Область техники изобретения
Данное изобретение относится к новому антибактериальному белку, новому гену, кодирующему белок, способу экспрессии рекомбинантной ДНК с получением нового антибактериального белка.
Предпосылки к созданию изобретения
Широко распространенное применение антибиотиков в течение последних десятилетий для лечения инфекционных заболеваний человека, домашних животных и аквакультурных видов привело к постепенному отбору бактериальных штаммов, резистентных к таким лекарственным препаратам. Проблема микробной резистентности к антибиотикам вызывает особую тревогу системы здравоохранения, которая в течение десятилетий располагала указанными сильнодействующими соединениями для борьбы с заболеваниями и предупреждения распространения патогенных бактерий в окружающей среде. Данная ситуация благоприятствует интенсивному поиску новых антибиотиков, которые отличались бы от используемых сегодня антибиотиков механизмом действия и, следовательно, к которым не имелось бы генов резистентности, отобранных в результате предшествующего и нынешнего применения антибиотиков. Поиски источников новых антибактериальных веществ также затрагивают морскую среду и беспозвоночных животных, живущих в морской среде. В последние годы, антимикробные вещества были выделены из морских беспозвоночных животных. Выделенные вещества включают в себя некоторые антимикробные полипептиды из камчатского краба, мелкого краба и моллюсков. Выделенные полипептиды проявляют антибактериальную активность против различных грамположительных и грамотрицательных бактерий. Гликопротеиды с антибактериальной активностью выделены из морского зайца и двустворчатого моллюска. Одним хорошо известным антибактериальным белком является лизоцим. Однако лизоцим проявляет антибактериальную активность в отношении чрезвычайно ограниченного числа видов грамположительных бактерий, таких как микрококк. Хорошо известно, что белки с активностью, подобной активности лизоцима, встречаются у многих морских беспозвоночных животных, включая морских двустворчатых. Предположительно, такие белки вовлечены в защитные реакции организма, а также ассоциированы с пищеварительными процессами у морских двустворчатых. Бактерии представляют собой источники пищи для таких фильтраторов, как морские двустворчатые, а вещества, способные гидролизовать бактерии, выполняют функции как защиты, так и пищеварения. Хотя известно, что экстракт из внутренних органов арктического гребешка, как было показано, оказывает антибактериальное действие на Aeromonas salmonicida и Vibrio salmonicida in vitro, считается, что утомительный способ получения таких экстрактов в коммерчески значимых объемах препятствует коммерческому использованию таких экстрактов для применения их в уничтожении патогенных организмов рыб. Соответственно, если можно было бы выделить и идентифицировать лизоцимный антибактериальный компонент из внутренних органов арктического гребешка и ген можно было бы выделить из этого источника, это сделало бы возможным экспрессию лизоцимного антибактериального белка, и в данной области техники могла бы быть создана важная система для продукции нового антибактериального белка в количествах, которые сделали бы такой антибактериальный белок реальным кандидатом для применения для ингибирования и уничтожения бактериальных патогенов.
Таким образом, объектом данного изобретения является новый выделенный и охарактеризованный антибактериальный белок, полученный из исландского гребешка - Chlamys islandica.
Другим объектом данного изобретения является новый, выделенный и охарактеризованный ген, экспрессирующий новый лизоцимный белок, обладающий антибактериальной активностью.
Еще одним объектом данного изобретения является способ экспрессии нового гена, полученного из исландского гребешка Chlamys islandica, в системе микроба-хозяина.
Следующим объектом данного изобретения является новая конструкция для экспрессии антибактериального лизоцимного белка.
Другие аспекты, объекты и некоторые преимущества данного изобретения станут очевидными из следующего описания и прилагаемой формулы изобретения.
Краткое описание изобретения
В соответствии с одним аспектом данного изобретения авторами изобретения обнаружен новый антибактериальный белковый хламизин В, выделенный из внутренностей исландского гребешка, который проявляет антибактериальную активность как по отношению к грамположительным, так и грамотрицательным бактериям.
