Настоящее изобретение относится к биологически совместимому материалу для использования в трансплантатах, а также к получению и применению такого материала.
Пересадка недостающей или поврежденной животной (особенно человеческой) ткани, включая органы, в последние четыре десятилетия приобрела характер обычной терапии в клинической медицине. Эти методы пересадки варьируются, например, от использования полиэтилентерефталата, поставляемого под товарным знаком ДАКРОН фирмой Дюпон, для замены поврежденных кровеносных сосудов до использования подкожной вены ноги в качестве аутотрансплантата к артериям, блокированным по обходу, и до трансплантации сердца от одного человека другому.
Трансплантация органов подверглась значительному развитию с получением иммунодепрессантов, позволяющих достичь высоких результатов при относительно умеренной стоимости. Быстро возросла необходимость в пересадке органов. В настоящее время во всем мире осуществляется более 20000 пересадок органов в год, что, однако, составляет только около 15% от необходимого количества, как оценивается по текущим критериям. Отношение предложение/спрос донорских органов всех типов нельзя решить на основе существующих источников. Возможно, это наилучшим образом иллюстрируется на примере удовлетворения потребности в пересадке сердца. Первая пересадка сердца, осуществленная доктором Барнардом в 1967 году, вызвала громадный интерес во всем мире. В течение года в 22 странах была осуществлена 101 пересадка сердца 64 различными хирургическими бригадами. Разочарование, последовавшее после получения плохих результатов, привело к тому, что к началу 1970 годов во всем мире осуществлялось менее 30 пересадок в год. Однако введение в практику иммунодепрессии циклоспоринами революционизировало пересадку сердца так, что большинство хирургических центров теперь ожидает успешных результатов в пересадке сердца из числа более чем 80% трансплантатов в год (и выживаемости пациентов). По мнению экспертизы, этот процент выживаемости может быть повышен и далее. Успех данной методики, несомненно, предопределил то, что профессиональные медики и публика вообще стали более уверенными в том, что пересадка сердца представляет реальную альтернативу смерти, и все большее число пациентов подвергается данной методике. В настоящее время в мире ежегодно осуществляется свыше 2000 пересадок сердца.
Сегодня наибольшая вероятность смерти при пересадке сердца приходится на период ожидания пригодного донорского органа. Если искусственное сердце позволяет получить для таких пациентов аппараты краткосрочной помощи, то аппараты длительного пользования предполагают наличие большего числа центров по пересадке сердца и доноров. Потенциальное количество людей, которым помогла бы выжить пересадка сердца, никогда не устанавливалось научными методами, однако опубликованные оценки необходимости в пересадке сердца варьируются в широких пределах от 50 до 250 миллионов человек в год в зависимости от выбранных критериев, возраста реципиента, заболевания и так далее. Какой бы ни была фактическая цифра, уже совершенно ясно, что существующее в настоящее время количество доноров не в состоянии решить данную потребность. Аналогичные замечания можно привести и в отношении пересадки почек и печени и весьма вероятно, что вскоре, в результате широко применяемой теперь терапевтической методики трансплантации поджелудочной железы или островка Лагерганса для лечения диабета, первейшей заботой станет сокращение и этой ткани.
Существуют и другие недостатки, ассоциируемые с современным состоянием трансплантационной терапии. Конечно, донорские органы не всегда пригодны для использования при пересадке, так как многие доноры сами являются жертвами какого-то несчастного случая (например, дорожной аварии), который вызвал смерть в результате повреждения определенного органа, иного, нежели тот, который подлежит трансплантации; трансплантируемый орган может быть поврежден в результате иного случая или связанной с этим трудности.
Кроме того, из-за непредсказуемости в пригодности донорских органов терапию трансплантатами часто нельзя планировать в качестве обычной операции, И слишком уж часто хирургические бригады и администрация больниц должны реагировать на момент идентификации донорского органа и заниматься несоциальной деятельностью, что добавляется к административным и персональным трудностям.
В отношении трансплантатов сердца, печени и легких, если происходит отторжение ткани, пересадку обычно нельзя осуществить до тех пор, пока за короткий промежуток времени не появится другой пригодный донор.
Помимо вышеприведенных медицинских трудностей, существующая практика трансплантации в некоторых случаях ассоциируется с социальными затруднениями. Во-первых, могут быть религиозные возражения в пересадке органов от потенциальных доноров, в частности у народов, верящих в перевоплощение. Несомненно, существует целый ряд других этических и социальных трудностей, встречающихся при пересадке органов от умерших людей, в частности в некоторых странах, где требуется согласие на подобную операцию. Наконец, вызывает беспокойство появление коммерческой торговли донорами с живой почкой, в особенности в некоторых регионах третьего мира, и было бы желательно запретить или ослабить такую торговлю.
Как подчеркивалось выше, общепринятая трансплантационная хирургия со всеми ее недостатками заключается в пересадке органов от одного животного определенного вида (как правило, человека) другому животному этого же вида. Такие трансплантации называются аллотрансплантатами. Из-за трудностей в получении общепринятого аллотрансплантата, как отмечается выше, внимание должно быть уделено возможности использования ксенотрансплантатов при пересадке. Ксенотрансплантация является общим термином, используемым для имплантации тканей, включая клетки и органы, через видовые барьеры.
Уже есть несколько примеров успешного использования ксенотрансплантатов в терапевтической пересадке. Например, последние годы свидетельствуют об использовании свиной ткани для замены аортального клапана, свиной кожи для лечения пациентов с сильными ожогами, а также пупочной вены коровы в качестве трансплантата для замены вен. Все эти ксенотрансплантаты имеют одну общую особенность: они обеспечивают только механическую замену ткани. Однако используемая ткань биологически является нефункциональной. Причиной этого является то, что иммунные процессы, происходящие в человеке, немедленно (в течение минут или часов) разрушают целлюлярную целостность тканей от большинства видов. Такие ксенотрансплантаты известны как дискордантные ксенотрансплантаты.
Свирепость такого разрушения является филогенетически ассоциированной. Так, ткань от шимпанзе, примата, являющегося близким родственником человека, может сохраняться в человеке аналогичным образом, что и аллотрансплантат; такой ксенотрансплантат известен как конкордантный ксенотрансплантат.
Хотя можно подумать, что конкордантные ксенотрансплантаты могут ответить на вопрос относительно трудностей с аллотрансплантатами, на практике это не совсем так. Шимпанзе намного мельче человека, а органы шимпанзе, как правило, не функционируют в человеке. В отношении почек данную проблему можно решить за счет трансплантации почек других шимпанзе с целью замены поврежденных почек человека, однако в отношении печени и сердца это невозможно. Кроме того, шимпанзе медленно размножаются в природе и плохо вылавливаются, а потребность в шимпанзе в качестве экспериментальных животных (в особенности в настоящее время, для исследования синдрома приобретенного иммунного дефицита (СПИД)) означает, что вновь спрос обгоняет предложение. Кроме того, может существовать некая социальная трудность, связанная с общественным мнением относительно использования других приматов в качестве доноров ксенотрансплантатов.
Поэтому внимание вновь обращается на дискордантные ксенотрансплантаты. Вообще полагают, что причиной того, почему дискордантные ксенотрансплантаты так быстро терпят неудачу, является существование в реципиентных видах "встречающихся в природе" антител против еще не определенных антигенов донорских видов (Shons et al., (Europ.Surg.Res., 5, 26-36 1973)). Антитела названы "встречающимися в природе", поскольку они присутствуют в живых существах, которые не имеют никакого иммунологического признака от донорского вида.
Быстрое отторжение, известное как гиперострое антитело -опосредованное отторжение трансплантата органа, хорошо документировано. В начале 60-х годов, когда обычным лечением стала пересадка почек, было замечено, что трансплантированные почки время от времени отторгались реципиентом, хотя операция все еще продолжалась. Во время операции по пересадке почка, как правило, становится красной и твердой по консистенции вскоре после того, как сшиты сосуды реципиента и донора. Такие трансплантаты часто вырабатывают мочу почти сразу же. В форме отторжения, когда трансплантат разрушается в то время, как пациент все еще находится на столе (гиперострое отторжение), деструктивные процессы начинаются в первые несколько минут или вскоре после трансплантации. Когда это происходит, почка становится голубоватой и пятнистой, а затем застойной. Консистенция органа также изменяется. Как правило, трансплантат становится отечным, не происходит выработки мочи и вновь трансплантированная почка затем немедленно отторгается. Стало ясно, что в данный процесс вовлечена гуморально-опосредованная иммунологическая реакция между предварительно образованными циркулирующими антителами в реципиенте и антигенами в донорской почке. Единственным путем устранения данной проблемы при аллотрансплантации является проверка перед трансплантацией того, что никаких антител не существует в реципиенте против донорских клеток. С увеличением знания тестирования на такие антитела (известного как перекрестная проба) стало ясно, что эта генерализация относительно того, что антитело в реципиенте вступает во взаимодействие против антигенов в доноре, не является правильной, и что гиперострое отторжение трансплантата, когда оно вовлекает трансплантаты между отдельными пациентами одного и того же вида, ограничивается существованием специфических видов антитела, известного как положительная перекрестная проба с тепловыми лимфоцитотоксинами (лимфоцитотоксический тест); и почти несомненно, эти антитела принадлежат к подклассу иммуноглобулина Г (ИгГ). Кроме того, присутствие этих антител всегда обусловлено методикой пред существующей сенсибилизации либо в результате ранних переливаний крови, либо в результате беременности, либо, наиболее часто, в результате предшествующего повреждения трансплантата.
Полагают, что механизм гиперострого отторжения ксенотрансплантата аналогичен механизму гиперострого отторжения аллотрансплантата, как подчеркивалось выше. Обширная литература по механизму отторжения ксенотрансплантата, она охватывает около 83 лет. В течение этого периода времени только три публикации касались предположения механизма отторжения ксенотрансплантата, который не вовлекает антитела. Предположение основывалось на вовлечении в отторжение ксенотрансплантата альтернативного метаболического пути активации комплемента. Предположение впервые появилось в 1976 году в работе Schilling et al., (Surgery, Gynaecology and Obstetrics 142; 29-32 (1976)). Предположение было выдвинуто вновь в 1988 и 1989 годах (те же данные были опубликованы дважды) Miyagawa et al., (Transplantation, 46(6) 825-830 (1988) и Transplantation Proceedings, 21(1), 520-521 (1989)). Однако результаты не были убедительными, поскольку оба эксперимента испытывали, в основном, одну и ту же ошибку. Выбранная заявителем модель представляла собой модель ксенотрансплантата, в которой не существовали антитела перекрестных видов. Однако сейчас кажется, что анализы, использованные для обнаружения антител перекрестных видов, были неправильными, и что выводы, сделанные в данных работах, основывались на недостоверных данных.
Большинство шагов, предпринимаемых в настоящее время экспериментально для устранения или ослабления отторжения ксенотрансплантатов, включает химиотерапевтическое вмешательство в иммунную систему реципиента, в значительной степени на неспецифической основе, например с использованием циклоспорина А и других иммунодепрессантов, путем плазмофореза, путем обработки фактором яда кобры, абсорбции антитела белком A Staphylococcus и так далее. Этот подход естественно следует из химиотерапии, которая поддерживает аллотрансплантаты.
Настоящее изобретение предполагает совершенно другой подход: вместо неспецифического вмешательства в иммунную систему реципиента изобретение позволяет осуществить совместное введение вещества, которое оказывает воздействие на донорский трансплантат, рассматриваемый как "свой", посредством определенных компонентов иммунной системы реципиента. В особенно предпочтительных вариантах изобретения саму донорскую ткань модифицируют до появления иммунологичности к реципиенту, которому в некоторых отношениях надлежит быть "самим".
Также обнаружено, что гиперострое отторжение ксенотрансплантата необязательно опосредовано антителами. Этот вывод возникает на основе двух наблюдений. Во-первых, в отсутствие антитела, но в присутствии комплемента, наблюдается гиперострое отторжение; во-вторых, в присутствии антитела, но в отсутствие комплемента, не наблюдается никакого гиперострого отторжения.
