ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к выделению, очистке и рекомбинантному или химическому синтезу белков, которые влияют на выживаемость, пролиферацию, дифференцировку или созревание кроветворных клеток, в особенности прогениторных клеток тромбоцитов. Настоящее изобретение, в частности, относится к клонированию и экспрессии нуклеиновых кислот, кодирующих белковые лиганды, способные к связыванию и активации mpl, члена суперсемейства цитокиновых рецепторов. Настоящее изобретение далее относится к использованию этих белков самих по себе или в сочетании с другими цитокинами для лечения заболеваний кроветворения, включая тромбоцитопению.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
I. Система кроветворения
Кроветворная система производит зрелые высокоспецифичные клетки крови, как известно, необходимые для жизнедеятельности всех млекопитающих. Эти зрелые клетки включают: эритроциты, специализированные для транспорта кислорода и углекислого газа; Т- и В-лимфоциты, ответственные за клеточный и опосредованный антителами иммунные ответы; кровяные пластинки или тромбоциты, специализированные для образования кровяных тромбов; и гранулоциты и макрофаги, специализированные как ловушки или как вспомогательные клетки для борьбы с инфекцией. Гранулоциты далее подразделяются на: нейтрофилы, эозинофилы, базофилы и тучные клетки, специализированные типы клеток, обладающие дискретными функциями. Необходимо отметить, что все эти специализированные клетки происходят из одного общего примитивного типа клеток, обозначаемых как плюрипотентные (или тотипотентные) стволовые клетки, первично обнаруживаемые в костном мозге (Dexter et al., Ann. Rev. Cell Biol., 3: 423-441 [1987]).
Зрелые высокоспециализированные клетки крови должны производиться непрерывно в большом количестве в течение всей жизни млекопитающего. Подавляющему большинству этих специализированных клеток предназначено сохраняться функционально активными только в течение от нескольких часов до нескольких недель (Cronkite et al., Blood Cells, 2: 263-284 [1976]). Таким образом, непрерывное обновление зрелых клеток крови, самих примитивных стволовых клеток, так же как и любых промежуточных или предназначенных для дифференцировки прогениторных клеточных линий, лежащих между примитивными и зрелыми клетками, являются необходимыми для поддержания постоянного нормального состояния клеток крови, требующегося млекопитающему.
В центре гемапоэтической системы лежит плюрипотентная стволовая клетка(клетки). Этих клеток относительно немного по количеству и они подлежат самообновлению путем пролиферации с получением дочерних стволовых клеток или трансформируются в течение серии этапов дифференцировки в возрастающе зрелые, ограниченно дифференцированные клетки, однозначно образующие высокоспециализированную зрелую клетку(клетки) крови.
Например, определенные мультипотентные прогениторные клетки, обозначаемые как CFC-Mix, происходящие от стволовых клеток, подвергаются пролиферации (самообновлению) и развитию с получением колоний, содержащих все различные миелоидные клетки: эритроциты, нейтрофилы, мегакариоциты (предшественники тромбоцитов), макрофаги, базофилы, эозинофилы и тучные клетки. Другие прогениторные клетки лимфоидной линии дифференцировки подвергаются пролиферации и развитию в Т-клетки или В-клетки.
Помимо этого между CFC-Mix прогениторными клетками и миелоидными клетками лежат прогениторные клетки другого уровня, промежуточно комитированные по отношению к своему потомству. Эти ограниченно дифференцированные прогениторные клетки классифицируются на основании своего потомства, которое они производят. То есть известными прямыми предшественниками миелоидных клеток являются: эритроидные колониеобразующие единицы (КОЕ-Э) для эритроцитов, гранулоцит/макрофаг колониеобразующие клетки (ГМ-КОК) для нейтрофилов и макрофагов, мегакариоцит колониеобразующие клетки (Мег-КОК) для мегакариоцитов, эозинофил колоние-образующие клетки (Эоз-КОК) для эозинофилов и базофил колоние-образующие клетки (Баз-КОК) для тучных клеток. Другими клетками предшественниками, промежуточными между плюрипотентными стволовыми клетками и зрелыми клетками крови, являются известные клетки (см. выше), или клетки, которые, вероятно, будут открыты, имеющие различную степень ограниченной дифференцировки и способности к самообновлению.
Основополагающим принципом нормальной гемапоэтической клеточной системы, по-видимому, является пониженная способность к самообновлению, выражающаяся в виде потери мультипотентности и приобретения ограниченной дифференцировки и созревания. Таким образом, у одного конца спектра гемапоэтической клетки лежит плюрипотентная стволовая клетка, проявляющая способность к самообновлению и дифференцировке во все различные подлежащие специфической дифференцировке прогениторные клетки. Эта способность является основой терапии костно-мозговыми трансплантантами, где примитивные стволовые клетки восстанавливают популяцию полной гемапоэтической системы клеток. С другого конца спектра лежат высокоограниченной дифференцировки прогениторные клетки и их потомства, которые потеряли способность самообновления, но приобрели функциональные способности зрелых клеток.
Пролиферация и развитие стволовых клеток и ограниченно-дифференцированных прогениторных клеток находится под тщательным контролем различных гемапоэтических факторов роста или цитокинов. Роль этих факторов in vivo сложна и недостаточно понятна. Некоторые факторы роста, такие как интерлейкин-3 (ИЛ-3), способны стимулировать как мультипотентные стволовые клетки, так и комитированные прогениторные клетки нескольких дифференцирующихся линий, включая, например, мегакариоциты. Первоначально предполагалось, что другие факторы, такие как гранулоцит/макрофаг колоние-стимулирующий фактор (ГМ-КСФ), ограничиваются в своем действии на ГМ-КОК. Однако позднее, было обнаружено, что ГМ-КСФ также влияет на пролиферацию и развитие interalia мегакариоцитов. Таким образом, было обнаружено, что ИЛ-3 и ГМ-КСФ обладают частично перекрывающимися активностями, хотя с различающейся силой действия. Недавно было обнаружено, что интерлейкин-6 (ИЛ-6) и интерлейкин-11, хотя и не обладают заметным влиянием на Мег-колониеобразование сами по себе, действуют синергично вместе с ИЛ-3 на стимуляцию созревания мегакариоцитов (Yonemura et al., Exp. Hematol., 20:1011-1016 [1992]).
Следовательно, гемапоэтические факторы роста могут влиять на рост и дифференцировку одной или более дифференцирующихся линий, могут перекрываться с другими факторами роста во влиянии на отдельные прогениторные клеточные линии или могут действовать синергично с другими факторами.
Также вероятно, что гемапоэтические факторы роста могут проявлять свои эффекты на различных стадиях развития клетки от тотипотентной стволовой клетки, различных комитированных прогениторных клеток с ограниченной дифференцировкой, до зрелых клеток крови. Например, эритропоэтин (эпо), по-видимому, вызывает пролиферацию только зрелых эритроидных прогениторных клеток. ИЛ-3, по-видимому, проявляет свой эффект ранее, влияя на примитивные стволовые клетки и промежуточные ограниченно-дифференцированные прогениторные клетки. Другие факторы роста, такие как фактор стволовых клеток (ФСК), могут влиять на развитие даже более примитивных клеток.
Из последующего будет понятно, что новые гематопоэтические факторы роста, которые влияют на выживание, пролиферацию, дифференцировку или созревание любых клеток крови или их предшественников, были бы полезны, особенно для содействия восстановления ослабленной гематопоэтической системы, что является следствием заболевания или радиационной или химиотерапии.
II. Мегакариоцитопоэз - продукция тромбоцитов
Регуляция мегакариоцитопоэза и продукции тромбоцитов рассмотрены Mazur, Exp. Hematol., 15: 248 [1987] и Hoffman, Blood, 74: 1196-1212 [1989]. Вкратце, плюрипотентные стволовые клетки костного мозга дифференцируются в мегакариоцитарные, эритроцитарные и миелоцитарные клеточные линии. Предполагается, что существует иерархия комитированных мегакариоцитарных прогениторных клеток среди стволовых клеток и мегакариоцитов. Были идентифицированы, по крайней мере три класса мегакариоцитарных прогениторных клеток, а именно: мегакариоциты очаг-формирующей единицы (ОФЕ-МК), мегакариоциты колоние-формирующей единицы (КФЕ-МК) и мегакариоцитарные прогениторные клетки легкой плотности (МК-КФЕ-ЛП). Созревание мегакариоцитов само по себе представляет непрерывность развития, которое было разделено на стадии на основании обычных морфологических критериев. Самыми ранними распознаваемыми членами мегакариоцитарной (МК или Мег) семьи являются мегакариобласты. Эти клетки, первоначально от 20 до 30 мкм в диаметре, обладают базофильной цитоплазмой и слегка неправильным ядром с утраченным, каким-либо образом, ретикулярным хроматином и несколькими нуклеолями. Поздние мегакариоцитобласты могут содержать до 32 ядер (полиплоидия), но цитоплазма остается разреженной и незрелой. Как только наступает созревание, ядро становится более сегментированным и пикнотическим, цитоплазма увеличивается в количестве и становится более ацидофильной и гранулярной. Большинство зрелых клеток этого семейства могут дать появление высбождаемых тромбоцитов на своей периферии. Обычно менее чем 10% мегакариоцитов находятся на стадии бласта и более чем 50% являются зрелыми. По условной морфологической классификации, обычно применяемой для мегакариоцитарной серии, мегакариоцитобласт является самой ранней формой; промегакариоцит или базофильный мегакариоцит является промежуточной формой; и зрелый (ацидофильный, гранулярный или продуцирующий тромбоциты мегакариоцит) является последней формой. Зрелые мегакариоциты распространяют филаменты цитоплазмы в синусоидальное пространство, где они отрываются и фрагментируются на отдельные тромбоциты (Williams et al., Hematology, 1972).
В мегакариоцитопоэзе, как представляется, участвуют несколько регуляторных факторов (Williams et al., Br. J. Haematol., 52:173 [1982] и Williams et al., J. Cell Phisiol., 110: 101 [1982]). Ранний уровень мегакариоцитопоэза постулирован как митотический, имеющий отношение к пролиферации клетки и инициации колонии из КФЕ-МК, но не к количеству тромбоцитов (Burstein et al., J. Cell Phisiol., 109:333 [1981] и Kimura et al., Exp. Hematol., 13: 1048 [1985]). Поздняя стадия созревания является не митотической, связанной с ядерной полиплоидизацией и цитоплазматическим созреванием и, по-видимому, регулируется по механизму обратной связи, зависящему от количества периферических тромбоцитов (Odell et al.. Blood, 48: 765 [1976] и Ebbe et al., Blood, 32: 787 [1968]).
Обсуждалось существование отдельного и специфического колоние-стимулирующего фактора мегакариоцитов (МК-КСФ) (Mazur, Exp. Hematol., 15: 340-350 [1987]). Однако большинство авторов полагают, что процессы, столь жизненно важные для выживания, как продукция тромбоцитов, должны были бы регулироваться цитокином(ами), ответственным исключительно за этот процесс. Гипотеза, что существует мегакариоцит/тромбоцит специфический цитокин(ы), представляла основу для поисков в течение более чем 30 лет, но на настоящий день ни одного цитокина, подобного уникальному МК-КСФ (ТРО), не было очищено, секвенировано и установлено в исследовании.
Хотя, было сообщено, что МК-КСФ был частично очищен при экспериментально полученной тромбоцитопении (Hill et al., Exp. Hematol., 14: 752 [1986]), из кондиционированной среды почки эмбриона человека [КС] (McDonald et al., J. Lab. Clin. Med., 85: 59 [1975]), из экстрактов мочи человека с пластической анемией и идиопатической тромбоцитопенической пурпурой (Kawakita et al., Blood, 6: 556 [1983]) и из плазмы (Hoffman et al., J. Clin. Invest., 75: 1174 [1985]), в большинстве случаев их физиологическая функция до сих пор неизвестна.
В качестве потенциаторов мегакариоцитов была использована кондиционированная среда клеток селезенки, активированных митогеном из фитолакки американской (PWM-SpCM), и клеточной линии миеломоноцитов мыши WEHI-3 (WEHI-3CM). PWM-SpCM содержит факторы, усиливающие рост КФЕ-МК (Metcalf et al., Pro. Natl. Acad. Sci., USA, 72:1744-1748 [1975]; Quesenberry et al., Blood, 65:214 [1985]; Iscove, N.N., Hemapoetic Cell Differentiation, ICN-UCLA Simposia on Molecular and Cell Biology, Vol. 10, Golde et al., eds. [New York, Academy Press] pp 37-52 [1978], один из которых является интерлейкином-3 (ИЛ-3), мультидифференцирующим колоние-стимулирующим фактором (мульти-КСФ) (Burstein, Blood Cells, 11:469 [1986]). Другие факторы в этой среде еще не были определены и выделены. WEHI-3 является миеломоноцитарной клеточной линией мыши, секретирующей сравнительно большое количество ИЛ-3 и небольшое количество ГМ-КСФ. Было обнаружено, что ИЛ-3 потенцирует рост широкого круга гемапоэтических клеток (Ihle et al., J. Immunol., 13:282 [1983]). Было также обнаружено, что ИЛ-3 обладает синергизмом со многими известными гематопоэтическими гормонами или факторами роста (Barteliaez et al., J. Cell Physiol., 122:362-369 [1985] и Warren et al., Cell, 46:667-674 [1988]), включая и эритропоэтин (ЭП), и интерлейкин-1 (ИЛ-1), в индукции самых ранних мультипотентных предшественников и в образовании очень больших смешанных гематопоэтических колоний.
В кондиционированной среде легких, кости, клеточных линиях макрофагов, перитонеальных экссудатных клетках мыши и в эмбриональных клетках почки человека были обнаружены другие источники потенциаторов мегакариоцитов. Несмотря на некоторые противоречивые данные (Mazur, Exp. Hematol., 15:340-350 [1987]), существует доказательство (Geissler et al., Br.J.Haematol., 60: 233-238 [1985]), что скорее активированные Т-лимфоциты, чем моноциты играют усиливающую роль в мегакариоцитопоэзе. Эти данные свидетельствуют о том, что секреты активированных Т-лимфоцитов, такие как интерлейкины, могут быть регуляторными факторами в развитии МК (Geissler et al., Exp. Hematol., 15: 845-853 [1987]). Род исследований мегакариоцитопоэза при использовании очищенного эритропоэтина ЭПО (Vainchenker et al., Blood, 54:940 [1979]; McLeod et al., Nature, 261: 492-4 [1976]; Williams et al., Exp. Hematol., 12: 734 [1984]) указывает, что этот гормон обладает усиливающим действием на образование колонии МК. Это было также показано и на свободной от сыворотки, и на содержащей сыворотку культурах, и в отсутствии вспомогательных клеток (Williams et al., Exp. Hematol., 12: 734 [1984]). Было постулировано, что ЭПО участвует в большей степени на одно- или двух клеточной стадии мегакариоцитопоэза в противоположность к действию PWM-SрСМ, который участвует на четырехклеточной стадии развития мегакариоцита. Взаимодействие этих факторов на обеих, ранней и поздней, фазах развития мегакариоцита остается требующим разъяснения.
Из данных, полученных в нескольких лабораториях, предполагается, что только мультидифференцирующие факторы, которыми являются ГМ-КСФ и ИЛ-3, и, в меньшей степени, ИЛ-6, стимулирующий фактор В-клеток, обладают индивидуально МК-колоние-стимулирующей активностью, (Ikebuchi et al., Proc. Natl. Acad. Sci. USA, 84:9035 [1987]). Совсем недавно несколько авторов доложили, что ИЛ-11 и фактор ингибирующий лейкемию (ФИЛ) действуют синергично вместе с ИЛ-3 на увеличение размеров мегакариоцитов и плоидию (Yonemura et al., British Journal of Hematology, 84:16-23 [1993]; Burstein et al., J. Cell. Physiol., 153:305-312 [1992]; Metcalf et al.. Blood, 76:50-56 [1990]; Metcalf et al.. Blood, 77:2150-2153 [1991]; Bruno et al., Exp. Hematol., 19:378-381 [1991]; Yonemura et al., Exp. Hematol., 20:1011-1016 [1992]).
Другие материалы, представляющие интерес, включают: Eppstein et al., US Patent No. 4962091; Chong, US Patent No. 4879111; Fernandes et al., U.S. Patent No. 4604377; Wissler et al., US Patent No. 4512971; Gottlieb, US Patent No. 4468379; Bennet et al., US Patent No. 5215895; Kogan et al., US Patent No. 5250732; Kimura et al., Eur. J. Immunol., 20(9): 1927-1931 [1990]; Secor et al., J. of Immunol., 144 (4): 1484-1489 [1990]; Warren et al., J. of Immunol., 140(1): 94-99 [1988]; Warren et al., Exp. Hematol., 17(11): 1095-1099 [1989]; Bruno et al., Exp. Hematol., 17(10): 1038-1043 [1989]; Tanikawa et al., Exp. Hematol., 17 (8): 883-888 [1989]; Koike et al., Blood, 75 (12): 2286-2291 [1990]; Lotem, Blood, 75(5): 1545-1551 [1989]; Rennik et al., Blood, 73(7): 1828-1835 [1989]; и Clutterbuck et al.. Blood, 73(6): 1504-1512 [1989].
III. Тромбоцитопения
Тромбоциты являются ключевым элементом в механизме свертывания крови. Обеднение уровня циркулирующих тромбоцитов, называемое тромбоцитопения, наблюдается при различных клинических состояниях и заболеваниях. Тромбоцитопению обычно определяют, когда содержание тромбоцитов ниже 150×109 на литр. Большинство причин заболевания тромбоцитопенией может быть подразделено на три категории на основании продолжительности жизни тромбоцитов, а именно: (1) нарушенная продукция тромбоцитов костным мозгом, (2) секвестрация тромбоцитов в селезенке (сплиномегалия), или (3) увеличенное разрушение тромбоцитов в периферическом круге кровообращения (т.е. аутоиммунная тромбоцитопения или химио- и радиационная терапия). Кроме того, у пациентов, получающих большие объемы быстро вводимых обедненных тромбоцитами продуктов крови, тромбоцитопения может развиться из-за разведения.
Клиническое проявление кровотечения при тромбоцитопении зависит от тяжести тромбоцитопении, ее причины и возможной взаимосвязи с нарушениями свертываемости. В общем, пациенты со счетом тромбоцитов между 20 и 100×109 на литр имеют риск усиленного посттравматического кровотечения, когда этот счет тромбоцитов ниже 20×109 на литр, может быть спонтанное кровотечение. Эти последние пациенты являются кандидатами для трансфузии тромбоцитов, сопровождающуюся иммунным или вирусным риском. Для любых данных степеней тромбоцитопении кровотечение имеет тенденцию быть более тяжелым, когда причиной является пониженная продукция, чем при увеличенном распаде тромбоцитов. В последней ситуации ускоренный оборот тромбоцитов приводит к появлению в круге циркуляции более молодых, больших и гемастатически более эффективных тромбоцитов. Тромбоцитопения может быть следствием различных нарушений, кратко описанных ниже. Более детальное описание может быть найдено у Schafner, A.I., Thrombocytopenia and Disorders of Platelet Function, Internal Medicine, 3rd Ed., John J. Hutton et al., Eds., Little Brown and Co., Boston/Toronto/London [1990].
(а) Тромбоцитопения вследствие пониженной продукции тромбоцитов
Причины врожденной тромбоцитопении включают конституциональную апластическую анемию (синдром Фанкони) и врожденную амегакариоцитарную тромбоцитопению, которая может быть связана с пороком развития скелета. Приобретенные нарушения продукции тромбоцитов являются вызванными или гипоплазией мегакариоцитов, или неэффективным тромбопоэзом. Мегакариоцитарная гипоплазия может быть следствием различных состояний, включая костную аплазию (включая идиопатические формы или миелосупрессию химиотерапевтическими агентами или радиационной терапией), миелофиброз, лейкемия и инвазия в костный мозг опухоли или гранулемы. В некоторых ситуациях токсины, инфекционные агенты или лекарственные препараты могут влиять на тромбоцитопению относительно избирательно; примеры включают перемежающуюся тромбоцитопению, вызванную алкоголем или определенными вирусными инфекциями, и умеренную тромбоцитопению, связанную с введением тиазидовых диуретиков. Наконец, неэффективный тромбоцитопоэз, вторичный по отношению к мегалобластическим процессам (дефицит фолиевой кислоты или B12), может также вызывать тромбоцитопению, обычно сопровождающуюся анемией и лейкопенией.
Текущее лечение тромбоцитопениоза, вызванного сниженной продукцией тромбоцитов, зависит от определения и обращения, определяющей причины недостаточности костного мозга. Вливания тромбоцитов обычно резервируются для пациентов с серьезными осложнениями с кровотечениями или для восполнения при хирургических операциях, поскольку изоиммунизация может привести далее к рефрактерности к вливаниям тромбоцитов. Кровотечения слизистой, проявляющиеся при тяжелой тромбоцитопении, могут быть уменьшены путем перорального или внутривенного введения антифибринолитических агентов. Однако тромботические осложнения могут развиваться, если применяются антифибринолитические агенты у пациентов с диссиминированными внутривенными коагуляциями (ДВК).
(b) Тромбоцитопения вследствие захвата в селезенке
Увеличение селезенки, возникшее вследствие любой причины, может сопровождаться мягкой, до умеренной тромбоцитопенией. Это в значительной степени пассивный процесс (гиперспленизм) захвата тромбоцитов, в противоположность активному разрушению тромбоцитов селезенкой в результате иммуноопосредованной тромбоцитопении, обсужденной ниже. Хотя самой распространенной причиной гиперспленизма является застойное увеличение селезенки из-за гипертонии воротной вены, вследствие алкогольного цирроза, другие формы застоя, инфильтрационная или лимфопролиферативная спленомегалия также связаны с тромбоцитопенией. Счет тромбоцитов в результате только гиперспленизма в целом не падает ниже 50×109 на литр.
(c) Тромбоцитопения вследствие неиммуноопосредованного разрушения тромбоцитов
Тромбоцитопения может быть следствием ускорения разрушения тромбоцитов при различных неиммунологических процессах. Нарушения этого типа включают диссиминированную внутривенную коагуляцию, внутрисосудистые протезные приспособления, экстракорпоральную циркуляцию крови, тромботические микроангиопатии, такие как тромботическая тромбоцитарная пурпура. Во всех этих ситуациях циркулирующие тромбоциты, которые задерживаются либо на искусственной поверхности, либо на нарушенной интиме сосуда, либо утилизируются на этих участках, либо разрушаются и затем преждевременно очищаются ретикуло-эндотелиальной системой. Заболевание или нарушение, при котором может возникать диссиминированная внутрисосудистая коагуляция (ДВК), изложено в деталях Braunwald et al. (eds), Harrison’s Priciples of Internal Medicine, 11th Ed., p.1478, McGrow Hill [1987]. Внутривенные протезные приспособления, включая сердечные клапаны и внутриаортальные баллоны, могут быть причиной мягкой, до умеренной, деструктивной тромбоцитопении и перемежающейся тромбоцитопении; у пациентов, подлежащих сердечно-легочному шунтированию или гемодиализу, могут быть следствием утилизации или повреждения тромбоцитов при экстракорпоральном кровообращении.
(d) Иммунная тромбоцитопения, индуцированная лекарственными препаратами
Более чем 100 лекарственных препаратов вовлечены в иммунологически опосредованную тромбоцитопению. Охарактеризованы, однако, только хинидин, хинин, золото, сульфонамиды, цефалотины и гепарин. Вызванная лекарственными препаратами тромбоцитопения часто является очень тяжелой и обычно наблюдается стремительно в течение нескольких дней, пока пациент принимает предрасполагающую медикацию.
(e) Иммунная (аутоиммунная) тромбоцитопеническая пурпура (ИТП)
ИТП у взрослых является хроническим заболеванием, характеризующимся аутоиммунным разрушением тромбоцитов. Аутоантитела являются обычно IgG, хотя было доложено и о других иммуноглобулинах. Несмотря на то, что обнаруженные при ИТП аутоантитела связаны на мембране тромбоцитов с GPIIbIIIa, в большинстве случаев специфичность антигена тромбоцитов не идентифицирована. В ретикулоэндотелиальной системе селезенки и печени наблюдается внесосудистая деструкция сенсибилизированных тромбоцитов. Хотя более чем половина всех случаев ИТП является идиопатической, многие пациенты заболевают ревматическим или аутоиммунным заболеваниями (например, эритроматозной системной волчанкой), или лимфопролиферативными заболеваниями (например, хронической лимфоцитарной лейкемией).
(f) ВИЧ-индуцированная ИТП
ИТП является всевозрастающим обычным осложнением при инфекции ВИЧ (Morris et al., Ann. Intern. Med., 96: 714-717 [1982]) и может наблюдаться на различных стадиях развития заболевания и у пациентов с диагносцированным синдромом приобретенного иммунодефицита (СПИД), у которых имеется СПИД-зависимый комплекс, и у инфицированных ВИЧ, но без синдрома СПИД. ВИЧ инфекция является передаваемым заболеванием, однозначно характеризующимся выраженным дефицитом клеточной иммунной функции, так же как и сопровождающимися сопутствующими инфекциями и злокачественными новообразованиями. Первичные иммунологические нарушения, являющиеся следствием ВИЧ инфекции, - это прогрессирующее уменьшение и функциональная ослабленность Т-лимфоцитов, экспрессирующих клеточный поверхностный гликопротеин CD4 (Lane et al., Ann. Rev. Immunol., 3: 477 [1985]). Потеря CD4 хелперной/индукторной Т-клеточной функции, очевидно, подчеркивает глубокий дефект в клеточном и гуморальном иммунитете, приводящий к сопутствующим инфекциям и злокачественности, характеризующей СПИД (Н. Lane, выше). Хотя механизм ИТП, связанной со СПИДом, неизвестен, представляется, что он отличен от механизма ИТП, не ассоциированной с ВИЧ инфекцией. (Walsh et al., N. Eng. J. Med., 311: 635-639 [1984]; и Ratner, Am. J. Med., 86: 194-198 [1989]).
IV. Современная терапия тромбоцитопении
Терапевтические подходы к лечению пациентов с тромбоцитопенией определяются тяжестью и быстротой клинической ситуации. Лечение одинаково для ВИЧ-зависимой и не зависисмой от ВИЧ тромбоцитопении, и хотя применяется множество терапевтических подходов, терапия остается противоречивой.
Счет тромбоцитоов у пациентов с диагносцированной тромбоцитопенией успешно увеличивается под воздействием глюкокортикоидной (например, преднизолона) терапии, однако у большинства пациентов вылечивание неполное или наблюдаются рецидивы, когда дозировка глюкокортикоида снижается или лечение прерывается. Опираясь на пациентов, имеющих ВИЧ-связанную ИТП, некоторые исследователи предположили, что терапия глюкокортикоидами может привести к предрасположенности к СПИДу. Глюкокортикоиды обычно используются, если счет тромбоцитов падает ниже 20×109/литр или когда наблюдаются спонтанные кровотечения.
Для пациентов с рефрактерностью к глюкокортикоидам успешно применяли соединение 4-(2-хлорфенил)-9-метил-2-[3-(морфолинил)-3-пропанон-1-ил]6Н-тиено[3,2,f][1,2,4]триазоло-[4,3,а][1,4]диазепин (WEB 2086) для лечения тяжелых случаев ИТП, не связанной с ВИЧ. Пациентов, имеющих счет тромбоцитов 37000 - 58000/ мкл, лечили WEB 2086 и через две недели лечения счет тромбоцитов увеличился до 14000-190000/ мкл. (ЕР 361077 и Lohman et al., Lancet, 1147 [1988]).
Хотя оптимальное лечение приобретенной амегакариоцитарной тромбоцитопенической пурпуры (ПАТП) не определено, было показано, что антитимоцитарный глобулин (АТГ) лошадиной антисыворотки к ткани тимуса человека дает продолжительную полную ремиссию (Trimble et al., Am. J. Hematol., 37: 126-127 [1991]). Однако в недавней публикации отмечают, что гематопоэтические эффекты АТГ могут быть отнесены к тимеросалу, который действует преимущественно на белки как несущий ртуть (Panella et al., Cancer Research, 50: 4429-4435 [1990]).
Было сообщено о хороших результатах по спенэктомии. При спенэктомиии удаляется основной участок разрушения тромбоцитов и основной источник продукции аутоантител у многих пациентов. Эта операция приводит к длительной, не требующей лечения ремисси у большого количества пациентов. Однако поскольку хирургическое вмешательство, в общем, у иммунокомпромисных пациентов избегается, спленэктомия рекомендуется только в тяжелых случаях тромбоцитопении (например, при тяжелой ВИЧ-зависимой ИТП), у пациентов, которые не способны отвечать в течение 2-3 недель на лечение глюкокортикоидами, или при не достижении длительного ответа, после прерывания лечения глюкокортикоидами. Основываясь на современных научных данных, не ясно, предрасполагает ли спленэктомия пациентов к СПИДу.
Помимо преднизолоновой терапии и спленэктомиии определенные цитотоксические агенты, а именно винкристин и азидотимидин (AST, zid.ovud.ine), также проявляют обещания при лечении ИТП, индуцированной ВИЧ; однако эти результаты являются предварительными.
Из последующего будет понятно, что одним из путей лечения тромбоцитопении могло бы быть получение агента, способного ускорять дифференциацию и созревание мегакариоцитов или их предшественников в формах, продуцирующих тромбоциты. Были потрачены значительные усилия по поиску такого агента, называемого обычно как "тромбопоэтин" (ТП). Другие названия ТП, обычно встречаемые в литературе, включают: фактор, стимулирующий тромбоцитопоэз (ФСТ); колоние-стимулирующий фактор мегакариоцитов (МК-КСФ); мегакариоцит-стимулирующий фактор и потенциатор мегакариоцитов. Активность ТП обнаружили достаточно рано, уже в 1959 г. (Rak et al., Med. Exp., 1: 125), но и по настоящее время продолжаются попытки охарактеризовать и очистить этот агент. Несмотря на существующие данные по частичной очистке ТП-активного полипептида (см., например, Tayrien et al., J. Biol. Chem., 262: 3262 [1987] и Hoffman et al., J. Clin. Invest. 75: 1174 [1985]), некоторые авторы постулировали, что ТП по своей сути не является отдельно существующей реальностью, но скорее это просто полифункциональное проявление известных гормонов (ИЛ-3, Sparrow et al.. Prog. Clin. Biol. Res., 215: 123 [1986]). Независимо от ее формы или природы молекула, проявляющая тромбопоэтическую активность, представляла бы значительную терапевтическую ценность. Хотя ни один белок не был однозначно идентифицирован как ТП, значительным интересом окружено недавнее открытие того, что mpl, естественный рецептор цитокинов, может опосредовать тромбопоэтический сигнал.
V. Mpl является мегакариоцитопоэтическим рецептром цитокинов
Предполагается, что пролиферация и созревание гематопоэтических клеток тесно регулируются факторами, которые положительно или отрицательно модулируют пролиферацию и дифференцировку полидифференцирующихся плюрипотентных стволовых клеток. Эти эффекты опосредуются путем высокоаффинного связывания внеклеточного белкового фактора со специфическими поверхностными клеточными рецепторами. Эти поверхностные рецепторы проявляют определенную гомологию и обычно классифицируются как члены суперсемейства цитокиновых рецепторов. Члены этого семейства включают рецепторы для: ИЛ-2 (β и γ-цепи) (Hatakeayma et al., Science, 244: 551-556 [1989]; Takeshita et al., Science, 257: 379-382 [1991]), ИЛ-3 (Itoh et al.. Science, 247: 324-328 [1990]; Gorman et al., Proc. Natl. Accad. Sci. USA, 87: 5459-5463 [1990]; Kitamura et al., Cell, 66: 1165-1174 [1991a]; Kitamura et al., Proc. Natl. Acad. Sci. USA, 88: 5082-5086 [1991b]), ИЛ-4 (Mosley et al., Cell, 59: 335-348 [1989]), ИЛ-5 (Takaki et al., EMBO J., 9: 4367-4374 [1990]; Tavernier et al.. Cell, 66: 1175-1184 [1991]), ИЛ-6 (Yamasaki et al., Science, 241: 825-828 [1988]; Hibi et al., Cell, 63:1149-1157 [1990]), ИЛ-7 (Goodwin et al., Cell, 60: 941-951 [1990]), ИЛ-9 (Renault et al., Proc. Natl. Acad. Sci. USA, 89: 5690-5694 [1992]), гранулоцит-макрофаг колоние-стимулирующий фактор (ГМ-КСФ) (Gearing et al., EMBO J., 8: 3667-3676 [1991]; Hayashida et al., Proc. Natl. Acad. Sci. USA, 244: 9655-9659 [1990]), гранулоцит колоние-стимулирующий фактор (Г-КСФ) (Fukunaga et al., Cell, 61: 341-350 [1990а]; Fukunaga et al., Proc. Natl. Acad. Sci. USA, 87: 8702-8706 [1990b]; Larsen et al., J. Exp. Med., 172: 1559-1570 [1990]), ЭП (D’Andrea et al.. Cell, 57: 277-285 [1989]; Jones et al., Blood, 76: 31-35 [1990]), фактор ингибирования лейкемии (ФИЛ) (Gearing et al., EMBO J., 10: 2839-2848 [1991]), онкостатин М (ОСМ) (Rose et al., Proc. Natl. Acad. Sci. USA, 88: 8641-8645 [1991]) и также рецепторы пролактина (Boutin et al., Proc. Natl. Acad. Sci. USA, 88: 7744-7748 [1988]; Edery et al., Proc. Natl. Acad. Sci. USA, 86: 2112-2116 [1989]), гормона роста (ГР) (Leung et al.. Nature, 330: 537-543 [1987]) и цилиарный нейротропный фактор (ЦНТФ) (Davis et al., Science, 253: 59-63 [1991]).
Члены суперсемейства цитокиновых рецепторов могут быть сгруппированы в три функциональные категории (для обзора см. Nicola et al., Cell, 67:1-4 [1991]). Первый класс включает рецепторы с одной цепью, такие как рецептор эритропоэтина (ЭП-Р) или рецептор колоние-стимулирующего фактора гранулоцитов (Г-КСФ-Р), которые связывают лиганд с высоким сродством благодаря внеклеточному домену, а также генерируют внутриклеточный сигнал. Второй класс рецепторов, так называемых α-субъединичных, включает рецептор интерлейкина-6 (ИЛ-6-Р), рецептор гранулоцит-макрофаг колоние-стимулирующего фактора (ГМ-КСФ-Р), рецептор интерлейкина-3 (ИЛ-3-Рα) и других членов суперсемейства цитокиновых рецепторов. Эти α-субъединицы связывают лиганды с низким сродством, но не способны передавать внутриклеточный сигнал. Высокоаффинные рецепторы способны к передаче сигнала, генерированного гетеродимером, между α-субъединицей и членом третьего класса цитокиновых рецепторов, обозначенного β-субъединицей, а именно βс, общей β-субъединицей для трех α-субъединиц ИЛ-3-Рα и ГМ-КСФ-Р.
Доказательство того, что mpl является членом суперсемейства цитокиновых рецепторов вытекает из гомологии последовательности (Gearing, EMBO J., 8: 3667-3676 [1988]; Bazan, Proc. Natl. Acad. Sci. USA, 87: 6834-6938 [1990]; Davis et al., Science, 253: 59-63 [1991] и Vigon et al., Proc. Natl. Acad. Sci. USA, 89: 5640-5644 [1992]) и его способности передавать пролиферативные сигналы.
Выведенная последовательность белка при молекулярном клонировании c-mpl мыши обнаруживает, что этот белок является гомологичным к другим цитокиновым рецепторам. Внутриклеточный домен содержит 465 аминокислотных остатков и составляет два субдомена, каждый с четырьмя высококонсервативными цистеинами, и конкретную структуру в N-терминальном субдомене и в С-терминальном субдомене. Лиганд-связывающий внеклеточный субдомен предположительно должен иметь складчатую структурную геометрию, подобную двойной β-бочонкообразной. Этот дублированный внеклеточный домен является высокогомологичным сигнальной передающей цепи, общей для ИЛ-3, ИЛ-5 и ГМ-КСФ рецепторов, так же как и низкоаффинному связывающему домену ФИЛ (Vigon et al., Oncogene, 8:2607-2615 [1993]). Таким образом, mpl может принадлежать к низкоаффинному лиганд-связывющему классу цитокиновых рецепторов.
Сравнение mpl мыши и зрелого mpl P человека обнаружило, что эти два белка проявляют 81% идентичности в последовательности. Более специфические N-концевой и С-концевой внеклеточные субдомены проявляют 75 и 80% идентичности в последовательности соответственно. Наиболее консервативным регионом является цитоплазматический домен, проявляющий 91% идентичности аминокислот, с последовательностью из 37 аминокислот, полностью идентичных в области около трансмембранного домена у обоих видов. Таким образом, сообщается, что mpl является одним из наиболее консервативных членов суперсемейства цитокиновых рецепторов (Vigon, там же).
Доказательство того, что mpl является функциональным рецептором, способным к передаче пролиферативного сигнала, вытекает из конструкции химерного рецептора, содержащего внеклеточный домен из цитокинового рецептора, имеющего высокое сродство к известным цитокинам, вместе с цитоплазматическим доменом mpl. Поскольку не доложено ни об одном известном лиганде к mpl, было необходимым сконструировать химерный высокоаффинный лиганд-связывающий внеклеточный домен из класса цитокиновых рецепторов, таких как ИЛ-4-Р или Г-КСФ-Р. Vigon et al., там же, соединил внеклеточный домен Г-КСФ-Р с обоими трансмембранным и цитоплазматическим доменами c-mpl. ИЛ-3 зависимая клеточная линия BAF/B03 (Ba/F3), была трансфецирована химерой Г-КСФ-P/mpl вместе с контролем Г-КСФ-Р полной длины. Клетки, трансфецированные химерой, росли одинаково хорошо в присутствии цитокина ИЛ-3 или Г-КСФ. Подобным образом клетки, трансфецированные Г-КСФ-Р также хорошо росли и в присутствии ИЛ-3, и Г-КСФ. Все клетки гибли в отсутствии факторов роста. Подобные эксперименты были проведены Skoda et al., EMBO J., 12(7): 2645-2653 [1993], в которых оба, внеклеточный и трансмембранный домены рецептора ИЛ-4 человека (РЧ-ИЛ-4), были слиты в цитоплазматический домен mpl мыши, и трансфецированы в ИЛ-3-зависимую Ba/F3 клеточную линию мыши. Клетки Ba/F3, трансфецированные диким типом РЧ-ИЛ-4, нормально делились в присутствии видоспецифического либо ИЛ-4, либо ИЛ-3. Ba/F3 клетки, трансфецированные РЧ-ИЛ-4/mpl, нормально делились в присутствии РЧ-ИЛ-4 (в присутствии или отсутствии ИЛ-3), показывая, что у Ba/F3 клеток mpl цитоплазматический домен содержит все элементы, необходимые для передачи пролиферативного сигнала.
Эти эксперименты с химерами показывают способность к проведению пролиферативного сигнала у цитоплазматического домена mpl, но молчащего, не обращающего внимания, может ли связать лиганд внеклеточный домен mpl. Эти данные включают, по крайней мере, две возможности, а именно, mpl представляет рецептор с одной цепью (первый класс), подобный ЭП-Р или Г-КСФ-Р, или представляет сигнал-передающую β-субъединицу (третий класс), требующую α-субъединицу, подобно ИЛ-3 Skoda et al., там же).
VI. Лиганд mpl является тромбопоэтином (ТП)
Как описано выше, было предположено, что сыворотка содержит уникальный фактор, называемый иногда как тромбопоэтин (ТП), который действует синергично с различными другими цитокинами по стимуляции роста и созревания мегакариоцитов. Несмотря на значительные усилия, приложенные многочисленными группами, никакого подобного природного фактора не было выделено из сыворотки или любого другого источника. Несмотря на то, что неизвестно, способен ли mpl непосредственно связывать мегакариоцит-стимулирующий фактор, недавние эксперименты показывают, что mpl участвует в передаче пролиферативного сигнала фактора или факторов в сыворотке пациентов с апластическим костным мозгом (Mathia et al., Blood, 82(5):1395-1401 [1993]).
Доказательство того, что уникальный колониеобразующий фактор, отличный от ИЛ-1α, ИЛ-3, ИЛ-4, ИЛ-6, ИЛ-11, СКФ, ЭП, Г-КСФ и ГМ-КСФ, передает пролиферативный сигнал через mpl, вытекает из исследования распределения экспресии с-mpl у примитивных и комитированных гематопоэтических клеточных линий, и из антисенсорных (с использованием нуклеотидов с обратной последовательностью) исследований на одной из этих клеточных линий.
Используя обратную транскриптазу (RT)-PCR в иммуноочищенных гематопоэтических клетках человека, Mathia et al., там же, показал, что сильный сигнал мРНК mpl был обнаружен только в CD34+ очищенных клетках, мегакариоцитах и тромбоцитах.
CD34+, очищенные от костного мозга (КМ) клетки, представляют около 1% всех клеток КМ и обогащены примитивными и комитированными прогениторными клетками всех дифференцирующихся линий (а именно, эритроидными, грануломакрофагами и мегакариоцитами).
Было показано, что олигодезоксинуклеотиды mpl с обратной последовательностью подавляют образование колоний мегакариоцитов из плюрипотентных CD34+, культивируемых на сыворотке пациентов с апластическим костным мозгом (богатый источник колоние-стимулирующей активности мегакариоцитов [МК-КСА]). Те же самые олигодезоксинуклеотиды с обратной последовательностью не обладали действием на образование эритроидных или грануломакрофагальных колоний.
Непосредственно ли связывает mpl лиганд и действует ли фактор сыворотки, вызывающий мегакариоцитопоэз, через mpl, до сих пор неизвестно. Было предположено, однако, что, если mpl действует непосредственно, связывая лиганд, его аминокислотная последовательность была бы подобна высококонсервативной и имеются виды обладающие перекрестной реактивностью с определенной идентичной последовательностью между внеклеточными доменами mpl человека и мыши (Vigon et al., там же [1993]).
VII. Предметы изобретения
С точки зрения последующего будет понятно, что существует настоящая и постоянная потребность в способе выделения и идентификации молекул, способных стимулировать пролиферацию, дифференциацию и созревание гематопоэтичеких клеток, в особенности, мегакариоцитов или их предшественников для терапевтического использования при лечении тромбоцитопении. По-видимому, такая молекула представляет лиганд mpl и, следовательно, существует необходимость для дальнейшего выделения такого лиганда (лигандов), для оценки его роли (ролей) для роста и дифференцировки клеток.
Соответственно предметом настоящего изобретения является получение фармацевтически чистой молекулы, способной стимулировать деление, дифференциацию и/или созревание мегакариоцитов в зрелую форму, продуцирующую тромбоциты.
Другим предметом является получение молекулы в виде, пригодном для терапевтического использования при лечении гематопоэтических нарушений, в особенности тромбоцитопении.
Далее, предметом настоящего изобретения является выделение, очистка и специфическое определение лигандов, способных к связыванию in vivo с рецептором суперсемейства цитокинов, известному как mpl, для передачи пролиферативного сигнала.
Еще другим предметом является получение молекул нуклеиновых кислот, кодирующих подобные белковые лиганды, и использование этих молекул нуклеиновых кислот для производства mpl связывающих лигандов в рекомбинантных культурах клеток, для диагностики и терапевтического использования.
Еще одним предметом является получение производных и модифицированных видов белковых лигандов, включая варианты аминокислотной последовательности, варианты типов гликопротеидов и их ковалентные производные.
Дополнительным предметом является получение сплавленных видов полипептидов, объединяющих лиганд mpl и гетерологический белок, и их ковалентные производные.
Еще дополнительным предметом является получение варианта вида полипептида, сочетающего лиганд mpl с аминокислотными добавлениями и заменами из последовательности ЭП для получения белка, способного регулировать деление и рост обеих прогениторных клеток тромбоцитов и красных кровяных клеток.
Еще одним дополнительным предметом является приготовление иммуногенов для увеличения антител против лигандов mpl или их объединенных видов, так же, как и получение антител, способных к связыванию подобных лигандов.
Эти и другие предметы изобретения будут очевидны для обычного работника при рассмотрении деталей в целом.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Предметы изобретения получены путем получения выделенного белка млекопитающих, стимулирующего мегакариоцитопоэтическую пролиферацию и созревание, обозначаемого "mpl лиганд" (МЛ) или "тромбопоэтин" (ТП), способного стимулировать пролиферацию, созревание и/или дифференциацию мегакариоцитов в зрелую форму, продуцирующую тромбоциты.
Этот существенно гомогенный белок может быть очищен из природного источника способом, включающим: (1) контактирование источника плазмы, содержащего молекулу лиганда mpl для очистки на иммобилизованном рецепторном полипептиде, специфического mpl или mpl слитого белка, иммобилизованного на подложке, в условиях, когда очищаемая молекула лиганда mpl селективно адсорбируется на иммобилизованном полипептидном рецепторе, (2) промывание иммобилизованного рецепторного полипептида и его подложки для удаления неадсорбированного материала и (3) смывание молекулы mpl лиганда с иммобилизованного рецепторного полипептида, к которому он был адсорбирован, буфером элюции. Природным источником, предпочтительно, является плазма млекопитающих или моча, содержащая лиганд mpl. Млекопитающее необязательно является апластическим, и иммобилизованный рецептор представляет собой слияние mpl и IgG.
Предпочтительным белком, стимулирующим мегакариоцитопоэтическую пролиферацию и созревание, необязательно, является выделенный, существенно гомогенный лиганд mpl полипептид, полученный синтетическим или рекомбинантным способом.
Полипептидный "mpl лиганд" или "ТП" по настоящему изобретению, предпочтительно, имеет, по крайней мере, более 70% идентичной последовательности по отношению к аминокислотной последовательности высокоочищенного, существенно гомогенного полипептидного лиганда mpl свиньи и, по крайней мере, 80% идентичной последовательности по отношению к "ЭП-домену" полипептидного лиганда mpl свиньи. Выборочно лигандом mpl настоящего изобретения является зрелый лиганд mpl человека (МЛч), обладающий зрелой аминокислотной последовательностью, представленной на фиг.1 (Посл. No.1), или вариант, или его посттранскрипционная модифицированная форма, или белок, имеющий около 80% идентичной последовательности по отношению к зрелому лиганду mpl человека. Необязательно, вариантом лиганда mpl является фрагмент, особенно амино-конец (N-конец), или фрагмент "ЭП-домена" зрелого лиганда mpl человека (МЛч). Предпочтительно, фрагментом амино-конца остается существенно вся последовательность МЛ человека между первым и четвертым остатками цистеина, но могущая содержать существенные вставки, делеции или замены вне этого региона. В соответствии с этим осуществлением фрагмент полипептида может быть представлен формулой:
Х-МЛч(7-151)-Y
где МЛч(7-151) представляет аминокислотную последовательность ТП человека (ТПч) от Цис7 до Цис151 включительно; Х представляет аминогруппу Цис7 или одну или более амино концевого(ых) аминокислотного(ых) остатка(ов) зрелого МЛч, или аминокислотный остаток, распространяющийся, кроме того, на такие, как Мет, Тир или лидерную последовательность, содержащую, например, места для протеолитического расщепления (а именно, Фактор Ха или тромбина); и Y представляет карбокси концевую группу Цис151 или одну или более концевого(ых) аминокислотного(ых) остатка(ов) зрелого МЛч, или, помимо этого, их расширение.
Полипептидный лиганд mpl или его фрагмент необязательно могут быть слиты в гетерологический полипептид (химеру). Предпочтительным гетерологическим полипептидом является цитокин, колоние-стимулирующий фактор или интерлейкин или его фрагмент, в особенности кит-лиганд (КЛ), ИЛ-1, ИЛ-3, ИЛ-6, ИЛ-11, ЭП, ГМ-КСФ или ФИЛ. Предпочтительным выборочным гетерологическим полипептидом является цепь иммуноглобулина, в особенности IgG1 человека, IgG2, IgG3, IgG4, IgE, IgD, IgM или их фрагменты, в особенности, включающие домены тяжелой цепи IgG.
Другим аспектом настоящего изобретения является получение композиции, включающий выделенный агонист mpl, обладающий биологической активностью и, предпочтительно, способный стимулировать включение меченых нуклеотидов (а, именно, 3H-тимидина) в ДНК ИЛ-3 зависимых Ba/F3 клеток, трансфецированных mpl человека. Агонист mpl необязательно является биологически активным лигандом mpl, но предпочтительной является его способность стимулировать включение 35S в циркулирующие тромбоциты в исследовании восстановления тромбоцитов мыши.
Подходящие mpl агонисты включают МЛч153, МЛч(Р153А, Р154А), МЛч2, МЛч3, МЛч4, МЛм, МЛм2, МЛм3, МЛп и МЛп2 или их фрагменты.
В другом воплощении настоящее изобретение описывает выделение антитела, способного связываться с лигандом mpl. Выделенное антитело, способное связываться с лигандом mpl, может быть, необязательно, соединено со вторым полипептидом, и это антитело или его соединение могут быть использованы для выделения и очистки лиганда mpl из источников, как описано выше для иммобилизованного mpl. В дальнейшем аспекте настоящего воплощения изобретение описывает способ определения лиганда mpl in vivo, включающего контактирование антитела с образцом, в особенности с образцом сыворотки, подозреваемого на содержание лиганда, и регистрацию связывания, если оно наблюдается.
В дальнейшем воплощении изобретение описывает выделение молекулы нуклеиновой кислоты, кодирующей лиганд mpl или его фрагменты, молекула нуклеиновой кислоты которого, необязательно, может быть мечена детектируемым радикалом, и молекула нуклеиновой кислоты имеет последовательность, которая комплементарна к, или гибридизована в умеренных до высоко строгих условий с, молекулой нуклеиновой кислоты, имеющей последовательность, кодирующую mpl лиганд. Предпочтительными молекулами нуклеиновой кислоты являются те, которые кодируют лиганд mpl человека, свиньи и мыши, и включают РНК и ДНК, и геномную и кДНК. В дальнейшем аспекте настоящего воплощения молекула нуклеиновой кислоты представляет ДНК, кодирующую лиганд mpl и, далее, включающую реплицируемый вектор, в котором ДНК оперативно сцеплена с контрольной последовательностью, распознаваемой хозяином, которого трансформировали вектором. ДНК необязательно представляет собой кДНК, имеющую последовательность, представленную на фиг.1 5’-3’ (Посл. No.2), 3’-5’, или их фрагменты. Настоящий аспект далее включает клетки хозяина, предпочтительно, СНО клетки, трансформированные вектором, и способ использования ДНК для стимуляции продукции лиганда mpl, предпочтительно включающий экспрессию кДНК, кодирующую лиганд mpl, в культуре трансформированных клеток хозяина, и обратного получения лиганда mpl из клеток хозяина, или культуру клеток хозяина. Лиганд mpl, получаемый подобным способом, предпочтительно является лигандом mpl человека.
Изобретение далее включает способ лечения животных, имеющих гематопоэтическое нарушение, в особенности тромбоцитопению, включающий введение млекопитающему терапевтически активного количества лиганда mpl. Лиганд mpl, необязательно, вводится в комбинации с цитокином, в особенности с колоние-стимулирующим фактором или интерлейкином. Предпочтительные колоние-стимулирующие факторы или интерлейкины включают: кит-лиганд (КЛ), ФИЛ, Г-КСФ, ГМ-КСФ, М-КСФ, ЭП, ИЛ-1, ИЛ-3, ИЛ-6 и ИЛ-11.
Изобретение далее включает способ выделения и очистки ТП (МЛ) из ТП-продуцирующих микроорганизмов, включающий:
(1) разрушение или лизис клеток, содержащих ТП,
(2) необязательное отделение растворимого материала от нерастворимого материала, содержащего ТП,
(3) солюбилизацию ТП из нерастворимого материала с помощью солюбилизирующего буфера,
(4) отделение солюбилизированного ТП от другого растворимого и нерастворимого материала,
(5) укладывание ТП в окислительно/восстановительном буфере, и
(6) отделение правильно уложенного ТП от неуложившегося ТП.
Способ описывает солюбилизацию нерастворимого материала, содержащего ТП, хаотропным агентом, когда хаотропный агент выбирается среди солей гуанидина, тиоцианата натрия или мочевины. Способ далее описывает то, что солюбилизированный ТП отделяется от другого растворимого и нерастворимого материала при помощи одного или более этапов, выбранных среди центрифугирования, гель-фильтрации и хроматографии с обращенной фазой. Этап укладывания способа получения описывает окислительно/востановительный буфер, содержащий как окисляющий, так и восстанавливающий агент. В общем, окислителем является кислород или соединение, содержащее, по крайней мере, одну дисульфидную связь, и восстановителем является соединение, содержащее, по крайней мере, одну свободную сульфгидрильную группу. Предпочтительно, окислитель выбирается из окисленного глютатиона (GSSG) и цистина, и восстановитель выбирается из восстановленного глютатиона (GSH) и цистеина. Более предпочтительно окислителем является окисленный глютатион (GSSG), и восстановителем является восстановленный глютатион (GSH). Также является предпочтительным, чтобы молярное отношение окислителя было равно или больше, чем для восстановителя. Окислительно/восстановительный буфер дополнительно содержит детергент, предпочтительно выбираемый из CHAPS и CHAPSO, присутствующий, по крайней мере, на уровне 1%. Окислительно/восстановительный буфер дополнительно содержит NaCl, предпочтительно в ряду концентрации от около 0,1-0,5 М, и глицерин, предпочтительно в концентрации выше, чем 15%. рН окислительно/восстановительного буфера лежит, предпочтительно, в ряду от около рН 7,5-рН 9,0, и этап укладки проводится при 4 градусах в течение 12-48 часов. Этап укладки дает биологически активный ТП, у которого дисульфидная связь образуется между ближайшем к амино концу Цис и Цис, ближайшем к карбокси концу ЭП домена.
Изобретение далее включает способ очистки биологически активного ТП из микроорганизма, включающий:
(1) лизис, по крайней мере, внеклеточной мембраны микроорганизма,
(2) обработку лизата, содержащего ТП хаотропным агентом,
(3) укладку ТП и
(4) отделение примесного и неправильно сложившегося ТП от правильным образом сложившегося ТП.
Краткое описание чертежей
Фиг.1 показывает выведенную аминокислотную последовательность (Посл. No.1) mpl лиганда (МЛч), кДНК и кодирующей нуклеотидной последовательности (Посл. No.2). Нуклеотиды пронумерованы с начала каждой линии. 5’ и 3’ нетранслируемые области отмечены буквами снизу. Аминокислотные остатки пронумерованы выше последовательности, начинающейся от Сер 1 лиганда mpl (МЛ) зрелой белковой последовательности. Связи предполагаемого эксона 3 отмечены стрелками, и возможный участок N-гликозилирования отделен перегородкой. Остатки цистеина отмечены пунктиром над последовательностью. Подчеркнутая последовательность соответствует N-концевой последовательности mpl лиганда, очищенного из плазмы свиньи.
Фиг.2 показывает способ определения, использованного для включения 3H-тимидина в mpl лиганд. Для определения наличия mpl лиганда в различных источниках клетки mplPBa/F3 оставляли голодать без ИЛ-3 в течение 24 часов в увлажняемом инкубаторе при 37°С в присутствие 5% СO2 и воздуха. После голодания без ИЛ-3 клетки поместили в культуральные плашки с 96 ячейками с или без разведении образцов и культивировали в течение 24 часов в клеточном культуральном инкубаторе. В каждую ячейку добавляли по 20 мкл среды RPMI, свободной от сыворотки, содержащей 1 мкСi 3H-тимидина и оставляли на 6-8 часов. Затем клетки собирали на фильтровальной пластине с 96 ячейками и промывали водой. Затем подсчитывали счет на фильтрах.
Фиг.3 показывает действие проназы, ДТТ и нагревания на способность АПС стимулировать пролиферацию клеток Ba/F3-mpl. Для гидролиза АПС проназой проназу (Boehringer Mannheim) или бычий сывороточный альбумин привязали к Affi-гель10 (Biorad) и отдельно инкубировали с АПС в течение 18 часов. После этого гель удаляли путем центрифугирования, а супернатанты исследовали. АПС также прогревали до 80°С в течение 4 минут или обрабатывали 100 мкМ ДТТ, после чего диализовали против PBS.
Фиг.4 показывает элюцию активности mpl лиганда с колонок с Фенил-Toyoperl, Blue-Sepharose и Ultralink-mpl. Фракции 4-8 с mpl аффинной колонки были фракциями с пиком активности, элюировавшейся с колонки.
Фиг.5 показывает SDS-гельэлектрофорез на полиакриламидном геле (SDS-PAGE) фракций, элюировавшихся с Ultralink-mpl колонки. К 200 мкл из каждой фракции 2-8 было добавлено по 1 мл ацетона, содержащего 1 mM HCl при -20°С. Через 3 часа при -20°С образцы центрифугировали и полученный осадок промыли 2 раза ацетоном при -20°С. После этого ацетоновый осадок растворили в 30 мкл SDS-солюбилизирующего буфера, обработали 100 мкМ ДТТ и прогревали при 90°С в течение 5 минут. Образцы затем были разогнаны на 4-20% полиакриламидном геле и белки визуализировали прокрашиванием серебром.
Фиг.6 показывает элюцию активности mpl лиганда после SDS-PAGE. Фракция 6 после mpl-аффинной колонки была разогнана на 4-20% SDS-полиакриламидном геле при невосстанавливающих условиях. После электрофореза гель разрезали на 12 равных долей и электроэлюировали, как описано в примерах. Электроэлюированные образцы диализовали в PBS и исследовали при 1/20 разведения. Mr стандарты, использованные для калибровки геля, представляли собой 12 стандартов Novex Mark.
Фиг.7 показывает влияние АПС, лишенную лиганда mpl, на мегакариоцитопоэз у человека. АПС, лишенная лиганда mpl, была получена путем пропускания 1 мл через 1 мл mpl-аффинной колонки (700 мкг mpl-IgG/мл NHS-Superose, Pharmacia). Культуру периферических стволовых клеток человека обработали 10% АПС или 10% АПС, лишенного mpl лиганда, и культивировали в течение 12 дней. Мегакариоцитопоэз подсчитывали, как описано в примерах.
Фиг.8 показывает действие mpl-IgG на стимуляцию АПС мегакариоцитопоэза у человека. Культуру периферических стволовых клеток человека обработали 10% АПС и культивировали в течение 12 дней. На 0, 2 и 4 дни добавили mpl-IgG (0,5 мкг) или ANP-R-IgG (0,5 мкг). Через 12 дней подсчитывали мегакариоцитопоэз, как описано в примерах. Графики построены по средним значениям дубликатов образцов, в скобках действительные данные дублей.
Фиг.9 показывает обе нити фрагмента 390 пар оснований геномной ДНК человека, кодирующий mpl лиганд. Показаны выведенная аминокислотная последовательность "эксона 3" (Посл. No:3), кодирующей последовательности (Посл. No: 4) и его комплемента (Посл. No: 5).
Фиг.10 показывает выведенную аминокислотную последовательность зрелого лиганда mpl человека (МЛч) (Посл. No: 6) и зрелого эритропоэтина человека (ЭПч) (Посл. No: 7). Предсказанная аминокислотная последовательность для лиганда mpl человека расположена в ряд с последовательностью эритропоэтина человека. Идентичные аминокислоты отделены перегородкой, а разрывы, введенные для оптимального расположения, отмечены штриховкой. Места для возможного N-гликозилирования подчеркнуты прямой чертой для МЛч и ломаной линией для ЭПч. Два цистеина, важных для активности эритропоэтина, отмечены жирным пунктиром.
Фиг.11 показывает выведенную аминокислотную последовательность изоформ зрелого лиганда mpl человека МЛч (Посл. No: 6), МЛч2 (Посл. No: 8), МЛч3 (Посл. No: 9) и МЛч4 (Посл. No: 10). Идентичные аминокислоты отделены перегородкой, а разрывы, введенные для оптимального расположения, отмечены штриховкой.
Фиг.12А, 12В и 12С показывают влияние лиганда nipl человека на пролиферацию Ba/F3 клеток (А), количество мегакариоцитопоэзов у человека in vitro, подсчитанное при использовании меченого IgG моноклонального антитела мыши, специфического к гликопротеину GPIIbIIIa мегакариоцита (В), и определение тромбоцитопоэзов у мыши, исследуемое по восстановительному синтезу тромбоцитов (С).
Клетки двести девяносто три были трансфецированы согласно CaPO4 методу (Gorman, С in DNA Cloning: A New Approach 2:143-190 [1985]), только вектором pRK5, pRK5-МЛЧ или pRK5-МЛЧ153 в течение ночи (pRK5-MЛЧ153 был получен введением стоп кодона после остатка 153 МЛЧ методом РПЦ). Затем среду уравновесили на 36 часов и исследовали ее на стимуляцию пролиферации Ba/F3-mpl клеток, как описано в Примере 1 (А) или мегакариоцитопоэза у человека in vitro (В). Мегакариоцитопоэзы подсчитывали, используя меченное 125IIgG моноклональное антитело мыши (НР1-ID) к специфическому гликопротеину GPIIbIIIa, как описано (Grant et al., Blood, 69: 1334-1339 [1987]). Действие частично очищенного рекомбинантного МЛ (рМЛ) на продукцию тромбоцитов in vivo (С) определяли, используя исследование восстановительного синтеза тромбоцитов, описанное McDonald, Т.Р. Рrос. Sоc. Ехр. Biol. Med. 144: 1006-1012 (1973). Частично очищенный рМЛ получили из 200 мкл кондиционированной среды, содержащей рекомбинантный МЛ. Среду пропустили через колонку с 2 мл Blue-Sepharose, уравновешенную PBS, колонку промыли PBS и элюировали PBS, содержащим по 2 М мочевины и NaCl, каждого. Фракции с активностью диализовали в PBS и создали концентрацию 1 мг/мл БСА, свободным от эндотоксина. Образцы содержали менее чем одну единицу эндотоксина/мл. Мышам инъецировали по 64000, 32000 или 16000 единиц рМЛ или только наполнитель. Каждая группа состояла из шести мышей. Представлено среднее и стандартное отклонение в каждой группе. Значение р было определено по парному Т-тесту сравнения медиан.
