Изобретение относится к плазмидным векторам, содержащим ДНК-последовательности, кодирующим усеченные и химерные формы белка p185neu, и к их применению в ДНК-вакцинации против Her-2/neu (ErbB-2)-позитивных опухолей, экспрессирующих белок p185neu. Плазмиды по изобретению способны вызывать защитный иммунный ответ, основанный на индукции антител и/или Т-лимфоцитов, в отношении опухолей, экспрессирующих белок p185neu. Дополнительно изобретение относится к фармацевтическим композициям, содержащим такие плазмиды, и к их применению в профилактике и лечении p185neu-позитивных опухолей.
Предпосылки к созданию изобретения
Неопластические клетки зачастую отличаются от нормальных клеток тем, что экспрессируют несколько аномальных белков. Вследствие этой аномальной экспрессии некоторые белки могут действовать как антигены, ассоциированные с опухолью (TAA). Это происходит потому, что иммунная система организма-хозяина может распознать эти аномалии и вызвать иммунную реакцию, которая могла бы защитить организм-хозяин от появления и развития опухоли. Являясь мишенью противоопухолевой иммунотерапии, TAA должны:
играть патогенентическую роль на определенной стадии неопластического роста;
определяться иммунной системой, даже когда опухоль дает начало клоновым вариантам, которые больше не экспрессируют гликопротеины главного комплекса гистосовместимости (HLA);
распознаваться как антителами, так и Т-лимфоцитами.
В последние годы некоторые TAA были обнаружены в карциномах человека. Среди них p185neu, белковый продукт онкогена Her-2/neu (ErbB2), является особенно подходящей мишенью для иммунотерапии (Lollini и Forni, 2003, Trends Immunol. 24: 62). p185neu представляет собой мембранный рецептор I класса рецепторов семейства тирозинкиназ, к которому также относится рецептор эпидермального фактора роста (EGF-R или ErbB-1) и другие родственные рецепторы (ErbB-3, ErbB-4), которые играют ключевую роль в пролиферации и дифференцировке клеток (Hynes и Stern, 1994, BBA 1198: 165).
p185neu рецепторный белок может быть разделен на три домена: внеклеточный домен (EC домен), трансмембранный домен (TM домен) и внутрицитоплазматический домен (IC домен). Недавно была опубликована кристаллографическая структура EC домена белка p185neu человека и крыс.Было описано, что этот домен состоит из четырех субдоменов (I/L1, II/CR1, III/L2 и IV/CR2) в общей сложности из приблизительно 630 аминокислот. Дополнительно было показано, что белок p185neu имеет жесткую структуру, которая позволяет ему взаимодействовать с другими рецепторами ErbB, образовывать димеры и вызывать трансдукцию пролиферативного сигнала, даже если этот белок не связывает лиганды непосредственно (Cho et al., 2003, Nature 421: 756).
Онкоген Her-2/neu (ErbB2) участвует в нормальных процессах эмбрионального органогенеза и эпителиального роста, хотя у взрослых он экспрессируется только в незначительных количествах (Press et al., 1990, Oncogene 5: 953). У людей гиперэкспрессия этого онкогена главным образом вызвана амплификацией гена. Онкоген Her-2/neu (ErbB2) гиперэкспрессируется примерно в 30% случаев карциномы молочной железы, и такая гиперэкспрессия связана с более быстрой опухолевой прогрессией (Slamon et al., 1989, Science 244: 707). Среди различных стратегий, которые были предложены, ДНК-вакцинация, по-видимому, является эффективным способом стимуляции иммунного ответа в отношении Her-2/neu-позитивных опухолей. Несмотря на то, что белок p185neu является «собственным» антигеном, т.е. белком, который обычно присутствует в организме, у пациентов с p185neu-позитивной карциномой молочной железы зачастую проявляется как клеточный, так и гуморальный иммунный ответ (Signoretti et al., 2000, J. Natl. Cancer Inst. 23: 1918; Disis et al., 1994, Cancer Res. 54: 16; Peoples et al., 1995, Proc. Natl. Acad. Sci. USA 92: 432). Одной из задач противоопухолевой иммунотерапии, направленной на белок p185ncu, является повышение интенсивности уже имеющегося иммунного ответа у пациентов, или генерирование иммунного ответа у пациентов, у которых этот ответ не выявлен. Тот факт, что белок p185neu является «собственным» антигеном, подразумевает, что вакцина должна быть способной преодолевать состояние иммунотолерантности.
Авторы настоящей патентной заявки первыми использовали и обосновали эффективность ДНК-вакцинации в активации иммунной защиты как в отношении спонтанных карцином молочной железы, так и трансплантируемых Her-2/neu-позитивных опухолей. Эти исследования подтвердили, что профилактика и лечение предраковых патологических изменений является достижимой целью. В частности, в экспериментах, целью которых является профилактика развития спонтанных опухолей молочных желез, которые возникают у трансгенных мышей, под действием Her-2/neu онкогена крыс (мыши FVB/neuT и мыши BALB-neuT), было показано, что плазмида, кодирующая ЕС и ТМ домены белка p185neu крыс, способна вызывать более эффективную защиту по сравнению с плазмидой, кодирующей полную длину белка p185neu, или плазмидой, кодирующей только ЕС домен (секретируемый антиген) (Amici et al., 2000, Gene Ther. 7: 703; Rovero et al., 2000, J. Immunol. 165: 5133). Аналогичные данные были получены Chen et al., (1998, Cancer Res. 58: 1965). Кроме того, было показано, что эффективность вакцинации ДНК плазмидами резко возрастает, если за ней следует очень короткий электрический импульс при внутримышечном введении плазмид (Quaglino et al., 2004, Cancer Res. 64: 2858). Другими авторами было показано, что плазмиды, кодирующие полную длину белка p185neu, при необходимости видоизмененные таким образом, что он не обладает тирозинкиназной активностью, являются эффективными для профилактики возникновения опухолей после трансплантации p185neu-позитивных злокачественных клеток (Wei-Zen et al., 1999, Int. J. Cancer 81: 748). Было доказано, что те же плазмиды настолько эффективны даже тогда, когда не содержащие лидерного сигнального пептида, отвечающего за процессинг белка в эндоплазматическом ретикулуме, они приводят к цитоплазматической локализации антигена p185neu. В тех случаях, когда используют плазмиды, кодирующие белок p185neu, который расположен в мембране, благодаря присутствию лидерного сигнального пептида, защитные действия зависят от гуморального иммунного ответа. И наоборот, иммунный ответ, опосредованный Т-лимфоцитами, наблюдается в тех случаях, если вакцина не содержит лидерного сигнального пептида, и, следовательно, белок p185neu расположен в цитоплазме трансфектированных клеток, а не на их плазматической мембране (Pilon et al., 2001, J. Immunol. 167: 3201). Кроме того, комбинированная вакцинация, полученная использованием как плазмид с лидерным сигнальным пептидом, так и плазмид, у которых этот лидерный сигнальный пептид был удален, является более эффективной для защиты от опухолевого роста (Piechocki et al., 2001, J. Immunol, 167; 3367). Это демонстрирует существование синергетического эффекта между гуморальным и клеточным ответами при профилактике p185neu-позитивных карцином (Reilly et al., 2001, Cancer Res. 61: 880).
