ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к области биотехники и, в частности, касается способа превращения запасных веществ трансгенных семян двудольных растений на основе принципа источник - сток в композиции, содержащей один или несколько представляющих интерес генных продуктов. В изобретении также раскрываются композиции на основе прорастающих семян, включающие как минимум один генный продукт в семядолях прорастающих семян или в среде, окружающей прорастающие семена или проростки.
УРОВЕНЬ ТЕХНИКИ
В настоящее время способы получения терапевтически активных белков млекопитающих путем выделения и очистки таких белков непосредственно из самих млекопитающих вследствие высокого риска заражения почти полностью были заменены технологией рекомбинантных ДНК, которая обеспечивает новые способы получения требуемых в промышленности белков млекопитающих в больших количествах. Методы рекомбинантной ДНК обеспечивают большой выбор хозяев и систем экспрессии. Если нужны белки млекопитающих, то предпочитают использовать эукариотические системы экспрессии, чтобы получить гликозилированные формы этих белков.
Наиболее эффективную систему экспрессии для получения гликозилированных белков в большом объеме и с низкими затратами обеспечивают грибы. Грибы с успехом применяются для получения их собственных белков в больших количествах, однако они менее эффективны при получении гетерологических белков, таких как белки млекопитающих терапевтического назначения.
В настоящее время грибы используют в основном для широкомасштабной продукции гомологических промышленных ферментов, тогда как для получения гликозилированных гетерологических белков исследования сосредоточились на поисках альтернативных систем экспрессии. Поскольку профили гликозилирования и процессы укладки в растительных системах экспрессии близки таковым у млекопитающих, то растительные системы экспрессии являются наиболее экономически эффективными из всех имеющихся. Основной интерес был направлен на продукцию фармацевтических продуктов и важных для промышленности ферментов. Можно указать на получение интерферона, энкефалинов, фактора роста эпидермиса и сывороточного альбумина человека в растениях табака и/или картофеля. Уровни экспрессии в трансгенных растениях были достаточно низкими.
Обычно чужеродные гены в растениях экспрессируют под контролем сильных тканеспецифичных растительных промоторов в развивающихся органах растений. Типичными примерами являются промотор запасных белков семян или клубнеспецифичный промотор пататина в клубнях картофеля. В качестве альтернативы используют культуры клеток или органов и культуры водорослей, а также еще более сложные системы, включая системы на основе вирусов растений, в которых искомый ген встроен в геном вируса и экспрессируется одновременно с вирусными белками. Самые передовые системы основываются на индуцибельной экспрессии и секреции рекомбинантного белка в среду.
В патентах US 5,693,506 и US 5,994,628 раскрыта продукционная система для получения чужеродных белков в прорастающих семенах однодольных растений. Рекомбинантные белки экспрессируют под контролем сильного промотора амилазы в клеточной культуре или в прорастающих семенах, а белок секретируется в питательную среду или экстрагируется из солода. В семенах однодольных растений запасные вещества в основном состоят из крахмала эндосперма. Запасы белка у однодольных растений небольшие по сравнению с семенами двудольных растений. Кроме того, если даже уровень экспрессии амилазы в отдельных клетках высок, в семенах она экспрессируется лишь в небольшом числе клеток. Эти клетки ограничиваются щитком зародыша и алейроновым слоем эндосперма. Существует большая потребность в более экономичных системах, в которых бы использовались белки и масла из запасных веществ семян двудольных растений. В патентах US 5,543,576 и US 5,714,474 раскрыт способ, в котором трансгенные семена добавляют в корма без предварительного проращивания и в размолотом виде в качестве дополнительного источника ферментов.
В патенте US 5,670,349 раскрыто применение индуцируемых при повреждении промоторов HMGR/HMG2 для экспрессии рекомбинантных белков в листьях табака, собранных свежими в поле. Нанесение повреждений на свежий или хранившийся растительный материал путем иссечения или смятия вызывает быстрое усиление экспрессии. Однако содержание белка в листьях табака невелико по сравнению с тем, что можно получить из семян двудольных растений. Кроме того, срок годности у свежих листьев несравним с сухими семенами в отношении времени, пространства и/или затрат на хранение. Использование свежего растительного материала типа листьев, собранных в поле, также является важным источником микробного заражения, что представляет серьезную проблему в технологии ферментации.
В патентных заявках WO 94/11519 и WO 97/32986 раскрыты способы и установки для получения продуктов деградации и превращения у растений с помощью процесса солодования. В этих способах предлагается использовать ферменты глиоксисом, которые катализируют превращение жирных кислот в ацетил-СоА. В норме ацетил-СоА идет на образование органических кислот, которые экспортируются из глиоксисом и поступают в другие метаболические пути, такие как дыхание и синтез сахара, что предлагается заменить генами, кодирующими ферменты пути, ведущего к полигидроксиалканоатам, которые полезны для получения биоразрушаемых термопластиков.
Следует заметить, что даже если процесс солодования известен и применяется главным образом для переработки однодольных растений, этот процесс ограничивается переработкой крахмала. Раскрытые в патентах WO 94/11519 и WO 97/32986 способы, даже если они предполагают использование запасных веществ двудольных растений, ограничиваются применением ферментов и путей, ведущих к образованию сахара и получению энергии, и способами переключения этих продуктов из дыхательных путей на ресинтез.
Даже если процесс солодования известен, как и способы переключения дыхательных путей у растений на продукцию полигидроксиалканоатов, получение белковых продуктов типа структурных белков и ферментов из запасных веществ двудольных растений не было разработано.
Следует заметить, что существует большой спрос на белковые продукты, поэтому главной целью настоящего изобретения является решение проблемы превращения запасных веществ у растений в различные белковые продукты, из которых затем можно получать другие продукты.
Главная задача настоящего изобретения - обеспечить новый, более практичный, экономически эффективный, экологически благоприятный способ и продукционную систему для получения генных продуктов, особенно белковых генных продуктов, в семядолях трансгенных семян двудольных растений. Другим преимуществом настоящего изобретения является то, что оно предусматривает продукционную систему закрытого типа, в которой генный продукт можно вырабатывать в помещении, а не в поле. Это дает возможность осуществлять настоящий способ в условиях, практически свободных от заражения.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Цели настоящего изобретения достигаются путем переключения регуляторных последовательностей белков, временно накапливающихся при прорастании, на продукцию требуемых генных продуктов.
Характеристики настоящего изобретения изложены в Формуле изобретения.
ПЕРЕЧЕНЬ ФИГУР
На фигуре 1А показано проросшее трансгенное семя, экспрессирующее ген GUS. Ген GUS находится под контролем промотора гена теплового шока сои. Справа показан нетрансформированный контрольный проросток.
На фигуре 1В показано проросшее трансгенное семя, экспрессирующее ген GUS. Ген GUS находится под контролем промотора эндопептидазы.
На фигуре 1С показано проросшее трансгенное семя, экспрессирующее ген GUS. Ген GUS находится под контролем салицилат-индуцируемого промотора.
На фигуре 1D показано проросшее трансгенное семя, экспрессирующее ген GUS. Ген GUS находится под контролем промотора 35S.
На фигуре 2 показано, что бесцветный субстрат β-глюкуронид (100 мкг/мл) приобретает синюю окраску в результате ферментативного превращения его в глюкопиранозидуроновую кислоту и агликон под действием фермента GUS, вырабатываемого в проросших трансгенных семенах.
На фигуре 3 показаны различные стадии прорастания семян Brassica campestris через указанное число дней после начала прорастания.
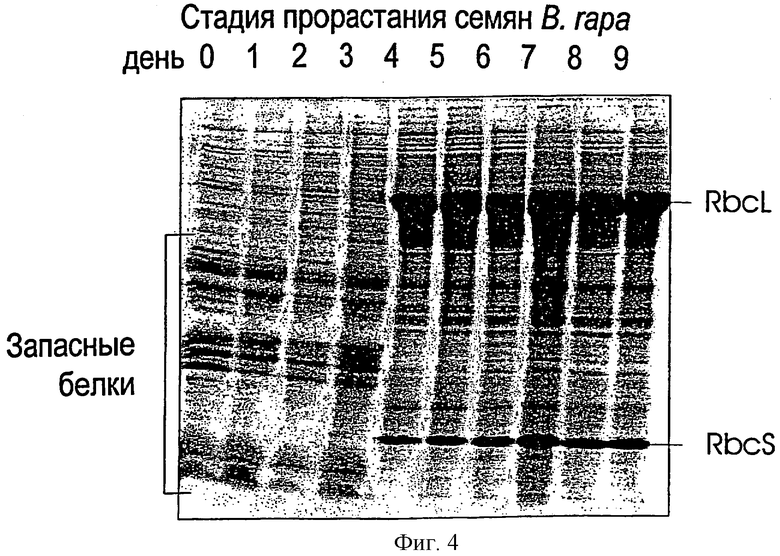
На фигуре 4 представлен SDS-PAGE-гель прорастающих семян. Образцы отбирали ежедневно, начиная с сухих семян. На окрашенном Кумасси геле отмечены запасные белки и субъединицы фермента rubisco. Как видно из фигуры, продуцируемое за день количество белка rubisco значительно повышается через 3 дня после прорастания.
