Изобретения относятся к биотехнологии и марикультуре, в частности к способам получения посадочного материала морских красных водорослей, и могут быть использованы для культивирования красных водорослей в промышленных масштабах.
Получение проростков и спор как посадочного материала при экстенсивном и интенсивном культивировании красных водорослей, используемых для получения агара и каррагинана, а также для питания некоторых видов моллюсков на ранних стадиях их выращивания, очень актуально в связи с массовым культивированием этих водорослей и отсутствием в мировой практике быстрых и экономичных способов получения посадочного материала.
Известно несколько способов получения посадочного материала при культивировании водорослей. Наиболее распространенный из них - это фрагментирование взрослых слоевищ (Титлянов Э.А., Титлянова Т.В., Скрипцова А.В. Первый опыт плантационного культивирования неприкрепленной формы агароносной водоросли грацилярии в России. Биология моря. 1995. Т. 21, №2. С. 146-156).
Существенным недостатком этого метода является потеря до 30% урожая за счет использования наиболее качественных растений для новых посадок.
Другой способ - получение проростков в массовом количестве из спор дикорастущих водорослей (Rojas R.H., Leon N.M., Rojas R.O. Practical and descriptive techniques for Gelidium rex (Gelidiales. Rhodophyta) culture. Hydrobiologia. 1996. Vol.326/327. P. 367-370).
Однако из естественных зарослей водорослей можно получать посадочный материал только один раз в году, вследствие этого данный способ не нашел широкого применения в практике.
Наиболее близким к заявляемым техническим решениям по количеству существенных признаков и заявляемому результату является способ получения посадочного материала, включающий получение жизнеспособного материала из верхней или периферической частей водорослей (шириной 1-6 мм, которые разрезают на кусочки диаметром 3-4 мм), выдерживание их в питательной среде при освещении люминесцентными лампами (пат. США №6858430, A 01 G 33/00, опубл. 2005 г.).
К недостаткам данного способа следует отнести его сложность и длительность, поскольку культивирование ведется в несколько стадий, при использовании в качестве питательной среды агаровых сред. Сначала культивирование ведут до образования каллюса, а затем уже из каллюса получают соматические эмбрионы (проростки), которые и являются посадочным материалом. Длительность способа получения посадочного материала составляет, по крайней мере, 10,5 недель.
Задача, решаемая заявляемыми изобретениями, - упрощение и сокращение времени получение посадочного материала из водорослей за счет проведения культивирования в жидкой постоянно перемешиваемой культуре.
Поставленная задача решается тем, что в известном способе получения посадочного материала из водорослей, включающем получение жизнеспособного материала из верхушечной или краевой частей водоросли и культивирование его в питательной среде при освещении люминесцентными лампами, согласно изобретению, в качестве питательной среды используют жидкую питательную среду, а в качестве жизнеспособного материала используют микрофрагменты размером не более 1 мм2. Культивирование микрофрагментов ведут по меньшей мере в течение 3-х недель в питательной среде при постоянном перемешивании и аэрировании, с еженедельным обновлением питательной среды.
Культивирование в жидкой питательной среде позволяет значительно упростить и ускорить способ получения посадочного материала (проростков), поскольку дает возможность выращивать проростки непосредственно из ткани водоросли, а не из каллюса. В качестве жидкой питательной среды могут использоваться обычно широко распространенные, в частности Фон Стоша (Von Stosch, 1964. Virkung von Jod und Arsenit auf Meeresalgen in Kultur. Proc. Int. Seaweed Symp. 4, 142-150) или среда Провазоли (Provasoli L., 1964. Growing marine seaweeds. In D. Deviaville and J. Feldmann (eds). Proc. Int. Seaweed Symp. 4. Pergamont Press, Oxford: 9-17).
Выращивание проростков из микрофрагментов в отличие от известных способов культивирования водорослей, в которых в качестве жизнеспособного материала берут фрагменты водоросли, позволяет экономить до 30% урожая.
Установлено, что использование в качестве жизнеспособного материала микрофрагментов водоросли размером не более 1 мм2 обеспечивает оптимальную скорость роста проростков. Увеличение размера исходного микрофрагмента более 1 мм2 ведет к увеличению в таком микрофрагменте отношения субмеристематической к меристематической ткани и, как следствие, к уменьшению начальной скорости роста проростков.
