня, до начала экспрессии целевого гена. Использованный здесь термин "подходящая ростовая среда" обозначает среду, содержащую питательные и др. компоненты, необходимые для роста клеток и экспрессии гена модифицированного Фактора VII. Среда обычно включает источники углерода, азота, незаменимые аминокислоты, углеводы, витамины, соли, фосфолипиды, белок и факторы роста. Для продукции гамма-карбоксилированного модифицированного Фактора VII среда будет содержать витамин К, желательно в концентрации 0,1-5 мкг/мл. Затем проводится подбор препаратов для селекции клеток, которые устойчиво экспрессируют селектируемый маркер. Для клеток, которые были трансфицированы амплифицируемым селектируемым маркером, концентрация препарата может быть повышена, чтобы отобрать большее число копий клонированных последовательностей, тем самым повышая уровни экспрессии. Затем клоны устойчивых трансфицированных клеток скринируются на экспрессию модифицированного Фактора VII.
Предпочтительные линии клеток млекопитающих для использования в настоящем изобретении включают клетки COS-1 (ATCC CRL 1650), клетки почек новорожденного хомяка (ВНК) и 293 (ATCC CRL 1573; Graham et al., J. Clin. Virol. 36: 59-72 (1977)). Предпочтительной линией клеток ВНК является штамм tk-ts13 (Waechter and Baserge, Proc. Natl. Acad. Sci. USA 79:1106-1110 (1982), включено здесь в виде ссылки), в дальнейшем отнесенный к ВНК 570. Линия клеток ВНК 570 зарегистрирована в Американской коллекции типов культур (ATCC), 12301 Parklawn Dr., Роквилль, Мэриленд 20852, под номером CRL 10314. Линия клеток ВНК tk-ts13 также известна в ATCC под номером CRL 1632. Кроме того, в рамках настоящего изобретения могут быть использованы и другие клеточные линии, включая клетки Rat Hep I (гепатома крысы; ATCC CRL 1600), клетки Rat Hep II (гепатома крысы; ATCC CRL 1548), клетки ТСМК (ATCC CCL 139), клетки легких человека (ATCC НВ 8065), клетки NCTC 1469 (ATCC CCL 9.1), клетки СНО (АТСС CCL 61) и клетки DUKX (Urtaub and Chasin, Proc. Natl. Soc. Sci. USA 77:4216-4220 (1980).
В рамках настоящего изобретения для получения модифицированного Фактора VII может быть использована технология трансгенных животных. Предпочтительнее продуцировать белки в молочных железах самок млекопитающих. Экспрессия в молочной железе и последующая секреция целевого белка в молоко преодолевают многие трудности, связанные с выделением белков из других источников. Молоко легко собирается, доступно в больших количествах и хорошо охарактеризовано биохимически. Более того, основные белки молока присутствуют в нем в высоких концентрациях (обычно 1-15 г/л). С коммерческой точки зрения очевидно, что предпочтительнее использовать в качестве хозяина виды, которые дают большое количество молока. В то время как могут использоваться мелкие животные, такие как мыши и крысы (которые являются предпочтительными при подтверждении принципиального этапа), в рамках настоящего изобретения предпочтительнее использовать домашних млекопитающих, включая (но не только) свиней, коз, овец и крупный рогатый скот. Овцы являются особо предпочтительными благодаря таким факторам, как предшествующая история трансгенеза у этого вида, выход молока, стоимость и высокая доступность оборудования для сбора овечьего молока. Для сравнения факторов, влияющих на выбор вида хозяина, см. WIPO Publication WO 88/00239. Обычно желательно выбирать ту породу животного-хозяина, которая разводится для получения молока, такую как овца породы East Friesland, или интродуцировать молочную породу размножением трансгенной линии на более позднем этапе. В любом случае должно использоваться известное животное и в хорошем состоянии.
Для получения экспрессии в молочной железе используют промотор гена молочного белка. Группу генов молочного белка составляют гены, кодирующие казеин (см. US Patent No 5304489, включенный здесь в виде ссылки), бета-лактоглобулин, альфа- лактоглобулин и кислый белок сыворотки. Предпочтительным является промотор бета-лактоглобулина. В случае гена овечьего бета-лактоглобулина, как правило, будет использована проксимальная 5'-фланкирующая последовательность из 406 нуклеотидов, хотя предпочтительными являются более значительные фрагменты 5'-фланкирующей последовательности, примерно до 5 тыс. нуклеотидов, такие как сегмент ДНК в 4,25 тыс. нуклеотидов, окружающий 5'-фланкирующий промотор и некодирующую часть гена бета-лактоглобулина. См. Whitelaw et al. , Biochem. J. 286:31-39 (1992). Применимы также и подобные фрагменты промоторной ДНК других видов.
Другие области бета-глобулинового гена могут также включаться в конструкцию, т.к. могут экспрессироваться геномные области гена. Принято считать, что, например, конструкции, лишенные интронов, экспрессируют плохо по сравнению с теми, которые содержат такие последовательности ДНК (см. Brinster et al. , Proc. Natl. Acad. Sci. USA 85:836-840 (1988); Palmiter et al., Proc, Natl. Acad. Sci. USA 88:478-482 (1991); Whitelaw et al., Transgenic Res. 1: 3-13 (1991); WO 89/01343; и WO 91/02318, каждый включен здесь в виде ссылки). В этом отношении обычно предпочтительно использовать, где возможно, геномные последовательности, содержащие все или некоторые нативные интроны гена, кодирующего целевой белок или полипептид, следовательно, дополнительное включение, по крайней мере, некоторых интронов, например, из бета-лактоглобулинового гена, является предпочтительным. Одной из таких областей служит сегмент ДНК, который обеспечивает сплайсинг интрона и полиаденилирование РНК от 3'-некодирующего участка гена овечьего бета-лактоглобулина. При замене на природные 3'-некодирующие последовательности гена этот овечий бета-лактоглобулиновый сегмент может как усилить, так и стабилизировать уровни экспрессии целевого белка или протеина. В рамках других воплощений область, окружающая ATG инициации модифицированного Фактора VII, заменяется соответствующими последовательностями гена специфического белка молока. Такая замена обеспечивает предполагаемое ткане-специфическое инициационное окружение для того, чтобы усилить экспрессию. Удобно заменить полностью пре- про- и 5'-некодирующие последовательности модифицированного Фактора VII последовательностями, например, гена BLG, хотя можно заменять и меньшие области.
Для экспрессии модифицированного Фактора VII у трансгенных животных сегмент ДНК, кодирующий модифицированный Фактор VII, оперативно связывается с дополнительными сегментами ДНК, необходимыми для его экспрессии, чтобы образовать экспрессионные единицы. Такие дополнительные сегменты включают вышеупомянутый промотор, а также последовательности, которые обеспечивают терминацию транскрипции и полиаденилирование мРНК. Экспрессионные единицы затем будут включать сегмент ДНК, кодирующий секреторную сигнальную последовательность, оперативно связанную с сегментом, кодирующим модифицированный Фактор VII. Секреторная сигнальная последовательность может быть последовательностью нативного Фактора VII или какого-либо другого белка, такого как белок молока. См. , например, von Heinje, Nucl. Acids Res. 14:4683-4690 (1986); Meade et al., US Patent No 4873316, которые включены здесь в виде ссылок.
Конструирование экспрессионных единиц у трансгенных животных удобно осуществлять включением последовательности модифицированного Фактора VII в плазмиду или фаговый вектор, содержащий дополнительные сегменты ДНК, хотя экспрессионная единица может быть сконструирована с помощью других последовательностей лигирования. Особенно удобно создать вектор, содержащий сегмент ДНК, кодирующий белок молока, и заменить кодирующую последовательность молока последовательностью полипептида модифицированного Фактора VII, тем самым создавая смешение генов, которое включает экспрессионные контрольные последовательности гена белка молока. В любом случае клонирование экспрессионных единиц в плазмидах или других векторах благоприятствует амплификации последовательности модифицированного Фактора VII. Амплификацию удобно проводить в бактериальной клетке-хозяине (например, Е. coli), таким образом, векторы будут включать источник репликации и селектируемый маркер в хозяйские бактериальные клетки.
Экспрессионную единицу затем вводят в оплодотворенные яйцеклетки (включая эмбрионы на ранней стадии развития) выбранного вида-хозяина. Введение гетерологичной ДНК может быть выполнено одним из нескольких способов, включая микроинъекцию (например, US Patent No 4873191), ретровирусную инфекцию (Jaenisch, Science 240:1468-1474 (1988)) или направленную интеграцию с использованием стволовых клеток эмбриона (ES) (рассмотрено Bradley et al., Bio/Technology 10: 534-539 (1992)). Яйцеклеткам, имплантированным в фаллопиевы трубы или матки псевдобеременных самок, дают развиваться до срока. Потомок, несущий введенную ДНК в зачаточном состоянии, может передавать ДНК своему потомству нормальным способом, по Менделю, с образованием трансгенного стада.
Специалистам известны обычные процедуры для получения трансгенных животных известны. См., например, Ноаnп et al., Манипулирование мышиным зародышем: лабораторное руководство. Cold Spring Harbor Laboratory, 1986; Simons et al. , Bio/Technology 6:179- 183 (1988); Wall et al., Biol. Reprod. 32: 645-651 (1985); Buhler et al., Bio/Technology 8:140-143 (1990); Ebert et al. , Bio/Technology 9: 835-838 (1991); Krimpenfort et al., Bio/Technology 9: 844-847 (1991); Wall et al., J. Cell. Biochem. 49:113-120 (1992); US Patent No 4873191 и 4873316; WIPO - WO 88/00239, WO 90/05188, WO 92/11757; и GB 87/00458, которые включены здесь в виде ссылок. Техника введения последовательностей чужеродной ДНК млекопитающим и в эмбриональные клетки других видов изначально отрабатывалась на мышах. См., например, Gordon et al., Proc. Natl. Acad. Sci. USA 77:7380- 7384 (1980); Gordon and Ruddle, Science 214:1244-1246 (1981); Palmiter and Brinster, Cell 41:343-345 (1985); Binster et al., Proc. Natl. Acad. Sci. USA 82:4438-4442 (1985) и Hogan et al., (там же). Впоследствии эти приемы были адаптировангы для использования на более крупных животных, включая домашних животных (см., например, публикации WIPO - WO 88/00239, WO 90/05188 и WO 92/117577; и Simons et al., Biotechnology 6: 179-183 (1988). Итак, в большинстве известных до настоящего времени эффективных способов генерации трансгенных мышей или домашних животных несколько сот линейных молекул ДНК, представляющих интерес, вводят в одно из проядер оплодотворенной яйцеклетки в соответствии с разработанными приемами. Может быть использована также инъекция ДНК в цитоплазму зиготы. Можно также использовать получение трансгенных растений. Экспрессия бывает генерализованной или направленной на отдельный орган, такой как бугор. См. Hiatt, Nature 344: 469-479 (1990); Edelbaum et al. , J. Interferon Res. 12:449-453 (1992); Sijmons et al., Bio/Technology 8:217-221 (1990); и публикация Европейского патентного бюро ЕР 255378.
Модифицированный Фактор VII, полученный в соответствии с настоящим изобретением, может быть очищен аффинной хроматографией на колонке с антителом к анти-фактору VII. Особо предпочтительным является использование кальцийзависимых моноклональных антител, как описано у Wakabayashi et al., J. Biol. Chem. 261: 11097-11108 (1986) и Thim et al., Biochem. 27:7785-7793 (1988), включенных здесь в виде ссылок. Дополнительная очистка может быть осуществлена традиционными методами химической очистки, такими как высокоэффективная жидкостная хроматография. Известны и другие методы очистки, включая осаждение цитратом бария, которые могут быть использованы для очистки модифицированного Фактора VII, описанного здесь (см., как правило, Scopes R. , Очистка белков, Springer-Verlag, N.Y., 1982). Предпочтительным является достаточно чистый модифицированный Фактор VII, по меньшей мере, 90-95% гомогенности, самым предпочтительным - 98-99% гомогенности или более, используемый для фармацевтических целей. Очищенный частично или по желанию до гомогенного состояния, модифицированный Фактор VII может использоваться в терапевтических целях.
В рамках одного из воплощений изобретения модифицированный Фактор VII расщепляется в сайте активации для того, чтобы превратить его в двухцепочечную форму. Активация может осуществляться в соответствии с известными в этой области процедурами, такими как описанные Osterud et al., Biochemistry 11: 2853-2857 (1972); Thomas, US Patent No 4456591; Hedner and Kisiel, J. Clin. Invest. 71:1836-1841 (1983); Kisiel and Fujikawa, Behring Inst. Mitt. 73:29-42 (1983), которые включены здесь в виде ссылок. Образующаяся молекула затем преобразуется в лекарственную форму и вводится, как описано ниже.