Изобретение основано на обнаружении того, что активация комплемента играет главную роль в гиперостром отторжении ксенотрансплантата, независимо от того, сопровождается или нет такая активация связыванием молекул соответствующих антител. Активацию альтернативного метаболического пути комплемента можно индуцировать целым рядом клеточных продуктов. Эти продукты не ограничиваются чужеродными клетками, такими как бактерии или ксенотрансплантаты, однако существуют на многих клетках. Так, в принципе, многие клетки человека могут активировать альтернативный метаболический путь комплемента, вызывая обширное автоиммунное отторжение. Если это не происходит, то от того, что существует целый ряд белков, сводящих на нет комплемент, присутствующих в естественном состоянии в сыворотке и на поверхности клеток. Эти молекулы (упоминаемые как "гомологичные факторы рестрикции комплемента") предотвращают полную активацию само-комплемента либо путем классического, либо путем альтернативного метаболического пути под действием продуктов собственно клеток, тем самым препятствуя автоиммунному отторжению "своего". Функционирование таких молекул иллюстрируется при ночной пароксизмальной гемоглобинурии. При таком заболевании "мембранный якорь" по крайней мере одной из этих молекул (фактор распада) отсутствует. Таким образом, белок не удерживается в мембране эритроцитных клеток и отсоединяется от клетки, активируя альтернативный метаболический путь комплемента, после чего лизируется, вызывая тем самым гемоглобинурию.
В соответствии с первым вариантом настоящего изобретения предлагается способ трансплантации животной ткани в реципиент, причем ткань имеет происхождение от донора отличающегося от реципиента вида, донорский вид представляет собой дискордантный вид в отношении реципиента, отличающийся тем, что трансплантируют ткань в реципиент и получают в сочетании с трансплантируемой тканью один или более гомологичных факторов рестрикции комплемента, активных в реципиентном виде, для предотвращения полной активации комплемента.
Слово "ткань", употребляемое в данном описании, означает любой биологический материал, подлежащий трансплантации, и включает органы (особенно внутренние жизненные органы, такие как сердце, легкое, печень и почку, поджелудочную железу и щитовидную железу), роговицу, кожу, кровеносные сосуды и другую соединительную ткань, клетки, включая кровяные и гемопоэтические клетки, островки Лангерганса, клетки мозга и клетки от эндокринных и других органов, а также общие воды организма (например, плазменно-белковый консервант), и все они могут быть кандидатами для трансплантации от одного вида другому.
"Дискордантный вид" - это вид (как правило, васкуляризованный), ксенотрансплантат которого в реципиент обычно дает начало гиперострому отторжению, то есть отторжению в пределах минут или часов, а не дней (Calne, Transplant. Proc., 2:550, 1970). Такие гиперострые отторжения должны быть хорошо известны специалистам в данной области знания и могут иметь место в пределах 24 часов, в пределах 6 часов или даже в пределах одного часа после трансплантации.
Комплемент и его активация в настоящее время хорошо известны и описаны в работе Roitt, Essential Immunology (Fifth Edition, 1984) Blackwell Scientific Publications, Oxford. Активность, приписываемая комплементу (С’), зависит от функционирования девяти белковых компонентов (от С1 до С9), действующих последовательно, из которых первый состоит из трех основных субфракций, называемых C1q, C1r и C1s. Комплемент может быть активирован классическим или альтернативным метаболическим путем, причем оба пути будут описаны вкратце.
При классическом метаболическом пути антитело связывается с С1, субъединица C1s которой приобретает эстеразную активность и вызывает активацию и перенос к сайтам на мембране или иммунном комплексе вначале С4, а затем С2. Этот комплекс обладает активностью "С3-конвертазы" и расщепляет С3 в растворе с получением маленького пептидного фрагмента С3а и остаточной молекулы С3b, которые имеют совершенно разные функции. С3а обладает активностью анафилатоксина и не играет никакой другой роли в каскаде амплификации комплемента. С3b является мембранно-связанным и может вызывать иммунную адгезию комплекса антиген-антитело-С3b, тем самым облегчая последующий фагоцитоз.
При альтернативном метаболическом пути активность конвертазы С3 представлена благодаря С3bВ, активацию которого можно запустить внешними агентами, в частности микробными полисахаридами, такими как эндотоксин, действующий независимо от антитела. Конвертазу образуют под действием Фактора I на комплексе С3b и Фактора В. Этот комплекс образует петлю с положительной обратной связью, в которой продукт распада С3 (С3b) помогает образовать более чем один фермент расщепления.
Как в классическом, так и в альтернативном метаболическом пути уровень С3b поддерживают под действием инактиватора С3b (Фактора I). С3b легко объединяется с Фактором Н с образованием комплекса, который разрушается Фактором I и теряет свои гемолитические и иммунно-адгезивные свойства.
Классический и альтернативный пути являются общими после осуществления стадии С3. С5 расщепляется с получением фрагментов С5а и С5b. С5а обладает активностью анафилатоксина и дает начало хемотаксису полиморфно-ядерных нейтрофилов. С5b связывается в виде комплекса с С6 и С7, образуя термоустойчивый сайт на мембране, который пополняет конечные компоненты С8 и С9 с образованием мембранно-атакующего комплекса (MAC). Он представляет собой кольцевую структуру, вставленную в мембрану и простирающуюся из нее, которая образует трансмембранный канал, полностью проницаемый электролитами и водой. Благодаря высокому внутреннему коллоидно-осмотическому давлению происходит результирующий приток ионов натрия и воды, что приводит к лизису клеток.
Гомологичные факторы рестрикции комплемента (HCRFS), пригодные в настоящем изобретении, могут вообще интерферировать с любой частью каскада активации комплемента. HCRFS может интерферировать исключительно с той частью, которая составляет классический метаболический путь, или исключительно с той частью, которая составляет альтернативный метаболический путь, или, более обычно, может интерферировать с той частью, которая является общей как для классического, так и для альтернативного метаболических путей. Предпочтительно, чтобы регулятор HCRFS интерферировал с общей частью пути метаболизма. HCRFS может быть идентичен натуральному HCRF или может просто иметь соответствующую функцию. Синтетические и полусинтетические HCRFS, включая полученные методами рекомбинантных ДНК, а также их варианты, включены в понятие термина HCRFS.
Как упомянуто выше, гомологичные факторы рестрикции комплемента представляют собой вещества, которые регулируют действие комплементного каскада таким образом, чтобы понизить или предотвратить его литическую активность; они используются животным телом для мечения ткани как "своей" с тем, чтобы можно было избежать аутоиммунной реакции. В предлагаемом изобретении в принципе можно, чтобы HCRFS был либо мембранно-связанным, или свободным в сыворотке, хотя на практике было бы предпочтительнее иметь HCRFS мембранно-связанным на клетках ткани ксенотрансплантата. Таким путем этому фактору легче быть "в ассоциации" с тканью трансплантата. Предпочтительные HCRFs включают путативные факторы клеточной мембраны, включая рецептор C3b/C4b (CR1), рецептор С3.dg (CR2), стимулятор гемолиза (фактор распада, DAF), инактиватор С3b и белок мембранного кофактора (МСР). Путативные сывороточные HCRF включают Фактор Н, стимулятор гемолиза (DАF)и С4-связующий белок (C4bp). Эти HCRFs все регулируют вниз активность комплемента путем интерференции на стадии С3. Гомологичный фактор рестрикции (HRF), который блокирует у С8, также является путативным мембранным фактором.
Многие, но не все, гены для пригодных HCRFS расположены в RCA (регулятор активации комплемента), локус которого картирует с диском q32 хромосомы 1 (Rey-Campos et al., J.Exp Med. 167: 664-669 (1988)).
Хотя существует некоторая путаница с номенклатурой и расположением HCRFs, факторы С4ВР, CRI, DAF и фактор Н идентифицированы Rey-Campos et al., (выше), а также в их более ранней работе (J.Exp.Med, 166: 246-252 (1987)). Белок мембранного кофактора (МСР) обрабатывают как синоним С4-связующего белка (С4bр), и может быть, что эти два фактора являются либо родственными, либо идентичными. Pother и Till ("The Complement System", Springer-Verlag, Berlin (1988)) приводят регуляторные факторы конвертазы С3 в разделе 1,2.3.2; они установили равенство между С4-связующим белком (С4bр) и стимулятором гемолиза, а также между фактором Н, BIH-белком и акселератором инактиватора С3b. Нет сомнения в том, что номенклатура, локализация и характеристика HCRFS будут продолжать развиваться, однако следует понять, что настоящее изобретение предполагает использование всех HCRFS в соответствии с требованием пригодности и предпочтения.
Другие отсылки на HCRFS включают следующие:
Фактор I (также известный ранее как инактиватор С3b или KAF): Tamura and Nelson (J.Immunol., 99: 582-589 (1967);
Фактор Н: Pangburn et al., (J.Exp.Med., 146: 257-270 (1977);
С4-связующий белок: Fujita et al., (J.Exp.Med., 148: 1044-1051 (1978));
DAF (известный так же как CD55): Nicholson-Weller et al., (J.Immunol., 129: 184 (1982));
Белок мембранного кофактора (МСР; также известный как CD46 и впервые описанный как qp45-70 и далее известный как qp66/56): Seya et al./ (J.Exp.Med., 163: 837-855 (1986));
CRI (также известный как CD35): Medof et al., (J.Exp.Med., 156: 1739-1754 (1982)) и Ross et al., (J.Immunol., 129: 2051-2060 (1982));
CR2 (также известный как CD21, рецептор 3d/EBV и р140: Iida et al., (J.Exp.Med., 158: 1021-1033 (1983)) и Weis et al., (PNAS 81: 881-885 (1984)).
Ниже приведено распределение ткани некоторых белков RCA:
CR1: Мембрана (ограниченная): эритроциты; моноциты; большинство клеток В и некоторые клетки Т; полиморфно-ядерные лейкоциты; фолликулярные дендритные клетки; подоциты;
CR2: Мембрана (ограниченная): большинство клеток В; фолликулярные дендритные клетки; некоторые эпителиальные клетки и несколько линий Т-клеток;
МСР: Мембрана (широкая): все периферические кровяные клетки (кроме эритроцитов); эпителиальные, эндотелиальные и фибробластные клеточные линии; трофобласт и сперма;
DAF: Мембрана (широкая): все периферические кровяные клетки; эпителиальные, эндотелиальные и фибробластные клеточные линии; трофобласт и сперма;
С4bр: Плазма: печеночный синтез;
Н: Плазма: печеночный синтез; фибробластные и моноцитарные линии клеток.
Что касается белков, вовлеченных в гомологичную рестрикцию на уровне мембраноатакующего комплекса, использование которого также предусматривается настоящим изобретением, то существует общее согласие (однако, еще не доказательство) в форме белковой последовательности, что следующие белки массой 65 кДа (или около этого) являются идентичными: С8-связующие белки (Schonermark et al., J.Immunol., 136: 1772 (1986)); гомологичный фактор рестрикции (HRF) (Zaiman et al., Immunology, 83: 6975 (1986)), а также МАС-ингибирующий белок (MIP) (Watts et al., (1988)).
Белок C8bp/HRF/MIP присоединен к поверхности клетки при помощи гликолипидного якоря, как и CD59 и DAF: известно, что эти белки функционально отсутствуют при ночной пароксизмальной гемоглобинурии.
Белок массой 18-20 кДа также вовлечен на уровне MAC. Полагают, что следующие белки являются идентичными (а может быть и нет):
P-18 (Sudita et al., (J.Biochem., 104: 633 (1988));
HRF - 20 (Okada et al.,(Intl.Immunol., 1 (1989));
CD59 (Davies et al., (J.Exp.Med., (сентябрь 1989)));
а также мембранный ингибитор реакционноспособного лизиса (MIRL) (Hologuin et al., (J.Clin.Invest., 84: 7 (1989))).
Доказательством путативной идентичности этих белков является то, что белковые и/или кДНК-последовательности для CD59 и HRF-20 показаны идентичными: вероятно, что они те же, что и P-18/MIRL. Следует отметить, что существует некоторая гомология последовательности CD59/HRF-20 с последовательностью мышиного LY-6 антигена, который вовлечен в активацию Т-клеток (Gronx et al., (J.Immunol., 142: 3013 (1989))).
SP-40.40 также вовлечен в регуляцию MAC (Kivszbaum et al., EMBO 8, 711 (1989)).
Предпочтительно, чтобы HCRFS вмешивался в активацию комплемента на стадии С3. Оба МСР и DAF блокируют петлю с положительной обратной связью при альтернативном метаболическом пути активации С3, и они составляют предпочтительные HCRFS.