Культивирование посадочного материала осуществляют путем выдержки в жидкой питательной среде в течение по меньшей мере 3-х недель при постоянном перемешивании и аэрировании. Именно данное время выдержки в постоянно перемешиваемой культуре позволяет получить проростки достаточных размеров, которые в дальнейшем могут использоваться не только как посадочный материал, но и как корм для выращивания моллюсков, в частности морского уха. Выдержка менее 3 недель не позволяет получить проростки, которые могли бы использоваться как в качестве корма, так и в качестве посадочного материала. Для получения проростков из микрофрагментов субмеристематической ткани целесообразно выдерживать водоросли 5-6-ти недель, поскольку субмеристематическая ткань начинает спороносить только на 5-й неделе.
Проведение культивирования водорослей в постоянно перемешиваемой и аэрируемой культуре необходимо для постоянного подтока свежей порции питательных веществ, поддержания жизнедеятельности и роста проростков.
Еженедельная смена жидкой питательной среды дает возможность избежать массового развития бактерий, а также поддерживать в жидкой питательной среде оптимальную концентрацию ингредиентов, обеспечивающих оптимальные условия роста проростков.
Для получения микрофрагментов из водоросли используют верхушечную или краевую меристематическую ткань шириной или длиной не более 1 мм в зависимости от морфологических особенностей водорослей.
Использование верхушечной или краевой меристематической ткани позволяет быстро и просто получить проростки. Кроме того, меристематическая ткань не несет на себе эпифитов, что способствует получению чистого незараженного посадочного материала. Для получения микрофрагментов меристематической ткани оптимальным является отрезание от таллома водоросли полоски шириной не более 1 мм, поскольку только в этом случае в образце присутствует достаточно большое количество меристематических клеток, что значительно влияет на скорость роста проростков в начальной стадии культивирования и, как следствие, позволяет получить проростки достаточных размеров и хорошего качества и сократить длительность получения проростков. Более того, это сберегает исходные слоевища водорослей, поскольку позволяет избежать использования для получения посадочного материала значительного количества исходной биомассы, что в конечном результате ведет к упрощению способа получения посадочного материала.
Для рационального использования исходного сырья и получения дополнительного количества посадочного материала для приготовления микрофрагментов используют также субмеристематическую ткань, следующую непосредственно за меристематической тканью.
Использование субмеристематической ткани позволяет получить дополнительное количество посадочного материала в виде тетраспор. Таким образом, заявляемый способ позволяет получать проростки и тетраспоры красных водорослей в любое время года и в любом количестве независимо от периода спороношения водоросли и использовать их как первичный посадочный материал.
Способ осуществляется следующим образом.
Пример 1.
Берут молодые слоевища (5-10 см длиной) спорофитных растений красной водоросли Palmaria palmata (фиг.1А), бритвой или острым ножом отрезают верхнюю половину слоевища, от которой отделяют краевую часть ткани шириной 1 мм (краевую меристему), затем отрезают следующую за ней часть слоевища полосой 1 мм ширины (субмеристематическая ткань). После этого обе полоски ткани рассекают на кусочки с площадью поверхности не более 1 мм2 (в основном от 0,5 до 1 мм2) и помещают по 0.1 г микрофрагментов в разные стеклянные сосуды (объемом 2 литра каждый) с жидкой питательной средой Фон Стош (Von Stosch, 1964), приготовленной на отфильтрованной и стерилизованной морской воде. Сосуды помещают в термостатированную комнату, где поддерживают температуру 10°С. Сосуды освещают люминесцентными лампами белого цвета. Освещенность на поверхности воды составляет не менее 70 мк моль/см2 ·сек. Фотопериод - 12 часов света: 12 часов темноты. Среда в сосуде постоянно аэрируется и микрофрагменты слоевища перемешиваются пузырьками воздуха. Стеклянные сосуды и среда заменяются новыми еженедельно. По мере роста фрагментов, когда их биомасса увеличивается до 5-6 г, половина из них переносится в другой сосуд.
Микрофрагменты культивируются в течение 5 недель. Изменения биомассы (м) и относительной скорости роста (с) микрофрагментов из слоевища красной водоросли Palmaria palmata в культуре представлены в Табл.1. За это время биомасса микрофрагментов, приготовленных из краевой меристемы слоевища, увеличивается более чем в 2000 раз (Табл. 1). Для наглядности на фиг.1 представлены фотографии, отображающие процесс культивирования проростков, где 1А - используемые слоевища Palmaria palmata 1Б - проросток Palmaria palmata после 5-ти недель культивирования, 1В - микроталлом из субмеристематической ткани после 5-ти недель культивирования, 1Г - тетраспорангии после 5-ти недель культивирования микрофрагментов из субмеристематической ткани.
