Изобретение относится к способу микробиологического получения аминокислот согласно пп.1-20 формулы изобретения, экспорт-генов по пп.21-26, регуляторных генов по пп.27 и 28, генных структур по пп.29 и 30, векторов по пп.31-33, трансформированных клеток по пп.34-40; мембранных протеинов по пп.41 и 42, а также к применениям по пп.43-48.
Аминокислоты представляют большой хозяйственный интерес, причем аминокислоты имеют разнообразное применение. Так, например, L-лизин, как и L-треонин, и L-триптофан используют в качестве кормовой добавки, L-глутамат в качестве пряности, L-изолейцин и L-тирозин в фармацевтической промышленности, L-аргинин и L-изолейцин в качестве медикамента или L-глутамат и L-фенилаланин в качестве исходного вещества для синтеза чистых химикатов.
Предпочтительным методом получения этих самых различных аминокислот является биотехнологическое получение при помощи микроорганизмов, так как этим способом прямо получается биологически эффективная и оптически активная форма соответствующей аминокислоты и можно применять простое и недорогое сырье. В качестве микроорганизмов применяют, например, Corynebacterium glutamicum и его родственников ssp. flavum и spp. lactofermentum (Liebl и др.)., Int J System Bacteriol (1991) 41:255-260), как и Escherichia coli и родственные бактерии.
Эти бактерии продуцируют обычным образом аминокислоты, но только в необходимом для роста количестве, так, что, следовательно, никаких избыточных аминокислот не образуется и не выделяется. Это обосновано тем, что в клетке биосинтез аминокислот многократным образом контролируется. Следовательно, уже известны самые различные способы, чтобы повышать образование продукта выключением контрольных механизмов. При этих процессах применяют, например, аналоги аминокислот, чтобы выключать эффективное регулирование биосинтеза. Так, описан способ, при котором используют штаммы Corynebacterium, которые резистентны против аналогов L-тирозина и L-фенилаланина (патент Японии 19037/1976 и 39517/1978). Описаны также способы, при которых применяют резистентные против аналогов L-лизина или также L-треонина бактерии, чтобы преодолевать контрольные механизмы (ЕР 0205849 В1, патент Украины, приложение к патенту Великобритании 2152509 А).
Далее, известны также сконструированные путем рекомбинантных ДНК-методов микроорганизмы, при которых также повышается регуляция биосинтеза тем, что гены, которые больше не кодируют тормозимые конечные энзимы с обратными связями, клонируют и экспрессируют. Так, например, известна продуцирующая L-лизин, рекомбинантная бактерия с кодированной плазмидой, резистентной к обратной связи аспартаткиназой (ЕР 0381527). Описана также рекомбинантная, продуцирующая L-фенилаланин бактерия с резистентной к обратной связи префенатдегидрогеназой (патент Японии 124375/1986, ЕР 0488424). Кроме того, избыточной экспрессией генов, которые не кодируют чувствительные к обратной связи энзимы синтеза аминокислот, достигают также повышенных выходов аминокислоты. Так, например, образование лизина улучшают путем повышенного синтеза дигидропиколинатсинтазы (ЕР 0197335). Повышенным синтезом треониндегидратазы также достигают улучшенного образования треонина (ЕР 0436886 A1).
Дальнейшие попытки повышения производства аминокислот направлены на улучшенное получение клеточных первичных метаболитов центрального обмена веществ. Так, известно, что достигнутая рекомбинантными методами избыточная экспрессия транскетолазы делает возможным улучшенное образование продукта L-триптофана, L-тирозина или L-фенилаланина (ЕР 0600463 А2). Далее, снижение активности фосфоэнолпируваткарбоксилазы в Corynebacterium приводит к улучшенному образованию ароматических аминокислот (ЕР 0331145).
