Изобретение относится к биофармакологии, биотехнологии, в частности к генной и белковой инженерии, и представляет собой сконструированную in vitro плазмидную ДНК, обеспечивающую биосинтез фактора свертываемости крови VII человека, и линию клеток-продуцента, содержащей эту плазмиду и дающей высокий выход целевого белка.
Фактор коагуляции VII человека представляет собой редкий белок плазмы - гликопротеин с молекулярным весом порядка 50 кДа, который циркулирует в крови в форме одноцепочечного зимогена. Одноцепочечный фактор VII может быть превращен в активный двуцепочечный фактор VIIa при помощи нескольких различных протеаз плазмы, например, фактора XIIa. Такая активация приводит к образованию двухцепочечного белка, который далее участвует в процессе свертывания крови, приводящем к образованию фибринового сгустка.
Лекарственная форма фактора VII (Новосевен) используется при лечении пациентов, страдающих дефицитом факторов VII и IX, пациентов с врожденной и приобретенной гемофилией с антителами к факторам свертывания VIII и IX, а также больных, страдающих болезнью Виллебранда. Кроме того, фактор VII эффективен для подавления кровотечений различного происхождения.
В настоящее время существует несколько биотехнологических систем, позволяющих получать указанный белок. Известна плазмида pLN174, содержащая ген фактора VII (патент США 6329176), и продуцент на ее основе. Однако уровень экспрессии белка, достигаемый при использовании только указанной плазмиды, весьма низок (0,18 мкг/мл). Соэкспрессия с протеазой КЕХ-2 дает более высокое значение - 0,90 мкг/мл, но при этом имеется вероятность получения неактивных форм белка, которые не обладают коагуляционной активностью, за счет протеолитической деградации (согласно патенту США 5580560).
Кроме того, существует ряд продуцентов, позволяющих получать модифицированный фактор VII, отличающийся от природного заменой одной или нескольких аминокислот (см. патенты США 6960657, 691123, 6905683; заявки РФ 2004112768, 2003131077, 2003110419), но при использовании таких белков для получения лекарственных препаратов сохраняется опасность возникновения иммунного ответа.
Наиболее близким аналогом настоящего изобретения является плазмида, содержащая ген фактора VII, и продуцент на ее основе, использованный для получения этого белка в клетках млекопитающих, полученные способом, предложенном в патенте РФ 2122583. Синтез фактора в этом изобретении основан на получении фрагментов ДНК, кодирующей целевой белок, с последующей их сборкой в единую последовательность. Недостатком указанной плазмиды является трудоемкость сборки из нескольких элементов требуемой последовательности и, как следствие, высокая себестоимость, а также небольшой выход целевого белка (порядка 1 мкг/мл).
Таким образом, основным недостатком имеющихся продуцентов является низкий выход целевого белка.
Настоящее изобретение решает задачу конструирования плазмидной ДНК, содержащей последовательность рекомбинантного фактора VII, и создания высокопродуктивной линии клеток-продуцента, позволяющей получать рекомбинантный фактор VII с выходом 40 мкг/мл. Такой выход в десятки раз превышает выходы существующих в настоящее время продуцентов.
Таким образом, технический результат заявленного изобретения - повышение выхода белка фактора VII полученным продуцентом.