Каждый микрофрагмент вырастает в проросток длиной 2-4 см и шириной 0.5-2 см. Из краевой меристемы одного растения заявляемым способом получают от 100 до 200 проростков. Полученные проростки могут быть использованы как посадочный материал для культивирования водорослей как пища для моллюсков, или их меристематическая кайма может быть использована для получения проростков второй генерации.
Микрофрагменты краевой меристемы, полученные из выращенных пятинедельных растений (вторая генерация) при их культивировании описанным методом, растут с той же скоростью, что и микрофрагменты, полученные из меристемы материнского растения. Культивирование из микрофрагментов второй генерации вели в течение 3-х недель при тех же условиях, что и при получении посадочного материала из материнских водорослей. Результаты представлены в (Табл. 1). Таким образом, вторая генерация проростков, выращенных представленным методом, уже составляет около 20000 молодых талломов, полученных из одного материнского растения за 2 месяца культивирования.
Биомасса микрофрагментов, полученных из субмеристематической ткани за 5 недель культивирования увеличилась в 150 раз (Табл. 1). Средний выросший фрагмент в конце эксперимента представлял собой кусочек ткани, не имеющий меристематических клеток и спороносящий по всей поверхности (фиг.1В). Выход созревших спор из тетраспорангиума (фиг.1Г) начинался после пятой недели культивирования и заканчивался к концу шестой недели (Пример 2). Споры могут быть использованы как посадочный материал для получения мужских и женских гаметофитных растений.
Пример 2.
Пример 2 отличается от Примера 1 тем, что микрофрагменты из меристемы выращиваются не 5, а 6 недель. За это время общая биомасса фрагментов увеличивается в 2000 раз, каждый проросток вырастает до 3-5 см длиной и 1-3 см шириной (фиг.1Б). Скорость роста проростков в течение шестой недели выращивания составляет около 7% в сутки (Табл. 1). Биомасса микрофрагментов из субмеристематической ткани увеличилась более чем в 150 раз, все фрагменты спороносили.
Пример 3.
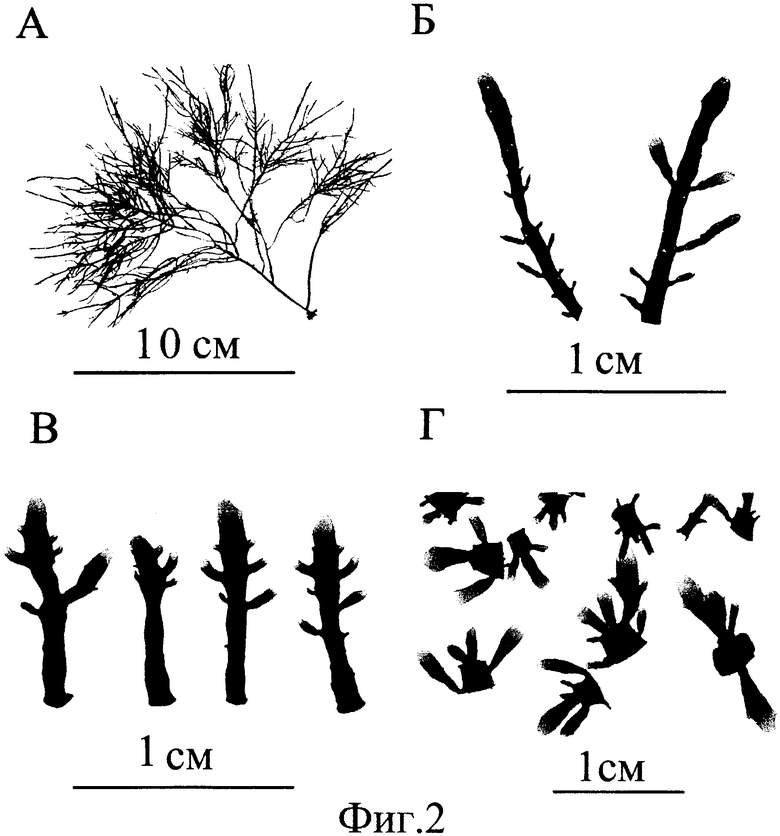
Берут взрослые стерильные растения красной водоросли гелидиума {Gelidium spp.) (фиг.2А) и отрезают верхушечные части длиной около 1 см (фиг.2Б), очищают их жесткой волосяной щеткой от водорослей-эпифитов и промывают под струей стерилизованной морской воды. От верхушечных частей отрезают все видимые точки роста с апикальной (верхушечной) меристемой шириной до 1 мм. Для другой пробы отрезают часть слоевища до 1 мм шириной, следующую за точкой роста (субмеристема). Микрофрагменты слоевища с меристематической и с субмеристематической тканью (каждая проба по 0.1 г) помещают в разные стеклянные сосуды (2 литра) с жидкой питательной средой Провазоли (Provasoli, 1964), приготовленной на фильтрованной и стерилизованной морской воде. Сосуды помещают в термостатированную комнату, где поддерживают температуру 15°С. Сосуды освещают металл-галогеновыми лампами. Освещенность на поверхности воды составляла 120 мк моль/см2 ·сек при фотопериоде 12 час света: 12 час темноты. Среда в сосуде постоянно аэрировалась, а микрофрагменты перемешивались пузырьками воздуха. Стеклянные сосуды и питательная среда заменялись еженедельно.