Эти многочисленные попытки повышения производительности, в целом, направлены на то, чтобы преодолеть ограничение цитосолического синтеза аминокислот. Но в качестве другого ограничения, в принципе, принимают во внимание также экспорт образованных во внутренней части клеток аминокислот в питательную среду. Поэтому имеются отдельные исходные смеси для улучшения экспорта и рентабельности производства аминокислот. Так, проницаемость клеток при Corynebacterium повышали дефицитом биотина, обработкой детергентами или обработкой пенициллином. Однако эти вспомогательные меры были успешными только при производстве глутамата, в то время, как синтез других аминокислот улучшить таким способом не смогли. Были выделены также штаммы бактерий, при которых активность системы секреции была повышена на основании химической или физической мутации. В результате этого получали, например, Corynebacterium glutamicum-штамм, который благодаря улучшенной активности секреции особенно пригоден, в частности, для производства L-лизина (патент ФРГ 4203320).
В целом, все проведенные до сих пор опыты повышения секреции образованных во внутренней части клеток аминокислот отличаются тем, что повышенный отток аминокислот на основании выбранных неправильных или неспецифических методов мог быть достигнут только случайно. Только в немецкой заявке на патент №19523279.8-41 описан способ, который позволяет совершенно целенаправленно повышать секрецию образованных во внутренней части клеток аминокислот, причем экспрессия кодирующих импорт аминокислот генов повышалась. Лежащий в основе этого принципа действий вывод о том, что клетки используют импортные протеины для экспорта аминокислот, как и тот факт, что микроорганизмы по природе не образуют и не выделяют никаких избыточных аминокислот, заставляет предполагать, что специфические для переноса аминокислот экспорт-гены или экспорт-протеины не существуют, а что из клетки аминокислоты экскретируются через другие системы экспорта.
Известные до сих пор системы экспорта экспортируют ядовитые ионы металлов, токсические антибиотики и более высокомолекулярные токсины. Эти системы экспорта имеют относительно сложную конструкцию: как правило, мембранные протеины участвуют как мембраны цитоплазмы, которые, однако, способствуют только частичной реакции экспорта, так что предположительно для переноса требуются дополнительные, экстрацитоплазматические вспомогательные протеины (Dinh, Т. и др., A family of extracytoplasmic proteins that allow transport of large molecules across the outer membranes of gram-negative bacteria. J. Bacteriol. 1994, 176:3825-3831). Затем известно, что при зависимой от секреции системы экспорта для внеклеточных протеинов существенными являются по меньшей мере 6 различных компонентов протеинов для экспорта. Этот уровень техники предполагает, что и соответствующие для экспорта аминокислот, но до сих пор неизвестные системы состоят из многих компонентов протеинов или многие гены обеспечивают экспорт аминокислот. Ссылкой на это могли бы быть описанные Vrljic и др. (J Bacteriol (1995) 177:4021-4027) различные, дефектные в экспорте лизина мутанты.
Теперь неожиданно было найдено, что за экспорт аминокислот несет ответственность соответственно только один специфический ген, так что согласно изобретению впервые представляется в распоряжение способ микробиологического получения аминокислот, при котором целенаправленно повышают экспрессию экспорт-гена и/или активность экспорт-носителя, продуцирующего соответствующую аминокислоту микроорганизма. Получающаяся в результате этого способа действий повышенная экспрессия или активность экспорт-носителя приводит к повышенной норме секреции, так что экспорт соответствующей аминокислоты повышен. Измененные таким образом микроорганизмы накапливают также повышенное количество соответствующей кислоты в культуральной среде.
Для повышения активности экспорт-носителей повышают, в частности, эндогенную активность продуцирующего аминокислоту микроорганизма.
Повышения активности фермента можно, например, достичь тем, что в результате изменения каталитического центра происходит повышенное превращение субстрата или прекращается действие ферментных ингибиторов. Повышенную активность фермента можно вызывать также повышением синтеза фермента, например, усилениями гена или исключением факторов, которые подавляют биосинтез фермента. Эндогенную активность экспорт-носителей повышают преимущественно мутацией эндогенного экспорт-гена. Подобные мутации можно производить или классическими методами нецеленаправленно, как, например, облучением ультрафиолетовыми лучами или растворяющими мутации химикалиями, или целенаправленно при помощи методов генетической инженерии, как делеция (делеции), инсерция(ии) и/или обмен нуклеотидов (обмены нуклеотидов).