Было доказано, что вакцинация плазмидой, кодирующей EC и TM домены (EC-TM плазмида), является эффективной не только для профилактики развития спонтанных p185neu-позитивных карцином, но также для лечения опухолевых масс диаметром 2 мм, вовлекая ряд эффекторных механизмов иммунной системы (клетки T-хелперы и Т-киллеры, антитела, макрофаги, нейтрофилы, естественные киллеры, Fc рецепторы, IFN-гамма и перфорины), которые координированно способствуют отторжению опухоли (Curcio et al., 2003, J. Clin. Invest. 111: 1161).
Описание изобретения
Были сконструированы особые плазмиды, кодирующие полноразмерный ТМ домен, и уменьшающиеся участки EC домена белка p185neu крыс. Усеченные плазмиды, полученные делецией NH2-концевых 240 пар оснований (bp) или кратные величине этой длины, использовали в экспериментах, нацеленных на предотвращение роста трансплантируемых опухолевых клеток, гиперэкспрессирующих белок p185neu крыс (TUBO клеток). Более того, ряд плазмид, кодирующих химерные формы белка p185neu, были созданы путем добавления NH2-концевых участков кДНК ErbB2 человека к последовательностям, кодирующим усеченные формы белка крыс для реконструкции полной последовательности белка.
Защитное действие, достигаемое вакцинацией плазмиды, кодирующей полноразмерные домены EC и TM, главным образом обусловлено антигенной защитой, тогда как защита, достигаемая использованием плазмид, кодирующих усеченные формы белка p185neu крыс, во многих случаях не связана со значительной продукцией антител.
Исходя из результатов экспериментов in vivo были выбраны плазмиды, способные индуцировать сильный иммунный ответ, как гуморальный, так и опосредованный Т-лимфоцитами.
В первом аспекте изобретение относится к плазмидам, содержащим кодирующую последовательность фрагмента белка p185neu, которая выбрана из группы, состоящей из SEQ ID NO: 1-5; или последовательность, кодирующую химерный белок p185neu, которая выбрана из группы, состоящей из SEQ ID NO: 6-12 (эталонные последовательности для генов, кодирующих белки p185neu человека и крыс депонированы в банке генов (Gene Bank) под регистрационными номерами M11730 и X03362 соответственно).
ДНК-последовательности, кодирующие усеченные и химерные формы белка p185neu по изобретению, могут быть вставлены в любые плазмидные векторы, подходящие для использования у млекопитающих, в частности у людей. Помимо вышеуказанных кодирующих последовательностей плазмиды могут содержать функциональные элементы для контроля транскрипции, в частности промоторы, предпочтительно CMV промотор, расположенный перед кодирующей последовательностью, последовательности инициации транскрипции и стоп-последовательности; селективный маркер, предпочтительно гены резистентности к ампициллину или канамицину; CpG мотивы; сайт полиаденилирования; и на случай энхансеры или активаторы транскрипции. Элементы для контроля транскрипции должны подходить для использования векторов у млекопитающих, в частности у людей.
В другом аспекте изобретение относится к фармацевтической композиции, содержащей ДНК-плазмиду, определенную выше, вместе с фармацевтически приемлемыми носителями и эксципиентами. Альтернативно, композиции могут содержать смеси двух или нескольких различных плазмид, кодирующих как усеченные, так и химерные формы белка p185neu. Для ДНК-вакцинации предпочтительно используют фармацевтические композиции в форме, подходящей для парентерального введения, предпочтительно в форме инъекционного раствора. Принципы и способы ДНК-вакцинации известны специалистам в этой области и описаны, например, Liu, 2003; J. Int. Med. 253: 402.
Использование плазмид, кодирующих усеченные и химерные формы p185neu, для профилактической и терапевтической вакцинации против p185neu-позитивных (Her-2/neu-, ErbB-2-позитивных) опухолей имеет целый ряд преимуществ, которые повышают их эффективность. Для плазмид, кодирующих усеченные формы, такими преимуществами являются:
1) возможность получить вакцину, кодирующую только определенные участки TAA, в отношении которых требуется развитие иммунного ответа; при использовании этой вакцины меньше вероятность развития аутоиммунных процессов;
2) эксклюзивная индукция некоторых выбранных форм иммунного ответа, т.е. опосредованного антителами или Т-лимфоцитами;
3) возможность получать вакцины, которые объединяют множественные эпитопы, имеющие определенную иммуногенность, путем связывания друг с другом фрагментов кДНК, которые кодируют различные усеченные формы, необязательно последовательно.
Использование химерных плазмид, полученных комбинацией усеченных форм p185neu животных различных видов, позволяет:
a) вакцинировать плазмидами, кодирующими белковые детерминанты тех же видов, подвергаемых иммунизации, например людей, что может вызывать специфический высокоаффинный ответ;
b) объединять плазмиды, кодирующие антигенные детерминанты одних и тех же иммунизируемых видов, с последовательностями кДНК, кодирующими антигенные детерминанты других видов, антигенные детерминанты, демонстрирующие значительное сходство, но отличающиеся тем, что антигенные детерминанты других видов вызывают более интенсивный иммунный ответ, следовательно, преодолевая состояние толерантности. Эти аллогенные детерминанты, которые распознаются как частично экзогенные, действуют как вспомогательные детерминанты, облегчающие индукцию более интенсивного ответа и высвобождение цитокинов;
c) объединять плазмиды, кодирующие антигенные детерминанты одинаковых видов, с последовательностями кДНК, которые, кодируя детерминанты других видов, у некоторых индивидуумов могут образовывать гетероклитические детерминанты, которые связываются с высокой аффинностью с молекулами HLA и индуцируют более интенсивные иммунные реакции, имеющие более высокую аффинность;
d) получать вакцину, которая объединяет преимущества a) с преимуществами b) и c).
ДНК-плазмиды, соответствующим образом полученные в соответствии с изобретением, используют для профилактики и лечения людей или животных с высоким риском развития p185neu-позитивных карцином или пациентов с p185neu-позитивными первичными опухолями, рецидивами или метастазированием этих опухолей. Профилактика может быть первичной, когда опухоль еще не выражена; вторичной, когда опухоль находится на ранних стадиях, таких как предраковые патологические изменения; третичной, если наблюдается рецидив опухоли или процесс метастазирования.
Опухоли, поддающиеся лечению плазмидами или композициями по изобретению, прежде всего представляют собой опухоли эпителиального происхождения, в частности, аденокарциномы легких, яичников и молочных желез; сквамозные карциномы тканей головы и шеи, и, в более общем смысле, опухоли, экспрессирующие белок p185neu.