Фиг.13 сравнивает действие изоформ и вариантов лиганда mpl человека при исследовании пролиферации Ba/F3 клеток. Были исследованы МЛч, mock, МЛч2, МЛч(Р153А, Р154А) и МЛч153 при различных разведениях, как описано в Примере 1.
Фиг.14А, 14В и 14С показывают выведенную аминокислотную последовательность (Посл. No: 1) лиганда mpl человека (МЛч) или ТП человека (ТПч) геномной ДНК человека, кодирующей последовательность (Посл. No: 11). Нуклеотиды и аминокислотные остатки пронумерованы с начала каждой линии.
Фиг.15 показывает SDS-PAGE (электорофорез в полиакриламидном геле в присутствии додецилсульфата натрия) очищенного 293-рМЛч332 и очищенного 293-рМЛч153.
Фиг.16 показывает нуклеотидную последовательность: кодирующей кДНК (Посл. No: 12) и выведенную аминокислотную последовательность (Посл. No: 13) открытой рамки считывания изоформы МЛ мыши. Это изоформа зрелого лиганда mpl мыши содержит 331 аминокислотный остаток, на четыре меньше, чем естественный полной длины МЛм, и поэтому обозначена как МЛм2. Нуклеотиды пронумерованы с начала каждой линии. Аминокислотные остатки пронумерованы поверх последовательности, начинающейся с Сер 1. Участки возможного N-гликозилирования подчеркнуты. Остатки цистеина отмечены пунктиром над последовательностью.
Фиг.17 показывает последовательность кДНК (Посл. No: 14) и предсказанной белковой последовательности (Посл. No: 15) данной изоформы МЛ мыши (МЛм). Нуклеотиды пронумерованы с начала каждой линии. Аминокислотные остатки пронумерованы поверх последовательности, начинающейся с Сер 1. Данная изоформа зрелого лиганда mpl мыши содержит 335 аминокислотных остатков, и представляется, что является лигандом mpl полной длины, обозначенным МЛм. Сигнальная последовательность обозначена подчеркнутой штриховкой и вероятные места расщепления обозначены стрелкой. 5’ и 3’ нетранслируемые области отмечены расположенными ниже буквами. Две обнаруженные делеции как результат альтернативного сплайсинга (МЛм2 и МЛм3) подчеркнуты. Четыре цистеиновых остатка отмечены штриховкой. Семь возможных мест N-гликозилирования отделены перегородкой.
Фиг.18 сравнивает выведенную аминокислотную последовательность МЛч3 изоформы МЛ человека (Посл. No: 9) и изоформы МЛ мыши, обозначенной МЛм3 (Посл. No: 16). Предсказанная аминокислотная последовательность лиганда mpl человека расположена вдоль последовательности лиганда mpl мыши. Идентичные аминокислоты отделены перегородкой, а разрывы введены для оптимального расположения в ряд и отмечены штриховкой. Аминокислоты пронумерованы с начала каждой линии.
Фиг.19 сравнивает предсказанную аминокислотную последовательность зрелого МЛ, изоформ МЛ мыши (Посл. No: 17), МЛ свиньи (Посл. No: 18) и МЛ человека (Посл. No: 6). Аминокислотные последовательности расположены вдоль с разрывами, отмеченными штриховкой, вставленные для оптимального расположения. Аминокислоты пронумерованы с начала каждой линии, с разделенными перегородкой идентичными аминокислотами. Места возможного N-гликозилирования отмечены затемненными перегородками, а цистеиновые остатки обозначены пунктиром. Структура с консервативными двухосновными аминокислотами, которая представляет место для потенциального расщепления протеазами, подчеркнута. Обнаруженная четвертая аминокислотная делеция, наблюдающаяся у всех трех видов (МЛ2) отделена жирной перегородкой.
Фиг.20 показывает последовательность кДНК (Посл. No: 19) и предсказанную последовательность (Посл. No:18) изоформы зрелого белка МЛ свиньи (МЛс). Данная изоформа лиганда mpl свиньи содержит 332 аминокислотных остатка и, предположительно, является лигандом mpl свиньи полной длины, обозначаемым МЛс. Нуклеотиды пронумерованы с начала каждой линии. Аминокислотные остатки пронумерованы поверх последовательности, начинающейся с Сер 1.
Фиг.21 показывает последовательность кДНК (Посл. No: 20) и предсказанную последовательность (Посл. No: 21) изоформы зрелого белка МЛ свиньи (МЛс2). Данная изоформа лиганда mpl свиньи содержит 228 аминокислотных остатков и представляет форму с четырьмя делениями остатков по сравнению с лигандом mpl полной длины, обозначаемая МЛс2. Нуклеотиды пронумерованы с начала каждой линии. Аминокислотные остатки пронумерованы поверх последовательности, начинающейся с Сер 1.
Фиг.22 сравнивает выведенную аминокислотную последовательность МЛ свиньи с полной длиной изоформы МЛс (Посл. No: 18) и изоформу МЛ свиньи, обозначенную МЛс2 (Посл. No: 21). Предсказанная аминокислотная последовательность для МЛ расположена в ряд с последовательностью МЛс2. Идентичные аминокислоты разделены перегородками, а разрывы, введенные для оптимального расположения, отмечены штриховкой. Аминокислоты пронумерованы с начала каждой линии.
Фиг.23 показывает существенные свойства плазмиды p3V15. ID.LL.MLORF ("полная длина" или ТП332), используемой для трансфекции хозяина CHO-DP12 клеток для продукции СНО-rhТП332.
Фиг.24 показывает существенные свойства плазмиды p3V15.ID.LL.MLEPO-D ("усеченной" или ТП153), используемой для трансфекции хозяина CHO-DP12 клеток для продукции СНО-rhТП153.
Фиг.25А, 25В и 25С показывает действие Е.colirhin (Мет-1, 153) на тромбоциты (А), красные кровяные клетки (В) и белые кровяные клетки (С) обычных мышей. Двум группам по шесть самок С57 В6 мышей инъецировали ежедневно либо PBS буфер, либо 0,3 мкг Е.coli-rhTn (Meт-1, 153) (100 мкл/п.к.). На 0 день и на 3-7 дни забирали по 40 мкл крови из глазного синуса. Данную кровь немедленно разводили в 10 мл коммерческого разбавителя и получали полный счет крови на Serrono Baker Heiaatology Analyzer 9018. Данные представлены в виде среднее ± стандартная ошибка определения.
Фиг.26А, 26В и 26С показывают действие Е.coli-rhTu (Мет-1, 153) на тромбоциты (А), красные кровяные клетки (В) и белые кровяные клетки (С) облученных в сублетальной дозе мышей. Две группы по 10 самок С57 В6 мышей облучили в сублетальной дозе 750 cGy гамма облучением из источника с 137Сs, которые ежедневно получали в виде инъекций либо PBS буфер, либо 3,0 мкг Е.coli-rhТГКМет-1, 153) (100 мкл/п.к.). На 0 день и в последующие промежуточные сроки отбирали по 40 мкл крови из глазного синуса. Данную кровь немедленно разводили в 10 мл коммерческого разбавителя и получали полный счет крови на Serrono Baker Hematology Analyzer 9018. Данные представлены в виде среднее ± стандартная ошибка определения.
Фиг.27А, 27В и 27С показывает действие СНО-rhТП332 на тромбоциты (А), красные кровяные клетки (эритроциты) (В) и белые кровяные клетки (лейкоциты) (С) обычных мышей. Двум группам по шесть самок С57 В6 мышей инъецировали ежедневно либо PBS буфер, либо 0,3 мкг СНО-rhТП332,) (100 мкл/п.к.). На 0 день и на 3-7 дни забирали по 40 мкл крови из глазного синуса. Данную кровь немедленно разводили в 10 мл коммерческого разбавителя и получали полный счет крови на Serrono Baker Hematology Analyzer 9018. Данные представлены в виде среднее ± стандартная ошибка определения.
Фиг.28 показывает кривую дозозависимого ответа на различные формы рТПч, полученные из различных клеточных линий. Построены кривые дозозависимых ответов на рТПч из следующих клеточных линий: ТПч332 из СНО (полной длины из клеток яичника китайского хомячка); ТПчМет-1 153 (производное Е.coli, усеченной формы с N-концевым метионином); ТПч332 (ТП полной длины из 293 клеток); без Мет 155 Е.coli (усеченная форма [рТПч155] из Е.coli без конечного метионина). Группам по 6 самок С57В6 мышей ежедневно инъецировали в течение 7 дней рТПч в зависимости от группы. Каждый день отбирали по 40 мкл крови из глазного синуса для полного счета крови. Данные, представленные выше, представляют максимальные эффекты, наблюдаемые при различных введениях, за исключением (мет 153 Е.coli), которые проявляются на 7 день введения. В вышеупомянутой "мет 153 Е.coli" группе максимальный эффект наблюдался на 5 день. Данные представлены в виде среднее ± стандартная ошибка определения.
Фиг.29 показывает сравнение кривых ответов дозовой зависимости активностей рТПч полной длины и "обрезанной" формы рТПч, продуцируемой СНО клетками, с усеченной формой из Е.coli. Группам из 6 самок С57В6 мышей ежедневно вводили по 0,3 мкг рТПч различных типов. На 2-7 дни забирали кровь из глазного синуса для полного подсчета крови. Обработанные группы следующие: ТП153 усеченная форма ТП из Е.coli; ТП332 (смешанная фракция), ТП полной длины, содержащий приблизительно 80-90% ТП полной длины и 10-20% обрезанной формы; ТП332(30К фракция) = очищенной обрезанной фракции из оригинального "смешанного" препарата; ТП332(70К фракция) = очищенно фракции ТП полной длины из оригинального "смешанного" препарата. Данные представлены в виде среднее ± стандартная ошибка определения.
Фиг.30 представляет картинку, показывающую исследование KIRA ELISA для определения ТП. Чертеж показывает химеру MPL/Rse.gD и соответствующие части родительских рецепторов, так же как и окончательную конструкцию (правая часть чертежа) и плавную диаграмму (левая часть чертежа), показывающую соответствующие этапы исследования.
Фиг.31 представляет плавную диаграмму исследования KIRA ELISA, показывающую каждый этап в определении.
Фиг.32A-32L описывает нуклеотидную последовательность (Посл. No:22) вектора p3VI17.ID.LL экспрессии, использованного для экспрессии Rse.gD в Примере 17.
Фиг.33 является схематическим представлением приготовления плазмиды pMP1.
Фиг.34 является схематическим представлением приготовления плазмиды рМР21.
Фиг.35 является схематическим представлением приготовления плазмиды рМР151.
Фиг.36 является схематическим представлением приготовления плазмиды рМР202.
Фиг.37 является схематическим представлением приготовления плазмиды рМР172.
Фиг.38 является схематическим представлением приготовления плазмиды рМР210.
Фиг.39 является таблицей пяти лучших экспрессии ТП клонов из плазмиды рМР210 банка плазмид (Посл. No: 23, 24, 25, 26, 27 и 28).
Фиг.40 является схематическим представлением приготовления плазмиды рМР41.
Фиг.41 является схематическим представлением приготовления плазмиды рМР57.
Фиг.42 является схематическим представлением приготовления плазмиды рМР251.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
1. Определения
В основном, следующие слова или фразы имеют указанное определение, когда используются в описаниях, примерах и формуле изобретения.
"Хаотропный агент" относится к соединению, которое в водном растворе и подходящей концентрации может вызвать изменения в пространственной конфигурации или конформации белка, по крайней мере, частично разрушая силы, ответственные за поддержку нормальной вторичной или третичной структуры белка. Подобные соединения включают, например, мочевину, гуанидин-НСl и тиоцианат натрия. Для оказания конформационного воздействия на белок обычно требуются высокие концентрации, 4-9 М, этих соединений.
"Цитокин" - термин, характерный для белков, выделяемых одной клеточной популяцией, которые действуют на другие клетки как межклеточные медиаторы. Примерами подобных цитокинов являются лимфокины, монокины и обычные полипептидные гормоны. К цитокинам относятся: гормон роста, инсулиноподобные факторы роста, гормон роста человека, N-метионил гормон роста человека, бычий гормон роста, паратиреоидный гормон, тироксин, инсулин, проинсулин, релаксин, прорелаксин, гормоны гликопротеинового ряда, такие как фолликулостимулирующий гормон (ФСГ), тиреостимулирующий гормон (ТСГ) и лютеинизирующий гормон (ЛГ), гематопоэтический фактор роста, фактор роста печени, фактор роста фибробластов, пролактин, плацентарный лактоген, фактор некроза опухолей α (ФНО-α или ФНО-β), мюллериан-ингибирующая субстанция, ассоциированная с гонадотропином белок мыши, ингибин, активин, эндотелиальный фактор роста сосудов, интегрин, фактор роста нейронов, такой как ФРН-β, фактор роста тромбоцитов, трансформирующие факторы роста (ТФР), такие как ТФР-α и ТФР-β, инсулиноподобный фактор роста-1 и -2, эритропоэтин (ЭП), остеоиндуктивные факторы, интерфероны, такие как интерферон-α, -β и -γ, колоние-стимулирующие факторы (КСФ), такие как макрофаг-КСФ (М-КСФ), гранулоцит-макрофаг-КСФ (ГМ-КСФ), и гранулоцит-КСФ (Г-КСФ), интерлейкины (ИЛ), такие как ИЛ-1, ИЛ-1α, ИЛ-2, ИЛ-3, ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-7, ИЛ-8, ИЛ-9, ИЛ-11, ИЛ-12 и другие полипептидные факторы, включая КСФ, ФИЛ и kit-лиганд. Как здесь использовано, последующие термины предназначены для охвата белков из природных источников или рекомбинантных форм клеточных линий. Подобным же образом термины предназначаются для охвата биологически активных эквивалентных производных, а именно, различающихся в аминокислотной последовательности по одной или более аминокислот, или типу, или степени гликозилирования.
"Лиганд mpl", "полипептидный лиганд mpl", "МЛ", тромбопоэтин" или "ТП" используются здесь взаимозаменяемо и включают любой полипептид, который проявляет способность связывания с mpl, членом супергруппы цитокиновых рецепторов, и обладающий биологическими свойствами МЛ, как показано ниже. Примером биологической особенности является способность стимулировать включение меченых нуклеотидов (а именно, 3H-тимидина) в ДНК ИЛ-3 зависимых Ba/F3 клеток, трансфецированных mpl P человека. Другим примером биологической особенности является способность стимулировать включение 35S в циркулирующие тромбоциты в исследовании восстановительного синтеза тромбоцитов. Данное определение заключает в себе полипептид, выделенный из источника mpl лиганда, такого как апластическая плазма свиньи, описанная здесь, или другого вида животных, включая человека, или полученный рекомбинантными путями или методами синтеза, и содержит различные формы, включая функциональные производные, фрагменты, аллели, изоформы и его аналоги.
"Фрагмент лиганда МЛ" или "фрагмент ТП" - это часть естественно обнаруживаемого зрелого лиганда mpl полной длины, или последовательности ТП, имеющей делецию по одному или более аминокислотных остатков, или лишенных углеводных частей. Делеция аминокислотного остатка (остатков) может наблюдаться в любом месте белка, включая и N-терминальный, и С-терминальный конец, или внутри. Фрагмент имеет, по крайней мере, одно биологическое свойство, общее с лигандом mpl. Фрагменты лиганда mpl обычно будут иметь порядковую последовательность как минимум 10, 15, 20, 25, 30, или 40 аминокислотных остатков, которые идентичны последовательности лиганда mpl, выделенного из млекопитающего, включая лиганд, выделенный из апластической плазмы свиньи, или лиганд человека, или лиганд мыши, в особенности их ЭП-домен. Характерными примерами N-концевых фрагментов являются МЛч или ТП (Met-1 1-153).
"Варианты лиганда mpl" или "варианты последовательности лиганда mpl", как здесь определено, означает биологически активный лиганд mpl, как определено ниже, имеющий менее чем 100% идентичности в последовательности с лигандом mpl, выделенным из рекомбинантной культуры клеток или апластической плазмы свиньи, или лиганд человека, имеющий выведенную последовательность, описанную в фиг.1 (Посл. No:l). Обычно биологически активный лиганд mpl будет обладать аминокислотной последовательностью, имеющей, по крайней мере, 70% идентичности с аминокислотной последовательностью лиганда mpl, выделенного из апластической плазмы свиньи или зрелого лиганда мыши, или человека, или их фрагментов, (см. фиг.1 [Посл. No:l]), предпочтительно, по крайней мере, около 75%, более предпочтительно, по крайней мере, 80%, еще более предпочтительно, не менее 85%, и еще более предпочтительно, по крайней мере, 90%, и наиболее предпочтительно, по крайней мере, 95%.
"Химерный лиганд mpl" - это полипептид, содержащий лиганд mpl полной длины, или один или более их фрагментов, слитых или связанных со вторым гетерологическим полипептидом, или с одним или более их фрагментами. Химера обладает, по крайней мере, одним биологическим свойством, общим с лигандом mpl. Второй полипептид обычно является цитокином, иммуноглобулином или их фрагментом.
"Выделенный лиганд mpl", "высоко очищенный лиганд mpl" и "существенно гомогеннный лиганд mpl" используются взаимозаменяемо и обозначают лиганд mpl, который был очищен из источника для лиганда mpl или был получен рекомбинантным способом или методами синтеза и был существенно очищен от других пептидов или белков (1), с получением, по крайней мере, 15 и, предпочтительно, 20 аминокислотных остатков С-концевой или внутренней аминокислотной последовательности, используя секвенатор с вращающимся реактором, или лучший коммерчески доступный аминокислотный секвенатор, или при помощи опубликованных способов согласно модификациям на дату заполнения данной заявки, или (2) до гомогенного состояния при помощи SDS-PAGE в невосстанавливающих или восстанавливающих условиях, используя Coomassie blue или, предпочтительно, прокрашивание серебром. Гомогенное состояние здесь означает менее чем 5% загрязнение другими белками.
"Биологическое свойство", когда использовано в связи с "лигандом mpl" или "выделенным лигандом mpl", обозначает наличие тромбопоэтической активности, или наличие in vivo эффекторной или антигенной функции, или активности, которая прямо или косвенно связана или вызвана лигандом mpl (так или иначе в естественной или денатурированной конформации) или его фрагментом. Эффекторные свойства включают в себя: связывание mpl и любая связывающая активность переносчика, агонизм или антагонизм mpl, в особенности в передаче пролиферативного сигнала, включая репликацию, функцию регуляции ДНК, модуляцию биологической активности других цитокинов, активацию рецепторов (особенно цитокиновых), деактивацию, прямую и обратную регуляцию, регуляцию роста клеток и дифференциацию и тому подобное. Антигенная функция означает обладание эпитопом или антигенным сайтом, который способен перекрестно реагировать с антителами, образовавшимися против природного лиганда mpl. Главное антигенное свойство полипептидного лиганда mpl состоит в том, что он связывается по сродству, по крайней мере, около 106 л/моль, с антителом, вырабатывающимся против лиганда mpl, выделенного из апластической плазмы свиньи. Обычно полипептид связывается по сродству, по крайней мере, около 107 л/моль. Наиболее предпочтительно, чтобы антигенно активный полипептидный лиганд mpl являлся полипептидом, который связывается с антителами, вырабатывающимися против лиганда mpl, имеющего одну из описанных выше эффекторных функций. Антитела, используемые для определения "биологической активности", являются поликлональными антителами кролика, вырабатывающимися на препарат лиганда mpl, выделенного из рекомбинантной культуры клеток или из апластической плазмы свиньи с полным адьювантом Freund'a при подкожной инъекции препарата и усилении иммунного ответа путем внутрибрюшинной инъекции препарата, до выхода на плато титра антител к лиганду mpl.
"Биологически активный", когда использован в связи либо с термином "лиганд mpl", либо с "выделенный лиганд mpl", обозначает лиганд mpl или полипептид, который проявляет тромбопоэтическую активность, или обладает эффекторной функцией лиганда mpl, выделенного из апластической плазмы свиньи или экспрессированного в рекомбинантной клеточной культуре, описанной здесь. Основной известной эффекторной функцией лиганда mpl или его полипептида является связывание с mpl и стимуляция включения меченных нуклеотидов (3H-тимидина) в ДНК ИЛ-3 зависимых клеток Ba/F3, трансфецированных mpl P человека. Другой известной эффекторной функцией лиганда mpl или его полипептида является способность стимулировать включение 35S в циркулирующие тромбоциты в исследовании обратного связывания тромбоцитов у мыши. Еще одной известной эффекторной функцией лиганда mpl является способность стимулировать in vitro мегакириоцитопоэз у человека, который можно подсчитать путем использования радиоактивно меченных антител, специфических к гликопротеину GPllbllla мегакариоцита.
"Процент идентичности аминокислотной последовательности", принимая во внимание последовательность лиганда mpl, определяется здесь в виде процентного соотношения аминокислотных остатков выбранной идентичной последовательности к остаткам последовательности лиганда mpl, выделенного из апластической плазмы свиньи, или лиганда мыши или человека, имеющего аминокислотную последовательность установленного происхождения, описанную на фиг.1 (Посл. No:1), после выпрямления последовательности и введения разрывов, если необходимо, для достижения максимального процента идентичности последовательности и, не учитывая любые консервативные замены, как часть идентичной последовательности. Никакие из N-концевых, или С-концевых, или внутренних расширений, делеций или вставок в последовательности лиганда mpl не должны быть истолкованы как влияющие на идентичность последовательности или гомологию. Таким образом, рассмотренные примеры биологически активных полипептидов лиганда mpl должны иметь идентичные последовательности, включая: препро-mpl лиганд, про-mpl лиганд и зрелый mpl лиганд.
"Микросеквенирование лиганда mpl" может быть выполнено любым подходящим обычным способом, если способ достаточно чувствителен. В одном из таких способов высокоочищенные полипептиды, полученные из SDS геля, или на конечной стадии ЖХВР секвенируются непосредственно путем автоматизированного расщепления (фенилизотиоцианатного) по Edman, используя модель секвенатора 470А Applied Biosystems с газовой фазой, снабженным 120А фенилтиогидантоиновым (ФТГ) аминокислотным анализатором. Кроме этого, фрагменты лиганда mpl, полученные химическим путем (а именно, с CNBr, гидроксиламином, 2-нитро-5-тиоцианобензо-атом) или ферментативным (а именно, с трипсином, клострипаном, стафилококковой протеазой) расщеплением с последующей очисткой фрагментов (а именно, путем ЖХВР), могут быть секвенированы подобным же образом.
ФТГ аминокислоты исследовали при помощи системы данных ChromPerfect (Justice Innovations, Palo Alto, CA). Интерпретация последовательностей провели на компьютере VAX 11/785 Digital Equipment Co. как описано Henzel et al., J. Chromatography, 404: 41-52 [1987]. Равные доли фракций ЖХВР, необязательно, могут быть подвергнуты электрофорезу в 5-20% SDS-PAGE, перенесены под воздействие электротока на PDVF мембрану (ProBlott, AIB, Foster City, СА) и окрашены Кумасси бриллиантовым синим (Matsurdiara J. Biol. Chem., 262: 10035-10038 [1987]. Идентифицированный по окраске специфический белок вырезают из блота, а N-концевую последовательность подвергают обработке при помощи секвенатора с газовой фазой, описанного выше. Для внутренних последовательностей белка, фракции ЖХВР высушивают под вакуумом (SpeedVac), ресуспендируют в подходящем буфере и подвергают взаимодействию с цианоген бромидом, Лиз-специфическим ферментом Lys-C (Wako Chemicals, Richmond, VA), или Acп-N (Boehringer Mannheim, Indianapolis, IN). После гидролиза полученные белки секвенируются в смеси или после разделения на ЖХВР при разгонке на С4 колонке в градиенте пропанола и 0,1% TFA, перед секвенированием в газообразной фазе.
"Тромбоцитопения" определяется как количество тромбоцитов в крови ниже 150×109 на литр.
"Тромбопоэтическая активность" определяется как биологическая активность, которая включает ускорение пролиферации, дифференциации и/или созревания мегакариоцитов или предшественников мегакариоцитов в тромбоцитообразующие формы этих клеток. Эта активность может быть измерена в различных исследованиях, включая исследование восстановительного синтеза тромбоцитов мыши in vivo, исследование индукции клеточного поверхностного антигена тромбоцитов, измеренное путем антитромбоцитарного иммуноанализа (anti-GPllbllla) для мегакариобластической клеточной лейкемии человека (СМК), и исследование индукции полиплоидизации в линии мегакариобластических клеток (DAMI).
"Тромбопоэтин" (ТП) определяется как соединение, имеющее тромбопоэтическую активность или способное увеличивать количество тромбоцитов в сыворотке млекопитающих. ТП преимущественно способен увеличивать количество эндогенных тромбоцитов, по крайней мере, на 10%, более предпочтительно, на 50%, и наиболее предпочтительно, способен увеличивать количество тромбоцитов у человека, более чем до 150×109 на литр крови.
"Нуклеиновая кислота выделенного лиганда mpl" - это РНК или ДНК, содержащая более чем 16 и предпочтительно 20 или более последовательных оснований нуклеотидов, которые кодируют биологически активный лиганд mpl или его фрагмент, являющийся комплементарным к РНК или ДНК, или гибридизующимся с РНК или ДНК и сохраняющим стабильную связь при умеренных, до жестких условий. Данные РНК или ДНК свободны, по крайней мере, от единственного источника примесной нуклеиновой кислоты, с которой она обычно связана в природных условиях и, предпочтительно, существенно свободна от любой другой РНК или ДНК млекопитающих. Фраза "свободна, по крайней мере, от единственного источника примесной нуклеиновй кислоты, с которым она обычно связана" включает место, в котором обычно находятся нуклеиновые кислоты в источнике, или природную клетку, но в различном хромосомном положении, или в противном случае сцеплены с последовательностью нуклеиновых кислот, обычно не встречающихся в источнике клеток. Примером нуклеиновой кислоты выделенного лиганда mpl является РНК или ДНК, которые кодируют биологически активный лиганд mpl, обладая, по крайней мере 75% идентичности в последовательности, более предпочтительно, по крайней мере, 80%, еще более предпочтительно 85%, и еще более предпочтительно 90%, и наиболее предпочтительно, 95% идентичности в последовательности с лигандом mpl человека, мыши или свиньи.
"Контрольные последовательности", когда относятся к экспрессии, означают последовательности ДНК, необходимые для экспрессии операционно сцепленной к кодирующей последовательности в конкретном организме хозяина. Контрольные последовательности, применимые, например, для прокариот, включают промотор, необязательно операторную последовательность, участок связывания рибосом и, возможно, другие, еще недостаточно изученные последовательности. Клетки эукариотов, как известно, используют промоторы, сигналы полиаденилирования и усилители.
"Операционно сцепленный", когда относится к нуклеиновым кислотам, означает, что нуклеиновые кислоты находятся в функциональной взаимосвязи с другими последовательностями нуклеиновых кислот. Например, ДНК для предпоследовательности или секреторного лидера операционно сцеплена с ДНК для полипептида, если он экспрессирован как препротеин, который принимает участие в секреции полипептида; промотор или усилитель является операционно сцепленным с кодирующей последовательностью, если она влияет на транскрипцию последовательности; или участок связывания рибосомы, операционно сцеплен с кодирующей последовательностью, если он расположен так, чтобы усиливать трансляцию. В общем, "операционно сцепленный" означает, что сцепленные последовательности ДНК соприкасаются, и в случае секреторного лидера соприкасаются и в фазе считывания. Однако, усилители не должны соприкасаться. Сцепливание сопровождается лигацией в сайтах, подходящих для рестрикции. Если такие сайты не существуют, синтетические олигонуклеотидные адаптеры или линкеры используются в соответствии с общепринятым опытом.
"Экзогенная", когда относится к элементу, означает последовательность нуклеиновой кислоты, которая является чужеродной для клетки или гомологичной для клетки, но в таком положении в нуклеиновой кислоте клетки хозяина, в котором элемент обычно не обнаруживается.
"Клетка", "линия клеток" и "культура клеток" взаимозаменяемо используются здесь, и подобные обозначения включают всех потомков клетки или линии клетки. Таким образом, например, термины, такие как "трансформанты" и "трансформированные клетки", включают в себя предмет первичной клетки и происходящие из нее культуры вне зависимости от количества пересевов. Также понятно, что все потомки не могут быть совершенно идентичны по составу ДНК благодаря преднамеренным или случайным мутациям. Потомки мутантов, имеющие такие же функции или биологическую активность, как просеивается для обычно трансформированной клетки, также включены. Где предполагаются отличающиеся обозначения, они будут удалены из контекста.
"Плазмиды" - автономно реплицирующиеся молекулы кольцевой ДНК, проявляющие независимую репликационную природу, и обозначены здесь маленькой буквой "р", которая ставится до и/или после заглавной буквы и/или номера. Исходные плазмиды здесь либо коммерчески доступны, открыто доступны без ограничений, либо могут быть получены из таких же доступных плазмид в соответствии с описанными способами. Помимо этого в технике известны другие эквивалентные плазмиды, которые будут доступны для обычного работника.
"Ограниченное ферментативное расщепление", когда относится к ДНК, означает каталитическое расщепление фосфодиэфирных связей ферментом, который реагирует только по определенным положениям или сайтам в последовательности ДНК. Такие ферменты названы "рестрикционными эндонуклеазами". Каждая рестрикционная эндонуклеаза узнает специфическую последовательность ДНК, называемую "рестрикционный сайт", который проявляет двустороннюю симметрию. Различные рестрикционные ферменты, используемые здесь, коммерчески доступны, и применены условия для реакции, кофакторы и другие требования, как установлено поставщиками ферментов. Рестрикционые ферменты обычно обозначены сокращениями, составленными из заглавной буквы и следующими за ней другими буквами, представляющими микроорганизм, из которого каждый рестрикционный фермент изначально был получен, и затем номер, обозначающий конкретный фермент. Обычно используют около 1 мкг плазмид или фрагмента ДНК с около 1-2 единицами фермента в около 20 мкл буферного раствора. Подходящие буферы и количество субстрата для конкретного рестрикционных ферментов определяются производителем. Обычно используют инкубацию в течение около 1 часа при 37°С, но их можно изменять в соответствии с указаниями поставщика. После инкубации белки или полипептиды удаляются путем экстракции фенолом и хлороформом, а расщепленная нуклеиновая кислота извлекается из водной фракции путем осаждения этанолом. Расщепление рестрикционным ферментом может быть продолжено гидролизом бактериальной щелочной фосфатазой концевых 5’фосфатов для предотвращения появления двух рестрикционно расщепленных концов фрагмента ДНК из "циркуляризующейся" (образующей кольцо) или формирующей замкнутую петлю, которая могла бы препятствовать инсерции (вставке) другого фрагмента ДНК в рестрикционный сайт. Если не установлено другое, расщепление плазмид не следует за 5’концевым дефосфорилированием. Общепринятые способы и реактивы для дефосфорелирования подробно описаны в разделах 1.56-1.61 Sambrook et al., Molecular Cloning: A Laboratory Manual [New-York: Cold Spring Harbor Laboratory Press, 1989].
"Выход" или "выделение" данного фрагмента ДНК из расщепленного рестрикционного остатка означает выделение расщепленного остатка путем электрофореза на полиакриламиде или агарозном геле, идентификацию интересующего фрагмента путем сравнения его подвижности по отношению к маркерным фрагментам ДНК с известным молекулярным весом, выделяя из геля секцию, содержащую интересующий фрагмент, и разделяя гель и ДНК. Эти способы хорошо известны. Например, см. в Lawn et al., Nucleic Acids Res., 9: 6103-6114 [1981] и Goeddel et al., Nucleic Acids Res., 8: 4057 [1980].
"Southern анализ" или "Southern блоттинг" - это метод, при помощи которого подтверждается присутствие последовательности ДНК в остатке ДНК или в композиции, содержащей ДНК, после расщепления рестрикционной эндонуклеазой путем гибридизации с известным меченым олигонуклеотидом или фрагментом ДНК. Southern анализ, как правило, включает в себя электрофоретическое разделение фрагментов ДНК и агарозного геля, денатурацию ДНК после электрофоретического разделения и перенос ДНК на нитроцеллюлозу, нейлон или другую подходящую мембранную подложку для анализа с радиоактивно меченным, биотинилированным или ферментативно-меченым зондом, как описано в разделах 9.37-9.52 Sambrook et al., там же.
"Nothern анализ" или "Nothern блот" - метод, используемый для идентификации последовательностей РНК, которые гибридизуются с известным зондом, таким как олигонуклеотид, фрагмент ДНК, кДНК или ее фрагмент, или фрагмент РНК. Зонд мечен радиоактивным изотопом, таким как 32P, или путем биотинилирования, или ферментативно. Анализируемую РНК обычно электрофоретически разделяют на агарозе или полиакриламидном геле, переносят на нитроцеллюлозу, нейлон или другую подходящую мембрану и гибридизируют с зондом, используя обычные способы, хорошо известные в методах, подобных тем, которые описаны в разделах 7.39-7.52 Sambrook et al., там же.
"Лигирование" - процесс образования фосфодиэфирных связей между двумя фрагментами нуклеиновых кислот. Для лигирования двух фрагментов концы данных фрагментов должны быть совместимы друг с другом. В некоторых случаях концы будут совместимы непосредственно после эндонуклеазного расщепления. Однако может возникнуть необходимость сначала преобразовать свободно колеблющиеся концы, обычно получающиеся в процессе эндонуклеазного расщепления, для выравнивания концов, чтобы придать им совместимость для лигирования. Для выравнивания концов ДНК обрабатывают в подходящем буфере, по крайней мере, 15 минут при 15°С с примерно 10 частями Klenow фрагмента ДНК-полимеразы I или Т4 ДНК-полимеразы в присутствии четырех дезоксирибонуклеотид трифосфатов. Затем ДНК очищают путем экстракциии фенол-хлороформом и осаждением этанолом. Фрагменты ДНК, которые должны быть лигированы, помещают в раствор в примерно эквимолярных количествах. Раствор также содержит АТФ, лигазный буфер и лигазу, такую как Т4 ДНК-лигаза в количестве примерно 10 единиц на 0,5 мкг ДНК. Если эта ДНК должна быть лигирована в вектор, вектор сначала выпрямляют путем расщепления подходящей рестрикционной эндонуклеазой (эндонуклеазами). Затем выпрямленный в линию фрагмент связывают с бактериальной щелочной фосфатазой или фосфатазой из кишечника теленка для предотвращения самолигации во время этапов лигации.
"Получение" ДНК из клеток означает выделение плазмидной ДНК из культуры клеток хозяина. Обычно для получения ДНК используется метод крупно- и мелкомасштабного получения плазмид, описанный в разделах 1.25-1.33 Sambrook et al., там же. После получения ДНК она может быть очищена при помощи способов, хорошо известных в технике, таких как описанные в разделе 1.40 Sambrok et al., там же.
"Олигонуклеотиды" - это короткие по длине, одиночные или двухнитевые полидезоксинуклетиды, которые химически синтезированы согласно известным химическим способам, таким как фосфотриэфирный, фосфитный или фосфорамидный, используя твердофазные технологии, такие, как описано в ЕР 266032, опубликованном 4 мая 1988 г., или через дезоксинуклеозид Н-фосфонатные интермедиаты, как описано Froeler et al., Nucl. Acids Res., 14: 5399-5407 [1986]). Далее способы включают реакцию полимеразной цепи, описанную ниже, и другие аутопраймерные способы и синтез олигонуклеотидов на твердых субстратах. Все эти методы описаны Engels et al., Agnew. Chem. Int. Ed. Engi., 28: 716-734 (1989). Данные способы применяются, если известна полная последовательность нуклеиновых кислот в гене или доступна последовательность нуклеиновых кислот, комплементарная кодирующей нити. Альтернативно, если мишень последовательности аминокислот известна, можно вывести возможные последовательности нуклеиновых кислот, используя известные и предпочтительные кодирующие остатки для каждого аминокислотного остатка. Олигонуклеотиды затем очищают на полиакриламидном геле.
"Реакция полимеразной цепи" или "РПЦ" относится к методу или способу, в которой небольшие количества специфического фрагмента нуклеиновой кислоты, РНК и/или ДНК размножают, как описано в Патенте США No.4683195, опубликованном 28 июля 1987 г. В целом, если требуется для доступности информация о последовательности из концов и интересующей области или помимо требующейся могут быть созданы подобные олигонуклеотидные праймеры; эти праймеры для амплификации будут идентичны или близки последовательности нити противоположного направления шаблона. 5’ нуклеотиды двух праймеров могут совпадать с концами амплифицируемого материала. РПЦ может быть использована для размножения специфических последовательностей РНК, специфических последовательностей ДНК из всей геномной ДНК и кДНК, считываемой со всей последовательности РНК клетки, бактериофага или плазмиды, и т.д. В целом, см. Mullis et al., Cold Spring Harbor Symp. Quant Biol., 51: 263 [1987]; Eriich, ed., PCR Technology, (Stockton Press, NY, 1989). Как использовано здесь, РПЦ предположительно является одной, но не единственной, примером способа полимеразной реакции нуклеиновой кислоты для размножения исследуемого образца нуклеиновой кислоты, включающего использование известной нуклеиновой кислоты в виде праймера и нуклеиновой кислоты полимеразы для умножения или производства специфических фрагментов нуклеиновой кислоты.
"Строгие условия" - это те, когда (1) для промывки применяют низкая ионная сила и высокая температура, например, 0,015 М NaCl/0,0015 M цитрата натрия/0,1% NaDodSO4 (SDS) при 50°С, или (2) применяют в течение гибридизации денатурирующий агент, такой как формадид, например, 50% (об./об.) формадид с 0,1% бычьим сывороточным альбумином/01%, Фиколл/0,1% поливинилпирролидон/50 mM фосфатного буфера при рН 6,5 с 750 mM NaCl, 75 тМ цитрата натрия при 42°С. Другим примером является использование 50% формадида, 5×SSC (0,75 М NaCl, 0,075 М цитрата натрия), 50 mM фосфата натрия (рН 6,8), 0,1% пирофосфата натрия, 5×раствор Denhardt’a, обработанная ультразвуком ДНК сперма лосося (50 мкг/мл), 0,1% SDS, и 10% сульфат декстрана при 42°С, с промывкой в 0,2×SSC и 0,1% SDS.
"Умеренно строгие условия" описаны Sambrook et.al., там же, и включают в себя использование промывающего раствора и гибридизационных условий (а именно, температура, ионная сила и %SDS), менее строгих, чем описанные выше. Примерами умеренно строгих условий являются такие условия, как инкубация в течение ночи при 37°С в растворе, содержащем: 20% формадида, 5×SSC (150 мМ NaCl, 15 мМ тринатрия цитрата), 50 мМ фосфата натрия (рН 7,6), 5×раствора Denhardt’a, 10% сульфата декстрана и 20 мкл/мл денатурированной порезанной ДНК спермы лосося, с последующим промыванием фильтров в 1×SSC при около 37-50°С. Опытный работник разберет, как довести температуру, ионную силу и т.д. при необходимости согласования таких факторов, как длина зонда и подобное.
"Антитела" (AT) и "Иммуноглобулины" (ИГ) - это гликопротеины, имеющие одинаковые структурные характеристики. Во время того как антитела проявляют специфичность в связывании со специфическими антигенами, иммуноглобулины включают и антитела, и другие подобные антителам молекулы, которые лишены антигенной специфичности. Полипептиды последнего вида, например, продуцируются лимфатической системой на низких уровнях и на повышенном уровне при миеломах.
"Естественные антитела и иммуноглобулины" представляют обычно гетеротетрамерные гликопротеины массой около 150000 дальтон, состоящие из двух идентичных легких (Л) цепей и двух идентичных тяжелых (Т) цепей. Каждая легкая цепь связана с тяжелой цепью посредством одной ковалентной дисульфидной связью, хотя количество дисульфидных связей между тяжелыми цепями различается у различных изотипов иммуноглобулинов. Каждая тяжелая и легкая цепь также имеет обычно расположенные внутри цепи дисульфидные мостики. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VT), следующий после многочисленных постоянных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VЛ) и постоянный домен на другом его конце; постоянный домен легкой цепи располагается вдоль первого постоянного домена тяжелой цепи, а легкий вариабельный домен располагается вдоль вместе с вариабельным доменом тяжелой цепи. Предполагается, что конкретные аминокислотные остатки образуют границу раздела между легкой и тяжелой цепью вариабельных доменов (Clothia et al., J. Mol. Biol., 186: 651-663 [1985]; Novotny and Haber, Proc. Natl. Acad. Sci. USA, 82: 4592-4596 [1985]).
Термин "вариабельный" относится к тому факту, что определенные части вариабельного домена обширно различаются в последовательности между антителами и используются для связывания и (придания) специфичности каждого конкретного антитела по отношению к его конкретному антигену. Однако вариабильность не равным образом распределена по вариабельным доменам антител. Она концентрируется в трех фрагментах, называемых областями определения комплементарности (ООК) или гипервариабельными областями в обеих: и в легкой цепи, и в тяжелой цепи вариабельных доменов. Более высококонсервативная часть вариабельных доменов называется рабочей рамкой (РР). Вариабельные домены естественных тяжелой и легкой цепей, каждая, содержит четыре области РР, в значительной степени, адаптирована к β-складчатой конфигурации, соединенных тремя ООК, которые образуют петли соединений и в некоторых случаях образуют часть β-складчатой структуры. ООК в каждой цепи поддерживаются вместе в тесной близости к РР области и к ООК из другой цепи, вносящей вклад в образование участка связывания антигена на антителах (см. Kabat et al., Sequences of Proteins of Immunological Interest, National Institute of Health, Bethesda, MD, [1987]). Постоянные домены не вовлечены непосредственно в связывание антитела к антигену, но проявляют различные эффекторные функции, такие как участие антитела в зависимой от антител клеточной токсичности.
Расщепление антител папаином дает два идентичных фрагмента связывания антигена, называемых "Fab" фрагментами, каждый с одним участком связывания антигена, и оставшимся "Fc" фрагментом, чье имя отражает его способность легко кристаллизоваться. Обработка трипсином дает F(ab’)2 фрагмент, который имеет две соединенных области связывания антигена, и по-прежнему способный к перекрестному связыванию антигена.
"Fv" представляет минимальный фрагмент антитела, который содержит полный участок распознавания и связывания антигена. Эта область состоит из димера вариабельного домена одной тяжелой и одной легкой цепи в тесной нековалентной связи. Именно в этой конфигурации, в которой взаимодействуют три ООК каждого из вариабельных доменов, устанавливается участок связывания антигена на поверхности VT-VЛ димера. Все вместе шесть ООК придают антителу специфичность связывания антигена. Однако даже в одном вариабельном домене (или половине Fv, содержащей только три ООК, специфических к антигену) имеется способность для распознавания и связывания антигена, хотя при более низком сродстве, чем у полного участка связывания.
Fab фрагмент также содержит постоянный домен легкой цепи и первый постоянный домен (СН1) тяжелой цепи. Fab’ фрагменты отличаются от Fab фрагментов добавлением нескольких остатков в карбокси-конце тяжелой цепи СН1 домена, включая один или более цистеинов из области петли антитела. Fab’-SH является здесь обозначением для Fab’, в котором остаток(и) цистеина постоянного домена несут три свободных тиольных группы. Фрагменты F(ab’)2 антитела обычно продуцируются в виде пар Fab’ фрагментов, которые имеют цистеины в петле между ними. Известно также другое химическое соединение фрагментов антитела.
"Легкие цепи" антител (иммуноглобулины) из любого вида позвоночных могут быть определены к одному или двум четко различающимся типам, называемым каппа и ламбда (λ), на основании аминокислотных последовательностей их постоянных доменов.
Иммуноглобулины могут быть разделены на различные классы в зависимости от их аминокислотных последовательностей постоянного домена тяжелых цепей. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и несколько из этих классов могут быть подразделены далее на подклассы (изотипы), а именно: IgG-1, IgG-2, IgG-3 и IgG-4; IgA-1 и IgA-2. Постоянные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов названы α, дельта, эпсилон, γ и μ соответственно. Структуры субъединиц и трехмерная конфигурация различных классов иммуноглобулинов хорошо известна.
Термин "антитело" используется в самом широком смысле и специфически охватывает одни моноклональные антитела (включая агонисты и антагонисты антител), композиции антител с полиэпитопной специфичностью, так же как и фрагменты антител (а именно, Fab, F(ab’)2 и Fv) и так далее, пока они проявляют желаемую биологическую активность.
Термин "моноклональное антитело", как здесь использовано, относится к антителу, полученному из популяции существенно гомогенных антител, то есть индивидуальные антитела, составляющие популяцию, являются идентичными за исключением возможных естественно наблюдающихся мутаций, которые могут присутствовать в минорных количествах. Моноклональые антитела являются высокоспецифическими, поскольку направлены против одного антигенного участка. Более того, в противоположность к общепринятым (поликлональным) препаратам антител, которые обычно включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело является направленным против одного детерминанта на антигене. В дополнение к их специфичности моноклональные антитела выгодны в том, что они синтезируются гибридомной культурой, не загрязненной другими иммуноглобулинами. Модифицированный "моноклон" отмечает характер антитела как получаемого из существенно гомогенной популяции антител, и не должно быть истолковано, как требующееся получение антител любым другим конкретным способом. Например, моноклональные антитела, которые должны использоваться в соответствии с настоящим изобретением, могут быть получены гибридомным методом, впервые описанным Kohler & Milstein, Nature, 256: 495 (1975), или получены методами рекомбинации ДНК (см., например, Патент США No.4816567 [Cabilly et.al.]).
Моноклональные антитела здесь включают в себя "химерные" антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующей последовательности антител, происходящих от определенных видов или относящихся к конкретному классу или подклассу антител, в то время как остаток цепи (цепей) идентичен с, либо гомологичен соответствующей последовательности антител, происходящих от другого вида, либо относящихся к другому классу или подклассу антител, так же, как и фрагменты таких антител, пока они проявляют желаемую биологическую активность (Патент США No.4816567 (Cabilly et al.); и Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855 [1984]).
"Очеловеченные" формы нечеловеческих (например, мышиных) антител представляют химерные иммуноглобулины, иммуноглобулиновые цепи или их фрагменты (такие, как Fv, Fab, Fab’, F(ab’)2 или другие связывающие антиген подпоследовательности антител), которые содержат минимальную последовательность, происходящую из нечеловеческого иммуноглобулина. В основной части очеловеченные антитела являются иммуноглобулинами человека (антитела реципиента), в которых участки из области, определяющей комплементарность (ООК) реципиента, заменены участками из (ООК) нечеловеческих видов (антитела донора), таких как мышь, крыса или кролик, обладающими желаемой специфичностью, сродством и емкостью. В некоторых случаях остатки рабочей рамки у Fv иммуноглобулина человека заменены соответствующими нечеловеческими остатками. Более того, очеловеченные антитела могут включать в себя остатки, которые не находят ни в антителах реципиента, ни во внесенных ООК, или последовательностях рабочей рамки. Эти модификации сделаны для дальнейшего усовершенствования и упорядочивания представления антител. В основном, очеловеченные антитела будут включать существенно все из, по крайней мере, одного и обычно двух изменяемых доменов, в которых все или существенно все ООК области соответствуют тем нечеловеческим иммуноглобулинам, и все или существенно все FR области являются теми согласованным последовательностям иммуноглобулина человека. Очеловеченные антитела оптимально будут также включать, по крайней мере, часть постоянной области иммуноглобулина (Fc), которая типична для иммуноглобулина человека. Для дальнейших подробностей см.: Jones et al., Nature, 321: 522-525 [1986]; Reichmann et al., Nature, 332: 323-329 [1988]; и Presta, Curr. Op. Struct. Biol., 2: 593-596 [1992]).
"He иммуногенный у человека" означает, что при условиях контактирования полипептида на фармацевтически приемлемом носителе и в терапевтически эффективных количествах с подходящей тканью человека состояние чувствительности или устойчивости к полипептиду вплоть до второго введения полипептида после надлежащего латентного периода (а именно, от 8 до 14 дней) не проявляется.
II. Предпочтительные воплощения изобретения
Предпочтительные пептиды настоящего изобретения являются существенно гомогенным полипептидом(ами), касаясь как лиганда(ов) mpl или тромбопоэтина (ТП), которые проявляют свойство связывания с mpl, членом суперсемейства цитокиновых рецепторов, обладающие биологическим свойством стимуляции включения меченых нуклеотидов (3H-тимидина) в ДНК ИЛ-3 зависимых Ba/F3 клеток, трансфецированных mpl P человека. Более предпочтительным лигандом(ами) mpl является выделенный белок(ки), обладающие гематопоэтической, в особенности мегакариоцитопоэтической или тромбоцитопоэтической активностью - а именно, способные к стимуляции пролиферации, созреванию и/или дифференциации незрелых мегакариоцитов или их предшественников в зрелые продуцирующие тромбоциты формы. Большинство предпочтительных полипептидов настоящего изобретения являются лигандом(ами) mpl человека, включая их фрагменты, обладающие гематопоэтической, мегакариоцитопоэтической или тромбоцитопоэтической активностью. Этот лиганд(ы) mpl человека необязательно лишены гликозилирования. Другие предпочтительные лиганды mpl человека являются "доменом ЭП" МЛч, касаясь как МЛч153 или ТПч153, усеченной формы МЛч, так и касаясь МЛЧ245 или ТПЧ245, и зрелым полипептидом полной длины, имеющим аминокислотную последовательность, показанную на фиг.1 (Посл. No:1), касаясь как МЛч, МЛч332 или ТПч332 и вариантов биологически активных заменителей МЛч(R153А, R154A).
Необязательно предпочтительными полипептидами настоящего изобретения являются биологически или иммунологически активные варианты лигандов mpl, выбранные среди МЛч2, МЛч3, МЛч4, МЛм, МЛм2, МЛм3, МЛс иМЛс2.
Необязательно предпочтительными полипептидами настоящего изобретения является биологически активный вариант(ы) лиганда mpl, который имеет аминокислотную последовательность, обладающую, по крайней мере, 70% идентичной аминокислотной последовательности с лигандом mpl человека (см. фиг.1 [Посл. No: 1]), лиганд mpl мыши (см. фиг.16 [Посл. No: 12 и 13]), рекомбинантный лиганд mpl свиньи (см. фиг.19 [Посл. No: 18]), или лиганд mpl свиньи, выделенный из апластической плазмы свиньи, предпочтительно, по крайней мере, 75%, более предпочтительно, по крайней мере, 80%, еще более предпочтительно, по крайней мере, 85%, и еще более предпочтительно, по крайней мере, 90%, и наиболее предпочтительно, по крайней мере, 95%.
Лиганд mpl, выделенный из апластической плазмы свиньи, имеет следующие характеристики:
(1) Частично очищенный лиганд элюируется с колонки при гель фильтрации как PBS; PBS, содержащим 1% SDS или PBS, содержащим 4М MgCl2, при Mr 60000-70000;
(2) Активность лиганда нарушается проназой;
(3) Лиганд стабилен при низком рН (2,5), SDS до 0,1% и 2М мочевине;
(4) Лиганд является гликопротеином, что основано на его способности связываться к различными лектиновыми колонками;
(5) Высокоочищенный лиганд элюируется при SDS-PAGE в невосстанавливающих условиях с Mr 25000-35000. Меньшее количество активности также элюируется при Mr 18000-22000 и 60000;
(6) Высокоочищенный лиганд разделяется при восстанавливающих условиях при SDS-PAGE в виде дублета с Mr 28000 и 31000;
(7) Амино-концевая последовательность полос при 18000-22000, 28000 и 31000 является одинаковой SPAPPACDPRLLNKLLRDDHVLHGR (Посл. No: 29); и
(8) Лиганд связывается и элюируется со следующих колонок
Blue-Sepharose,
CM Blue-Sepharose,
MONO-Q,
MONO-S,
Лентил-лектин-Сефароза,
WGA-Sepharose,
Кон-А-Сефароза,
Эфир 650м Toyoperl,
Бутил 650 м Toyoperl,
Фенил 650 м Toyoperl, и
Фенил-Сефароза.
Более предпочтительными полипептидами лиганда mpl являются те, которые кодируют геномную или кДНК, имеющую аминокислотную последовательность, описанную на фиг.1 (Посл. No: 1).
Другие предпочтительные естественно встречающиеся биологически активные полипептиды лиганда mpl настоящего изобретения включают лиганд препро-mpl, лиганд про-mpl, зрелый лиганд mpl, фрагменты лиганда mpl и их гликозилированные варианты.
Еще другие предпочтительными полипептидами настоящего изобретения включают последовательности вариантов и химер лиганда mpl. Обычно предпочтительной последовательностью вариантов и химер лиганда mpl являются биологически активные варианты лиганда mpl, которые имеют аминокислотную последовательность, обладающую, по крайней мере, 70% идентичностью последовательности с лигандом mpl человека, или лигандом mpl, выделенным из апластической плазмы свиньи, предпочтительно, по крайней мере, 75%, более предпочтительно, по крайней мере, 80%, еще более предпочтительно, по крайней мере, 85%, и еще более предпочтительно, по крайней мере, 90%, и наиболее предпочтительно, по крайней мере, 95%. Примером предпочтительного варианта лиганда mpl является N-концевой домен варианта МЛч (называемый как "ЭП-домен" из-за его гомологии с эритропоэтином). Предпочтительный ЭП-домен МЛч включает около 153 первых аминокислотных остатков зрелого МЛч и обозначен как МЛч153. Предпочтительная последовательность варианта МЛч, необязательно, включает одну, в которой один или более основной или два основных аминокислотных остатка(ков) в С-концевом домене являются замененными на неосновной аминокислотный остаток(ки) (например, гидрофобный, нейтральный, кислый, ароматический, Гли, Про и подобный). Предпочтительная последовательность С-концевого домена варианта МЛч включает одну, в которой остатки Арг 153 и 154 заменены на остатки Aла. Этот вариант обозначен как МЛч332 (R153A, R154A). Альтернативно предпочтительный вариант МЛч включает либо МЛч332 или МЛч153, в которых аминокислотные остатки 111-114 (QLPP или LPPQ) удалены или заменены различными тетрапептидными последовательностями (например, AGAG или подобная). Последующие мутанты с делениями обозначены как Δ4МЛч332 или Δ4МЛч153.
Предпочтительной химерой является слияние между лигандом mpl или его фрагментом (определенным ниже) с гетерологическим полипептидом или его фрагментом. Например, МЛч153 может быть слит с фрагментом IgG для улучшения времени полужизни в плазме или с ИЛ-3, Г-КСФ или ЭП для получения молекулы с увеличенной тромбопоэтической или химерной гематопоэтической активностью.
Альтернативно предпочтительным химерным лигандом mpl человека является "химера МЛ-ЭП домен", включающий N-концевые от 153 до 157 остатки, замененные на один или более, но не все, остатки ЭП человека, расположенные в ряд, как показано на фиг. 10 (Посл. No: 7). В настоящем воплощении химера МЛч была бы около 153-166 остатков длиной, в которых отдельные или сгруппированные в блок остатки из последовательности ЭП человека добавлены или заменены на последовательность МЛч в положениях, соответствующих расположению в ряд, показанному на фиг. 10 (Посл. No: 6). Примерная блок-последовательность, вводимая в N-конецевую часть МЛч, включала бы один или более участков N-гликозилирования в положениях (ЭП) 24-26, 38-40 и 83-85; один или более предсказанных алифатичеких α-спиральных складок в положениях (ЭП) 9-22, 59-76, 90-107 и 132-152; и другие высококонсервативные области, включая N-концевую и С-концевую области, и положения остатков (ЭП) 44-52 (см., например. Wen et al., Blood, 82: 1507-1516 [1993] и Boissel et al., J. Biol. Chem., 268(21): 15983-15993 [1993]). Предполагается, этот "химера МЛ-ЭП домен" будет иметь смешанную тромбопоэтическую-эритропоэтическую (ТЭП) биологическую активность.
Другие предпочтительные полипептиды настоящего изобретения включают фрагменты лиганда mpl, имеющие порядковую последовательность, по крайней мере, 10, 15, 20, 25, 30 или 40 аминокислотных остатков, которые идентичны последовательности лиганда mpl, выделенного из апластической плазмы свиньи или лиганда mpl человека, описанных здесь (см., например, Таблицу 14, Пример 24). Предпочтительный фрагмент лиганд mpl является лигандом человека, где МЛ[1-Х], где Х является 153, 164, 191, 205, 207, 217, 229 или 245 (см. фиг. 1 [Посл. No: 1] для последовательности остатков 1-Х). Другие предпочтительные фрагменты лиганда mpl включают те, которые получены в результате химического или ферментативного гидролиза или расщепления очищенного лиганда.
Другим предпочтительным аспектом изобретения является способ очистки молекулы лиганда mpl, включающий контактирование источника лиганда mpl, содержащего молекулы лиганда mpl, с иммобилизованным рецепторным полипептидом, особенно mpl, или слитого полипептида mpl, в условиях, вследствие которых очищаемые молекулы лиганда mpl являются селективно адсорбированными на иммобилизованном рецепторном полипептиде, промывание иммобилизованной подложки для удаления неадсорбированного материала, и элюцию очищаемых молекул с иммобилизованного рецепторного полипептида буфером элюции. Когда иммобилизованный рецептор является предпочтительно слитым mpl-IgG, источником, содержащим лиганд mpl, может быть плазма.
Альтернативно, источником, содержащим лиганд mpl, является рекомбинантная культура клеток, где концентрация лиганда mpl и в культуральной среде, и в лизате клеток, в целом, выше, чем в плазме или другом природном источнике. В этом случае вышеописанный mpl-IgG иммуноаффинный способ, хотя, по-прежнему, полезный, обычно не необходим, могут быть применены известные в технике более традиционные способы очистки белков. В кратком виде предпочтительный способ очистки, обеспечивающий получение существенно гомогенного лиганда mpl, включает: удаление частичек остатков как клеток хозяина, так и фрагментов лизата, например, центрифугированием или ультрафильтрацией; белок, необязательно, может быть сконцентрирован при помощи коммерчески доступного фильтра для концентрации белка; последующее разделение лиганда от других примесей при помощи одного или более этапов, выбранных из: иммуноаффинной, ионообменной (например, ДЭАЭ или на матриксе, содержащем карбоксиметильные или сульфгидрильные группы), Blue-Sepharose, CM Blue-Sepharose, MONO-Q, MONO-S, Лентил лектин-Сефарозе, WGA-Sepharose, Кон А-Сефарозе, Эфир-Toyoperl, Бутил-Toyoperl, Фенил-Toyoperl, протеин А Сефарозе, SDS-PAGE, ЖХВР с обращенной фазой (а именно, силикагель с добавленными алифатическими группами) хроматографии, или хроматографии на молекулярных ситах из Сефадекса, или хроматографией по размеру; и осаждение этанолом или сульфатом аммония. На любом из предшествующих этапов для ингибирования протеаз может быть включен ингибитор протеаз, такой как метилфенилсульфонилфторид (МФСФ).
В другом предпочтительном воплощении настоящее изобретение описывает выделение антитела, способного связываться с лигандом mpl. Предпочтительное выделенное антитело к лиганду mpl является моноклональным (Kohler and Milstein, Nature, 256: 495-497 [1975]; Campbell, Laboratory Techniques in Biochemistry and Molecular Biology, Burdon et al., Eds, Volume 13, Elsevier Science Publishers, Amsterdam [1985]; и Huse et al., Science, 246: 1275-1281 [1989]). Предпочтительным выделенным антителом к лиганду mpl является то, которое связывается к лиганду mpl со сродством, по крайней мере, около 106 л/моль. Более предпочтительно, антитело связывается со сродством, по крайней мере, около 107 л/моль. Более предпочтительно, антитело, вырабатывается против лиганда mpl, имеющего одну из вышеописанных эффекторных функций. Выделенное антитело, способное к связыванию с лигандом mpl, может быть, необязательно, сплавлено со вторым полипептидом, и антитело, или их сплав, может быть использован для выделения и очистки лиганда mpl из источника, описанного выше для иммобилизованного полипептида mpl. В дальнейшем предпочтительном аспекте настоящего воплощения изобретение описывает способ для обнаружения лиганда mpl in vitro или in vivo, включающего контактирование антитела с образцом, в особенности с образцом сыворотки, подозреваемым на содержание лиганда, и обнаружение, если наблюдается связывание.
