Изобретение относится к новым модифицированным белкам, способным образовывать стабильные С3 конвертазы, к ДНК последовательностям, кодирующим такие белки, и к их использованию в качестве терапевтических агентов, особенно для использования для снижения уровней белков схемы комплемента.
Функционирование системы комплемента в иммунных реакциях людей и других позвоночных играет основную роль в таких эффекторных функциях, как фагоцитоз, цитолиз и возбуждение клеток, которые индуцируют локальные воспалительные реакции [15]. Такие свойства необходимы для исключения таких заражающих патогенов, как бактерии, но вовсе нежелательны, когда их действие направлено против тканей организма-хозяина /например, при повреждении постишемической реперфузии [3]/ или против чужеродного терапевтического материала /например, гиперострое отторжение ксенотрансплантатов [7]/. Были предприняты попытки исключить эти нежелательные свойства за счет использования производных регуляторных протеинов комплемента, функция которых состоит в подавлении активации комплемента [10, 18].
Система комплемента включает протеины как на поверхности клеток /рецепторы и регуляторы/, так и в жидкой фазе /плазма крови и другое внеклеточное окружение/. Критической стадией для возбуждения реакции является протеолитическое превращение С3 во фрагменты C3b и С3а. Фрагмент С3а представляет анафилатоксин, который, подобно C5a, притягивает тучные клетки к очагу заражения, что приводит к локальному выделению гистамина, вазодиляции, и к другим воспалительным эффектам. Образующиеся С3b обладают способностью связываться с поверхностью вокруг очага их образования. Затем эти С3b фокусируют атаку за счет цитолитических компонентов комплемента /С5-С9/.
Связанные с поверхностью С3b и их продукты разложения также функционируют как лиганды для С3 рецепторов, участвуя, например, в фагоцитозе [15]. Существуют две различные схемы активации комплемента, причем обе они приводят к превращению С3 в С3b и к последующей реакции. Классическим путем является обычно возбуждение комплексов антитело - антиген, инициируя ряд ферментов, включающий белки С1q, С1r, C1s, С2 и С4. Альтернативная схема зависит от активационной петли, включающей сам С3 и требующей участия факторов В и Д.
Превращение С3 в С3b /или C3i / дает продукт, который может объединяться с фактором В, образуя С3bВ /или С3iВ/. Эти комплексы подвергаются воздействию фактора D с образованием С3bВb, что представляет собой С3 конвертазу, которая способна увеличить превращение С3 в С3b, что приводит к увеличению числа С3bBb, и еще более увеличивает конверсию С3. В определенных обстоятельствах комплекс С3bВb стабилизируется за счет ассоциации с позитивным регуляторным пропердином /Р/. Однако такая петля положительной обратной связи обычно бывает ограничена до медленных актов за счет регуляторных протеинов, особенно фактора Н и фактора I.
Фактор Н /и структурно родственные связанные с клетками молекулы/ /i/ удаляет В и Bb с С3b, и /ii/ действует как кофактор для фактора I, который расщепляет С3b до iC3b, тем самым предотвращая какую бы то ни было рекомбинацию с фактором В и образование большего количества конвертаз. Этот путь направлен на усиленное образование С3b на поверхности, например, клеточных стенок бактерий, которые связывают образующиеся С3b и мешают их регуляции за счет факторов Н и I. Образующиеся С3b способны также связываться с эндогенными клетками. Поэтому эндогенные клеточные поверхности, обычно доступные комплементу, оказываются дополнительно защищены за счет таких связанных с мембраной регуляторов, как МСР, DАF и CR1, действующих аналогично фактору Н.
Существует несколько редко встречающихся в природе условий, при которых не может происходить нормальная регуляция в жидкой фазе, и спонтанная С3 конверсия в конечном счете приводит к общему исключению С3 из циркуляции: /i/ генетический дефицит фактора Н или фактора I [13], /ii/ наличие антител /нефритные факторы/, которые связываются в С3bВb и препятствуют диссоциации [4] , и /iii/ контакт с протеином яда кобры, называемым фактором яда кобры /CVF/, который объединяется с фактором В и образует конвертазу, которая не содержит С3b, и не подвержена воздействию факторов Н и I [14]. Это иллюстрирует нормальную физиологическую важность регуляции в сторону уменьшения /даунрегуляции/ комплемента в отсутствие специфической активации.
Существуют также обстоятельства, при которых наблюдается специфическая активация, но оказывается нежелательной, особенно, если она направлена против тканей хозяина /например, повреждение тканей при ишемии или хирургическом вмешательстве/ или против чужеродного материала, преднамеренно используемого для терапевтических целей /такого, как ксенотрансплантаты, искусственные органы или мембраны для диализа/. Активация комплемента приводит к нежелательной атаке и дальнейшему поражению, так что в таких случаях желательно блокировать или ингибировать такую активацию и реакцию.
Существующие подходы для предотвращения повреждений за счет комплемента направлены на использование даун-регулирующих протеинов /СR1, МСР, DАF и факторов Н и I/ для ингибирования активации комплемента. Ингибиторы комплемента подобные фактору I, фактору Н и растворимым производным связанных с мембраной протеинов СR1, DAF MCP, подавляют петлю амплификации в жидкой фазе в альтернативной схеме. Поэтому были предприняты попытки использовать эти молекулы, в частности СR1 /который, по-видимому, является наиболее эффективным/ для уменьшения связанных с комплементом поражений в моделях физиологических ситуаций [10, 18].
Фактор Н эндогенно присутствует в плазме крови в высоких концентрациях /обычно 0,3-0,5 мг/мл [15]/, и хотя повышенные уровни ингибиторов ослабляют жидкофазные реакции, их эффективность мала, и поэтому приходится вводить ин виво большие количества очищенных протеинов /например, более 5 мг/кг веса тела растворимого CR1/. Кроме того, альтернативная схема активируется поверхностями, где влияние фактора Н уже ослаблено. Хотя это не обязательно попутно снижает активности других ингибиторов, предполагают, что те же факторы, по-видимому, не могут быть столь же полно или универсально эффективны.
Фактор яда кобры /CVF/ обладает свойством выработки стабильной С3 конвертазы, которую можно использовать экспериментально для снижения количества комплемента у животных ин виво и в других образцах /например, в плазме крови человека/ ин витро. CVF эффективен /например, 40 мкг/кг могут разрушить активность комплемента у мыши [16]/. Однако существуют недостатки, которые делают его неподходящим для терапевтического использования при лечении людей.
Во-первых, его получают из яда кобры /малодоступный источник и опасности при работе с препаратом/, что требует тщательной очистки от нейротоксинов яда. Существуют также очевидные трудности в получении материала. Эту проблему нельзя легко решить за счет клонирования и экспрессии гена ex vivo, так как существуют посттрансляционные модификации, которые наблюдаются у змей /специфический протеолитический процессинг/, которые могут оказаться трудно /или невозможно/ воспроизводимыми ин витро. Кроме того, ферменты и условия расщепления, необходимые для такого процессинга, в настоящее время неизвестны. Во-вторых, протеин является чужеродным /для человека/ и поэтому иммуногенным. Это делает невозможным его повторное использование для терапевтических целей, так как требует декомплементации пациента в течение многих недель /например, для обеспечения приживаемости ксенотрансплантатов/.
Хотя CVF имеет некоторую структурную и функциональную гомологичность с С3 человека [17], он также имеет существенные отличия в этих двух аспектах /например, в строении цепей, сайтах биосинтеза, нечувствительности к регуляторам комплемента, в образовании стабильной С3 конвертазы/. Он не получен из змеиного эквивалента /от кобры/ С3, который, как известно, был клонирован и секвенирован и который по структуре и функциям напоминает С3 человека более сильно, нежели CVF [8].
CVF представляет ядоспецифический продукт животного, находящегося на значительной эволюционной дистанции от гомо сапиенс. Именно поэтому нежелательно использовать генетические манипуляции для модификации этого протеина для получения продукта, который приемлем человеку для неиммуногенного использования.
Был разработан альтернативный подход, который основан на том, чтобы обойти физиологическую регуляцию, и вместо ингибирования активации комплемента вызывает суперактивацию системы. Это имеет два применения. Во-первых, это можно использовать ин виво для активации комплемента до тех пор, пока не израсходуется один или более из компонентов, что приводит к потере способности вызывать локальные реакции на любое последующее заражение /например, на ксенотрансплантаты/. Во-вторых, нерегулируемую суперактивацию можно преднамеренно локализовать на конкретной мишени /например, на вирус или инфицированную вирусом клетку/ для повышения чувствительности такой направленной, осуществляемой за счет комплемента деструктивной реакции.
Термин "регуляторы активации комплемента" используют здесь по отношению ко всем протеинам, которые действуют как ингибиторы амплификации С3 конверсии, и его значение не следует ограничивать теми протеинами, гены которых расположены в RCA генетическом локусе. Однако он не должен включать такие "ап-регуляторы" /усиливающие регуляторы/, как пропердин. Термин "С3 конверсия" определяют как конверсию С3 в C3b и С3а, если нет других указаний, а термин "С3 конвертаза" /или просто "конвертаза"/ определяют как фермент /обычно комплекс двух или более протеиновых компонентов; например, C3bBb, C3iBb, CVFBb или С4b2а/, который является катализатором этой реакции.
Таким образом, в первом аспекте в настоящем изобретении предложен протеин, действующий по схеме нативного комплемента, модифицированный таким образом, что этот протеин способен образовывать стабильную С3 конвертазу.
Под термином "нативный" подразумевают встречающийся в природе. Таким образом, это определение охватывает любые протеины, действующие по схеме нативного комплемента, модифицированные как указано ранее. Их не следует ограничивать протеинами специфических видов. Другими словами, модифицированный протеин человека можно использовать в качестве стабильной С3 конвертазы и в других видах млекопитающих. Обычно используют модифицированные протеины, действующие по схеме комплемента, из того же самого вида.
Модификации С3 ДНК кодирующей последовательности, например, за счет сайтнаправленного мутагенеза могут привести к получению такого варианта С3, который устойчив к регуляторным протеинам комплемента, сохраняя при этом позитивные функциональные свойства /расщепление до C3b за счет С3 конвертазы/ и характеристики структурной целостности /правильная структура цепи и наличие тиолэстерной связи/. Описываемое здесь изобретение относится к генетически модифицированным формам протеинов нативных комплементов, например к С3 человека, C3b фрагмент которого приобретает свойство устойчивости к физиологической комплементной регуляции. Благодаря такой устойчивости эти молекулы могут вырабатывать стабилизированные формы соответствующей С3 конвертазы, которые продуцируют усиленную конверсию С3 в C3b, и последующие продукты разложения в физиологических условиях /например, ин виво/.
В предпочтительном варианте в изобретении предложен модифицированный С3 протеин человека, который устойчив к расщеплению под действием фактора I.
Этого можно достичь модифицируя остатки протеина по протеолитическим сайтам.
Наиболее предпочтительный вариант изобретения относится к модифицированному протеину С3 человека, где протеин модифицирован за счет замены либо Arg-1303, Аrg-1320, либо их обоих, другой аминокислотой. Такой другой аминокислотой может быть тирозин, цистеин, триптофан, глутамин, глутаминовая кислота или глицин. Предпочтительно заменять Аrg-1303 на глутаминовую кислоту или глицин /менее предпочтительно на глутамин/. Предпочтительно Arg-1320 заменять на глутамин.
Другие подходы для получения подходящих модифицированных протеинов настоящего изобретения включают:
i/ пониженную подверженность ингибирующему воздействию фактора Н и родственных протеинов /например, МСР, DAF, CR1/. Для примера в С3 человека остатки 767-776 и 1209-1271 вовлечены в связывание фактора Н [20, 24], и замена одного или более из этих остатков или других остатков, также связанных с действием этих протеинов, может уменьшить связывание одного или более из этих регуляторных протеинов.
ii/ Пониженную степень диссоциации C3bВb. Можно осуществить мутации, которые усилят взаимодействие между C3b и Вb. Это приведет как к снижению спонтанной декомпозиции энзима, так и уменьшит эффективность фактора Н /и родственных регуляторов/ в вытеснении Вb из C3b.
