Настоящая заявка представляет собой частичное продолжение патентной заявки США №09/012453 с притязанием на приоритет предварительной заявки №60/072156.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение включает в себя способы и составы для модификации соединений, получаемых в результате вторичных процессов обмена веществ в растениях. Также настоящее изобретение включает в себя клетки растений с модифицированным уровнем содержания вторичных продуктов обмена веществ и семена растений с модифицированным уровнем содержания вторичных продуктов обмена веществ. В одном из вариантов осуществления изобретения изменяется содержание вторичных продуктов обмена веществ, нежелательных с точки зрения питательных свойств, в растениях, клетках растений и семенах растений. В другом варианте осуществления изобретения изменяется уровень содержания продуктов, входящих в фенилпропаноидные и сахарно-спиртовые процессы обмена веществ в растениях, клетках растений и семенах растений. Помимо этого настоящее изобретение включает в себя генетические конструкции и векторы, которые могут быть использованы для изменения содержания вторичных продуктов обмена веществ в клетках и семенах растений. Дополнительно настоящее изобретение относится к кормовым смесям на основе модифицированных зерновых культур и корму для животных, содержащему модифицированные зерновые культуры, в частности к кормовым смесям на основе зерновых культур с уменьшенным или измененным содержанием вторичных продуктов обмена веществ.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В результате вторичных процессов обмена веществ в растениях вырабатываются разнообразные соединения. Несмотря на свое общепризнанно малое значение для обмена веществ в растениях, вторичные процессы обмена веществ приводят к образованию уникальных биохимических соединений, часть из которых считается нежелательными с точки зрения питательных свойств или даже токсичными. Вторичные процессы обмена веществ и соединения, получаемые в результате них, являются характерными признаками биологических индивидуумов или родов. Таким образом, управление вторичными процессами обмена веществ может быть использовано для получения новых комплексов биохимических соединений или растительных тканей с измененным содержанием вторичных продуктов обмена веществ. В частности, управление вторичными процессами обмена веществ с целью изменения содержания нежелательных с точки зрения питательных свойств или токсичных вторичных продуктов обмена веществ может найти уникальные варианты применения в пищевой и кормовой промышленностях.
В предпочтительном варианте управление вторичными процессами обмена веществ должно осуществляться без вмешательства в биохимические процессы, которые считаются важными для роста и жизнеспособности растительных клеток. Биохимические процессы и участвующие в них химические соединения, имеющие первоочередное значение для роста и жизнеспособности растений, рассматриваются как первичные процессы обмена веществ и их продукты. В общем случае считается, что первичные процессы обмена веществ включают в себя такие биохимические процессы, которые приводят к образованию первичных сахаров (например, глюкозы), аминокислот, жирных кислот, нуклеотидов и производных полимеров (полисахаридов, таких как крахмал, белки, липиды, РНК, ДНК и т.д.) (Yeoman, Yeoman, Tansley Review No. 90, Manipulating Secondary Metabolism in Cultured Plant Cells (Управление вторичными процессами обмена веществ в клетках культивируемых растений). New Phytologist, 134, стр.553-569, 1996).
Таким образом, считается, что первичными процессами обмена веществ являются те процессы обмена веществ, которые влияют на жизнеспособность и рост всех клеток растения, в то время как вторичные процессы обмена веществ могут быть определены как те биохимические процессы, которые не имеют значения для всех клеток растения. Например, вторичные процессы обмена веществ определяют такие свойства растения, как цвет, вкус, морфологию и т.п. Вторичные процессы обмена веществ также приводят к образованию различных химических соединений, которые распознаются насекомыми или участвуют в патогенной реакции растений. Некоторые из этих соединений могут быть полезны для жизнеспособности отдельных растений в условиях дикой природы, но при культивировании окажутся вредными для качества урожая или будут накладывать ограничения на спектр возможных применений такой растительной продукции. Некоторые вторичные продукты обмена веществ представляют собой уникальные соединения, которые вырабатываются отдельными растениями в результате особых биохимических процессов. Биохимические механизмы, задействованные во вторичных процессах обмена веществ, не столь разнообразны, как в случае первичных процессов обмена веществ, вследствие чего количество механизмов образования вторичных продуктов обмена веществ в растениях обычно также ограничено. Вторичные продукты обмена веществ, как правило, характерны больше для индивидуальных растений, чем универсальные биохимические соединения, участвующие в первичных процессах обмена веществ.
Результатом многочисленных попыток управлять первичными процессами обмена веществ стало получение растительных клеток с измененным содержанием крахмала или масел (липидов). Однако серьезное вмешательство в первичные процессы обмена веществ может привести к отрицательным последствиям. Например, несмотря на то что изменение состава липидов может принести положительные результаты, полное удаление липидов, несомненно, отрицательно скажется на жизнеспособности растений. Управление первичными процессами обмена веществ не всегда оказывается успешным, поскольку результаты всех таких попыток могут быть сведены до минимума вследствие активизации альтернативных биохимических механизмов. Таким образом, первичные процессы обмена веществ в растениях редко поддаются управлению таким образом, чтобы результат вмешательства был предсказуемо полезным и заметным в условиях культивирования.
Существуют примеры успешного изменения первичных процессов обмена веществ, в результате чего были получены новые фенотипы, причем в этих случаях изменения касались биохимического состава, а не уменьшения или полного удаления отдельных веществ. Обычно такие результаты достигались путем эктопического изменения степени экспрессии растительного гена, например достижения избыточной экспрессии гена в определенных растительных тканях, нерегулируемой экспрессии гена, а также путем ингибирования отдельного гена с помощью антисенсибилизирующей РНК, рибозим или ко-супрессии. Однако результаты этих экспериментов были трудно предсказуемы заранее.
Степень экспрессии растительного фермента может быть изменена на большом количестве уровней, в том числе на уровне экспрессии его гена, трансляции, видоизменения белка, а также аллостерического управления активностью белка. Поэтому эктопическое изменение экспрессии растительного гена, участвующего в первичных процессах обмена веществ, может оказаться недостаточным для преодоления биохимических факторов регулирования этих процессов. Кроме того, разнообразие альтернативных механизмов первичных процессов обмена веществ также создает серьезные трудности на пути управления этими процессами, поскольку все эти процессы имеют первоочередное значение для роста и жизнеспособности растений. В силу вышеназванных причин попытки внести изменения в первичные процессы обмена веществ очень редко приводят к созданию желаемого фенотипа. Более того, наблюдение за такими генетически модифицированными растениями в полевых условиях или под влиянием разнообразных климатических условий чаще всего приводило к выводу о том, что предсказанное свойство растений не проявляется или же производительность культуры снижается. Таким образом, модифицирование первичных процессов обмена веществ с целью получения желаемых фенотипов требует тщательного анализа их механизмов или отдельных стадий этих процессов.
Управление вторичными процессами обмена веществ было затруднено вследствие слабого понимания биохимии происходящих процессов, недостаточной информации о генах, проявляющихся в ходе этих процессов, и общей сложности взаимосвязей между биохимическими механизмами.
Следует, однако, отметить, что изменение вторичных процессов обмена веществ может стать эффективным способом получения новых фенотипов, включая фенотипы с измененными уровнями содержания продуктов вторичного обмена веществ, например, тех продуктов, которые считаются нежелательными с точки зрения питательных свойств растений. Поэтому механизмы вторичных процессов обмена веществ являются важным направлением исследований по генетической модификации растений.
Объектами исследований, направленных на изменение уровней содержания конечных продуктов вторичных процессов обмена веществ, были два механизма, по которым эти процессы реализуются в растениях. Способы, использовавшиеся для управления этими механизмами, не принесли желаемых результатов. Например, фенилпропаноидный механизм участвует в образовании лигнина и считается вторичным процессом обмена веществ. Биосинтез лигнина является частью общего фенилпропаноидного процесса биосинтеза, в результате которого образуются, по крайней мере, три первичных фенольных предшественника, а именно кумаровая, феруловая и синапиновая кислоты, производные которых полимеризируются с образованием лигнина и прочих фенольных соединений (см. Фиг.2).
В настоящее время в ходе исследований, направленных на модификацию вторичного фенилпропаноидного процесса обмена веществ, гены многих ферментов, участвующих в образовании мономеров лигнина, рассматриваются в качестве возможных объектов воздействия с целью понижения уровня лигнина с помощью методов антисенсибилизирования или ко-супрессии (патенты США №№5451514 и 5633439, а также WO 93/05160 и WO 94/08036). К этим генам относятся гены, которые кодируют дегидрогеназу циннамилового спирта, оксиметилтрансферазу кофеиновой кислоты и лиазу фенилаланина аммония. Вышеуказанные методы направлены на снижение содержания лигнина, в результате чего предполагается достичь благоприятного эффекта с точки зрения технологических и пищевых свойств растений.
Однако снижение уровня содержания лигнина путем антисенсибилизирующего или ко-супрессивного воздействия на один из генов, участвующих в фенилпропаноидном механизме обмена веществ, может иметь ряд нежелательных последствий. Среди таковых могут быть повышенная склонность к болезням, замедление роста или снижение механической прочности растительных волокон, и, как следствие, снижение агрономической ценности растений. Исследования показали, что ингибирование фермента аммонийфенилаланинлиазы приводит к появлению большого количества нежелательных фенотипов (Elkind и др., Abnormal Plant Development and Down-Regulation of Phenylpropanoid Biosynthesis in Transgenic Tobacco Containing a heterologous Phenytalanine Ammonia Liase Gene (Аномалии развития и замедление фенилпропаноидного биосинтеза в трансгенном табаке, содержащем гетерологический ген аммонийфенилаланинлиазы), Proc. Natf. Acad. Sci. USA, 87, стр.9057-9061, 1990). Фермент аммонийфенилаланинлиазы воздействует на первичный продукт обмена веществ фенилаланин, являющийся аминокислотой. Результаты этих экспериментов показывают, что изменение вторичных процессов обмена веществ путем модификации одного из первичных продуктов обмена веществ, участвующих в одном из вторичных механизмов обмена веществ, может привести к появлению непредсказуемых и нежелательных фенотипов. Следовательно, выбор биохимической стадии (стадий) в пределах одного из вторичных процессов обмена веществ имеет принципиальное значение для получения растений с нормальным набором фенотипов, но пониженным уровнем содержания одного из вторичных продуктов обмена веществ. Кроме того, использование антисенсибилизирующей РНК или ко-супрессии может оказаться недостаточным для того, чтобы обеспечить промышленно приемлемое снижение содержания одного из вторичных продуктов обмена веществ или приемлемую избирательность такого снижения по отношению к нужному продукту. Наконец, ингибирование генов, кодирующих основные функции ферментов, может оказать влияние и на степень экспрессии связанных с ними генов. Поэтому исследования, направленные на уменьшение содержания фенольных соединений, считающихся нежелательными с точки зрения питательных свойств, и на уменьшение содержания лигнина с помощью антисенсибилизирующей РНК или путем ко-супрессии без вредных побочных эффектов, не привели к сколько-нибудь заметному прогрессу в данной области.
Другим примером неудачной попытки изменить механизм обмена веществ стала модификация биосинтеза глюкозинолаты в рапсе (canola). Были опубликованы сведения о попытках изменить содержание глюкозинолаты в продуктах, приготовленных из рапса, путем управления механизмом биосинтеза глюкозинолаты. Одним из предложенных способов модификации биосинтеза глюкозинолаты было создание нового дополнительного биомеханизма, также потребляющего серу, участвующую в образовании глюкозинолат, или снижение уровня триптофана, участвующего в образовании глюкозинолат, путем его трансформации в триптамин (Engineering Altered Glucosinolate Biosynthesis by Two Alternative Strategies (Две альтернативные стратегии биосинтеза искусственно модифицированной глюкозинолаты), авт. Ibrahim, Chavadej, De Luca, в сб. Genetic Engineering of Plant Secondary Metabolism (Генетическая модификация вторичных процессов обмена веществ в растениях), 1994, Plenum Publishing Corporation, New York, USA).
Однако указанный способ оказался малоэффективным для уменьшения содержания глюкозинолаты в продуктах из семян рапса. Следовательно, этот способ непригоден для уменьшения негативного влияния глюкозинолат на питательные свойства рапса. Глюкозинолаты образуются в листьях растений и затем переходят в семена. Соответственно вышеуказанный метод был основан на предположении о том, что простым уменьшением доступного количества одного из первичных продуктов обмена веществ (серы и аминокислоты триптофана), участвующих в образовании глюкозинолат, можно уменьшить количество образующихся глюкозинолат. Однако первичные глюкозинолаты в семенах растений представляют собой алифатические глюкозинолаты, которые не используют аминокислоту триптофан для образования побочных цепочек. Кроме того, результаты этих экспериментов (например, Chavadej и др., Proc. Nad. Acad. Sci. USA, 91, стр.2166-2170, 1994) показали, что трансгенные растения, содержащие фермент, который может изменить содержание первичной аминокислоты триптофана, не содержали восстановленных глюкозинолат в семенах, а содержание алифатических глюкозинолат в их семенах соответствовало или даже превышало уровень глюкозинолат в нетрансгенных растениях. По этой причине общий объем глюкозинолат, образовывавшихся в семенах растений, не уменьшался даже несмотря на то, что восстанавливалась даже второстепенная компонента (индоловые глюкозинолаты). Генетические исследования показали, что растения с пониженным уровнем глюкозинолат можно выводить обычными методами селекции и что существует большое количество локусов, ответственных за низкое содержание глюкозинолат в крестоцветных. Биосинтез глюкозинолаты включает в себя большое количество биохимических превращений, и для создания способа снижения общего количества глюкозинолат необходимо влиять на те стадии этого процесса, которые являются общими для всех или большинства механизмов образования глюкозинолат. Следовательно, общий метод подавления образования глюкозинолат в крестоцветных должен учитывать возможное присутствие разных ферментов и субстратов, участвующих в биосинтезе глюкозинолат.
Оказалось, однако, что ферменты, использующиеся для достижения такой модификации биохимического процесса, также оказывали влияние на первичные продукты обмена веществ (аминокислоту триптофан и минерал серу), вследствие чего любое заметное изменение содержания этих соединений в клетке растения могло привести к отрицательному эффекту. Соответственно предложенный метод оказался неспособен оказывать выборочное воздействие на процесс вторичного обмена веществ. Естественно, изменение содержания триптофана вполне ожидаемо приведет к большому числу нежелательных эффектов. Таким образом, модификация процесса на уровне первичных продуктов обмена веществ не привела к желаемому эффекту снижения уровня глюкозинолаты в рапсе, что лишний раз подтверждает сложность модификации первичных процессов обмена веществ.
Таким образом, для изменения биохимического состава растительных тканей будет необходим общий метод изменения вторичных процессов обмена веществ. Такое изменение может включать в себя, например, снижение содержания соединений, нежелательных с точки зрения питательных свойств, изменение профиля вторичных продуктов обмена веществ, изменение технологических качеств растительных тканей, изменение уровня содержания соединений, представляющих интерес для промышленности или фармацевтики, выведение растений с измененными вкусовыми качествами, структурой тканей или внешним видом, получение растений с измененным составом вторичных продуктов обмена веществ, ответственных за привлечение насекомых, устойчивость растения к болезням или иные биологические процессы, зависящие от вторичных продуктов обмена веществ, или получение растений, показатели роста которых выгодно отличаются от соответствующих показателей обычных растений благодаря измененному составу вторичных продуктов обмена веществ.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение представляет собой способ оказания воздействия на образование вторичных продуктов обмена веществ. Данный способ включает в себя изменение доступного количества субстрата, который необходим для реализации определенного механизма вторичного обмена веществ и для образования конечного продукта обмена веществ, в частности соединений, образующихся за 1-5 стадий биохимической реакции до образования конечного продукта. Оказание воздействия на субстраты за несколько стадий до образования конечного продукта реакции позволяет избежать проблем, связанных с модификацией продуктов обмена веществ, которые участвуют также и в первичных процессах обмена веществ, поскольку данное воздействие имеет место уже после вступления субстрата в первичный процесс обмена веществ. Таким образом, данный способ представляет собой новое средство для целенаправленного уменьшения или изменения состава вторичных продуктов обмена веществ за счет идентификации тех продуктов предыдущих реакций, принимающих участие во вторичных процессах обмена веществ, которые не служат субстратами для первичных процессов обмена веществ.
Данный способ также может включать в себя изменение доступного количества субстрата в зависимости от конкретной растительной ткани, например, для того, чтобы изменить содержание определенного вещества только в семенах.
Таким образом, данный способ представляет собой новое средство для целенаправленного уменьшения или изменения состава вторичных продуктов обмена веществ за счет идентификации тех продуктов предыдущих реакций, принимающих участие во вторичных процессах обмена веществ, которые не служат субстратами для первичных процессов обмена веществ.
Один из вариантов осуществления изобретения представляет собой метод выведения генетически модифицированных растений, включающий в себя:
А) введение в клетку растения, пригодную для модификации и последующей регенерации, кассеты с последовательностью ДНК, которая содержит, помимо последовательности ДНК, необходимой для осуществления модификации и селекции в клетках растений, такую последовательность ДНК, которая осуществляет под контролем промотора, находящегося в растительных клетках, кодировку белка, способного модифицировать использование субстрата во вторичных процессах обмена веществ, причем данный субстрат не является первичным продуктом обмена веществ из группы, включающей в себя глюкозу, аминокислоты, жирные кислоты и нуклеотиды;
Б) регенерацию растения с измененным уровнем содержания, по крайней мере, одного из вторичных продуктов обмена веществ.
Другой вариант осуществления изобретения представляет собой способ выведения генетически модифицированных семян растений, включающий в себя выращивание растений, полученных в результате стадий А и Б вышеописанного способа, в условиях, благоприятных для развития семян.
Рекомбинантная ДНК вводится на хромосомном уровне в геном продуктивного растения таким образом, чтобы она могла передаваться последующим поколениям.
Дополнительные варианты осуществления изобретения представляют собой векторы, которые могут быть использованы для модификации растений, растения и семена, модифицированные в соответствии с вышеописанным способом, а также пищевые продукты на основе зерновых культур или производных продуктов.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг.1: Схематическое представление общей схемы способа изменения произвольного механизма вторичного обмена веществ.
Фиг.2: Схематическое представление общего фенилпропаноидного механизма обмена веществ и образования синапина.
Фиг.3: Начало и развитие синтеза синапина в развивающихся семенах. Тонкослойная хроматофафия семян рапса (Brassica napus сорт Westar).
Фиг.4: Количественный анализ накопления синапина в развивающихся семенах методом жидкостной хроматографии высокого давления.
Фиг.5: Определение способности развивающихся семян синтезировать синапин путем введения радиоактивного холина через плодоножки удаленных стручков с введением радиоактивных индикаторов в синапин спустя от 7 до 43 дней после опыления.
Фиг.6: Определение способности развивающихся семян синтезировать синапин и инфильтрация изолированных семян с помощью радиоактивного раствора холина с введением радиоактивных индикаторов в синапин спустя от 43 до 64 дней после опыления.
Фиг.7: Накопление синтезированного синапина в семядоле, стержнях завязи и оболочки семени растущих семян в процентах от общего количества меченого синапина в семени.
Фиг.8: Содержание синапина в семядоле и стержнях завязи развивающихся семян в пересчете на единицу массы образца ткани.
Фиг.9: Определение содержания синапина в семядоле и стержнях завязи развивающихся семян в расчете на одно семя, т.е. либо стержень, либо пару семядолей.
Фиг.10А и 10Б: Нуклеотидная последовательность открытой рамки считывания оксидазы холина (SEQ. №3).
Фиг.11: Выведенная последовательность аминокислот открытой рамки считывания оксидазы холина (SEQ. №4).
Фиг.12: Диаграмма вектора трансформации растения pHS 731, содержащего ген СОХ, подконтрольный тканеизбирательному промотору.
Фиг.13: Снижение содержания синапина в семенах рапса (Brassica sp.) путем повышения степени экспрессии гена СОХ.
Фиг.14: Диаграмма вектора трансформации растения pHS 981, содержащего ген BADH, подконтрольный тканеизбирательному промотору.
Фиг.15: Снижение содержания синапина в семенах Brassica sp. путем повышения степени экспрессии гена BADH.
Фиг.16: Изменение содержания фенолов в семенах Brassica sp. путем повышения степени экспрессии генов СОХ и BADH.
Фиг.17: Нуклеотидная последовательность гена декарбоксилазы феруловой кислоты синтетического В. pumulis, оптимизированного для экспрессии в растительных клетках (SEQ. №1).
Фиг.18: Выведенная последовательность аминокислот белка, закодированного открытой рамкой считывания декарбоксилазы феруловой кислоты синтетического В. pumulis (SEQ. №2).
Фиг.19: Рестрикционная карта вектора трансформации растения, содержащего ген декарбоксилазы феруловой кислоты, подконтрольный нерегулируемому промотору 35S.
Фиг.20: Рестрикционная карта вектора трансформации растения, содержащего ген декарбоксилазы феруловой кислоты, подконтрольный промотору напину селективному по отношению к семенам.
Фиг.21: Накопление фитиновой кислоты в процессе развития семян.
Фиг.22: Различия в накоплении фитиновой кислоты в различных тканях развивающихся семян.
Фиг.23: Процентное содержание усвоенного меченого мио-инозита во фракциях фитиновой кислоты, липидов, стенок клетки, растворимых в TFA, и отходов деления клеток.
Фиг.24: Последовательность амплифицированного фрагмента ДНК гена мио-инозит-оксиметилтрансферазы (SEQ. №5).
Фиг.25: Рестрикционная карта вектора pSIMTБ, содержащего кассету терминатора промотор 35S - IMT - GUS - Nos в pRD400 5.
Фиг.26: Рестрикционная карта вектора pNIMT, содержащего кассету терминатора промотор 35S - IMT - GUS - Nos в pRD400.
Фиг.27: PCR-анализ трансгенных растений, содержащих ген IMT.
Фиг.28: Northern blot-анализ растений с выраженным геном IMT.
Фиг.29: Гистограмма снижения содержания фитиновой кислоты в трансгенных растениях.
Фиг.30: Таблица, иллюстрирующая снижение содержания фитиновой кислоты в растениях F1, F2 и F3, выращенных в полевых условиях и содержащих вектор pSIMT.
Фиг.31: Таблица, иллюстрирующая снижение содержания фитиновой кислоты в растениях F1 и F2, выращенных в полевых условиях и содержащих вектор pNIMT.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано на избирательном снижении или изменении содержания предшественников вторичных продуктов обмена веществ и отличным от первичных продуктов обмена веществ. Указанным образом можно избежать возможного негативного влияния изменения первичных процессов обмена веществ при получении равноценного результата. Таким образом, настоящее изобретение позволяет избежать управления образованием соединений, которые могут относиться к первичным продуктам обмена веществ.
В предпочтительном варианте использования настоящего изобретения доступное количество субстрата изменяется с помощью фермента, например, являющегося гетерологическим по отношению к указанной растительной клетке и способного оказывать воздействие на указанный субстрат, а также удаляющего нужный субстрат из доступного набора субстратов или изменяющего доступное количество данного субстрата в данном наборе. Обобщенное представление о способе в соответствии с настоящим изобретением и соотношении между первичными и вторичными процессами обмена веществ проиллюстрировано на Фиг.1. Использование гетерологического фермента для изменения потока продуктов реакций в процессе вторичного обмена веществ приводит к изменению нужного продукта вторичного обмена веществ. При этом уровень содержания конечных продуктов, получаемых в результате некоторых стадий ферментной реакции в процессе вторичного обмена веществ, понижается, в то время как на других стадиях процесса продукт накапливается в значительных количествах, что ингибирует ферменты, ответственные за образование данного продукта. Такое ингибирование по принципу обратной связи, в свою очередь, влияет на образование конечного продукта всего вторичного процесса обмена веществ. Таким образом, изменение уровней содержания продуктов отдельных вторичных процессов обмена веществ и связанных с ними процессов достигается за счет введения новых ферментов, которые изменяют величину потока биохимических веществ (например, субстратов и продуктов реакций) в процессе реакции. Повышение степени экспрессии гетерологических ферментов в растительных тканях привело к метаболической трансформации предшественников процессов обмена веществ в вещества, которые накапливаются в растительных клетках, не вызывая нежелательных эффектов.
В некоторых вариантах осуществления изобретения в результате образуются ценные продукты или вещества, оказывающие благоприятное воздействие на растительную клетку.
В одном из вариантов осуществления изобретения нужный предшественник впоследствии модифицируется путем добавления нового фермента. В предпочтительном аспекте изобретения предполагается выбор фермента, активность которого гетерологична растительной клетке, в которой он проявляется. Например, таким ферментом может быть фермент, который в обычных условиях связан с вторичным процессом обмена веществ в тканевых растительных клетках. Используемый фермент может быть растительного, животного или же бактериального происхождения и модифицирован для достижения нужной степени экспрессии в растительной клетке. Выбранный гетерологический фермент может быть использован для модификации предшественника с целью изменения его количества, доступного для образования вторичного продукта обмена веществ. Использование фермента, являющегося гетерологическим по отношению к клетке, в которой он проявляется, позволяет избежать влияния биохимических регулирующих факторов и модифицировать вторичные процессы обмена веществ предсказуемым образом. Особый интерес в настоящем изобретении представляет модификация продуктов вторичного процесса обмена веществ, относящегося к сахарным спиртам, а также фенилпропаноидного вторичного процесса обмена веществ.
Дополнительным примером использования способа в соответствии с настоящим изобретением является изменение уровней содержания модифицированных сахаров, таких как глюкоза и нежелательные с точки зрения питательных свойств сукрозилглюкозиды (например, стахиоза и рафиноза). Галактоза переводится в галактинол, являющийся одним из предшественников вышеуказанных глюкозидов сукрозила, нежелательных с точки зрения питательных свойств. Такая промежуточная форма галактозы представляет собой конъюгированную модификацию UDP-галактозы. В методе в соответствии с настоящим изобретением образование и накопление UDP-галактозы (и ее последующее превращение в галактинол) предотвращаются с помощью фермента, являющегося гетерологическим по отношению к растительным клеткам, который может быть использован для изменения уровня содержания UDP-галактозы. Фермент UDP-галактоза-4-эпимераза (gaLE) участвует в одном из основных стадий галактозного обмена веществ в живых системах. Он является катализатором превращения UDP-галактозы в UDP-глюкозу. Ген данного фермента может быть получен из организма человека, дрожжей или бактерий. В настоящем изобретении предполагается использование бактериальной кодировки фермента.
В результате экспрессии данного гетерологического фермента в растительной клетке доступное для реакции количество UDP-галактозы снижается, и образуется полезное соединение - UDP-глюкоза. Авторы изобретения полностью уверены в том, что экспрессия фермента UDP-галактоза-4-эпимераза приведет к уменьшению количества биологически синтезируемого галактинола, который является одним из предшественников гликозидов сукрозы, нежелательных с точки зрения питательных свойств. В дополнение к снижению скорости накопления нежелательных гликозидов сукрозы действие нововведенного фермента приведет к увеличению доступного для реакции количества UDP-глюкозы с последующим образованием сукрозы. Предполагается, что сукроза будет участвовать и усиливать интенсивность других процессов обмена веществ, в которых она необходима либо как источник углерода для увеличения производительности растений (например, содержания белков, липидов, повышенной урожайности и т.п.), либо сама по себе.
В рамках настоящего изобретения могут быть реализованы и другие варианты использования данного способа. В частности, с его помощью можно изменять уровни содержания различных производных сахаров, таких как глюкоза-1-фосфат и глюкоза-6-фосфат, используя фермент фосфоглюкомутазу (pgm). Этот фермент является катализатором взаимопревращения глюкоза-1-фосфата и глюкоза-6-фосфата (G-1-P и G-6-P) в процессе биосинтеза и потребления сукрозы. Этот фермент играет основную роль в биосинтезе и потреблении сукрозы, крахмала и гликогена и присутствует во всех организмах. Ген этого фермента может быть получен из различных эукариотических и бактериальных источников (например, Agrobacterium).
G-6-P является основным исходным материалом для множества взаимопревращений между сахарами, в результате одного из которых образуется мио-инозит-1-Р, который является основным субстратом и одним из факторов образования соответственно фитиновой кислоты и нежелательных для питания сукрозилгликозидов. Экспрессия данного фермента, как ожидается, снизит уровень содержания G-6-P и, соответственно, уменьшит вышеуказанное негативное влияние на питательные свойства. Таким образом, в рамках настоящего изобретения могут быть осуществлены разнообразные модификации процессов обмена веществ.
Способ в соответствии с настоящим изобретением не ограничивается каким-либо конкретным вторичным процессом обмена веществ или каким-либо конкретным растением. Данный способ может быть использован для модификации вторичных процессов обмена веществ, которые являются общими для многих ценных культурных растений, в том числе однодольных и двудольных, но также может быть применен и к отдельному вторичному продукту обмена веществ, который важен только для одного конкретного вида.
Биохимической основой для способа в соответствии с настоящим изобретением является концепция регулирования активности фермента за счет манипуляции доступным количеством необходимого субстрата. В общем случае скорости ферментных реакций зависят от доступного количества субстрата. Иными словами, скорость образования продукта на основе фермента пропорциональна доступному количеству субстрата. Уменьшение концентрации субстрата приводит к снижению количества образующегося продукта. Кроме того, многие ферменты ингибируются конечным продуктом реакции, в результате чего избыток конечного продукта приводит к торможению ферментной реакции. Следовательно, изменение количества доступного субстрата или продукта ферментной реакции может быть использовано для регулирования общего количества вещества, образующегося в результате биохимической реакции.
