Данное изобретение относится к изоформам GnRH (рилизинг-фактора гонадотропина).
GnRH-I (в литературе обычно обозначаемый как GnRH) является небольшим, имеющим длину 10 аминокислот пептидом (декапептидом) из гипоталамуса. Аминокислотная последовательность GnRH-I (SEQ ID NO:1) может быть представлена следующим трехбуквенным кодом:
pGlu-His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-Gly-NH2
или соответствующим однобуквенным кодом, где рЕ является пироглутаминовой кислотой, а # обозначает амид:
pE H W S Y G L R P G#.
GnRH-I действует при гипофизе, вызывая увеличение высвобождения биологически активного фолликулостимулирующего гормона (FSH) и лютеинизирующего гормона (LH) в крови, которые, в свою очередь, стимулируют развитие яичек в растущем животном мужского пола и синтез мужских стероидных гормонов. В растущем животном женского пола стимулируется развитие яичников, а также развитие фолликулов в яичнике, синтез женских стероидных гормонов и овуляция.
Известно, что GnRH-I, если он связан с белком-носителем, может быть использован для вакцинации животных. Такая вакцинация может проводиться по различным причинам, все из которых связаны с природной функцией GnRH-I. Как известно, резкое снижение LH и/или FSH в крови ингибирует продуцирование мужских стероидных гормонов или андрогенов и спермы в яичке мужской особи и образование женских стероидных гормонов или прогестагенов и эстрогенов и созревание фолликулов в яичнике женской особи. Такое уменьшение количества андрогенов, прогестагенов и эстрогенов в крови до уровня, сравнимого с уровнем, получаемым удалением яичек или яичников посредством кастрации, может быть достигнуто эффективной иммунизацией животного против GnRH-I. В животных-самцах во многих случаях яички затем развиваются, по-видимому, медленно или вообще не развиваются, не синтезируют андрогены (мужские стероидные гормоны) и не образуют сперматозоиды. В животных-самках активность яичников, по-видимому, уменьшается, не синтезируются эстрогены и прогестагены (женские стероидные гормоны) и ингибируется созревание фолликулов и овуляция.
Недавно сообщалось, что в головном мозге приматов присутствует вторая форма GnRH (GnRH-II) (Lescheid et al. Endocrinol. 138 (1997) 5618-5629), и ген для этой второй молекулы GnRH-II был клонирован из геномной библиотеки человека (GnRH-II, (SEQ ID NO:2) (White et al. PNAS USA 95 (1998) 305-309). GnRH-I (SEQ ID NO:1) млекопитающих почти не экспрессируется вне головного мозга. В этом отношении известны несколько исключений. GnRH-I присутствует в эндометрии женщин с менструальным циклом (Casan et al. Fertil. Steril. 1998, (70, 102-106) и экспрессируется во время беременности в плаценте человека (Kelly et al. DNA Cell Biol. 1991, 10, 411-421). мРНК GnRH была обнаружена в яичнике, яичке, тимусе, плаценте и гипоталамусе крысы (Oikawa et al., Endocrinology, 1990, 127, 2350-2356). Экспрессия GnRH была обнаружена в иммунной ткани (селезенке, тимусе и лимфоцитах) свиней (Weesner et al., Life Sci, 1997, 61, 1643-1649).
GnRH-II экспрессируется во многих тканях вне головного мозга и обнаруживается в особенно высоких концентрациях в почках, костном мозгу и предстательной железе. Присутствие GnRH-II в разнообразных тканях, иных, чем головной мозг, предполагает, что GnRH-II может иметь множественные функции. Кроме того, строго консервативная структура пептида GnRH-II во всех различных видах позвоночных животных предполагает, что этот нейропептид обладает жизненно важными биоактивностями. Однако до сих пор функции GnRH-II практически были неизвестными. Несколько типов дифференцированных лимфоцитов, таких как Т- и В-лимфоциты, и тучные клетки, продуцируют GnRH и GnRH-подобные пептиды. Значительные количества последнего типа клеток присутствуют в почке, костном мозгу и предстательной железе, возможно, способствуя высокой экспрессии GnRH-II в этих тканях. GnRH-II, по-видимому, в меньшей степени участвует в репродукции, чем GnRH-I. В случае гипогонадной мыши, мыши, которая не имеет гена GnRH-I, продуцирующие GnRH-II клетки присутствуют в том же самом распределении, что и у нормальной мыши, но это не является достаточным для индукции нормального гонадного развития у этой мыши (Chen et al. FEBS Letters 435 (1998) 199-203). Однако у макаков в лютеиновой фазе менструального цикла обнаружили заметное увеличение в плазме концентраций лютеинизирующего гормона после внутривенного введения GnRH-II, но это увеличение не могло быть индуцировано во время средней фолликулярной фазы (Lescheid et al. Endocrinol. 138 (1997) 5618-5629).
Данное изобретение относится к пептиду, пригодному для индукции иммунной реакции против форм GnRH, также называемого рилизинг-фактором лютеинизирующего гормона (LHRH). Данное изобретение относится также к иммуногенным композициям и вакцинам, фармацевтическим средствам и другим медицинским препаратам на основе такого пептида. Далее, данное изобретение относится к применению такой вакцины или медицинского препарата в способе иммунизации млекопитающего против GnRH для воздействия на репродуктивные и бихевиористические (поведенческие) характеристики этого млекопитающего и в способе улучшения качества туши свиней. Данное изобретение относится также к пептиду, пригодному для индуцирования селективной иммуногенной реакции против GnRH-I или GnRH-II. Кроме того, данное изобретение относится к антителу против GnRH-I и/или GnRH-II, композициям, содержащим эти антитела, и применению этих пептидов в фармацевтических композициях или в приготовлении лекарственного средства для лечения рака предстательной железы.
Далее, данное изобретение относится к пониманию того, что посредством обеспечения пептидных последовательностей, которые позволяют дифференцировать (различать) различные типы GnRH, может достигаться более адекватное и эффективное применение вариации или различия в иммунологической реакции на различные типы GnRH. Более конкретно, данное изобретение относится к пониманию того, что могут быть достигнуты улучшения в эффективности и селективности вакцин против GnRH-I. Иммунизация против GnRH-I является эффективной в нейтрализации GnRH-I и приводит к уменьшенным уровням гонадотропина и блокированию синтеза половых стероидных гормонов. Однако ничего неизвестно о каких-либо физиологических эффектах антител, индуцированных против GnRH-I, на функцию GnRH-II. Поскольку GnRH-II в основном синтезируется и секретируется в почках, антитела, индуцированные против GnRH-I, которые перекрестно реагируют с GnRH-II, могут влиять на функцию почек. Для устранения возможных побочных эффектов иммунизации GnRH-I на функцию почек было бы желательным нацеливание антигенной реакции иммунокастрационной вакцины специфически на GnRH-I и исключение возможных вредных побочных действий, обусловленных нейтрализацией не-гонадного GnRH-II.
Если требуется устранить только репродуктивную способность вида, часто с сопутствующим ей половым поведением, было бы предпочтительным стремление к получению иммунокастрационной вакцины, специфически нейтрализующей GnRH-I. Таким образом, желательно достичь селективной иммунизации против гонадотропин-высвобождающего гормона (гормонов), предпочтительно селективной против GnRH-I.
В ветеринарной медицине 100%-ная эффективная иммунизация против GnRH-I могла бы использоваться для стерилизации, например, небольших домашних животных, таких как самцы и самки кошек и собак, или для лечения агрессивности у самцов собак и быков обычной вакцинацией вместо радикальной хирургии, такой как кастрация или овариэктомия. Другими возможными причинами для иммунизации против GnRH-I являются предотвращение периода течки у самок животных, таких как собаки, кошки и коровы, и беспокойного состояния у самцов животных, откармливаемых на убой.
В здравоохранении человека иммунизация против GnRH-I и/или GnRH-II может быть использована в лечении рака предстательной железы и рака молочной железы и, если требуется, в лечении некоторых форм злокачественной опухоли гипофиза. В случае рака предстательной железы может быть желательной нейтрализация как GnRH-I, так и GnRH-II, так как последняя изоформа является высоко экспрессируемой в ткани предстательной железы.
Другое применение вакцины против GnRH-I находится в области животноводства, в частности откорма свиней на убой. Мясо самцов половозрелых свиней (хряков) имеет типичный запах, так называемый запах или привкус хряка. В яичках половозрелой свиньи образуются многие С19-дельта-16-стероиды, которые запасаются в жировой ткани этого животного (Patterson, J. Sci Food Agric. 19, 31-38 (1968); Brooks en Pearson, J. Anim. Sci. 62, 632-645 (1986); Claus, Zeitschrift. Tierzüchtg. Züchtungsbiol 93, 38-47 (1976); Claus, Acta Endocrinol. (Copenh.) 91, Suppl. 225, 432-433 (1979)). Эти стероиды ответственны в основном за образование неприятного подобного запаху мочи запаха при нагревании мяса (Fuchs, Swedish J. Agric. Res. 1 233-237 (1971); Bonneau, Livest. Prod. Sci 9, 687-705 (1982)). Из-за этого неприятного запаха мясо самцов половозрелых свиней обычно является непригодным для употребления и негодным для экспорта. Поскольку приблизительно 10% самцов убойных свиней являются уже половозрелыми перед временем убоя, это потенциально влечет за собой большую потерю для свиноводства на промышленной основе.