Еще далее, в предпочтительном воплощении изобретение описывает выделение молекулы нуклеиновой кислоты, кодирующей лиганд mpl или его фрагмент, молекула нуклеиновой кислоты которого может быть мечена или не мечена по обнаруживаемому радикалу, и молекула нуклеиновой кислоты, имеющая последовательность, которая комплементарна к, или гибридизуется в строгих или умеренно строгих условиях с молекулой нуклеиновой кислоты, имеющей последовательность, кодирующую лиганд mpl. Предпочтительной нуклеиновой кислотой лиганда mpl является РНК или ДНК, которая кодирует биологически активный лиганд mpl, обладающий, по крайней мере, 75% идентичной последовательности, более предпочтительно, по крайней мере, 80%, еще более предпочтительно, по крайней мере, 85%, и еще более предпочтительно, по крайней мере, 90%, и наиболее предпочтительно, по крайней мере, 95% идентичной последовательности с лигандом mpl. Более предпочтительной молекулой выделенной нуклеиновой кислоты является последовательность ДНК, кодирующая биологически активный лиганд mpl, выбранная из: (а) ДНК, основанная на кодирующей области гена лиганда mpl млекопитающих (а именно, ДНК, включающая последовательность нуклеотидов, представленная на фиг.1 (Посл. No: 2) (или комплемент или ее фрагмент) в условиях с низкой до умеренной жесткости. Таким образом, дальнейший аспект настоящего изобретения включает ДНК, которое гибридизуется в условиях с низкой до умеренной жесткостью с ДНК, кодирующей полипептиды лиганда mpl.
В дальнейшем предпочтительном воплощении настоящего изобретения молекулой нуклеиновой кислоты является кДНК, кодирующая лиганд mpl, и далее, включающая реплицирующийся вектор, в котором кДНК оперативно сцеплена с контрольной последовательностью, распознающейся хозяином, трансформированным вектором. Данный аспект далее включает клетки хозяина, трансформированные вектором, и способ использования кДНК для воздействия на продукцию лиганда mpl, включающий экспрессию кДНК, кодирующую лиганд mpl в культуре трансформированных клеток хозяина, и выделение лиганда mpl из культуры клеток хозяина. Лиганд mpl, полученный таким образом, является предпочтительно, существенно гомогенным лигандом mpl человека. Предпочтительными клетками хозяина для продуцирования лиганда mpl являются клетки яичника хомяка (СНО).
Изобретение далее включает предпочтительный способ для лечения млекопитающего, имеющего иммунологическое или гематопоэтическое заболевание, в особенности, тромбоцитопению, включающий введение млекопитающему терапевтически эффективного количества лиганда mpl. Лиганд mpl, необязательно, вводится в сочетании с цитокином, в особенности, с колоние-стимулирующим фактором или интерлейкином. Предпочтительные колоние-стимулирующие факторы или интерлейкины включают: kit-лиганд, ФИЛ, Г-КСФ, ГМ-КСФ, М-КСФ, ЭП, ИЛ-1, ИЛ-2, ИЛ-3, ИЛ-5, ИЛ-6, ИЛ-7, ИЛ-8, ИЛ-9 или ИЛ-11.
III. Способы получения
Продукция тромбоцитов, с давних пор исследуемая некоторыми авторами, находится под контролем многочисленных специфических дифференцирующих гуморальных факторов. Было постулировано, что две различных активности цитокина, обозначаемых как колоние-стимулирующий фактор (мег-КСФ) и тромбопоэтин, регулируют мегакариоцитопоэз и тромбоцитопоэз (Williams et al., J. Cell Physiol., 110: 101-104 [1982]; Williams et al., Blood Cells, 15: 123-133 [1989]; и Gordon et al. Blood, 80: 302-307 [1992]). В соответствии с этой гипотезой мег-КСФ стимулирует пролиферацию прогениторных мегакариоцитов, тогда как тромбопоэтин сначала воздействует на созревание более дифференцированных клеток и, в конечном счете, на высвобождение тромбоцитов. Начиная с 1960-х годов данные об индукции и обнаружении обеих активностей - мег-КСФ и тромбопоэтина в плазме, сыворотке и моче животных и человека, после тромбоцитопенических приступов, хорошо документированы (Odell et al., Proc. Soc. Exp. Biol. Med., 108: 428-431 [1961]; Nakeff et al., Acta Haematol., 54: 340-344 [1975]; Specter, Proc. Soc. Exp. Biol., 108: 146-149 [1961]; Schreiner et al., J. Clin. Invest., 49: 1709-1713 [1970]; Ebbe, Blood, 44: 605-608 [1974]; Hoffman et al., N. Engi. J. Med., 305: 533 [1981]; Straneva ey al., Exp. Hematol., 17: 1122-1127 [1988]; Mazur et al., Exp. Hematol., 13: 1164 [1985]; Mazur et al., J. Clin. Invest., 68: 733-741 [1981]; Sheiner et al., Blood, 56: 183-188 [1980]; Hill et al., Exp. Hematol., 20: 354-360 [1992]; и Hegyi et al.. Int. J. Cell. Cloning, 8: 236-244 [1990]). Было сообщено, что эти активности проявляют специфичность по отношению к дифференцирующимся линиям и отличны от активностей известных цитокинов (Hill R.J. et al., Blood, 80: 346 [1992]; Erickson-Miller C.L. et al., Brit. J. Haematol., 84: 197-203 [1993]; Staneva J.E. et al., Exp. Hematol., 20: 4750 [1992]; и Tsukada J. et al., Blood, 81: 866-867 [1993]). После этого попытки очистить мег-КСФ или тромбопоэтин из тромбоцитопенической плазмы или мочи были безуспешными.
Согласно с вышеупомянутыми наблюдениями, описывающими тромбоцитопеническую плазму, было обнаружено, что апластическая плазма свиньи (АПС), полученная от облученных свиней, стимулирует мегакариоцитопоэз у человека in vitro. Было обнаружено, что данная стимуляторная активность предотвращается растворимым внеклеточным доменом с-mpl, подтверждая, что АПС является потенциальным источником естественного лиганда mpl (МЛ). В настоящее время лиганд mpl успешно очищен из плазмы АПС, а информация об аминокислотной последовательности была использована для выделения кДНК МЛ мыши, свиньи и человека. Эти последовательности МЛ гомологичны эритропоэтину и обладают и мег-КСФ, и тромбопоэтино-подобной активностью.
1. Очистка и идентификация mpl лиганда из плазмы
Как было изложено выше, апластическая плазма различных видов, как сообщалось, содержит активности, которые стимулируют гематопоэз in vitro, однако, как сообщалось ранее, никакого гематопоэтического стимуляторного фактора из плазмы не выделяли. Один источник апластической плазмы является тот, который получают из облученных свиней. Данная апластическая плазма свиньи (АПС) стимулирует гематопоэз у человека in vitro. Для определения, содержит ли АПС лиганд mpl, ее эффект был исследован по измерению включения 3H-тимидина в Ba/F3 клетки, трансфецированные mpIP (Ba/F3-mpl) человека, способом, показанном на фиг.2. АПС стимулирует включение 3H-тимидина в Ba/F3-mpl клетки, но не в контрольные Ba/F3 клетки (т.е., нетрансфецированные mpl P человека). Кроме этого, никакой другой активности в обычной плазме свиньи не наблюдали. Эти результаты отмечают, что АПС содержит фактор или факторы, которые передают пролиферативный сигнал через mpl рецептор и, следовательно, могут быть естественным лигандом для данного рецептора. Это было далее подтверждено обнаружением того, что обработка АПС растворимым mpl-IgG подавляет стимуляторное действие АПС на Ba/F3-mpl клетки.
Активность АПС, по-видимому, представляет белок, поскольку проназа, ДТТ или нагревание разрушают активность АПС (фиг.3). Активность также является недиализуемой. Однако активность была стабильной при низких значениях рН (рН 2,5 в течение 2,5 часов), и было показано связывание и элюция с различных лектинаффинных колонок, отмечая, что она являлась гликопротеином. В дальнейшем, чтобы пролить свет на структуру и идентификацию данной активности, она была очищена из АПС по сродству, используя mpl-IgG химеру.
АПС обработали согласно протоколу, изложенному в Примерах 1 и 2. В кратком виде mpl лиганд очистили, используя хроматографию по гидрофобному взаимодействию (ГХ), хроматографию с иммобилизованным красителем и хроматографию mpl по сродству. Выход активности на каждом этапе показан на фиг.4, а степень очистки представлена в Таблице 1. Общий выход активности после mpl-аффинной колонки составлял приблизительно 10%. Фракции с пиком активности (F6) после mpl-аффинной колонки имели согласно определению специфическую активность 9,8×106 единиц/мг. Общая очистка из 5 литров АПС составляла приблизительно 4×106 раз (от 0,8 единиц/мг до 3,3×106 единиц/мг), с 86×10° степени потери по белку (от 250 г до 3 мкг). Была определена специфическая активность лиганда, элюированного с mpl-аффинной колонки, составляющая 3×106 единиц/мг.
ТАБЛИЦА 1
Очистка mpl лиганда
Белок определяли по методу Bradford. Концентрацию белка во фракциях 5-7 с элюированным mpl измеряли на основе интенсивности окраски серебром SDS-геля. Одну единицу активности определяли как способность вызывать 50% эффекта от максимальной стимуляции пролиферации Ba/F3-mpl клеток.
Анализ фракций после элюции с mpl-аффинной колонки на SDS-PAGE (4-20%, Novex гель), разогнанных при восстанавливающих условиях, выявил наличие нескольких белков (фиг.5). Белки, которые окрашивались серебром более интенсивно, дали разрешение при кажущемся Mr 60000, 55000, 30000, 28000 и 18000-22000. Для определения, какой из этих белков стимулирует пролиферацию Ba/F3-mpl клеток, белки элюировали из геля как описано в Примере 2.
Результаты данных экспериментов показали, что большая активность элюируется с нарезанного геля, который включал белки с Mr 28000-32000, меньшая активность элюировалась из области геля с 18000-22000 (фиг.6). Единственные белки, видимые в данной области имели Mr 30000, 28000 и 18000-22000. Для определения и получения белковой последовательности белков, проявившихся в этой области геля (а именно, полосы при 30, 28 и 18-22 kDa), три этих белка подвергли электроблоттингу на PVDF и секвенировали, как описано в Примере 3. Полученная аминокислотная последовательность представлена в Таблице 2.
ТАБЛИЦА 2
Амино-концевые последовательности mpl лиганда
Анализ с применением компьютера выявил новизну этих аминокислотных последовательностей. Поскольку все три последовательности были близкими, было предположено, что 30, 28 и 18-22 kDa белки являются родственными и, возможно, являются различными формами одного и того же нового белка. Более того, этот белок(и) был вероятным кандидатом естественного mpl лиганда, поскольку активности, разогнанные на SDS-PAGE, были в тех же самых областях (28000-32000) 4-20% геля. Помимо этого частично очищенный лиганд мигрирует с Mr 17000-30000, когда его подвергают гель-хроматографии на колонке с Superose 12 (Pharmacia). Было предположено, что различные Mr формы лиганда являются результатом протеолиза или различий в гликозилировании или других пост- или пре-трансляционных модификаций.
Как описано ранее, антисенсная РНК mpl человека отменяет мегакариоцитопоэз в культуре клеток костного мозга человека, обогащенных CD 34+ прогениторными клетками без воздействия на дифференцировку других дифференцирующихся клеточных линий (Methia et al., там же). Эти результаты подтверждают, что mpl рецептор может играть роль в дифференцировке и пролиферации мегакариоцитов in vitro. Для дальнейшего прояснения роли лиганда mpl в мегакариоцитопоэзе были сравнены эффекты АПС и АПС, лишенной лиганда mpl, на мегакариоцитопоэз человека in vitro. Действие АПС на мегакариоцитопоэз человека определяли используя модификацию исследования мегакариоцитопоэза в жидкой суспензии, описанной в Примере 4. В данном исследовании периферические стволовые клетки человека (ПСК) обработали АПС до и после mpl-IgG аффинной хроматографии. Стимуляцию мегакариоцитопоэза по активности GPIIbIIIa подсчитывали при помощи 125I-анти-IIbIIIа антитела (фиг.7). Как показано на фиг.7, 10% АПС вызывает приблизительно 3-кратную стимуляцию, тогда как АПС, лишенная лиганда mpl, не обладала эффектом. Примечательно, что АПС, лишенная лиганда mpl, не индуцировала пролиферацию Ba/FS-mpI клеток.
В другом эксперименте растворимый mpl-IgG человека, добавленный в 0, 2 и 4 дни к культуре, содержащей 10% АПС, нейтрализовал стимуляторный эффект АПС на мегакариоцитопоэз человека (фиг.8). Данные результаты отмечают, что mpl лиганд играет роль в регуляции мегакариоцитопоэза у человека и, следовательно, может быть использован для лечения тромбоцитопении.
2. Молекулярное клонирование mpl лиганда
Основываясь на аминокислотной последовательности амино-конца, полученной из белков 30, 28 и 18-22 kDa (см. Таблицу 2 выше), были сконструированы два олигонуклеотидных праймерных пула, использованных для амплификации геномной ДНК свиньи методом РПЦ. Было рассуждено, что, если аминокислотная последовательность амино-конца кодируется единым эксоном, тогда было бы ожидаемо, что правильный PCR продукт будет длиной 69 пар оснований. Фрагмент ДНК данного размера был найден и субклонирован в pGEMT. Последовательность олигонуклеотидного РПЦ праймера и полученных трех клонов представлены в Примере 5. Аминокислотная последовательность (PRLLNKLLR [Посл. No: 33]) пептида, промежуточно кодируемого праймерами, была идентичной к той полученной при белковом секвенсе амино-конца лиганда свиньи (см. остатки 9-17 для последовательности белков 28 и 30 kDa свиньи выше).
Синтетический олигонуклеотид, полученный на основе последовательности РПЦ фрагмента, использовали для скрининга геномной библиотеки ДНК человека. Был сконструирован и синтезирован олигонуклеотид из 45 оснований, обозначенный как pR45, на основе последовательности фрагмента РПЦ. Данный нуклеотид имел следующую последовательность:
5’GCC-GTG-AAG-GAC-GTG-GTC-ACG-AAG-CAG-TTT-ATT-TAG-GAG-TCG 3’ (Посл. No: 34).
Данный нуклеотид использовали для скрининга геномной библиотеки ДНК человека в λgem12 при низкой строгости гибридизации, и отмывочных условиях в соответствии с Примером 6. Положительные клоны были отобраны, плашки очищены и анализированы методами рестрикционного картирования и southern блоттинга. Фрагмент 390 пар оснований EcoRI-Xbal, который гибридизовали с 45-и членом, субклонировали в pBluescript SK-. Секвенирование ДНК этого клона подтвердило, что данная ДНК кодирует выделенный лиганд mpl человека, гомологичный свиному. Последовательность ДНК человека и выведенная аминокислотная последовательность представлены на фиг.9 (Посл. No: 3 и 4). Предсказанные положения интронов геномной последовательности, также отмеченные стрелками, определяют естественный эксон ("эксон 3").
Основываясь на последовательности "эксона 3" человека (Пример 6), был синтезирован олигонуклеотид, соответствующий 3’ и 5’ концам последовательности эксона. Эти два праймера были использованы в выделенный mpl лиганд человека РПЦ реакции, использующей в качестве матрицы кДНК, полученную из различных тканей человека. Ожидаемые размеры правильного продукта РПЦ составляли 140 пар оснований. После анализа продукта РПЦ в 12% полиакриламидном геле был обнаружен фрагмент кДНК ожидаемого размера в библиотеке кДНК, полученной из почек взрослого человека, клеток 293 эмбриональной почки человека, и кДНК, полученной из эмбриональной печени человека.
Следующей была просеяна библиотека кДНК эмбриональной печени (7×106 клонов) в ламбда DR2 с тем же самым 45-и членным олигонуклеотидом, используемым для скрининга геномной библиотеки человека и для библиотеки кДНК эмбриональной печени при низкой строгости гибридизационных условий. Один клон с включением 1,8 тыс. оснований был отобран для дальнейшего анализа. Используя процедуры, описанные в Примере 7, были получены нуклеотидная и выведенная аминокислотная последовательность лиганда mpl человека (МЛч). Эти последовательности представлены на фиг.1 (Посл. No: 1 и 2).
3. Структура лиганда mpl человека (МЛч)
Последовательность кДНК лиганда mpl человека (МЛч) (фиг.1 [Посл. No: 2]) содержит 1774 нуклеотида, следующих за окончанием поли(А). Он содержит 215 нуклеотидов 5’ нетранслируемой последовательности, и 3’ нетранслируемую область из 498 нуклеотидов. Предполагаемый инициирующий кодон при (216-218) положении нуклеотидов располагается внутри согласованной последовательности, благоприятной для инициации трансляции у эукариот. Открытая рамка считывания составляет 1059 нуклеотидов длиной и кодирует полипептид из 353 аминокислотных остатков, начинающийся на 220 нуклеотидном положении. N-конец предсказанной аминокислотной последовательности является высокогидрофобным и, вероятно, соответствует сигнальному пептиду. Компьютерный анализ предсказанной аминокислотной последовательности (von Heijne et al., Eur. J. Biochem., 133: 17-21 [1983]) отмечает места потенциального расщепления для сигнальной пептидазы между остатками 21 и 22. Расщепление в этой позиции дало бы зрелый полипептид из 332 аминокислотных остатков, начинающийся с амино-концевой последовательности, полученной из лиганда mpl, очищенного из плазмы свиньи. Предсказанный молекулярный вес негликозилированного лиганда из 332 аминокислотных остатков составляет около 38 kDa. Имеется 6 потенциальных участков N-гликозилирования и 4 цистеиновых остатка.
Сравнение последовательности лиганда mpl с последовательностью банка данных Genbank выявило 23% идентичности между 153 аминоконцевыми остатками зрелого mpl лиганда человека и эритропоэтином (фиг. 10 [Посл. No: 6 и 7]). Если принять во внимание консервативные замены, данная область МЛч проявляет 50% сходства с эритропоэтином человека (ЭПч). Оба ЭПч и МЛч содержат четыре цистеина. Три из 4 цистеинов консервативны у МЛч, включая первый и последний цистеины. Эксперименты по точечно направленному мутагенезу показали, что первый и последний цистеины эритропоэтина образуют дисульфидную связь, которая требуется для функционирования (Wang, F.F. et al., Endocrinology, 116: 2286-2292 [1983]). По аналогии, первый и последний цистеины МЛч могут также образовывать значимую дисульфидную связь. Ни одно из гликозилирумых участков не является консервативным у МЛч. Все возможные участки гликозилирования МЛч расположены в карбокси-концевой половине полипептида МЛч.
Подобно ЭПч, мРНК МЛч не содержит ни согласованной последовательности полиаденилирования AAUAAA, ни регуляторного элемента AUUUA, который присутствует в 3’ нетранслируемой области многих цитокинов, и, тем самым, влияет на стабильность мРНК (Shaw et al., Cell, 46: 659-667 [1986]). Northern блот анализ выявил низкий уровень единого в 1,8 тыс. оснований РНК транскрипта МЛч и в эмбриональной, и во взрослой печени. После долгого выдерживания можно было определить и во взрослой почке слабую полосу того же самого размера. Для сравнения, эритропоэтин человека экспрессировался в эмбриональной печени, во взрослых почке и печени (Jacobs et al., Nature, 313: 804-809 [1985] и Boudurant et al., Molec. Cell. Biol., 6: 2731-2733 [1985]) в ответ на гипоксию.
Важность С-концевой области МЛч остается необходимой для разъяснения. Основываясь на наличии шести возможных мест для N-связанного гликозилирования и способности лиганда связываться с лектин-аффинной колонкой, эта область МЛч, по-видимому, является гликозилированной. В некоторых экспериментах по элюции с геля наблюдалась активность при разрешении с Mr около 60000, которая может представлять гликозилированную молекулу полной длины. С-концевая область, следовательно, может стабилизировать и увеличивать время полужизни циркулирующего МЛч. В случае эритропоэтина негликозилированная форма обладает полной биологической активностью in vitro, но имеет заметно уменьшенное время полужизни в плазме по сравнению с гликозилированным эритропоэитном (Takeuchi et al., J. Biol. Chem., 265: 12127-12130 [1990]; Narhi et al., J. Biol. Chem., 266: 23022-23026 [1991] и Spivack et al., Blood, 7: 90-99 [1989]). С-концевой домен МЛч содержит две последовательности из двух основных аминокислот [Арг-Арг структуру в положениях 153-154 и 245-246], которые могли бы служить как возможные участки для процессинга. Расщепление по этим участкам могло бы быть ответственным за образование 30, 28 и 18-22 kDa форм МЛ, выделенного из АПС. Важно, что последовательность Apr153-Apr154 наблюдается сразу же после эритропоэтин-подобного домена МЛ. Эти наблюдения отмечают, что МЛ полной длины может представлять собой предшественник, который подлежит ограниченному протеолизу с образованием зрелого лиганда.
4. Изоформы и варианты mpl лиганда человека
Изоформы, или альтернативно сплайсинговые формы, mpl лиганда человека были определены методом РПЦ во взрослой печени человека. В кратком виде, были синтезированы праймеры, соответствующие каждому концу, так же как и выбранным областям, кодирующим последовательность МЛч. Эти праймеры были использованы в ОТ-РПЦ для умножения РНК взрослой печени человека, как описано в Примере 10. В добавление к полной форме, обозначенной МЛч, были обнаружены или выведены три других формы, обозначенные МЛч2, МЛч3 и МЛч4. Выведенная зрелая аминокислотная последовательность всех четырех изоформ представлена на фиг.11 (Посл. No: 6, 8, 9, и 10). МЛч3 имеет 116 нуклеотидную делецию в положении 700, которая приводит и к аминокислотной делеции, и к сдвигу рамки. Теперь кДНК кодирует зрелый полипептид, который является длиной в 265 аминокислот, и происходит от последовательности МЛч со 139 аминокислотного остатка. Наконец, МЛч4 имеет как делецию в 12 (пар) нуклеотидов после 618 положения нуклеотида (также обнаруженную в последовательности у мыши и свиньи [см. ниже]), так и делецию в 116 пар оснований, найденную у МЛч3. Хотя у (обозначенной МЛч2) человека не было выделено клонов с делецией только в 12 пар оснований (после 619 нуклеотида), эта форма, предположительно, существует, поскольку такая изоформа была идентифициована и у мыши, и у свиньи (см. ниже) и к тому же она была идентифицирована в конъюгации со 116 нуклеотидной делецией в МЛч4.
Для определения, действительно ли МЛ полной длины необходим для биологической активности, были сконструированы и вариант МЛч с замещением, в котором последовательность из двух основных аминокислот Apr153-Apr154 заменена на два остатка аланина, и усеченная форма МЛч, "ЭП-домен. Используя РПЦ, как описано в Примере 10, был сконструирован замененный вариант с двумя основными аминокислотами в последовательности Apr153-Apr154, обозначенный как МЛч(В.153А, R154A). Используя РПЦ, была также создана усеченная форма "ЭП-домена" МЛч153, путем введения стоп кодона после Арг153.
5. Экспрессия рекомбинантного лиганда mpl человека (рМЛч) в транзиторно трансфецированных клетках эмбриональной почки человека (293)
Для подтверждения того, что клонированная кДНК человека кодирует лиганд для mpl, лиганд был экпрессирован в клетки 293 млекопитающих, применяя экспрессию векторами рRК5-МЛч или pRK5-МЛч153, в условиях непосредственного подавления раннего промотера цитомегаловируса. Было обнаружено, что супернатанты из транзиторно трансфецированных клеток 293 эмбриональной почки человека стимулируют включение 3H-тимидина в Ba/F3-mpl клетки, но не в родительские Ba/F3 клетки (фиг.12А). Среда от клеток 293, трансфецированных только pRK вектором, не содержит данной активности. Добавление mpl-IgG к среде предотвращает стимуляцию (данные не представлены). Эти результаты показывают, что клонированная кДНК кодирует функциональный МЛ человека (МЛч).
Для определения, способен ли "ЭП-домен" сам по себе связываться и активировать mpl, была экспрессирована усеченная форма МЛч, рМЛч153, в клетках 293. Было обнаружено, что супернатанты трансфецированных клеток обладают активностью, близкой к той, которая имеется в супернатантах клеток, экспрессирующих МЛч полной длины (фиг.12А), показывая, что С-концевой конец МЛ не требуется для связывания и активации с-mpl.
7. Мегакариоцитопоээ и mpl лиганд
Было предположено, что мегакариоцитопоэз регулируется на многих клеточных уровнях (Williams et al., J. Cell Physiol., 110: 101-104 [1982] и Williams et al., Blood Cells, 15: 123-133 [1989]). Это основано, главным образом, на наблюдении, что определенные гематопоэтические факторы роста стимулируют пролиферацию прогениторов мегакариоцитов, тогда как другие, в основном, проявляют действие на созревание. Данные, представленные здесь, подтверждают, что МЛ действует как пролиферативный фактор, так и как фактор созревания. То, что МЛ стимулирует пролиферацию прогениторов мегакариоцитов, подтверждается несколькими путями доказательств. Первое, АПС стимулирует и пролиферацию, и созревание мегакариоцитов человека in vitro, и эта стимуляция полностью подавляется mpl-IgG (фиг.7 и 8). Более того, ингибирование формирования колоний с-mpl антисенсными нуклеотидами (Methia et al., Blood, 82: 1395-1401 [1993]), и обнаружение того, что с-mpl может передавать пролиферативный сигнал в клетки, которые трансфецированы (Skoda et al., EMBO, 12: 2645-2653 [1993] и Vigon et al., Oncogene, 8: 2607-2615 [1993]), также отмечает, что МЛ стимулирует пролиферацию. Видимая экспрессия с-mpl в течение всех стадий дифференциации мегакариоцитов (Methia et al., Blood, 82: 1395-1401 [1993]) и способность рекомбинантного МЛ быстро стимулировать продукцию тромбоцитов in vivo отмечает, что МЛ также действует на созревание. Способность к активации у рекомбинантного МЛ создает возможности для более тщательной оценки его роли в регуляции мегакариоцитопоэза и тромбоцитопоэза, так же как и его способность влиять на другие гематопоэтические дифференцирующиеся линии.
8. Выделение гена лиганда mpl (ТП) человека
Клоны геномной ДНК человека гена ТП выделили методом просеивания геномной библиотеки человека в X-Geml2 с pR45 в условиях с низкой строгостью или в условиях с высокой строгостью с фрагментом, соответствующим 3’ половине кДНК человека, кодирующей лиганд mpl. Было выделено два перекрывающихся ламбда клона протяженностью 35 тыс. оснований. Были клонированы и секвенированы два перекрывающихся фрагмента (BamHl и EcoRI), содержащие полный ген ТП (см. фиг.14А, 14В и 14С).
Структура гена человека состоит из 6 эксонов по 7 тыс. оснований геномной ДНК. Связи всех соединений эксон/интрон состоят из согласованных структур, характерных для генов млекопитающих (Shapiro, M.B., et al., Nucl. Acids Res. 15: 7155 [1987]). Эксон 1 и эксон 2 содержат 5’ нетранслируемую последовательность и начальные четыре аминокислоты сигнального пептида. Оставшиеся для секреторного сигнала и первые 26 аминокислот зрелого белка кодируются 3 эксоном. Полный карбоксильный домен и 3’ нетранслируемый, так же как и -50 аминокислот эритропоэтин-подобного домена, кодируются эксоном 6. Четыре аминокислоты, включенные в делецию, наблюдающиеся между МЛч-2 (ТП-2), кодируются у 5’ конца эксона 6.
Анализ геномной ДНК человека при помощи Sauthern блота отметил, что ген ТП имеется в единственной копии. Хромосомное расположение гена было определено с помощью флуоресцентной гибридизации in situ (FISH), которой картировали хромосому 3q27-28.
9. Экспрессия и очистка ТП из клеток 293
Получение и очистка МЛ или ТП из клеток 293 подробно описана в Примере 19. В кратком виде, кДНК, соответствующая полному ТП с открытой рамкой, была получена при помощи РПЦ, используя pRK5-hmpl I. Продукт РПЦ очистили и клонировали между участками рестрикции Clal и Xbal плазмиды pRK5tkneo (производный вектор pRK5, модифицированный для экспрессии гена устойчивости к неомицину, в условиях подавления промотора киназы тимидина) для получения вектора pRK5tkneo.ORF (вектора, кодирующего полную открытую рамку считывания).
Второй вектор, кодирующий гомологичный домен ЭП, был получен таким же образом, но используя различные РПЦ праймеры для получения окончательной конструкции, названной pRK5-tkneoSn-A.
Эти две конструкции были трансфецированы в клетки эмбриональной почки человека при помощи СаРO4 метода и были отобраны клоны устойчивые к неомицину, которые выращивали до конфлуентного состояния. Экспрессия MЛ153 или МЛ332 в кондиционированной среде из данных клонов была оценена при помощи исследования пролиферации Ba/F3-mpl.
Очистку рМЛч332 провели, как описано в Примере 19. В кратком виде, 293-рМЛч332 кондиционированную среду нанесли на колонку с Blue-Sepharose (Pharmacia), которую сразу же промыли буфером, содержащим 2 М мочевину. Колонку элюировали буфером, содержащим 2 М мочевину и 1 М NaCl. Объем элюции после колонки с Blue-Sepharose затем непосредственно нанесли на колонку с WGA-Sepharose, промыли 10 объемами колонки буфера, содержащего 2 М мочевину и 1 М NaCl, и элюировали тем же буфером, содержащим 0,5 М N-ацетил-D-глюкозамин. Элюат с колонки с WGA-Sepharose нанесли на С4-ЖХВР колонку (Synchron, Inc.) и элюировали непрерывным градиентом пропанола. При электрофорезе в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE) очищенный 293-рМЛч332 мигрирует в геле в виде широкой полосы в области 68-70 kDa (см. фиг.15).
Очистку рКЛч153 также провели, как описано в Примере 19. В кратком виде, 293-pMЛч153 кондиционированную среду пропустили через колонку с Blue-Sepharose, как описано для рМЛч332. Элюат с колонки с Blue-Sepharose непосредственно нанесли на mpl-аффинную колонку, как описано выше. Элюированный с mpl-колонки рМЛч153, очистили до гомогенного состояния, используя С4-ЖХВР колонку, разогнав при тех же условиях, что и для рМЛч332. На SDS-PAGE очищенный рМЛч153 разогнался на 2 большие и 2 малые полосы с Mr около 18000-22000 (см. фиг.15).
10. Лиганд mpl мыши
Фрагмент ДНК, соответствующий области кодирования лиганда mpl человека, полученный при помощи РПЦ, очистили и пометили в присутствии 32Р-dАТФ и 32Р-dАТФ. Данные зонды использовали для скрининга 106 клонов библиотеки кДНК печени мыши в λGT10. Был выделен и секвенирован клон мыши (фиг.16 [Посл. No: 12 и 13]), содержащий 1443 пар оснований включения. Предполагаемый кодон инициации в положении 138-141 нуклеотидов находился в согласованной последовательности, благоприятной для инициации трансляции у эукариот (Kozak, M.J. Cell Biol., 108:229-241 [1989]). Последовательность определяется открытой рамкой считывания в 1056 нуклеотидов, которые предсказывают первичный продукт трансляции в 352 аминокислоты. Окаймляют данную открытую рамку считывания 137 5’ нуклеотидов и 247 3’ нуклеотидов несчитываемой последовательности. Окончание поли(А) после 3’ нетранслируемой области отсутствует, что отмечает, что клон, по-видимому, не завершен. N-конец предсказанной аминокислотной последовательности является очень гидрофобным и, по-видимому, представляет сигнальный пептид. Компьютерный анализ (von Heijne, G. Eur. J. Biochem., 133: 17-21 [1983]) отметил возможные места для разрыва для сигнальной пептидазы между 21 и 22 остатками. Разрыв в данном месте мог дать зрелый полипептид из 331 аминокислоты (35 kDa), идентичный МЛм331 (или МЛм2 по причине, описанной выше). Последовательность содержит 4 цистеина, все консервативны в последовательности человека, и 7 возможных участков N-гликозилирования, 5 из которых являются консервативными в последовательности человека. Снова, как у МЛч, все семь возможных участков N-гликозилирования расположены в С-концевой половине белка.
Когда сравнили с МЛ человека, была обнаружена значительная идентичность и нуклеотидной, и выведенной аминокислотной последовательности у "ЭП-доменов" данных МЛ. Однако когда выведенные аминокислотные последовательности МЛ человека и мыши расположили в ряд, выяснилось, что последовательность мыши имеет тетрапептидную делецию между остатками 111 и 114, соответствующую 12 нуклеотидной делеции после 618 положения нуклеотида, видимую у обеих кДНК, человека (см. выше) и свиньи (см. ниже). Поэтому были исследованы дополнительные клоны для обнаружения возможных изоформ МЛ мыши. Один клон кодировал выведенную 335 аминокислотную последовательность полипептида, содержащего "утерянный" тетрапептид LPLQ. Данная форма представляется МЛ мыши полной длины и обозначена как МЛм или МЛм335. Нуклеотидная и выведенная аминокислотная последовательность МЛм представлены на фиг.17 (Посл. No: 14 и 15). Данный клон кДНК содержит 1443 пары оснований, оканчивающихся поли(А) окончанием. Он проявляет открытую рамку считывания из 1068 пар оснований, окаймленных 134 основаниями 5’ и 241 основанием 3’ нетранслируемой последовательности. Предполагаемый кодон инициации лежит в положении нуклеотидов 138-140. Открытая рамка считывания кодирует предсказанный белок из 356 аминокислот, 21 первых которого являются высокогидрофобными и, по-видимому, функционируют как секреторный сигнал.
Наконец, третий выделенный и секвенированный клон мыши, было найдено, что он содержит 116 нуклеотидную делецию, соответствующую МЛч3. Эта изоформа мыши, следовательно, была обозначена МЛм3. Сравнение выведенных аминокислотных последовательностей этих двух изоформ показано на фиг.18 (Посл. No: 9 и 16).
Общая идентичность аминокислотной последовательности между МЛ человека и мыши (фиг.19 [Посл. No: 6 и 17]) составляет 72%, но эта гомология распределена неравномерно. Область, обозначенная как "ЭП-домен" (аминокислотные остатки 1-153 для последовательности человека и 1-149 для мыши) является более консервативной (86% гомологии), чем карбокси-концевая область белка (62% гомологии). Это может далее указывать, что только "ЭП-домен" является важным для биологической активности белка. Интересно, относительно двух структур из двух основных аминокислот, обнаруженных у МЛч, только структура из двух основных аминокислот, непосредственно следующая за "ЭП-доменом" (положение остатков 153-154) в последовательности человека, присутствует в последовательности мыши. Это находится в соответствии с возможностью, что МЛ полной длины может представлять белок предшественник, который подлежит ограниченному протеолизу с получением зрелого лиганда. Альтернативно, протеолиз между Арг153-Apr154 может ускорять выведение МЛч.
Вектор экспрессии, содержащий полную кодирующую последовательность МЛм, временно трансфецировали в клетки 293, как описано в Примере 1. Кондиционированная среда от этих клеток стимулировала включение 3H-тимидина в Ba/F3 клетки, экспрессирующие mpl и человека и мыши, но не оказывала действия на родительскую (лишенную mpl) линию клеток. Это свидетельствует, что клонированная кДНК МЛ мыши кодирует функциональный лиганд, который способен активировать оба МЛ рецептора (mpl), мыши и человека.
11. Лиганд mpl свиньи
кДНК МЛ свиньи (МЛс) выделили путем RACE РПЦ, как описано в Примере 13. кДНК из 1342 пар оснований, продукт РПЦ, была обнаружена в почках и субклонирована. Было секвенировано несколько клонов и обнаружено 332 аминокислотных остатка для кодирования лиганда mpl свиньи, обозначенных как МЛс (или МЛс332), имеющих нуклеотидную и выведенную аминокислотную последовательность, показанную на фиг. 20 (Посл. No: 18 и 19).
Была снова идентифицирована вторая форма, обозначенная МЛс2, кодирующая белок с 4 делециями аминокислотных остатков (228 аминокислотных остатков) (см. фиг.21 [Посл. No: 21]. Сравнение аминокислотных последовательностей МЛс и МЛс2 показало, что последняя форма является идентичной за исключением того, что тетрапептид QLPP, соответствующий остаткам 111-114 включительно, был удален (см. фиг.22 [Посл. No: 18 и 19]). Выпадение четырех аминокислот, обнаруженное в кДНК МЛ и у мыши, и у свиньи, наблюдается точно в том же положении в предсказанных белках.
Сравнение предсказанных аминокислотных последовательностей зрелых МЛ человека, мыши и свиньи (фиг.19 [Посл. No: 6, 17 и 18] отмечает, что в общем последовательности идентичны и составляют 72 процента между мышью и человеком, 68 процентов между мышью и свиньей и 73 процента между свиньей и человеком. Гомология существенно выше в амино-концевой половине МЛ (гомологичный домен ЭП). Этот домен от 80 до 84 процентов идентичен между любыми двумя видами, тогда как карбокси-концевая половина (углеводный домен) идентичен только на 57-67 процентов. Структура из двух основных аминокислот, которая могла бы представлять участок для расщепления протеазами, присутствует в карбоксильном конце гомологичного домена эритропоэтина. Эта структура является консервативной у трех видов в данном положении (фиг.19 [Посл. 6, 17 и 18]). Второй участок с двумя основаниями присутствует в положении 245 и 246 у человека и не присутствует в последовательности у мыши или свиньи. Последовательность МЛ мыши и свиньи содержит 4 цистеина, все консервативны в последовательности человека. Имеется семь участков возможного N-гликозилирования в лиганде мыши и шесть в МЛ свиньи, 5 из них консервативны с последовательностью человека. Все возможные участки N-гликозилирования расположены снова в С-концевой половине белка.
12. Экспрессия и очистка ТП из клеток яичника китайского хомячка (СНО)
Использованные векторы экспрессии для трансфекции СНО клеток обозначены: pSVIS.ID.LL.MLORF (полная длина или ТП332), и p3V15.ID.LL.MLEPO-D (усеченный или ТП153). Имеющие отношения к делу особенности этих плазмид представлены на фиг.23 и 24.
Процедуры трансфекции описаны в Примере 20. В кратком виде, кДНК, соответствующая полной открытой рамке считывания ТП, получена путем РПЦ. Продукт РПЦ очищали и клонировали между двумя сайтами рестрикции (Clal и Sall) плазмид p3V15.ID.LL. для получения вектора pSV15.ID.LL.MLORF. Вторую конструкцию, соответствующую гомологичному домену ЭП, получили тем же путем, но используя другой обратный праймер (EPOD.Sal). Конечную конструкцию для вектора, кодирующего гомологичный домен ЭП у ТП назвали pSV15.ID.LL.MLEPO-D.
Эти две конструкции линеаризовали с Notl и трансфецировали в клетки яичника китайского хомячка (клетки CHO-DP12, Европейский патент 307247, опубликованный 15 марта 1989 г.) путем электропорации. Были электропорированы 107 клеток при помощи BRL аппарата для электоропорации (350 вольт, 350 мФ, низкая емкость) в присутствии 10, 25 или 50 мг ДНК, как описано (Andreason, G.L. J. Tissue Cult. Meth. 15, 56, [1993]). На следующий день после трансфекции клетки расслоили в DHFR селективной среде (DMEM-F12 с высоким содержанием глюкозы 50:50 без глицина, 2 мМ глютамин, 2-5% диализованной телячьей сыворотки). Через 10-15 дней отдельные колонии перенесли в платы с 96 ячейками и оставили расти до конфлуентности. Эксперессию MA153 или МЛ332 определяли в кондиционированной среде из данных клонов, используя исследование пролиферации Ba/F3-mpl клеток (описано в Примере 1).
Способ очистки и выделения ТП из культуральной жидкости выращенных клеток СНО описан в Примере 20. В кратком виде, культуральную жидкость после выращивания клеток (КЖК) наносили на колонку с Blue Sepharose (Pharmacia) в соотношении приблизительно 100 л КЖК на литр гранул носителя. Затем колонку промывали 3-5 объемами колонки буфером, содержащим 2,0 М мочевину. Затем элюировали ТП 3-5 объемами колонки буфером, содержащим и 2,0 М мочевину, и 1 М NaCl.
Объем элюата с колонки с Blue Sepharose, содержащий ТП, наносили затем на колонку Wheat Germ Lectin Spharose (Сефарозу с лектином проростков пшеницы) (Pharmacia), уравновешенную элюирующим буфером Blue Sepharose, в соотношении от 8 до 16 объемов элюата на мл объема гранул Blue Sepharose. Затем колонку промыли 2-3 объемами буфера уравновешивания. Затем элюировали ТП 2-5 объемами буфера, содержащего 2,0 М мочевину и 0,5 М N-ацетил-D-глюкозамин.
Затем элюат с колонки с Weat Germ Lectin, содержащий ТП, подкислили и добавили С12Е8 до конечной концентрации 0,04%. Конечный объем нанесли на колонку С4 с обращенной фазой, уравновешенную 0,1% TFA, 0,04% С12Е8 при загрузке, приблизительно, от 0,2 до 0,5 мг белка на мл носителя.
Белок элюировали в две фазы линейным градиентом ацетонитрила, содержащего 0,1% TFA и 0,04% С12Е8, и создали объем основе данных SDS-PAGE.
Объем с С4 развели и диафильтровали против, приблизительно, 6 объемов буфера на Amicon YM или подобной ультрафильтрационной мембране, имеющей отсечение по молекулярному весу от 10000 до 30000 дальтон. Конечный диафильтрат затем может быть прямо обработан или далее сконцентрирован путем ультрафильтрации. Диафильтрат/концентрат обычно доводят до конечной концентрации с помощью 0,01% Твин-80.
Весь или часть диафильтрата/концентрата, эквивалентную 2-5% рассчитанного объема колонки, затем наносят на колонку с Сефакрил S-300 HR (Pharmacia), уравновешенную буфером, содержащим 0,01% Твин-80, и хроматографируют. Фракции, содержащие ТП, которые свободны от агрегатов и продуктов деградации, затем объединили на основе данных SDS-PAGE. Конечный объем профильтровали и хранили при 2-8°С.
13. Способы трансформирования и индукции синтеза ТП в микроорганизме и выделение, очистка и укладывание полученного здесь ТП
Конструирование вектора экспрессии ТП в Е.coli описано в подробностях в Примере 21. В кратком виде, плазмиды рМР21, рМР151, рМР41, рМР57 и рМР202 все созданы для экспрессии первых 155 аминокислот ТП вниз по течению маленьким лидером, который варьирует для различных конструкций. Лидеры получают главным образом для инициации трансляции высокого уровня и быстрой очистки. Плазмиды рМР210-1, -Т8, -21, -22, -24, -25 созданы для экспрессии первых 153 аминокислот ТП вниз по течению инициации метионина и различаются только по кодону, используемому для первых 6 аминокислот ТП, тогда как плазмида рМР251 является производной от рМР210-1, в которой карбоксильный конец ТП увеличен на две аминокислоты. Все вышеуказанные плазмиды будут производить внутриклеточную экспрессию высокого уровня ТП в Е.coli в условиях индукции триптофанового промотера (Yansura, D.G. et al.. Methods in Enzymology (Goeddel, D.V., Ed.) 185:54-60, Academic Press, San Diego [1990]). Плазмиды pMP1 и рМР172 являются промежуточными в конструкциях вышеуказанных плазмид внутриклеточной экспрессии ТП.
Вышеуказанные плазмиды экспрессии ТП были использованы для трансформации Е.coli, применяя метод теплового шока с CaCl2 (Mandel, M. et al., J. Mol. Biol., 53:159-162 [1970]) и другие способы, описанные в Примере 21. В кратком виде, трансформированные клетки сначала выращивали при 37°С до достижения оптической плотности культуры при (600 нм) приблизительно 2-3. Затем культуру разводили и после выращивания при аэрации добавляли кислоту. Культуру затем оставляли для продолжения роста при аэрации на следующие 15 часов, по истечение которых клетки собирали путем центрифугирования.
Способы выделения, очистки и укладывания, приведенные ниже, для получения биологически активного, сложившегося ТП человека или его фрагментов, описанные в Примерах 22 и 23, могут быть применены для получения любого варианта ТП, включая N- и С-концевые расширенные формы. Другие способы, подходящие для укладывания рекомбинантного или синтетического ТП, могут быть найдены в следующих патентах: Builder et al., Патент США 4511502; Jones et al.. Патент США 4512922; Olson, Патент США 4518526 и Builder et al., Патент США 4620948; для общего описания способов выхода и укладывания для различных рекомбинантных белков, экспрессированных в нерастворимом виде в Е.coli.
А. Получение нерастворимого ТП
Микроорганизм подобный Е.coli, экспрессирующий ТП, кодируемый любыми подходящими плазмидами, ферментировали в условиях, в которых ТП откладывается в нерастворимых "рефрактильных или преломляющих тельцах". Необязательно, клетки сначала промывали в буфере для разрушения клеток. Обычно, около 100 г клеток ресуспендировали в 10 объемах буфера разрушения клеток (а именно, 10 мМ Трис, 5 мМ ЭДТА, рН 8) при помощи, например, гомогенизатора типа Политрон, и клетки центрифугировали при 5000×g в течение 30 минут. Затем клетки лизировали, используя любые общепринятые способы, такие как тонический шок, озвучивание, циклическое продавливание, химические или ферментативные способы. Например, промытый вышеупомянутый осадок клеток может быть ресуспендирован в других 10 объемах буфера для разрушения клеток при помощи гомогенизатора, а затем клеточную суспензию пропускали через LH Cell Disrupter (LH Iceltech, Inc.) или через Microfluidizer (Microfluidics International) в соответствии с инструкциями. Частички вещества, содержащего ТП, затем отделяли от жидкой фазы и, необязательно, промывали любой подходящей жидкостью. Например, суспензия клеточного лизата может быть центрифугирована при 5000×g в течение 30 минут, ресуспендирована и, необязательно, центрифугирована во второй раз для получения осадка рефрактильных телец. Отмытый осадок может быть использован немедленно или, необязательно, сохранен при заморозке (при, например, -70°С).
В. Солюбилизация и очистка мономерного ТП
Нерастворимый ТП в осадке рефрактильных телец затем солюбилизировали в буфере солюбилизации. Буфер солюбилизации содержит хаотропный агент, и обычно создают буфер при основных значениях рН, и содержит восстанавливающий агент для увеличения выхода мономерного ТП. Примерные хаотропные агенты могут включать мочевину, гуанидин-НСl и тиоцианат натрия. Предпочтительным хаотропным агентом является гуанидин-НСl. Концентрация хаотропного агента обычно 4-9 М, предпочтительно, 6-8 М. рН солюбилизирующего буфера поддерживается любым подходящим буфером в ряду рН от около 7,5 до 9,5, предпочтительно 8,0-9,0, и наиболее предпочтительно, 8,0. Предпочтительно, солюбилизирующий буфер также содержит восстанавливающий агент для способствования образования мономерной формы ТП. Подходящий восстанавливающий агент включает органические вещества, содержащие свободный тиол (RSH). Примерные восстанавливающие агенты включают дитиотреитол (ДТТ), дитиоэритрит (ДТЭ), меркптоэтанол, глютатион (GSH), цистеамин и цистеин. Предпочтительным восстанавливающим агентом является дитиотреитол (ДТТ). Буфер солюбилизации может, необязательно, содержать мягкий окисляющий агент (например, молекулярный кислород) и подходящую соль для образования мономерного ТП через сульфитолизиз. В этом воплощении конечный ТП-3-сульфонат далее складывается в присутствии окислительно-восстановительного буфера (например, GSH/GSSH) для образования надлежащим образом уложенного ТП.
Белок ТП обычно далее очищали используя, например, центрифугирование, хроматографию при гель фильтрацию и хроматографию на колонке с обращенной фазой.
Согласно иллюстрации следующие процедуры дают удовлетворительный выход мономерного ТП. Осадок рефрактильных телец ресуспендировали в около 5 объемах по весу буфера солюбилизации (20 мМ Трис, рН 8, с 6-8 М гуанидина и 25 мМ ДТТ) и перемешивали в течение 1-3 часов или в течение ночи при 4°С для солюбилизации белка ТП. Высокие концентрации гуанидина также применимы, но в целом приводят к несколько пониженному выходу по сравнению с гуанидином. После солюбилизации раствор центрифугировали при 30000×g в течение 30 минут для получения прозрачного супернатанта, содержащего денатурированный мономерный белок ТП. Затем супернатант хроматографировали путем гель фильтрации на колонке с Супердекс 200 (Pharmacia, 2,6×60 см) при скорости тока 2 мл/мин, и элюировали белок 20 мМ Na фосфата, рН 6,0, в присутствие 10 мМ ДТТ. Фракции, содержащие мономерный денатурированный белок ТП, элюировавшиеся между 160 и 200 мл, объединяли. Далее белок ТП очищали на полупрепаративной колонке С4 с обращенной фазой (2×20 см VYDAC). Образцы наносили при скорости 5 мл/мин на колонку, уравновешенную 0,1% ТФК (трифторуксусной кислотой) с 30% ацетонитрилом. Белок элюировали линейным градиентом ацетонитрила (30-60% за 60 минут). Очищенный восстановленный белок элюируется при приблизительно 50% ацетонитрила. Данный материал использовали для укладывания с целью получения биологически активного варианта ТП.
С. Укладывание ТП для получения биологически активной формы
После солюбилизация и дальнейшей очистки ТП получали биологически активную форму путем укладывания денатурированного мономерного ТП в окислительно-восстановительном буфере. Благодаря высокой активности ТП (полумаксимальная стимуляция в исследовании на Ba/F3 клетках достигается при приблизительно 3 пг/мл) было возможно получить биологичкски активный материал, используя многие различные буферы, детергенты и окислительно-восстановительные условия. Однако в большинстве условий получали только небольшое количество надлежащим образом уложенного материала (<10%). Для коммерческого производственного способа желательно иметь выход уложенного материала, по крайней мере, 10%, более предпочтительно, 30-50%, и, наиболее предпочтительно, >50%. Было обнаружено, что многочисленные различные детергенты, включая Тритон Х-100, додецил-бета-мальтозид, CHAPS, CHAPSO, SDS, саркозил, Твин 20 и Твин 80, Цвиттергент 3-14 и другие для получения, по крайней мере, некоторого надлежащим образом уложенного материала. Однако среди них наиболее предпочтительными детергентами были те, из семейства CHAPS (CHAPS и CHAPSO), которые, как было обнаружено, наилучшим образом работали при реакции укладывания, и ограничивали агрегацию белка и неправильное образование дисульфидных связей. Наиболее предпочтительным был уровень CHAPS более чем около 1%. Хлорид натрия требовался для лучшего выхода, с оптимальным уровнем между 0,1 М и 0,5 М. Было предпочтительным присутствие ЭДТА (1-5 мМ) в окислительно-восстановительном буфере для ограничения уровня катализируемого металлом окисления (и агрегации), которое наблюдалось в некоторых приготовлениях. Концентрация глицерина более чем 15% создавала оптимальные условия для укладки. Для максимального выхода было обязательным иметь окислительно-восстановительную пару в окислительно-восстановительном буфере, содержащую оба, и окисленный, и восстановленный органический тиол (RSH). Подходящее окислительно-восстановительные пары включают меркаптоэтанол, глютатион (GSH), цистеамин, цистеин и их соответствующие окисленные формы. Предпочтительной окислительно-восстановительной парой были глютатион (GSH) : окисленный глютатион (GSSH) или цистеин : цистин. Наиболее предпочтительной окислительно-восстановительной парой был глютатион (GSH) : окисленный глютатион (GSSH). В целом, высокий выход наблюдали, когда молярное отношение окисленного члена окислительно-восстановительной пары было равно или превышало молярное отношение восстановленного члена окислительно-восстановительной пары. Для укладывания этих вариантов ТП было оптимальным значение рН между 7,5 и около 9. Органические растворители (например, этанол, ацетонитрил, метанол) в концентрациях 10-15% или ниже были индифферентными. Высокие уровни органических растворителей увеличивали количество неправильно уложенных форм. В общем, применимыми были Трис и фосфатный буферы. Инкубация при 4°С также давала повышенный уровень уложенного надлежащим образом ТП.
Выход при укладывании 40-60% (на основании количества восстановленного и денатурированного ТП, использованного в реакции укладки) является типичным для приготовления ТП, который был очищен путем использования первого С4 этапа. Активный материал может быть получен и из менее очищенных препаратов (например, сразу после колонки Супердекс 200, или после первичной экстракции рефрактильных телец), хотя выход является меньшим, из-за увеличенного осаждения и влияния не-ТП белков, в течение процесса укладки ТП.
Поскольку ТП содержит четыре остатка цистеина, возможно получить три различных дисульфидных версии этого белка:
версия 1: дисульфидная связь между остатками цистеина 1-4 и 2-3;
версия 2: дисульфидная связь между остатками цистеина 1-2 и 3-4;
версия 3: дисульфидная связь между остатками цистеина 1-3 и 2-4.
В течение первичного исследования определения условий для укладки методом хроматографии с обращенной фазой на С4 было разделено несколько различных пиков, содержащих белок ТП. Только один из этих пиков имел значительную биологическую активность, согласно определению при использовании Ba/F3 клеток. В соответствии с этим условия укладки были оптимизированы для предпочтительного выхода этой версии. В этих условиях неправильно уложившиеся версии составляли менее чем 10-20% от общего мономерного ТП, полученного на этапе солюбилизации.
Путем масс-спектрометрии и секвенирования белка была определена картина дисульфидных связей для биологически активного ТП, представляющая 1-4 и 2-3 связи, где цистеины пронумерованы по порядку с амино-конца. Картина перекрестных связей находится в соответствии с картиной известных дисульфидных связей родственной молекулы эритропоэтина.
D. Биологическая активность рекомбинантного уложенного ТП
Уложенный и очищенный ТП обладает активностью в исследованиях и in vitro, и in vivo. Например, в исследовании на клетках Ba/F3 полумаксимальную стимуляцию включения тимидина в Ba/F3 клетки для ТП (Мет-1 1-153) достигали при 3,3 пг/мл (0,3 пМ). В основанном на mpl рецепторе исследовании ELISA полумаксимальную активность наблюдали при 1,9 нг/мг (120 пМ). У обычных и миелосупрессированных животных, полученных при близком к летальному облучению рентгеновскими лучами, уложенный ТП (Мет-1 1-153) был высоко активен (активность наблюдали в дозах меньше, чем 30 нг/мышь) по стимуляции продукции новых тромбоцитов. Близкая биологическая активность была обнаружена и для других форм ТП, уложенных в соответствии с вышеуказанными способами (см. фиг.25, 26 и 28).
14. Способы измерения тромбопоэтической активности
Тромбопоэтическая активность может быть измерена в различных исследованиях, включая исследование лиганда mpl на клетках Ba/F3, описанного в Примере 1, исследование восстановительного синтеза тромбоцитов у мыши in vivo, исследование индукции клеточного поверхностного антигена тромбоцитов по измерению методом антитромбоцитарного иммуноопределения (анти-GРIIbIIIа) на мегакариобластической клеточной линии (СМК) лейкемии человека (см. Sato et al., Brit. J. Haematol., 72: 184-190 [1989]) (см. также исследование мегакариоцитопоэза в жидкой суспензии, описанное в Примере 4), и индукцию полиплоидизации на мегакариоцитобластической клеточной линии (DAMI) (см. Ogura et al.. Blood, 72(1): 49-60 [1988]). Созревание мегакариоцитов из незрелой формы, главным образом, не синтезирующих ДНК клеток, в морфологически идентифицируемые мегакариоциты вовлекает процесс, который включает появление цитоплазматических органелл, приобретение мембранных антигенов (GPIIbIIIa), внутреннюю репликацию и высвобождение тромбоцитов, как описано в предпосылках. Специфически дифференцирующий стимулятор линии (например, лиганд mpl) созревания мегакариоцитов, можно было бы ожидать, будет индуцировать, по крайней мере, некоторые из этих изменений у незрелых мегакариоцитотов, приводящих к высвобождению тромбоцитов и облегчению тромбоцитопении. Таким образом, исследование было построено так, чтобы измерять появление этих параметров у незрелых мегакариоцитарных клеточных линий, например, СМК и DAMI клеток. Исследование СМК определяет появление специфических маркеров тромбоцитов, GPIIbIIIa, и выход тромбоцитов. В определении DAMI (Пример 15) измеряется эндорепликация, поскольку увеличение плоидности является отличительным знаком зрелых мегакариоцитов. Распознаваемые мегакариоциты имеют значение плоидности 2N, 4N, 8N, 16N, 32N, и т.д. Наконец, исследование восстановительного синтеза тромбоцитов у мышей in vivo (Пример 16) полезно для демонстрации того, что введение исследуемого соединения (здесь лиганда mpl) приводит к увеличению количества тромбоцитов.
Были разработаны два дополнительных исследования in vitro для измерения активности ТП. Первое представляет исследование активации рецептора киназы (KIRA) методом ELISA, в котором клетки СНО трансфецированы химерой mpl-Rse, и измеряется фосфорилирование тирозина Rse методом ELISA после выдерживания части mpl химеры с лигандом mpl (см. Пример 17). Второе представляет метод ELISA, основанный на определении рецептора, в котором пластинки для ELISA, покрытые кроличьей сывороткой против человеческого IgG, захватывают химерные рецепторы человека mpl-IgG, которые связывают исследуемый лиганд mpl. Биотинилированное моноклональное антитело кролика к лиганду mpl (ТП155) использовали для определения связанного лиганда mpl, которое измеряли, используя стрептавидин-пероксидазу, как описано в Примере 18.
18. Биологический ответ нормальных и сублетально облученных мышей на введение ТП in vivo
Обеим, нормальным и облученным в сублетальной дозе мышам ввели усеченный и полной длины ТП, выделенный из клеток яичника китайского хомячка (СНО), Е.coli и эмбриональных клеток почки человека (293). Обе формы ТП, полученные от этих трех хозяинов, стимулировали продукцию тромбоцитов у мышей, однако ТП полной длины, выделенный из СНО, по-видимому, вызывает самый сильный ответ in vivo. Данные результаты отмечают, что надлежащее гликозилирование карбокси-концевого домена может быть необходимым для оптимальной активности in vivo.
(а) Е.соli-рТПч(Mет-1,153)
Форма "Мет" домена ЭП (Мет в -1 положении +153 первых остатков ТП человека), продуцированный в Е.coli (см. Пример 23), ежедневно инъецировали обычным самкам С57 В6 мышей, как описано в объяснениях к фиг.25А, 25В и 25С. Эти чертежи показывают, что продуцируемая в Е.coli, негликозилированная усеченная форма ТП, уложенная, как описано выше, способна стимулировать приблизительно двукратное увеличение продукции тромбоцитов у обычных мышей без воздействия на популяцию красных и белых клеток крови.
Эти самые молекулы, инъецированные ежедневно сублетально облученным (137Сs) самкам С57 В6 мышей, как описано в подписях к фиг. 26А, 26В и 26С, стимулировали выход тромбоцитов и уменьшали период крайнего упадка, но не влияли на эритроциты или лейкоциты.
(b) СНО-рТПч332
Форма ТП полной длины, продуцируемая в клетка СНО, вводимая ежедневно обычным самкам С57 В6 мышей, как описано выше в подписях к фиг.27А, 27В и 27С, вызывала приблизительно пятикратное увеличение продукции тромбоцитов у обычных мышей без воздействия на популяцию эритроцитов или лейкоцитов.
(c) СНО-рТПЧ332; Е. coli-рТПч (Мет-1, 153); 293-рТПч332 и
Е.coli-pTПч155
Была построена кривая дозового ответа на введение обычным мышам рТПч из различных клеточных линий (СНО-рТПч332; Е.coli-рТПч(Мет-1, 153); 293-рTПч332 и Е.соli-рТПч155), как описано в подписи к фиг. 28. Этот чертеж показывает, что все исследуемые формы молекул стимулируют продукцию тромбоцитов, однако форма полной длины, продуцируемая в клетках СНО, имеет наибольшую активность in vivo.
(d) CHO-pTПч153, СНО-рТПч "обрезанный" и СНО-рТПч332
Была также построена кривая дозового ответа на введение обычным мышам различных формы рТПч, продуцируемых в клетках СНО (СНО-рТПч153, СНО-рТПч "обрезанный" и СНО-рТПч332), как описано в подписи к фиг. 29. Этот чертеж показывает, что все тестируемые формы молекул из клеток СНО стимулируют продукцию тромбоцитов, но что форма полной длины 70 Kda имеет наибольшую активность in vivo.
16. Общий рекомбинантный препарат и варианты лиганда rnpl
Лиганд mpl, предпочтительно, получали согласно обычным рекомбинантным способам, которые включают продукцию полипептида лиганда mpl, путем культивирования клеток, трансфецированных для экспрессии нуклеиновой кислоты лиганда mpl (обычно путем трансфецирования клеток вектором экспрессии) и выделения полипептида из клеток. Однако необязательно является предсказуемым, что лиганд mpl может быть получен гомологичной рекомбинацией или по способам получения рекомбинанта, используя контрольные элементы, введенные в клетки, уже содержащие ДНК, кодирующую лиганд mpl. Например, сильный промотерный/усилительный элемент, ингибитор или экзогенный элемент, модулирующий транскрипцию, могут быть введены в геном предполагаемых клеток хозяина в достаточной близости и ориентации для влияния на транскрипцию ДНК, кодирующую желаемый полипептид лиганда mpl. Контрольный элемент не кодирует лиганд mpl, скорее ДНК является близкой по природе к геному клетки хозяина. Следующий элемент отсеивает клетки, создающие рецепторный полипептид по настоящему изобретению, или для увеличения, или для уменьшения уровня экспрессии по желанию.
