Эта заявка испрашивает приоритет предварительной заявки U.S. 61/040456, поданной 28 марта 2008 г.
Область техники, к которой относится изобретение
Настоящее изобретение предоставляет способы для амплифицирования генного локуса без применения антибиотиков. В частности, изобретение относится к способу амплифицирования in vivo последовательностей ДНК, кодирующих полипептид, представляющий интерес, к клетке, несущей множественные копии указанной амплифицированной ДНК последовательности, и вектору, несущему ДНК конструкт, который применяется в этом способе. Кроме того, настоящее изобретение относится к способу продуцирования белка, представляющего интерес, например фермента, путем культивирования клеток, как описано выше.
Уровень техники
Экспрессионная и рекомбинантная продукция экзогенных полипептидов является широко используемой методикой. Хорошо известно, что клетки могут быть трансформированы нуклеиновыми кислотами, кодирующими экзогенные полипептиды, представляющие интерес, для экспрессии и продуцирования больших количеств желаемых полипептидов. В некоторых вариантах применения, способы используются для получения очень больших количеств полипептидов по сравнению с тем, что могло бы быть продуцировано естественным путем в первоначальном организме. Действительно, экспрессия экзогенных последовательностей нуклеиновых кислот, также как и сверхэкспрессия эндогенных последовательностей, широко используется в современной биотехнологии.
Несмотря на прогресс в молекулярной биологии и белковой инженерии, остается необходимость в новых способах и композициях, которые увеличивают уровень экспресии полипептидов в клетках-хозяевах.
Сущность изобретения
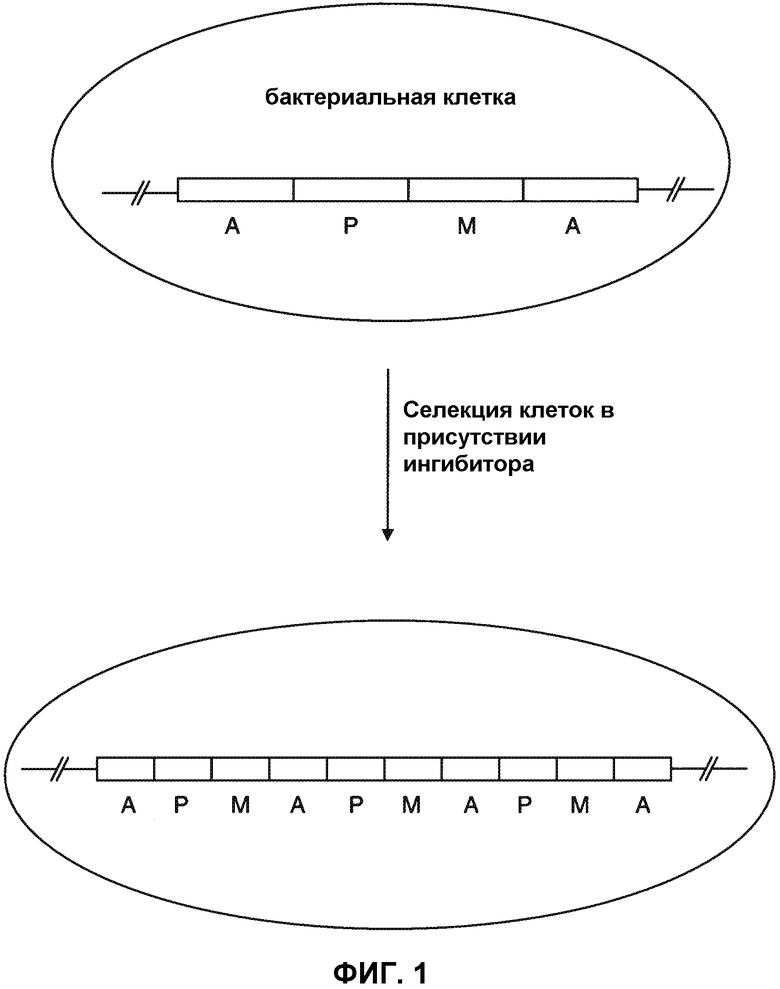
Здесь представлен способ амплифицирования геномного локуса. В определенных вариантах осуществления, способ может включать: а) приведение популяции бактериальных клеток-хозяев с ингибитором необходимого фермента, где бактериальные клетки-хозяева содержат геномный локус со структурой A1-P-M-A2, где А1 и А2 являются прямыми повторами, Р содержит кодирующую последовательность для белка, представляющего интерес, и М содержит кодирующую последовательность для необходимого фермента; и б) отбор клеток, которые являются устойчивыми к действию ингибитора; где клетки, которые являются устойчивыми к действию ингибитора, имеют множественные копии амплификационной единицы. Бактериальная клетка-хозяин может быть клеткой Bacillus sp., хотя и другие бактериальные штаммы, например, Streptomyces sp., предусмотрены. В некоторых вариантах осуществления, полипептид, представляющий интерес, представляет собой субтилизин, например, субтилизин с последовательностью SEQ ID NO: 8, или его зрелой формой, представленной в SEQ ID NO: 12. В определенных случаях, способ избегает использования антибиотических маркеров и антибиотиков и предоставляет альтернативу системам амплификации, основанным на антибиотиках. В определенных вариантах осуществления, необходимый фермент представляет собой аминокислотную последовательность фермента, например фермента дикого типа, который является эндогенным для клетки. В отдельных вариантах осуществления, бактериальная клетка-хозяин, используемая в способе, может содержать или не содержать инактивированный эндогенный ген, кодирующий необходимый фермент, где инактивированный ген может быть в отличном геномном локусе от геномного локуса со структурой: A1-P-M-A2. В определенных случаях, необходимый фермент может быть аланинрацемазой, например, SEQ ID NO: 11, и ингибитор может быть β-хлоро-D-аланин или циклосерин, хотя могут быть использованы и другие комбинации фермент/ингибитор. В некоторых вариантах осуществления, амплификационная единица включает последовательность, представленную в SEQ ID NO:7.
Амплификационная единица обеспечивает экспрессию необходимого фермента, кодируемого областью М. В определенных вариантах осуществления М может включать кодирующую последовательность для необходимого фермента и промотер, функционально связанный с кодирующей последовательностью, где промотер является нативным для кодирующей области необходимого фермента. В определенных вариантах осуществления, кодирующая последовательность и промотер могут быть эндогенными для клетки-хозяина. Амплификационная единица может также обеспечивать экспрессию белка, представляющего интерес, кодируемого областью Р. В определенных вариантах осуществления, кодирующая последовательность Р может быть функционально связана с эндогенным или не-эндогенным промотером, который представлен в прилегающем прямом повторе (А1). В других вариантах осуществления, промотер для Р может быть не представлен в прилегающем прямом повторе. Скорее, промотер может быть представлен в области Р.
В некоторых вариантах осуществления, изобретение предоставляет бактериальную клетку-хозяина, содержащую геномный локус, включающий амплификационную единицу со структурой: A1-P-M-A2, где А1 и А2 являются прямыми повторами, Р содержит кодирующую последовательность для белка, представляющего интерес, и М содержит также предоставленную кодирующую последовательность для необходимого фермента. В этом варианте осуществления, амплификационная единица обеспечивает значительную экспрессию необходимого фермента. Бактериальная клетка-хозяин может быть клеткой Bacillus sp., хотя и другие бактериальные штаммы, например, Streptomyces sp., предусмотрены. В некоторых вариантах осуществления, полипептид, представляющий интерес, представляет собой субтилизином, например, субтилизин с последовательностью SEQ ID NO: 8, или его зрелой формой, представленной в SEQ ID NO: 12. В определенных случаях, способ избегает использования антибиотических маркеров и антибиотиков и предоставляет альтернативу системам амплификации, основанным на антибиотиках. В определенных вариантах осуществления, необходимый фермент представляет собой аминокислотную последовательность фермента, например, фермента дикого типа, который является эндогенным для клетки. В отдельных вариантах осуществления, бактериальная клетка-хозяин, используемая в способе, может содержать или не содержать инактивированный эндогенный ген, кодирующий необходимый фермент, где инактивированный ген может быть в отличном геномном локусе от геномного локуса со структурой: A1-P-M-A2. В определенных случаях, необходимый фермент может быть аланинрацемазой, например, SEQ ID NO: 11, и ингибитором может быть β-хлоро-D-аланин или циклосерин, хотя могут быть использованы и другие комбинации фермент/ингибитор. В некоторых вариантах осуществления, амплификационная единица включает последовательность, представленную в SEQ ID NO:7.
В других вариантах осуществления, бактериальная клетка-хозяин по изобретению содержит геномный локус, включающий множественные копии амплификационной единицы со структурой: A1-P-M-A2, где A1 и A2 являются прямыми повторами, Р содержит первую кодирующую последовательность для белка, представляющего интерес, и М содержит вторую кодирующую последовательность, кодирующую последовательность для необходимого фермента. В некоторых вариантах осуществления, амплификационная единица включает полинуклеотидную последовательность, представленную в SEQ ID NO:7. В некоторых вариантах осуществления, первая кодирующая последовательность функционально связана с промотером, который представлен в прямом повторе A1. В определенных вариантах осуществления, бактериальная клетка-хозяин содержит геномный локус, включающий множественные копии амплификационной единицы, описываемой формулой: (A1-P-M)n-A2, где n равно по крайней мере 2, A1 и A2 являются прямыми повторами, Р содержит кодирующую последовательность для белка, представляющего интерес, и М кодирует необходимый фермент, где кодирующая последовательность М функционально связана в эндогенным или не-эндогенным промотером. В одном варианте осуществления, кодирующая последовательность М и промотер могут быть эндогенными для клеток-хозяев. В других вариантах осуществления, бактериальные клетки-хозяева содержат геномный локус, включающий множественные копии, например, по крайней мере 2 копии, амплификационной единицы с последовательностью SEQ ID NO:7. Амплификационная единица обеспечивает экспрессию как белка, представляющего интерес, например, субтилизина, так и необходимого фермента. В некоторых вариантах осуществления, экспрессируемый полипептид, представляющий интерес, представляет собой субтилизин FNA, представленный в SEQ ID NO: 8, или его зрелой формой, представленной в SEQ ID NO: 12, и необходимый фермент представляет собой аланин-рацемазу, представленной в SEQ ID NO: 11. В определенных вариантах осуществления, промотер, функционально связанный с кодирующей последовательностью Р, может быть частью прилегающего прямого повтора (A1). В другом варианте осуществления, промотер, функционально связаный с кодирующей последовательностью области Р, находится в области Р, а не в прилегающем прямом повторе.