Примерами вторичных процессов обмена веществ, которые могут быть изменены с использованием настоящего изобретения, являются изопреноидный биосинтез, алкалоидный биосинтез, терпеноидный биосинтез, фенольный биосинтез, сахарно-спиртовой биосинтез и прочие вторичные процессы обмена веществ, в результате которых образуются нежелательные для питания или промышленно ценные вещества. К вторичным продуктам обмена веществ, содержание которых может быть изменено с помощью настоящего изобретения, относятся нежелательные с точки зрения питательных свойств фенольные соединения, такие как синапин или глюкозинолаты в крестоцветных, продукты сахарного спирта, например фитиновая кислота или стахиоза и рафиноза, госсипол в хлопке, никотин, хлорогеновая кислота, конденсированные танины или иные нежелательные вторичные продукты обмена веществ. Хлорогеновая кислота присутствует в сое, хлопке, подсолнечнике и является производной от кофеиновой кислоты, которая образуется в фенилпропаноидном процессе обмена веществ. К другим соединениям, нежелательным для питательных свойств растений, относятся сапонины, которые присутствуют во многих растениях, включая люцерну. Сапонины представляют собой высокомолекулярные гликозиды, состоящие из сахарной компоненты, связанной с тритерпеном или стероидным агликоном. Известно, по крайней мере, три класса сапонинов, а именно гликозиды тритерпена, стероидные гликозиды и стероидные алкалоидные гликозиды. Одним из исходных материалов для биосинтеза сапонинов в растениях является сквален. Другие важные классы вторичных продуктов обмена веществ, таких как фитостерины, карденолиды, кукурбитацины, квассиноиды и лимониды, также являются производными сквалена. Избыток сапонинов в рационе питания животных связан с эффектом, известным как бомбаж.
Настоящее изобретение иллюстрируется на примере использования данного способа во многих не связанных вторичных процессах обмена веществ. Однако, поскольку для специалистов будет очевидна возможность использования данного способа для модификации любого конкретного вторичного процесса обмена веществ, ниже представлен только общий способ осуществления настоящего изобретения.
Один из вариантов осуществления настоящего изобретения представляет собой способы и композиции ДНК, служащие для изменения содержания синапина и связанных с ним фенольных соединений в растениях. Другой вариант осуществления настоящего изобретения представляет собой растительные клетки с модифицированным содержанием фенолов и семена растений с пониженным содержанием фенолов, в частности крестоцветные растения с пониженным содержанием синапина. Еще один вариант осуществления настоящего изобретения представляет собой способы и композиции ДНК, служащие для изменения содержания в растительных клетках фитиновой кислоты, которая образуется в результате сахарно-спиртового вторичного процесса обмена веществ. Другой вариант осуществления настоящего изобретения представляет собой растительные клетки с модифицированным содержанием фитиновой кислоты и семена растений с пониженным содержанием фитиновой кислоты. Другой вариант осуществления настоящего изобретения представляет собой семена растений с пониженным содержанием фитиновой кислоты, пригодным для кормовых целей, семена растений с пониженным содержанием фитиновой кислоты, подходящим для приготовления переработанных продуктов, а также растительные клетки с модифицированным сахарно-спиртовым (инозитным) процессом обмена веществ.
Каждый вторичный процесс обмена веществ в растениях связан с ограниченным количеством ферментов и субстратов для этих ферментов, которые характерны только для него или используются характерным только для него образом. В результате вторичного процесса обмена веществ могут образовываться различные соединения, которые потребляются или присутствуют во всех тканях растения. Следовательно, для осуществления способа, позволяющего модифицировать вторичные процессы обмена веществ, используются уникальные ферменты, которые выборочно изменяют поток компонент реакций в процессе обмена веществ. В качестве примера осуществления способа в соответствии с настоящим изобретением были модифицированы два разных вторичных механизма обмена веществ. Ниже приведена информация о данных механизмах, позволяющая понять сущность способа в соответствии с настоящим изобретением.
(А) ВТОРИЧНЫЙ ФЕНИЛПРОПАНОИДНЫЙ МЕХАНИЗМ ОБМЕНА ВЕЩЕСТВ
В результате процессов вторичного фенилпропаноидного обмена веществ в растениях образуются различные фенольные соединения. Фенилпропаноидный обмен веществ включен во многие физиологические процессы, происходящие в растениях, в том числе ответственные за сопротивляемость заболеваниям, защиту от ультрафиолетового излучения и регулирование роста растений. Продукты этого процесса необходимы для биосинтеза суберина и лигнина, которые входят в состав растительных тканей и участвуют в образовании растительных волокон, что приводит к неполному усвоению углеводов в составе зерновых культур или продуктов на их основе. Считается, что лигнин участвует в образовании нерастворимых волокон, вследствие чего высокое содержание лигнина в зерне или зерновых продуктах приводит к неполному усвоению такой пищи, в особенности у нежвачных животных. Поэтому растительные фенолы, в частности фенольные предшественники лигнина, могут рассматриваться как факторы, неблагоприятные с точки зрения питательных свойств корма животных, и снижение содержания или соответствующее изменение растительных фенолов, в частности фенолов, участвующих в образовании лигнина, позволит получать высококачественные корма.
Растительные фенолы также участвуют в образовании других соединений, многие из которых также нежелательны с точки зрения питательных свойств.
Настоящее изобретение включает в себя способ уменьшения содержания фенольных соединений, которые считаются нежелательными с точки зрения питательных свойств, в растительных клетках, семенах и кормах. В частности, настоящее изобретение предполагает снижение уровня содержания горького и нежелательного с точки зрения питательных свойств соединения синапина (или синапоилхолина), образующегося в растительных клетках. Считается также, что синапин образует комплексы с белками в кормах, что снижает количество белка, усвоенного животными. Кроме того, горький вкус может также повлиять на качество корма. Наконец, при включении синапиносодержащих кормов в рацион кур, несущих яйца с коричневой скорлупой, синапин приводит к появлению у этих яиц неприятного запаха, напоминающего запах рыбы. Соотношение синапиносодержащих кормов в рационе таких пород кур должно быть не более 10%, что ограничивает использование синапиносодержащих кормов в рационе этих кур. Следовательно, уменьшение содержания синапина в семенах крестоцветных и производных от них кормов является важной задачей.
Синапин образуется в растениях в результате пролонгированного фенилпропаноидного процесса обмена веществ. Механизм процесса, определенный в рамках настоящего изобретения, включает в себя стадии биохимических реакций, приводящие к образованию синапиновой кислоты, а также биохимические механизмы, ответвляющиеся от различных стадий реакции, таких как механизмы, приводящие к образованию мономеров лигнина и других биохимических веществ, являющихся производными от фенилпропаноидного процесса обмена веществ.
В фенилпропаноидном процессе обмена веществ ароматическая аминокислота L-фенилаланин выступает в роли субстрата для фермента аммонийфенилаланинлиазы (PAL). Под влиянием фермента PAL образуется коричная (В-фенилакриловая) кислота, которая является субстратом для циннамат-4-гидроксилазы (С4Н). Продуктом С4Н является р-кумаровая кислота, которая является предшественником многих флаваноидных соединений, некоторые из которых могут также образовываться при воздействии на L-тирозин ферментом аммонийтирозинлиазы (TAL). Кумаровая кислота может также служить предшественником лигнина. При воздействии на р-кумаровую кислоту ферментом р-кумарат-3-гидроксилаза (С3Н) образуется кофеиновая кислота. Кофеиновая кислота в процессе обмена веществ переводится кофеат/5-гидроксиферулат-3-гидроксилазой (ОМТ) в феруловую кислоту. Феруловая кислота представляет собой один из трех известных первичных фенольных мономеров, участвующих в биосинтезе лигнина. Феруловая кислота также может выступать в роли субстрата для фермента ферулат-5-гидроксилазы (F5H) с образованием 5-гидроксиферуловой кислоты, которая может быть впоследствии модифицирована ферментом ОМТ с образованием синапиновой кислоты. Синапиновая кислота является одним из основных фенольных лигниновых мономеров. Также эта кислота может быть конъюгирована ферментом UDP-глюкоза-синапоилтрансферазой (SGT) с образованием синапоилглюкозы. Синапоилглюкоза служит в качестве субстрата для синапоилглюкоза-синапоилхолинтрансферазы (SCT) с образованием синапоилхолина или синапина. Две последних стадии преобладают в крестоцветных, а синапин, накапливающийся в семенах рапсовых, является основным неполимерным фенольным соединением, присутствующим в зрелых семенах. Обобщенный механизм фенилпропаноидного обмена веществ представлен на Фиг.2. Следует отметить, что в некоторых видах растений в фенилпропаноидном обмене веществ могут принимать участие дополнительные ферменты.
Синапин (синапоилхолин) является наиболее распространенным фенольным соединением, содержащимся в семенах крестоцветных, причем в В. napus он может достигать уровня 4% (Blair, Reichert, 1984, J. Sci. Food Agric., 35, стр.29). Биосинтез синапина осуществляется в незрелых семенах, которые еще имеют светло-зеленую окраску, но при созревании семян уменьшения его уровня не наблюдается (Vogt и др., 1993, Arch. Biochem. Biophys., 300, стр.622). При развитии рассады (прорастающих семян) уровень содержания синапина снижается в результате реакции эстеразы, продуктами которой являются синапиновая кислота и холин. Есть неподтвержденные предположения о том, что снижение уровня синапина может обеспечивать холин для фосфолипидного синтеза в процессе прорастания семян (Strack и др., 1981, Z. Naturforsch., 36 с., стр.215). Однако тот факт, что многие некрестоцветные не содержат синапин в семенах, позволяет подвергнуть сомнению то, что синапин является важным соединением для развития или прорастания семян вообще.
Из образцов Arabidopsis thaliana (Chaple и др., 1992, The Plant Cell, 4, стр.1413-1424) были выделены мутанты с дефектами общего механизма фенилпропаноидного обмена веществ. Несмотря на свой исключительный интерес для физиолгических, биохимических и генетических исследований, эти мутировавшие растения не имеют сельскохозяйственной ценности вследствие негативных последствий мутации, в частности повышенной чувствительностью к ультрафиолетовому излучению, объясняющейся экспрессией мутировавшего фенотипа во всем растении, в особенности в листьях. Эти мутанты, обозначаемые SIN1 (мутанты биосинтеза синапоилмалата), демонстрируют замедленный синтез эфиров синапиновой кислоты, уменьшая содержание синапина (синапоилхолинового эфира), вследствие чего изменяется мономерный состав лигнина в растениях. Изменение содержания лигнина может привести к развитию растений с уникальным составом лигнина и, следовательно, иным строением растительных волокон.
Далее Chapple и соавторы в своей статье касаются возможности использования мутировавших растений наподобие SIN1 для снижения или изменения уровня лигнина в семенах важных для сельского хозяйства крестоцветных, таких как семена рапса, но не указывают пути, по которому это может быть осуществлено. Далее в статье делается утверждение, что снижение уровня синапина в семенах рапса является важной задачей, однако также не указывается способа, с помощью которого это можно осуществить на основе мутации SIN1. При очевидной повышенной чувствительности SIN1 к ультрафиолетовому излучению такая мутация не может дать значительного положительного результата для сельского хозяйства. Тем не менее, несмотря на то что в литературе не описано способов снижения уровня синапина в семенах, мутация SIN1 является важным научным доказательством того, что синапин не является важным соединением для роста и развития растений и что семена с пониженным уровнем синапина могут расти и развиваться. Однако мутация SIN1 не дает достаточной основы для разработки способа снижения уровня содержания синапина в культивируемых крестоцветных без сопутствующего вредного эффекта повышенной чувствительности к ультрафиолетовому излучению.
Помимо фенольных соединений, нежелательных с точки зрения питательных свойств растений, таких как синапин, многие продукты фенилпропаноидного обмена веществ участвуют в образовании лигнина. Биосинтез лигнина является составной частью общего процесса фенилпропаноидного биосинтеза, в результате которого образуются, по крайней мере, три первичных фенольных промежуточных продукта, а именно кумаровая, феруловая и синапиновая кислоты, продукты которых при полимеризации образуют лигнин и другие фенольные соединения.
Биохимия образования лигнинов из этих промежуточных продуктов чрезвычайно сложна и включает в себя большое количество других ферментов, таких как кофеиновая кислота / оксиметилтрансфераза 5-гидроксиферуловой кислоты (СОМТ), кофеоил-СоА-редуктаза (ССоАОМТ), дегидрогеназа циннамилового спирта (CAD), циннамоил-СоА-редуктаза (CCR), пероксидаза, 4-кумарат-СоА-лигаза (4CL) и бета-глюкуронидаза, характерная для кониферина (CBG). В процессе могут принимать участие и другие ферменты, поэтому полностью биохимия образования лигнина не ясна.
Лигнин представляет собой сложный полимер, состоящий в основном из соединенных между собой участков указанных мономерных фенольных соединений в различных пропорциях и с различными связями между ними в зависимости от видов растительных клеток и растений. Лигнины являются важными компонентами стенок растительных клеток, ткани которых должны иметь высокую механическую прочность или обеспечивать пропускание воды. Кроме того, считается, что лигнины участвуют в механизмах сопротивления болезням. Общим для двудольных является гваяцил-сирингиловый лигнин, состоящий как из гваяциловых участков, являющихся производными феруловой кислоты, так и из сирингиловых остатков, являющихся производными синапиновой кислоты. Соответственно, для образования лигнина произвольного типа необходимы и синапиновая, и феруловая кислоты. В частности, способность древесины к разложению или переработке (например, при получении целлюлозы), по-видимому, сильно зависит от мономерного состава лигнина, которая, в свою очередь, определяется в основном доступным для реакции количеством определенных мономеров лигнина. Считается, что присутствие 5-оксиметиловых групп в сирингиловых лигнинах, образующихся из синапоила, уменьшает сшивание полимера, поскольку лигнин, образованный в основном из сирингиловых участков, легче поддается переработке, чем лигнин, образованный в основном из гваяциловых участков, являющихся производными феруловой кислоты. Следовательно, способы повышения уровня сирингилового лигнина позволят получать корма с повышенной усваиваемостью вследствие более слабого сшивания лигнина.
Усваиваемость фуражных зерновых культур зависит от содержания волокон, потому что сшитые лигнины более устойчивы к разложению и способствуют физическому связыванию компонент стенок клеток.
Из вышесказанного следует, что попытки улучшить свойства зерновых кормов путем понижения уровня или удаления синапина или изменения общего фенольного состава должны быть направлены на семена в процессе их развития. Для изменения биохимического механизма образования синапина именно в семенах растений наиболее приемлем молекулярно-генетический подход вследствие отсутствия зародышевой плазмы, которая обеспечивает характерно низкий уровень синапина в естественных условиях.
Один из вариантов осуществления настоящего изобретения представляет собой способ изменения уровня содержания фенольных соединений в семенах и уровней содержания синапиновой кислоты и связанных с ней фенольных соединений. Данный способ основан на введении новых ферментов, которые изменяют количество доступных соединений, выступающих в роли предшественников и субстратов для ферментов фенилпропаноидного процесса обмена веществ. Снижение содержания или изменение доступного количества этих соединений приводит к смещению нормального биохимического процесса фенилпропаноидного обмена веществ и изменению уровней содержания образующихся фенилпропаноидных соединений.
В качестве примера осуществления данного способа можно привести использование нового фермента холиноксидазы (СОХ) для уменьшения запаса холина в растительных клетках, в частности в семенах. Помимо всего прочего, холин участвует в образовании синапина из синапоилглюкозы. Уменьшение запасов холина приводит к изменению величины запасов предшественников синапина (холина и синапоилглюкозы) и, следовательно, изменяет состав промежуточных продуктов, образованных на более ранних стадиях ферментной реакции фенилпропаноидного процесса обмена веществ, включая синапиновую кислоту. В результате в семенах значительно снижается содержание синапина и изменяется содержание фенолов. Известно, что окисление холина холиноксидазой приводит к образованию перекиси водорода. Образование перекиси водорода в растительных клетках считается благоприятным фактором, что является дополнительным доводом в пользу увеличения экспрессии холиноксидазы в растениях. В качестве иллюстрации дополнительного аспекта способа в соответствии с настоящим изобретением может быть приведено использование второго фермента, бетаинальдегиддегидрогеназы (BADH), для ускорения превращения продукта холиноксидазы - бетаинальдегида - в глицинбетаин, который выполняет функции защиты от стресса. Соединение бетаин полезно во многих областях пищевой промышленности, а также может быть использовано в качестве добавки, ускоряющей рост растений.
Следовательно, положительный эффект изменения содержания фенолов в растительных клетках и селективного снижения содержания синапина проявляется за счет снижения содержания одного только промежуточного продукта фенилпропаноидного обмена веществ. Для специалистов в данной области будет понятно, что другие ферменты также могут быть использованы в рамках данного способа для изменения содержания других конечных продуктов фенилпропаноидного обмена веществ.
Другим примером в рамках настоящего изобретения является использование новых ферментов для подавления феруловой кислоты, которая также образуется в результате фенилпропаноидного обмена веществ. Из трех основных мономеров лигнина феруловая кислота считается ответственной за придание структурной жесткости и прочности стенкам клеток за счет сшивания пентозных цепочек, арабиноксиланов и гемицеллюлоз, в результате чего стенки клеток становятся менее чувствительны к разложению ферментами. Таким образом, феруловая кислота как компонента лигнина играет важную роль для механической прочности растительных тканей. Изменение уровня содержания феруловой кислоты особым образом может принести ряд преимуществ.
Феруловая кислота синтезируется в рамках общего фенилпропаноидного процесса, как описано выше, и служит предшественником различных веществ, образующихся в этом процессе (обзоры даны: J.P.N. Rosazza и др., Journal of Industrial Microbiology, 15, стр.457-471, 1995; R. Whetten, R. Sederoff, Plant Cell, 7, 1001-1013,1995; R.A. Dixon, N.L. Paiva, Plant Cell, 7, 1085-1097, 1995).
Способ в соответствии с настоящим изобретением предоставляет возможность модулирования уровней содержания феруловой кислоты с целью изменения количества других соединений, образующихся в результате реакции фенилпропаноидного обмена веществ, в частности синапина. Один из аспектов настоящего изобретения представляет собой способ изменения уровня содержания феруловой кислоты в растительных клетках. Данный способ основан на введении гетерологических ферментов, которые приводят к усвоению феруловой кислоты. Некоторые из этих ферментов могут также приводить к образованию промышленно важных соединений. Например, используется фермент декарбоксилаза феруловой кислоты, которая приводит к образованию соединения винилгваякол, которое может быть использовано в качестве промышленного сырья и может накапливаться в растительных клетках, не вызывая нежелательных эффектов. Снижение содержания или изменение количества доступной феруловой кислоты приводит к смещению нормального биохимического процесса фенилпропаноидного обмена веществ и изменению уровней образующихся различных фенилпропаноидных соединений.
Способ в соответствии с настоящим изобретением не ограничивается фенилпропаноидным обменом веществ. В качестве примера использования настоящего изобретения можно привести модификацию вторичного процесса обмена веществ, в результате которого образуются сахарные спирты.
(Б) ВТОРИЧНЫЙ САХАРНО-СПИРТОВОЙ МЕХАНИЗМ ОБМЕНА ВЕЩЕСТВ
Настоящее изобретение также включает в себя способы и композиции ДНК для изменения содержания сахарных спиртов в растительных клетках. В результате применения способа в соответствии с настоящим изобретением в растительных клетках изменялись содержания соединений, являющихся производными сахарных спиртов, в частности фитиновой кислоты, стахиозы, рафинозы, сукрозилгликозидов, уронидов, пентоз, фосфоинозитидов и гликофосфокерамидов. В некоторых вариантах осуществления настоящего изобретения были получены семена растений с пониженным содержанием фитиновой кислоты. Настоящее изобретение также предоставляет возможность получения семян растений с пониженным содержанием фитиновой кислоты, делающим их пригодными для использования в кормовых целях, семян растений с пониженным содержанием фитиновой кислоты, делающим их пригодными для приготовления модифицированных кормов, а также растительных клеток с модифицированным инозитным обменом веществ. Другие соединения, являющиеся производными процесса сахарно-спиртового обмена веществ, также изменяются в результате использования настоящего изобретения, причем среди этих соединений находятся вредные с точки зрения питательных свойств производные сахарного спирта, такие как стахиоза и рафиноза. Особую важность для кормовых целей представляют семена растений с модифицированным содержанием фитатов.
Фитаты являются важными компонентами состава многих семян, составляя обычно 2-4% массы семени, а в некоторых случаях достигая 10%. Наличие высокого содержания фитатов в рационе связывалось с потерей аппетита, снижением приплода и другими негативными факторами. Эти эффекты, вероятнее всего, связаны со способностью фитатов связывать цинк. Комплексы фитиновой кислоты с другими компонентами состава семян называются фитином. Высокое содержание фитина также связывается с негативными эффектами.
В дополнение к негативному влиянию фитиновой кислоты на эффективность кормов ее присутствие в составе кормов также приводит к ряду негативных последствий для экологии. Фосфор, связанный с фитиновой кислотой, обычно не усваивается нежвачными животными, вследствие чего его приходится добавлять в рацион животных, увеличивая тем самым расходы на их содержание. Фосфор, связанный с фитиновой кислотой, выводится из организма нежвачных животных с испражнениями, и последующее разложение фекалий бактериями приводит к попаданию этого фосфора в окружающую среду. Высокое содержание фитиновой кислоты в зерновых кормах приводит к выделению большого количества фосфора с экскрементами. Постоянно увеличивающееся поголовье скота зачастую приводит к эвтрофикации водных ресурсов и другим экологическим проблемам, связанным с загрязнением фосфором. Экологи прогнозируют рост масштабов этих нежелательных последствий, что может в будущем стать основным фактором, сдерживающим увеличение поголовья скота. Таким образом, способы снижения уровня содержания фитиновой кислоты в экскрементах могут стать ключом к решению этих экологических проблем.
Несмотря на то что реальные расходы, связанные с добавлением фосфора в рацион животных, составляют лишь незначительную долю общих расходов, экологические последствия выделения фосфора с экскрементами требуют дополнительных расходов на их устранение, которых можно избежать за счет снижения содержания фитатов в кормах.
Наличие бактериальной фауны в рубце жвачных животных может привести к выделению фосфора, содержащегося в фитиновой кислоте, и соответственно увеличению его доступного количества. В результате этого количество фосфора, которое необходимо добавлять в рацион жвачных животных, значительно меньше, чем у нежвачных животных. Однако количество фитиновой кислоты в семенах большинства растений выше реально необходимого, поэтому даже экскременты жвачных животных представляют собой серьезную угрозу для экологии.
На сегодняшний день наиболее распространенные способы снижения содержания фитатов в растительных клетках в основном направлены на разложение фитатов с помощью ферментов фитазы, содержащихся в кормах. Несмотря на то что такие способы приводят к увеличению доступного количества фосфора, они не позволяют получать зерна или корма с пониженным содержанием фитиновой кислоты, а лишь корма, в которых общее количество доступного фосфора, содержащегося в фитиновой кислоте, выше. Следовательно, использование фермента фитазы целесообразно только в плане увеличения доступности фосфора в фитиновой кислоте.
Фермент фитаза обычно встречается в микроорганизмах, таких как плесень (Aspergillis), бактерии (Bacillus, Pseudomonas), а также дрожжи (Saccharomyces). Удаление фитатов путем обработки растительных материалов смесями ферментов, содержащих микробную фитазу, описано, например, в патенте США №5554399. Большинство бактерий синтезируют целый ряд ферментов фитазы, в частности сам фермент фитазу, а также различные фосфатазы, которые впоследствии разлагают фитиновую кислоту. Полнота высвобождения доступного фосфора из фитиновой кислоты может зависеть от наличия большого количества различных ферментов.
Альтернативным подходом к модификации растений с целью получения микробной фитазы является способ, описанный в патенте США 5593963 и состоящий в повышении степени экспрессии фермента фитазы в трансгенных растениях под контролем регулирующих последовательностей мономеров, среди которых есть такие, которые могут управлять степенью экспрессии либо нерегулируемым образом, либо в зависимости от стадии процесса или от ткани, в которой он происходит. Можно получать растительные клетки, в частности клетки семян растений, содержащие фермент фитазу, способную высвободить часть фосфора, содержащегося в фитиновой кислоте.
Однако простое высвобождение фосфора из фитиновой кислоты не может полностью решить проблему снижения содержания фосфора в отходах и влияния фитиновой кислоты, потенциально вредного с точки зрения питательных свойств кормов. Содержание фитиновой кислоты в форме комплексов или фитатов не снижается, что представляет собой более серьезную проблему. В литературе приводятся средства высвобождения фосфора из фитиновой кислоты, но не указывается способов контроля над уровнем фитиновой кислоты, образующейся в результате обмена веществ.
Исходя из вышесказанного, удобный способ, который позволил бы контролировать уровень содержания фитиновой кислоты, нашел бы широкое применение в производстве сельскохозяйственных кормов. Например, семена растений с пониженным содержанием фитиновой кислоты могли бы стать основой кормов с повышенным количеством доступных минеральных веществ. Семена растений с пониженным содержанием фитиновой кислоты также могли бы использоваться в качестве кормов с низким содержанием фитина и, следовательно, более питательных и легче усваиваемых за счет снижения количества комплексов фитиновой кислоты.
Помимо своей повышенной питательной ценности, корма с пониженным содержанием фитиновой кислоты привели бы к снижению выделения фосфора в окружающую среду и, следовательно, меньшему загрязнению последней. Несмотря на то что способы высвобождения фосфора из фитиновой кислоты позволят снизить уровень фосфора в экскрементах и необходимое количество фосфора, добавляемого в корм, реальный уровень фитиновой кислоты и, следовательно, количество потенциально доступного для усвоения фосфора превышают требования к содержанию фосфора в кормах для многих животных.
Следовательно, способы управления содержанием фитиновой кислоты могут найти применение в производстве кормов для скота, а также дадут возможность получить новые ценные корма и кормовые смеси.
Из вышесказанного становится очевидным, что способ регулирования количества фитатов, образующихся в процессе развития семян, был бы полезен и позволил бы решить проблемы, связанные с загрязнением окружающей среды фосфором и отрицательным влиянием фитиновой кислоты на питательные свойства кормов. Кроме того, был бы особенно полезен такой генетический механизм, который был бы применим к широкому разнообразию растений. Настоящее изобретение позволяет решить все эти проблемы.
Для оценки области применения настоящего изобретения необходимо понимание биологического механизма, ответственного за образование фитиновой кислоты. Несмотря на то что механизм биосинтеза фитиновой кислоты в настоящее время ясен не полностью, его многие ключевые моменты известны. Фитиновая кислота является гексафосфатной производной мио-инозита. В биохимическом процессе синтеза фитиновой кислоты мио-инозит, являющийся сахарным спиртом, участвует исключительно в качестве исходного субстрата. Это соединение мио-инозит, также часто называемое инозитом, также играет центральную роль в образовании других производных мио-инозита и эпимеров. Некоторые из этих производных, такие как сукрозилгликозиды, также оказывают негативное влияние на питательные свойства кормов, и, следовательно, понижение содержания этих соединений в растениях желательно.
Мио-инозит представляет собой сахарный спирт, присутствующий в растительных клетках в больших количествах. Однако простая изоляция одной из гидроксильных групп мио-инозита делает его непригодным для участия в биосинтезе фитиновой кислоты и других процессах обмена веществ. Изоляция мио-инозита может быть осуществлено in vivo путем метилирования различными метилтрансферазами в определенных точках. Например, мио-инозит можно перевести в ононит путем монометилирования в точке 6. Метилирование в точке 5 приводит к образованию секвоита, который переводится путем эпимеризации в пинит. Последний не может использоваться в биосинтезе фитиновой кислоты. Метилированные производные мио-инозита, как известно, придают растениям полезные качества, в частности устойчивость к стрессу, и участвуют в переносе растворенных веществ и стабилизации мембранных белков. Таким образом, настоящее изобретение предполагает модифицирование инозита с помощью гетерологических ферментов, обычно не участвующих в сахарно-спиртовом обмене веществ.
Биохимия образования фитатов до сих пор не полностью ясна, но, тем не менее, известно по крайней мере два возможных механизма этого явления и большое количество участвующих в нем различных ферментов. Фитаты присутствуют в большинстве растительных тканей, но их присутствие особенно заметно в семенах и пыльце, которая играет очевидно важную роль в накоплении фосфора. В связи с этим оказалось достаточно проблематичным разработать способ модифицирования процесса биосинтеза фитатов обычными способами, что главным образом объясняется как раз неполным пониманием биохимии этого процесса.
Для оптимизации процесса образования фитиновой кислоты настоящее изобретение предусматривает способы и составы ДНК, которые кодируют фермент(ы), модифицирующие мио-инозит, не позволяя таким образом ему участвовать в биосинтезе фитиновой кислоты. Настоящее изобретение предполагает использование гетерологического гена метилтрансферазы для избирательного метилирования мио-инозита в семенах, в особенности в тех тканях семян, которые ответственны за биосинтез фитиновой кислоты. (Под термином гетерологический подразумевается, что данный фермент в нормальных условиях не связан с биосинтезом фитатов в указанной растительной клетке.) Настоящее изобретение использует гетерологический фермент, полученный из галофитового растения и не встречающийся в обычных культурах наподобие зерновых, соевых, хлопке, люцерне, пшенице, ячмене, ржи, сорго, подсолнечнике, масличных рапсовых (Brassica) и других культивируемых растениях.
Образование метилинозита из мио-инозита также дает дополнительную выгоду, поскольку образуется безвредное соединение, не оказывающее отрицательного воздействия на растительную клетку. Таким образом, образование фитиновой кислоты тормозится за счет снижения доступных запасов мио-инозита.
Фитаты являются одним из основных негативных факторов, влияющих на питательные свойства зерновых кормов. Эти отрицательные эффекты включают в себя связывание минералов, образование комплексов с белком и т.д., но в особенности это касается выведения избыточного фосфора с экскрементами в виде фитиновой кислоты, которая перерабатывается в окружающей среде и приводит к загрязнению последней фосфором. Следовательно, способы снижения содержания фитиновой кислоты в растительных клетках важны в кормовой промышленности. В рыбоводстве высокий уровень содержания фитиновой кислоты также представляет серьезную проблему, поскольку растительный белок используется в качестве корма для сельди. В связи с этим способы снижения содержания фитиновой кислоты в растительных клетках, в особенности клетках тех растительных тканей, которые используются в качестве кормов, были бы желательны и нашли бы широкое применение. Способы снижения содержания фитиновой кислоты в растительных клетках широкого круга растений, используемых в качестве кормов, были бы особенно ценны для кормовой промышленности.
Растительный ген, кодирующий мио-инозит-оксиметил трансферазу (IMT), был выделен из ледяника (Mesembryanthemum crystallinum) и успешно использован для перевода мио-инозита в пинит в гетерологических трансгенных растениях (патент США №5563324). Этот растительный ген был введен в клетку под контролем нерегулируемого промотора с целью увеличения устойчивости растений к стрессу за счет избыточного образования метилированной производной мио-инозита - ононита, который впоследствии может быть эпимеризован в пинит. В настоящем изобретении растительный ген вводится в клетку под контролем промотора, избирательного по отношению к тканям семян, для изменения инозитного процесса обмена веществ в семенах.