Таким образом, изобретение рассматривает способ получения лиганда mpl, включающий введение модуляторного элемента транскрипции в геном клеток, содержащих в молекуле нуклеиновой кислоты лиганда mpl, в достаточной близости и ориентированным на молекулу нуклеиновой кислоты, для влияния на их транскрипцию, с дальнейшим необязательным этапом, включающим культивирование клеток, содержащих модуляторный элемент транскрипции и молекулу нуклеиновой кислоты. Изобретение также рассматривает клетку хозяина, содержащую естественную молекулу нуклеиновой кислоты лиганда mpl, операбельно сцепленную к экзогенной контрольной последовательности, распознаваемой клеткой хозяина.
А. Выделение ДНК, кодирующей полипептид лиганда mpl
ДНК, кодирующая полипептид лиганда mpl, может быть получена из любой библиотеки кДНК, полученной из ткани, подозреваемой на содержание мРНК лиганда mpl и экспрессирующей ее на детектируемом уровне. Ген лиганда mpl может также быть получен из библиотеки геномной ДНК или путем олигонуклеотидного синтеза in vitro из полной нуклеотидной или аминокислотной последовательности.
Библиотеки просеивали с зондами, разработанными для идентификации интересующего гена или белка, кодируемого им. Для библиотек экспрессируемых кДНК подходящие зонды включают моноклональные или поликлональные антитела, которые распознают и специфически связываются с лигандом mpl. Для библиотек кДНК подходящие зонды включают олигонуклеотиды из около 20-80 оснований длиной, которые кодируют известные или подозреваемые участки кДНК лиганда mpl из того же или другого вида; и/или комплементарные или гомологичные кДНК или их фрагменты, которые кодируют тот же или подобный ген. Подходящие зонды для скрининга библиотек кДНК включают, но не ограничиваются этим, олигонуклеотиды, кДНК, или их фрагменты, которые кодируют тот же или подобный ген, и/или гомологичные геномные ДНК или их фрагменты. Скрининг кДНК или геномной библиотеки с выбранным зондом может быть проведен, используя обычные процедуры, как описано в Частях 10-12, Sambrook et al., там же.
Альтернативным способом для выделения гена, кодирующего лиганд mpl, является использование техники РПЦ, как описано в разделе 14, Sambrook et al., там же. Этот способ требует использование олигонуклеотидного зонда, который гибридизуется с ДНК, кодирующей лиганд mpl. Стратегии для выбора нуклеотидов описаны ниже.
Предпочтительным практикуемым способом настоящего изобретения является использование тщательного отбора нуклеотидных последовательностей для просеивания библиотек кДНК из различных тканей, предпочтительно человека или почки свиньи (взрослой или эмбриона), или линий клеток печени. Например, библиотека кДНК клеточной линии эмбриональной печени человека была просеяна с олигонуклеотидными зондами. Альтернативно, могут быть просеяны с олигонуклеотидными зондами геномные библиотеки человека.
Олигонуклеотидные последовательности, выбранные в качестве зондов, должны быть достаточной длины и достаточно недвусмысленны, чтобы минимизировать фальшивые положительные ответы. Действенная нуклеотидная последовательность(и) обычно разрабатывается на основе областей лиганда mpl, которые имеют наименьший избыточный кодон. Олигонуклеотиды могут быть получены из одного или многих положений. Использование созданных нуклеотидов является особенно важным, когда просеивается библиотека вида, у которого предпочтительно используемый кодон неизвестен.
Олигонуклеотид должен быть помечен таким образом, чтобы его можно было обнаружить при гибридизации с ДНК просеиваемой библиотеки. Предпочтительным способом мечения является использование АТФ (а именно, γ32P) и полинуклеотидкиназы для включения радиоактивной метки в 5’ конец олигонуклеотида. Однако могут быть использованы другие способы мечения олигонуклеотидов, включая, но не ограничивая этим, биотинилирование или ферментативное мечение.
Особенный интерес представляет нуклеиновая кислота лиганда mpl, которая кодирует полный полипептид лиганда mpl. В некоторых предпочтительных воплощениях последовательность нуклеиновой кислоты включает естественную сигнальную последовательность лиганда mpl. Нуклеиновую кислоту, имеющую все последовательности кодонов белка, получили при скрининге выбранной кДНК или геномных библиотек, используя выведенную аминокислотную последовательность.
В. Аминокислотная последовательность вариантов естественного лиганда mpl
Аминокислотные последовательности вариантов лиганда mpl получили путем введения подходящих нуклеотидных изменений в ДНК лиганда mpl или путем синтеза in vitro желаемого полипептида лиганда mpl. Такие варианты включают, например, форму с делецией, или с вставкой, или заменой остатков внутри аминокислотной последовательности лиганда mpl свиньи. Например, части карбокси-конца зрелого лиганда mpl полной длины могут быть удалены при протеолитическом расщеплении как in vivo, так и in vitro или путем клонирования и экспрессии фрагмента или ДНК, кодирующего лиганд mpl полной длины для получения биологически активного варианта. Предусматривается применение любой комбинации из делеции, вставки и замены для достижения конечной конструкции, чтобы конечная конструкция проявляла желаемую биологическую активность. Изменения аминокислот также могут изменить посттрансляционные процессы лиганда mpl, такие как изменение порядка или положения гликозилируемых сайтов. Для разработки вариантов аминокислотной последовательности лиганда mpl положение сайта мутации и природа мутации будут зависеть от модификации характеристики (характеристик) лиганда mpl. Участки мутации могут быть модифицированы индивидуально или в сериях, например, путем: (1) сначала замена выбранными консервативными аминокислотами и затем более радикально выбранными в зависимости от достигаемого результата, (2) удаление целевого остатка или (3) вставка остатков того же класса или другого, приспособленного к участку расположения, или комбинации выборов 1-3.
Способ, полезный для идентификации определенных остатков или области полипептида лиганда mpl, который предпочтителен для локализации мутагенеза, назван "мутагенез сканирования аланина", как описано Cunningham and Wells, Science, 244: 1081-1085 [1989]. Здесь, остаток или группу целевых остатков определяли (а именно, заряженные остатки, такие как арг, асп, гис, лиз и глю) и заменяли на любую, но предпочтительно нейтральную или отрицательно заряженную аминокислоту (наиболее предпочтительно, аланин или полиаланин) для воздействия на взаимодействие аминокислот с окружающей водной средой внутри или снаружи клетки. Затем те домены, демонстрирующие функциональную чувствительность к заменам, усовершенствовали путем дальнейшего введения или других вариантов у доменов или участков для замены. Таким образом, несмотря на то, что участок для введения вариации аминокислотной последовательности предопределен, природа мутации per se (как таковой) не требует быть предопределенной. Например, для оптимизации выполнения мутации в данном участке проводили сканирование по ала(нину) или проводили случайный мутагенез по целевому кодону или области, а экспрессированные варианты лиганда mpl просеивали для оптимальной комбинации желаемой активности.
Существует две принципиальные возможности варьирования в конструкции аминокислотной последовательности вариантов: расположение участка мутации и природа мутации. Например, варианты полипептида лиганда mpl включают варианты форм последовательности лиганда mpl и могут представлять естественно встречающиеся аллели (которые не будут требовать манипуляций с ДНК лиганда mpl), или предопределенные мутантные формы, полученные путем мутирования ДНК, либо для достижения аллели, либо не обнаруживаемый в природе вариант. В целом, расположение и природа выбранной мутации будут зависеть от модифицируемой характеристики лиганда mpl.
Аминокислотная последовательность делеций в целом охватывает от около 1 до 30 остатков, более предпочтительно от около 1 до 10 остатков, и они обычно являются смежными. Альтернативно, аминокислотная последовательность делеций может также включать часть или полный карбокси-концевой гликозилированный домен. Аминокислотная последовательность делеции может также включать одну или более из первых 6 амино-концевых остатков зрелого белка. Аминокислотная последовательность делеции, необязательно, охватывает один или более остатков в одной из областей петли, которая расположена "узлами спирали". Смежные делеции обычно получаются по четным номерам остатков, но единичные или нечетные номера делеции находятся внутри этой рамками. Для модификации активности лиганда mpl могут быть введены делеции в области с низкой гомологией между лигандами mpl, которая приходится на большую часть идентичной последовательности. Или делеции могут быть введены в области с низкой гомологией между полипептидами лиганда mpl человека и лиганда mpl других млекопитающих, которые обладают наибольшей идентичностью последовательности к лиганду mpl человека. Делеции у полипептида лиганда mpl млекопитающего в области существенной гомологии с лигандом mpl другого млекопитающего будут более вероятными, чтобы более значимо модифицировать биологическую активность лиганда mpl. Количество порядковых делеции будет выбрано таким образом, чтобы сохранить четвертичную структуру лиганда mpl в изменяемом домене, например, бета-складчатую или альфа-спиральную.
Аминокислотная последовательность вставок включает амино- и/или карбокси-концевые слияния, различающиеся по длине от одного остатка до полипептида, содержащего сотню или более остатков, так же как и вставки внутри последовательности одной или многих аминокислотных остатков. Вставки внутри последовательности (то есть вставки внутри последовательности зрелого лиганда mpl) могут различаться в размере, в целом, от около 1 до 10 остатков, более предпочтительно, от 1 до 5, наиболее предпочтительно, от 1 до 3. Примерное предпочтительное слияние состоит из лиганда mpl или его фрагмента и другого цитокина или его фрагмента. Образцы конечных вставок включают зрелый лиганд mpl с N-концевым остатком метионина, артифактом прямой экспрессии зрелого лиганда mpl в рекомбинантной клеточной культуре, и слияния гетерологичной N-концевой сигнальной последовательности с N-концом молекулы зрелого лиганда mpl для усиления секреции зрелого лиганда mpl из рекомбинантного хозяина. Такие сигнальные последовательности обычно получают из и благодаря гомологии с предполагаемым видом клетки хозяина. Подходящие последовательности включают STII или Ipp для Е.coli, альфа фактор дрожжей и вирусные сигналы, такие как герпес gD, для клеток млекопитающих.
Другие варианты вставок молекулы лиганда mpl включают сплавление N- и С-концов лигада mpl иммуногенными полипептидами (а именно, не эндогенными по отношению к хозяину, которому эта сплавка вводится), например, бактериальный полипептид, такой как бета-лактамаза, или фермент, кодируемый trp локусом Е.coli, или белок дрожжей, и С-концевая сплавка с белком, имеющим длинную продолжительность полужизни, такого как постоянная область иммуноглобулина (или другая область иммуноглобулина), альбумин, или ферритин, как описано в WO 89/02922, опубликованном 6 апреля 1989 г.
Третья группа вариантов является вариантами аминокислотных замен. Эти варианты имеют, по крайней мере, один удаленный аминокислотный остаток в молекуле лиганда mpl, и другой остаток, вставленный на ее место. Участки наибольшего интереса для заместительного мутагенеза включают участки, идентифицированные как активные участки(ок) лиганда mрl, и участки, где аминокислоты, обнаруженные у других аналогов, являются существенно различными в терминах объема боковой цепи, заряда, или гидрофобности, но, где имеется также высокая степень идентичности в выбранном участке, среди лигандов mpl у различных видов, и/или внутри одного лиганда mpl различных членов близких животных.
Другим участком, представляющим интерес, является тот, в котором конкретные остатки лиганда mpl получены от различных членов семьи и/или видов животных, с одним идентичным членом. Эти участки, в особенности те, выпавшие из последовательности, по крайней мере, из трех других идентично консервативных участков, заменены сравнительно консервативным образом. Такие консервативные замены представлены в Таблице 3 под заголовком предпочтительных замен. Если такие замены приводят к изменениям в биологической активности, тогда вводили более существенные замены, обозначенные как примерные замены в Таблице 3, или как описано далее в ссылках на классы аминокислот, и полученные продукты просеяли.
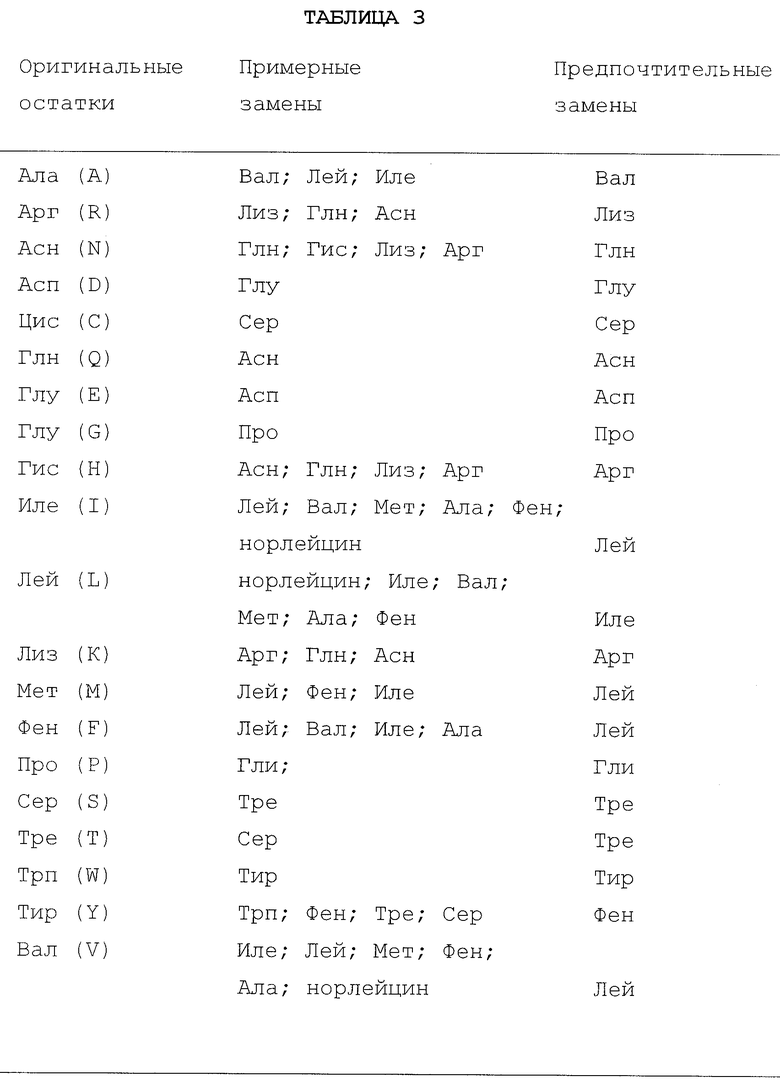
Существенные модификации в функции или иммунологической идентичности лиганда mpl сопровождались выбором замен, которые существенно различаются по своему действию на поддержание (а) структуры основы полипептида в области замен, например, в виде складок или спиральной конформации, (б) заряд или гидрофобность молекулы в участке мишени, или (в) объем боковой цепи. Естественно обнаруживаемые остатки разделены на группы, основанные на общих свойствах боковой цепи:
(1) гидрофобные: норлейцин, Мет, Ала, Вал, Дей, Иле;
(2) нейтральные гидрофобные: Цис, Сер, Тре;
(3) кислые: Асп, Глу;
(4) основные: Асн, Глн, Гис, Лиз, Арг;
(5) остатки, которые влияют на ориентацию цепи: Гли, Про;
(6) ароматические: Трп, Тир, Фен.
Неконсервативные замены повлекут за собой замены членов одного из этих классов на другой. Такие замененные остатки также могут быть введены в участки консервативных замен или, более предпочтительно, в оставшиеся (неконсервативные) участки.
В одном воплощении изобретения является желательным инактивировать один или более участков расщепления протеазой, который присутствует в молекуле. Эти участки идентифицировали по проверке кодирующей аминокислотной последовательности в случае трипсина, например, на остатки аргинина или лизина. Когда участки расщепления протеазой идентифицировали, их превратили в неактивные для протеолитического расщепления путем замены целевого остатка на другой остаток, предпочтительно основной остаток, такой как глутамин, или гидрофобный остаток, такой как серин; путем удаления остатка или путем вставки остатка пролина сразу после остатка.
В другом воплощении любые остатки метионина, другие чем начальный остаток метионина сигнальной последовательности, или любой остаток, расположенный внутри трех N-концевых или С-концевых остатков по отношению к каждому такому остатку метионина, заменен на другой остаток (предпочтительно в соответствии с Таблицей 3) или удален. Альтернативно, около 1-3 остатков ввели по соседству с таким участком.
Любые остатки цистеина, не участвующие в поддержании правильной конформации лиганда mpl, также могут быть заменены, обычно на серин, для увеличения стабильности молекулы к окислению и предотвращения аберрантного перекрестного связывания. Было обнаружено, что первый и четвертый цистеины в ЭП домене, пронумерованные с амино-конца, необходимы для поддержания надлежащей конформации, но что второй и третий - нет. Соответственно второй и третий цистеины в ЭП домене могут быть заменены.
Молекулы нуклеиновой кислоты, кодирующие аминокислотную последовательность варианта лиганда mpl, получили различными способами, известными в методологии. Эти способы включают, но не ограничиваются выделением из природного источника (в случае естественно встречающейся аминокислотной последовательности вариантов) или получением путем опосредованного олигонуклеотидом (или сайт-направленного) мутагенеза, мутагенеза РПЦ, и кассетного мутагенеза ранее полученного варианта или инвариантной версии полипептида лиганда mpl.
Опосредованный олигонуклеотидом мутагенез является предпочтительным способом для получения вариантов ДНК лиганда mpl с заменой, делецией и вставкой. Эти способы хорошо известны в технике, как описано Adelman et al., DNA, 2: 183 [1983]. В кратком виде, ДНК лиганда mpl изменили путем гибридизации олигонуклеотида, кодирующего желаемую мутацию с матрицей ДНК, где матрица представляет однонитевую форму плазмиды или бактериофага, содержащего неизмененую или нативную последовательность ДНК лиганда mpl. После гибридизации использовали ДНК полимеразу для синтеза полной второй комплементарной нити к матрице, которая будет, таким образом, включать олигонуклеотидный праймер и будет кодировать выбранное изменение в ДНК лиганда mpl.
В целом, использовали олигонуклеотиды, по крайней мере, 25 нуклеотидов длиной. Оптимальный нуклеотид будет иметь от 12 до 15 нуклеотидов, которые полностью комплементарны матрице по обеим сторонам нуклеотида(ов), кодирующих мутацию. Это дает уверенность, что нуклеотиды гибридизуются надлежащим образом к однонитевой молекуле ДНК матрицы. Нуклеотиды легко синтезировали, используя способы, известные в технике, такие как описаны Crea et al., Proc. Natl. Acad. Sci. USA, 75: 5765 [1978].
Матрицы ДНК могут быть получены при помощи тех векторов, которые либо происходят от вектора бактериофага М13 (применимы коммерчески доступные векторы М13тр18 и М13тр19), либо те векторы, которые содержат однонитевой фаг репликации, как описано Viera et al., Meth. Enzymol., 153: 3 [1987]. Таким образом, та ДНК, которая будет мутировать, может быть вставлена в один из этих векторов для создания однонитевой матрицы. Получение однонитевой матрицы описано в Секциях 4.21-4.41 Saiabrook et al., Molecular Cloning: A Laboratory Mannual (Cold Spring Harbor Laboratory Press, NY 1989).
Альтернативно, матрица однонитевой ДНК может быть получена путем денатурирования двухнитевой ДНК плазмиды (или другой), используя обычную технику.
Для изменения естественной последовательности ДНК (для получения аминокислотной последовательности варианта, например) нуклеотид гибридизовали с однонитевой матрицей при подходящих условиях гибридизации. Затем добавляли фермент, полимеризующий ДНК, обычно Klenow-фрагмент ДНК полимеразы I для синтеза комплементарной нити матрицы, используя нуклеотиды в качестве праймеров для синтеза. Таким образом, формировали гетеродуплексную молекулу, такую, что одна нить ДНК кодирует мутантную форму лиганда mpl, а другая нить (оригинальная матрица) кодирует естественную, неизмененную последовательность лиганда mpl. Затем гетеродуплексную молекулу трансформировали в подходящую клетку хозяина, обычно прокариота, такого как Е.coli JM101. После выращивания клеток их перенесли на агарозные плашки и просеяли, используя нуклеотидный праймер, радиоактивно меченный 32-фосфатом для идентификации бактериальных колоний, которые содержат мутантную ДНК. Мутированную область затем удаляли и помещали в подходящий вектор для получения белка, главным образом вектор экспрессии типа, обычно используемого для трансформации подходящего хозяина.
Способ, непосредственно описанный выше, может быть модифицирован так, чтобы гетеродуплексную молекулу создавали в обеих нитях плазмиды, содержащей мутацию(и). Модификации следующие: однонитевой олигонуклеотид ренатурировали к однонитевой матрице, как описано выше. Объединяли смесь трех дезоксирибонуклеотидов, дезоксирибоаденозина (дАТФ), дезоксирибогуанозина (дГТФ) и дезоксириботимидина (дТТФ) с модифицированным тио-дезоксирибоцитозином, так называемом дСТФ-(аS) (который может быть поставлен Amersham Corporation). Эту смесь добавляли к матрично-олигонуклеотидному комплексу. При добавлении ДНК полимеразы к данной смеси получали нить ДНК, идентичную матрице, за исключением мутированных оснований. Помимо этого эта новая нить ДНК будет содержать дСТФ-(аS) вместо дСТФ, которые служат для их защиты от расщепления рестрикционной эндонуклеазой.
После разрывания одной нити матрицы двухнитевого гетеродуплекса при помощи подходящего рестрикционного фермента нить матрицы может быть расщеплена ExoIII нуклеазой или другой подходящей нуклеазой после области, которая содержит участок(и), предназначенные для мутагенеза. Затем реакцию останавливали, оставляя молекулу, которая является только частично однонитевой. Затем образовывали полный двухнитевой гетеродуплекс ДНК, используя ДНК полимеразу в присутствии всех четырех дезоксирибонуклеотидтрифосфатов, АТФ и ДНКлигазу. Данная гомодуплексная молекула затем может быть трансформирована в подходящую клетку хозяина, такого как Е.coli JM101, как описано выше.
ДНК, кодирующая мутанты лиганда mpl с более чем одной замененной аминокислотой, может быть получена одним из нескольких путей. Если аминокислоты расположены близко вместе в полипептидной цепи, они могут быть мутированы одновременно, используя один олигонуклеотид, который кодирует все желаемые замены аминокислот. Если, однако, аминокислоты расположены на некотором удалении друг от друга (разделены более чем около десяти аминокислот), значительно более трудно получить единый олигонуклеотид, который кодирует все желаемые изменения. Вместо этого могут быть использованы один или два альтернативных способа.
В первом способе для каждой заменяемой аминокислоты получали отдельный олигонуклеотид. Затем олигонуклеотиды одновременно ренатурировали к однонитевой матрице ДНК, а вторая нить ДНК, которую синтезировали по матрице, будет кодировать желаемые замены аминокислот.
Альтернативный способ включает два или более циклов мутагенеза для получения желаемого мутанта. Первый цикл таков, как описано для одного мутанта: для матрицы использовали дикий тип ДНК, олигонуклеотид, кодирующий первую желаемую замену(ы) аминокислоты ренатурировали к данной матрице и затем получали гетеродуплексную молекулу ДНК. Во втором цикле мутагенеза использовали мутантную ДНК, полученную в первом цикле мутагенеза по матрице. Таким образом, матрица содержит одну или более мутаций. Затем к данной матрице ренатурировали олигонуклеотид, кодирующий желаемую аминокислотную замену(ы), и полученная теперь нить ДНК кодирует мутации из обоих, первого и второго циклов мутагенеза. Настоящая конечная ДНК может быть использована в качестве матрицы в третьем цикле мутагенеза, и т.д.
Мутагенез РПЦ также подходит для получения аминокислотных вариантов полипептида лиганда mpl. Поскольку следующее обсуждение касается ДНК, понятно, что эта техника также применима и к РНК. Техника РПЦ в целом относится к следующим процедурам (см. Eriich, там же, раздел R. Higuchi, стр.61-70). Когда использовали небольшие количества матрицы в качестве начального материала в РПЦ, для этого можно было использовать праймеры из соответствующих областей матрицы ДНК, которые незначительно различались в последовательности, для получения сравнительно больших количеств специфических фрагментов ДНК, которые различались от последовательности матрицы только в положениях, где праймеры различались с матрицей. Для введения мутации в ДНК плазмиды один из праймеров сконструировали для перекрывания положения мутации и содержания мутации; последовательность другого праймера должна быть идентичной фрагменту секвенирования противоположной нити плазмиды; но эта последовательность может быть расположена где-нибудь далеко в ДНК плазмиды. Предпочтительно, однако, чтобы последовательность второго праймера располагалась в 200 нуклеотидах от первого, так чтобы в конце полной амплифицируемой области ДНК, привязанной к праймеру, могла быть легко секвенирована. Амплификация РПЦ, использующая подобные пары праймеров, только описывает результаты в популяции фрагментов ДНК, которые различаются по положению мутации, определенной праймером, и возможно, по другим положениям, поскольку копирование матрицы до некоторой степени склонно к ошибке.
Если отношение матрицы к материалу продукта крайне низкое, имеется огромное большинство фрагментов продуктов ДНК для включения желаемой мутации(и). Данный материал продукта использовали для замены соответствующей области в плазмиде, которая служит в качестве матрицы РПЦ, при использовании обычной технологии ДНК. Мутации по раздельным положениям могут быть введены одновременно, путем использования второго мутантного праймера, или проведения второй РПЦ с различными мутантными праймерами и лигированием двух конечных фрагментов РПЦ одновременно к фрагменту вектора при трех- (или более) частей лигации.
В специфическом примере мутагенеза РПЦ матричную плазмиду ДНК (1 мкг) линеаризовали путем расщепления с рестрикционной эндонуклеазой, которая имеет уникальный участок распознавания ДНК плазмиды вне амплифицируемой области. Данный материал 100 нг добавили к РПЦ смеси, содержащей буфер РПЦ, который содержит четыре дезоксинуклеотидтрифосфата и который включен в наборы GeneAmp® (полученные от Perkin-Elmer Cetus, Norwalk, CT and Emeryville, CA), и 25 пмоль каждого олигонуклеотидного праймера до конечного объема 50 мкл. Реакционную смесь покрыли 35 мкл минерального масла. Реакционную смесь денатурировали в течение 5 минут при 100°С, быстро поместили на лед и затем добавили под пленку минерального масла 1 мкл ДНК полимеразы Thermus aquaticus (Taq) (5 единиц/мкл, полученную от Perkin-Elmer Cetus). Затем реакционную смесь поместили в ДНК Thermal Cycler (полученный от Perkin-Elmer Cetus), запрограммированный следующим образом:
2 мин 55°С,
30 сек 72°С, затем 19 циклов:
30 сек 94°С,
30 сек 55°С,
30 сек 72°С.
В конце программы реакционный флакон удалили из температурного циклизатора, а водную фазу перенесли в новый флакон, экстрагировали фенол/хлороформом (50:50 по объему) и осадили этанолом, ДНК выделили согласно обычным способам. Данный материал сразу же подвергли подходящей обработке для включения в вектор.
Другой способ получения вариантов - кассетный мутагенез, основан на технике, описанной Wells et al., Gene, 34: 313 [1985]. Начальным материалом является плазмида (или другой вектор), включающий мутируемую ДНК лиганда mpl. Идентифицируют предназначенный для мутации кодон ДНК лиганда mpl. Здесь должно находиться специальное место для рестрикционной эндонуклеазы, на каждой стороне участка(ков), определенного для мутации. Если таких участков для рестрикции не существует, они должны быть получены, используя вышеописанный опосредованный олигонуклеотидами способ мутагенеза, для введения их в подходящее месторасположение в ДНК лиганда mpl. После этого участки рестрикции были введены в плазмиду, плазмиду отрезали по этим участкам для их линеаризации. Используя обычные способы, синтезировали двухнитевой олигонуклеотид, кодирующий последовательность ДНК между участками рестрикции, но содержащий желаемую мутацию(и). Две нити синтезировали раздельно, а затем гибридизовали вместе, используя обычные способы. Данный двухнитевой олигонуклеотид называется кассетой. Эта кассета сконструирована так, чтобы иметь 3’ и 5’ концы, которые совместимы с концами линеаризованной плазмиды таким образом, чтобы непосредственно лигировать плазмиду. Данная плазмида теперь содержит мутированную последовательность ДНК лиганда mpl.
С. Введение нуклеиновой кислоты в реплицирующийся вектор
Нуклеиновую кислоту (например, кДНК или геномная ДНК), кодирующую естественный полипептид или вариант полипептида лиганда mpl, ввели в реплицирующийся вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии. Многие векторы являются доступными, поэтому выбор подходящего вектора будет зависеть от: (1) будет ли он применяться для амплификации ДНК или для экспрессии ДНК, (2) размера нуклеиновой кислоты для введения в вектор, (3) трансформируемой вектором клетки хозяина. Каждый вектор содержит различные компоненты в зависимости от его функции (амплификация ДНК или экспрессия ДНК) и клетки хозяина, с которой он должен быть совместим. Компоненты вектора обычно включают, но не ограничиваются этим, одно или более следующее: сигнальную последовательность, образец репликации, один или более маркеров генов, усилительный элемент, промотер и конечную транскрипционную последовательность.
(i) Компонент сигнальной последовательности Лиганд mpl по настоящему изобретению может быть экспрессирован не только непосредственно, но также в сплавленном виде с гетерологическим полипептидом, предпочтительно сигнальной последовательностью, или другим полипептидом, имеющим специфический участок для расщепления в N-конце зрелого белка или полипептида. В целом, сигнальная последовательность может быть компонентом вектора, или она может быть частью ДНК лиганда mpl, которая введена в вектор. Выбранная гетерологическая сигнальная последовательность должна быть такой, которая распознается и подвергается процессингу (то есть расщепляется сигнальной пептидазой) клеткой хозяина. Для клеток хозяина прокариота, которые не распознают, не подвергают процессингу сигнальную последовательность естественного лиганда mpl, сигнальную последовательность заменяли на выбранную сигнальную последовательность прокариотов, например, из группы щелочной фосфатазы, пенициллиназы, Ipp, или лидеры устойчивого к нагреванию энтеротоксина II. Для секреции дрожжей естественная сигнальная последовательность может быть заменена на, например, инвертазу дрожжей, альфа фактор или лидеры кислой фосфатазы, лидер глюкоамилазы С.albicans (EP 362179, опубликованный 4 апреля 1990 г.), или сигнал, описанный в WO 90/13646, опубликованном 15 ноября 1990 г. У клеток млекопитающих экспрессия естественной сигнальной последовательности (то есть предпоследовательности лиганда mpl, которая обычно ведет к секреции лиганда mpl из его естественных клеток млекопитающего in vivo), является удовлетворительной, хотя могут быть применимы другие сигнальные последовательности млекопитающих, такие как сигнальные последовательности из других полипептидов лиганда mpl, или из той же формы лиганда mpl других видов животных, сигнальных последовательностей из лиганда mpl, сигнальных последовательностей из секретируемых полипептидов того же или близких видов, так же как и секреторные лидеры вируса, например, gD сигнал герпеса симплекс.
(ii) Точка начала компонента репликации
Оба, экспрессируемый и клонируемый, векторы содержат последовательность нуклеиновой кислоты, которая дает возможность вектору реплицироваться в одних или более выбранных клетках хозяина. В целом, в клонируемых векторах данная последовательность является единственной, которая дает возможность вектору реплицироваться независимо от хромосомной ДНК хозяина и включает точку начала репликации или автономно реплицирующуюся последовательность. Такие последовательности хорошо известны для различных бактерий, дрожжей и вирусов. Точка начала репликации из плазмиды pBR322 применима для большинства Грам-отрицательных бактерий, точка начала плазмиды 2 μ применима для дрожжей, а различные вирусные точки начала (SV40, полиома, аденовирус, VSV или BPV) применимы для клонирующих векторов в клетках млекопитающих. В целом, точка начала компонента репликации не требуется для экспрессии векторов у млекопитающих (обычно, может быть использована точка начала SV40 только потому, что она содержит ранний промотер).
Большинство векторов экспрессии являются "челночными" векторами, то есть они способны к репликации в, по крайней мере, одном классе организмов, но могут быть трансфецированы для экспрессии в другой организм. Например, вектор клонировали в Е.coli, и затем тот же вектор трансфецировали в дрожжи или клетки млекопитающего для экспрессии, даже не обращая внимания на то, способен ли он к независимой репликации в хромосомах клетки хозяина.
ДНК также может быть амплифицирована путем введения в геном хозяина. Это легко выполняется, используя виды Bacillus в качестве хозяина, например, путем включения в вектор последовательности ДНК, которая комплементарна к последовательности, обнаруженной в геномной ДНК Bacillus. Трансфецирование Bacillus вектором приводит к гомологической рекомбинации с геномом и встраиванию в ДНК лиганда mpl. Однако выделение геномной ДНК, кодирующей лиганд mpl, является более сложным, чем экзогенно реплицируемого вектора, поскольку требуется расщепление ферментом рестрикции для вырезания ДНК лиганда mpl.
(iii) Отбор компонентов гена
Экспрессируемые и клонируемые векторы должны содержать селекционный ген, также называемый селектируемый маркер. Этот ген кодирует белок, необходимый для выживания или роста трансформированной клетки хозяина в селективной культуральной среде. Клетки хозяина, нетрансформированные вектором, содержащим селекционный ген, не выживут в культуральной среде. Обычно селекционные гены кодируют белки, которые (а) придают устойчивость к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, (б) дополняют ауксотрофную недостаточность, или (в) обеспечивают критическими питательными веществами, не доступными из сложной среды, например ген, кодирующий рацемазу D-аланина для Bacilli.
Один пример селекционой схемы, использующей лекарственные препараты для задержки роста клеток хозяина. Те клетки, которые успешно трансформировали гетерологическим геном, экспрессируют белок, придающий устойчивость к лекарственному препарату и, таким образом, выживают в селекционном режиме. В примерах такой доминантной селекции используют препараты неомицин (Southern et al., J, Molec. Appi. Genet., 1: 327 [1982]), микофеноловую кислоту (Mulligan et al., Science, 209: 1422 [1980]) или гигромицин (Sugden et al., Mol. Cell. Biol., 5: 410-413 [1985]). В трех примерах, приведенных выше, участвуют бактериальные гены в условиях контроля эукариотами передачи устойчивости к подходящему препарату G418 или неомицину (генетицин), xgpt (микофеноловая кислота) или гигромицину соответственно.
Примерами других подходящих селектируемых маркеров для клеток млекопитающих являются те, которые дают возможность идентификации клеточных компонентов для извлечения нуклеиновой кислоты лиганда mpl, такие как дигидрофолат редуктаза (ДГФР) или тимидин киназа. Трансформированные клетки млекопитающих помещали в условия давления отбора, в которых выживают только особенно адаптированные трансформированные клетки благодаря захвату маркера. Давление отбора на трансформированные клетки, примененное путем культивирования в условиях, в которых концентрация селекционного агента в среде последовательно изменяется, приводит посредством этого к амплификации обоих генов, селектируемого гена и ДНК, которая кодирует полипептид лиганда mpl. Амплификация - это процесс, посредством которого гены, имеющие большую потребность для продукции белков, критических для роста, многократно повторяются в тандеме в хромосомах последовательно размножающихся рекомбинантных клеток. Таким образом, синтезировали увеличенное количество лиганда mpl из амплифицированной ДНК.
Например, клетки, трансформированные селекционным геном ДГФР, сначала идентифицировали при культивировании всех трансформантов в культуральной среде, которая содержит метотрексат (Mtx), конкурентный антагонист ДГФР. При применении дикого типа ДГФР получали и размножали подходящее количество клеток хозяина из яичника китайского хомячка (СНО), дефицитных по активности ДГФР, как описано Uriaub and Chasin, Proc. Natl. Acad. Sci. USA, 77: 4216 [1980]. Затем трансформированные клетки выдерживали с увеличивающимся уровнем Mtx. Это приводило к синтезу множества копий гена ДГФР и соответственно множества копий другой ДНК, содержащей экспрессируемые векторы, такие как ДНК, кодирующая лиганд mpl. Техника амплификации может быть использована с любым более или менее подходящим хозяином, например, АТСС No. CCL61 СНО-К1, несмотря на наличие эндогенной ДГФР, если, например, применен мутантный ген ДГФР, который высоко устойчив к Mtx (ЕП 117060). Альтернативно, клетки хозяина (в частности, хозяина дикого типа, который содержит эндогенную ДГФР) трансформировали или трансформировали совместно с последовательностью ДНК, кодирующей лиганд mpl, диким типом белка ДГФР и другим селектируемым маркером, таким как аминогликозид 3’ фосфотрансфераза (АФТ), могут быть отобраны при выращивании клеток в среде, содержащей селекционный агент для селектируемого маркера, такого как аминогликозидный антибиотик, например канамицин, неомицин, или G418. См. Патент США No.4965199.
Походящий селекционный ген для использования на дрожжах представляет ген trp1, присутствующий в плазмиде Yrp7 (Stinchomb et al.. Nature, 282: 39 [1979]; Kingsman et al., Gene, 7: 141 [1979]; или Tschemper efc al., Gene, 10: 157 [1980]). Ген trp1 производит селектируемый маркер для мутантного штамма дрожжей, лишенных способности расти в отсутствии триптофана, например, АТСС No. 44076 или РЕР4-1 (Jones, Genetics, 85: 12 [1977]). Наличие повреждения trp1 в геноме дрожжевых клеток хозяина создает эффективную окружающую среду для обнаружения трансформации при выращивании в отсутствии триптофана. Подобным образом, Лей2-дефицитный штамм дрожжей (ФТСС No.20622 или 38626) дополнен известными плазмидами, несущими ген Лей2.
(iv) Промотерный компонент
Экспрессия и клонирование векторов, обычно содержащих промотеры, которые распознаваемы организмом хозяина и операбельно сцеплены с нуклеиновой кислотой лиганда mpl. Промотеры представляют нетранслируемую последовательность, располагающуюся в обратном направлении (5’) к начальному кодону структурного гена (в общем, от 100 до 1000 пар оснований), который контролирует транскрипцию и трансляцию конкретной последовательности нуклеиновой кислоты, такой как последовательность нуклеиновой кислоты лиганда mpl, к которой они операбельно сцеплены. Такие промотеры обычно разделяют на два класса, индуцируемые и конституитивные. Индуцируемые промотеры являются промотерами, которые инициируют повышенные уровни транскрипции с ДНК под их контролем в ответ на некоторые изменения в условиях культивирования, например наличие или отсутствие питательного вещества, или изменение в температуре. В настоящее время известно большое количество промотеров, распознаваемых разнообразными потенциальными клетками хозяевами. Эти промотеры опреративно сцепляют к ДНК, кодирующей лиганд mpl, путем удаления промотера из источника ДНК при расщеплении рестрикционным ферментом и включения выделенной последовательности промотера в вектор. И естественные промотерные последовательности лиганда mpl и многие гетерологичные промотеры могут быть использованы для непосредственной амплификации и/или экспрессии ДНК лиганда mpl. Однако предпочтительны гетерологические промотеры, поскольку они в целом позволяют большую транскрипцию и более высокий выход экспрессии лиганда mpl по сравнению с естественным промотером лиганда mpl.
Промотеры, подходящие для использования с хозяевами прокариотами, включают β-лактамазную или лактозную системы промотеров (Chang et al.. Nature, 275: 615 [1978]; и Goeddel et al.. Nature, 281: 544 [1979]), системы промотеров щелочной фосфатазы или триптофана (trp) (Goeddel, Nucleic Acida Res., 8: 4057 [1980] и ЕП 36 776) и гибридные промотеры, такие как tac промотер (deBoer et al., Proc. Natl. Acad. Sci. USA, 80: 21-25 [1983]). Однако применимы другие известные бактериальные промотеры. Их нуклеотидные последовательности опубликованы, таким образом, позволяя опытному работнику операбельно пришить их к ДНК, кодирующей лиганд mpl (Siebenlist et al. Cell, 20: 269 [1980]), используя линкеры или адаптеры для получения любого требуемого участка рестрикции. Промотеры для использования в бактериальных системах также будут содержать Shine-Dalgarno (S.D.) последовательность, операбельно пришитую к ДНК, кодирующую полипептид лиганда mpl.
Известны промотерные последовательности для эукариот. Фактически все гены эукариот имеют AT-обогащенную область, расположенную приблизительно от 25 до 30 оснований в противоположном направлении от участка, где инициируется транскрипция. Другая последовательность найдена от 70 до 80 основания в противоположном направлении от начала транскрипции многих генов представляет СХСААТ область, где Х может быть любым нуклеотидом. На 3’ конце у многих генов эукариот присутствует последовательность ААТААА, которая может являться сигнальной, в добавление к поли А концу 3’ конца кодирующей последовательности. Все эти последовательности применимы для введения в векторы экспрессии эукариот.
Примеры подходящих последовательностей векторов для использования с клетками хозяевами дрожжей включают промотеры для 3-фосфоглицерокиназы (Hitzeman et al., J. Biol. Chem., 255: 2073 [1980]), или другого гликолитического фермента (Hess et al., J. Adv. Enzyme Res., 7: 149 [1980]; и Holland, Biochemistry, 17: 4900 [1978]), такие как энолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицеромутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа.
Другими промотерами дрожжей, которые представляют индуцируемые промотеры, имеющие дополнительное преимущество в транскрипции, контролируемой условиями выращивания, являются промотерные области для алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, расщепляющими ферментами, связанными с метаболизмом азота, металлотионеин, глицеральдегид-3-фосфатдегидрогеназа и ферменты, ответственные за утилизацию мальтозы и галактозы. Подходящими векторами и промотерами для использования в экспрессии у дрожжей являются далее описанные Hitzeman et al., ЕП 73657А. Усилители дрожжей также имеют преимущество при использовании с промотерами дрожжей.
Транскрипция лиганда mpl из векторов в клетках хозяина млекопитающего контролируется, например, при помощи промотеров, полученных из геномов вирусов, таких как вирус полиомы, вирус оспы домашних птиц (Патент Великобритании 2211504, опубликованный 5 июля 1989 г.), аденовирус (такой как аденовирус 2), вирус бычьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и, наиболее предпочтительно, вакуолизирующий обезьяний вирус 40 (SV 40), из гетерологических промотеров млекопитающих, например промотера актина или промотера иммуноглобулина, промотеров тепловой устойчивости и из промотера, обычно ассоциированного с последовательностью лиганда mpl, при условии, что такие промотеры совместимы с системами клеток хозяина.
Ранние и поздние промотеры вируса SV40 получены общепринятым способом в виде рестрикционных фрагментов SV 40, которые также содержат вирусную точку начала репликации SV 40. Fiers et al. Nature, 273: 113 [1978]; Mulligan and Berg, Science, 209: 1422-1427 [1980]; Pevlakis et al., Proc. Natl. Acad. Sci. USA, 78: 7398-7402 [1981]. Немедленный ранний промотер цитомегаловируса человека получен общепринятым способом в виде HindIII E фрагмента рестрикции. Greenaway et al. Gene, 18: 355-360 [1982]. Система экспрессии ДНК млекопитающего хозяина, использующая вирус бычьей папилломы в качестве вектора описана в Патенте США No.4419446. Модификация этой системы описана в Патенте США No.4601978, см. также Gray et al., Nature, 295: 503-508 [1982] об экспрессии кДНК, кодирующей иммунный интерферон в клетках обезьян; Reyes et al., Nature, 297: 598-601 [1982] об экспрессии кДНК (3-интерферона человека в клетках мыши под контролем тимидинкиназного промотера из вируса герпеса симплекс; Canaani and Berg, Proc. Natl. Acad. USA, 79: 5166-5170 [1982] об экспрессии гена β1-интерферона человека в культуре клеток мыши и кролика; и German et al., Proc. Natl. Acad. Sci. USA, 79: 6777-6871 [1982] об экспрессии бактериальной CAT последовательности в CV-1 клетках почки обезьяны, фибробластов эмбриона цыпленка, клетках яичника китайского хомячка, клетках HeLa и клетках NIH-3T3 мыши, используя повтор длинного конца вируса саркомы Рауса в виде промотера.
(v) Компонент усилительного элемента
Транскрипцию ДНК, кодирующую лиганд mpl по настоящему изобретению, высшими эукариотами часто усиливают путем введения усилительной последовательности в вектор. Усилители представляют цис-действующие элементы ДНК, обычно около от 10 до 300 пар оснований, которые действуют на промотер для увеличения его транскрипции. Усилители являются относительно ориентированными и независимыми в положении, обнаруживая 5’ (Laimins et al., Proc. Natl. Acad. Sci. USA, 78: 993 [1981]) и 3’ (Lusky et al., Mol. Cell Bio., 3: 1108 [1983]) локализацию в транскрипционной единице, внутри интрона (Banerji et al., Cell, 33: 729 [1983]), так же как и в самой кодирующей последовательности (Osborne et al., Mol. Cell Bio., 4: 1293 [1984]). Многие усилительные последовательности из генов млекопитающих теперь известны (глобин, эластаза, альбумин, α-фетопротеин и инсулин). Обычно, однако, используют усилители из вирусов клеток эукариот. Примеры включают усилитель SV 40 из поздней области точки начала репликации (пары оснований 100-270), усилитель раннего промотера цитомегаловируса, усилитель полиомы из поздней области точки начала репликации и усилители аденовируса. См. также Yaniv, Nature, 297: 17-18 [1982] об усилительных элементах для активации промотеров эукариот. Усилитель может быть врезан в вектор по 5’ или 3’ положению к кодирующей последовательности mpl, но предпочтительно расположен в 5’ участке от промотера.
(iv) Транскрипция компонента терминации
Экспрессируемые векторы, используемые в клетках хозяевах эукариот (дрожжах, грибах, насекомых, растениях, животных, человеке или ядерных клетках из других многоклеточных организмов) будут также содержать последовательности, необходимые для остановки транскрипции и для стабилизации мРНК. Такие последовательности из 5’, иногда 3’ нетранслируемых областей эукариот или вирусных ДНК или РНК широко доступны. Эти области содержат сегменты нуклеотидов, транскрибируемых в виде фрагментов полиаденилирования в нетранслируемой части мРНК, кодирующей лиганд mpl.
(vii) Конструкция и анализ векторов
Конструкция подходящих векторов, содержащих один или более вышеперечисленных компонентов, включает обычную технику лигации. Выделенные плазмиды или фрагменты ДНК расщепили, получили хвосты и соединили в формы, желаемые для получения требуемых плазмид.
Для анализа и подтверждения правильности последовательностей в сконструированных плазмидах, использовали лигационную смесь для трансформации Е.coli K12 штамма 294 (АТСС No.31446), а удачные трансформанты, где необходимо, отбирали на устойчивость к ампициллину или к тетрациклину. Плазмиды, полученные из трансформантов, анализировали путем расщепления рестрикционной эндонуклеазой, и/или секвенировали по методу Messing et al., Nucleic Acids Res., 9: 309 [1981], или по методу Maxam et al., Methods in Enzymology, 65: 499 [1980].
(viii) Переходные векторы экспрессии
Особенно полезными в практике настоящего изобретения являются векторы, которые предусмотрены для переходной экспрессии ДНК в клетках млекопитающих, кодирующей полипептид лиганда mpl. В целом, переходная экспрессия включает в использование экспрессионных векторов, которые способны эффективно реплицироваться в клетках хозяина, так чтобы клетки хозяина накопили множество копий экспрессированного вектора и, в свою очередь, синтезировали высокий уровень желаемого полипептида, кодируемого экспрессированным вектором. Sambrook et al., там же, стр.16.17-16.22. Переходные системы экспрессии, содержащие подходящий вектор экспрессии и клетку хозяина, допускают удобную положительную идентификацию полипептидов, кодируемых клонируемой ДНК, так же как и быстрый скрининг подобных полипептидов с желаемыми биологическими или физиологическими свойствами. Таким образом, системы переходной экспрессии являются особенно полезными в изобретении для целей идентификации аналогов и вариантов полипептида лиганда mpl, которые имеют биологическую активность полипептида лиганда mpl.
(ix) Примеры удобных векторов клеток позвоночных
Другие способы, векторы и клетки хозяина, применимые для синтеза лиганда mpl в рекомбинантных культурах клеток, позвоночных описаны в Gething et al., Nature, 293: 620-625 [1981]; Mantel et al., Nature, 281: 40-46 [1979]; Levinson et al., ЕП 117060; ЕП 117058. Особенно полезная плазмида для экспрессии лиганда mpl в культуре клеток млекопитающих является pRK5 (ЕП 307247; Патент США No.5258287) или pSVI6B (Публикация РСТ No. WO 91/08291).
D. Отбор и трансформация клеток хозяина
Подходящими клетками хозяина для клонирования или экспрессии векторов в настоящей работе являются прокариоты, дрожжи или высшие эукариоты, описанные выше. Подходящие прокариоты включают эубактерии, такие как Грам-отрицательные или Грам-положительные, например, Е.coli, Bacilli, такие как В.subtilis, виды Pseudomonas, такие как Р. aeruginosa, Salmonella typhimurium, или Serrata marcescans. Одним предпочтительным клонирующим хозяином Е.coli является Е.coli 294 (АТСС No.31446), хотя применимы также другие штаммы, такие как Е.coli В, Е.coli X1776 (АТСС No.31537) и Е.coli W3110 (АТСС No.27325). Эти примеры скорее иллюстративны, чем ограничивающие. Предпочтительная клетка хозяин должна секретировать минимальное количество протеолитических ферментов. Альтернативно, применимы способы клонирования in vitro, например РПЦ или реакции других полимераз нуклеиновой кислоты.
Помимо прокариот подходящими хозяевами для кодирующих лиганд mpl векторов являются прокариотические микробы, такие как нитевидные грибы или дрожжи. Наиболее широко используемыми среди низших эукариотических хозяев микроорганизмами являются Saccharomyces cerevisiae, или обычные пекарские дрожжи. Однако большое количество других родов, видов и штаммов широко доступно и применимо здесь, таких как Schizosaccharomyces pombe (Beach and Nurse, Nature, 290: 140 [1981]; ЕП 139383, опубликованный 2 мая 1985 г.), в виде хозяев Kluyveromyces (Патент США No.4943529), такие как, например, К. lactis (Louvencourt et al., J. Bacteriol., 737 [1983]), К. fragilis, К. bulgaricus, К. termotolerans, и К. marxianus, yarrowia [ЕП 402226], Pichia pastoris (ЕП 183070; Sreekrishna et al., J. Basic Micribiol., 28: 265-278 [1988], Candida, Nrichoderma reesia (ЕП 244234), и нитевидные грибы, такие как, например, Neurospora, Penicillium, Tolypocladium (WO 91/00357, опубликованный 10 января 1991 г.), и в виде хозяев, такие как A. nidulans (Ballance et al., Biochem. Biophys. Res. Commun., 112: 284-289 [1983]; Tiburn et al., Gene, 26: 205-221 [1983]; Yelton et al., Proc. Natl. Acad. Sci. USA, 81: 1470-1474 [1984]) и A. niger (Kelly and Hynes, EMBO J., 4: 475-479 [1985]).
Подходящие клетки-хозяева для экспрессии гликозилированного лиганда mpl происходят от многоклеточных организмов. Такие клетки-хозяева способны к сложному процессингу и гликозилирующей активности. В принципе, любые культуры клеток высших эукариотов работоспособны, будь то культура позвоночных или беспозвоночных. Примеры клеток беспозвоночных включают растительные и клетки насекомых. Были определены многочисленные штаммы и варианты бакуловирусов и соответствующие клетки - хозяева пермессивных (допускающих) насекомых от таких хозяев, как Spodoptera frugiperda (гусеница), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (фруктовая мушка) и Bombix mori. См., например, Lukow et al., Bio/Technology, 6: 47-55 [1988]; Miller et al.. Genetic Engineering, Setlow et al., eds., Vol.8 (Plenum Publishing, 1986) pp.277-279; Maeda et al., Nature, 315: 592-594 [1985]. Свободно доступны разнообразные штаммы вирусов для трансфекции, например, L-1 вариант Autographa californica NPV и штамм Bm-5 Bombix mori NPV, подобные вирусы могут быть использованы здесь в качестве вирусов в соответствии с настоящим изобретением, конкретно для трансфекции клеток Spodoptera frugiperda.
Могут быть применены в качестве хозяев культуры растительных клеток хлопка, кукурузы, картофеля, соевых бобов, петунии, помидоров и табака. Обычно растительные клетки трансфецируют при инкубации с определенным штаммом бактерии Agrobacterium tumefaciens, который ранее обработали для содержания ДНК лиганда mpl. В течение инкубации культуры растительной клетки с A. tumefaciens, ДНК, кодирующая лиганд mpl, переносится в растительную хозяйскую клетку таким образом, что она становится трансфецированной и будет при определенных условиях экспрессировать ДНК лиганда mpl. Помимо этого доступны регуляторные и сигнальные последовательности, совместимые с растительными клетками, такие как промотер нопалин синтетазы и сигнальная последовательность полиаденилирования. Depicker et al., J. Mol. Appl. Gen., 1: 561 [1982]. Кроме этого, сегменты ДНК, выделенные из области противоположного направления Т-ДНК гена 780, способна к активации или увеличению уровня транскрипции экспрессируемых в растениях генов в рекомбинантной ДНК-содержащей растительной ткани, ЕП 321196, опубликованный 21 июня 1989 г.
Однако наибольший интерес к клеткам позвоночных, в последние годы размножение клеток позвоночных в культуре (тканевая культура) стало рутинным способом (Tissue Culture, Academic Press. Kruse and Patterson, editors [1973]). Примером полезной линии клетки хозяина млекопитающего являются линия CV 1 почки обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); линия эмбриональной почки человека (293 или клетки 293, субклонироанные для роста в суспензионой культуре, Gracham et al., J. Gen Virol., 36:59 [1977]); клетки почки детеныша хомяка (ВНК, ATCC CCL 10); клетки яичника китайского хомячка/-DHFR (СНО, Urlaub and Chasin, Proc. Natl. Acad. Sci. USA, 77:4216 [1980]); клетки сертоли мыши (ТМ4, Mather, Biol. Reprod., 23:243-251 [1980]); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки зеленой африканской макаки (VERO-76, ATCC CRL-1587); клетки цервикальной карциномы человека (HELA, ATCC CCL 2); клетки почки собаки (MDCR, ATCC CCL 34); клетки печени крыс buffalo (BRL ЗА, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (Нер G2, НВ 8065); опухоли молочных желез мыши (ММТ 060562, ATCC CCL51); клетки TRI (Mather et al.. Annals N.Y. Acad. Sci., 383: 44-68 [1982]); клетки MRC 5; клетки FS4; и линия гепатомы человека (Нер G2).
Клетки хозяина трансфецировали и преимущественно трансформировали вышеописанными векторами экспрессии или клонирования по настоящему изобретению и культивировали в общепринятой питательной среде, модифицированной соответственно для индуцирования промотерами, отбора трансформантов, или амплифицирования генов, кодирующих желаемую последовательность.
Трансфекция обозначает захват вектора экспрессии клеткой хозяином независимо от того, кодирует он или нет в действительности любую экспрессируемую последовательность. Обычному квалифицированному работнику известны многочисленные способы трансфекции, например способ с СаРO4 или электропорация. Успешная трансфекция распознается, когда наблюдается любой показатель операции того, что данный вектор находится внутри клетки.
Трансформация означает введение ДНК в организм таким образом, что ДНК способна к репликации либо как внехромосомный элемент, либо встроенная в хромосому. В зависимости от используемых клеток хозяина проведенная трансформация использует обычные способы, подходящие для таких клеток. Обработка кальцием включает использование хлорида кальция, как описано в разделе 1.82 Sambrook et al., там же, главным образом, используя прокариотов, или другие клетки, которые содержат существенный барьер из клеточной стенки. Инфекцию Agrobacterium tumefaciens использовали для трансформации определенных клеток растений, как описано Shaw et al. Gene, 23: 315 [1983] и WO 89/05859, опубликованном 29 июня 1989 г. Помимо этого растения можно трансфецировать, используя ультразвуковую обработку, как описано в WO 91/00358, опубликованном 10 января 1991 г. Для клеток млекопитающих, не имеющих такой клеточной стенки, предпочтителен способ осаждения фосфатом кальция по Graham anad van der Eb, Virology, 52: 456-457 [1978]. Общие аспекты систем трансформации клеток хозяев млекопитающих описаны Axel в Патенте США No.4399216, изданном 16 августа 1983 г. Трансформации дрожжей обычно проводятся в соответствии со способом Van Solingen et al., J. Bact., 130: 946 [1977] и Hsiao et al., Proc. Natl. Acad. Sci. USA, 76: 3829 [1979]. Однако могут быть использованы другие способы для введения ДНК в клетки, такие как инъекция ядер, электропорация или сплавление протопластов.
Е. Культивирование клеток хозяев
Прокариотические клетки, использованные для получения полипептида лиганда mpl по настоящему изобретению, культивировали в подходящей среде, как описано в общем у Sambrook et al., там же.
Клетки хозяева млекопитающих, использованные для получения лиганда mpl по настоящему изобретению, можно культивировать в различных средах. Коммерчески доступные среды, такие как Ham’s F10 (Sigma), минимальная достаточная среда ([MEM], Sigma), RPMI-1640 (Sigma) и модифицированная Дульбекко среда Игла ([DMEM], Sigma) применимы для культивирования клеток хозяина. Кроме этого, могут быть использованы в качестве культуральной среды для клеток хозяина любые из сред, описанных Ham and Walles, Meth. Enz., 58: 44 [1979], Barnes and Sato, Anal. Biochem., 102: 255 [1980], патенты США No.4767704; 4657866; 4927762; или 4560655; WO 90/03430; WO 87/00195; патент США Re. 30985; или совместно ожидаемые U.S.S.N. 07/592 107 или 07/592 141, оба заполненные 3 октября 1990 г., все описания которых включены здесь в виде ссылок. Любые из этих сред могут быть дополнены, если необходимо, гормонами и/или другими факторами роста (такими как инсулин, трансферрин или фактор роста эпидермиса), солями (такими как хлорид натрия, кальция, магния и фосфаты), буферами (такими как HEPES), нуклеозидами (такими как аденозин или тимидин), антибиотиками (такими как препарат Gentamycin™), следовыми элементами (определенными как неорганические соединения, обычно присутствующие в конечных концентрациях в микрограммовой области) и глюкоза или эквивалентный источник энергии. Любые другие необходимые добавки также могут быть включены в подходящих концентрациях, которые должны быть известны опытным работникам. Культуральные условия, такие как температура, рН и подобные, и те, ранее примененные для отбора клеток хозяина для экспрессии, будут очевидны обычному квалифицированному работнику.
Клетки хозяина, отнесенные к настоящему изобретению, включают в себя клетки в культуре in vitro, так же как и клетки животного хозяина.
F. Определение амплифицированного/экспрессированного гена
Амплификация и/или экспрессия гена может быть измерена подсчитыванием транскрипции мРНК непосредственно в образце, например, общепринятым Southern блоттингом, northern блоттингом (Thomas, Proc. Natl. Acad. Sci. USA, 77:5201-5205 [1980]), дот блоттингом (анализ ДНК), или гибридизацией in situ, используя походящие меченые зонды, основанные на описанных здесь последовательностях. Могут быть использованы различные метки, наиболее распространенные радиоизотопы, а именно 32Р. Однако может быть применена другая техника, такая как использование модифицированных биотином нуклеотидов для введения в полинуклеотид. Биотин далее служит в качестве участка связывания авидина или антитела, которое может быть мечено широким разнообразием меток, таких как радионуклиды, флуоресцентные метки, ферменты или подобные им. Альтернативно, могут быть применены антитела, которые распознают специфические дуплексы, включая дуплексы ДНК, дуплексы РНК и гибридные дуплексы ДНК-РНК, или дуплексы ДНК-белок. Эти антитела, в свою очередь, могут быть мечены, и определение может быть проведено там, где дуплекс связался с поверхностью так, что при образовании дуплекса на поверхности, можно определить наличие антитела, связанного в дуплексе.
Альтернативно, для непосредственного подсчета экспрессии продукта гена экспрессия гена может быть измерена при помощи иммунологических способов, таких как иммуногистохимическое окрашивание кусочков ткани и исследование клеточной культуры или жидкостей тела. При технике иммуногистохимического окрашивания образцы клеток приготавливают, обычно применяя дегидратацию и фиксацию, с последующим реагированием с меченым антителом, специфическим для привязанного продукта гена, где метки обычно визуально обнаруживаемы, такие как ферментативные метки, флуоресцентные метки, люминисцентные метки, и тому подобные. Особенно чувствительная техника окрашивания, пригодная для использования в настоящем изобретении, описана Hsu et al. Am. J. Clin. Path., 75: 734-738 [1980].
Антитела, применимые для иммуногистохимического окрашивания и/или исследования образцов жидкостей, могут быть как моноклональными, так и поликлональными, и могут быть получены у любого млекопитающего. Удобно, когда антитела можно получить против естественного полипептида лиганда mpl или против синтетического пептида на основе последовательности ДНК, полученной здесь, как описано далее ниже.
G. Очистка полипептида лиганда mpl
Лиганд mpl предпочтительно выделяют из культуральной среды как секреторный полипептид, хотя он также может быть выделен из лизата хозяйских клеток, если непосредственная экспрессия не сопровождается секреторным сигналом.