Микрофрагменты культивировались в течение 6 недель. Результаты увеличения биомассы и скорости роста микрофрагментов из талломов гелидиума в аэрируемой культуре (исходный вес одного образца 0.1 г) представлены в Табл. 2. За время культивирования биомасса микрофрагментов из тканей, содержащих меристему, увеличилась в 450 раз, а из субмеристематической ткани в 920 раз (Табл. 2). После шести недель культивирования фрагменты представляли собой проростки (молодые растения) без ризоидов. Проростки, полученные из тканей, содержащих меристему, имели в среднем по 4 пролификации (веточки) на боковых сторонах (фиг.2В). Проростки, выросшие из субмеристематических частей, образовали значительно большее количество веточек (6-10) как на боковых поверхностях, так и на поверхностях заросших ран (фиг.2Г). Скорость роста меристематических (апикальных) фрагментов в начале эксперимента составляла 2.8±0.5%/сут и 3.8±0.4%/сут в конце эксперимента, максимальная скорость - 4.2%/сут. У субапикальных (субмеристематических) фрагментов 2.1±0.7%/сут, 6.7±1.2%/сут и 7.9%/сут, соответственно (Табл. 2).
Таким образом, в среднем из одного материнского растения было получено около 400 проростков обоих типов. Полученные проростки использовались как посадочный материал при культивировании гелидиума в интенсивно перемешиваемой культуре или для получения проростков гелидиума второй генерации.
Пример 4.
В этом эксперименте проростки гелидиума выращивались в тех же условиях, что и в Примере 3, однако только в течение 5 недель. За это время общая биомасса проростков из меристематической верхушки увеличилась в 3,7 раза и скорость роста в течение последней пятой недели выращивания составляла 4.0% в сутки, а из субмеристематической ткани в 7,8 раза и 5.8% соответственно. Проростки после пяти недель выращивания могли быть использованы в дальнейшем культивировании водоросли с перемешиванием и аэрированием.
Поставленная задача решается также тем, что в известном способе получения посадочного материала из водорослей, включающем получение жизнеспособного материала из верхушечной части водоросли и культивирование его в питательной среде при освещении люминесцентными лампами, согласно изобретению. В качестве питательной среды используют жидкую питательную среду, а в качестве жизнеспособного материала используют клеточные агрегаты, которые получают из ткани, содержащей меристематические и субмеристематические клетки, для чего верхушечную часть водоросли замораживают в течение 10-30 мин, затем их оттаивают при комнатной температуре. Оттаявавшие водоросли помещают в питательную среду на одну-две недели, после чего берут живую ткань, содержащую меристематические и субмеристематические клетки и измельчают ее до клеточных агрегатов, которые затем помещают в питательную среду и культивируют в течение по меньшей мере 3-х недель при постоянном перемешивании, аэрировании и еженедельном обновлении питательной среды.
Культивирование в жидкой питательной среде позволяет значительно упростить и ускорить способ получения посадочного материала (проростков), поскольку дает возможность выращивать проростки непосредственно из ткани водоросли, а не из каллюса. В качестве жидкой питательной среды могут использоваться, в частности, среда Фон Стоша (Von Stosch, 1964) или Провазоли (Provasoli, 1964).
Использование клеточных агрегатов из замороженной-оттаявшей меристематической ткани дает значительное увеличение посадочного материала в виде проростков.
Глубокая заморозка значительно позволяет упростить отбор жизнеспособной меристемы для дальнейшего получения проростков.