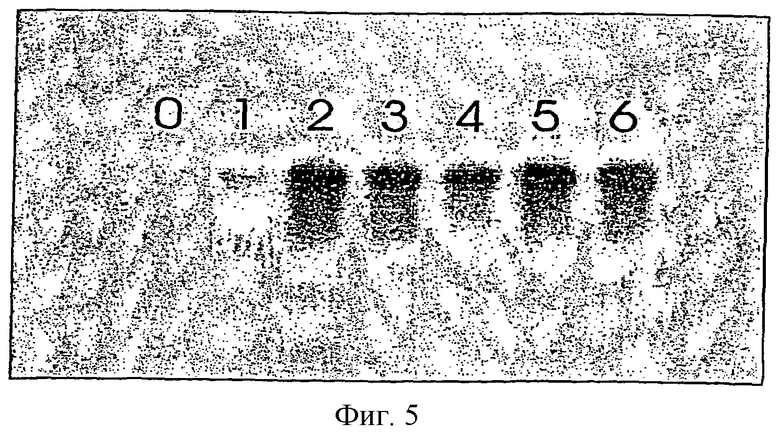
На фигуре 5 представлен результат Northem-гибридизации с РНК-зондами прорастающих семян. Образцы отбирали ежедневно, начиная с сухих семян.
Определения
В настоящем изобретении большинство терминов имеют те же значения, которые они имеют в области технологии рекомбинантных ДНК, молекулярной биологии и ботаники, особенно в тех областях, что связаны с получением трансгенных растений и генных продуктов. Некоторые термины, однако, применяются в несколько другом виде и более подробно разъясняются ниже.
Термин "семянные растения" означает Spermatophyta - наиболее высокоразвитые растения, отличающиеся наличием сложных структур и полностью развитых органов - корней, стеблей, листьев, цветков и сосудистой системы. Семянные растения подразделяются на 2 класса - покрытосемянных и голосемянных. Покрытосемянные подразделяются на подклассы однодольных и двудольных. В зародыше зрелых семян однодольных имеется только одна семядоля, которая редуцирована до всасывающего органа - щитка. У двудольных в зародыше имеется две семядоли, служащие запасающими органами. Усвоенные вещества, идущие на отложение запасов в семядолях двудольных растений, поступают из материнского растения через сосудистую систему в оболочку семян, а затем в семядоли. Оболочка семян является материнской тканью, которая не имеет симпластических контактов с зародышем. Усвоенные вещества сначала поступают в апоплазматическое пространство, а затем в зародыш, где они распространяются симпластически и утилизируются при синтезе запасных веществ.
Термин "семена двудольных растений" означает семена, получаемые из двудольных растений, содержащие запасы питательных веществ из белков, жиров и углеводов, находящихся в семядолях, в отличие от семян однодольных растений, у которых запасы питательных веществ состоят главным образом из углеводов и содержат лишь небольшое количество других соединений. В настоящем изобретении термин "семена двудольных растений" означает целые семена или их фрагменты или части семян, полученные при измельчении семян, предпочтительно в сухом виде. Следует заметить, что настоящее изобретение касается двудольных растений, тогда как однодольные растения не пригодны для достижения целей настоящего изобретения.
К наиболее предпочтительным родам растений, которые могут применяться для получения генных продуктов, в соответствии со способом настоящего изобретения, относятся двудольные растения с высоким содержанием белка или жира в семенах, но не ограничиваются ими. Типичными примерами являются роды Brassicaceae, Fabaceae и Polygonaceae. Из рода Brassicaceae, например, к ним относятся следующие виды: Brassica napus, Brassica campestris, Camelina sativa, Sinapis alba, из рода Fabaceae - Lupinus angustifolius, Phaseolus vulgaris, Glysine max. Vicia faba, Lens culinaris, Pisum sativum, Vigna mungo и Medicago sativa. Из рода Polygonaceae - виды Polygonum. Подсолнечник Helianthus annuus потенциально тоже полезен из-за высокого содержания жира в семенах.
Выражение по принципу источник - сток означает такую продукционную систему или блок, которая разделяется на 2 отдельные стадии: стадию накопления, на которой происходит запасание белков, углеводов и жиров в семенах, и стадию мобилизации продукции, на которой накопленные запасы, в норме мобилизующиеся для запуска роста растения, вместо этого мобилизуются или переключаются на продукцию одного или нескольких продуктов чужеродных генов при включении системы экспрессии. Стадия накопления включает выращивание трансгенных растений практически совсем без экспрессии генного продукта в поле, тогда как продукция генных продуктов ограничивается проращиванием в закрытом помещении. Это достигается специфичностью систем экспрессии, как описано более подробно в спецификации.
Термин "окружающая среда" означает водный буферный раствор в жидком, полужидком или твердом виде, содержащий вещества, способные запускать, усиливать, задерживать или продлевать самую благоприятную стадию проращивания. "Окружающая среда" может содержать факторы, индуцирующие прорастание, факторы, усиливающие определенные процессы, и/или факторы, ослабляющие некоторые другие процессы, наряду с питательными веществами. "Окружающая среда" может представлять собой влажный отсек или комнату.
Термин "субстрат" означает твердую, полужидкую или жидкую композицию или среду, содержащую, по меньшей мере, одно соединение или вещество, которое можно превратить в другое соединение или вещество с более желательными свойствами с помощью композиций настоящего изобретения, включающих один или несколько искомых генных продуктов в качестве катализаторов.
Выражение "происходящая из семян композиция" означает неочищенную смесь, полученную способом настоящего изобретения из прорастающих семян двудольных растений, которая включает один или несколько синтезированных de novo генных продуктов, экспрессируемых в семядолях прорастающих семян двудольных растений, включая ростки, или секретируемых в среду, окружающую прорастающие семена или ростки. "Происходящая из семян композиция" может использоваться как таковая путем приведения ее в контакт с субстратом. Естественно, происходящая из семян композиция может быть высушена и/или обработана соответствующими методами, а также выделена и/или очищена перед употреблением. Альтернативно в качестве субстрата может применяться реагент, содержащий одно или несколько соединений, способных модифицировать один или несколько генных продуктов в этой композиции. Например, можно добавлять замещающие реагенты или модифицировать профили укладки генных продуктов либо удалять или добавлять фрагменты или части для того, чтобы получить более стабильный продукт. "Происходящие из семян композиции" можно формулировать, то есть добавлять соответствующие добавки типа средств, способствующих грануляции, наполнителей, смазывающих веществ и т.д. Происходящие из прорастающих семян композиции настоящего изобретения также можно добавлять в корма животным для улучшения их переваривания.
Выражение "практически стерильная продукционная система типа источник - сток" означает, что семена двудольных растений можно стерилизировать с поверхности такими способами, как облучение или химическая стерилизация. Проращивание может проводиться в или на предварительно стерилизованной среде, с помощью методов, применяемых при асептической работе и/или при работе в стерильных или сверхчистых условиях. Тот факт, что все производство может проводиться в стерильных условиях, означает, что устраняются главные причины гибели или непрорастания семян - грибковое или бактериальное заражение семян. Более того, и конечный продукт становится практически свободным от заражения и не поддается деградации со стороны примесей, обладающих протеолитической активностью. Таким образом, предложенная продукционная система особенно подходит для получения фармацевтических продуктов, требующих высоких стандартов гигиены.
Выражение "проращивание трансгенных семян двудольных растений" означает создание условий, способствующих прорастанию. Необязательно он включает хранение, сушку, стерилизацию, добавление воды, обеспечение семян индуцирующими факторами, усиливающими и/или ослабляющими факторами, а также дополнительное питание. К вспомогательным средствам относятся как физические, так и химические средства. Например, дополнительное питание включает источники N и/или С, витамины, минералы, микроэлементы и т.п. Факторы роста, факторы прорастания, индукторы, факторы, усиливающие одни и ослабляющие другие процессы во время синтеза de novo, к ним относятся, к примеру, такие гормоны растений, как ауксин и цитокинин. К физическим средствам относятся, к примеру, кратковременное или продолжительное освещение, стерилизация и/или нагрев.
Термин "экспрессионная система" означает конструкцию из ДНК, включающую одну или несколько последовательностей ДНК, кодирующих один или несколько искомых генных продуктов, которая функционально соединена с регулирующими экспрессию последовательностями - энхансерами, промоторами и/или терминаторами. Экспрессионная система предпочтительно индуцируется или может быть индуцирована при прорастании семян, но она практически не проявляется или может не проявляться, когда двудольные растения размножают и выращивают в полевых условиях. Предпочтительно регуляторные последовательности содержат промотор, который можно назвать промотором "camera obscura", то есть промотор, активизирующийся при прорастании, когда семена хранятся без освещения в темной комнате. Это не исключает применение освещения. Наоборот, подвергая продукционную систему типа "источник - сток" вспышкам света, можно усиливать выход искомого генного продукта посредством индукции и повышения активности специфических промоторов.
В экспрессионных системах используют такие регуляторные последовательности, которые особенно активны при временном синтезе de novo белков, накапливающихся в семядолях при прорастании.
Индуцибельные экспрессионные системы содержат индуцибельные регуляторные последовательности, отобранные из регуляторных последовательностей тех генов, которые кодируют белки, временно синтезируемые de novo при прорастании и накапливающиеся в семядолях и эндосперме растения.
Такие экспрессионные системы получают путем селекции соответствующих регуляторных последовательностей. Селекционный метод включает следующие стадии:
(a) выделение тотальной РНК из семядолей и/или листьев двудольного растения;
(b) получение кДНК на основе РНК путем амплификации с помощью одного праймера, специфичного к данному растению, и второго праймера, специфичного к тому белку, который синтезируется de novo при прорастании и накапливается в семядолях или эндосперме растения;
(c) сбор и рассев клонов, содержащих кДНК;
(d) сравнение результатов с клонами, полученными из выделенной из листьев РНК;
(е) выделение клонов, содержащих кДНК, проявляющую высокую активность при прорастании семян двудольного растения.
Термин "генный продукт" в настоящем изобретении означает белки и полипептиды, включая структурные белки и/или ферменты. Обычно "генным продуктом" является пептид, содержащий, по меньшей мере, 2 аминокислоты, соединенные пептидной связью. Пептиды, содержащие 2-10 аминокислот, называют олигопептидами, а пептиды, содержащие более 10 аминокислот, называют полипептидами. Полипептид может представлять собой цепь полиаминокислоты, например, цепь полилизина, состоящую только из молекул лизина, но может представлять собой и белок, белковый комплекс, либо часть или фрагмент белка.