В другом варианте осуществления, изобретение охватывает культуру бактериальных клеток, которая содержит среду для выращивания и популяцию бактериальных клеток-хозяев, содержащих по крайней мере один, по крайней мере две или более копий амплификационной единицы со структурой A1-P-M-A2, где A1 и A2 являются прямыми повторами, Р содержит первую кодирующую последовательность для белка, представляющего интерес, и М содержит вторую кодирующую последовательность для необходимого фермента. Как описано выше, амплификационная единица обеспечивает экспрессию как белка, представляющего интерес, например, субтилизина, так и необходимого фермента. В некоторых вариантах осуществления, экспрессируемый полипептид, представляющий интерес, представляет собой субтилизин FNA, представленный в SEQ ID NO: 8, или его зрелой формой, представленной в SEQ ID NO: 12, и необходимый фермент представляет собой аланин рацемазу, представленную в SEQ ID NO: 11. В еще одном варианте осуществления, культура бактериальной клетки может быть использована в способе продуцирования белка, который включает: содержание культуры соответствующих клеток при условиях, подходящих для продуцирования белка, представляющего интерес, кодируемого кодирующей последовательностью. В определенных вариантах осуществления, этот способ может дополнительно включать выделение белка, представляющего интерес, из культуральной среды.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1 схематически иллюстрирует некоторые особенности вариантов осуществления, описанных здесь.
Фиг. 2 представляет карту плазмиды pBSFNAalr (SEQ ID NO:2).
Фиг. 3 показывает кПЦР калибровочную кривую.
Фиг. 4 является графиком, показывающим уровень экспрессии субтилизина FNA в различных штаммах-хозяевах.
ОПРЕДЕЛЕНИЯ
Если здесь не определено иначе, все технические и научные термины, используемые здесь, имеют то значение, которое обычно понятно обычному специалисту в области техники, к которой относится данное изобретение. Хотя любые способы и материалы, сходные или эквивалентные описанным здесь могут быть применены на практике или при тестировании настоящего изобретения, описаны предпочтительные способы и материалы.
Все патенты или публикации, включая все последовательности, раскрытые в таких патентах или публикациях, упоминаемые здесь, являются однозначно включенными посредством ссылки.
Области числовых значений включают числовые значения, определяющие область. Если не указано иначе, нуклеиновые кислоты написаны слева направо в 5'-3' ориентации; аминокислотные последовательности написаны слева направо в ориентации от амина к карбоксилу, соответственно.
Заголовки, представленные здесь, не являются ограничениями различных аспектов или вариантов осуществления изобретения. Соответственно, термины, определенные сразу ниже, являются более полно определенными в ссылках ко всей спецификации.
Если не определено иначе, все технические и научные термины, используемые здесь, имеют то значение, которое обычно понятно специалисту в области техники, к которой относится данное изобретение. Singleton, et al., DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOGY, 2d ED., John Wiley and Sons, New York (1994) и Hale & Markham, THE HARPER COLLINS DICTIONARY OF BIOLOGY, Harper Perennial, N.Y. (1991) предоставляют специалисту общие значения множества терминов, использованных здесь. Хотя определенные термины определены ниже для ясности и легкости ссылок.
Термин «рекомбинантный» относится к полинуклеотиду или полипептиду, который не представлен в природных условиях в клетке-хозяине. Рекомбинантная молекула может содержать две или более встречающихся в природе последовательности, которые связаны вместе путем, который не встречается в природе. Рекомбинантная клетка содержит рекомбинантный полинуклеотид или полипептид.
Термин «гетерологичный» относится к элементам, которые в норме не связаны друг с другом. Например, если клетка-хозяин продуцирует гетерологичный белок, этот белок в норме не продуцируется этой клеткой-хозяином. Таким же образом, промотер, который функционально связан с гетерологичной кодирующей последовательностью, является промотером, который функционально связан с кодирующей последовательностью, которая обычно не является функционально связанной в клетке-хозяине дикого типа. Термин «гомологичный», по отношению к полинуклеотиду или белку, относится к полинуклеотиду или белку, которые встречаются в природных условиях в клетке-хозяине.
Термины «белок» или «полипептид» здесь применяются взаимозаменяемо.
«Сигнальная последовательность» представляет собой последовательность аминокислот, представленную в N-концевой части белка, которая обеспечивает секрецию зрелой формы белка из клетки. Определение сигнальной последовательности является функциональным. Зрелая форма внеклеточного белка не имеет сигнальной последовательности, которая отрезается в процессе секреции.
Термин «нуклеиновая кислота» включает ДНК, РНК, их одноцепочечные или двухцепочечные формы или их химические модификации. Термины «нуклеиновая кислота» и «полинуклеотид» здесь используются взаимозаменяемо.
«Вектор» относится к полинуклеотиду, предназначенному для введения нуклеиновых кислот в одну или более клеток-хозяев. В определенных вариантах осуществления, вектор может автономно реплицироваться в различных клетках-хозяевах и включает: векторы для клонирования, экспрессионные векторы, «челночные» векторы, плазмиды, фаговые частицы, кассеты и тому подобное. В других вариантах осуществления, вектор может интегрировать в геном клетки-хозяина.
«Промотер» представляет собой регуляторную последовательность, которая инициирует транскрипцию нижележащей по направлению 5'-3' нуклеиновой кислоты.
Термин «функционально связанный» относится к организации элементов, которая позволяет им быть связанными функционально. Например, промотер функционально связан с кодирующей последовательностью, если он контролирует транскрипцию этой последовательности.
Термин «селективный маркер» относится к белку, допускающему экспрессию в клетках-хозяевах, который предусматривает легкость селекции тех клеток-хозяев, которые содержат введенную нуклеиновую кислоту или вектор. Примеры селективных маркеров включают, но не ограничиваются, антибактериальные препараты (например, гигромицин, блеомицин или хлорамфеникол) и/или гены, которые придают метаболическое преимущество, такое как трофическое преимущество для клетки-хозяина.
Термины «выделен», «изолирован» или «очищен из», как они используются здесь, относятся к белку, клетке, нуклеиновой кислоте или аминокислоте, которые отделены от по крайней мере одного другого компонента, с которым они являются обычно ассоциированы.
Как используется здесь, термин «трансформированный», «стабильно трансформированный» и «трансгенный», использованные по отношению к клетке, означают, что клетка несет неприродную (например, гетерологичную) последовательность нуклеиновой кислоты, интегрированную в ее геном или в качестве эписомальной плазмиды, которая сохраняется во многих поколениях.
Как используется здесь, термин «экспрессия» относится к процессу, в котором полипептид продуцируется на основе нуклеотидной последовательности гена. Процесс включает как транскрипцию, так и трансляцию.
Термин «введенный» в контексте помещения последовательности нуклеиновой кислоты в клетку, обозначает «трансфекция», или «трансформация» или «трансдукция» и содержит ссылку на включение последовательности нуклеиновой кислоты в эукариотическую или прокариотическую клетку, где последовательность нуклеиновой кислоты может быть включена в геном клетки (например, хромосомную, плазмидную, пластидную или митохондриальную ДНК), конвертирована в автономный репликон, или временно экспрессирована (например, трансфецированная мРНК).
Термин «гибридизация» относится к процессу, в котором цепь нуклеиновой кислоты соединяется с комплементарной цепью путем спаривания оснований, как известно в области техники. Нуклеиновая кислота считается «селективно гибридизуемой» по отношению к последовательности нуклеиновой кислоты, если две последовательности специфически гибридизуются друг с другом при условиях средней и высокой жесткости гибридизации и отмывок. Условия средней и высокой жесткости гибридизации и отмывок хорошо известны (см., например, Ausubel, et al., Short Protocols in Molecular Biology, 3rd ed., Wiley & Sons 1995 and Sambrook et al., Molecular Cloning: A Laboratory Manual, Third Edition, 2001 Cold Spring Harbor, N.Y.). Один пример условий высокой жесткости включает гибридизацию при примерно 42°С в 50%-ном растворе формамида, 5X SSC, 5X раствор Denhardt, 0,5% SDS и 100 мкг/мл денатурированной ДНК-носителя, с последующей отмывкой два раза в 2X SSC и 0,5% SDS при комнатной температуре и два дополнительных раза в 0,1X SSC и 0,5% SDS при 42°C.
«Кодирующая последовательность» представляет собой часть ДНК, которая кодирует полипептид.
«Экспрессионная кассета», как используется здесь, означает ДНК конструкцию, содержащую область, кодирующую белок, которая функционально связана с подходящей контрольной последовательностью, которая делает возможной эффективную экспрессию белка в соответствующей клетке-хозяине. Такие контрольные последовательности могут включать промотер для эффективной транскрипции, опциональную последовательность оператора для контроля транскрипции для получения мРНК, последовательность, кодирующую соответствующие участки связывания рибосомы на мРНК, и энхансеры или другие последовательности, которые контролируют терминирование транскрипции и трансляции.
Полипепид или полинуклеиотид, который является «нативным для клетки-хозяина», имеет аминокислотную или нуклеиотидную последовательность, которая является такой же, как и представленная в неизмененной клетке-хозяине. В отдельных случаях, клетка может содержать рекомбинантную нуклеиновую кислоту, содержащую полинуклеотид (например, кодирующую последовательность), который является нативным для клетки. В этих случаях, клетка содержит рекомбинантную нуклеиновую кислоту, содержащую полинуклеотид с нуклеотидной последовательностью, которая также присутствует в неизмененной версии клетки-хозяина (например, клетка-хозяин, которая не содержит какого-либо генного нокаута), в другом локусе. В отдельных случаях, клетка может содержать рекомбинантную нуклеиновую кислоту, кодирующую полипептид, который является нативным для клетки. В этих случаях, клетка содержит рекомбинантную нуклеиновую кислоту, содержащую полипептид с аминокислотной последовательностью, которая является такой же, как и полипептид, найденный в неизмененной версии клеток-хозяев (например, клетка-хозяин, которая не содержит какого-либо генного нокаута). Термин «эндогенный» является синонимом термину «нативный».
«Нативный промотер», по отношению к кодирующей последовательности, которая функционально связана со своим нативным промотером, относится к промотеру дикого типа клетки-хозяина, который функционально связан с кодирующей последовательностью в этой клетке.
Термин «прямые повторы» относится к по крайней мере двум элементам последовательности, которые представлены в одинаковой ориентации и которые могут подвергаться гомологичной рекомбинации в клетке. Прямые повторы имеют идентичные или почти идентичные нуклеотидные последовательности (например, по крайней мере 98% или 99% идетичность последовательности) на протяжении по крайней мере 50 нуклеотидов, например, по крайней мере 100, по крайней мере 200 или по крайней мере 500 или более нуклеотидов.
