Данное изобретение касается цис-регуляторных элементов и созданных из них химерных промоторов с активностью в растениях, индуцируемой патогенами или элиситорами. Кроме того, изобретение касается клетки-хозяина, трансгенной растительной клетки, трансгенной растительной ткани, а также трансгенного растения и его семян. Также данное изобретение относится к технологии создания трансгенного растения, в частности, стойкого к действию патогена.
Заболевания растений, вызываемые грибками, вирусами, нематодами и бактериями, приводят к большим потерям урожая во всем мире, ухудшают качество продукции и требуют использования дорогостоящих химических средств защиты растений, так как природные механизмы защиты растений, с помощью которых они способны защититься от большей части потенциальных возбудителей болезней или замедлить и ограничить их распространение, зачастую недостаточны. Для создания растений, резистентных к действию вышеназванных патогенов, можно использовать методы генной инженерии. Такие растения обладают повышенной стойкостью, так как они способны вырабатывать протеины (эффекторы), вызывающие сильную реакцию сопротивления, или молекулы, являющиеся токсичными для патогенов или подавляющие их рост и вирулентность; причем выработка происходит именно в месте заражения (контакта патогена и растения).
Эффекторами, вызывающими сильную реакцию сопротивления растения, являются, например, протеины самоактивирующихся генов устойчивости (R-генов) или гены авирулентности, которые вызывают активацию эндогенных генов устойчивости растения. К сильным реакциям сопротивления относятся гиперинтенсивная реакция (HR), контролируемая смерть клеток ткани хозяина в месте контакта, укрепление стенки клетки благодаря лигнификации и образованию каллуса, образование фитоалексинов и выработка PR-протеинов (патогенез-зависимых). Так как для этих реакций требуется большое количество энергии, и они могут привести к отмиранию (некрозу) клеток растения, их активация должно строго контролироваться. Это относится и к экспрессии токсичных для патогенов протеинов или пептидов, если конститутивная экспрессия таких протеинов или пептидов оказывает отрицательное действие на растение или его агрономические свойства, например, урожайность. В рамках трансгенных технологий обеспечить контроль можно путем использования промоторов с нужными свойствами.
Например, добиться индуцированной патогенами экспрессии таких трансгенов, как эффекторы, можно путем использования различных известных природных промоторов, индуцируемых патогенами, например, промотора PR1 (Rushton et al., 1996). В частности, разработка микроматричной технологии привела к идентификации большого количества индуцируемых патогенами промоторов (WO 03/00898, WO 02/50293 или JP 2003284566).
Однако, подобные природные индуцируемые патогенами промоторы могут демонстрировать крайне неспецифические действия, так как они могут активировать многочисленными разнообразными стимулами. Это связано с модульной структурой промотора, состоящего из нескольких разных цис-регуляторных элементов, которые объединяются самые разные сигналы в сложный экспрессионный профиль. Следовательно, природные индуцируемые патогенами промоторы отличаются и нежелательными действиями, например, в определенных тканях или сильной фоновой активностью.
Например, индуцируемые патогенами промоторы дефензинов из пшеницы являются активными во время развития и всхожести семян (Kovalchuk et al., 2010). Уже упомянутый промотор PR1 индуцируется не только патогенами, но и старением (Morris et al. 2000)
Другие исследования показали, что природный, индуцируемый ржавчиной промотор Fis1 из льна после трансформации не годился для особого регулирования экспрессии самоактивирующихся форм гена устойчивости к ржавчине L6 в Linum usitatissimum, при котором помимо стойкости к ржавчине не возникали бы отрицательные с агрономической точки зрения свойства, например, карликовость (Howies et al., 2005). Одним из способов повышения требуемой специфичности промотора является идентификация цис-регуляторных элементов, ответственных за нужную индукцию, и создание химерных промоторов из таких цис-регуляторных элементов (Venter, 2007). Напротив, мотивы, отвечающие за другие стимулы, удаляются. Промоторы многих индуцируемых патогенами генов изучены подробнее, причем были идентифицированы многие цис-регуляторные элементы, которые могут влиять на патоген-специфическую индукцию (Strittmatter et al., 1996; Eulgem et al., 2000; Kirsch et al., 2000, 2001; Himmelbach et al., 2010). Другими примерами являются цис-регуляторные элементы D-бокс, S-бокс или W-бокс (WO 00/29592), идентифицированные путем пошаговой мутации природных индуцируемых патогенами промоторов, или участки сканирования линкером LS10 и LS7 (Lebel et al., 1998). В частности, хорошо изучен W-бокс, причем его сердцевинную последовательность TTGAC(C/T) можно использовать для поиска дополнительных вариантов W-бокса в природных патоген-индуцируемых промоторах.