Выделенный из исландского гребешка ген хламизина В, что является другим объектом данного изобретения, представляет ген, используемый для конструирования экспрессирующего вектора, посредством которого достигается экспрессия хламизина В в организме-хозяине.
Подробное описание изобретения
Новый белок согласно данному изобретению (в дальнейшем упоминаемый как "хламизин В") может быть получен из отходов переработки арктического гребешка (Chlamys islandica) посредством способа, при котором внутренности гребешка продавливали через винтовой пресс. Затем выдавленный материал (в дальнейшем упоминаемый как "экстракт внутренностей гребешка") собирали. Потом экстракт внутренностей гребешка осветляли осаждением соляной кислотой. После отстаивания полученного экстракта получали коричневый супернатант. Присутствующий в супернатанте хламизин В затем выделяли с помощью общепринятых способов выделения белков. Во время очистки определяли содержание хламизина В, измеряя лизоцимную активность при рН 5,2. Полученный супернатант смешивали с буферным раствором, содержащим ацетат аммония, и подвергали катионообменной колоночной хроматографии. Из фракций, адсорбированных на катионообменной колонке, фракции, которые характеризовались ферментативной активностью, объединяли. Полученную объединенную фракцию фракционировали и концентрировали, используя ультрафильтрацию. Объединенную фракцию сначала концентрировали, применяя мембрану, которая отсекает вещества с расчетной молекулярной массой 105 Дальтон. Затем растворенное вещество, полученное при описанной фильтрации, впоследствии концентрировали, используя мембрану, позволяющую отсекать вещества с расчетной молекулярной массой 104 Дальтон. Раствор, удержанный при описанной фильтрации, диализовали и повторно подвергали катионообменной колоночной хроматографии. Фракции, адсорбированные на колонке, могут быть элюированы по растущему градиенту концентрации ацетата аммония. Элюированные фракции, которые характеризовались наивысшей ферментативной активностью, объединяли и подвергали аффинной колоночной хроматографии. Элюированные с аффинной колонки фракции, которые характеризовались наиболее высокой ферментативной активностью, объединяли и концентрировали с помощью ультрафильтрации. Затем концентрированный белковый раствор подвергали гель-фильтрации, чтобы выделить хламизин В. Измерение ферментативной активности фракций, полученных с помощью вышеупомянутых способов выделения и очистки, может быть проведено с использованием клеток убитых Micrococcus luteus.
Ген хламизина В получали выделением мРНК из вида Chlamys islandica, из которой затем синтезировали кДНК. На основании аминокислотных последовательностей, полученных из очищенного белка хламизина В, в соответствии с традиционными методами, получали вырожденные олигонуклеотиды и последние использовали в качестве праймеров при амплификации части гена хламизина В. Продукт PCR (ПЦР) гена хламизина В длиной 290 пар оснований секвенировали, используя праймеры PCR как праймеры для секвенирования. Данные о последовательности служили основой для конструирования двух внутригенных олигонуклеотидов, которые использовали как праймеры, применяя ранее описанную кДНК в качестве матрицы. Полученный продукт длиной 490 пар оснований и дополнительный полиА хвост секвенировали, а кДНК и данные о последовательности кДНК использовали при конструировании двух новых олигонуклеотидных праймеров, которые использовали при PCR-амплификации целого гена хламизина В из кДНК или геномной ДНК (выделенной из мускула гребешка). Продукт PCR клонировали в плазмидный вектор, а конечные продукты PCR и, впоследствии, клонированные продукты секвенировали. На основании последовательности кДНК, зрелый транскрипт гена хламизина В из Chlamys islandica содержал 490 нуклеотидов помимо хвоста из аденинрибонуклеотидов.
Следующие примеры приведены в иллюстративных целях и для рекомендации специалисту в данной области, как осуществлять и применять изобретение. Предлагаемые примеры никоим образом не предназначены для ограничения изобретения.
Пример 1
Новый белок, хламизин В, выделяли из отходов переработки арктического гребешка (Chlamys islandica) в соответствии со следующим способом.