Термин «ингибитор» относится к соединению, которое обратимо ингибирует фермент, как посредством конкурентного ингибирования, так и неконкурентного ингибирования (например, аллостерически).
Термин «необходимый фермент» является ферментом, который является необходимым для роста клеток.
Термин «экспрессионная кассета, которая обеспечивает значительную экспрессию необходимого фермента» относится к экспрессионной кассете, которая обеспечивает экспрессию необходимого фермента на уровне более чем 50% (например, по крайней мере 70%, по крайней мере 90% или по крайней мере 100%, до 1000%) от уровня эндогенного необходимого фермента, если ген эндогенного необходимого фермента является геном дикого типа (т.е. не-инактивированным) в клетке.
Термин «аланин рацемаза» относится к ферменту, который катализирует взаимопревращение L-аланина в D-аланин. Аланин рацемаза имеет активность, описанную в EC 5.1.1.1, в соответствии с IUBMB номенклатурой ферментов. Ген, кодирующий аланин рацемазу, может быть обозначен как "air", "alrA" или "dal" ген.
Другие определения терминов могут возникнуть на всем протяжении спецификации.
ПОДРОБНОЕ ОПИСАНИЕ ПРИМЕРОВ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Как приведено выше, описан способ амплификации геномного локуса. Несколько основных особенностей этого способа проиллюстрированы на Фиг. 1. Со ссылкой на Фиг. 1, бактериальные клетки-хозяева, используемые в способе, могут содержать геномный локус, включающий амплификационную единицу со структурой A1-P-M-A2, где A1 и A2 являются прямыми повторами, Р содержит кодирующую последовательность для белка, представляющего интерес, и М содержит кодирующую последовательность для фермента, который является необходимым для клетки. Подразумевается, что формула A1-P-M-A2 охватывает геномный локус, который содержит прямые повторы, которые ориентированы в любом направлении по отношению к Р и М.
Амплификационная единица обеспечивает экспрессию в клетке белка, представляющего интерес, и необходимого фермента. В определенных случаях, область Р и область М могут независимо включать экспрессионную кассету (а именно, кодирующую последовательность, функционально связанную с промотером) для белка, представляющего интерес, и необходимого фермента, соответственно. В некоторых вариантах осуществления, амплификационная единица включает первую экспрессионную кассету для экспрессии белка, представляющего интерес, и вторую кассету для экспрессии необходимого фермента. В других вариантах осуществления, кодирующая последовательность области Р может быть функционально связана с промотером, который находится в прилегающем прямом повторе. В этом варианте осуществления и, как будет обсуждаться в более подробных деталях ниже, комбинированная нуклеотидная последовательность прямого повтора и кодирующая последовательность для необходимого фермента могут быть эндогенными для клетки, а именно присутствующей в геноме клетки-хозяине. В определенных вариантах осуществления, промотер, функционально связанный с кодирующей последовательностью области М, может быть эндогенным или не-эндогенным для такой кодирующей последовательности. В определенных вариантах осуществления, кодирующая последовательность области М может управляться промотером, функционально связанным с кодирующей последовательностью в Р.
Как является очевидным, ориентация Р и М в любой нуклеиновой кислоте, описанной здесь, может быть обратной ориентацией (а именно, A1-M-P-A2). При этой обратной ориентации и в определенных вариантах осуществления, кодирующая последовательность области М может быть функционально связана с промотером в прямом повторе A1. Альтернативно, кодирующая последовательность области М может быть функционально связана с промотером, который находится в области М.
Популяция таких клеток подвергается воздействию ингибитора необходимого фермента, и отбираются клетки, которые являются устойчивыми к действию ингибитора (то есть, клетки, которые могут расти и делиться в присутствии ингибитора до образования колоний). Как показано ни Фиг. 1, отобранные клетки имеют геномный локус, содержащий множественные копии амплификационной единицы, этот геномный локус может быть описан формулой (A1-P-M)n-A2, где A1 и A2 являются прямыми повторами, n равно по крайней мере 2 и n может быть, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, или по крайней мере 10, например, в пределах от 10 до 50, или от 50 до 100, или более. Отобранные клетки не имеют мутаций в кодирующей последовательности необходимого фермента (например, в сайте связывания ингибитора) или в промотере, связанном с кодирующей последовательностью, по сравнению с неотобранными клетками. Точнее, отобранные клетки имеют увеличенное количество копий амплификационной единицы, что позволяет клеткам расти в присутствии ингибитора. В определенных случаях, популяция клеток может быть подвергнута нескольким раундам селекции, с использованием в каждом раунде селекции последовательно увеличивающейся концентрации ингибитора (а именно, с последовательным удваиванием концентрации ингибитора). В некоторых вариантах осуществления, амплификационная единица A1-Р-М-A2 содержит полинуклеотидную последовательность, представленную в SEQ ID NO:7



где повторяющиеся единицы A1 и A2 подчеркнуты, полинуклеотидная последовательность, кодирующая белок, представляющий интерес, а именно субтилизин FNA, выделена полужирным шрифтом, и полинуклеотидная последовательность, кодирующая необходимый фермент, например, аланин рацемазу, выделена курсивом. Последовательности промотеров показаны заглавными полужирными буквами.
Поскольку клетки-хозяева, полученные этим способом, содержат больше копий первой экспрессионной кассеты, клетки могут продуцировать больше белка, представляющего интерес, кодируемого первой экспрессионной кассетой, чем клетки-хозяева, которые имеют единственную копию амплификационной единицы A1-Р-М-A2. В конкретных вариантах осуществления, результирующие клетки-хозяева могут продуцировать по крайней мере на 20%, по крайней мере на 40%, по крайней мере на 60%, по крайней мере на 80%, по крайней мере на 100%, по крайней мере в два раза, по крайней мере в три раза, по крайней мере в пять раз или по крайней мере в десять раз, до примерно в 100 раз больше белка по сравнению с во всем остальном идентичными клетками-хозяевами, которые имеют единственную копию амплификационной единицы A1-Р-М-A2.
Концентрация ингибитора, используемая в тематическом способе, может изменяться в зависимости от используемого необходимого фермента и эффективности ингибитора. В определенных вариантах осуществления, ингибитор может быть в концентрации в пределах от 1 мкМ до 100 мМ, например, в пределах от 5 мкМ до 10 мМ, от 20 мкм до 1 мМ, хотя концентрации ингибитора вне этих пределов также представимы. Ингибитор может быть добавлен в жидкую культуру или может быть представлен в твердой среде (например, среда с агаром) в зависимости от того, на какой среде растут бактерии. Как замечено выше, популяция клеток может быть подвергнута нескольким раундам селекции, при использовании в каждом раунде селекции последовательно увеличивающейся концентрации ингибитора (а именно, с последовательным удваиванием концентрации ингибитора).
В определенных вариантах осуществления, амплификационная единица не содержит маркера устойчивости к антибиотику, и отбор клеток может проводиться в среде без антибиотика.
Первая и вторая экспрессионные кассеты и клетки-хозяева описаны более подробно ниже.
Экспрессионные кассеты
Как замечено выше, амплификационная единица обеспечивает экспрессию полипептида, представляющего интерес, и необходимого фермента. Собственно, амплификационная единица обычно содержит по крайней мере две экспрессионные кассеты: первая экспрессионная кассета для экспрессии полипептида, представляющего интерес, и вторая экспрессионная кассета для экспрессии необходимого гена. Каждая экспрессионная кассета содержит, в функциональной связи: промотер, кодирующую последовательность, и терминатор. В определенных случаях, область Р амплификационной единицы может содержать первую экспрессионную кассету, и область М амплификационной единицы может содержать вторую экспрессионную кассету. В других случаях, как отмечено выше, прямой повтор, прилегающий к области Р, может содержать промотер, функционально связанный с кодирующей последовательностью области Р. В определенных случаях, непрерывная последовательность области Р и прямой повтор, прилегающий к области Р, могут быть эндогенными для клетки-хозяина (то есть, содержаться в геноме клетки-хозяина). В конкретном варианте осуществления кодирующая последовательность области М может быть функционально связана с промотером области Р.
Каждая экспрессионная кассета, рассмотренная здесь, может содержать следующие элементы в функциональной связи: промотер, кодирующую последовательность и последовательность терминатора, где экспрессионная кассета является достаточной для продуцирования белка в клетке-хозяине. Как будет рассмотрено более подробно ниже, кодирующая последовательность первой экспрессионной кассеты может кодировать рекомбинантный белок, например терапевтический белок или так называемый «индустриальный фермент». В конкретных вариантах осуществления, эта кодирующая последовательность может кодировать белок, имеющий сигнальную последовательность, которая обеспечивает секрецию белка из клетки. Как указано выше и как будет описано более подробно ниже, вторая экспрессионная кассета обеспечивает экспрессию необходимого фермента.
Выбор промотеров, терминаторов и сигнальных последовательностей, если таковые используются, значительно зависит от используемых клеток-хозяев. Клетки-хозяева включают клетку-хозяина Bacillus sp., клетки-хозяева Streptomyces sp., E. Coli и другие бактериальные клетки-хозяева. Как указано выше, в примерах осуществления, могут быть использованы клетки-хозяева Streptomyces, в которых сигнальная последовательность, если используется, может быть celA сигнальной последовательностью. В определенных случаях, celA сигнальная последовательность может быть сигнальной последовательностью, кодируемой геном целлюлазы А S. Lividans, CelA, как описано Kluepfel et al. (Nature Biotechnol. 1996 14:756-759). В других примерах осуществления, в которых используется клетка-хозяин Bacillus, сигнальная последовательность может быть последовательностью аминокислот, которая способна направлять гибридный белок в секреторные пути клетки-хозяина Bacillus. В определенных случаях, сигнальная последовательность, которая может быть использована, содержит сигнальную последовательность белков, которые секретируются Bacillus клетками-хозяевами дикого типа. Такие сигнальные последовательности включают сигнальные последовательности, кодируемые генами α-амилазы, протеазы, например aprE или субтилизин Е, или β-лактамазы. Типичные сигнальные последовательности включают, но не ограничиваются, сигнальные последовательности, кодируемые геном α-амилазы, геном субтилизина, геном β-лактамазы, геном нейтральной протеазы (например, nprT, nprS, nprM), или геном prsA из любого подходящего вида Bacillus, включая, но не ограничиваясь, B. stearothermophilus, B. licheniformis, B. clausii, B. subtilis и B. amyloliquefaciens. В одном варианте осуществления, сигнальная последовательность кодируется геном aprE из B. subtilis (как описано в Appl. Microbiol. Biotechnol. 2003 62:369-73). Дополнительные сигнальные пептиды описаны в Simonen and Palva (Microbiological Reviews 1993 57: 109-137) и в других ссылках.