Когда лиганд mpl экспрессировали в другой рекомбинантной клетке, отличающейся по происхождению от клетки человека, лиганд mpl полностью свободен от белков или полипептидов человеческой природы. Однако по-прежнему необходимо очищать лиганд mpl от других рекомбинантных белков или полипептидов клетки для получения препарата, который существенно гомогенный, как сам по себе лиганд mpl. На первом этапе культуральную среду или лизат центрифугировали для удаления частичек остатков клетки. Затем разделяли мембранную фракцию и растворимые белки. Альтернативно, может быть применен коммерчески доступный фильтр для концентрирования белков (например, фильтровальная ячейка Amicon или Millipore Pellicone). Лиганд mpl затем может быть очищен из фракции растворимых белков и из мембранной фракции лизата культуры в зависимости, является ли лиганд mpl мембранно-связанным. Согласно этому лиганд mpl очищали от сопутствующих растворимых белков и полипептидов путем высаливания и обмена или хроматографическими способами, используя различные гелевые матриксы. Эти матриксы включают: акриламид, агарозу, декстран, целлюлозу и другие, принятые для очистки белков. Примерные хроматографические процедуры, применимые для очистки белков, включают: иммуноаффиные (например, анти-hmpl лиганд Mab), рецептероаффинные (например, mpl-IgG или протеин А Сефароза), хроматография по гидрофобному взаимодействию (ГХ), (например, эфир, бутил, или фенил Toyopearl), хроматография на лектине (например, Кон А-Сефароза, лентил-лектин-Сефароза), разделение по размеру (например, Сефадекс Г-75), катион- и анионообменная колонка (например, ДЕАЕ или карбоксиметил- и сульфопропил-целлюлоза), и жидкостная хроматография с высоким разрешением с обращенной фазой (ОФ-ЖХВР) (см., например, Urdal et al., J. Chromatog., 296:171 [1984], где использовали два последовательных этапа ОФ-ЖХВР для очистки рекомбинантного ИЛ-2 человека). Другие этапы очистки, необязательно, включают: осаждение этанолом, осаждение сульфатом аммония, хроматофокусирование, препаративный SDS-PAGE и подобные.
Варианты лиганда mpl, у которых остатки удалены, вставлены или заменены, получали тем же самым образом, как и естественный лиганд mpl, принимая во внимание любые существенные изменения в свойствах, случающиеся у вариантов. Например, приготовление лиганда mpl, сплавленного с другим белком или полипептидом, например бактериальным или вирусным антигеном, облегчает очистку; может быть использована иммуноаффинная колонка, содержащая антитело к антигену для адсорбции сплавленного полипептида. Иммуноаффинная колонка, такая как колонка с поликлональным анти-mpl лигандом кролика, может быть применена для адсорбции варианта лиганда mpl, путем связывания его к, по крайней мере, одному иммунному эпитопу. Альтернативно, лиганд mpl может быть очищен путем аффинной хроматографии, используя очищенный mpl-IgG, привязанный к (предпочтительно) иммобилизованной смоле, такой как Аффи-гель 10 (Bio-Rad, Richmond, CA) или подобной, способом, хорошо известным в технике. Для ингибирования протеолитического расщепления в процессе очистки может быть использован ингибитор протеаз, такой как фенилметилсульфонилфторид (PMSF), а антибиотики могут быть включены для предотвращения роста случайных загрязнений. Опытный в технике работник поймет, что способы очистки, применимые для естественного лиганда mpl или его вариантов могут потребовать модификации для оценки характера изменений лиганда mpl или его вариантов при экспрессии в рекомбинантной культуре клеток.
Н. Ковалентные модификации полипептида лиганда mpl
Ковалентные модификации полипептида лиганда mpl включены в рамки настоящего изобретения. И естественный лиганд mpl, и последовательности аминокислотных вариантов лиганда mpl можно ковалентно модифицировать. Один тип ковалентной модификации, включенный в рамки настоящего изобретения, представляет собой фрагмент лиганда mpl. Фрагменты варианта лиганда mpl, имеющие до около 40 аминокислотных остатков, могут быть ковалентно получены путем химического синтеза или путем ферментативного или химического расщепления полипептида лиганда mpl полной длины. Другой тип ковалентных модификаций лиганда mpl или его фрагментов представляет введение в молекулу путем реагирования с целевыми аминокислотными остатками лиганда mpl или его фрагментами, органического агента, образующего производное, которое способно реагировать с выбранной боковой цепью или с N- или С-концевыми остатками.
Остатки цистеина чаще всего реагируют с α-галоацетатами (и соответствущими аминами), такими как хлоруксусная кислота или хлорацетамид, с получением карбоксиметильных или карбоксиамидометильных производных. Остатки цистеина также модифицируют в реакции с бромтрифторацетоном, α-бромо-β-(5-имидазоил)пропионовой кислотой, хлорацетилфосфатом, N-алкилмалеимидами, 3-нитро-2-пиридилдисульфидом, метил-2-пиридилдисульфидом, р-хлормеркурибензоатом, 2-хлормеркури-4-нитрофенолом или хлоро-7-нитробензо-2-окса-1,3-диазолом.
Остатки гистидина модифицируют в реакции с диэтилпирокарбонатом при рН 5,5-7,0, поскольку этот агент относительно специфичен для боковой цепи гистидина. Применимым является также парабромфенацилбромид; реакцию предпочтительно проводят в 0,1 М какодилате натрия при рН 6,0.
Лизинил и аминоконцевые остатки реагируют с янтарным или другими ангидридами карбоновой кислоты. Получение производных с этими агентами оказывает действие по изменению заряда остатков лизина. Другие подходящие агенты для получения производных аминосодержащих остатков включают имидоэфиры, такие как метилпиколинимидат; пиридоксальфосфат; пиридоксаль; хлороборогидрат; тринитробензенсульфоновая кислота; O-метилизомочевина; 2,4-пентандион и катализируемые трансаминазой реакции с глиоксилатом.
Остатки аргинина модифицируют в реакции с одним или несколькими общепринятыми реагентами, среди которых фенилглиоксаль, 2,3-бутандион, 1,2-циклогександион и нингидрин. Получение производных остатков аргинина требует, чтобы реакцию проводили в щелочных условиях из-за высокого значения pKa гуанидиновой функциональной группы. Более того, эти реагенты могут реагировать с группами лизина, так же, как и с ипсилон-аминогруппами аргинина.
Можно получить особенно интересные специфические модификации остатков тирозина с введенными спектральными метками в остатки тирозина в реакции с ароматическими диазониевыми соединениями или тетранитрометаном. Наиболее широко для образования видов O-ацетилтирозина и 3-нитро производных используются N-ацетилимидазол и тетранитрометан соответственно. Для получения меченых белков при использовании в радиоиммуноопределении иодируют остатки тирозина, используя 125I или 131I, для чего подходит вышеописаный способ с хлорамином Т.
Карбоксильные боковые группы (аспартил или глютамил) селективно модифицируют в реакции с карбодиимидом (R-N=C=N-R’), где R и R’ представляют различные алкильные группы, такие как 1-циклогексил-3-(морфолинил-4-этил)карбодиимид или 1-этил-3-(4-азония-4,4-диметилпентил)карбодиимид. Более того, остатки аспартила и глютамила превращают в аспарагинильные или глютаминильные остатки в реакции с ионом аммония.
Производные с бифункциональными агентами применимы для перекрестного сшивания лиганда mpl с нерастворимым в воде матриксом подложки или поверхности для использования в способах очистки антител анти-mpl лиганда и наоборот. Обычно используемые перекрестносшивающие агенты включают, например, 1,1-бис(диазоацетил)-2-фенилэтан, глютаровый альдегид, эфиры N-гидроксисукцинимида, например эфиры с 4-азидосалициловой кислотой, гомобифункциональные имидоэфиры, включая эфиры дисукцинимидила, такие как 3,3’-дитиобис(сукцинимидилпропионат) и бифункциональные малеимиды, такие как бис-N-малеимидо-1,8-октан. Агенты для получения производных, такие как метил-3-[(р-азидофенил)дитио]пропиоимидат дают фотоактивируемые интермедиаты, которые способны к образованию перекрестных связей в присутствии света. Альтернативно, активированные нерастворимые в воде матриксы, такие как активированные цианогенбромидом углеводы и активированные субстраты, описанные в Патентах США No.3969287; 3691016; 4195128; 4247642; 4229537 и 4330440, применяются для иммобилизации белков.
Глютаминильные и аспарагинильные остатки часто дезамидируют до соответствующего глютамильного и аспартильного остатков соответственно. Эти остатки дезаминируются в нейтральных или щелочных условиях. Дезаминированные формы этих остатков попадают в рамки настоящего изобретения.
Другие модификации, включая гидроксилирование пролина и лизина, фосфорилирование гидроксильных групп остатков серина или треонина, метилирование α-аминогрупп боковых цепей лизина, аргинина и гистидина (Т.Е.Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, pp.79-86 [1983]), ацетилирование N-концевого амина и амидирование любых концевых карбоксильных групп.
Другой тип ковалентной модификации полипептида лиганда mpl, включенного в рамки настоящего изобретения, состоит в изменении естественной картины гликозилирования полипептида. Под изменением имеется в виду удаление одного или более углеводных радикалов, обнаруженных у естественного лиганда mpl, и/или добавление одного или более участков гликозилирования, которые не присутствуют в естественном лиганде mpl.
Гликозилирование пептидов обычно является N- или С-связанным. N-связанное относится к присоединению углеводного радикала к боковой цепи остатка аргинина. Последовательности трипептидов аспарагин-Х-серин и аспарагин-Х-треонин, где Х представляет любую аминокислоту кроме пролина, являются участками узнавания для ферментативного присоединения углеводного радикала к боковой цепи аспарагина. Таким образом, присутствие и того и другого из этих трипептидных последовательностей в полипептиде создает потенциальные участки гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров N-ацетилгалактозамина, галактозы или ксилозы к гидроксиаминокислоте, наиболее обычно к серину или треонину, хотя может использоваться 5-гидроксипролин или 5-гидроксилизин.
Добавление участков гликозилирования к полипептиду лиганда mpl удобно достигается путем изменения аминокислотной последовательности так, что она содержит один или более остатков серина в вышеописанной трипептидной последовательности (для N-связанных участков гликозилирования). Изменения можно также сделать путем добавления или замещения на, одного или более остатков серина или треонина к естественной последовательности лиганда mpl (для O-связанных участков гликозилирования). Для облегчения последовательность аминокислот лиганда mpl предпочтительно изменяют путем изменения на уровне ДНК, в частности, путем мутации ДНК, кодирующей полипептид лиганда mpl по предварительно выбранным основаниям, так, чтобы те, полученные кодоны, будут транслировать желаемые аминокислоты. Мутацию(и) ДНК можно получить, используя способы, описанные выше под заголовком "Аминокислотная последовательность вариантов лиганда mpl."
Другие способы увеличения количества углеводных радикалов в лиганде mpl представляют собой химическое или ферментативное пришивание гликозидов к полипептиду. Эти способы имеют преимущество в том отношении, что они не требуют получения полипептида в клетке хозяине, который имеет возможности для N- и O-связанного гликозилирования. В зависимости от использованного способа пришивания сахар(а) могут быть присоединены к (а) аргинину и гистидину, (б) свободной карбоксильной группе, (в) свободным сульфгидрильным группам, таким как у цистеина, (г) свободным гидроксильным группам, таким как у серина, треонина или гидроксипролина, (д) ароматическому остатку, такому как у фенилаланина, тирозина или триптофана, или (е) аминогруппе глютамина. Эти способы описаны в WO 87/05330, опубликованному 11 сентября 1987 г., и у Alpin and Wriston, CRC Crit. Rev. Biochem., pp.259-306 [1981].
Удаление углеводного радикала, присутствующего у полипептида лиганда mpl, может быть выполнено химически или ферментативно. Химическое дегликозилирование требует выдерживания полипептида с соединением трифторметансульфоновой кислоты или равноценного соединения. Эта обработка приводит к расщеплению большинства или всех сахаров за исключением сочлененного сахара (N-ацетилглюкозамина или O-ацетилгалактозамина), оставляя полипептид неизмененым. Химическое дегликозилирование описано Hakimuddin et al., Arch. Biochem. Biophys., 259: 52 [1987] и Edge et al., Anal. Biochem., 118: 131 [1981]. Ферментативного расщепления углеводных остатков полипептида можно достигнуть при использовании различных эндо- и экзогликозидаз, как описано Nhotakura et al., Meth. Enzymol., 138: 350 [1987].
Гликозилирование по потенциальным участкам гликозилирования можно предотвратить при использовании соединения туникамицин, как описано Duskin et al., J. Biol. Chem., 257: 3105 [1982]. Туникамицмн блокирует образование белок-N-гликозидной связи.
Другой тип ковалентной модификации лиганда mpl включает пришивание полипептида лиганда mpl к одному из разнообразных небелковых полимеров, например полиэтиленгликолю, полипропиленгликолю или полиоксаалкиленам, по способам, описанным в Патентах США No.4640835; 4496689; 4301144; 4670417; 4791192 или 4179337. Полипептиды лиганда mpl, ковалентно пришитые к вышеупомянутым полимерам, обозначены здесь как пегилированные (от PEG=ПЕГ - полиэтиленгликоль, пришитые к ПЕГ, прим. переводчика) полипептиды лиганда mpl.
Будет понятно, что будет необходим определенный скрининг полученных вариантов лиганда mpl для отбора оптимального варианта для связывания с mpl, имеющего иммунологическую и/или биологическую активность, определенную выше. Можно просеять по стабильности в рекомбинантной культуре клеток или в плазме (например, по отношению к протеолитическому расщеплению), высокому сродству к членам mpl, устойчивости к окислению, по способности секретироваться в повышенном количестве, и тому подобное. Например, изменения в иммунологическом характере полипептида лиганда mpl, такое как сродство к данному антителу, определяется по иммуноопределению конкурентного типа. Другие потенциальные модификации свойств белка или полипептида, такие как окислительно/восстановительная или температурная стабильность, гидрофобность, или восприимчивость к протеолитической деградации, определяются по способам, хорошо известными из техники.
17. Общие способы приготовления антител к лиганду mpl
Приготовление антител
(i) Поликлональные антитела
Поликлональные антитела к полипептидам лиганда mpl или фрагментам обычно вырабатываются у животных путем многократных подкожных (п/к) или внутрибрюшинных (в/б) инъекций лиганда mpl и адъюванта. Может быть приемлемо присоединение лиганда mpl или фрагмента, содержащего целевую аминокислотную последовательность, к белку, который иммуногенен у вида, которого иммунизируют, например keyhole limpet гемоцианин, сывороточный альбумин, бычий тироглобулин или ингибитор трипсина из соевых бобов, используя бифункциональный или модифицирующий агент, например малеимидобензоилсульфосукцинимидный эфир (конъюгация через остатки цистеина), N-гидроксисукцинимид (через остаток лизина), глютаровый альдегид, ангидрид янтарной кислоты, SOCl12, или R1N=C=NR, где R и R1 являются алкильными группами.
Животных иммунизируют против полипептида лиганда mpl или фрагментов иммуногенными конъюгатами или производными путем сочетания 1 мг или 1 мкг пептида или конъюгата (для кроликов или мышей соответственно) с 3 объемами полного адъюванта Freund’а и внутрикожными инъекциями раствора во многих местах. Через месяц животное реиммунизируют 1/5 или 1/10 исходного количества пептида или конъюгата с полным адъювантом Freund’a путем подкожной инъекции во многих местах. Спустя 7 или 14 дней у животного берут кровь и исследуют сыворотку на титр антител к лиганду mpl. Животных реиммунизируют, пока титр не выйдет на плато. Предпочтительно, животное реиммунизируют конъюгатом того же лиганда mpl, но конъюгированного к различным белкам и/или при помощи различных перекрестно-сшивающих реагентов. Конъюгаты также могут быть получены в рекомбинантной культуре клеток в виде сплавленных белков. Агрегирующие агенты, такие как alum, также используют для усиления иммунного ответа.
(ii) Моноклональные антитела
Моноклональные антитела получают из популяции существенно гомогенных антител, то есть отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных природно наблюдающихся мутаций, которые могут присутствовать в минорных количествах. Таким образом, модификация "моноклональное" отмечает характер антитела как не присутствующего в смеси дискретных антител.
Например, моноклональные антитела к лиганду mpl по изобретению могут быть получены, используя гибридомный способ, впервые описанный Kohler and Milstein, Nature, 256: 495 [1975], или могут быть получены по способу с рекомбинантной ДНК (Патент США No.4816576 [Cabilyet al.]).
В гибридомном способе мыши или другое подходящее животное-хозяин, такое как хомяк, иммунизируют, согласно вышеописанному здесь, для выявления лимфоцитов, которые продуцируют или способны продуцировать антитела, которые будут специфически связывать белок, использованный для иммунизации. Альтернативно, лимфоциты можно иммунизировать in vitro. Лимфоциты затем сплавляют с клетками миеломы, используя подходящий сплавляющий агент, такой как полиэтиленгликоль, для образования клеток гибридомы (Coding, Monoclonal Antibodies: Principles and Practice, pp.59-103 [Academic Press, 1986]).
Полученные таким образом гибридомные клетки высеваются и выращиваются в подходящей культуральной среде, которая предпочтительно содержит одно или более соединений, которое ингибирует рост или выживаемость несплавленных, родительских миеломных клеток. Например, если родительские миеломные клетки лишены фермента гипоксантин-гуанидин-фосфорибозил-трансферазы (HGPRT [ГГФРТ] или HPRT [ГФРТ)], культуральная среда для гибридом обычно будет включать гипоксантин, аминоптерин и тимидин (HAT среда), вещества которой препятствуют росту ГГФРТ-дефицитных клеток.
Предпочтительные клетки миеломы - это те, которые эффективно сплавляются, поддерживают стабильный высокий уровень экспрессии антитела путем отбора продуцирующих антитела клеток и являются чувствительными к среде, такой как HAT среда. Среди этих предпочтительных клеточных линий миеломы находится линия миеломы мышей, таких как происходящие от МОРС-21 или МРС-11 опухоли мыши, доступные в Salk Institute Cell Distribution Center, San Diego, Калифорния США, и SP-2 клетки, доступные из American Type Culture Collection, Rockville, Мэриленд, США. Линии клеток миеломы человека и гетеромиеломы мыши и человека также описаны как продуцирующие моноклональные антитела человека (Kozbor, J. Immunol., 133: 3001 [1984]; Broder et al., Monoclonal Antibody Production Techniques and Applications, pp.51-63, Marcel Dekker, Inc., New York, 1987).
Культуральная среда, в которой растут гибридомные клетки, исследуется на продукцию моноклональных антител, направленных против лиганда mpl. Предпочтительно, связывающая специфичность моноклональных антител, продуцируемых клетками гибридомы, определяется по иммунопреципитации или по исследованию связывания in vitro, такому как радиоиммуноисследование (RIA), или в исследовании на ферментативено-пришитом иммуноадсорбенте (ELISA).
Сродство связывания моноклонального антитела можно, например, определить из анализа Скетчарда (Scatchard) no Munson and Pollard, Anal. Biochem., 107: 220 [1980].
После идентификации гибридомных клеток, которые продуцируют антитела желаемой специфичности, сродства, и/или активности, клоны можно субклонировать путем процедуры ограниченного разведения и выращивания по обычным способам, (Goding, там же). Подходящая культуральная среда для этой цели включает, например, модифицированную Дульбекко среду Игла или среду RPMI-1640. Помимо этого клетки гибридомы могут расти in vivo в виде асцитных опухолей у животных.
Моноклональные антитела, секретируемые субклонами, удобно выделяются из культуральной среды, асцитной жидкости или сыворотки путем общепринятых способов для очистки иммуноглобулинов, таких как, например, протеин А-Сефароза, хроматография на гидроксилаппатите, гельэлектрофорез, диализ или хроматография по сродству.
ДНК, кодирующую моноклональные антитела по настоящему изобретению, легко выделяется и секвенируется, используя общепринятые способы (например, путем использования олигонуклеотидных зондов, которые способны к специфическому связыванию с генами, кодирующими тяжелые и легкие цепи антител мыши). Гибридомные клетки по изобретению служат как предпочтительный источник подобной ДНК. Для установления синтеза моноклональных антител в рекомбинантной клетке хозяина однажды выделенная ДНК может быть включена в вектор экспресии, который затем трансфецируют в клетку-хозяин, такую как COS клетки обезьяны, клетки яичника китайского хомячка (СНО) или клетки миеломы, которые не продуцируют иным способом белок иммуноглобулина. ДНК также можно модифицировать, например, путем замены кодирующей последовательности для тяжелой и легкой цепей постоянного домена человека в гомологичном месте последовательности мыши (Cabilly et al., там же; Morrison, et al., Proc. Natl. Acad. Sci. USA, 81: 6851 [1984]), или путем ковалентного присоединения к иммуноглобулину, кодирующему последовательность всех или части кодирующей последовательности неиммуноглобулинового полипептида.
Обычно такие неиммуноглобулиновые полипептиды заменяют на постоянные домены антитела по изобретению или их заменяют на вариабельные домены объединенного участка антитела по изобретению для создания химерного бивалентного антитела, содержащего один объединенный участок антигена, имеющего специфичность к лиганду mpl, и другой объединенный участок антитела, имеющего специфичность к другому антигену.
Химерные или гибридные антитела также можно получить in vitro, используя известные способы из химии синтеза белков, включая те, которые используют перекрестно-сшивающие агенты. Например, иммунотоксины можно сконструировать, используя реакцию обмена дисульфидами, или путем образования тиоэфирной связи. Примеры подходящих реагентов для этих целей включают иминотиолат и метил-4-меркаптобутиримидат.
Для применения в диагностике антитела по изобретению обычно будут мечены обнаруживаемым радикалом. Обнаруживаемый радикал может быть одним из любых, которые способны производить либо непосредственно, либо опосредованно, детектируемый сигнал. Например, обнаруживаемый радикал может быть радиоизотопом, таким как 3H, 13С, 32P, 35S или 125I, флуоресцирующим или хемилюминисцирующим соединением, таким как флуоресцеин, изотиоцианат, родамин или люциферин; радиоактивной изотопной меткой, такой как, например, 125I, 32P, 14С или 3H, или фермент, такой как щелочная фосфатаза, бета-галактозидаза или пероксидаза хрена.
Любой известный в технике способ может быть применен для раздельного присоединения антитела к обнаруживаемому радикалу, включая способы, описанные Hunter, et al., Nature, 144: 945 [1962]; David, et al., Biochemistry, 13: 1014 [1974]; Pain, et al., J. Immunol. Meth., 40: 219 [1981]; и Nygren, J. Histochem. and Cytochem., 30: 407 [1982].
Антитела по настоящему изобретению можно применять в любых известных способах исследования, таких как исследование конкурентного связывания, прямое и непрямое исследование типа сэндвич, и иммунопреципитационное исследование. Zola, Monoclonal Antibodies: A Manual of Techniques, pp.147-158 (CRC Press, Inc., 1987).
Исследование конкурентного связывания основано на способности стандарта (которым может быть лиганд mpl или его иммунологически реактивная часть) конкурировать с тестируемым исследуемым образцом (лигандом mpl) по связыванию с ограниченным количеством антител. Количество лиганда mpl в тестируемом образце обратно пропорционально количеству стандарта, который связан с антителами. Для облегчения определения количества стандарта, который будет связан, антитела обычно переводят в нерастворимое состояние перед или после конкурентного связывания, так чтобы стандарт и исследуемое вещество, которое связывается с антителом, можно было легко отделить от стандарта и от исследуемого вещества, которое осталось несвязанным.
Исследование типа сэндвич включает использование двух антител, каждое способное к связыванию с различными иммуногенными частями, или эпитопами определяемого белка (лиганда mpl). В исследовании типа сэндвич тестируемый исследуемый образец связывается первым антителом, которое иммобилизовано на твердой подложке, и после этого второе антитело связывается с исследуемым образцом, таким образом образуя нерастворимый комплекс из трех частей. David and Greene, Патент США No.4376110. Второе антитело может быть само по себе мечено детектируемым радикалом (прямое исследование типа сэндвич), или может быть измерено, используя анти-иммуноглобулиновое антитело, которое мечено детектируемым радикалом (непрямое исследование типа сэндвич). Например, первый тип исследования типа сэндвич представляет исследование ELISA, в этом случае детектируемым радикалом является фермент (например, пероксидаза хрена).
(iii) Очеловеченные антитела и антитела человека
Способы с очеловеченными и нечеловеческими антителами хорошо известны в технике. В общем, очеловеченное антитело имеет введенные в него аминокислотные остатки из источника, который является нечеловеческим. Эти нечеловеческие аминокислотные остатки часто обозначают как "импортные" остатки, которые обычно берут из "импортного" вариабельного домена. Очеловечивание можно, по существу, провести, следуя способу Winter с соавторами (Jones et al., Nature, 321: 522-525 [1986]; Reichmann et al., Nature, 332: 323-327 [1988]; Verhoeyen et ai.. Science, 239: 1534-1536 [1988]), путем замены CDRs (остатков постоянного домена, прим. переводчика) грызунов или последовательности CDR на соответствующую последовательность антитела человека. Таким образом, такие "очеловеченные" антитела являются химерными антителами (Cabilly et al., там же), где заменено существенно меньше на соответствующую последовательность из нечеловеческих видов, чем интактный вариабельный домен человека. В действительности, очеловеченные антитела являются обычно антителами человека, в которых некоторые остатки CDR и, возможно, некоторые FR остатки (рабочей рамки) замещаются остатками из аналогичных участков антител грызунов.
Выбор вариабельных доменов человека, обоих, легкого и тяжелого, для использования с получении очеловеченных антител очень важен для того, чтобы снизить антигенность. В соответствии с так называемым "best-fit" (наилучшего соответствия) способом последовательность вариабельного домена антитела грызуна просеивается против полной библиотеки известных последовательностей вариабельных доменов человека. Последовательность человека, которая наиболее близка к таковой у грызунов, затем принимается в качестве рабочей рамки (FR -РР) для очеловеченного антитела (Sims et al., J. Immunol., 151: 2296 [1993]; Chotia and Lesk, J. Mol. Biol., 196: 901 [1987]). Другие способы использования конкретной рабочей рамки происходят от согласованной последовательности всех антител человека конкретной подгруппы легких и тяжелых цепей. Та же рабочая рамка может быть использована для нескольких очеловеченных антител (Carter et al., Proc. Nati. Acad. Sci. USA, 89: 4285 [1992]; Presta et al., J. Immunol., 151: 2623 [1993]).
Далее важно, чтобы антитела были очеловечены с сохранением высокого сродства и других благоприятных биологических свойств. Для достижения этой цели в соответствии с предпочтительными способами очеловеченные антитела получают при помощи анализа родственных последовательностей и различных задуманных очеловеченных продуктов, используя трехмерные модели родственных и очеловеченных последовательностей. Трехмерные модели иммуноглобулина широко доступны в технике и знакомы для опытного работника. Доступны компьютерные программы, которые иллюстрируют и представляют возможные трехмерные конформационные структуры выбранных кандидатов последовательностей иммуноглобулинов. Рассмотрение данных представлений позволяет проанализировать вероятную роль остатков в функционировании предполагаемой последовательности иммуноглобулина, то есть анализ остатков, которые влияют на способность предполагаемого иммуноглобулина связываться с антигеном. В этом направлении остатки FR (рабочей рамки) могут быть выбраны и объединены с согласованной и импортной последовательностью так, чтобы получить желательные характеристики антитела, такие как увеличенное сродство к антигену(ам)-мишени. В целом, остатки CDR, непосредственно и наиболее существенно участвуют во влиянии на связывание антигена. Для дальнейших подробностей см. Заявки США, Серийные No.07/934373, поданную 21 августа 1992 г., которая является составной частью заявки с серийным No.07/715272, поданной 14 июня 1991 г.
Альтернативно, в настоящее время возможно получить трансгенных животных (например, мышей), которые способны при иммунизации продуцировать полный репертуар антител человека в отсутствии эндогенной продукции иммуноглобулинов. Например, описано, что гомозиготная делеция в гене области соединения (JH) тяжелой цепи антигена у химерных и линии зародышевых мутантных мышей приводит к полному ингибированию эндогенной продукции антитела. Пересадка всего ряда гена зародышевой линии иммуноглобулина человека в такую зародышевую линию мутантных мышей приведет к продукции антител человека при замене антигена. См., например, Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90: 2551-255? [1993]; Jakobovita et al., Nature, 362: 255-258 [1993]; Bruggermann et al. Year in Immuno., 7: 3 [1993]. Антитела человека также можно продуцировать в проявляемых фагом библиотеках (Hoogenboom and Winter, J. Mol. Biol., 227: 381 [1991]; Marks et al., J. Mol. Biol., 222: 581 [1991]).
(iv) Биспецифические антитела
Биспецифические антитела являются моноклональными, предпочтительно очеловеченными или антителами человека, которые обладают способностью связываться, по крайней мере, с двумя различными антигенами. Способы для получения биспецифических антител известны в технике.
По традиции, продукция рекомбинантами биспецифических антител основана на совместной экспрессии двух пар иммуноглобулинов тяжелой цепи - легкой цепи, где две тяжелые цепи имеют различную специфичность (Millstein and Cuello, Nature, 305: 53-539 [1983]). Из-за случайности разнообразия тяжелой и легкой цепей иммуноглобулинов эти гибридромы (квадромы) продуцируют смесь возможных 10 различных молекул антител, из которых только одна имеет правильную биспецифическую структуру. Очистка правильной молекулы, которую обычно проводят при помощи этапов аффинной хроматографии, является более чем затруднительной и дает низкий выход продукта. Близкие способы описаны в публикациях РСТ No. WO 93/08829 (опубликоваая 13 мая 1993 г.) и у Trauneker et al., EMBO J., 10: 3655-3659 [1991].
В соответствии с различными и более предпочтительными подходами вариабельные домены антитела с желаемой специфичностью связывания (комбинированные участки антитело-антиген) сплавляют с последовательностями постоянных доменов иммуноглобулина. Сплавляют предпочтительно внутри постоянного домена тяжелой цепи иммуноглобулина, включающей, по крайней мере, часть петли областей СН2 и СН3. Предпочтительно иметь первую постоянную область тяжелой цепи (СН1), содержащей участок, необходимый для связывания легкой цепи, представляющий, по крайней мере, одно из сплавлений. ДНК, кодирующую сплавленную тяжелую цепь иммуноглобулина и, если желательно, легкую цепь иммуноглобулина, вводят в отдельный вектор экспрессии и совместно трансфецируют в подходящий организм хозяина. Этим обеспечивается большая гибкость в подстраивании взаимных соотношений трех фрагментов полипептидов в воплощении, когда используются неравные части цепей трех полипептидов в конструкции, дающей оптимальный выход. Однако возможно ввести кодирующую последовательность для двух или трех цепей полипептидов в один вектор экспрессии, когда экспрессия, по крайней мере, двух полипептидных цепей в равных отношениях приводит к высокому выходу или когда соотношения не имеют особого значения. В предпочтительном воплощении данного подхода биспецифические антитела составляют тяжелую цепь гибридного иммуноглобулина с одной специфичностью связывания в первой ручке, и гибридной паре тяжелая цепь - легкая цепь иммуноглобулина (обеспечивающей вторую специфичность связывания) в другой ручке. Было обнаружено, что эти асимметрические структуры облегчают отделение желаемых биспецифических соединений от нежелательных комбинаций цепей иммуноглобулинов, поскольку наличие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает легкий путь для разделения. Эти подходы описаны в нерешенной заявке, серийный No.07/931811, поданной 17 августа 1992 г.
Для дальнейших подробностей получения биспецифических антител см., например, Suresh et al., Methods in Enzymology, 121: 210 [1986].
(v) Гетероконъюгированные антитела
Гетероконъюгированные антитела также входят в объем настоящего изобретения. Гетероконъюгированные антитела состоят из двух ковалентно связанных антител. Такие антитела, например, было предположено, нацеливают иммунную систему на нежелательные клетки (Патент США No.4676980) и для лечения инфекции ВИЧ (публикации РСТ No.WO 91/00360 и WO 92/00373; ЕР 03089). Гетероконъюгатные антитела можно получить, используя способы ковалентного перекрестного связывания. Подходящие перекрестно-сшивающие агенты хорошо известны в технике и описаны в Патенте США No.4676980 по порядку номеров способов перекрестного пришивания.
IV. Терапевтическое использование мегакариоцитопоэтического белка лиганда mpl
Биологически активный лиганд mpl, обладающий гематопоэтической эффекторной функцией, обозначен здесь как мегакариоцитопоэтический или тромбопоэтический белок (ТП), может быть использован в стерильном фармацевтическом препарате для стимуляции мегакариоцитопоэтической или тромбопоэтической активности у пациентов, страдающих от тромбоцитопении, в результате нарушения продукции, захвата или повышенного распада тромбоцитов. Ассоциированную с тромбоцитопенией гипоплазию костного мозга (например, апластическую анемию после хемотерапии или трансплантации костного мозга) можно эффективно лечить соединениями настоящего изобретения, так же как и такие нарушения, как диссиминированная внутрисосудистая свертываемость (DIC=ДВС), иммунная тромбоцитопения (включая ВИЧ-индуцированную ИТП и не-ВИЧ-индуцированную ИТП), хроническая идиопатическая тромбоцитопения, наследственная тромбоцитопения, миелодисплазия и тромботическая тромбоцитопения. Кроме этого, данные мегакариоцитопоэтические белки могут быть полезны при лечении миелопролиферативных тромбоцитарных заболеваний, так же как и тромбоцитоза после воспалительных состояний и при дефиците железа.
Предпочтительное использование мегакариоцитопоэтического или тромбоцитопоэтического белка (ТП) по настоящему изобретению представляет: миелотоксическая хемотерапия при лечении лейкемии или солидных опухолей, миелоаблативная (подавляющая миелоидные линии клеток - прим. переводчика) хемотерапия при аутологичной или аллогенной пересадке костного мозга, идиопатическая апластическая анемия, наследственная тромбоцитопения и иммунная тромбоцитопения.
Еще другие нарушения, пригодные для лечения мегакариоцитопоэтическими белками по настоящему изобретению, включают дефекты или повреждения тромбоцитов в результате приема лекарств, яда или активации на искусственных поверхностях. В этих случаях можно использовать настоящие соединения для стимуляции "шеддинга - конечной стадии тромбоцитопоэза" новыми "неповрежденными" тромбоцитами. Для более полного перечисления пригодных применений см. выше, в особенности раздел (a)-(f) и цитируемые здесь ссылки.
Мегакариоцитопоэтические белки по настоящему изобретению можно применять сами по себе или в комбинации с другими цитокинами, гематопоэтинами, интерлейкинами, факторами роста или антителами для лечения вышеобозначенных нарушений и заболеваний. Таким образом, настоящие соединения можно применять в сочетании с другими белками или пептидами, имеющими тромбопоэтическую активность, включая Г-КСФ, ГМ-КСФ, ФИЛ, М-КСФ, ИЛ-1, ИЛ-3, эритропоэтин (ЭП), кит-лиганд, ИЛ-6 и ИЛ-11.
Мегакариоцитопоэтические белки по настоящему изобретению приготавливают в смеси с фармацевтически приемлемым носителем. Данные терапевтические составы можно вводить внутривенно или через нос или легкие. Композиции можно также вводить парентерально или подкожно по желанию. При систематическом введении терапевтические композиции должны быть апирогенными и представлять парентерально применимый раствор, считающийся со значением рН, изотоничности и стабильности. Эти условия известны опытному в технике сотруднику. В кратком виде дозовые составы соединений по настоящему изобретению приготавливают для хранения или применения путем смешивания соединений, обладающих желаемой степенью чистоты, с физиологически применимыми носителями, наполнителями или стабилизаторами. Такие материалы являются нетоксичными для реципиента в применяемых дозировках и концентрациях и включают буферы, такие как фосфатный, цитратный, ацетатный и другие соли органических кислот; антиоксиданты, такие как аскорбиновая кислота; пептиды низкого молекулярного веса (менее чем около 10 остатков), такие как полиаргинин, белки, такие как сывороточный альбумин, желатин, или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глютаминовую кислоту, аспарагиновую кислоту или аргинин; моносахара, дисахара и другие углеводы, включая целлюлозу или ее производные, глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахароспирты, такие как маннитол или сорбитол; противоионы, такие как натрий и/или неионные сурфактанты (поверхностно-активные вещества), такие как Твин, Pluronics или полиэтиленгликоль.
Около от 0,5 до 500 мг соединения или смеси мегакариоцитопоэтического белка как в свободной кислоте, или в виде основания, или в виде фармацевтически приемлемой соли смешивается с физиологически приемлемым разбавителем, носителем, наполнителем, связывающим, консервантом, стабилизатором, ароматизатором, и т.п., как предусмотрено для общепринятой фармацевтической практики. Количество активного ингредиента в данной композиции таково, чтобы получить подходящую дозировку в ряду положенных.
Стерильная композиция для инъекции может быть составлена в соответствии с общепринятой фармацевтической практикой. Например, может быть желательно разведение или суспендирование активного соединения в разбавителе, таком как вода или встречающемся в природе растительном масле, подобном сезамовому, арахисовому или хлопковому, или синтетическом жирном разбавителе, подобном этилолеату или подобному. Могут быть включены буферы, консерванты, антиоксиданты и подобные в соответствии с принятой фармацевтической практикой.
Подходящие примеры долговременно высвобождающихся препаратов включают полупроницаемые матриксы или твердые гидрофобные полимеры, содержащие полипептид, матрикс которых представляет вид заключенных в форму частиц, например пленку, или микрокапсулы. Примеры долговременно высвобождающихся матриксов включают полиэфиры, гидрогели (например, поли(2-гироксиметилметакрилат), как описано Langer et al., J. Biomed. Mater. Res., 15: 167-277 [1981] и Langer, Chem. Tech., 12: 98-105 [1982], или поли(винилалкоголь)), полилактиды (Патент США No. 3773919, ЕП 58481), сополимеры L-глютаминовой кислоты и гамма этил-L-глютамата (Sidman et al., Biopolymers, 22: 547-556 [1983]), нераспадающийся этиленвинилацетат (Langer et al., там же), распадающийся сополимер молочной кислоты и гликоловой кислоты, такой как Lupron Depot™ (инъекционные микросферы, состоящие из сополимера молочной кислоты-гликоловой кислоты и ацетата лейпролида), и поли-D-(-)-3-гидроксимасляная кислота (ЕП 133988).
Хотя полимеры, такие как этилен-винил ацетат и полимер молочной кислоты-гликолевой кислоты, позволяют высвобождаться молекулам более 100 дней, определенные гидрогели высвобождают белки за более короткий период времени. Когда заключенные в капсулы белки остаются в теле на более длительное время, они могут денатурировать или агрегировать в результате выдерживания во влажных условиях при 37°С, приводя к потере биологической активности и возможным изменениям в иммуногенности. Можно спланировать рациональную стратегию для стабилизации белка, зависящую от вовлеченных механизмов. Например, если обнаруживается, что механизм агрегации представляет внутримолекулярное образование S-S связей через дисульфидный взаимообмен, то стабилизацию можно достигнуть путем модификации сульфгидрильных остатков, лиофилизацией из кислых растворов, контролем содержания влаги, используя подходящие добавки и разрабатывая специальные композиции полимерных матриксов.
Долговременно высвобождающие композиции мегакариоцитопоэтического белка также включают заключенный в липосомальную ловушку мегакариоцитопоэтический белок. Липосомы, содержащие мегакариоцитопоэтические белки, получают по таким способам: DE 3218121; Epstein et al., Proc. Natl. Acad. Sci. USA, 82: 3688-3692 [1985]; Hwang et al., Proc. Natl. Acad. Sci. USA., 77: 4030-4034 [1980]; ЕП 36676; ЕП 143949; ЕП 142641; Японская патентная заявка 83-118008; Патенты США No.4485045 и 4544641; и ЕП 102324. Обычно липосомы являются униламеллярного типа, небольшие (около 200-800 Ангстрем), в которых содержание липидов больше чем 30 мол.% холестерина, выбранные пропорции подобраны для оптимальной терапии мегакариоцитопоэтическим белком.
Дозировка будет определяться наблюдающим врачом при рассмотрении различных известных факторов, модифицирующих действие препаратов, включая тяжесть и тип заболевания, вес тела, пол, диету, время и путь приема, другие механизмы и другие имеющие отношение клинические факторы. Обычно дневной режим будет варьировать от 0,1 до 100 мкг/кг веса тела. Предпочтительно, дозировка будет варьировать от 0,1 до 50 мкг/кг веса тела. Более предпочтительно, начальная доза будет варьировать от 1 до 5 мкг/кг/день. Необязательно, уровень дозировки будет таким же, как для других цитокинов, в особенности, Г-КСФ, ГМ-КСФ и ЭП. Терапевтически эффективные дозировки можно определять по способам in vitro или in vivo.
Примеры
Очевидно, что любой специалист исходя из предыдущего описания и иллюстрирующих его примеров, может осуществить и использовать настоящее изобретение во всей его полноте без каких-либо дополнительных описаний. Поэтому нижеприведенные рабочие примеры описывают лишь предпочтительные варианты настоящего изобретения и не должны рассматриваться как некое ограничение изобретения.
Пример 1
Частичная очистка свиного лиганда mpl
Обедненную тромбоцитами плазму собирали от нормальных свиней или анемичных свиней с аплазией. Аплазию у свиней индуцировали путем облучения всего тела животного излучением в 900 сантигрэй (cGy) с использованием линейного ускорителя при потоке энергии 4 МэВ. Облученным свиньям в течение 6-8 дней вводили внутримышечные инъекции цефазолина. Затем у животных под общей анестезией удаляли кровь в полном объеме и эту кровь гепаринизировали и центрифугировали при 1800×g в течение 30 минут, в результате чего получали обедненную тромбоцитами плазму. Пик мегакариоцитстимулирующей активности наблюдался через 6 дней после облучения.
Плазму, полученную от облученных свиней с аплазией, обрабатывали 4 М NaCl и размешивали при комнатной температуре в течение 30 минут. Полученный преципитат удаляли путем центрифугирования при 3800 об/мин в Sorvall RC3B, а супернатант загружали в колонку с Phenyl-Toyopearl (220 мл), уравновешенную в 10 мМ NaPO4, содержащий 4М NaCl. Колонку промывали этим буфером до тех пор, пока А280 не достигал значения менее 0,05, после чего колонку элюировали dH2O. Элюированный пик белка разводили dH2O так, чтобы его проводимость составляла 15 мСим, после чего его загружали в колонку с Blue-Sepharose, уравновешенную (240 мл) в PBS. Затем колонку промывали 5 колоночными объемами из PBS и 10 мМ NaPO4 (pH 7,4), содержащего 2М мочевины. Белки элюировали с колонки 10 мМ NaPO4 (pH 7,4), содержащим 2М мочевины и 1 М NaCl. Элюированный пик ьелка обрабатывали 0,01%-ным октилглюкозидом (н-октил β - D глюкопиранозидом) и 1 мМ каждого из EDTA и Pefabloc (Boehinger Mannheim) и непосредственно загружали в тандемно соединенные колонки с CD4 - IgG (Capon D.J/ et al. Nature 337: 525-531 (1989)) и mpl-IgG Ultralink (Pierce) (см. ниже). После загрузки образца, колонки с CD4-IgG (2 мл) удаляли, а колонку с mpl - IgG (4 мл) промывали 10 объемами каждого из PBS и PBS, содержащего 2М NaCl, и элюировали 0,1М глицином-HCl (pH 2,25). Фракции собирали в 1/10 объема 1 М Трис-HCl (pH 8,0).
Анализ элюированных фракций с mpl-аффинной колонки, проведенный с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ) (4-20%, Novex gel) в условиях восстановления, выявил присутствие несколько белков (фиг.5). Белки, которые окрашивались серебром с наибольшей интенсивностью, имели кажущуюся молекулярную массу Мг 66000, 55000, 30000, 28000 и 140000. Для того что определить, которые из этих белков стимулируют пролиферацию Ba/F3-mp1- клеточных культур, эти белки элюировали из геля, как описано ниже в Примере 2.
Ultralink аффинная колонка
10-20 мг mpl-IgG или CD4-IgG в PBS объединяли с 0,5 г смолы Ultralink (Pierce), как описано в инструкциях производителей.
Коструирование и экспрессия mpl-IgG
Химерную молекулу, содержащую полный внеклеточный домен mpl человека (аминокислоты 1-491) и Fc-фрагмент молекулы IgGI человека, экспрессировали в клетках 293. кДНК-фрагмент, кодирующий аминокислоты 1-491 mpl человека, получали с помощью полимеразной цепной реакции (PCR) из кДНК мегакариоцитов СМК и секвенировали. Clal-сайт вводили в 5’-конец, а BstEll-сайт вводили в 3’-конец. Этот фрагмент клонировали выше IgGl-Fc-кодирующей области в векторе Bluescript, между сайтами Clal и BstEll, после частичного гидролиза PCR-продукта ферментом BstEll, поскольку в ДНК, кодирующей внеклеточный домен mpl, присутствуют два других BstEll-сайта. BstEll-сайт, введенный в 3’-конец PCR-продукта mpl, конструировали так, чтобы Fc-область имела ту же рамку считывания, что и внеклеточный домен mpl. Эту конструкцию субклонировали в вектор pPK5-tkneo между сайтами Clal и Xbal, и трансфицировали в эмбриональные клетки почки человека 293 методом с использованием фосфата кальция. Эти клетки отбирали в 0,4 мг/мл G418 и отдельные клоны выделяли. Экспрессию mpl-IgG из выделенных клонов оценивали с помощью чел. Fc-специфического ELISA. Наиболее высокоэкспрессирующий клон имел уровень экспрессии 1-2 мг/мл mpl-IgG.
Клетки, эксперессируюцие mpl Ba/F3
кДНК, соответствующую полной кодирующей области mpl F, клонировали в pPKS-tkneo, который затем линеаризовали ферментом Notl, и трансфецировали в IL-3-зависимую клеточную линию Ba/Fe путем электропорации (1×107 клеток, 9605F, 250В). Через три дня проводили отбор в присутствии 2 мг/мл G-418. Клетки отбирали в виде пулов либо получали отдельные клоны путем серийного разведения в 96-луночных планшетах. Отобранные клетки выдерживали в RPM1, содержащей 15% FBS, 1 мг/мл G-418, 20 мМ глутамина, 10 мМ HEPES и 100 мкг/мл Pen-Strep. Экспрессию mpl P в отобранных клонах оценивали флуоресцентным методом разделения клеток (FAC-анализ) с использованием кроличьих поликлональных антител против mpl Р.
Анализ на присутствие лиганда mpl Ba/F3
Анализ на присутствие лиганда mpl проводили, как показано на фиг.2. Для определения присутствия mpl-лиганда от различных источников клетки mpl P-Ba/F3 культивировали в среде, не содержащей IL-3, в течение 24 часов при клеточной плотности 5×105 кл./мл во влажной камере при 37°С в атмосфере 5% СO2 и воздуха. После выращивания клеток в среде, не содержащей IL-3, эти клетки высевали в 96-луночные планшеты при плотности 50000 клеток в 200 мкл среды в присутствии или в отсутствие разведенных образцов и культивировали в течение 24 часов в термостате для клеточных культур. Затем в течение последних 6-8 часов к каждой лунке добавляли бессывороточную RPMl-среду (20 мкл), содержащую 1 мкКи 3H-тимидина. После этого клетки собирали на в 96-луночных планшетах с фильтром GF/C, и 5 раз промывали водой. Фильтры подвергали анализу в счетчике Packard Top Count в присутствии 40 мкл сцинтилляционной жидкости (Microscint 20).
Пример 2
Высокоочищенный свиной лиганд mpl
Протокол гель-элюирования
Равные количества аффиноочищенного mpl-лиганда (фракция 6, элюированная с mpl-IgG-колонки) и 2× буфера Лэммли смешивали при комнатной температуре в отсутствие восстановителя и как можно быстро загружали на 4-20% полиакриламидный гель Novex. Образец не нагревали. В качестве контроля на соседней дорожке проводили электрофорез буфера без лиганда. Гель подвергали электрофорезу при 135 вольт в течение приблизительно 2 1/4 часа при 4-6°С. Сначала пропускали буфер при комнатной температуре. Затем гель удаляли из лотка и пластину с одной стороны геля удаляли.
Реплики геля на нитроцеллюлозе приготавливали следующим образом. Кусок нитроцеллюлозы, смоченный дистиллированной водой, осторожно помещали верхней стороной на поверхность геля так, чтобы не образовывались пузырьки воздуха. На нитроцеллюлозе и пластине геля делали ориентировочные метки для того, чтобы реплики можно было точно репозиционировать после окрашивания. Примерно через 2 минуты нитроцеллюлозу осторожно удаляли, а гель оборачивали полимерной пленкой и помещали в холодильник. Нитроцеллюлозу окрашивали красителем для полного белка Biorad gold (золотым) сначала путем ее размешивания в 3×10 мл 0,1% Твин 20+0,5 М NaCl+0,1 M Трис-HCl (рН 7,5) в течение приблизительно 45 минут, а затем в 3×10 мл очищенной воды в течение 5 минут. Затем добавляли золотой краситель и оставляли для развития окраски до тех пор, пока полосы в стандартах не становились видимыми. После этого реплики промывали водой, помещали поверх полиэтиленовой пленки на геле, осторожно совмещали с ориентировочными метками. На пластине геля были размечены положения стандартов Novex и проведены линии, указывающие места для вырезания. Затем нитроцеллюлозу и полиэтиленовую пленку удаляли и гель вырезали вдоль указанных линий острым лезвием бритвы. Разрезы продолжались за пределы дорожек геля для образцов так, чтобы они могли быть использованы для определения положений срезов при окрашивании геля. После удаления срезов оставшийся гель окрашивали серебром и положения стандартов и меток измеряли. Молекулярные массы, соответствующие положениям вырезания, определяли исходя из стандартов Novex.
12 Срезов геля помещали в ячейки двух аппаратов для электроэлюирования модели Biorad 422. В этих ячейках использовали мембраны, отсекающие молекулярные массы 12-14 кДа. В качестве элюирующего буфера использовали 50 мМ бикарбоната аммония +0,05% ДСН (приблизительно рН 7,8). Один литр буфера охлаждали примерно в течение 1 часа перед его использование в холодном помещении при 4-5°С. Срезы геля элюировали при 10 мА/ячейка (начальное напряжение 40 В) в холодном помещении при 4-6°С. Элюцию проводили примерно 4 часа. Затем ячейки осторожно вынимали и жидкость поверх фрита удаляли пипеткой. Затем камеру для элюирования вынимали и всю жидкость поверх мембраны удаляли пипеткой. Жидкость в мембране удаляли пипеткой Pipetman и сохраняли. Затем в мембрану добавляли 50 мкл очищенной воды, размешивали и удаляли до тех пор, пока не растворялись все кристаллы ДСН. После этого промывки объединяли с вышеупомянутой сохраненной жидкостью. Полный объем образца для элюции составлял 300-500 мкл на срез геля. Образцы помещали в 10-миллиметровую 12-14 К-отсекающую диализную трубку Spectrapor 4, которую предварительно в течение нескольких часов пропитывали очищенной водой. Диализ проводили в течение ночи при 4-6°С против 600 мл забуференного фосфатом физиологического раствора (PBS-приблизительно 4 мМ в калии) на 6 образцов. Затем на следующее утро буфер заменяли и диализ продолжали еще в течение 2,5 часов. Затем образцы удаляли из диализационных мешков и помещали в пробирки для микроцентрифугирования. Эти пробирки на 1 час помещали на лед, центрифугировали в течение 3 минут при скорости (об/мин) на отметке 14К и супернатанты осторожно удаляли из осажденного ДСН. Эти супернатанты помещали примерно на 1 час на лед и еще раз центрифугировали в течение 4 минут. Затем супернатанты разводили в физиологическом растворе, забуференном фосфатом, и подвергали анализу на активность. Оставшиеся образцы замораживали при -70°С.
Пример 3
Микросеквенирование лиганда mpl свиньи
Фракцию 6 (2,6 мл) из mpl-IgG-аффинной колонки концентрировали на Microcon-10 (Amicon). Во избежание абсорбции лиганда mpl на Microcon мембрану промывали 1% ДСН и к фракции 6 добавляли 10% ДСН. После концентрирования фракции 6 на Microcon (20 мкл) добавляли буфер для образца (20 мкл) (2х) и полученный объем (40 мкл) загружали на одну дорожку 4-20% градиента акриламидного геля (Novex). Затем гель подвергали электрофорезу в соответствии с протоколом Novex. После этого до проведения электроблотирования гель уравновешивали в течение 5 минут в 10 мМ САРS-буфере (CAPS=3-(циклогексиламино)-1-пропансульфоновая кислота), рН 11/0, содержащем 10%-ный метанол. Электроблоттинг на мембранах Immobilon-PSQ (Millipore) проводили в течение 45 минут при постоянном токе 250 мА в ячейке для электрофоретического переноса (32) BioRad Trans-Blot. PVDF-мембрану окрашивали 0,1% кумасси синим Р-250 в 40%-ном метаноле, 0,1%-ной уксусной кислоте в течение 1 минуты, после чего краситель отмывали в течение 2-3 минут с использованием 10%-ной уксусной кислоты в 50%-ном метаноле. Белки, которые были видны в 18000-35000-области блота, имели Mr=30000, 28000 и 22000.
Полосы белков 30, 28 и 22 кДа подвергали секвенированию. Для этого осуществляли автоматическое секвенирование на секвенаторе модели 470А (Applied Biosystem), снабженном анализатором РТН, работающем в реальном масштабе времени. Указанный секвенатор был модифицирован для введения 80-90% образца (Rodriguez, J. Chromatogr., 350: 217-225 [1985]). Для регулирования оптической плотности в УФ-диапазоне к растворителю А добавляли ацетон (≈12 мкл/л). Электроблотированные белки секвенировали в кассете Blott. Пики интегрировали с помощью программного обеспечения Justic Innovations с использованием интерфейсов Nelson Analitycal 970. Интерпретацию последовательностей проводили на приборе vAX 5900 (Henzel et al., J. Chromatogr., 404: 41-52 [1987]). N-концевые последовательности (с использованием однобуквенного кода; неопределенные остатки приведены в скобках) и количества полученного материала (в квадратных скобках) представлены в Таблице 4.

Пример 4
Анализ на мегакариоцитопоэз в жидкой суспензионной культуре
Периферические стволовые клетки человека (PSC) (полученные с согласия пациентов) 5-кратно разводили 1М М-средой (Gibco), и центрифугировали в течение 15 минут при 800×g при комнатной температуре. Клеточный осадок ресуспендировали в 1 MDM и покрывали слоем поверх 60% Percoll (плотность 1,077 г/мл) (Pharmacia), после чего центрифугировали в течение 30 минут при 800×g. Мононуклеарные клетки небольшой плотности отсасывали у границы и промывали 2×1 MDM, после чего их высевали при плотности (1-2)·106 кл./мл в среду 1 MDM, содержащую 30% FBS (конечный объем 1 мл), в 24-луночные планшеты для культивирования тканевых культур (Costar). Затем добавляли (до 10%) АРР или АРР, не содержащий лиганд mpl, и культуры выращивали в течение 12-14 дней во влажной камере при 37°С в атмосфере 5% CO2 и воздуха. После этого культуры инкубировали в присутствии 10% АРР с 0,5 мкг добавленного mpl-IgG, добавленного в дни 0, 2 и 4. Удаление лиганда mpl из АРР осуществляли путем пропускания АРР через mpl-IgG-аффинную колонку.
Для количественной оценки мегакариоцитопоэза в этих жидких суспензионных культурах использовали модифицированный метод Solberg и др. И радиоактивно меченные мышиные моноклональные антитела IgG (HP1-1D) к Gpllbllla (поставляемые Dr.Nichols, Mayo Clinic). 100 мкг HP1-D (см. Grant, В. Et al., Blood 69: 1334-1339 [1987]) подвергали радиоактивному мечению 1 мКи Na125 с использованием Enzzmobeads (Biorad, Richmond, CA) в соответствии с инструкциями производителей. Радиоактивно меченные HP1-1D хранили при 70°С в PBS, содержащем 0,01% октилгликозид. Типичная специфическая активность составляла 1-2-10° им./мин/мкг (>95%, осажденных 12,5%-ной трихлоруксусной кислотой).
Жидкие суспензионные культуры приготавливали в тройном дубликате для каждой экспериментальной стадии. Через 12-14 дней выдерживания 1 мл культур переносили в 1,5-миллилитровые пробирки Эппендорфа, затем центрифугировали при 800×g в течение 10 минут при комнатной температуре, и полученные клеточные осадки ресуспендировали в 100 мкл PBS, содержащего 0,02% EDTA и 20% телячьей сыворотки. К ресуспендированным культурам добавляли 10 нг 125l-HPI-ID в 50 мкл аналитического буфера и инкубировали в течение 60 минут при комнатной температуре (КТ), периодически встряхивая. После этого клетки собирали путем центрифугирования при 800×g в течение 10 минут при КТ и промывали 2х буфером для анализа. Клеточный осадок помещали на 1 минуту в гамма-счетчик для подсчета (Packard). Неспецифическое связывание оценивали путем добавления 1 мкг немеченого HP1-1D с последующим выдерживанием смеси в течение 60 минут перед добавление меченого HP1-1D. Специфическое связывание определяли как общую величину 125l-НРl-lD-связывания минус связывание в присутствии избыточного количества немеченого HP1-1D.
Пример 5
Олигонуклеотидне PCR-праймеры
Исходя из амино-концевой аминокислотной последовательности, полученной для белков 30, 28 и 18-22 кДа, конструировали вырожденные олигонуклеотиды в качестве PCR-праймеров (PCR-полимеразная цепная реакция) (см. Таблицу 5). Было синтезировано два пула праймеров: один смысловой 20-мерный пул, кодирующий аминокислотные остатки 2-8 (mpl 1) и антисмысловой 21-мерный пул, комплементарный последовательностям, кодирующим аминокислоты 18-24 (mpl 2).
Свиную геномную ДНК, выделенную из лимфоцитов свиной периферической крови, использовали в качестве матрицы для PCR. 50 мкл реакционной смеси содержали: 0,8 мкл свиной геномной ДНК в 10 мМ Трис-НСl (рН 8,3), 50 MM KCl, 3 мМ MgCl2, 100 мкг/мл BSA, 400 мкМ dNTP, 1 мкМ каждого праймерного пула и 2,5 ед. полимеразы Тао. Первоначальную денатурацию матрицы проводили при 94°С в течение 8 минут с последующими 35 циклами по 45 сек при 94°С, 1 мин при 55°С и 1 мин при 72°С. Конечный цикл проводили в течение 10 минут при 72°С. PCR-продукты разделяли с помощью электрофореза на 12%-ном полиакриламидном геле и визуализировали путем окрашивания этидийбромидом. Разумно предположить, что если амино-концевая аминокислотная последовательность кодируется одним экзоном, то правильный PCR-продукт, очевидно, составляет 69 п.о. ДНК-фрагмент этого размера элюировали из геля и сублокировали в pGEMT (Promega). Последовательности трех клонов показаны ниже:
Свиные геномные ДНК-фрагменты длиной 69 п.о.
gem T3
5`CCAGCGCCGC CAGCCTGTGA CCCCCGACTC СТАААТАААС TGCCTCGTGA
3`GGTCGCGGCG GTCGGACACT GGGGGCTGAG GATTTATTTG ACGGAGCACT
TGACCACGTT CAGCACGGC [69 п.о.] Посл. NO: 37
ACTGGTGCAA GTCGTGCCG Посл.NO: 38
gem T7
5`CCAGCACCTC CGGCATGTGA CCCCCGACTC СТАААТАААС TGCTTCGTGA
3`GGTCGTGGAG GCCGTACACT GGGGGCTGAG GATTTATTTG ACGAAGCACT
CGACCACGTC CATCACGGC [69 п.о.] Посл.NO: 39
GCTGGTGCAG GTAGTGCCG Посл.NO: 40
Пример 6
Ген лиганда mpl человека
Исходя из результатов, полученных, как описано в Примере 5, конструировали и синтезировали 45-мерный дезоксиолигонуклеотид, обозначенный pR45, для скрининга геномной библиотеки. Указанный 45-мер имел следующую последовательность:
5’ GCC-GTG-AGG-GAC-GTG-GTC-GTC-ACG-AAG-CAG-TTT-ATT-TAG-GAG-TCG 3’ (Посл.NO: 28)
Этот олигонуклеотид подвергали 32Р-мечению с использованием (γ32P)-ATP и киназы Т4 и использовали для скрининга геномной ДНК-библиотеки в λgеm 12 в жестких условиях гибридизации и промывания (см. Пример 7). Положительные клоны собирали, бляшки очищали и анализировали путем рестрикционного картирования и Саузерн-блоттинга. Клон №4 отбирали для дополнительного анализа.
BamHl-Xbal-фрагмент (2,8 кв), который гибридизировался с 45-мером, субклонировали в вектор pBluescript SK-. Частичное ДНК-секвенирование этого клона осуществляли с использованием олигонуклеотидных праймеров, специфичных к ДНК-последовательности свиного лиганда mpl. Полученная последовательность подтвердила, что была выделена ДНК, кодирующая человеческий гормон лиганда mpl свиньи. В этой последовательности был обнаружен рестрикционный EcoRl-сайт, что позволило авторам этой заявки выделить 390 п.о. -EcoRl-Xbal-фрагмент из 2,8кв-ВатН1-Xbal-фрагмента для его субклонирования в вектор pBluescript SK-.
Обе нити этого фрагмента секвенировали. Человеческая ДНК-последовательность и выведенная аминокислотная последовательность показаны на фиг.9 (Посл. NOS 3 и 4). Предсказанные положения интронов в геномной последовательности также показаны стрелками и определяют предполагаемый экзон ("экзон 3").