К оттаявшему экстракту внутренностей гребешка (30 литров) добавляли соляную кислоту до конечного рН 4,8. Экстракт оставляли при комнатной температуре (24°С) для оседания полученных хлопьев. Через четыре дня получали сформированный коричневый супернатант (15,6 литров). Затем к супернатанту добавляли 55 литров 10 мМ аммоний-ацетатного буфера (рН 5,0) и помещали в холодную комнату (6-8°С). Все дальнейшие стадии при выделении хламизина В проводили при 6-8°С.
Разбавленный супернатант (70,6 литров) разделяли на две порции и использовали в отдельных экспериментах на колонке с S сефарозой FF (11,3×10 см), уравновешенной 10 мМ ацетатом аммония (рН 5,0). После промывания колонки 9 литрами названного буфера, за которым следовали 5,5 литров 10 мМ аммоний-ацетатного буфера (рН 6,0), абсорбированные фракции элюировали 0,4 М аммоний-ацетатным буфером (рН 6,0). Характеризующиеся ферментативной активностью фракции из описанных двух экспериментов объединяли. Скорость элюирования во время хроматографии через сефарозу S составляла 40 см/ч.
Объединенные фракции, составленные из двух вышеописанных экспериментов на сефарозе S, сначала в отдельном эксперименте подвергали ультрафильтрации через устройство для ультрафильтрации амикона СН2, снабженное диском из пористого волокна амикона HIP 100-20. Фильтрацию проводили при средней скорости тока 75 литров/м2/час и рабочем давлении 110×103 Па. Полученное в описанных экспериментах растворенное вещество объединяли и впоследствии подвергали ультрафильтрации, используя мембрану из пористого волокна амикона HIP 10-43 при средней скорости тока 30 литров/м2/час. Удержанное вещество, полученное в результате описанной ультрафильтрации, в дальнейшем концентрировали через ультрафильтр РМ 10, используя перемешивающую ячейку. Удержанный при проведенной фильтрации раствор (40 мл) центрифугировали (14000×g в течение 20 минут) и дважды диализовали против 10 мМ аммоний-ацетатного буфера (рН 5,0).
Затем полученный диализованный белковый раствор повторно подвергали колоночной хроматографии через сефарозу FF (1,6×10 см). После нанесения образца, гель промывали 20 мМ аммоний-ацетатным буфером (рН 5,0) и абсорбированные белки элюировали при ступенчатом увеличении концентрации ацетата аммония. Собирали главный пик ферментативной активности, элюировавшийся при 0,5 М ацетата аммония. Полученный белковый раствор диализовали против 10 мМ аммоний-ацетатного буфера (рН 6,0) и наносили на колонку с сефарозой Blue CL-6B (1,6×13 см), уравновешенной названным буфером. Собирали пик ферментативной активности, элюированный 0,18 М ацетатом аммония. Белки в этом элюате концентрировали ультрафильтрацией. Полученный раствор подвергали гель-фильтрации на колонке с сефакрилом S 200 HR (2,6×93 см), уравновешенным 0,1 М аммоний-ацетатным буфером рН 5,0. Собирали элюированные фракции, содержащие ферментативную активность. Аликвоты из каждой фракции подвергали электрофорезу в полиакриламидном геле в присутствии додецилсульфата натрия (SDS PAGE), используя быструю систему (Amersham Pharmacia Blotech) с предварительным приготовлением быстрых гомогенных гелей с высокой плотностью. Стандартный набор пептидов фирмы Pharmacia (область Mr 2512-16949 Дальтон) использовали как стандартные пептиды и гели красили серебром, как описано W. Ansorge (1983) в Electrophoresis'82, рр 235-242. Фракции, показавшие одну белковую зону ˜10 кДа при SDS PAGE, объединяли и белок концентрировали ультрафильтрацией, используя мембранный диск YM2.
Указанным выше способом получали 1,45 мг хламизина В из 30 литров экстракта внутренностей гребешка. Количество белка определяли, используя белковый анализ Bio-Rad в соответствии с Bradford (1976) Anal. Biochem., 72, рр 248-254. Чистоту препарата хламизина В контролировали с помощью SDS PAGE, используя 10% гель в бис-трис-буфере NOVEX NuPAGE и рабочий буфер MES SDS в соответствии с протоколами NuPAGE-электрофореза. Гель окрашивали кумасси G-250, используя набор коллоидного голубого красителя NOVEX. В результате получали одну окрашенную зону.