Эти мутации желательны для уменьшения степени как спонтанной, так и осуществляемой за счет фактора Н декомпозиции C3bВb. Даже в отсутствие фактора Н срок полураспада комплекса C3bВb в жидкой фазе составляет всего около 10 минут при 37oС в присутствии пропердина [6].
iii/ Остатки 752-761 С3 человека участвуют в связывании фактора В. Это высококонсервативный участок С3, и близкородственная последовательность обнаружена в С4. Так как С4 связывает гомолог С2 фактора В, близкое сходство этого участка для С3 и С4, наряду с его высокой консервативностью в С3, является дальнейшим подтверждением его роли в С3 в качестве сайта связывания фактора В. Таким образом, изменения в этом участке могут оказывать влияние на афинность В и на стабильность C3bВb.
iv/ Устойчивость к другим регуляторам активации комплемента, таким как СR1, DАF и МСР, также была бы желательна. Тип действия этих регуляторов совершенно схож с действием фактора Н, так что при этом не потребуется дополнительного мутагенеза. Аналогично некоторые патогенные организмы экспрессируют свои собственные ингибиторы активации комплемента, которые часто структурно и функционально гомологичны фактору Н /например, секреторный пептид вируса Vaccinia/. Эти молекулы защищают вторгающихся от иммунной реакции, и было бы желательно иметь возможность атаковать их направленными энзимами - С3-конвертазами, устойчивыми к этим защитным свойствам.
v/ Мутации, которые повышают стабилизацию С3 конвертазы за счет пропердина. Активность пропердина состоит в стабилизации C3bВb комплекса, уменьшая спонтанную и зависящую от фактора Н диссоциацию. Такая стабилизация неэффективна в жидкой фазе, но, по-видимому, более важна в усилении процесса, коль скоро он уже начался на подходящей активирующей поверхности [5]. Повышение активности /за счет повышения афинности/ может нарушить баланс в жидкой фазе и тем самым промотировать спонтанную С3 конверсию. Это должно быть особенно эффективно в сочетании с другими модификациями, описанными ранее.
vi/ Мутации, которые предохраняют C3bВb от обладания С5 конвертазной активностью. При использовании для предотвращения активного С3 от циркуляции нежелательным побочным эффектом может быть выработка больших количеств анафилактических пептидов. Наиболее эффективным из них является С5а, который отщепляется от С5 С3 конвертазными энзимами. Эта реакция, вероятно, зависит от афинности конвертазы для другой молекулы С3b [11], и поэтому может служить предметом для подавления за счет мутаций для С3, которые удаляют это взаимодействие.
vii/ Повышенная активность С3 конвертазы. Активный сайт C3bВb С3 конвертазы расположен в Вb части. Предположительная функция С3 компонента состоит в придании активной конформации на Вb и/или в связывании и ориентировании субстрата, на который необходимо воздействовать, за счет Вb. Это неизвестно, но в любом случае может найтись возможность для повышения активности конвертазы за счет мутаций в С3.
viii/ Экспрессия в функциональной форме. Дикого типа С3 требует конверсии в C3b до того, как он может объединиться в новый С3 конвертазный комплекс. При использовании ин виво, требование для превращения в C3b /или C3i/ будет замедлять действие модифицированного С3. Поэтому было бы желательно либо вводить протеин в форме, способной к немедленному образованию конвертазы, либо вводить заранее сформированные комплексы конвертазы. Поэтому выгодно создавать C3b-подобный реагент экс виво. Этого можно достичь ин витро /например, за счет протеолиза/.
ix/ Модификации нативного протеина, которые служат для введения новых сайтов расщепления, таких как пептидные участки, необходимые для связывания фактора В, сохраняют, а те, которые необходимы исключительно для связывания фактора Н, можно специфически удалить. Так, например, можно ввести сайты таким образом, чтобы C3b-подобную форму модифицированных С3 можно было бы далее расщепить до форм, которые все еще связывают фактор В, но которые менее подвержены инактивации за счет факторов Н и I.
х/ Модификации в других участках, которые могут повлиять на C3b взаимодействия с фактором В и/или фактором Н.
Настоящее изобретение основано на опровержении традиционного подхода за счет промотирования С3 конверсии для расщепления С3 и тем самым выведения системы из строя. Дополнительное применение изобретения состоит в возможности промотировать С3 конверсию по конкретному сайту и тем самым заставить комплементзависимый эффекторный механизм атаковать специфическую мишень.
Вот почему конечным эффектом является повышение степени С3 конверсии при введении модифицированного протеина в физиологическую среду /например, в кровь/, содержащую регуляторы активации комплемента. Затем эту активность используют либо для истощения этой среды нативного С3, либо для локализации С3 конверсии целевой мишени.
Аналог С3, С3b-фрагмент которого устойчив к действию фактора I /например, производное, описанное в примере 1/, будет связывать фактор В, который затем будет расщепляться фактором D, и, в конечном счете, будет диссоциировать в неактивную форму. В отсутствие инактивации за счет фактора I модифицированный C3b будет способен повторно связывать новые молекулы фактора В и тем самым промотировать его инактивацию. Поэтому другим возможным применением описываемых в настоящем изобретении модификаций должна быть инактивация альтернативной схемы за счет исключения активности фактора В. Аналогичный подход можно также использовать для модификации С4 для промотирования исключения С2 и тем самым исключить классическую схему активации комплемента.
Настоящее изобретение включает любые другие протеазы, которые используют способом, аналогичным с C3bBb ферментом, что приводит к расщеплению С3 до C3b, независимо от присутствия регуляторов активации комплемента.
Настоящее изобретение включает также ДНК последовательности, которые кодируют протеин настоящего изобретения, а также ДНК конструкции, включающие такие ДНК последовательности.
Термин "ДНК последовательности" включает все другие последовательности нуклеиновых кислот, которые за счет дегенеративности генетического кода также кодируют данную аминокислотную последовательность или которые практически гомологичны этой последовательности. Поэтому такие последовательности также включены в объем настоящего изобретения.
Последовательности нуклеиновых кислот, которые "практически гомологичны", также входят в объем настоящего изобретения. Термин "существенная гомологичность" может относиться либо к уровню нуклиновых кислот, либо к уровню аминокислот. На уровне нуклеиновых кислот последовательности, обладающие существенной гомологичностью, могут рассматриваться как последовательности, которые гибридизуются с последовательностями нуклеиновых кислот настоящего изобретения в жестких условиях /например, при 35-65oС в солевом растворе около 0,9 М/. На аминокислотном уровне последовательность протеина можно рассматривать как практически гомологичную другой последовательности протеина, если значительное количество составляющих аминокислот демонстрируют гомологичность. Гомологичными могут быть по крайней мере 55%, 60%, 70%, 80%, 90%, 95% или даже 99% аминокислот в порядке возрастающей предпочтительности.
Как обсуждалось ранее, протеины настоящего изобретения можно использовать для достижения эффектов локализованной активации комплемента. Одним из способов достижения этого является конъюгация протеина с фрагментом, который будет связываться по нужной мишени. Таким образом, в другом аспекте настоящее изобретение обеспечивает конъюгат, включающий протеин настоящего изобретения, связанный со специфическим связывающим фрагментом, например со специфическим связывающим протеином. Примером такого протеина может служить антитело или антиген, связывающие его фрагмент.
Протеины настоящего изобретения предназначены для введения субъекту для выработки желательного терапевтического эффекта. С этой точки зрения настоящее изобретение обеспечивает также:
а/ протеин настоящего изобретения для использования при терапевтическом лечении;
b/ использование протеина или конъюгата настоящего изобретения для получения медикамента для использования для снижения уровней протеина, действующего по схеме комплемента, и, в частности, для предотвращения отторжения чужеродного материала;
с/ фармацевтическую композицию, включающую один или более из протеинов или конъюгатов изобретения вместе с одним или более из фармацевтически приемлемых носителей и/или эксципиентов; и
d/ способ уменьшения уровней протеина, действующего по схеме комплемента, у млекопитающего, который включает введение этому млекопитающему протеина настоящего изобретения предпочтительно в форме фармацевтической композиции.
Фармацевтические композиции могут присутствовать в форме единичных доз, содержащих заранее определенное количество активного ингредиента на дозу. Такая единица может содержать как минимум, например, 1 мг активного ингредиента и предпочтительно 2-3 мг. Верхний предел, который может содержать такая доза, будет зависеть от многих факторов, таких как подлежащее лечению состояние, способ введения и возраст пациента, вес и его состояние, а также экономические соображения. В качестве примера, единичная дозовая форма может содержать 10 мг и даже 100 мг активного ингредиента.
Протеины настоящего изобретения можно использовать ин виво для разрушения системы комплемента. Такая необходимость может возникнуть в следующих случаях:
а/ Для предотвращения осуществляемой за счет комплемента деструкции или повреждения трансплантата, в частности ксенотрансплантата /материала, трансплантированного от другого человека или от животного/ и особенно дискордантных ксенотрансплантатов /когда виды донора и реципиента дискордантно связаны/. Реципиента следует "декомплементировать" до операции и поддерживать в этом состоянии до тех пор, пока трансплантат не приживется, либо не будет заменен более совместимым органом.
Начальную обработку можно осуществить за несколько дней до трансплантации. В моменты кризисов отторжения может потребоваться дополнительная декомплементация. Такое лечение может сопровождаться использованием антигистаминных реагентов для контроля за общими воспалительными реакциями /например, за вазодиляцией/, подобно результатам образования С3а и/или С5а.
Декомплементация может оказаться выгодной при использовании искусственных органов или тканей /например, мембран для диализа искусственной почки/, которые активируют систему комплемента. Как было указано ранее, протеин можно вводить либо в инактивированной форме, функционально C3b-подобной форме или преформе активной С3 конвертазы /подобно C3bВb/. Их можно вводить любым способом, за счет которого активированная конвертаза сможет встретить циркулирующие С3 /например, внутривенно, подкожно и т.д./.
Другой альтернативной является экс виво обработка, например, за счет трансфузии циркулирующей жидкости через матрицу, содержащую активную конвертазу. Этот способ имеет то преимущество, что позволяет анафилактические пептиды /С3а и C5a/ и другие низкомолекулярные медиаторы воспалений /например, гистамин и окись азота/ удалять /например, за счет диализа/ до того, как декомплементированная кровь /или плазма/ будет возвращена в организм пациента.
b/ Для предотвращения осуществляемых за счет комплемента повреждений после хирургических операций. Осуществляют декомплементацию пациента, как указано ранее, предпочтительно перед операцией /но при необходимости после нее/ и поддерживают это состояние до тех пор, пока не уменьшается опасность дополнительных внутренних осложнений за счет зависимых от комплемента иммунных реакций.
с/ Для минимизации связанных с комплементом повреждений, возникающих за счет травм нехирургического происхождения. В этих случаях декомплементацию следует осуществлять после исходного повреждения, но композиции и способы введения, по-видимому, должны быть в основном сходны с описанными ранее. Это может быть особенно полезным, если выздоровление включает повторную перфузию ишемических тканей за счет циркуляции /например, ишемия миокарда, обморожения, ожоги и т.д./.
d/ Для минимизации связанных с комплементом повреждений, возникающих за счет взаимодействий типа антитело - антиген. Защитные реакции, осуществляемые за счет комплемента, особенно нежелательны при автоиммунных заболеваниях, которые могут включать гломерулонефриты, гемолитическую анемию, миастению гравис и артриты, вызванные коллагеном типа II. Блокирование функций системы комплемента во время серьезных случаев заболеваний может облегчить состояние.
е/ Для того чтобы сделать специфическую патогенную мишень более чувствительной к действию иммунных механизмов, осуществляемых за счет комплемента. При таком подходе целью является не использование суперактивной С3 конвертазы для достижения полного исчерпания С3, но вместо этого использование конвертазы локально для концентрирования С3 конверсии у желательной цели. Такой целью может быть патогенный организм, например бактерия, вирус или другие паразиты, или вредные клетки или ткани хозяина, например опухолевые клетки или инфицированные вирусом клетки. С3 конвертазу можно локализовать у мишени за счет локального введения /например, за счет прямой инъекции, возможно в среду, которая задерживает его распространение в общую циркуляцию/, или за счет объединения с направляющим к мишени фрагментом, например антителом. Таким образом, модифицированный протеин можно связать со специфическим иммуноглобулином либо за счет химической сшивки протеинов, либо за счет присоединения к ДНК кодирующим последовательностям и последующей экспрессии /и выделения/ слитого белка /например, в случае IgG, либо тяжелая, либо легкая цепи могут быть соединены с С3 и могут совместно экспрессироваться с С3, либо обе цепи можно объединить в один полный слитый полипептид/, или за счет введения специфических кодирующих последовательностей /например, для доменов, похожих на "лейциновую молнию/" в ДНК обоих партнеров слияния /например, модифицированного С3 и специфического антитела/ так, что продукты экспрессии, будучи смешаны вместе, самоассоциируют с образованием стабильных конъюгатов. Затем слитый белок можно ввести локально или в общую циркуляцию.
Липосомы /содержащие антитела на поверхности с модифицированным протеином, либо на поверхности либо внутри липосомы/ и/или вирионы /например, сконструированные так, чтобы экспрессировать протеины на своей поверхности/ также можно использовать для совместной доставки антитела и модифицированного протеина. Такой подход можно использовать непосредственно, отдельно или в сочетании с другими обработками, на любой стадии развития заболевания. Это может быть особенно подходящим для исключения любых раковых клеток, которые остаются в циркуляции после хирургического удаления опухоли. Конъюгаты антитело - модифицированный протеин также можно использовать экс виво для исключения патогенных тканей. Так например, для уничтожения лейкемичных клеток в извлеченном костном мозге с последующим возвращением оставшихся здоровых клеток пациенту.
В другом варианте лимфоциты, которые не соответствуют МНС типам реципиента /основной комплекс гистосовместимости/, можно исключить из костного мозга перед трансплантацией. Кроме того, модифицированный протеин можно связать с антигеном, и такую комбинацию использовать либо ин виво, либо экс виво для подавления лимфоцитов с нежелательной реакционной способностью /например, против трансплантата или собственной ткани/.
Ту же самую технологию можно применить при лечении других видов, используя либо производные модифицированного протеина человека, либо аналоги, сконструированные специально для этих видов.
Предпочтительными характеристиками для каждого из аспектов настоящего изобретения являются те же, что и для каждого аспекта Mutatis Mutandis.
Далее настоящее изобретение будет описано со ссылкой на следующие примеры, которые не следует рассматривать как ограничивающие изобретение. Примеры сопровождаются чертежами, на которых:
Фиг. 1 представляет предсказанную последовательность протеина С3 человека, как закодировано в РС3 /использован стандартный однобуквенный код для аминокислот/.