Исследование предсказанной аминокислотной последовательности подтвердило, что сериновый остаток является первой аминокислотой зрелого лиганда mpl, как было определено из прямого анализа аминокислотной последовательности. Предполагается, что непосредственно выше от этого кодона находится сигнальная последовательность, участвующая в секреции зрелого лиганда mpl. Область, кодирующая сигнальную последовательность, очевидно, прерывается интроном в положении нуклеотида 68.
Этот экзон, вероятно, оканчивается в положении нуклеотида 196 в направлении 3`. Поэтому этот экзон кодирует последовательность в 42 аминокислот, из которых 16, очевидно, являются частью сигнальной последовательности, а 26 являются частью зрелого лиганда inpl человека.
Пример 7
Полноразмерная кДНК лиганда mpl человека
Исходя из последовательности "экзон 3" человека (Пример 6) синтезировали два невырожденных олигонуклеотида, соответствующие 3`- и 5`-концам последовательности "экзон 3" (Таблица 6).
Таблица 6
Невырожденные чел. кДНК-олигонуклеотидные PCR-праймеры
Эти два праймера были предназначены для полимеразной цепной реакции, которую проводили в условиях, описанных в Примере 5 с использованием в качестве матрицы ДНК от различных чел.кДНК-библиотек или 1 нг кДНК-клона (Quick DNA, Clonetech) от различных тканей. Ожидаемый размер правильного PCR-продуктов на 12%-ном полиакриламидном геле, ДНК-фрагмент ожидаемого размера был обнаружен в кДНК-библиотеках, полученных от почки взрослого индивидуума, из клеток фетальной почки 293 и ДНК, полученной из фетальной печени человека (Clonetech кат.№7171-1).
кДНК-библиотеку от фетальной печени в DR2 (Clonetech кат. №Н 1151 х) скринировали с использованием того же самого 45-мерного олигонуклеотида, который был использован для скрининга геномной библиотеки. Олигонуклеотид метили (γ32Р)-АТР с использованием полинуклеотид-киназы Т4. Библиотеку скринировали в гибридизационных условиях низкой жесткости. Фильтры предварительно гибридизировали в течение 2 часов, а затем гибридизировали с зондом в течение ночи при 42°С в 20%-ном формамиде, 5×SSC, 10х растворе Денхардта, 0,05 М фосфата натрия (рН 6,5), 0,1% пирофосфате натрия, 50 мкг/мл обработанной ультразвуком ДНК спермы лосося в течение 16 часов. Затем фильтры промывали в 2×SSC, а затем один раз в 0,5×SSC, 0,1% ДСН при 42°С. Фильтры экспонировали с рентгеновской пленкой (Kodak) в течение ночи. Положительные клоны собирали, бляшки очищали и размер вставки определяли посредством полимеразной цепной реакции с использованием олигонуклеотидов, фланкирующих BamHl-Xbal-франмент, клонированный в λ DR2 (Clonetech кат.№6475-1). В качестве источника матрицы использовали 5 мкл исходной фаговой культуры. Начальную денатурацию проводили 7 минут при 94°С с последующими 30 циклами амплификации (1 мин при 94°С, 1 мин при 52°С и 1,5 мин при 72°С). Конечное удлинение проводили в течение 15 минут при 72°С. Клон №FL2b имел 1,8 ко-вставку и был отобран для последующего анализа.
"Спасение" плазмиды pDR2 (Clonetech, DR2 & pDR2 cloning and Expression System Library Protocol Handbook, p.42), содержащейся λ ВР2-фаговых плечах, осуществляли, как описано в инструкциях производителей (Clonetech, DR2 & pDR2 cloning and Expression System Library Protocol Handbook, p.29-30). Рестрикционный анализ плазмиды pDR2-FL2b, проведенный с использованием ферментов BamHl и Xbal, выявил присутствие рестрикционного BamHl-сайта внутри вставки приблизительно в положении 650. Гидролиз плазмиды ферментами BarnHl-Xbal разрезал вставку на два фрагмента, один 0,65 кв и один 1,15 кв. ДНК-последовательность определяли с использованием матрицы трех различных классов, происходящей от плазмиды pDR2-FL2b. ДНК-секвенирование двухцепочечной плазмидой ДНК осуществляли с помощью автоматического флуоресцентного ДНК-секвенатора АВ1373 (Applied Biosystems, Foster City, California) с использованием стандартной схемы секвенирования с применением меченных красителем дидезоксинуклеозидтрифосфат-терминаторов (краситель-терминатор) и синтезированных традиционным способом праймеров для "прогулки по геному" (Sanger et al., Proc. Nati.Acad.Sci. USA, 74: 5463-5467 [1977]; Smith et ai., Nature, 321: 674-679 [1986]). Прямое секвенирование PCR-амплифицированных фрагментов, полученных из плазмиды, осуществляли с помощью секвенатора АВ1373 с использованием обычных праймеров и меченый терминатор. Одноцепочечную матрицу генерировали с использованием вектора М13 Janus (DNASTAR, Inc., Madison, Wisconsin) (Burland et al., Nucl. Acids res., 21: 3385-3390 [1993]). BamHI-Xbal (1,15 кв) - и BamHl (0,65 кв)-фрагменты выделяли из плазмиды pDR2, концы застраивали с помощью ДНК-полимеразы Т4 в присутствии дезоксинуклеотидов, а затем субклонировали в Smal-сайт М13 Janus. Секвенирование осуществляли в соответствии со стандартной методикой, предусматривающей использование меченных красителем универсального и обратного праймеров М13 либо праймеров для "прогулки по геному" и красителей-терминаторов. Обычное неавтоматическое секвенирование проводили в соответствии со стандартной методикой на одноцепочечной ДНК М13 с использование праймеров для "прогулки по геному" и традиционной техники дидезокси-терминации (Sanger et al., Proc. Natl.Acad.Sci. USA, 74: 5463-5467 [1977] с применением 32Р-меченного α-dATP и Секвеназы (United States Biochemical Corp. Cleveland, Ohio). Сборку ДНК-последовательности осуществляли с использованием секвенатора v2.1bl2 (Gene Codes Corporation. Ann Arbor, Michigan). Нуклеотидная и выведенная последовательности hML представлены на фиг.1 (Посл.NO: 1).
Пример 8
Выделение гена лиганда mpl человека (ТРО)
Геномные чел. ДНК-клоны гена ТРО выделяли путем скрининга геномной библиотеки в X-Geml2 с использованием pR45, ранее описанного олигонуклеотидного зонда, а условиях низкой жесткости (см. Пример 7), либо в условиях высокой жесткости с использованием фрагмента, соответствующего 3’-половине чел.кДНК, кодирующей лиганд mpl (от BamHl-сайта до 3’-конца). Были выделены два перекрывающиеся клона лямбда протяженностью 35 ко. Два перекрывающиеся фрагмента (BamHl и EcoRl), содержащие полный ген ТРО, были субклонированы и секвенированы. Структура этого гена человека состояла из 6 экзонов в 7 ко. геномной ДНК (фиг.14А, В и С). Границы всех экзон-интронных сочленений соответствовали консенсусному элементу, установленному для генов млекопитающих (Shapiro M.B., Nucl.Acids Res. 15:7155 [1987]). Экзон 1 и экзон 2 содержат 5-нетранслируемую последовательность и начальные четыре аминокислоты сигнального пептида. Оставшийся секреторный сигнал и первые 26 аминокислот зрелого белка кодировались в экзоне 3. Полный карбокси-домен и 3’-нетранслируемая область, а также -50 аминокислоты эритропоэтин-подобного домена кодировали в экзоне 6.. Четыре аминокислоты, входящие в делецию, наблюдаемую в hML-2 (hTPO-2), кодировались в 5'-конце экзона 6.
Пример 9
Кратковременная экспрессия лиганда mpl человека (hML)
В целях субклонирования полноразмерной вставки, содержащейся в pDR2-FL2b, плазмиду полностью гидролизовали ферментом Xbal, а затем частично ферментом Bamhl. ДНК-фрагмент, соответствующий 1,8 ко-вставке подвергали гель-очистке и субклонировали в pRK5 (pRK5-hmpl 1) (см. Патент США №5258287 для конструирования pRK5) под контролем предраннего промотора цитомегаловируса. ДНК из конструкции pRK5-hmpl 1) получали REG-методом и трансфицировали в человеческие эмбриональные клетки почки 293, поддерживаемые в модифицированной по способу Дульбекко среде Игла (DMEM), в которую были добавлены: питательная смесь F-12, 20 мМ Hepes (pH 7,4) и 10% фетальная коровья сыворотка. Клетки были трансфицированы кальцийфосфатным методом, как описано (Corman С. [1985] в DNA Cloning: A Practical Approach (Glover, D.M., ed.) Vol.11, pp.143-190, IRL Press, Washington, D.C.). Через 36 часов после трансфекции супернатант трансфицированных клеток анализировали на пролиферативную активность (см. Пример 1). Супернатант клеток 293, трансфицированных лишь одним вектором pRK, не обнаруживал стимуляции ни клеток Ba/F3, ни клеток Ba/F3-mpl (фиг.12А). Супернатант от клеток, трансфицированных pRK5-hmpl 1, не дал какого-либо эффекта в отношении клеток Ba/F3, но обнаруживал значительную стимуляцию пролиферации клеток Ba/F3-mpl (фиг.12А), что свидетельствует о том, что эта кДНК кодирует функционально активный лиганд mpl человека.
Пример 10
Изоформы hML2, hML3 и hML4 чел. лиганда
Для идентификации форм hML, продуцированных в результате альтернативного сплайсинга, синтезировали праймеры, соответствующие каждому концу кодирующей последовательности hML.
Эти праймеры использовали для ампликации РНК печени взрослого человека с помощью полимеразной цепной реакции при комнатной температуре (RT-PCR). Кроме того, были сконструированы и аналогичным образом использованы внутренние праймеры, фланкирующие выбранные нужные области (см. ниже). Прямое секвенирование концов PCR-продукта выявило одну последовательность, которая точно соответствовала последовательности кДНК, выделенной из библиотеки фетальной печени человека (см. фиг.1 (Посл.NO: 1)). Однако область, расположенная возле С-конца ЕРО-домена (в середине PCR-продукта), обнаруживала комплексные последовательности, позволяющие предположить существование вариантов возможного сплайсинга в этой области. Для выделения этих вариантов сплайсинга праймеры, проиллюстрированные в Таблице 7 и фланкирующие нужную область, были использованы в PCR в качестве матриц для кДНК печени взрослого человека.
Таблица 7
PCR-праймеры для выделения изоформы ML человека
PCR-продукты субклонировали по тупым концам в М13. Секвенирование отдельных субклонов выявило существование, по крайней мере, трех изоформ ML. Одна из них hML (обозначаемая также hML332) является самой длинной и точно соответствует последовательности, выделенной из библиотеки фетальной печени. Последовательности изоформ чел. лиганда mpl от самой длинной до самой короткой (hML-4) показаны на фиг.11 (Посл.NO 6, 8,9 и 10).
Пример 11
Конструирование и кратковременная экспрессия изоформ чел. лиганда Mpl и его вариантов замещения hML2, hML3 и hML (R153A, R154A)
Изоформы hML2 и hML3 и субституционные варианты (варианты замещения) hML (R153A, R154A) конструировали из hML с использованием рекомбинантной PCR-техники, описанной Russell Higuchi, в PCR Protocols, A guide to Methods and Applications Acad. Press, M.A.Innis, D.H.Gelfand, J.J.Sninsky & T.J.White Editors.
Во всех конструкциях "фланкирующие" праймеры были такими, как показано в Таблице 8, а "перекрывающиеся" праймеры были такими, как показано в Таблице 9.

Все PCR-ампликации проводили с использованием готовой к употреблению ДНК-полимеразы (Stratagene) при следующих условиях: начальную денатурацию матрицы осуществляли при 94°С в течение 7 минут, а затем проводили 30 циклов: 1 мин при 94°С; 1 мин при 55°С; 1,5 мин при 72°С. Конечный цикл продолжался 10 минут при 72°С. Полученный PCR-продукт гидролизовали Clal-Xbal-ферментами, подвергали очистке на геле и клонировали в pRK5tkneo. Клетки 293 трансфицировали различными конструкциями, описанными выше, а супернатант от этих клеток анализировали на пролиферацию Ba/F3-mpl. В этом анализе hML-2 и hML-3 не показали заметной активности, однако активность hML (R153A, R154A) оказалась аналогичной активности hML, что свидетельствует о том, что для активности не требуется процессинга в этом двухосновном сайте (см.фиг.13).
Пример 12
кДНК мышиного лиганда mpl, mML, mML-2, inML-3
Выделение кДНК mML
ДНК-фрагмент, соответствующий полной кодирующей области чел.лиганда mpl, получали с помощью PCR, а затем подвергали гель-очистке и мечению методом случайного праймирования в присутствии 32P-dATP и 32P-dCTP. Этот зонд использовали для скрининга кДНК-библиотеки от мышиной печени в GT10 (Clontech кат. №М 3001а). Дубликатные фильтры гибридизировали в 35% формамиде, 5×SSC, 10× растворе Денхардта, 0,1% ДСН, 0,05 М фосфате натрия (рН 6,5), 0,1% пирофосфате натрия, 100 мкг/мл обработанной ультразвуком ДНК спермы лосося, в присутствии зонда в течение ночи. Фильтры промывали в 2×SSC, а затем один раз промывали в 0,5×SSC, 0,1% ДСН при 42°С. Гибридизирующий фаг очищали методом бляшек, и кДНК-вставки сублокировали в EcoRl-сайт плазмиды Bluescript SK-. Клон "LD", содержащий 1,5 кb-вставку, отбирали для последующего анализа и обе нити секвенировали, как описано выше для чел. кДНК ML. Нуклеотидная и выведенная аминокислотная последовательности клона LD представлены на фиг.14 (SEQ ID NOS 1 и 11). Выведенная последовательность зрелого ML от этого клона имела длину в 331 аминокислотных остатков и была идентифицирована как mМL331 (или mML-2 из соображений, указанных ниже). Значительная степень идентичности обеих нуклеотидной и выведенной аминокислотной последовательностей наблюдалась в ЕРО-подобном домене этих ML. Однако при сравнении выведенных первичных аминокислотных последовательностей человеческого и мышиного ML было обнаружено, что мышиная последовательность имеет делецию в четыре пептида, расположенную между чел. остатками 111-114, и соответствующую делецию в 12 нуклеотидов, расположенной за нуклеотидом в положении 618 и наблюдаемой в обеих как челов. (см. выше), так и свиной (см. ниже) кДНК. В связи с этим для обнаружения возможных мышиных ML-изоформ были исследованы дополнительные клоны. Один клон "14" имел 1,4 кb-вставку, соответствующую выведенной аминокислотной последовательности из 335 аминокислот, в которой отсутствовал тетрапептид LPLQ. Эта форма, которая, очевидно, является полноразмерным мышиным ML, была обозначена mML или mМL335. Нуклеотидная и выведенная аминокислотная последовательности для ML представлены на фиг.16 (SEQ ID NOS 12 и 13). И наконец, был выделен и секвенирован клон "12". Этот клон имел делецию в 116 нуклеотидов, соответствующую чел.МL3, а поэтому он был обозначен ML-3. Сравнение выведенных аминокислотных последовательностей этих двух изоформ показано на фиг.16.
Экспрессия рекомбинантных mML.
Экспрессирующие векторы для мышиного ML получали в основном, как описано в Примере 8. Клоны, кодирующие mMl и mML-2, субклонировали a pRK5-tkneo, экспрессирующий вектор млекопитающего, который обеспечивает экспрессию под контролем промотора CMV и сигнала полиаденилирования SV40. Полученные экспрессирующие векторы. MMlpRKtkneo и mML2pRKtkneo были кратковременно трансфицированы в клетки 293 методом с использование фосфата кальция. После кратковременной трансфекции среды кондиционировали в течение пяти дней. Клетки поддерживали в среде DMEM с высоким содержанием глюкозы и с добавленной 10%-ной фетальной телячьей сывороткой.
Экспрессия мышиного mpl (mmpl) в клетках Ba/F3.
Стабильные клеточные линии, экспрессирующие с-mpl, бьши получены путем трансфекции mmpl-pRKtkneo в основном так же, как описано в Примере 1 для человеческого mpl. Для этого экспрессирующий вектор (20 мкг, линеаризованный), содержащий полную последовательность, кодирующую mpl (Skoda R.C. et al., EMBO J. 12: 2645-2653 (1993)), трансфицировали в клетки Ba/F3 путем электропорации (5×106 клеток, 250 Вольт, 960 мкФ) с последующим отбором резистентности к неомицину с использованием 2 мг/мл G418. Экспрессию mpl оценивали посредством проточной цитометрии с использованием антисыворотки против мышиных mpl-IgG. Клетки Ba/F3 поддерживали в среде RPM1 1640 от клеток WEH1-3B, используемых в качестве источника 1L-3. Супернатанты от клеток 293, кратковременно трансфицированных обоими mML и mMl-2, анализировали в ВаF3-клетках, трансфицированных обоими mmpl и hrnpl, как описано в Примере 1.
Пример 13
кДНК свиного лиганда mpl, pML и рМ1-2
кДНК свиного ML (pML) выделяли с помощью PACE-PCR. Для этого были сконструированы olioo dT-праймер и 2 специфических праймера исходя из последовательности экзона гена свиного ML, кодирующего амино-конец ML и выделенного из сыворотки свиньи с аплазией. Затем из различных тканей апластических свиней получали и амплифицировали кДНК. PCR-продукт кДНК, состоящий из 1342 п.о. и обнаруженный в ткани почки, был субклонирован. Несколько клонов было секвенировано, в результате чего было обнаружено, что они кодируют зрелый свиной лиганд mpl (не включая полный сигнал секреции). Было установлено, что кДНК кодирует 332 аминокислоты зрелого белка (pML332), имеющего последовательность, показанную на фиг.18 (SEQ ID NOS: 9 и 6).
Метод
Выделение гена pML и кДНК.
Геномные клоны гена свиного ML были выделены путем скрининга геномной библиотеки свиньи в EMBL3 (Clontech Inc.) с pR45. Библиотеку скринировали методом, в основном описанном в Примере 7. Несколько клонов было выделено и экзон, кодирующий аминокислотную последовательность, идентичную последовательности, полученной из очищенного ML, секвенировали. КДНК свиного ML получали в соответствии с модифицированной схемой для PAGE-PCR. Исходя из последовательности свиного гена ML были сконструированы два ML-специфических праймера. Полиаденилированную МРНК выделяли из почечной ткани апластических свиней, как описано выше. КДНК получали путем обратной транскрипции с использованием BamdT-праймера:
(BamdT: 5’ GACTCGAGGATCCATCGATTTTTTTTTTTTTTTTT 3’
(Посл. NO: 55),
направленного против полиаденозин-хвоста мРНК. Начальную серию PCR-амплификации (28 циклов в течение 60 сек при 95°С, в течение 60 сек при 58°С и в течение 90 сек при 72°С) проводили с использованием Мг-специфического прямого праймера
(h-forward-1: 5’ GCTAGCTCTAGAAATTGCTCCTCGTGGTCATGCTTCT 3’)
(Посл. NO: 43)
и праймера BAMAD
(BAMAD: 5’ GACTCGAGGATCCATCG 3’)
(Посл. NO: 56)
в 100 мл реакционной смеси (50 мМ КСl, 1,5 VgCl, 10 мМ Трис, рН 8,0, 0,2 мМ dNTPs и 0,05 ед./мл полимеразы Perkin (Elmer Amplitaqst)). Затем PCR-продукт гидролизовали ферментом Clal, экстрагировали фенолом-хлороформом (1:1), осаждали этанолом и лигировали в вектор Bluescript (0,1 мг) (Stratagene Inc.), который был разрезан ферментами Clal и Kpnl. После 2-часового инкубирования при комнатной температуре одну четвертую смеси для лигирования добавляли непосредственно во вторую серию PCR (22 цикла, как описано выше), проводимую с использованием второго ML-специфического праймера forward-1.
(forward-1: 5’ GCTAGCTCTAGAAGCCCGGCTCCTCCTGCCTG 3’
(Посл. NO: 57)
и Т3-21 (олигонуклеотид, который связывается с последовательностью, смежной с областью множественного клонироания в векторе Bluescript SK-):
(5’ CGAAATTAACCCTCACTAAAG 3’
(Посл. NO: 58)
Полученный PCR-продукт гидролизовали ферментами Xbal и Clal и субклонировали в Bluescript SK-. Несколько клонов от независимых PCR-реакцией секвенировали.
И снова идентифицировали вторую форму (обозначенную pML-2), кодирующую белок с делецией в 4 аминокислотных остатка (328 аминокислотных остатков) (см. фиг.21 (SEQ ID NO: 21)). Сравнение аминокислотных последовательностей pML и pML-2 показало, что эти последовательности являются идентичными за исключением того, что в форме pML-2 отсутствует тетрапептид QLPP, соответствующий остаткам 111-114, которые были делетитрованы (см. фиг.22 (SEQ ID NOS: 18 и 21)). Таким образом, делеции из 4 аминокислот наблюдались в мышиной, человеческой и свиной кДНК, причем в тех же самых положениях предсказанных белков.
Пример 14
Анализ клеток СМК на ТРО (тромбопоэтин) - индуцирование экспрессии тромбоцитарного антигена GPIIbIIIa
Клетки СМК поддерживали в среде RMPI 1640 (Sigma), в которую были добавлены 10%-ная фетальная коровья сыворотка и 10 мМ глутамина. Для получения препаратов для анализа клетки собирали, промывали и ресуспендировали при плотности 5×105 кл./мл в бессывороточной среде GIF, в которую было добавлено 5 мг/л коровьего инсулина, 10 мг/л апо-трансферрина, 1х микроэлементов. Затем в каждую лунку плоскодонного 96-луночного планшета добавляли стандартный ТРО или экспериментальные образцы при соответствующем разведении в 100 мкл объемах. После этого, в каждую лунку добавляли 100 мкл СМК-клеточной суспензии и планшеты инкубировали в течение 48 часов в инкубаторе при 37°С в атмосфере 5% CO2. После инкубирования планшеты подвергали центрифугированию при 1000 об/мин при 4°С в течение 5 минут. Супернатанты отбрасывали и в каждую лунку, добавляли FITC-конъюгированное (FITC-флюоресцеинизотиоцианат) моноклональное антитело 2D2 к GPIIbIIIa. После 1-часового инкубирования при 4°С планшеты снова центрифугировали при 100 об/мин в течение 5 минут. Супернатанты, содержащие несвязанные антитела, отбрасывали и в каждую лунку добавляли 200 мкл 0,1% BSA-PBS-промывки. Стадию промывки 0,1% BSA-PBS повторяли три раза. Затем клетки анализировали FASCAN с использованием стандартного однопараметрического анализа с измерением относительной интенсивности флюоресценции.
Пример 15
Анализ клеток DAMI на тромбопоэтин (ТРО) путем измерения эндомитотической активности этих клеток на 96-луночных планшетах для микротитрования
Клетки DAMI поддерживали в среде IMDM + 10% лошадиной сыворотки (Gibco) с добавлением в эту среду 10 мМ глутамина, 100 нг/мл пенициллина G и 50 мкг/мл стрептомицина. Для получения препаратов для анализа клетки собирали. Промывали и ресуспендировали (при плотности 1×106 кл./мл) в IMDM + 1% лошадиной сыворотки. К DAMI-клеточной суспензии в 96-луночном круглодонном планшете добавляли 100 мкл ТРО-стандарта или экспериментальных образцов. Затем клетки инкубировали в течение 48 часов при 37°С в инкубаторе в присутствии 5% СО2. После инкубирования планшеты подвергали центрифугированию на центрифуге Sorvall 6000B при 1000 об/мин в течение 5 минут при 4°С. Супернатанты отбрасывали и повторяли стадию промывки 200 микролитрами PBS-0,1% BSA. Затем клетки фиксировали путем добавления 200 мкл охлажденного льдом 70% этанола-РВ и ресуспендировали путем аспирации. После 15 минут инкубирования при 4°С планшеты центрифугировали при 2000 об/мин в течение 5 минут и в каждую лунку добавляли 150 мкл 1 мг/мл РНКазы, содержащие 0,1 мг/мл пропидийиодида и 0,05% Твина 20. После 1-часового инкубирования при 37°С оценивали изменения в составе ДНК с помощью проточной цитометрии. Количественное измерение полиплоидии проводили по следующему уравнению
Нормализованный коэффициент полиплоидии (NPR)=
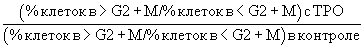
Пример 16
Тромбопоэтин (ТРО) в in vivo-анализе (анализ на повторное связывание мышиных тромбоцитов)
In vivo-анализ для определения (35S) продуцирования тромбоцитов
Мышам C57BL6 (полученным из Charies River) инъецировали внутрибрюшинно (IP) 1 мл козьей антисыворотки против мышиных тромбоцитов (6 амп.) (на день 1) для продуцирования тромбоцитопении. На дни 5 и 6 мышам вводили две внутрибрюшинные инъекции фактора или PBS для контроля. На день 7 мышам вводили внутрибрюшинно инъекцию 30 мкКи Na
Пример 17
KIRA-ELISА-анализ на тромбопоэтин, проводимый путем измерения фосфорилирования mpl-Rse.gD-химерного рецептора
Рецептор чел. mрl был описан Vigon и др. (PNAS, USA, 89: 5640-5644 91992)). Для использования в KIRA-ELISA-анализе, описанном в настоящей заявке, конструировали химерный рецептор, содержащий внеклеточный домен (ECD) рецептора mpl и трансмембранный (ТМ) и внутриклеточный (ICD) домены Rse (Mark et al., J. of Biol. Chem. 269(14): 10720-10728 (1994)). С карбокси-концевым флагеллярным полипептидом (т.е. Rse.gD). См. фиг.30 и 31, представленные для схематического описания анализа.
(a) Получение захватывающего агента
Моноклональное антитело к gD (клон 5В6) было продуцировано против пептида, гликопротеина D, вируса простого герпеса (Paborsky et al., Protein Engineering 3(6): 547-553 [1990]). Очищенный исходный препарат доводили до концентрации 3,0 мг/мл в забуференном фосфатом физиологическом растворе (PBS), рН 7,4 и 1/0 мл-аликвоты хранили при -20°С.
(b) Получение антитела против фосфотирозина
Моноклональное антитело против фосфотирозина (клон 4G10) закупали у UBI (Lake Placid NY) и биотинилировали с использованием высокоразветвленного биотин-N-гидроксисукцинамида (Biotin-X-NHS, Research Organics, Cleveland, ОН)
(с) Лиганд
Лиганд mpl получали с использованием техники рекомбинантных ДНК, описанной в настоящей заявке. Очищенный лиганд mpl хранили в виде маточного раствора при 4°С.
(d) Получение нуклеиновой кислоты Rse.gD
Синтетические двухцепочечные олигонуклеотиды были использованы для реконстрирования последовательности, кодирующей С-концевые 10 аминокислот (880-890) чел-Rse с добавлением еще 21 аминокислоты, содержащих эпитоп для антитела 5В6 и стоп-кодон. В Таблице 10 представлена конечная последовательность синтезированной части гибридного гена

Синтетические ДНК лигировали с кДНК, кодирующей аминокислоты 1-880 чел-Rse, в Pstl-сайте, начинающегося с нуклеотида 2644, опубликованной кДНК-последовательности чел-Rse (Mark et al. Journal of Biological Chem. 269(14): 10720-10728, и Hindl I-сайты в полилинкере экспрессирующего вектора pSV17.ID.LL (см. фиг.32 A-L; SEQ ID NO: 22), для создания экспрессирующей плазмиды pSV17.ID.Rse.gD. Короче говоря, экспрессирующая плазмида включает в себя дицистронный первичный транскрипт, который содержит последовательность, кодирующую DHFR и ограниченную 5'-сайтом сплайсинта (донорным сайтом) и 3'-сайтом сплайсинга (акцепторным сайтом), а за этой последовательностью находится последовательность, которая кодирует Rse.gD. Полноразмерный (несплайсированный) транскрипт содержит DHFR в качестве первой открытой рамки считывания, а поэтому продуцирует белок DHFR, что позволяет проводить отбор стабильных трансформантов.
(е) Получение нуклеиновой кислоты mpl-Rse.gD
Экспрессирующую плазмиду pSV.ID.Rse.gD., продуцированную как описано выше, модифицировали в целях получения плазмиды pSV. ID.M.tmRd.6, которая содержит кодирующую последовательность ECD чел.mрl (аминокислоты 1-491), соединенную с трансмембранным доменом и внутриклеточным доменом Rse.gD (аминокислоты 429-911). Для присоединения кодирующей последовательности части внеклеточного домена чел.mрl к кодирующей последовательности части Rse проводили двухстрадийную реакцию клонирования посредством PCR с использованием синтетических олигонуклеотидов, как описано Mark и др. (J. Biol. Chem. 267: 26166-26171 91992)). Праймерами, использованными в первой PCR-реакции, были: M1
(5’ TCTCGCTACCGTTTACAG 3’)
(Посл. NO: 61)
и М2
(5’ CAGGTACCCACCAGGCGGTCTCGGT 3’)
(Посл. NO: 62)
с mpl-кДНК-матрицей; и R1
(5’ GGGCCATGACACTGTCAA 3’)
(Посл. NO: 63)
и R2
(5’ GACCGCCACCGAGACCGCCTGGTGGGTACCTGTGGTCCTT 3’)
(Посл. NO: 64)
с Rse-кДНК-матрицей. Pvull-Smal-фрагмент этого участка соединения использовали для конструирования полноразмерного химерного рецептора.
(f) Трансформация клеток
Плазмиду pSV. ID.M. tmRd.6, которая была линеаризована в уникальном Notl-сайте в плазмидном остове, вводили в клетки DP12.CHO (ЕР 307247, опубликованный 15 марта 1989) путем электропорации. После экстракции фенолом/хлороформом ДНК осаждали этанолом и ресуспендировали в 20 мкл 1/10 Трис-ЭДТК. Затем 10 мкг ДНК инкубировали с 107 клетками СНО Р12 в 1 мл PBS на льду в течение 10 минут, после чего подвергали электропорации при 400 Вольт и 330 мкФ. Клетки возвращали на 10 минут на лед, а затем высевали на неселективную среду. Через 24 часа, клетки переносили на среду, не содержащую нуклеозид, для отбора на стабильные DHFR+-клоны.
(g) Отбор трансформированных клеток для использования в KIRA-ELISA
Клоны, экспрессирующие MPL/Rse.gD, идентифицировали путем Вестерн-блоттинга цельных клеточных лизатов после фракционирования с помощью ДСН-ПААГ с использованием антитела 5В6, которое обнаруживает метку эпитопа gD.
(h) Среды
Клетки культивировали в F12/DMEM (Gibco/BPL, Life Technologies, Grand Island, NY). В эти среды добавляли 10% диафильтрованного FBS (HyClone, Logan, Utah), 25 мМ HEPES и 2 мМ L-глутамина.
(i) KIRA-ELISA
Mpl-Rse.gD-трансформированные клетки DP12.CHO высевали в лунки (3×104 клеток на лунку) плоскодонного 96-луночного планшета в 100 мкл среды и культивировали в течение ночи при 37°С в 5% СO2. На следующее утро, супернатанты декантировали, а планшеты слегка промокали бумажной салфеткой. Затем в каждую лунку добавляли 50 мкл среды, содержащей либо экспериментальные образцы, либо 200, 50, 12,5, 3,12, 0,78, 0,19, 0,048 или 0 нг/мл лиганда mpl. Клетки стимулировали в течение 30 минут при 37°С, супернатанты декантировали, а планшеты еще раз слегка промокали бумажной салфеткой. Для лизиса клеток и солюбилизации химерных рецепторов в каждую лунку добавляли 100 мкл буфера для лизиса. Буфер для лизиса состоял из 150 мМ NaCl, содержащего 50 мМ HEPES (Gibco), 0/5% Тритон-Х (Gibco), 0,01% тимерозал, 30 КИЕ/мл апротинина (ICN Biochemicals, Aurora, ОН), 1 мМ 4-(2-аминоэтил)-бензолсульфонилфторида гидрохлорида (AEBSF; ICN Biochemicals), 50 мкМ лейпептина (ICN Biochemicals) и 2 мМ ортованадата натрия (Na3VO4; Sigma Chemicals Co, St. Lous, MO), pH 7,5. Затем планшеты слегка размешивали на шейкере для планшетов (Belico, Instruments, Vineland, NJ) в течение 60 минут при комнатной температуре.
Когда клетки были солюбилизированы, ELISA-микропланшеты (Nunc Maxis, Inter Med, Denmark), сенсибилизированные в течение ночи при 4°С моноклональным антителом 5В6 против gD (5,0 мкг/мл в 50 мМ карбонатного буфера, рН 9,6, 100 мкл/лунка), декантировали, промокали бумажной салфеткой и блокировали 150 мкл/лунка блокирующим буфером (PBS, содержащим 0,5% BSA (Intergen Company, Purchase, NJ) и 0,01% тимерозал) в течение 60 минут при комнатной температуре с легким помешиванием. Через 60 минут анти-gD5В6-сенсибилизированные планшеты промывали 6 раз промывочным буфером (PBS, содержащим 0,05% Твин-20 и 0,01% тимерозал) с использованием автоматического устройства планшетов (ScanWasher 300, Skatron Insrtuments, Inc., Sterling, VA).
Лизат, содержащий солюбилизированный MPL/Rse.gD, переносили из культурального планшета для микротитрования в анти-gD 5В6-сенсибилизированные и блокированные ELISA-лунки (85 мкл/лунка) и инкубировали в течение 2 часов при комнатной температуре, слегка помешивая. Несвязанные mpl-Rse.gD удаляли путем промывания промывочным буфером и в каждую лунку добавляли 100 мкл биотинилированных 4G10 (против фосфотирозина), разведенных 1:18000 в буфере для разведения (PBS, содержащем 0,5% BSA, 0,05% Твин-20, 5ММ EDTA и 0,01% тимерозал), т.е. 56 нг/мл. После 2-часового инкубирования при комнатной температуре планшеты промывали и в каждую лунку добавляли 100 мкл конъюгированного с пероксидазой хрена (НРРО) стрептавидина (Zymed Laboratores, S. San Francisco, CA), разведенного в соотношении 1:60000 в буфере для разведения. Планшеты инкубировали в течение 30 минут при комнатной температуре, слегка помешивая при этом. Свободный авидиновый конъюгат отмывали и в каждую лунку добавляли 100 мкл свежеприготовленного субстратного раствора (тетраметилбензидин) (ТМВ); 2-компонентный субстратный набор; Kirkegaard & Perry, Gaithersburg, MD). Смесь оставляли на 10 минут для протекания реакции, после чего развитие окраски прекращали путем добавления 100 мкл/лунка 1,0 М Н3РО4. Оптическую плотность при 450 нм определяли по сравнению с оптической плотностью при 650 нм (АВ450/650) с использованием планшет-ридера (vmax) (Molecular Devices, palo Alto, CA), контролируемого Macintosh Centris 650 (Apple Computers, Cupertino, CA) и программным обеспечением Deltasoft (Biometallics, Inc., Princeton, NJ).
Стандартную кривую строили путем стимулирования клеток dp12.trkA,B или C.gD 200, 50, 12,5, 0,78, 0,19, 0,048 или 0 нг/мл лиганда mpl и представляли (с использованием программы Deltasoft) в виде графика зависимости ТРО (нг/мл) от (АВ450/650)±ср. отклонение. Концентрации образца получали путем интерполяции их оптической плотности на стандартной кривой и выражали в виде активности ТРО (нг/мл).
Было установлено, что mpl-лиганд способен активировать химерный рецептор mpl-Rse.gD лиганд-специфическим способом в зависимости от концентрации. Кроме того, анализ KIRA-ELISA на mpl-Rse.gD, как было установлено, пригоден почти для 100% чел.сыворотки (показано) или 100% плазмы (не показано), что позволяет легко проводить скрининг пациента и рК-образцов.
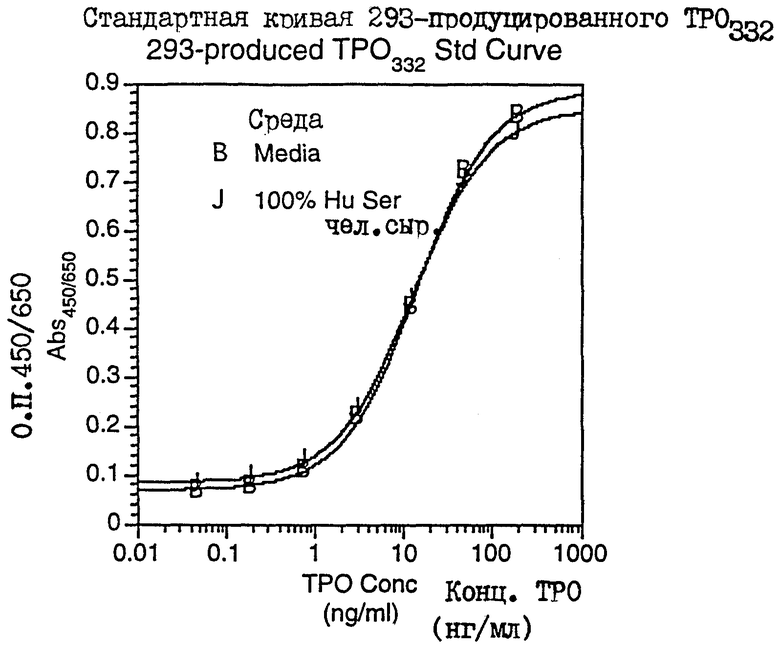
Суммарное значение ЕС50 для ТРО
Summary of TPO EC50’s
Пример 18
ELISA-анализ, проведенный с использованием рецептора для тромбопоэтина (ТРО)
ELISA-планшеты сенсибилизировали F(ab’)2-фрагментами кроличьего антитела против человеческого IgG (Fc) в карбонатном буфере (рН 9,6) при температуре 4°C в течение ночи. Затем планшеты блокировали 0,5% альбумином бычьей сыворотки в PBS при комнатной температуре в течение одного часа. Сбор, полученный из ферментера и содержащий химерный рецептор mpl-IgG, был добавлен в планшеты, после чего полученную смесь инкубировали в течение 2 часов. Двукратные серийные разведения (0,39-25 нг/мл) стандарта (ТРО332, продуцированного в клетках 293 с использованием концентрации, определенной путем количественного аминокислотного анализа) и серийно разведенные образцы в 0,5%-ном альбумине бычьей сыворотки и 0,05% Твине 20 добавляли в планшеты и полученную смесь инкубировали в течение 2 часов. Связанный ТРО детектировали с помощью очищенных белком А биотинилированных кроличьих антител к TPO155, которые были продуцированы в E.coli (время инкубирования = 1 час), а затем с помощью стрептавидин-пероксидазы (время инкубирования = 30 минут) и 3,3`,5,5`-тетраметилбензидина, используемых в качестве субстрата. Оптическую плотность считывали при 450 нм. Между стадиями планшеты промывали. Для данных анализа стандартную кривую строили с использованием программы Kaleidagraph для построения кривой с четырьмя параметрами. Концентрации образцов вычисляли с помощью стандартной кривой.
Пример 19
Экспрессия и очистка ТРО из клеток 293
1. Получение экспрессирующих векторов клеток 293
кДНК, соответствующую полной открытой рамке считывания ТРО, получали с помощью полимеразной цепной реакции, используя в качестве праймеров соответствующие олигонуклеотиды (табл.11).
Таблица 11
PCR-праймеры 293
PRKS-hrnpl I (описанную в Примере 9) использовали в качестве матрицы для реакции в присутствии готовой к употреблению (Stratagene) ДНК-полимеразы. Первоначальную денатурацию осуществляли в течение 7 минут при температуре 94°С, а затем с помощью 25 циклов амплификации (в течение 1 минуты при 94°С, 1 минуты при 55°С и 1 минуты при 72°С). Конечное удлинение проводили в течение 15 минут при температуре 72°С. PCR-продукт очищали и клонировали между рестрикционными сайтами Clal и Xbal плазмиды pRK5tkneo, а вектор, происходящий от pRK5, модифицировали так, чтобы он экспрессировал ген резистентности к неомицину под контролем тимидинкиназного промотора с получением открытой рамки считывания вектора pRK5tkneo.ORF. Вторую конструкцию, соответствующую гомологичному домену еро, получали тем же самым способом, но с использованием Cla.FL.F в качестве прямого праймера и соответствующего обратного праймера:
Arg.STOP.Xba: 5’ TCT AGA TCT AGA TCA CCT GAC GCA GAG GGT GGA СС 3’
(Посл.NO: 66)
Конечную конструкцию обозначали pRKS-tkneoEPO-D. Последовательность обеих конструкций проверяли способом, описанным в Примере 7.
2. Трансфекция эмбриональных клеток почек человека
Эти 2 конструкции трансфицировали в эмбриональные клетки почек человека посредством метода с использованием СаРO4, как описано в Примере 9. Через 24 часа после трансфекции отбор клонов, резистентных к неомицину, инициировали в присутствии 0,4 мг/мл G418,10 и через 15 дней отдельные колонии переносили в 96-луночные планшеты, а затем оставляли для культивирования до состояния сплошности. Экспрессию ML153 или ML332 в кондиционированной среде от этих клонов анализировали с помощью анализа на пролиферацию Ba/F3-mpl (как описано в Примере 1).
3. Очистка rhML332.
293-rhМL332-кондиционированную среду наносили на колонку с Blue-Sepharose (Pharmacia), которую уравновешивали в 10 мМ фосфата натрия при рН 7,4 (буфер А). После этого колонку промывали 10 колоночными объемами 2М буфера А и 2М буфера В. Затем колонку элюировали буфером А, содержащим 2М NaCl и 1М NaCl. Сбор после элюирования Blue-Sepharose сразу наносили на колонку с WGA-Sepharose, уравновешенную в буфере А. Колонку с WGA-Sepharose промывали промывали 10 колоночными объемами 2М буфера А, содержащего 2М NaCl и 1М NaCl, а затем элюировали тем же буфером, содержащим 0,5М N-ацетил-D-глюкозамин. Элюат WGA-Sepharose наносили на ВРЖХ-колонку с С4 (Synchrom, Inc.), уравновешенную в 0,1% трифторуксусной кислоте. ВРЖХ-колонку с С4 элюировали ступенчатым пропаноловым градиентом (0-25, 25-35 и 35-70%). Выделенный rhML332 подвергали элюированию в 28-30% пропаноловом градиенте. Очищенный с помощью электрофореза в ПААГ с ДСН rhМL332 перемещался в виде широкой полосы в 68-80 кДа-области геля (см.фиг.15).
4. Очистка rhML153
293-rhМL153-кондиционированную среду подвергали разделению на колонке Biue-Sepharose, как описано в методе для rhМL332. Элюат Blue-Sepharose сразу наносили на mpl-аффинную колонку, как описано выше. RhML153, проэлюированный на mpl-аффинной колонке, очищали до получения гомогенной массы с использованием ВРЖХ-колонки с С4 при тех же самых условиях, описанных для rhМL332. RhML153, очищенный с помощью электрофореза в ПААГ с ДСН, разделяли на 2 мажорных и 2 минорных полосы с массой (Mr), равной около 18000-21000 (см. фиг.15).
Пример 20
Экспрессия и очистка ТРО из СНО
1. Очистка экспрессируюцих векторов СНО
Сконструированные экспрессирующие векторы, используемые в электропорации, описаны ниже:
pSVI5. ID.LL.MLORF (полноразмерная или hТРО332), и pSVl5.ID.LL.MLEPO-D (усеченная или hТРО153). Соответствующие отличительные особенности этих плазмид представлены на фиг.23 и 24.
2. Получение экспрессируюцих векторов СНО
кДНК, соответствующую полной открытой рамке считывания hTPO, получали с помощью полимеразной цепной реакции с использованием олигонуклеотидных праймеров, указанных в Таблице 12.
Таблица 12
PCR-праймеры для эрепрессирующего вектора в СНО
PRKS-hmpI I (описанную в Примере 7 и 9) использовали в качестве матрицы для реакции в присутствии готовой к употреблению (Stratagene) ДНК-полимеразы. Первоначальную денатурацию осуществляли в течение 7 минут при 94°С, а затем с использованием 25 циклов амплификации (в течение 1 минуты при 94°С, 1 минуты при 55°С и 1 минуты при 72°С). Конечное удлинение проводили в течение 15 минут при температуре 72°С. PCR-продукт очищали и клонировали между рестрикционными сайтами Clal и Sall плазмиды pSVI5.ID.LL с получением вектора pSVI5.ID.LL.MLORF. Вторую конструкцию, соответствующую домену ЕРО, получали тем же самым способом, но с использованием Cla.FL.F2 в качестве прямого праймера и обратного праймера, указанного ниже:
EPOD.Sal 5’ AGT CGA CGT CGA CTC ACC TGA CGC AGA GGG TGG ACC 3’ (Посл. NO: 68)
Конечную конструкцию обозначали pSVI5.ID.LL.MLEPO-D. Последовательность обеих конструкций проверяли, как описано в Примере 7 и 9.
В основном кодирующие последовательности для полноразмерного и усеченного лиганда вводили в сайт множественного клонирования экспрессирующего вектора pSVI5.ID.LL клеток CHO. Этот вектор содержал область раннего промотора/энхансера вируса SV40, модифицированный сайт сплайсинга, содержащий мышиную кДНК DHFR; сайт множественного клонирования для введения гена нужного (описанные в этом случае последовательности ТРО) сигнала полиаденилирования SV40; сайт инициации репликации и ген бета-лактамазы для отбора плазмиды и ее амплификации в бактерии.
3. Методика получения стабильных клеточных линий CHO, экспрессируюцих рекомбинантные ТРО332 и TPO153 человека
а. Описание родственных клеточных линий CHO
Хозяйские клеточные линии CHO (яичник китайского хомячка), используемые для экспрессии молекул ТРО, описаны в настоящем изобретении как CHO-DP12 (см. ЕР 307247, опубликованный 15 марта 1989 г). Клеточную линию млекопитающего клонально отбирали в результате трансфекции родственной линии (CHO-KI DUX-BII (DHFR)-, полученной от Dr.Frank Lee Станфордского университета с разрешения Dr. L.Chaisi) вектором, экспрессирующим препроинсулин, с получением клонов с пониженными требованиями инсулина. Эти клетки также являлись отрицательными по DHFR, а клоны могут быть отобраны на присутствие последовательностей DHFR-кДНК-вектора путем культивирования на среде, не содержащей нуклеотидных добавок (глицина, гипоксантина и тимидина). В основном была использована система селекции стабильной экспрессии клеточных линий CHO.
b. Метод трансфекции (электроперация)
Клеточные линии, экспрессирующие ТРО332 и TPO153, получали путем трансфекции клеток DP12 путем электропорации (см., например, Adreason, G.L. J.Tiss. Cult.Meth., 1556 (1993)) с помощью линеаризованных плазмид pSVI5.ID.LL.MLORF или pSVI5.ID.LL.MLEPO-D соответственно. Три реакционных смеси рестриктирующих ферментов брали для каждого разрезания плазмиды; 10, 25 и 50 мкг вектора гидролизовали ферментом NOTI стандартными методами, используемыми в молекулярной биологии. Лишь один рестрикционный сайт находился в векторе в области линеаризации 3 и в области, находящейся вне области транскрипции лиганда ТРО (см. фиг.23). Инкубирование 100 мкл реакционной смеси осуществляли в течение ночи при температуре 37°С. На следующий день смесь один раз экстрагировали фенолом/хлороформом/изоамиловым спиртом (50:49:1), а затем осаждали этанолом, помещая на сухой лед. Процедуру проводили приблизительно в течение одного часа. Образовавшийся осадок собирали путем 150-минутного микроцентрифугирования, а затем осушали. Лианеризованную ДНК ресуспендировали в 50 мкл среды DMEM-F12 (1:1) Хема, содержащей стандартные антибиотики и 2 мМ глутамин.
Клетки DP112, выращенные в суспензионной среде, собирали, один раз промывали в той среде, что была использована для ресуспендирования ДНК, и наконец, ресуспендировали в этой среде при концентрации 107 клеток на 750 мкл. Аликвоты клеток (750 мкл) и каждую лианеризованную ДНк-смесь вместе инкубировали при комнатной температуре в течение одного часа, а затем переносили в камеру BRL для элетропорации. Каждую реакционную смесь подвергали элетропорации в стандартном устройстве BRL для элетропорации при 350 В, 330 мкФ и низкой емкости. После проведения элетропорации клетки оставляли в этом устройстве на 5 минут, а затем помещали на лед и осуществляли дополнительное инкубирование в течение 10 минут. Затем клетки, обработанные путем элетропорации, переносили в 60-мм чашки для культивирования клеток, содержащие 5 мл стандартной полной ростовой среды для клеток СНО (высокомолекулярная глюкоза; DMEM-F12 (50:50), не содержащая глицина, с добавлением 1х GHT, 2 мМ глутамина и 5%-ную бычью фетальную сыворотку), после чего культивировали в течение ночи в инкубаторе для культивирования клеток в присутствии 5% СO2.
С. Метод селекции и скрининга
На следующий день клетки отделяли от планшетов с помощью трипсина стандартными способами, а затем переносили в 150-мм планшеты с клеточными культурами, содержащие DHFR-селективную среду (среда DMEM-F12 (1:1) Хема, описанную выше и содержащую 2%-ную или 5-ную диализованную фетальную бычью сыворотку, но не содержащую глицина, гипоксантина и тимидина; была использована стандартная DHFR-селективная среда). Затем клетки от каждого 60-мм планшета переносили в 5/150-мм планшеты. Полученные клетки инкубировали в течение 10-15 дней (с одной заменой среды) при температуре 37°С в присутствии 5% СO2 до тех пор, пока не были получены клоны, имеющие размеры, достаточные для их переноса в 96-луночные планшеты с использованием стерильных желтых наконечников на Pipettmann, установленной при 50 мл. Эти клетки оставляли для культивирования до состояния сплошности (обычно 3-5 дней), после чего планшеты трипсинизировали и 2 копии от первоначальных планшетов были снова продуцированы. Две этих копии в течение небольшого промежутка времени хранили в холодильнике, причем клетки в каждой лунке разводили 50 микролитрами 10% фетальной бычьей сыворотки в диметилсульфоксиде. Образцы 5-дневной кондиционированной бессывороточной среды анализировали исходя из конфлуэнтных лунок третьего планшета для экспрессии ТРО посредством анализа на активность клеток Ba/F. Наиболее высокоэкспрессирующие клоны, подтвержденные с помощью данного анализа, доводили еще раз до состояния сплошности в 150-мм Т-колбах, а затем переносили в группу клеточных культур для адаптации суспензии, повторного анализа и создания банка.
d. Амплификация
Несколько клеточных линий с наиболее высоким титром, отобранных способом, описанным выше, подвергали амплификации с использованием стандартного метотрексата с получением клонов с более высоким титром. Клоны клеток СНО размножали и высевали в 10-см планшеты при 4 концентрациях метотрексата (т.е. 50, 100, 200 и 400 нМ) при двух или трех количествах клеток (105,5×105 и 106 клеток на лунку). Затем культуры инкубировали при температуре 37°С в присутствии 5% CO2 и образованные клоны переносили в 96-луночные планшеты для последующего анализа. Несколько клонов с высокими титрами, отобранных в результате селекции, снова обрабатывали высокими концентрациями метотрексата (т.е. 600, 800, 1000 и 1200 нМ) и в соответствии со способом, описанным ранее, клоны, обладающие резистентностью, оставляли, а затем переносили в 96-луночные планшеты и анализировали.
4. Культивирование стабильных клеточных линий СНО, экспрессирукщих рекомбинантные ТРО332 и ТРО153 человека
Собранные клетки оттаивали и клеточную популяцию размножали с помощью стандартных методов культивирования клеток либо в бессывороточной среде, либо в среде содержащей сыворотку. После размножения до получения достаточной клеточной плотности клетки промывали для удаления культуральной среды. Клетки культивировали с помощью любого стандартного способа, включающего в себя: периодическое культивирование, культивирование с подпиткой или непрерывное культивирование при 25-40°С при нейтральном рН в присутствии растворенного О2, по крайней мере, при 5% до тех пор, пока не накапливался конститутивно секретируемый ТРО. Затем жидкую культуру отделяли от клеток путем механического перемешивания, например центрифугирования.
5. Очистка рекомбинантного ТРО человека из клеточной суспензии СНО
Собранную суспензионную клеточную культуру (HCCF) непосредственно наносили на колонку Fast Flow (Pharmacia) с Blue-Sepharose 6, уравновешенную 0,01 М фосфата натрия (рН 7/4) и 0,15 М хлорида натрия в отношении приблизительно 100 литров HCCF на 1 литр смолы и при линейной скорости потока приблизительно 300 мл/час/см2. Затем колонку промывали 3-5 колоночными объемами уравновешенного буфера, после чего 3-5 колоночными объемами 0,01 М фосфата натрия (рН 7,4) и 2,0 М мочевины. Затем ТРО элюировали 3-5 колоночными объемами 0,01 М фосфата натрия (рН 7,4), 2,0 М мочевины и 1,0 М хлорида натрия.
Пул Blue-Sepharose, содержащий ТРО, наносили на колонку с Сефарозой 6МВ и лектином из зерен пшеницы (Wheat Germ Lectin Sepharose 6MB, (Pharmacia), уравновешенную 0,01 М фосфата натрия (рН 7,4), 2,0 М мочевине и 1,0 М хлориде натрия в отношении 8-16 мл пула Wheat Germ Lec. На 1 мл смолы при скорости потока равной приблизительно 50 мл/час/см2. После этого колонку промывали 2-3 колоночными объемами уравновешенного буфера. Затем ТРО элюировали 2-5 колоночными объемами 0,01 М фосфата натрия при рН 7,4, 2,0 М мочевины и 0,5 М N-ацетил-D-глюкозамина.
Пул лектина зерен пшеницы доводили до конечной концентрации 0,04% С12Е8 и 0,1% трифторуксусной кислоты (TFA). Полученный пул наносили на обращенно-фазовую колонку с С4 (Vydac 214TP1012), уравновешенную в 0,1% трифторуксусной кислоте (TFA) и 0,04% С12Е8 при загрузке, равной приблизительно 0,2-0,5 мг белка на один миллиметр смолы, и при скорости потока, равной 157 мл/час/см2.
Белок элюировали в двухфазном линейном градиенте ацетонитрила, содержащего 0,1%-ную трифторуксусную кислоту и 0,04% C12E8. Первую фазу обрабатывали линейным градиентом 0-30% ацетонитрила в течение 15 минут, а вторую фазу обрабатывали линейным градиентом 30-60% ацетонитрила в течение 60 минут. ТРО элюировали приблизительно 50%-ным ацетонитрилом. Полученный пул подвергали электрофорезу в ПААГ с ДСН.
Пул С4 разводили двумя объемами 0,01 М фосфата натрия при рН 7,4 и 0,15 М хлорида натрия, а затем диафильтровали против приблизительно 6 объемов 0,01 М фосфата натрия (рН 7,4) и 0,15 М хлорида натрия на мембране для ультрафильтрации Amicor УМ или ей подобной мембране, имеющих молекулярные массы в 10000-30000 Да. Полученный диафильтрат может быть сразу обработан путем ультрафильтрации с последующим концентрированием. Смесь диафильтрата и концентрата доводили до конечной концентрации 0,01% Твин-80.
Всю эту смесь или ее часть с 2-5% вычисленным колоночным объемом наносили на колонку HP с Sephacryl S-300 (Pharmacia), уравновешенную в 0,01 М фосфате натрия (рН 7,4), 0,15 М хлориде натрия и 0,01% Твине-80, а затем хроматографировали на скорости потока приблизительно 17 мл/ч/см2. Фракции, содержащие ТРО и не содержащие продуктов агрегации и продуктов протеолитического расщепления, объединяли в пул с помощью электрофореза в ПААГ с ДСН. Полученный пул фильтровали через 0,22-мкл фильтр Millex-GY или ему подобный фильтр, а затем хранили при температуре 2-8°С.
Пример 21
Трансформация и индуцирование синтеза белков ТРО в E.coli 1. Конструирование векторов экспрессии ТРО в E.coli
Все плазмиды рМР21, рМР151, рМР41, рМР57 и рМР202 конструировали так, чтобы они экспрессировали первые 155 аминокислот ТРО, расположенных ниже от небольшой лидерной последовательности, которая расположена среди различных конструкций. Главным образом, лидерные последовательности обеспечивают высокий уровень инициации трансляции и быструю очистку. Плазмиды рМР210-1, -Т8, -21, -22, -24, -25 конструировали так, чтобы они экспрессировали первые 153 аминокислоты ТРО, расположенных ниже от инициирующего метионина и отличающиеся друг от друга только лишь использованием кодонов первых 6 аминокислот ТРО, в то время как плазмида рМР251 представляет собой производное рМР210-1, в котором карбокси-конец ТРО удлинен на две аминокислоты. Все вышеуказанные плазмиды продуцируют высокие уровни внутриклеточной экспрессии ТРО E.coli после индуцирования триптофанового промотора (Yansura, D.G. et al., Methods in Enzymology (Goeddel D.V. Ed) 185: 54-60, Academic Press, San Diego [1990]). Плазмиды pMPI и pMP172 являются промежуточными в конструкции вышеуказанных плазмид внутриклеточной экспрессии ТРО.
(а) Плазмида pMPI
Плазмида pMPI представляла собой секретирующий вектор для первых 155 аминокислот ТРО и была сконструирована путем лигирования всех 5 фрагментов ДНК, как показано на фиг.33. Первые из них являлись вектором pPho21, в котором небольшой Mlul-BamHl-фрагмент был удален. pPho21 представляет собой производное phGHl (Chano, C.N. et al., Gene 55: 189-196 [1987]), в котором ген гормона роста человека заменяли на ген phoA E.coli, а рестрикционный Mlul-сайт конструировали в к-последовательность, кодирующую сигнальную последовательность STll у аминокислот 20-21.
Следующие два фрагмента, Hinfl-Pstl-фрагмент (258 п.о.) ДНК от pRK5-hmpl 1 (Пример 9), кодирующий аминокислоты ТРО 19-103 и нижеследующую синтетическую ДНК, кодирующую аминокислоты 1-18:
5’CGCGTATGCCAGCCCGGCTCCTCCTGCTTGTGACCTCCGAGTCCTCAGTAAACTGCTTCGTG
ATACGGTCGGGCCGAGGAGGACGAACACTGGAGGCTCAGGAGTCATTTGACGAAGC ACTGA-5’
(Посл. NO: 69)
(Посл. NO: 70)
предварительно лигировали с использованием ДНК-лигазы Т4 и второй отрезок гидролизовали ферментом Pstl. Четвертый фрагмент представлял собой Pstl-Haelll-фрагмент (152 п.о.) от плазмиды pRKShmpll, кодирующей аминокислоты 104-155 ТРО. Последний являлся Stul-BamHl-фрагментом (412 п.о.) от плазмиды рро108, содержащей лямбда-терминатор транскрипции, как описано выше (Scholtissek, S. Et al., NAP 15: 3185 [1987]).
(b) Плазмида рМР21
Плазмиду рМР21 конструировали так, чтобы она экспрессировала первые 155 аминокислот ТРО с помощью лидерной последовательности из 13 аминокислот, содержащей часть сигнальной последовательности STll. Эту плазмиду конструировали путем лигирования всех трех ДНК-фрагментов, как показано на фиг.34, причем первый из них являлся вектором pVEG31, в котором небольшой Xbal-Sphl-фрагмент был удален. Вектор pVEG31 представлял собой производное pHGH207-l (de Boer, H.A. et al., in Promoter Structure and Function (Rodrigues, P.L. и Chamberlain, M.J., Eo), 462, Praege, New York [1982]), в котором ген гормона роста человека заменен геном васкулярного эндотелиального фактора роста (идентичный фрагмент вектора может быть получен от плазмиды pHGH207-1).
Вторая часть лигирования представляла собой синтетический ДНК-дуплекс, имеющий последовательность, указанную ниже:
5’-CTAGAATTATGAAAAAGAATATCGCATTTCTTCTTAA
TTAATACTTTTTCTTATAGCGTAAAGAAGAATTGCGC-5’
(Посл. NO: 71)
(Посл. NO: 72)
Последняя часть представляла собой Mlul-Sptl-фрагмент (1072 п.о.) от рМР1, кодирующий 155 аминокислот ТРО.
(c) Плазмида рМР151
Плазмиду рМР151 конструировали для экспрессии первых 155 аминокислот ТРО, расположенных ниже лидерной последовательности, содержащей 7 аминокислот сигнальной последовательности STll, 8 гистидиновых остатков и сайт расщепления фактор Ха. Как показано на фиг.35, рМР151 конструировали путем лигирования всех трех ДНК-фрагментов, первый из которых представлял собой вышеописанный вектор pVEG31, из которого был удален небольшой Xbal-Sptl-фрагмент. Вторая плазмида представляла собой синтетический ДНК-дуплекс, имеющий последовательность, указанную ниже:
5’-CTAGAATTATGAAAAAGAATATCGCATTTCATCACCATCACCATCACCATCACATCGAAG GTCGTAGCC
TTAATACTTTTTCTTATAGCGTAAAGTAGTGGTAGTGGTAGTGGTAGTGTAGCTTC CAGCAT-5’
(Посл. NO: 73)
(Посл. NO: 74)
Последняя плазмида представляла собой Boll-Sptl-фрагмент (1064 п.о.) от pMP11, кодирующий 154 аминокислоты ТРО. Плазмида pMP11 идентична плазмиде рМР1 за исключением того, что в сигнальной последовательности STll были заменены несколько кодонов (этот фрагмент может быть получен из рМР1).