Заморозку ведут в течение 10-30 минут. Ведение заморозки в течение менее 10 минут не обеспечивает глубокого замораживания исходного материала и в результате не позволяет получить качественный исходный материал - отделенную от других тканей меристему. Проведение процесса заморозки более 30 минут нецелесообразно, поскольку это никоим образом не сказывается на качестве получения исходных клеточных агрегатов.
Стадия оттаивания обеспечивает получение клеточных агрегатов нужного размера при измельчении водорослей.
Выдержка оттаявших водорослей в питательной среде в течение одной - двух недель позволяет не только подрастить оставшуюся живой меристематическую ткань, но и способствует визуальному проявлению четких контуров между живой и мертвой тканями, так как последняя за это время обесцвечивается. Выдержка менее одной недели не обеспечивает достижения вышеуказанного эффекта и, как следствие, осложняет отделение меристематической ткани и замедляет способ получения жизнеспособного материала, а выдержка более двух недель уже не оказывает влияния на процесс разделения живой и мертвой ткани. Более того, это способствует относительно большему росту субмеристематической ткани, которая отрицательно влияет на количество получаемых проростков из клеточных агрегатов.
Культивирование посадочного материала осуществляют путем выдержки в жидкой питательной среде в течение по меньшей мере 3-х недель при постоянном перемешивании и аэрировании. Именно данное время выдержки позволяет получить проростки достаточных размеров, которые в дальнейшем могут использоваться не только как посадочный материал, но и как корм для выращивания моллюсков, в частности морского уха. Выдержка менее 3-х недель не позволяет получить проростки, которые могли бы использоваться в качестве корма.
Проведение культивирования вородослей в постоянно перемешиваемой и аэрированной культуре необходимо для постоянного подтока свежей порции питательных веществ, поддержания жизнедеятельности и роста проростков.
Еженедельная смена жидкой питательной среды не дает возможности массового развития бактерий в культуре, а также поддерживает в жидкой питательной среде оптимальную концентрацию ингредиентов, обеспечивающих оптимальные условия роста проростков.
Способ осуществляют следующим образом.
Пример 5.
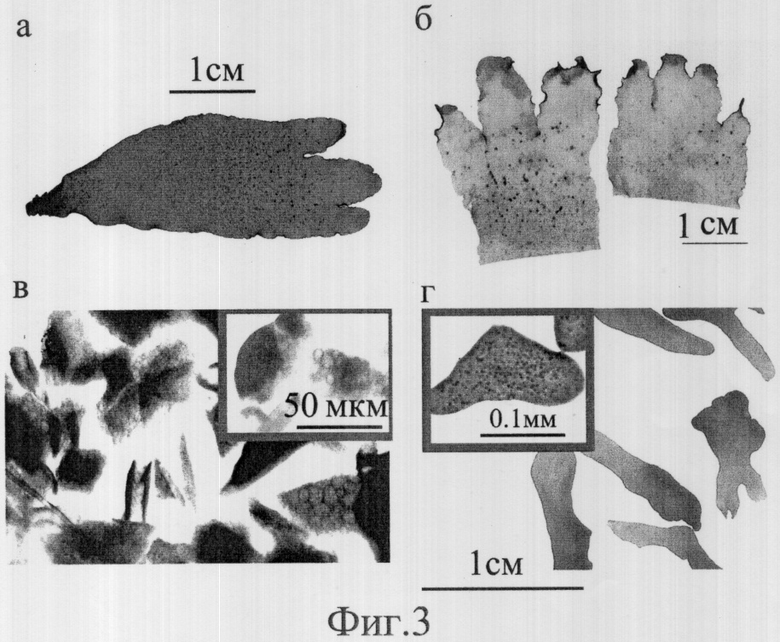
Берут взрослые стерильные спорофитные или мужские гаметофитные растения красной водоросли Palmaria palmata и отрезают верхние половины слоевищ. Эти отрезанные части заворачивают в алюминиевую фольгу и помещают в цилиндр, сделанный из папье-маше (˜ 5 мм толщины). Цилиндр с обеих сторон закрывают резиновыми или корковыми пробками и помещают в шкаф глубокого охлаждения (-80°С) на 10 минут. Затем пакет с водорослями вынимают из цилиндра и размораживают при комнатной температуре. Талломы освобождают от фольги и помещают в сосуды с фильтрованной и стерилизованной морской водой на одну неделю при температуре +10°С и освещенности 20 мкмоль/м2 × сек. Через неделю морскую воду в сосудах заменяли питательной средой. Водоросли выдерживались в питательной среде в течение одной недели. За этот период паренхиматозная ткань слоевищ, убитая замораживанием, обесцвечивалась, а оставшиеся живыми клетки краевой меристемы и меристематических "островков", рассеянных по таллому и сохранивших натуральный цвет, продолжали делиться. После того, как граница между "мертвой" паренхимой и "живой" меристемой становилась видимой, слоевище разделялось на две части: содержащую краевую меристему и содержащую меристематические "островки". Части, содержащие краевую меристему, измельчались (иссекались) микротомным ножом или опасной бритвой на плотной резине или силиконе на клеточные агрегаты, содержащие от нескольких десятков до нескольких тысяч живых и мертвых клеток. Клеточные агрегаты с общей массой 100 мг помещались в 2-литровые стеклянные сосуды, заполненные на 2/3 питательной средой, и выдерживались в течение 6 недель. Клеточные агрегаты перемешивались и аэрировались в сосудах пузырьками воздуха. Культура клеточных агрегатов пальмарии содержалась при температуре +9-10°С, освещенности 70-20 мкмоль/м2 × сек и фотопериоде 12:12 час (свет: темнота).