Подробное описание изобретения
Конструирование плазмид, кодирующих усеченные формы белка p185 neu крыс
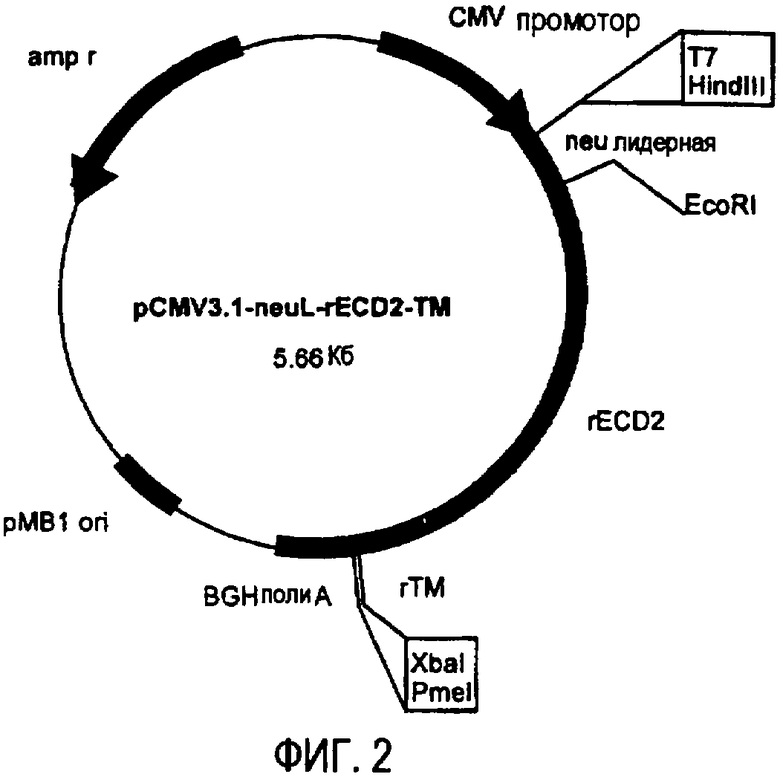
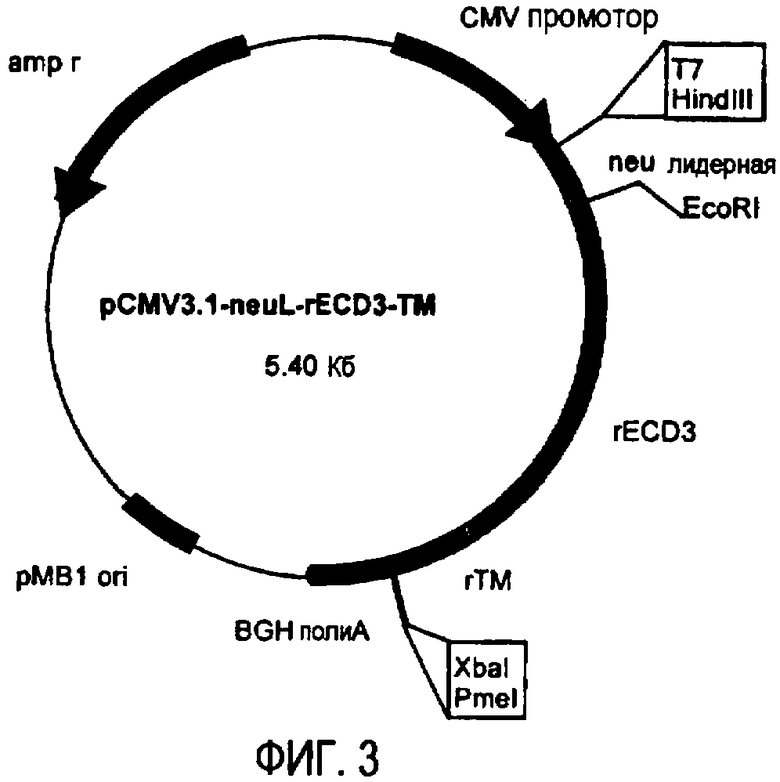
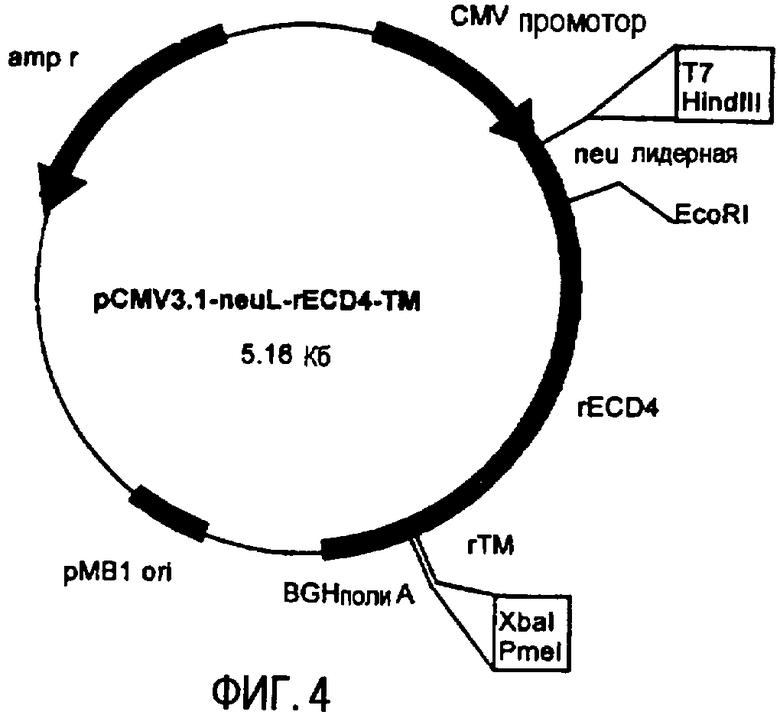
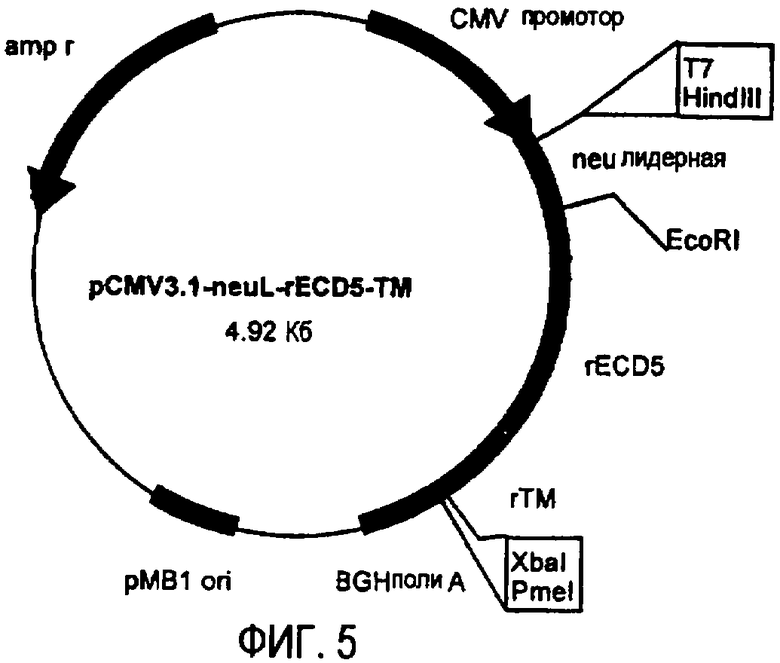
Остов плазмиды pCMV3.1 (полученной в лаборатории авторов изобретения на основе pcDNA3.1 от фирмы Invitrogen, Milan, Italy) использовали для получения ДНК плазмид, кодирующих полноразмерный ТМ домен и уменьшающиеся участки EC домена белка p185neu крысы. pCMV3.1 содержит 5' UTR нуклеотидную последовательность Her-2/neu крысы (которая транскрибирована, но не транслирована) и лидерную последовательность (neuL). Секреторный сигнальный ДНК фрагмент белка p185neu крысы получали ферментативной амплификацией ДНК с использованием вектора pCMV-EC-TM (Amici et al., 2000, Gene Ther., 7: 703; Rovero et al., 2000, J. Immunol., 165: 5133) в качестве матрицы, праймера T7 в качестве смыслового олигонуклеотида (олигонуклеотид #1) и олигонуклеотида (олигонуклеотид #2), имеющего терминальный сайт EcoRI в качестве антисмыслового олигонуклеотида. После очистки и обработки ферментами рестрикции HindIII и EcoRI, амплифицированный фрагмент клонировали в плазмиду pCMV3.1, которую расщепляли теми же ферментами, получая, таким образом, pCMV3.1-neuL. В дальнейшем семь различных последовательностей, кодирующих фрагменты EC домена и полноразмерный TM домен белка p185neu крысы, вставляли в рамку вектора pCMV3.1-neuL, расщепленного ферментами рестрикции EcoRI и XbaI. Новые плазмиды, полученные таким образом, обозначили pCMV3.1-neuL-rEC1-TM (-70 аминокислот) (фиг.1), pCMV3.1-neuL-rEC2-TM (-150 аминокислот) (фиг.2), pCMV3.1-neuL-rEC3-TM (-230 аминокислот) (фиг.3), pCMV3.1-neuL-rEC4-TM (-310 аминокислот) (фиг.4), pCMV3.1-neuL-rEC5-TM (-390 аминокислот) (фиг.5), pCMV3.1-neuL-rEC6-TM (-470 аминокислот) (фиг.6) и pCMV3.1-neuL-rEC7-TM (-550 аминокислот) (фиг.7). Фрагмент, кодируемый первой из этих плазмид, на 70 аминокислот короче, включая секреторную сигнальную аминокислотную последовательность. Все другие фрагменты прогрессивно укорачиваются на 80 аминокислот.
Эти фрагменты были получены ферментативной амплификацией ДНК с использованием семи различных смысловых олигонуклеотидов, имеющих концевой сайт рестрикции EcoRI (олигонуклеотиды #3-#9), и антисмыслового олигонуклеотида (олигонуклеотид #10), способного распознавать сайт, называемый «pcDNA3.1/BGH сайт обратного праймирования» (830-850 нуклеотидов) на 3' конце множественного сайта клонирования pCMV3.1. В качестве ДНК матрицы для ПЦР использовали вектор pCMV-EC-TM (Amici A. et al. 2000, Gene Ther. 7: 703; Rovero et al., 2000, J. Immunol. 165: 5133). После ферментативного расщепления ферментами рестрикции EcoRI и XbaI продукты амплификации клонировали в плазмиду pCMV3.1-neuL.