Как сказано в предыдущем абзаце, термин "генный продукт" также может означать продукты на основе нуклеотидов типа мРНК. Этот термин также охватывает и другие желательные биохимические соединения или вещества, получаемые, к примеру, путем биотрансформации, когда один или несколько генных продуктов, находящихся в происходящих из семян композициях настоящего изобретения, действуют как катализаторы при контакте с субстратом, включающим соединение или вещество, которое может быть превращено в требуемый продукт или трансформировать генный продукт должным образом.
С другой стороны, "производные генных продуктов" получают путем биотрансформации, при которой один или несколько исходных "генных продуктов" настоящего изобретения подвергаются модификации при реакции с реагентом, входящим в состав субстрата. Генные продукты можно сделать более стабильными или более активными, изменяя их вторичную и/или третичную структуру, например, путем разворачивания или сворачивания аминокислотной цепи, удаления или добавления фрагментов, комплексирования, добавления структур или замещающих групп к этим генным продуктам. Такие "производные генных продуктов" покрываются термином "генные продукты" настоящего изобретения. Обычно генные продукты, полученные способом настоящего изобретения, являются гетерологичными чужеродными белками. Это значит, что "генные продукты" не вырабатываются растениями дикого типа. Другими словами, они не являются своими для этих растений. Естественно, собственные продукты семян тоже можно получать способом настоящего изобретения. При этом особенно удобно повысить продукцию определенных, фармацевтически полезных, гомологичных, собственных продуктов данного растения.
Выражение "выделить с или без дальнейшей обработки генного продукта, экспрессированного в прорастающих трансгенных семенах двудольных растений" означает, что семядоли и ростки можно получить в целом виде, выделяя их из окружающей жидкой среды с дальнейшей обработкой или без нее. В качестве альтернативны можно выделить вмещающую их среду. Сохранность "происходящих из семян композиций" можно улучшить посредством высушивания, экстрагирования, фильтрования, измельчения и т.д. Многие из этих способов применяются одновременно для прекращения прорастания. Нагревание, сушка, измельчение, отделение, экстрагирование, выжимание и фильтрование обычно применяются для прекращения прорастания до того, как скорость синтеза de novo и/или концентрация требуемых генных продуктов начинает снижаться. Прекращение прорастания важно потому, что если оно продолжается слишком долго, то искомые, синтезированные de novo, белки начинают использоваться для выработки других веществ, необходимых растущему растению.
Запасные вещества, накопленные в прорастающих семенах двудольных растений, в число которых входят свободные аминокислоты и другие биохимические субстраты, составляют модуль возобновляемых источников сырья, тогда как экспрессия требуемого трансгена путем регуляции транскрипции, функционирующая в семядолях при прорастании и последующем росте, образует модуль продукции (стока). Таким образом, настоящее изобретение прежде всего касается продукционной системы, состоящей из двух модулей - источника и стока, то есть двух раздельных стадий: стадии накопления сырья и стадии продукции, на которой источник сырья используется для синтеза de novo генных продуктов и их производных, или продуктов, модифицируемых генным продуктом из источника сырья, полученного на первой стадии экспрессионной системы.
Цель настоящего изобретения заключается в разработке нового способа получения генных продуктов в семенах двудольных растений на основе продукционной системы типа источник - сток. Продукционная система включает две отдельные стадии. На первой стадии используются универсальные возобновляемые источники сырья - белки и жиры, которые накапливаются в виде запасных веществ при выращивании двудольных растений и необязательно могут быть собраны в виде семян. Другая стадия включает мобилизацию запасных веществ в прорастающих трансгенных растениях для продукции одного или нескольких генных продуктов путем синтеза de novo требуемых генных продуктов с добавлением или без добавления дополнительных факторов питания и обеспечения физических и/или химических средств, а также генной инженерии для индукции прорастания и усиления и/или ослабления процессов синтеза de novo, включая транскрипцию, экспрессию и/или секрецию.
В настоящем изобретении на первой стадии используются универсальные запасные вещества - белки, жиры и/или углеводы, которые имеются в двудольных растениях, но отсутствуют у однодольных, у которых запасные вещества состоят в основном из двух родственных форм крахмала - амилозы и амилопектина. У однодольных растений крахмал мобилизуется под действием амилаз во время прорастания и последующего роста молодого растения. При этом образуется сахароза, которая используется как источник энергии и соединений углерода, но исходных материалов для синтеза de novo, особенно белков, у них намного меньше, чем у двудольных растений, вследствие чего прорастающие семена двудольных образуют несравненно лучшую продукционную систему типа источник - сток.
В настоящем изобретении семена трансгенных двудольных растений собирают с поля, транспортируют и хранят как обычно. Жизнеспособность семян при хранении можно поддерживать в течение длительного времени, обычно несколько лет. Температура и влажность семян являются главными факторами, определяющими жизнеспособность семян при хранении. Обычно семена хранят при температуре 0-5°С. В норме влажность семян при хранении составляет 4-10%. Если влажность семян составляет меньше 20%, они будут дышать и выделять тепло. При плохой вентиляции семена могут погибнуть от выделяемого тепла. Если влажность семян больше 20%, то это может вызвать их порчу из-за роста микроорганизмов.
Высыхание при созревании является нормальным процессом в развитии семян, при котором семена переходят в состояние метаболического покоя. Однако семена многих видов способны к прорастанию без высыхания. Семена помидоров прорастают, если их вынуть из созревающего плода и поместить в воду. У некоторых других видов, включая Brassica campestris, запасы крахмала быстро уменьшаются, а концентрация некоторых сахаров и олигосахаридов повышается при высыхании семян. К тому же некоторые гидрофильные белки сильно экспрессируются при высыхании семян. Они обладают способностью притягивать воду и, по-видимому, играют важную роль в защите от вредных эффектов высыхания. Зрелые сухие семена сохраняют способность к прорастанию при хранении от нескольких дней до многих лет. Как неочищенный исходный материал без добавок семена двудольных растений могут стабильно храниться в течение нескольких лет.
У двудольных растений основным видом запасных липидов являются триацилглицерины, хранящиеся в субклеточных органеллах в виде жировых включений. Содержание жира в семенах может достигать 64% у клещевины и 48% у рапса. Запасные жиры синтезируются из сахарозы, поступающей в развивающиеся семена. В цитозоле сахароза подвергается превращению до гексозофосфосфата, который переносится в пластиды или превращается в глицерол-3-фосфат (остов глицерина). В пластидах из гексозофосфосфата, малата и ацетата синтезируются жирные кислоты. В эндоплазматическом ретикулуме (ЭР) остов глицерина подвергается этерификации с образованием триацилглицеринов.
У двудольных растений основным видом аминокислот, переносимых по флоэме, являются глутамин и аспарагин. В оболочке семян перед переносом в семядоли часть глутамина и аспарагина превращается в другие аминокислоты. В семядолях аминокислоты используются для синтеза запасных белков, которые хранятся в белковых включениях. На ранних стадиях развития семян клетки семядолей содержат одну или две большие вакуоли, которые потом заполняются запасными белками и постепенно фрагментируют, образуя отдельные небольшие белковые включения. Обычно семена двудольных растений содержат два или больше различных запасных белков. Например, Brassica campestris в целом содержит 24% белка, в состав которого входят 2S-альбумин, напин, 11S-глобулин и круциферин. Напин составляет 20%, а круциферин - 60% от общего содержания белка в семенах. В настоящем изобретении целесообразно использовать гены, кодирующие эти белки.
В общем случае запасные белки кодируются мультигенными семействами. Общей чертой белков просвета эндоплазматического ретикулума эукариот является наличие консервативного карбоксиконцевого тетрапептида KDEL, который служит сигналом удержания в ЭР и всегда находится последним на С-конце. Нет никаких данных, что может понадобиться больше 4 концевых аминокислот (Pelham H.R.B., TIBS 15: 483-486, 1990), хотя и были утверждения, что для эффективного удержания может иметь значение состав кислотных остатков среди последних 20 аминокислот (Wandelt C.I. et al. Plant J., 2: 181-192, 1992). Предполагается, что механизм удержания скорее всего опосредован рецепторами и включает связывание сигнала KDEL с мембранным рецептором и быстрый возврат комплекса сигнал-рецептор в ЭР, где он высвобождается в виде везикулы (Pelham H.R.B., TIBS 15: 483-486, 1990).
Генетически модифицированные белки не всегда находятся в своем естественном клеточном компартменте. Например, при экспрессии в трансгенном табаке обогащенный Met фазеолин обнаруживается только в ЭР (эндоплазматическом ретикулуме), цистернах Гольджи и везикулах Гольджи. В норме фазеолин накапливается в матриксе запасающих белок вакуолей растущих семян табака. Эти данные означают, что обогащенный Met фазеолин подвергается деградации в везикулах Гольджи или сразу после попадания в запасающие белок вакуоли (Hoffinan L.M. et al. Plant Mol. Biol, 11: 717-730, 1988). Сообщалось о высоком уровне аккумуляции запасного белка семян Vicia faba вицилина, содержащего дополнительную последовательность KDEL на С-конце, в табаке и люцерне (Wandelt C.I. et al.. Plant J., 2: 181-192, 1992). При применении конструкций с модифицированными генами, слитыми с промотором 35S вируса мозаичности цветной капусты, белок накапливается в ЭР листьев трансгенных растений. Соответственно представляется вероятным, что при образовании генетически модифицированных запасных белков удержание в ЭР предотвращает их деградацию и что ДНК-конструкции с промотором 35S вируса мозаичности цветной капусты можно применить в настоящем изобретении.