Молекулы модифицированного Фактора VII настоящего изобретения и фармацевтические составы на их основе особенно удобны для введения людям при лечении различных состояний, включая внутрисосудистую коагуляцию. Например, хотя тромбоз глубоких вен и легочную эмболию можно лечить традиционными антикоагулянтами, модифицированный Фактор VII, описанный здесь, может быть использован для того, чтобы предотвратить возникновение тромбоэмболических осложнений у пациентов с установленным высоким риском, таких как перенесшие хирургическую операцию или страдающие застойной сердечной недостаточностью. Кроме того, модифицированный Фактор VII может действовать как антагонист опосредованной фактором ткани индукции коагуляции, тем самым блокируя продукцию тромбина и последующее отложение фибрина. Модифицированный Фактор VII как таковой может оказаться полезным в подавлении активности фактора ткани, приводя, в частности, к подавлению свертывания крови, тромбоза или отложения тромбоцитов.
Молекулы модифицированного Фактора VII настоящего изобретения могут быть особенно полезны в лечении гиперплазии внутренней оболочки или рестеноза вследствие острого сосудистого нарушения. Острыми сосудистыми нарушениями являются те, которые возникают быстро (т.е. от дней до месяцев) по сравнению с хроническими сосудистыми нарушениями (например, атеросклерозом), которые развиваются в течение всей жизни. Острые сосудистые нарушения часто возникают при хирургических процедурах, таких как реконструкция сосудов, при которых используются приемы ангиопластики, эндартерэктомии, атерэктомии, шунтирования и т.п. Гиперплазия может также возникать как отсроченная реакция в ответ, например, на шунтирование или трансплантацию органа. Поскольку модифицированный Фактор VII обладает более избирательным, чем гепарин действием, обычно связывая только фактор ткани в сайтах повреждения, и благодаря тому, что модифицированный Фактор VII не разрушает другие коагуляционные белки, он будет более эффективным, чем гепарин и с меньшей вероятностью вызовет нежелательные кровотечения при профилактическом применении для предотвращения тромбоза глубоких вен. Доза модифицированного Фактора VII для предотвращения тромбоза глубоких вен находится в пределах от примерно 50 мкг до 500 мг в день, более типично 1-200 мг/день, предпочтительно 10-175 мг/день для пациента весом 70 кг, введение должно начинаться, по крайней мере, за 6 часов до хирургического вмешательства и продолжаться до перехода пациента на амбулаторный режим. Доза модифицированного Фактора VII для лечения рестеноза будет различаться для каждого пациента, но в общем будет находиться в упомянутых выше пределах.
Последние достижения в лечении коронарных сосудистых заболеваний включают использование механического вмешательства для того, чтобы или удалить, или переместить виновную в нарушении бляшку с целью восстановления адекватного тока крови в коронарных артериях. Несмотря на использование разнообразных форм механического вмешательства, включая баллонную ангиопластику, редукционную атерэктомию, установку сосудистых стентов, лазерную терапию или иссечение распадающейся ткани, эффективность этих приемов ограничивается примерно 40% рестенозов, возникающих в течение 6 месяцев после лечения.
Считается, что рестеноз является результатом сложного взаимодействия биологических процессов, включая отложение тромбоцитов и тромбообразование, высвобождение хемотаксисных и митогенных факторов и миграцию и пролиферацию клеток сосудистой гладкой мышцы во внутреннюю оболочку расширенного артериального сегмента.
Подавление аккумуляции тромбоцитов в сайтах механического повреждения может лимитировать скорость рестеноза у человека. Терапевтическое использование моноклонального антитела к тромбоциту GpIIb/IIIa способно снизить уровень рестеноза у человека (Califf et al., N. Engl. J. Med. 330:956-961 (1994)). Антитело может связываться с рецептором GpIIb/IIIa на поверхности тромбоцитов и тем самым подавлять аккумуляцию тромбоцитов. Эти данные предполагают, что подавление аккумуляции тромбоцитов в сайте механического повреждения коронарной артерии человека оказывает положительное воздействие на окончательный результат лечения. Поскольку в сайтах острых сосудистых нарушений происходит аккумуляция тромбоцитов, то генерация тромбина в этих сайтах может быть ответственна за активацию тромбоцитов и их оследующее накопление.
Как показано в последующих примерах, модифицированный Фактор VII настоящего изобретения способен связывать находящийся на поверхности клетки фактор ткани. Например, DEGR- Фактор VIIa связывает поверхностный фактор ткани с одинаковым или более высоким сродством, чем Фактор VIIa дикого типа. Однако DEGR-Фактор VIIa не имеет ферментной активности, но связывается с фактором ткани и действует как конкурентный антагонист Фактора VIIa дикого типа, тем самым ингибируя последующие этапы внешнего метаболического пути коагуляции, приводящие к генерации тромбина.
Молекулы модифицированного Фактора VII настоящего изобретения, поддерживающие связывание фактора ткани, подавляют аккумуляцию тромбоцитов в сайте сосудистого нарушения путем блокирования продукции тромбина и последующего отложения фибрина.
Благодаря способности DEGR-Фактора VII блокировать генерацию тромбина и ограничивать отложение тромбоцитов в сайте острого сосудистого нарушения, молекулы модифицированного Фактора VII, которые обладают активностью связывания фактора ткани, но лишены ферментной активности, могут использоваться для подавления сосудистого рестеноза.
Таким образом, составы и методы настоящего изобретения имеют широкую область применения. Например, они полезны для предотвращения или подавления рестеноза, сопровождающего типично механическое вмешательство, проведенное с целью либо удаления, либо замены виновного в нарушении материала бляшки при лечении повреждений коронарных или перифирических сосудов, таких, которые сопутствуют и/или следуют за баллонной ангиопластикой, редукционной атерэктомией, установкой сосудистых стентов, лазерной терапией, иссечением распадающихся тканей и т.п.
Соединения будут типично вводиться за 24 часа до вмешательства и в течение 7 дней и более после него. Введение может проводиться разнообразными способами, как описано в дальнейшем. Соединения настоящего изобретения могут также вводиться системно и местно при установке сосудистых имплантатов (например, покрытием искусственных или модифицированных природных имплантатов артериальных сосудов) в сайтах анастомоза, хирургической эндартерэктомии (типично для эндартерэктомии сонной артерии), шунтирования и т.п. Модифицированные Факторы VII и VIIa находят также применение для подавления гиперплазии внутренней соудистой оболочки, раннего склероза и венозной окклюзии, связанной с трансплантацией органов, например, возникающей после трансплантации костного мозга. При лечении установленного тромбоза глубоких вен и/или легочной эмболии ударная и поддерживающая дозы модифицированного Фактора VII в зависмости от веса пациента и тяжести заболевания изменяются в пределах от примерно 50 мкг до 500 мг/день, более типично 1-200 мг/день и более предпочтительно 10-175 мг/день для пациента весом 70 кг. Благодаря сниженной вероятности нежелательных кровотечений при инфузии Фактора VII можно заменить им гепарин или снизить дозу гепарина в ходе или после хирургического вмешательства в сочетании с тромбэктомией или эмболэктомией.
Составы модифицированного Фактора VII настоящего изобретения очень полезны для предотвращения кардиогенной эмболии и в лечении тромботических ударов. Благодаря малой вероятности нежелательных кровотечений и избирательности действия модифицированный Фактор VII можно давать перенесшим удар и он может предотвратить распространение закупоривающих артериальных тромбов. Количество введенного модифицированного Фактора VII будет меняться для каждого пациента в зависимости от характера и тяжести удара, но дозы в основном будут находиться в пределах, предложенных ниже.
Фармацевтические составы модифицированного Фактора VII, включающие DEGR-Фактор VII и представленные здесь, окажутся полезными в лечении острого инфаркта миокарда благодаря способности модифицированного Фактора VII подавлять коагуляцию in vivo. Модифицированный Фактор VII может быть дан в качестве адъюванта с активатором тканевого плазминогена или стрептокиназой в период острой фазы инфаркта миокарда и может ускорить tPA-индуцированный тромболиз. При остром инфаркте миокарда пациенту дается ударная доза, по крайней мере, от 50 мкг до 500 мг/день, более типично 1-200 мг/день и более предпочтительно 10-175 мг/день для больного весом 70 кг в качестве ударной и поддерживающей дозы. Модифицированный Фактор VII дается до, в сочетании или вскоре после введения тромболитического агента, такого как активатор тканевого плазминогена.
Модифицированный Фактор VII настоящего изобретения полезен в лечении коагулопатии потребления (DIC) и других проявлений, связанных с грам-отрицательной бактеремией. Для пациентов с DIC характерны широко распространенные микроциркуляторные тромбы и частые острые кровотечения, которые возникают от истощения необходимых факторов свертывания. Благодаря своей избирательности действия модифицированный Фактор VII не будет, как традиционные коагулянты, осложнять кровотечения, связанные с DIC, но задержит или подавит образование дополнительных микроваскулярных фибриновых отложений. Таким образом, модифицированный Фактор VII данного изобретения, включая DEGR-Фактор VII и DEGR-Фактор VIIa, полезен для подавления отложений фибрина, связанных с эндотоксемией и эндотоксиновым шоком и, следовательно, смягчает эффекты, вызываемые грам-отрицательной бактеремией. Показано, что DEGR-Фактор VIIa у мышей проявляет дозозависимый эффект при блокировании отложений фибрина в почках и легких, а у бабуинов увеличивает выживаемость леченых животных. В случае острой бактеремии, эндотоксемии или DIC пациенту дается ударная доза, по крайней мере, от примерно 50 мкг до 500 мг/день, более типично 1-200 мг/день и более предпочтительно 10-175 мг/день при весе пациента 70 кг, после этого поддерживающие дозы в пределах от 50 мкг до 500 мг/день, типично 1-200 мг/день при весе пациента 70 кг.
Фармацевтические составы предназначены для парентерального, местного или локального введения при профилактической и/или терапевтической обработке. Предпочтительнее фармацевтические составы вводить парентерально, т.е. внутривенно, а также подкожно или внутримышечно. Таким образом, это изобретение обеспечивает составы для парентерального введения, которые включают раствор молекул модифицированного Фактора VII в приемлемом носителе, предпочтительно в водном. Могут использоваться разнообразные водные носители, например вода, забуференная вода, 0,4% физиологический раствор, 0,3% глицин и т.п. Молекулы модифицированного Фактора VII также могут быть заключены в липосому в качестве лекарственной формы для доставки в (или нацеливания на) сайты повреждения. Липосомные препараты, в общем, описанные, например, в US 4837028, US 4501728 и US 4975282, включенных здесь в виде ссылок. Составы могут стерилизоваться стандартными хорошо известными приемами. Получаемые в результате водные растворы могут переноситься в упаковку для использования или фильтроваться в асептических условиях и лиофилизоваться, причем лиофилизованные препараты перед введением смешиваются со стерильным водным раствором. Составы могут содержать фармацевтически приемлемые вспомогательные вещества для приближения к физиологическим условиям, такие как поддерживающие рН и буферные агенты, поддерживающие тонус агенты и т.п., например ацетат натрия, лактат натрия, хлористый натрий, хлористый калий, хлористый кальций и т.д. Концентрация модифицированного Фактора VII в этих лекарственных формах может широко меняться, а именно от менее чем 0,5%, обычно 1% или, по крайней мере, около того, до 15-20% по весу и будет выбираться прежде всего по объему жидкости, вязкости и т. д. в соответствии с особенностями выбранного способа введения.
Таким образом, типичный фармацевтический состав для внутривенного вливания мог бы содержать 250 мл стерильного раствора Рингера и 10 мг модифицированного Фактора VII. Применяемые методы приготовления вводимых парентерально препаратов известны профессионалам и более детально описаны, например, в Ремингтонской фармацевтической науке, 16-ое издание, Mack Publishing Company, Истон, Пенсильвания (1982), которая включена здесь в виде ссылки.