Для контроля и предупреждения этих потерь почти всех самцов-поросят кастрируют, когда они очень маленькие, причем используют хирургическую процедуру, которая обычно выполняется без какой-либо формы анестезии. Не говоря уже о неблагоприятном аспекте отношения к животным в случае такой кастрации, эта кастрация приводит также к инфекциям, ингибированию роста и конечному качеству туши, худшему, чем качество туши интактного животного, по меньшей мере до тех пор, пока не подвергнутое такой кастрации животное на развило еще привкуса хряка (Walstra, Livest. Prod. Sci.1, 187-96 (1974)).
Благоприятная для животного альтернатива состоит в снижении концентрации GnRH-I в гипофизе свиньи посредством иммунизации против GnRH-I, так называемой иммунокастрации. Это снижение уровней GnRH-I приводит к уменьшению концентраций биологически активных FSH и LH, что, в свою очередь, будет задерживать развитие яичек в растущих животных и ингибировать синтез тестикулярных стероидных гормонов, в том числе андростенона, тестостерона и эстрогенов. Этот способ предотвращает привкус хряка у самцов свиней во время убоя и делает ненужной хирургическую кастрацию, так как уровни андростенона уменьшаются до низких или недетектируемых уровней (Oonk et al., 1995, Livestock production Science 42, 63-71).
Строгое требование для приемлемой вакцины против привкуса хряка заключается в том, что почти во всех свиньях развитие яичек замедляется до такой степени, что привкус хряка не наблюдается во время убоя, и что в случае, если вакцина не уменьшает развитие яичка в животном, это может быть легко детектировано по слишком большому размеру яичка в сравнении с успешно иммунокастрированными свиньями.
В существующей литературе и предыдущих патентных заявках, относящихся к свойствам антифертильности вакцин против GnRH-I, результаты вакцинации, по-видимому, часто являются вариабельными. В большинстве описанных исследований либо небольшой процент вакцинированных животных не отвечает на вакцинацию, либо большие дозы вакцины, многочисленные вакцинации или коммерчески неприемлемые адъюванты требуются для получения желательного эффекта (Hoskinson et al., 1990, Austr. J. Biotech. 4, 166-170; Falvo et al., (1986) J. Anim. Sci. 63:986-994; Clarke et al, 1998, Endocrinology 139, 2007-2014; Adams T.E and B.M Adams, Feedlot performance of steers and bulls active immunized against Gonadotropine-Releasing Hormone, J. Anim. Sci. 1992, 70:1691-1698; Brown et al. Immunization of sheep against GnRH-I early in life: effects of reproductive function and hormones in rams, Journal of reproduction and Fertility (1994) 101, 15-21; Ferro et al., Immunological castration using a Gonadotropine-releasing Hormone analogue conjugated to PPD, Food and agricultural immunology, 1995, 7, 259-272; U.S. patent 4,608,251; Int. patent appl. WO 88/05308).
Некоторые исследования предполагали эффективность 100% вакцины против GnRH-I, но эта вакцина не испытывалась на большом числе животных (Ladd et al (1994), Development of an antifertility vaccine for pets based on active immunization against Luteinizing Hormone releasing hormone. Biology of Reproduction 51, 1076-1083; J.G. Manns and S.R. Robbins (1997). Prevention of boar taint with a recombinant based GnRH vaccine, In: Boar taint in entire male pigs. Proceedings of a meeting of the EAAP working group "Production and Utilisation of Meat from Entire Male Pigs", EAAP Publication No. 92, 137-140); другие исследования сообщают эффективность вакцины в виде средней величины для обработанных животных, так как индивидуальные величины сильно не различаются между иммунизированными животными и необработанными контролями (Bonneau et al., J. Anim. Sci. 72, 14-20 (1994); Hennesy et al., 1997. Elimination of boar taint: a commercial boar taint vaccine for male pigs. In: Bonneau, M., Lundström, K. and Malmfors, B. (Eds.), Boar taint in entire male pigs. Wageningen Pers, Wageningen, EAAP Publication No. 92, 141-145).
Трудность в приготовлении этого типа вакцин, возможно, обусловлена явлением толерантности. "Свои" вещества, такие как гормоны, не узнаются как чужеродные, а переносятся иммунной системой животного. Обычно против "своих" веществ не индуцируются антитела. Для того, чтобы вакцина была успешной, она должна быть достаточно чужеродной. Только в том случае, когда вакцина является достаточно чужеродной, иммунная система не будет переносить эту вакцину и будет индуцироваться образование антител. Однако, напротив, антитела должны быть все еще способными узнавать гормон, и, следовательно, эта вакцина не может быть слишком "чужеродной".
Поскольку эти условия, по-видимому, являются взаимоисключающими, до недавнего времени не было уверенности в том, могут ли вообще быть получены такие вещества. Одна из попыток получения содержащих GnRH-подобный пептид вакцин состояла в замене Gly в положении 6 декапептида GnRH-I правовращающей аминокислотой (D-Trp; Chaffaux et al., Recueil de Medicine Veterinaire 161 (2), 133-145, 1985). Однако было продемонстрировано, что вакцинный препарат, содержащий этот модифицированный GnRH-пептид, функционировал даже хуже, чем нормальный декапептид GnRH-I (Европейская патентная заявка 464124 А).
Недавно авторы изобретения определенно показали, что можно индуцировать эффективную реакцию в виде образования антител во всех индивидуумах, вакцинированных против GnRH-I (Meloen et al., Vaccine 12, 741-746 (1994)). В этих экспериментах свиней вакцинировали дважды GnRH-I-вакциной, которая отклоняется от классического типа GnRH-I-вакцины (GnRH-I, связанного с белком-носителем, в адъюванте Фройнда), а именно содержащей тандемный GnRH-I вакциной (Европейский патент с номером 0464124). В этой публикации описан пептид, который отличается тем, что он содержит по меньшей мере 2 последовательности GnRH-I в тандеме (SEQ ID NO:3) в соответствии с общей формулой
Z1-Glx-His-Trp1-Ser-Tyr-Gly-Leu-Arg-Pro[-Gly-X-Gln-His-Trp2-Ser-Tyr-Gly-Leu-Arg-Pro]n-Gly-Z2,
в которой аминокислоты обозначены в соответствии с трехбуквенным кодом, Trp1 и Trp2 являются триптофаном (Trp) или формилированным триптофаном (N(индол)формилтриптофан), n равно числу, имеющему величину по меньшей мере 1, Х является либо прямой связью, либо спейсерной группой между Gly и Gln, Z1-Glx обозначает либо pGlu (пироглутаминовую кислоту), либо Gln, имеющий присоединенный к нему хвост, содержащий одну или несколько дополнительных аминокислот, и Gly-Z2 обозначает либо Gly-NH2, либо Gly, имеющий присоединенный к нему хвост, содержащий одну или несколько дополнительных аминокислот. В этой общей формуле Х может быть прямой связью между аминокислотами глицином и глутамином, т.е. эти аминокислоты связаны друг с другом непосредственно без промежуточного звена (посредством обычной пептидной связи). Изобретение содержащей тандемный GnRH-I вакцины включает в себя также пептиды, в которых последовательности GnRH-I связаны между собой через спейсеры. Природа спейсерной группы может сильно варьироваться, от одной или более аминокислот до более короткой или более длинной цепи и групп или молекул других соединений. В приведенной выше общей формуле Z1-Glx предпочтительно представляет pGlu (пироглутаминовую кислоту), но может представлять также Gln, имеющий присоединенный к нему хвост, содержащий одну или несколько дополнительных аминокислот, например, для использования для связывания этого пептида с белком-носителем. В приведенной выше общей формуле Gly-Z2 обозначает, например, Gly-NH2, или Gly, имеющий присоединенный к нему хвост, содержащий одну или несколько дополнительных аминокислот, например, для использования для связывания этого пептида с белком-носителем. Предпочтительно, Gly-Z2 представляет Gly-Cys-NH2, причем С-концевой цистеин добавляется в связи с возможным связыванием этого пептида с белком-носителем.