Подходящие промотеры и терминаторы для использования в Bacillus и Streptomyces клетках-хозяевах известны и включают: промотеры и терминаторы генов apr (алкалиновая протеаза), npr (нейтральная протеаза), amy (α-амилаза) и β-лактамазы, также как и ген левансукразы B. subtilis (sacB), ген альфа-амилазы B. licheniformis (amyh), ген мальтогенной амилазы B. stearothermophilus (amyM), ген альфа-амилазы B. amyloliquefaciens (amyQ), ген пенициллиназы B. licheniformis (penV), B. subtilis xylA и xylB гены, промотеры и терминаторы, описанные в WO 93/10249, WO 98/07846, и WO 99/43835. Экспрессионные кассеты для использования в Streptomyces клетках-хозяевах могут быть сконструированы с использованием промотеров и терминаторов, описанных в Hopwood et al (Genetic Manipulation of Streptomyces: A Laboratory Manual; Cold Spring Harbor Laboratories, 1985), Hopwood et al (Regulation of Gene Expression in Antibiotic-producing Streptomyces. In Booth, I. and Higgins, C. (Eds) Symposium of the Society for General Microbiology, Regulation of Gene Expression, Cambridge University Press, 1986 pgs. 251-276), Fornwald et al (Proc. Natl. Acad. Sci. 1987 84: 2130-2134), Pulido et al (Gene. 1987 56:277-82); Dehottay et al (Eur. J. Biochem. 1987 166:345-50), Taguchi (Gene. 1989 84:279-86), Schmitt-John et al (Appl. Microbiol. Biotechnol. 1992 36:493-8), Motamedi (Gene 1995 160:25-31) и Binnie (Protein Expr. Purif. 1997 11:271-8), например. В одном варианте осуществления, промотер А4 может быть применен, который описан в WO 06/054997, который является включенным в эту заявку посредством ссылки.
В определенных вариантах осуществления, любая из последовательностей может быть кодон-оптимизированной для экспрессии полипептида, представляющего интерес, в использованных клетках-хозяевах. Поскольку таблицы использования кодонов, перечисляющие использование каждого кодона во многих клетках, известны в области техники (см., например, Nakamura et al, Nucl. Acids Res. 2000 28: 292) или легко выводимы, такие нуклеиновые кислоты могут быть легко получены, дающие аминокислотную последовательность белков, которые будут экспрессированы.
Системы для экспрессии рекомбинантных белков в клетках-хозяевах Streptomyces и Bacillus хорошо известны в области техники и не нуждаются в более детальном обсуждении, чем представлено выше.
Первая экспрессионная кассета
Первая экспрессионная кассета может содержать промотер и полинуклеотид, кодирующий белок, представляющий интерес (то есть, кодирующую последовательность), где промотер и полинуклеотид функционально связаны таким образом, что выделенная нуклеиновая кислота обуславливает транскрипцию полинуклеотида и продукцию белка, представляющего интерес.
Кодируемый белок, представляющий интерес, может быть так называемый «индустриальный фермент», терапевтический белок, репортерный белок, пищевая добавка или продукт питания и тому подобное.
В одном варианте осуществления белок, представляющий интерес, может быть ферментом, таким как карбогидраза, такая как разжижающая и осахаривающая α-амилаза, алкалиновая α-амилаза, β-амилаза, целлюлаза, декстраназа, α-глюкозидаза, α-галактозидаза, глюкоамилаза, хемицеллюлаза, пентозаназа, ксиланаза, инвертаза, лактаза, наринганаза, пектиназа или пуллуланаза; протеаза, такая как кислая протеаза, щелочная протеаза, бромелаин, фицин, нейтральная протеаза, папаин, пепсин, пептидаза, реннет, реннин, химозин, субтилизин, термолизин, аспарагиновая протеаза, или трипсин; липаза или эстераза, такая как триглицеридаза, фосфолипаза, преджелудочная эстераза, фосфатаза, фитаза, амидаза, иминоацилаза, глутаминаза, лизоцим, или пенициллин ацилаза; изомераза, такая как глюкоза изомераза; оксидоредуктаза, например, аминокислотная оксидаза, каталаза, хлоропероксидаза, глюкоза оксидаза, гидроксистероидная дегидрогеназа или пероксидаза; лиаза, такая как ацетолактат декарбоксилаза, аспарагиновая β-декарбоксилаза, фумараза или гистадаза; трансфераза, такая как циклодекстрин гликозилтрансфераза; или лигаза, например. В конкретных вариантах осуществления, белок может быть аминопептидазой, карбоксипептидазой, хитиназой, кутиназой, дезоксирибонуклеазой, α-галактозидазой, β-галактозидазой, β-глюкозидазой, лакказой, маннозидазой, мутаназой, пектинолитическим ферментом, полифенолоксидазой, рибонуклеазой или трансглутаминазой, например.
В определенных вариантах осуществления, белок, представляющий интерес, кодируемый первой экспрессионной кассетой, является детергент-аддитивным белком, то есть белком (а именно, ферментом), который: а) секретируется из клетки и б) добавляется в моющее средство для стирки. Примеры детергент-аддитивных белков включают протеазы, например, субтилизины, α-амилазы и липазы. Субтилизины, а именно экстрацеллюларные щелочные сериновые протеазы, представляют особый интерес. Субтилизины могут иметь аминокислотную последовательность, которая присутствует в геноме дикого типа (то есть субтилизин может быть субтилизином, встречающимся в природе) или могут быть вариантом субтилизина, встречающегося в природе, и таким образом могут содержать аминокислотную последовательность, которая по крайней мере на 80%, крайней мере на 90%, крайней мере на 95% или крайней мере на 98% идентична субтилизину, кодируемому в геноме дикого типа. Примеры субтилизинов включают: Alcanase® (Novozymes), FNA™ (Genencor), Savinase® (Novozymes), Purafect™ (Genencor), KAP™ (Kao), Everlase™ (Novozymes), Purafect OxP™ (Genencor), FN4™ (Genencor), BLAP S™ (Henkel), BLAP X™ (Henkel), Esperase® (Novozymes), Kannase™ (Novozymes) and Prosperase™ (Genencor). В других вариантах осуществления, субтилизин может быть субтилизин 168, субтилизин BPN', субтилизин Carlsberg, субтилизин DY, субтилизин 147 или субтилизин 309 (см., например, EP414279B, WO89/06279 и Stahl et al., J. Bacteriol. 1984 159:811-818). В некоторых вариантах осуществления субтилизин, кодируемый первой экспрессионной кассетой, является FNA

Пре-про-область субтилизина показана курсивом, и область зрелого белка показана полужирным шрифтом (SEQ ID NO: 12). Пример полинуклеотида, который кодирует FNA, является следующим:

Примеры субтилизинов и других протеаз, которые могут быть использованы здесь, включают описанные в WO 99/20770; WO 99/20726; WO 99/20769; WO 89/06279; RE 34606; U.S. Patent No. 4914031; U.S. Patent No. 4980288; U.S. Patent No. 5208158; U.S. Patent No. 5310675; U.S. Patent No. 5336611; U.S. Patent No. 5399283; U.S. Patent No. 5441882; U.S. Patent No. 5482849; U.S. Patent No. 5631217; U.S. Patent No. 5665587; U.S. Patent No. 5700676; U.S. Patent No. 5741694; U.S. Patent No. 5858757; U.S. Patent No. 5880080; U.S. Patent No. 6197567; и U.S. Patent No. 6218165. Субтилизины в целом рассмотрены в подробностях в Siezen (Protein Sci. 1997 6:501-523), и детергент-аддитивные субтилизины рассмотрены в Bryan (Biochim. Biophys. Acta 2000 1543:203-222), Maurer (Current Opinion in Biotechnology 2004 15:330-334) и Gupta (Appl Microbiol Biotechnol. 2002 59: 15-32). Конкретные субтилизины, представляющие интерес, имеют активность, описанную в EC 3.4.4.16, в соответствии с номенклатурой ферментов IUBMB.
В других вариантах осуществления, белок, представляющий интерес, может быть терапевтическим белком (то есть, белком, имеющим терапевтическую биологическую активность). Примеры подходящих терапевтических белков включают: эритропоэтин, цитокины, такие как интерферон-α, интерферон-β, интерферон-γ, интерферон-o, и гранулоцит-CSF, GM-CSF, факторы коагуляции, такие как фактор VIII, фактор IX, и человеческий белок C, антитромбин III, тромбин, растворимый IgE рецептор α-цепь, IgG, IgG фрагменты, IgG гибридные белки, IgM, IgA, интерлейкины, урокиназа, химаза, мочевинный ингибитор трипсина, IGF-связывающий белок, эпидермальный фактор роста, релиз-фактор гормона роста, аннексин V гибридный белок, ангиостатин, васкулярный эндотелиальный фактор-2 роста, ингибирующий фактор-1 миелоидных предшественников, остеопротегерин, α-1-антитрипсин, α-фето протеины, ДНказа II, kringle 3 плазминогена человека, глюкоцероброзидаза, TNF связывающий белок 1, фолликул-стимулирующий гормон, цитотоксический T-лимфоцит ассоциированный антиген 4-Ig, трансмембранный активатор и модулятор кальция и циклофилин лиганд, растворимый рецептор TNF Fc гибридный белок, глюкагон-подобный белок 1 и антагонист IL-2 рецептора. Белки антител, например, моноклональные белки, которые могут быть гуманизированы, представляют особенный интерес.
В дополнительных вариантах осуществления, белок, представляющий интерес, может быть репортерным белком. Такие репортерные белки могут быть оптически детектируемые или цветогенными, например. В этом варианте осуществления, белок может быть β-галактозидазой (lacZ), β-глюкоронидазой (GUS), люциферазой, щелочной фосфотазой, нопалин синтазой (NOS), хлорамфеникол ацетилтрансферазой (CAT), пероксидазой хрена (HRP) или флуоресцентным белком, например, зеленым флуоресцентным белком (GFP), или его производными.