Экспрессию экспорт-гена повышают увеличением числа копий гена и/или усилением регуляторных факторов, которые положительно влияют на экспрессию экспорт-гена. Так, усиление регуляторных элементов можно осуществить предпочтительно на уровне транскрипции тем, что, в частности, повышаются транскрипционные сигналы. Этого можно достичь, например, в результате того, что путем изменения предвключенной структурному гену последовательности промоторов промотор повышает свою эффективность или тем, что промотор полностью меняется на более эффективные промоторы. Можно осуществлять также усиление транскрипции путем соответствующего влияния подчиненного экспорт-гену регуляторного гена, как поясняется дальше. Но наряду с этим возможно также усиление перевода, причем улучшается устойчивость m-РНК.
Для повышения числа копий гена экспорт-ген встраивают в конструкцию гена или в вектор, предпочтительно в вектор с более низким числом копий. Конструкция гена содержит, в частности, подчиненные экспорт-гену регуляторные последовательности генов, предпочтительно такие, которые усиливают экспрессию гена. Регуляторные последовательности генов проявляют особенно приведенную в табл.1 последовательность аминокислот или их кодирующую вариации аллелей последовательность нуклеотидов или последовательность нуклеотидов от нуклеотида 954 до 82 по табл.2 или одинаково действующую, в основном, последовательность ДНК. Вариации аллелей или одинаково действующие последовательности ДНК включают особенно функциональные производные, которые получают делецией (делециями), инсерцией (инсерциями) и/или замещением (замещениями) нуклеотидов из соответствующих последовательностей, причем, однако, активность регуляторного протеина или функция регуляторного протеина сохраняются или даже повышены. Так, мутацией регуляторной последовательности гена можно оказывать такое влияние на связывание регуляторного протеина с ДНК регулирующего экспорт-гена, что в результате этого усиливается транскрипция и, следовательно, повышается экспрессия гена. Затем можно придавать экспорт-гену в качестве регуляторных последовательностей также и так называемые "усилители", которые через улучшенное взаимодействие между РНК-полимеразой и ДНК вызывают также повышенную экспрессию экспорт-гена.
Для встройки экспорт-гена в генную конструкцию экспорт-ген, выделенный преимущественно из штамма микроорганизмов рода Corynebacterium и с содержащей экспорт-ген генной конструкцией, трансформируют в продуцирующий соответствующую аминокислоту штамм микроорганизмов, особенно Corynebacterium. Выделение и трансформацию соответствующего транспортного гена осуществляют общепринятыми методами. В случае выделения и клонирования транспортного гена из Corynebacterium используют, например, метод гомологичной комплементации экспортдефектного мутанта (J. Bacteriol (1995) 177:4021-4027). Если невозможно прямое клонирование структурного гена, то возможно сначала проведение также инсерции последовательностей вектора в транспортный ген, чтобы затем выделять его через "спасательный плазмид" в форме неактивных фрагментов. Для способа согласно изобретению особенно подходят гены из C.glutamicum ATCC 13032 или C.glutamicum ssp. flavum ATCC 14067 или также C.glutamicum ssp. lactofermentum ATCC 13869. После выделения генов и их рекомбинации в пробирке с известными векторами (Appl Env Microbiol (1989) 55:684-688; Гены 102 (1991) 93-98), в продуцирующих аминокислоту штаммах осуществляют трансформацию электропорацией (Lieb и др. (1989) FEMS Microbiol Lett 65:299-304) или конъюгацией (Шефер и др. (1990) J Bacteriol 172:1663-1666). Для переноса применяют предпочтительно переносчики с низким числом копий. В качестве клеток хозяев применяют предпочтительно такие продуценты аминокислоты, которые в синтезе соответствующих аминокислот дерегулированы и/или содержат повышенное количество метаболитов центрального обмена веществ.
После выделения экспорт-гены получают с нуклеотидными последовательностями, которые кодируют указанную в табл.3 последовательность аминокислот, или вариации их аллелей, соответственно которые имеют нуклеотидную последовательность от нуклеотида 1016 до 1725 по табл.2 или, в основном, действующую одинаково последовательность ДНК. И здесь вариации аллелей или одинаково действующие последовательности ДНК охватывают, в частности, функциональные производные в указанном выше для регуляторных последовательностей смысле. Эти экспорт-гены применяют предпочтительно в способе согласно изобретению.