Из WO 96/40755 известно, что тандемный-димерный принцип, примененный к варианту молекулы GnRH-I, приводил к вакцине, которая была высокоэффективной в нескольких мягких адъювантах, а именно Specol и двойной масляной эмульсии, и была также эффективной в низких дозах. В этом случае вариант молекулы GnRH-I был образован заменой шестой аминокислоты Gly этого декапептида правовращающей (D-) аминокислотой, D-Lys, после чего полученный пептид димеризовали и связывали с обычным соединением-носителем, овальбумином. Таким образом, в то время как вакцина, использующая D-аминокислотные замены Gly в положении 6 исходного пептида, и декапептид с единственной молекулой GnRH-I с D-аминокислотой уменьшали иммуногенность в сравнении с исходной последовательностью GnRH-I (Chaffaux et al., Recueil de Medicine Veterinaire 161 (2), 133-145, 1985), такие замены D-аминокислотой, примененные к тандемной-димерной вакцине GnRH-I, были способны генерировать даже более иммуногенные GnRH-I-вакцинные препараты. Тем не менее этот способ вакцинации требовал повторной дозы этой вакцины для полной эффективности. Необходимость дополнительной бустер-дозы для достижения по существу 100% эффективной вакцинации млекопитающих против GnRH-I является недостатком этих известных пептидов. Авторы изобретения нашли также, что в определенных случаях применение этого тандемного димера (D-Lys6) GnRH-I (т.е. тандемного димера GnRH-I с заменой D-Lys6 или без замены) приводило к очень низким, но все еще измеряемым количествам тестостерона, что является нежелательным и является недостатком тандемного димера (D-Lys6) GnRH-I.
Данное изобретение относится к пептидным последовательностям, которые обеспечивают альтернативы тандему (D-Lys6) GnRH-I при использовании в вакцинах, обеспечивающих создание вакцин, которые являются эффективными для иммунокастрации.
Одним аспектом данного изобретения является определение степени, до которой могут варьироваться аминокислоты в тандемной GnRH-I-последовательности, в то время как полученный замещенный тандемный GnRH-I является все еще способным производить иммуногенную реакцию на GnRH-I, достаточную для иммунокастрации. Таким образом, данное изобретение относится к генерированию пептидной последовательности, которая может индуцировать продуцирование антител против GnRH-I, которые являются также достаточно конкурентными как по количеству, так и по активности. Данное изобретение не ограничивается только тандемными пептидами per se, должно быть понятно, что сходные дифференцирующие реакции могут быть индуцированы вариациями с единственным декапептидом или даже с последовательностями GnRH, которые были укорочены или из которых были делетированы конкретные аминокислоты или которые были любым другим путем модифицированы или дериватизованы, например, добавлением неприродных аминокислот, или D-аминокислот, в частности, такими модификациями, которые приводят к уменьшенному сходству с GnRH-I или GnRH-II, но увеличивают иммуногенность GnRH.
Следующий аспект данного изобретения относится к пептидной последовательности, которая селективно индуцирует продуцирование антител против GnRH-I, индуцируя небольшую иммунную реакцию или вызывая отсутствие иммунной реакции против GnRH-II. Предпочтительным вариантом является пептидная последовательность, которая не только селективно индуцирует продуцирование антител против GnRH-I, но является также эффективной в иммунокастрации, тогда как иммунная реакция против GnRH-II уменьшается или отсутствует.
Авторы данного изобретения обнаружили, что различные аминокислоты тандемной GnRH-I-пептидной последовательности могут быть заменены, что приводит к уменьшению сходства с собственным гормоном с сохранением в то же самое время или даже увеличением способности этого пептида индуцировать GnRH-I-связывающие антитела. Определенные замены аминокислот в GnRH-I-пептидной последовательности приводят также к селективной иммунной реакции против GnRH-I и к пониженной или отсутствующей иммунной реакции против GnRH-II.
В аспекте данного изобретения определенные модифицированные тандемные GnRH-I-пептидные последовательности обеспечивают вакцины, которые не только способны уменьшать рост яичек у самцов животных, но также способны существенно уменьшать уровни тестостерона до такой степени, что они не могут быть определены общепринятыми способами.
Далее, вакцины, приготовленные из этих пептидных последовательностей, проявляют активность, которая в большинстве случаев исключает необходимость второй бустер-иммунизации (ревакцинации), требующейся в случае общепринятого тандема (D-Lys6) GnRH-I, для достижения по существу 100%-ной активности. Активность или эффективность 100% в контексте данного изобретения определяется как уровень тестостерона, который является по существу недетектируемым общепринятыми способами после единственной вакцинации.
Одним из наиболее примечательных признаков данного изобретения является то, что антитела, индуцированные этими альтернативными GnRH-вакцинами, дифференцируют GnRH-I и GnRH-II. Таким образом, пептиды данного изобретения проявляют увеличенную или сохраняемую активность против GnRH-I при одновременной пониженной или отсутствующей иммунной реакции против GnRH-II. Это делает возможным развитие получения пептидов, которые проявляют противоположное действие, т.е. они проявляют увеличенную или сохраняемую активность против GnRH-II при одновременном проявлении уменьшенной или отсутствующей иммунной реакции против GnRH-I. Такая GnRH-II-специфическая реакция может быть использована для содействия супрессии имплантации эмбриона в млекопитающих, когда беременность является нежелательной или только мало желательной. Кроме того, с использованием такой GnRH-II-специфической реакции можно понижать уровни FSH, что приводит к пониженной общей фертильности (способности к оплодотворению).
Данное изобретение относится в одном аспекте к пептиду, который содержит модифицированную последовательность тандемного декапептида GnRH-I, в результате чего вакцинация этим пептидом в подходящей дозе позволяет получить уровень тестостерона, который является по существу недетектируемым.
Данное изобретение относится также к пептиду, который содержит по меньшей мере две связанных последовательности декапептида GnRH-I, необязательно связанных через спейсер, который делает возможной иммуногенную реакцию, позволяющую эффективно дифференцировать GnRH-I и GnRH-II.
Пептиды данного изобретения являются достаточно сходными с данным гормоном, но в то же самое время более «чужеродными» для иммунной системы и имеют увеличенную способность индуцировать продуцирование антител, направленных против данного гормона.
Признаком данного изобретения является то, что индивидуальные тандемные звенья могут быть димеризованы для дополнительного усиления их иммуногенности без потери возможности связывания данного пептида или пептидной композиции с белковым соединением-носителем.
Могут быть использованы способы димеризации и связывания этого тандема с носителем, сходные со способами, описанными в WO 96/40755. Также предполагается, что пептиды, содержащие только часть пептидных последовательностей GnRH-I или GnRH-II, могут быть использованы в данном изобретении. Примерами их являются нонапептиды и ундекапептиды.
Линкеры для применения в пептидах в соответствии с данным изобретением могут быть выбраны из линкеров, описанных в другом месте в данной заявке, или таких линкеров, как SMCC-линкеры или другие линкеры, известные в данной области.
Линкеры применяют для связывания двух или более димеризованных пептидных последовательностей. Аминокислотой, которую используют для замены аминокислоты в тандемных пептидных последовательностях, является предпочтительно аминокислота, которая является относительно простой, такая как аланин. Таким образом, в предпочтительном варианте этой отличающейся аминокислотой является аланин. Другие аминокислоты могут быть также использованы для замены аминокислоты в тандемной декапептидной последовательности. Предпочтительно проводят только консервативные замены. Консервативными заменами являются замены аминокислот, в которых «объемные» аминокислоты заменяют «объемными» аминокислотами, ароматические аминокислоты заменяют ароматическими аминокислотами, и т.д. Эти концепции хорошо известны специалистам с квалификацией в данной области.
Спейсеры могут быть помещены между пептидами в соответствии с данным изобретением. Это позволяет получать мультимеры. Подходящие спейсеры известны в данной области.