Фиг. 2 представляет кДНК последовательность в РС3 /использован стандартный однобуквенный код для дезоксинуклеотида для смысловой цепочки, записанный как 5'-3'/.
Фиг. 3 представляет визуализацию модифицированных протеинов настоящего изобретения.
Фиг.4 представляет влияние различных мутаций на С3 человека, при которых заменяют Аrg 1303 или Аrg 1320 на расщепление по этим сайтам за счет фактора I.
Примечание:
1. [35S] - биосинтетически меченные образцы.
2. Реакции ведут при нормальной ионной силе.
3. Иммуноосаждение за счет анти-С3.
4. ПААГ - ДСН (гель - электрофорез в полиакриламидном геле с додецилсульфатом Nа) в восстанавливающих условиях.
5. Авторадиография.
Фиг. 5 представляет повышенную устойчивость С3 человека, содержащего мутацию, при которой Аrg 1303 заменяют на Сln 1303, к инактивации за счет факторов I и Н.
Фиг.6 представляет анализ расщепления С3 конвертазы, мутированной по аминокислотным остаткам 752-754 и 758-760.
Это фотография результатов Вестернблоттинга из 7,5% полиакриламидного ПААГ-ДСН геля /восстановительные условия/ после электрофоретического переноса на нитроцеллюлозу, зондирования овечьими анти-С3 человека антителами и выработки антител к антиовечьему иммуноглобулину, соединенному с пероксидазой хрена, и усовершенствованной хемилюминесценции /способ и реагенты для детектирования от Amersham, Великобритания/, зарегистрированный на ренгеновской пленке. Реакции расщепления и процедуру детектирования осуществляют в соответствии со способом примера 4 со ссылкой на результаты, представленные на фиг.3.
Ключ:
Полосы 1-4: дикого типа С3 /экспрессированы в COS клетках)
Полосы 5-8: мутантные С3 /остатки 752-754 заменены на Сlу - Ser-Gly и остатки 758-760 также заменены на Сlу--Сlу /экспрессированы в COS клетках)
Полосы 1,5: без добавления
Полосы 2,6: +CVFBb
Полосы 3,7: + факторы Н+I
Полосы 4,8: +CVFBb + факторы Н+I
Стрелками указаны полосы:
А: С3 альфа-цепь
В: С3 альфа'-цепь
С: С3 бета-цепь
Д: 68 кDа продукт расщепления С3 альфа'-цепи
Е: тяжелая цепь IgG
Фиг. 7 представляет анализ расщепления радиомеченого фактора В за счет фактора D в присутствии дикого типа и мутантных С3 /С3i/.
Представлена фотография авторадиографии ПААГ-ДСН геля.
Все образцы, содержащие фактор D и 125I-меченый фактор В, были инкубированы в течение 3 часов при 37oС.
Образцы в пронумерованных полосах включают также:
1. Один буфер
2. 1/125 дикого типа С3
3. 1/25 дикого типа С3
4. 1/5 дикого типа С3
5. 1/25 мутант С3 /остатки 1427 Gln, 1431 Asp и 1433 Gln/
6. 1/5 мутант С3
7. Неразбавленный мутант С3
Полосы, указанные стрелками:
А. Нерасщепленный 125I-меченый фактор В /93 кДа/
B. 60 кДа продукт расщепления /"Bb"/
С. 33 кДа продукт расщепления /"Ва"/
Фиг. 8 представляет ПААГ-ДСН исследования, иллюстрирующие образование конъюгата между С3i и IgG.
Это штамм Coomassic 4% акриламидного ПААГ-DСН геля, проведенного в невосстанавливающих условиях. Пронумерованные полосы содержат образцы:
1. РДР - IgG
2. С3
3. РДР - IgG + С3i реакционная смесь
Стрелками обозначены:
А. Вероятный C3i - IgG конъюгат /350 кДа/
В. С3i /200 кДА/
С. IgG /150 КДа/
Фиг. 9 демонстрирует тот факт, что конъюгат направляет С3 конвертазную активность против эритроцитов овцы. /Этот график показывает % подвергшихся лизису эритроцитов овцы после нанесения на них разбавлений либо C3i-IgG-конъюгата, PDP-IgG, или C3i с последующей промывкой, выработкой С3 конвертаз с пропердином и факторами В и D и, наконец, осуществлением лизиса за счет NGPS в CFD/EDTA, как указано в разделе методики. Только конъюгат осуществляет лизис, и этот лизис зависит от дозы/.
Нижеследующие стандартные методики и определения применимы во всех примерах.
Все компоненты комплемента, которые упоминаются, относятся к комплементу человека, если только нет других указаний; использована стандартная терминология для всех протеинов и полученных из них фрагментов /например, как указано в ссылке [15]/. Кроме того, термин "C3i " относится к любой молекулярной форме С3 без интактной тиолэстерной связи, но сохраняющей С3а полипептид на альфа-цепи.
кДНК С3 человека и кодирующая последовательность пронумерованы, как представлено на фиг.2, с использованием нумерации, принятой для EMBL нуклеотидной базы данных /из ссылки [2]/. Представленная последовательность представляет нашу конструкцию /'РС3'/, в которой отсутствуют первые 11 нуклеотидов 5' нетранслируемого участка, о котором сообщается в ссылке [2], и, следовательно, первое основание пронумеровано как 12. Предполагаемый кодон инициирования представлен нуклеотидами 61-63, кодон для аминотерминального серинового остатка бета-цепи представлен нуклеотидами 127-129, а кодон для аминотерминального серинового остатка альфа-цепи представлен нуклеотидами 2074-2076.
Последовательность протеинов пронумерована в соответствии с нумерацией последовательности-предшественника, представленной на фиг. 1, которая представляет предсказанную трансляцию ДНК последовательности /ожидается, что аминокислоты 1-22 содержат сигнальную последовательность, которая удаляется во время биосинтеза, и, как ожидается, аминокислоты 668-671 удаляются, когда предшественник расщепляется на альфа- и бета-цепи/.
Следующие сокращения имеют следующие значения:
CVF фактор яда кобры, ELIS А иммуноадсорбентный анализ со связанным энзимом;
E.coli - Escherichia coli;
к.п.о. - килопароснований;
HSV - 1-вирус герпес симплекс типа 1;
РВS - фосфатом буферированный раствор;
COS-1 представляет клеточную линию, полученную из клеток почки обезьяны.
Рестрикционные эндонуклеазы: - AflII, DraI, DraIII, EcoRI, HindIII, NaeI, XbaI, NheI. EcoRV.
Стандартные методики
Описание стандартных процедур, применяемых в молекулярной биологии, таких как выделение плазмид, электрофорез в агарозном геле и ДНК лигирование, можно найти в ссылке [21]. Двухцепочечную ДНК секвенируют, используя набор Sequenase Version 2.0, поставляемый "United States Biochemicals".
С3 экспрессию измеряют в анализе ELISA, используя пластиковые пластины, предварительно покрытые афинно-очищенными поликлональными овечьими античеловеческими С3, к которым добавлены образцы культуральных надосадочных жидкостей. Связанные С3 определяют с помощью моноклональных крысиных антител к С3, конъюгированных с щелочной фосфатазой, и хромогенным субстратом, р-нитрофенолфосфатом. Результаты анализов прокалиброваны за счет С3 очищенной плазмы человека.
Способы очистки протеинов комплемента и CVF, и для получения афинно-очищенных анти-С3 антител, использованных в анализах, можно найти в ссылке [28]. Эквивалентные реагенты можно закупить в Sigma chemical company LTD.
С3 кДНК кодирующая последовательность
С3 кДНК кодирующую последовательность настоящего изобретения конструируют из двух сегментов, выделенных из библиотеки статистически примированных кДНК печени человека, содержащихся в векторе pGEM4 / Promega/. Пять олигодезоксинуклеотидов, соответствующих известным сегментам в последовательности, кодирующей С3 человека, радиометят за счет Т4 полинуклеотидкиназы, и [γ - 32P] АТР, и используют для зондирования переноса на фильтр библиотеки с агарозных пластин. Выделяют два клона, содержащие вставки примерно в 4 к.п. о. Расщепление рестрикционными эндонуклеазами, гибридизация со специфическими олигодезоксинуклеотидными зондами и частичное секвенирование демонстрируют, что один из них /'А13'/ включает 5'-конец 5,1 к.п.о. месседжа, тогда как другой простирается до 3'-конца.
Поэтому эти вставки перекрываются примерно на 3 к.п.о. включая уникальный сайт EcoRI рестрикционного фермента. Неполный 5' отрезок А13 вырезают с помощью ЕсоRI и NheI и заменяют полным сегментом, выделенным из В44 за счет переваривания с помощью ЕсоRI и XbaI. Оба отрезка очищают с помощью гель-электрофореза в агарозе с низкой температурой плавления перед тем, как лигируют вместе с Т4 ДНК лигазой до получения вектора /'PGC3'/, содержащего 5,1 к.п.о ДНК, кодирующей полный протеин - предшественник С3.
Линкерные последовательности в направлении 5' до С3 кодирующего участка содержат два ATG, которые представляют сайты ложного начала трансляции, поэтому их удаляют за счет "гэп-пед-плазмидного" мутагенеза, как раскрыто в способе примера 1, используя олигодезоксинуклеотид РL-АТС-3 /таgggаgасс ggааgсттgс сстстссстс тgтссстстg т/, который исключает примерно 50 пар оснований линкера/адапторной ДНК, не изменяя при этом С3 кодирующей последовательности. Этот мутированный вектор в 7,7 к.п.о, который содержит 5,1 к.п.о. С3 кДНК последовательности плюс 2,6 к.п.о последовательности из PGEM4 вектора /Promega/ называют РС3.
С3 кодирующий участок PGC3 плазмиды полностью секвенируют и выявляют всего 4 отличия от ранее опубликованной человеческой С3 /"S " аллель/ кДНК последовательности [2].
/i/ изменения С2481-->G, и С2805-->Т не меняют кодирования;
/ii/ Т1001-->С кодирует ранее описанную НАV 4-1-/лейцин 314-->пролин/ полиморфную форму [20]; и
/iii/ G2716-->А кодирует валин 886--> изолейцин, который не был ранее обнаружен в С3 человека, хотя Ile найден в этом положении в мышином и крысином С3.
Последовательность настоящего изобретения включает стартовый и стоп кодоны с полной сигнальной последовательностью и должны поэтому кодировать функциональный С3.
Уровни вплоть до 1,7 мкг/мл экспрессированные дикого типа С3 в надосадочных жидкостях культур СOS-l клеток /трансфектированных с помощью липофектамина и рсDNА3 /Invitrogen/ вектора экспрессии/ были определены с помощью ELISA. Клетки, трансфектированные одним только вектором рсDNА3, не продуцируют детектируемых количеств С3. Более того, анализ продуктов, экспрессированных за счет реакций расщепления с последующим иммуноосаждением, ПААГ-DСН и иммуноблоттингом демонстрирует, что:
i/ продукт первичной трансляции был правильно процессирован в зрелую двухцепочечную форму;
ii/ этот продукт был, подобно нативному С3, расщепляем до C3b за счет С3 конвертазы /CVFBb/; и
iii/ экспрессированный протеин был, подобно нативному С3, нерасщепляем факторами Н плюс I, но становился расщепляемым после конверсии до С3b за счет С3 конвертазы. Это подтверждает, тот факт, что исходную плазмиду можно транслировать в функциональный С3.
Для альтернативного описания конструирования и экспрессии С3 кодирующей последовательности см. ссылку [25].
Пример 1
Получение С3, который имеет аргининовые остатки по обоим сайтам расщепления фактора I /положения аминокислот 1303 и 1320/, превращенные в глутаминовые остатки для предотвращения расщепления C3b фрагмента за счет фактора I.
а/ Мутагенез
Были использованы мутагенные олигодезоксинуклеотиды
QRI1 /саастgсссаgссаааgстссааgатсасс/,
QBI2 /gссаgсстсстgсаатсаgааgаgассааg/ и
AFL 4149 /таатаааттсgассттааggтсассатаааас/,
а также соответствующие антисмысловые олигодезоксинуклеотиды
QPI1n /ggтgатсттggаgстттggстgggсаgттg/,
QRI2n /сттggтстсттстgаттgсаggаggстggс/ и
AFL 4149n /gттттатggтgассттааggтсgаатттатта/.
QRI1 и QRI1n определяют замену аргинина на глутамин по сайту расщепления фактора I у аминокислотного остатка 1303 в последовательности С3 предшественника /за счет изменения G3968С3969 на АА в кДНК последовательности/, и QRI2 и QRI2n осуществляют то же самое замещение по сайту расщепления фактора у аминокислотного остатка 1320 /за счет замены нуклеотида G4019 на А/.
AFL 4149 и АFL 4149n вводят сайт расщепления для рестрикционной эндонуклеазы AflII в положении 4149 в кДНК последовательности /за счет замены С4149 на Т/, без изменения кодируемой аминокислотной последовательности. Эти два праймера используют в качестве маркеров, что обеспечивает успешную идентификацию мутагенеза на основании расщепления ДНК продукта за счет AflII.