Вакцинация плазмидой pCMV3.1-neuL-rEC4-TM, а также вакцинация плазмидой pCMV3.I-neuL-rEC-TM, которая кодирует полноразмерные домены EC и TM, в 100% случаев защищает мышей BALB/c от развития опухолей, индуцированных инокуляцией клеток TUBO. С другой стороны, вакцинация плазмидами pCMV3.1-neuL-rEC1-TM, pCMV3.1-neuL-rEC2-TM и pCMV3.1-neuL-rEC3-TM, которые кодируют первые три усеченные формы белка p185neu, защищает 70-80% мышей BALB/c. Плазмида pCMV3.1-neuL-rEC5-TM, которая кодирует пятую усеченную форму, в 50% случаев защищает мышей BALB/c, тогда как плазмиды pCMV3.1-neuL-rEC6-TM и pCMV3.1-neuL-rEC7-TM, которые кодируют шестую и седьмую усеченные формы, не индуцируют защитную реакцию. Полученные результаты показывают, что клеточный ответ, активированный усеченными формами белка pI85neu, локализованными в цитоплазме, является достаточным для профилактики опухолей. Однако сопутствующая активация клеточного и гуморального ответа позволяет получить более эффективное лечение (Rielly et al., 2001, Cancer Res. 61: 880). Для достижения продукции антител вакцинация должна проводиться плазмидой, кодирующей полноразмерные ЕС и ТМ домены белка p185neu. Вакцинация плазмидой pCMV3.1-neuL-rEC4-TM, которая кодирует четвертую усеченную форму p185neu, лишенную аминокислот 1-310, сохраняет способность обеспечить полную защиту, но ее антительный ответ в 10 раз ниже по сравнению с антительным ответом при использовании плазмиды pCMV3.1-neuL-rEC-TM (Таблица 1).
Конструирование химерных плазмид, содержащих ДНК человека и крысы, способных кодировать семь различных слитых форм белка p185 neu (HuRT1-7)
Большинство эпитопов, представленных HLA, расположено на первом субдомене (I/L1) белка p185neu. Следовательно, для индукции специфического иммунного ответа в отношении этих эпитопов были сконструированы химерные плазмиды, кодирующие последовательности ErbB2 белка человека, которые все больше удлиняются, начиная с NH2-конца (крайняя часть EC домена). Эти новые плазмиды, обозначенные HuRT (Human Rat Transmembrane), получали путем добавления недостающих участков кДНК ErbB2 человека к последовательностям, кодирующим усеченные формы крысиного белка p185neu.