Составы, содержащие модифицированный Фактор VII, могут вводиться при профилактической и/или терапевтической обработке. При терапевтическом применении составы вводятся больному, как описано выше, в количестве, достаточном для лечения или, по крайней мере, для того, чтобы остановить болезнь и ее осложнения. Количество, адекватное достижению этого, определяется как "терапевтически эффективная доза". В этом случае эффективное количество зависит от тяжести заболевания или нарушения, а также от веса и общего состояния пациента, но обычно изменяется в пределах 0,05-500 мг модифицированного Фактора VII в день для пациента весом 70 кг, причем чаще используются дозы 1-200 мг модифицированного Фактора VII в день. Следует помнить, что материалы настоящего изобретения могут, как правило, использоваться при серьезных заболеваниях или нарушениях, т.е. в угрожающих жизни или потенциально угрожающих ситуациях. В таких случаях с учетом минимизации посторонних веществ и общим отсутствием иммуногенности модифицированного Фактора VII у человека возможно и может считаться желательным введение пациенту врачом значительного избытка составов модифицированного Фактора VII.
При профилактическом применении составы, содержащие модифицированный Фактор VII, вводятся пациенту, восприимчивому к заболеванию, или при опасности заболевания или нарушения для того, чтобы усилить у пациента собственные антикоагулятивные возможности. Такое количество определяется как "профилактически эффективная доза". При таком использовании точные количества снова зависят от состояния здоровья пациента и его веса, но, как правило, изменяются в пределах 0,5-500 мг при весе пациента 70 кг, более часто 1-200 мг/70 кг.
Можно проводить однократное или многократные введения составов с дозами и режимом, выбранным лечащим врачом. Для амбулаторных пациентов, которым необходимы ежедневные поддерживающие дозы, модифицированный Фактор VII может быть введен, например, непрерывной инфузией с использованием портативного насоса.
Местная доставка модифицированного Фактора VII может быть осуществлена, например, путем перфузии, с помощью двойных баллонных катетеров, стентов, гидрогелей, используемых для покрытия баллонных катетеров, или другими хорошо известными методами. В любом случае фармацевтические лекарственные формы должны обеспечить количество модифицированного Фактора VII данного изобретения, достаточное для эффективного лечения пациента.
Пример I. Экспрессия Фактора VII с мутацией Ser344 -> А1а344.
Для того чтобы генерировать Фактор VII с мутацией Ser344 -> А1а344 в активном центре плазмиду FVII(565+2463)/pDX (US Patent No 4784950, включенный здесь в виде ссылки; плазмида зарегистрирована в Американской коллекции типов культур под номером 40205) расщепляли с Хbа I и Крn I с образованием фрагмента в 0.6 тысяч нуклеотидов, включающего кодирующую серин-344 область. Этот фрагмент клонировали в Xba I, Kpn I-расщепленный М13mр19, как показано на фиг. Эта процедура и последующие этапы, описанные ниже, обычно осуществляются в соответствии со стандартными протоколами (как описано, например, Maniatis et al. , Молекулярное клонирование. Лабораторное руководство, Cold Spring Harbor Laboratory Press, Колд-Спринг-Харбор, Нью-Йорк (1982), включено здесь в виде ссылки).
Мутагенез проводили на матрице М13 в соответствии с методами Золлера и Смита, см. выше, используя мутагенный олигонуклеотид ZC1656 (5' TGG GCC ТСС GGC GTC ССС СТТ 3') и "универсальный" второй праймер ZC87 (5" ТСС CAG ТСА CGA CGT 3'). Продукты реакции скринировали с использованием обработанного киназой ZC1656. Позитивные бляшки собирали, получали матричную ДНК и секвенировали от Pst I-сайта при 1077 до Kpn I-сайта при 1213. Анализ последовательности подтвердил наличие желаемой мутации. Мутантный клон обозначили 1656.
Затем с использованием клона 1656 был сконструирован экспрессионный вектор. Последовательность с внесенной мутацией выделяли из вектора М13 в виде фрагмента Pst I-Kpn I с 0,14 тыс. нуклеотидов. Этот фрагмент лигировали к фрагменту Hind III-Xba I с 1,7 тыс. нуклеотидов из FVII (565+2463)/pDX, к фрагменту Xba I-Pst I c 0,5 тыс. нуклеотидов из FVII (565+2463)/pDX и фрагменту Kpn I-Hind III с 4,3 тыс. нуклеотидов из FVII (565+2463)/pDX, как показано на фиг. Присутствие желаемой мутантной последовательности подтверждали расщеплением мутантного клона и клона дикого типа с Pst I, вставкой мутантного Фактора VII в М13 с Kpn b Xba I, получением саузерн-блотов расщепленной ДНК и зондированием блотов с радиоактивномеченным ZC1656.
Клетки почек новорожденного хомяка линии ВНК 570 (зарегистрированные в Американской коллекции типов культур клеток под номером 10314) трансфицировали двумя изолятами (обозначенными #544 и #545) экспрессионного вектора 1656. Клетки получали разбавлением 1:10 клеток ВНК 570 в пяти 10 см-планшетах с неселективной средой (модифицированной средой Игла (DMFM), содержащей 10% эмбриональную бычью сыворотку и 1% смесь антибиотиков PSN (GIBCO Life Technologies, Гайтерсбург, Мэриленд). Через 24 часа, когда клетки достигли 20-30% слияния, их сотрансфицировали с одним изолятом экспрессионного вектора, кодирующего мутацию 1656, плазмидой р486 (включающей аденовирус 5 ori, энхансер от SV40, основной поздний промотор аденовируса 2, лидерную последовательность аденовируса 2, сайты 5'- и 3'-сплайсинга, DHFRr-кДНК и сигнал полиаденилирования от SV40 в pML-1 (Lusky and Botchan, Nature 293:79-81 (1981)) и 10 мкг ДНК-носителя (фрагментированной под действием ультразвука ДНК спермы лося), как показано в таблице 1. В пробирки на 15 мл вносили ДНК, затем добавляли 0,5 мл буфера HEPES-2X (25 г HTHTS, 40 г NaCl, 1,8 г KCl, 0,75 г дигидрата Na2HPO4, 5 г декстрозы, разбавленные до объема 2,5 л дистиллированной водой, с рН 6,95-7,0) и перемешивали. В каждой пробирке осаждали ДНК добавлением 0,5 мл 0,25 М раствора хлористого кальция при барботировании воздуха через раствор ДНК в буфере HEPES с помощью пастеровской пипетки. Пробирки встряхивали, инкубировали при комнатной температуре в течение 15 минут, снова встряхивали. Смесь, содержащую ДНК, прибавляли по каплям с помощью пипетки в планшеты с клетками. Планшеты встряхивали и инкубировали при 37oС в течение 4-6 часов. После инкубации в каждый планшет добавляли 2 мл 20% глицерина в смеси Tris-буфера и физиологического раствора (0,375 г KCl, 0,71 г Na2HPO4, 8,1 г NaCl, 3,0 г Tris-HCl, 0,5 г сахарозы, разбавленные до общего объема 1 л, с рН 7,9). Планшеты встряхивали и выдерживали 2 минуты при комнатной температуре. Затем из планшетов удаляли среду, заменяли ее 2 мл смеси Tris-буфера с физиологическим раствором. Планшеты выдерживали 2 минуты при комнатной температуре, затем смесь буфера и физиологического раствора заменяли 10 мл неселективной среды. Планшеты инкубировали при 37oС в течение двух дней.
После двухдневной инкубации клетки разбавляли селекционной средой (DMEM, содержащая 10% диализованную эмбриональную бычью сыворотку, 1% смесь антибиотиков PSN и 150 нМ метотрексат) и с разбавлением 1:100, 1:250 и 1:500 помещали в макси-планшеты. Планшеты инкубировали при 37oС в течение недели. Через неделю среду удаляли и заменяли селекционной средой, в планшетах контролировали образование колоний.
Через восемь дней после образования колоний произвольно отбирали двенадцать колоний из планшетов с разведением 1:500 трансфекции изолятов #544 и #545. Каждый клон помещали в ячейку 6-ячеечного планшета и выращивали в селекционной среде. Через 7 дней планшеты сливали и каждый клон помещали в десятисантиметровый планшет с селекционной средой. Клоны, описанные выше, и контрольные клетки, трансфицированные с целью экспрессии Фактора VII дикого типа, были метаболически помечены 358-метионинцистеиновой смесью для мечения белков (NEN Dupont Biotechnology Systems, Уилмингтон, Дэлавер). Клоны выращивали и подготавливали к эксперименту с импульсной меткой в селективной среде. Клетки смывали смесью физиологический раствор-фосфатный буфер (Sigma, Сент-Луис, Миссури) и в течение 4 часов подвергали импульсу от 35S-Cys-35S-Met в 20 мкКи/мл. Через 4 часа собирали супернатант и клетки. Клетки лизировали в основном, как описано Lenk and Penman (Cell 16:289-302 (1979)), и 400 мкл каждого лизата проверяли со стафилококком на безвредность (Sigma, Сент-Луис, Миссури).
Образцы метаболически меченных клеток подвергали радиоиммунопреципитации (RIP) инкубацией с 6 мкл поликлональной антисыворотки анти-Фактора VII в течение 4 часов. К каждому образцу добавляли 60 мкл промытого белка А стафилококка, и образцы встряхивали при 4oС в течение 1,5 часа. Образцы центрифугировали, супернатант отделяли. Осадки промывали дважды 0,7 М буфером RIPA (10 мМ Tris, pH 7,4, 1% дезоксихолевая кислота (Calbiochem Corp., Ла-Джолла, Калифорния), 1% Тритон Х-100, 0,1% SDS, 5 мМ EDTA, 0,7 M NaCl) и один раз 0,15 М буфером RIPA (10 мМ Tris, pH 7,4, 1% дезоксихолевая кислота (Calbiochem Corp. , Ла-Джолла, Калифорния), 1% Тритон Х-100, 0,1% SDS, 5 мМ EDTA, 0,15 М NaCl). К каждому образцу добавляли краситель в 100 мкл буфера SDSx1 (50 мМ Tris-HCl, pH 6,8, 100 мМ дитиотреит, 2% SDS, 0,1% бромфенол синий, 10% глицерин), образцы кипятили в течение 5 минут, после чего центрифугировали, чтобы удалить белок А. Прогоняли 50 мкл каждого образца в полиакриламидном геле. Результаты показали, что 9 из 10 клонов секретировали модифицированный Фактор VII.
Пример II. Антикоагулянтная активность модифицированного Фактора VII.
Способность белка модифицированного Фактора VII подавлять свертывание крови измеряли в одностадийном анализе с использованием Фактора VII дикого типа в качестве контроля. Рекомбинантные белки получали в основном так, как описано выше, из клеток, культивированных в среде, содержащей 5 мкг/мл витамина К. Различные количества модифицированного Фактора VII (из клона 544) или рекомбинантного Фактора VII дикого типа разбавляли 50 мМ Tris-буфером с pH 7,5, 0,1% BSA до объема 100 мкл. Смеси инкубировали со 100 мкл Фактор VII-дефицитной плазмы (George King Bio-Medical Inc., Оверленд-Парк, Канзас) и 200 мкл тромбопластина С (Dade, Майами, Флорида; содержит тромбопластин кроличьего мозга и 11,8 мМ Са++). Анализ свертывания проводили в автоматическом таймере коагуляции (MLA Electra 800, Medical Laboratory Automation Inc. , Плизантвилле, Нью-Йорк), время свертывания преобразовывали в единицу активности Фактора VII с использованием стандартной кривой, полученной для нормальной объединенной человеческой плазмы (с допущением, что она содержит одну единицу активности Фактора VII на мл; получена объединением обработанных цитратом сывороток от здоровых доноров) с разведением от 1:5 до 1: 640. По этому методу препараты модифицированного Фактора VII не обладали детектируемой коагулянтной активностью. Таблица 2 показывает результаты анализа, выраженные во времени свертывания, для контрольных (нетрансфицированных), кондиционных для клеток ВНК сред (+/- витамин К), Фактора VII дикого типа и двух изолятов клеток, экспрессирующих модифицированный Фактор VII. Активность Фактора VII представлена как уменьшение времени свертывания по сравнению с контрольными образцами.
Для того чтобы определить влияние модифицированного Фактора VII на субстраты фактора плазмы, препараты одифицированного Фактора VII и рекомбинантного Фактора VII дикого типа или нативного Фактора VII инкубировали либо с Фактором X, либо с Фактором IX, их активацию контролировали анализом свертывания или электрофорезом в полиакриламидном геле.
Пример III.
Способность модифицированного Фактора VII связывать Фактор ткани
Способность. модифицированного Фактора VII конкурировать с Фактором VII дикого типа за фактор ткани и подавлять его вертывающую активность была оценена одностадийным анализом свертывания в присутствии лимитирующего количества фактора ткани (тромбопластина).