Мутагенез осуществляют, используя метод "разорванной плазмиды". Порцию PGC3 /'UPGC3'/, обогащенную уридином вместо тимидина, получают, выращивая в E. coli штамме CJ236 в присутствии 0,25 мкг/мл уридина. Эту плазмиду расщепляют SmaI и выделяют из агарозного геля продукт 7,2 к.п.о. /'US1'/ для удаления 0,5 к.п.о. фрагмента из С3 последовательности /остатки 1463-1747/. Остальные компоненты геппед плазмиды /'СN2'/ получают расщепляя PGC3 за счет DraIII плюс NaeI и дважды очищая фрагмент 5,1 к.п.о. с помощью электрофореза на агарозном геле. 200 нг DN 2 смешивают с примерно 500 нг USl в 50 мкл Н2О, нагревают до 100oС и медленно охлаждают до температуры ниже 50oС, перед тем, как добавляют 20 мкл до 25 мкл 2ХТ7 буфера /100 мМ Tris /HCl/ pH 7,4, 14 мМ MgCl2, 100 мМ NaCl, 2 мМ дитиотреитола и 1 мМ каждого из АТР, dАТР, dСТР, dТТР и dGТР/ плюс 10 нмолей каждого 5'-фосфорилированного мутагенного праймера /в одной реакции используют QRI1, QRI2 плюс AFL 4149, в другой реакции используют QRI1n, ОRI2n плюс AFL 4149n/. Полученные смеси снова нагревают в течение 5 минут и медленно охлаждают /за 30-60 минут/ до 20oС. При температуре 0oС добавляют 10 единиц Т7 ДНК полимеразы плюс 80 единиц Т4 ДНК лигазы. Полученную смесь /полный объем 50 мкл/ инкубируют вначале при 0oС в течение 5 минут, затем при комнатной температуре в течение 5 минут и, наконец, при 37oС в течение 3 часов. По 1 мкл каждой смеси используют для трансформации 100 мкл суперкомпетентной XL1 E.coli /Stratagene/ в соответствии с инструкциями изготовителей.
Устойчивые к ампициллину колонии скринируют на АflII расщепление, и удачные мутанты выращивают в 100 мл культурах, из которых выделяют плазмиды и осуществляют секвенирование /используя секвенирующий праймер С3ра-3876, сттсатggтgттссаат cст, соответствующий нуклеотидам 3876-3895 С3 кДНК/ для характеристики мутаций по сайту расщепления фактора I.
Альтернативную схему для "гэппед плазмидного" мутагенеза можно найти в ссылке [26, 27].
b/ Перенос мутантной ДНК в эукариотный вектор экспрессии
С3 кодирующие фрагменты из мутантных плазмид вырезают двойным расщеплением с помощью HindIII и NaeI. Включают также DraI для инактивации оставшейся плазмиды. С3 кодирующую последовательность очищают на агарозном геле и лигируют в рсДНК3 вектор /pcDNA3, Invitrogen/, который был линеаризован за счет HindIII и ЕсоRV энзимов и дефосфорилирован фосфорилазой кишечника теленка. Смеси лигирования используют для трансформации суперкомпетентной XL1 E.coli, которую затем высевают на культуральные пластины, содержащие ампициллин.
Случайно выбранные /три из четырех/ устойчивые к ампициллину колонии выращивают в 2-3 мл культуры, и выделяют плазмидную ДНК. Плазмиды, содержащие правильную вставку, определяют с помощью расщепления плазмидной ДНК рестрикционными эндонуклеазами EcoRI, HindIII и AflII. Соответствующие колонии выращивают в 100 мл культур, и плазмиды выделяют стандартными способами. Эти мутанты исходно конструируют из PGC2, и поэтому они сохраняют два ATG в направлении 5' к кодирующему участку. Этот участок /плюс 5' 3 к.п.о. С3 кодирующей последовательности/ вырезают HindIII плюс EcoRI, и заменяют за счет лигирования тем же самым фрагментом, вырезанным из РС3. Реконструированные таким образом векторы получают стандартными способами и используют для трансфекции COS клеток.
с/ экспрессия дикого типа и мутантных С3
Мутантные и дикого типа С3, кратковременно экспрессированные из плазмид, трансфектируют в СОS-1 клетки, используя липофектамин /G1BCO/ в соответствии с инструкциями изготовителей. Обычно 1-1,5•105 клеток на ячейку стандартных культуральных пластин с 6 ячейками трансфектируют 2-4 мкг плазмид, используя 9 мкл реагента липофектамина. Надосадочные жидкости анализируют на С3 секрецию, и обычно выходы 0,3-1,7 мкг на мл надосадочной жидкости получают через 3-6 дней после трансфекции.
Результаты
а/ Получение мутантов
Были выделены следующие мутанты, названные в соответствии с мутагенными олигодезоксинуклеотидными последовательностями, которые были встроены:
/i/ 3 мутанта как с QRI1, так и с QRI2 мутациями, плюс АFL 4149: С3М-26, С3М-58 и С3М-61;
/ii/ 1 мутант с QRIl и QRI2, но без AFL4149: С3М-8; и
/iii/ 1 мутант с QRI2 и AFL 4149, но без QRI1: С3М-51 /который используют в примере 3/
b/ Подтверждение того, что функциональные эффекты связаны с мутациями, специфически введенными по сайтам расщепления фактора I.
Секвенирование подтверждает отсутствие других изменений в 178-350 основаниях вокруг мутированного участка каждого из мутантов. Последовательность одного мутанта, полученного таким образом, С3М-51 /см. пример 3/ была проанализирована по всему разрыву /основания 2463-5067/, использованному в мутагенезах, и не было обнаружено никаких других отклонений от последовательности дикого типа.
Кроме того, представительное секвенирование полных 2922 оснований из всех мутантов не выявило никаких одноточечных мутаций, которые могли бы быть вызваны ошибками за счет полимеразной активности. Все экспрессированные мутанты демонстрируют двухцепочечную структуру и расщепление под действием С3 конвертазы, характерное для нативного С3. Итак, по-видимому, использованные мутанты не содержат каких-либо нежелательных изменений, хотя они и не были полностью секвенированы.
Пример 2
Получение С3, который содержит аргининовый остаток по одному сайту расщепления фактора I /положение аномикислоты 1303/, превращенному в глутаминовый остаток.
Используют способ примера 1 за исключением того, что при мутагенезе используют только мутагенные олигодезоксинуклеотиды AFL4149 плюс QRI1 или AFL4149n плюс QRI1n /то есть без QRI2 или QRI2/.
Результаты:
а/ Полученные мутанты
Мутанты с QRI1 и AFL4149, но без QRI2 были выделены: С3М-123, 27. Экспрессируют мутант С3М-123 по способу примера 1. Этот протеин расщепляют CVFBb. C3b-подобный продукт оказывается относительно /по сравнению с диким типом/ устойчив к расщеплению в положении 1303 за счет факторов I и Н, но все еще может расщепляться в положении 1320. Это C3b производное поэтому частично устойчиво по отношению к фактору I.
Пример 3
Получение С3, который имеет аргининовый остаток по одному сайту расщепления фактора I /положение аминокислоты 1320/, превращенному в глутаминовый остаток.
Используют способ примера 1 за исключением того, что при мутагенезе используют только мутагенные олигодезоксинуклеотиды AFL4149 плюс QRI2 или AFL4149n плюс QRI2n /то есть без QRI1 или QRI1n/. Кроме того, в результате использования способа примера 1 получают также один мутант с QRI2 и AFL4149, но без QRI1.
Результаты.
а/ Полученные мутанты
Выделены 3 мутанта с QRI2 и AFL4149, но без QRI1: С3М-51, C3M-Q2, C3M-Q13. Мутант С3М-51 Экспрессируют по способу примера 1. Этот протеин расщепляется CVFBb. C3b-подобный продукт легко не расщепляется в положении 1320 под действием факторов I и Н, но его все еще можно расщепить в положении 1303. Поэтому это C3b производное частично устойчиво по отношению к фактору I.
Пример 4
Анализ функциональных эффектов мутаций.
Надосадочные жидкости /100-400 мкл/ с трансфектированных COS клеток инкубируют при 37oС в течение 2 часов с COS клетками, которые были трансфектированы рсDNА3, содержащими вставки:
1/ C3 последовательности без мутаций;
2/ мутантных С3М-123 /кодирующих Arg1303-->Gln/;
3/ мутантных С3М-26 /кодирующих Arg1303-->Gln, Arg1320-->Gln/; и
4/ мутантных С3М-51 /кодирующих Arg1320-->Gln/.
По 200 мкл надосадочных культур, взятых через 3 дня после трансфектации, предварительно обрабатывают 2 мМ фенилметан-сульфонилфторида /0oС, 15 минут/, а затем инкубируют при 37oС в течение 2 часов со следующим:
А/ без добавления;
В/ предварительно полученной C3 конвертазой, CVFBb /10 мкл из 200 мкл, содержащих 6,6 мкг CVF, 100 мкг фактора В и 1,4 мкг фактора D в фосфатном буферированном растворе /PBS/, содержащем 10 мМ MgCl2, предварительно инкубированном при 37oС в течение 15 минут/;
С/ факторами Н /5 мкг/ и I /1 мкг/; и
Д/ CVFB плюс факторы Н и I.
Затем все это иммуноосаждают за счет добавления 0,6 мкг афинно-очищенного овечьего анти-C3 человеческого иммуноглобулина при комнатной температуре и добавлением через 1 час 20 мкл 5% суспензии промытых формалином фиксированных клеток Streptococcus sp. группы С /протеин С// Sigma/. Через 45 минут при комнатной температуре частицы промывают один раз в PBS, 5 мМ NаN3, и один раз в 20 мМ Tris/HCl, 137 мМ NaCl, 0,1% /объем/объем/ Tween 20, рН 7,6, перед элюированием в 1% DCH /2% 2-меркаптоэтаноле /90-100oС, 5 мин/. Эти элюаты разделяют с помощью ПААГ-DCH, осуществляют электроблоттинг на нитроцеллюлозе, и С3 полосы детектируют, зондируя афинно-очищенным овечьим анти С3 человека иммуноглобулином, а затем соединенным с пероксидазой хрена обезьяним антиовечьим иммуноглобулином /Sigma/, и детектируют, используя субстраты /"Enhanced Chemiluminescence" усиленная хемилюминесценция/, субстраты, поставляемые Amersham. Представлена фотография, полученная после 2-минутного экспонирования рентгеновской пленки. Видимые С3-полученные полосы указаны отмеченными стрелками, и описаны отдельные образцы /1-4, А-Д/, отчетливая полоса на примерно 50 кДа /между полосами 46 и 68 кДа/ присутствует во всех образцах и представляет тяжелую цепь иммуноглобулина, использованного при иммуноосаждении и детектированного с помощью соединенного с пероксидазой хрена обезьяньего антиовечьего иммуноглобулина.
Результаты /см. фиг.3/
1. Все необработанные образцы /1-А, 2-А, 3-А, 4-А/ содержат полосы правильной миграции для альфа- и бета-цепей С3, что указывает на то, что все мутанты экспрессированы и посттрансляционно процессированы правильно. Присутствие полос 43 и 46 кДа в этих образцах указывает на наличие некоторой фактор Н+фактор I-подобной активности в культуральной среде. Спонтанный гидролиз С3 в течение трехдневного периода биосинтеза приводит к получению C3i, который расщепляется под действием этой активности. В не подвергшихся мутации С3 это вызывает наличие полос 43 кDa и 75 кДа /полоса на 75 кДа невидима из-за того, что она /i/ перекрыта полосой 75 кДа бета-цепи, и /ii/ антитело, которое использовали для проявления вестернблотов, обладает очень малой активностью по отношению к этой части С3 альфа-цепи - ее наличие затем подтверждается за счет повторного зондирования крысиными моноклональными антителами, "Клоном-3", которые специфичны для этого участка/. Добавление факторов Н и I без СVFВb /1-С, 2-С, 3-С, 4-С/ не расщепляет оставшиеся С3, что указывает на то, что они представляют активные С3 /тиолэстер интактен/.
2. Не подвергавшийся мутации С3 [1] расщепляют за счет CVFBb, и продукт C3b расщепляют далее эндогенными энзимами в 1-В или добавленными факторами Н и I в 1-Д. Полоса 43 кДа демонстрирует расщепление по Аrg1320, а полоса 67 кДа /обнаруживаемая при более длительных экспозициях/ указывает на расщепление по Аrg1303.
3. Мутантный С3М-123 /Arg1303-->Gln/ расщепляется за счет CVFBb, и полученный продукт относительно устойчив к активности, подобной эндогенным факторам Н и I /2-В/, при различных количествах альфа-цепи /C3b/, но все еще остается расщепляемым при добавлении дополнительных количеств факторов Н и I /2-Д/. Наличие продукта 43 кДа демонстрирует расщепление по Аrg1320 /слабая полоса на 71 кДа, представляющая другой фрагмент альфа'-цепи, становится видимой при более длительном воздействии/, но полоса 68 кДа отсутствует, что свидетельствует о том, что этот мутант устойчив к расщеплению по мутированному Gln1303.