Процесс получения пинита, описанный в Патенте США №5563324, придавал растениям устойчивость к соли при нерегулируемой экспрессии этого гена в ряде трансгенных табачных культур. Аналогично, нерегулируемая экспрессия бактериальной маннит-1-Р-дегидрогеназы в растительных клетках ускоряла образование маннита, сахарного спирта, или полиола, а также повышала устойчивость растений к соляному стрессу. Следовательно, Патент США №5563324 описывает получение сахарных спиртов с помощью фермента, который способен образовывать его из сахаров, характерных для растительных клеток, как способ придания растениям устойчивости к соляному стрессу.
Однако Патент США №5563324 не дает никаких рекомендаций по поводу использования гена, способного модифицировать мио-инозит для предотвращения его участия в качестве предшественника других соединений, среди которых есть и производные, свойственные для клеток. Тем не менее, Патент США №5563324 дает убедительное доказательство того, что модифицирование мио-инозита не приводит ни к каким негативным последствиям для растений, и, следовательно, регулирование уровня содержания мио-инозита не должно оказывать на них вредного воздействия.
Соответственно, модифицирование мио-инозита геном метилтрансферазы не рассматривается как возможный вредный фактор для растительных клеток, и метилированный продукт этой ферментной реакции известен как безвредный для растительных клеток.
Однако мио-инозит является основной компонентой для многих аспектов роста и развития растений. Помимо своей роли как предшественника фитиновой кислоты, мио-инозит также участвует в биосинтезе уронида и пентозы, присутствует в фосфоинозитидах в мембранах растительных клеток и в других сложных растительных липидах, включая гликофосфокерамиды. Кроме того, он является предшественником многих других естественных изомеров инозита, многие из которых, равно как и сам мио-инозит, распределяются как метиловые эфиры индивидуальным образом в различных растениях.
Роль мио-ниозита в общем обмене веществ растений велика, и модифицирование его запасов могло бы иметь непредсказуемые последствия для сельского хозяйства. Хотя Патент США №5563324 описывает нерегулируемую экспрессию гена метилтрансферазы, он не приводит никаких доказательств того, что полученные в результате этого растения будут иметь сельскохозяйственную ценность. В Патенте США №5563324 делается следующее утверждение: «Даже если нововведенные гены не приведут к повышению производительности растений в сельскохозяйственных условиях, трансгенные растения, содержащие такие гены, будут полезны для исследовательских целей, в частности для изучения того, как изменение внутренних процессов, происходящих в растениях (например, осмотического регулирования), влияют на производительность растений в полевых условиях.» (Колонка 2, строки 60-65.) Следовательно, Патент США №5563324 не предусматривает регулирование уровня содержания мио-инозита для понижения содержания фитиновой кислоты и не предполагает получение таким образом растений, имеющих полезные агротехнические свойства и, следовательно, значимых для сельского хозяйства. Поэтому Патент США №5563324 описывает экспрессию гена, модифицирующего мио-инозит, во всех растениях только с целью повышения устойчивости растений к стрессу или для дальнейших научных исследований.
В соответствии с этим авторы полностью уверены в том, что настоящее изобретение содержит описание нового подхода к снижению уровня фитиновой кислоты. Использование гена метилтрансферазы позволяет селективно снижать уровень содержания фитиновой кислоты. Настоящее изобретение не основывается на уже существующих методах, таких как повышение экспрессии ферментов фитазы, которые не учитывают проблему избытка фитиновой кислоты в зернах или зерновых кормах. В ходе работы, проведенной в рамках настоящего изобретения, было обнаружено, что уровни содержания фитиновой кислоты можно регулировать путем изменения уровней мио-инозита, доступного для осуществления биосинтеза фитиновой кислоты. Было также показано, что экспрессия гена метилтрансферазы в тканях семян приводит к снижению содержания фитиновой кислоты без каких-либо видимых изменений в остальных характеристиках растения. Очевидно, что ограничение экспрессии в тканях семян гена метилтрансферазы, ответственного за биосинтез фитатов, резко снижало уровень содержания фитиновой кислоты в созревших семенах, не вызывая дополнительных изменений в растении, даже в случае выращивания в экстремальных климатических или полевых условиях. Использование тканеизбирательного промотора дает большое количество преимуществ по сравнению с существующими способами, включая ограничение содержания фермента в тканях семян, при котором не оказывается воздействия на мио-инозитный обмен веществ в других тканях.
Целью настоящего изобретения является исключить использование мио-инозита в качестве исходного материала для биосинтеза фитиновой кислоты путем изменения способа проведения метаболизма таким образом, чтобы мио-инозит накапливался в резерве обмена веществ, где он конвертируется в безвредные для растения соединения. Способ основан на введении новых ферментов, причем в одном из способов осуществления настоящего изобретения таковым является ген метилтрансферазы, который уменьшает или изменяет запас мио-инозита, участвующего в образовании фитиновой кислоты. Понижение или изменение уровня содержания этого предшественника приводит к сдвигу нормального биохимического механизма биосинтеза фитиновой кислоты и вызывает снижение количества образующейся фитиновой кислоты.
Биохимической основой настоящего изобретения является концепция регулирования активности фермента за счет изменения доступного количества субстрата. В общем случае скорости ферментных реакций зависят от наличия субстрата. Фермент вызывает образование продукта реакции со скоростью, которая в общем случае пропорциональна доступному количеству субстрата. Снижение концентрации субстрата приводит к снижению уровня образования конечных продуктов.
Настоящее изобретение использует данный подход в сочетании с новым ферментом - метилтрансферазой, который уменьшает доступное количество субстрата, инозита, который участвует в образовании фитиновой кислоты. В рамках настоящего изобретения может быть использовано любое количество различных ферментов, которые могут оказывать влияние на запас мио-инозита в растениях. Данный способ основан на введении какого-либо фермента, который каким угодно образом изменяет величину запаса мио-инозита для биосинтеза фитиновой кислоты.
В отличие от патента США №5563324 настоящее изобретение имеет целью модифицировать количество образующихся фитатов селективно в семенах растения путем экспрессии гена, способного модифицировать мио-инозит так, чтобы сделать его неспособным участвовать в образовании фитиновой кислоты. Неожиданным открытием было то, что даже нерегулируемая экспрессия гена, модифицирующего мио-инозит, приводит к снижению уровня фитатов в семенах. Однако в наиболее предпочтительном варианте осуществления настоящего изобретения для снижения уровня фитатов в семенах растений используется промотор, избирательно действующий на семена, для того, чтобы обеспечить сохранение агротехнических качеств растения и чтобы растение оставалось нормальным в любом отношении, за исключением пониженного уровня фитатов в семенах. Таким образом, в отличие от патента США №5563324 настоящее изобретение не предполагает выведение растений, устойчивых к стрессу, или интересных для исследовательских целей.
Описываемый способ дает большое количество преимуществ по сравнению с существующими способами, относящимися к снижению уровня фитиновой кислоты путем экспрессии ферментов фитазы. К таким преимуществам относятся получение растений с измененным уровнем содержания фитиновой кислоты в отдельных тканях, растений с семенами, имеющими пониженное содержание фитиновой кислоты, а также зерновых кормов с пониженным содержанием фитиновой кислоты. Выгода от использования таких растений, семян и зерновых кормов состоит в лучшем усвоении кормов и эффективности последних, в понижении содержания фосфора в кормах и, соответственно, улучшеннии экологических показателей таких кормов и возможности их использования в таких областях кормовой промышленности, в которых до этого их применение было ограничено вследствие наличия нежелательной с точки зрения питательных свойств фитиновой кислоты.
Описываемый способ может применяться для модификации многих видов растений, таких как одно- и двудольные, а также зерновые и масличные культуры. В особенности же данный способ эффективен для масличных и крестоцветных растений.
Результатом такой генной модификации являются растительные клетки с измененным сахарно-спиртовым обменом веществ, в частности инозитным обменом веществ, а также клетки с пониженным содержанием фитиновой кислоты, в частности с пониженным содержанием фитатов. Таким образом, способ в соответствии с настоящим изобретением предоставляет ценный инструмент изменения вторичного обмена веществ, относящегося к сахарным спиртам.
Хотя вариант использования настоящего изобретения, включающий изменение сахарно-спиртового обмена веществ, например получение растительных клеток с пониженным содержанием фитиновой кислоты, уже был продемонстрирован на примере клеток рапса, зерновые культуры также обладают повышенным содержанием фитиновой кислоты, вследствие чего снижение ее содержания в клетках зерновых растений также возможно.
Настоящее изобретение включает в себя способы, генетические структуры и векторы, которые могут быть полезны для модифицирования содержания фитатов в растительных клетках и семенах, в частности, для снижения уровней фитатов в растительных клетках. Кроме того, настоящее изобретение включает в себя способы и составы ДНК для снижения уровня фитатов в семенах растений. Также настоящее изобретение дает возможность получать модифицированные зерновые смеси и корма для животных на основе таких смесей, в частности зерновых смесей с пониженным содержанием фитатов.
Снижение уровня содержания фитиновой кислоты осуществляется с использованием описанных здесь способов и составов ДНК. Способ включает в себя развитие так называемого метаболического шунтирования, или использование нового биохимического механизма, в результате чего предшественник фитина участвует в другой биохимической реакции, и количество образующейся фитиновой кислоты снижается. В результате одного из вариантов осуществления настоящего изобретения образуется продукт/продукты тупикового механизма реакции, являющиеся безвредными для указанной клетки растения. Соответственно, авторы полностью уверены в том, что шунтирование промежуточных мио-инозитных соединений в такие соединения никоим образом не ухудшит качества растений.
В соответствии с другим аспектом настоящего изобретения предлагаются векторы и рекомбинантные конструкции ДНК для выведения семян растений с измененным содержанием фитиновой кислоты, в частности семян растений с пониженным содержанием фитиновой кислоты. Кроме того, другой аспект настоящего изобретения включает в себя семена растений, которые могут быть использованы для получения зерновых кормов со сниженным содержанием фитатов. За счет пониженных уровней содержания фитатов указанные корма благоприятны для введения в рацион животных, в особенности в тех случаях, когда фитиновая кислота рассматривается как экологически неблагоприятный или нежелательный с точки зрения питательных свойств фактор. Также настоящее изобретение включает в себя корма, полученные из генетически модифицированных семян, которые можно использовать в рационах различных животных, включая корма для птицы, свиней, крупного рогатого скота и рыбы.
Способ включает в себя введение нового фермента, способного изменять содержание мио-инозита в семенах растений, который может применяться напрямую для любых типов растений. Один и тот же ген может быть использован в разных растениях, поскольку изменение мио-инозита в них происходит аналогичным образом. Способ не основан на использовании технологий супрессии генов, которые зависят от типа последовательности ДНК и, следовательно, требуют специальных генов для каждого отдельного гена или вида растений. Следовательно, настоящее изобретение может использоваться для всех видов растений. В рамках данного способа могут использоваться многочисленные ферменты, полученные из бактериальных, растительных или животных источников. Одни и те же генетические конструкции могут использоваться для широкого разнообразия различных растений.
Следовательно, в соответствии с общим аспектом настоящего изобретения предлагается способ снижения содержания фитатов в семенах растений, включающий в себя построение вектора трансформации растения, который, помимо выборочного маркера, позволяющего идентифицировать растительные клетки, трансформированные с помощью данного вектора, содержит фермент, способный осуществлять переработку мио-инозита, что делает последний непригодным для участия в образовании фитиновой кислоты.
В контексте настоящего изобретения переработка инозита включает в себя гидроксилирование, изомеризацию, метилирование, расщепление или иную биохимическую модификацию, которая делает мио-инозит неспособным к восприятию воздействия соответствующего фермента, который в обычных условиях воздействует на мио-инозит с образованием фитиновой кислоты. Следовательно, переработка мио-инозита включает в себя любую модификацию, которая эффективно уменьшает количество доступных запасов мио-инозита в качестве субстрата для биосинтеза фитиновой кислоты.
В качестве примера осуществления настоящего изобретения в одном из его аспектов можно привести ген, кодирующий мио-инозит-метилтрансферазу. Этот фермент под контролем промотора, избирательно проявляющегося в клетках семян растений, способен уменьшить количество доступных запасов мио-инозита для образования фитатов в указанных семенах растений. В соответствии с другим аспектом настоящего изобретения, указанная метилтрансфераза способна переводить мио-инозит в безвредное вещество.
Растительные клетки, полученные с использованием способа в соответствии с настоящим изобретением, имеют неизменный первичный обмен веществ и фенотипы, неотличимые от фенотипов немодифицированных растений, за исключением изменения продукта или нескольких продуктов вторичного механизма обмена веществ.
Для того чтобы обеспечить неизменность первичного обмена веществ, в настоящем изобретении предлагается способ оказания воздействия на образование вторичных продуктов обмена веществ селективно в зависимости от вида ткани путем изменения доступного количества субстрата, который характерен именно для этого механизма обмена веществ и необходим для образования конечного продукта этого механизма, в особенности такогосубстрата, который участвует в обмене веществ за 1-5 стадий до образования конечного продукта. Таким образом, достигается снижение содержания отдельных продуктов вторичного обмена веществ.
В контексте настоящего изобретения соединения, используемые в качестве предшественников фенилпропаноидного вторичного обмена веществ, включают в себя, среди прочих, коричную кислоту, р-кумаровую кислоту, кофеиновую кислоту, феруловую кислоту, 5-гидроксиферуловую кислоту, синапиновую кислоту, холин, а также различные синапоиловые соединения, которые включают в себя, не ограничиваясь, синапоилглюкозу, синапоилмалат и синапоилхолин. В контексте настоящего изобретения продукты фенилпропаноидного вторичного обмена веществ включают в себя, среди прочих, флавоноиды, лигнины, различные мономерные и полимеризованные фенольные соединения, а также синапин. Выбирая определенный фермент, можно вносить сколь угодно большое количество изменений в содержание продуктов фенилпропаноидного обмена веществ. Ферменты, используемые для модифицирования или оказания влияния на соединения, участвующие в обмене веществ в качестве предшественников, могут иметь бактериальную, растительную или животную природу. Ферменты могут быть как встречающимися в естественных условиях, так и выведенными за счет мутации известного фермента с целью изменения его субстрата и, следовательно, проявления им других свойств.
Ферменты, используемые в рамках настоящего изобретения, могут быть способны модифицировать соединения, участвующие в качестве предшественников в процессе фенилпропаноидного вторичного обмена веществ, путем изомеризации, конъюгации, фосфорилирования, гидроксилирования, окисления, дегидрирования, метилирования или иной подобной биохимической реакции (включая связывание или секвестрирование), а также могут быть способны разлагать соединения, участвующие в качестве предшественников в процессе фенилпропаноидного вторичного обмена веществ, например гидролазы, декарбоксилазы, оксидазы, эстеразы или другие ферменты, способные разлагать L-фенилаланин, коричную кислоту, р-кумаровую кислоту, кофеиновую кислоту, феруловую кислоту, 5-гидроксиферуловую кислоту, синапиновую кислоту, синапоилглюкозу, синапоилмалат и холин (участвующий в образовании синапина из синапоилглюкозы). В одном аспекте данного способа используются ферменты, образующие стрессозащитные вещества из соединений, участвующих в качестве предшественников в образовании продуктов обычного фенилпропаноидного обмена веществ. Аналогичным образом уровень конечного продукта фенилпропаноидного обмена веществ может быть снижен за счет использования фермента для избирательного снижения содержания дополнительного субстрата, который не образуется в процессе фенилпропаноидного обмена веществ, но необходим для образования продукта этого обмена веществ. Соответственно, в рамках настоящего изобретения могут быть изменены как соединения, участвующие в качестве предшественников, так и любые иные соединения, необходимые для образования продуктов фенилпропаноидного обмена веществ.
В контексте настоящего изобретения соединения, используемые в качестве предшественников сахарно-спиртового вторичного обмена веществ, включают, среди прочих, мио-инозит, галактинол и UDP-производные сахаров. Продукты сахарно-спиртового вторичного обмена веществ включают, не ограничиваясь, фитиновую кислоту, стахиозу, рафинозу, сукрозилгликозиды, урониды, пентозы, фосфоинозитиды и гликофосфокерамиды. В контексте настоящего изобретения фермент, используемый для изменения соединений, участвующих в сахарно-спиртовом вторичном обмене веществ, включают ферменты, спобные к гидроксилированию, изомеризации, метилированию, расщеплению или любой иной биохимической модификации (например, связывание или секвестрирование), которая делает сахарный спирт неспособным к восприятию воздействия соответствующего фермента, который в обычных условиях воздействует на него. В качестве примера осуществления настоящего изобретения по отношению к сахарно-спиртовому вторичному обмену веществ показано, как метилирование инозита, являющегося предшественником фитиновой кислоты, уменьшает количество доступного инозита для биосинтеза фитиновой кислоты.
Для специалистов в данной области понятно, что предшественники конечного продукта вторичного обмена веществ представляют собой либо те биохимические вещества, которые непосредственно участвуют в образовании этого продукта вторичного обмена веществ, либо те вещества, которые участвуют в биохимической реакции, приводящей к образованию определенного продукта вторичного обмена веществ, причем ни те, ни другие вещества не являются продуктами первичного обмена веществ.
Ферменты, используемые для оказания воздействия на соединения, действующие как предшественники продуктов вторичного обмена веществ, предпочтительно должны превращать эти соединения в вещества, являющиеся безвредными для растительных клеток и, следовательно, могущие накапливаться до значительных уровней без изменения свойств растительных клеток. Такие ферменты могут также образовывать из указанных соединений, являющихся предшественниками, вещества, оказывающие благоприятное воздействие на растительную клетку, например стрессозащитные вещества. Эти ферменты также могут образовывать вещества, пригодные к использованию в химической, фармацевтической или пищевой промышленности. В рамках настоящего изобретения могут быть использованы один или более таких ферментов в одном или более качестве для снижения уровня содержания отдельных веществ, являющихся предшественниками продукта вторичного обмена веществ.
Ферменты, используемые в контексте настоящего изобретения, могут быть получены из разных источников разными методами. Например, специалист в данной области всегда сможет определить, на какой предшественник в рамках вторичного обмена веществ необходимо оказать воздействие. Химическое строение всех этих соединений хорошо известно или может быть раскрыто, поэтому для определения фермента, которым необходимо воздействовать на химически подобные субстраты, достаточно проанализировать данные, имеющиеся в соответствующей литературе. Таким образом, в контексте настоящего изобретения может быть определен подходящий фермент и выделен и использован соответствующий ген. Кроме того, могут быть использована комбинаторная химия или аналогичные приемы поиска искусственных белков, оказывающих нужный тип воздействия, в качестве основы для получения синтетического гена. Ферменты, используемые в рамках настоящего изобретения, могут быть модифицированы с использованием общеизвестных способов.
Как пример модифицирования фермента можно привести ген, кодирующий фермент, способный оказывать воздействие на химически родственные вещества, участвующие в качестве предшественников во вторичном обмене веществ. Этот ген можно модифицировать и/или выделить для усиления его действия с помощью таких способов, как сайт-специфическая модификация, мутация или селекция измененных ферментов в гетерологической системе, например в бактериальных или грибковых клетках. Этот процесс может включать в себя экспрессию гена, кодирующего указанный измененный фермент в бактериальных клетках, неспособных усвоить указанные вещества, выбор бактерий, в которых этот фермент проявляется и которые способны расти за счет этих веществ как только источника питания или источника углерода, а также восстановление активности фермента, способного оказывать воздействие на определенное вещество, участвующее в качестве предшественника во вторичном обмене веществ. Таким образом, можно получить фермент, избирательно воздействующий на определенное соединение, участвующее в качестве предшественника в определенном процессе вторичного обмена веществ. В альтернативном варианте могут быть выбраны бактерии для выращивания на определенных соединениях, участвующих в качестве предшественников во вторичном обмене веществ. Специалист в данной области легко сможет понять, что, выбирая мутированные бактерии или иные клетки, выращенные на соединениях, участвующих в качестве предшественников во вторичном обмене веществ, можно определить конкретные ферменты, способные превратить указанные соединения в безвредные вещества. Этот подход может включать в себя прямую селекцию определенных ферментов или пошаговую модификацию с селекцией нужных ферментов.
Аналогичным образом, использование известных ферментов, в настоящее время применяемых на промышленной основе, например, препаратов из экстракта грибка Trichoderma, способных разлагать лигнин в процессе получения целлюлозы, дает возможность получить подходящий фермент, например, для изоляции ферментов, участвующих в разложении фенольных соединений. Специалистам в данной области будет понятно, что гены, кодирующие ферменты, используемые в целлюлозно-бумажной промышленности для переработки лигнина, могут быть использованы и в рамках способа в соответствии с настоящим изобретением, либо напрямую, либо после их модифицирования, для воздействия на одно из соединений, участвующих в качестве предшественника в фенилпропаноидном обмене веществ. Соответственно предполагается, что в рамках настоящего способа может также быть использовано большое количество разнообразных ферментов. Таким образом, данный способ не ограничивается источником или особенностями используемого фермента.
Например, в одном из аспектов данного способа фермент, проявляющийся в растительных клетках, воздействует на соединение, используемое в качестве предшественника или субстрата для продукта фенилпропаноидного обмена веществ, в результате чего образуется субстрат, который более не подвержен воздействию фермента, который в обычных условиях использовал бы его в качестве предшественника в фенилпропаноидном обмене веществ. Соответственно уровень содержания данного продукта фенилпропаноидного обмена веществ снижается за счет уменьшения доступного количества предшественника, необходимого для образования конечного продукта. В результате растительная клетка может использоваться либо как таковая, либо для регенерации растения с измененным уровнем содержания продуктов фенилпропаноидного обмена веществ. Этот путь может найти применение в большом количестве промышленных процессов, не ограничиваясь кормовой промышленностью, целлюлозно-бумажной промышленностью или выведением растений с измененным составом фенольных соединений, включая растения с избытком некоторых продуктов фенилпропаноидного обмена веществ. Деревья с измененным уровнем продуктов фенилпропаноидного обмена веществ могут использоваться в целлюлозно-бумажной промышленности. Растения с измененным уровнем продуктов фенилпропаноидного обмена веществ могут использоваться в кормовой промышленности.
Настоящий способ также основан на регенерации и использовании растительных клеток или тканей с измененными свойствами, в частности растительных тканей, используемых в кормовой промышленности или иных промышленных процессах. Вероятно, изменения в фенилпропаноидном обмене веществ, осуществление которых предполагается в рамках настоящего изобретения, приведут также и к положительным последствиям для агротехнических качеств растений, выведенных таким образом, в особенности в тех аспектах изобретения, в которых введение нового фермента приводит к образованию стрессозащитных веществ. Другими положительными последствиями применения данного способа могут стать повышенная устойчивость к заболеваниям и ультрафиолетовому излучению, механическая прочность и т.д.
В еще одном варианте осуществления настоящего изобретения способ включает в себя стадию выращивания указанной растительной клетки и регенерирования растения с измененными продуктами фенилпропаноидного обмена веществ.
Далее один из вариантов осуществления настоящего изобретения включает в себя использование указанного растения в промышленных целях, например производстве химикатов, приготовлении корма для скота или в целлюлозно-бумажной промышленности.
Гены, кодирующие ферменты, способные воздействовать на соединения, используемые в качестве предшественников в фенилпропаноидном обмене веществ, могут быть естественного происхождения, синтетическими или комбинированными. Для опытного профессионала в данной области очевидно, что кодирующая последовательность гена может быть модифицирована для того, чтобы обеспечить высокую степень экспрессии гена в растительных клетках. Это может быть достигнуто за счет изменения последовательностей кодонов для более адекватного использования кодона, естественно встречающегося в растительных клетках, в которых данный ген должен быть экспрессирован. Кроме того, становится очевидно, что можно создать определенные рестрикционные сайты для удобного управления кодирующей последовательностью. Также предполагается, что введение различных последовательностей, таких как трансляционных ускорителей, интронов и т.п., может использоваться для достижения адекватной экспрессии кодирующей последовательности. Ферменты могут быть также модифицированы путем изменения сайтов, связывающих субстраты, или с помощью иной модификации первичной последовательности аминокислот, усиливающей активность фермента. Все эти манипуляции широко известны в практике и будут понятны специалистам.
В другом аспекте настоящего изобретения фермент контролируется тканеизбирательным промотором, в частности промотором, действующим в тканях семян. Такой промотор представляет собой промотор, который действует исключительным или предпочтительным образом, вызывая экспрессию находящихся под его контролем последовательностей только в тканях семян растений. Соответственно уровни определенного продукта фенилпропаноидного обмена веществ изменяются селективно по отношению к ткани растения, в то время как в других тканях растения уровни этого продукта фенилпропаноидного обмена веществ остаются на нормальном уровне. Ткань, в которой фермент проявляется селективным образом, используется для приготовления кормов или в ином промышленном процессе.
Для специалиста в данной области очевидно, что в рамках настоящего изобретения может быть использовано любое количество тканеизбирательных промоторов. В частности, промотор, селективный по отношению к тканям семян, используется для изменения продуктов фенилпропаноидного обмена веществ в культурах, зерна которых используются для приготовления кормов. В других культурах могут быть использованы другие тканеизбирательные промоторы в зависимости от той части растения, в которой необходимо изменить уровень фенольных соединений. Например, промотор, селективный по отношению к корням или клубням, может быть использован для изменения содержания фенолов или лигнинов в корнеплодах или клубневых культурах, таких как картофель, репа, маниока и т.п. В тех случаях, когда в качестве кормов используются листья или стебли растений, можно использовать промотор, селективный по отношению к соответственно листьям или стеблям, для ограничения экспрессии ферментов, способных воздействовать на соединения, участвующие в фенилпропаноидном обмене веществ в качестве продуктов предыдущей реакции. Другие тканеизбирательные промоторы могут быть использованы в растениях, например деревьях, идущих на переработку в целлюлозно-бумажной промышленности. Таким образом, в рамках настоящего изобретения может найти применение и промотор, который ограничивает активность фермента в древесных тканях дерева.
В одном особом аспекте настоящего изобретения предполагается экспрессия кодирующей последовательности фермента, ответственного за усвоение холина в растительных клетках. Экспрессия указанного фермента уменьшает запасы доступного холина, который участвует в образовании синапина из синапоилглюкозы. Некоторые растения, в особенности крестоцветные, вырабатывают синапин, являющийся нежелательным с точки зрения питательных свойств растения. Считается, что синапин синтезируется путем глюкозной компоненты синапоилглюкозы на холин. Снижение содержания синапина как нежелательного для питательных свойств растений вещества увеличивает кормовую ценность семян и получаемых из них кормов. Использование фермента, модифицирующего холин, например холиноксидазы, может также найти применение в рамках настоящего изобретения.
Известно, что холин при окислении переходит в бетаин в растениях, имеющих значение для пищевой промышленности, например пшенице, шпинате, сахарной свекле и ячмене (Rhodes, Hanson, 1993, Annual Review of Plant Physiology and Plant Molecular Biology (Ежегодный обзор физиологии и молекулярной биологии растений), 44, стр.357-384)). Катализаторами этой окислительной цепочки являются два фермента (в общем виде известные как система окисления холина), а продукты этой цепочки приведены ниже:
Холин→бетаинальдегид→глицинбетаин (бетаин)
Первая стадия этой цепочки катализируется в бактериях и тканях животных холиндегидрогеназой (CDH) или холиноксидазой (СОХ) (Rozwadowski, Khachatourians, Selvaraj, 1991, Journal of Bacteriology, 173, стр.472-478) и холинмонооксидазой (СМО) в растениях (Rhodes, Hanson, 1993). Вторая стадия этой цепочки катализируется бетаинальдегиддегидрогеназой (BADH) (Rhodes, Hanson, 1993). Соответственно, использование фермента, способствующего усвоению холина, может уменьшить доступные запасы холина общим или специфическим образом с участием нерегулируемых, индуктируемых или тканеизбирательных промоторов. Снижение содержания доступного холина уменьшает количество образующегося синапина. Кроме того, снижение уровня синапина может вызвать изменение содержания других продуктов фенилпропаноидного обмена веществ, что приведет к изменению содержания фенольных соединений в растительных тканях.
В другом особом варианте осуществления настоящего изобретения бактериальный ген холиноксидазы (СОХ) (Rozwadowski, Khachatourians, Selvaraj, 1991, Journal of Bacteriology, 173, стр.472-478) используется под контролем промотора, селективного по отношению к семенам. Экспрессия холиноксидазы в семенах переводит холин, который является одним из предшественников синапина в семенах рапса, в безвредное вещество бетаин через образование бетаинальдегида, который постепенно переводится в бетаин под действием фермента СОХ. Семена, содержащие рекомбинантную ДНК, отличаются пониженным содержанием синапина и измененным содержанием других продуктов фенилпропаноидного обмена веществ. Корма, получаемые из таких семян, ценны с точки зрения их питательных свойств.
В другом особом варианте осуществления настоящего изобретения ген СОХ также используется под контролем промотора, селективного по отношению к семенам, но для ускорения образования бетаина из бетаинальдегида используется второй фермент - BADH. В этом варианте фермент BADH (Boyd L.A., L. Adam, L.E. Pelcher, A. McHughen, R. Hirji, G. Selvaraj, 1990, Gene, 103, стр.45-52) проявляется под контролем промотора, селективного по отношению к семенам. Бетаин - соединение, встречающееся в съедобных частях многих пищевых и кормовых растений и также иногда используемое в качестве добавок к кормам и пище людей. Помимо этого бетаин придает растениям стрессозащитные свойства.
В другом особом варианте осуществления настоящего изобретения ген СОХ используется в клетках крестоцветных растений (Brassica) под контролем промотора, селективного по отношению к семенам, и для образования бетаина из бетаинальдегида используется второй фермент - BADH, также под контролем промотора, селективного по отношению к семенам.