Как замечено выше, кодирующая последовательность может кодировать гибридный белок. В некоторых из этих вариантов осуществления, гибридный белок может обеспечивать секрецию белка, представляющего интерес, из клетки-хозяина, в которой он экспрессируется, и, сам по себе, может содержать сигнальную последовательность, функционально связанную с N-концом белка, представляющего интерес, где сигнальная последовательность содержит последовательность аминокислот, которая направляет белок в секреторную систему клетки-хозяина, что приводит к секреции белка из клетки-хозяина в среду, в которой растут клетки-хозяева. Сигнальная последовательность отщепляется от гибридного белка перед секрецией белка, представляющего интерес.
Вторая экспрессионная кассета
Вторая экспрессионная кассета обеспечивает экспрессию необходимого фермента, где, как указано выше, необходимый фермент требуется клетке для клеточного роста. В определенных вариантах осуществления, необходимый фермент может быть условно необходимым, то есть требоваться клетке для клеточного роста только при определенных условиях (то есть, в отсутствие экзогенного соединения, которое нейтрализует какой-либо недостаток необходимого фермента). В определенных случаях, клетки, испытывающие недостаток активности условно необходимого фермента (который может быть создан путем инактивирования гена, кодирующего фермент, или путем контактирования клеток с ингибитором фермента), могут выращиваться в культуре путем добавления экзогенного соединения, которое в определенных условиях может быть продуктом фермента или альтернативного источника углерода. Так, в определенных случаях, необходимый фермент, используемый во второй экспрессионной кассете, может быть ферментом, который, когда отсутствует в клетке, придает клетке ауксотрофность к определенному соединению или неспособность утилизировать один или более специфических источников углерода.
Примеры таких комбинаций необходимый фермент/ингибитор известны и включают, например: ферменты, которые вовлечены в синтез аминокислот и их соответствующие ингибиторы; и ферменты, вовлеченные в утилизацию специфических источников углерода и их соответствующие ингибиторы. Примеры таких комбинаций необходимый фермент/ингибитор представлены ниже. Инактивация гена, кодирующего фермент, вовлеченный в синтез аминокислот, приводит к ауксотрофности по этой аминокислоте. Подобным образом, инактивация гена, кодирующего фермент, вовлеченный в утилизацию специфических источников углерода, приводит к ауксотрофности по этому источнику углерода. Фермент не расщепляет ингибитор. Более того, ингибитор обратимо и специфически ингибирует каталитическую активность фермента как конкурентно, так и не конкурентно.
В одном варианте осуществления, фермент может быть S-аденозил-метионин синтазой (кодируемой metE; Genbank accession no. U52812; см. Yocum et al, Cloning and characterization of the metE gene encoding S-adenosylmethionine synthetase from Bacillus subtilis. J. Bacterid. 1996 178:4604), которая может быть ингибирована циклолейцином (Chiang et al Molecular characterization of Plasmodium falciparum S-adenosylmethionine synthetase. Biochem J. 1999 344:571-6), также как и аналогами метионина, аналогами пурина, 8-азагуанидином и азатиоприном (Berger et al Characterisation of methionine adenosyltransferase from Mycobacterium smegmatis and M. tuberculosis BMC Microbiol. 2003; 3: 12). Инактивация гена S-аденозил-метионин синтазы вызывает метиониновую ауксотрофтность.
В другом варианте осуществления, фермент может быть 3-изопропилмалат дегидрогеназой, которая катализирует превращение 3-карбокси-2-гидрокси-4-метилпентаноата в 3-карбокси-4-метил-2-оксопентаноат. Этот фермент кодируется leuB, и leuB-дефицитные штаммы являются ауксотрофами. 3-изопропилмалат дегидрогеназа может быть ингибирована, например, О-изобутенил оксалилгидроксаматом (Singh et al The High-resolution Structure of LeuB (Rv2995c) from Mycobacterium tuberculosis Journal of Molecular Biology 2005 346: стр. 1-11).
В другом варианте осуществления, фермент может быть диаминопимелат декарбоксилазой, которая катализирует превращение мезо-2,6-диаминогептандиоата в L-лизин, и кодируется lysA. lysA-дефицитные штаммы будут лизиновыми ауксотрофами. Ингибиторы диаминопимелат декарбоксилазы включают аналоги диаминопимелиновой кислоты, включая, но не ограничиваясь: лантионин сульфоксиды, мезо- и LL-изомеры лантионин сульфона, лантионин, N-модифицированные аналоги, включая N-гидроксидиаминопимелат 4 и N-гидроксидиаминопимелат 5 (см. Kelland et al J. Biol. Chem. 1986 Analogs of diaminopimelic acid as inhibitors of meso-diaminopimelate decarboxylase from Bacillus sphaericus and wheat germ 261: 13216-13223).
В другом варианте осуществления, фермент может быть глютамил-тРНК-редуктазой, которая катализирует синтез 5-амино левулиновой кислоты, и кодируется геном hemA. hemA-дефицитные штаммы являются ауксотрофными по 5-амино левулиновой кислоте или хемину. Этот фермент может быть ингибирован глютамицином (Schauer et al Escherichia coli Glutamyl-tRNA Reductase J. Biol. Chem. 2002 277: 48657-48663). В дополнительном варианте осуществления, фермент может быть D-аланин рацемазой, и кодироваться alr (также известным как dal). Alr-дефицитные штаммы являются ауксотрофными по D-аланину, который необходим для биосинтеза клеточной стенки. Ингибиторы D-аланин рацемазы включают, но не ограничиваются, D-циклосерин, β-хлор-D-аланин и O-карбамил-D-серин (см., например, Manning et al, Inhibition of Bacterial Growth by β -chloro-D-alanine PNAS 1974 71: 417-421). В некоторых вариантах осуществления, вторая экспрессионная кассета содержит полинуклеотид, например
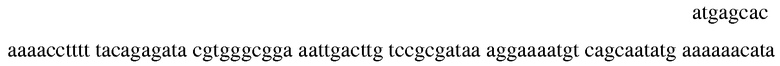

который кодирует D-аланин рацемазу

Другие необходимые ферменты, для которых могут быть использованы ингибиторы, включают: ксилоза изомеразу (xylA), глюконат киназу (EC 2.7.1.12), глюконат пермеазу (gntK или gntP), глицеролкиназу, глицерол дегидрогеназу, например, glpP, glpF, glpK, или glpD или арабиноза изомеразу (araA), например.
В определенных вариантах осуществления, вторая экспрессионная кассета обеспечивает значительную экспрессию необходимого фермента, при которой необходимый фермент продуцируется на уровне более чем 50% (например, по крайней мере примерно 70%, по крайней мере примерно 90% или по крайней мере примерно 100%, до по крайней мере примерно 1000%) от уровня эндогенного необходимого фермента, если в клетке ген для эндогенного необходимого фермента является геном дикого типа (т.е. не-инактивированным).
В определенных вариантах осуществления, необходимый фермент, кодируемый областью М, может быть встречающимся в природе, то есть имеющим аминокислотную последовательность необходимого фермента дикого типа. В других вариантах осуществления необходимый фермент, кодируемый областью М, может быть вариантом фермента, встречающегося в природе, то есть может иметь аминокислотную последовательность по крайней мере на 80% идентичную, по крайней мере на 90% идентичную, по крайней мере на 95% идентичную, по крайней мере на 98% идентичную или по крайней мере на 99% идентичную необходимому ферменту, встречающемуся в природе.
В определенных вариантах осуществления, необходимый фермент может быть встречающейся в природе аминокислотной последовательностью и, в определенных вариантах осуществления, может быть эндогенным для клетки-хозяина тем, что имеет аминокислотную последовательность, которая кодируется геномом клетки-хозяина, до каких-либо инактивирующих мутаций.
В определенных вариантах осуществления, нуклеотидная последовательность экспрессионной кассеты (то есть, промотер, кодирующая последовательность и терминатор) могут быть геном, который является эндогенным для клетки-хозяина, до каких-либо инактивирующих мутаций. Такой ген может быть в отличном геномном локусе от локуса экспрессионной кассеты.
Хотя это и не требуется для осуществления на практике настоящего способа, эндогенный ген необходимого фермента (т.е. ген, который содержится в клетке-хозяине и не включает пока локус A1-P-M-A2) может быть инактивирован посредством мутации. Способы для специфической инактивации бактериальных генов, например, путем делеции, замены или вставки, хорошо известны в области техники.
Клетки-хозяева
Бактериальные клетки-хозяева, используемые здесь, могут быть грамположительными или грамотрицательными, и включают, но не ограничиваются: Bacillus sp. бактерии, например, Bacillus clausii, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacillus coagulans, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus stearothermophilus, Bacillus subtilis, или Bacillus thuringiensis бактерии; Streptomyces sp. бактерии, например, S. lividans, S. carbophilus, S. helvaticus, S. rubiginosus или S. murinus bacteria, Pseudomonas sp. бактерии и E. coli. В определенных случаях, бактериальные клетки-хозяева могут быть клетками штамма, который имеет историю использования для продуцирования белков, который имеет GRAS статус от FDA, то есть общепризнан безопасным (Generally Recognized as Safe).
Клетки-хозяева B. subtilis включают, но не ограничиваются, описанные в патентах U.S. 5264366 и 4760025 (RE 34606), также как и 1A6 (ATCC 39085), 168 (IAOl), SB19, W23, Ts85, B637, PB1753 до PB1758, PB3360, JH642, 1A243 (ATCC 39087), ATCC 21332, ATCC 6051, MI113, DEl00 (ATCC 39094), GX4931, PBT 110, и PEP 211 штамм (см., например, Hoch et al, Genetics 1973 73:215-228; U.S. Patent No. 4450235; U.S. Patent No. 4302544; и EP 0134048). Применение B. subtilis в качестве экспрессионного хозяина также описано Palva et al. и др. (см., Palva et al, Gene 1982 19:81-87; также см. Fahnestock and Fischer, J. Bacterid. 1986 165:796-804; и Wang et al, Gene 1988 69:39-47), например.
В определенных вариантах осуществления клетки-хозяева Bacillus могут быть сконструированы для максимизирования экспрессии белка, и, как таковые, могут содержать инактивирующее изменение по крайней мере в одном из следующих генов degU, degS, degR и degQ. См., Msadek et al. (J. Bacteriol. 1990 172:824-834) и Olmos et al, (MoI. Gen. Genet. 1997 253:562-567). Один штамм из видов Bacillus subtilis и несет degU32(Hy) мутацию. В другом варианте осуществления, клетка-хозяин Bacillus может содержать мутацию или делецию в scoC4, (см., Caldwell et al., J. Bacteriol. 2001 183:7329-7340); spoIIE (см., Arigoni et al, Mol. Microbiol. 1999 31: 1407-1415); oppA или другом гене в opp опероне (см., Perego et al, Mol. Microbiol. 1991 5:173-185).