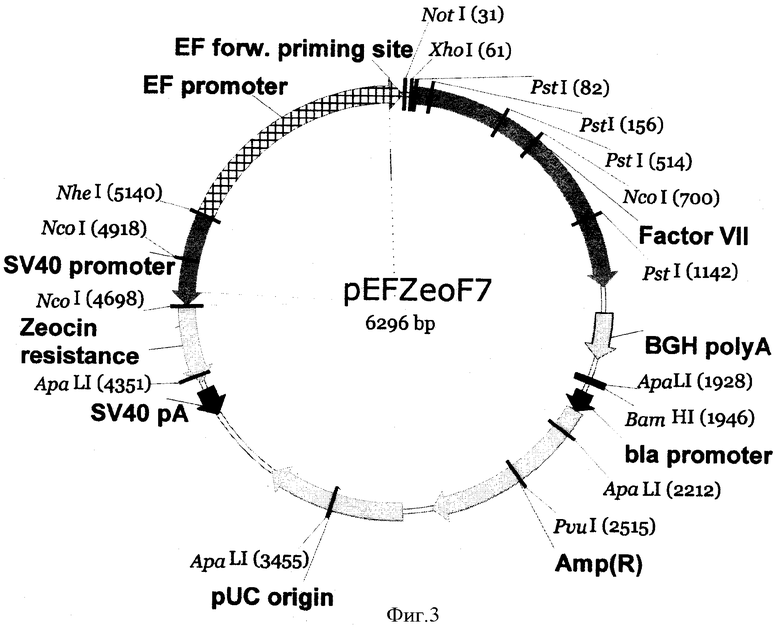
Поставленная задача решается за счет конструирования рекомбинантной плазмидной ДНК pEFZeoF7, состоящей из 6296 п.о., со следующими признаками:
- имеет молекулярную массу 4,16 МДа;
- кодирует полипептид фактора VII;
- включает: XhoI-BgIII/BamHI-фрагмент ДНК плазмиды pEFZeoF7;
ген Amp(R), обеспечивающий устойчивость трансформированных плазмидой pEFZeoF7 клеток к ампициллину; ген, обеспечивающий устойчивость плазмидой pEFZeoF7 клеток к зеоцину; уникальные участки узнавания следующих рестриктаз - NotI (31 п.о.), XhoI (61 п.о.), PstI (1142 п.о.), BamHI (1946 п.о.), PvuI (2515 п.о.).
Заявленное изобретение поясняется на следующих фигурах, где:
на Рфиг.1 представлены:
А - нуклеотидная последовательность ОТ-ПЦР фрагмента, содержащая кДНК гена коагуляционного фактора VII человека: обозначены фланкирующие рестрикционные сайты, открытая рамка считывания подчеркнута; Б - соответствующая ей аминокислотная последовательность фактора VII человека, выделен лидерный пептид.
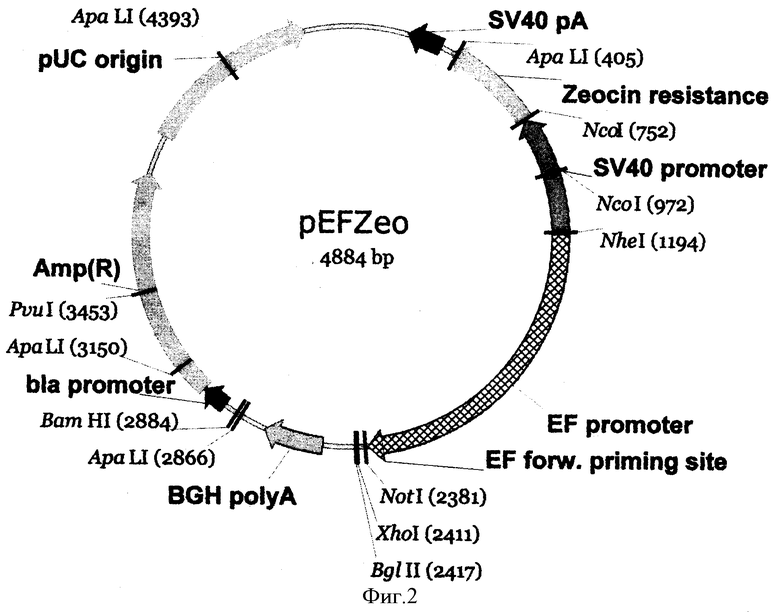
На фиг.2 - карта вектора pEFZeo.
На фиг.3 - карта рекомбинантной плазмидной ДНК pEFZeoF7.