4. Мутант С3М-26 /Arg1303-->Gln, Аrg1320-->Gln/ расщепляется за счет CVFBb и C3b-подобный продукт /альфа/ оказывается устойчивым к подобной активности эндогенных факторов Н и I /3-В/. Он оказывается также очень устойчивым к дополнительным факторам Н и I /3-Д/ по сравнению с немутированным С3 [1] и другими мутантами [2 и 4]. Присутствует небольшое количество продукта 46 кДа, что указывает на некоторое расщепление по мутированному Gln1303 /сопровождающий 68 кДа фрагмент также становится видимым при более длительном воздействии/. Присутствует немного или вовсе не детектируется полоса 43 кДа, которая могла бы соответствовать любому расщеплению по Gln1320. Поэтому мутация Аrg-->Gln в положении 1303 менее эффективна, нежели мутация в положении 1320 при предотвращающем расщепление за счет фактора I. Такое медленное остаточное расщепление может также происходить в мутанте С3М-123 /Аrg1303-->Gln/, но промежуточная полоса 46 кДа вероятно быстро процессирует до 43 кДа за счет дальнейшего расщепления по немутированному Arg1320.
5. Мутант С3М-51 /Аrg1320-->Gln/ расщепляем за счет CVFBb, а полученный продукт расщепляется под действием подобной эндогенному фактору Н и I активности /4-В/, и за счет дополнительных факторов Н и I /4-Д/ продукт 46 кДа /и слабая полоса 68 кДа/ указывают на расщепление по Arg1303. Однако отсутствие полосы 43 кДа свидетельствует о том, что расщепление по мутированному Gln1320 отсутствует.
Пример 5
Сравнение различных аминокислотных замещений в положении 1303
1. Введение
Предыдущие примеры описывали мутации по Аrg 1303 и Аrg 1320 на глутаминовые остатки. Обе мутации придавали устойчивость к расщеплениям в этих положениях под действием фактора I. Однако существует небольшая, но детектируемая степень расщепления по Gln 1303. Поэтому некоторое количество других аминокислотных замещений в этом положении было осуществлено и протестировано. Расщепления происходят в убывающем порядке эффективности, когда остатком 1303 является: Аrg>Туr>/Cys или Trp/Gln>Gly/ или Cly/. Эти результаты оказались неожиданными, так как /i/ все происходящие в природе расщепления за счет фактора I человека происходят С-терминально по отношению к аргининовым остаткам, так что можно было бы предположить, что ферменту необходим аргинин; и /ii/ если он действительно расщепляет по другим остаткам, то можно предсказать, что они должны быть электростатически сходны с аргинином, т. е. должны быть основным остатком /Lys или His/ /например, трипсин селективно расщепляет С-конец до Аrg, Lys или His/, так что нельзя предсказать расщепления тирозинового замещения.
Поэтому замещение Аrg 1303 глицином или глутаминовой кислотой предпочтительно для целей создания производных С3, устойчивых к инактивации за счет фактора I.
2. Методики
2.1. Мутагенез: в качестве дегенеративного мутагенного праймера используют:
саастgсссаgс(gт)(аg)(сg)аgстссааgатсасс /буквы в скобках обозначают смесь оснований в этом положении/. Мутанты конструируют либо способом с использованием "гэппед-плазмид" /как было указано ранее в примерах/, либо с помощью "мегапраймерного метода" /V. Picard et al. Nuс. Asid Res., 22: 2587-91 /1994//, в котором праймером в обратном направлении был праймер сассаggаастgаатстаgатgтgтссстс, а праймером в прямом направлении был праймер gттттатggтgассттааggтсgааттт атта. Все мутации осуществляют на матрицах, в которых С3-кодирующая ДНК уже подвергнута мутации, так что аминокислотный остаток 1320 является глутамином, а рестрикционный сайт для AflII встроен в положение 4149 /как указано ранее в примерах/, что подтверждает ДНК секвенирование.
2.2. Экспрессия: мутанты были экспрессированы в СOS клетках с помощью pcDNA3 вектора, как указано ранее в примерах, биосинтетически помечен [35S] метионином в не содержащей сыворотки среде.
2.3. Анализ: надосадочную жидкость обрабатывают CVFBb /образующимся в реакции CVF с факторами В и D в магнийсодержащем буфере/ и факторами Н и I с последующим иммуноосаждением за счет анти-С3, и выделением с помощью электрофореза на DСН-полиакриламидном геле, осуществляемом в восстанавливающих условиях /как указано ранее в примерах/. Этот гель фиксируют, обрабатывают реагентом "Amplidy" /Amersham/, сушат и экспонируют на авторадиографической пленке до получения результата, представленного на чертеже.
3. Результаты
Расщепление в положении 1303 /сайт 1/ без расщепления по 1320 /сайт 2/ /который был мутирован до глутамина/ за счет фактора I приводит к образованию полос 46 и 68 кДа. Можно видеть, что расщепление происходит в порядке: arg(R)>tyr(Y)>cys(C) u trp(W)>gln(Q)>gly(G) u glu(E). Дикий тип /аргинин в обоих положениях/ расщепляется по обоим положениям с образованием фрагментов 43 кДа /слишком мал, чтобы обнаружить его в геле/ и 68 кДа.
4. Рисунок
Результаты представлены на фиг.4. Остатки в сайте 1 /положение 1303/ и сайте 2 /положение 1320/ указаны над соответствующими полосами.
Пример 6
Демонстрация повышенной устойчивости к инактивации за счет факторов I и Н после мутации arg 1303 в gln.
1. Введение
В предыдущих примерах было продемонстрировано, что конверсия либо arg 1303, либо arg 1320 в глутамин делает этот сайт устойчивым к расщеплению с помощью фактора I. Мутации обоих сайтов создают молекулу, которая устойчива к расщеплению по любому сайту. Здесь далее демонстрируется, что мутация argl303 только до gln /без изменения на arg 1320/ приводит к значительной устойчивости по сравнению с диким типом, к функциональной инактивации за счет факторов I и Н.
2. Методы
2.1. Экспрессия: получение мутации arg 1303-->gln описано в предыдущем примере. Осуществляют трансфекцию в СНО - обычная лабораторная клеточная линия, полученная из клеток яичников китайского хомячка методом с использованием кальций-фосфата и стабильные трансфектанты отбирают на основании устойчивости к С418 /"Geneticin", доступный от Sigma/. Надосадочные жидкости клеточных культур собирают, и экспрессированный С3 частично очищают за счет осаждения натрийсульфатом /10-20% /вес/объем/ фракции/, и ионообменной хроматографии на Q-сефарозе и моно-Q-сефарозе /A.W. Dodds Methods Enzymol 223:46 /1993//.
2.2. Анализ: Овечьи эритроциты покрывают S016 моноклональными антителами /R. A. Harrison and P.J. Lachmann Handbook of Experimental Immunology 4 Ed. Chpt, 39 /1986/ и 4,4 мл 5% /объем/объем/ суспензии затем инкубируют с примерно 10 мкг С2, 24 мкг С4 и 1 мкг Сl /очищенные человеческие компоненты/ в течение 10 минут при 37oС в CFD /R.А. Harrison and P.J. Lachmann/. 0,8 мл этой смеси инкубируют затем в течение 105 минут с 0,25 мл, содержащими полуочищенный мутантный или дикого типа С3 и ЕDТА до конечной концентрации 12,5 мМ. Затем клетки промывают в CFD, и используют в CFD, содержащей 0,1% /вес/объем/ желатина /СFD-гель/. Для подтверждения того, что отложились аналогичные количества дикого типа или мутантного C3b, осуществляют радиолигандное связывание 125I-меченым клона 4 моноклональным анти-С3 антителом.
Для анализа 40 мкл 5% суспензии клеток разбавляют в 250 мкл СFD-геля, и 50 мкл аликвоты инкубируют с 50 мкл CFD-гельсодержащими разбавлениями факторов I и Н до конечной концентрации 100, 10, 1 и 0 мкг/мл каждый, при 37oС в течение 30 минут. 0,9 мл CFD затем добавляют, клетки осаждают центрифугированием и дважды промывают порциями по 1 мл CFD. Затем эти клетки снова суспендируют в 100 мкл CFD-геля, содержащего 100 мкг/мл фактора В, 100 мкг/мл пропердина, 1 мкг/мл фактора D и 0,3 мМ NiCl2. Спустя 10 минут при 37oС добавляют 0,9 мл CFD, содержащего 10 мМ ЕDТА и 2% /объем/объем/ нормальной сыворотки морских свинок. После еще 30 минут при 37oС нелизированные клетки осаждают центрифугированием, и степень лизиса определяют, измеряя поглощение надосадочной жидкости на 412 нм. Поглощение, эквивалентное 100% лизиса, определяют для аликвот клеток, лизированных в воде, и далее определяют процент лизиса.
С помощью такого анализа определяют способность отложившихся С3 образовывать функциональную C3bВbР конвертазу. Конверсия до C3b предотвращает образование конвертазы и последующий лизис в смеси сыворотка/EDTA.
3. Результаты
Результат, представленный на чертеже, указывает, что по сравнению с диким типом необходимо десятикратное превышение фактора I и фактора Н для прекращения гемолитической активности аrg 1303-->gln мутанта. Поэтому такая мутация выгодна для создания производного С3, продукт которого C3b устойчив по отношению к инактивации за счет факторов Н и I. Эффект может быть связан либо с большей устойчивостью к расщеплению в положении 1303 /если arg мутирован до gln/, либо с большей устойчивостью к расщеплению в положении 1320, если расщепление вначале происходит в положении 1303.
4. Чертеж
Полученные результаты представлены на фиг.5. По оси Х отложены концентрации факторов Н и I. Q1 представляет 1303-->gln мутацию, % лизиса определяют способом, раскрытым в разделе методики.
Обсуждение
Отличительными особенностями С3 человека в отношении описываемых здесь модифицированных вариантов являются следующие:
i/ У молекулы есть производное, функционально похожее на C3b, в том плане, что оно может объединяться с функционально активным фактором В человека, который затем может быть расщеплен фактором D человека с образованием энзима, способного расщеплять С3 человека.
ii/ Аминокислотные последовательности более гомологичны С3 человека нежели С3 любых других видов, для которых в настоящее время эти последовательности известны, или с любыми другими известными в настоящее время последовательностями. Структурные характеристики С3, которые присутствуют в протеине дикого типа, но не обязательно в модифицированных производных, включают следующие:
а/ ДНК кодирующая последовательность и транслированная последовательность протеина для варианта С3 человека, использованного в примерах настоящего изобретения и здесь описываемого, представлены на фиг.2 и 1 соответственно. Эта последовательность протеина отличается от опубликованной последовательности [2] только двумя аминокислотами /подробности представлены в примерах/. Можно предположить, что гораздо больше вариантов совместимы с С3 функциями, даже если большинство не будет присутствовать в популяции.
b/ Продукт первичной трансляции протеолитически процессирован в две дисульфидно-связанные цепочки, альфа /остатки 672 - 1663/ и бета /остатки /23-667/, причем удалена сигнальная последовательность /остатки 1-22/.
с/ Зрелый протеин содержит тиолэстерную связь между остатками Суs1010 и Gln1013.
d/ С3 конвертазы расщепляют С3 с удалением С3а /остатки 672-748/. Эта реакция сопровождается разрушением тиолэфирной связи.
е/ В присутствии фактора Н фактор I расщепляет C3b между остатками Arg 1303 и Ser 1304, и между Arg 1320 и Ser 1321.
Модификации нативной С3 молекулы
Замена Arg 1303 на Gln
Эту модификацию осуществляют по одному сайту расщепления C3b за счет фактора I. Эффект состоит в уменьшении степени расщепления за счет фактора I в этом положении. Замену на глутамин выбирают с тем, чтобы удалить положительный заряд аргинина, который, по-видимому, важен для активности сериновой протеазы фактора I, причем при этом сохраняется гидрофильный характер и аналогичный размер боковой цепи, что может свести к минимуму любые нарушения третичной структуры протеина. Доказательством, подтверждающим это предположение, является то, что эта мутация не предотвращает процессинг в двухцепочечную структуру, образование тиолэстера или расщепление С3 за счет С3 конвертазы. Мутацию Arg 1303 в другую аминокислоту можно осуществить аналогично, или даже более эффективно, что продемонстрировано в примере 5.
Возможно также уменьшить такое расщепление за счет мутации Ser 1304 /с другой стороны сайта расщепления/ или других остатков, вовлеченных во взаимодействие фермент - субстрат.
Замена Аrg 1320 на Gln
Эту модификацию осуществляют по другому сайту расщепления C3b за счет фактора I. Результатом является резкое уменьшение /практически, исключение/ степени расщепления за счет фактора I в этом положении. Замену на глутамин осуществляют на основании указанных выше критериев, и эта мутация также не предотвращает процессинг в двухцепочечную структуру, образование тиолэстера или расщепление С3 за счет С3 конвертазы. И снова, мутации до другой аминокислоты могут привести к тому же результаты, что и мутация Ser 1321 или других остатков, вовлеченных во взаимодействие энзим - субстрат.
Если эти две мутации, Arg 1303 - Gln и Аrg 1320 - Gln, осуществлены вместе, возникает защита С3 от инактивации, и, следовательно, сохраняется его способность образовывать часть активной C3bBb конвертазы. Другие мутации /включая комбинации мутаций/, которые исключают обе реакции расщепления, также можно использовать /например, Arg 1303Gln или Arg 1303 Gly можно использовать в сочетании с Аrg 1320 Gln/.