Описание чертежей
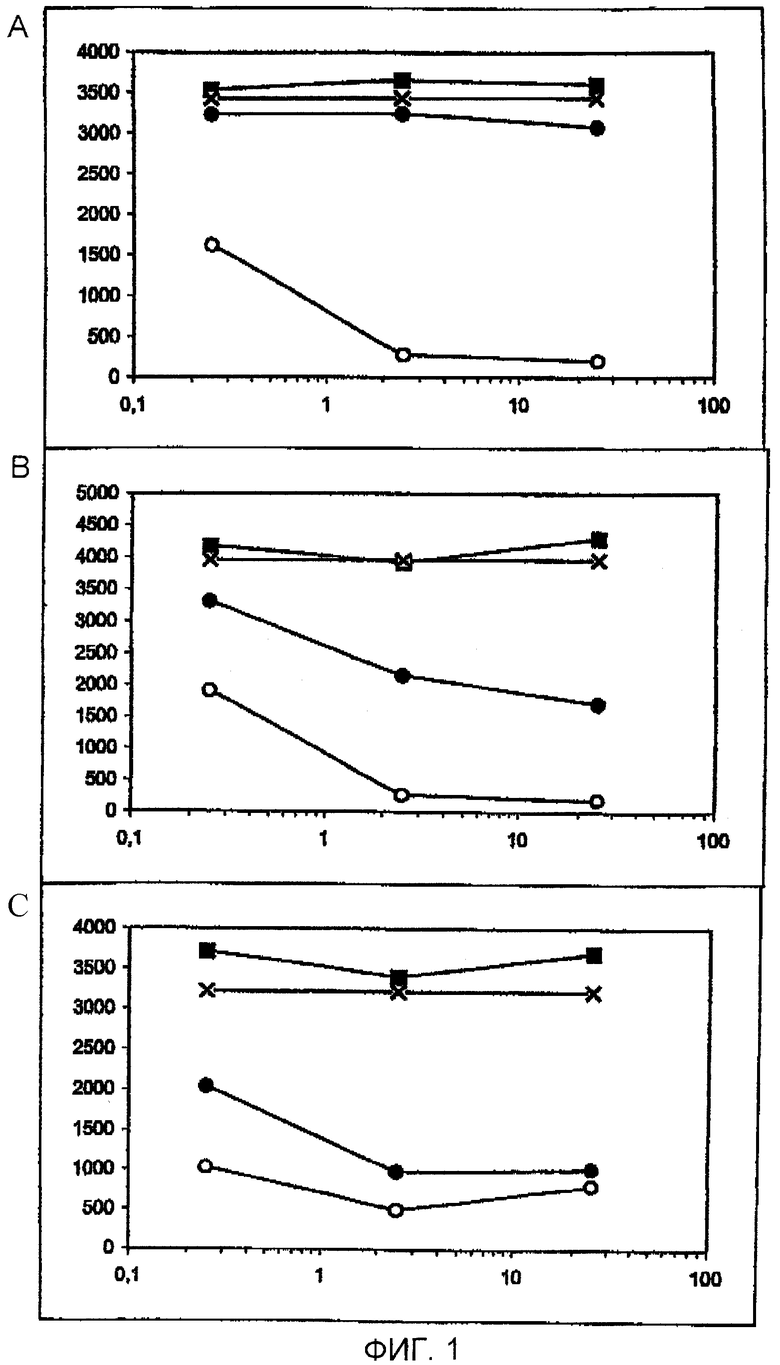
Фигура 1: Конкуренция за связывание йодированного GnRH с антисывороткой с использованием GnRH (светлые кружки), GnRH-II (темные кружки), контрольного пептида (темные квадраты) и без пептида (звездочка). Сыворотку разводили 1/10000 и добавляли увеличивающиеся концентрации пептидов (0,25, 2,5, 25 пмоль) на лунку. В графиках А-С показаны сыворотки с увеличивающейся связывающей способностью в отношении GnRH-II. Горизонтальная ось: количество пептида (пмоль на лунку); вертикальная ось: связывающая способность (имп/мин).
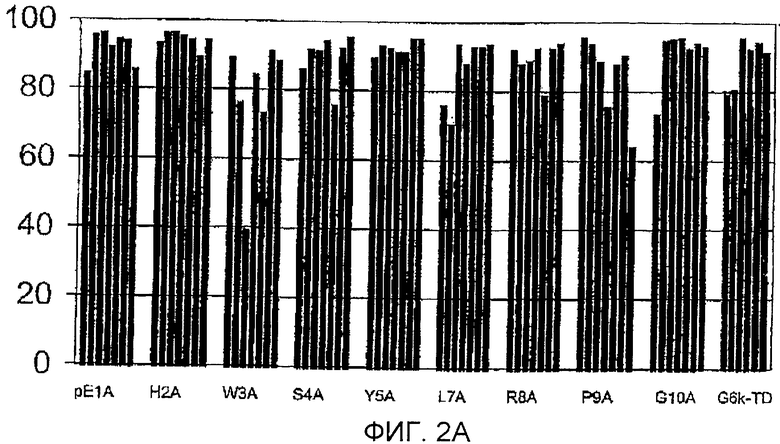
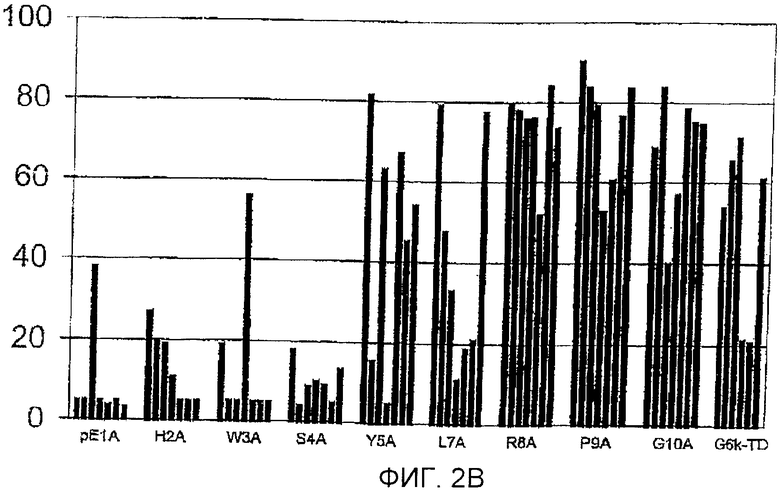
Фигура 2: Процент йодированного GnRH-I, вытесняемого GnRH-I (фигура 2А) и GnRH-II (фигура 2В) для сывороток индивидуальных животных (каждый столбец обозначает одно животное) после иммунизации тандемными димерными пептидами G6k-GnRH с заменами аланином, указанными под каждым кластером столбцов. Антисыворотку, полученную спустя 3 недели после бустер-иммунизации (10 wpv (недели после вакцинации)), разводили 1:100-1:10000. GnRH-I и GnRH-II для вытеснения добавляли в концентрации 250 пмоль на мл.
Фигура 2А
Горизонтальная ось: пептиды, используемые для иммунизации.
Вертикальная ось: процент вытеснения йодированного GnRH-I нейодированным GnRH-I.
Фигура 2В
Горизонтальная ось: пептиды, используемые для иммунизации.
Вертикальная ось: процент вытеснения йодированного GnRH-I нейодированным GnRH-II
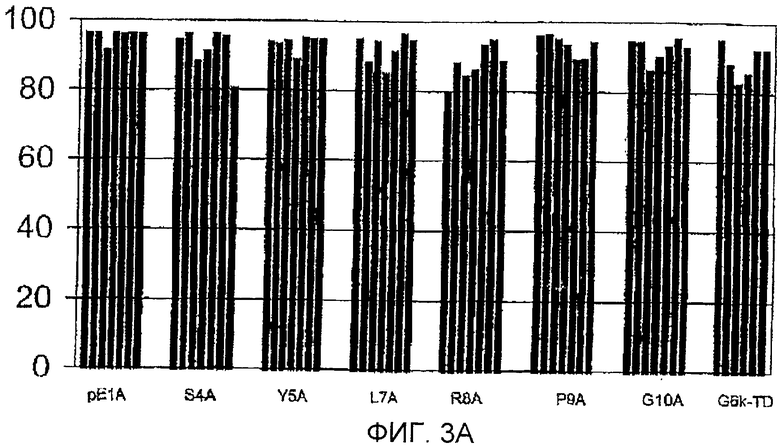
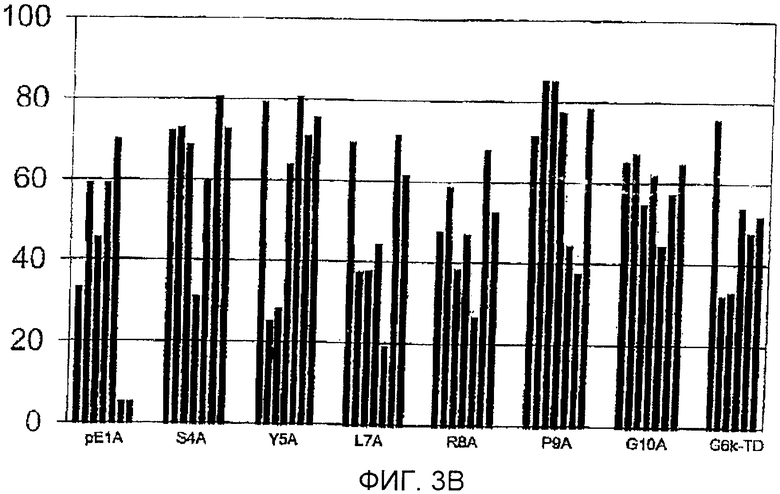
Фигура 3: Процент йодированного GnRH-I, вытесняемого GnRH-I (фигура 3А) и GnRH-II (фигура 3В) для сывороток индивидуальных животных (каждый столбец обозначает одно животное) после иммунизации тандемными димерными пептидами G6k-GnRH с заменами аланином, указанными под каждым кластером столбцов. Антисыворотку получали во время бустер-иммунизации (7 wpv), разводили 1:100-1:10000. GnRH-I и GnRH-II для вытеснения добавляли в концентрации 250 пмоль на мл. Данные сывороток Н2А и W3A не включены, так как титры антител были слишком низкими для измерения вытеснения.
Фигура 3А
Горизонтальная ось: пептиды, используемые для иммунизации.
Вертикальная ось: процент вытеснения йодированного GnRH-I нейодированным GnRH-I.
Фигура 3В
Горизонтальная ось: пептиды, используемые для иммунизации.
Вертикальная ось: процент вытеснения йодированного GnRH-I нейодированным GnRH-II.