Круциферин - гексамер из 6 субъединиц, каждая из которых состоит из одной цепи, кодируемой одним геном (Rodin J. & Rask L., Physiol. Plant. 79: 421-426, 1990). Например, у Brassica napus гены круциферина гомологичны примерно на 60%. Напин - второй по значению белок семян у Brassica napus. Как и субъединицы круциферина, являющиеся продуктами отдельных генов, напин состоит из двух разных полипептидных цепей, соединенных дисульфидными мостиками. Круциферин и напин синтезируются на мембране ЭР в виде предшественников, содержащих адресный сигнал ЭР (Ericson M.L. et al., J. Biol. Chem., 261: 14576-14581, 1986).
Как указано выше, круциферин и напин содержат адресный сигнал ЭР на N-концах. Белки поступают в ЭР в развернутом виде и сигнальная последовательность отщепляется вскоре после попадания в ЭР. В последние годы стало ясно, что не все белки, попадающие в ЭР, подвергаются дальнейшему процессингу и поступают в вакуоли. В просвете ЭР постоянно имеется большое число белков, способствующих созреванию секреторных белков на начальных стадиях. Многие из этих белков также описаны у млекопитающих (Pelham H.R.B., TIBS 15: 483-486, 1990), значит, гликозилирование генных продуктов, особенно белков протекает сходным образом у млекопитающих и у растений. Например, было показано, что у растений ауксин-связывающий белок остается в ЭР (Inohara N. et al., Proc. Natl. Acad. Sci. USA 86: 3564-3568, 1989).
Обычно при прорастании первыми появляются протеиназы из группы SH-зависимых протеиназ, которые действуют на нерастворимые нативные запасные белки. У Vigna mungo синтез SH-зависимой протеиназы в семядолях начинается сразу после вымачивания и усиливается до 4-го дня, после чего снижается (Okamoto Т. & Minamikawa Т., Plant Physiol., 152 (6). 675-682, 1998). Короткие пептиды, образующиеся в результате эндопептидазной активности, отщепляются от нативных белков, что в свою очередь повышает их подверженность действию других протеиназ.
Протеиназы второй группы неактивны в отношении нативных запасных белков, но гидролизируют короткие пептиды до олигопептидов. В то же самое время активируются карбоксипептидазы и из пептидов отщепляются аминокислоты. Этот протеолиз происходит в белковых включениях, из которых олигопептиды и аминокислоты выходят в цитозоль, где олигопептиды подвергаются дальнейшей деградации до аминокислот под действием амино-, ди- или трипептидаз. Аминокислоты, высвобождаемые из запасающих органов, подвергаются дальнейшему метаболизму с образованием главных транспортных форм аминокислот - аспарагина и глутамина. Запасные белки являются источником аминокислот для синтезируемых de novo белков во время прорастания.
Вторая стадия способа настоящего изобретения заключается в прорастании семян, при котором сырьевые материалы, накопленные при выращивании в запасных веществах семян двудольных растений, мобилизуются для выработки требуемых генных продуктов. Продолжительность прорастания относительно коротка, а затраты низкие. Объем продукции практически неограничен, так как сырьевые материалы получают из возобновляемых источников, а необходимая энергия поступает от солнца.
В настоящем изобретении прорастание означает этап развития семян двудольных растений, начиная с поглощения воды, усиление дыхания и синтез макромолекул, мобилизация запасных веществ и субклеточные структурные изменения. Прорастание в физиологическом смысле - это короткий этап, который завершается началом удлинения зародышевой оси. В настоящем изобретении прорастание также включает фазу роста после начала удлинения зародышевой оси. Семена обычно поглощают воду в количестве, в два-три раза превышающем их сухой вес, но на более поздней стадии развития ростков требуется большее количество воды. Прорастание включает множество явлений, таких как усиление дыхания, синтез макромолекул и субклеточные структурные изменения. Прорастание в общем завершается удлинением корешка, которое происходит путем увеличения размера клеток без деления. Выделяют 2 типа ростков, отличающихся судьбой семядолей после прорастания. В подземном типе прорастания семядоли не выходят из-под земли при появлении ростков. Гипокотиль остается коротким, а эпикотиль удлиняется и выпускает первые настоящие листья из-под земли. В надземном типе прорастания семядоли выносятся на поверхность растущим гипокотилем. Семядоли обычно становятся фотосинтетически активными до появления настоящих листьев.
При прорастании семян в природе или в поле запасные вещества семядолей обычно используются для запуска физиологической активности после стадии покоя в виде сухих семян. Жизненные функции быстро усиливаются, включая экспрессию генов. Из запасных веществ семядолей высвобождаются значительные количества аминокислот и других биологических соединений при прорастании, которые затем идут на рост.
Стерилизованные с поверхности семена также способны к прорастанию при инкубации в воде или водном растворе, содержащем соединения, способствующие выработке требуемого генного продукта. Во время прорастания активируется экспрессия генов для получения нужного генного продукта. Для проращивания семена сначала вымачивают до содержания влаги в 40-50%, выдерживая их в чанах с водой, в которых поддерживают температуру согласно типу семян и организуют аэрацию сжатым воздухом. После вымачивания семена для проращивания обычно переносят в камеры с влажностью 100%. В типичных случаях поддерживается температура 10-30°С при продолжительности инкубации от 2 до 10 дней, предпочтительно от 3 до 6 дней, и наиболее предпочтительно от 4 до 5 дней.
В настоящем изобретении прорастающие семена двудольных растений содержат трансгенные семядоли, то есть семядоли двудольных растений, стабильно трансформированные последовательностью ДНК, кодирующей требуемый генный продукт. Семядоли являются составной частью зародыша и служат запасающим органом. Семядоли имеют относительно простую физиологическую структуру и набор клеточных типов. После прорастания начинается мобилизация большей части запасных веществ семядолей. Три основных вида запасных веществ в семядолях - белки, жиры и углеводы - подвергаются ферментативным превращениям в подвижные формы во время роста ростков. Запасные белки расщепляются постепенно целым рядом протеиназ, экспрессируемых в клетках семядолей. Протеиназы синтезируются de novo структурными генами, которые регулируются промоторными участками, функционирующими в семядолях.
При прорастании и последующем росте ростков мобилизация запасных липидов начинается с липолиза в жировых отложениях, когда из триацилглицерина образуется глицерин и свободные жирные кислоты. В запасающих клетках семян глицерин и свободные жирные кислоты превращаются в сахарозу посредством сложных реакций, в которых участвуют десятки ферментов, а также такие органеллы, как глиоксисомы и митохондрии. Сахароза поступает в вакуоль или в зародыш.
На стадии мобилизации/продукции накопленные запасы белков, углеводов и жиров служат источником исходных материалов и подвергаются превращению в коммерчески привлекательные продукты вместо нормальных белков, вырабатываемых при формировании растения. Мобилизация начинается при добавлении к семенам жидкой, полужидкой или твердой среды, содержащей воду, в которую необязательно добавляют физические или химические средства, включая питание, индуцирующие прорастание факторы, а также факторы, способные усиливать и/или ослаблять определенные процессы при продукции (транскрипцию, экспрессию и/или секрецию). При этом мобилизация запускает прорастание и продукция продолжается до появления зеленых листьев.
Известно, что заражение представляет серьезную проблему в технологии ферментации. При промышленной продукции значительная часть (20-25%) выпускаемых партий оказывается зараженной. При проведении таких процессов, в которых используется свежий растительный материал, как получение крахмала из картофеля или мокрый помол зерна, тоже сталкиваются с проблемами роста микроорганизмов. Такой свежий растительный материал, как собранные в поле листья, является главным источником заражения, что может вызвать серьезные потери конечной продукции.
В настоящем изобретении описывается система, в которой исходным материалом служат сухие семена. В поле происходит лишь ограниченная экспрессия чужеродных генов, вместо этого экспрессия проводится на предприятии, в контролируемых условиях, экологически безопасным образом. Сухие семена двудольных растений легче поддаются стерилизации с поверхности, чем однодольные растения, потому что они обычно имеют плотную, гладкую оболочку в отличие от зерен однодольных растений. Стерилизация может проводиться путем обработки такими химикатами, как гипохлорит, перекись водорода, или с помощью других дезинфицирующих соединений. Поскольку семена двудольных растений имеют гладкую поверхность в отличие от морщинистой текстуры семян однодольных, стерилизация семян двудольных значительно более эффективна. Вследствие этого продукция требуемых чужеродных или собственных генных продуктов может проводиться в стерильных условиях, что дает большое преимущество, особенно когда искомым генным продуктом является терапевтически активный белок, еще и потому, что можно избежать деградации конечного продукта микроорганизмами.
Генный продукт можно выделить практически свободным от заражения, в виде композиции, происходящей из сухих семян растения, содержащей один или несколько генных продуктов, накопленных при прорастании трансгенных семян двудольного растения, причем предпочтительно выделить в точке максимального накопления путем остановки процесса прорастания. Генный продукт можно получить в виде смеси из проросших семян двудольного растения для дальнейшей обработки или в стабильном при хранении виде. Композиция может содержать, к примеру, белки, ферменты, пептиды, гормоны, факторы роста, вакцины, аминокислоты, витамины и/или антибиотики.
В настоящем изобретении применяется искусственная система экспрессии, чтобы направить синтез на продукцию нужных генных продуктов, при этом аминокислоты, углеродные соединения и другие генные продукты, высвобождаемые при прорастании, служат источником сырья для синтеза de novo трансгенного продукта. Природные или собственные генные продукты - протеиназы, липазы, амилазы и др., образовавшиеся в прорастающих семенах, могут служить субстратами для трансгенного фермента или вырабатывать нужные биохимические соединения.