Изменение содержания холина в крестоцветных растениях обеспечивает снижение содержания синапина и в дальнейшем изменение различных продуктов фенилпропаноидного обмена веществ. Авторы полностью уверены, что изменение уровней содержания холина в семенах приводит к повышению уровня синапоилглюкозы или других синапоиловых соединений, например синапоилмалата, который, в свою очередь, изменяет уровень содержания синапиновой кислоты, являющейся предшественником лигнина. Следовательно, в данном случае задействован биохимический механизм регулирования, в общем случае называемый ингибированием конечного продукта, в результате которого изменяются уровни содержания различных продуктов фенилпропаноидного обмена веществ. Соответственно, предотвращение образования синапина приводит к изменению уровня большого числа продуктов, образующихся в ходе фенилпропаноидного обмена веществ до его завершающей стадии - образования синапина.
Вторичный процесс фенилпропаноидного обмена веществ может также быть изменен путем использования фермента, проявляющегося в растительных клетках и действующего на феруловую кислоту. Феруловая кислота является еще одним предшественником фенилпропаноидного обмена веществ. Соответственно снижается уровень содержания феруловой кислоты. Снижение доступного количества феруловой кислоты приводит к изменению уровней содержания фенольных соединений, включая синапин в крестоцветных.
Гены, кодирующие ферменты, способные воздействовать на феруловую кислоту, могут быть естественными, синтетическими или комбинированными. Для опытных специалистов очевидно, что кодирующая последовательность гена может быть модифицирована таким образом, чтобы ген мог проявляться в растительных клетках в значительной степени.
В соответствии с другим аспектом настоящего изобретения ген, кодирующий фермент, способный воздействовать на феруловую кислоту, используется под контролем тканеизбирательного промотора. Соответственно уровень содержания феруловой кислоты изменяется в определенной ткани растения, причем в других тканях ее содержание остается на нормальном уровне. Ткань растения, в которой фермент проявляется селективным образом, используется для приготовления кормов, в пищевой промышленности или для других промышленных целей.
Ферменты, которые могут использоваться в этих вариантах осуществления настоящего изобретения, включают такие ферменты, которые способны модифицировать феруловую кислоту, и, в частности, такие ферменты, которые приводят к образованию из феруловой кислоты ценных соединений. Для специалиста в данной области очевидно, что в рамках данного способа могут использоваться и другие ферменты, способствующие усвоению феруловой кислоты. Известными источниками ферментов, способствующих усвоению феруловой кислоты, являются следующие микроорганизмы: Bacillus megaterium, В. Subtilis, Corynebacterum glutamicum, виды Enterobacter, Pseudomonas, Streptomyces, Aspergillis, Candida, Fusarium, hanseluna, Penicillum, Rhodotorula, Saccharomyces. Этот перечень далеко не полон, но иллюстрирует разнообразие известных в литературе источников ферментов, способствующих усвоению ферулатов. Таким образом, настоящий способ не ограничен конкретным источником фермента, способствующего усвоению феруловой кислоты. В рамках настоящего изобретения предполагается изменение бактериально кодированного гена для достижения оптимальной экспрессии фермента в растительных клетках. Способы такого изменения хорошо известны в литературе.
В качестве примера такого фермента можно привести декарбоксилазу феруловой кислоты, получаемую из В. pumulis. Ген этого фермента был выделен, и его последовательность была установлена в работе Zago и др., Applied and Environmental Microbiology, 61, стр.4484-4486, 1995. Последовательность этого гена может быть модифицирована для достижения более полного соответствия растительному гену при сохранении необходимой последовательности аминокислот. Этот фермент приводит к образованию винилгваякола из феруловой кислоты путем декарбоксилирования. Таким образом, в данном примере с использованием гена, способного воздействовать на феруловую кислоту, образуется полезное соединение винилгваякол (VG). Этот фермент не требует дополнительных факторов и имеет молекулярную массу приблизительно 22 кДа.
В другом особом варианте осуществления настоящего изобретения фермент ферулат-декарбоксилазы используется под контролем тканеизбирательного промотора, в результате чего некоторые ткани растения имеют измененный обмен веществ по феруловой кислоте.
Изменение феруловой кислоты приводит к снижению ее содержания и в дальнейшем к изменению различных продуктов фенилпропаноидного обмена веществ.
Настоящее изобретение также предполагает получение растительных тканей, в частности семян растений, которые могут использоваться в кормовой или пищевой промышленности, причем рассматривается как непосредственное использование семян в качестве корма, так и использование модифицированного корма из обработанных семян. Снижение содержания фенолов в семенах растений было бы желательно с точки зрения производства кормов, а генетически модифицированные семена с измененным содержанием фенолов, в частности сниженным содержанием нежелательных с точки зрения питательных свойств фенолов, имели бы большую ценность для пищевой промышленности.
В дополнение к непосредственному использованию цельных или частично измолотых семян в качестве кормов существуют и другие способы получения кормов. Для крестоцветных особое значение имеет способ получения кормов в процессе переработки семян масличных крестоцветных. В общем случае масло получают из семян масличных культур либо с использованием растворителя, либо без такового. Жмых и прочие твердые отходы, получаемые в ходе этого процесса, используются для приготовления кормов, и обычно в них содержится основная часть фенолов в семенах крестоцветных. Способ приготовления кормов с пониженным содержанием фенолов дает возможность получения нового состава кормов, в настоящее время не получаемого из семян крестоцветных.
Настоящее изобретение предполагает использование генетически модифицированных семян, полученных в соответствии со способом приготовления кормов с пониженным содержанием фенолов.
Описываемый способ имеет большое количество преимуществ с точки зрения изменения фенотипа растения. Кроме того, данный способ позволяет получать различные соединения с высокой промышленной ценностью. Способ применим к любым растениям или растительным тканям.
Указанный способ дает большие преимущества по сравнению с существующими способами управлением фенилпропаноидным обменом веществ. Способ не основан на мутациях, таких как мутация SIN1, которая проявляется во всем растении и связана с чувствительностью к ультрафиолетовому излучению. Способ не использует генетических механизмов, разработанных с целью блокировки экспрессии гена, таких как использование антисенсибилизирующей ДНК или ко-супрессии, которые требуют клонирования растительных генов из фенилпропаноидного обмена веществ. Способ также не модифицирует никаких генов, которые свойственны растению. Соответственно, не оказывается никакого нежелательного воздействия на какие-либо процессы, происходящие в ходе фенилпропаноидного обмена веществ, в частности образование флавоноидов, выработку веществ, защищающих растение от ультрафиолетового излучения, активизацию защитных реакций на заболевания или иные биохимические функции. Использование тканеизбирательных промоторов позволяет изменять содержание фенолов в любой отдельно взятой ткани растения при сохранении их содержания в других тканях.
Описываемый метод применим к большому числу растений, но имеет особое значение для крестоцветных. Использование этого способа по отношению к таким растениям дает дополнительные преимущества, такие как образование стрессозащитных или ценных для промышленности веществ в результате введения фермента. Очевидно, что метод может быть использован для оказания воздействия на любое количество реакций в ходе фенилпропаноидного обмена веществ, и что аналогичным образом могут быть изменены и другие биохимические механизмы.
Настоящее изобретение также применимо для модификации других вторичных процессов обмена веществ. Например, в одном из вариантов осуществления настоящего изобретения существуют способы и структуры ДНК, служащие для выведения растений с измененным сахарно-спиртовым обменом веществ, в частности с измененным содержанием фитатов в клетках семян. Фитаты образуются из сахарно-спиртового мио-инозита, который также является предшественником или промежуточным продуктом для образования других компонент, включая стахиозу, рафинозу, сукрозилглюкозиды, урониды, пентозы, фосфоинозитиды и гликофосфокерамиды. Кроме того, настоящее изобретение включает в себя способы и составы ДНК для снижения содержания фитатов в семенах крестоцветных. Также вариант осуществления настоящего изобретения включает в себя семена растений с пониженным содержанием фитиновой кислоты, приемлемым для кормовых целей, семена растений с пониженным содержанием фитиновой кислоты, приемлемым для приготовления модифицированных кормов, а также растительные клетки с модифицированным инозитным обменом веществ.
Настоящее изобретение включает в себя способы и конструкции ДНК для выведения растений с измененным содержанием фитатов в клетках семян. Также настоящее изобретение включает в себя способы и составы ДНК для снижения содержания фитатов в семенах однодольных, двудольных, масличных и крестоцветных. Помимо этого, настоящее изобретение включает в себя семена растений с пониженным содержанием фитиновой кислоты, приемлемым для кормовых целей, семена растений с пониженным содержанием фитиновой кислоты, приемлемым для приготовления модифицированных кормов, а также растительные клетки с модифицированным инозитным обменом веществ.
Настоящее изобретение предполагает экспрессию генов, кодирующих ферменты, которые воздействуют на сахарно-спиртовой предшественник фитиновой кислоты, а именно на мио-инозит, что приводит к тому, этот предшественник участвует в другой биохимической реакции или переводится в плохоперерабатываемые соединения, которые могут накапливаться в растительных клетках без вреда для последних. Таким образом достигается снижение содержания фитиновой кислоты. Другие сахарные спирты также могут быть объектом применения способов, описанных в настоящем изобретении. Фермент, используемый для воздействия на мио-инозит, может быть бактериальным, животным или растительным. Он может быть как естественного происхождения, так и искусственно полученным путем мутации известного фермента с целью изменения его субстрата и таким образом получения новых свойств этого фермента.
В предпочтительном варианте фермент, используемый для оказания воздействия на мио-инозит, должен приводить к образованию соединений, которые безвредны для растительных клеток и могут накапливаться в больших количествах без отрицательных последствий для них. В рамках настоящего изобретения могут использоваться один или несколько ферментов, причем для снижения уровня мио-инозита могут также использоваться более одного различного фермента.
Ферменты, используемые в рамках настоящего изобретения, способны модифицировать мио-инозит путем изомеризации, конъюгации, фосфорилирования, гидроксилирования, метилирования или любой иной биохимической процедуры, или же разлагать мио-инозит или иные сахарные спирты.
Ферменты, используемые в контексте настоящего изобретения, могут быть получены из большого количества источников или с использованием большого количества способов. Например, специалисту в данной области не составит труда определить, какой фермент необходим для оказания нужного воздействия с использованием химически подобных субстратов. Соответственно такой фермент может быть определен, а его ген выделен и использован в контексте настоящего изобретения. Ферменты, используемые в рамках настоящего изобретения, могут быть модифицированы с использованием общеизвестных способов.
Способ в соответствии с настоящим изобретением также основан на использовании трансформации с целью введения гена, кодирующего нужный фермент, в клетки растений. Трансформация растительной клетки может быть осуществлена большим количеством способов. Общепринятые способы включают в себя системы, основанные на Agrobacterium, с использованием либо бинарных, либо коинтегральных плазмид и A. tumifaciens, и А. Rhyzogenes (например, Патенты США №№4940838 и 5464763), биолистического подхода (например, Патенты США №№4945050 и 5015580), микроинъекции (например, Патент США №4743548), прямого выбора ДНК протопластами (например, Патенты США №№5231019 и 5453367) или способа игольчатых усиков (например, Патент США №5302523). В контексте настоящего изобретения может быть использован любой способ генетической трансформации растительной клетки.
Способ также основан на восстановлении и использовании растительных клеток или тканей с измененными свойствами, в частности тканей семян растений, используемых в кормовых целях.
Таким образом получают растительные клетки с измененным сахарно-спиртовым обменом веществ. Из сахарных спиртов образуется большое количество соединений, но особое значение имеет фитиновая кислота или фитаты. Способы снижения содержания фитатов в определенных растительных клетках имеют большое значение для приготовления кормов.
Гены, кодирующие ферменты, способные воздействовать на мио-инозит, могут быть естественными, синтетическими или комбинированными. Для опытного специалиста в данной области очевидно, что кодирующая последовательность гена может быть модифицирована с целью более активной его экспрессии в растительных клетках. Этого можно достичь, изменяя последовательности кодонов для более полного соответствия кодонам, свойственным для той клетки, в которой ген должен проявляться. Кроме того, очевидно, что могут быть созданы специальные рестрикционные сайты для обеспечения более удобного управления кодирующей последовательностью. Также предполагается, что для обеспечения адекватной экспрессии кодирующей последовательности может быть использовано введение различных последовательностей, например трансляционных промоторов, интронов и т.п. Все эти манипуляции являются общепринятыми в практике и будут легко доступны опытным специалистам в данной области.
Очевидно также, что для ограничения экспрессии фермента, способного модифицировать мио-инозит в тех такнях, где вырабатывается фитиновая кислота, например в семенах, может быть использован тканеизбирательный промотор. Подходящими промоторами, селективными по отношению к тканям семян, являются напин, получаемый из Brassica napus, фазеолин, получаемый из Phaseolis, а также любые иные промоторы, действующие только в семенах.
В частности, в рамках настоящего изобретения предполагается использование мио-инозит-оксиметилтрансферазы, например, такой, которая получается из Mesembryanthemum crystallinum.
Настоящее изобретение также предполагает использование генетически модифицированных семян, полученных путем экспрессии ферментов, способных воздействовать на мио-инозит и контролируемых промотором, селективным по отношению к семенам. Соответственно снижается уровень содержания фитиновой кислоты в семенах. При обработке таких семян в результате получается корм для животных с более низким содержанием фитиновой кислоты. Соответственно, данный способ также позволяет получать корма с пониженным содержанием фитиновой кислоты.
Настоящее изобретение предоставляет способ получения крестоцветных растений с пониженным содержанием фитиновой кислоты. В частности, в рамках настоящего изобретения предполагается использование мио-инозит-оксиметилтрансферазы, например, такой, которая получается из Mesembryanthemum crystallinum, для понижения уровня содержания фитиновой кислоты в крестоцветных культурах. Для опытного специалиста в данной области очевидно, что в результате использования настоящего изобретения возможно получение кормов с пониженным содержанием фитиновой кислоты на основе семян крестоцветных.
Известно, что наличие фитиновой кислоты в зерновых кормах отрицательно сказывается на качестве разводимой рыбы. Однако некоторые корма, например на основе семян рапса, содержат благоприятное для разведения рыбы сочетание белков. Высокое содержание фитиновой кислоты в семенах рапса ограничивает ее применение в рационе искусственно разводимой рыбы. Следовательно, для этой отрасли сельского хозяйства имеет большое значение использование кормов с пониженным содержанием фитиновой кислоты, полученных из семян крестоцветных.
Таким образом, настоящее изобретение может использоваться для получения семян с модифицированным содержанием фитиновой кислоты и зерновых кормов с пониженным содержанием фитиновой кислоты. Модифицированные семена и корма могут использоваться в рационе самых различных животных, например птицы, свиней, рыб, жвачных и нежвачных. Кроме того, из предыдущего описания очевидно, что для модификации мио-инозита в рамках настоящего изобретения может использоваться большое количество различных ферментов. Очевидно также, что для снижения уровня фитиновой кислоты до нужного уровня могут использоваться различные комбинации этих ферментов в сочетании с промоторами, селективными по отношению к семенам.
Применение настоящего изобретения по отношению к вторичному процессу сахарно-спиртового обмена веществ не ограничивается приведенными здесь примерами растений. Действительно, способ в соответствии с настоящим изобретением может использоваться для модификации любых растений. Данный способ применим к любым культурным растениям, поскольку используемый фермент оксиметилтрансфераза гетерологичен по отношению ко всем основным культурным растениям, имеющим значение для сельского хозяйства. Следовательно, любые культурные растения, используемые в качестве кормов для животных, или семена которых используются целиком или частично в данной области, могут быть модифицированы с помощью метода в соответствии с настоящим изобретением для снижения уровня фитиновой кислоты. Способ применим к любым культурным растениям, включая однодольные (например, хлебные злаки, рис, пшеница, ячмень, рожь) и двудольные (например, соевые, хлопок, люцерна, Brassica, лен, подсолнечник и т.д.).
Данный способ обеспечивает модифицирование как фенилпропаноидного, так и сахарно-спиртового процессов обмена веществ, а также представляет собой пример того, каким образом в соответствии с настоящим изобретением надежным и предсказуемым образом может быть достигнуто снижение содержания определенных соединений, участвующих в обмене веществ, в частности соединений, нежелательных с точки зрения питательных свойств.
Для специалистов в данной области очевидно, что в рамках настоящего изобретения может быть изменен любой вторичный процесс обмена веществ. Настоящее изобретение было применено в качестве примера по отношению к двум вторичным процессам обмена веществ и дало предсказуемые и ощутимые результаты. В широком понимании настоящего изобретения возможно снижение компонент, вредных с точки зрения питательных свойств, в частности, в клетках рапса, хлебных злаков, риса и хлопка.
В качестве части способа в соответствии с настоящим изобретением опытный специалист в данной области должен оценить количество образующегося конкретного вторичного продукта обмена веществ. Такой специалист легко определит, что для этого необходимо глубокое знание тканей, в которых синтезируется данный вторичный продукт обмена веществ или его предшественник, помимо наличия информации о стадии, на которой этот процесс происходит. Такая информация будет являться критерием для выбора промотора, который бы контролировал экспрессию фермента, действующего на вторичный продукт обмена веществ или его предшественник. Таким образом, опытный специалист легко поймет, что оказание воздействия на указанный вторичный продукт обмена веществ требует наличия фермента, способного воздействовать на предшественник указанного вторичного продукта обмена веществ, в ткани, где указанный вторичный продукт обмена веществ вырабатывается, а также на той стадии, на которой он вырабатывается. Следовательно, фермент, оказывающий воздействие на предшественник указанного вторичного продукта обмена веществ, должен проявляться в момент синтеза указанного предшественника или вблизи этого момента.
Далее специалисту необходимо провести биохимический анализ ткани, по отношению к которой данное изобретение должно быть применено. Такой анализ может включать в себя определение уровней различных соединений в данной ткани на протяжении всего развития процесса, компартментализацию или субклеточную локализацию процесса биосинтеза, период времени в процессе развития, когда данное соединение образуется впервые или перестает синтезироваться, а также другую соответствующую биохимическую информацию. Проводя такой анализ, опытный специалист сможет выбрать промотор, который обеспечивал бы приемлемую степень экспрессии фермента для того, чтобы уровень содержания вторичного продукта обмена веществ в ткани растения изменялся нужным образом. Следует учитывать, что даже незначительные изменения содержания предшественника могут оказаться достаточными для достижения желаемого эффекта.
Например, как часть варианта осуществления настоящего изобретения, относящегося к фитиновой кислоте, для опытного специалиста очевидно, что базовое знание тканей, в которых синтезируется фитиновая кислота, а также информация о стадии, на которой происходит образование фитиновой кислоты, являются критериями выбора промотора для контроля за экспрессией фермента, воздействующего на мио-инозит. Таким образом, специалисту станет понятно, что для оказания воздействия на мио-инозит будет необходим фермент, который способен воздействовать на мио-инозит в той ткани, где образуется фитиновая кислота, а также на той стадии, на которой фитиновая кислота образуется. Следовательно, фермент, оказывающий воздействие на мио-инозит, должен проявляться в момент образования фитиновой кислоты или вблизи него. Степень экспрессии этого фермента может быть модифицирована с целью достижения нужного уровня содержания фитиновой кислоты. Величина снижения этого уровня может варьироваться от нескольких процентов до почти полного удаления фитиновой кислоты из ткани. Для опытного специалиста очевидно, что снижение содержания фитиновой кислоты, в особенности в некоторых культурных растениях, где ее содержание высоко, может потребовать более высокой степени экспрессии гена, модифицирующего мио-инозит, чем в тех культурах, где уровень фитиновой кислоты ниже. Управление степенью экспрессии генов различными способами широко известно на практике. В частности, такими способами являются введение последовательностей ДНК, которые ускоряют трансляцию, сильных промоторов, которые обеспечивают высокую степень транскрипции, или присоединение последовательностей ДНК, например присоединение к матричным или каркасным областям, в которых надежно обеспечивается высокая степень транскрипции трансгенов.
Данный вариант осуществления настоящего изобретения также предполагает получение растительных тканей, в частности семян, которые пригодны для производства кормов. В рамках настоящего изобретения предусматривается как прямое использование семян для кормовых целей, так и использование модифицированных кормов из обработанных семян. Снижение уровня фитиновой кислоты в семенах растений желательно для производства кормов, вследствие чего генетически модифицированные семена с измененным содержанием фитиновой кислоты найдут широкое применение в кормовой промышленности.
В некоторых случаях в качестве кормов для животных используется цельное зерно или зерно, прошедшее минимальную обработку, но не помол. Например, в качестве кормов с минимальной предварительной обработкой часто используются зерна кукурузы. Однако некоторые зерна кукурузы проходят определенную обработку, и продукты такой обработки, например клейковина, также включаются в рацион животных. Снижение уровня содержания фитиновой кислоты было бы полезно для обоих типов кормов из кукурузы, в особенности с точки зрения экологии, поскольку наличие фитиновой кислоты в кормах приводит к избыточному содержанию фосфора в экскрементах животных. Другие зерновые, используемые в качестве кормов, такие как пшеница, ячмень или овес, также могут быть серьезно улучшены благодаря настоящему изобретению. Следовательно, настоящее изобретение может быть использовано по отношению к большому количеству важных для сельского хозяйства культур. Ген, который в рамках описания настоящего изобретения был приведен в качестве примера изменения уровня содержания мио-инозита, может оказаться эффективным по отношению к любым культурным растениям, поскольку мио-инозит содержится во всех таких культурах и является единственным известным предшественником фитиновой кислоты. Таким образом, было хорошо продемонстрировано использование гена метилтрансферазы для изменения мио-инозита. Для опытного специалиста очевидно, что кодирующая область, либо определенная последовательность ДНК промотора, либо нетранслированная основная последовательность, терминатор и т.п. могут быть модифицированы с целью достижения оптимальной степени экспрессии гена в растительных клетках определенных растений. Все это находится в пределах профессиональных возможностей опытного специалиста в данной области. Авторы полностью уверены в том, что настоящее изобретение может применяться по отношению ко всем растениям, которые могут быть генетически трансформированы, включая те растения, семена которых используются непосредственно в кормовых целях, или же растения, семена которых перед использованием для приготовления кормов проходят определенную обработку.
Следовательно, в результате использования данного способа возможно получение цельных зерен, имеющих пониженное содержание фитиновой кислоты.
В дополнение к непосредственному использованию цельных или частично размолотых зерен в кормовых целях существуют многие другие способы получения зерновых кормов. Для получения клейковины, помимо прочих способов, используется мокрый помол зерновых. Помимо кормов из обработанных зерен, в соответствии со способом, предусмотренным настоящим изобретением, могут быть получены цельные зерна с пониженным содержанием фитиновой кислоты. Настоящее изобретение также может применяться для получения кормов на основе клейковины с пониженным содержанием фитиновой кислоты.
Кроме основных кормовых культур, таких как зерновые, масличные культуры также обеспечивают значительную долю производства кормов. Для масличных культур особое значение имеет получение кормов в процессе переработки их семян на масло. В общем случае масло получают из семян масличных культур либо с использованием растворителя, либо без такового. Жмых и прочие твердые отходы, получаемые в ходе этого процесса, используются для приготовления кормов, и обычно в них содержится основная часть фитиновой кислоты.
Наиболее распространенным способом переработки семян масличных культур является выделение масла с последующим восстановлением жмыха или других остаточных компонент семян. Настоящее изобретение найдет применение и в этом процессе, в результате чего получаемые таким образом корма будут более ценными с точки зрения их питательных свойств, чем обычные корма, благодаря пониженному содержанию фитиновой кислоты.
Следовательно, способ приготовления кормов с пониженным содержанием фитиновой кислоты дает возможность получить новые составы кормов, в настоящее время не получаемые из семян масличных культур. Основные масличные культуры, использование которых в кормовых целях будет более эффективным благодаря настоящему изобретению, включают в себя крестоцветные масличные культуры, такие как Brassica napus, Brassica capa, Brassica juncea, Brassica nigra, Brassica alba, Crambe, Eruca sativa и другие, которые также могут быть важными для промышленности и приготовления кормов. Некрестоцветные масличные культуры, которые часто используются для приготовления кормов, включают в себя соевые, хлопок, кукурузу, сафлор, подсолнечник и горох.
Настоящее изобретение предполагает использование генетически измененных семян масличных культур, полученных за счет экспрессии ферментов, способных оказывать воздействие на мио-инозит под контролем промотора, селективного по отношению к семенам растений. В результате этого уровень фитиновой кислоты в семенах таких растений снижается, и после их обработки получаются корма также с пониженным уровнем содержания фитиновой кислоты.
Нижеприведенные примеры иллюстрируют использование данного способа, но никоим образом не ограничивают область применения настоящего изобретения.
Пример 1: Общие способы, используемые для идентификации продуктов фенилпропаноидного обмена веществ: Определение содержания синапина - конечного продукта фенилпропаноидного обмена веществ - в семенах крестоцветных.
Очевидно, что опытный специалист в данной области может адаптировать иллюстрируемые ниже способы к определению содержания продуктов фенилпропаноидного обмена веществ, используя информацию, содержащуюся в многочисленных публикациях по аналитической химии. В первом примере использования данного способа на основе имеющихся в литературе способов иллюстрируется использование простого опыта по идентификации синапина в растительной ткани по отношению к ткани семян крестоцветных растений (Chappie, C.C.S., Т. Vogt, B.E. Ellis, C.R. Somerville, 1992, Plant Cell, 4, стр.1413-1424). Метод тонкослойной хроматографии (TLC) был настроен на определение продукта фенилпропаноидного обмена веществ - синапина. Этот метод основан на выделении растительного экстракта и нанесении известного количества синапина на кварцевые пластины (например, типа Whatman 60A SilicaG). Синапин очищают с использованием общеизвестных способов (например, D. Strack, 1977, Z. Pflanzenphysiol., 84, стр.139-145). Экстракт выделялся путем перемещения растворителя на основе n-бутанола, уксусной кислоты и воды в соотношении соответственно 10:2:3 на расстояние приблизительно 10 см от исходного положения. Экстракты из семян приготовляли путем вымачивания предварительно взвешенных образцов в течение одной ночи в плотно закрытой трубке, содержащей 100-500 мкл раствора метанола (метанол 98%, уксусная кислота 2%) с последующим добавлением такого же количества метанола и размолом. Надосадочная жидкость была получена путем центрифугирования со скоростью 12000 об/мин и экстрагирована смесью хлороформа и воды (400 мкл:100 мкл), центрифугирована, как описано выше, после чего липидная фаза была удалена, а водяную фаза высушивали на воздухе или в вакууме. Образцы растворяли в воде и наносили на пластинки тонкослойного хроматографа. Перед выдержкой в восходящем растворителе на эти пластинки наносили эмпирически определенное количество образца. Сухое взвешивание производили на предварительно высушенных образцах.
Пластинки тонкослойного хроматографа изучали в ультрафиолетовом излучении, в котором синапин имел вид блестящей капли. Предварительно очищенный образец использовали в качестве эталона, как в чистом виде, так и в виде искусственной смеси с экстрактом из зерен, для того чтобы убедиться, что очищенный синапин мигрировал вместе с синапином в экстракте из зерен в тех случаях, когда он присутствовал в сочетании с компонентами указанных экстрактов. При визуализации радиоактивно помеченного синапина пластинки тонкослойного хроматографа сканировали в сканере марки Ambis4000 фирмы Scanalytics, который позволяет количественно визуализировать источники радиации.
После этого содержание синапина уточняли методом жидкостной хроматографии высокого давления (HPLC). Аналогичные вышеописанным, но не содержащие экстракта воды и хлороформа метаноловые экстракты растительного материала подвергали HPLC-анализу на хроматографе фирмы Varian с колонной Nucleosil C18AB. Объем инъекции обычно составлял 10 мкл, а подвижная фаза представляла собой смесь растворителя А (2% уксусная кислота в воде) и растворителя В (2% уксусная кислота в ацетонитриле). В обычных условиях протекания в подвижной фазе раствора содержалось 90-80% растворителя А в течение 17 минут, после чего его содержание снижали до 10% на период 1 мин. В колонне содержание растворителя А составляло 10% в течение 4-х мин, после чего колонну промывали и балансировали раствором, содержащем 90% растворителя А. Данные условия могут варьироваться для ускорения отделения элюированных веществ. Например, содержание растворителя А может изменяться от 90% в первоначальный момент до 80% на период 15 минут, затем до 70% на период 15 минут, затем до 10% на 2 минуты, после чего выдерживаться еще 2 минуты на уровне 10% и уравновешиваться до уровня 90%. Поглощение ультрафиолетового излучения контролировали детектором на основе диодной матрицы. Для анализа образцов измеряли поглощение ультрафиолетового излучения на длине волны 330 нм. Строили эталонную кривую очищенного синапина, которая впоследствии использовали для оценки количества синапина в растительных экстрактах. Для опытных специалистов в данной области очевидно, что условия проведения HPLC-анализа (например, пропорции и градиенты растворителей и различные виды последних) могут варьироваться. Желательно определить условия, которые соответствуют используемому оборудованию и сложности образца. В литературе опубликовано большое количество вариантов метода HPLC для анализа фенольных соединений, включая синапин.
С помощью радиоактивных индикаторов получали более точную отметку времени начала синтеза синапина и определяли пропорцию вновь образованного синапина в различных частях растения. Метод радиоактивных индикаторов может также использоваться для определения способности отдельных частей семян растений (например, семядоли, завязи или оболочки) синтезировать синапин из внешних источников холина. Радиоактивно помеченные продукты могут быть после этого исследованы на содержание синапина, например, методом тонкослойной хроматографии с последующим сканированием на сканере Ambis4000.
Метод HPLC использовался для проведения количественного анализа общего содержания синапина в семенах и отдельных частях семян. Тонкослойную хроматографию использовали для количественного анализа содержания синапина на различных стадиях развития семян, а метод радиоактивных индикаторов в сочетании с TLC и последующим подсчетом радиоактивных точек использовался для определения момента начала синтеза синапина и его распределения в семенах и различных тканях семени, таких как семядолях, стержне и оболочке.