Результаты изменения биомассы и относительная скорость роста клеточных агрегатов меристемы пальмарии, выделенной методом замораживания-оттаивания, приведены в табл. 3. Фотографии, подтверждающие процесс культивирования проростков, представлены на фиг.3, где 3А - слоевище Palmaria palmata, использованное в эксперименте, 3Б - половинки слоевища после замораживания-оттаивания и выдерживания в питательной среде в течение двух недель, 3В - клеточные агрегаты из замороженной-оттаявшей ткани краевой меристемы, 3Г - проростки Palmaria palmata, полученные из клеточных агрегатов, содержащих клетки краевой меристемы, в течение шести недель культивирования. На вставке - спороносящие микроталломы, полученные из клеточных агрегатов, не содержащих меристематические клетки.
За шесть недель культивирования клеточные агрегаты, содержащие живые клетки, частично (50-70%) образовали проростки, состоящие из паренхиматозной и меристематической тканей (фиг.3Г), частично (30-50%) они не образовали проростков, но наиболее крупные из них, содержащие от нескольких сотен до нескольких тысяч клеток, образовали спорангии с тетраспорами по всей поверхности агрегата микроталлома (фиг.3Г). За шесть недель культивирования биомасса образцов, содержащих клеточные агрегаты краевой меристемы, увеличилась в 25 раз, наибольшая скорость прироста биомассы в культуре была 14.7% в сутки (табл. 3). Проростки, выращенные из клеточных агрегатов краевой меристемы, представляли собой слоевища размером от 0.1 до 1 см (фиг.3Г). Из краевой меристемы одного слоевища пальмарии было получено примерно 500 проростков. Полученные проростки использовались для дальнейшего культивирования пальмарии в бассейнах или использовались для получения следующей генерации проростков из клеточных агрегатов. Тетраспоры, полученные в культуре клеточных агрегатов, могли также служить посадочным материалом для культивирования пальмарии.
Пример 6.
В этом примере использовались части замороженных-оттаявших слоевищ Palmaria palmata, содержащие меристематические "островки" (Пример 5, фиг.3Б). Клеточные агрегаты из этих тканей получали тем же способом, как описано в Примере 5. Клеточные агрегаты из меристематических "островков" культивировали в жидкой питательной среде, как описано выше, в течение 7 недель. Наибольшая скорость роста биомассы в культуре составляла 12.9% в сутки. За это время биомасса образцов увеличилась в 7 раз. Проростки, выращенные из клеточных агрегатов, были размером от 0.1 до 1 см длиной и ничем не отличались от проростков, выращенных из краевой меристемы (Пример 5, фиг.3Г). Из меристемы "островков" одного слоевища Р. palmata можно получить около 100 проростков. В культуре клеточных агрегатов содержались также спороносящие микроталломы. Использование полученного материала такое же, как и в Примере 5.
Пример 7.
Процесс получения посадочного материала представлен на фиг.4, где 4А - исходная водоросль, 4Б - верхушки слоевища красной водоросли гелидиум (Gelidium spp.) после замораживания-оттаивания и выдерживания в жидкой питательной среде в течение двух недель, 4В - клеточные агрегаты, полученные из замороженных-оттаявших верхушек талломов гелидиума, содержащие меристематические клетки, 4Г - проростки гелидиума, полученные из клеточных агрегатов замороженных-оттаявших верхушек талломов, а в таблице 3 приведены изменения биомассы и относительная скорость роста клеточных агрегатов из верхушечной меристемы гелидиума, выделенной методом замораживания-оттаивания.