Время свертывания определяли одностадийным анализом одобно тому, как описано в примере II. В смешанных экспериментах использовали лимитированное количество фактора ткани, постоянное количество Фактора VII дикого типа и увеличивающиеся количества генетического варианта Фактора VII. Подавление прокоагулянтной активности Фактора VII/VIIa проявилось бы как увеличение времени свертывания в опытах с нарастающими количествами генетического варианта Фактора VII.
Величину активности Фактора VII в тестируемых образцах вычисляли в процентах по стандартной кривой, полученной на основе измерений активности Фактора VII в нормальной объединенной плазме. Стандартную кривую активности Фактора VII получали с использованием серии разведений нормальной объединенной плазмы в фосфатном буфере (PBS) в пределах от 1:5 до 1:640. С этой целью было принято допущение о том, что нормальная плазма содержит приблизительно 500 нг/мл Фактора VII и это было принято за единицу активности. Смесь, состоящую из 100 мкл Фактор VII-дефицитной плазмы, 100 мкл разведенной плазмы и 200 мкл тромбопластина-С (Dade, Майами, Флорида), использовали для измерения времени свертывания с использованием автоматического таймера MLA Electra 800. Для получения стандартной кривой результаты представляли в графическом виде как процент активности (1:5=100% активности) против времени свертывания в секундах.
По условиям анализа было необходимо, чтобы в состав среды, содержащей дикий тип и вариант Фактора VII, входило не менее одного процента сыворотки. Разведения в PBS были сделаны таким образом, чтобы падение времени свертывания происходило в соответствии со стандартной кривой. Типичным было минимальное разведение 1: 2. Окончательный объем составил 100 мкл. В экспериментах использовали два различных варианта человеческого Фактора VII с замещением Ser344 -> Ala, обозначенные как клоны "#10" и "#6". Результаты, приведенные в таблице 3, показывают, что по мере увеличения количества варианта Фактора VII снижался процент активности Фактора VIIa.
Эксперименты показали, что Фактор VII с замещением Ser344 - > Ala конкурировал с нативным Фактором VII по дозозависимому профилю и ингибировал прокоагулянтную активность нативного Фактора VII/VIIa. Можно было заключить, что вариант человеческого Фактора VII с заменой Ser344 -> Ala конкурирует с нативным человеческим Фактором VIIa и, следовательно, подавляет активацию Фактора Х и/или IX в человеческой плазме.
Пример IV.
Реакция Фактора VII с PPAck.
Рекомбинантный Фактор VII был продуцирован в трансфицированных клетках почек новорожденного хомяка. Белок очищали и активировали, как описано Thim (Biochemistry 27:7785-7793 (1988)), Brinkons et al., (Proc. Natl. Acad. Sci. USA 86:1382-1386 (1989)) и Bjoern and Thim (Res. Disci. No 269, 564
(1986)), которые включены здесь в виде ссылок. Клеточную среду собирали, фильтровали и разбавляли, чтобы снизить концентрацию солей. Затем разбавленную среду фракционировали анионообменной хроматографией, используя для элюции буфер, содержащий хлористый кальций. Фракцию, содержащую Фактор VII, собирали и затем очищали иммунохроматографией с использованием моноклонального антитела к кальцийзависимому антифактору VII. Дополнительную очистку проводили, используя двукратно анионообменную хроматографию, в ходе которой Фактор VII элюировали соответственно растворами хлористого кальция и хлористого натрия. Фактор VIIa получали в последнем элюате.
Рекомбинантный Фактор VIIa (1 мкМ) в 50 мМ Tris-HCl, 100 мМ хлористом натрии и 5 мМ хлористом кальции, с рН 7,4 инкубировали с 20 мкМ PPAck (D-фенилаланил-пролил-аргинил-хлорметилкетон; Calbiochem, Ла-Джолла, Калифорния) в течение 5, 20 и 60 минут. Затем добавляли буфер, содержащий хромогенный субстрат S-2288 (п-нитроанилид D-изолейцил-L-пролил-1-аргинина; Kabi Vitrum AB, Молндал, Швеция) для того, чтобы получить 2,5-кратное разведение и конечную концентрацию 0,3 мМ для S-2288. Измеряли выделение п- нитроанилина и сопоставляли с результатом, полученным с необработанным Фактором VIIa в качестве контроля. Полученные данные указывают на то, что в изученных условиях Фактор VIIa инактивируется полностью через приблизительно 60 минут.
Пример V.
Генерирование DEGR-Фактора VIIa,
Рекомбинантный человеческий Фактор VIia был получен, как описано в примере IV. Рекомбинантный человеческий Фактор VIIa в 10 мМ глициновом буфере с рН 8,0, 10 мМ хлористом кальции и 50 мМ хлористом натрии разбавляли до концентрации 1,5 мг/мл. К Фактору VIIa прибавляли 10-кратный избыток Dansyl-L-Glu-Gly-Arg- хлорметилкетона (Calbiochem, Ла-Джолла, Калифорния 92037), который был растворен в дистиллированной воде. После двухчасовой инкубации при 37oС к смеси добавляли второй 10-кратный молярный избыток DEGRck и инкубировали еще 2 часа при 37oС. Затем к Фактору VIIa прибавляли третий 10-кратный молярный избыток DEGRck и инкубировали приблизительно 16 часов при 4oС. Для удаления свободного DEGRck образец DEGR-Фактора VII тщательно диализовали против Tris-забуференного физиологического раствора (0,05 М Tris-HCl, 0,1 М NaCl, рН 7,5) при 4oС. Введением свободного DEGRck в смесь, содержащую Фактор Ха, получена стандартная кривая для измерения количества свободного DEGRck в растворе по мере ингибирования хромогенной активности Фактора Ха. Анализ смеси с DEGR-Фактором VIIa показал, что после экстенсивного диализа отношение свободного DEGRck к DEGR-Фактору VIIa составило менее 0,5%, тем самым подтверждая, что ингибирование, наблюдаемое для DEGR-Фактора VIIa в различных тест-системах, приведенных ниже, не связано с присутствием свободного DEGRck.
Пример VI.
Генерация Фактора Ха на клетках крысиной гладкой мышцы. Клетки сосудистой гладкой мышцы были проанализированы на наличие поверхностного фактора измерением способности клеток стимулировать превращение Фактора Х в Фактор Ха с использованием специфичного для Фактора Ха хромогенного субстрата.
Клетки крысиной сосудистой гладкой мышцы (Clowes et al., J. Clin. Invest. 93: 644-561 (1994)) помещали в 96-ячеечные планшеты (American Scientific Products, Чикаго, Иллинойс), 8000 клеток на ячейку в ростовой среде (табл. 4).
Таблица 4.
500 мл модифицированной среды Игла (DMEM) (GIBCO-BRL, Гейтерсбург, Мэриленд) 10% эмбриональная телячья сыворотка (Hyclone, Логан, Юта) 1 мМ пируват натрия (Irvine, Санта-Ане, Калифорния) 0,29 мг/мл L-глутамина (Hazelton, Ленекса, Канзас) 1xPSN; (100•= 5 мг/мл пенициллина, 5 мг/мл стрептомицина, 10 мг/мл неомицина (GIBCO-BRL, Гейтерсбург, Мэриленд)
Через 48 часов инкубации при 37оС среду заменяли на сывороточную свободную среду (Таблица 5).
Таблица 5.
250 мл модифицированной среды Игла (DMEM)
250 мл среды Ham's F-12 (Fred Hutchinson Cancer Research Center, Сиэтл, Вашингтон)
1 мМ пируват натрия
29 мг/мл L-глутамина
20 мкМ трансферрин (JRH, Ленекса, Канзас)
5 мкМ инсулин (GIBCO-BRL)
16 нг селена (Aldrich, Мильвоки, Висконсин)
1 мг/мл бычьего сывороточного альбумина (Sigma, Сент-Луис, Миссури).
Клетки инкубировали 72 часа при 37oС. После инкубации к клеткам добавляли или 10 нг/мл PDGF-BB (человеческого рекомбинантного фактора роста тромбоцитов), или 10% эмбриональную телячью сыворотку для стимуляции экспрессии фактора ткани (Taubman et al., J. Clin. Invest. 91:547-552 (1993)). В параллельную серию клеток не добавляли ни PDGF, ни сыворотку для того, чтобы контролировать подлинную активность нестимулированных клеток. После 6-часовой инкубации к клеткам прибавляли рекомбинантный человеческий Фактор VIIa с окончательной концентрацией 10 нМ. Одна серия клеток не получала Фактор VIIa в качестве отрицательного контроля. Клетки инкубировали 2 часа при 37oС, промывали буфером HEPES (10 мМ HEPES, 137 мМ NaCl, 5 мМ CaCl2, 11 мМ глюкоза, 0,1% BSA). После промывания клетки инкубировали 5 минут с 50 мкл на ячейку 200 нМ очищенного Фактора Х плазмы человека в Tris-забуференном физиологическом растворе с 5 мМ CaCl2. В каждую ячейку добавляли 25 мкл 0,5 М EDTA и 25 мкл 800 мкМ раствора хромогенного субстрата S-2222 (Kabi Pharmacia, Франклин, Огайо). Планшеты инкубировали 40 минут при комнатной температуре, затем анализировали при 405 нм с использованием прибора THERMOMAX (Molecular Services, Менло-Парк, Калифорния).
Таблица 6 показывает увеличение поглощения для ячеек, обработанных Фактором VIIa, по сравнению с контрольными ячейками (без добавления Фактора VII). Увеличение поглощения является прямым измерением уровня Фактора Ха, генерированного в ячейках, и последующего расщепления им хромогенного субстрата, высвобождающего хромофор. Эти данные демонстрируют также, что уровень хромогенной активности в клетках, предварительно обработанных или PDGF-BB, или 10% эмбриональной телячьей сывороткой (FCS), был выше, чем в нестимулированных клетках.
Эти результаты отчетливо показывают, что на поверхности клеток крысиной сосудистой гладкой мышцы наблюдается Фактор VIIa-зависимая активация Фактора Х в Фактор Ха.
Пример VII.
Ингибирование поверхностноклеточной хромогенной активности DEGR-Фактором VIIa.
Клетки крысиной сосудиситой гладкой мышцы помещали в планшеты с 96 ячейками, как описано выше. Клетки культивировали в течение 72 часов в бессывороточной среде, как описано выше; затем обрабатывали 10% эмбриональной телячьей сывороткой в течение 6 часов для стимуляции экспрессии фактора ткани. После стимуляции в каждую ячейку добавляли только буфер (контроль), 10 нМ Фактор VIIa или 10 нМ Фактор VIIa+100 нМ DEGR-Фактор VIIa. Клетки инкубировали 2 часа при 37oС, затем промывали буфером HEPES. После промывания клетки инкубировали в течение 5 минут с 50 мкл на ячейку 200 нМ Фактора Х в Tris-забуференном физиологическом растворе с добавкой 5 мМ хлористого кальция. В каждую ячейку прибавляли 25 мкл 0,5 М EDTA и 25 мкл хромогенного субстрата S-2222 (80 мкМ). Клетки инкубировали при комнатной температуре в течение 40 минут. Хромогенную активность анализировали при 405 нм, как описано выше.
Таблица 7 показывает хромогенную активность в ячейках, обработанных только Фактором VIIa, и торможение стимуляции, когда DEGR-Фактор VIIa соинкубировали с Фактором VIIa. Эти результаты демонстрируют, что DEGR-Фактор VIIa действует как конкурентный антагонист связывания Фактора VIIa, тем самым подавляя активацию Фактора Х в Фактор Ха и последующее расщепление хромогена S-2222.
Пример VIII.
Дозозависимое ингибирование DEGR-Фактором VIIa поверхностной хромогенной активности на клетках крысиной гладкой мышцы.
Клетки крысиной сосудистой гладкой мышцы помещали в планшеты с 96 ячейками при 4000 клеток на ячейку в ростовой среде с добавкой 1% эмбриональной телячьей сыворотки (как в Таблице 4, но без 10% эмбриональной телячьей сыворотки). Через 5 дней среду удаляли и к клеткам прибавляли или возрастающие концентрации только Фактора VIIa, или 10 нМ Фактор VIIa с возрастающими концентрациями DEGR-Фактора VIIa. Клетки инкубировали в течение часов при 37oС. После инкубации клетки промывали, затем инкубировали с 50 мкл 200 нМ Фактора Х в Tris-забуференном физиологическом растворе в течение 5 минут при комнатной температуре. В каждую ячейку добавляли 25 мкл 0,5 М EDTA и 25 мкл 800 мкМ S-2222 (Kabi Pharmacia), планшеты инкубировали 40 минут при комнатной температуре. Хромогенную активность анализировали при 405 нм со считывающим устройством для микропланшетов, как описано выше.