Метод радиоактивных индикаторов для определения момента начала синтеза синапина из холина проводился не на отдельных семенах, а на цельных стручках, для того чтобы как можно более адекватно воспроизвести в эксперименте условия живого растения. Стручки, находящиеся на различных стадиях развития, получали из растений, выращенных в контролируемых условиях, обычно при дневной температуре 20°С, ночной температуре 15°С, при цикле день/ночь в соотношении 16/8 часов. Стебельковую часть стручка погружали в раствор, содержащий холин, помеченный изотопом 14С, приобретенным у фирмы New England Nuclear (совокупная удельная активность 0,2 мКи/мл, концентрация 7,4 МБк/мл, что соответствует 0,2 мКи/мл, содержание холина 3,7 мкмоль/мл). Стручки, стебельковые части которых были смочены в трубке, содержащей 1,8 мл среды на водной основе, например среды Мурашига-Скуга с половинной силой и 1 мкмоль нерадиоактивного холина, а также 1,8-9 мкл радиоактивного холина, выращивали в инкубаторе в течение от 24 до 72 часов с циклом свет/темнота в соотношении 16/8 часов при температуре 20°С. Условия освещения составляли 51 микро-Эйнштейн на кв. м. в сек. Семена, полученные из этих стручков, подвергали анализу методом тонкослойной хроматографии. Из этих образцов получали экстракт в растворе метанола, а затем в экстракте хлороформа и воды, описанном выше.
При анализе более зрелых семян в дополнение к вышеуказанным методам применяли инфильтрацию семян, отделенных от стручков. 20 семян инфильтрировали в 500 мкл среды, содержащей радиоактивный холин, описанный выше, в низком вакууме в течение 15 мин при комнатной температуре 23°С. Затем радиоактивную среду удаляли и заменяли на 500 мкл нерадиоактивной среды вышеуказанного состава, за исключением холина с изотопом 14С, после чего проводили выращивание материала при 23°С в течение 24 часов. Экстракт из семян получали вышеописанным способом.
Растительный материал может быть также экстрагирован с помощью смеси хлороформа, метанола и муравьиной кислоты (CMF) в соотношении соответственно 5:12:3. Обычно добавляют 2 мкл CMF в расчете на 1 мг образца, образец размалывают и выдерживают при комнатной температуре (21-23°С) в течение ночи (16 часов), после чего центрифугируют со скоростью 12 000 об/мин в течение 5 минут. Надосадочную жидкость собирают и тщательно смешивают с аналогичным объемом CMF, выдерживают на протяжении 20 мин и центрифугируют, как описано выше. После этого всю надосадочную жидкость собирают в одной емкости, вышеуказанным способом приготавливают экстракт хлороформа и воды и водную фракцию анализируют, как описано выше.
Пример 2: Определение локализации синтеза продукта фенилпропаноидного обмена веществ в отдельной ткани.
В данном примере определяется время развития синтеза продукта фенилпропаноидного обмена веществ. Этот пример приведен для иллюстрации синтеза синапина и его накопления в тканях семян крестоцветных. Использовали развивающиеся семена Brassica napus. Тонкослойная хроматография экстрактов стручков, выращенных из опыленных вручную цветков, была проведена для определения времени начала накопления синапина. Образцы забирали через 7, 14, 16, 18, 20, 22, 24, 26, 28, 20, 32 и 34 дня после опыления, а также после созревания семян. Экстракты приготовляли способами, описанными в примере 1, а содержание синапина визуализировали с помощью метода TLC. Синапин проявлялся начиная с 18 дней после опыления до стадии созревания семян, а созревающие семена содержали наибольшее синапина, и, следовательно, в развивающихся семенах происходило чистое накопление синапина.
Ткань стенки стручка, с которого снимали семена, не содержала синапина, равно как его присутствие не было обнаружено в более молодых семенах, взятых из образцов на 7-й, 14-й и 16-й день после опыления. Анализ показал, что начало накопления синапина в семенах приходится примерно на 18-й день после опыления, однако, поскольку данный метод основан на регистрации ультрафиолетовой люминесценции, присутствие очень малого количества в более молодых семенах, если бы оно имело место, не могло быть обнаружено. На Фиг.3 показан пример TLC-анализа образцов до 28-го дня после опыления, а на второй пластинке показаны результаты анализа вплоть до созревания. Поскольку вышеприведенные результаты не были количественными, проводился HPLC-анализ экстрактов из цельных семян, получанных с помощью раствора метанола, как описано в примере 1. Результаты, представленные на Фиг.4, подтверждают, что синтез синапина начинается на 20-й день после опыления и постоянно усиливается вплоть до созревания. Количество синапина в зрелых семенах составляло 0,8% от сухого веса семян, а поскольку масло составляет до 45% массы семян, то в обезжиренном зерновом корме этот показатель составит 1,5%.
Пример 3: Определение временных и пространственных параметров синтеза продукта фенилпропаноидного обмена веществ - синтез синапина в зависимости от развития семян.
На основании информации, приведенной в примере 2, было показано, что синапин синтезируется в развивающихся семенах. Также было очевидно, что в процессе развития семян разложение синапина было минимально. Для подтверждения результатов этого эксперимента развивающиеся семена выращивали в присутствии холина, меченного изотопом 14С. Холин является предшественником синапина в его синтезе из синапоилглюкозы, который является завершающей стадией фенилпропаноидного обмена веществ в растениях. Присутствие радиоактивного предшественника приводит к образованию меченного синапина. Следовательно, можно количественно оценить биосинтез синапина. Как показано на Фиг.5, синтез синапина (внедрение 14С-холина в синапин) не наблюдается на протяжении 10 дней после опыления и впервые регистрируется на 14-й день после опыления. Синтез синапина (т.е. образование конечного продукта синапина из его предшественников - синапоилглюкозы и холина) происходит на 14-й день после опыления и вплоть до почти полного созревания. Соответственно, начало накопления синапина приходится на 14-й день после опыления, и развивающиеся семена продолжают вырабатывать и накапливать синапин вплоть до созревания.
Хотя нерадиоактивный анализ начинает регистрировать накопление синапина на 18-й день после опыления, радиационный эксперимент более чувствителен и позволяет регистрировать наличие биосинтеза даже в малых объемах. Следовательно, как показывает радиационный эксперимент, синтез синапина начинается в небольших количествах на 14-й день после опыления. Инфильтрационные эксперименты показали, что способность синтезировать синапин сохраняется и в более зрелых семенах (Фиг.6).
Помимо определения сроков накопления продуктов фенилпропаноидного обмена веществ опытному специалисту далее следует определить ткани, в которых происходит биосинтез данного продукта. В этой части примера приводится определение накопления синапина в семядоле, стержне и оболочке семян. Методом радиоактивных индикаторов исследовали цельные стручки возрастом от 18 до 42 дней после опыления. Стебельковую часть срезанных стручков погружали в раствор, содержащий холин, помеченный изотопом 14С, на 24-72 часа, и три вышеуказанные части семян разделяли и отдельно исследовали на содержание синапина. Результаты измерений приводятся на Фиг.7, где показано, что наибольшее количество синапина в расчете на одно семя содержится в семядоле, за ним по убывающей следует стержень и оболочка. Экстракты синапина, полученные из семядоли и стержня семян возрастом от 25 дней после опыления до созревания, показали, что в пересчете на сухой вес содержание синапина в стержне составило 50% от содержания синапина в семядоле (Фиг.8). Однако, поскольку масса семядоли значительно больше массы стержня (в шесть раз на момент созревания), содержание синапина в ней составляет до 90% всего синапина, содержащегося в семенах (Фиг.9).
Пример 4: Генетическая трансформация растения с геном, кодирующим фермент, способный воздействовать на предшественник фенилпропаноидного обмена веществ.
В данном примере фермент холиноксидаза (СОХ), который воздействует на предшественник синапина, проявляется в растительной клетке. Фермент холиноксидаза внедряется в вектор трансформации растения под контролем промотора, селективного по отношению к семенам. Холин является предшественником синапина в его синтезе из синапоилглюкозы, вследствие чего уменьшение запаса синапина снижает количество образующегося синапина.
Генетическая трансформация Brassica napus (рапс), являющегося крестоцветным растением, с помощью конструкции холиноксидазы (СОХ), селективной по отношению к семенам. Последовательность ДНК гена холиноксидазы показана на Фиг.10, а прогнозируемая последовательность аминокислот показана на Фиг.11. Чтобы экспрессия этого гена была селективной по отношению к семенам, использовали последовательность промотора напина (Kohno-Murase, J., М. Murase, Y. Ichikawa, J. Imamura, 1994, Plant Molecular Biology, 26, стр.1115-1124). Последняя плазмида pHS731 была сконструирована в результате серии клонирований и субклонирований в соответствии со стандартными методиками, описанными справочниках, например Maniatis, Т., Frittsch, E.F., Sambrook, J., 1982, Molecular Cloning: A Laboratory Manual (Лабораторный справочник по молекулярному клонированию), Cold Spring Harbor, New York. Основная цепь вектора начинается от RD400 (Datia, R.S.S., Hammerlindl, J.K., Panchuk, В., Pelcher, L.E., Keller, W., 1992, Gene, 211, стр.383-384), который в результате модификации содержит вместо маркера растений RD400 NosP-NptII ген слияния между gus и npt (Gus:npt). Последовательность Gus-npt была описана ранее (Datla, R.S.S., Hammerlindl, J.K., Pelcher, L.E., Crosby, W.L., G. Selvaraj, 1991, Gene, 101, стр.239-246). Последовательность Gus-npt была клонирована на фрагменте 2.5 Kb, который содержит терминатор Nos (сигнал polyA) на конце последовательности npt, из pGKK14 (Datla и др., 1991) в промежуточную плазмиду, потерявшую полностью Т-ДНК в RD400. Промотор CaMV35S с частичным дублированием, а также основная последовательность мозаичного вируса люцерны из pBI525 (Datia, R.S.S., F. Bekkaoui, J.K. Hammerlindl, G. Pilate, D.I. Dunstan, W.L. Crosby, 1993, Plant Science, 94, стр.139-149) были клонированы как фрагмент HindIII-BamHI на 5'-окончании последовательности Gus:npt. Сайты для HindIII, BamHI и соседние сайты XbaI были удалены с заполнением окончаний Klenow-фрагментом полимеразы I с образованием pHS722. Функциональная область lac-alpha, содержащая множество сайтов клонирования из pTZ19R (Mead, D.A., E. Szczesna-Skorupa, В. Kemper, 1986, Protein Engineering, 1, стр.67-74), была получена как фрагмент 0.26 Kb в результате реакции цепочки полимеразы. Этот фрагмент был изменен с помощью MunI и EcoRI и введен в сайт EcoRI pHS722 с образованием pHS723. Нуклеотидная последовательность участка, Т-ДНК вектора была расшифрована, и неповторяющиеся сайты во множественных сайгах клонирования были идентифицированы методом рестрикционного анализа. PHS725 является производной, полученной от pHS723, содержащей открытую рамку холиноксидазы, исходно относящейся к pKRIl (Rozwadowski и др., 1991), через серию промежуточных векторов, что привело к появлению удобных сайтов или таких особенностей, как сигнал сигнал Poly A мозаичного вируса цветной капусты. Вектор pHS725 обеспечивает наличие в фоновой холиноксидазе pHS723 открытой рамки считывания с сигналом PolyA от CaMV на 3'-окончании. 5'-участок открытой рамки считывания, содержащий промотор CaMV35S, был замещен на промотор напин из промежуточной плазмиды с образованием pHS731. Указанная промежуточная плазмида содержала производную pUC19, включающую в себя промотор напин (1.1 Kb) в виде кассеты HindIII-НапинР-BamHI, предоставленной Kohno-Murase (Kohno-Murase и др., 1994), которая была лигирована в виде вставки HindIII-BamHI в окно HindIII-BamHI плазмиды RD400 (Datla и др., 1992). В результате была получена плазмида pHS974. Кассета открытая рамка считывания BamHI-BADH -PolyA -KpnI была выделена в виде фрагмента 2.1 Kb из pRKJ6A (pRKJ6A был подробно описан в диссертации R.K. Jain, 1995, Genetic Engineering of Osmolyte Biosynthesis in Plants: Manipulation of Betaine Aldehyde Dehydrogenase Gene Expression Tobacco (Генная инженерия биосинтеза осмолита в растениях: управление экспрессией гена бетаинальдегиддегидрогеназы в табаке), Ph.D. Thesis, University of Saskatchewan, Saskatoon, Canada) и лигирована в окно BamHI-Kpn I плазмиды pHS974 с образованием вектора pHS981, который был использован при получении плазмиды pHS731. На Фиг.12 показана полученная в результате плазмида pHS731, которая содержит с правого и левого краев сегмент промотор напин - открытая рамка считывания СОХ - сигнал CaMV PolyA. Нуклеотидная последовательность сегмента промотор напин - открытая рамка считывания СОХ - сигнал CaMV PolyA была определена на основании ее компонент. Дополнительным подтверждением именно такой формы последовательности является то, что предшественник pHS723 содержит целиком pBIN19 (Bevan, M., 1984, Nucleic Adds Research, 12, стр.8711-8721), включая левый и правый края и всю область за их пределами. Полная нуклеотидная последовательность pBIN19 приводится в литературе. Следовательно, вышеописанная генная конструкция СОХ может быть легко получена для клонирования в другие векторы. Вектор плазмиды pHS731 в Е.Coli DH5 был внесен 22 января 1997 г. в Американскую коллекцию типовых культур (АТСС), расположенную по адресу 12301 Parklawn Drive, Rockville MD, USA 28052, под номером АТСС Designation 98300.
На основании опубликованной информации (DeLisle, A.J., Crouch, M.L, 1989, Plant Physiology, том 91, стр.617-623; Hoglund, A.-S., Т. Rodin, E. Larsson, L. Rask, 1992, Plant Physiology, том 98, стр.509-515), а также наших результатов по промотору напин, было предсказано, что экспрессия гена напина наступит на средней стадии развития семян рапса.
Вектор pHS731 был введен в штамм Agrobacterium MP90 стандартным методом тройного скрещивания с последующей трансформацией Brassicac Agrobacterium в качестве посредника. Данная трансформация была проведена в общих чертах, как описано в работе Moloney и др., Plant Cell reports, 8, стр.238-242.
Штамм Agrobacterium tumifaciens GV 3101/pMP90 (Koncz C., Schell, J., 1986, Mol. Gen. Genet., 204, стр.383-396), в котором содержится вектор pHS731, использовался для исследования генетических трансформаций. Стационарная фаза бактериальной культуры в питательной среде LB (Difco, США) в количестве 100 мл была собрана путем центрифугирования и повторно взвешена в 10 мл свежей питательной среды LB, содержащей 1% диметилсульфоксида (Sigma, США) в качестве криозащитного вещества. Дозы, кратные 200 мкл, содержали при температуре -20°С перед использованием для трансформирования, при котором одну кратную дозу бактериальной среды вводили в 2 мл питательной среды Brain Heart Infusion Broth (Difco, США), содержащей 2% сукрозы, 50 мкл ацетосирингона, рН 5,6, после чего проводили выращивание в течение ночи при 28°С. Плотность клеток бактерий составляла примерно 1×109 мл-1.
Искусственно культивированные семядоли подвергали воздействияю Agrobacterium, которые содержали вектор трансформации растения в соответствии с методом, описанным в работе Moloney и др.,. Plant Cell reports, 8, стр.238-242. После этого поверхность среза черешка выращенных семядолей на короткое время помещали в бактериальную культуру. Культивированные растения вводили в среду совместного культивирования таким образом, чтобы поверхности среза находились в контакте со средой. 10 растений были помещены в отдельные чашки Петри размером 100×15 мм. Чашки для совместного культивирования были запечатаны пластической оберткой марки Stretch'n Seal™. Чашки выдерживали в течение трех дней в камере для выращивания культур при температуре и цикличности освещения, описанных выше, для завершения стадии зародышеобразования в семенах. После этого ростки пересаживали в выделяющую среду.
Спустя 3-4 недели выдержки в выделяющей среде прорастающие зеленые побеги (предположительно трансформированные) удаляли и переносили в свежую выделяющую среду для дальнейшего выращивания. По достижении побегами размера 1,5-2,0 см их переносили в среду для пускания корней. Предположительно трансгенные побеги изолировали для экспрессии гена gus в основном так, как это описано в работе Jefferson, R.A., 1987, Plant Mol. Biol.Rep., 5, стр.387-405. Наличие синеватого окрашивания рассматривалось как доказательство трансформации.
Подтверждение трансформации было получено путем проведения опытов NPTII и PAT, Southerm blot-экспериментов, PCR (реакция цепочки полимеразы), а также анализом потомства. Трансгенные семена Brassica napus сорта Westar, содержащие вектор pHS731, ссылка №202622, были внесены 22 января 1997 г. в Американскую коллекцию типовых культур (АТСС), расположенную по адресу 12301 Parklawn Drive, Rockvilte MD, USA 28052, под номером АТСС Designation 97854.
После этого определяли экспрессию гена холиноксидазы. Регенерации были подвергнуты несколько независимых трансгенных линий. Наличие фермента СОХ показывали с использованием двойной ферментной реакции. Фермент СОХ образует из холина бетаинальдегид, который может быть легко обнаружен опытным путем его NAD-зависимого окисления ферментом BADH. Снижение содержания NAD обнаруживалось по изменению поглощения на длине волны 340 нм. Для стандартизации опыта используется препарат, содержащий различное количество промышленно производимого фермента СОХ (например, фирмой Sigma) и постоянное количество фермента BADH, получаемого из Е.coli (50 Ед, 1 Ед=1 нмоль восстановленного NAD в 1 мин на 1 мг белка) и получаемый из бактериального штамма с повышенной экспрессией BADH. В результате стандартизируются условия реакции и определяется ход эталонной кривой, которая ограничивается ферментом СОХ, а не BADH. После этого опыт проводится на растительных экстрактах, полученных из трансгенных линий. Существуют способы обогащения или очищения BADH (Falkenberg, P., Strom, A.R., 1990, Biochemica Biophysica Acta, 1034, стр.253-259).
Экстракты из растений получали следующим образом. Листья растений весом приблизительно 100 мг замораживали в жидком азоте и размалывали вместе с охлажденным до льда буферным экстрактом в пропорции двух объемов на вес образца. После этого образцы центрифугировали со скоростью 10 000 об/мин и надосадочную жидкость снова центрифугировали. Центрифугирование повторяли до тех пор, пока частицы твердой фазы не перестали обнаруживаться визуально. В случае семян (20 семян на образец) после первого центрифугирования добавляли 20 мг активированного угля, а затем процедуру доводили до завершения, как описано выше. Буферный экстракт с рН 8,0, доведенный КОН, содержит на 100 мл 1,92 г состава HEPES, 0,2 мл 0,5-М EDTA, 10 мл глицерина и деионизированную воду. Перед опытом добавляли DTT из общего состава концентрацией 1 М до конечной концентрации 25 мМ, а также полный набор ингибитора протеазы (Boehringer-Mannheim, Каталог: 1697-498) из общего состава 10Х в буфере HEPES. Буфер в ферментном опыте (BADH) содержал 50 мМ HEPES-KOH, рН 8,0, 1 мМ EDTA, а также свежий добавленный DTT до конечной концентрации 1 мМ. Дублирующий опыт проводился с помощью реакции, содержавшей 50 мкл буфера BADH, 50 мкл 10-мМ состава NAD, 50 мкл образца растения, 30 мкл или эквивалент 50 Ед BADH, а также деионизованную воду до общего объема 450 мкл. Реакционную смесь получали перемешиванием в течение 20 мин исходных, ранее проконтролированных компонент. Для достижения фоновых изменений в поглощении на длине волны 340 нм добавляли 50 мкл хлорида холина, после чего проводили спектрофотометрические измерения в течение 10-20 мин. Для получения эталонной кривой образец растения в смесь реагентов не вводился, а вместо него добавляли различные количества очищенного СОХ. Единицы рассчитывали с помощью программируемого регистрирующего спектрофотометра модели Beckman DU65, Опытный специалист в данной области сможет модифицировать данный эксперимент в соответствии с доступным оборудованием на основании биохимических данных, имеющихся в литературе по экстинкционному коэффициенту NAD. Восстановление NAD является стандартным опытом, проводимым с дегидрогеназами.
Пример 5: Анализ трансгенных семян на содержание фенолов.
В этом примере семена анализировали на содержание синапина. Семена трансгенных растений, полученных в примере 4, были пророщены для получения собственного потомства, и эти линии потомства анализировали на предмет наличия или отсутствия сегрегации фермента бета-глюкуронидазы (GUS), который кодирует трансген. Этот ген (Datia, R.S.S., Hammerlindl, J.K., Pelcher, L.E., Crosby, W.L, G. Selvaraj, 1991, Gene, 101, стр.239-246) присутствует в правом и левом краях Т-ДНК вектора рН5731 и, следовательно, выступает в качестве удобного маркера при генетическом анализе сегрегации. Отсутствие генетической сегрегации означало гомозиготную природу растений, от которого получено потомство. Те линии, которые демонстрировали сегрегацию, были гемизиготны, и сегреганты, не имеющие фермента GUS, сохранялись в них как контролирующие механизмы, не содержащие гена СОХ. Таким образом, в опыте присутствовали наборы гомозиготных линий, содержащих СОХ, и линий, не содержащих трансген. Эти линии анализировали на содержание синапина методом жидкостной хроматографии высокого давления. Результаты, представленные на Фиг.13, четко показывают, что трансгенные сегреганты имеют значительно сниженное содержание синапина в отличие от их нетрансгенных аналогов. Нетрансгенные сегреганты представляют собой наилучший механизм контроля, поскольку происходят из тех же исходно трансгенных растений. Данные результаты говорят о применимости описанного способа. Для опытного специалиста очевидно, что можно обеспечить разные величины снижения содержания синапина путем изменения степени, времени и локализации экспрессии гена, а также путем поиска наиболее благоприятных вариантов среди индивидуальных трансгенных линий, демонстрирующих желаемые результаты вследствие большого набора факторов, среди которых могут быть позиционные эффекты и порядковый номер/количество копии.
Пример 6: Получение стрессозащитных веществ путем исключения участия предшественника в фенилпропаноидном обмене веществ.
В этом примере стрессозащитное вещество бетаин получали с помощью комбинации ферментов СОХ и BADH под контролем промотора, селективного по отношению к семенам растений. В первой части данного примера получали трансгенный Brassica napus, содержащий ген BADH под контролем промотора, селективного по отношению к семенам. Проводили анализ этих растений на степень экспрессии BADH.
Экспрессия бетаинальдегиддегидрогеназы в семенах Brassica napus. Ген BADH, полученный из Е. coli (betB) выделяют и селективно проявляют в семенах путем сцепления с промотором напином в В. napus (Boyd и др., Gene, 103, стр.45-52, 1990). Плазмиду RD400 ((Datla, R.S.S., Hammerlindl, J.K., Panchuk, В., Pelcher, LE., Keller, W., 1992, Gene, 211, стр.383-384) использовали в качестве вектора, из которого получали конечную производную pHS974, содержащую в пределах своих правых и левых краев Т-ДНК открытую рамку считывания betB от своего сайта ATG под контролем промотора напина и сигнала PolyA мозаичного вируса цветной капусты. Кассета напин-betB-PolyA (3.3 Kbp) содержит в приведенном порядке сайт HindIII, промотор напин, сайт BamHI, betB ORF, EcoRI сайт - сигнал PolyA ДНК мозаичного вируса цветной капусты, сайт KpnI и сайт EcoRI. Промотор напин был предоставлен Kohno-Murase (Plant Molecular Biology, 26, стр.1115-1124, 1994), а сигнал PolyA исходно был получен из плазмиды pJIT 117 (Guerineau, F., Woolston, S., Brooks, L, Mullineaux, 1988, Nucleic Acids Research, 16, стр.11380). Конечная плазмида pHS981 (15 Kbp) представлена на Фиг.14. Эту плазмиду вводили в Agrobacterium tumefaciens GV3101 [рМР90] (Koncz, С., Schell, J., 1986, Molecular and General Genetics, 204, стр.383-396), и полученный в результате этого штамм использовался для генетической трансформации Brassica napus.
Было получено несколько трансгенных линий, и семена на различных стадиях развития анализировали на степень экспрессии фермента BADH. Данный параметр имел максимум около 35 дней после опыления, и у зрелых семян сохранялась остаточная активность фермента. Удельная активность фермента BADH в зависимости от стадии развития показывает, что начало экспрессии фермента BADH сопутствует синтезу синапина.
Линии, в которых проявлялся ген BADH, скрещивали с линиями, имевшими ген СОХ согласно примеру 4. Растения, содержащие ген СОХ, и растения, содержащие гены BADH и СОХ, анализировали на содержание синапина и общее содержание фенолов. Результаты представлены на Фиг.15 (содержание синапина) и Фиг.16 (общее содержание фенолов). Семена для анализа получали от растений, выращенных в полевых условиях летом 1999 г. Как показано на Фиг.15, совместная экспрессия генов СОХ и BADH приводит к дополнительному уменьшению накопления синапина по сравнению со случаем, когда проявлялся только ген СОХ. Как показано на Фиг.16, общее содержание фенолов в трансгенных растениях снижается по сравнению с эталонными.
Пример 7: Нуклеотидная последовательность синтетического гена декарбоксилазы феруловой кислоты.
В данном примере опубликованную ранее в литературе последовательность декарбоксилазы феруловой кислоты, полученной из Bacillus pumilus (Zago и др., Applied and Environmental Microbiology, 61, стр.4484-4486,1995), использовали для построения гена, оптимизированного для экспрессии в растительных клетках. Открытую рамку считывания декарбоксилазы феруловой кислоты синтезировали путем лигирования синтетических олигонуклеотидов в соответствии с опубликованной в литературе последовательностью. Олигонуклеотиды синтезировали в основном в соответствии с кодонной предпочтительностью сильно проявляющихся генов Brasses napus.
Синтетические олигонуклеотиды имели протяженность приблизительно 60 нуклеотидов. Структура олигонуклеотидных дуплексов включала в себя на 5'-окончании BamHI-когезивное окончание (5' GATC-) и когезивный терминатор EcoRI (3'-TTAA-5') на 3'-окончании. Собирали индивидуальные дуплексы, в результате чего была получена полномерная открытая рамка считывания с сайтами 5' BamHI и 3' EcoRI. Реакции лигирования проводили по стандартному варианту. Продукты лигирования, в свою очередь, лигировали в вектор клонирования.
При лигировании вышеуказанного синтетического гена с расщепленным BamHI-EcoRI pBlueprint SK-(Statagene) клоны со вставкой 0.5 Kb идентифицировали путем экранирования плазмиды ДНК от клонов Е.coli. Экранировали потенциальных кандидатов и определяли нуклеотидные последовательности двух из этих клонов. Было показано, что каждый из данных клонов имеет точечную мутацию, причем эти две точечные мутации в двух выбранных клонах были разнесены в пространстве. Данное пространственное разделение мутаций позволило реконструировать немутированный ген путем комбинации двух немутированных частей гена из этих клонов. Полученный в результате клон получил обозначение pGS97bl. Нуклеотидная последовательность этого синтетического гена представлена на Фиг.17. Прогнозируемая последовательность аминокислот представлена на Фиг.18.
Функциональность ситнетического гена проверяли с помощью простого эксперимента. Декарбоксилаза феруловой кислоты переводит феруловую кислоту в 4-винилгваякол (4-VG). 4-VG имеет отчетливый запах гвоздики и считается единственным из наиболее важных компонент, определяющих запах этого растения. Штамм Escherichia coli, содержащий pGS97bl, при выращивании в присутствии 1 мМ феруловой кислоты в среде выращивания дал отчетливый запах гвоздики, в то время как культура, выращенная без феруловой кислоты, или культура, содержавшая только данный вектор, не давали подобного запаха. HPLC-анализ культур, выращенных в присутствии феруловой кислоты, подтвердил удаление данной компоненты. На основании этих результатов был сделан вывод о том, что pGS97bl является функциональным клонированным геном.
Пример 8: Генетическая трансформация растения с геном, кодирующим декарбоксилазу феруловой кислоты под контролем нерегулируемого промотора.
В данном примере фермент декарбоксилаза феруловой кислоты, клонированный в pGS97bl, был введен в вектор трансформации растения RD400 (Datla, R.S.S., Hammerlindl, J.K., Panchuk, В., Pelcher, L.E., Keller, W., 1992, Gene, 211, стр.383-384), который был модифицирован таким образом, чтобы содержать вместо селекционного маркера растения NosP-NptII ген слияния между gus и npt (Gus:npt). Последовательность Gus-npt была описана ранее (Datla, R.S.S., Hammerlindl, J.K., Pelcher, L.E., Crosby, W.L, G. Selvaraj, 1991, Gene, 101, стр.239-246). Ген декарбоксилазы феруловой кислоты был введен под контролем промотора 35S, и полученную плазмиду использовали для трансформации образцов табака по стандартной процедуре.
Рестрикционная карта данного вектора представлена на Фиг.19. Вектор pGS97bl, представленный на Фиг.19, аналогичен вектору pHS731 со следующими исключениями. На той же основной цепи вектора pHS731 кассета HindIII-напинР-BamHI замещена кассетой промотор 35S мозаичного вируса цветной капусты - основная последовательность мозаичного вируса люцерны (описано в R.S.S. Datia, F. Bekkaoui, J.K. Hammerlindl, G. Pilate, D.I. Dunstan, W.L. Crosby, Improved High-Level Constitutive Foreign gene Expression In Plants Using an AMV RNA4 Untranslated Leader Sequence, Plant Science, 94, стр.139-149, 1993). Данный участок промотора на Фиг.19 сокращенно обозначается 35S. За 35S следует открытая рамка считывания ферулатдекарбоксилазы, связанная bamHI на 5'-окончании и EcoRI на 3'-окончании вместо открытой рамки считывания СОХ в pHS731.
Были выращены трансгенные растения, в которых проявляется ген декарбоксилазы феруловой кислоты. Эти растения были исследованы на содержание фенолов и вырабатывание винилгваякола.
В независимо выращенных трансгенных образцах табака, содержащих ген декарбоксилазы феруловой кислоты, при Western blot-анализе был обнаружен иммунореактивный полипептид ожидаемого размера с поликлональными антителами, вырабатывающимися в ответ на наличие декарбоксилазы феруловой кислоты. Нетрансформированные растения не содержали иммунореактивного пептида. Эти данные подтверждают трансгенность экспрессии белка декарбоксилазы феруловой кислоты в этих растениях.
Пример 9: Генетическая трансформация растения с геном, кодирующим декарбоксилазу феруловой кислоты под контролем тканеизбирательного промотора.