Берут взрослые стерильные растения агароносной красной водоросли Gelidium spp. и отрезают верхушечные части длиной 2-3 см (Фиг.4А), очищают жесткой волосяной щеткой от эпифитов, промывают струей стерилизованной морской воды и промокают бумажной салфеткой. Отрезанные верхушки растений заворачивались в алюминиевую фольгу и помещались в цилиндр, сделанный из папье-маше (˜ 5 мм толщины) и закрывали пробками с обеих сторон. Цилиндр с образцами помещался в шкаф глубокого охлаждения (-80°С) на 30 минут. После замораживания образцы вынимались из цилиндра и размораживались при комнатной температуре в течение 10 минут, затем освобождались от фольги и помещались в чашки Петри с фильтрованной и стерилизованной морской водой и выдерживались при температуре плюс 10°С и освещенности 20 мкмоль × м-2 × сек-1 в течение одной недели. После этого образцы помещались в питательную среду (Von Stosch, 1964) и выдерживались в течение 2-х недель с ежедневной заменой питательной среды в чашках. За этот период паренхиматозная ткань обесцвечивалась, а меристематические верхушечные (апикальные) клетки оставались живыми и продолжали делиться. Затем верхушечные кусочки длиной 0.5-1 мм отрезались от мертвой ткани и измельчались (иссекались) острым ножом на мельчайшие фрагменты (клеточные агрегаты, содержащие от 10 до 10000 клеток). Образцы с клеточными агрегатами (0.1 г) помещались в 2-литровые стеклянные сосуды с питательной средой, которая перемешивалась и аэрировалась пузырьками воздуха. Культивирование вели в течение 3-х недель. Стеклянные сосуды и питательная среда заменялись еженедельно. За три недели культивирования биомасса образцов увеличилась в 8 раз, а максимальная скорость прироста биомассы в сутки составляла 13.2% (табл. 3). Проростки представляли собой клеточные агрегаты с пролификациями длиной от 50 до 500 мкм (рис. 4Г). Всего из одного верхушечного фрагмента получали в среднем 4 проростка, а из одного растения - около 500 проростков. Проростки использовались для дальнейшего культивирования гелидиума в бассейнах с аэрируемой и перемешиваемой средой.
Таким образом, заявляемые технические решения решают проблемы массового и быстрого получения проростков из красных водорослей для экстенсивного, плантационного и интенсивного культивирования красных водорослей в любое время года. Авторами впервые предложено получать проростки из меристемической, а тетраспоры из субмеристемической тканей водорослей в интенсивно перемешиваемой жидкой культуре. Разработанные способы позволяют также исключить в методике культивирования водорослей такой этап, как сохранение посадочного материала для будущей посадки, что не только упрощает процесс культивирования водорослей, но и значительно удешевляет его.
Gelidium spp.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ПОСАДОЧНОГО МАТЕРИАЛА КРАСНОЙ ВОДОРОСЛИ ГЕЛИДИУМ (GELIDIUM) С РИЗОИДАМИ (ВАРИАНТЫ) | 2006 |