HCRFS предлагается в ассоциации с трансплантируемой тканью. Это означает, что HCRFS вводят таким образом, чтобы ткань трансплантата была мечена как "своя", однако другой чужеродный материал, такой как поражающие бактерии, так не метят. Один или более HCRFS можно вводить в ткань трансплантата парентерально, но локально. Однако на практике это может быть непредпочтительным из-за трудности обеспечения адекватной локализации HCRFS на ткани трансплантата, а также из-за другой трудности, заключающейся в том, что HCRFS может быть введен реципиенту неоднократно после того, как получен трансплантат, и эту трудность можно устранить за счет использования специальных систем доставки фармацевтических средств.
Вообще, было бы более удобно получить HCRFS таким образом, чтобы он интегрировал с клеточной мембраной на донорской ткани. Хотя могут быть какие-то легкие инфекции трансплантированной ткани, вызывающие пригодную экспрессию, на сегодняшний день наиболее предпочтительным путем достижения данной цели является трансгенный путь для донорской ткани, состоящий в том, что ткань содержит и экспрессирует нуклеиновую кислоту, кодирующую один или более HCRFS, активных в реципиентном виде, когда она трансплантирована в реципиент. Такая трансгенная ткань может продолжать экспрессию HCRFS неограниченное время. Генетически HCRFS может происходить от реципиентного вида или, менее предпочтительно, от близкого вида, для которого возможны конкордантные ксенотрансплантаты.
Хотя, в принципе, трансгенная донорская ткань может происходить от культуры клеток, предпочтительно, чтобы донорская ткань происходила от трансгенного животного. Трансгенное животное должно экспрессировать (или быть способным экспрессировать) HCRFS по крайней мере в трансплантируемой ткани. Однако даже и это неважно, поскольку существует возможность привязать HCRFS к клеточным мембранам донорской ткани с помощью определенного связующего (такого, как гибридное моноклональное антитело (Milstein и Cuello, Nature, 305: 537 (1983)) или рецептора.
Реципиентный вид является главным образом человеческим, но не исключительно. Другие приматы также могут быть пригодными реципиентами, как и многие другие виды, если позволяют соображения экономики и этики.
Донорским видом может быть любой пригодный вид, который отличается от реципиентного вида и который, принимая во внимание физиологию реципиентного вида, способен продуцировать приемлемую для трансплантации ткань. Для человеческих реципиентов пригодны свиные доноры, хотя пригодны и другие виды.
В соответствии со вторым вариантом настоящего изобретения предлагаются трансплантируемые животные клетки или ткань донорского вида, причем клетки или ткань ассоциируются с одним или более гомологичными факторами рестрикции комплемента, активными в искомом реципиентном виде, для предотвращения полной активации комплемента, при этом донорский вид является дискордантным видом относительно реципиентного вида.
В соответствии с третьим вариантом настоящего изобретения предлагается трансгенное животное, имеющее трансплантируемую ткань, которая не позволяет произойти отторжению ксенотрансплантата при трансплантации в иммунную систему по крайней мере одного дискордантного вида. Дискордантный вид - это такой вид, который обычно гиперостро отторгает ксенотрансплантат из животного.
Следовательно, настоящее изобретение предусматривает использование животной ткани, полученной от донорского вида, и одного или более гомологичных факторов рестрикции комплемента, активных в реципиентном виде, причем донорский вид является дискордантным видом относительно реципиентного вида, при получении ткани, трансплантируемой в реципиентный вид.
В соответствии с четвертым вариантом настоящего изобретения предлагается трансгенное животное, имеющее клетки, способные экспрессироватъ гомологичный фактор рестрикции комплемента другого вида. Гомологичный фактор рестрикции, как правило, активен в виде, являющемся дискордантным относительно вида трансгенного животного. Клетки могут быть от одной определенной ткани, предпочтительно описанной со ссылкой на первый вариант изобретения, или от более чем одной, или от всех тканей, и в этом случае животное может стать донором более чем одной ткани. Такое трансгенное животное может рассматриваться как коллекция нетрансформированных (в смысле непролиферативных) клеток.
В соответствии с пятым вариантом настоящего изобретения предлагается нетрансформированная клетка животного, способная экспрессировать один или более гомологичных факторов рестрикции комплемента, активных в виде, являющемся дискордантным относительно клетки животного.
В соответствии с шестым вариантом настоящего изобретения предлагается рекомбинантная ДНК, включающая ДНК, кодирующую по крайней мере один гомологичный фактор рестрикции комплемента, и одну или более последовательностей с тем, чтобы кодирующая ДНК была экспрессирована нетрансформированной животной клеткой. Животной клеткой может быть клетка трансгенного животного, генетически инкорпорирующего конструкцию. В качестве альтернативы клеткой может быть культивированный орган дли другая ткань, как например, островок Лагерганса.
В соответствии с седьмым вариантом настоящего изобретения предлагается генетическая конструкция, пригодная для инкорпорирования в генетический материал животного, с получением трансгенного животного, конструкция включает ДНК, кодирующую по крайней мере один гомологичный фактор рестрикции комплемента и одну или более последовательностей, с тем, чтобы кодирующая ДНК была экспрессирована по крайней мере в некоторых клетках трансгенного животного, генетически инкорпорирующего конструкцию. Такая генетическая конструкция может быть в форме мини-хромосомы, известной как YAC. Как указано выше, гомологичный фактор рестрикции комплемента является активным, как правило, в виде, дискордантном относительно вида трансгенного животного.