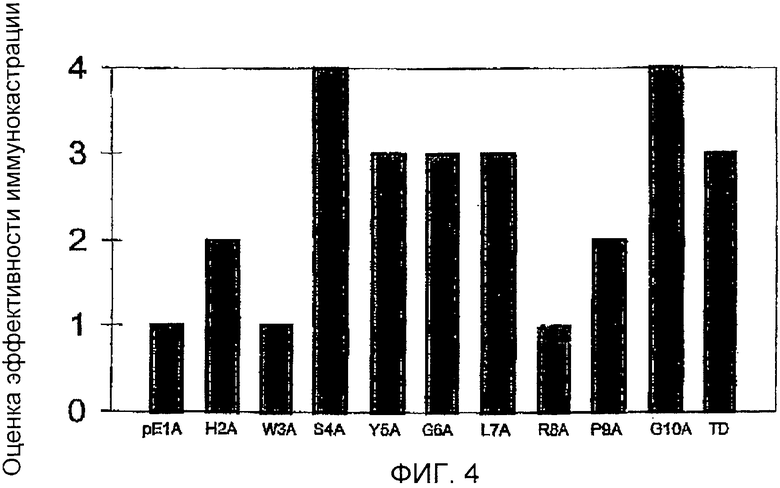
Фигура 4: Оценка эффективности иммунокастрации нескольких групп свиней, иммунизированных GnRH-тандемными димерными пептидами (62 мкг), конъюгированными с овальбумином и Specol, используемыми в качестве адъюванта. Исходный пептид был GnRH-тандемным (Cys-OH) димером (аббревиатура Cys-OH). Из этого пептида все обнаруживающие аланин при сканировании пептиды использовали для иммунизации. Эффективность иммунокастрации оценивали по шкале 1-4 (1 = иммунокастрация не происходила, 2 = меньшинство свиней были иммунокастрированными, 3 = большинство свиней были иммунокастрированными, 4 = все свиньи были иммунокастрированными).
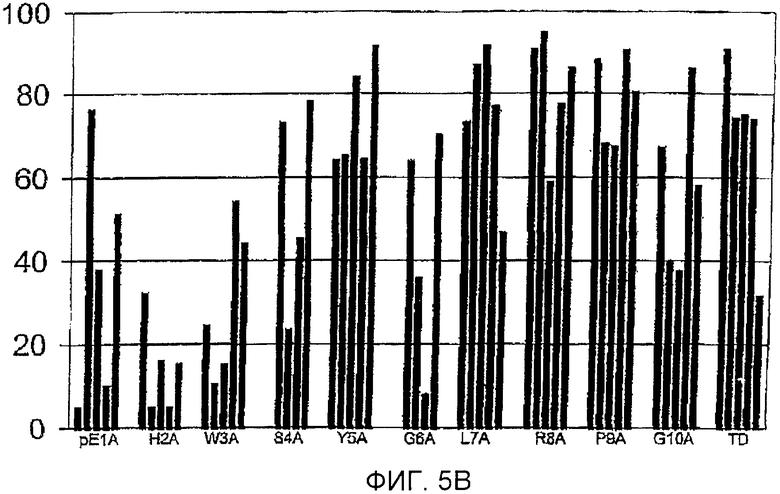
Фигура 5: Процент йодированного GnRH-I, вытесняемого GnRH-I (фигура 5А) и GnRH-II (фигура 5В), для сывороток индивидуальных животных (каждый столбец обозначает одно животное) после иммунизации тандемными димерными пептидами GnRH с заменами аланином, указанными под каждым кластером столбцов. Антисыворотку, полученную спустя 3 недели после бустер-иммунизации (10 wpv), разводили 1:100-1:10000. GnRH-I и GnRH-II для вытеснения добавляли в концентрации 250 пмоль на мл.
Фигура 5А
Горизонтальная ось: пептиды, используемые для иммунизации.
Вертикальная ось: процент вытеснения йодированного GnRH-I нейодированным GnRH-I.
Фигура 5В
Горизонтальная ось: пептиды, используемые для иммунизации.
Вертикальная ось: процент вытеснения йодированного GnRH-I нейодированным GnRH-II.
Подробное описание
Согласно признаку данного изобретения пептиды данного изобретения имеют общую формулу, изображенную посредством (SEQ ID NO:4) или следующим однобуквенным кодом:
pEHWSYkLRPGQHWSYkLRPGC#
В этой общей формуле Q обозначает Gin и перед ним может находиться X, где Х обозначает спейсер. Несколько из этих аминокислот были заменены другими аминокислотами. В этих формулах положение замены изображается жирным шрифтом или подчеркнутыми буквами. Прописные буквы обозначают левовращающие аминокислоты, строчные буквы обозначают правовращающие аминокислоты, например К: L-Lys; k: D-Lys. Затем их иммуногенную реакцию определяли при связывании с носителем, обычно овальбумином, но могут быть использованы и другие носители, такие как KLH (гемоцианин моллюска), БСА.
Будет понятно, что последовательность GnRH-II-пептида (SEQ ID NO:2) определяет, какие аминокислоты могут быть заменены таким образом, чтобы все еще было возможным эффективное дифференцирование на основе иммунной реакции этих двух последовательностей, GnRH-I и GnRH-II.
Для определения дифференцирования антител, индуцированных GnRH-вакцинами данного изобретения, между GnRH-I и GnRH-II, выполняли анализ связывания GnRH-антитела для определения, связываются ли антитела, индуцированные против GnRH-I-тандемного димерного пептида или его аналогов с заменами аланином, с GnRH-II или не могут связываться с GnRH-II. Разведения сыворотки предынкубировали с GnRH-I, GnRH-II, контрольным пептидом или без пептидов. Затем добавляли йодированный GnRH-I для конкуренции с предынкубированными пептидами за связывание с этими антителами.
Эту процедуру выполняли для сыворотки, собранной до и после бустер-иммунизации, так как специфичность антител может увеличиваться после бустер-иммунизации.
Когда пептиды данного изобретения использовали в виде конъюгатов с овальбумином (OVA-конъюгатов) и сравнивали с контролями, все показали эффективность в иммунокастрации молодых самцов свиней. Сравнение с известным конъюгатом G6k-GnRH-тандемный димер-OVA (см. таблицу 1) показало, что пептиды данного изобретения обнаруживали сравнимую или сходную эффективность, даже хотя их сходство с «собственным» гормоном GnRH-I уменьшалось. Пептиды данного изобретения дают иммуногенную реакцию, которая позволяет эффективно дифференцировать GnRH-I и GnRH-II. Эти пептиды приводили к маленьким яичкам и низким уровням тестостерона. Более конкретно, пептидами, приводящими к маленьким яичкам и низкому уровню тестостерона, являются R8A, G10A и S4A. Предпочтительными пептидами на основе иммунологической селективности между GnRH-I и GnRH-II являются S4A и рЕ1А.
В предпочтительном варианте пептид выбран из группы, состоящей из pEHWAYkLRPGQHWAYkLRPGC# (SEQ ID NO:5), pEHWSYkLAPGQHWSYkLAPGC# (SEQ ID NO:6) и pEHWSYkLRPAQHWSYkLRPAC# (SEQ ID NO:7). Более предпочтительно он выбран из группы, состоящей из pEHWSYkLAPGQHWSYkLAPGC# (SEQ ID NO:6) и pEHWSYkLRPAQHWSYkLRPAC# (SEQ ID NO:7).
В пептидах согласно данному изобретению димеризация тандемных звеньев может, например, происходить через карбоксильный конец или через амино-конец. Два тандемных звена могут быть, например, димеризованы посредством дисульфидного или простого тиоэфирного мостика. Для димеризации тандемных последовательностей может быть использован Cys в положении 21 или Cys может быть синтезирован перед глутаминовой кислотой в положении 1. Другие способы димеризации или мультимеризации GnRH-тандемных звеньев могут быть также найдены в предыдущем уровне техники. Если Cys в положении 21 участвует в димеризации и, следовательно, недоступен для связывания, можно также использовать другую аминокислоту тандема, которая может быть связана. Если димеризация или мультимеризация приводит к потере доступных сайтов, с которыми может конъюгироваться соединение-носитель, достаточно ограничить выбор заменяющих аминокислот аминокислотой с подходящей боковой цепью. Такой заменяющей аминокислотой может быть, например, L- или D-Lys, L- или D-Glu или другая аминокислота, содержащая боковую цепь, которая делает возможным связывание с соединением-носителем. Как L-, так и D-замены были испытаны, и было обнаружено, что они оказывают одно и то же действие.
Более конкретно, примером такого предпочтительного пептида в соответствии с данным изобретением является димер D-Lys6-тандем-GnRH (SEQ ID NO:8) следующей формулы:
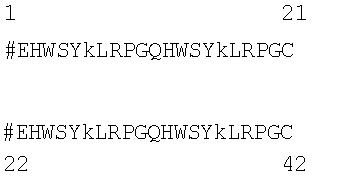
В этом примере варианта данного изобретения можно заменить одну из аминокислот этого тандемного димера другой аминокислотой.