Аминокислотный анализ последовательности белка хламизина В проводили анализом состава после вакуумного гидролиза. Гидролизаты анализировали на автоматическом аминокислотном анализаторе модели 421 (Applied Biosystems).
Получены следующие результаты
N-концевые аминокислотные последовательности хламизина В анализировали с помощью автоматической деградации по Эдману (Edman), используя белковый секвенатор модели 477А (Applied Biosystems) и секвенирующую систему HP G 1005A. В результате получена предварительная последовательность из 30 аминокислот.
Для определения аминокислотных последовательностей пептидных фрагментов после расщепления бромцианом, хламизин В восстанавливали и алкилировали 4-винилпиридином (Fridman М., Nilsen R., Cornwell III G.G., Husby G.& Sletten K. (1970) J. Biol. Chem., 245, pp 3868-3871. Расщепление бромцианом проводили на восстановленном и алкилированном хламизине В и полученные полипептиды разделяли с помощью ВЭЖХ с обращенной фазой, используя колонку Pep-S C2/C18 (Amersham Pharmacia Biotech), как описано Foss, G.S., Nilsen, R. & Sletten, K. (1988) Scand. J. Immunol., 47, pp 348-354. Пики собирали и брали для деградации по Эдману. В результате были получены аминокислотные последовательности, представленные в SEQ ID No:1 и 2. SEQ ID No:1 согласуется с предварительной N-концевой последовательностью. В SEQ ID No: 2 никакие дополнительные аминокислоты после последней глутаминовой кислоты не были определены, что указывает на достижение С-конца.
Последовательности записывали слева направо в направлении от амино к карбоксильному концу в соответствии со стандартной номенклатурой, последовательности аминокислотных остатков обозначали однобуквенным кодом, как указано ниже.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID No:l
Характеристики последовательности
a) длина: 25 аминокислот
b) тип: аминокислота
c) тип молекулы: пептид
Описание последовательности.

SEQ ID No: 2
Характеристики последовательности
a) длина: 29 аминокислот
b) тип: аминокислота
c) тип молекулы: пептид
Описание последовательности.

Пример II
Исследование антибактериальной активности
Антибактериальную активность хламизина В, полученного в примере I, испытывали против различных грамположительных и грамотрицательных бактерий. Тестирование антибактериальной активности основано на классическом способе микротитрационного разведения среды, рекомендованном Национальным комитетом лабораторной безопасности и стандартов (NCLSS), который описан D. Amersterdam (1996) в "Antibiotics in Laboratory Medicine", Fourth Edition, pp 52-111.
Способ тестирования антибактериальной активности
Исследования бульонов проводили в стерильных, 96-луночных, круглодонных, полипропиленовых микротитрационных планшетах Costar 3790 (Corning Costar Corporation). Серийные дублированные разведения хламизина В делали в 0,01% уксусной кислоте (аналитической степени чистоты, Merck), 0,2% бычьем сывороточном альбумине (Cat. No. A-7511, Sigma). Концентрация хламизина В в опытных образцах составляла от 250 мкг/мл до 1,95 мкг/мл. Выращенные в течение ночи бактериальные культуры в среде разбавляли для получения 103-106 колониеобразующих единиц/мл. В каждую лунку добавляли 100 мкл бактериальной суспензии и добавляли по 11 мкл опытного образца с различными концентрациями и инкубировали при 12°С или комнатной температуре в течение 24-48 часов. С помощью электронного смешивающего устройства Ika-Vibra-VXR (Janke & Kenkel) осуществляли смешивание культур во время инкубации. После инкубации исследовали степень клеточной пролиферации путем измерения поглощения при 620 нм, используя спектрофотометр SPECTRAmax Plus Microplate (Molecular Devices). Минимальную ингибиторную концентрацию (MIC) определяли как наименьшую концентрацию, при которой оптически не наблюдали никакого бактериального роста. Аликвоты из лунок, соответствующие MIC, помещали на питательный агар и после 48 часов инкубации определяли количество колониеобразующих единиц (CFU). Минимальная бактериальная концентрация (МВС) соответствовала наименьшей концентрации, которая полностью подавляла образование колоний.