На фиг.4 представлены:
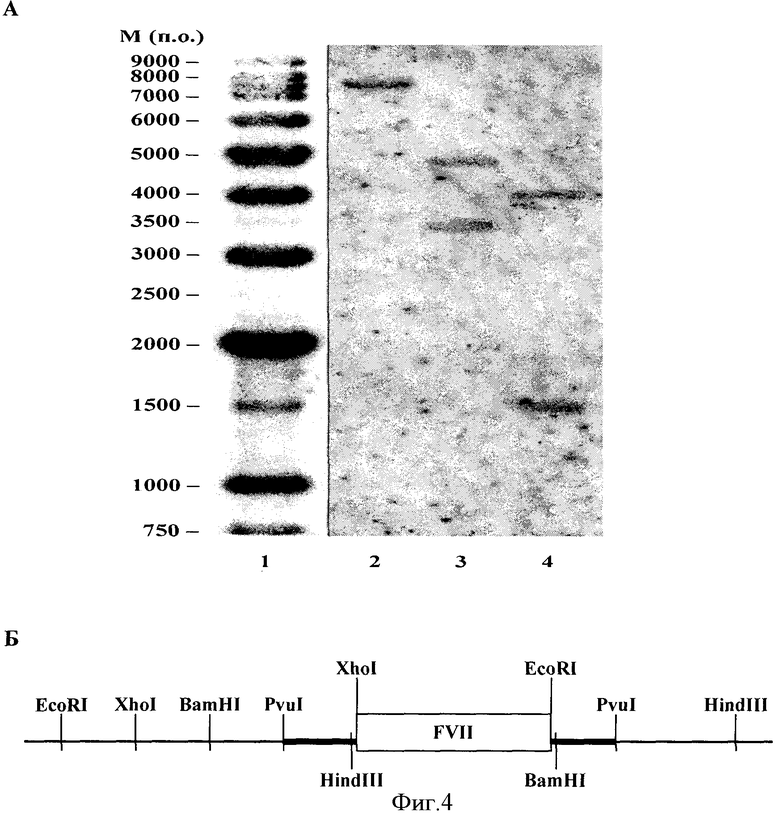
А - гибридизация по Саузерну рестрицированной геномной ДНК с радиоактивно меченой последовательностью, кодирующей ген FVII (1 - радиоактивно меченый маркер молекулярного веса ДНК; 2 - геномная ДНК, обработанная парой рестриктаз HindIII-XhoI; 3 - геномная ДНК, обработанная парой рестриктаз EcoRI-BamHI; 4 - геномная ДНК, обработанная парой рестриктаз XhoI-EcoRI); Б - предположительная рестрикционная карта сайта интеграции гена FVII в геном клеток.
Заявленное изобретение поясняется примерами его исполнения.
Пример 1.
Конструирование рекомбинантной плазмидной ДНК pEFZeoF7.
Нуклеотидную последовательность, соответствующую кДНК гена коагуляционного фактора VII человека (фиг.1 А), получают генно-инженерным методом. Для этого из 5×106 клеток гепатомы человека НЕР2 выделяют мРНК, например, с использованием набора Oligotex Direct mRNA Kit (QIAGEN, Франция) согласно протоколу производителя. Выделенные элюаты мРНК используют для реакции обратной транскрипции с последующей амплификацией (ОТ-ПЦР), которую проводят, например, с помощью набора Superscript™ One-Step RT-PCR (Invitrogen, США) по инструкции производителя с использованием специфических олигонуклеотидных праймеров
F7for 5′-CTATACTCGAGCAGGGGCAGCACTGCAGAG-3′ и
F7rev 5′-CTATAGGATCCGACAGTTCGACGCAGCCTTG-3′.
Для этого готовят реакционную смесь 50 мкл согласно протоколу к набору, содержащую 10 пг - 1 мкг РНК, 0,2 мкМ каждого из праймеров, реакционный буфер, обратную транскриптазу (SuperScript™ II H-Reverse Transcriptase) и термоустойчивую ДНК-полимеразу (Platinum® mix с блокирующим антителом), входящие в состав набора, и проводят реакцию по следующей схеме: синтез первой цепи кДНК (45-55°С, 15-30 мин), денатурация (94°С, 2 мин), 35-40 циклов амплификации (94°С, 15 с; 55°С, 30°С; 72°С, 90°С), завершающая достройка продуктов ПНР (72°С, 5-10 мин).