Первые пять усеченных плазмид, кодирующих полноразмерный ТМ домен и уменьшающиеся фрагменты EC домена белка p185neu крысы, обрабатывали ферментами рестрикции HindIII и EcoRI. Пять различных фрагментов кДНК человека, полученных с помощью ПЦР и расщепленных на концах, клонировали в эти пять усеченных плазмид таким образом, чтобы рамка считывания не сдвигалась. Фрагменты кДНК, кодирующие вставляемые участки белка p185neu человека, включая 5' UTR область и секретируемый сигнал для прохождения через эндоплазматический ретикулум, получали амплификацией с использованием плазмиды pcDNA3.1erbB2 в качестве матрицы. Шесть олигонуклеотидов использовали в качестве праймеров. Смысловой олигонуклеотид является одинаковым для всех шести праймеров и соответствует праймеру T7 (олигонуклеотид #1), тогда как пять антисмысловых олигонуклеотидов были расщеплены таким образом, чтобы они распознавались кДНК ErbB2 онкогена человека в постоянно удлиняющихся участках последовательности и имели EcoRI сайт рестрикции на их 3' концах (олигонуклеотиды #11-#15). После очистки и расщепления ферментами рестрикции HindIII и EcoRI амплифицированные фрагменты вставляли в соответствующие плазмиды (pCMV3.1-rEC1-TM, pCMV3.1-rEC2-TM, pCMV3.1-rEC3-TM, pCMV3.1-rEC4-TM, pCMV3.1-rEC5-TM), которые предварительно были расщеплены теми же ферментами рестрикции. Таким образом, было получено пять новых плазмид (pCMV3.1-HuRT1-5), которые кодируют химерные белки длиной 689 аминокислот, из которых 2 аминокислоты (Glu-Phe) лежат в сайте рестрикции EcoRI, используемом для соединения молекул ДНК человека и крысы. Белки, кодируемые этими химерными плазмидами, отличаются друг от друга увеличивающимися участками белка p185neu человека и уменьшающимися участками белка p185neu крысы.
Для получения химерных плазмид, кодирующих шестую и седьмую усеченные формы белка p185neu крысы, были сконструированы две новые плазмиды, в которых можно было использовать сайты клонирования, отличные от EcoRI, как, например, сайт рестрикции EcoRI, находящийся в положении 1450 генной последовательности ErbB2 человека. Плазмиду pCMV3.1 модифицировали путем использования синтетической последовательности, состоящей из смыслового олигонуклеотида (олигонуклеотид #16) и антисмыслового олигонуклеотида (олигонуклеотид #17), таким образом, чтобы один из двух сайтов рестрикции для фермента PmeI был удален, а сайты рестрикции для ферментов рестрикции HindIII и NheI, расположенные на их множественном сайте клонирования, были инвертированы. Полученный таким образом новый плазмидный остов обозначили pCMV3.1H/N. Фрагменты для шестой и седьмой усеченных форм белка p185neu крысы получали амплификацией, используя плазмиду pCMV-EC-TM (Amici et al., 2000, Gene Ther., 7: 703; Rovero et al., 2000, J, Immunol., 165: 5133) в качестве матрицы и два различных смысловых олигонуклеотида с сайтом рестрикции NheI на их концах (олигонуклеотиды #18 и #19), и антисмысловой олигонуклеотид #10.
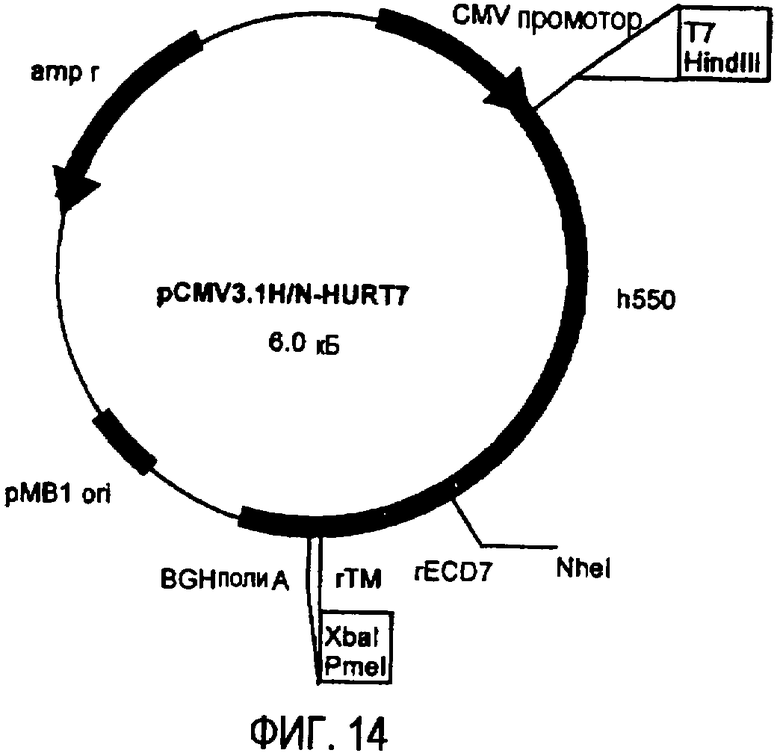
После ферментативного расщепления ферментами рестрикции NheI и PmeI продукты амплификации клонировали в плазмиду pCMV3.1H/N, получая, таким образом, новые плазмиды pCMV3.1H/N-rEC6-TM и pCMV3.1H/N-rEC7-TM. Фрагменты кДНК, кодирующие вставляемые участки белка p185neu человека, для создания химерных плазмид pCMV3.1H/N-HuRT6 и pCMV3.1H/N-HuRT7, получали амплификацией с использованием плазмиды pcDNA3.1erbB2 в качестве матрицы, праймер T7 в качестве смыслового олигонуклеотида (олигонуклеотид #1), и двух праймеров, сконструированных так, чтобы они распознавали кДНК человека в соответствующих положениях и имели на своих концах сайт рестрикции NheI (олигонуклеотиды #20 и #21), в качестве антисмысловых олигонуклеотидов.
После очистки и расщепления ферментами рестрикции HindIII и NheI амплифицированные фрагменты вставляли в соответствующие плазмиды (pCMV3.1H/N-rEC6-TM; pCMV3.1H/N-rEC7-TM), которые предварительно расщепляли теми же ферментами рестрикции. Таким образом были получены две новые химерные плазмиды pCMV3.1H/N-HuRT6 и pCMV3.1H/N-HuRT7, которые кодируют белки длиной 689 аминокислот, из которых 2 аминокислоты (Val-Ser) лежат в сайте рестрикции NheI, используемом для соединения молекул ДНК человека и крысы.