Действие хламизина В испытывали против различных бактерий. В таблице, ниже, суммированы результаты проведенных экспериментов. Хламизин В, исследованный при низких концентрациях в культуре, ингибирует рост патогенных бактерий рыб (см. фигуру ниже). Аликвоты из лунок, соответствующие величинам MIC, помещали на питательный агар и после 48 часов инкубации определяли количество колониеобразующих единиц. Полученные значения MIC и МВС составляли 0,6-1,25 мкМ и 1,25-2,5 мкМ соответственно.
Хламизин В оказался менее эффективным по отношению к другим испытанным бактериям, см. таблицу ниже. При концентрации 2,5 мкМ в культуре хламизин В ингибировал рост L. monocytogeneses после 96 часов инкубации, В. cereus, S. epidermidls и Е. faecalis, всех после 24 часов инкубации. При указанной концентрации в культуре хламизин В не оказывал действия на рост других исследованных бактерий. Увеличение концентрации хламизина В в культуре до 10 мкМ при рН 5,7 приводило к полному ингибированию роста Е. coil, P. aeruglnose, P. mirabilis, S. epidermidis и Е. faecalis после 24 часов инкубации. В описанных экспериментах выращенные в течение ночи культуры бактерий разбавляли в 104 раз до того, как бактериальную суспензию использовали в анализе. В описанных экспериментах после 24-часовой инкубации дельта поглощения при 620 нм в контрольных лунках составляла 0,25-0,4. Это давало величину MIC для хламизина В на названных бактериях ˜10 мкМ.
Полученные результаты показывают, что хламизин В проявляет антибактериальное действие относительно грамположительных и грамотрицательных бактерий, включая патогенные бактерии рыб, которые вызывают разрушение пищи, и бактерии, которые вызывают инфекции у человека.
Действие хламизина В на бактерии:
SDS-PAGE-электрофорез и секвенирование аминокислотной последовательности очищенного белка хламизина В выявили зрелый и активный белок приблизительно из 100 аминокислот, начиная с серина или аланина (соответствующего положению 18 в продукте, кодированном кДНК) и заканчивающийся глутаминовой кислотой (соответствующей положению 121 в белке, кодированном кДНК). Это позволяет предположить, что активный (ферментативный и антибактериальный) белок содержит 104 аминокислоты, хотя не выявлено никакого участка расщепления для удаления 16 аминокислот с С-конца белка. Высказано предположение, что предполагаемый процессированный продукт из 104 аминокислот имеет М.м.=11402 Да, pI=6,70 и общий заряд минус 2.
Белковая последовательность предполагаемого хламизина В, кодированная кДНК, находится в полном соответствии с аминокислотной последовательностью, полученной из активного белка (один N-концевой фрагмент из 25 остатков и один С-концевой фрагмент из 29 остатков).