К экспорт-гену с предварительно включенным промотором или без него или с соответственным регуляторным геном или без него могут быть предварительно включены и/или дополнительно включены одна или несколько ДНК последовательностей, так что ген содержится в структурном гене.
Клонированием экспорт-генов получают плазмиды соответственно вектора, которые содержат экспорт-ген и, как уже упомянуто выше, используют их для трансформации продуцента аминокислоты. Получаемые трансформацией клетки, у которых речь идет предпочтительно о трансформированных клетках Corynebacterium, содержат ген в реплицируемой форме, т.е. в дополнительных копиях на хромосоме, причем копии гена интегрируют гомологичной рекомбинацией на любых местах генома, и/или на плазмиде, соответственно - векторе.
Известно множество последовательностей, которые кодируют мембранные протеины неизвестной функции. Благодаря получению экспорт-генов согласно изобретению, как, например, экспорт-гена с последовательностью нуклеотидов от нуклеотида 1016 до 1725 по табл.2, или соответствующих экспорт-протеинов, как, например, с последовательностью аминокислот по табл.1, можно теперь идентифицировать мембранные протеины, функцией которых является транспорт аминокислот, путем сравнения последовательностей. Идентифицированный таким способом экспорт-ген можно применять затем для улучшения производства аминокислот по способу согласно изобретению.
Известные из уровня техники мембранные протеины, как правило, имеют 12, частично также 4 трансмембранные спирали. Теперь неожиданно было найдено, то соответствующие для экспорта аминокислот, соответственно подходящие мембранные протеины имеют 6 трансмембранных спиралей (см. например, приведенную в табл.3 последовательность аминокислот экспорт-протеина, при которой 6 трансмембранных областей обозначены подчеркиванием). Тем самым здесь представлен еще не описанный до сих пор и, следовательно, новый класс мембранных протеинов.
Примеры осуществления:
а) Клонирование экспорт-гена и клонирование регулятора из Corynebacterium glutamicum.
Хромосомные ДНК из С. glutamicum R127 (FEMS Microbiol Lett (1989) 65:229-304) были описаны как у Шарца и др. (Bio/Technology (1990) 9:84-87) и выделены. Последние расщепляли с ограничительным ферментом Sau3А и разделяли градиентным центрифугированием сахарозы, как описано у Sambrook и др. (Molecular Cloning, A Laboratory manual (1989) Cold Spring Harbour Laboratory Press). Отдельные фракции анализировали методом гель-электрофореза относительно их размера, и фракцию с размером фрагмента около 6-10 кв использовали для сшивания с вектором pJCI. Для этого вектор pJCI с BamHI линеаризировали и дефосфорилировали. 5 нг от этого лигировали с 20 нг хромосомных фрагментов размером 6-10 кв. С общей сшивочной вставки трансформировали экспорт-дефектный мутант NA8 (J Bacteriol (1995) 177:4021-4027) путем электропорации (FEMS Microbiol Lett (1989) 65:299-304). Трансформанты селекционировали на LBHIS (FEMS Microbiol Lett (1989) 65:299-304) с 15 мкг канамицина на 1 мл. Эти трансформанты подвергали обширным анализам на плазмиды, причем выбирали отдельно 200 из всех 4500 полученных клонов, и определяли количество в них плазмидов и размер. В среднем, приблизительно половина исследованных резистентных к канамицину клонов несла рекомбинантный плазмид с вставкой среднего размера 8 кв. Тем самым получается вероятность 0,96 для присутствия каждого х-любого гена из С. glutamicum в сформированном банке генов. Все 4500 полученных трансформантов испытывали отдельно на обратное получение секреции лизина. Для этого применяли описанную Vrljic систему для индукции выделения L-лизина в Corynebacterium glutamicum (J Bacteriol (1995) 177:4021-4027). Для этого изготовляли так называемые индикаторные пластинки с минимальной средой, которые на 1 л содержали 20 г (NH4)2SO4, 5 г мочевины, 1 г КН2РO4, 1 г К2НРO4, 0,25 г МgSO4 × 7Н2О, 42 г морфолинопропансульфокислоты, 1 мл СаСl2 (1 г/100 мл), 750 мл дистиллированной воды, 1 мл микросолей Сg, 1 мл биотина (20 мг/100 мл), рН 7, 4% глюкозы, 1,8 мг протокатехиновой кислоты, 1 мг FeSO4 × 7Н2О, 1 мг MnSO4 × Н2О, 0,1 мг ZnSO4 × 7Н2О, 0,02 мг CuSO4, 0,002 мг NiCl2 × 6Н2О, 20 г агар-агара, а также 107 клеток/мл лизин-ауксотроф С. glutamicum мутанта 