Таблица 8 показывает дозозависимое повышение хромогенной активности по мере увеличения количеств Фактора VIIa, добавленных в ячейки. При добавлении в клетки смеси DEGR-Фактора VIIa и 100 нМ Фактора VIIa (Таблица 9) наблюдалось дозозависимое ингибирование хромогенной активности. При молярном отношении DEGR-Фактора VIIa к Фактору VIIa, равном 1:1, ингибировалось приблизительно 95% хромогенной активности. Эти данные позволяют предположить, что в этом эксперименте в культуре клеток гладкой мышцы у DEGR-Фактора VIIa было значительно более высокое сродство к поверхностному фактору ткани, чем у нативного Фактора VIIa. Если бы DEGR-Фактор VIIa и Фактор VIIa имели равное сродство к фактору ткани, то в том случае, когда две молекулы были добавлены к клеткам в равном молярном соотношении, уровень наблюдаемого ингибирования не был бы таким высоким.
Таблица 9 показывает дозозависимое ингибирование DEGR-Фактором VIIa хромогенной активности Фактора Ха на клетках крысиной гладкой мышцы. Увеличивающиеся концентрации DEGR-Фактора VIIa соинкубировали с 10 нМ Фактором VIIa, хромогенную активность Фактора Ха определяли с использованием хромогенного субстрата S-2222.
Пример IX
Ингибирование образования Фактора Ха DEGR-Фактором VIIa в анализе растворимого фактора ткани
Превращение Фактора Х в Фактор Ха с использованием очищенного рекомбинантного растворимого фактора ткани установлено с помощью хромогенного анализа. Фактор ткани был экспрессирован и выделен из Saccharomyces cerevisae (Shigematsu et al., J. Bioi. Chem. 267:21329-21337 (1992)). Растворимый фактор ткани был очищен и охарактеризован д-ром W. Kisiel (Университет Нью- Мексико). Реакционная смесь содержала 65,9 мкл растворимого фактора ткани (2,2 мкМ), 29,0 мкл PCPS (1 мМ, Sigma, Сент-Луис, Миссури), 29,5 мкл человеческого Фактора Х (4,1 мкМ), 2,77 мл буфера Хенкса (25 мМ Tris, рН 7,4, 150 мМ NaCl, 2,7 мМ KCl, 5 мМ CaCl2, 0,1% BSA). В каждую из 96 ячеек планшета добавляли 40 мкл смеси фактора ткани и Фактора X, 25 мкл Фактора VIIa, разведенного TBS, и 25 мкл DEGR-Фактора VIIa, разведенного TBS. Для контроля использовали 40 мкл смеси фактора ткани и Фактора X; контроль включал 40 мкл смеси фактора ткани и Фактора X, 20 мкл Фактора VIIa, разбавленного TBS, и 25 мкл только TBS. К реакционной смеси в ячейки добавляли 10 мкл хромогенного субстрата S-2222 (4 мМ), смесь инкубировали 2-10 минут при комнатной температуре. Результаты анализировали при 405 нм со считывающим устройством для микропланшетов, как описано выше.
Стандартная кривая получена с использованием возрастающих концентраций Фактора VIIa, добавленного в отсутствие DEGR-Фактора VIIa. Результаты, представленные в таблице 10, показывают, что существует дозозависимое усиление хромогенной активности по мере увеличения количества Фактора VIIa, добавленного к реационной смеси. Одновременное добавление изменяющихся количеств DEGR-Фактора VIIa и 100 нМ Фактора VIIa привело к понижению хромогенной активности (таблица 11). Эти данные демонстрируют, что DEGR-Фактор VIIa действует как конкурентный антагонист связывания нативного Фактора VIIa с растворимым фактором ткани, тем самым ингибируя образование Фактора Ха, что определяется по снижению хромогенной активности по отношению к хромогенному субстрату S-2222.
Таблица 10.
Стимуляция хромогенной активности Фактора Ха возрастающими концентрациями Фактора VIIa, добавленного к растворимому фактору ткани. Изменение оптической плотности измерено с использованием хромогенного субстрата S-2222.
Пример X.
Подавление коагуляции DEGR-Фактором VIIa.
Стандартные анализы свертывания с целью изучения влияния DEGR-Фактора VII на время свертывания проводили следующим образом. 100 мкл нормальной плазмы бабуина, полученной обработкой с цитратом натрия в качестве антикоагулянта, добавляли к 100 мкл возрастающих концентраций DEGR-Фактора VIIa, разведенного TBS (20 мМ Tris, pH 7,4 и 150 мМ NaCl). Образцы смешивали и недолго инкубировали при 37oС. Затем добавляли к автоматическому счетчику времени коагуляции Electra 800 (Medical Laboratories Automation, Плизантвилле, Нью-Йорк). После инкубации 200 мкл препарата фактора ткани, содержащего 20 мМ хлористый кальций, добавляли к препаратам DEGR-Фактора VIIa. Препарат фактора ткани готовили в виде экстракта свежезамороженной мозговой ткани бабуина в физиологическом растворе и характеризовали по способности инициировать коагуляцию плазмы бабуина. Была выбрана концентрация фактора ткани, которая обеспечивала время свертывания порядка 40 секунд.
Данные, представленные в таблице 12, демонстрируют дозозависимое увеличение времени при добавлении DEGR-Фактора VIIa. Такая низкая доза, как 1 мкг/мл, вызывала значительное увеличение времени свертывания.
Пример XI.
Подавление накопления тромбоцитов DEGR-Фактором VIIa. Исследована способность DEGR-Фактора VIIa подавлять накопление тромбоцитов в сайтах артериального тромбоза при механическом повреждении у нечеловеческих приматов. Использовали модель эндартерэктомии аорты у бабуинов, в основном, как описано Lumsden et al., Blood 81:1762-1770 (1993). Часть артерии бабуина длиной 1-2 см удаляли, инвертировали и выскабливали для удаления внутренней и примерно 50% средней оболочки. Артерию вновь инвертировали, канюлировали по обоим концам и помещали в экстракорпоральный шунт бабуина, тем самым освобождая посредством шунта механически поврежденную артерию бабуина от тока крови. Перед пуском крови через шунт животному впрыскивали внутривенно собственные тромбоциты, меченные 111 In. Уровень накопления тромбоцитов в поврежденной артерии определяли с помощью видеомонитора гамма-камеры.
В исследовании способности DEGR-Фактора VIIa ингибировать накопление тромбоцитов применяли струйное введение DEGR-Фактора VIIa или физиологического раствора в качестве контроля, которые проводили перед открытием шунта. Измерения поврежденных артерий проводили непрерывно в течение 60 минут. DEGR-Фактор VIIa в дозе 0,005 мг/кг ингибировал накопление тромбоцитов. При струйном введении 1 мг/кг подавление накопления тромбоцитов составило примерно 90% через 1 час после введения препарата. Эти результаты представлены на фиг.2.
Пример XII.
DEGR-Фактор VIIa подавляет сосудистый рестеноз, возникающий после баллонной ангиопластики у кроликов с атеросклерозом.
Исследована способность DEGR-FVIIa модулировать развитие патологических изменений после баллонной ангиопластики у кроликов породы Новозеландская белая (NZW) с атеросклерозом. Эта модель хорошо охарактеризована и считается подходящей для оценки действия соединений с антитромботическими свойствами на развитие сосудистых нарушений Gimple et al., Circulation 86:1536-1546 (1992); Rogosta et al., Circulation 89:1262-1271 (1994)}). Животная модель, использованная для оценки DEGR-FVIIa, в основном такая, как описанная Rogosta, там же.
Анестезию кроликам проводили внутримышечной инъекцией 5 мг/кг ксилазина и 35 мг/кг кетамина. Проксимальные бедренные артерии подвергали веносекции ниже паховой связки с наложением проксимальной и дистальной лигатуры. Выделенные сегменты канюлировали с иглами 27 размера. Выходные отверстия создавали проколом иглы. Выделенные сегменты промывали физиологическим раствором от оставшейся крови, сушили воздухом, пропускаемым со скоростью 80 мл/мин в течение 8 минут. После высушивания воздухом выделенные сегменты снова промывали физиологическим раствором и удаляли лигатуры. Гемостаз поддерживали необтурирующим местным давлением. Сегменты ограничивали металлическими зажимами. Локальный спазм снимали местной обработкой 1% ксилокаином. На следующий день после операции животных переводили на диету, включающую 1% холестерин и 6% масло земляного ореха, на один месяц до баллонной ангиопластики. Для снятия постоперационной боли давали 10 мг/кг тиленола перорально в течение 3-5 дней. После хирургической процедуры давали 1 см3 амбипена в течение 3-5 дней.
Доставка тестируемого препарата животным включала начальное струйное введение непосредственно перед баллонной ангиопластикой, сопровождаемое непрерывной системной инфузией через внутреннюю шейную вену с использованием осмотического насоса. Продолжительность инфузии препарата составила 3 суток. Контрольные животные получали 150 МЕ/кг гепарина струйно до баллонной ангиопластики, затем следовала инфузия физиологического раствора. Животным, получавшим DEGR-FVIIa, проводиди струйное введение с дозой 1 мг/кг, сопровождавшееся инфузией 50 мкг/кг/час.
При установке осмотических насосов для непрерывной системной инфузии у животных проводили анестезию, как описано выше, и поддерживали ее в течение всей процедуры дополнительными 1М инъекциями кетамина и ксилазина. Через разрез вдоль средней линии шеи выделяли отслаиванием правую внутреннюю шейную вену, дистальный конец лигировали. Силиконовую трубку (РЕ-160) вводили в правую внутреннюю шейную вену. Подкожный тоннель был создан для прохода силиконовой трубки. Эту трубку соединяли с осмотическим насосом. Осмотический насос имплантировали подкожно на спине у кролика. Правую общую сонную артерию выделяли отслаиванием, дистальный конец лигировали. Посредством сосудистого скальпеля устанавливали интубатор 5F и перемещали его до соединения с дугой аорты. Кровь брали для определения гемостатических параметров, содержания препаратов и холестерина. Внутриартериально впрыскивали 20 мг ксилокаина. Контрольную ангиографию шейной артерии проводили посредством катетера Бермана 5F, расположенного под аортальным разветвлением с использованием 3-4 мл ренографина, введенного вручную через 3 секунды.
После удаления катетера Бермана в нисходящую аорту вводили 0,014-дюймовый проволочный направитель и располагали его над аортальным разветвлением. Под рентгеноскопическим контролем вводили баллонный катетер для ангиопластики соответствующего размера от 2,0 до 2,5 мм, продвигали его над проволочным направителем и вводили в стеноз. Баллон накачивали до 6 атм в течение 60 секунд вручную. Проводили три накачивания с интервалами в 60 секунд. Эту процедуру выполняли в обеих бедренных артериях у каждого животного.
После баллонной дилатации ангиопластический катетер извлекали и снова вводили катетер Бермана в положение 3 см над аортальным разветвлением. Для уменьшения спазма внутриартериально вводили 20 мг лидокаина. Постоперационная ангиограмма была получена, как описано выше. Растр (1 см) располагали на уровне бедренной артерии для того, чтобы рассчитать фактический диаметр. Затем удаляли катетер. Правую сонную артерию лигировали шелком 3-0, рану ушивали послойно. Амбипен и ацетаминофен давали, как описано выше.
Протромбиновое время и концентрацию DEGR-FVIIa в крови определяли непосредственно перед струйным введением препарата, через 1 час после струйного введения и через 3 суток в конце непрерывного введения. Получали 1-2 мл обработанной цитратом плазмы и определяли протромбиновое время и уровни антигена.
Стандартный анализ свертывания использовали для того, чтобы контролировать протромбиновое время у контрольных и обработанных DEGR-FVIIa животных следующим образом. 25 мкл тестируемой плазмы кролика, обработанной цитратом натрия в качестве антикоагулянта, добавляли к 150 мкл TBS (20 мМ Tris, pH 7,4, 150 мМ NaCl). Образцы перемешивали и присоединяли к автоматическому коагуляционному таймеру Electra 800 (Medical Laboratories Automation, Плизантвилле, Нью-Йорк). После инкубации к плазме добавляли 200 мкл препарата тромбопластина (Sigma Chemical), содержащего 25 мМ хлористый кальций. Выбирали концентрацию тромбопластина, при которой время свертывания контрольной кроличьей плазмы составило примерно 20 секунд.