Пример III
Идентификация и клонирование гена хламизина В
Выделение мРНК, синтез кДНК и амплификация гена хламизина В
мРНК выделяли из вида Chlamys Islandica, используя систему PolyATract System® 1000 (Promega), из которой синтезировали кДНК с использованием набора для синтеза кДНК Smart™PCR (Clontech). На основе аминокислотных последовательностей, полученных из очищенного белка хламизина В, изготовляли вырожденные олигонуклеотиды ChlaN4 и ChlaCT и утилизировали как праймеры при амплификации части гена хламизина В, применяя набор кДНК-РСТ Advantage® (Clontech). PCR-продукт хламизина В из 290 пар оснований секвенировали, следуя инструкциям набора для термо-секвеназноготм радиомеченного терминаторного циклического секвенирования (Amershaim), с использованием праймеров PCR в качестве праймеров для секвенирования. Сведения о последовательности послужили основанием для конструирования двух внутригенных олигонуклеотидов. Полученные олигонуклеотиды, CmarF и CmarR, применяли в качестве праймеров в реакциях 3'- и 5'-RACE соответственно, проводимых согласно набору для амплификации кДНК Marathonтм (Clontech), используя ранее описанную кДНК как матрицу. Полученные 3'- и 5'-RACE-продукты, представляющие продукт кДНК из 490 пар оснований и дополнительный полиА-хвост, секвенировали и сведения о последовательности кДНК 3'- и 5'-концов учитывали в конструкции двух новых олигонуклеотидных праймеров ChlaBF и ChlaBR, которые использовали при PCR-амплификации целого гена хламизина В из кДНК или геномной ДНК (выделенной из мускула одного гребешка), и продукты PCR клонировали в плазмидный вектор, содержащийся в наборе для клонирования pCR-ScriptтмAmp SK(+) (Stratagene). Конечные продукты PCR и впоследствии клонированные продукты секвенировали и проводили соответствующий компьютерный анализ последовательности.
Последовательность гена хламизина В, последовательность кодированного белка и характеристики последовательностей
Последовательность кДНК из 490 нуклеотидов и полиА-хвост ([а]n) и кодированный белковый продукт хламизина В из Chlamys islandica, содержащий 137 аминокислот. Установленный сигнальный пептид из 17 остатков записан курсивом, а последовательность из 104 аминокислот предполагаемого зрелого активного белка выделена. Полагают, что С-концевые 16 остатков удаляются с помощью неизвестного механизма. Нуклеотидные варианты, которые, как полагают, представляют аллельные варианты, представлены строчными буквами выше последовательности кДНК, и полученные аминокислотные варианты представлены ниже белковой последовательности. Высоко консенсусный 3'-сигнал полиаденилирования подчеркнут, а потенциальный 5'-связывающий рибосому участок, перекрывающий инициирующий кодон, подчеркнут и выделен.
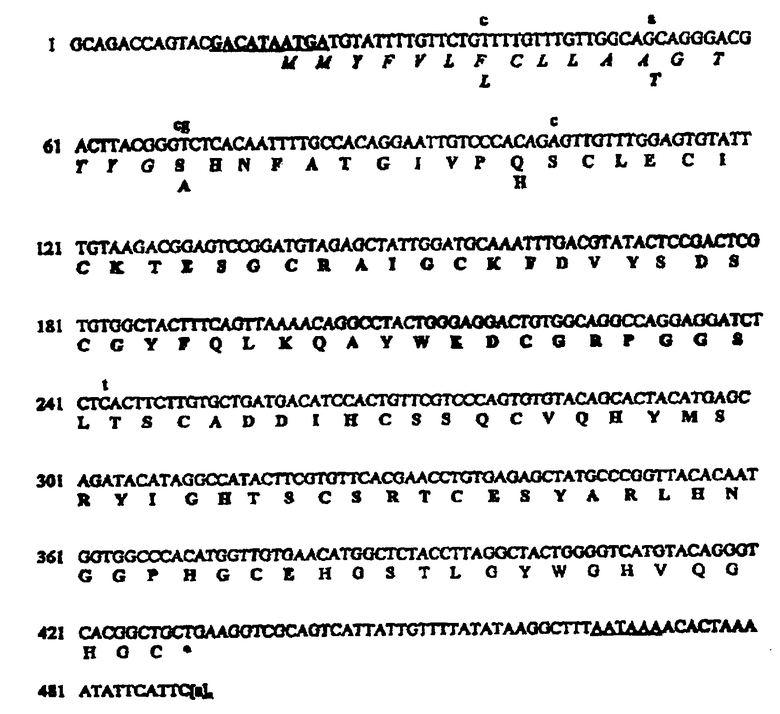
Основанный на последовательности кДНК зрелый транскрипт гена хламизина В из Chlamys islandica содержит 490 нуклеотидов плюс хвост из аденинрибонуклеотидов. Открытая рамка считывания из 411 нуклеотидов может кодировать белок из 137 аминокислот, начиная с двух последовательных метионинов. Последовательности кДНК и геномные последовательности демонстрируют альтернативные нуклеотиды в положениях 36 (Т или С), 52 (G или А), 69 (G или С), 70 (G или Т), 102 (G или С) и 243 (С или Т). Эти чередования, обнаруженные в кДНК, а также в геномных копиях гена хламизина В, позволяют предположить, что Chlamys islandica содержит аллельные копии гена и аналогичные количества транскриптов, очевидно, образуются из аллелей. В результате нуклеотидных вариантов появляются следующие аминокислотные варианты: Phe или Leu в остатке 7, Ala или Thr в остатке 12, Ser или Ala в остатке 18 и Gln или His в остатке 28. Нуклеотидные вариации 69 и 243 не оказывают влияния на кодируемые аминокислоты. Белок из 137 аминокислот, из которого 17 N-концевых остатков, которые, как было предсказано, образуют расщепляемый сигнальный пептид независимо от остатка (серин или аланин) в положении 18, имеет предсказанную молекулярную массу (М.м.) 14964 Да, общий заряд составляет минус 2 и рассчитанная изоэлектрическая точка (pI) соответствует 6,90.