(d) Плазмида рМР202
Плазмида рМР202 являлась аналогичной экспрессирующему вектору рМР151 за исключением того, что сайт расщепления фактором Ха в лидерной последовательности заменен сайтом расщепления тромбином. Как показано на фиг.36, рМР202 конструировали путем лигирования всех трех ДНК-фрагментов. Первый из них представлял собой вышеописанный вектор pVEG31, в котором был удален небольшой Xbal-Sptl-фрагмент. Второй представлял собой синтетический ДНК-дуплекс, имеющий последовательность, указанную ниже:
5’-CTAGAATTATGAAAAAGAATATCGCATTTCATCACCATCACCATCACCATCACATCGAA CCAACGTAGCC
TTAATACTTTTTCTTATAGCGTAAAGTAGTGGTAGTGGTAGTGGTAGTGTAGCTT GGTGCAT-5’
(Посл. NO: 75)
(Посл. NO: 76)
Последняя плазмида представляла собой Bgll-Sptl-фрагмент (1064 п.о.) плазмиды рМР11, описанной выше.
(e) Плазмида рМР172
Плазмида рМР172 является секретирующим вектором для первых 153 аминокислот ТРО и промежуточным звеном для конструирования рМР210. Как показано на фиг.37 рМР172 получали путем совместного лигирования трех ДНК-фрагментов, первый из которых являлся вектором pLS321amB, в котором был удален небольшой EcoRl-Hindlll-фрагмент. Второй представлял собой EcoRl-Hbal-фрагмент (946 п.о.) вышеописанной плазмиды рМР11. Последняя часть представляла собой синтетический ДНК-дуплекс, имеющий нижеследующую последовательность:
5`-TCCACCCTCTGCGTCAGGT (Посл. NO: 77)
GGAGACGCAGTCCATCGA-5’ (Посл. NO: 78)
(f) Плазмида рМР210
Плазмиду рМР210 конструировали для экспрессии первых 153 аминокислот ТРО после метионина, инициирующего трансляцию. Эту плазмиду использовали в виде банка плазмид, в котором первые 6 кодонов ТРО рандомизированы в третьем положении каждого кодона, а затем конструировали, как показано на фиг.38, путем лигирования всех трех ДНК-фрагментов. Первый фрагмент представлял собой ранее описанный вектор pLEG31, в котором был удален небольшой Xbal-Sptl-фрагмент. Второй представлял собой синтетический ДНК-дуплекс, который сначала обрабатывали ДНК-полимеразой 1 (фрагмент Кленова), а затем гидролизовали ферментами Xbal и Hinfl, и наконец, этот дуплекс, кодирующий метионин инициации, рандомизировали в первых шести кодонах ТРО.
5’-GCAGCAGTTCTAGAATTATGTCNCCNGCNCCNCCNGCNTGTGACCTCCGA
ACACTGGAGGCTGTTCTCAGTAAA (Посл. NO: 79) CAAGAGTCATTTGACGAAGCACTGAGGGTACAGGAAG-5’ (Посл. NO: 80)
Третий фрагмент представлял собой Hinfl-Sptl-фрагмент (890 п.о.) плазмиды рМР172, кодирующей аминокислоты 19-153 ТРО.
рМР210-плазмидный банк, приблизительно 3700 клонов, повторно трансформировали в LB-планшеты с высокомолекулярным тетрациклином (50 мкг/мл) для отбора клонов, инициирующих трансляцию (Yansura, D.G. et al., Methods: A Companion to Methods in Enzymology 4:151-158 [1992]). Из 8 колоний, платированных в планшеты с высокомолекулярным тетрациклином, 5 наиболее продуцирующих колоний в промежутке между экспрессией ТРО подвергали ДНК-секвенированию; результаты приведены на фиг.39 (SEQ ID NOS: 23, 24, 25, 26, 27 и 28)
(g) Плазмида рМР41
Плазмиду рМР41 конструировали для экспрессии первых 155 аминокислот ТРО, слитого с лидерной последовательностью, состоящей из 7 аминокислот сигнальной последовательности STll, за которым расположен сайт расщепления фактором Ха. Эту плазмиду конструировали как показано на фиг.40 путем глигирования всех трех частей ДНК, первая из которых представляла собой ранее описанный вектор pVEG31, в котором был удален небольшой Xbal-Sptl-фрагмент. Второй представлял собой следующий синтетический ДНК-дуплекс:
5` CTAGAATTATGAAAAAGAATATCGCATTTATCGAAGGTCGTAGCC (Посл. NO: 81)
TTAATACTTTTTCTTATAGCGTAAATAGCTTCCAGCAT-5’ (Посл. NO: 82)
Последний фрагмент в лигировании представлял собой Bgll-Sphl-фрагмент (1064 п.о.) ранее описанной плазмиды рМ11.
(h) Плазмида рМР57
Плазмида рМР57 экспрессирует первые 155 аминокислот ТРО, расположенные ниже лидерной последовательности, состоящей из 9 аминокислот сигнальной последовательности STll и двухосновного сайта Lys-Arg. Этот двухосновной сайт обеспечивает удаление лидерной последовательности посредством протеазо ArgC, Данную плазмиду конструировали, как показано на фиг.41 путем совместного лигирования трех ДНК-фрагментов. Первый из этих фрагментов представлял собой ранее описанный вектор pLEG31, в котором был удален небольшой Xbal-Sptl-фрагмент. Второй фрагмент представлял собой следующий синтетический ДНК-дуплекс:
5’ CTAGAATTATGAAAAAGAATATCGCATTTCTTCTTAAACGTAGCC (Посл. NO: 83)
TTAATACTTTTTCTTATAGCGTAAAGAAGAATTTGCAT-5’ (Посл. NO: 84)
Последний фрагмент после лигирования представлял собой Bgll-Sphl-фрагмент (1064 п.о.) ранее описанной плазмиды рМ11.
(i) Плазмида рМР251
Плазмида рМР251 представляла собой производное рМР210-1, где на карбокси-концах были расположены две дополнительные аминокислоты ТРО. Как показано на фиг.42, эту плазмиду конструировали путем лигирования двух ДНК, а первый фрагмент представлял собой ранее описанный вектор рМР21, в котором был удален небольшой Xbal-Apal-фрагмент. Второй фрагмент после лигирования представлял собой Xbal-Apal-фрагмент (316 п.о.) плазмиды рМР210-1.
2. Трансформация и индуцирование векторов экспрессии ТРО в E.coli
Вышеуказанные ТРО-экспрессирующие плазмиды использовали для трансформации штамма 44С6 E.coli (w3110 tоnАΔ rpoHts lоnΔсlрРΔ galE) с помощью метода теплового шока с использованием CaCl2 (Handel, et al., J.Mol.Biol., 53:159-162, [1970]). Трансформироованные клетки сначала культивировали при температуре 37°С в LB-среде, содержащей 50 мкг/мл карбенициллина, до тех пор пока оптическая плотность (600 нм) культуры не достигла приблизительно 2-3. Затем LB-культуру разводили 20×М9-среде, содержащей 0,49% казаминокислоты (мас./об.) и 50 мкг/мл карбенициллина. После 10-часового культивирования при температуре 30°С с аэрацией к этой культуре добавляли индол-3-акриловую кислоту до конечной концентрации 50 мкг/мл. После этого культуру выращивали еще 15 часов при температуре 30°С с аэрацией и в этот период времени клетки собирали посредством центрифугирования.
Пример 22
Продуцирование биологически активного ТРО (Met-1 1-153) в E.coli
Нижеприведенные процедуры для продуцирования биологически активного восстановленного ТРО (Met-1 1-153) могут быть осуществлены по аналогии с другими вариантами ТРО, включая N- и С-концевые удлиненные формы (см. Пример 23).
А. Восстановление нерастворимого ТРО (Met-1 1-153)
ТРО, экспрессирующий клетки E.coli (Met-1 1-153) и кодированный плазмидой рМР210-1, подвергали ферментации, как описано выше. Приблизительно 100 г клеток ресуспендировали в 1 литре (10 объемов) буфера для дезинтеграции клеток (10 мМ, 10 мМ Трис, 5 мМ ЭДТК (рН 8) с использованием гомогенизатора Polytron, а затем клетки центрифугировали при 5000 g в течение 30 минут. Промытый клеточный осадок снова ресуспендировали в 1 л буфера для дезинтеграции клеток с использованием гомогенизатора Polytron, а затем клеточную суспензию пропускали через дезинтегратор клеток LH (LH Inceltech, Inc.) или через псевдоожижитель Microfluidizer (Microfluidics International) в соответствии с инструкциями изготовителя. Полученную суспензию центрифугировали при 5000 g в течение 30 минут, а затем ресуспендировали и центрифугировали еще раз с получением промытого осадка рефрактивных телец. Промытый осадок использовали сразу или предварительно замораживали при -70°С.
В. Солюбилизация и очистка мономерного ТРО (Met-1 1-153)
Вышеуказанный осадок ресуспендировали в 5 объемах 20 мМ Трис (рН 8), содержащего 6-8 М гуанидин и 25 мМ DTT (дитиотреитол), затем перемешивали в течение 1-3 часов или в течение ночи при температуре 4°С с получением солюбилизированного белка ТРО. Были также использованы высокие концентрации (6-8 М) мочевины, однако в результате получали более низкий выход по сравнению с гуанидином. После солюбилизации раствор центрифугировали при 30000 g в течение 30 минут и получали прозрачный супернатант, содержащий денатурированный мономерный белок ТРО. После этого супернатант хроматографировали посредством гель-фильтрации на колонке Superdex 200 (Pharmacia, 2,6×60 см) при скорости потока 2 мл/минуту, а затем элюировали 20 мМ фосфата натрия (рН 6) и 10 мМ дитиотреитола (DTT). Фракции, содержащие мономерный денатурированный белок ТРО, элюировали (160-200 мл) и объединяли. Кроме того, белок ТРО очищали с помощью полупрепаративной обращенно-фазовой хроматографии на колонках С4 (2×20 см; Vydac). Образец наносили при 5 мл/мин на колонку, уравновешенную в 0,1% трифторуксусной кислоте (TFA), элюируя 30%-ным ацетонитрилом. После этого белок элюировали градиентом ацетонитрила (30-60% в течение 60 минут). Затем очищенный и восстановленный белок элюировали приблизительно 50%-ным ацетонитрилом. Полученный продукт использовали для повторной укладки с получением биологически активного варианта ТРО.
С. Продуцирование биологически активного ТРО (Met-1 1-153)
Приблизительно 20 мг мономерного, восстановленного и денатурированного белка ТРО в 40 мл смеси 0,1%-ной трифторуксусной кислоты и 50%-ного ацетонитрила разводили в 360 мл буфера для укладки, необязательно содержащего реагенты:
50 мМ Трис;
0,3 М хлорид натрия;
5 мМ ЭДТК;
Детергент 2% CHAPS;
25%-ный глицерин;
5 Мм окисленный глутатион;
1 мМ восстановленный глутатион;
рН доведенный 8,3.
После перемешивания буфер для укладки мягко помешивали при температуре 4°С 12-48 часов для получения максимального выхода правильных дисульфид-связанных форм ТРО (см. ниже). Полученный раствор подкисляли трифторуксусной кислотой до конечной концентрации 0,2%, а затем фильтровали через фильтр в 0,45 микрон или 0,22 микрон, и наконец, добавляли 1,10 объема ацетонитрила. Этот раствор непосредственно с помощью насоса вводили в обращенно-фазовую колонку с С4, а затем очищали и уложенный ТРО элюировали с помощью той же самой градиентной программы, описанной выше. Уложенный биологически активный ТРО элюировали в присутствии приблизительно 45% ацетонитрила в тех же самых условиях. Неправильные дисульфид-связанные формы ТРО элюировались раньше. Конечный очищенный ТРО (Met-1 1-153), имеющий чистоту более чем 95%, о чем свидетельствовал анализ, проведенный с помощью электрофореза в ПААГ с ДСН, анализировали посредством обращенно-фазовой хроматографии на колонках С4. Для in vivo-исследований материал, очищенный на колонке С4, диализовали в физиологически совместимые буферы. Были также использованы изотонические буферы (10 мМ ацетат натрия (рН 5,5)), 10 мМ сукцинат натрия (рН 5,5) или фосфат натрия (рН 7,4)) содержащие 150 мМ хлорид натрия и 0,01% Твин 80.
Благодаря высокой активности ТРО в анализе Ba/F3 (половина максимальной стимуляции достигалась приблизительно при 3 пг/мл), был получен биологически активный материал с использованием многих других буферов, детергентов и окислительно-восстановительных условий. Однако даже в улучшенных условиях было получено лишь небольшое количество правильно уложенного материала (<10%). Для промышленного изготовления данного продукта желательно, чтобы его выход составлял, по крайней мере, 10%, более предпочтительно 30-50% и наиболее предпочтительно >50%. Многие другие детергенты (Тритон Х-100, додецил-бета-мальтозид, CHAPS, CHAPSO, ДСН, саркозил, Твин 20 и Твин 80, Цвиттергент 3-14 и др.) анализировали на эффективность поддержания высокого уровня самопроизвольного сорачивания белка. Из этих детергентов, как было обнаружено, только семейство CHAPS (CHAPS и CHAPSO) может быть использовано в реакции сворачивания в целях ограничения агрегации белка и образования неправильных дисульфидных связей. Наиболее предпочтительно, чтобы уровни CHAPS составляли более 1%. Для наилучшего выхода оптимальные уровни хлорида натрия должны составлять 0,1 М и 0,5 М. Присутствие EDTA (1-5 мМ) ограничивает количество катализируемого металлом окисления (и агрегации), которое наблюдается при использовании некоторых препаратов. Концентрации глицерина, составляющие более чем 15%, дают оптимальные условия для сворачивания молекулы. Для максимального выхода очень важно, чтобы присутствовали оба как окисленный и восстановленный глутатио, так окисленный и восстановленный цистеин в качестве окислительно-восстановительной пары реагентов. Более высокий выход наблюдается в основном в том случае, когда молярное количество окисленного реагента равно или превышает количество восстановленного реагента в окислительно-восстановительной паре. Оптимальное значение рН для сворачивания указанных ТРО-вариантов составляет в пределах от 7,5 до около 9. Органические растворители (например, этанол, ацетонитрил, метанол) допустимы при концентрациях 10-15% или ниже. Более высокие уровни органических растворителей повышают возможность образования неправильно уложенных форм. Обычно используются Трис и фосфатные буферы. Инкубирование при 4°С также дает более высокие уровни правильно уложенного ТРО.
Обычно для препаратов ТРО, которые были очищены в процессе первой С4-стадии, уровни восстановления укладки белка составляют 40-60% (исходя из количества восстановленного и денатурированного ТРО, используемого в реакции сворачивания. Активный материал может быть получен при менее чистых препаратах (например, полученных непосредственно после колонки с Сефадексом 200 или после начальной экстракции рефрактивных телец), хотя выходы в этом случае меньше из-за экстенсивной преципитации и воздействия других белков (не ТРО) во время процесса восстановления укладки белка.
Поскольку ТРО (Met-1 1-153) содержит 4 цистеиновых остатка, то могут быть продуцированы три различных дисульфидных варианта этого белка:
Вариант 1: дисульфиды между цистеиновыми остатками 1-4 и 2-3
Вариант 2: дисульфиды между цистеиновыми остатками 1-2 и 3-4
Вариант 3: дисульфиды между цистеиновыми отатками 1-3 и 2-4
В процессе первоначального исследования по определению условий восстановления укладки белка несколько различных пиков, содержащих белок ТРО, были разделены с помощью С4-обращенно-фазовой хроматографией. Как было определено с помощью анализа на клетках Ba/F3, лишь один из этих пиков обнаруживал значительную биологическую активность. Затем условия для восстановления укладки были оптимизированы для получения предпочтительно этого варианта. В этих условиях варианты неправильного свертывания из всего полученного мономерного ТРО составляли менее 10-20%.
С помощью масс-спектроскопии и секвенирования белка было установлено, что для биологического активного ТРО тип дисульфидных связей представляет собой 1-4 и 2-3 (т.е. вариант 1). Аликвоты различных С4-разрешенных пиков (5-10) гидролизовали трипсином (молярное отношение трипсина к белку составляло (1:25). Смесь для гидролиза анализировали с помощью лазерной масс-спектрометрии путем десорбционной ионизации с использованием матрицы как перед, так и после ДТТ-восстановления. После восстановления были детектированы массы, соответствующие наибольшему из триптических пептидов ТРО. В невосстановленных образцах некоторые из этих масс отсутствовали и наблюдались новые массы. Масса новых пиков соответствовала в основном сумме отдельных триптических пептидов, присутствующих в дисульфидной паре. Таким образом, можно со всей определенностью утверждать, что дисульфидный тип рекомбинантного вновь свернутого биологически активного ТРО соответствует варианту 1-4 и 2-3. Это предположение согласуется с известной дисульфидной картиной родственной молекулы эритропоэтина.
D. Биологическая активность рекомбинантного вновь сложенного ТРО (Met-1 1-153)
Вновь сложенный и очищенный ТРО (Met-1 1-153) обладал активностью как в in vivo, так и в in vitro-анализах. В Ba/F3, полумаксимальная стимуляция включения тимидина в клетки Ba/F3 достигалась при 3/3 пг/мл (0,3 пМ). В ELISA на основе рецептора mpl полумаксимальная активность наблюдалась при 1,9 нг/мл (120 пМ). У нормальных и миелосуппрессированных животных, полученных путем рентгеновского облучения дозами, близкими к летальной, ТРО (Met-1 1-153) обладал высокой активностью (эта активность наблюдалась уже при дозах 30 нг/мышь) к стимуляции продуцирования новых тромбоцитов.
Пример 23
Продуцирование других биологически активных вариантов ТРО в E.coli
В E.coli было продуцировано три различных варианта ТРО, которые представляли собой очищенные и вновь уложенные биологически активные формы, описанные ниже.
(1) MLF - 13 остатков от бактериальной сигнальной последовательности ST11 соединены с N-концевым доменом ТРО (остатки 1-155). Полученная последовательность представляла собой
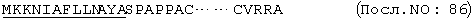
где лидерная последовательность подчеркнута, а C·······C означает Cys7-Cys151. Этот вариант был сконструирован для получения тиррозина для радиоактивного иодирования ТРО для исследований рецепторов и биологических исследований.
(2) H8MLF - 7 остатков от последовательности ST11, 8 остатков гистидина и последовательность фактор Ха-ферментативного расщепления IEGR были присоединены к N-концевому домену (остатки 1-155) ТРО. Эта последовательность представляет собой:

где лидерная последовательность подчеркнута, а C·······C означает Cys7-Cys151. Этот вариант при очистке и укладке может быть обработан фактором Ха, который будет расщеплять последовательность IEGR после аргининового остатка, в результате чего будет получен вариант ТРО в 155 остатков с натуральной сериновой N-концевой аминокислотой.
(3) T-H8MLF - получали, как описано выше для варианта (2), за исключением того, что тромбин-восприимчивая последовательность IEPR соединена с N-концевым доменом ТРО. В результате была получена последовательность:

где лидерная последовательность подчеркнута, а C·······C означает Цис7-Цис151. Этот вариант после очистки и укладки может быть обработан тромбином с получением N-концевого натурального варианта ТРО из 155 остатков.
А. Восстановление, солюбилизация и очистка мономерных биологически активных вариантов ТРО: (1), (2) и (3)
Все варианты были экспрессированы в E.coli. Большинство вариантов было однаружено в рефрактивных тельцах, как описано в Примере 22 для ТРО (Met-1 1-153). Идентичные процедуры для восстановления, солюбилизации и очистки мономерных ТРО-вариантов проводили, как описано в Примере 22. При этом условия восстановления укладки белка были идентичны условиям, использованным для ТРО (Met-1 1-153), где полный выход составлял 30-50%. После восстановления укладки ТРО-варианты очищали с помощью С4-обращенно-фазовой хроматографии в 0,1% ТГФ с использованием градиента ацетонитрила, как описано ранее. Все ТРО-варианты (в их непротеолизированной форме) имели биологическую активность в Ва/F3-анализах с полумаксимальной активностью 2-5 пМ.
В. Протеолитический процессинг вариантов (2) и (3) с образованием активных N-концевых ТРО (1-155)
Были сконструированы вышеуказанные ТРО-варианты (2) и (3), содержащие ферментативно отщепляемый лидерный пептид, расположенный перед нормальным N-концевым аминокислотным остатком ТРО. После укладки и очистки вариантов (2) и (3), как описано выше, каждый вариант подвергали гидролизу соответствующим ферментом. На стадии С4-обращенно-фазовой хроматографии ацетонитрил удаляли путем продувания раствора легким потоком азота. После этого два варианта обрабатывали либо фактором Ха, либо тромбином, как описано ниже.
В случае варианта (2) к не содержащему ацетонитрила раствору добавляли 1М Трис-буфер (рН 8) до конечной концентрации 50 мМ и, если необходимо, рН доводили до 8. Затем добавляли NaCl и CaCl2 до 0,1 М и 2 М соответственно. Фактор Ха (New England Biolabs) добавляли в таком количестве, что молярное отношение фермента к варианту составляло 1:25-1:100. Образец инкубировали в течение 1-2 часов при комнатной температуре для достижения максимального расщепления, которое оценивали по изменению миграции на ДСН-геле, отображающей потерю лидерной последовательности. Затем реакционную смесь очищали посредством С4-обращенно-фазовой хроматографии с использованием того же самого градиента и тех же самых условий, которые были описаны выше для очистки правильно уложенных вариантов. При этих условиях нерасщепленный вариант В отделялся от расщепленного варианта (2). Было показано что N-концевыми аминокислотами являются SPAPP, что свидетельствует о том, что отщепление N-концевой лидерной последовательности прошло успешно. Фактор Ха также генерировал вариабельные количества внутреннего расщепления в ТРО-доменен; причем расщепление наблюдалось после агрининового остатка в положении 118, в результате чего образовывалась дополнительная N-концевая последовательность TTAHKDP (SEQ ID NO 88). На нередуцирующих ДСН-гелях наблюдалась одна полоса приблизительно 17000 Да для варианта, расщепленного фактором Ха; а на редуцирующих гелях наблюдались две полосы с молекулярной массой приблизительно 12000 и 5000 Да, что соответствует расщеплению у аргинина 118. Эти наблюдения также подтверждают тот факт, что две части молекулы связаны вместе дисульфидной связью между 1 и 4 цистеиновыми остатками, как было выявлено в результате экспериментов по триптическому расщеплению, описанных выше. В Ва/F3-анализе очищенный ТРО-вариант (1-155) после удаления N-концевой лидерной последовательности и внутреннего расщепления имел полумаксимальную активность 0,2-0,3 пМ. Интактный вариант с лидерной последовательностью имел полумаксимальную активность 2-4 пМ.