Синтезированный таким образом фрагмент ДНК содержит последовательность кДНК гена коагуляционного фактора VII человека, фланкированную сайтами узнавания рестриктаз XhoI и BamHI. Продукт амплификации гидролизуют рестриктазами XhoI и BamHI, очищают электрофорезом в 1% агарозном геле, полосу ДНК величиной около 1450 п.о. выделяют из геля методом электроэлюции и осаждают ДНК из раствора этанолом.
Для приготовления вектора ДНК плазмиды pEF/Zeo (фиг.2) (3 мкг, 1 пмоль) обрабатывают в 40 мкл буфера Y (33 мМ трис-ацетат, рН 7,9, 10 мМ Mg-ацетат, 66 мМ К-ацетат 1, 0,5 мМ DTT, 0,1 мг/мл BSA) рестриктазами XhoI и BglII (10 ед.акт.) в течение 1 ч при 37°С. Полученный фрагмент ДНК величиной 3,9 т.п.о. после электрофоретического разделения в 1% агарозном геле выделяют из геля методом электроэлюции и осаждают ДНК из раствора этанолом.
1 мкг полученного векторного фрагмента лигируют с 3,5 мкг XhoI-BamHI фрагмента размером 1424 п.о., содержащего последовательность кДНК гена коагуляционного фактора VII человека, в 10 мкл буфера (20 мМ трис-HCl, рН 7,56, 10 мМ MgCl2, 0,2 мМ rATP, 10 мМ дитиотреита) с помощью 10 ед.акт. Т4-ДНК-лигазы в течение 4 ч при 22°С.
1 мкл полученной лигазной смеси используют для электротрансформации компетентных клеток Е.coli штамма DH5α, которую проводят, например, при помощи аппарата для электротрансформации ВТХ600 при зазоре между пластинами электропорационной кюветы 1 мм и напряжении разряда 1,4 кВ. После трансформации суспензию бактерий смешивают с питательной средой SOC, растят 1 час на +37°С и высевают на чашки Петри с LB-агаром, содержащим 50 мкг/мл ампициллина.
Первичный отбор клонов, содержащих нужную плазмиду, проводят методом "ПЦР с клонов" с использованием специфических праймеров:
F7for 5′-CTATACTCGAGCAGGGGCAGCACTGCAGAG-3′ и
F7rev 5′-CTATAGGATCCGACAGTTCGACGCAGCCTTG-3′,
с последующим анализом продуктов в 1% агарозном геле.
Отобранные положительные клоны используют для подроста в жидкой среде и выделения плазмидной ДНК, которую дополнительно анализируют на наличие вставки размером около 1,9 т.п.о. с помощью эндонуклеаз рестрикции XhoI и BamHI с последующим разделением продуктов гидролиза в 1% агарозном геле (фиг.3). Окончательное строение плазмид, содержащих XhoI-BglII/BamHI фрагмент около 1450 п.о., подтверждают определением нуклеотидной последовательности методом секвенирования по Сенгеру. По данным секвенирования отбирают ту плазмиду, нуклеотидная и соответствующая ей аминокислотная последовательности XhoI-BglII/BamHI фрагмента которой полностью идентичны последовательности кДНК гена коагуляционного фактора VII человека (фиг.1).
Пример 2.
Получение линии клеток BHK/F7, стабильно продуцирующих рекомбинантный коагуляционный фактор VII человека.
Линия ВНК - Baby Hamster Kidney - клетки почек детенышей хомячков [Virology, 1962, 16: 147-151; J.Natl. Cancer Inst., 1963, 30: 995-811].
Для получения линии клеток BHK/F7, стабильно продуцирующих рекомбинантный коагуляционный фактор VII человека (rFVII), клетки линии ВНК21 трансформируют плазмидной ДНК pEFZeoF7, например методом липофекции при помощи реагента Lipofectamine® (Invitrogen, США).