Пример 7
Различные мутации, которые уменьшают взаимодействие С3b/C3i с фактором Н
7.1. Введение
В других лабораториях были получены доказательства, основанные либо на эффектах синтетических пептидов /Ganu, V.S. and Muller-Eberhard, H.J., 1985, Complement 2: 27; Becherer, J.D. et al., 1992, Biochemistry 31: 1787-1794/ или ограниченного мутагенеза /Tanigu chi-Sidle, A. and Isenman, D.E., 1994, J. Immunol. 153:5285-5302/, позволившие предположить, что остатки 752-761 в первичной последовательности С3 транскрипта /см. фиг.1/ могут быть вовлечены во взаимодействие с фактором Н. Однако другие опубликованные доказательства предполагают, что только остатки 767-776 вовлечены во взаимодействие с фактором Н, тогда как остатки 752-761 важны для взаимодействия с фактором В /Fishelson 1991, Mol. Jmmunol, 28:545-552/. Мы подозреваем, что более интенсивный мутагенез этого участка может понизить афинность к фактору Н, и поэтому было бы желательно для целей создания С3 производных, чтобы он был устойчив по отношению к фактору Н. Кроме того, предполагается, что важными остатками, которые следует подвергнуть мутации, должны быть известные аминокислотные остатки /аспарагиновой и глутаминовой кислот/ и что было бы желательно изменить их на нейтральные остатки, которые, по-видимому, менее вероятно участвуют в сильных взаимодействиях. В этом примере осуществлена замена остатков 752-754 с Asp-Glu-Asp на Gly-Ser-Gly, в сочетании с изменением остатков 758-760 с Glu-Clu-Asn на Gly-Ser-Gly. Такой продукт демонстрирует пониженные характеристики расщепления, соответствующие снижению подверженности расщеплению под действием фактора Н. Это представляет доказательство того, что С3 можно модифицировать так, чтобы уменьшить связывание фактора Н и, следовательно, подверженность действию факторов Н и I. Эти модификации желательны для создания С3 конвертазы, которая стабильна в физиологических условиях.
7.2. Методика
Методики мутагенеза, экспрессии и анализов были описаны в предыдущих примерах. Мутагенный олигонуклеотид, который был синтезирован, имеет следующую последовательность:
аgтаасстgggттсgggсатсаттgсаggатсgggсатсgтттсс.
7.3. Результаты
Результаты реакций расщепления представлены на фиг.6. Они свидетельствуют о том, что:
1. Добавление CVFBb к дикого типа С3 приводит к исключению альфа-цепи /полоса 2/, так как C3b, который образуется, подвержен действию малых концентраций фактора I и Н в надосадочной жидкости культур. С3i, который был образован во время экспрессии или во время этой последующей инкубации был разрушен до iC3i таким же способом. Добавление экзогенных факторов I и Н /полосы 3 и 4/ поэтому не отличается от полос 1 и 2 соответственно, так как сама среда содержит достаточное количество активности факторов Н и I для осуществления полного расщепления.
2. Напротив, обработка мутанта С3 за счет CVFBb /полоса 6/ не приводит к исчезновению альфа-цепи. Имеется некоторое количество альфа', соответствующих C3b, но некоторое количество их или все они остаются, что указывает на то, что присутствие альфа-цепи является не просто результатом отсутствия расщепления за счет СVFBb. Остающаяся нерасщепленной альфа-цепь в полосе 2 может поэтому представлять С3i, который не был расщеплен под действием эндогенных активностей факторов Н и I, хотя возможно также, что часть из них представляет нативный С3, продолжающий существовать, если мутант обладает приобретенной частичной устойчивостью к СVFBb. Добавление высоких концентраций экзогенных факторов Н и I /полосы 7 и 8/ приводит к истощению альфа- и альфа'-цепей, указывая на то, что /i/ мутант не полностью устойчив по отношению к этим факторам, и что
/ii/ альфа-цепь, не расщепленная под действием СVFBb в полосе 2, получена, главным образом, из С3i /который расщепляем под действием факторов Н и I, но не под действием СVFBb/, скорее, нежели из нативного С3 /который расщепляем под действием CVFBb, но не под действием факторов Н и I/. Все еще расщеплены не все альфа-цепи, даже в полосе 8, вероятно, благодаря устойчивости к факторам Н и I.
Поэтому мутация остатков 752-754 и остатков 758-760 может создать молекулу С3, которая все еще будет расщепляться под действием С3 конвертазы, но которая частично устойчива действию факторов Н и I. Учитывая данные других публикаций, это наиболее вероятно, так как мутации модифицировали участок, который вовлечен во взаимодействие с фактором Н, и, следовательно, привели к уменьшению афинности для фактора Н.
Пример 8
Сайт в С3, который можно подвергнуть мутации для модификации взаимодействия С3i с фактором В
8.1. Введение
Предыдущие примеры продемонстрировали, что мутации до С3 могут модулировать взаимодействия с факторами Н и I. Для того чтобы обнаружить другие сайты в С3, которые могли бы взаимодействовать с фактором В, проведено сравнение известных последовательностей С3 молекул из различных видов, а также с доступными последовательностями для С4 и других гомологичных протеинов. Был идентифицирован участок, соответствующий остаткам 1427-1433 С3 человека, который может быть вовлечен во взаимодействие с фактором В /или его гомологом, С2, в случае С4/, но не обязательно, так как другие потенциальные функции включают образование тиолэстера, конверсию в C3b /или С4b форму/, взаимодействие с субстратом С3 и/или С5 в активности конвертазы и взаимодействие с фактором I и его кофакторами. Поэтому выбранные остатки были мутированы до соответствующих остатков на основании сравнительного анализа последовательностей, обнаруженных в другом гомологичном протеине, в данном случае в С5 человека. Таким образом, остаток 1427 был изменен с Arg на Gln, остаток 1431 с Lys на Asp, а остаток 1433 с Glu на Gln. Полученный мутант, как было обнаружено, подвержен расщеплению С3 конвертазой /CVFBb/, а С3b продукт расщепляем факторами Н и I. Однако этот мутант не поддерживает конверсию фактора В в Bb плюс Ва, что зависит от связывания фактора В с С3i /или C3b/. Поэтому имеется доказательство того, что мутация этого участка уменьшает взаимодействие с фактором В. Хотя это и нежелательно для создания суперактивной конвертазы С3, это обеспечивает указание на то, что другие модификации этого участка С3 также изменяют взаимодействие с фактором В, и некоторые из них смогут, вероятно, повысить афинность. Как следствие, такие мутации могут также повысить стабильность и активность бимолекулярного конвертазного энзима, C3bBb /или С3 Вb/.
8.2. Методики
Сопоставления, представленные в таблице, иллюстрируют, почему этот участок рассматривается как кандидат для осуществления мутаций. Предполагается, что характеры некоторых остатков высоко консервативны в С3 и С4, но сильно различаются в других протеинах. Остатки 1427, 1431 и 1433 были выбраны на основании того, что природа их зарядов может быть показателем групп, вовлеченных во взаимодействия протеин - протеин. В соответствующих остатках С5 человека были осуществлены изменения, так как они демонстрируют очень значительное отличие в электростатических характеристиках, но остаются в контексте некоторых других консервативных остатков, что может указывать на аналогичную локальную структуру.
Методики мутагенеза, экспрессии и анализа реакций расщепления С3 были описаны в предыдущих примерах /примеры 1-4/. Был синтезирован мутагенный олигонуклеотид с последовательностью:
тggтgттgассаатасатстссgастатсаgстggасаа.
Анализ функционального цикла фактора В
Экспрессированный продукт выделяют из COS клеточной среды за счет афинной очистки на колонке Clone-3-Sepharose, как указано в примере 9. Этот способ приводит к существенной конверсии формы разрушенного тиолэстера, С3i. Дикого типа С3 выделяют тем же способом, осуществляют циклы очистки на SDS - PAGE геле /восстанавливающие условия/ разбавлений дикого типа С3 /1/5, 1/25 и 1/125/, наряду с мутантным С3, и окрашивание серебром показывает, что мутант присутствует в концентрации, эквивалентной несколько менее чем 1/25, но гораздо более, нежели 1/125 разбавление для дикого типа. Те же самые разбавления используют для анализа функционального цикла фактора В. 5 мкл этих С3 инкубируют с 25 мкл СFD-6, содержащего 5 мкг/мл фактора Д, и примерно 1,6 мкг/мл 125I-меченого фактора В /примерно 1000-2000 число распадов /мин/мкл// в течение 3 часов при 37oС. Затем образцы анализируют с помощью ПААГ-DСН /восстанавливающие условия/ с авторадиографией высушенного геля. Результаты представлены на фиг.7.
8.3. Результаты
Как видно на фиг.7, отчетливое расщепление фактора В происходит даже при 1/125 разбавлении дикого типа С3 /С3i/. Напротив, не наблюдается заметного расщепления в присутствии мутантного С3, даже для неразбавленного варианта, когда концентрация должна быть выше, чем 1/125 для образца дикого типа. Поэтому этот мутант, по-видимому, обладает приобретенной способностью поддерживать расщепление фактора В, вероятнее всего за счет восстановления его связывающей афинности для фактора В. Вот почему именно этот участок С3 следует подвергать мутациям для модулирования взаимодействий между C3i /или C3b/ и фактором В и, вероятно? также стабильности конвертазы /C3iBb или C3bВb/.
Пример 9
Очистка экспрессированных мутантных С3 молекул
9.1. Введение
Этот пример демонстрирует, как мутантные молекулы С3 можно выделить из экспрессионной среды, например культуральной среды трансфектированных эукариотичных клеток. За счет простой афинной очистки молекулы С3 получают с достаточной степенью чистоты для функциональных тестов и для конъюгации с антителами способом, описанным в примере 10. Хотя элюирование из антител сопровождается гидролизом значительной части внешнего тиолэфира, продукт С3i все еще остается подходящим предшественником для создания активной С3 конвертазы, а также для получения C3i-антитело конъюгатов. Такой подход, по-видимому, также может быть полезным как часть подготовки, необходимой для использования ин виво.
9.2. Методика
Афинное очищение на клон-3 Сефарозе
Клон-3 представляет собой крысиные моноклональные антитела, которые специфичны для С3 и его производных, включая C3b и C3i /Lachmann, P.J. et al., 1980, J.Immunol. 41:503-515/. Доступны также другие моноклональные антитела к С3, и в некоторых случаях они были с успехом использованы для выделения С3 на малых количеств плазмы человека /Dodds, A.W., 1993, Methods Enzymol. 223: 46-61/ и поэтому их, по-видимому, также можно использовать для выделения молекул, экспрессированых экс виво. IqG фракцию соединяют с Sepharose СL-4В, используя цианогенбромид /методика описана у Harrison and Lachmann, 1986, Handook of Experimental Immunology 4 ed., Eds Weir, Herzenberg, Blackwell and Herzenberg; Blackwell, Oxford/. Надосадочные культуральные жидкости либо пропускают непосредственно через колонку этой смолы /рециркуляция/, либо вначале концентрируют, осаждая за счет 25% /вес/объем/ Nа2SO4, и затем повторно солюбилизируя и осуществляя диализ в PBS, 5 мМ NаN3. Затем колонку промывают последовательно / i / PBS, 5 мМ NaN3 и / ii/ РВS, содержащим 1 M NaCl. Связанный С3 элюируется за счет 50 мМ натрийборатного буфера, рН 10,5, и его немедленно нейтрализуют, собирая 0,9 мл фракции в 0,1 мл 1 М Tris /HC1, рН 7. Затем этот материал диализуют в РВ S, 5 мМ.
Получение С3, содержащих " His-Tag"
"His-Tag" представляет нить гистидиновых остатков, которые демонстрируют афинность для колонки, разрушающей ионы никеля. Этот способ используют с целью выделения экспрессированных протеинов. По-видимому, это может оказаться полезным для выделения экспрессированных мутантных С3 молекул, и поэтому был использован мутагенез со встраиванием для создания плазмиды, кодирующей С3 с хвостом из 6 гистидиновых остатков по карбоксильному концу /немедленно карбокси-конец до остатка 1663/. Такое положение для His-tag выбирают с тем, чтобы свести к минимуму вмешательство в синтез, складчатость, процессинг и образование дисульфидных связей образующегося С3. Остаток 1661 представляет собой цистеиновый остаток, который участвует в создании дисульфидной связи с более ранним остатком в последовательности /вероятно, Cys 1537; Dolmer, K. and Sottrup-Jensen, 1993, FEBS-Lett. 315:85-90/ и поэтому, по-видимому, может оказаться разумным осуществить вставку после этой структурной единицы. Мутацию вводят, используя методику "гэппед плазмиды", использованную в примере 1, используя синтезированный мутагенный олигонуклеотид, с последовательностью:
тgggтgссссаассатсатсатсатсатсаттgассасассссс.
Включение правильной последовательности подтверждается за счет ДНК секвенирования. Теперь эту ДНК последовательность можно перенести в вектор экспрессии. После трансфекции эукариотных клеток может оказаться возможным выделить экспрессированный С3 за счет афинности в колонке с ионами никеля или с помощью какой-либо другой матрицы со специфической афинностью для " His-Tag".
9.3 Результаты
Ряд мутантных С3 был выделен на Clone-3-Sepharose, включая те, которые описаны в примерах 1 и 2, экспрессированные в клетки СНО. Эти продукты сохраняют способность поддерживать расщепление фактора В за счет фактора D. Тот же самый метод используют для выделения мутанта, описанного в примере В2, экспрессированного в COS клетки. Серебряное окрашивание ПААГ-DСН гелей указывает, что выделенные продукты не являются чистыми на 100%, но достаточно часто их степень чистоты, по-видимому, больше или равна 50%. Это обусловлено исходными материалами, обычно содержащими менее 10 мкг/мл С3 в 10% /объем/объем/ сыворотке плода теленка плюс другие клеточные протеины. Кроме того, С3 не разлагаются в процессе выделения, а активность эндогенных факторов Н и I, по-видимому, оказывается удаленной.