В данном примере фермент декарбоксилазы феруловой кислоты, клонированный в pGS97bl, был введен в вектор трансформации растения RD400. Ген декарбоксилазы феруловой кислоты был введен под контролем промотора напина, селективного по отношению к семенам, полученного из В. napus, и полученную плазмиду использовали для трансформации образцов табака по стандартной процедуре. Рестрикционная карта данного вектора представлена на Фиг.20. Вектор pGS97b2, представленный на Фиг.20, аналогичен вектору pGS97b3, за исключением того, что на той же основной цепи вектора кассета HindIII-35S-BamHI замещена кассетой HindIII-промотор напин-BamHI, показанной на Фиг.12 для pHS731. Были выращены трансгенные растения, в которых проявлялся ген декарбоксилазы феруловой кислоты. Семена растений исследовали на содержание фенолов и вырабатывание винилгваякола.
Пример 10. Определение хода накопления фитиновой кислоты и избирательности этого процесса по отношению к ткани в развивающихся семенах крестоцветного растения Brassica napus.
Биосинтез фитиновой кислоты.
В данном примере была получена информация о биосинтезе фитиновой кислоты в семенах Brassica. Поскольку фитиновая кислота является гексафосфатной производной мио-инозита (фитиновая кислота обозначается IP6), в эксперименте определяли пропорции индивидуальных фосфорилированных производных компонент мио-инозита (т.е. IP1, IP2, IP3, IP4 и IP5) из общего количества фосфорилированных производных инозита. Чистую фитиновую кислоту, частично деградированную методом автоклавной обработки для получения различных фосфорилированных производных, использовали в качестве эталона для HPLC-анализа, проведенного в соответствии с работами Vernon, Bohnert, 1992, The EMBO Journal, 11, стр.2077-2085 и Vemon DM, Tarczynski MC, Jensen RG, Bohnert HJ, 1993, The Plant Journal, 4, стр.199-205.
Различные формы фосфорилированного инозита (т.е. IP1, IP2, IP3, IP4 и IP5), а также фитиновая кислота были легко различимы данным методом. При анализе образцов инозитфосфата, полученных из развивающихся семян Brassica, был зарегистрирован только один максимум, относящийся к IP6 (фитиновой кислоте). Этот результат говорит о том, что IP6 является основным инозитфосфатом, присутствующим в развивающихся семенах, и что другие промежуточные формы фосфорилированного инозита методом жидкостной хроматографии высокого давления не регистрируются. Следовательно, биосинтез фитиновой кислоты из мио-инозита, очевидно, происходит достаточно быстро и продуктивно.
Биосинтез фитиновой кислоты в процессе развития
В данной части примера определялся период развития семян, когда происходит накопление фитиновой кислоты. Способ определения содержания фитиновой кислоты в семенах следующий. Семена измельчают в ступке в присутствии жидкого азота, и получаемый порошок помещают в стерильную 15-мл трубку, содержащую 5 мл 0,5 М HCl. После удаления липидов при экстрагировании гексаном остающуюся фазу (водную среду с тканевым осадком) обрабатывают ультразвуком в течение 90 сек на 3-м уровне с использованием прибора для обработки жидкостей ультразвуком (модель XL2020, Heat Systems, Inc., Farmingdale, NY, USA). После центрифугирования жидкость помещают в чистую трубку для HPLC-анализа на содержание фитиновой кислоты.
Накопление фитиновой кислоты в процессе развития семян иллюстрируется на Фиг.21. Несмотря на то что фитиновая кислота впервые обнаруживается в семенах на достаточно ранних стадиях развития (12 дней после опыления), ее содержание практически не увеличивается вплоть до 22 дней после опыления. В течение 10 дней после своего первого появления в семенах фитиновая кислота достигает наивысшего уровня приблизительно 240 мкг на семя. В единицах сухого веса содержание фитиновой кислоты в зрелых семенах составляет примерно 3,2%. Эти результаты говорят о том, что снижение содержания фитиновой кислоты при наличии ее биосинтеза требует введения промотора, проявляющегося начиная примерно с 12 дней после опыления практически до созревания семени.
Дальнейшие исследования показали, что фитиновая кислота в основном накапливается в зародышевой ткани, а не в оболочке семени (Фиг.22). Семядоля содержит почти 90% фитиновой кислоты, присутствующей в семени, и 10% находится в зародышевом стержне. Эти данные дополнительно свидетельствуют о том, что различные зародышевые ткани семян являются наиболее подходящими для снижения содержания фитиновой кислоты путем генетической модификации биосинтеза фитиновой кислоты.
Пример 11. Определение миозитного обмена веществ в тканях развивающихся семян.
В данном примере иллюстрируется та часть запаса мио-инозита, которая используется для биосинтеза фитиновой кислоты. Хотя мио-инозит является предшественником фитиновой кислоты в растениях, он также участвует и в других анаболических механизмах, в которых образуются фосфатидинозит и компоненты клеточных стенок. Пример касается той части запаса мио-инозита, которая используется для биосинтеза фитиновой кислоты в развивающихся семенах. Меченый 3Н мио-инозит в семенах Brassica использовался для отслеживания in vivo распределения инозита в различных частях, экстрагированных с помощью различных растворителей. Способ импульсного маркирования in vivo развивающихся семян с помощью мио-инозита, меченого 3H, был следующим. Стручки, находящиеся на различных стадиях развития, удаляли с растений и срезы немедленно погружали в стерильную культурную среду объемом 10 мл, содержащую мио-инозит, меченый 3H с дозой 5 мкКи, в 50-мл трубках и проращивали при стандартных условиях роста (цикл 20°С в течение 16 ч при свете, 15°С в течение 8 ч без света) в течение 2 дней. Семена собирали для экстрагирования липидов, фитиновой кислоты, компонент клеточных стенок, растворимых в трифтороуксусной кислоте (TFA), а также отходов клеточных стенок. Уровень радиоактивности определялся отдельно для каждой части путем жидкой люминесценции.
Было сделано четыре типа экстрактов из импульсно-меченых семян для разделения компонент клеточных стенок, растворимых в воде (для анализа на фитиновую кислоту), растворимых в гексане (для анализа на липиды), растворимых в трифтороуксусной кислоте (TFA), а также отходы клеточных стенок. На Фиг.23 приводится список средних уровней радиоактивности каждой части на разных стадиях развития семян. Из этих данных следует, что более 20% общего количества индикатора в семенах находится в липидной фракции через 25-30 дней после опыления. Радиоактивность фракции клеточных стенок (клеточные стенки, растворимые в TFA и отходы клеточных стенок) составляла примерно 5% общего количества индикатора, накопившегося в семенах в процессе их развития. Радиоактивность воднорастворимой фракции исследовали методом HPLC для определения содержания фитиновой кислоты. Было обнаружено, что примерно 10% индикатора в воднорастворимом экстракте присутствует в максимуме фитиновой кислоты и около 30% в максимуме инъекции образца, соответствующем чистому мио-инозиту. Остальная часть индикатора, присутствующая в воднорастворимом экстракте, проявлялась в максимумах перед и после максимума фитиновой кислоты, идентифицировать которые по химической принадлежность не удалось.
На Фиг.23 представлены доли усвоенной меченой компоненты в фитиновой кислоте, липидах, клеточных стенках, растворимых в трифтороуксусной кислоте, и отходах клеточных стенок. Около 30% индикатора в продуктах мио-инозитного обмена веществ присутствует в фитиновой кислоте между 20 и 30 днями после опыления. При этом около 60% индикатора присутствует в липидах (фосфолипиды, содержащие инозит) и менее 10% в клеточных стенках.
Следовательно, часть общего запаса мио-инозита, используемая для биосинтеза фитиновой кислоты, составляет около 30% в период развития семян, когда синтез фитиновой кислоты происходит наиболее активно.
Пример 12. Клонирование гена, кодирующего фермент, способный воздействовать на мио-инозит.
В данном примере был выделен ген, способный воздействовать на мио-инозит. Этот ген представлял собой ген инозит-оксиметилтрансферазы, полученный из ледяника обыкновенного. Для выделения гена нижеописанным способом использовали клонирование обратной транскриптазой. Стандартные манипуляции с ДНК выполняли в соответствии с работой Maniatis и др., Molecular Cloning, A Laboratory Manual (Лабораторный справочник по молекулярному клонированию). Cold Spring Harbor, MY.
Общее количество ДНК экстрагировали из тканей листьев ледяника, обработанных в 500 мМ NaCl, после чего производили очистку Poly (A) + РНК. Эту мРНК использовали для обратной транскрипции с помощью обратной транскриптазы Superscript II (Promega, Madison, WI.) при следующих условиях. 3 мкг мРНК, растворенной в 20 мкл воды, использовали в качестве шаблона. РНК денатурировали при температуре 65°С в течение 5 мин с последующим охлаждением на льду. К смеси добавляли 3 мкл олиго-оТ (500 мкг/мл), 8 мкл буфера обратной транскриптазы 5Х, 4 мкл 0,1 М DTT и 2 мкл 10 мМ dNTPs. Смесь нагревали до 42°С, после чего вводили 3 мкл (60 единиц) обратной транскриптазы Superscript II. Реакцию проводили в течение 1 часа при 42°С. После этого добавлялся 1 мкл Rnase H (1,5 Ед./мкл) и реакцию продолжали в течение 30 мин при 37°С. Полученную в результате кДНК использовали в качестве шаблона для PCR-анализа при следующих условиях. Реакцию цепочки полимеразы проводили в объеме 100 мкл циклами 94°С в течение 1 мин, 55°С в течение 1 мин и 72°С в течение 1 мин с 2-сек удлинением каждого цикла и общим числом циклов 30. Праймеры, использованные в данной реакции, выбирали на опубликованной в литературе последовательности ДНК IMT (W банка генов М87340). В качестве прямого праймера использовали последовательность I.D. №6:

которая включает в себя сайт ВатН I на 5'-окончании (подчеркнут).
В качестве обратного праймера использовали последовательность I.D. №7:

которая включает в себя сайт Not I на 5'-окончании (подчеркнут).
Амплифицированный фрагмент ДНК усваивался с помощью ВатН I и Not I, после чего субклон ировался в вектор pSPORT I (BRL, Bethesda, MD.). Фрагмент цепочки полимеразы, усвоенный усваивался с помощью BamH I и Not I, клонировался в соответствующие сайты вектора pSPORT для получения pSportIMT. На Фиг.24 (последовательность I.D. №5) представлена последовательность амплифицированного фрагмента ДНК, идентичного опубликованной в литературе последовательности ДНК IMT, за исключением двух базовых различий в нетранслированном 3'-участке. Предсказанная белковая последовательность соответствует опубликованной информации, которая может быть получена из банка генов.
Пример 13: Построение вектора трансформации растения с геном, кодирующим фермент, способный воздействовать на мио-инозит: Использование гена мио-инозит-оксиметилтрансферазы ледяника.
В данном примере иллюстрируется построение вектора трансформации растения, содержащего ген, который кодируюет фермент, способный воздействовать на мио-инозит под контролем промотора, проявляющегося в растительных клетках. Приводимый в качестве примера вектор трансформации растения содержит ген мио-инозит-оксиметилтрансферазы под контролем промотора, проявляющегося в растительных клетках (35S). Построение вектора проводили следующим образом: на вектор pSportIMT воздейстовали с помощью Ват HI и Eco RI для высвобождения гена IMT. Фрагмент гена IMT клонировался в соответствующие сайты pBluescript SK (-) (Strategene, La Jolla, CA.), и полученную в результате плазмиду обозначали pBlueIMT. Плазмиду pBlueIMT метаболизировали с помощью Spe I и клонировали в вектор pBI 221 (CloneTech), предварительно расщепленный Xba I. Плазмиду, которая содержала ген IMT с правильной ориентацией относительно промотора 35S в pBI 221, выбирали и метаболизировали с помощью Hind III и Eco RI. Фрагмент терминатора 35S-IMT-Nos переносился в вектор pRD 400, в результате чего образовывался вектор трансформации растения p35SIMT.
Полученная в результате конструкция p35SIMT содержит кассету терминатора 35S-IMT-GUS-Nos в pRD400 и представлена на Фиг.25.
Пример 14: Построение вектора трансформации растения с геном, кодирующим фермент, способный воздействовать на мио-инозит под контролем промотора, селективного по отношению к семенам.
В данном примере в векторе трансформации растения построен ген, кодирующий фермент, способный воздействовать на мио-инозит под контролем промотора, селективного по отношению к семенам. Использованный ген был ген мио-инозит-оксиметилтрансферазы, а в качестве промотора был использован промотор напин, селективный по отношению к семенам. Полученный вектор обозначен pNIMT.
Вектор pNIMT строился следующим образом: фргамент ДНК IMT, усвоенный с помощью Spe I (пример 4), был лигирован в pDHl на сайт Xba I, в результате чего образовалась кассета напин-IMT-Nos. Данную кассету впоследствии вводили в pRD400. Вектор, полученный в результате этого, представлен на Фиг.26.
Пример 15: Трансформация Brassica napus (Westar) с помощью p35SIMT.
Вектор p35SIMT вводили в штамм Agrobacterium MP90 методом стандартного тройного скрещивания, после чего Brassica подвергали трансформации с Agrobacterium в качестве посредника. Трансформацию проводили так, как описано в примере 4. Растения, трансформированные с помощью p35SIMT, анализировали на содержание фитатов.
Пример 16: Молекулярный анализ трансгенных растений.
Трансгенные растения F0, содержащие кассету CaMV 35S-IMT, исследовали методами PCR, Southern blot, Northern blot, а также в опыте с ферментом IMT. Предварительный PCR-анализ показал, что все трансгенные растения содержали ген IMT (Фиг.27). Southern blot-гибридизация показала, что ген IMT интегрирован в геном Brassica с различным числом/номером копий. Общая РНК была экстрагирована из развивающихся растений для анализа методом северной гибридизации, как показано на Фиг.28.
Пример 17: Анализ трансгенных растений на фитиновую кислоту.
Содержание фитиновой кислоты определяли в зрелых семенах, полученных с трансгенных растений F0. Экстракты из семян получали аналогично примеру 10. Полученные данные говорят о том, что среднее количество фитиновой кислоты снижается в трансгенных растениях более чем на 15%. На Фиг.29 и 30 представлен обзор полученных данных. Среднее снижение содержания фитиновой кислоты в растениях, выращенных с помощью вектора pSIMT, составляет 15%. Семена F1 представляют собой смесь трансгенных и нетрансгенных семян, полученную в результате сегрегации, поэтому реальное снижение уровня фитиновой кислоты в пересчете на только трансформированные семена значительно выше. Семена F2 и F3 представляют собой гомозиготные линии, содержащие вектор pSIMT. Очевидно, что в полевых условиях гомозиготные линии, содержащие вектор pSIMT, демонстрируют снижение содержания фитиновой кислоты в семенах в среднем на 30%. Кроме анализа на фитиновую кислоту проводился Southern blot-анализ с целью определения количества копий введенных генов. Даже в растениях с единственной введенной копией гена наблюдалось значительное снижение содержания фитиновой кислоты (34%, растение №ТР 11). Следовательно, экспрессия фермента, способного модифицировать мио-инозит в тканях, ответственных за биосинтез фитиновой кислоты, приводит к снижению содержания фитиновой кислоты в семенах. Также очевидно, что признак пониженного содержания фитиновой кислоты может быть передан половым путем, поскольку он передавался по наследству. Таким образом, практический способ может включать в себя стандартные технологии разведения, если уже получено растение с низким уровнем фитиновой кислоты.
Пример 18: Трансформация Brassica napus с помощью pNIMT.
Вектор p35SNIMT вводился в штамм Agrobacterium MP90 методом стандартного тройного скрещивания с последующей трансформацией Brassica с Agrobacterium в качестве посредника. Трансформацию проводили аналогично примеру 4.
Растения, трансформированные с помощью вектора p35SNIMT, анализировали на пониженное содержание фитиновой кислоты аналогично примеру 17. На Фиг.31 представлена таблица данных трансгенных семян F1 и F2, содержащих вектор pNIMT, выращенных в полевых условиях. В растениях F1 вставка pNIMT сегрегирует, поэтому исследованные семена были смесью трансгенных и нетрансгенных сегрегантов. В поколении F2 большинство линий были гомозиготными или практически гомозиготными. При анализе семян F2, выращенных в полевых условиях, наблюдалось практически 40%-ное снижение содержания фитиновой кислоты. Таким образом, экспрессия фермента, способного уменьшать доступное количество мио-инозита для биосинтеза фитиновой кислоты селективно по отношению к ткани растения, приводит к значительному снижению содержания фитиновой кислоты.
Пример 19: Предотвращение образования и накопления UDP-галактозы с помощью фермента, являющегося гетерологическим по отношению к растительным клеткам.
Фермент UDP-галактоза-4-эпимераза (gaLE) участвует в одной из основных стадий реакции галактозного обмена веществ в живых системах. Данный фермент выступает в качестве катализатора превращения UDP-галактозы в UDP-глюкозу. Ген данного фермента может быть получен из организма человека, дрожжей и бактерий.
Целью опыта, иллюстрируемого этим примером, было достижение чрезмерно сильной экспрессии данного фермента в определенных тканях растения для того, чтобы увеличить запас UDP-глюкозы за счет запаса UDP-галактозы. Предполагалось, что в результате этого будет снижена интенсивность биосинтеза галактинола, который является предшественником нежелательных с точки зрения питательных свойств гликозидов сукрозы (глюкозидов RFO, например, рафинозы, стахиозы и т.п.). Достижение этой цели, помимо снижения скорости накопления нежелательных глюкозидов RFO, позволит увеличить доступное количество UDP-глюкозы и сукрозы. Как ожидается, сукроза будет участвовать, ускоряя их, в других механизмах обмена веществ, в которых она необходима либо для образования других продуктов обмена веществ, которые важны для растения, либо как источник углерода для увеличения продуктивности растения (например, уровень белков, липидов, урожайность и т.п.).
Данный пример приводится для иллюстрации эффективности использования бактериального фермента в высших растениях для метаболического превращения одной важной в другую, в результате чего образующийся субстрат будет активно использоваться растением в большом количестве важных метаболических превращений.
Пример 20: Изменение уровней сахарных производных с использованием фермента фосфоглюкомутазы (pgm).
Фермент фосфоглюкомутаза (pgm) является катализатором взаимного превращения глюкозы (Glc)-l-и (Cic)-6-фосфата в процессе синтеза и потребления сукрозы. Данный фермент играет ключевую роль в синтезе и потреблении сукрозы, крахмала и глюкогена и присутствует во всех живых организмах. Ген данного фермента может быть получен из большого количества эукариотических или бактериальных источников (например, Agrobacterium).
G-1-P и G-6-P являются важными веществами для большого количества основных процессов углеводного обмена веществ во всех живых системах. В частности, G-1-P является основным субстратом для образования UDP-глюкозы, которая, в свою очередь, является основным субстратом для биосинтеза сукрозы. С другой стороны, G-6-P является основным исходным материалом для различных взаимных преобразований сахаров, одно из которых - синтез мио-инозит-1-Р. Это соединение является основным субстратом и одним из факторов в синтезе соответственно фитиновой кислоты и RFO.
Целью опыта, иллюстрируемого в данном примере, было показать, что путем достижения чрезмерно сильной экспрессии бактериального гена pgm в растении можно манипулировать соотношением G-1-P и G-6-P в пользу той или иной фосфорилированного формы глюкозы (независимо от ткани растения). Правильно направляя действие фермента, например в развивающихся семенах, можно ожидать повышения содержания G-1-P, что было бы желательно с точки зрения образования UDP-или ADP-глюкозы и впоследствии сукрозы и других накапливаемых веществ, в частности белков, липидов или крахмала. С другой стороны, эффектом от снижения содержания G-6-P стало бы снижение уровней вышеуказанных нежелательных с точки зрения питательных свойств факторов.
Пример 21: Получение и регенерация трансгенных клеток кукурузы.
Культуры каллус 2-го типа получают из незрелых зиготных эмбрионов генотипа Hi-II (Armstrong и др., 1991, Maize Genet. Coop. Newslett., 65, стр.92-93). Незрелые эмбрионы изолируют в возрасте около 14 дней после опыления от початков, выращенных в теплицах от гибридов Hi-II типа А и Hi-II типа В, или же эмбрионов F2, полученных путем самоопыления или опыления сестринских растений Hi-II. Незрелые эмбрионы размером 1,5-3,5 мм выращивали на инициирующей среде, содержащей соли и витамины N6 (Chu и др., 1978, The N6 Medium and Its Application to Anther Culture of Cereal Crops (Среда N6 и ее применение в культуре зернового пыльника), Proc. Symp. Plant Tissue Culture, Peking Press, стр.43-56), 1,0 мг/л 2,4-дихлорофеноксиуксусной кислоты (2,4-D) 25 мМ L-пролина, 100 мг/л казеингидролизазы, 10 мг/л AgNO3, 2,5 г/л состава GERLITE (Schweizerhall, South Plainfield, NJ) и 20 г/л сукрозы (рН 5,8). Через 4-6 недель каллус пересаживают на поддерживающую среду (инициирующая среда без AgNO3 и с содержанием L-пролина, пониженным до 6 мМ). Селекция каллуса 2-го типа производится в течение 12-16 недель.
Для бластинга, или введения чужеродной ДНК в клетки растения (трансформация путем бомбардировки микрочастицами), 140 мкг ДНК плазмиды (например, 35S-PAT - промотор глобулин из кукурузы - IMT) осаждают на 60 мг сферические частицы золота, промытые в спирте (диаметр 1,5-3 мкм, Aldrich Chemical Co., Inc., Milwaukee, WI) путем добавления 74 мкл 2,5 М CaCl2 Н2О и 30 мкл 0,1 М спермидина (свободная основа) к 300 мкл ДНК плазмиды и воды. Непосредственнор после этого полученный раствор перемешивали со встряхиванием, и частицы золота с осажденными ДНК оседали. Получаемую в результате чистую надосадочную жидкость удаляют и частицы золота снова помещают в 1 мл абсолютного этанола для получения взвеси. Данную взвесь разбавляют абсолютным этанолом для получения раствора 15 мг золота с осажденными ДНК на 1 мл.
Использованная ДНК плазмиды в предпочтительном варианте содержит выборочный индикатор, такой как ген bar или pat, что придает растениям сопротивляемость фосфинотрицину или гербицидглюфозинатаммонию, а также генетическую конструкцию, способную изменить вторичный обмен веществ, такую как кодирующая область гена IMT, описанная на Фиг.24, или кодирующая область гена декарбоксилазы феруловой кислоты, описанная на Фиг.17, под контролем подходящего промотора, селективного по отношению к семенам, например промотора глобулина 1 из кукурузы или промотора зеина из кукурузы. В альтернативном варианте для достижения экспрессии гена IMT или декарбоксилазхы феруловой кислоты может использоваться нерегулируемый промотор, такой как, например, убихитин из кукурузы или актин из риса.
Примерно 600 мг ткани эмбриогенного каллуса распределяют по поверхности поддерживающей среды каллуса 2-го типа, не содержащей казеингидролизазу и L-пролин, но с добавлением 0,2 М сорбитола и 0,2 М маннита в качестве вещества, придающего осмотические свойства. Каллус выдерживают на этой среде в течение 4 часов в качестве предварительной обработки и после этого переносят на чашки для культур, содержащие бластинговую среду (осмотическая среда, сгущенная 20 г/л агара ТС (Phytotechnology Laboratories, LLC, Shawnee Mission, KS) вместо 7 г/л состава GERLITE. Гелий (бластинг) используют для ускорения движения взвешенных частиц золота с нанесенными ДНК по направлению к тканям, подлежащим трансформации. Используемый прибор описан в Патенте США №5141131, который включен в данное описание в качестве ссылки. Ткани покрывают изолирующей прокладкой из нержавеющей стали (отверстия размером 104 мкм) и помещают в частичный вакуум (635 мм рт.ст.) в камере прибора. Перед бластингом частицы золота с нанесенными ДНК разбавляют абсолютным этанолом в пропорции 1:1 и их движение в момент достижения каллуса ускоряется приложенным давлением гелия 1500 фунтов на кв. дюйм (102 атм), причем с каждым бластингом к ткани подводится 20 мкл суспензии ДНК на золотых частицах. Непосредственно после бластинга ткань помещают в осмотическую среду на 16-24 часа для восстановления. После этого ткань разделяют на мелкие части и помещают в выделяющую среду (поддерживающая среда, не содержащая казеингидролизазы и L-пролина, но содержащая 30 мг/л состава BASTA фирмы AgrEvo, Берлин, Германия). В течение 3 месяцев образцы ткани каждые 4 недели неселективно переносятся в новую выделяющую среду. По истечении 7 недель и до 22-й недели секции каллуса, размножающиеся на фоне ткани с ингибированным ростом, удаляют с нее и изолируют. Получаемую в результате ткань, обладающую сопротивляемостью к составу BASTA, еженедельно пересаживают на новую выделяющую среду. Соответствующие исследования помогают идентифицировать положительные трансгенные линии для последующего перенесения на регенеративную среду.
Регенерацию начинают с помещения ткани каллуса в индуктивную среду на основе цитокинина, состоящую из солей Мурашиге-Скуга (MS) и витаминов (Murashige, Skoog, 1962, Physiol. Plant., 15, стр.473-497), 30 г/л сукрозы, 100 мг/л мио-инозита, 30 г/л маннитола, 5 мг/л 6-бензиламинопурина (ВАР), 0,025 мг/л 2,4-дихлорофеноксиуксусной кислоты (2,4-0), 30 мг/л состава BASTA и 2,5 г/л состава GERUTE при рН 5,7. Культуры помещают под слабый свет (125 кандел-фут) на 1 неделю, после чего на одну неделю под сильный свет (325 кандел-фут). По истечении двухнедельного периода индуктирования ткани неселективно переносятся в регенерирующую среду, не содержащую гормонов, которая аналогична индукционной среде, за исключением того, что она не содержит 2,4-D и ВАР, и содержатся под сильным светом. Небольшие (1/5-3 см) ростки удаляют и помещают в кубы для разведения культур размером 150×25 мм, содержащие среду SH (соли и витамины SH в соответствии с работой Schenk, Hilderbrandt, 1972, Can. J. Sot, 50, стр.199-204, 10 г/л сукрозы, 100 мг/л мио-инозита, 5 мл/л FeEDTA и 2,5 г/л состава GERUTE, при рН 5,8).
Подросшие ростки помещают в 12-сантиметровые горшки, содержащие приблизительно 0,25 кг среды METRO-MIX 360 (The Scotts Co., Marysville, ОН) в теплице сразу после того, как они начинают расти и развивают достаточную для жизнедеятельности корневую систему. Выращивание производится с 16-часовым суточным световым периодом, освещение обеспечивается натриевыми и галогенными лампами высокого давления, поливка производится по мере надобности водой, содержащей сочетание трех независимых формул удобрений Peter Excel (Grace-Sierra Horticultural Products Company, Milpitas, CA). На стадии образования 6-8 листьев растения пересаживают в горшки объемом 22,7 л, содержащие примерно 4 кг среды METRO-MIX 360, и выращивают до стадии зрелой плодовитой трансгенной кукурузы. Семена, полученные от этих растений, содержат гены, введенные в незрелые эмбрионы, и при вырастании растений приводят к экспрессии белков, кодированных введенной ДНК.
Пример 22: Получение трансгенного риса.
Для получения эмбриогенного каллуса зрелые семена культурного сорта Japonica, Taipei 309 очищают, а поверхность их стерилизуют в 70% этаноле в течение 2-5 мин, после чего проводится их пропитка в течение 40-45 мин в 50% промышленном отбеливающем растворе (2,6% гипохлорит натрия) с добавлением нескольких капель мыла Liquinox. После этого семена промывают 3 раза в стерильной дистиллированной воде и помещают на фильтровальную бумагу перед переносом в среду индуктирования каллуса (среду NB). Среда NB состоит из макроэлементов N6 (Chu и др., 1978, The N6 Medium and Its Application to Anther Culture of Cereal Crops (Среда N6 и ее применение в культуре зернового пыльника), Proc.Symp. Plant Tissue Culture, Peking Press, стр.43-56), микроэлементов и витаминов В5 (Gamborg и др., 1968, Nutrient Requirements of Suspension Cultures of Soybean Root Cells (Требования к питательной среде для суспензионных культур клеток корней сои), Exp. Cell res., 50, стр.151-158), а также 300 мг/л казеингидролизазы, 500 мг/д L-пролина, 500 мг/л L-глютамина, 30 г/л сукрозы, 2 мг/л 2,4-дихлорофеноксиуксусной кислоты (2,4-D) и 2,5 г/л состава GERUTE (Schweizerhall, NJ), с рН, доведенным до 5,8. Зрелые семена, культивированные на среде индуктирования каллуса, выращивают в темноте при 28°С. После 3-недельного выращивания появляющийся первичный каллус из щитковой области зрелого эмбриона помещают в свежую поддерживающую среду NB.
Биолистическая трансформация растительной ткани служит для введения чужеродной ДНК. Около 140 мкг ДНК плазмиды осаждают на золотые частицы размером 1 мкм (Bio-Rad) общим весом 60 мг, как описано в примере 21. Используют плазмиду, содержащую промотор убихитин из кукурузы, который активизирует hpt (гидромицинфосфотрансферазу) и убихитин 1 из кукурузы, глобулин 1 из кукурузы, либо промотор зеин, который активирует ген IMT. Около 140 мкг ДНК плазмиды осаждают на золотые частицы размером 1 мкм (Bio-Rad) общим весом 60 мг, как описано в данном примере.
Для гелиевого бластинга активно растущие культуры эмбриогенного каллуса размером 2-4 мм подвергают обработке сильным веществом, придающим осмотические свойства. Такая обработка включает помещение каллуса на среду NB с 0,2 М маннита и 0,2 М сорбитола (Vain и др., 1993, Osmoticum Treatment Enhances Particle Bombardment-Mediated Transient and Stable Transformation of Maize (Ускорение переходной и стационарной трансформации маиса с промежуточной бомбардировкой частицами путем обработки веществом, придающим осмотические свойства), Plant cell Rep., 12, стр.84-88) на период 4 часов перед гелиевым бластингом. После обработки веществом, придающим осмотические свойства, культуры каллуса помещают в бластинговую среду (NB + 2% агар) и покрывают прокладкой из нержавеющей стали (230 мкм). Культуры каллуса подвергают бластингу с давлением гелия 2000 фунтов на кв. дюйм (136 атм) по два раза на каждую мишень. После бластинга каллус помещают обратно в среду с сильным веществом, придающим осмотические свойства, на ночь перед высаживанием на выделяющую среду, которая состоит из среды NB с 30 мг/л гидромицина. По истечении 2-х недель культуры помещают в свежую выделяющую среду с более высокой концентрацией выделяющего реагента, т.е. NB + 50 мг/л гидромицина (Li и др., An Improved Rice Transformation System Using the Biolistic Method (Улучшенная система трансформации риса с использованием биолистического метода), Plant Cell Rep., 12, стр.250-255).