Перед трансфекцией плазмидную ДНК pEFZeoF7 линеаризуют с помощью эндонуклеазы рестрикции PvuI и дополнительно очищают от компонентов реакционной смеси.
Накануне трансфекции высевают 6×104 клеток ВНК21 в каждую лунку 24-луночного планшета в 500 мкл среды DMEM, содержащей 10% бычьей фетальной сыворотки. На следующий день готовят смесь для трансфекции. Для этого, из расчета на одну лунку, смешивают 0,4 мкг плазмидной ДНК pEFZeoF7/PvuI с 25 мкл среды DMEM без сыворотки и добавляют 4 мкл реагента Plus® Reagent (Invitrogen, США), также смешивают 1 мкл реагента Lipofectamine® (Invitrogen, США) с 24 мкл среды DMEM без сыворотки. Затем объединяют обе смеси и оставляют на 15 минут при комнатной температуре для формирования комплексов (плазмидная ДНК в липосомах).
От клеток, достигших 60-80% конфлюентности, убирают среду с сывороткой и добавляют по 200 мкл среды DMEM без сыворотки и по 54 мкл смеси для трансфекции. Инкубируют клетки 3 часа при 37°С в СО2-инкубаторе, после чего добавляют 250 мкл среды, содержащей 20% сыворотки и 10 мкг/мл витамина К.
Для контроля эффективности трансформации отбирают среду через 24, 48 и 72 часа. Наличие rFVII в среде определяют методом твердофазного иммуноферментного анализа (ELISA) или иммуноблотинга с использованием моноклональных или поликлональных антител к коагуляционному фактору VII человека.
Через 72 часа после трансфекции клетки пересевают в разведении 1:10 в среду, содержащую селективный антибиотик зеоцин в концентрации 500 мкг/мл, для отбора клонов стабильно трансфицированных клеток. Смену среды, содержащей антибиотик, проводят регулярно каждые 3-4 дня. Начиная с 7-го дня, лунки анализируют на наличие клонов активно делящихся клеток, устойчивых к зеоцину.
Через 14-25 дней среду из лунок, содержащих клоны трансформированных клеток, анализируют на наличие rFVII методом твердофазного иммуноферментного анализа (ELISA). Лунки с положительными клонами, экспрессирующими rFVII, дополнительно расклонируют из расчета 30, 50 и 100 клеток на 96-луночный планшет. Через 14-25 дней среду из лунок, содержащих индивидуальные клоны трансформированных клеток, анализируют на наличие rFVII методом твердофазного иммуноферментного анализа (ELISA) или иммуноблотинга с использованием моноклональных или поликлональных антител к коагуляционному фактору VII человека. Для создания исходного банка отбирают клетки с уровнем экспрессии целевого белка - 20-40 мкг/мл среды, выращивают во флаконах и замораживают.
Полученный штамм обладает следующими свойствами:
- морфология: фибробластоподобная;
- способ культивирования: монослойный;
- условия культивирования: среда ЕМЕМ ил DMEM с 10% эмбриональной телячьей сыворотки или КРС и 1% NEAA; при пересеве для снятия клеток используется 0,25% трипсин: 0,02% версен (1:3), кратность рассева 1:3-1:6, оптимальная плотность 2,0-4,0×104 клеток/см2; криоконсервация проводится в ростовой среде с 5-10% DMSO при 1,0-1,5×106 клеток/см2; жизнеспособность после консервации - 92%;
- кариология: 2n=44, пределы изменчивости по числу хромосом 44-52, модальное число хромосом 49-50, количество маркеров - 1, крупная метацентрическая хромосома;
- особенности трансформированного штамма: содержание в геноме рекомбинантной плазмидной ДНК pEFZeoF7, устойчивость к ампициллину, устойчивость к зеоцину, продукция белка фактора VII человека.
Пример 3.
Определение наличия интеграции гена FVII в геном клеток ВНК
Чтобы определить наличие последовательности гена FVII, встроенного в геном клеток ВНК21, было проведено выделение суммарной геномной ДНК клеток штамма BHK/F7, трансфицированных рекомбинантной плазмидной ДНК pEFZeoF7, для последующего проведения ПЦР с использованием специфических праймеров (см. пример 1).