Бактериальные клетки, используемые в тематическом способе, могут быть созданы путем вставки рекомбинантной нуклеиновой кислоты в геном бактериальной клетки-хозяина. В определенных вариантах осуществления, клетки могут быть созданы путем гомологичной или не-гомологичной рекомбинации, с использованием способа, сходного с общепризнанными способами, такими как описанные в Jung et al (J. Gen. Appl. Microbiol. 1998 44 107-111); Tangney et al (FEMS Microbio. Lett. 1995 125: 107-114); Petit et al (EMBO J. 1992 11:1317-1326); US патент 5733753 и опубликованная патентная заявка US 20070134760.
Клетка-хозяин может или может не иметь инактивированный эндогенный ген, кодирующий необходимый фермент.
Способы продуцирования белка
Способы использования вышеописанных клеток также описаны. В определенных вариантах осуществления, тематические способы включают: культивирование популяции клеток для продуцирования белка, представляющего интерес, кодируемого экспрессионной кассетой. В определенных вариантах и как обсуждается выше, белок, представляющий интерес, может быть секретирован в культуральную среду. Конкретные варианты осуществления способа включают шаг выделения белка, представляющего интерес, из культуральной среды.
Белок, представляющий интерес, может быть выделен из среды для выращивания любым удобным способом, например, осаждением, центрифугированием, аффинной хроматографией, фильтрацией или любым другим способом, известным в области техники. Например, аффинной хроматографией (Tilbeurgh et al., (1984) FEBS Lett. 16:215); ион-обменными хроматографическими методами (Goyal et al., (1991) Biores. Technol. 36:37; Fliess et al., (1983) Eur. J. Appl. Microbiol. Biotechnol. 17:314; Bhikhabhai et al., (1984) J. Appl. Biochem. 6:336; и Ellouz et al., (1987) Chromatography 396:307), включая ион-обмен с применением материалов высокой разрешающей силы (Medve et al., (1998) J. Chromatography A 808:153; hydrophobic interaction chromatography (Tomaz and Queiroz, (1999) J. Chromatography A 865:123; two-phase partitioning (Brumbauer, et al., (1999) Bioseparation 7:287); высаживание этанолом; обратно-фазная HPLC; хроматография на силиконой или на катион-обменной смоле, такой как DEAE; хроматофокусирование; SDS-PAGE; высаживание сульфатом аммония; и гель-фильтрация с использованием, например, Сефадекс G-75, могут быть применены. В определенных вариантах осуществления, детергент-аддитивный белок может быть использован без очистки от других компонентов культуральной среды. В определенных вариантах осуществления, компоненты культуральной среды могут быть просто сконцентрированы, например, и потом использованы без дополнительной очистки белка от других компонентов среды для выращивания.
В некоторых вариантах осуществления, клетка может быть культивирована при условиях периодической или непрерывной ферментации. Классические способы периодической ферментации используют закрытую систему, где культуральная среда приготавливается перед началом цикла ферментации, среда инокулируется желаемым организмом(-ами), и ферментация происходит без последовательного добавления каких-либо компонентов к среде. В определенных случаях, рН и содержание кислорода, но не содержание источника углерода, в среде для выращивания может изменяться в течение периодического способа. Метаболиты и клеточная биомасса в периодической системе изменяются постоянно до того времени, когда ферментация останавливается. В периодической системе клетки обычно проходят через статичную lag фазу к log фазе высокого роста и в заключение к стационарной фазе, где коэффициент роста снижается или затухает. В случае невмешательства, клетки в стационарной фазе в конечном счете умирают. В общих чертах, клетки в log фазе продуцируют наибольшее количество белка.
Вариацией стандартной периодической системы является «загрузочно-периодическая» система. В этой системе питательные вещества (т.е. источник углерода, источник азота, соли, О2 или другие питательные вещества) добавляются только тогда, когда их концентрация в культуре падает ниже критического уровня. Загрузочно-периодические системы применимы, когда катаболитная репрессия склонна к ингибированию метоболизма клеток и где желательно иметь ограниченное количество питательных веществ в среде. Показатели актуальной концентрации питательного вещества в загрузочно-периодической системе оцениваются на основе изменений в измеряемых факторах, таких как рН, растворенный кислород и парциальное давление отработанных газов, таких как СО2. Периодическая и загрузочно-периодическая ферментации являются широко распространенными и известны в области техники.
Непрерывная ферментация является открытой системой, где определенная культуральная среда добавляется постоянно в биореактор и такое же количество кондиционной среды одновременно отбирается для процессирования. Непрерывная ферментация обычно поддерживает культуры при постоянной высокой плотности, где клетки преимущественно в log фазе роста.
Непрерывная ферментация позволяет модулирование одного фактора или любого количества факторов, которые влияют на рост клеток и/или концентрацию конечного продукта. Например, в одном варианте осуществления, лимитируюшее питательное вещество, такое как источник углерода или источник азота, поддерживается на фиксированном уровне и все другие параметры разрешены к изменению. В других системах, количество факторов, влияющих на рост, может быть изменено непрерывно, тогда как концентрация клеток, определяемая по мутности среды, сохраняется постоянной. Непрерывные системы преследуют цель поддержания устойчивых условий роста. Так, потеря клеток из-за извлеченной среды может быть уравновешена коэффициентом роста клеток в ферментации. Способы модулирования питательных веществ и факторов роста для процессов непрерывной ферментации, также как и техники для максимизирования уровня образования продукта, хорошо известны.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Нижеследующие примеры приведены для того, чтобы продемонстрировать и дополнительно проиллюстрировать определенные предпочтительные варианты осуществления и аспекты настоящего изобретения, и не должны быть истолкованы как ограничивающие объем изобретения.
Материалы и методы
Экспериментальные техники, использованные для манипуляций с ДНК, были стандартными техниками в области молекулярной биологии (Sambrook et al. Molecular cloning: A Laboratory Manual). Плазмиды были приготовлены и вставки очищены с помощью наборов Qiagen (Qiagen Inc.). Рестрикционные эндонуклеазы и другие ферменты были закуплены у Roche Applied Science (Indianapolis, IN) и использовались, как рекомендовано производителями. Компетентные клетки B. subtilis были приготовлены, как описано Ferrari E. и B. Miller (Bacillus expression: a Gram-Positive Model. In Gene Expression Systems: Using Nature for the Art of Expression. 1999. Academic Press, N.Y.).
ПЦР реакции проводились с ферментом Herculase (Stratagene) в соответствии с инструкциями производителей. Реакции содержали 200 нМ каждого праймера, 1 Ед. Herculase и 200 мкМ каждого dNTP. Термоциклер PxE Thermal Cycler от Hybaid (Thermo) использовался со следующим циклом: денатурация при 94°C в течение 3 мин, далее 30 циклов денатурация при 94°C в течение 30 с, отжиг при 55°C в течение 30 с и протяжка при 72°C в течение 1 мин/1 тпн, подлежащих амплификации. ПЦР реакции затем анализировались на 0,8% агарозном э-геле от Invitrogen.
Геномная ДНК была приготовлена с использование пробирок Eppendorf Phase Lock Gel (Eppendorf) и их протокола.
D-аланин, D-циклосерин и β-хлор-D-аланин были получены от Sigma.
Субтилизиновая проба
Субтилизиновые пробы проводились как ранее описано (Estell, D. V., Graycar, T. P., Wells, J. A. (1985) J. Biol. Chem. 260, 6518-6521) в 0,1 M Tris буфере, pH 8,6, содержащем 1,6 мМ N-сукцинил-L-Ala-L-Ala-L-Pro-L-Phe-p-нитроанилид (Vega Biochemicals). Проба измеряет увеличение в поглощении при 410 нм/мин из-за высвобождения р-нитроанилина. Пробы проводились при условиях фоновой величины. Единица протеазы определена как количество фермента протеазы, которое увеличивает поглощение стандартного раствора, описанного выше, при 410 нм на 1 адсорбционную единицу (AU) в минуту при 25°С в кювете с длиной пути 1 см.
Бактериальные штаммы
Escherichia coli MM294: endA thiA hsdR17 supE44
Штаммы Bacillus subtilis BG2190 (alr-) и BG2189 (alr- CmR) были описаны Ferrari и Yang (1985) (Isolation of an alanine racemase gene from Bacillus subtilis and its use for plasmid maintenance in B. subtilis. Biotechnology, 3, 1003-1007 [1985]).
B. subtilis штамм BG3594: nprE aprE spoIIE degU32 oppA
B. subtilis штамм BG3594comK: Это B. subtilis BG3594, содержащий xylR-PxylA-comK конструкт, как описано в WO 02/14490, который позволяет сделать этот штамм суперкомпетентным (а именно более чем 1% клеточной популяции трансформируемым хромосомальной Bacillus ДНК).
B. subtilis штамм CP3490: Этот штамм является штаммом B. subtilis BG3594comK, в котором alr нокуатирован (та же мутация, что и в BG2190).
B. subtilis штамм CP35491: Этот штамм является штаммом B. subtilis BG3594, в котором alr нокуатирован (та же мутация, что и в BG2190).
B. subtilis MDT01-138: Этот штамм яаляется штаммом B. subtilis BG3594 с амплифицируемой кассетой со следующей структурой: aprE 5'- субтилизин FNA-хлорамфеникол- aprE 5'. Он был амплифицирован в Cm25.
B. subtilis штамм CP4010: Этот штамм является штаммом B. subtilis BG3594comK с амплифицируемой кассетой со следующей структурой: aprE 5'- субтилизин FNA - alr-aprE 5'. Он был амплифицирован с использованием β-хлор-D-аланина.
B. subtilis штамм CP4020: Этот штамм яаляется штаммом BG3594 с амплифицируемой кассетой со следующей структурой: aprE 5'- субтилизин FNA - alr-aprE 5'. Он был амплифицирован с использованием β-хлор-D-аланина.
B. subtilis штамм Hyper1: имеет амплифицированную кассету, кодирующую субтилизин FNA и содержащую хлорамфеникольный маркер.
CP3591: Этот штамм яаляется BG3594 с одной копией субтилизин FNA под aprE промотером в oppA локусе (то есть, всего 1 копия субтилизина).
CP3592: Это CP3591 с одной дополнительной копией субтилизин FNA под aprE промотером между ybdL иd ybdM генами (то есть, всего 2 копии субтилизина).
CP3593: Это CP3592 с одной дополнительной копией субтилизин FNA под aprE промотером в pps локусе (то есть, всего 3 копии субтилизина).