В соответствии с восьмым вариантом настоящего изобретения предлагается способ получения трансгенного животного, включающий инкорпорирование в генетический материал животного ДНК, кодирующей по крайней мере один гомологичный фактор рестрикции комплемента и одну или более последовательностей для того, чтобы кодирующая ДНК экспрессировалась по крайней мере в некоторых клетках трансгенного животного.
Способы получения трансгенных животных становятся все более распространенными, и подробные стадии могут быть взяты из известных общепринятых источников. Например, в заявке РСТ WO-A-8800239 раскрываются стадии, необходимые, в принципе, для конструирования трансгенного животного.
Фактический способ инкорпорирования конструкции в клетки трансгенного животного может быть выполнен микроинъекцией, сперма-опосредованным введением или любым другим пригодным образом. Предварительные генетические манипуляции могут быть осуществлены в прокариоте, как, в основном, и предпочитается.
ДНК, кодирующую HCRFS, либо получают в форме кДНК, либо выводят с использованием традиционных методик клонирования. ДНК, кодирующая стимулятор гемолиза (DAF), вероятно наилучшим образом охарактеризована и описана в работе Medof et al., (PNAS, 84: 2007-2011 (1987)).
Физическая карта кластера генов RCA приведена в работе Rey-Campos et al., (1988) (выше). Варианты DAF и их получение методами рекомбинантных ДНК раскрываются в заявке на Европатент А-0244267; такие варианты могут быть использованы в настоящем изобретении.
Из-за лучшего представления генетики DAF и известной последовательности кДНК, кодирующей DAF, этот фактор является предпочтительным гомологичным фактором рестрикции комплемента.
Другие предпочтительные признаки 2-7 вариантов настоящего изобретения аналогичны признакам первого варианта, mutatis mutandis.
Изобретение будет теперь проиллюстрировано следующими примерами. В примерах отсылка сделана на чертежи, на которых:
фиг.1А-1Е показывают последовательные записи ЭКГ для сердца кролика, трансплантированного новорожденным поросятам в соответствии с Примером 1;
фиг.2 показывает результат радиомммуноанализа, который говорит о том, что используемые в Примере 1 свиньи не имеют значительных количеств антивидовых антител;
фиг.3 показывает некоторые стадии белкового электрофореза, используемого в Примере 4;
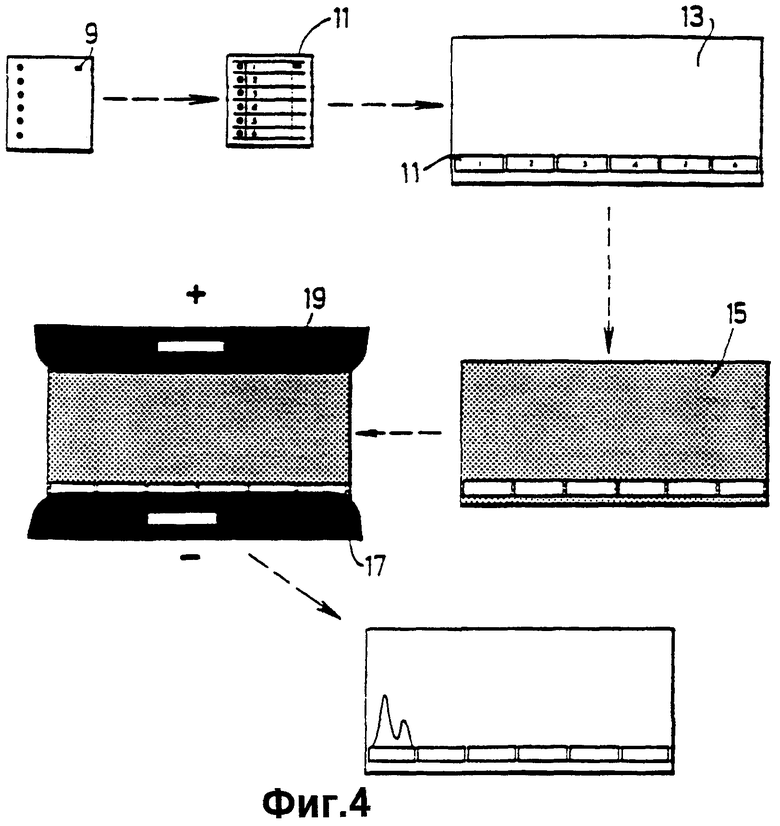
фиг.4 показывает определенные стадии двумерного перекрестного электрофореза, используемого в Примере 4;
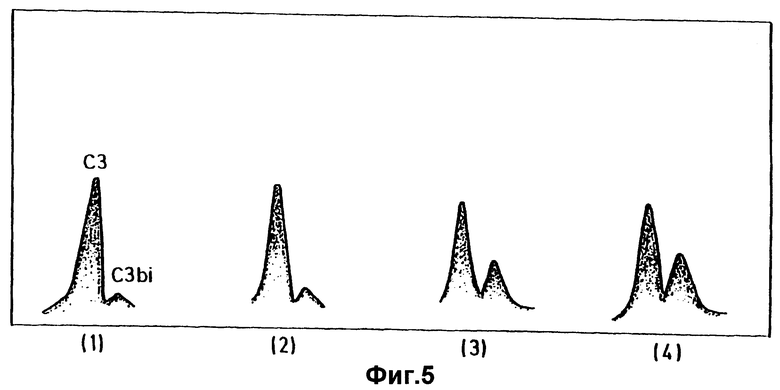
фиг.5 показывает "20-Rockets", полученные в результате проведения Примера 4 ;
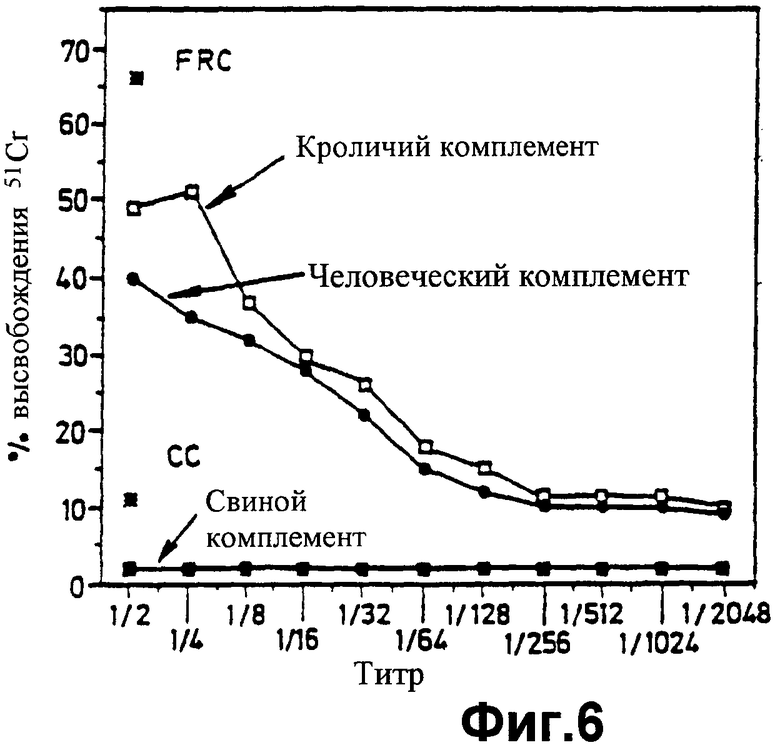
фиг.6 показывает результат анализа клеточного лизиса с радиоактивным хромом в Примере 5;
фиг.7 иллюстрирует титры литических противохомячковых антител от крысиного реципиента трансплантата сердца хомячка перед трансплантацией (день 0) и на 5, 7 и 9 дни после трансплантации, как описано в Примере 6;
фиг.8 показывает графически оптические плотности (OD) фракцией G200; гистограмма иллюстрирует титры в каждой фракции литических противохомячковых антител от крысиного реципиента сердца хомячка, как описано в Примере 6;
фиг.9 показывает блоттинг по Саузерну ДНК, экстрагированной из линий клеток Т5, b10 и DB3, как описано в Примере 7;
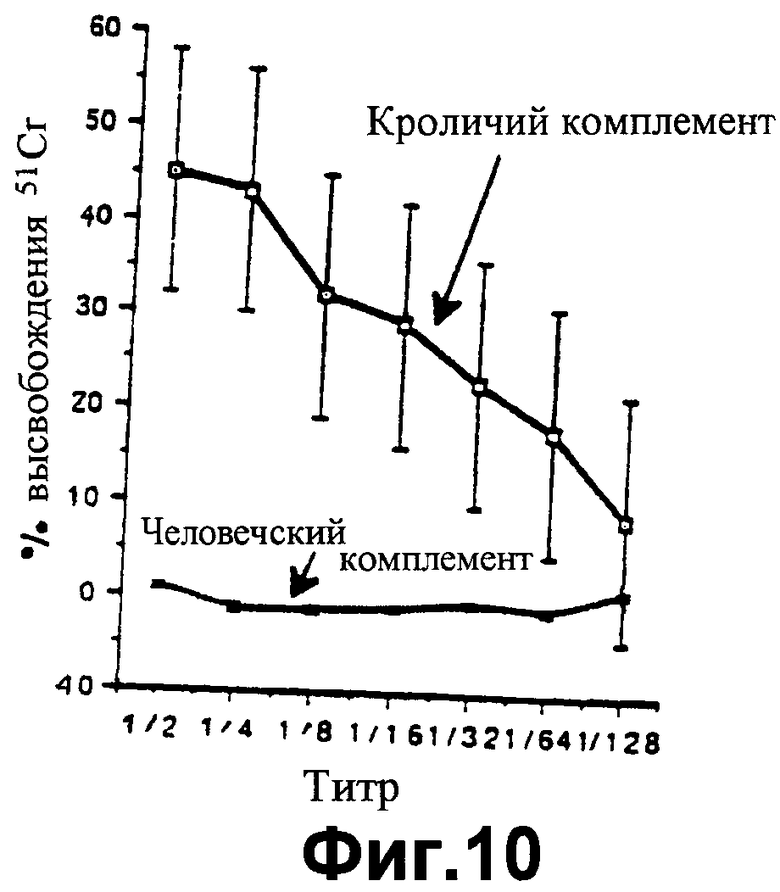
фиг.10 показывает значения высвобождения 51Сr, показательные для линии клеток Т5 человека, подверженных лизису кроличьим комплементом, но не человеческим комплементом, в присутствии свиных противочеловеческих антител, как описано в Примере 7;
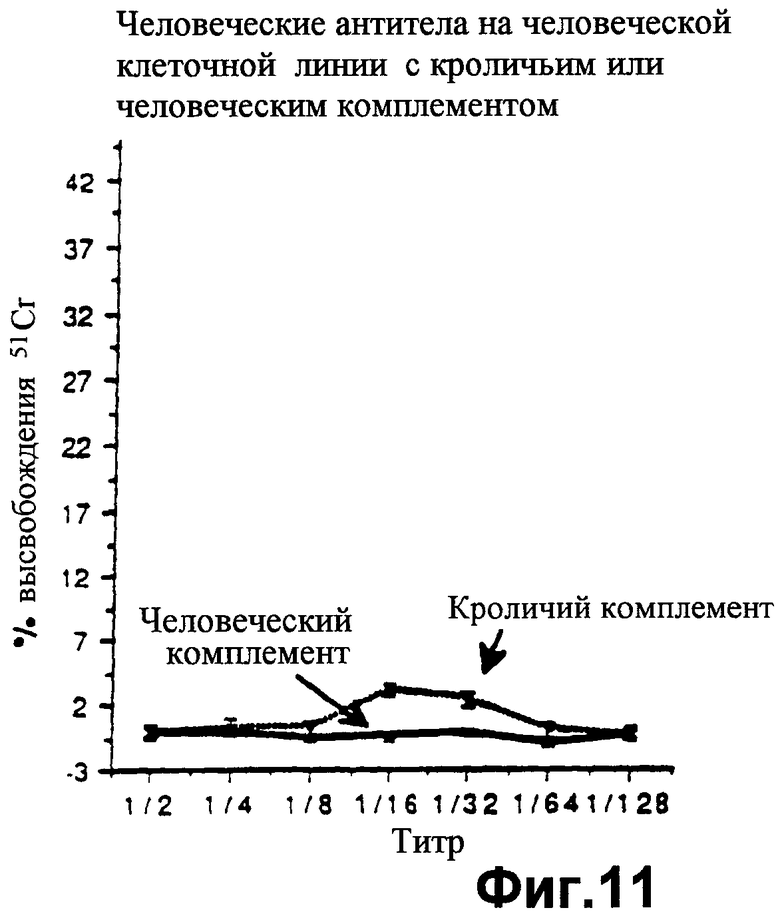
фиг.11 показывает значения высвобождения, показательные для неэффективности человеческих антител лизировать линию клеток Т5 человека либо человеческим, либо кроличьим комплементом, как описано в Примере 7;
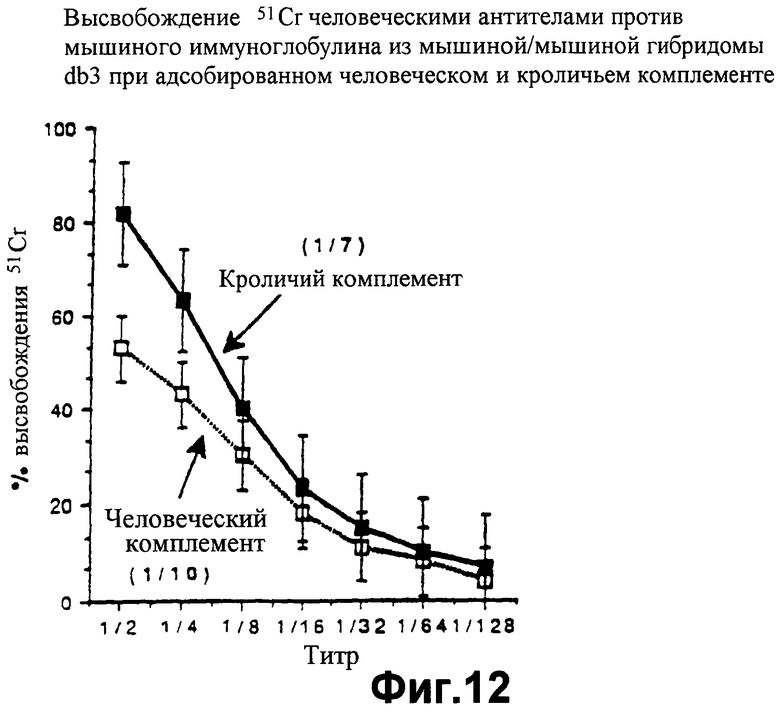
фиг.12 показывает значения высвобождения 51Сr, которые демонстрируют, что человеческие антитела могут лизировать гибридому типа "мышь-мышь" (DB3) в присутствии и кроличьего комплемента, и человеческого комплемента, как описано в Примере 7;
фиг.13 показывает высвобождение 51Сr, показывающее, что гибридная человеко-мышиная линия клеток В10 лизируется человеческими антителами в присутствии кроличьего комплемента, но не лизируется человеческими антителами в присутствии человеческого комплемента, как описано в Примере 7;
фиг.14 показывает потребление 3Н-аденина (в импульсах в минуту) клетками СНО, которое говорит о том, что эти клетки уничтожаются иммунной крысиной сывороткой в присутствии человеческого комплемента или кроличьего комплемента, как описано в Примере 8;
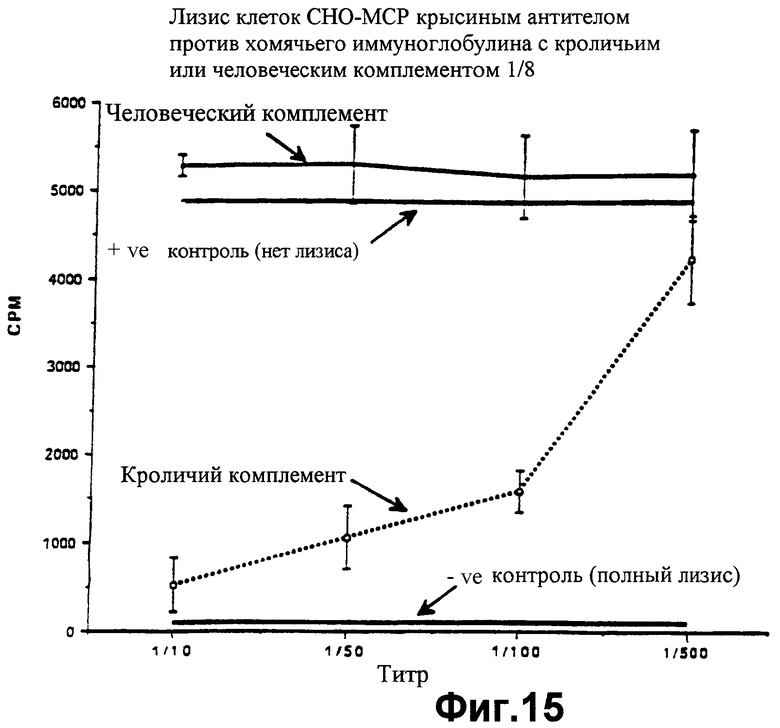
фиг.15 показывает потребление 3Н-аденина (в импульсах в минуту) клетками СНО, трансфецированными человеческим белком мембранного кофактора (МСР), которое говорит о том, что эти клетки уничтожаются иммунной крысиной сывороткой в присутствии кроличьего комплемента, однако не уничтожаются этой иммунной крысиной сывороткой в присутствии человеческого комплемента, как описано в Примере 8;
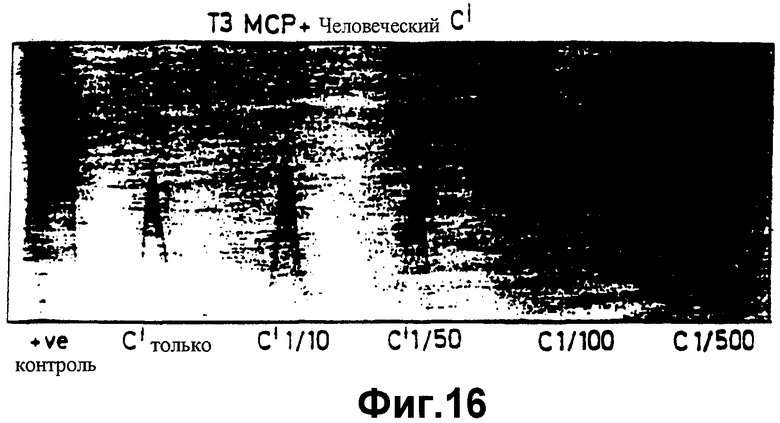
фиг.16 показывает "2D-rockets", говорящие о том, что при обстоятельствах, описанных в отношении фиг.15, компонент С3 человеческого комплемента не расщепляется с образованием С3b, как описано в Примере 8;
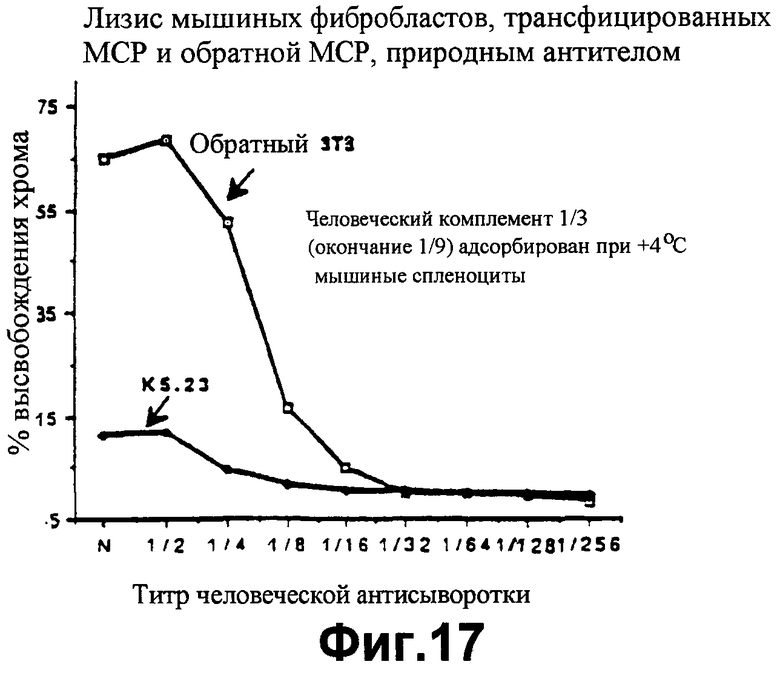
фиг.17 показывает значения высвобождения 51Сr, являющиеся показателем мышиных фибробластов 3Т3, лизируемых встречающимися в природе антителами в присутствии человеческого комплемента, и защитного эффекта экспрессии человеческого МСР мышиными клетками;
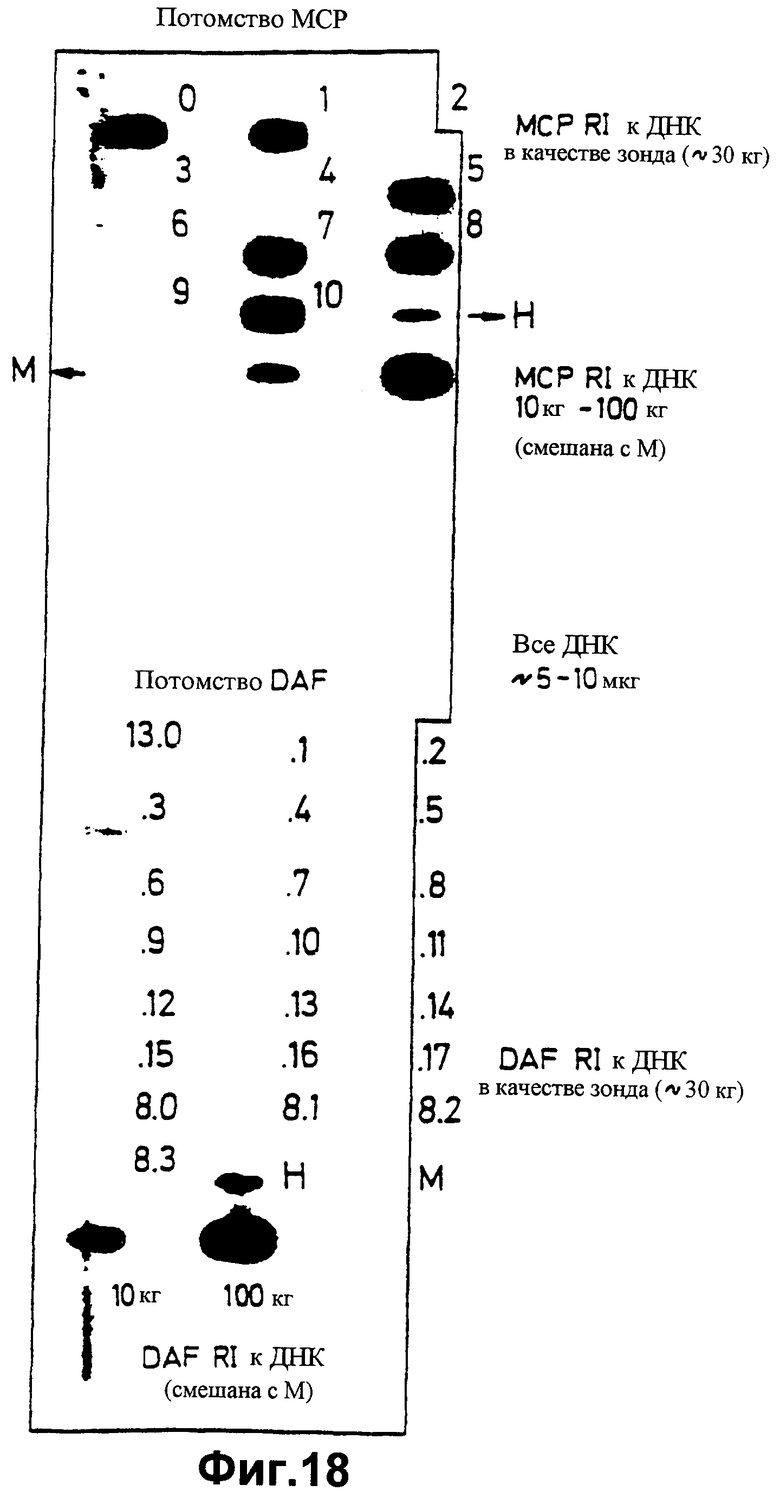
фиг.18 показывает слот-блот анализ ДНК трансгенных мышей второй генерации с использованием меченой кДНК МСР (верхней) или меченой кДНК DAF в качестве зонда.
ПРИМЕР 1. Отторжение ксенотрансплантата происходит в отсутствие любых антивидовых антител.