Другие пептиды или пептидные последовательности или связанные пептидные последовательности, в которых присутствуют мономеризованные, димеризованные или мультимеризованные GnRH-тандемные звенья, которые содержат аминокислотные замены, являются также частью данного изобретения.
Далее, данное изобретение относится к композиции, которая содержит пептид, сделанный иммуногенным. Как известно квалифицированному в данной области специалисту, имеются различные способы получения иммуногенной формы вещества, которое само по себе не является иммуногенным. Одной из возможностей является связывание пептида данного изобретения с подходящим белком-носителем. Подходящим белком-носителем является овальбумин, KLH или БСА. В тандемном пептиде для химического связывания может быть удобным образом использован цистеин при N-или С-конце. В тандемном димерном пептиде связывание может быть также выполнено с использованием простой или модифицированной боковой цепи (D-)лизина, (D-)глутамина или любой другой модифицированной аминокислоты, заменяющей аминокислоты данной пептидной последовательности. Подходящие способы связывания и белки-носители хорошо известны специалистам с обычной квалификацией в данной области.
Согласно данному изобретению предпочтительной является композиция, которая отличается тем, что она содержит иммуногенный конъюгат белка, такого как овальбумин, и пептида или пептидной композиции.
Композиция данного изобретения может быть использована в форме вакцины. Для этой цели композиция может быть приготовлена в форме, которая пригодна для введения. Посредством введения вакцины данного изобретения генерируется иммуногенная реакция против GnRH, предпочтительно иммуногенная реакция против GnRH-I.
Таким образом, данное изобретение относится также к способу иммунизации млекопитающего против GnRH-I посредством вакцинации млекопитающего вакциной в соответствии с данным изобретением. В предпочтительном варианте данное изобретение относится к способу селективной иммунизации млекопитающего против GnRH-I вакциной в соответствии с данным изобретением.
Конечно, вакцинный препарат данного изобретения может комбинироваться по меньшей мере с одним иммуноадъювантом. Подходящие иммуноадъюванты известны специалистам с квалификацией в данной области. Предпочтительным адъювантом согласно данному изобретению может быть Specol или двойная масляная эмульсия, но могут быть также использованы другие адъюванты, которые не индуцируют побочных реакций или индуцируют лишь слабые побочные реакции. Данное изобретение может быть использовано в способах для иммунизации индивидуумов, выбранных из большого диапазона позвоночных животных и, в частности, млекопитающих, против GnRH-I. В предпочтительном варианте данного изобретения эта вакцина может вводиться в одной дозе, которая имеет ту же самую эффективность, что и известные в настоящее время вакцины, которые должны вводиться в виде двух доз. Иммунизация против GnRH-I, предпочтительно селективная, могла бы, например, использоваться для стерилизации, например, небольших домашних животных, таких как самцы и самки кошек и собак, или для лечения агрессивности у самцов собак и быков. Другими возможными причинами для иммунизации против GnRH-I с использованием данного изобретения являются предотвращение периода течки у самок животных, таких как собаки, кошки и коровы, и предупреждение или лечение беспокойного состояния у самцов животных, откармливаемых на убой. В здравоохранении человека иммунизация против GnRH, предпочтительно селективная против либо GnRH-I, либо GnRH-II, может быть использована в лечении рака предстательной железы и рака молочной железы и, если требуется, в лечении некоторых форм рака гипофиза.
Предпочтительным вариантом является способ улучшения качества туши свиней, в котором свиней вакцинируют вакцинным препаратом в соответствии с данным изобретением. Данное изобретение иллюстрируется следующей экспериментальной частью.
I. Иммунокастрация свиней
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
Ацетонитрил (ACN) имел чистоту, требующуюся для HPLC-S-градиента, N-метилпирролидон (НМП), диизопропилэтиламин (DIEA), диметилформамид (ДМФ), трифторуксусная кислота (ТФУ) и пиперидин имели чистоту, требующуюся для пептидного синтеза, и все были получены из Biosolve (Valkenswaard, NL). N-гидроксибензотриазол (HOBt) и 2-(1Н-бензотриазол-1-ил-1,1,3,3-тетраметилуронийгексафторфосфат (HBTU) получали из Richelieu Biotechnologies Inc. (Hamon, Canada). Бензотриазол-1-ил-окси-трис-пирролидинофосфонийгексафторфосфат (РуВОР) получали из Novabiochem (Laufelfingen, Switzerland). Тиоанизол (ТА), этандитиол (EDT), диметилсульфоксид (ДМСО), пентан и диметиламинопиридин (DMAP) с проаналитической чистотой получали из Merck (Darmstadt, Germany). Диэтиловый эфир очищали на колонке активированного, щелочного оксида алюминия перед использованием. Производные аминокислот и смолы получали из Bachem Feinchemicalien AG (Bubendorf, Switzerland).
Синтез множественных пептидов (MPS)
Микролаборатория 2200 Гамильтона была запрограммирована для подачи промывных растворителей и реагентов к штативу с 40 индивидуальными колонками на 4 мл с фильтром, содержащими 30 ммоль смолы для пептидного синтеза. Эти колонки дренировали после каждой стадии с использованием вакуума. Цикл связывания был основан на Fmoc-химии с использованием стадий двойного связывания:
1. Промывка НМП (1 мл)
2. 30% (об/об) пиперидин/НМП (3 мин, 0,5 мл)
3. 30% (об/об) пиперидин/НМП (17 мин, 0,5 мл)
4. Промывка НМП (5×1 мл)
5. Двойное связывание (2×30 мин)
6. Промывка НМП (2×1 мл)
Стадия связывания: Fmoc-аминокислоту в НМП (0,4 М, 0,25 мл), HBTU/HOBt (0,45 М, 0,22 мл) в ДМФ и DIEA (2 М, 0,2 мл) в НМП переносили в реакционный сосуд и давали реагировать в течение 30-50 минут. Реакционную смесь дренировали (сливали) и процедуру связывания повторяли еще один раз.
После связывания последней аминокислоты Fmoc-группу отщепляли смесью 30% пиперидин/НМП, пептиды промывали, ацетилировали в течение 30 минут с использованием смеси НМП/уксусный ангидрид/DIEA 10/1/0,2, промывали снова и сушили. Пептиды освобождали от защитных групп и отщепляли от смолы в течение 2 часов в смеси 1,5 мл ТФУ/фенол/ТА/вода/EDT 10/0,75/0,5/5,0/0,25 (реагенте К). Смесь для отщепления фильтровали, смолу промывали 0,5 мл ТФУ и пептид осаждали добавлением 13 мл смеси пентан/диэтиловый эфир 1/1. После центрифугирования осадок экстрагировали снова смесью пентан/диэтиловый эфир. Осадок сушили, растворяли в смеси ACN/вода 1/1 и лиофилизировали. Эта процедура дает в зависимости от молекулярной массы 25-70 мг пептида.
Синтезированные пептидные последовательности в однобуквенном коде аминокислот суммированы в таблице 1.
Аминокислотная последовательность в однобуквенном коде синтезированных пептидов
Аналитическая ВЖХ
Для анализа пептидов авторы использовали систему LC-MS (электрораспыление), которая состоит из двух насосов модели 510 Waters, контроллера градиента модели 680 Waters, автоинжектора 712 Waters WISP и фотодиодного матричного детектора 991 Waters. Использовали масс-спектрометр Micromass Quattro II sq, который применяли в режиме положительного иона. Продукты анализировали в линейном градиенте от 10% ACN/вода с 0,05% ТФУ до 70% ACN/вода с 0,05% ТФУ в течение 30 минут на колонке Waters Delta Pak С18-100Å (3,9×150 мм, 5 мм) при 1 мл/мин при 215 нм. Все продукты имели чистоту между 40 и 70% в соответствии с площадью пиков.
Димеризация
Неочищенные продукты димеризовали растворением продуктов в 20% ДМСО в воде. рН доводили до 5-6 1% NH4НСО3, поддерживающим прозрачный раствор. Коррекцию рН выполняли 1% уксусной кислотой. После перемешивания при комнатной температуре в течение по меньшей мере 5 часов продукты хранили при -20°С до очистки.
Препаративная ВЖХ
Очистки пептидов проводили с использованием жидкостного хроматографа Waters Prep 4000, оборудованного модулем RCM Waters с двумя картриджами PrepPak плюс предохранительный картридж (40×210 мм или 25×210 мм), заполненными материалом delta-Pak С18-100Å (15 мм). Обычно очистки проводили с использованием тех же самых элюентов, что и в аналитической ВЖХ, но со скоростью градиента 0,5% АСМ/мин и скоростью тока 40 или 100 мл/мин. Пептиды детектировали при 215-230 нм с использованием спектрофотометра 486 Waters с препаративной ячейкой. Пептиды лиофилизировали, и было определено, что чистота была по меньшей мере 90%.