Метод ELISA (иммуноферментный анализ) использовали для определения концентрации DEGR-FVIIa в образцах плазмы контрольных и обработанных DEGR-FVIIa кроликов. Анализ включал разведение моноклонального антитела к человеческому анти-FVII (Dr. W. Kisiel, Университет Нью-Мексико) до 20 мкг/мл в 0,1 М карбонатном буфере, pH 9,6 и добавление 100 мкл на ячейку в планшеты с 96 ячейками. Затем планшеты инкубировали при 4oС в течение ночи, после чего промывали дважды водным буфером (PBS, pH 7,4, содержащий 0,05% Tween 20). Подавление неспецифических сайтов связывания достигали при использовании 200 мкл блокирующего буфера на ячейку (PBS, pH 7,4, содержащий 0,05% Tween 20 и 1% DSA), инкубации при 37oС в течение 2 часов и последующей обработки промывным буфером.
После блокирования добавляли серию стандартных разведении DEGR-FVIIa, изменяющихся в пределах 20-0,027 нГ/мл, наряду с серией разведении тестируемой кроличьей плазмы (1:100 до 1:4000 в блокирующем буфере) по 100 мкл на ячейку. В качестве отрицательного контроля использовали плазму неиммунизированных кроликов. Затем планшеты инкубировали в течение 1 часа при 37oС, после чего обрабатывали промывным буфером.
Определяли DEGR-FVIIa добавлением 100 мкл на ячейку разбавленного 1:1000 кроличьего поликлонального антитела к человеческому анти-FVII (Dr. Kisiel, Университет Нью-Мексико) в блокирующем буфере. Планшеты инкубировали 1 ч при 37oС, затем пятикратно обрабатывали промывным буфером. Специфическое связывание антитела определяли с использованием 100 мкл на ячейку разведения 1: 2000 конъюгата козье антитело к кроличьему IgG - пероксидаза (Таgо, Inc.). Планшеты инкубировали 1 ч при 37oС и шестикратно обрабатывали промывным буфером. В заключение добавляли 100 мкл раствора субстрата (0,42 мг/мл дигидрохлорида о-фенилендиамина (OPD) в 0,2 М цитратном буфере с рН 5,0, содержащем 0,3% перекись водорода). Через 1-3 минуты при комнатной температуре цветную реакцию останавливали добавлением 100 мкл на ячейку 1н. серной кислоты и содержимое планшетов анализировали при 490 нм с использованием микропланшетного спектрофотометра. Концентрации DEGR-FVIIa в образцах плазмы определяли сопоставлением получаемых значений А490 со значениями стандартной кривой для DEGR-FVIIa.
Результаты анализа образцов плазмы на протромбиновое время и уровни антигена DEGR-FVIIa приведены в таблице 13 и таблице 14 соответственно. Данные представлены для каждого животного в отдельности. В таблице 15 приведена сводка средних величин времени свертывания. Во всех случаях обработанные с DEGR-FVIIa животные имели повышенное значение протромбинового времени в точке, соответствующей 1 часу после струйного введения; в точке, соответствующей третьему постоперационному дню, оно почти достигало уровней предварительной обработки. Анализ антигена DEGR-FVIIa также выявил его высокий уровень в точке, соответствующей 1 часу, изменяющийся в пределах 2-6 мкг/мл, и уровни намного ниже в точке, соответствующей третьему дню. Уровни DEGR-FVIIa, измеренные через 1 час, соответствовали предполагаемому повышению протромбинового времени, установленному обработкой нормальной кроличьей плазмы с DEGR- FVIIa in vitro и определением протромбинового времени анализом стандартного разведения тромбопластина.
Через три недели после ангиопластики непосредственно перед умерщвлением повторяли контрольную ангиограмму, как описано выше, используя левую сонную артерию. Через вертикальный абдоминальный разрез изолировали дистальную аорту и вставляли перфузионную канюлю над аортальным разветвлением. Дистальную аорту промывали 50 мл физиологического раствора, затем фиксировали in vivo введением в течение 15 минут при 120 мм рт.ст. 500 мл раствора Histochoice (AMRESCO, Солон, Огайо). Как только начиналась перфузия, животных умертвляли сверхдозой нембутала (3 мл (65 мг/мл) раствора пентобарбитала внутривенно). Сегмент (5 см) бедренной артерии иссекали билатерально. Для оптической микроскопии ткань сохраняли в растворе Histochoice.
Для того чтобы обнаружить появление повреждения в сайте баллонной ангиопластики, из иссеченных бедренных артерий вырезали 3 мм срезы, заливали парафином, брали срезы областей артерии с множественными поражениями. Готовили препараты срезов на предметных стеклах, окрашивали гематоксилином, эозином и красителем Van Giemson. С использованием программы Биоквант проводили морфометрический анализ для получения данных о просвете, внутренней и средней оболочке артерий. С помощью морфометрического анализа срезов от поврежденных артерий проводили измерения всей площади просвета; площади внутренней оболочки, полученной измерением области во внутреннем эластическом слое и вычитанием соответствующей площади просвета каждого среза; площади средней оболочки, определенной измерением области внутри внешнего эластического слоя и вычитанием площади внутри внутреннего эластического слоя. Измерения повреждений внутренней оболочки (интимы) бедренных артерий у контрольных и обработанных DEGR-FVIIa животных показали, что наблюдалось значительное снижение размера интимы у животных, обработанных DEGR-FVIIa (таблица 16). В противоположность этому, измерения медиальной области не выявили заметных отличий между этими двумя группами.
Данные ангиографических измерений представлены в таблице 17 в виде значений среднего светового диаметра (среднего диаметра просвета) (MLD) ± стандартное отклонение для контрольных и обработанных DEGR-FVIIa животных для всех трех временных точек: непосредственно перед ангиопластикой, сразу после и через 21 день после ангиопластики. Не отмечено значительных различий в значениях MLD между контрольными и обработанными DEGR-FVIIa животными ни до, ни сразу после ангиопластики. Однако заметное повышение значения MLD наблюдалось у животных, обработанных DEGR-FVIIa, на 21-ый день после ангиопластики.
Пример XIII.
Подавление DEGR-Фактором VIIa генерации поверхностного Фактора Х на SMCs бабуина.
Проведен поверхностноклеточный хромогенный анализ, в основном, как описано выше в Примере VIII, для определения эффективности DEGR-FVIIa в подавлении связывания FVIIa с поверхностноклеточным фактором ткани и последующего превращения Фактора Х в Фактор Ха на монослойных клетках гладкой мышцы (SMCs) бабуина. Метод является модификацией методик, описанных Sakai et al., J. Biol. Chem. 264: 9980-9988 (1989); Wildgoose et al., Proc. Natl. Acad. Sci. USA 87:7290-7294 (1990). Бабуиновые SMCs получали из Университета Вашингтона (Сиэтл, Вашингтон) и культивировали из имплантатов аорты. Бабуиновые SMCs вносили в культуральные чашки с 96 ячейками в концентрации 8000 клеток на ячейку в 200 мкл на ячейку среды DMEM с добавкой 10% эмбриональной телячьей сыворотки и выдерживали в этой среде в течение 4 дней при 37oС в атмосфере 5% СO2. При анализе удаляли 110 мкл культуральной среды и в ячейки добавляли возрастающие концентрации FVIIa в комбинации с DEGR-FVIIa. Была получена стандартная кривая для концентрации FVIIa, изменяющейся в пределах 5-0,4 нМ. Для оценки ингибиторного действия DEGR-FVIIa на активность FVIIa в тестируемые ячейки добавляли возрастающие концентрации DEGR-FVIIa в присутствии постоянного количества (5 нМ) FVIIa. И FVIIa, и DEGR-FVIIa разбавляли буфером HEPES (10 мМ HEPES, 137 мМ NaCl, 4 мМ KCl, 5 мМ CaCl2, 11 мМ глюкоза, 0,1% BSA) и 10 мкл 10• исходных растворов, добавленных к клеткам. Клетки инкубировали с тестируемыми соединениями в течение 2 часов при 37oС, затем трижды промывали буфером HEPES. Затем в каждую ячейку прибавляли 50 мкл 200 нМ раствора Фактора Х в Tris-буфере (25 мМ Tris, pH 7,4, 150 мМ NaCl, 2,7 мМ KCl, 5 мМ CaCl2, 0,1% BSA). Через 4 минуты при комнатной температуре добавляли 25 мкл 0,5 М EDTA для того, чтобы остановить превращение Фактора Х в Ха. Прибавляли 25 мкл на ячейку 0,8 М Фактор Ха-специфичного хромогенного субстрата S-2222 в Tris-буфере и через 60 минут измеряли поглощение при 405 нм в микропланшетном считывающем устройстве Thermomax (Molecular Devices Corp., Менло-Парк, Калифорния).
Результаты, представленные на Фиг.3, демонстрируют дозозависимое увеличение амидолитической активности в ячейках, обработанных FVIIa (незаштрихованные квадраты). Повышение поглощения является прямым измерением уровня Фактора Ха, генерированного в ячейках, и последующего расщепления им хромогенного субстрата. Добавление возрастающих количеств DEGR-FVIIa при постоянном количестве FVIIa (5 нМ) вызвало дозозависимое снижение амидолитической активности (закрашенные квадраты). Эквимолярные количества DEGR-FVIIa и FVIIa способны ингибировать более 90% хромогенной активности. Даже при сниженном 10-кратно уровне DEGR-FVIIa еще наблюдается 40% ингибирование хромогенной активности Фактора Ха. Эти результаты подтверждают заключение о том, что DEGR-FVIIa является чрезвычайно сильным антагонистом активации Фактора Х в Ха в присутствии FVIIa на поверхности интактного клеточного монослоя SMCs.
Пример XIV. Влияние DEGR-Фактора VIIa на возникновение сосудистого тромбоза и сосудистых нарушений у бабуинов.
Была исследована спосбность человеческого DEGR-Фактора VIIa ингибировать фактор ткани (TF) и опосредованное активированным Фактором VII (FVIIa) образование сосудистых нарушений (VLF), индуцированных механическим повреждением сосудов у нечеловеческих приматов.
DEGR-Фактор VIIa вводили внутривенно в течение 7 дней (5 животным) или 30 дней (1 животному), начиная непосредственно перед возникновением механического сосудистого повреждения. Измерения образования сосудистого повреждения проводили на 30-ый день. Результаты, полученные у 5 обработанных животных, сопоставляли с данными для 5 параллельных контролей с инфузией наполнителя в буфере.
Базовые значения получали при определении у животных: а) количества тромбоцитов, нейтрофилов, моноцитов и эритроцитов; б) уровня плазменного фибриногена; в) уровней активности плазменных факторов коагуляции VII, VIIa, Х и V наряду с антигенностью FVII; г) базовый образец плазмы для определения антител к анти-фактору VIIa.
При анестезии галотаном и стерильных операционных условиях животные, меченные аутологичными 111 In-тромбоцитами, получали внутривенные вливания с использованием всей системы непрерывного внутривенного введения (начальное струйное введение 1 мг/кг, сопровождавшееся непрерывным внутривенным вливанием 50 мкг/кг/час). Животным проводили хирургическую эндартерэктомию сонной артерии, билатеральную ангиопластику плечевой или бедренной артерии с помощью катетера Фогарти.
DEGR-FVIIa вводили в течение 7 или 30 дней непрерывной инфузией через венозный катетер с использованием всей системы. Через 30 дней после хирургического вмешательства животному давали галотановый наркоз и проводили фиксацию in situ методом перфузии под давлением с 4% параформальдегидом, содержащим 0,1% глутаровый альдегид, в течение 30 минут. Затем отбирали сегменты (содержащие ранее поврежденные сайты) с использованием методик Marker et al. , Circulation 83:41-44 (1991); Hanson et al., Hypertension 18:1170-1176 (1991). Образцы подвергали вторичной фиксации in vitro (4% параформальдегидом, содержащим 0,1% глутаровый альдегид), замораживали и подготавливали для морфометрического анализа области поражения.
Исследовали 11 нормальных зрелых бабуинов (Paio anubis). Шесть животных получали вливания (50 мкг/кг/час), остающееся пять служили контролем, не получавшим DEGR-FVIIa. Животные были дегельминтизированы и находились под наблюдением, чтобы избежать заболеваний, в течение трех месяцев перед использованием. Все процедуры были утверждены Институциональным комитетом по уходу и использованию животных и находились в соответствии с процедурами и методами, изложенными в "Руководстве Национального института здоровья по уходу и использованию лабораторных животных", и подобными институциональными правилами. Инвазивные процедуры проводили под анестезией галотаном после индукции кетамином (10 мг/кг внутримышечно) и валиумом (0,5 мг/кг внутривенно). Для последующей кратковременной иммобилизации при проведении постоперационных экспериментальных процедур использовали гидрохлорид кетамина (5-20 мг/кг внутримышечно).