В результате этих манипуляций были получены следующие плазмиды:
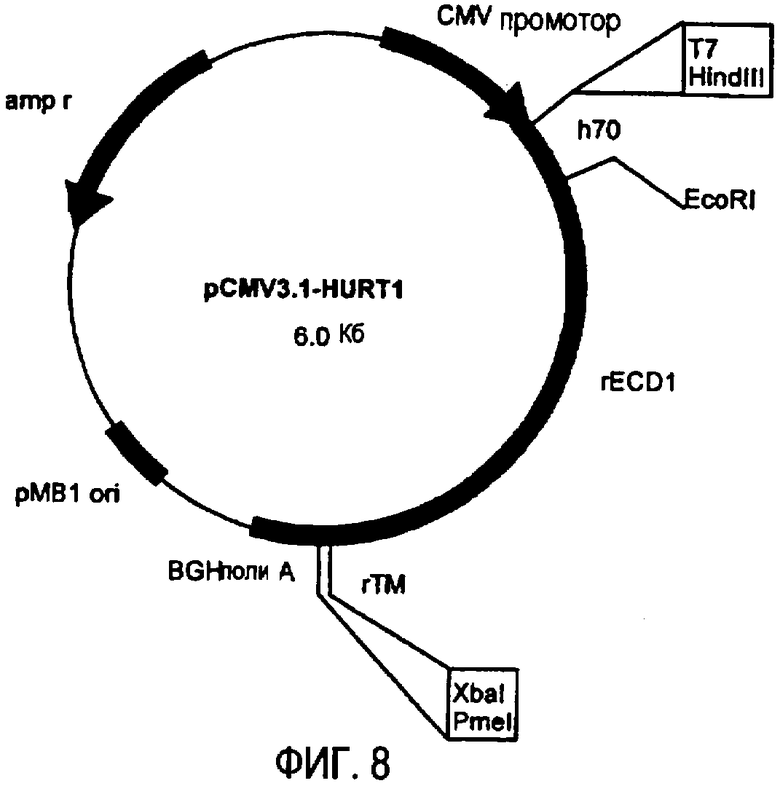
плазмида pCMV3.1-HuRT1 (фиг.8), кодирующая 70 аминокислот ЕС домена белка p185neu человека, 2 аминокислоты, принадлежащие сайту EcoRI, и 618 аминокислот белка p185neu крысы,
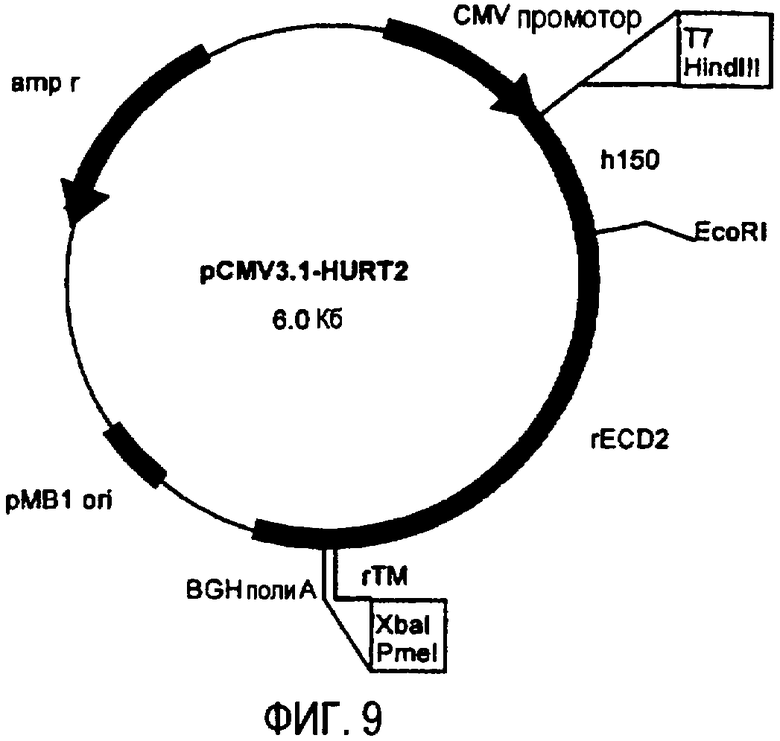
плазмида pCMV3.1-HuRT2 (фиг.9), кодирующая 150 аминокислот ЕС домена белка p185neu человека и 538 аминокислот белка p185neu крысы,
плазмида pCMV3.1-HuRT3 (фиг.10), кодирующая 230 аминокислот ЕС домена белка p185neu человека и 458 аминокислот белка p185neu крысы,
плазмида pCMV3.1-HuRT4 (фиг.11), кодирующая 310 аминокислот ЕС домена белка p185neu человека и 378 аминокислот белка p185neu крысы,
плазмида pCMV3.1-HuRT5 (фиг.12), кодирующая 390 аминокислот ЕС домена белка p185neu человека и 298 аминокислот белка p185neu крысы,
плазмида pCMV3.1H/N-HuRT6 (фиг.13), кодирующая 470 аминокислот ЕС домена белка p185neu человека и 218 аминокислот белка p185neu крысы,
плазмида pCMV3.1H/N-HuRT7 (фиг.14), кодирующая 550 аминокислот ЕС домена белка p185neu человека и 138 аминокислот белка p185neu крысы.
Непрямое доказательство мембранной экспрессии химерных белков человека и крысы, кодируемых этими плазмидами, было получено путем иммунизации мышей семью новыми плазмидами (pCMV3.1-HuRT1-5 и pCMV3.1H/N-HuRT6-7). В сыворотках всех вакцинированных мышей находятся специфические антитела к химерному белку p185neu человека и крысы. Более того, животные, вакцинированные плазмидами, кодирующими семь различных химерных белков, защищены от летального заражения клетками TUBO и/или опухолевыми клетками, гиперэкспрессирующими белок p185neu человека (D2F2-E2 клетки).
ПРИМЕРЫ
Пример 1: Конструирование плазмиды pCMV3.1-HuRT5
Плазмиду pCMV3.1-rEC5-TM, кодирующую пятую усеченную форму белка p185neu крысы, расщепляли ферментами рестрикции HindIII и EcoRI (BioLabs, Beverly, MA) для удаления области 5' UTR и последовательности neuL.
Полоса ДНК 4794 п.н., соответствующая плазмиде pCMV3.1-rEC5-TM, лишенная 5' UTR области и последовательности neuL, была разделена с помощью электрофореза в агарозном геле и элюирована с использованием набора для гель-экстракции Qiaquick (Qiagen, Italy). кДНК для 5' UTR области, лидерной последовательности и последовательности, кодирующей недостающую часть ErbB2 гена человека, получали с помощью ПЦР. pcDNA3.1ErbB2 плазмиду использовали в качестве матрицы, праймер T7 (олигонуклеотид #1) использовали в качестве смыслового олигонуклеотида, а праймер с сайтом рестрикции EcoRI на 5' конце (олигонуклеотид #15) использовали в качестве антисмыслового олигонуклеотида. Для проведения ПЦР использовали реактивы и корректирующую Taq полимеразу Finnzymes (CELBIO, Milan, Italy). После ПЦР амплифицированную ДНК очищали и осаждали стандартными способами, ресуспендировали в 50 мкл H2O и расщепляли ферментами рестрикции HindIII и EcoRI. Фрагмент кДНК, кодирующий релевантную часть ErbB2 человека, и линеаризированную плазмиду pCMV3.1-rEC5-TM клонировали реакцией лигирования, используя T4 ДНК лигазу (BioLabs, Beverly, MA).
Затем продукт лигирования использовали для трансформации DH5α штамма бактерии E.coli, компетентные клетки которого были получены с помощью хлористого кальция.