Рекомбинантная продукция хламизина В
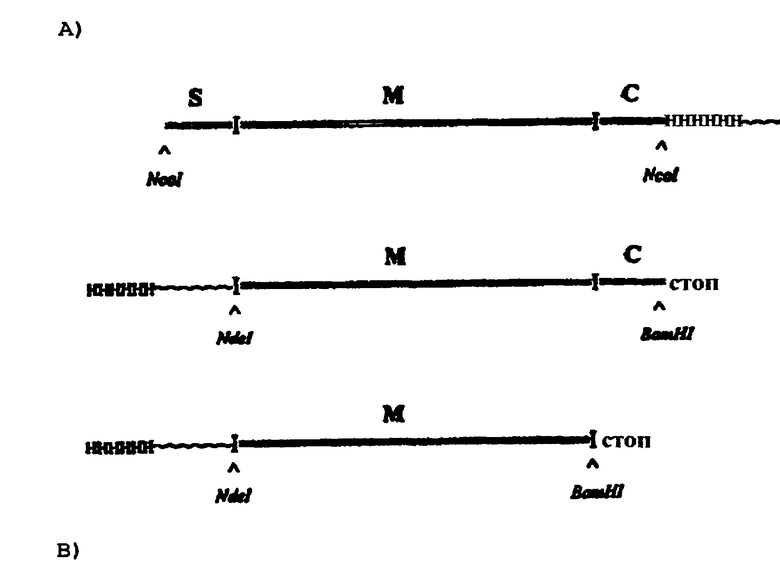
Использовали PCR для продукции трех различных конструкций гена хламизина В, содержащих кодирующие участки для целого продукта гена хламизина В, включая сигнальный пептид плюс зрелый активный белок плюс карбоксиконцевой участок (SMC), зрелый активный белок плюс карбоксиконцевой участок (МС), или зрелый активный белок (М). Следующие олигонуклеотиды использовали как праймерные пары в описанных конструкциях хламизина В: Sig + Cnco для SMC-продукта, Mnde + Cbam для МС-продукта и Mnde + Mbam для М-продукта. Полученные олигонуклеотиды были сконструированы с включением соответствующих последовательностей узнавания фермента рестрикции для последующей рестрикции продуктов PCR. SMC-кодирующая ДНК была вырезана посредством Ncol, a Ndel и BamHI использовали в комбинации, чтобы вырезать оба продукта PCR, кодирующие МС или М. Одновременно, плазмидный рЕТ15b (Novagen) был рестриктирован теми же ферментами и использован в качестве вектора для процессированных продуктов PCR после лигирования с индивидуальными конструкциями ДНК. После трансформации клеток Е. coli и последующих выделений плазмид из клеток, отобранных в результате роста в присутствии ампициллина, рекомбинантные плазмиды рЕТ-15b, несущие вставки SMC, МС и М, были секвенированы для подтверждения вставленных последовательностей и лигирований с сохранением рамки считывания с содержащими вектор последовательностями, кодирующими конец с 6 остатками гистидина и участок расщепления тромбина. Как было предсказано, сплавы SMC с С-концевым пептидом слияния и МС и М с N-концевым пептидом имеют молекулярные массы 19,8 кДА, 15,4 кДА и 13,6 кДа соответственно. Наконец, три подтвержденные рекомбинантные конструкции были введены в клетки Е. coli AD494 (DE3) (Novagen) для экспрессии продуктов хламизина В. После индукции IPTG [1 мМ] в течение двух часов при 21°С клетки собирали и экстракты подвергали электрофорезу SDS-PAGE, используя 10% гель в бис-трис-буфере NuPage™ (Novex) и MES-буфер, с последующим окрашиванием кумасси для определения присутствия рекомбинантных белков. Как видно из следующей таблицы, соответствующие клеточные экстракты показывают присутствие специфических белковых зон, имеющих кажущиеся молекулярные массы приблизительно 20 кДа для SMC, 16 кДа для МС и 14 кДа для М. Молекулярные массы названных специфических белков полностью соответствуют предсказанным значениям. Таким образом, три формы хламизина В продуцировались в высоких уровнях клетками AD494 (DES). Также заслуживающим внимания является присутствие форм хламизина В, МС и М в периплазме, несмотря на отсутствие сигнальных пептидов в этих конструкциях.