Для определения количества копий гена FVII, встроившихся в геном клеток, была проведена гибридизация по Саузерну рестрицированной геномной ДНК с радиоактивно меченой последовательностью, кодирующей ген FVII. Для анализа использовались уникальные сайты рестрикции, фланкирующие данную последовательность. Полученные результаты свидетельствуют о наличии единичной копии гена FVII в геноме данного клона (фиг.4).
Таким образом, с помощью заявленной плазмидной ДНК и линии клеток-продуцента возможно получать рекомбинантный белок фактора VII с высоким выходом, который в дальнейшем можно импользовать для получения лекарственных препаратов.
Пример 4. Конструирование вектора pEFZeo.
Описание вектора.
(Карта вектора представлена на фиг.2)
Вектор pEFZeo представляет собой рекомбинантную плазмидную ДНК, состоящую из 4884 п.о., со следующими признаками:
включает:
последовательность начала инициации репликации плазмидной ДНК (pUC origin), обеспечивающую наработку плазмидной ДНК в прокариотической клетке;
ген "Amp(R)" под контролем прокариотического промотора (bla promoter), обеспечивающий устойчивость трансформированных плазмидой pEFZeo клеток Е.coli к ампициллину и позволяющий селективный отбор и наращивание различных генно-инженерных конструкций на основе данного вектора в прокариотической системе;
ген "zeocin resistance" под контролем эукариотического раннего SV40 промотора и SV40 сайта полиаденилирования, обеспечивающий устойчивость к зеоцину трансфицированных плазмидой pEFZeo (или плазмидной конструкцией на основе pEFZeo) эукариотических клеток и позволяющий получение стабильной линии эукариотических клеток, содержащих в своем геноме встроенную плазмидную последовательность ДНК;
последовательность, содержащая уникальные участки узнавания рестриктаз NotI (2381 п.о.), XhoI (2411 п.о.) и BglII (2417 п.о.), предназначенная для встраивания гена, кодирующего белковый продукт;
эукариотический промотор гена 1α субъединицы фактора элонгации человека (EF promoter) и последовательность полиаденилирования гена гормона роста быка (BGH polyA), обеспечивающие высокоэффективную экспрессию белкового продукта в эукариотческих клетках;
уникальные участки узнавания следующих рестриктаз - NotI (2381 п.о.), XhoI (2417 п.о.), BglII (2417 п.о.), NheI (1194 п.о.), BamHI (2884 п.о.), PvuI (3453 п.о.).
1) ген "zeocin resistance" под контролем эукариотического раннего SV40 промотора и SV40 сайта полиаденилирования, обеспечивающий устойчивость к зеоцину, был получен следующим образом.
Синтетическая ДНК последовательность, соответствующая Sh ble гену (Streptoalloteichus hindustanus bleomycin gene; Calmels et al., 1991, Drocourt et al., 1990), была получена с помощью ПЦР из перекрывающихся олигонуклеотидных фрагментов; конечный ПЦР-продукт, величиной 437 п.о., фланкированный AvrII и Bsp120I сайтами узнавания эндонуклеаз рестрикции, обрабатывался соответствующими рестриктазами и лигировался в вектор pSI (Promega), рестрицированный AvrII и NotI, соответственно. Корректность сборки подтверждалась секвенированием по Сенгеру. Результирующая конструкция pSIZeo содержала ген ble под контролем эукариотического раннего SV40 промотора и SV40 сайта полиаденилирования.