Эндартерэктомию сонной аотерии проводили посредством разреза по средней линии шеи с использованием приемов, описанных Hanson et al., Hypertension 18:1170-1176 (1991); Krupski et al., Circulation 84:1749-1757 (1991), включенных здесь в виде ссылок. Эндартерэктомию использовали в качестве модели сосудистого нарушения вследствие ее клинической важности и потому, что, как было показано, VLF, вызванное эндартерэктомией нормальных артерий, является воспроизводимым. Кратко, обычную сонную артерию иссекали из окружающих тканей от ключицы проксимально до бифуркации сонной артерии дистально. Обычную артерию пережимали с использованием атравматических сосудистых зажимов на каждом конце выделенного сосуда через 3 минуты после струйного введения сульфата гепарина (100 МЕ/кг внутривенно; Elkins-Simm Inc., Черри-Хилл, Нью-Джерси) и отделяли 1 см, проксимальный к дистальному пережиму. Затем проксимальный артериальный сегмент выворачивали над изогнутым зажимом. После того как достигалось максимальное выворачивание, в выделенном в просвете сегменте накладывали одну пару полипропиленовых швов (7-0) на любую из сторон проксимально и вторую пару дистально. Затем проводили эндартерэктомию, начиная в 1 см от отделенного конца вывернутого сосудистого сегмента и продолжая на измеренное расстояние в 1 см. Эта процедура включает механическое удаление нормальной внутренней оболочки и частичное уплотнение средней оболочки с использованием зажима и хирургического микроскопа (увеличение 32х). После эндартерэктомии сосуду возвращали его обычную конфигурацию и налагали анастомоз "конец-к-концу" с помощью полипропиленового шва (7-0) и постоянной техники при 2,5-кратном увеличении, рану зашивали послойно.
При морфометрическом анализе исследовали срезы, залитые парафином и обработанные красителем, для компонентов соединительной ткани (коллаген, эластин) и смесью гематоксилин-эозин, с использованием фотоскопа (Zeiss), соединенного с системой обработки изображения (Thomas Optical Measurement Systems, Колумбус, Джорджия), состоящей из камеры микроскопа CCD высокого разрешения (580 линий), присоединенной к монитору высокого разрешения (700 линий), компьютеру IBM 386, 80 МБ с графическим вводом высокого разрешения для получения и хранения изображения. Количественную обработку изображения проводили с использованием орфометрического програмного обеспечения (Optimas, Bioscan Inc., Эдмондс, Вашингтон). В артериальных поперечных срезах анализировали общую площадь пролиферативного повреждения неоинтимы и соответствующую площадь артериальной средней оболочки. Для статистического анализа сравнение между группами проводили с использованием t-теста Стьюдента (двустороннего для парных и непарных данных).
Результаты показали, что площадь интимы значительно снижена у животных, обработанных DEGR-Факторм VIIa в течение 7 дней и наблюдавшихся до 30 дней, по сравнению с контрольными животными, которые не получали DEGR-Фактор VIIa (фиг.4). Подобные результаты получены у животных, обработанных DEGR-Фактором VIIa в течение 30 дней и наблюдавшихся до 30 дней.
Предварительные исследования с моделью баллонной ангиографии плечевой артерии не наводили на мысль о заметном преимуществе терапии с DEGR-Факторм VIIa. Однако на бабуинах не было показано, что эта модель является протромботической, в которой ключевую роль играет фактор ткани.
Исследования баллонного повреждения бедренной артерии у бабуинов действительно показали статистически значимое преимущество DEGR-Фактора VIIa по сравнению с контролем, как показано на фиг.5.
Пример XV.
Влияние DEGR-Фактора VIIa на tPA-индуцированный тромболиз.
Постоянное образование коронарных тромбов при остром инфаркте миокарда опосредовано, в первую очередь, фактором ткани (TF) в комплексе с Фактором VIIa через внешний коагуляционный метаболический путь. Было изучено влияние дополнительного ингибирования коагуляционного каскада в различных точках внешнего метаболического пути на эффективность тромболиза, индуцированного активатором тканевого плазминогена (tPA).
Тридцати шести собакам с индуцированными электрическим током коронарными тромбами в ходе тромболиза с tPA (1 мг/кг, дольше 20 минут) назначали одну из четырех дополнительных обработок: 9 получали клещевой антикоагулянтный пептид (ТАР), селективный ингибитор Фактора Ха, в дозе 30 мкг/кг/мин в течение 90 минут. Комплекс TF-Фактор VIIa подавляли рекомбинантным ингибитором метаболизма фактора ткани (TFRI) (100-150 мкг/кг/мин в течение 90 минут) у 9 собак и DEGR-FVIIa (1-2 мг/кг струйно) в качестве конкурентного антагониста активированного Фактора VIIa у 9 собак. В качестве контроля 9 собак получали физиологический раствор. Собаки находились под наблюдением по поводу реокклюзии в течение 120 минут после тромболиза. Данные о влиянии этих агентов на эффективность тромболиза приведены в таблице 18 (данные представлены в виде средних значений ± SD).