Целью настоящего изобретения является получение трансгенного растения, содержащего, по меньшей мере, одну систему экспрессии, включающую, по меньшей мере, одну последовательность ДНК, кодирующую, по меньшей мере, один искомый генный продукт, причем эта последовательность функционально соединена с регулирующими экспрессию последовательностями, которые индуцируются или могут быть индуцированы при прорастании, но практически не проявляются или не могут проявиться при выращивании двудольного растения в полевых условиях. В экспрессионных системах настоящего изобретения могут применяться промоторы, активные в темноте (без освещения), которые в настоящем изобретении именуются промоторами camera obscura, включая более или менее светонезависимые промоторы rubisco.
Настоящее изобретение дает возможность получать различные искомые генные продукты, а также другие негенные биохимические соединения. Экспрессируемый при прорастании трансген образует первый генный продукт - РНК, из которой можно получить другие РНК-продукты сразу или после трансформации. Обычно стадия РНК непродолжительна, а генный продукт получают в виде белкового продукта. В основном существует два вида белковых продуктов - ферменты и неферментные продукты, которые часто являются структурными белками, вакцинами, гормонами и т.д. Ферменты способны модифицировать внутриклеточные субстраты самого растения, которые превращаются в искомые биохимические соединения. С другой стороны, можно добавить внеклеточный субстрат, который будет превращен в другое биохимическое соединение. После этого белок, обладающий или не обладающий ферментативной активностью, можно привести в контакт с реагентом и получить производное этого белка.
В настоящем изобретении используют гетерологичные чужеродные гены или последовательности ДНК, предназначенные для оптимальной экспрессии в прорастающих семядолях двудольных растений. Характеристики последовательностей генов растения и гетерологичных генов проверяют, чтобы экспрессировались чужеродные трансгенные гены из гетерологичных источников. У растений полезно сравнить структуру этих генов с известными генами растений. Поисковые работы в этой области проводились на генах эндотоксина (cry) из Bacillus thuringiensis. Для усиления экспрессии богатых AT генов cry в растениях необходимо произвести некоторые модификации в исходной последовательности генов.
У растений предпочтительной последовательностью начала транскрипции является AACAATGG (наиболее консервативные положения нуклеотидов выделены жирным шрифтом). Из числа стоп-кодонов TGA, TAG, TAA первый имеет небольшое предпочтение, а TAA почти не используется у однодольных растений. Кодоны в последовательности гена можно преобразовать в кодоны, свойственные растениям, с помощью таблиц частоты употребления кодонов. После преобразования последовательность ДНК следует проанализировать на присутствие сайтов, способных уменьшить экспрессию в растениях. Обычно сигналами полиаденилирования у растений служат ААТААА, ААТААТ и их варианты: ААССАА, АТАТАА, ААТСАА, АТАСТА, АТАААА, ATGAAA, AAGCAT, АТТААТ, АТАСАТ, ААААТА (наиболее консервативные положения нуклеотидов выделены жирным шрифтом). Предполагаемые сайты сплайсинга, которые следует убрать из последовательности, - это CAN7-9AGTNNA. Кроме того, в последовательности ДНК не должно быть последовательности АТТТА, которая предположительно является элементом деградации мРНК.
Последовательности терминации транскрипции можно получить из различных генов, экспрессируемых в семенах, или из других генов растений, например из гена Rubisco. Можно использовать сигналы терминации из различных бактерий, например из Agrobacterium, а также из генов nos или ocs. Ген nos кодирует нопалин-синтетазу (Depicker A. et al. (1982) J. Mol. Appl. Genet. 1: 561-573; Shaw C.H. et al. (1984) Nucl. Acids Res. 12: 7831-7846; и An G. et al. (1986) Mol. Gen. Genet. 203: 245-250). Ген ocs кодирует октопин-синтетазу (Koncz С. et al. (1983) EMBO J. 3: 1597-1603; Dhaese P. et al. (1983) EMBO J. 2: 419-426).
РНК-полимераза II транскрибирует белок-кодирующие гены у растений практически таким же образом, как у других эукариотических организмов. В участках инициации транскрипции (промоторах) эукариотических клеток находятся очень близкие в своей основе регуляторные элементы. Рамка ТАТА находится на 25-40 нуклеотидов впереди, а кодон инициации трансляции ATG - на 40-80 нуклеотидов позади от сайта инициации транскрипции. В зрелой мРНК за кодоном ATG следует открытая рамка считывания, которая оканчивается одним из трех стоп-кодонов: UGA, UAG или UAA.
Для экспрессии трансгенов в растениях можно использовать промоторы из различных источников. Многие из них происходят из Agrobacterium ssp. и из вирусов растений, как конститутивный промотор nos из Agrobacterium и промотор 35S вируса мозаичности цветной капусты. Тканеспецифичные промоторы полезны в тех случаях, когда требуется экспрессия в конкретных тканях. Обычно тканеспецифичные промоторы к тому же подвергаются онтогенетической регуляции таким образом, что они активны только на определенных стадиях развития ткани. В настоящем изобретении желательно, чтобы промотор обладал специфичностью к прорастающим семенам.
В прорастающих семядолях фермент rubisco (рибулозо-1,5-бифосфаткарбоксилаза/ оксигеназа) составляет до 50% от общего белка. Он является наиболее распространенным белком растений и катализирует реакцию конденсации между молекулой СО2 и рибулозо-1,5-бифосфатом с образованием двух молекул 3-фосфоглицерата. Rubisco находится на стромальной поверхности тилакоидных мембран. Фермент состоит из 8 больших субъединиц, кодируемых в геноме хлоропластов, и 8 малых субъединиц. Малая субъединица кодируется ядерным мультигенным семейством (гены rbcS). Малая субъединица синтезируется на цитоплазматических полисомах в виде предшественника. Она переносится в пластиды и подвергается процессингу перед сборкой голофермента. Экспрессия генов rbcS специфична к ткани и стадии развития и контролируется освещением через систему фитохрома.
В настоящем изобретении в наиболее предпочтительных генных продуктах или системах продукции белка применяются промоторы гена малой субъединицы rubisco. Их можно выделить из выращенных в темноте (camera obscura) или без освещения семядолей Brassica campestris. Этот вид промотора способен продуцировать чужеродные белки на очень высоком уровне. Наши работы показали, что около 50% от всех синтезированных de novo белков, прорастающих в темноте семядолей В. campestris, составляют белки малых и больших субъединиц rubisco. Содержание мРНК природного транскрипта гена rbcS в прорастающих семенах достигает 500 пг в 1 мкг тотальной РНК. Максимальный уровень экспрессии достигается примерно через 60 часов после начала вымачивания. Близкие результаты отмечались в отношении уровня мРНК природных транскриптов генов rbcS у Brassica napus (Fiebig С. et al., Bot. Acta 103 (3): 258-265, 1990). Прорастающие семена можно освещать видимым светом в короткие или длинные промежутки времени для повышения экспрессии. Можно добавлять моно-, ди- и олигосахариды, предпочтительно сахарозу или глюкозу, чтобы репрессировать активность природного промотора rbcS. Если из трансгенного промотора удалить реагирующий на сахарозу элемент, то он не будет репрессироваться сахарозой. Чем меньше вырабатывается фермента rubisco, тем больше остается свободных аминокислот для экспрессии трансгена. Также можно применять антисмысловые методы для подавления экспрессии эндогенного гена rbcS - экспрессировать антисмысловой ген rbcS в прорастающих семенах.
Поскольку rubisco является самым распространенным растительным белком, его промотор применялся предпочтительно в настоящем изобретении, однако экспрессионная система настоящего изобретения никоим образом не ограничивается применением данного промотора. Трансгенные семядоли для продукции белков преимущественно содержат промоторную систему для продукции белков, предпочтительно индуцируемый светом промотор rubisco, обладающий высокой активностью в семядолях. Можно получить другие сильно экспрессирующие промоторы rubisco путем скринирования кДНК rubisco, экспрессируемой в семядолях, особенно в отсутствие освещения. кДНК можно получить методом RT-ПЦР с помощью олиго-Т-праймеров и последовательностей на 5'-конце кодирующей последовательности сильно консервативного фермента rubisco. Можно секвенировать кДНК с высоким уровнем экспрессии в семядолях и использовать вариабельную 3'-нетранслируемую область (UTR) в качестве зонда для выявления уровня экспрессии в тканях растения. Можно воспользоваться базами данных по последовательностям кДНК и выделить промоторы, экспрессирующиеся только в семядолях, различными методами. Аналогичным образом можно использовать информацию по кДНК для получения других, в сильной степени специфичных к семенам и активируемых и/или индуцируемых при прорастании промоторов, которые в настоящем изобретении именуются промоторами camera obscura.
Многие протеиназы и липазы специфически экспрессируются в семядолях прорастающих семян. Одной из наиболее изученных является сульфгидрил-эндопептидаза (SH-EP) Vigna mungo, которая синтезируется de novo в семядолях вскоре после вымачивания. Обычно уровень мРНК SH-EP повышается вплоть до 3-го или 4-го дня. Промоторный участок SH-EP (номер в банке генов: EMBL X51900) соединяют с геном GUS, чтобы облегчить измерение уровня экспрессии при прорастании семян двудольных растений.