Способ очистки за счет "His-Tag" включает более мягкие условия элюирования из колонки, содержащей ионы никеля. Так например, можно использовать ЕDТА. Поэтому применение этого способа к С3 должно обеспечить выделение без разрушения внутренних тиолэстерных связей.
Пример 10
Конъюгирование C3i с антителом, и использование для того, чтобы направить активность С3 конвертазы против конкретных клеток.
10.1 Введение
Одним аспектом настоящего изобретения является то, что стабильные С3 конвертазы, полученные из мутантных С3 молекул, будут вызывать усиленную С3 конверсию, что, если это локализовать в конкретном необходимом центре, они будут промотировать комплементзависимую атаку этой цели. Удобным подходом для направления реакции на цель является соединение мутантной С3 молекулы, а также ее С3i или C3b производного, с любым антителом, специфичным для нужной мишени. В этом примере демонстрируется рабочая методика для образования таких конъюгатов, которая применима к мутантным молекулам С3i или C3b и может быть использована на материале, афинно-очищенном из экспрессионной системы, даже если в этом процессе был разрушен тиолэстер С3. За счет соединения C3i с антителом, которое специфически связывается с овечьими эритроцитами, показано далее, что такой конъюгат фиксирует C3i на поверхности эритроцита, так что может быть получена конвертаза, C3iBbP, которая инициирует лизис этих клеток, когда компоненты другого комплемента поступают в форме нормальной сыворотки морской свинки /в ЕDТА для предотвращения нового образования С3 конвертаз/. Следовательно, конъюгирование с антителом можно использовать для направления С3i молекулы для инициирования комплементзависимой атаки конкретного типа клеток. В этом примере используют дикого типа C3i из плазмы человека, который образует С3 конвертазу ин витро. Ин виво дикого типа C3i и C3b разрушаются факторами Н и I. Поэтому мутантный С3, сконструированный в соответствии с целями настоящего изобретения, должен быть устойчив к действию факторов Н и I и поэтому, образуя стабильную С3 конвертазу, был бы выгоден в физиологическом контексте.
10.2. Методика
/i/ создание и выделение С3i-антитело конъюгата
В качестве антитела используют IgG фракцию, выделенную из сыворотки поликлональных кроличьих антиовечьих эритроцитов. 1.1 мг инкубируют с 75 нмолями SPDP в конъюгационном буфере, рН 7,5 /20 мМ KH2PO4, 0,12 М NaCl/, в течение 2 часов при комнатной температуре. РДР-IgG выделяют за счет гель-фильтрации на колонке Superose-6 /Pharmacia/ в фосфатном буфере, рН 7,4, содержащем 0,5 М NaCl. Восстановление образца дитиотреитолом используют для оценки 4 РДР групп, соединенных на молекулу IgG. C3i приготавливают за счет обработки очищенного С3 0,1 М метиламином, рН 7,2 /2 часа при 37oС/. Избыток метиламина удаляют за счет гель-фильтрации с последующим диализом в конъюгационном буфере. 18 нмолей C3i смешивают с 1,7 нмолями PDp-IgG в 1,26 мл конъюгационного буфера и инкубируют в течение 1 дня при комнатной температуре, а затем 1,5 дня при 4oС. На фиг.8 представлен окрашенный Кумасси синим ПААГ-DСН конъюгационной реакционной смеси, демонстрирующий появление образцов примерно 350 кДа, которые не присутствовали ни в PDP-IgG, ни в C3i. Эти образцы, частично очищают за счет гель-фильтрации на колонке Сефароза-6 в фосфатном буфере, рН 7,4, содержащем 0,5 М NaCl, а затем диализуют в PBS. Это элюируется перед С3, в объеме, из которого молекулярный вес 300-400 кДа можно оценить за счет калибровки глобулярными стандартами молекулярного веса. Концентрации конъюгата, свободного антитела и неприсоединенного С3 оценивают из ПААГ-DСН геля с окрашиванием Кумасси /невосстанавливающие условия/. Двумерный ПААГ-DСН /первое измерение невосстановленное, второе измерение восстановленное/ выявляет картину совместимую с картиной для 1:1 конъюгата между IgG и C3i.
/ii/ Демонстрация того, что конъюгат С3-антитело можно использовать для направления активности конвертазы против конкретной клетки. 20 мкл разбавлений конъюгата /0 /без конъюгата/, 1/100, 1/50, 1/10/ инкубируют со 100 мкл примерно 1% /объем/объем/ эритроцитов овец /предварительно промытых CFC/ в течение 1 часа при 37oС.
Параллельные инкубирования осуществляют с эквивалентными количествами PDP - IgG /без С3/ и одного только С3. Затем клетки промывают 4 раза в CFD и снова суспендируют до 100 мкл в CFD-G. 50 мкл этого подвергают лизису под действием 150 мкл Н2О с последующим добавлением 800 мкл CFC, содержащего 10 мМ EDTA и 2% /объем/объем/NGPS. Другие 50 мкл покрытых конъюгатом клеток инкубируют в течение 15 минут при 37oС с 50 мкл CFD-G, содержащим 190 мкг/мл фактора В, 2 мкг/мл фактора D, 20 мкг/мл пропердина и 0,6 мМ NiCl2, с последующим лизисом за счет 900 мкл CFD, содержащим 10 мМ ЕDТА и 2% /объем/объем/ NGPS. Через 30 минут при 37oС клетки осаждают центрифугированием /2000•g, около 3 минут/ и измеряют оптическое поглощение надосадочной жидкости на 412 нм, используя образцы, обработанные водой в качестве стандарта - 100% лизиса и буфер без клеток в качестве 0, рассчитывают % лизиса, что представлено на фиг. 9. Конъюгат осуществляет лизис в зависимости от дозы, тогда как ни PDP -IgG, ни один только C3i не вызывают какого-либо заметного лизиса, превышающего значения, наблюдаемые в отсутствие какой-либо обработки такого типа.
10.3. Результаты
Используемый способ доказывает успешность соединения С3 с IgG на основании того, что:
1. Появляется полоса соответствующего размера /около 350 кДа/ для 1:1 С3:IgG конъюгата, что показано за счет ПААГ-DCA на фиг. 8.
2. Двумерный ПААГ-DСН /первое измерение невосстановленное, второе измерение восстановленное/ указывает, что эти образцы содержат как IgG, так и С3i.
3. Характеристики элюирования этих образцов при гель-фильтрации снова соответствуют молекуле примерно в 350 кДа.
4. Конъюгат демонстрирует гемолитическую активность, которой нет ни у PDP-IgG, ни у С3i /фиг.9/.
Кроме того, гемолитический анализ /фиг.9/ показывает, что:
1. Специфическое антиовечий эритроцит антитело локализует C3i в мембране мишеневых клеток /овечьих эритроцитов/, что предотвращает их вымывание /в противоположность свободным C3i/.
2. Конъюгат сохраняет активность C3i в том плане, что он остается способным образовывать С3 конвертазу за счет взаимодействия с пропердином и факторами В и D.
3. Эта конвертаза может инициировать комплементзависимую атаку мишени, в данном случае за счет активации литической схемы /С5-9/ для лизиса эритроцитов.
Дополнительные данные из других лабораторий показывают, что фактор яда кобры можно соединить с антителом и что эти конъюгаты могут направить активацию комплемента против клеток конкретного типа /Vogel, 1988, Targeted. Diagn. Ther. , 1: 191-224; Muller, B. and Muller - Rucholtz, W., 1987, Leuk. Res. 11:461-468, Parker, C.J., White, V.F. and Falk, R.J., 1986, Complement 3: 223-235; Petrella, E. C. , et al., 1987 Journal. Jmmunol. Methods 104: 159-172).
Эти данные подтверждают утверждение, что С3, модифицированные таким образом, что они способны образовывать стабильную С3 конвертазу, подобно фактору яда кобры, можно использовать для того, чтобы направлять осуществляемые за счет комплемента реакции, что выявлено в настоящем изобретении.
Пример 11
Демонстрация того, что мутантные С3 молекулы вызывают метаболизм фактора В в нормальной сыворотке человека
11.1. Введение
Основной целью описываемого здесь изобретения являются инактивации комплемента из биологических жидкостей. Настоящее изобретение описывает способы получения С3 молекул, которые устойчивы к даун-регуляции за счет факторов Н и I. В этом случае они будут связывать фактор В и вырабатывать активные С3 конвертазы. Активность таких конвертаз демонстрируется в гемолитическом анализе, который использован в примере 6. Поэтому такая конвертаза будет потреблять С3. Если эта конвертаза нестабильна, она будет диссоциировать без значительной С3 конверсии. Однако это позволит обеспечить связывание свежего фактора В и его конверсию в Bb и Ва. Таким образом, мутантный С3 будет промотировать расход фактора В, приводящий в конечном счете к невозможности осуществления альтернативного пути и неспособности амплифицировать классическую схему стимуляции. Если образуется стабильная С3 конвертаза, метаболизм фактора В можно понизить, но при этом увеличится расход С3, поэтому желательны обе ситуации. В этом примере демонстрируется тот факт, что мутантные молекулы С3, которые модифицированы так, чтобы придать им устойчивость к фактору I, но без какой-либо модификации в отношении стабильности конвертазы, промотируют ускорение метаболизма фактора В в сыворотке человека. Напротив, дикого типа С3 не вызывает заметного метаболизма, предположительно это связано с тем, что дикого типа C3i быстро деградирует под действием факторов Н и I.
11.2. Методика
Мутанты получают следующим образом:
Q1R2 Аrg 1303 заменяют на Gln /пример 2/
Q1Q2 Аrg 1303 заменяют на Gln плюс Аrg 1320 заменяют на Gln /пример 1/
E1Q2 Arg 1303 заменяют на Glu, плюс Аrg 1320 заменяют на Gln /пример 5/
Все эти мутанты экспрессируют в СНО клетки, а затем очищают за счет осаждения Na2SO4, с последующей афинной очисткой на Clone-3-Sepharose, по способу примера В3. Аналогичным образом выделяют дикого типа С3 /R1R2 / по данным SDS -PAGE с окрашиванием серебром, концентрация Q1 находится между 1/5 и 1/25 от дикого типа, концентрация Q1Q2 составляет около 1/5 дикого типа, а концентрация Е1Q2 находится между 1/25 и 1/125 для дикого типа. Вероятно,
все препараты содержат основную часть тиолэстер-разрушенных молекул /C3i/.
10 мкл этих С3 препаратов инкубируют с 10 мкл раствора 10% /объем/объем/ нормальной сыворотки человека в PBS, содержащем 1 мМ MgCl2 и примерно 300 нг 125I-меченого фактора В /примерно 2-300000 распадов в минуту/ в течение 1 часа при 37oС. Затем 5 мкл анализируют с помощью SDS-PAGE /восстанавливающие условия/. Высушенный гель экспонируют на авторадиографическую пленку для того, чтобы указать положения полос, соответствующих интактному фактору В и его продуктам расщепления. Их иссекают и подсчитывают для точного определения степени расщепления. Значения, полученные в одном только буфере, вычитают как фоновые значения /что охватывает не только фоновое расщепление, но также продукты деградации и другие примеси, которые присутствуют в радиолигандном препарате/.
11.3. Результаты
Полученные степени расщепления фактора В представлены далее:
1/25 дикий тип 1,49%
1/5 дикий тип 2,74%
Q1R2 6,19%
Q1Q2 7,41%
E1Q2 6,42%
Поэтому все устойчивые к фактору I мутанты продуцируют большие уровни расщепления фактора В, нежели эквивалентные количества дикого типа С3 /C3i/. При больших дозах или более длительном инкубировании можно достичь полного исключения альтернативной схемы.
Сокращения, которые были использованы в предыдущих примерах: CFD комплемент фиксирующий разбавитель /определенный у Harrison and Lachmann, 1986, Handbook of experimental Immunology 4 ed., Ed. Weir, Herzenberg, Blackwell and Herrenberg; Blackwell, Oxford /; CFD-G, CFD, содержащий 0,1% /вес/объем/ желатина; РВS, буферированный фосфатом физиологический раствор; NGPS, сыворотка нормальных морских свинок; ПААГ-DСH электрофорез в DСH - полиакриламидном геле; SPDP N-сукцинимидил-3[2-пиридилдитио]пропионат.
Литература
1. Bergmann, M. & Fruton, J.S. (1941) Adv. Enzymol., 1:63-98.
2. de Bruijn, M.H. & Fey, G.H. (1985), Proc. Natl. Acad. Sci. U.S.A. 82: 708-712.