Плотные эмбриогенные культуры каллуса, имеющие бело-желтую окраску, восстановленные на среде NB + 50 мг/л гидромицина, регенерировали в пререгенерирующей среде (PR) + 50 мг/л гидромицина. Среда PR состоит из среды NB с 2 мг/л бензиламинопурина (ВАР), 1 мг/л нафталинуксусной кислоты (NAA) и 5 мг/л абсцизиновой кислоты (АВА). После двухнедельного выдерживания культуры в темноте каллус переносился в регенерирующую среду (RN). Состав среды RN - среда NB с 3 мг/л ВАР и 0,5 мг/л NAA. Культуры каллуса выращивали на среде RN в течение 2-х недель при температуре 28°С под сильным люминесцентным светом (325 кандел-фут). После того как образовывающиеся ростки достигали длины 2 см, они пересаживались в коробки цвета маренго, содержащие 1/2 среды МС (Murashige, Skoog, 1962, A Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Cultures (Измененная среда для быстрого роста и биологических опытов на культурах тканей табака), Physiol. Plant., 15, стр.473-497 с 1/2 витаминов В5, 10 г/л сукрозы, 0,05 мг/л среды NAA, 50 мг/л гидромицина и 2,5 г/л состава GERLITE, рН 5,8). Крупные растения длиной 8-15 см с хорошо развитой корневой системой пересаживали в почву (metromix: верхняя почва=1:1) и выращивали в теплице (температурный цикл 29/24°С для дня/ночи, влажность 50-60%, суточный световой период 12 ч.). Ростки риса развиваются и дают продуктивный урожай трансгенного риса. Семена, полученные от этих растений, содержат гены, введенные в каллус риса, и при вырастании в зрелые растения приводят к экспрессии в них белков, кодированных введенной ДНК.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИНАРНЫЙ ВЕКТОР pMOG413, БИНАРНЫЙ ВЕКТОР pMOG429, СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ИЛИ РАСТИТЕЛЬНЫХ ОРГАНОВ, СОДЕРЖАЩИХ УВЕЛИЧЕННОЕ КОЛИЧЕСТВО ФИТАЗЫ (ВАРИАНТЫ), И КОРМОВАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ) | 1991 |
|
RU2128228C1 |
| СЕМЕНА ТРАНСГЕННЫХ РАСТЕНИЙ С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ЛИЗИНА | 2006 |
|
RU2460281C2 |
| МОДИФИЦИРОВАННЫЕ ФИТАЗЫ | 2003 |
|
RU2329301C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОЛОГИЧНОГО БЕЛКА В ПРОРАСТАЮЩИХ СЕМЕНАХ ДВУДОЛЬНЫХ РАСТЕНИЙ | 2000 |
|
RU2273665C2 |
| (ЦИТОХРОМ Р450)-МОНООКСИГЕНАЗЫ | 1998 |
|
RU2237714C2 |
| ПРОМОТОР ЦЕЛОГО СЕМЕНИ | 2010 |
|
RU2561463C2 |
| ГЕНЫ-МИШЕНИ ДЛЯ НАПРАВЛЕННОГО ВОЗДЕЙСТВИЯ НА АЗОТФИКСАЦИЮ ДЛЯ УЛУЧШЕНИЯ КАЧЕСТВ РАСТЕНИЙ | 2018 |
|
RU2805085C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЛИПИДОВ | 2012 |
|
RU2656996C2 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И СПОСОБЫ ПОЛУЧЕНИЯ МАСЛА С ИЗМЕНЕННЫМ СОСТАВОМ ИЗ СЕМЯН | 2007 |
|
RU2392795C2 |
| Способы получения липидов | 2011 |
|
RU2636344C2 |
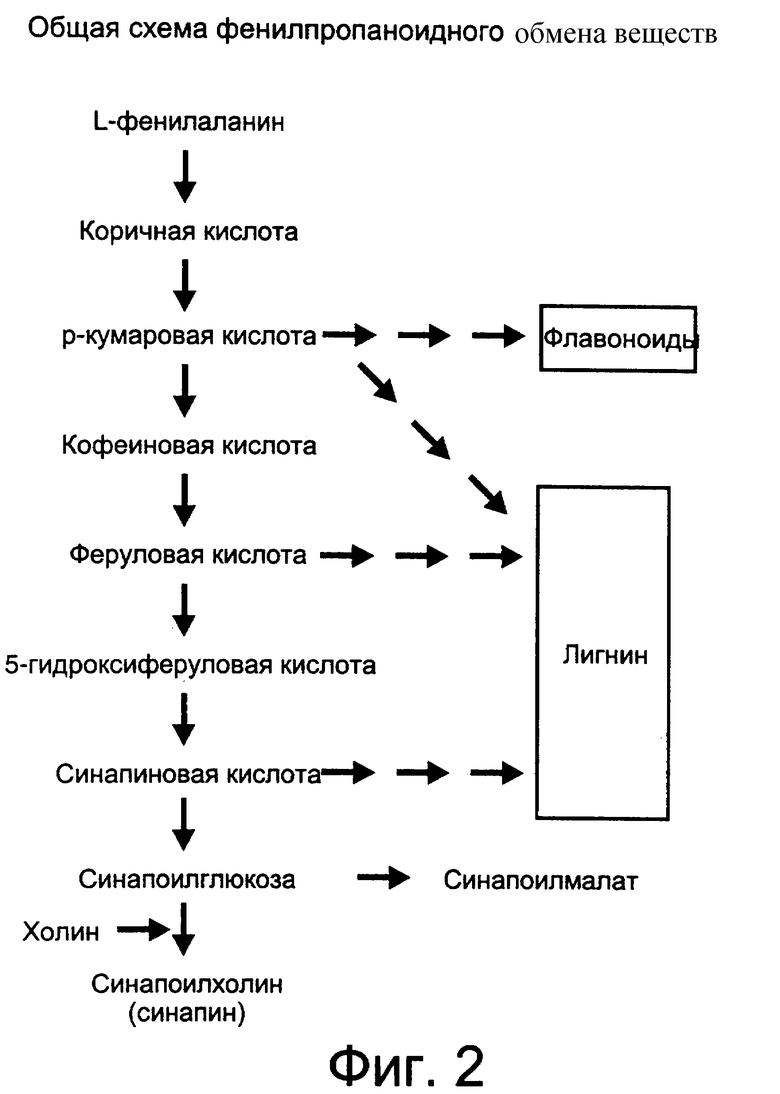
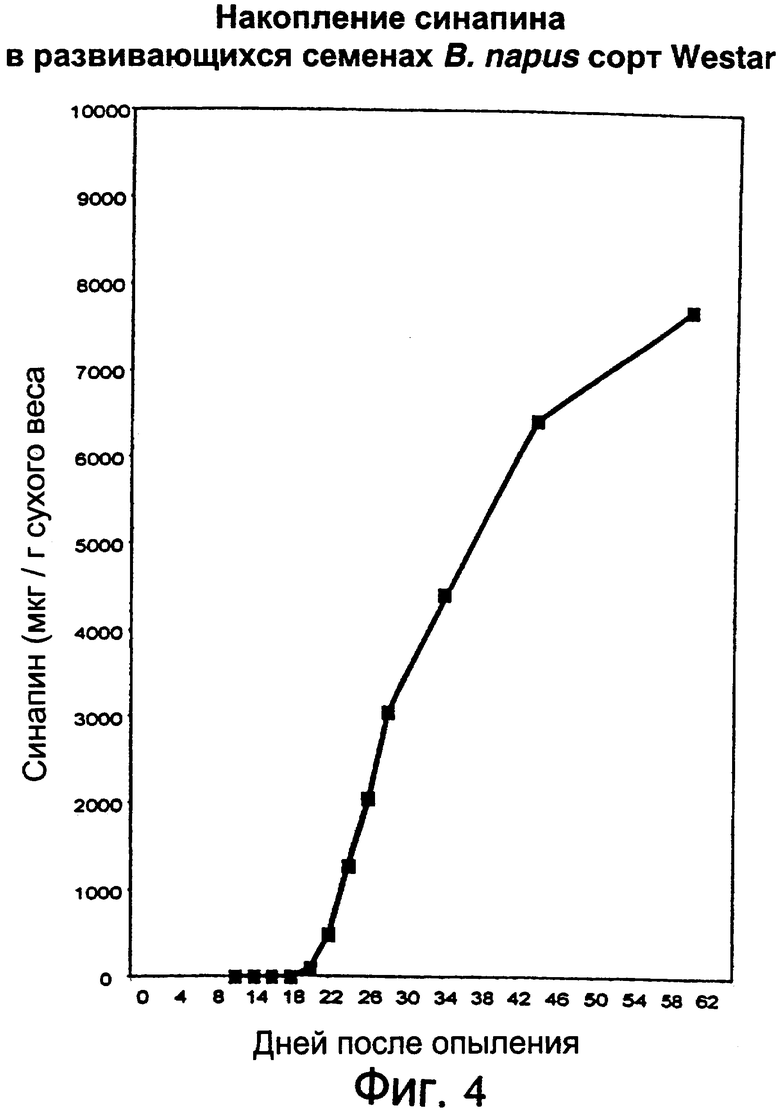
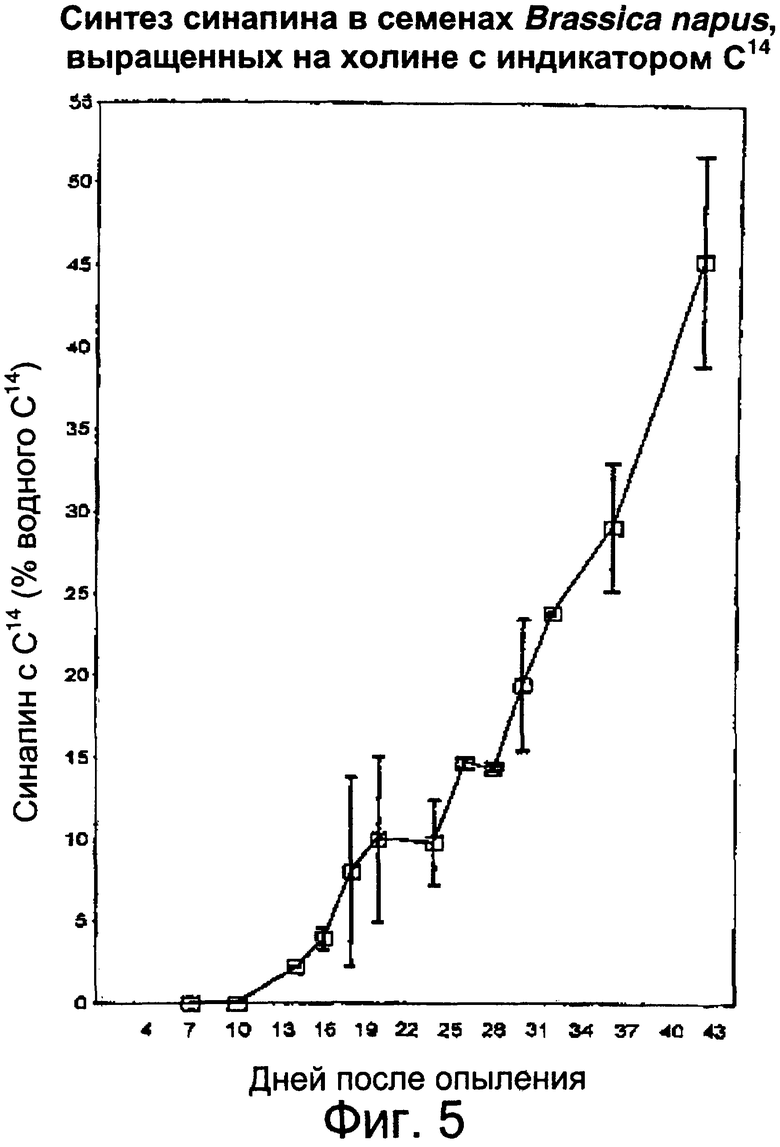
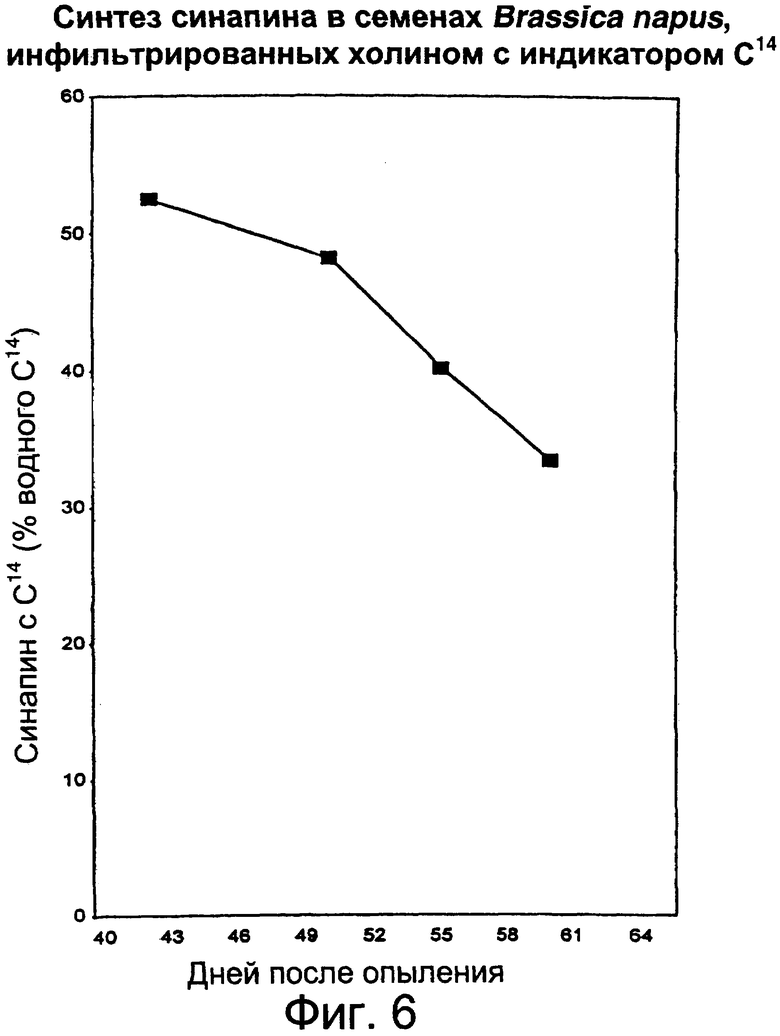
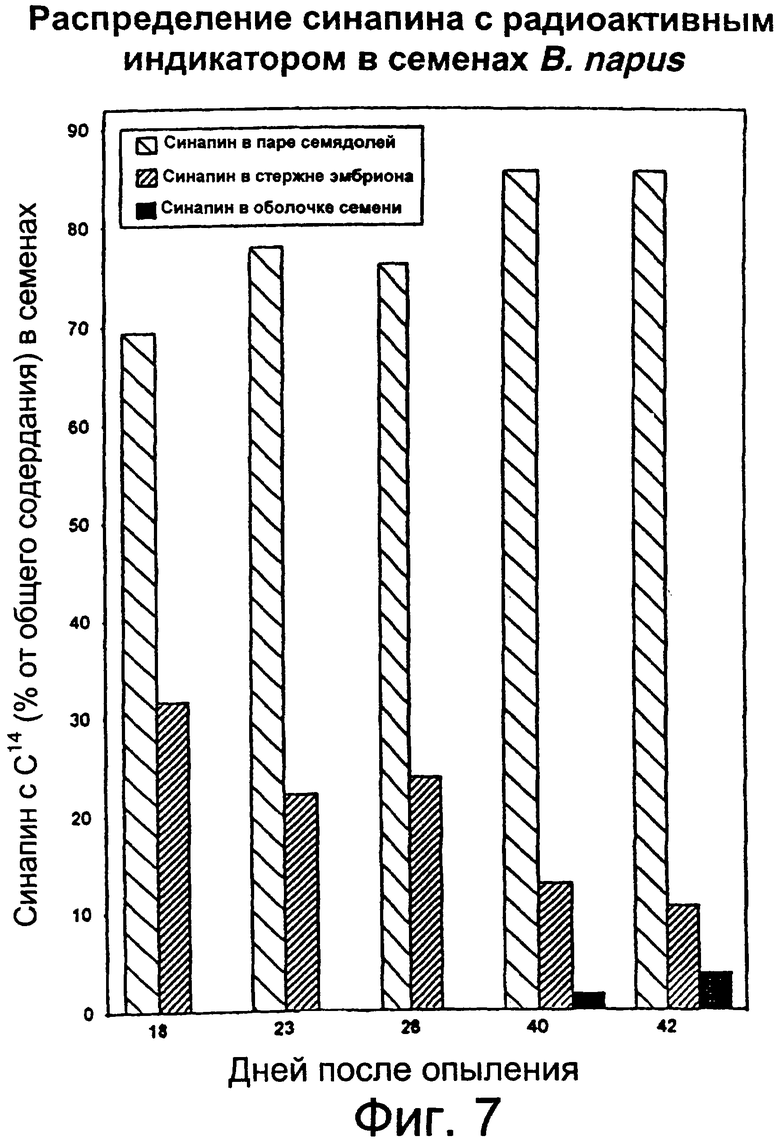
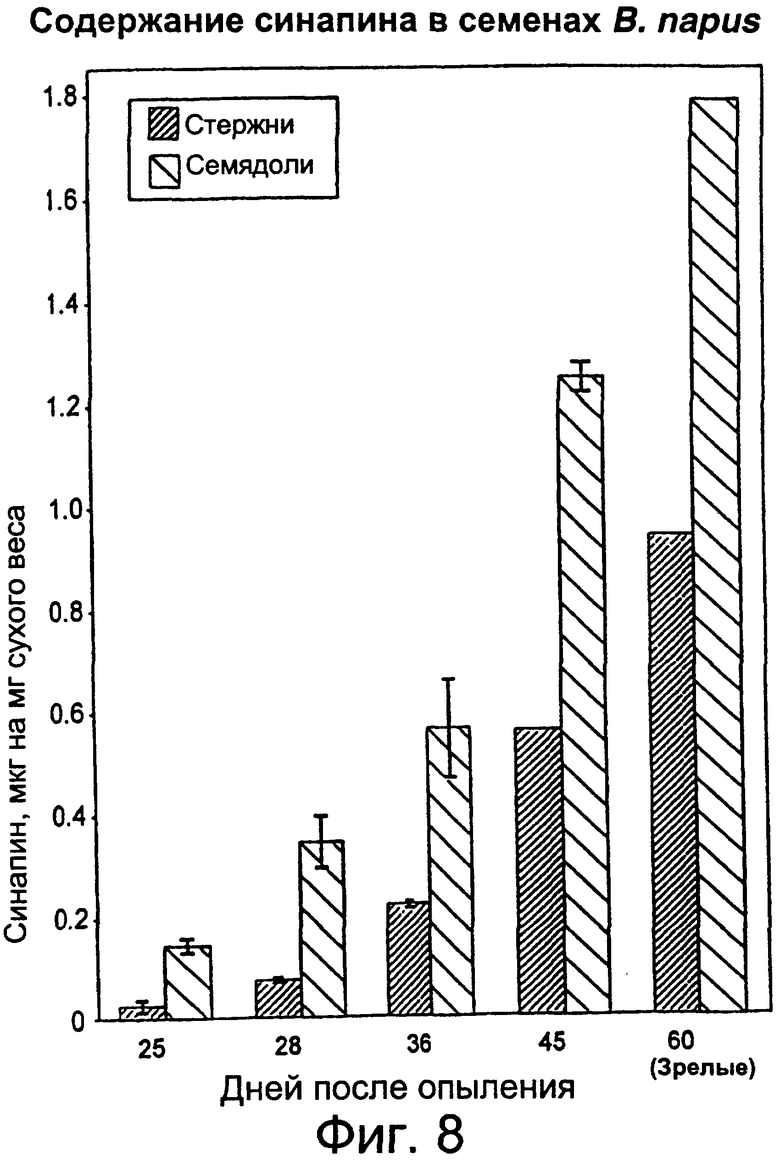
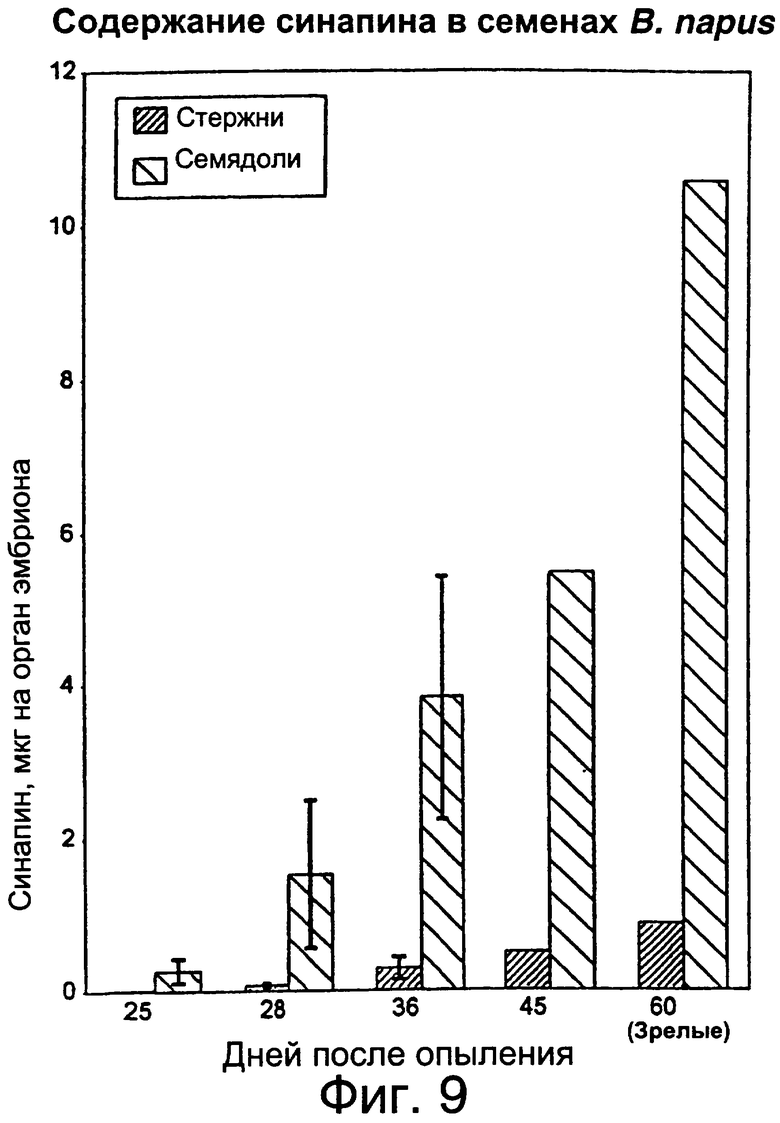

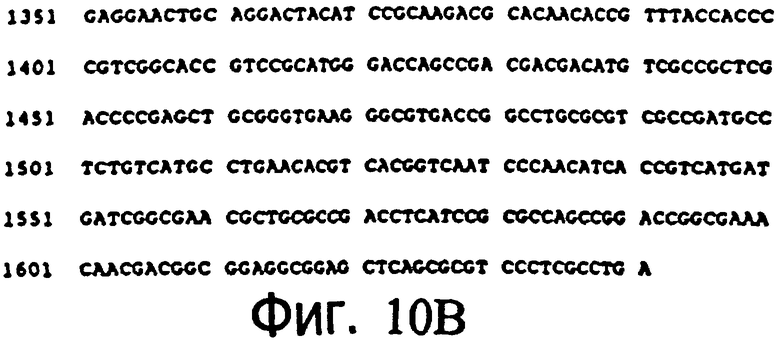

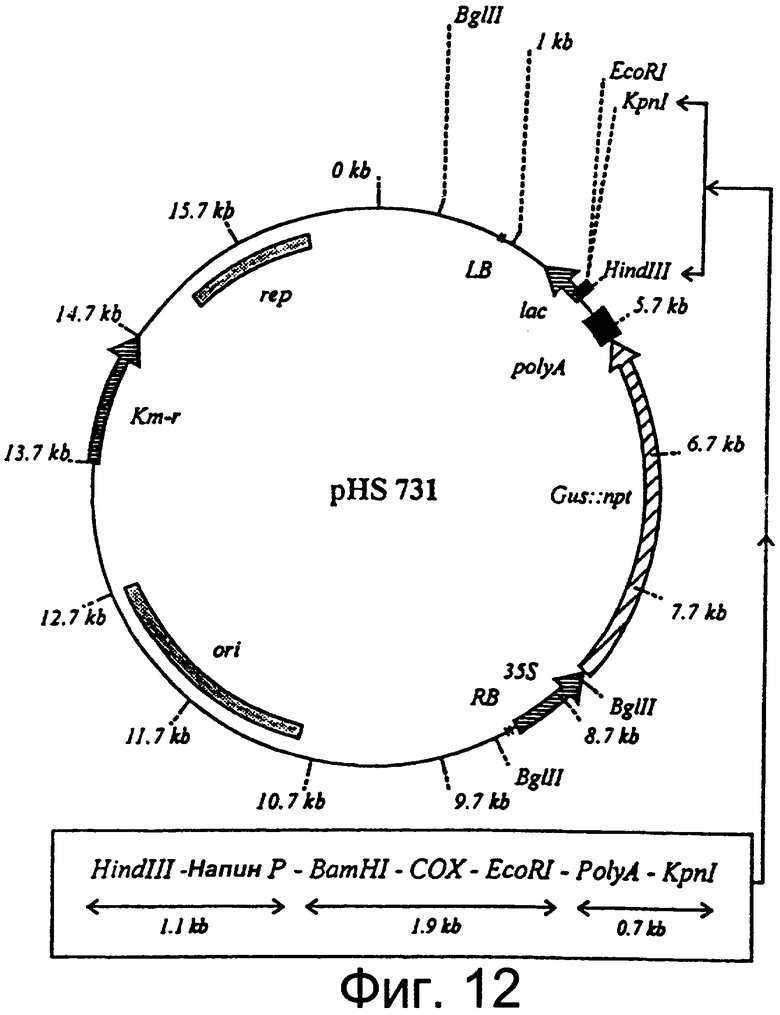
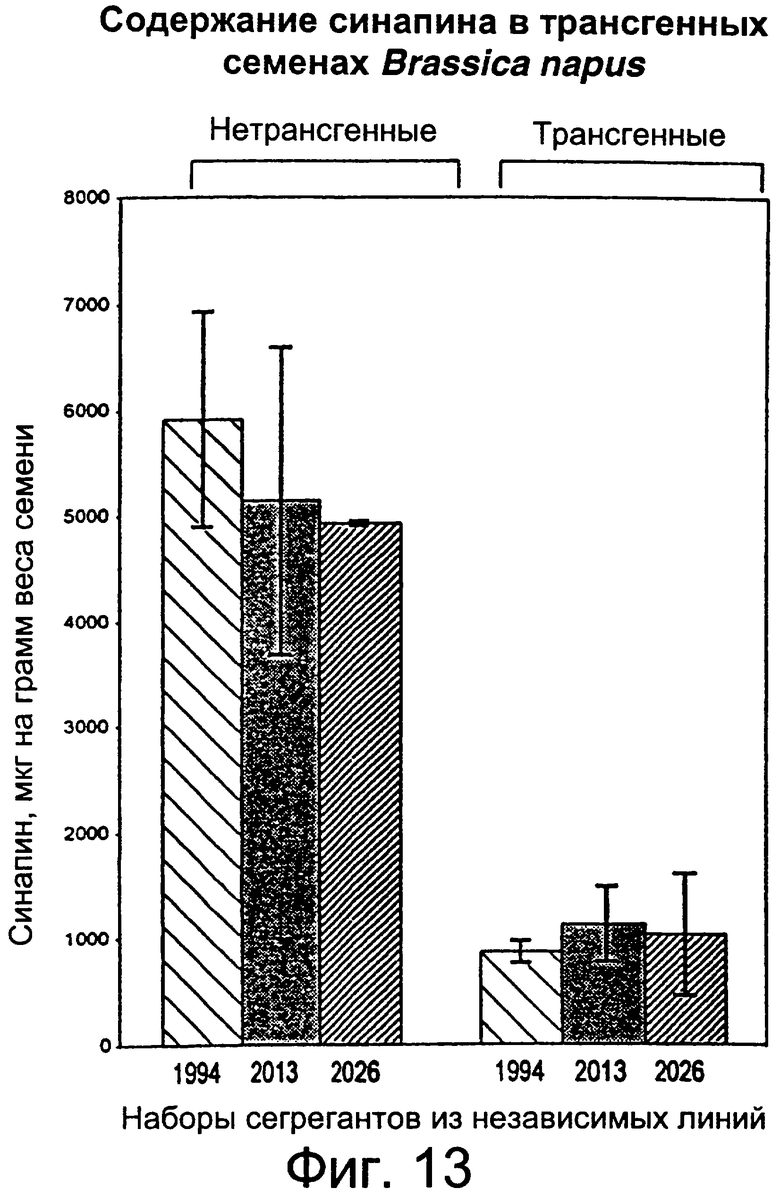
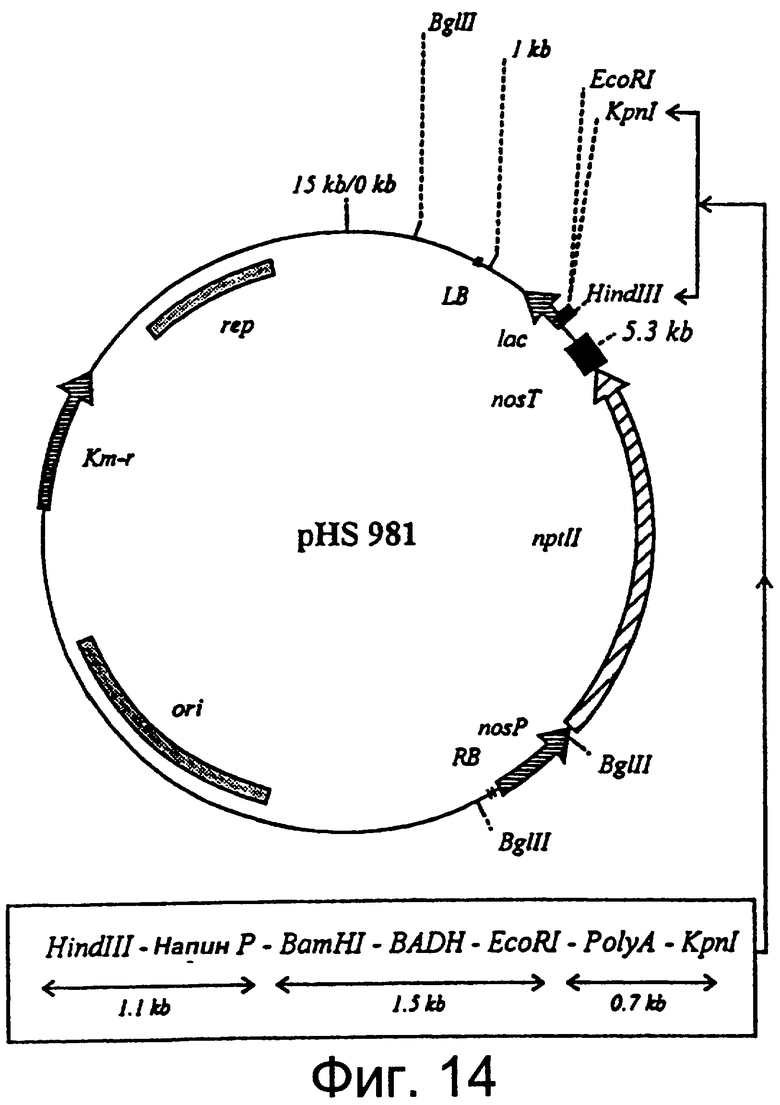
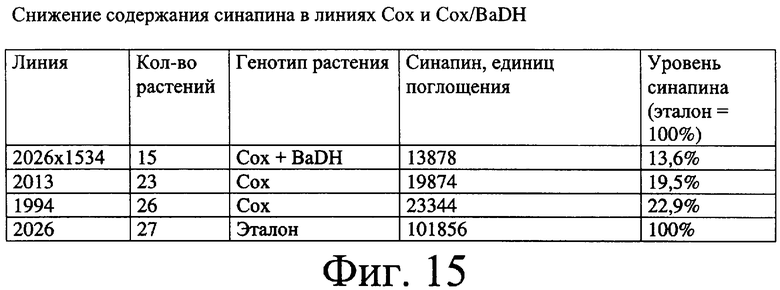