Вообще животные не могут выжить без циркулирующих иммуноглобулинов. Они продуцируются лимфоцитами в ответ на антигенные раздражители. В ранний период новорожденности, однако, пассивно переносимый материнский иммуноглобулин действует в качестве временной замены этого самопродуцируемого антитела. Этот пассивно переносимый иммуноглобулин придает защиту молодняку тем временем, как приобретается, ранний иммунный опыт. У млекопитающих этот пассивный перенос материнского иммнолобулина обычно происходит как трансплацентарно, так и через молозиво. У нескольких видов, однако, структура плаценты такова, что никакое материнское антитело не может переноситься этим путем. Свинья представляет собой один такой вид. Все материнское антитело получается из молозива. Таким образом, новорожденные, вскормленные грудью поросята являются в принципе неиммуноглобулиновыми.
Большие белые свиньи, отобранные во время рождения, помещаются в деревянные загоны, нагреваемые бутылями с горячей водой, и они не получают доступа к груди. Два поросенка от каждого опороса отбирают для каждого эксперимента. Эти животные весят приблизительно 1 кг после рождения.
Молодых новозеландских белых кроликов весом приблизительно 300 г используют в качестве доноров. Этих доноров анестезируют гипнолом и диазепамом, грудную клетку вскрывают и полую вену катетеризируют при помощи иглы 19 калибра. Холодовую (+4°С) кардиоплегию (Thomas №2) инфузируют до тех пор, пока сердце прекратит биться, и начинают перфузировать кардиоплегией. Охлаждение также прилагают наружно холодовой кардиоплегией непосредственно из шприца. Затем сердце кроликов извлекают с использованием хирургических методик и сохраняют в растворе для кардиоплегии при температуре +4°С. Эти меры предосторожности необходимы, так как доказано, что сердце кролика очень чувствительно к ишемическому нарушению.
Реципиентных поросят анестезируют первоначально аспирацией Галотана/02. Затем в вену молочной железы вставляют иглу для внутривенных вливаний (23 калибр), анестезию поддерживают внутривенным кетамином. Поросят одновременно гидратируют внутривенным физиологическим раствором. Пробы крови сыворотки и ЭДТК отбирают после трансплантации.
Сердце кролика трансплантируют в шею поросят в соответствии с методом Heron (Acta Pathol.Microbiol.Scand., 79:366-372 (1971)). Аорту анастомизируют концом к боку (6-0 пролин) к сонной артерии, а легочную артерию анастомизируют к яремной вене. Все другие сердечные сосуды лигируют. Сердца начинают биться в течение нескольких минут после удаления зажимов. Частоту сердечных сокращений регистрируют с помощью диаскопа/ЭКГ-монитора. Шею поросенка не закрывают в течение экспериментов, сердца сохраняют влажными, покрывая липкой пленкой.
Результаты ЭКГ приведены на фиг.1А-1Е. Кривая, приведенная на фиг.1А, показывает нормальное сердечное сокращение сразу же после трансплантации. Нарушение происходит примерно через двадцать минут (фиг.1В) и в пределах часа (фиг.1D), когда нельзя обнаружить сердечное сокращение, что говорит о гиперостром отторжении трансплантата.
Следовательно, данный пример демонстрирует то, что гиперострое отторжение дискордантного ксенотрансплантата происходит даже в отсутствие антител.
ПРИМЕР 2. Новорожденные поросята, используемые в Примере 1, не имеют никакого антивидового антитела.
Кроличий противосвиной иммуноглобулин Г (ИгГ) радиоиодируют методом Greenwood et al., Biochemical Journal 89: 114-123 (1963), модифицированным Davies и Howard (не опубликовано).
Следующие ингредиенты прибавляют в полистирольные пробирки (L P2, 6 см×1 см) в быстрой последовательности: 25-50 мкл белка (при концентрации 1 мг/мл); 3-4 мкл Na125I (100 мккуб.дюйм/мл); 10 мкл хлорамина-Т (*4 мг/мл; 0,5 М); натрий-фосфатный буфер (рН 7,5).
* должен быть приготовлен заново перед употреблением.
Эти компоненты смешивают в течение 30 секунд, постоянно перемешивая. Затем быстро прибавляют следующие компоненты: 50 мкл DL-тирозина (солевой раствор в 0,5 мл натрий-фосфатного буфера, рН 7,5); 300 мкл 2% бычьего сывороточного альбумина (БСА) /забуференного фосфатом физиологического раствора (ЗФР)/ азида.
Затем меченый белок выделяют из непрореагировавшего иода путем использования маленькой колонки (8 см×1,0 см) Сефадекса G-25, средний градиент которой выполнен в ЗФР/азиде. Смесь для реакции иодинирования количественно переносят в полученную колонку G-25 и элюируют ЗФР/азидом. Шестикапельные фракции собирают в полистироловые пробирки (L P2). Колонку элюируют до тех пор, пока оба пика белка и (125I) иода не станут элюированными, и измеряют радиоактивность во всех фракциях. Радиоактивность, инкорпорированную в белок, можно вычислить следущим образом:
величина радиоактивности в белке = первоначальное общее количество - количество в иодидных пиках.
Радиоактивно меченый ИгГ (упоминается теперь как "изотоп") затем используют для анализа на (свиные) антитела у новорожденных поросят следующим образом:
Материалы
ЗФР + 0,01% азид - оксоид;
ЗФР/БСА 1% - БСА-Сигма;
изотоп - полная молекула кроличьего противосвиного ИгГ с 12-18×103 циклами/мин;
инактивированные теплом сыворотки (56°С в течение 30 минут);
антикоагулированные кровяные пробы.
Метод
1. Получают 1% суспензию кроличьих красных кровяных клеток в ЗФР, и 100 мкл количества суспензии прибавляют в пробирки. Клетки раскручивают до супернатанта,
2. Последовательные разбавления инактивированных сывороток получают в ЗФР/БСА от взрослой свиньи (положительный контроль), новорожденного поросенка (испытуемая проба) или кролика (отрицательный контроль). 0,025 мл количества прибавляют к красным клеткам с удвоением. Пробирки инкубируют при температуре 4°С в течение 4 часов.
3. После инкубации пробирки промывают три раза в ЗФР/БСА. Затем в каждую пробирку прибавляют 0,05 мл изотопа и инкубирование продолжают в течение ночи при температуре 4°С.
4. Пробирки промывают повторно три раза и подсчеты количества в течение 1 минуты осуществляют на счетчике гамма-квантов.
5. Результаты отложены как количество циклов в минуту в зависимости от титра.