Получение конъюгатов
Для конъюгации посредством гидрохлорида N-этил-N-(3'-диметиламинопропил)карбодиимида (EDC) с альбумином куриного яйца (овальбумином) (OVA) равные веса как пептида, так и белка-носителя растворяли раздельно в воде milli Q и оба раствора хорошо смешивали. Затем 10-кратный избыток в расчете на весовые эквиваленты EDC растворяли в воде milli Q. Затем этот раствор медленно добавляли к раствору пептид/OVA при непрерывном перемешивании, рН этого конечного раствора равен 5. После по меньшей мере 6 часов медленного встряхивания этот продукт диализовали (предел MW 10000) против 300-кратного избытка воды milli Q в течение двух дней. Воду сменяли два раза в день. Нагрузку рассчитывали из сравнительного аминокислотного анализа конъюгата и белка-носителя. Аминокислотный анализ выполняли с использованием системы Waters Pico-Tag, после гидролиза в мини-компьютере (рабочей станции) Pico-Tag с использованием 6 н. HCl при 150°С в течение 1 часа и дериватизации фенилизотиоцианатом.
Согласно аминокислотному анализу эти конъюгаты содержали между 0,3 и 0,5 мг пептида на мг белка-носителя, за исключением конъюгата Р9А-G6k-GnRH-тандемного димера, который содержал только 0,16 мг пептида на мг овальбумина.
Конъюгат G6k-GnRH-тандем-димер/OVA имеет аббревиатуру G6k-TD. Конъюгаты с заменами в виде аланина имеют аббревиатуру, использующую название аминокислоты в природной последовательности GnRH, которая заменяется, ее положения и А для замены аланином. Например, конъюгат рЕ1А-G6k-GnRH-тандем-димер OVA имеет аббревиатуру рЕ1А.
Приготовление эмульсии
Specol (Special Oil Phase, ID-DLO, Lelystad, The Netherlands), состоящий из двух детергентов в легком минеральном масле, использовали в качестве масляной фазы (Bokhout et al., 1981). Эмульсии типа вода в масле (WIO) готовили с использованием Ultra Turrax (Janke and Kunkel, Staufen, Germany) с перемешивающим стержнем. Масляную фазу Specol (5 частей об/об) вносили в стеклянный сосуд на 25 мл и медленно добавляли водную фазу (4 части об/об), состоящую из конъюгата в воде milli Q, при перемешивании эмульсии. После добавления водной фазы эмульсию перемешивали в течение полминуты при той же самой скорости вращения (15000 об/мин). Эмульсии выдерживали в течение ночи при 4°С для проверки стабильности и вводили животным на следующий день.
Животные
Этот эксперимент проводили с самцами-поросятами в возрасте приблизительно 10 недель. Кроссбредных поросят содержали в клетках с решетчатой половиной пола и им предоставляли доступ ad libitum к корму и воде.
Иммунизация
Поросят случайным образом относили к вариантам обработки, 6 или 7 поросят на каждый вариант обработки. Всех животных инъецировали 2 мл эмульсии, содержащей димеризованные конъюгаты тандемного GnRH (т.е. 62 мкг пептида), или эмульсии без антигена. Инъекции вводили внутримышечно в шею в начале эксперимента (день 0) и спустя 7 недель (7 wpv (недель после вакцинации)).
Измерения и взятие проб крови
Животных взвешивали в день 0 и спустя 7 и 13 недель после вакцинации. Размеры яичка определяли измерением длины яичка измерительным циркулем (vernier calipers) в день 0 и спустя 7, 10 и 13 недель после этого. Размеры яичек регистрировали в виде среднего размера для обоих яичек.
Пробы крови брали посредством прокалывания яремной вены в те же самые дни, когда измеряли размеры яичек, а также спустя 4 недели после начальной иммунизации. Пробы крови выдерживали в течение ночи при 4°С и на следующий день сыворотку получали центрифугированием (1500 g, 15 мин). Пробы сыворотки хранили при -20°С до анализа.
Оценка после убоя
После убоя яички извлекали, освобождали от эпидидимиса и взвешивали. Веса яичек регистрировали в виде среднего размера из величин обоих яичек.
Антитела к пептидам
Антитела к пептидам, используемые для иммунизации, определяли с использованием ELISA. Пептиды наносили в виде покрытия в лунки микротитрационного планшета с использованием глутардиальдегида (GDA). GDA наносили в виде покрытия на поверхность лунок инкубированием с 0,2% GDA в 0,1 М фосфатном буфере (рН 6) в течение 4 часов при комнатной температуре. Планшеты промывали 3 раза в течение 10 минут 0,1 М фосфатным буфером (рН 8). Один микрограмм пептида в 100 мл фосфатного буфера (0,1 М, рН 8) наносили в виде покрытия на лунку инкубированием в течение 3 часов при 37°С. Планшеты хранили при -20°С до использования. Оттаявшие планшеты промывали 3 раза в течение 10 минут водой milli Q, содержащей 8,2 г NaCl, 1,15 г Na2HPO4·2Н2O, 0,20 г NaH2PO4·2H2O и 5 мл 10% раствора Твина 80 в воде на литр воды.
Серийным разведениям антипептидных сывороток давали реагировать с нанесенными в виде покрытия пептидами в течение 1 часа при 25°С. После промывания 3 раза в течение 10 минут козьи антитела против свиного IgG, связанные с пероксидазой хрена (Dako, Glastrup, Denmark), вводили в качестве второго антитела в течение 1 часа и в качестве субстрата использовали ABTS (Boehringer, Mannheim, Germany) (250 мл (2 г/100 мл) в 10 мл субстратного буфера, к которому добавляли 20 мл Н2О2 (3% раствор)). Поглощение измеряли при 405 нм.
Антитела к GnRH
Антитела к GnRH определяли, как описано Meloen et al. (Vaccine 12, 741-746 (1994)). Серийным разведениям антисывороток свиней давали реагировать для связывания с 125I-GnRH. Титры выражали в виде процентного связывания 125I-GnRH в конкретном разведении сыворотки.
Тестостерон
Уровни тестостерона в сыворотке определяли с использованием набора Coat-a-Count kit, приобретенного из DPC-лабораторий, Los Angeles, CA.
РЕЗУЛЬТАТЫ
Размер яичка и вес яичка
Спустя семь недель после первой иммунизации уже можно было наблюдать эффекты иммунокастрации посредством измерения размеров яичек. Три варианта обработки (R8A, G10A и G6k-TD) едва ли показали какое-либо приращение (<10 мм) среднего размера яичка во время бустер-иммунизации. Эти варианты обработки были успешными, с весом яичка при убое, равным 70 или менее граммов. Другие варианты обработки, которые были эффективными в отношении низких весов яичек, являются рЕ1А и S4A, тогда как в группе Y5A, L7A и Р9А 2 животных, 1 и 1 животное не отвечали на иммунизацию соответственно (таблица 2). Индивидуальный вес яичка иммунокастрированных животных не превышал 70 граммов, что приводило к явному различию между иммунокастрированными и не-иммунокастрированными животными.
Эффективность различных обработок в отношении веса яичка
Реакция в виде антител
Средние титры антител против пептидов, использованных для иммунизации, представлены в таблице 3. Средний титр антител свиней, обработанных Н2А и Р9А, ниже, чем титр антител других обработок.
Титры антител индивидуальных животных против различных пептидов находятся в диапазоне от 2 до 4. У обработанных животных, которые не были иммунокастрированными, наблюдали самый низкий титр антипептидных антител. Животные, обработанные Н2А и W3A, не были иммунокастрированными, но присутствовали значительные антипептидные титры.
Проценты связывания антител против GnRH при разведении сыворотки 1/2000 были недетектируемыми или низкими в группах Н2А и W3A. Однако при разведении сыворотки 1/200 антитела были детектируемы в сыворотках всех животных в группе Н2А и трех животных группы W3A.
Животные с низкими средними титрами антител к GnRH не были иммунокастрированными. Высокие титры антител действительно всегда приводили к успешно иммунокастрированным животным. Вес яичек животных с промежуточными титрами антител варьировался от 15 до 300 граммов.
Средние титры антител к GnRH на обработку показали явную связь с весом яичка (медианой) на обработку (таблица 3).
Тестостерон
Уровни тестостерона всех успешно обработанных животных были низкими и уменьшались после второй иммунизации. Однако большинство (n=31) животных отражали эффект кастрации так рано, как спустя четыре недели после начальной иммунизации, обнаруживая недетектируемые уровни тестостерона. Вес яичек этих животных варьировался от 8 до 36 граммов.
Таким образом, большинство животных были эффективно иммунизированы после введения единственной дозы.
Хотя уровни тестостерона при убое были низкими для всех иммунокастрированных животных, два животных с весами яичка 65 и 70 граммов имели значимые уровни тестостерона 3,80 и 1,18 пмоль/мл соответственно.
Пептиды, которые приводили к низкому весу яичка, в комбинации с недетектируемыми уровнями тестостерона, рассматриваются как наиболее эффективные пептиды для иммунокастрации свиней. Этими пептидами являются S4A, R8A и G10A.