Полученные таким образом клоны анализировали щелочным лизисом для определения клонов, содержащих химерную плазмиду pCMV3.1-HuRT5.
pCMV3.1-HuRT5 затем анализировали методом секвенирования по Сенгеру, используя автоматизированный секвенатор ABI PRISM 310 Genetic Analyzer (Applied Biosystem) для подтверждения того, что вставка участка последовательности человека, кодирующей ген ErbB2, в плазмиду, кодирующую пятую усеченную форму белка p185neu крысы, была сделана правильно и без сдвига рамки считывания.
Перечень олигонуклеотидов:
#1 T7 праймер (SEQ ID No: 13)
#2 neu лидерный антисмысловой EcoRI (SEQ ID No: 14)
#3 rECD1 смысловой EcoRI (SEQ ID No: 15)
#4 rECD2 смысловой EcoRI (SEQ ID No: 16)
#5 rECD3 смысловой EcoRI (SEQ ID No: 17)
#6 rECD4 смысловой EcoRI (SEQ ID No: 18)
#7 rECD5 смысловой EcoRI (SEQ ID No: 19)
#8 rECD6 смысловой EcoRI (SEQ ID No: 20)
#9 rECD7 смысловой EcoRI (SEQ ID No: 21)
#10 pcDNA3.1/BGH реверсированный сайт прайминга (SEQ ID No: 22)
#11 His-Myc смысловой EcoRI mut (SEQ ID No: 23)
#12 His-Myc антисмысловой EcoRI (SEQ ID No: 24)
#13 70 erbB2 антисмысловой EcoRI (SEQ ID No: 25)
#14 150 erbB2 антисмысловой EcoRI (SEQ ID No: 26)
#15 230 erbB2 антисмысловой EcoRI (SEQ ID No: 27)
#16 310 erbB2 антисмысловой EcoRI (SEQ ID No: 28)
#17 390 erbB2 антисмысловой EcoRI (SEQ ID No: 29)
#18 HindΙΙΙ-NheΙ смысловой (SEQ ID No: 30)
#19 HindΙΙΙ-NheΙ антисмысловой (SEQ ID No: 31)
#20 rECD6 смысловой HheΙ (SEQ ID No: 32)
#21 rECD7 смысловой HheΙ (SEQ ID No: 33)
#22 470 erbB2 антисмысловой HheΙ (SEQ ID No: 34)
#23 550 erbB2 антисмысловой HheΙ (SEQ ID No: 35)
Пример 2: испытание in vivo
Животные
Во всех экспериментах использовали самок мышей линии BALB/c в возрасте 7 недель. Животные поступили из лаборатории Charles River Laboratories (Calco, Milan, Italy), где они были выведены в асептических условиях и в соответствии с правилами, установленными Европейским союзом.
Внутримышечное введение с последующей in vivo электропорацией
Во избежание боли и нежелательного сокращения большеберцовых мышц каждую мышь анестезировали внутрибрюшинной инъекцией 300 мкл авертина, раствор, состоящий из 0,58 г трибромэтанола (Sigma-Aldrich) и 310 мкл трет-амилового спирта (Aldrich) в 39,5 мл деионизированной H2O. Большеберцовые мышцы анестезированных мышей брили и в каждую мышцу вводили 20 мкл раствора, содержащего 25 мкг ДНК. Раствор, содержащий ДНК, готовили непосредственно перед использованием в соответствии с инструкциями Dr. F. Pericle (Valentis, Inc., The Woodlands, Texas, USA). Этот раствор содержал плазмидную ДНК в концентрации 1,25 мг/мл, поли-L-глутамат в виде натриевой соли (Sigma-Aldrich, S.r.l., Milan, Italy) в концентрации 6 мг/мл, хлорид натрия в концентрации 150 мМ (Fluka, BioChemika, Buchs, Switzerland), дистиллированную воду, свободную от эндотоксина (Nucleare Free Water, Promega Corporation) до конечного объема 1 мл. Примерно через 5 минут после инокуляции два электрических импульса, интенсивностью 375 В/см2 и продолжительностью 25 мсек каждый, генерированных с помощью устройства Electro Square Porator electroporator (T820, BTX, San Diego, CA, USA) прикладывали к обеим большеберцовым мышцам мышей, используя два стальных электрода, расположенных на расстоянии 3 мм в четырехугольном расположении латерально на голени. Генную иммунизацию с помощью электропорации проводили дважды каждому животному за 21 и 7 дней до инокуляции опухолевых клеток.
Инокуляция опухолевых клеток
На левую сторону каждой мыши инокулировали 0,2 мл суспензии, содержащей 2×105 клеток TUBO.
Оценка опухолевого роста in vivo
Рост опухоли оценивали каждую неделю путем пальпации, а размер опухоли измеряли по двум перпендикулярным диаметрам мерной линейкой. Опухолью считали новообразование размером более 1 миллиметра. Рост опухоли контролировали в течение 100 дней от инокуляции опухоли или до превышения размера опухоли 10 миллиметров в диаметре, время, в которое животных скарифицировали. Полученные результаты показали, что химерная плазмида pCMV3.1-HuRT5 обладает способностью защищать вакцинированных мышей BALB/c от летальной инокуляции клеток TUBO в 100% случаев (Таблица 2).
Оценка anti-p18S neu антител, присутствующих в сыворотке вакцинированных животных
За день до инокуляции опухолевых клеток у животных, вакцинированных химерной плазмидой pCMV3.1-HuRT5, брали кровь. Сыворотки анализировали для определения наличия крысиных антител к p185neu. Сыворотки инкубировали в течение 45 минут при 4°C с клетками, гиперэкспрессирующими p185neu крыс. После промывания раствором, называемым промывочным буфером, который состоит из фосфатно-солевого буфера (PBS), содержащего 0,2% бычьего сывороточного альбумина (BSA, Sigma, Milan, Italy) и 0,1% азида натрия (NaN3, Sigma, Milan, Italy), образцы инкубировали в течение 20 минут при 4°C с антителом к иммуноглобулину мыши, конъюгированным с ФИТЦ, промывали промывочным буфером и анализировали с помощью цитофлуориметра FACScan (Becton Dickinson Immunocytometry Systems, Mountain View, California, USA). Одновременно те же клетки инкубировали с понижающимися концентрациями моноклонального антитела к c-ErbB2/c-neu (Ab4, Oncogene) таким образом, чтобы вывести зависимость между интенсивностями флуоресценции, полученными в результате цитофлуориметрического анализа, и концентрациями анти-p185neu антител в сыворотках животных. Полученные данные показывают, что у всех вакцинированных животных высокие уровни антител к p185neu крысы и, следовательно, химерная плазмида pCMV3.1-HuRT5 являются эффективными для индукции отторжения трансплантируемых p185neu-позитивных опухолей и активации специфического антительного ответа.