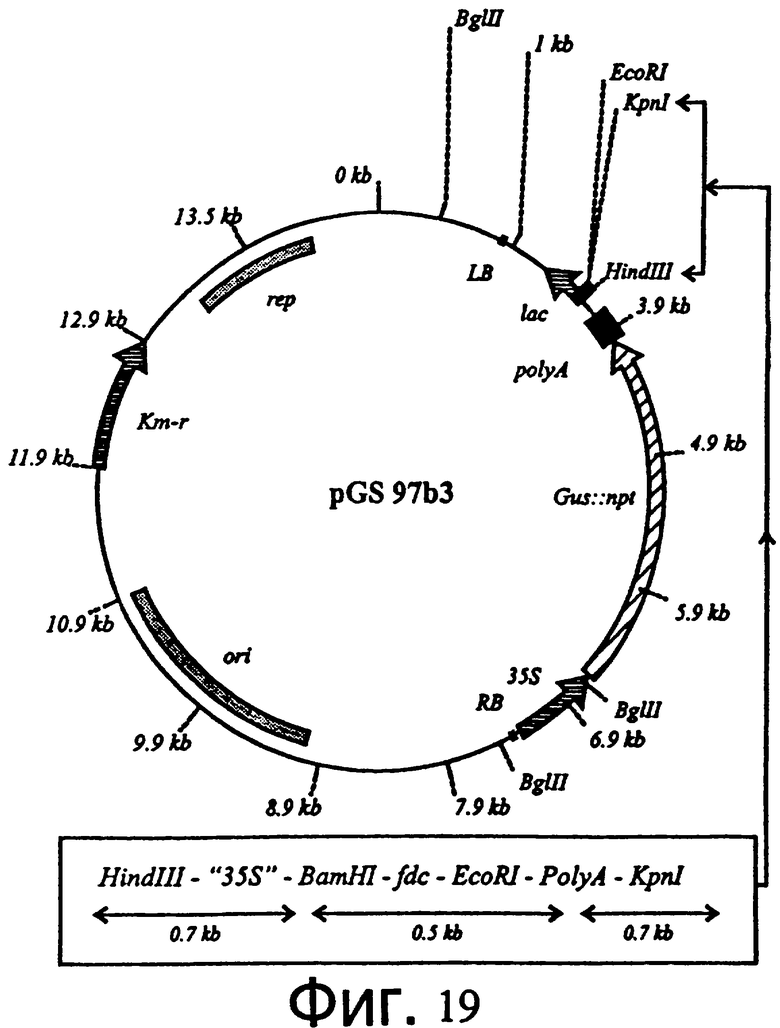
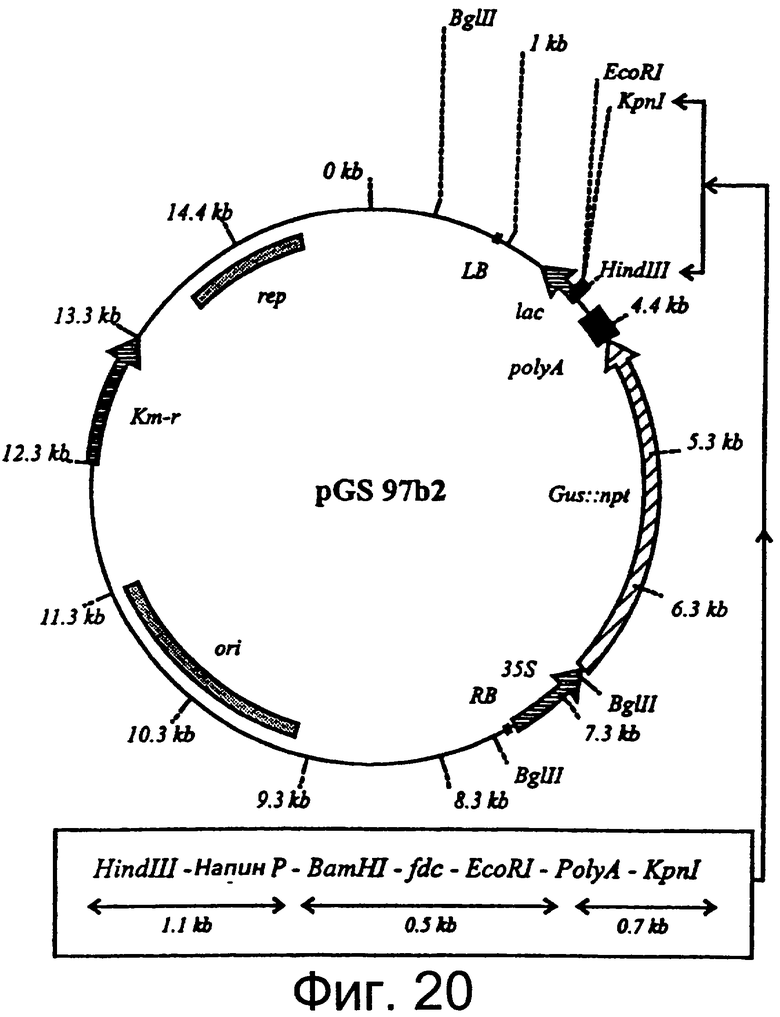
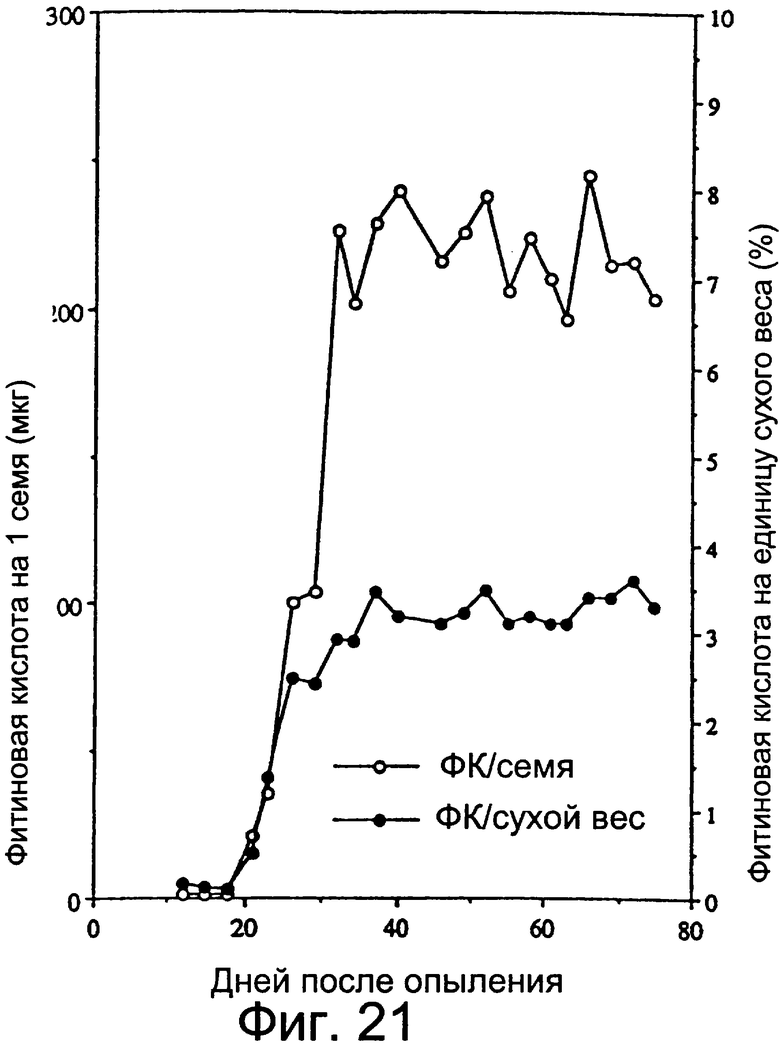
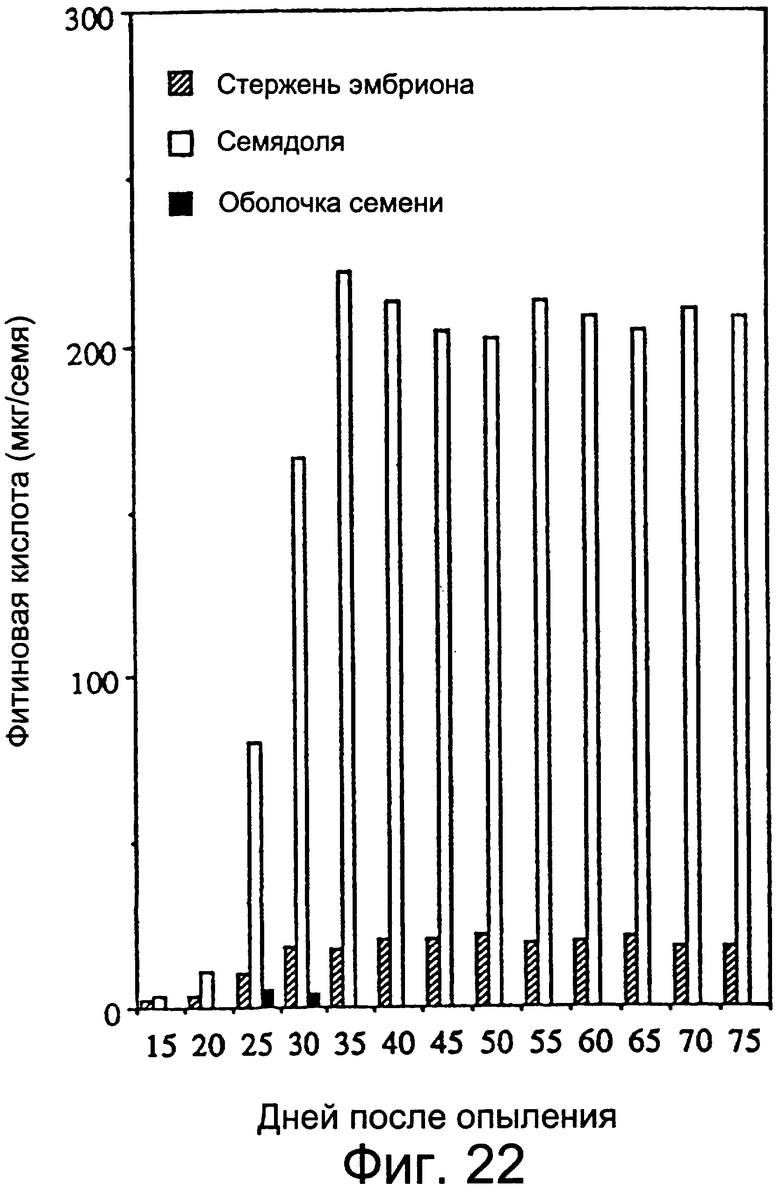
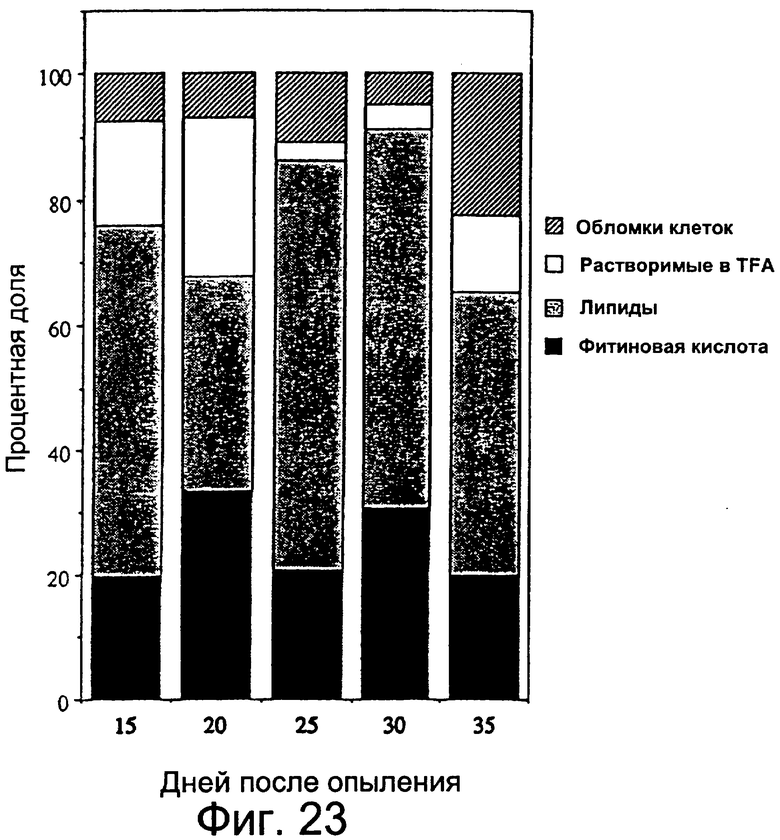
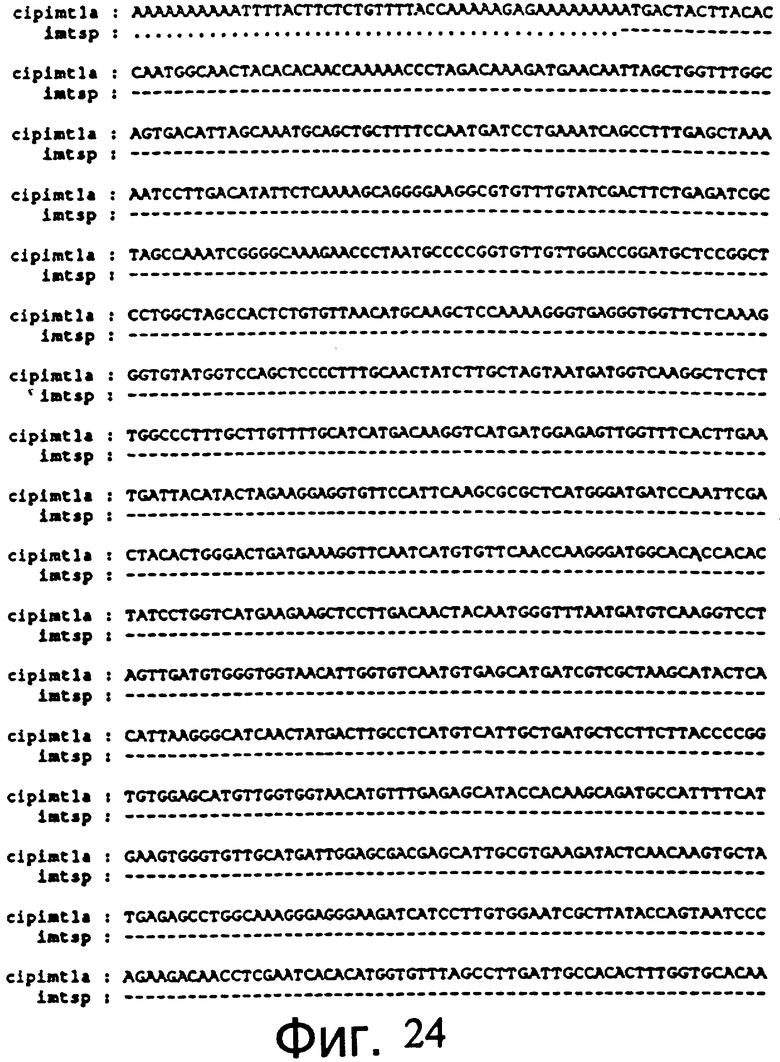

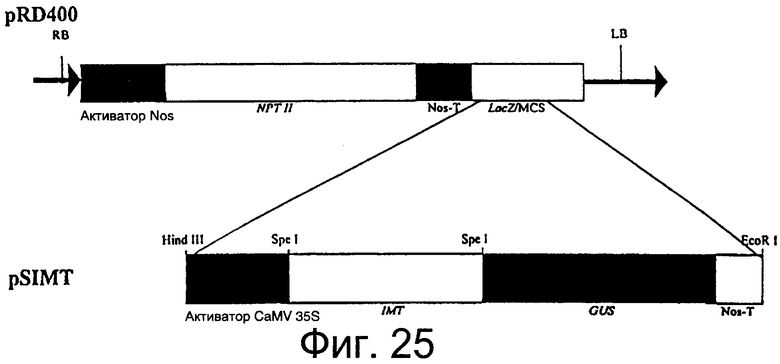
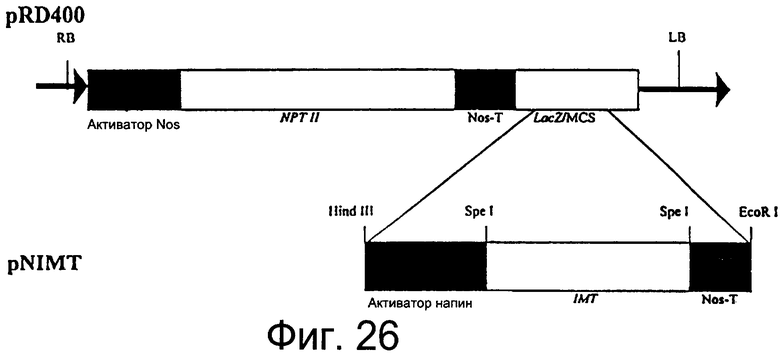

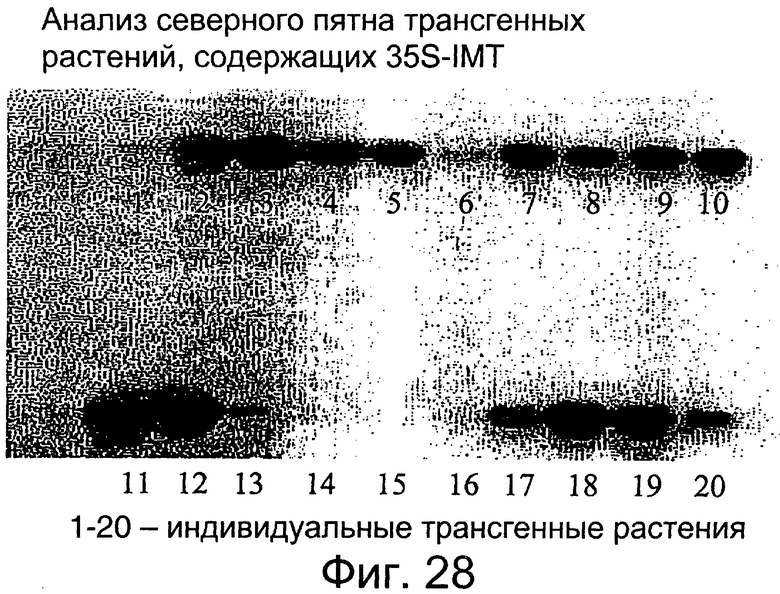
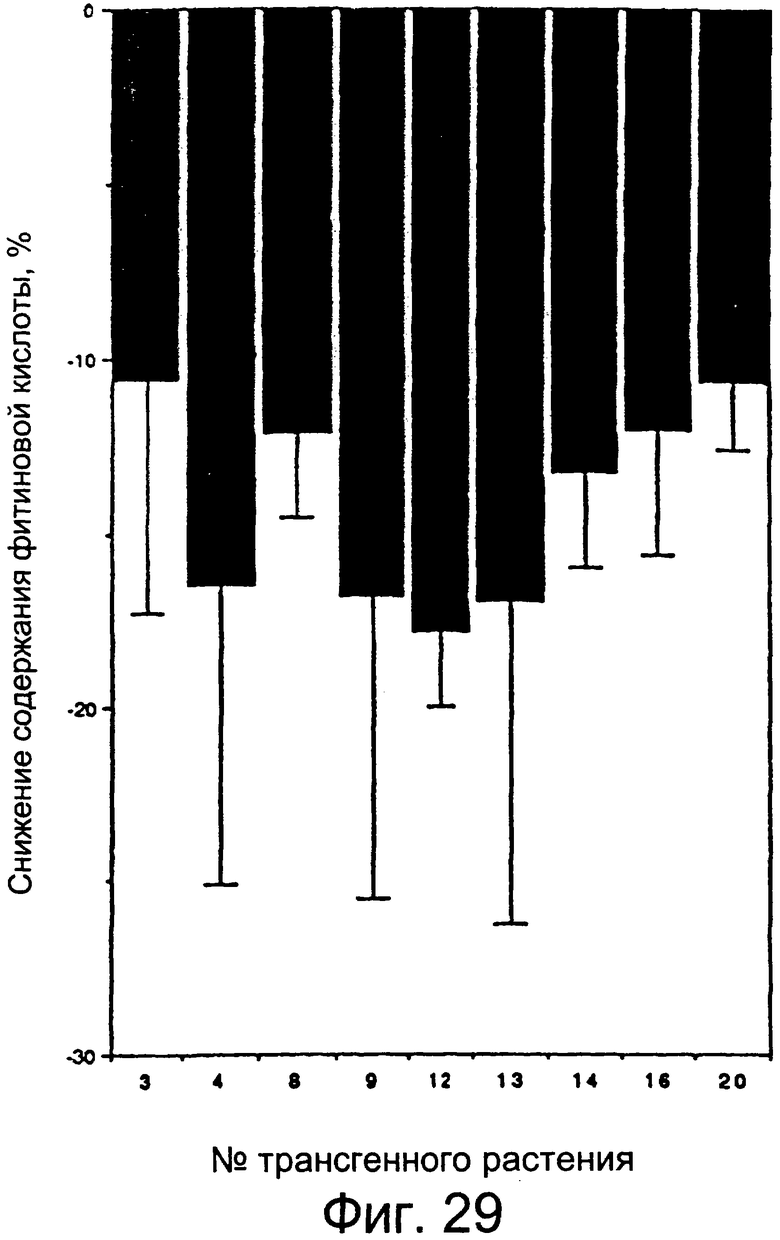

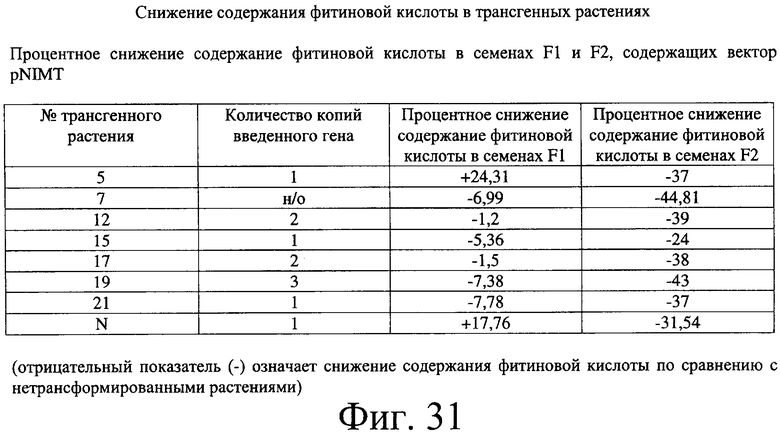
Изобретение относится к биотехнологии, в частности представляет собой способ получения генетически модифицированных растений, имеющих измененный уровень содержания одного из продуктов вторичных процессов обмена веществ. Выбирают последовательность нуклеиновой кислоты, которая кодирует фермент, не встречающийся во вторичном процессе обмена веществ, способный модифицировать использование промежуточного субстрата во вторичном процессе обмена веществ, связанном с питательным профилем растений. Конструируют рекомбинантную молекулу, содержащую вышеуказанную последовательность, которая используется для трансформации растительной клетки. Далее регенерируют генетически модифицированное растение из указанной растительной клетки. Используют полученное растение при приготовлении корма для животных. Изобретение позволяет получать растения с высокой пищевой ценностью, а также с высокими технологическими качествами. 3 н. и 32 з.п. ф-лы, 31 ил.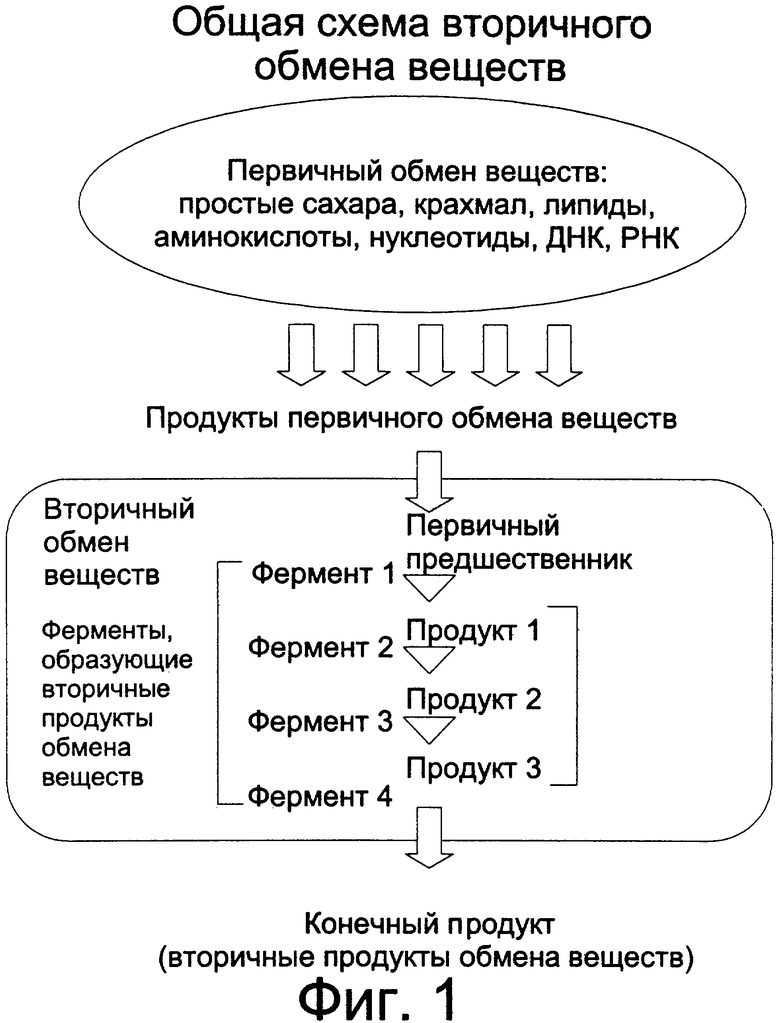
выбора последовательности нуклеиновой кислоты по ее способности кодировать фермент, способный модифицировать использование промежуточного субстрата во вторичном процессе обмена веществ, связанном с питательным профилем растения, причем указанный фермент естественным образом не встречается в указанном вторичном процессе обмена веществ;
трансформации растительной клетки указанного растения экспрессионной кассетой, содержащей указанную последовательность нуклеиновой кислоты, оперативно связанную с промотором, избирательным по отношению к семенам; и
регенерации генетически модифицированного растения из указанной растительной клетки, причем указанное генетически модифицированное растение характеризуется улучшенным питательным профилем относительно дикого типа указанного растения.
выращивания указанного генетически модифицированного растения в условиях, допускающих формирование семени, и регенерации указанного семени.
(а) промотор избирательный по отношению к семенам и
(б) последовательность нуклеиновой кислоты, оперативно связанную с указанным промотором, избирательным по отношению к семенам, причем указанная последовательность нуклеиновой кислоты кодирует фермент, способный модифицировать использование промежуточного субстрата во вторичном процессе обмена веществ, связанном с питательным профилем растения, указанный фермент естественным образом не встречается в указанном вторичном процессе обмена веществ.
| WO 9723599 03.07.1997.CHAPPLE C | |||
| ET AL.: “An Arabidopsis mutant defective in the general phenylpropanoid pathway” PLANT CELL, vol.4, no | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| JP 08266179 A 15.10.1996.RU 94046212 A1 20.10.1996. | |||