3. Crawford-MH еt al. (1988) Circulation. 78:1449-58.
4. Daha, M.R. & van Es, L.A. (1982) Immunol. 43:33-38.
5. Farries, T. C. , Lachmann, P.J. & Harrison, R.A. (1988) Biochem. J. 252:47-54.
6. Rarries, T. C. , Lachmann, P.J. & Harrison, R.A. (1988) Biochem. J. 253:667-75.
7. Forty, J. , Hasan, R., Cary, N., White, D.J. & Wallwork, J. (1992) Transplant. Proc. 24:488-9.
8. Fritzinger, D.C. et al. (1992) J. Immunol. 149:3554-3562.
9. Harrison.R.A. & Lachmann, P.J. (1980) Mol. Immunol. 17:9-20.
10. Kalli, K.R., Hsu, P. & Fearon, D.T. (1994) Springer Semin. Immununopathol. 15:417-431.
11. Kinoshita, T., Takata, Y., Kozono, H., Takeda, J., Hong, K.S. & Inoue, K. (1988) J. Immunol. 141:3895-901.
12. McNearney, T.A., Odell, C., Holers, V.M., Spear, P.G., Atkinson, J. P. (1987) J. Exp. Med. 166:1525-35.
13. Nicol, P.A.E. & Lachmann, P.J. (1973) Immunol. 24:259-275.
14. Pangburn, M.K. & Muller-Eberhard, H.J. (1984) Springer Semin. Immunopathol. 7:163-92.
15. Rother, K. & Till, G. O. (eds) (1988) "The complement System" (Springer-Verlag Berlin Heidelberg, Germany).
16. Van den Berg, C.W., Aerts, P.C. & Van Dijk, H. (1991) J. Immunol. Methods 136:287-294.
17. Vogel, C.W., Smith, C.A. & Muller-Eberhard, H.J. (1984) J. Immunol. 133:3235-41.
18. Weisman, H.F. еt al. (1990) Science 249:146-51.
19. Wu, R. (ed.) (1993) Methods Enzymol. 217: ch.s 12-14 (Academic Press, San Diego, U.S.A.).
20. Botto, M., Fong, K.Y., So, A.K., Koch, C. & Walport, M.J. (1990) J. Exp. Med. 172:1011-7.
21. Sambrook, J. , Fritsch, E.F. & Maniatis, T. (1989) "Molecular Cloning. A Laboratory Manual" second edition (Cold Spring Harbor Laboratory Press).
22. Fishelson, Z.(1991) Mol. Immunol. 28:545-52.
23. Taniguchi-Sidle, A. & Isenman, D.E. (1993) Mol. Immunol. 30:54.
24. Lambris, J. D., Avila, D., Becherer, J.D. & Muller, Eberhard, H.J. (1988) J. Biol. Chem. 263:12147-50.
25. Taniguchi-Sidle, A. and Isenman, D.E. (1992) J. Biol. Chem. 267: 635-643.
26. Hofer, В. and Kuhlein, B. (1993) Methods Enzymol. 217:173-189.
27. Morinaga, Y. , Franceschini, Т., Inouye, S. and Inouye, M. (1984) Biotechnology 2:636-639.
28. Harrison, R.A. and Lachmann, P.J. (1986) "Handbook of Experimental Immunology" (eds Weir, Herzenberg, Blackwell and Herzenberg; Blackwell, Oxford) 4th ed.
29. Kotwal, G.J. and Moss, В., Nature (1988) 335 (6186):176-8.8
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОТЕИН, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ К ОБРАЗОВАНИЮ С3-КОНВЕРТАЗЫ, ДНК, ВЕКТОР, КОНЪЮГАТ И ИХ ИСПОЛЬЗОВАНИЕ | 1997 |
|
RU2238322C2 |
| АНТИТЕЛА ПРОТИВ C3b И СПОСОБЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СВЯЗАННЫХ С КОМПЛЕМЕНТОМ НАРУШЕНИЙ | 2008 |
|
RU2473563C2 |
| ВАРИАНТЫ CRIG С СОЗРЕВШЕЙ АФФИННОСТЬЮ | 2009 |
|
RU2553517C2 |
| ГЕННАЯ ТЕРАПИЯ | 2016 |
|
RU2740038C2 |
| ЛЕЧЕНИЕ КОМПЛЕМЕНТ-ОПОСРЕДУЕМЫХ РАССТРОЙСТВ | 2017 |
|
RU2768982C2 |
| СПОСОБ ТРАНСПЛАНТАЦИИ ЖИВОТНОЙ КЛЕТКИ, ТКАНИ ИЛИ ОРГАНА РЕЦИПИЕНТУ И СРЕДСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1990 |
|
RU2195956C2 |
| СПОСОБ И НАБОР ДЛЯ ИММУНОФЕРМЕНТНОГО ОПРЕДЕЛЕНИЯ АКТИВНОСТИ C1 ИНГИБИТОРА, ПРОЯВЛЯЕМОЙ В ОБОИХ ПУТЯХ АКТИВАЦИИ КОМПЛЕМЕНТА | 2011 |
|
RU2464568C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К КОНФОРМАЦИОННОМУ ЭПИТОПУ С3 КОМПОНЕНТА КОМПЛЕМЕНТА ЧЕЛОВЕКА, ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК (ВАРИАНТЫ), ЭКСПРЕССИОННЫЙ ВЕКТОР, СОДЕРЖАЩИЙ ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК (ВАРИАНТЫ), И ШТАММ КЛЕТОК ЯИЧНИКОВ КИТАЙСКОГО ХОМЯЧКА CHO-humC34-ПРОДУЦЕНТ ДАННОГО ГУМАНИЗИРОВАННОГО АНТИТЕЛА | 2016 |
|
RU2630647C1 |
| АНАЛОГИ ФАКТОРА КОМПЛЕМЕНТА В И ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2639521C2 |
| СПОСОБ И НАБОР ДЛЯ ИММУНОФЕРМЕНТНОГО ОПРЕДЕЛЕНИЯ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ ФАКТОРА Н КОМПЛЕМЕНТА ЧЕЛОВЕКА | 2014 |
|
RU2565454C1 |
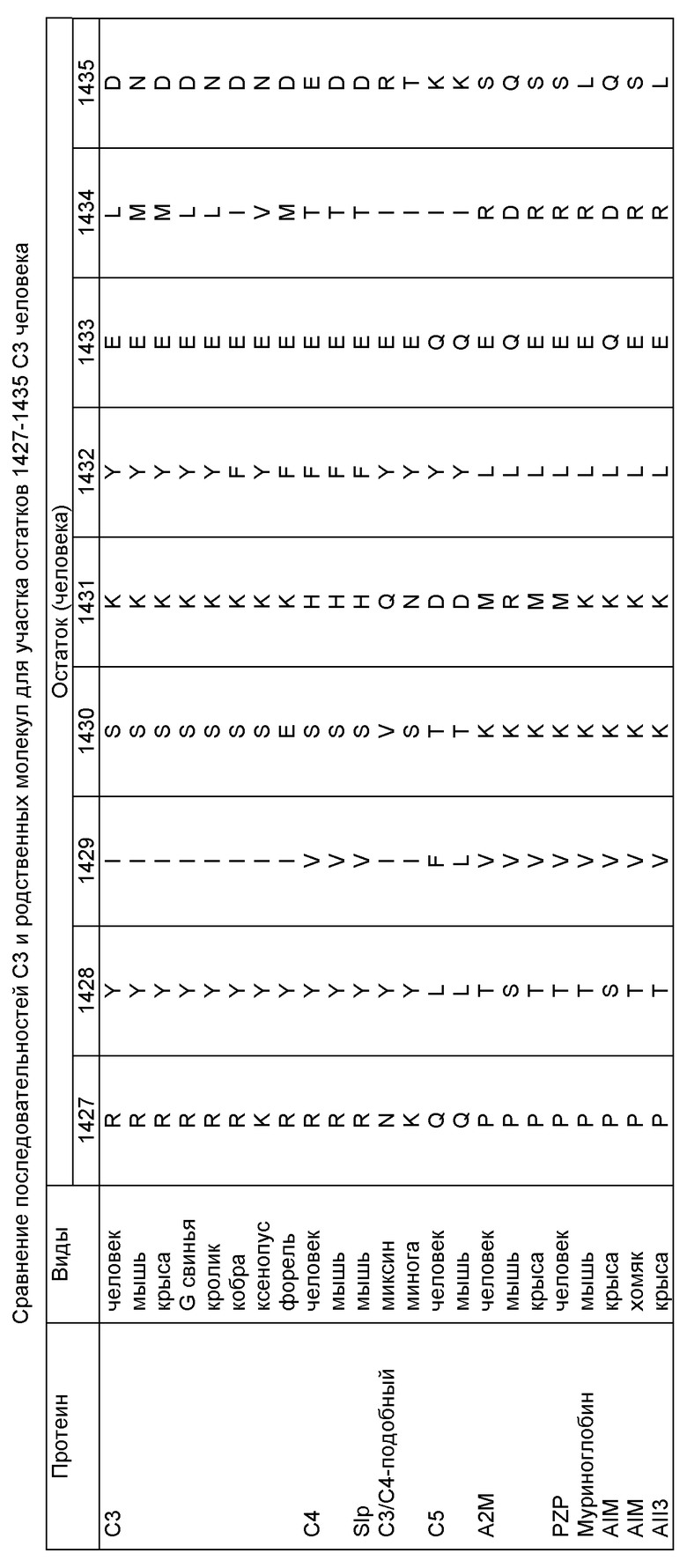











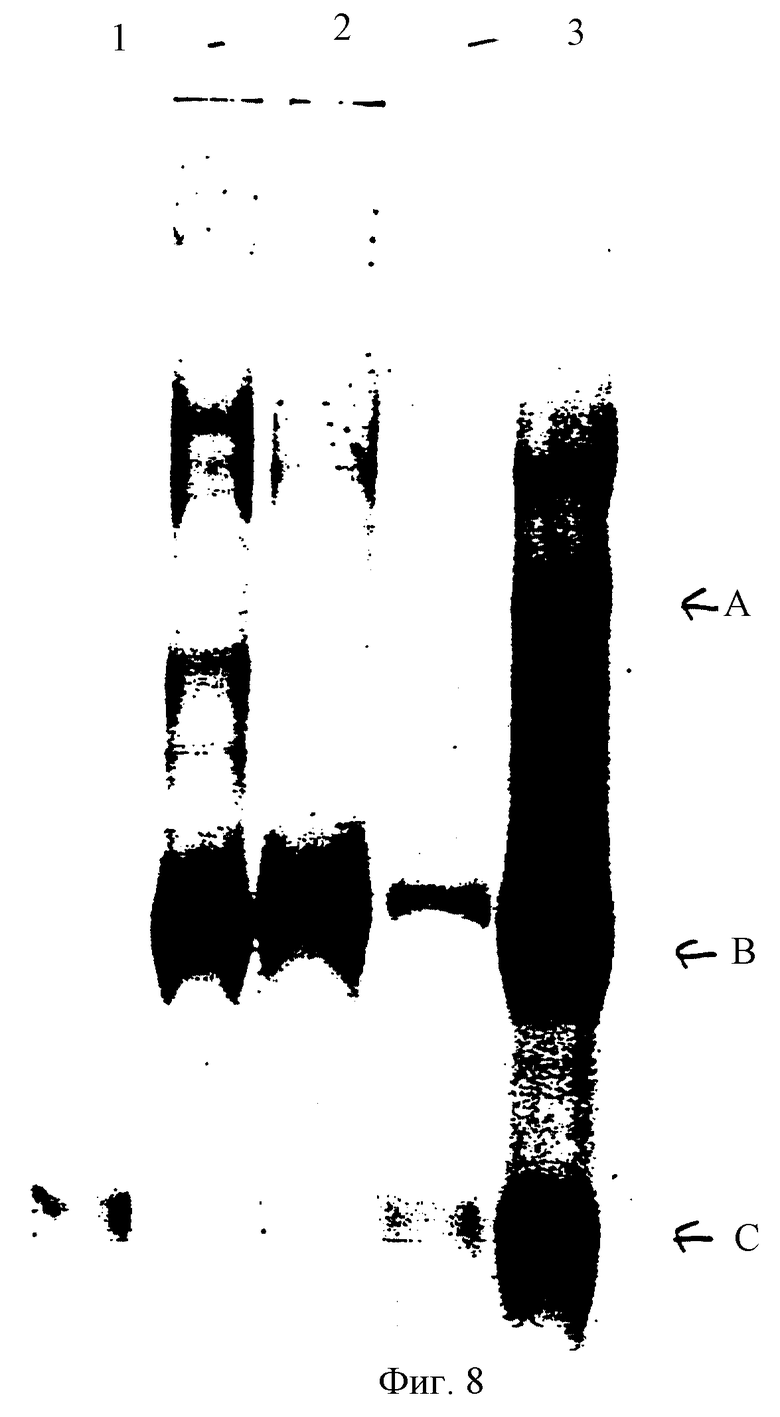

Изобретение относится к нативным белкам схемы комплемента, модифицированным таким образом, что белок способен образовывать стабильную С3 конвертазу. Предпочтительный модифицированный белок представляет модифицированный С3 белок человека. В аминокислотной последовательности С3 белки заменены Arg 1303 и/или Arg 1320, или аминокислотными остатками 752-754 и/или 758-780, или 1427, 1431 и/или 1433. Предложены также ДНК последовательности, кодирующие такие белки. Описаны конъюгаты, содержащие такие белки, связанные с антителом. Фармацевтическая композиция включает конъюгат и фармацевтически приемлемый носитель. При введении млекопитающему модифицированного белка или фармацевтической композиции происходит уменьшение количества белка системы комплемента. Изобретение позволяет повысить степень С3 конверсии. 5 с. и 12 з.п.ф-лы, 9 ил., 1 табл.