2) Фрагмент ДНК, соответствующий эукариотическому промотору гена 1α субъединицы фактора элонгации человека (EF promoter), был наработан методом ПЦР с геномной ДНК человека с использованием специфических олигонуклеотидных праймеров:
prEFforwNheI 5' GAGGAGCTAGCTTCGTGAGGCTCCGGTG 3'
prEFrevNotI 5' GAGAAGCGGCCGCGACCACGTGTTCACGACA 3'
ПЦР-продукт, величиной 1204 п.о., лигировался в вектор pUC19, рестрицированный SmaI, с образованием конструкции pUCEF. Синтетическая ДНК последовательность, соответствующая полилинкеру, была получена из двух перекрывающихся олигонуклеотидных праймеров;
PolyLinkfbrwNotI
5' GGCCGCTTCGAAGGTACCAGCACAGTGGACTCGAGAGATCTGGCCGGCTG 3'
PolyLinkrevBsp120I
5' GGCCCAGCCGGCCAGATCTCTCGAGTCCACTGTGCTGGTACCTTCGAAGC 3'
конечный продукт, величиной 54 п.о., фланкированный NotI и Bsp120I сайтами узнавания эндонуклеаз рестрикции, лигировался в вектор pUCEF, рестрицированный NotI, соответственно. Скрининг проводили методом ПНР с клонов с использованием стандартных векторных праймеров M13fbrw и М13rev, с последующей рестрикцией ПЦР-продуктов XhoI и электрофоретическим анализом в 1.2% агарозном геле. Отобранные конструкции pUCEFNB pUCEFBN содержали полилинкерную последовательность в различной ориентации, фланкированную NotI и BglII сайтами узнавания эндонуклеаз рестрикции, соответственно. Корректность сборки подтверждалась секвенированием по Сенгеру.
3) Фрагмент ДНК, соответствующий последовательности полиаденилирования гена гормона роста быка BGH polyA (Woychik, R.P. et al., Nucleic Acids Res. 10 (22), 7197-7210 (1982)) был наработан методом ПЦР с геномной ДНК быка с использованием специфических олигонуклеотидных праймеров:
BGHforwBglII
5' GGAGGAGATCTGTGCCTTCTAGTTGCCAG 3'
BGHrevSphI
5' GAAGAGCATGCCTGCTATTGTCTTCCC 3'
конечный ПЦР-продукт, величиной 216 п.о., фланкированный BglII и SphI сайтами узнавания эндонуклеаз рестрикции, обрабатывался соответствующими рестриктазами и лигировался в вектор pUCEFNB, рестрицированный BamHI и SphI, соответственно. Корректность сборки результирующей конструкции pUCEFpolyA подтверждалась секвенированием по Сенгеру.
4) NheI-SphI Фрагмент ДНК конструкции pUCEFpolyA, величиной 1848 п.о., содержащий эукариотический EF промотор, полилинкерную последовательность и последовательность полиаденилирования BGH polyA, переклонировали в вектор pGEM-1m (вектор pGEM-1 с делегированным SmaI-PvuII фрагментом), рестрицированный NheI и SphI, соответственно. Acc65I-BamHI Фрагмент ДНК конструкции pSIZeo, величиной 1043 п.о., содержащий ген ble под контролем эукариотического раннего SV40 промотора и SV40 сайта полиаденилирования, переклонировали в полученную на предыдущем этапе конструкцию pGEMEF, рестрицированную Bsp1407I и BamHI, соответственно. Результирующая конструкция - вектор pEFZeo - содержала все необходимые элементы и была использована для экспрессии различных рекомбинантных продуктов в эукариотических клетках.

Изобретение относится к области биотехнологии и касается получения белка фактора VII методом рекомбинантных ДНК. Сконструирована рекомбинантная плазмидная ДНК для экспрессии фактора свертывания крови VII в клетках млекопитающих, представляющая собой продукт лигирования фрагмента кДНК гена фактора VII человека, фланкированного сайтами узнавания рестриктазами XhoI и BamHI, с большим XhoI/BglII фрагментом вектора pEFZeo, включающим гены устойчивости к ампициллину и зеоцину. В результате трансформации клеток ВНК новой рекомбинантной плазмидой получена линия клеток BHK/F7, продуцирующая рекомбинантный белок фактора VII с выходом до 40 мкг/мл. 2 н.п. ф-лы, 4 ил.
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| KOVARIK P | |||
| ET AL | |||
| EMBO J., 2001, 20(102), 91-100 | |||
| RU 2004113441 A, 20.05.2005. | |||