|
RU2318374C1 |
| КЛЕТОЧНЫЕ КЛОНЫ КАМБИЯ, СПОСОБ ИХ ВЫДЕЛЕНИЯ И КОНСЕРВАЦИИ | 2006 |
|
RU2398874C2 |
| РЕКОМБИНАНТНЫЕ КЛЕТКИ РАСТЕНИЙ, СПОСОБ ИХ ПОЛУЧЕНИЯ И СПОСОБ ПОЛУЧЕНИЯ БЕЛКА-МИШЕНИ С ИХ ИСПОЛЬЗОВАНИЕМ | 2014 |
|
RU2636462C2 |
| СПОСОБ РЕГЕНЕРАЦИИ МЕРИСТЕМ | 1995 |
|
RU2120739C1 |
| Способ регенерации растений земляники | 1988 |
|
SU1692409A1 |
| СПОСОБ КЛОНАЛЬНОГО МИКРОРАЗМНОЖЕНИЯ РАСТЕНИЙ | 1994 |
|
RU2080780C1 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ ЧЕРНОМОРСКОЙ КРАСНОЙ ВОДОРОСЛИ GELIDIUM SPINOSUM (GREV.) BORN. ET THUR (RHODOPHYTA) | 2017 |
|
RU2691579C2 |
| Способ получения посадочного материала картофеля и среда для размножения регенерированных растений | 1981 |
|
SU1025373A1 |
| Способ клонального микроразмножения растений сем. Betulaceae | 2016 |
|
RU2627194C1 |
| Способ культивирования растений in vitro разных таксономических групп | 2023 |
|
RU2804965C1 |

Изобретения относятся к биотехнологии и марикультуре, в частности к культивированию красных водорослей в промышленных масштабах. Способ включает получение жизнеспособного материала из верхушечной части водоросли, культивирование его в питательной среде при освещении люминесцентными лампами. В качестве питательной среды используют жидкую питательную среду, а в качестве жизнеспособного материала используют микрофрагменты водоросли размером не более 1 мм2. Культивирование ведут в течение, по меньшей мере, 3-х недель (до 5-6-ти недель) при постоянном перемешивании, аэрировании и еженедельном обновлении питательной среды. Для получения микрофрагменгов водоросли берут верхушечную или краевую меристематическую ткань, либо субмеристематическую ткань. Вариантом способа является то, что в качестве жизнеспособного материала используют клеточные агрегаты, которые получают из тканей, имеющих меристематические и субмеристематические клетки. Для этого верхушечные части водоросли замораживают в течение 10-30 мин, затем их оттаивают и помещают, по меньшей мере, на одну неделю в питательную среду. Далее живую ткань водоросли измельчают, полученные клеточные агрегаты помещают в питательную среду и культивируют в течение, по меньшей мере, 3-х недель при постоянном перемешивании, аэрировании и еженедельном обновлении питательной среды. Разработанные способы позволяют уменьшить количество этапов культивирования водорослей и значительно упростить процесс. 2 н. и 3 з.п. ф-лы, 4 ил., 3 табл.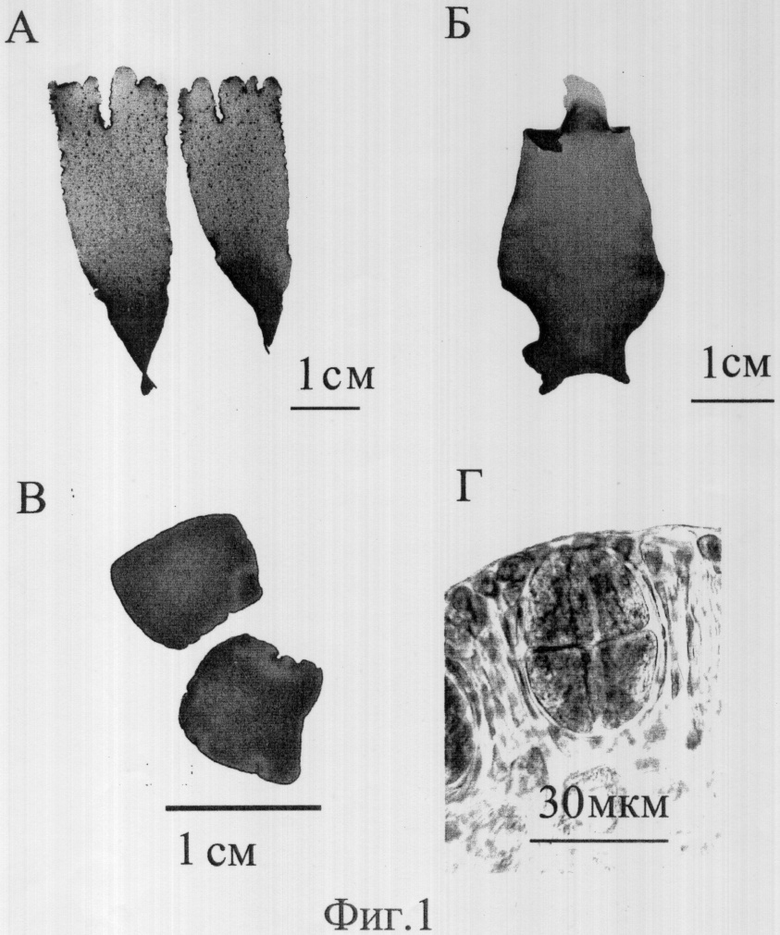
| US 6858430 B1, 22.02.2005 | |||
| Способ культивирования черноморской красной водоросли GRacILaRIa VeRRUcoSa (HUDS) PapeNF | 1987 |
|
SU1634708A1 |
| Способ культивирования морской красной водоросли рода грацилярия | 1988 |
|
SU1551292A1 |
| Способ культивирования морской красной водоросли рода грацилярия | 1989 |
|
SU1799537A1 |