CP3594: Это CP3593 с одной дополнительной копией субтилизин FNA под aprE промотером в nprE локусе (то есть, всего 4 копии субтилизина).
Плазмиды
pDALsub1 была описана в Ferrari et al. 1985. Эта плазмида экспрессирует alr (Ferrari and Yang, Biotechnology, 3, 1003-1007 [1985]).
pBSFNACm (Seq ID NO: 1): эта плазмида является производной pBluescript (Alting-Mees,M.A. and ShortJ.M. pBluescript II: gene mapping vectors. Nucleic Acids Res. 17 (22), 9494 (1989)), содержащей f1 (IG) - межгенную область фага f1; rep (pMB1) - pMB1 репликон, ответственный за репликацию фагмиды; bla (ApR) - ген, кодирующий β-лактамазу, который придает устойчивость к ампициллину; lacZ - 5'-концевую часть гена lacZ, кодирующую N-концевой фрагмент β-галактозидазы; полипептидную экспрессионную кассету, содержащую aprE 5' область, ген, кодирующий субтилизин (FNA), ген устойчивости к хлорамфениколу из pC194 с его промотером, повтор aprE 5' области. Эта плазмида используется для интеграции экспрессионной кассеты в aprE 5' область.
pBSFNAalr (Seq ID NO:2): эта плазмида является производной pBSFNACm, описанной выше. В этой плазмиде B. subtilis alr ген с его собственным промотером замещает ген устойчивости к хлорамфениколу. Эта плазмида используется для интеграции экспрессионной кассеты в aprE 5' область.
Среды
LB и LB агар (LA), как описано в Ausubel, F.M. et al. (eds) "Current Protocols in Molecular Biology". John Wiley and Sons, 1995. LBG 1% является LB средой, дополненной 10 г/л глюкозы. LBSM является LB агар, дополненной 1,6% обезжиренного молока. FNII среда, использованная для исследования продуцирования протеазы, описана в WO 05052146A2. Alr- штаммы выращивались на LB агаре + 100 мг/л D-аланина.
При необходимости хлорамфеникол, ампициллин, циклосерин или β-хлор-D-аланин были добавлены на чашку или в жидкую среду.
Количественный ПЦР (кПЦР)
кПЦР для определения количества копий гена, кодирующего полипептид, представляющий интерес, который должен был продуцироваться (например, субтилизин), был проведен на ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster City, CA). Набор TaqMan® Gene Expression Master Mix был использован, следуя инструкциям производителя (Applied Biosystems, Foster City, CA).
Ферментации
Для тестирования штаммы выращивались в 5 мл LBG 1% в 10-мл пробирках, при 37°С и 250 об/мин. При OD600 примерно ~1, 2,5 мл культуры было использовано для инокуляции 25 мл FNII среды в 250-мл встряхиваемой колбе Эрленмейера. Встряхиваемые колбы были инкубированы при 37°С и 250 об/мин, и образцы питательной среды отбирались регулярно для измерения активности субтилизина.
ПРИМЕР 1
Определение порога чувствительности Bacillus subtilis BG3594 и BG3594, pDALsub1 к β-хлор-D-аланину (CDA)
В первой фазе концентрация β-хлор-D-аланина (CDA), необходимая для ингибирования роста B. subtilis, была определена путем высевания разведений в LB среде штамма BG3594 на LB-агар чашки, содержащие различные концентрации CDA. Как показано в Таблице 1, тогда как BG3594 все еще могут расти при концентрациях 20 мг/л, рост полностью ингибируется при концентрациях 50 мг/л. Во второй фазе pDalsub1 была трансформирована в BG3594 для определения, может ли сверхэкспрессия alr восстановить рост на ингибирующих концентрациях CDA. Как показано в Таблице 1, наличие alr-экспрессирующей плазмиды делает возможным рост на всех протестированных концентрациях CDA. Этот результат показывает, что штамм, содержащий хромосомно-кодируемую экспрессионную кассету «полипептид, представляющий интерес, - alr» может расти на концентрациях CDA более чем 20 мг/л только если произошла амплификация. Другие ингибиторы аланин рацемазы, такие как циклосерин, могут быть использованы вместо CDA.
Устойчивость BG3594 и BG3594, pDALsub1 к возрастающим концентрациям β-хлор-D-аланину на LB агар чашках
** Нет роста.
ПРИМЕР 2
Конструирование штаммов, содержащих хромосомно-кодируемую экспрессионную кассету «полипептид, представляющий интерес, - маркер» (штаммы BG4010 (comK) и BG4020)
Плазмида pBSFNAalr (SEQ ID NO:2) была сконструирована из pBSFNACm (SEQ ID NO:1) следующим образом. Ген B. subtilis alr c его собственным промотером был ПЦР-амплифицирован с использованием хромосомной ДНК в качестве матрицы. Были использованы праймеры EcoRIDrdIalrF (имеющий сайт DrdI; gaagaattcg actaggttgt cttttcgtta gacatcgttt ccctttagc; SEQ ID NO:3) и SmaI-alrR (имеющий сайт SmaI; ggttcccggg ttaattgctt atatttacct gcaataaagg; SEQ ID NO:4). ПЦР продукт был расщеплен DrdI/SmaI и ре-лигирован с большим фрагментом BsmI/StuI-расщепленной pBSFNACm. Лигирование было трансформировано в E. coli штамм MM294 и высеяно на 50 мг/л карбенициллин. Четыре колонии из каждой трансформации были инокулированы в 5 мл LB + 50 мг/л карбенициллин для выделения плазмиды. Результирующий конструкт был назван pBSFNAalr (SEQ ID NO:2).





pBSFNAalr была расщеплена NotI/ScaI (буфер H), что привело к четырем фрагментам, разделяемым на геле. Размеры фрагментов были следующие: 3660-2263-1105-174. Наибольший фрагмент, содержащий кассету 5'-FNA-alr-5', был очищен из геля и лигирован сам на себя. Лигирование было проведено с использованием набора «быстрого лигирования» (Roche). Полученный кольцевой фрагмент ДНК был подвергнут реакции катящегося кольца (набор Amersham) и трансформирован в компетентные клетки либо CP3590 или CP3591 и высеян на LA + 1,6% обезжиренного молока, либо BG3594 или BG3594comK и высеян на LA + 1,6% обезжиренного молока + 50 мг/л CDA. Штаммы с правильным конструктом растут на этих чашках и окружены ореолом вследствие просветления обезжиренного молока из-за экспрессируемой протеазы.
Кассета может быть введена в любой штамм посредством трансформации хромосомной ДНК в штамм, названный в параграфе выше, и селекции на CDA 50. Амплификация кассеты была осуществлена путем высевания штамма штрихом на увеличивающиеся количества CDA (до 200 мг/л). Амплифицированный штаммы имеют больший ореол в случае высевания на 1,6% обезжиренное молоко, демонстрируя, что пассажи штамма на увеличивающиеся количества CDA приводят к амплификации кассеты.
ПРИМЕР 3
Определение количесства копий СУБТИЛИЗИН генов в штамме BG4020 методом кПЦР
Хромосомная ДНК была экстрагирована из штаммов CP3591, CP3592, CP3593, BG4020, BG4020, амплифицирована и концентрация Hyperl ДНК была измерена, образцы были разбавлены до получения одной и той же концентрации во всех образцах. Одинаковое количество ДНК для каждого штамма далее было использовано в кПЦР реакции с праймерами к гену субтилизина (FNA-R2; ccagtgtagc cttgagag; SEQ ID NO:5 и FNA-F2; acaatgagca cgatgagc; SEQ ID NO:6).
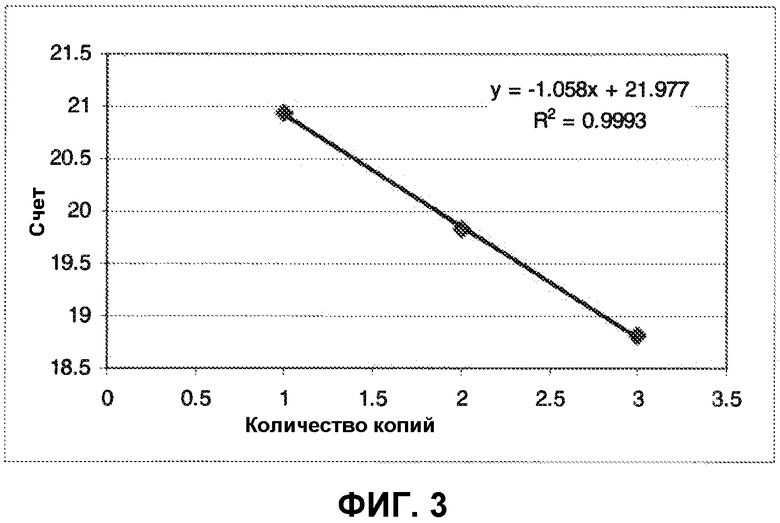
Штаммы CP3591, CP3592 и CP3593, с 1, 2 или 3 копиями гена, соответственно, были использованы для построения калибровочной кривой (Фиг. 3). Эта калибровочная кривая была использована для определения количества копий субтилизина в BG4020, амплифицированном BG4020 и MDT01-138.
Таблица 2 показывает данные, полученные в кПЦР реакциях для штаммов BG4020, BG4020-амплифицированный и Hyperl. Соответствующее количество копий гена было получено по калибровочной кривой, представленной на Фиг. 3. Результаты показывают, что амплификация с alr маркером (BG4020-амплифицированный) является такой же эффективной, как и амплификация с маркером хлорамфеникола (Hyperl); было определено, что оба штамма BG4020-амплифицированный и Hyperl имеют одинаковое количество копий, а именно 4. Нецелое число копий в 4020 может быть результатом для популяции клеток, которая не гомогенно содержит одно и то же количество копий амплифицированного гена; и значение 2,4 представляет собой среднее.