Результаты приведены на фиг.2. Кроличью сыворотку используют в качестве отрицательного контрольного значения, а сыворотку взрослой (то есть вскормленной грудью) свиньи используют в качестве положительного контрольного значения. Можно видеть, что уровень содержания свиного антитела в организме вскормленной грудью свиньи 2 сравним с уровнем в отрицательной контрольной пробе.
ПРИМЕР 3. Демонстрация релевантности комплемента С3 к разрушению ксенотрансплантата
Комплемент-дефицитных морских свинок, имеющих происхождение от линии, описанной Burger et al., (Eur.J.Iimnunol., 16: 7-11 (1986)), используют в качестве доноров для трансплантации сердца с использованием, в основном, аналогичной методики, которая описана для кроличьих-к-свиным ксенотрансплантатов в Примере 1. Крыс анестезируют эфирной аспирацией и сердца охлаждают кардиоплегией и отсекают, как описано выше. Доноров-морских свинок анастезируют валиумом и внутримышечным гипнолом. Сердца имплантируют в шею, как описано выше. Для контрольных морских свинок, то есть тех, которые имеют нормальные уровни комплемента, отторжение трансплантата обычно имеет место в пределах нескольких минут, что устраняет необходимость в закрывании шеи. У экспериментальных животных шею закрывают и сердца подвергают мониторингу двумя ежесуточными пальпациями. Нормальные ЭКГ наблюдают в течение нескольких часов после хирургического вмешательства, что показывает на отсутствие гиперострого отторжения.
ПРИМЕР 4.
А. Свиные лимфоциты и клетки почки активируют человеческий комплемент альтернативным путем метаболизма
В соответствии с методикой Grabar и Williams (Biochem. Biophys.Acta, 10: 193 (1953)) агарозные гели I вливают в 8×8 см стеклянные чашки (фиг.3). Требуется 10 мл гелевой смеси, и она состоит из 5 мл 2% агарозы и 5 мл веронального буфера (VB), (VB представляет собой 75 мМ барбитон натрия, 10 мМ ЭДТК, 10 мМ NаN3, рН 8,5). Агарозу и VB смешивают при температуре 60°С непосредственно перед использованием. Гели выливают и охлаждают на уравнительной платформе. После осаждения гель состоит из 1% агарозы и имеет глубину около 1,5 мм.
Лунки 3 диаметром 3 мм разрезают на расстоянии около 1 см от конца геля. Каждая лунка может содержать около 8 мкл пробы. Проба не имеет специального препарата, кроме добавленного бромофенолового синего для окрашивания пробы. После аппликации пробы гель осторожно помещают на платформу сосуда для электрофореза. Затем хлопчатобумажные тампоны, пропитанные в VB (буфере для образца), осторожно накладывают вдоль кромки геля, ближайшего к лункам, а другой тампон накладывают на противоположный край агарозы. (Важно гарантировать, чтобы концы тампонов были погружены в резервуары с забуференной смесью). Затем ток 25-30 мА пропускают через гель до тех пор, пока альбумин (визуализируемый с помощью связанного бромфенола) не достигнет положительного (анодного) тампона. Процесс занимает приблизительно от 2,5 часа до 3 часов. Если два или более гелей необходимо обработать одновременно и параллельно, тогда прилагаемый ток следует увеличить по мощности соответствующим образом с тем, чтобы два геля потребляли ток величиной 50 мА, а три геля потребляли ток 75 мА, и так далее.
Когда электрофорез завершен, на что указывает перемещение маркера 9 на основе белка яйца, который визуализируют бромфеноловым синим, гель извлекают из сосуда для проведения электрофореза.
В. Двухмерный перекрестий иммуноэлектрофорез (2-D Rockets)
Полоски 11 (фиг.4), содержащие электрофорезированные белки из стадии (А), отрезают и укладывают на один конец новой стеклянной чашки 13. Затем 1:1 смесь 15 2% агарозы: УВ, содержащую около 1% антисыворотки к белку, подлежащему визуализации, выливают на чашку и отстаивают. Антисыворотку прибавляют к смеси агарозы и VB, когда последняя охладится до температуры около 50°С.
"Ракетную" чашку затем подвергают электрофорезу так, как описано выше, причем конец геля, содержащий полоски первого размера, соединяют через хлопчатобумажный тампон с отрицательным электродом (катодом) 17, а противоположный конец соединяют с анодом 19. Гели электрофорезируют в течение ночи при подводимом токе, величина которого зависит от размера гелей; 10 мА нужно для каждой 8 см длины геля с тем, чтобы гель длиной 16 см потреблял ток 20 мА, и так далее.
Белки разделяют электрофорезом в первом размере, а подсчитывают и визуализируют электрофорезом во втором размере; ниже описывается методика окрашивания для целей визуализации.
С. Размягчение и окрашивание гелей
Эта методика аналогична как для традиционного иммуноэлектрофореза, так и для ракет. Окрашиваемый гель покрывают 1 слоем неволокнистой бумаги POSTL IP (товарный знак), изготовленной Adiard Evans and Co., которую предварительно смачивают водой. Затем гель покрывают 5 слоями абсорбирующего бумажного полотенца. Конструкцию размягчают в течение 1 часа, после чего всю бумажную массу удаляют, и процесс повторяют.
После вторичного размягчения гель сушат в потоке теплого воздуха и затем пропитывают с помощью ЗФР в течение по меньшей мере 1 часа с тем, чтобы удалить неосажденный белок. Гель сушат снова и окрашивают в течение 10 минут в растворе 0,5% (в отношении массы к объему) кумассии бриллиантовой голубой G250, 45% воды, 45% метанола и 10% уксусной кислоты.
Затем гель освобождают от окраски путем непрерывной промывки в 20% метаноле, 6% уксусной кислоте до тех пор, пока фон не станет чистым. После этого гель сушат в теплом воздухе.
Фиг.5 воспроизводит высушенный гель. Ракета 1 представляет собой отрицательный контрольный образец, содержащий 50 мкл нормальной человеческой сыворотки (HHS) и 25 мкл VBS, включая EGTA. EGTA представляет собой хелатор, который отщепляет кальций; кальций является существенным компонентом для активации комплемента классическим метаболическим путем, и поэтому присутствие EGTA гарантирует то, что комплемент можно активировать только альтернативным путем метаболизма. Левосторонний (больший) пик представляет С3, а правосторонний (меньший) пик представляет C3bi, продукт распада активированного С3. Поэтому в контрольном образце меньшее количество C3bi указывает только на незначительную величину активации комплемента.
В ракете 2 75% свиные эритроциты (отношение объемов) прибавляют в забуференную смесь. Наблюдается незначительное, но, вероятно, несущественное повышение уровня C3bi, говорящее о том, что свиные эритроциты только маргинально активируют человеческий комплемент альтернативным метаболическим путем. Причина такого плохого ответа непонятна.
В ракетах 3 и 4 75% свиные лимфоциты (отношение объемов) или 75% клетки свиной почки (отношение объемов) соответственно прибавляют в забуференную смесь. В каждом случае отмечается существенное повышение уровня C3bi, говорящее об активации человеческого комплемента свиными лимфоцитами.
ПРИМЕР 5. Свиные лимфоциты не лизируются человеческими антителами в присутствии свиного комплемента, однако лизируются в присутствии кроличьего или человеческого комплемента.
Метод с высвобождением хрома используют для мониторинга лизиса клеток, опосредованного человеческой сывороткой, в присутствии или свиного комплемента, или комплемента новорожденного кролика, или человеческого комплемента.
Материалы
Среда для выделения лимфоцитов - Flowlabs;
RPMI 1640 + 10% инактивированная фетальная телячья сыворотка;
ЗФР (без азида) - Oxoid;
Планшеты с V-образными лунками - Стерилин;
Лимфоциты комплемента новорожденного кролика - Сыворотки;
Метки - или человеческий или свиной комплемент (разбавления 1 + 7 в RPMI);
Инактивированные теплом сыворотки (56°С, 30 минут)
Метод
1. Дефибринированную цельную кровь свиньи, разбавленную 1:1 в ЗФР, наносят послойно на равный объем среды для выделения лимфоцитов Ficoll Hypaque. Пробирки центрифугируют при 1200 г в течение 80 минут при температуре 20°С.
2. Полученные свиные лимфоциты на границе раздела извлекают и промывают один раз в ЗФР. Супернатант ресуспендируют в RPMI 1640, и количество клеток доводят до 2×107 /мл.
3. 200 мккуб. дюйм 51Сr прибавляют к 2×107 осадка клеток и инкубируют при комнатной температуре в течение 1,5 часа.
4. Меченые клетки промывают дважды при 900 г в течение 5 минут и регулируют с получением конечного количества клеток, равного 1×106 /мл, в RPMI.
5. 0,05 мл количества инактивированных сывороток, подлежащих тестированию в качестве последовательных разведении с удвоением, вместе с контрольными образцами высевают на чашки. Разбавленный комплемент прибавляют в релевантные лунки в количестве 0,05 мл с последующим прибавлением 0,05 мл меченых клеток. Чашки инкубируют в течение 1 часа при температуре 30°С в печи с СO2.
6. После инкубации чашки центрифугируют в течение 15 минут при 900 г, 20°С с тем, чтобы осадить клетки. 100 мкл супернатанта удаляют в меченые пробирки и подсчеты в течение 1 минуты осуществляют на счетчике гамма-квантов.
7. Результаты отложены в % от количества первоначально меченых клеток в зависимости от титра.
Контрольные пробы
Контроль с полным высвобождением (FRC) - 50 мкл клеток + 100 мкл воды + 0,1% Твин;
Отрицательный контроль - 50 мкл клеток + 100 мкл RPMI;
Комплементный контроль (СС) - 50 мкл клеток + 50 мкл разбавленного комплемента + 50 мкл RPMI.
Результаты
Результаты испытания приведены на фиг.6. Можно видеть, что свиные лимфоциты лизируются человеческой сывороткой только в присутствии несвиного (то есть кроличьего или человеческого) комплемента, но не в присутствии свиного комплемента.
Вывод состоит в том, что один или более гомологичных факторов рестрикции комплемента, присутствующих на свиных клетках, успешно регулируют в сторону уменьшения действия свиного комплемента, но не действия человеческого или кроличьего комплемента.
ПРИМЕР 6.
Целью данного примера является показ того, что антитело может вызывать гиперострое отторжение трансплантата. Концепция, на которой основано данное изобретение, возникла в результате обнаружения, что гиперострое отторжение может иметь место в отсутствие антител против трансплантата, однако требует наличия функционального комплемента. Поскольку такое обнаружение является новым, в литературе нет описания экспериментов, которые формально показывали бы, что антитело может вызывать отторжение ксенотрансплантата. Так как в присутствии встречающегося в природе антитела трудно определить, играют ли эти антитела роль или нет, такой эксперимент нелегко осуществить. В данном примере роль антитела продемонстрирована путем обращения конкордантного ксенотрансплантата в дискордантный ксенотрансплантат с помощью введения антитела соответствующей специфичности. Используемыми в данном исследовании реципиентами являются самцы крыс линии PVG (RTIC) (Banting and Kingdom, Bicester, Oxon, UK) в возрасте от 3 до 6 месяцев и весом от 250 до 300 г. Донорами сердца являются сирийские хомячки, также полученные из Banting and Kingdom и весящие от 100 до 150 г. Трансплантацию сердца осуществляют в соответствии с методом Heron (см. ссылку в Примере 1). Сердца хомячков трансплантируют в шею крыс, соединяя аорту с сонной артерией, а легочную артерию - с яремной веной с помощью методики манжеты. Все другие сосуды лигируют. Сердца начинают биться через несколько минут после освобождения от васкулярных зажимов и подвергаются мониторингу наружной пальпацией. Все операции осуществляют на животных, анестезированных аспирацией галотана и кислорода.