| название | год | авторы | номер документа |
|---|---|---|---|
| р185-КОДИРУЮЩАЯ ДНК И ЕЕ ТЕРАПЕВТИЧЕСКИЕ ПРИМЕНЕНИЯ | 2004 |
|
RU2350652C2 |
| ICAM-4 И ЕГО ДИАГНОСТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ | 1998 |
|
RU2208230C2 |
| МОНОМОЛЕКУЛЯРНЫЙ ХИМЕРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР К РАКОВОМУ АНТИГЕНУ СА125 | 2018 |
|
RU2747095C2 |
| ВАРИАНТНАЯ ФОРМА УРАТ-ОКСИДАЗЫ И ЕЕ ИСПОЛЬЗОВАНИЕ | 2006 |
|
RU2435847C2 |
| ОЧИЩЕННЫЙ ИММУНОГЕННЫЙ БЕЛОК, ЕГО ФРАГМЕНТЫ И ПРОИЗВОДНЫЕ | 1999 |
|
RU2304585C2 |
| МОДИФИЦИРОВАННЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР ГРАНУЛОЦИТОВ ЧЕЛОВЕКА И СПОСОБ ЕГО ПОЛУЧЕНИЯ, ДНК, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР | 2000 |
|
RU2232772C2 |
| МУТАНТЫ ИНДУЦИРУЕМОЙ АКТИВАЦИЕЙ ЦИТИДИНДЕАМИНАЗЫ (AID) И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2537264C2 |
| РЕКОМБИНАНТНАЯ ГЕННАЯ КОНСТРУКЦИЯ ДЛЯ ИНДУКЦИИ ИММУННОГО ОТВЕТА, РЕКОМБИНАНТНЫЙ ПОЛИПЕПТИД И ВАКЦИНА | 2005 |
|
RU2396348C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПОЛИКЛОНАЛЬНЫХ АНТИТЕЛ, АНТИСЫВОРОТКА, СОДЕРЖАЩАЯ ПОЛИКЛОНАЛЬНЫЕ АНТИТЕЛА, И СПОСОБЫ ИММУНИЗАЦИИ | 1999 |
|
RU2358982C2 |
| АНТИГЕНЫ НЕЙССЕРИЙ | 1998 |
|
RU2347813C2 |



















Изобретение относится к биотехнологии и представляет собой плазмидный вектор для переноса ДНК, содержащий последовательность, кодирующую различные фрагменты онкобелка p185neu, способные индуцировать иммунный ответ в отношении опухолей, гиперэкспрессирующих p185neu. Изобретение относится также к фармацевтической композиции на основе вектора для профилактики или лечения пациентов с риском развития p185neu-позитивных опухолей или пациентов с первичными опухолями, метастазами или рецидивами p185neu-позитивных опухолей. Изобретение позволяет повысить эффективность профилактики или лечения пациентов с риском развития p185neu-позитивных опухолей или пациентов с первичными опухолями, метастазами или рецидивами p185neu-позитивных опухолей. 3 н. и 7 з.п. ф-лы, 14 ил., 2 табл.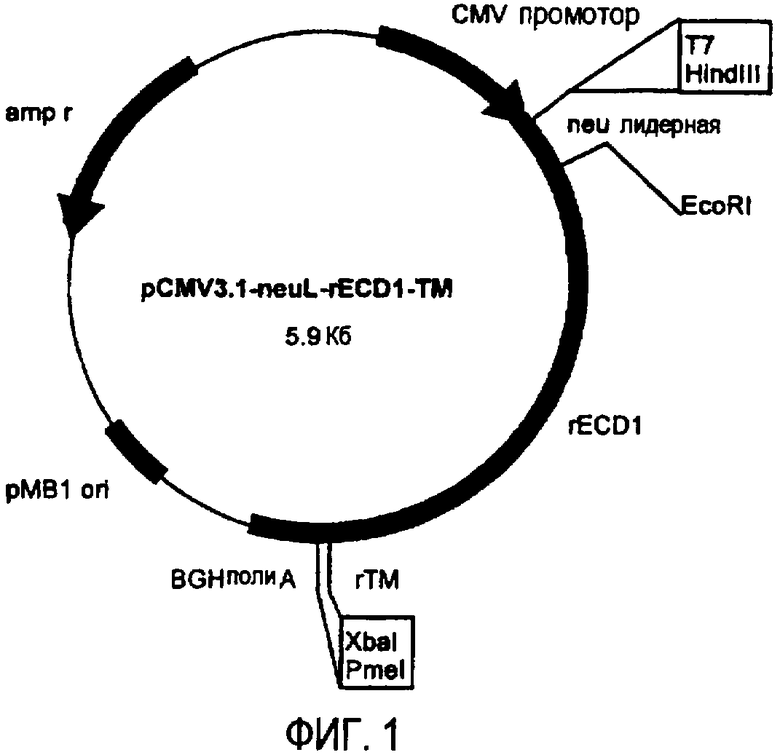
1. Плазмидный вектор для переноса ДНК, содержащий последовательность, выбранную из SEQ ID NO: 1, 2, 3, 4, 5, кодирующую фрагмент белка p185neu, или последовательность, выбранную из SEQ ID NO: 6, 7, 8, 9, 10, 11, 12, кодирующую химерный белок p185neu.
2. Плазмидный вектор по п.1, дополнительно содержащий промотор транскрипции.
3. Плазмидный вектор по п.2, где указанный промотор представляет собой CMV промотор.
4. Плазмидный вектор по пп.1 и 2, который подходит для использования у млекопитающих, в частности у людей.
5. Фармацевтическая композиция для профилактики или лечения пациентов с риском развития p185neu-позитивных опухолей, или пациентов с первичными опухолями, метастазами или рецидивами p185neu-позитивных опухолей, содержащая плазмидный вектор по пп.1-3 вместе с фармацевтически приемлемыми носителями и эксципиентами.
6. Композиция по п.5, подходящая для парентерального введения.
7. Композиция по п.6 в форме инъекционного раствора.
8. Композиция по п.5 в форме ДНК вакцины.
9. Применение плазмидного вектора по пп.1-4 для получения терапевтического агента, используемого для профилактики или лечения пациентов с риском развития p185neu-позитивных опухолей или пациентов с первичными опухолями, метастазами или рецидивами p185neu-позитивных опухолей.
10. Применение по п.9 для получения ДНК вакцины.
| WO 2004041065 А2, 21.05.2004 | |||
| US 5654406, 05.08.1997 | |||
| CHEN Y et | |||
| al | |||
| DNA vaccines encoding full-length or truncated Neu induce protective immunity against Neu-expressing mammary tumors, Cancer Res 1998, 58(9) p.1965-71 | |||
| ВЫДЕЛЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, ВЕКТОР, СПОСОБ ПОЛУЧЕНИЯ ГОМОГЕННОГО БЕЛКА gp350, ГОМОГЕННЫЙ БЕЛОК gp350, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ЕВV-СВЯЗАННОГО ЗАБОЛЕВАНИЯ ИЛИ СОСТОЯНИЯ | 1995 |
|
RU2178807C2 |