Таблица 2
ПРИМЕР 4
Продуцирование полипептида, представляющего интерес, штаммами, амплифицированными с Cm или alr-содержащей кассетой
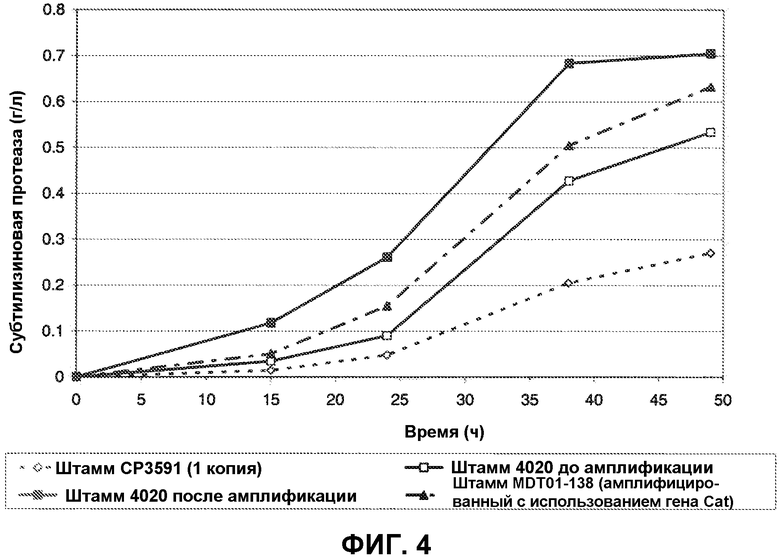
Штаммы для тестирования выращивались в 5 мл LBG 1% в 10-мл пробирке, при 37°С и 250 об/мин. При OD600 примерно ~1, 2,5 мл культуры было использовано для инокуляции 25 мл FNII среды в 250-мл встряхиваемой колбе Эрленмейера. Встряхиваемые колбы были инкубированы при 37°С и 250 об/мин, и образцы питательной среды отбирались регулярно для измерения активности субтилизина. Четыре штамма были протестированы этим путем: BG3591 (содержит одну копию гена субтилизина, не-амплифицируемую), MDT01-138 (амплифицированный с помощью хлорамфеникола), BG4020 (штамм, в который была введена кассета 5'-субтилизин-alr-5'), BG4020 амплифицированный (BG4020 штаммы, которые были пересеяны штрихом на возрастающие концентрации CDA). MDT0l-138 является штаммом, изогенным для Hyperl, - он содержит хлорамфеникол маркерный ген.
Количество субтилизиновой протеазы, продуцированной во встряхиваемых колбах каждым из этих штаммов, показано на Фиг. 4. Как следует из этих графиков, амплификация субтилизинового гена (BG4020 амплифицированный) приводит к более высокому продуцированию протеазы, чем для одно-копийного штамма (BG3591) или для неамплифицированного штамма (BG4020). В дополнение, штамм, полученный путем амплификации с использованием alr маркера (BG4020 амплифицированный), продуцирует больше протеазы, чем штамм, амплифицированный с использованием хлорамфеникол маркера (MDT01-138).
Эти результаты показывают, что ген alr может быть эффективно использован как не-антибиотический, не-эндогенный маркер для амплифицирования экспрессионной кассеты, кодирующей полипептид, представляющий интерес, и вследствие этого продуцирующей высокий уровень полипептида, представляющего интерес.
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННАЯ ПРОДУКЦИЯ БЕЛКА В BACILLUS | 2008 |
|
RU2515112C2 |
| СПОСОБЫ УЛУЧШЕНИЯ ЭФФЕКТИВНОСТИ БЕЛКОВ | 2008 |
|
RU2569106C2 |
| ЭКСПРЕССИОННЫЕ ВЕКТОРЫ ДЛЯ УЛУЧШЕННОЙ СЕКРЕЦИИ БЕЛКА | 2012 |
|
RU2661790C2 |
| CПОСОБ МЫТЬЯ ПОСУДЫ | 2007 |
|
RU2509152C2 |
| СИГНАЛЬНЫЕ ПЕПТИДЫ ТАТ ДЛЯ ПРОДУКЦИИ БЕЛКОВ В ПРОКАРИОТАХ | 2008 |
|
RU2487937C2 |
| Способы и композиции, содержащие варианты сериновой протеазы | 2012 |
|
RU2663114C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ЭНДОПЕПТИДАЗНОЙ АКТИВНОСТЬЮ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2011 |
|
RU2583293C2 |
| СПОСОБЫ УЛУЧШЕНИЯ МНОЖЕСТВЕННЫХ СВОЙСТВ БЕЛКА | 2008 |
|
RU2553550C2 |
| Рекомбинантная плазмида pBU-Sav, обеспечивающая синтез и секрецию белка щелочной протеазы в системе Bacillus subtilis | 2023 |
|
RU2816513C1 |
| ПРОТЕАЗЫ, СОДЕРЖАЩИЕ ОДНУ ИЛИ НЕСКОЛЬКО КОМБИНИРУЕМЫХ МУТАЦИЙ | 2009 |
|
RU2560978C2 |
Группа изобретений относится к способу амплифицирования геномного локуса в бактериальной клетке in vivo, бактериальной клетке-хозяину, его содержащей и способной его амплифицировать, а также культуре бактериальных клеток, способной амплифицировать геномный локус. Способ предусматривает введение локуса со структурой A1-P-M-A2 в популяцию бактериальных клеток-хозяев, приведение указанной популяции клеток в контакт с ингибитором фермента, необходимого для жизнеспособности клетки, культивирование и отбор клеток, которые являются устойчивыми к указанному ингибитору. В структуре локуса А1 и А2 являются прямыми повторами, Р содержит кодирующую последовательность для белка, представляющего интерес, способного экспрессироваться в бактериях, М содержит кодирующую последовательность для фермента, функционально связанную с промотором. Клетки, которые являются устойчивыми к указанному ингибитору, имеют множество копий указанной амплификационной единицы. Группа изобретений обеспечивает увеличение уровня экспрессии белка, представляющего интерес, в клетках-хозяевах. 3 н. и 11 з.п. ф-лы, 4 ил., 2 табл., 4 пр.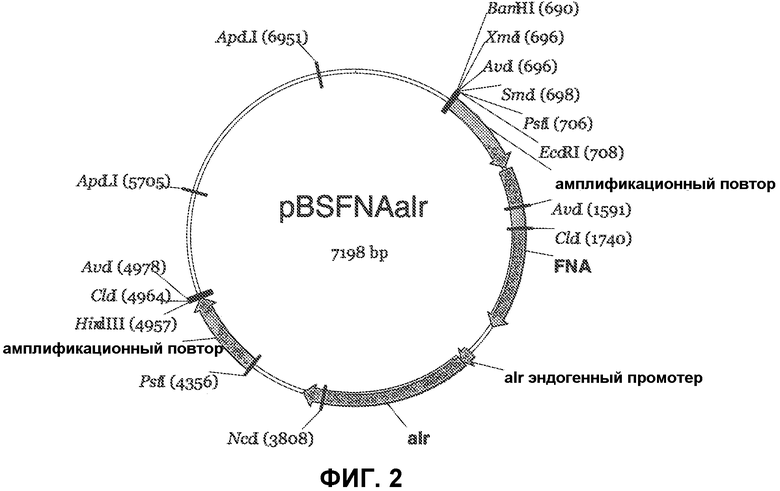
1. Способ амплифицирования геномного локуса в бактериальной клетке in vivo, включающий:
(а) введение локуса со структурой A1-P-M-A2 в популяцию бактериальных клеток-хозяев, где А1 и А2 являются прямыми повторами, Р содержит кодирующую последовательность для белка, представляющего интерес, способного экспрессироваться в бактериях, и М содержит кодирующую последовательность для фермента, функционально связанную с промотором, где указанный фермент является необходимым для жизнеспособности клетки;
(б) приведение указанной популяции бактериальных клеток-хозяев в контакт с ингибитором указанного необходимого фермента; и
(в) культивирование и отбор клеток, которые являются устойчивыми к указанному ингибитору,
где клетки, которые являются устойчивыми к указанному ингибитору, имеют множество копий указанной амплификационной единицы.
2. Способ по п. 1, где указанная бактериальная клетка-хозяин является клеткой Bacillus sp.
3. Способ по п. 1 или 2, где указанный необходимый фермент является ферментом дикого типа, который является эндогенным для указанной клетки.
4. Способ по п. 1 или 2, где указанные бактериальные клетки-хозяева дополнительно содержат инактивированный ген, кодирующий указанный необходимый фермент, где указанный инактивированный ген является эндогенным для указанной клетки и находится в геномном локусе, отличном от указанного геномного локуса со структурой A1-P-M-A2.
5. Способ по п. 1 или 2, где Р содержит экспрессионную кассету.
6. Способ по п. 1 или 2, где A1 содержит промотор, который является функционально связанным с кодирующей последовательностью из Р.
7. Способ по п. 1 или 2, где указанная кодирующая последовательность для указанного необходимого фермента является связанной с промотором, который является эндогенным для указанной кодирующей последовательности.
8. Способ по п. 1 или 2, где указанный белок, представляющий интерес, представляет собой субтилизин.
9. Способ по п. 1, где указанный необходимый фермент представляет собой D-аланин-рацемазу.
10. Способ по п. 1 или 2, где указанный необходимый фермент представляет собой D-аланин-рацемазу, и указанный ингибитор представляет собой β-хлор-D-аланин.
11. Бактериальная клетка-хозяин, содержащая геномный локус и способная амплифицировать его, где геномный локус включает множественные копии амплификационной единицы со структурой:
A1-P-M-A2,
где А1 и А2 являются прямыми повторами, Р содержит кодирующую последовательность для белка, представляющего интерес, способного экспрессироваться в бактериях, и М содержит кодирующую последовательность для фермента, функционально связанную с промотором, где указанный фермент является необходимым для жизнеспособности клетки.
12. Бактериальная клетка-хозяин по п. 11, где указанный белок, представляющий интерес, представляет собой субтилизин, и указанный необходимый фермент представляет собой аланин-рацемазу.
13. Бактериальная клетка-хозяин по п. 11, где указанная амплификационная единица имеет последовательность, представленную в SEQ ID NO: 7.
14. Культура бактериальных клеток, способная амплифицировать геномный локус и представляющая собой популяцию бактериальных клеток по любому из пп. 11-13 в совокупности с соответствующей средой для выращивания.
| US 2003044940 A1, 06.03.2003 | |||
| WO 2005042750 A1, 12.05.2005 | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| BRON P.A | |||
| ET AL | |||
| Use of the alr gene as a food-grade selection marker in lactic acid bacteria // Applied and Enviromental Microbiology, 2002, p | |||
| ВИНТОВОЙ ЗАТВОР ДЛЯ КОЛПАЧКОВ К РУЧКАМ ДЛЯ ПЕРА С РЕЗЕРВУАРОМ ДЛЯ ЧЕРНИЛ | 1926 |
|
SU5663A1 |
| JANNIERE L | |||
| ET AL | |||
| Stable gene amplification in the chromosome of Bacillus subtilis // Gene, 1985, 40, pp. | |||