Уровни тестостерона при убое свиней, которые не были иммунокастрированными животными, варьировались между 0,46 и 48,91 пмоль/мл.
Действие различных обработок на средний вес яичка, титр антител против пептидов, процентное связывание антител к
II. Замена D-аминокислоты L-аминокислотой
D-лизин (k) в положениях 6, 16, 27 и 37 в тандемном димере заменяли L-лизином (К) и полученный пептид испытывали (таблица 4).
G6k-GnRH-тандем: pEHWSYkLRPGQHWSYkLRPGC# (SEQ ID NO:9)
G6k-GnRH-тандем: pEHWSYKLRPGQHWSYKLRPGC# (SEQ ID NO:14)
Замена D-лизина L-лизином не изменяет эффективность вакцинного антигена.
III. Дифференцирование GnRH-I и GnRH-II
Конкурентный радиоиммуноанализ связывания антитела против GnRH выполняли для определения, связываются ли антитела, индуцированные против G6k-GnRH-тандемного (SEQ ID NO:9) димерного пептида или его аналогов с заменами аминокислот аланином, с GnRH-II или не связываются с GnRH-II. Серийные разведения предынкубировали либо с GnRH-I, либо с GnRH-II, контрольным пептидом или без пептида. Затем добавляли йодированный GnRH-I для конкуренции с предынкубированными пептидами за связывание с этими антителами.
Эту процедуру выполняли для сыворотки, собранной до и после бустер-иммунизации, так как специфичность антител может увеличиваться после бустер-иммунизации вследствие созревания антител.
Материалы и методы
Пробы сыворотки при взятии крови при 7 неделях и 10 неделях после вакцинации (при 3 неделях после бустер-иммунизации) разводили 1/100-1/10000 в ЗФР с 0,4% БСА (буфере для разведения). Пятьдесят мкл разведения сыворотки помещали в микротитрационные планшеты и добавляли 25 мкл раствора пептида (0, 0,25, 2,5 или 25 пмоль пептида в буфере для разведения на лунку). Этой смеси давали инкубироваться в течение 24 часов при 4°С. На следующий день добавляли 25 мкл йодированного GnRH (приблизительно 13000 имп/мин) и после выдерживания в течение ночи (4°С) несвязанный пептид отделяли от связанного пептида при помощи угля. После центрифугирования супернатант отделяли, считали и рассчитывали процент йодированного GnRH, связанного с антителами.
Результаты
Связывание антителами йодированного GnRH-I демонстрировали для сывороток всех обработок, за исключением сывороток Н2А и W3A, полученных при 7 неделях после иммунизации. Конкуренция с GnRH-I приводила к сильному снижению йодирования GnRH-I (см. фигуру 1), тогда как вытеснение йодированного GnRH-I посредством GnRH-II было высоковариабельным. Как и ожидалось, ни одна из сывороток не связывалась с контрольным пептидом.
Для сывороток при 10 неделях после вакцинации: В сыворотках почти всех обработок йодированный GnRH-I вытеснялся GnRH-I более чем на 80% (фигура 2А). Конкуренция с использованием GnRH-II за связывание антител с йодированным GnRH-I приводила к полному или частичному вытеснению йодированного GnRH-I почти во всех сыворотках обработок R8A, Р9А, G10A и G6k-TD и в половине сывороток обработок Y5A и L7A (фигура 2В). Однако антитела по существу всех животных групп рЕ1А, Н2А, W3A и S4A были полностью лишены связывания с GnRH-II и, следовательно, были специфическими для GnRH-I.
Результаты исследований вытеснения пре-бустерных (7 недель после вакцинации) сывороток отличались от результатов для 10-недельных сывороток. Опять сыворотки всех обработок (за исключением сывороток Н2А и W3A вследствие низких титров антител) узнавали GnRH-I, как определено вытеснением йодированного GnRH-I нейодированным GnRH-I (фигура 3А). Однако в противоположность 10-недельным после иммунизации сывороткам (фигура 2) большинство сывороток всех обработок, в том числе рЕ1А и S4A, действительно узнавали GnRH-II (фигура 3В). Таким образом, сыворотки при 7 неделях после иммунизации рЕ1А и S4A не были специфическими для GnRH-I, но специфичность в отношении GnRH-I, но не в отношении GnRH-II, полностью устанавливалась при 3 неделях после бустер-иммунизации (при 10 неделях после вакцинации).
Результаты связывания антител, полученные с пре-бустерными сыворотками, отличались от результатов, полученных для сывороток при 10 неделях после вакцинации. Антисыворотки обработок рЕ1А и S4A испытывали на их способность узнавать GnRH-I или GnRH-II (см. фигуру 3). Для обоих пептидов сыворотки, полученные через 10 недель после вакцинации, от 6 из 7 животных не связывались с GnRH-II. Из pE1A-сывороток, полученных до бустер-иммунизации, 4 из 7 животных обнаружили связывание с GnRH-II. Две сыворотки не узнавали GnRH-II, и одна сыворотка не обнаруживала связывания с йодированным GnRH. Результаты с пре-бустерными 34А-сыворотками противоречат результатам для сывороток при 10 неделях после вакцинации. Пре-бустерные сыворотки всех животных S4А-группы не узнавали GnRH-II, причем ингибирование связывающей способности йодированного GnRH является сходным как для GnRH-I, так и для GnRH-II.
IV. Иммунокастрация свиней конъюгатами GnRH-тандемных димеров
Подобно аланин-сканированию, описанному в примере 1, синтезировали пептиды с заменами аланином аминокислот GnRH-тандемного пептида (таблица 5). Пептиды димеризовали, очищали и конъюгировали с OVA, как описано ранее.

рЕ = пироглутаминовая кислота; * = ацетил; # = амид;
- = аминокислота в этом положении не отличается от аминокислоты в этом же положении в GnRH-тандеме.
Четыре или пять свиней на группу иммунизировали 62 мкг пептидного эквивалента конъюгатов в адъюванте Specol.
Эффективность иммунокастрации конъюгатов представлена на фигуре 4.
Титры антител, направленных против пептидов, которые использовали для иммунизации, были низкими для обработанных pE1A, H2A и R8A свиней, тогда как высокие титры антипептидных антител получили для групп W3A, S4A, Y5A и G10A. Титры антител против GnRH-I были в согласии с титрами антипептидных антител, за исключением титров антител свиней, обработанных W3A: высокие титры антипептидных антител были определены у этих животных, однако было обнаружено, что связывающая активность этих антител в отношении йодированного GnRH-I была низкой.
V. Дифференцирование GnRH-I и GnRH-II антисывороток против GnRH-тандемного димера
Связывание антител йодированного GnRH-I в радиоиммуноанализе наблюдали для сывороток при 10 неделях после вакцинации (разведенных 1/100-1/10000) всех животных, обработанных GnRH-тандемными димерными пептидами с заменами аланином разных аминокислот. Конкуренция с GnRH-I приводила к сильному уменьшению связывания йодированного GnRH-I. В сыворотках почти всех обработок йодированный GnRH-I вытеснялся GnRH-I более чем на 80% (фигура 5А).
Вытеснение йодированного GnRH-I с использованием GnRH-II было высоковариабельным среди групп.Оно приводило к полному или частичному вытеснению йодированного GnRH-I в почти всех сыворотках обработок Y5A, L7A, R8A, Р9А и TD (фигура 5В), тогда как для большинства сывороток остальных обработок определяли только частичное вытеснение йодированного GnRH-I при использовании GnRH-II или вообще не было никакого вытеснения.
















Изобретение относится к области биотехнологии и ветеринарии. Пептид содержит последовательность GnRH SEQ ID NO:1 или SEQ ID NO:2 и обеспечивает иммунный ответ, позволяющий дифференцировать GnRH-I и GnRH-II и содержание тестостерона, которое не определяется после вакцинации. Предложены также вакцины, содержащие такой пептид, антитело к нему и применение пептида для приготовления фармацевтической композиции. Изобретение может быть использовано в ветеринарии и животноводстве для вакцинации, связанной с природной функцией GnRH. 5 н. и 10 з.п. ф-лы, 5 ил., 5 табл.
pEHWAYkLRPGQHWAYkLRPGC#, pEHWSYkLAPGQHWSYkLAPGC# и pEHWSYkLRPAQHWSYkLRPAC#, предпочтительно выбран из группы, состоящей из pEHWSYkLAPGQHWSYkLAPGC# и pEHWSYkLRPAQHWSYkLRPAC#.
| WO 9640755, 19.12.1996 | |||
| Способ получения дисперсного азокрасителя | 1972 |
|
SU464124A3 |
| АНТАГОНИСТ ЛГ-РИЛИЗИНГ-ФАКТОРА (LHRH), КОМПЛЕКС, СПОСОБ ХИМИЧЕСКОЙ КАСТРАЦИИ И/ИЛИ ЛЕЧЕНИЯ ГОНАДОТРОПИНЗАВИСИМЫХ РАССТРОЙСТВ | 1994 |
|
RU2130464C1 |