Наряду с промоторами, которые регулируются тканеспецифичным образом, могут применяться индуцибельные промоторы, активируемые при различных воздействиях. У всех организмов, от бактерий до человека, встречаются гены одного класса, известные как гены теплового шока или стресса. Транскрипция этих генов начинается после стрессового воздействия (к примеру, теплового шока), а при трансляции этих транскриптов образуются белки, которые, вероятно, временно защищают клетки. Продукция мРНК и белков теплового шока является временным явлением: экспрессия генов теплового шока выходит на плато через несколько часов, а затем спадает. Если температура повышается медленно, а не скачком, то организм выдерживает температуры, которые могли бы оказаться летальными, то есть организм может адаптироваться к высокой температуре. Проросшие семена можно нагревать, чтобы повысить активность промоторов теплового шока. Было показано, что уровень мРНК теплового шока (ТШ) сои повышается от едва заметного до 15000-20000 копий через 2 часа после обработки при 40°С. В качестве индуцибельного промотора экспрессионной системы настоящего изобретения можно использовать промоторный участок ТШ, описанный в патенте US 5,447,858. Для измерения уровня экспрессии при прорастании семян промотор ТШ клонируют методом ПЦР и соединяют с геном GUS.
При мобилизации запасных веществ в прорастающих семенах синтезируются de novo и функционируют несколько типов гидролитических ферментов. В превращении запасных веществ в подвижные формы, например триацилглицерина в сахарозу, участвуют несколько сильно экспрессируемых генов типа изоцитратлиазы и малатдегидрогеназы. Промоторы из этих генов можно использовать для экспрессии трансгенов. Можно клонировать промотор, индуцируемый салицилатом, из растений табака Nicotiana tabacum с помощью ПЦР. Его можно использовать для экспрессии гетерологичных генов в прорастающих семенах.
Большим достоинством настоящего изобретения с точки зрения экологии является то, что чужеродный ген не экспрессируется при выращивании в полевых условиях. Ген размножается и отлагается вместе с запасными веществами, тогда как экспрессия продукта чужеродного гена может проводиться в закрытых условиях.
В принципе, по настоящему изобретению можно получить любой коммерчески привлекательный генный продукт. К ним относятся ферменты, гетерологичные для данного растения. Иными словами, они чужеродны для тех растений, которые их продуцируют. К ним же относятся ферменты, гомологичные для тех растений, которые их продуцируют. Эти гомологичные белки обычно подвергают суперэкспрессии методами рекомбинантной ДНК. К числу ферментов, представляющих интерес, относятся такие гидролазы, как протеазы, β-глюканазы, целлюлазы, геми-целлюлазы, фосфатазы, липазы, фосфолипазы, пектиназы, амилазы, амилогликозидазы, лизоцимы, пуллуланазы и хитиназы, а также пероксидазы, лиазы типа пектинолиазы, изомеразы типа глюкозоизомеразы, но ими можно не ограничиваться.
Также возможно получение структурных белков способом и с помощью продукционной системы настоящего изобретения. К интересным воплощениям настоящего изобретения относятся коллаген, желатин, спидроин, белок шелка, а также можно получать много других белковых генных продуктов, включая ингибиторы ферментов типа ингибитора трипсина, терапевтические пептиды - интерфероны, инсулин, нейропептиды, энкефалин, соматостатин и т.д. Также можно получать полиаминокислоты - полилизин и полиглутамат, а также волокна, мембраны, пленки, лекарственные препараты, антинакипины, или же пищевые добавки, вакцины типа LTB, факторы роста типа GM-CSF. Также могут быть синтезированы небелковые генные продукты, включая поли-β-гидроксибутират.
Поскольку синтез de novo при прорастании представляет быстро проходящую ситуацию, прорастание необходимо прервать или остановить до того, как количество искомого генного продукта начнет снижаться. Это достигается выделением требуемого генного продукта перед снижением его концентрации, останавливая прорастание либо путем нагревания в атмосфере N2, или же путем измельчения или срезания проросших семян двудольных растений. Если прорастание останавливают путем нагревания, то искомый генный продукт можно получить в виде неочищенного высушенного продукта, который хорошо хранится, а это значит, что таким же образом можно получить фермент в неочищенном, стабильном при хранении виде. Синтез искомого белка также можно остановить на пике путем измельчения или срезания прорастающих семян. Если прорастание останавливают путем срезания или измельчения, то генный продукт можно получить в виде неочищенной смеси и использовать в таком виде или подвергнуть дальнейшей обработке - формулировать, хранить, перевозить. Предпочтительно его используют именно в таком виде, например, в процессах биотрансформации. Естественно, искомый генный продукт можно выделить и очистить стандартными, хорошо известными методами.
Когда генный продукт используют для биотрансформации, то его приводят в контакт с субстратом - определенным соединением или смесью соединений, при этом генные продукты (часто это ферменты) реагируют с соединениями субстрата. При этом либо генный продукт (фермент) действует как катализатор и модифицирует соединения субстрата либо же субстрат или реагент содержит как минимум одно вещество или соединение, способное модифицировать генный продукт, например изменить его профиль укладки, осуществить гликозилирование или дегликозилирование, вставить замещающую группу, способствующую конъюгации этого генного продукта с другими продуктами.
В настоящем изобретении после прорастания, в условиях, способствующих выработке искомого генного продукта, происходит обогащение субстрата этим генным продуктом. Происходящие из семян композиции включают семядоли, гипокотили, корешки, а у растений с подземным типом прорастания - также эпикотили и иногда первичные листочки. Генный продукт можно экстрагировать или оставить в композиции.
Генным продуктом может быть белок, кодируемый геном, введенным в составе экспрессионной системы настоящего изобретения, или другое биохимическое соединение, вырабатываемое трансгенным ферментом, когда продукты собственных генов, запасенные в семенах, мобилизуются при прорастании и используются непосредственно для реакции с добавленным извне субстратом, осуществляя требуемую биотрансформацию субстрата.
Когда генный продукт экстрагируют из композиции, то его используют либо в свежем либо в высушенном виде. Сушка проводится с использованием сушильного шкафа при 40-80°С или в лиофильной сушке при -20-80°С. Высушенный материал можно измельчить или превратить в порошок. Перед экстракцией композицию можно разрушить механическим путем и/или обработать ферментом, получая весь белок в виде кашицы или раствора. Затем можно отделить остатки клеток любым удобным способом - центрифугированием, осаждением и/или фильтрованием.
В супернатанте или фильтрате обычно содержится от 1 до 40% искомого генного продукта от общего веса белка в среде, предпочтительно не менее 30%. Когда искомый продукт не растворяется в воде, его можно, к примеру, экстрагировать соответствующим растворителем. В качестве альтернативы применяются другие процессы, позволяющие ренатурировать, солюбилизировать и/или экстрагировать продукт без потери его активности.
После выделения искомого белка из водной среды генный продукт можно очистить стандартными способами. Поскольку генный продукт обычно составляет значительную часть общего белка в смеси, зачастую больше, чем любой другой белок, то очистка сильно упрощается. Кроме того, примеси в продукте после очистки вряд ли будут иметь физиологическое значение для любых применений генных продуктов, включая применение в терапии.
Когда генный продукт не экстрагируется из композиции, а остается в ней, его опять же можно получить в свежем или высушенном виде. Сушка проводится, как описано выше. Затем высушенный материал можно измельчить или превратить в порошок. Свежий материал можно размельчить механическим путем или обработать ферментом. И сухой и свежий материал может служить источником генного продукта. Если генный продукт является ферментом, можно добавить соответствующие субстраты в водную смесь или в сухую композицию или использовать в свежем виде.
В одном из воплощений настоящего изобретения проросшие семена, семядоли и/или ростки двудольных растений, содержащие один или несколько искомых белков, можно использовать в качестве кормовой добавки. Проросшие семена двудольных можно смешивать с обычными нетрансгенными семенами, чтобы получить требуемую концентрацию искомого генного продукта в корме для животных.
Происходящие из проросших семян смеси или композиции можно высушивать для хранения. Генные продукты можно выделять или оставлять в самом материале, например, когда рекомбинантный белок применяется как корм для животных. Генные продукты можно использовать для повышения содержания белка в кормах для животных или, к примеру, в качестве гормона роста или вакцины. Также можно получать лабильные или токсические вещества в строго контролируемых условиях и при этом эффективная продукция белка сочетается с экологическими аспектами и интересами сельского хозяйства.
Сведения, подтверждающие возможность осуществления изобретения
Более подробно изобретение описывается ниже. Представленные примеры и экспериментальные результаты служат для лучшего понимания и в качестве руководства для специалистов в этой области.
ПРИМЕР 1
Экспрессия гена GUS в прорастающих семенах Brassica campestris
Экспрессию GUS определяли гистохимическим методом. Для регуляции экспрессии GUS применяли 4 различных промотора. Использовали промотор теплового шока сои, промотор эндопептидазы Vigna mungo, промотор pr Nicotiana tabacum и промотор 35S вируса мозаики цветной капусты (CaMV). Последовательности промоторов получали методом ПЦР, используя в качестве матрицы тотальную ДНК растения.
Промоторы присоединяли к гену GUS с помощью фермента Ncol. Конструкции промотор-GUS клонировали в векторе для трансформации растений pGPTV-hpt (Becker et al., 1992, Plant Mol. Biol. 20: 1195-1197). В. campestris трансформировали по методике, описанной Kuvshinov V. et al. (Plant Cell Reports 18: 773-777, 1999). Трансгенные растения выращивали в теплице до образования семян. Трансгенные семена проращивали и использовали для гистохимического определения GUS.
Результаты представлены на фиг.1-5.
На фигуре 1А слева показано проросшее семя трансгенного растения В. campestris, экспрессирующее ген GUS. Ген GUS находится под контролем промотора теплового шока сои. Справа показан нетрансформированный контрольный росток.
На фигуре 1В показано проросшее семя трансгенного растения Brassica campestris, экспрессирующее ген GUS. Ген GUS находится под контролем промотора эндопептидазы.
На фигуре 1С показано проросшее семя трансгенного растения Brassica campestris, экспрессирующее ген GUS. Ген GUS находится под контролем салицилат-индуцируемого промотора.
На фигуре 1D показано проросшее семя трансгенного растения Brassica campestris, экспрессирующее ген GUS. Ген GUS находится под контролем промотора 35S.