49/3. Все первоначальные 4500 трансформантов отдельно при помощи зубочистки накалывали на индикаторные пластинки, соответственно с контролем первоначального невыделителя NA8 (J Bacteriol (1995) 177:4021-4027) и исходного штамма R127. Параллельно прививали соответственно 2 пластинки, из которых только одна содержала дополнительно 5 мМол L-метионина, чтобы таким образом индуцировать выделение лизина. Индикаторные пластинки инкубировали при 30°С и исследовали после 15, 24 и 48 ч. Всего получали таким образом 29 клонов, которые показывали на оставленной с метионином индикаторной пластинке ободок роста благодаря индикаторному штамму 49/3. Клоны разъединяли и затем снова, как описано выше, проверяли на обратное получение ободка роста. Этим способом получали два клона NA8 pMV8-5-24 и NA8 pMV6-3, которые снова получали способность выделять лизин.
Из этих клонов получали плазмидные препараты, которые описаны как у Шварца и др. (Bio/Technology (1990) 9:84-87).
Ретрансформацией в NA8 был подтвержден связанный с плазмидом эффект выделения L-лизина. Обе плазмиды были подвергнуты рестрикционному анализу. Плазмид pMV8-5-24 несет вставку 8,3 кв и pMV6-3 несет вставку 9,5 кв. Физическую карту вставки показывает фиг.1.
б) Субклонирование ДНК-фрагмента, который реконституирует экспорт лизина.
Из вставки плазмида pMV6-3, используя определенные места рестрикционных отрезков, изготовляли отдельные субклоны. Так, XhoI-SaII-фрагмент 3,7 кв, BamHI-фрагмент 2,3 кв и BamHI-фрагмент 7,2 кв лигировали с соответственно срезанным и обработанным вектором pJCI (Mol Gen Genet (1990) 220:478-480). С продуктами сшивания трансформировали непосредственно С. glutamicum NA8, трансформанты, как описано выше, проверяли на обратное получение выделения лизина, и присутствие субклонов подтверждалось получением плазмида и рестрикционным анализом. Таким способом получали штамм с плазмидом pMV2-3 как самый маленький субклон (фиг.1). Последний, способствующий экспорту лизина фрагмент содержит в качестве вставки BamHI-фрагмент 2,3 кв из pMV6-3.
с) Последовательность экспорт-гена лизина lys E и его регулятора lys G.
Последовательность нуклеотидов BamHI-фрагментов 2,3 кв осуществляли по методу дидеокси-разрыва цепи Зангера и др. (Рrос. Natl Acad Sci. США (1977) 74:5463-5467) и последовательных реакций с Auto-Read Sequencing kit фармация (Уппсала, Швеция). Электрофорезный анализ осуществляли при помощи автоматического лазерного флуоресцентного прибора для определения последовательности ДНК (A.L.F) Фармация-LKB (Piscataway, NJ, США). Полученную последовательность нуклеотидов анализировали при помощи пакета программ HUSAR (документ 3.0) Немецкого центра исследования рака (Гейдельберг). Последовательность нуклеотидов и результат анализа приведены в табл.2. Анализ дает два полных открытых растра для считывания (ORF) на отрезке с установленной последовательностью ДНК. ORFI кодирует протеин с длиной 236 аминокислот, ORF2 кодирует протеин с длиной 290 аминокислот. Произведенный от ORFI протеин показывает скопление гидрофобных аминокислот, которое характерно для мембранпостоянных протеинов. Подробный анализ распределения гидрофобных и гидрофильных аминокислот с программой PHD.HTM (Protein Science (1995) 4:521-533) показан в табл.3. Из этого следует, что протеин содержит шесть гидрофобных областей спиралей, которые пересекают мембрану. У этого протеина речь идет об искомом экспортере аминокислоты L-лизина. Поэтому соответствующий ген обозначают ниже как lysE. Он обозначен соответственно в табл.2. ORF2 транскрибируют в противоположном направлении к ORFI. Анализ последовательностей показывает, что ORF2 имеет высокую идентичность с регуляторными генами, которые объединяют как одно семейство (Ann. Rev Microbiol (1993) 597-626). Гены этого семейства положительным образом регулируют экспрессию самых различных участвующих в катабольных или анабольных процессах генов. Поэтому ниже ORF2 обозначают как lysG (Govern=регулирование). Вследствие этого подчинения и так как lysE можно было клонировать (см. а)) и субклонировать (см. б)) только вместе с lysG, lysG регулятор lysE участвует, следовательно, также и в экспорте лизина. Ген lysG и его образованная последовательность аминокислот показаны также в табл.2 или в табл.1.