Для варианта (3) использовали буфер для гидролиза, состоящий из 50 мМ Трис, рН 8; 2% CHAPS; 0,3 М NaCl; 5 мМ EDTA и человеческого или коровьего тромбина (Calbiochem) при соотношении (по массе) фермента и ТРО-варианта 1:25-1:50 соответственно. Гидролиз проводили при комнатной температуре в течение 2-6 часов. Процесс расщепления оценивали по ДСН-гелям, как было описано выше для реакции расщепления фактором Ха. За это время в основном достигалось более чем 90%отщепления лидерной последовательности. Полученный ТРО очищали на С4-обращенно-фазовых колонках, как описано выше и при аминокислотном секвенировании было выявлено, что этот ТРО имеет нужный N-аминоконец. При этом, как и в случае фактора Ха, описанного выше, наблюдались очень небольшие (<5%) количества внутреннего расщепления в той же самой аргинин-треониновой связи. Полученный ТРО-белок обладал высокой биологической активностью в Ва/F3-анализе, при этом полумаксимальные ответы наблюдались при 0,2-0,4 пМ белка. В ELISA-анализе на основе рецептора mpl этот белок имел полумаксимальный ответ при 2-4 нг/мл очищенного белка (120-240 пМ), тогда как интактный вариант, содержащий лидерную последовательность, имел в 5-10 раз меньшую активность в обоих анализах. Для in vivo-исследования ВЭЖХ-очищенный расщепленный белок диализовали в физиологически приемлемых буферах (150 мМ NaCl, 0/01% Твин 80 и 10 мМ сукцината натрия, рН 5,5 или 10 мМ ацетат натрия, рН 5,5 или 10 мМ фосфат натрия, рН 7,4). В случае ВЭЖХ-и ДСН-гелей очищенный белок сохранял стабильность в течение нескольких недель при 4°С. У нормальных и миелосуппрессированных мышей, этот очищенный ТРО с аутентичной N-концевой последовательностью обладал высокой активностью и стимулировал продуцирование тромбоцитов при дозах уже в 30 нг на мышь.
Пример 24
Синтетический лиганд mpl
Хотя человеческий лиганд mpl (hML) обычно получали с использованием рекомбинантных методов, однако он может быть также синтезирован путем ферментативного лигирования синтетических пептидных фрагментов в соответствии с методикой, описанной ниже. Синтетическое продуцирование hML позволяет вводить ненатуральным аминокислоты или синтетические функциональные группы, такие как полиэтиленгликоль. Ранее для эффективного лигирования сложных эфиров пептидов в водном растворе был сконструирован мутант субтилизина (сериновой протеазы) BPN, субтилигаза (S221C/P225A) (Abrahmsen et al., Biochem., 30: 4151-4159 [1991]). В настоящей работе было показно, что синтетические пептиды могут быть ферментативно лигированы последовательным методом в целях продуцирования ферментативно активных длинных пептидов, таких как рибонуклеаза A (Jackson et al., Science [1994]). Эта техника, которая более подробно описана ниже, позволяет осуществлять химических синтез белков, который ранее мог проводиться лишь с помощью технологии рекомбинантных ДНК.
Общая стратегия минтеза hМL153 с использованием субтилигазы представлена на Схеме 1. Начиная с полностью деблокированного пептида, соответствующего защищенному у N-конца С-концевому фрагменту белка, добавляли под действием субтилигазы активированный у С-конца сложным эфиром пептид. После завершения реакции продукт выделяли с помощью обращенно-фазовой ВЭЖХ и защитную группу у N-конца удаляли. Затем лигировали следующий пептидный фрагмент, после чего его деблокировали и процесс повторяли снова до тех пор, пока не был получен полноразмерный белок. Этот процесс аналогичен твердофазному пептидному синтезу, в котором защищенный у N-конца и активированный у С-конца пептид присоединяют к N-концу предыдущего пептида, в результате чего синтезируют пептид в направлении C→N. Однако вследствие того, что каждое присоединение добавляет до 50 остатков, а продукты выделяют после каждого лигирования, этот метод позволяет синтезировать белки с гораздо более высокой степенью чистоты и с приемлемыми выходами.
Схема I: Стратегия синтеза hML с использованием субтилигазы
На основании наших знаний относительно специфичности последовательности субтилигазы, а также аминокислотной последовательности биологически активного "эпо-домена" hML мы разделили hML153 на семь фрагментов длиной в 18-25 остатков. Для определения подходящих областей соединения для лигирования 18-25-метров были синтезированы тетрапептиды для теста на лигирование. Результаты таких тестов на лигирование представлены в Таблице 13.
Таблица 13
Тест на лигирование hML. Донорные и нуклеофильные пептиды при 10 мМ в 100 мМ трицина (рН 7,8) при 22°С. Затем добавляли лигазу до конечной концентрации 10 мкМ из 1,6 мг/мл маточного раствора (~70 мкМ) и смесь для лигирования оставляли на ночь для реакции. Выходы выражали как % лигирования по отношению к гидролизу донорных пептидов.
На основании этих экспериментов с помощью субтилигазы может быть проведено успешное лигирование пептидов, проиллюстрированных в Таблице 14. Для предупреждения самолигирования необходимо использовать подходящую защиту для N-конца каждого донорного сложноэфирного пептида. В качестве защитной группы мы выбрали изоникотинил (iNOC) (Veber et al., J.Org.Chem., 42: 3286-3289 [1977]), поскольку она является водорастворимой, может быть введена в последней стадии твердофазного синтеза и является стабильной в отношении безводного HF, который используется для деблокирования и отщепления пептидов от твердофазного полимера. Кроме того, эта группа может быть удалена из пептида после каждого лигирования в мягких восстановительных условиях (Zn/CH3CO2H) с получением свободного N-конца для последующего лигирования. Сложный эфир гликолат-лизил-амида (glc-K-NH2) был использован для активации С-конца на основании предыдущих экспериментов, которые показали, что он может быть эффективно ацилирован с помощью субтилигазы (Abrahmsen et al., Biochem., 30: 4151-4159 [1991]). INOC-защищенные и активированные glc-K-амидом пептиды могут быть синтезированы с использованием стандартных твердофазных методов, как показно на Схеме 2. Эти пептиды последовательно лигируют до тех пор пока не будет продуцирован полный белок, после чего конечный продукт подвергают реакции укладки in vitro. Исходя из гомологии с ЕРО дисульфидные пары, очевидно, образуются между цистеиновыми остатками 7 и 157 и между 28 и 85. Окисление дисульфидов может быть осуществлено путем простого размешивания восстановленного материала в течение нескольких часов в атмосфере кислорода. Свернутый материал затем очищают с помощью ВЭЖХ и фракции, содержащие активный белок, собирают и лиофилизуют. Альтернативно, дисульфиды могут быть различным образом защищены для контроля за последующим окислением между специфическими дисульфидными парами. Защита цистеинов 7 и 151 ацетомидометильными (acm) группами должна обеспечивать окисление 28 и 85. acm-Группы могут быть затем удалены, а остатки 7 и 151 окислены. И наоборот, остатки 28 и 85 могут быть асm-защищены и окислены, в случае если для правильной укладки необходимо последующее окисление. Цистеины 28 и 85 могут быть замещены, но не обязательно, другим натуральным или ненатуральным остатком, не являющимся Цис, для обеспечения правильного окисления цистеинов 7 и 151.
Таблица 14
Пептидные фрагменты, используемые для полного синтеза с использованием субтилигазы
Фрагмент
Последовательность
1 (Посл.NO: 110)
iNOC-HN-SPAPPACDLRVLSKLLRDSHVL-glc-K-NH2 (1-22)
2 (Посл.NO: 111)
iNOC-HN-HSRLSQCPEVHPLPTPVLLPAVDF-glc-K-NH2 (23-46)
3 (Посл.NO: 112)
iNOC-HN-SLGEWKTQMEETKAQDILGAVTL-glc-K-NH2 (47-69)
4 (Посл. NO: 113)
iNOC-HN-LLEGVMAARGQLGPTCLSSLL-glc-K-NH2 (70-90)
5 (Посл. NO: 114)
iNOC-HN-GQLSGQVRLLLGALQS-glc-K-NH2 (90-106)
6 (Посл. NO: 115)
iNOC-HN-LLGTQLPPQGRTTAHKDPNAIF-glc-K-NH2 (107-128)
7 (Посл. NO: 116)
iNOC-HN-LSFQHLLRGKVRFLMLVGGSTLCVR-CO2 (129-153)
Лигирование пептидов осуществляли при 25°С в 100 мМ трицина, рН 8 (свежеприготовленом и дегазированном путем вакуумной фильтрации через 5 мкм-фильтр). Обычно С-концевой фрагмент растворяли в буфере (2-5 мМ пептид) и добавляли 10х маточный раствор субтилигазы (1 мг/мл в 100 мМ трицина, рН 8) для доведения конечной концентрации фермента до ~5 мкМ. Затем добавляли 3-5 молярный избыток glc-K-активированного донорного пептида (в твердом виде), растворяли и полученную смесь оставляли при 25°С. За ходом лигирования следили с помощью аналитической обращенно-фазовой С18-ВЭЖХ (СН3СN/Н2O-градиент с 0/1% TFA). Продукты лигирования очищали с помощью препаративной ВЭЖХ и лиофилизовали. Снятие защитной изоникотинильной (iNOC) группы осуществляли путем размешивания HCl-активированной цинковой пыли с защищенным пептидом в уксусной кислоте. Цинковую пыль удаляли путем фильтрации, а уксусную кислоту выпаривали под вакуумом. Полученный пептид может быть затем непосредственно использован для последующей стадии лигирования и процесс повторяют. Синтетический hML153 может быть лигирован в соответствии с процедурами, аналогичными процедурам, описанным выше для синтетического или рекомбинантного hML154-332, в результате чего может быть продуцирован синтетический или полусинтетический полноразмерный hML.
Синтетический hML имеет много преимуществ по сравнению с рекомбинантным. Натуральные боковые цепи могут быть введены для повышения активности или специфичности. Полимерные функциональные группы, такие как полиэтиленгликоль, могут быть введены для повышения продолжительности действия. Например, полиэтиленгликоль может быть присоединен к лизиновым остаткам отдельных фрагментов (Таблица 14) до или после осуществления одной или нескольких стадий лигирования. Чувствительные к действию протеазы пептидные связи могут быть удалены или модифицированы для улучшения стабильности in vivo. Кроме того, для облегчения определения структуры могут быть синтезированы производные с тяжелыми атомами.
Схема 2: Твердофазный синтез пептидных фрагментов для сегментного лигирования
(а) Лизил-параметилбензгидриламиновую (МВНА) смолу 1 (0,063 мэк/г, Advanced Chem.Tech.) размешивали с бромуксусной кислотой (5 экв.) и диизопропилкарбодиимидом (5 экв.) в течение 1 часа при 25°С в диметилацетамиде (DMA), в результате чего получали бромоацетиловое производное 2. (b) Смолу интенсивно промывали DMA, а отдельные Вос-защищенные аминокислоты (3 экв., Bachem) этерифицировали путем размешивания с бикарбонатом натрия (6 экв.) в диметилформамиде (ДМФ) в течение 24 часов при 50°С, в s результате чего получали соответствующую гликолат-фенилаланиламидную смолу 3. Амино-ацетилированную смолу 3 промывали диметилформамидом (3х) и дихлорметаном (CH2Cl2) (3х) и после этой промывки смолу можно хранить при комнатной температуре в течение нескольких месяцев. Затем смолу (3) загружали в автоматический пептидный синтезатор (Applied Biosystems 430 А) и пептиды наращивали в соответствии со стандартыми процедурами твердофазного синтеза (5). (с) N-α-Boc-группу удаляли с помощью раствора 45%-ной трифторуксусной кислоты в СН2Сl2. (d) Последующие Вос-защищенные аминокислоты (5 экв.) предварительно активировали с использованием гексафторофосфата бензотриазол-1-ил-окси-трис-(диметиламино)-фосфония (ВОР, 4 экв.) и N-метилфорфолина (NMM, 10 экв.) в DMA и проводили реакцию в течение 1-2 часа. (е) Конечную N-α-Boc-группу удаляли (TFA/CH2Cl2), в результате чего получали 4 и вводили изоникотинильную (iNOC) защитную группу, как описано выше (4) посредством размешивания 4-изоникотинил-2-4-динитрофенил-карбоната (3 экв.) и NMM (6 экв.) в DMA в течение 24 часов при 25°С. (f) После расщепления и деблокирования пептида посредством обработки безводным HF (5% анизол/5% этилметилсульфид) в течение часа при 0°С получали iNOC-защищенный гликолат-Цис-амид-активированный пептид 5, который очищали с помощью обращенно-фазовой С18-ВЭЖХ (СН3СN/Н2О-градиент, 0,1% TFA). Идентичность всех субстратов подтверждалась масс-спектрометрией.
Дополнение к юридическому статусу
Настоящее изобретение имеет юридический статус в соответствии с вышеприведенным описанием и легкодоступными ссылками и исходными материалами. Тем не менее, заявители депонировали в Американской коллекции типовых культур (АТСС), Роквил, США, следующую клеточную линию:
Escherichia coli, DH10B-pBSK-hmpl 1 1,8, АТСС, рег. №CRL 69575, депонирована 24 февраля 1994.
Плазмида pSV15.ID.LL.MLORRF, АТСС рег. №CRL 75958, депонирована 2 декабря 1994.
Клетки CHO DP-12, ML 1/50 МСВ (мечен. №1594), АТСС рег. №CRL 11770, депонированы 6 декабря 1994.
Депонирование проводили в соответствии с Будапештским договором по Международному соглашению о депонировании микроорганизмов в целях проведения патентной экспертизы (Будапештский договор). Сохранение жизнеспособных культур гарантируется в течение 30 лет от даты депонирования. Эти микроорганизмы, в соответствии с Будапештским договором будут доступны широкой публике без всякого ограничения после выдачи патента на заявку. Доступность депонированного штамма не должна быть истолкована как лицензия на осуществление изобретения, поскольку это противоречит правам, предоставляемым любым правительством в соответствии с патентными законами данного государства.
Необходимо иметь в виду, что проиллюстрированные и описанные выше варианты осуществления изобретения следует рассматривать как предпочтительные, и в него могут быть внесены различные изменения, замещения и модификации, не выходящие, однако, за рамки существа и объема ниже следующей формулы изобретения. Поэтому следует отметить, что защита, предоставляемая патентной грамотой, ограничивается лишь прилагаемой формулой изобретения и ее эквивалентами.
Все цитируемые работы вводятся в настоящее описание посредством ссылки.



































































| название | год | авторы | номер документа |
|---|---|---|---|
| ДЕГРАДИРОВАННОЕ АНТИТЕЛО, ЯВЛЯЮЩЕЕСЯ АГОНИСТОМ TPO | 2001 |
|
RU2287534C2 |
| ПОЛИПЕПТИД ТРОМБОПОЭТИН (ТРО), ДНК, КОДИРУЮЩАЯ ПОЛИПЕПТИД ТРО (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДА (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБ ЛЕЧЕНИЯ (ВАРИАНТЫ), АНТИТЕЛО К ПОЛИПЕПТИДУ ТРО | 1995 |
|
RU2194074C2 |
| МОДИФИЦИРОВАННОЕ АГОНИСТИЧЕСКОЕ АНТИТЕЛО | 2001 |
|
RU2295537C2 |
| НОВЫЙ ЦИТОКИН ZALPHA11-ЛИГАНД | 2000 |
|
RU2258710C2 |
| ГЕМОПОЭТИЧЕСКИЙ БЕЛОК (ВАРИАНТЫ), ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, СПОСОБ ПОЛУЧЕНИЯ ГЕМОПОЭТИЧЕСКОГО БЕЛКА, СПОСОБ СЕЛЕКТИВНОЙ ЭКСПАНСИИ КРОВЕТВОРНЫХ КЛЕТОК, СПОСОБ ПРОДУЦИРОВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СПОСОБ СТИМУЛЯЦИИ ПРОДУЦИРОВАНИЯ КРОВЕТВОРНЫХ КЛЕТОК У ПАЦИЕНТА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБ ЛЕЧЕНИЯ | 1997 |
|
RU2245887C2 |
| СРЕДСТВА И СПОСОБ ПОЛУЧЕНИЯ ГЕМОПОЭТИЧЕСКОГО БЕЛКА | 1994 |
|
RU2233881C2 |
| СПОСОБЫ И ПРЕПАРАТЫ ДЛЯ СТИМУЛИРОВАНИЯ РОСТА И ДИФФЕРЕНЦИРОВКИ МЕГАКАРИОЦИТОВ | 1995 |
|
RU2158603C2 |
| НОВЫЙ ЛИГАНД РЕЦЕПТОРА ЦИТОКИНА ZCYTOR17 | 2003 |
|
RU2490276C2 |
| ПЕПТИД MPI БЕЗ СИГНАЛЬНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ, ДНК, ЕГО КОДИРУЮЩАЯ, ВЕКТОР, СПОСОБ ОПРЕДЕЛЕНИЯ СПОСОБНОСТИ К СЕКРЕТИРОВАНИЮ ТЕСТИРУЕМОГО ПЕПТИДА, СПОСОБ ВЫДЕЛЕНИЯ КДНК | 1998 |
|
RU2230751C2 |
| АНТИТЕЛА | 2005 |
|
RU2482131C2 |


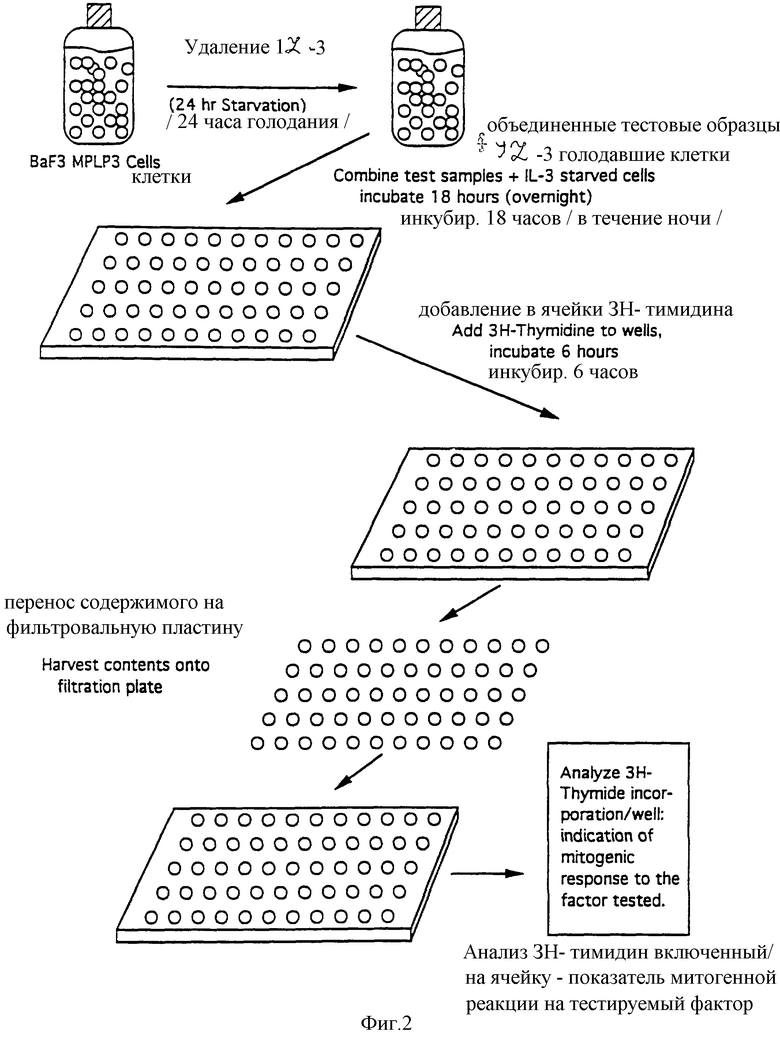
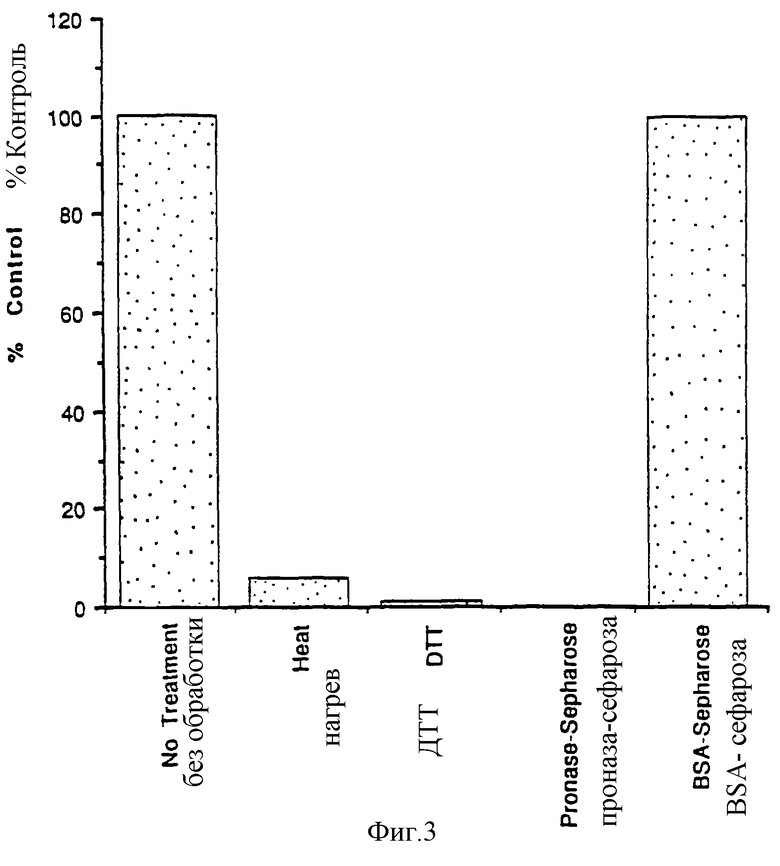
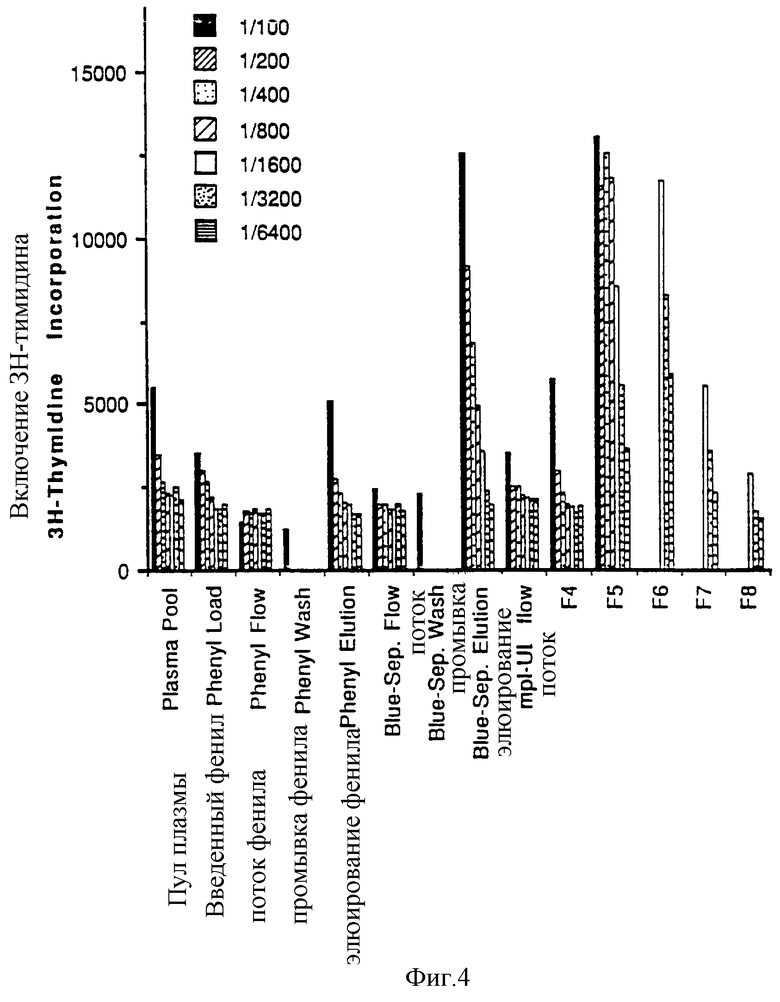
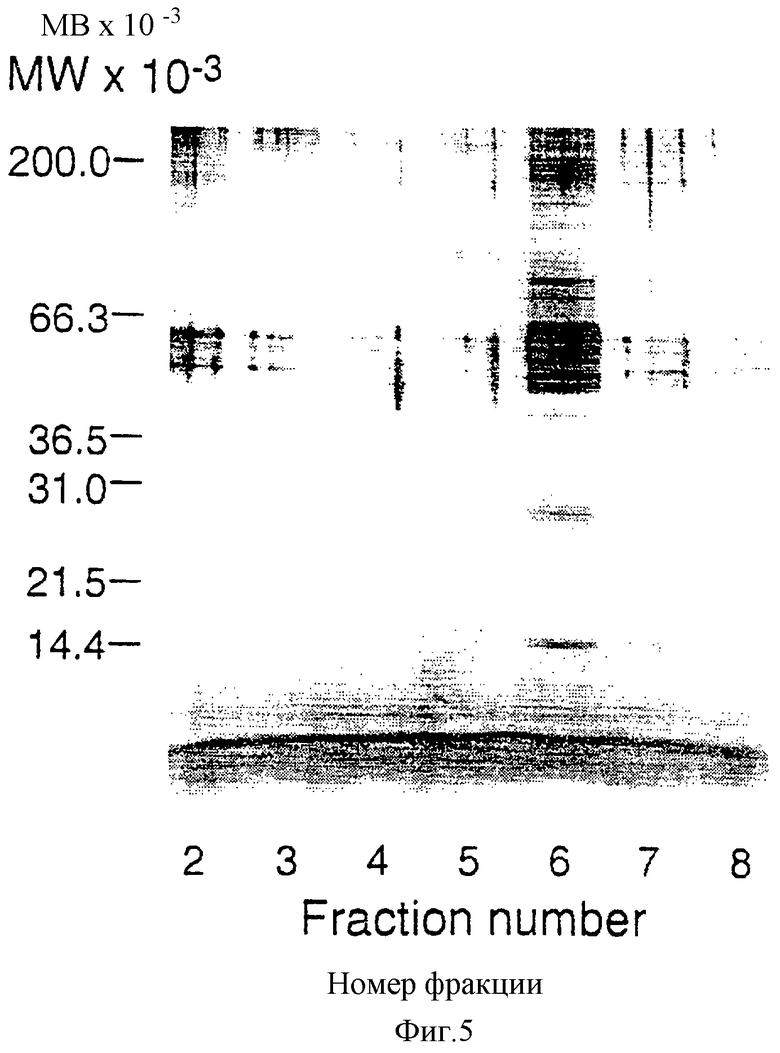
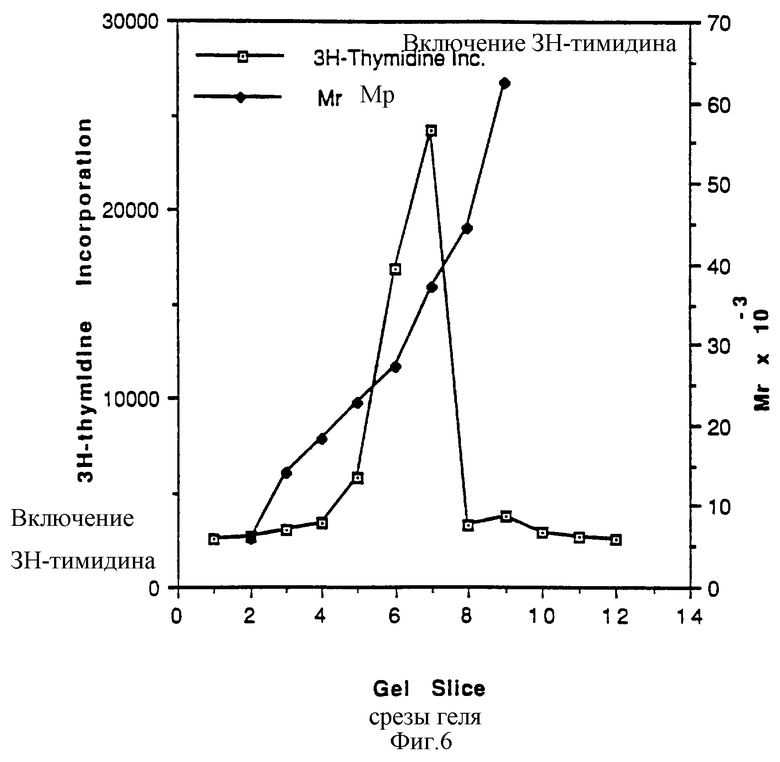
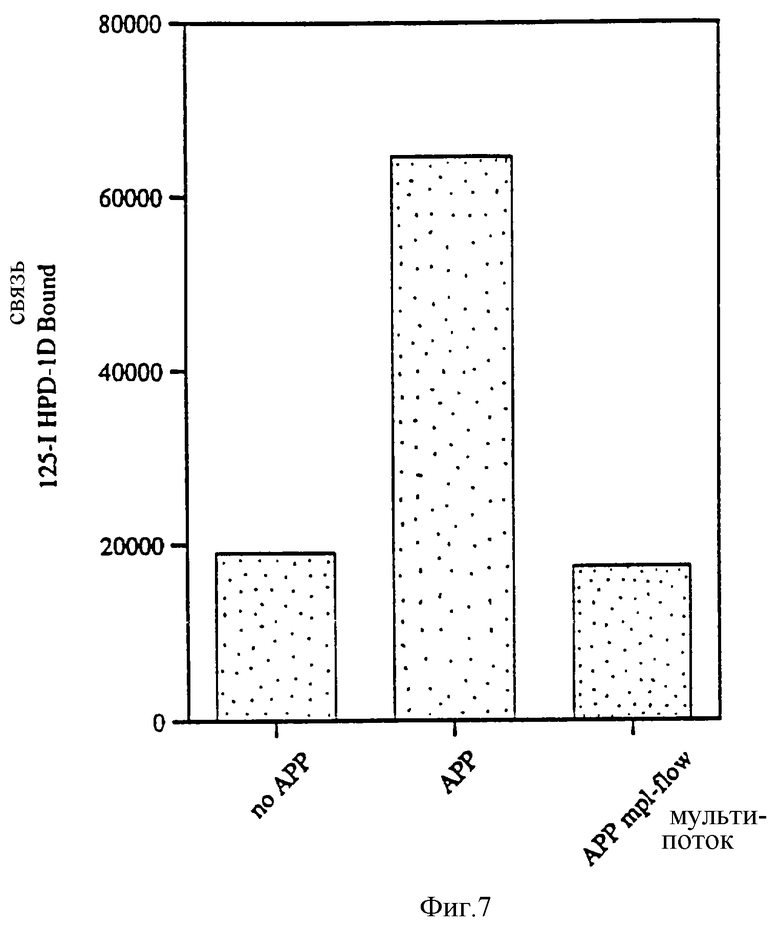
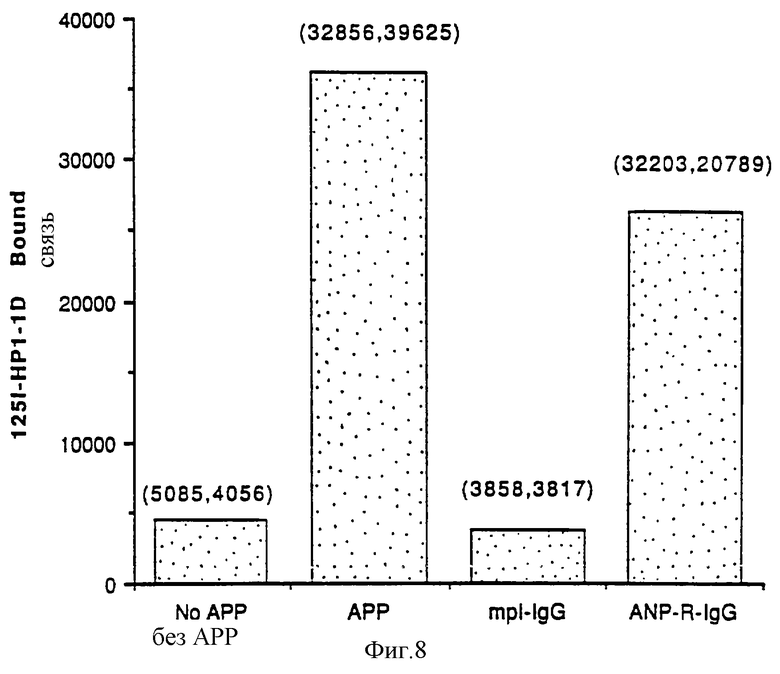

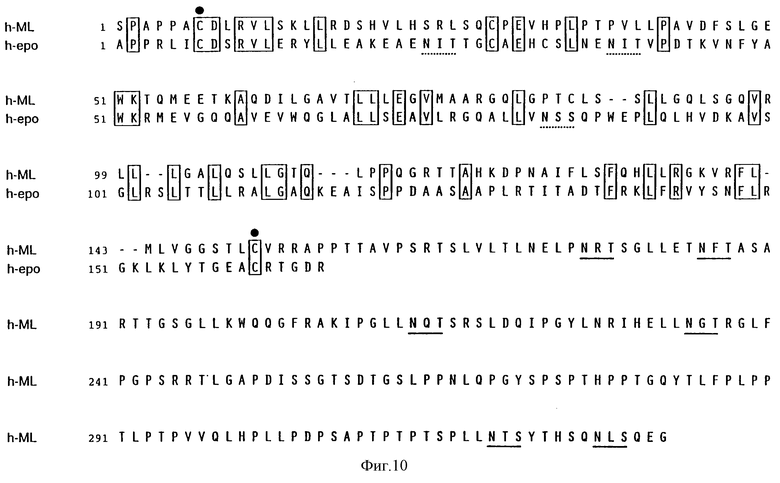

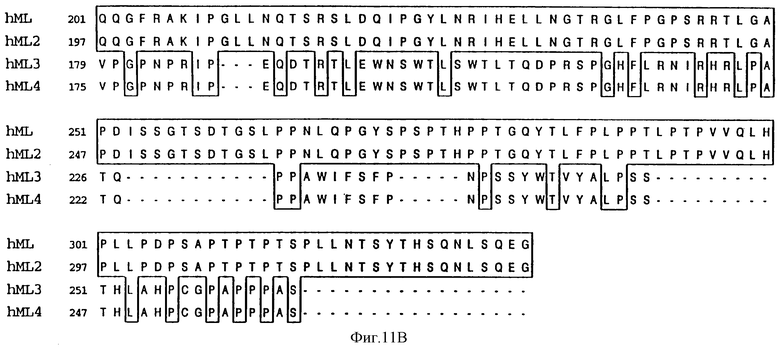
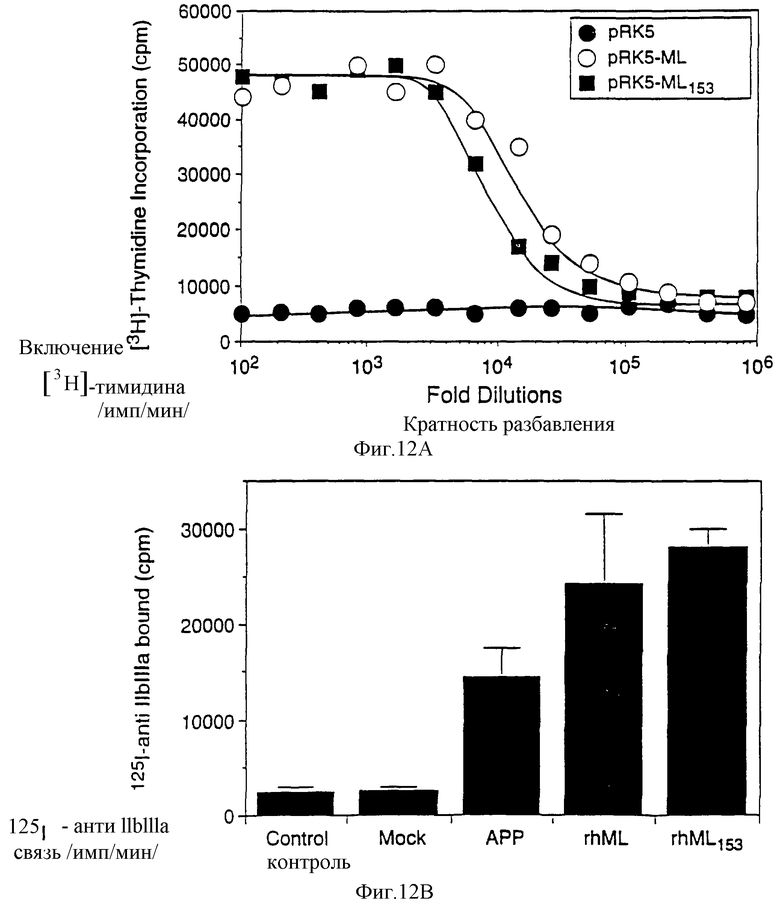
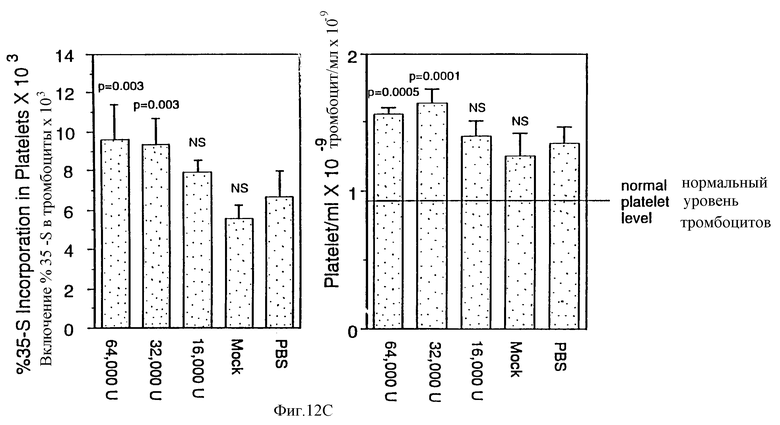
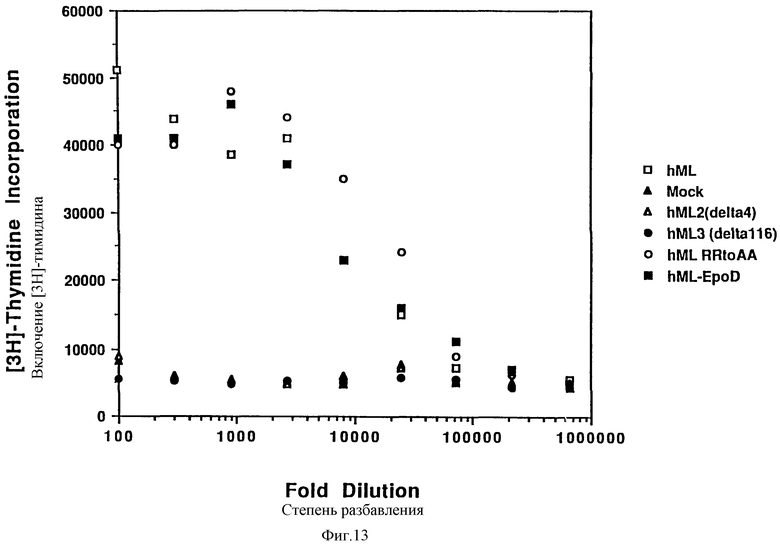
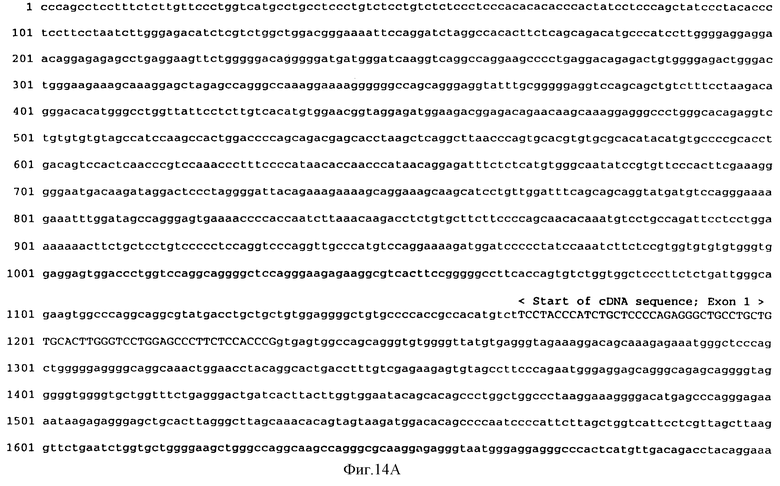









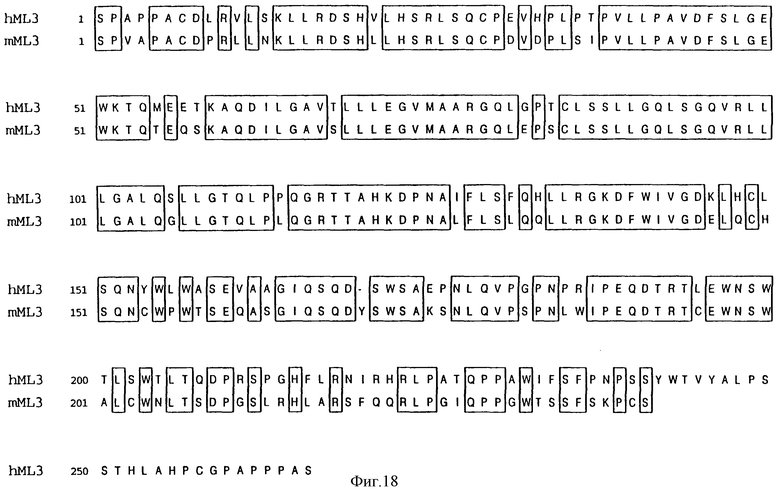






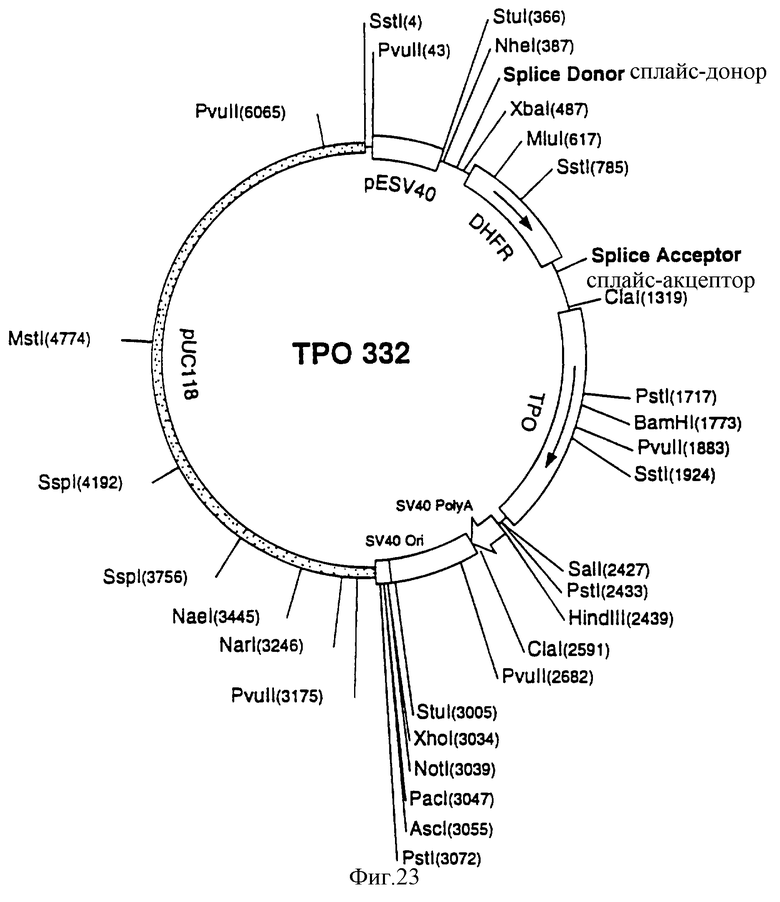
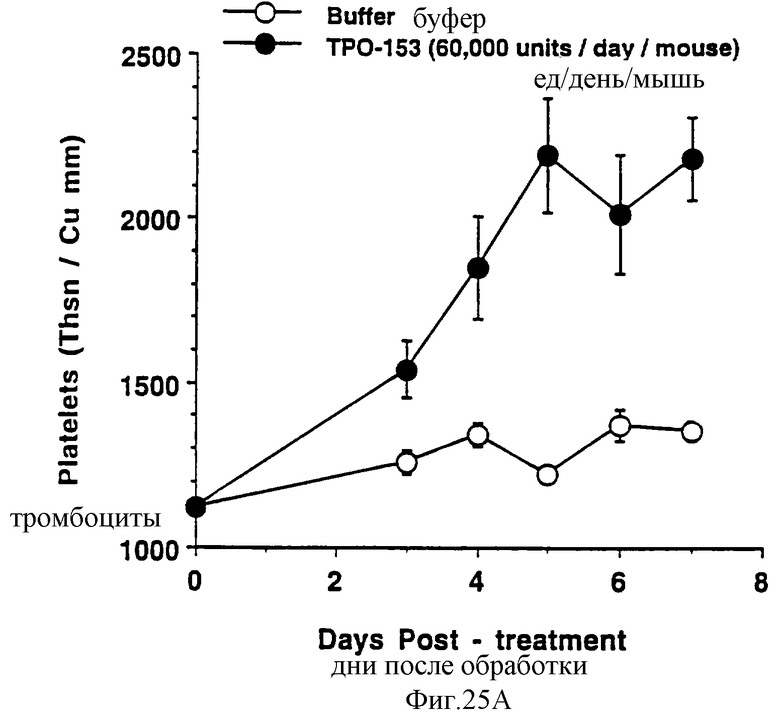
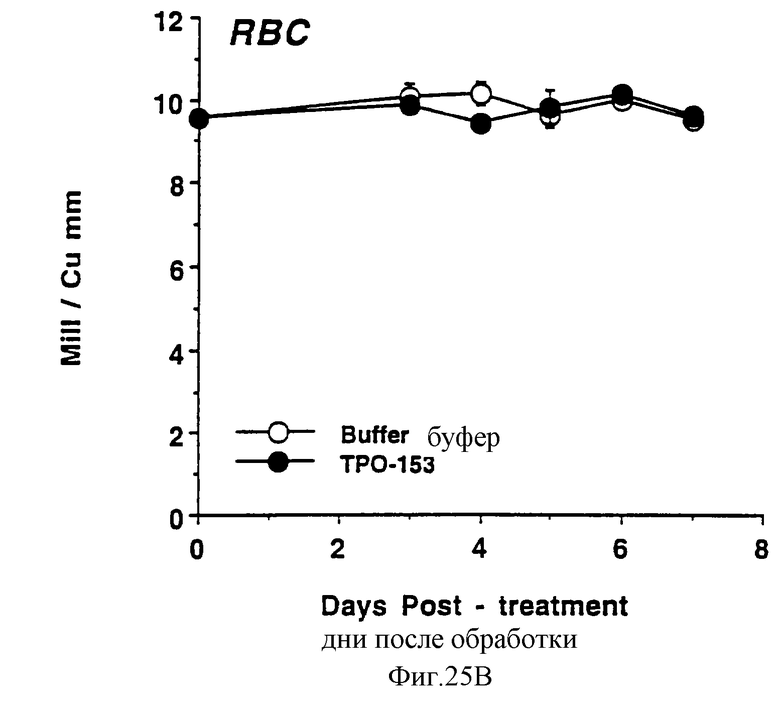
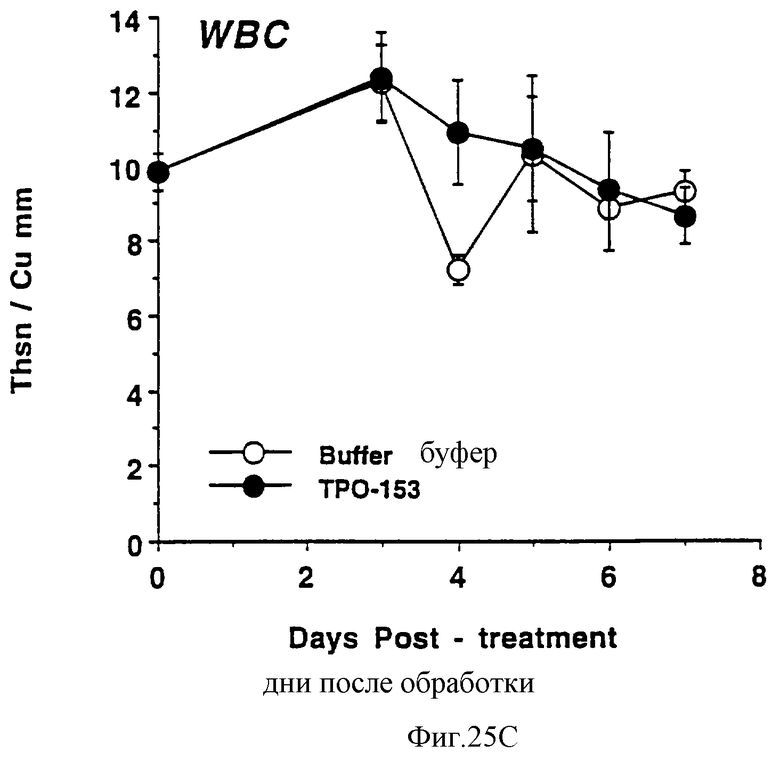
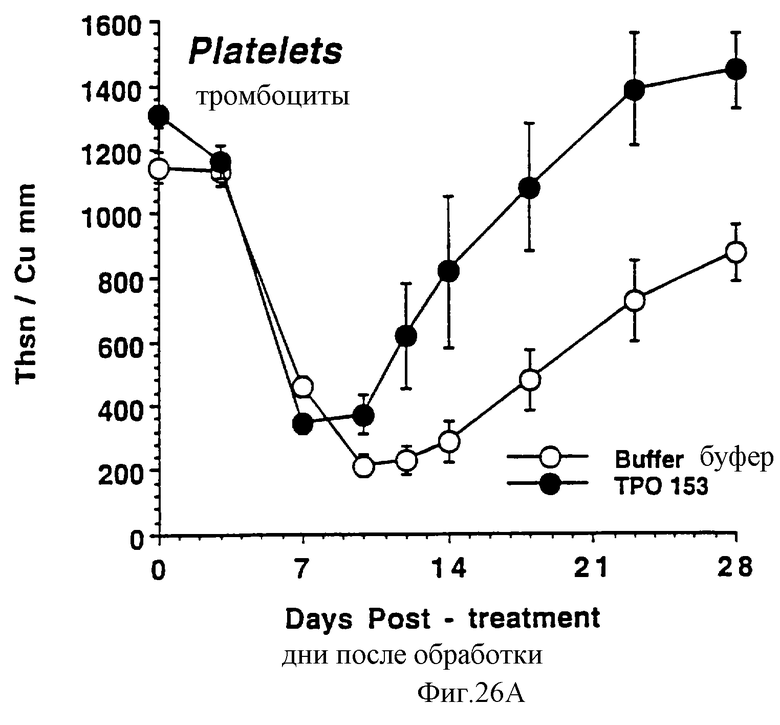
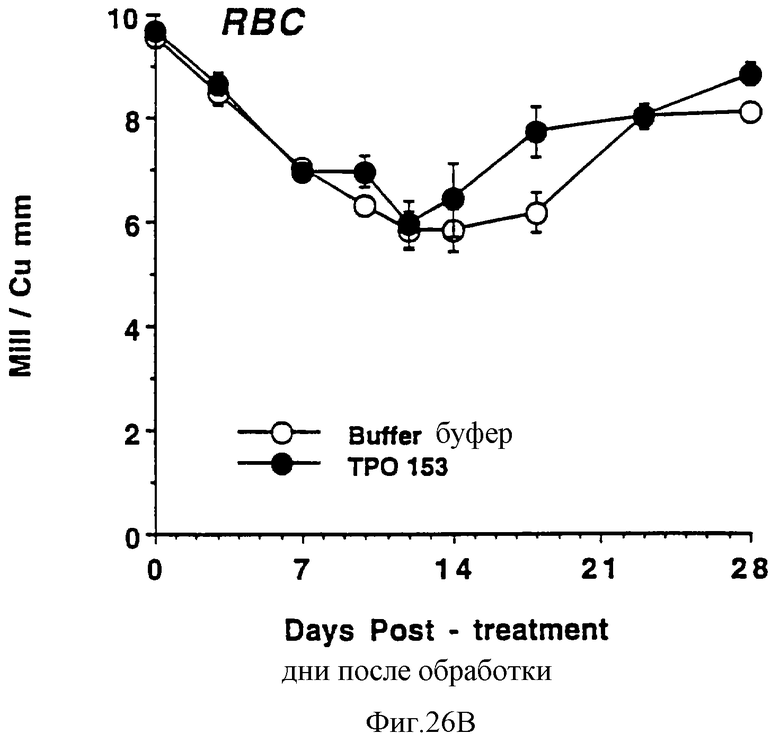
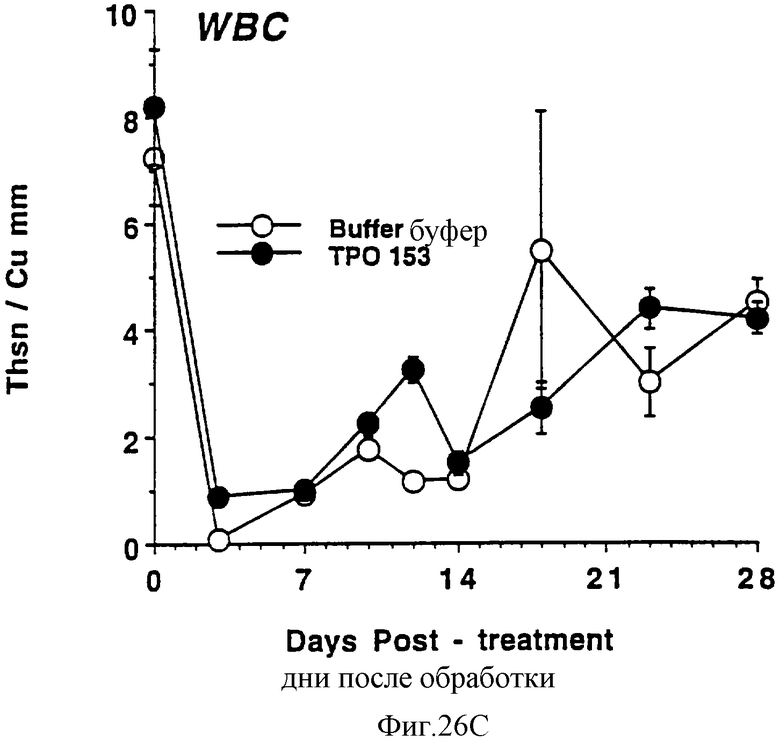
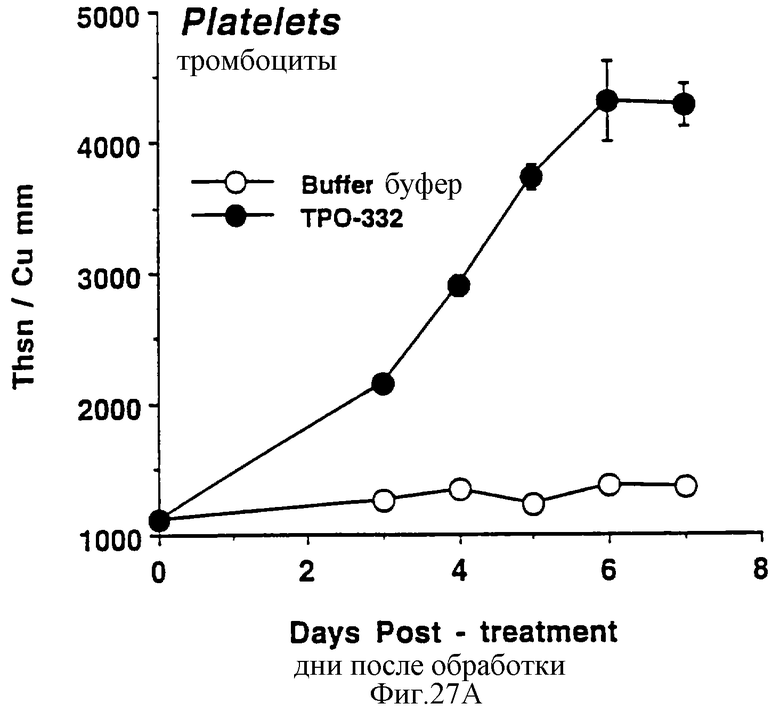
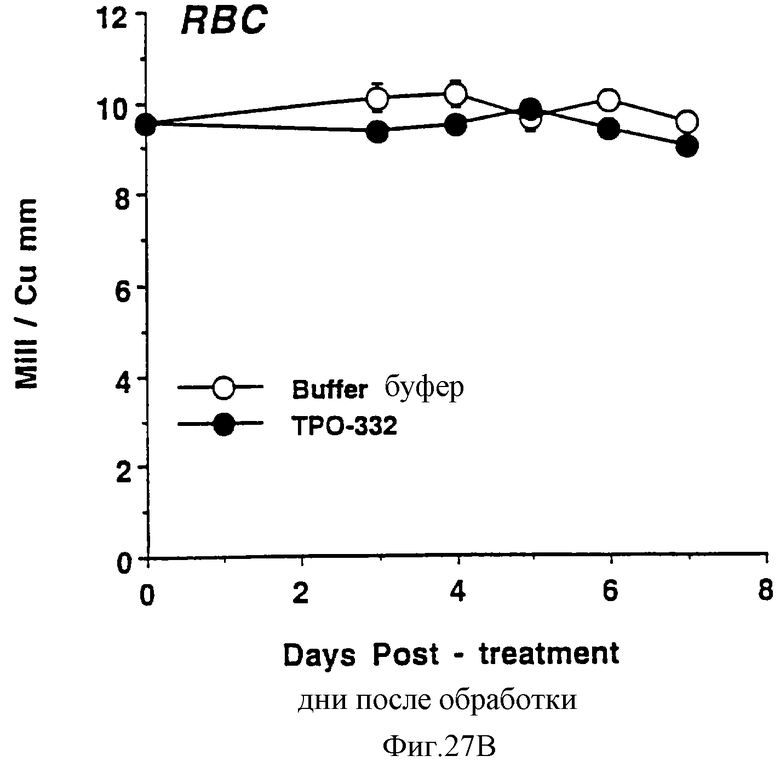
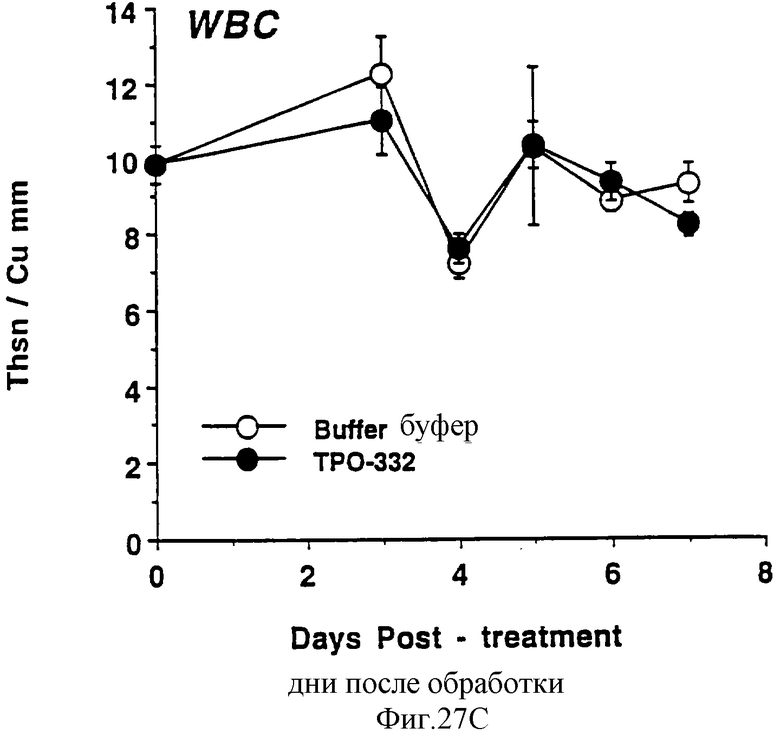
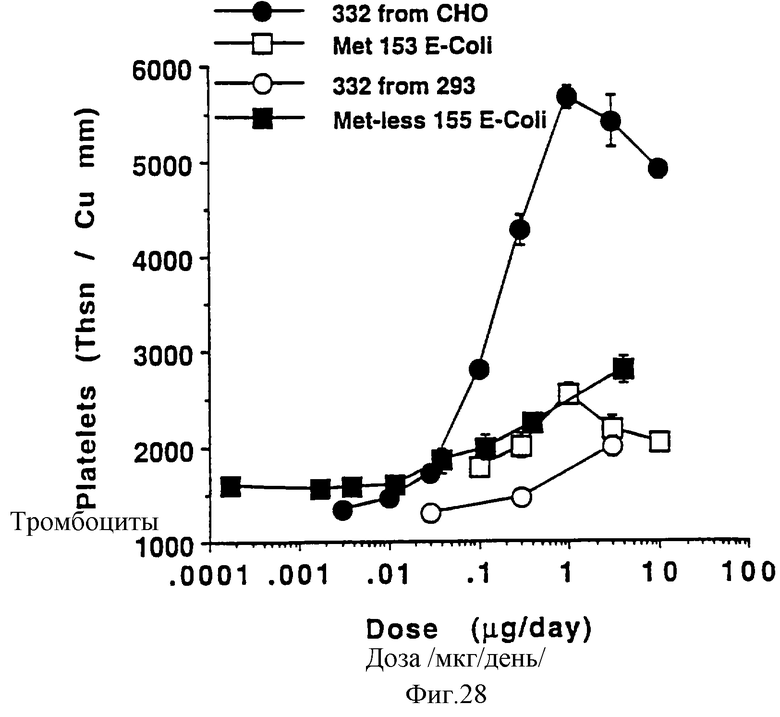
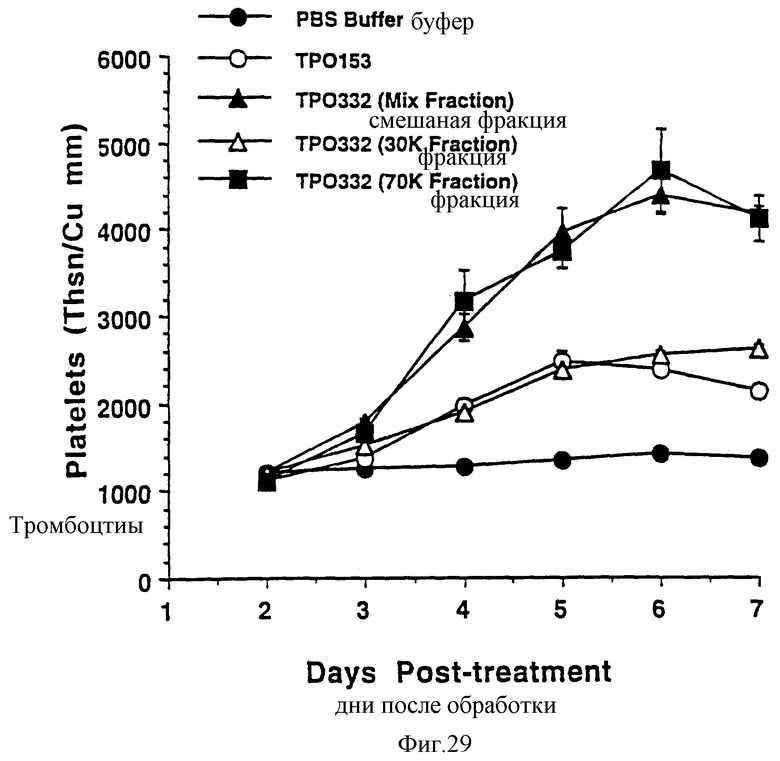
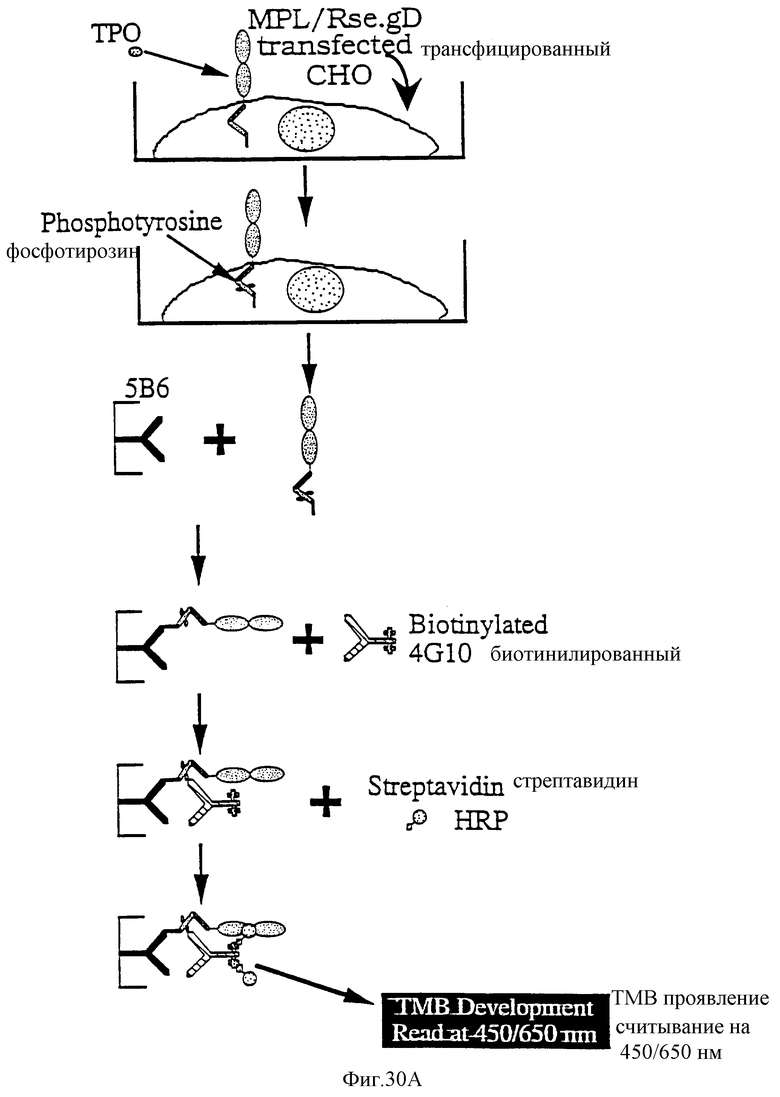
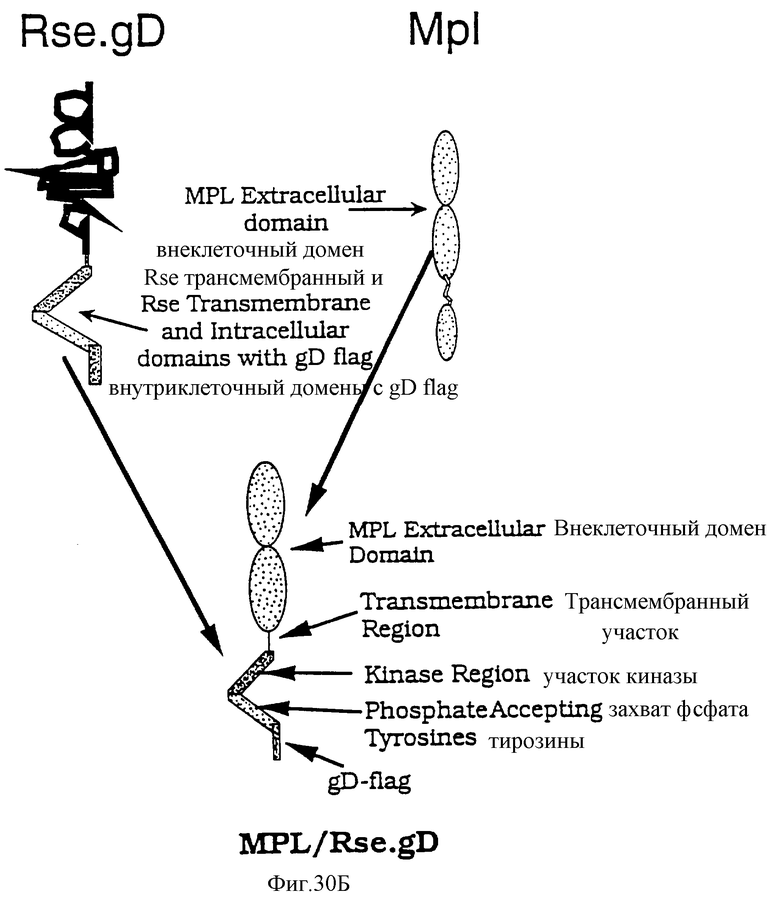
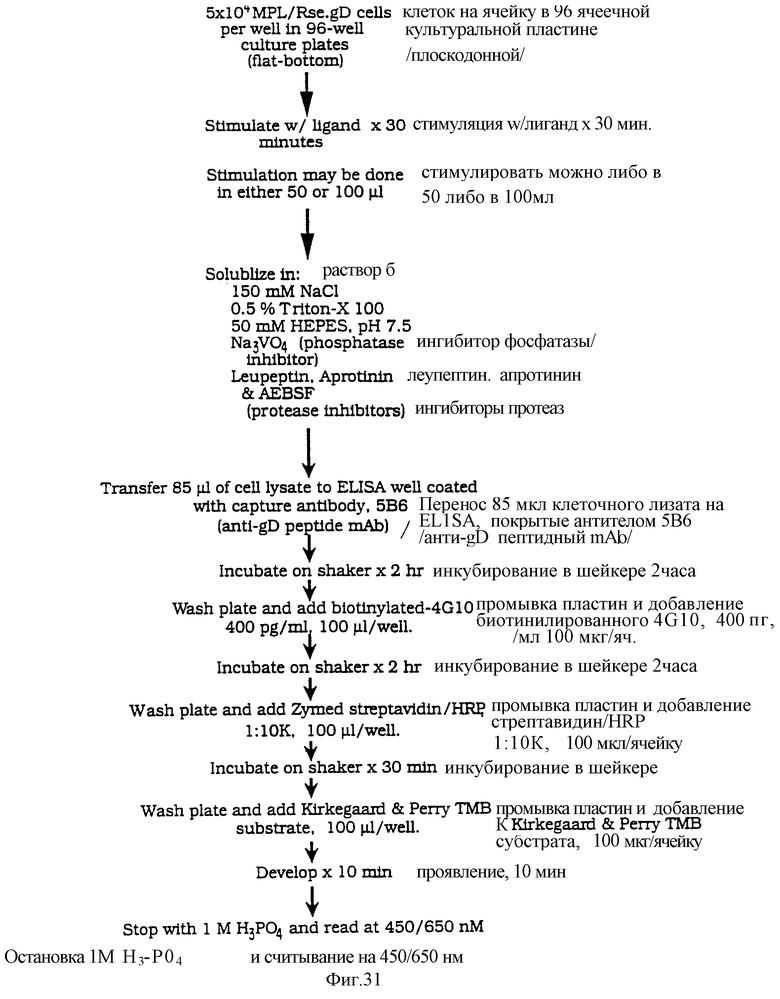
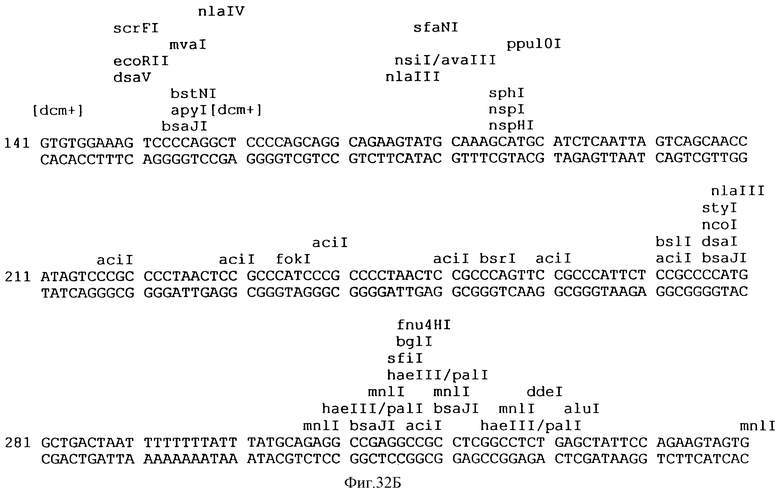
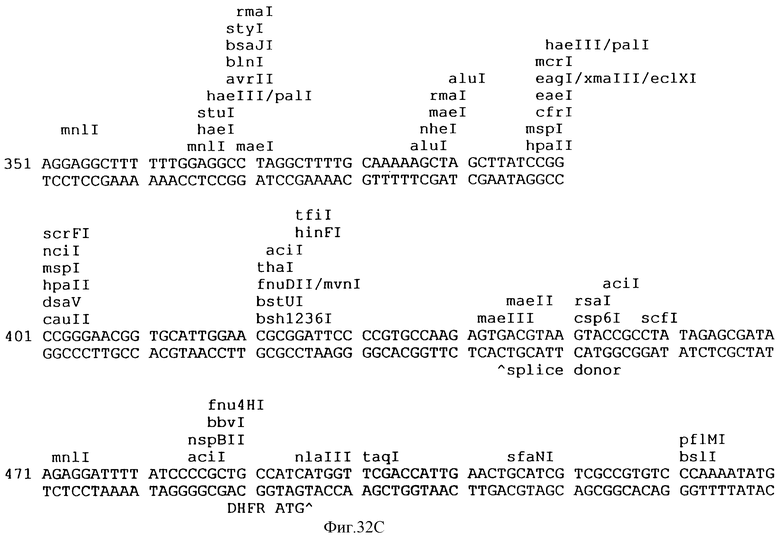
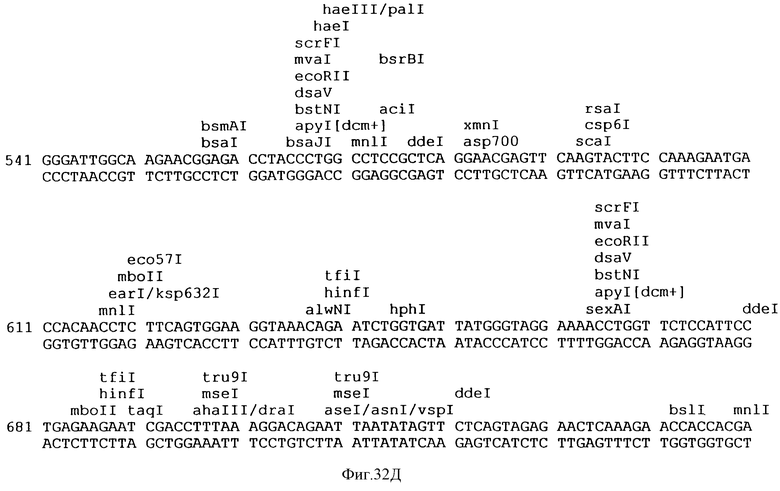
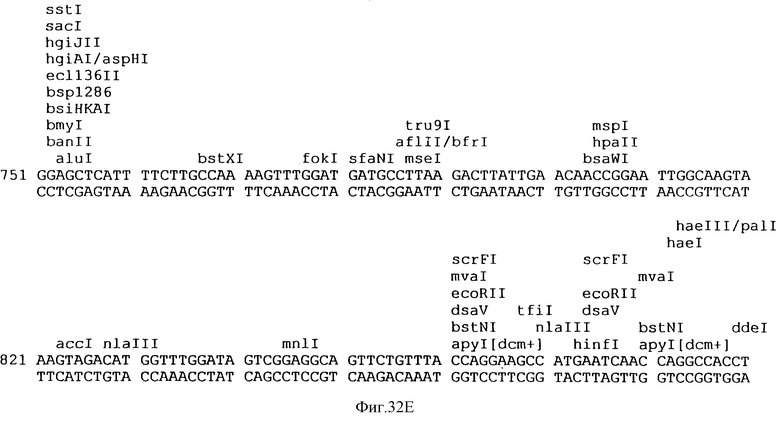

















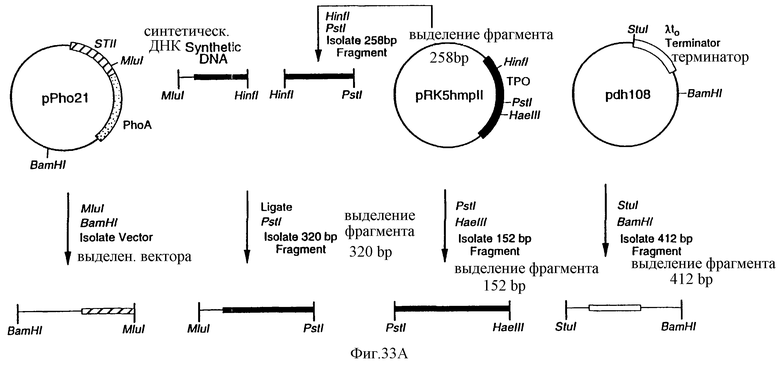
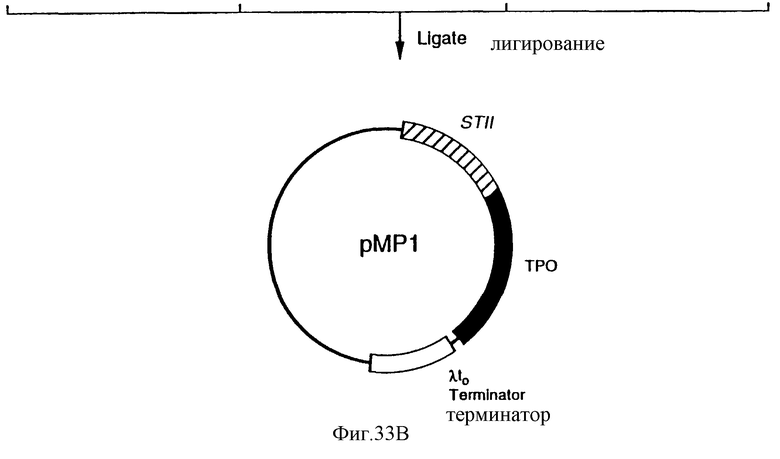
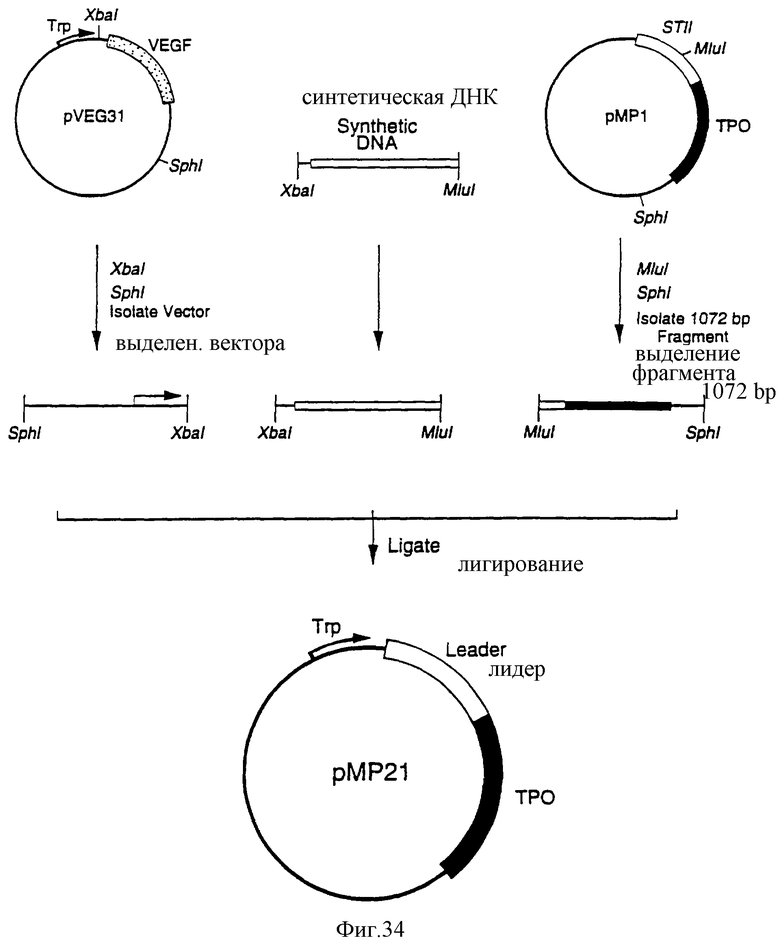
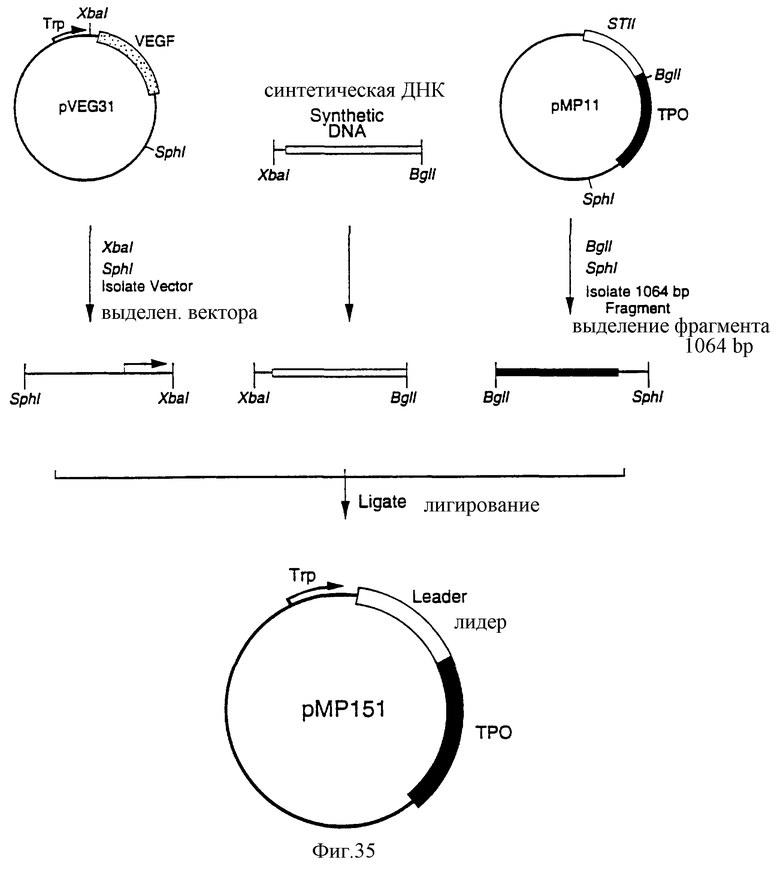
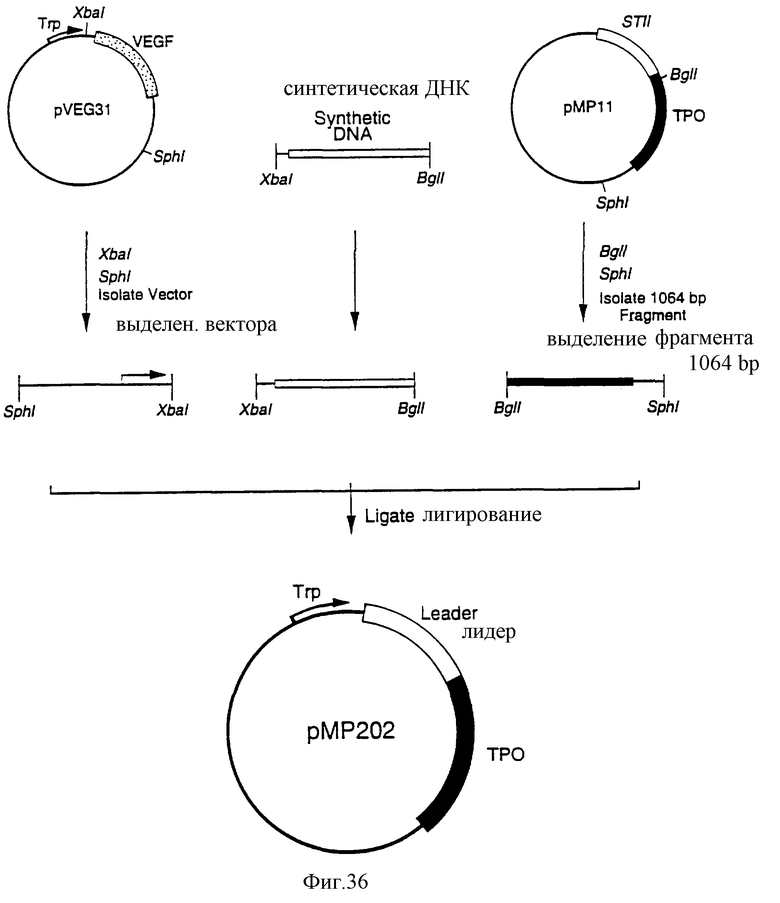
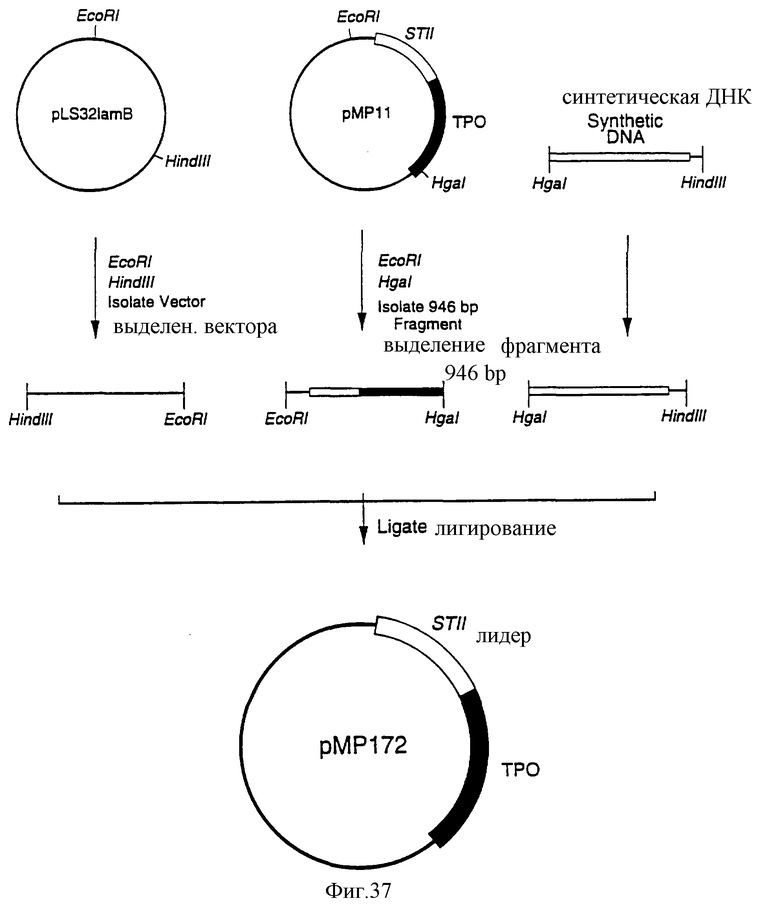
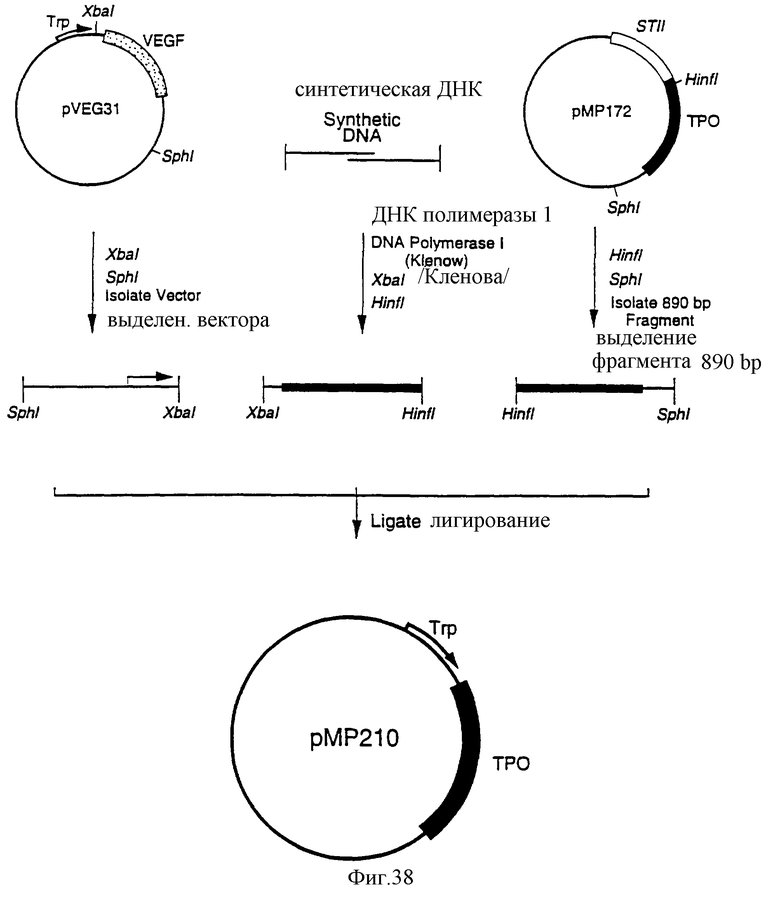

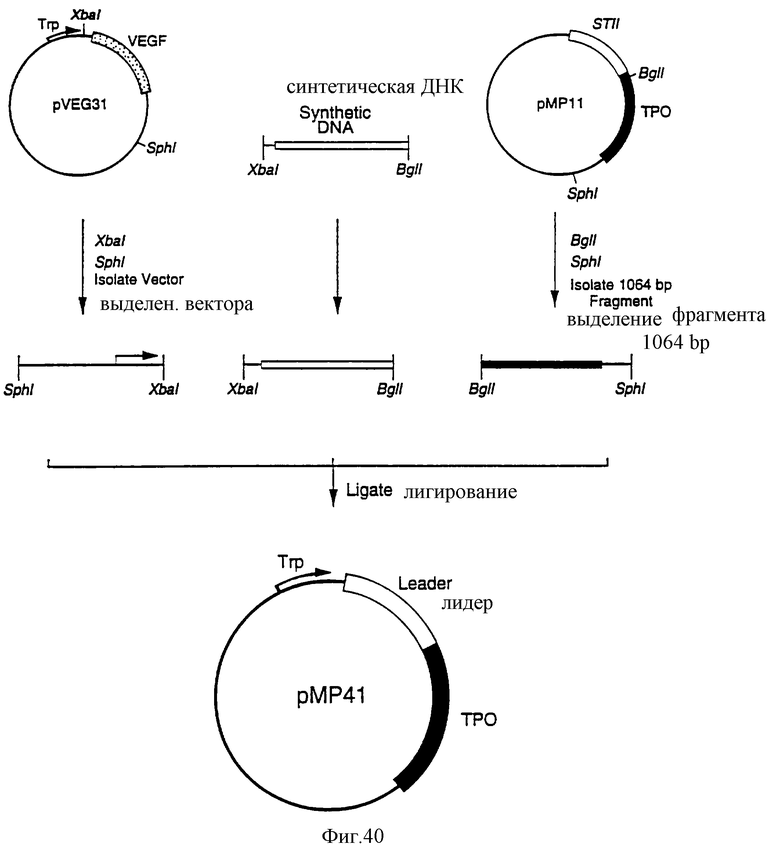
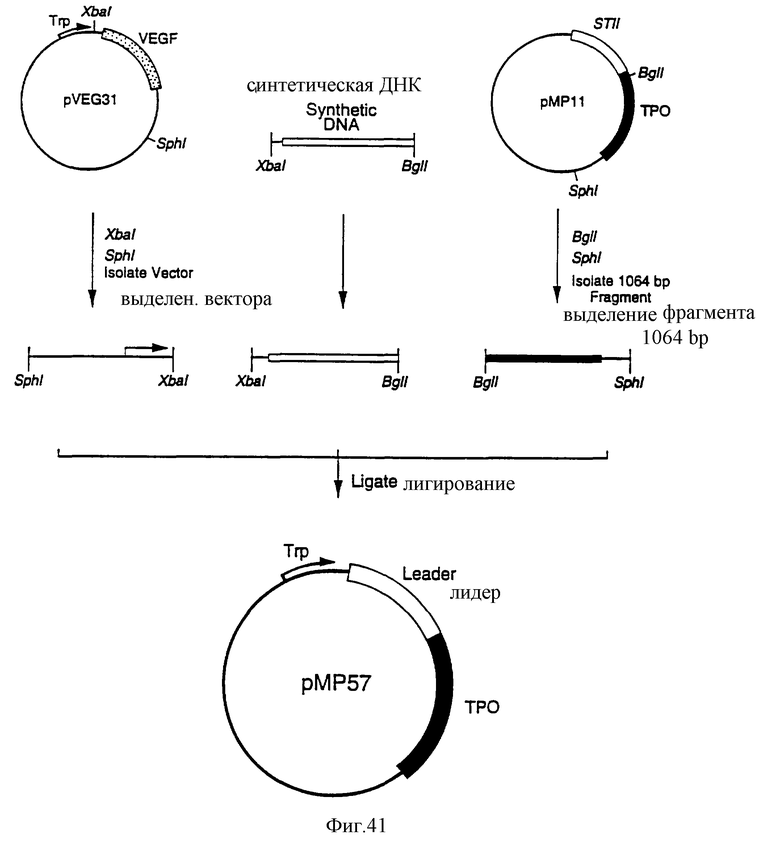
Изобретение относится к молекулярной биологии и может быть использовано в медицине. Описан существенно гомогенный полипептид лиганд mpI, представляющий собой полипептидный фрагмент формулы X-hTPO-Y, где hTPO имеет аминокислотную последовательность фрагментов человеческого ТРО (hML), X - аминоконцевая аминогруппа или аминокислотный(е) остаток(ки), Y - карбоксиконцевая карбоксигруппа или аминокислотный(е) остаток(ки), или химерный полипептид, или фрагмент полипептида, содержащий N-концевые остатки аминокислотной последовательности hML. Изобретение охватывает также НК, кодирующую полипептид, экспрессирующий вектор, включающий НК. Описаны способы получения полипептида с использованием клетки-хозяина, трансформированного вектором, и антитела к полипептиду. Описанны методы и средства с использованием активных агентов данного изобретения. Полипептид лиганд mpI влияет на репликацию, дифференциацию или созревание клеток крови, особенно мегакариоцитов и прогениторных клеток мегакариоцитов, что позволяет использовать полипептиды по изобретению для лечения тромбоцитопении. 7 с. и 14 з.п. ф-лы, 42 ил., 15 табл.
(а) полипептидного фрагмента, представленного формулой
X-hTPO-Y
где hTPO выбран из группы, состоящей из аминокислотной последовательности человеческого ТРО (hML) от Ser1 до Arg245, от Ser1 до Arg153 и от Cys7 до Cys151 включительно;
Х представляет собой аминоконцевую аминогруппу или аминокислотный(ые) остаток(ки), выбранные из группы
М,
МА,
МРА,
MPPA(SEQ ID NO: 117),
МАРРА (SEQ ID NO: 118),
MPAPPA (SEQ ID NO: 119),
MSPAPPA (SEQ ID NO: 120),
A,
PA,
PPA,
APPA (SEQ ID NO: 121),
PAPPA (SEQ ID NO: 122),
SPAPPA (SEQ ID NO: 123),
Y представляет собой карбоксиконцевую карбоксигруппу или аминокислотный(ые) остаток(ки), выбранные из группы
V,
VR,
VRR,
VRRA (SEQ ID NO: 124),
VRRAP (SEQ ID NO: 125),
VRRAPP (SEQ ID NO: 126),
VRRAPPT (SEQ I D NO: 127),
VRRAPPTT (SEQ ID NO: 128),
VRRAPPTTA (SEQ ID NO: 129),
VRRAPPTTAV (SEQ ID NO: 130),
VRRAPPTTAVP (SEQ ID NO: 131),
VRRAPPTTAVPS (SEQ ID NO: 132),
VRRAPPTTAVPSR (SEQ ID NO: 133),
VRRAPPTTAVPSRT (SEQ ID NO: 134),
VRRAPPTTAVRSRTS (SEQ ID NO: 135),
VRRAPPTTAVRSRTSL (SEQ ID NO: 136),
VRRAPPTTAVRSRTSLV (SEQ ID NO: 137),
VRRAPPTTAVRSRTSLVL (SEQ ID NO: 138),
VRRAPPTTAVRSRTSLVLT (SEQ ID NO: 139),
VRRAPPTTAVRSRTSLVLTL (SEQ ID NO: 140),
VRRAPPTTAVRSRTSLVLTLN (SEQ ID NO: 141),
VRRAPPTTA VRSRTSLVLTLNE (SEQ ID NO: 142),
VRRAPPTTAVRSRTSLVLTLNEL (SEQ ID NO: 143),
VRRAPPTTAVRSRTSLVLTLNELP (SEQ ID NO: 144),
аминоконцевую аминокислотную последовательность, содержащую один или несколько остальных остатков человеческого ML, представленных на фиг.1 (SEQ ID NO: 1), и его конъюгированной с ПЭГ формы;
(б) химерного полипептида, содержащего лиганд mpl, аминокислотная последовательность которого содержит аминокислотные остатки от 1 до Х на фиг.1 (SEQ ID NO: 1), где X представляет собой аминокислотный остаток в положении 153, 155, 164, 174, 191, 205, 207, 217, 229, 245 или 332, слитый с гетерологичным полипептидом; и
(в) полипептида, содержащего N-концевые остатки с 1 приблизительно по 153-157 hML, замещенного остатком человеческого ЕРО, добавленным или заменяющим N-концевые остатки hML в положениях, выбранных из группы, включающей 1, 3-6, 9, 13-15, 17-28, 30, 32-34, 36-41, 43-50, 53-59, 61-68, 71, 73, 75-80, 82-88, 90-96, 98-99, 102-103, 105-107, 110, 112-113, 115-119, 121-130, 132-133, 135, 137-140, 143-150 и 152-156 в соответствии с выравниванием, приведенным на фиг.10;
причем указанный полипептид стимулирует включение меченых нуклеотидов (3H-тимидина) в ДНК IL-3-зависимых клеток Ba/F3, трансфицированных человеческим mpl P; или стимулирует включение в циркулирующие тромбоциты 35S.
SPAPPACDLRVLSKLLRDSHVL (SEQ ID NO: 110),
HSRLSQCPEVHPLPTPVLLPAVDF (SEQ ID NO: 111),
SLGEWKTOMEETKAODILGAVTL (SEQ ID NO: 112),
LLEGVMAARGQLGPTCLSSLL (SEQ ID NO: 113),
GQLSGQVRLLLGALQS (SEQ ID NO: 114),
LLGTQLPPQGRTTAHKDPNAIF (SEQ ID NO: 115) или
LSFQHLLRGKVRFLMLVGGSTLCVR (SEQ ID NO: 116)
или их комбинации.
| Mc DONALD “Trombopoetin: ITS biology, purification and characterization”, J.Exper | |||
| hematology, 1988, v.6, №3, p.201-205 | |||
| Mc DONALD et al | |||
| "Studies on the purification of trombopoetin from kidney cell culture medium", J | |||
| of Laboratory and clinical medicine, 1985, v.106, №2, p.162-174. |