На фигуре 2 показано, что бесцветный буферный раствор, содержащий β-глюкуронид (100 мкг/мл), приобретает синюю окраску в результате ферментативного превращения этого субстрата в глюкопиранозидуроновую кислоту и агликон под действием фермента GUS, вырабатываемого в проросших трансгенных семенах.
На фигуре 3 показаны различные стадии прорастания семян Brassica campestris через указанное число дней после начала прорастания. Стерилизованные с поверхности семена проращивали в условиях in vitro с освещением в течение 16 ч при температуре 22°С или без освещения (в темноте) в течение 8 ч при температуре 18°С.
На фигуре 4 представлен SDS-PAGE-гель прорастающих семян. Образцы отбирали ежедневно, начиная с сухих семян. Материал гомогенизировали в жидком азоте и ресуспендировали в 50 мМ трис, рН 8,0, в присутствии 850 мМ NaCl. После центрифугирования прозрачный супернатант смешивали с буфером для образца. На окрашенном Кумасси геле отмечены запасные белки и субъединицы фермента rubisco. Через три дня после прорастания продуцируемое за день количество белка rubisco становится значительным.
На фигуре 5 представлен результат Northern-гибридизации прорастающих семян с помощью РНК-зондов. Образцы отбирали ежедневно, начиная с сухих семян. Антисмысловой зонд содержит 197 bp из экзона №3 и 153 bp из участка 3'-UTR кДНК малой субъединицы rubisco (не опубликовано). Зонд получали с использованием плазмиды pBluescript (промотор Т7), используя при реакции DIG-UTP. Вносили по 3 мкг тотальной РНК на дорожку. РНК выделяли с помощью набора RNeasy Plant Mini Kit фирмы Qiagen. Гибридизацию проводили с помощью буфера DIG Easy Hyb в основном согласно методике фирмы-изготовителя.
ПРИМЕР 2
Получение новых промоторов rbcS для экспрессионной системы
a) Выделение нового промотора rbcS
Секвенировали клоны кДНК rbc из 4-дневных трансгенных семядолей Brassica campestris. Клоны разделяли на 2 группы с близкими характеристиками на основании последовательности участков 3'-UTR.
Анализ методом SDS-PAGE ростков Brassica campestris. Семена Brassica campestris проращивали сериями от 1 до 7 дней. Пять пар семядолей измельчали в жидком азоте и ресуспендировали в буфере для лизиса образцов (50 мМ трис-HCl рН 8,5, 2% SDS, 0,1% бромфенола синего, 10% глицерина, 2% меркаптоэтанола. Образцы кипятили 10 мин на водяной бане и анализировали методом SDS-PAGE в 15%-ном геле по методу Laemmli (1970). Результаты показали, что экспрессия гена rbc начинается через 4 дня после вымачивания.
b) Очистка РНК из семядолей Brassica campestris
РНК очищали из 4-дневных семядолей с помощью набора RNeasy Kit фирмы Qiagen.
c) Синтез кДНК из тотальной РНК семядолей Brassica campestris
2 мкг тотальной РНК и 10 нг праймера notI-d(T)18 в 16 мкл воды инкубировали 5 мин при температуре 70°С. Смесь переносили на лед и добавляли 5 мкл 5-кратно концентрированного буфера для реакции, 2,5 мкл 5 мМ dNTP, 20 Ед. ингибитора рибонуклеазы и 200 Ед. обратной транскриптазы M-MLV. Реакционную смесь инкубировали 1 ч при температуре 42°С.
d) ПЦР-амплификация клонов кДНК, экспрессируемых в семядолях
ПЦР предварительно оптимизировали с помощью серии из 12 праймеров, причем 3'-праймеры состояли из тиминовых нуклеотидов и переменного динуклеотидного якоря, а 5'-праймеры представляли собой 49-мерный олигонуклеотид из начального участка экзона №3 гена rbc Brassica napus (номер доступа в GeneBank g 17849. Предварительная оптимизация показала, что из 3'-праймеров только GCT11 давал продукт ПЦР. Для клонирования этого продукта ПЦР был сконструирован 49-мерный олигонуклеотид, содержащий сайт рестрикции NotI. Праймеры перечислены в таблице 1.
Таблица 1. ПЦР-праймеры для выделения клонов кДНК rubisco
3'-Праймеры для предварительного скринирования:
1. 5'-ТТТТТТТТТТТСА-3' (Посл. №1)
2. 5'-ТТТТТТТТТТТСТ-3' (Посл. №2)
3. 5'-ТТТТТТТТТТТСС-3' (Посл. №3)
4. 5'-TTTTTTTTTTTCG-3' (Посл. №4)
5. 5'-TTTTTTTTTTTGA-3' (Посл. №5)
6. 5'-TTTTTTTTTTTGT-3' (Посл. №6)
7. 5'-TTTTTTTTTTTGC-3' (Посл. №7)
8. 5'-TTTTTTTTTTTGG-3' (Посл. №8)
9. 5'-ТТТТТТТТТТТАА-3' (Посл. №9)
10. 5'-ТТТТТТТТТТТАТ-3' (Посл. №10)
11. 5'-ТТТТТТТТТТТАС-3' (Посл. №11)
12. 5'-TTTTTTTTTTTAG-3' (Посл. №12)
5'-Праймер (Посл. №13):
5'-CGCGGATCCCACGGGTTTGTTTACCGTGAGCACGGAAGCACCCCCGGAT-3'
3'-Праймер для клонирования клонов кДНК гена rbc (Посл. №14):
5'-AAGGAAAAAAGCGGCCGCAATTTTTTTTTTTTTTTTTTTTTTTTTTTGC-3'
Для клонирования фрагментов кДНК гена rbc проводили ПЦР-амплификацию, используя в качестве матрицы 1 мкл реакционной смеси для синтеза кДНК, 100 нМ 5'-праймера, 100 нМ 3'-праймера, 100 мкМ dNTP и 1,25 Ед. ДНК-полимеразы Pfu в объеме 25 мкл. Полученный ПЦР-фрагмент клонировали в векторе pUK21 по сайтам NotI и BamHI. Семь индивидуальных клонов секвенировали. Исходя из последовательности, клоны разделили на 2 группы, состоящие из 3 и 4 представителей соответственно. Для дальнейшего анализа было отобрано по одному представителю из каждой группы. Их последовательности получили наименования utr2 (Посл. №15) и utr8 (Посл. №16).
Последовательность utr2 (Посл. №15):
TTCGCGTTGT AAGACATTTC АТАААТААТА ТСТАССТСАТ
ТТСАТТТССА TTTGTCTGTTT TCTTTGGCTTT TTGTTTCTGA
GGCATGTTAT ATCGGATTGT CAAGTGTCTG ATTTATGAAC
AACATGTAAT CTCTATATGC АТАТТТСТ
Последовательность utr8 (Посл. №16):
TTCGCTTTCA ТАТААТААТА ТСТТССТСАТ ТТСАТТТССА
ATAAGTCTGT ТТСТТТТТТС TCTTTGGATT TCTGTTACGA
GACTTTCTAT ATCGGATTGT AAAATGTCTG ATTTTATGAA
CATGTAATTT CGGCAAATA
е) Клонирование промотора 1 типа (65А) малой субъединицы rubisco из Brassica campestris
Фрагмент в 2,8 kb, содержащий промотор и ген, соответствующий UTR 65А (Посл. №24), амплифицировали с помощью 5'-праймера bnrbl (Посл. №17) (5'-GAATTCTAAC GACCCTTTTCCG-3'), комплементарного гену малой субъединицы rubisco В. napus, и 3'-праймера UTR2 (Посл. №18) (5'-GGCCACACTT GACAATCCGA TATAACATGC CTCA-3'), специфичного к UTR 65A (Посл. №24). Амплификацию проводили с помощью ДНК-полимеразы Pfx Platinum (Life Technologies) согласно рекомендациям изготовителя. Промоторные участки амплифицировали, используя в качестве матрицы полученный фрагмент. В качестве 5'-праймера использовали bnrb3 (Посл. №19) (5'-AAAAAGCTTC TAGACCCTTT TCCGTCATAA GTTTTATA-3'), который комплементарен гену малой субъединицы rubisco В. napus, а в качестве 3'-праймеров - RbSiB (Посл. №20) или RBNCO (Посл. №21) в отдельных реакциях. 3'-Конец RbSiB (5'-CAGGTCTCCC ATGCAGCTAA CTCTTCCTCC GTTGCT-3') комплементарен концевому участку предполагаемого пептида адресного сигнала хлоропластов гена малой субъединицы rubisco В. napus, а 5'-конец содержит сайт Есо31I, который после расщепления образует содержащий ATG и совместимый с сайтом NcoI конец для клонирования в экспрессионные векторы растений. RBNCO (Поел. №21) (5'-GAGGAAGCCATGGCT АСТТСТТ-3') комплементарен начальному участку кодирующей области гена малой субъединицы rubisco В. napus, за исключением замены двух нуклеотидов, при которой образуется сайт Ncol вокруг кодона ATG.
f) Анализ пула мРНК, кодирующей малую субъединицу рибулозо-1,5-бифосфаткарбоксилазы в листьях и семенах Brassies campestris
Тотальную РНК выделяли из зрелых листьев и 4-дневных ростков с помощью набора RNeasy (Qiagen). Матричную РНК дополнительно очищали на колонке с олиго((1Т)-целлюлозой согласно рекомендациям изготовителя (New England Biolabs). Для синтеза тотального пула кДНК использовали олигоТ-праймер (Посл. №22) (CUACUACUACUA GCGGCCGCTT ТТТТТТТТТТ ТТТТТТТТТТ TTTVN) и обратную транскриптазу вируса лейкемии мышей Молони (Promega). кДНК малой субъединицы rubisco амплифицировали за 2 цикла ПЦР (1 мин при 94°С, 1 мин при 55°С, 1 мин при 72°С) с помощью олигоТ-праймера и праймера, комплементарного концевому участку последнего экзона малой субъединицы rubisco (Посл. №23) (CAUCAUCAUCAU TCGACGATCA TCGGATTCGA CA). Продукт ПЦР обрабатывали урацил-ДНК-гликозилазой и клонировали в вектор pAMPI (CloneAMP pAMP System фирмы Life Technologies). Были проанализированы 102 клона путем секвенирования ДНК: 42 клона из проросших семян и 60 клонов из зрелых листьев. Были обнаружены 3 различных типа мРНК (I, II и III тип). Последовательности и процентное содержание различных типов последовательностей в проросших семенах и зрелых листьях представлены в таблице 2.