d) Идентифицирование неизвестного мембранного протеина из Escherichia coli путем сравнения последовательностей.
С устойчивыми последовательностями согласно табл.3 можно искать уже существующие банки последовательностей, чтобы идентифицировать действие образованных от секвентированных областей протеинов. Соответственно сравнивали последовательность аминокислот экспортера лизина из С. glutamicum при помощи пакета программ HUSAR (документ 3.0) Немецкого центра исследования рака (Гейдельберг) с образованными последовательностями протеинов всех депонированных там ДНК-последовательностей. Для единственной последовательности неизвестной до сих пор функции из E.coli получалась высокая гомология 39,3% идентичных аминокислот и 64,9% похожих аминокислот. Сравнение показано на фиг.2. Не охарактеризованный до сих пор открытый растр считывания из E.coli идентифицирован через этот способ как экспорт-ген аминокислоты.
e) Повышенный экспорт внутриклеточно аккумулированного L-лизина.
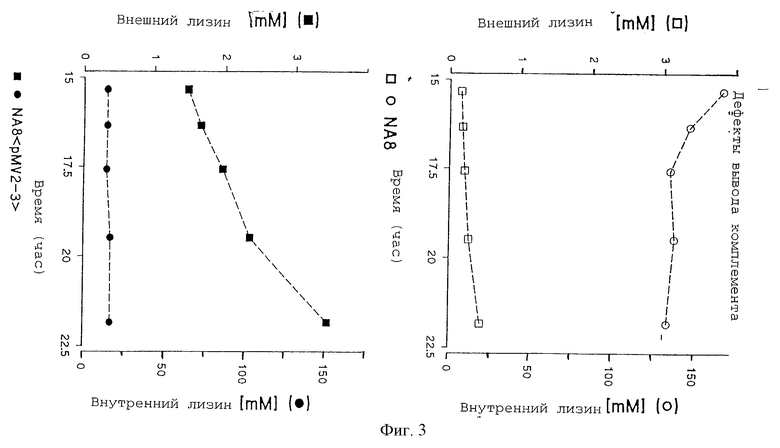
Штамм С. glutamicum NA8 (J Bacteriol (1995) 177:4021-4027) трансформировали с плазмидом pMV2-3 и сравнивали выделение L-лизина штаммов. Для этого соответственно отдельно прививали NA8 и NA8pMV2-3 в комплексной среде, описанной Vrljic и др. (J Bacteriol (1995) 177:4021-4027), и ферментационную среду CGXII (J Bacteriol (1993) 175:5595-5603). Среда содержала дополнительно 5 ммол L-метионина, чтобы индуцировать биосинтез внутриклеточного L-лизина. После культивирования в течение 24 ч при 30°С на ротационном аппарате для встряхивания при 140 об/мин проводили определения внутриклеточного и внешнего L-лизина. Для внутриклеточного определения осуществляли центрифугирования с силиконовым маслом (Methods Enzymology LV (1979) 547-567); определение аминокислот проводили при помощи жидкостной хроматографии высокого давления (J Chromat (1983) 266:471-482). Эти определения проводили в различные промежутки времени, как показано на фиг.3. В соответствии с использованным способом накопленный внутриклеточный лизин через pMV2-3 выделяется в повышенных количествах и накапливается. Как и следовало ожидать, соответственно сильно снижается также количество имеющегося внутри клеток L-лизина. Следовательно, использование открытого и описанного экспортера представляет собой способ значительно улучшить образование L-лизина.
f) Повышенное накапливание L-лизина путем lysE или lysEG.