Уровни противохомячковых литических антител измеряют следующим образом: 50 мкл 1% раствора эритроцитов хомячков добавляют к 50 мкл испытуемой сыворотки, которая последовательно разбавлена. 50 мкл 1 из 7 разбавлении комплемента новорожденного кролика (Sera Lab., Crawley Down, Sussex) прибавляют в раствор и инкубируют в течение 1 часа при температуре 37°С. Прибавляют 750 мкл разбавителя для фиксации комплемента и центрифугируют (Beckman MICROFUGE, 13000 об/мин, 4 минуты), после чего OD 415 измеряют в надсадочной жидкости. (Слово MICROFUGE является товарным знаком). Положительными и отрицательными контрольными образцами являются CFD и дистиллированная вода, прибавляемые к 1% раствору клеток, соответственно. Результаты измерения OD 415 откладывают в зависимости от титрования сыворотки на оси X. Как можно видеть из фиг.7, трансплантация сердца хомячка в крысу приводит к получению крысы, продуцирующей очень высокие титры литических противохомячковых антител. Сыворотку от некоторых этих крыс разделяют на ее компонентные белковые фракции колоночной хроматографией на колонке SEPHADEX G200 (Pharmacia GB Ltd, London) с использованием стандартной методики колоночной хроматографии ("Использование SEPHADEX при разделении, очистке и характеристике биологических материалов", Curling в журнале Exp.Physiol. and Biochem., 3 (1970) 417-484 (G.A. Kerkut Ed.) Academic Press, London and New York, 1970). (Слово SEPHADEX является товарным знаком). Каждую из 7 мл фракций, собранных из колонки, анализируют на литическую противохомячковую активность, как описано выше. Фиг.8 показывает, что, несмотря на то, что эти антитела индуцированы в результате трансплантации сердца, антивидовая активность сохраняется почти исключительно во фракции иммуноглобулина М. После испытания на активность фракции концентрируют с использованием ультрафильтров СХ10 (Pharmacia) до концентрации 0,5 мг/мл и сохраняют при температуре -70°С вплоть до использования.
С целью исследования их способности отторгать ксенотрансплантат в противоположность лизированию красных клеток, сердца хомячков трансплантируют в шеи интактивных крыс. Вскоре после того, как сердца начали биться, 2 мл чистой сыворотки или 0,5 мг очищенного иммуноглобулина, содержащего литические противохомячковые антитела, вводят внутривенно крысам. Несепарированная сыворотка и 0,5 мг иммуноглобулина М вызывают отторжение трансплантата хомячкового сердца в течение 15 минут. Результаты от введения иммуноглобулина Г не согласуется с некоторыми препаратами, вызывающими отторжение трансплантата, тогда как в других случаях трансплантат продолжает функционировать. Когда альбумин из колонки G200 вводят в качестве контрольного образца, трансплантаты сердца всегда выживают и отторгаются в течение обычного периода времени для данной модели, который составляет 3 суток. Данный эксперимент показывает, что связывание этого антитела с трансплантатом может индуцировать его гиперострое отторжение.
ПРИМЕР 7.
Данные, приводимые до сих пор в данной заявке, показывают, что отторжение ксенотрансплантата может вовлекать активацию комплемента либо альтернативным метаболическим путем, либо антитело-опосредованной активацией комплемента (классическим путем метаболизма). Кроме того, регуляторы комплемента на поверхности мишени ксенотрансплантата могут защитить его от отторжения гомологичным, но не гетерологичным комплементом. Решающей стадией активации, общей для обоих метаболических путей активации комплемента, является расщепление комплементного компонента С3. Это расщепление осуществляют С3-конвертазой, С4b2а (С3-конвертазой классического метаболического пути) или конвертазой С3bВb (С3-конвертазой альтернативного пути). Эти ферменты расщепляют С3 с образованием С3b, который, в свою очередь, может задействовать альтернативный метаболический путь для образования большего количества С3-конвертаз (петля с обратной связью). В результате система комплемента способна амплифицировать отложение С3b на "чужеродной" мишени. Однако множество С3b не взаимодействует успешно с чужеродной мишенью и остается в жидкой фазе и может поэтому беспорядочно связываться с клетками хозяина. С целью защиты этих клеток от воздействия беспорядочного связывания комплемента, контрольные белки вовлечены в инактивацию компонентов комплемента либо в жидкой фазе, либо в состоянии связывания с собственной тканью. Те гликопротеиды, которые участвуют в управлении С3, генетически все ассоциируются в пределах одного региона человеческой хромосомы 1, называемого локусом RCA (регуляторы активации комплемента). В данном примере заявитель показывает, что мышиные клетки, которые посредством методик синтеза приобрели человеческую хромосому I и экспрессируют белки локуса RCA на их поверхности, ведут себя при анализе in vitro отторжения ксенотрансплантата так, как если бы они были человеческими клетками, а не мышиными клетками.
Линии клеток
Т5 представляет собой трансформированную вирусом Эпштейна-Барр линию В-клеток миндалины, полученную методикой Bird et al., (Nature, 289: 300-301 (1981)). В10 представляет собой человеческую гибридому, продуцирующую противостолбнячное моноклональное антитело, которую получают в результате синтеза человеческой В-лимфобластоидной линии (BLL) с линией клеток мышиной миеломы X63-AG8.653 (Kierney et al., (J.Immunol., 123: I548-I550 (1979)). Линии клеток Т5 и В10 получают от Магистра наук К. Картера и Доктора Н.К. Хьюджес-Джонсона из Группы MRC MITI в Бабрахаме, Кембридж. DB3 представляет собой линию клеток мышиной гибридомы, которая продуцирует противопрогестероновое моноклональное антитело (Wright et al.. Nature, 295: 415-417 (1982)). Получают следующие олиго-нуклеотидные праймеры, специфичные для человеческой хромосомы I: (5’-CCACAGGTGTAACATTGTGT-3’) (SEQ ID NO: 1) и (5’-GAGATAGTGTGATCTGAGGC-3’) SEQ ID NO: 2; они, соответственно, являются расположенными выше и ниже в последовательности праймерами человеческого гена антитромбина 2 (АТЗ), который, как известно, находится на человеческой хромосоме I (Wu et al., Nucl.Acids Res., 17: 6433 (1989)). Олигонуклеотиды можно синтезировать хорошо известными специалистам методами.
Высокомолекулярную геномную ДНК получают с использованием метода Herrmann и Frischauf (Methods Enzymol., 152: 180-183 (1987)). Вкратце, 100×106 клеток от каждой культивированной клеточной линии лизируют с помощью 5 мл TNE (100 мМ-Трис, рН 7,5, 100 мМ Nad, 10 мМ ЭДТК 1% Саркозил) и обрабатывают свежей протеиназой К (100 мкг/мл). Препарат экстрагируют фенолом (насыщенным водой и уравновешенным против 0,1М Трис, рН 8), фенолхлороформом (1:1, объемн.) и затем изоамиловым спиртом:хлороформом (1:24, объемн.). ДНК получают этаноловой преципитацией и диализируют против ТЕ (10 мм Трис, рН 8,0, 1 мМ ЭДТК), доведенного до 100 мМ в NaCl, а также только ТЕ при температуре 4°С. Выделенную ДНК анализируют на 0,5% агарозном геле и концентрацию определяют по оптической плотности при 260 нм. Полимеразно - цепьевую реакцию (PCR) для каждой клеточной линии осуществляют в соответствии с описанием Saiki et al., (Science, 239: 487-491 (1989)). В объеме 100 мкл содержится 500 нг геномной ДНК, 1,2 нг каждого праймера и 2,5 единицы ДНК-полимеразы Taq (тип 3 Thermos acquaticus) (Cambio Ltd., Cambridge, UK) с использованием забуференного раствора, подаваемого с ферментом. Нуклеотиды (dNTPs) (Boehringer Mannheim Diagnostics and Biochemicals Ltd, Lewis/ East Sussex, UK) имеют концентрацию 2 мМ каждый. ДНК амплифицируют в течение 30 циклов с использованием программируемого термического контроллера (Genetic Research Instrumentation Ltd, Dunmow, Essex, UK): денатурация при температуре 93°С в течение 1 минуты: отжиг при температуре 55°С в течение 1 минуты: и вставка при температуре 72°С в течение 2 минут. 10 мкл реакционного продукта анализируют непосредственно на 2% агарозном геле в забуференном растворе Трис, борная кислота, ЭДТК. Размер продукта определяют путем сравнения с HincII-переваренной фаговой X-174-rf ДНК (Pharmacia LKB Biotechnology, Upsala, Sweden).
Культивированные клетки Т5, В10 и DB3 обрабатывают противоDAF (стимулятор гемолиза) моноклональным антителом (Kinoshita et al., (J.Exp.Med., 162: 75-92 (1985)) и меченым флуоресцином "вторым" антителом. Клетки (1×106) подвергают взаимодействию с мышиным противоDAF моноклональным антителом IAIO (ИгГ2а 10 мкг/мл в 100 мкл 10% фетальной плодной сыворотке, 0,1% азиде). IAIO (Kinoshita et al., (J. Immunol., 136:.3390-3395 (1986)) получают от Доктора М.Дэвица из Медицинского Центра Университета штата Нью-Йорк; Нью-Йорк, США. "Слепыми" контрольными пробами служат только буферы. После инкубации в течение 2 часов на льду клетки промывают 3 раза, ресуспендируют и инкубируют в 100 мкл буфера, содержащего 1 в 100 FITC-сопряженный козлиный F (ab’)2 противомышиный иммуноглобулин (с очищенными тяжелыми и легкими цепями по сродству и абсорбированными в человеческом иммуноглобулине) (Tago Immunochemicals Inc., Burlingame, California, USA) в течение одного часа на льду. Некоторые клетки инкубируют только со вторым антителом в качестве окрашивающих контрольных образцов. Поскольку DB3 представляет собой линию клеток, секретирующих мышиный ИгГ1, FITC-сопряженный овечий противомышиный ИгГ2А (I в 40, The Binding Site Ltd, Birmingham, UK) или козлиный противомышиный ИГ, предварительно абсорбированный равными объемами клеток DB3, также используют для того, чтобы элиминировать анти-ИгГ1 реактивность, происходящую при окрашивании клеток DB3. Все клетки тщательно промывают и ресуспендируют в 200 мкл забуференного раствора. DAF-положительные клетки обнаруживают с использованием аппарата Beckton Dickinson FACS-STAR для анализа клеточного сортера с возбуждением флуоресценции (FACS). (Выражение FACS-STAR является товарным знаком).
Метод полимеразно-цепьевой реакции (PCR) используют для обнаружения присутствия человеческой хромосомы 1 в трех различных линиях - культивированных клеток: Т5 (человеческая), В10 (человеко-мышиная) и DB3 (мышино-мышиная). Фиг.9 показывает, что после амплификации Т5 и В10 имеют размер полосы, равный 495 парам оснований, тогда как DB3 (то есть мышино-мышиный гибрид) вовсе не имеет полосы. Изложено, что продукты PCR с использованием праймеров АТЗ состоят из 2 аллелей размером 572 пары оснований (аллель 1) и 496 пар оснований (аллель 2) (Wu et al., выше). Полосы, обнаруженные в геномных ДНК Т5 и В10, соответствуют аллели 2. Это доказывает, что линия клеток В10 гибрида человеко-мышь содержит человеческую хромосому I.
FACS-анализ на присутствие DAF показывает, что большинство человеческих клеток Т5 (85,7%) окрашено положительно противоDAF моноклональным антителом. Аналогичный уровень (83,1%) положительных клеток обнаружен в клетках В10 гибрида мышь - человек. Клетки DB3 гибрида мышь-мышь проявляют идентичные характеры окрашивания для обоих противоDAF обработанных и необработанных препаратов. Однако, эта реакционноспособность противомышиного ИгГ1 пропадает, если
(1) используют FITC-сопряженный противомышиный ИгГ2а или
(2) вышеприведенный козлиный противомышиный ИгГ предварительно абсорбирует клетками DB3. Результаты показывают, что линия клеток В10 гибрида человек-мышь экспрессирует человеческий DAF на поверхности клеточной мембраны, как обнаружено специфическими противоDAF моноклональными антителами. Уровень экспрессии аналогичен уровню экспрессии относительно человеческой линии клеток Т5. Мышино-мышиная гибридомная линия клеток не экспрессирует человеческий DAF.