Последовательности трех различных типов мРНК приводятся ниже.
I тип (Посл. №24):
TTAATTCGCG TTGTAAGACA ТТТСАТАААТ ААТАТСТАСТ САТТТСАТТТ
CCATTTGTCT GTTTTCTTTG GCTTTTTGTT TCTGAGGCAT GTTATATCGG ATTGTCAAGT
GTCTGATTTA TGAACATGTA ATCTCTATAT TGCATATTTT CTTCTTGGAA ААААА
ААААА АААААААААА
II тип (Посл. №25):
TTAATTTGCT ATGACATTCA CATAATAATC TCTGCTCATT TCATTTCCAA TTGT
CTGTTT СТТТТСССТТ TGGTTTTCTG TTTCTCAGAC ATTCTATATC GGATTGTC
AAATGTGTGATT GTGAACATGT ААТСТСТАТА TTGCTTCTTC GTCTTGGTAAA
ААААААААА АААААААААА
III тип (Посл. №26):
TTAATTCGCT ТТСАТАТААТ ААТАТСТТСТ САТТТСАТТТ CCAATAAGTC TGTTT
СТТТТ TTTCTCTTTG GATTTCTGTT ACGAGACTTT CTATATCGGA TTGTAAAAT
GTCTGATTTTA TGAACATGTA ATTTCTAAAA AAAAAAAAAA AAAAAAAAAA AA
AAAAAAAA AAAAAA
g) Клонирование наиболее распространенного гена малой субъединицы rubisco из Brassica campestris
Один из генов малой субъединицы rubisco В. campestris составляет 56% от всех транскриптов, как показал количественный анализ последовательностей 102 клонированных UTR-клонов малой субъединицы rubisco. Этот клон получил наименование UTR56 (III тип. Посл. №26). Для клонирования соответствующего гена и его вышележащего участка были разработаны 4 праймера для 2 стадий ПЦР. На первой стадии ПЦР в качестве 5'-праймера использовали праймер bnrb4 (Посл. №27) (5'-GGCCATGAAT TC TAACGACC CTTTTCCGTC ATAAAAGT-3'), представляющий собой часть гена малой субъединицы rubisco В. napus (номер доступа х61097). В качестве 3'-праймера использовали праймер 56r6 (Посл. №28) (5'-CGCGATATAG AAATTACATG ТТСАТААААТ CAGACATTTT АС-3'), обладающий специфичностью к UTR56 (Посл. №26). Состав смеси для первой реакции ПЦР: 75 мМ трис, рН 8,0 при 25°С, 20 мМ (NH4)2S04, 0,01% твин 20, 2 мМ MgCI2, 200 мкМ dNTPs, по 0,2 мкМ праймеров, 0,025 ед./мкл ДНК-полимеразы Taq, 0,0016 ед./мкл ДНК-полимеразы Pfu и 100 пг/мкл ДНК В. campestris. Программа: сначала 4 мин при 94°С, затем 30 раз повторяли цикл из 30 с при 94°С, 30 с при 38°С, 3 мин при 72°С. Ожидаемый фрагмент в 2,8 kb очищали из агарозного геля. Ввиду отсутствия надежной информации о последовательности этот фрагмент амплифицировали повторно с помощью праймера bnrb5 (Посл. №29) (5'-GAGGTACCCG CGGCC GCGAA TTCTAACGAC CCTTTTCCGT CATAAAAG-3'), у которого 3'-конец комплементарен bnrb4 (Посл. №27), а 5'-конец имеет удлинение с сайтом расщепления NotI из 8 оснований, и праймера 56r7 (Посл. №30) (5'-GAGAATTCGG CGCGCCATAG AAATT ACATG ТТСАТААААТ CAGACA-3'), у которого 3'-конец комплементарен 56r6 (Посл. №28), а 5'-конец имеет удлинение с сайтом расщепления AscI из 8 оснований. Состав смеси для ПЦР был таким же, как в первой реакции ПЦР, за исключением того, что в качестве матрицы использовали очищенный в геле фрагмент от первой реакции ПЦР (25 пг/мкл) и проводили только 7 циклов.
Полученный продукт осаждали, расщепляли рестрикционными ферментами AscI и NotI и очищали в геле. Фрагмент клонировали в векторе pAN, представляющем собой модифицированный pNEB193 (сайт Pmel заменен на NotI), который был расщеплен AscI и NotI, обработан щелочной фосфатазой креветок и после тепловой инактивации фермента очищен в геле. Секвенировали 4 из полученных клонов, чтобы удостовериться, что они содержат последовательность UTR56.
Промоторный участок амплифицировали с помощью 5'-праймера bnrb5 (Посл. №29) и 3'-праймеров RbSiB (Посл. №20) RBNCO (Посл. №21) (см. выше). Условия ПЦР были такими же, как при амплификации целого гена, за исключением того, что в качестве матрицы использовали линеаризованную с помощью AscI плазмидную ДНК (5 нг/мкл), содержащую клоны, и проводили только 10 циклов.
ПРИМЕР 3
Сравнение различных конститутивных и индуцибельных промоторов для суперэкспрессии трансгена (GUS) в табаке и Brassica campestris
Проросшие семена замораживали в жидком азоте и измельчали. Экстрагировали белки фосфатным буфером с ЭДТА. Образцы центрифугировали, а в супернатантах определяли β-глюкуронидазную активность с помощью набора (Sigma). Концентрацию белка в образцах определяли с помощью реактива на белок (Biorad), используя в качестве стандарта БСА.
При измерении активности промоторов в прорастающих семенах, например промотора rubisco, наблюдалось экспоненциальное возрастание удельной активности при прорастании на протяжении 7 дней. Удельная активность промотора rubisco составила 0,25 на 4-й день, 0,5 на 5-й день, 1,6 на 6-й день и 7 на 7-й день.
Влияние ингибиторов развития хлоропластов на уровни белка и мРНК rubisco в прорастающих семенах
Исследование прорастающих натуральных семян В. campestris показало, что самая высокая концентрация белка малой субъединицы rubisco достигается через 3 дня после начала прорастания, а мРНК малой субъединицы rubisco - на один день раньше. Мы обнаружили, что добавление стрептомицина (100 мг/л) в культуральную среду через 48 часов после начала прорастания дает возможность ослабить синтез белка rubisco, но при этом уровень мРНК остается постоянным. Поскольку белок rubisco является основным белком в определенных типах прорастающих семядолей, то ослабление его синтеза будет полезным. Семена проращивали в аэрированной воде с добавлением стрептомицина. Образцы отбирали с интервалом в 12 часов. Содержание белка определяли методом SDS-PAGE, а уровень мРНК - методом точечной РНК-гибридизации.











Изобретение относится к генной инженерии и селекции растений. Получают двудольные растения, содержащие семена, включающие гетерологичные белки. Экспрессия гетерологичных белков становится возможной благодаря встраиванию в геном растения экспрессионной системы, содержащей регуляторные последовательности белков, синтезирующихся de novo, и индуцирующейся во время прорастания семян двудольного растения. Получение гетерологичного белка основано на принципе источник - сток путем мобилизации белковых запасов в семенах, накопленных во время культивирования растения. 17 з.п. ф-лы, 5 ил., 3 табл.
(a) получение и культивирование трансгенного двудольного растения, трансформированного одной или несколькими экспрессионными системами, включающими ген гетерологичного белка, находящийся под контролем регуляторных последовательностей;
(b) получение трансгенных семян из данного трансгенного двудольного растения;
(c) помещение трансгенных семян двудольного растения, содержащих одну или несколько экспрессионных систем, встроенных в геном растения, в условия, подходящие для прорастания, при котором происходит запуск экспрессионной системы;
(d) экспрессия одного или нескольких гетеролоичных белков во время прорастания под контролем регуляторных последовательностей;
(e) получение прорастающих семян и/или проростков двудольного растения;
(f) выделение экспрессированного гетерологичного белка,
характеризующийся тем, что регуляторные последовательности включают регуляторные последовательности белков, синтезирующихся de novo, и обеспечивают высокий уровень экспрессии во время прорастания семян двудольного растения.
(а) выделение тотальной РНК из семядолей и листьев двудольных растений;
(b) получение кДНК путем обратной транскрипции и амплификации этой РНК с помощью одного праймера, специфичного к данной РНК, и другого праймера, специфичного к тому белку, который экспрессируется на высоком уровне в семенах двудольных растений во время прорастания;
(c) сбор и рассев клонов, содержащих кДНК;
(d) сравнение клонов, содержащих кДНК, полученную из РНК, выделенную из семядолей и из листьев;
(e) выделение клонов, содержащих кДНК, проявляющую высокую активность в прорастающих семенах двудольного растения.
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| Экономайзер | 0 |
|
SU94A1 |
| US 5994628 А, 30.11.1999 | |||
| US 5608143 А, 04.03.1997. | |||