Из субклона pMV2-3, который содержит секвентированный 2374 вр ВamHI-фрагмент в pJCI (см. фиг.1), в соответствии с информацией о последовательности, лигировали несущий lysE 1173 вр PvuII-HindII-фрагмент в pZ1 (Appl Env Microbiol (1989) 55:684-688), и получали таким способом плазмид plysE. Этот плазмид и несущий lysElysG-плазмид pMV2-3 вводили электропорацией в С. glutamicum-штамм d, причем хромосомные области подвергнуты делении. Полученные штаммы С. glutamicum d pMV2-3, С. glutamicum d, plysE, C. glutamicum pJCI выбирали, как описано в пункте е), сначала на комплексную среду, затем культивировали в производственной минимальной среде CGXII вместе с 4% глюкозы и 5 ммол L-метионина, и пробы отбирали для определения накопленного L-лизина. Как видно из фиг.4, благодаря lysElysG достигают повышения накапливания лизина по сравнению с контрольным определением. Благодаря этому способу plysE достигает чрезвычайно высокого накапливания L-лизина от 4,8 до 13,2 ммол.
Пояснения к таблицам и чертежам.
Таблица 1: Последовательность аминокислот регулятора экспортера лизина из Corynebacterium glutamicum, с типичным для ДНК-связывающих протеинов Helix-Turn-Helix Motif.
Таблица 2 (три страницы): Последовательность нуклеотидов кодирующей экспортера лизина и регулятора экспорта лизина области из С. glutamicum.
Таблица 3: Последовательность аминокислот экспортера лизина из Corynebacterium glutamicum, с идентифицированными трансмембранными спиралями TMHI-TMH6.
Фиг.1: Полученные клонированием ДНК-фрагменты в pMV6-3 и pMV8-5-24, способствующие секреции лизина, и полученный из pMV6-3 субклон pMV2-3, который также способствует секреции лизина и был секвентирован. В, BamHI; Sm, SmaI; Sc, SacI; Sl, SalI; H, HindII; X, XhoI.
Фиг.2: Сравнение образованной последовательности аминокислот LysE из С. glutamicum (выше), с ген-продуктом неизвестной до сих пор функции из Escherichia coli (ниже), который идентифицирован как экспорт-носитель.
Фиг.3: Повышенный экспорт лизина путем pMV2-3 с С. glutamicum NA8. Выше, контроль с небольшим выделением и с накапливанием внутриклеточного лизина приблизительно до 150 ммол. Ниже, вызванное благодаря pMV2-3 высокое выделение только с небольшим накапливанием внутриклеточного лизина около 30 ммол.
Фиг.4: Повышение накапливания лизина в С. glutamicum благодаря lysElysG (pMV2-3) (средняя кривая), и обусловленное lysE (plysE) накапливание (верхняя кривая).









Изобретение относится к биотехнологии. Способ микробиологического получения аминокислот включает повышение активности экспорт-носителя и/или экспрессию экспорт-гена микроорганизма, продуцирующего желаемую аминокислоту. Согласно изобретению один специфический ген несет ответственность за экспорт данной аминокислоты и основной способ микробиологического получения аминокислот, включающий контролируемое повышение экспрессии экспорт-гена и/или активности экспорт-носителя микроорганизма, продуцирующего данную аминокислоту, проявляется в первый период времени. Повышенная экспрессия или активность экспорт-носителя, получающаяся в результате этого способа, увеличивает норму выделения и ускоряет поэтому транспорт желаемой аминокислоты. 6 с. и 24 з.п.ф-лы, 4 ил., 3 табл.
| J | |||
| of Bacteriology | |||
| Oct | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| Кулисный парораспределительный механизм | 1920 |
|
SU177A1 |
| J | |||
| of Bacteriology | |||
| Jul | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| Кулисный парораспределительный механизм | 1920 |
|
SU177A1 |
| US 4430430, 07.02.1984 | |||
| US 4346170, 24.08.1982. | |||