Рекомбинантный хламизин В, продуцируемый в клетках Е.coli AD494 (DES)
Три различные конструкции гена хламизина В получали посредством PCR и вставляли в плазмидный вектор рЕТ-15b для гетерологической экспрессии в клетках Е. coli. Конструкции для кодирующих последовательностей, их сливающиеся клетки на основе вектора и участки ферментативной рестрикции для инсерции в вектор проиллюстрированы на схеме А). Анализ SDS-PAGE после окрашивания белков в геле представлен на схеме В), представляющей белки из периплазмы на дорожках 2-5 и клеточные белки на дорожках 6-9 из SMC (3 и 7), МС (4 и 8) иМ (5 и 9) вместе с содержащими вектор контрольными клетками (2 и 6). На дорожке 1 находится внутренний стандарт с 12 образцами молекулярных масс Novex Mark.
Олигонуклеотидные праймеры, использованные для клонирования
Подчеркнутые последовательности представляют собой последовательности не хламизина В, добавленные к олигонуклеотиду, чтобы обеспечить участками ферментативной рестрикции, сигналами остановки транслокации, или лигирование при сохранении рамки считывания с пептидами слияния, кодированными вектором.

Специальные примеры представленного описания следует рассматривать главным образом как иллюстрацию. Различные изменения помимо описанного не должны вызывать сомнения у специалистов в данной области и такие изменения должны быть поняты как составляющие часть данного изобретения, поскольку они соответствуют идее и компетенции прилагаемой формулы изобретения.
Список последовательностей





Изобретение относится к биотехнологии. Предложены варианты белка, обладающего лизоцимной активностью. Описано антибактериальное средство на основе белка. Раскрыта молекула ДНК, кодирующая указанный белок, и прокариотическая клетка, содержащая указанную молекулу ДНК. Описан способ получения белка с использованием прокариотической клетки. Использование изобретения упрощает получение антибактериального средства в коммерчески значимых объемах. 7 н. и 6 з.п. ф-лы.
SHNFATGIVPQSCLECICKTESGCRAIGCKFDVYSDSCGYFQLKQAY WEDCGRPGGSLTSCADDIHCSSQCVQHYMSRYIGHTSCSRTCESYARLHN GGPHGCE необязательно по меньшей мере с одной из аминокислотных замен, выбранных из группы, состоящей из Ala в остатке 1, His в остатке 11 и Pro в остатке 101.
| MYRNES et al | |||
| Recovery of lysozyme from scallop waste | |||
| Preparative biochemistry | |||
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| СИНГЕР М., БЕРГ П | |||
| Гены и геномы | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Пер | |||
| с англ | |||
| - М.: Мир, 1998, т.2, с.286. | |||