Цитотоксические исследования активности клеток-киллеров с радиоактивным хромом осуществляют в отношении этих клеточных линий, как описано в Примере 5 выше. Фиг.10 показывает, что при инкубации свиных противочеловеческих антител с человеческой линией клеток Т5 прибавление кроличьего комплемента вызывает лизис, тогда как лизис не происходит при добавлении человеческого комплемента, так как, конечно, линия клеток Т5 обладает человеческими HCRFS. Это является подтверждением результатов Примера 5. Когда человеческие антитела используют на человеческой линии клеток, лизис не происходит как с человеческим комплементом, так и с кроличьим комплементом, что говорит об отсутствии аутоантител. Методика с радиоактивным хромом не позволяет проводить длительную инкубацию для обнаружения любых значительных уровней активации альтернативным метаболическим путем кроличьего комплемента человеческими клетками (фиг.11). Однако, когда человеческие антитела инкубируют с линией клеток мышино-мышиной гибридомы DB3 (фиг.12), киллинг (лизис) клеток достигается как кроличьим комплементом, так и человеческим комплементом, что демонстрирует возможность существования и функционирования человеческого комплемента в таком испытании. При использовании человеко-мышиного гибрида В10, обладающего человеческой хромосомой I и экспрессирующего по меньшей мере DAF, кроличий комплемент вызывает лизис клеточной линии, тогда как человеческий комплемент не в состоянии вызвать лизис клеточной линии (фиг.13). Объяснение такому феномену заключается в том, что человеческие HCRFS, экспрессируемые благодаря обладанию хромосомой I на мышино-человеческом гибриде, ингибируют активность человеческого комплемента.
ПРИМЕР 8.
Предыдущий пример показывает, что обладание хромосомой I может предотвратить отторжение клеток ксенотрансплантата. В то время как существуют сильные косвенные улики относительно того, что именно локус CRA защищает мышиную клетку от отторжения ксенотрансплантата, данный пример приводит формальное доказательство. В этом примере продемонстрирован эффект трансфекции нечеловеческих клеточных линий с помощью человеческого МСР и их взаимодействие с человеческим или кроличьим комплементом.
кДНК получают для МСР (человеческого белка мембранного кофактора) в соответствии с подробным описанием в работе Lublin et al., (J.Exp.Med., 168: 181-194 (1988)). Конструирование трансфецированных клеточных линий осуществляют с использованием экспрессирующей плазмиды SFFV.neo в соответствии с методикой, описанной Uhlbrigge et al., (Proc.Natl. Acad.Sci., 85:5649-5653 (1988)). Она содержит длинный концевой повтор 5’ вируса, образующего очаг птичьего лейкоза (вируса Фрейнда) (SFFV.LTR) (Clark и Mak (Nucl.Acids.Res., 10: 3315-3330 (1982)) и (Proc.Natl.Acad.Sci, 80: 5037-5041 (1983))). Клеточные линии получают из Американской Коллекции Типовых Культур, 12301, Parklawn Drive, Rockville, Maryland, USA. Используемыми клеточными линиями являются CHO-KI (ATCC CCL 61) и NIH/3Т3 (ATCC CPL 1658). Экспрессию гена подтверждают с использованием моноклонального антитела к МСР (Andrews et al., Ann.Hum.Genet., 49: 31-39 (1985) и анализа FACS, который уже описан выше. В некоторых случаях клеточные линии отбирают по принципу высокого уровня экспрессии МСР путем клеточного сортинга на FACS с использованием стандартной методологии.
Данный пример иллюстрирует эффект трансфекции клеток СНО с помощью МСР. Поскольку эти клеточные линии растут как монослои, киллинг клеток оценивают анализом поглощения терминального аденина, который описан в работе Воnо et al., (Immunology, 32: 221-226 (1977)). Вкратце, данный анализ включает инкубирование клеточных культур в плоскодонных стерильных 96-луночных планшетах с комплементом и антителом. К концу периода экспериментальной инкубации жизнедеятельность клеток оценивают по способности культуры принимать радиоактивный аденин. Жизнеспособные клетки принимают аденин, мертвые клетки не принимают; поэтому жизнеспособные клетки имеют высокие количества, мертвые клетки имеют низкие количества.
По аналогии со многими трансформированными клетками, СНО являются нечувствительными к антителам природного происхождения и воздействию альтернативного метаболического пути комплемента. Однако эти клетки чувствительны к тем антителам, которые, как продемонстрировано, вызывают отторжение ксенотрансплантата сердца хомячка, как описано в Примере 6. Поскольку клетки СНО получены от хомячка, эти антитела убивают клетки СНО как человеческим, так и кроличьим комплементом (фиг.14). Когда клетки СНО трансфецированы человеческим МСР, клетки могут быть лизированы только в присутствии кроличьего комплемента. Человеческий комплемент ингибируется благодаря присутствию человеческого МСР на поверхности линии клеток хомячка (фиг.15). Доказательством того, что распад клеток, подлежащих уничтожению, действительно обусловлен распадом С3-конвертазы, служит анализ распада человеческого С3 после инкубации клеток СНО под действием ракетного иммуноэлектрофореза, как описано в Примере 4 выше. Как можно заметить, никакого распада нет выше контрольных уровней комплемента (фиг.16).
Эти данные подтверждают, что полученные методами генной инженерии белки, регулирующие в сторону снижения комплемент на поверхности нечеловеческих клеток, защищают от механизмов гиперострого отторжения ксенотрансплантата те клетки, которые в качестве общего отличительного признака имеют потребность в расщеплении С3-компонента комплемента.
ПРИМЕР 9.
В соответствии с методикой Примера 8 клетки фибробласта мыши ЗТЗ трансфецируют кДНК, кодирующей МСР (МСР-клон К5.23). 51Сr прибавляют к клеткам, как описано в Примере 5. Один объем клеток затем инкубируют с одним объемом человеческого комплемента, инактивированного теплом, и одним объемом человеческого комплемента, предварительно абсорбированного при температуре 4°С с клетками мышиной селезенки с тем, чтобы удалить противомышиное антитело из человеческого комплемента. Смесь и последовательные разбавления комплементом высевают на чашки. Основные условия и отличительные признаки анализа с радиоактивным хромом аналогичны описываемым в Примере 5. Результаты относительно клона К5.23 приведены на фиг.17, который также показывает, в качестве контрольного опыта, эффект кДНК МСР, вводимой в обратной ориентации (в данном случае ее не транскрибируют). Правильно транскрибированная кДНК МСР придает защиту клеткам от киллинга, как видно по относительно низкому уровню высвобождения 51Сr, тогда как нетранскрибированные кДНК не придают защиту, на что указывают относительно высокие уровни высвобождения 51Сr.
ПРИМЕР 10.
Результаты, аналогичные описанным в Примере 8 выше, могут быть получены с использованием клеток LI/210 (мышиная лейкемическая клеточная линия), трансфецированных кДНК для DAF. кДНК получают для DAF в соответствии с описанием в работе Lublin и Atkinson (Ann.Rev.Immunol., 7: 35-58 (1989)).
ПРИМЕР 11.
кДНК для МСР получают и лигируют в плазмиду SFFV.neo, как в Примере 8 выше.
С использованием этого препарата ДНК трансгенные мыши получают в соответствии с описанием в работе "Манипулирование мышиными эмбрионами. Лабораторное Руководство" В.Ноgаn et al., Cold Spring Harbour Laboratory (1986). От десяти до пятнадцати мышей-самок (CBAxBlO)fI в возрасте 3-4 недель индуцируют к суперовуляту путем внутрибрюшинной инъекции 5 единиц сывороточного гонадотропина от беременных кобыл (поставляется коммерчески как Folligon) с последующей через 48 часов внутрибрюшинной инъекцией 5 единиц хорионического гонадотропина из мочи от беременных женщин (поставляется коммерчески как Chorulon). Самок оставляют для спаривания, в день иньецирования Chorulon, с самцами (CBAxB10)FI, а на следующий день самок с вагинальными пробками умерщвляют цервикальным (шейным) смешением и оплодотворенные яйцеклетки выделяют из их маточных труб.
От трех до четырех сотен яйцеклеток, выделенных таким образом, содержат два пронуклеуса, четко видимых с использованием дифференциальной интерференционной контрастной оптики Nomarski при 400-кратном увеличении. Один из двух пронуклеусов инъецируют приблизительно с 2000 копий ДНК-препарата, содержащего трансген кДНК МСР, при концентрациях от 0,5 до 2 нг/мкл.
Яйцеклетки, выжившие при микроинъекции, реимплантируют в маточные трубы самок (CBAxB10)FI, которые были спарены предыдущей ночью с вазоэктомизированными самцами и поэтому являлись псевдобеременными (то есть они овулированы, а их гормональное состояние является состоянием беременности, однако их собственные ооциты не оплодотворены). Приблизительно 30 микроинъецированных яйцеклеток переносят в маточные трубы каждого псевдобеременкого животного, под анестезией, либо в день микроинъецирования, либо на следующий день, когда яйцеклетки находятся на второй клеточной стадии. Происходит нормальная беременность, и от десяти матерей рождаются семнадцать мышей. Скрининг потомства осуществляют слот-блоттингом и/или Саузернблоттингом (см. Пример 8), а также полимеразно-цепьевой реакцией, анализом ДНК из клеток кожи хвоста, используя 32Р-меченые зонды и праймеры, которые распознают трансген. Одно животное из потомства, самец, является трансгенным для последовательности ДНК МСР.
ПРИМЕР 12.
Повторяют методику Примера 11 за исключением того, что кДНК для DAF, как описано в Примере 10, используют вместо кДНК для МСР. Двадцать три мыши рождаются от десяти матерей. Три из них (две самки, один самец), трансгенных для DAF, получают с использованием блоттинг-метода по Саузерну.
ПРИМЕР 13.
Самцу мыши, полученному в Примере 11, который содержит человеческий трансген кДНК МСР, позволяют вырасти до зрелости и спаривают с самкой (CBAxB10)FI. Получают потомство из одиннадцати животных. ДНК клеток хвоста от каждого члена потомства подвергают скринингу слот-блот-анализом, используя в качестве зонда меченую человеческую кДНК МСР, для определения того, наследован ли трансген или нет. Результаты анализа приведены в верхней части фиг.18. Можно видеть, что потомки 0, 1, 5, 7, 8 и 10 наследовали. (Предпринимают четыре контрольных анализа: с человеческой дНК (Н), мышиной ДНК (М), мышиной ДНК, смешанной с 10 пг человеческой МСР-меченой кДНК, а также мышиной ДНК, смешанной с 100 пг человеческой МСР-меченой кДНК).
ПРИМЕР 14.
Самцу мыши, полученному в Примере 12, который содержит человеческий трансген кДНК DAF, позволяют вырасти до зрелости и спаривают с самкой (CBAxB10)FI. В отношении каждого полученного потомка ДНК клеток хвоста подвергают скринингу слот-блоттингом, используя меченую кДНК человеческого DAF в качестве зонда, для определения того, наследован ли трансген или нет. Результаты анализа приведены в нижней части фиг.18. Можно видеть, что потомство 13.3 (самка) наследовало. (Осуществляют четыре контрольных пробы: человеческая ДНК (Н), мышиная ДНК (М), мышиная ДНК, смешанная с 10 пг человеческой меченой кДНК DAF, а также мышиная ДНК, смешанная со 100 пг меченой кДНК человеческого DAF (стимулятора гемолиза)).
Список последовательностей
SEQ ID NO: 1
Тип последовательности: Нуклеотидная
Длина последовательности: 20
Свойства: Расположенный выше праймер человеческого гена антитромбина 2 (АТЗ)
Последовательность: CCACAGGTGT AACATTGTGT 20
SEQ ID NO: 2
Тип последовательности: Нуклеотидная
Длина последовательности: 20
Свойства: Расположенный ниже праймер человеческого гена антитромбина 2 (АТЗ)
Последовательность: GAGATAGTGT GATCTGAGGC 20












Изобретение относится к области генетической инженерии, в частности к способу получения трансгенного животного. Способ предусматривает введение ДНК в генетический материал животного. При этом ДНК кодирует гомологичный фактор рестрикции комплемента дискордантного вида и последовательность, обеспечивающую экспрессию кодирующей ДНК в клетках трансгенного животного. Клетки, выделенные от такого трансгенного животного, можно трансплантировать животному указанного дискордантного вида без острого отторжения. Изобретение может быть использовано для получения материалов для трансплантации. 1 з.п. ф-лы, 18 ил.
| HAMMER R.E | |||
| et al., “Production of transgenic rabbits, sheep and pigs by microinjection” | |||
| Nature | |||
| Приспособление для установки двигателя в топках с получающими возвратно-поступательное перемещение колосниками | 1917 |
|
SU1985A1 |
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| WO 8800239, 14.01.1988. | |||