Из этих данных следует, что ингибирование внешнего метаболического пути либо Фактором Ха, либо блокадой комплекса TF-Фактор VIIa с DEGR-FVIIa или TFPI ускоряло tPA-индуцированный тромболиз. Селективное ингибирование Фактора Ха более эффективно поддерживает раскрытое состояние артерии после успешной реперфузии.
Пример XVI.
Модифицированный Фактор VIIa ингибирует образование внутри сосудистых тромбов, не оказывая воздействия на системную коагуляцию.
Для того чтобы определить, приводит ли ингибирование связывания Фактора VII с TF к антитромботическим эффектам, были инициированы изменения циклов кровотока (CFVs), вызываемые рецидивирующим тромбообразованием, путем помещения внешнего зажима вокруг кроличьих артерий с поврежденным эндотелием (метод Фоттса). После помещения зажима вокруг артерии CFVs развивались со средней частотой 11 ±2 циклов/час у 6 из 6 кроликов, в то время как скорость кровотока в сонной артерии составляла в среднем 5±2% от базовых значений при самом низком уровне CFVs. После наблюдения CFVs в течение 30 минут животные получали вливание человеческого рекомбинантного блокированного по активному центру (Phe-Phe-Arg-хлорметилкетон) Фактора VII (FVIlai) (0,1 мг/кг/мин в течение 10 минут). Фактор VIIai полностью снимал CFVs у 6 из 6 животных (частота CFV= 0 циклов/час; р<0,05; скорость кровотока через сонную артерию равна 106,9% от базовых значений; p=NS против базовой линии). Через 30 минут после ингибирования CFVs вводили человеческий рекомбинантный FVIIa в дозе 0,1 мг/кг/мин в течение 10 минут. Инфузия Фактора VIIa восстанавливала CFVs у всех животных, тем самым показывая, что связывание Фактора VIIa с TF было конкурентным. Значения протромбинового времени, времени активации и агрегации тромбоцитов ех vivo в ответ на ADP (аденозиндифосфат) и тромбин после инфузии FVIIai не отличались при сравнении с базовыми значениями. Таким образом, FVII-VIIa играет важную роль в инициировании тромбообразования in vivo. Введение Фактора VIIai вызывает на этой модели сильные антитромботические эффекты, не оказывая воздействия на системную коагуляцию.
Из всего упомянутого очевидно, что получены составы Фактора VII или VIIa с модифицированными каталитическим сайтами, способные связывать фактор ткани, однако не способные существенно активировать Факторы Х и IX. Поскольку модифицированный Фактор VII специфически прерывает каскад свертывания, не нарушая или не удаляя факторы свертывания, введение пепаратов модифицированного Фактора VII будет сопровождаться меньшими нежелательными побочными эффектами, чем те, которые известны по опыту современной терапии. Далее, модифицированный Фактор VII, описанный здесь, может быть легко получен рекомбинантными способами. Таким образом, эффективность, удобство и экономическая выгода от низких доз и снижения частоты введения, а также относительная потеря токсичности составляют преимущества, обеспечиваемые составами настоящего изобретения.
Хотя вышеупомянутое изобретение и описано достаточно подробно с привлечением иллюстраций и примеров для улучшения понимания, очевидно, что могут быть предприняты некоторые изменения и модификации в рамках прилагаемой формулы изобретения.

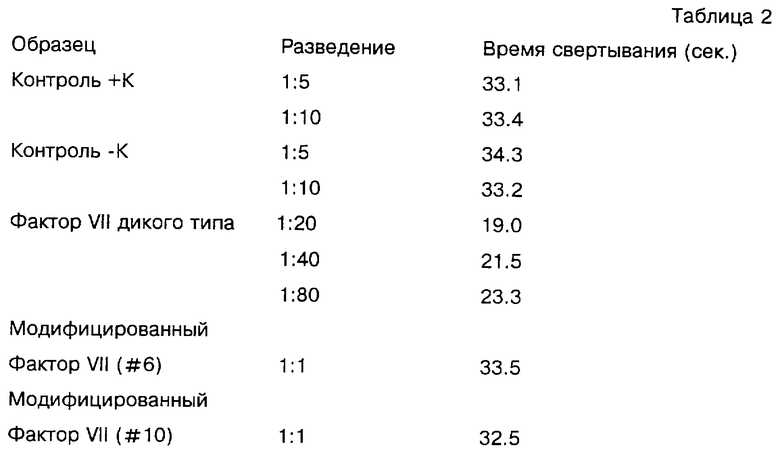
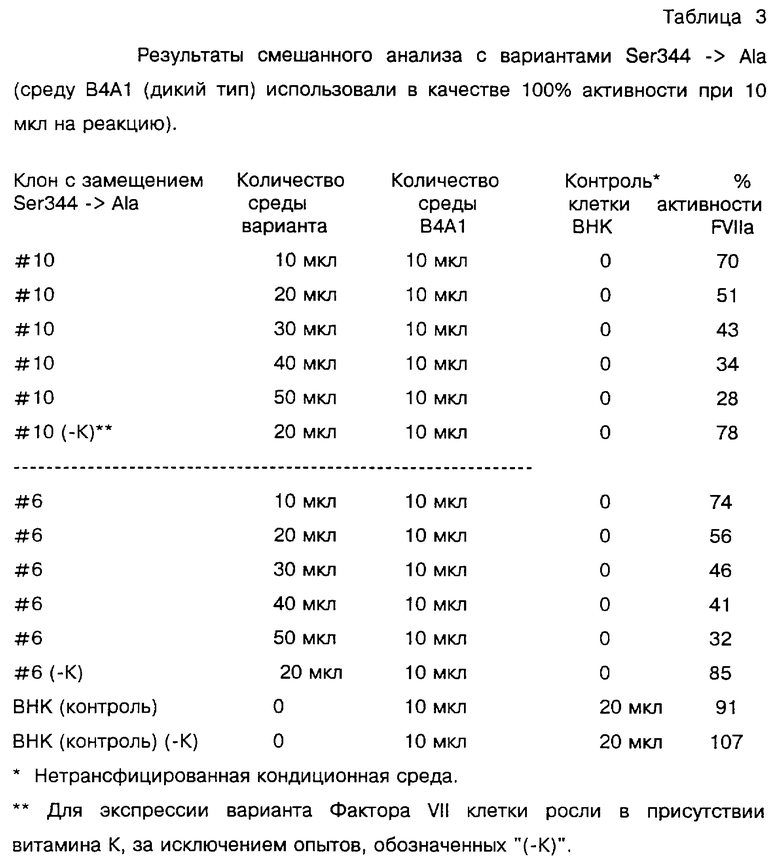






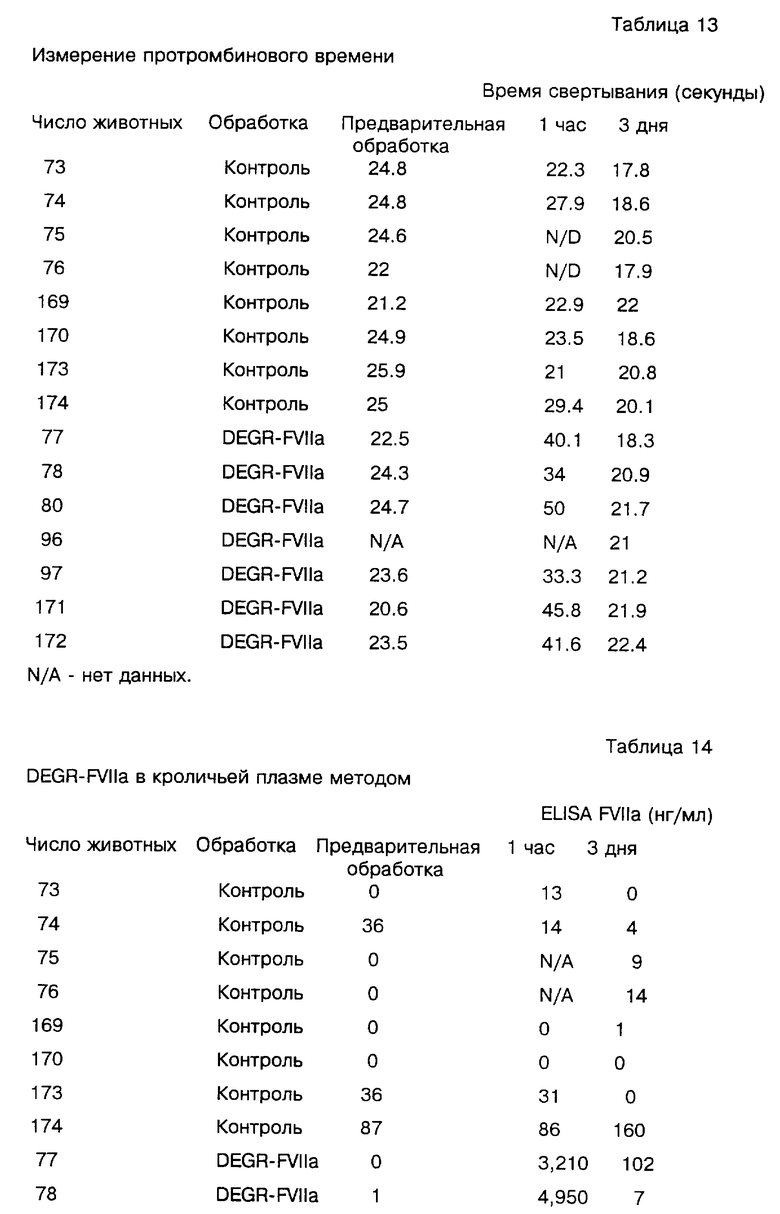

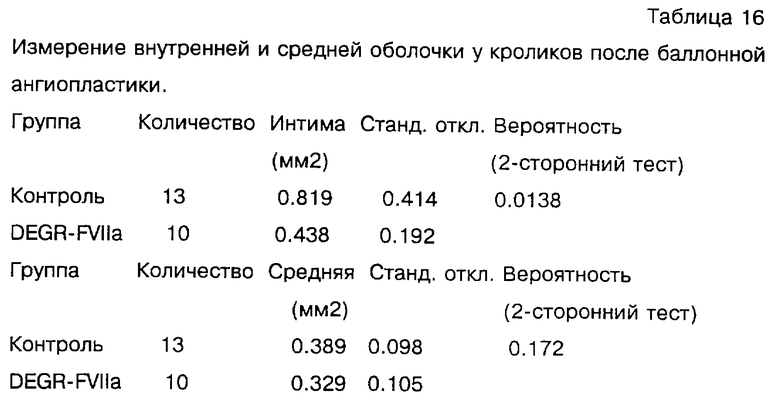
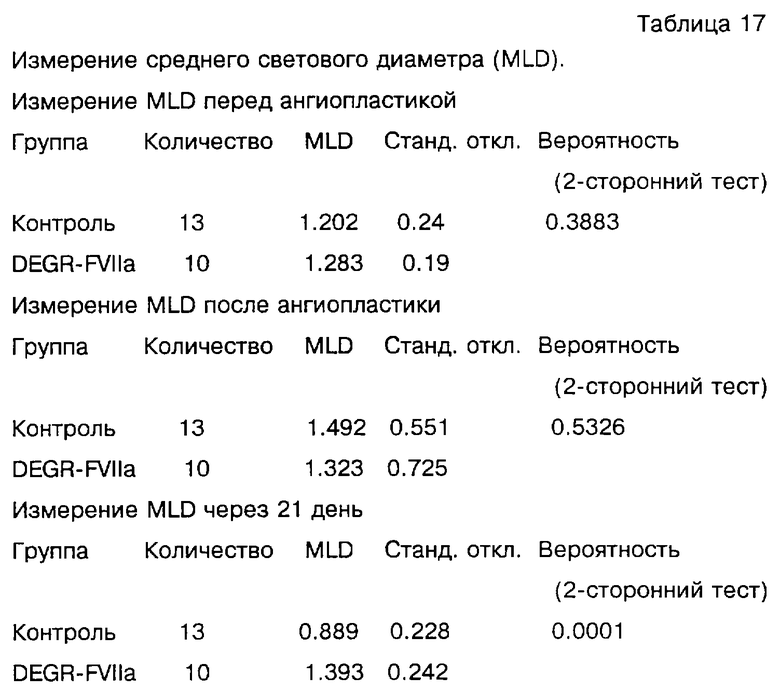

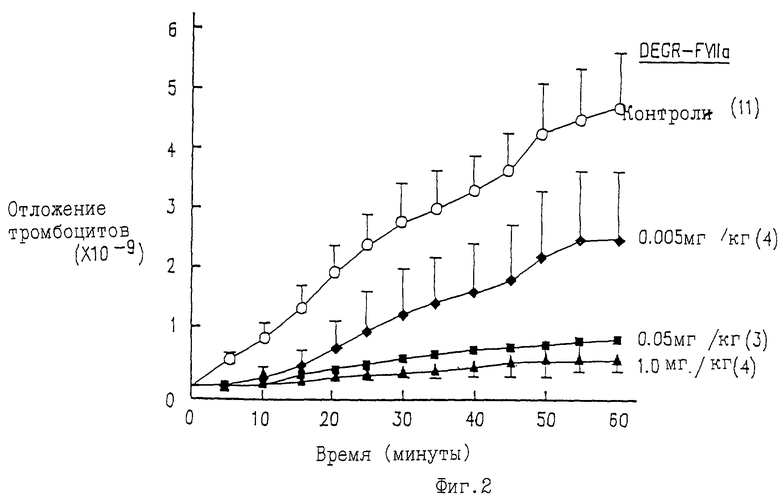
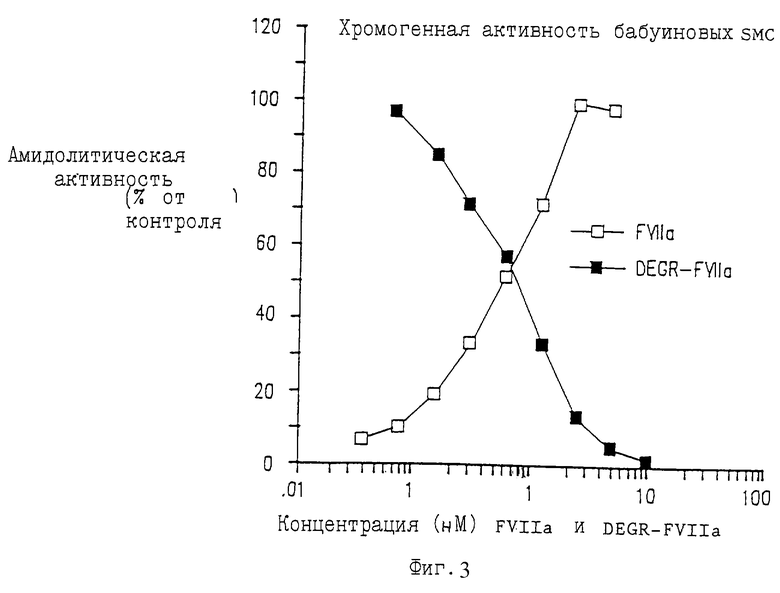
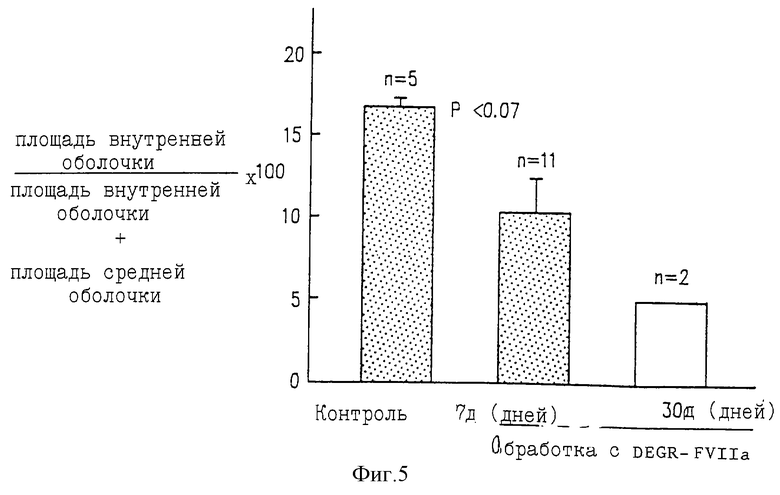
Изобретение относится к медицине, в частности к гематологии и к белкам, используемым в качестве антикоагулянтов, в частности к модифицированным формам Фактора VII, ингибирующим свертывание крови и фактор ткани. В изобретении заявляются способы для различного использования, в частности для подавления отложений тромбоцитов у пациента и лечения острой закупорки коронарной артерии, и состава, состоящего из Фактора VII, имеющего, по крайней мере, одну модификацию в своем активном центре, модификация которого существенно подавляет способность модифицированного Фактора VII активировать плазматический Фактор Х или IX. Изобретение позволяет получить препарат без нежелательных побочных эффектов, который может быть легко получен рекомбинантным путем. 4 с. и 19 з.п .ф-лы, 5 ил., 18 табл.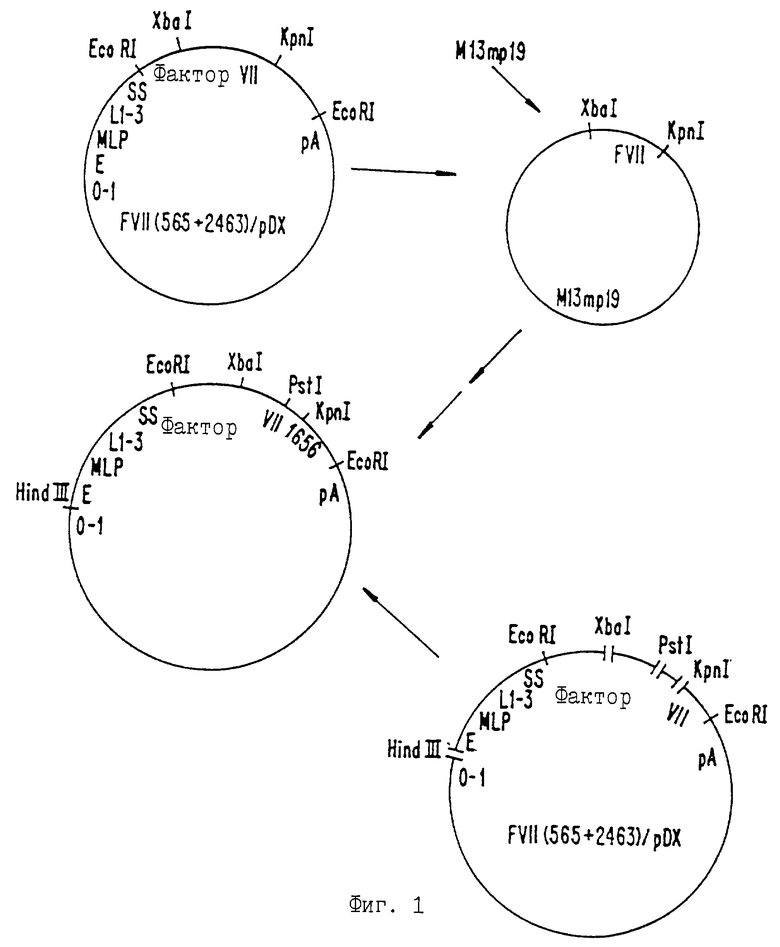
| US 4386025, 31.05.1983 | |||
| СПОСОБ ПОЛУЧЕНИЯ ЗИМОГЕННОЙ ФОРМЫ ЧЕЛОВЕЧЕСКОГО БЕЛКА С | 1989 |
|
RU2018535C1 |
| ЗАМЕЩЕННЫЕ ФЕНИЛАЦЕТИЛЕНЫ, ОБЛАДАЮЩИЕ СВОЙСТВОМ СПЕЦИФИЧЕСКИ ТОРМОЗИТЬ 5-ЛИПОКСИГЕНАЗУ | 1991 |
|
RU2014323C1 |