Кроме того, идентифицировать новые цис-регуляторные элементы можно методами биоинформатики с помощью таких программ, как MEME (Bailey and Elkan, 1994; Humphry et al., 2010) или BEST (Che et al., 2005). Одно из преимуществ при этом состоит в том, что цис-регуляторный элемент идентифицируется не как короткая отдельная последовательность, а как точно заданный мотив последовательности, с помощью которого собирается информация сразу о нескольких вариантах цис-регуляторного элемента, то есть о нескольких вариантах точки связывания транскрипционного фактора. В любом случае, такие биоинформационные подходы отличаются высокой вероятностью определения ложно-положительных последовательностей, поэтому их можно использовать только для предварительного отбора потенциальных последовательностей и мотивов. При этом абсолютно необходимо подтверждение и проверка пригодности использования в качестве цис-регуляторного элемента в целом и в качестве цис-регуляторного элемента, обеспечивающего индуцируемость патогеном, в частности. Кроме того, такие методики экспериментального анализа достаточно затратны.
Дальнейшее повышение специфичности промотора возможно путем сочетания различных цис-регуляторных элементов (Rushton et al., 2002). При этом к повышению активности (синергии) приводит не само сочетание, напротив, синергетический эффект возникает только при определенных сочетаниях, предсказать которые невозможно и нужно определять эмпирическим путем в каждом конкретном случае. Например, известны химерные промоторы с сочетаниями цис-регуляторных элементов D-бокс и S-бокс (WO 00/29592). Сила промотора и фоновая активность зависят от количества повторений элементов.
Еще одна проблема при разработке патоген-индуцируемых химерных промоторов заключается в их свойствах в различных видах растений. Хотя почти во всех исследованных видах растений удалось выявить индуцируемость патогеном, связанную с известными патоген-индуцируемыми химерными промоторами, они все же демонстрируют фоновую активность даже при отсутствии поражения патогеном. Фоновая активность колеблется в зависимости от вида растения, в котором используются химерные промоторы. То же относится к интенсивности индукции (частное от активности промотора в инфицированной и не инфицированной ткани) и к абсолютной активности промоторов (их силе). Например, из-за слишком высокой фоновой активности в не инфицированной ткани можно выявить лишь незначительную индуцируемость патогенов в инфицированной ткани.
В соответствии с современным уровнем знаний за вышеописанные колебания фоновой активности, интенсивности индукции, силы промотора, кинетики индукции и пространственного расширения активации промотора отвечают используемые цис-регуляторные элементы промотора (Rushton et al., 2002, Venter, 2007). Даже если известные химерные промоторы превосходят природные, все равно необходимо оптимизировать их, в частности, с точки зрения цис-регуляторных элементов и (или) их сочетаний. До сих пор недостаточно хорошо охарактеризованных цис-регуляторных элементов и подходящих сочетаний таких элементов, с помощью которых можно было бы конструировать химерные промоторы, которые обеспечивают точно заданную и контролируемую патоген-индуцированную экспрессию требуемых трансгенов сами по себе, а также в различных видах растений, причем экспрессия должна происходить только в результате заражения патогеном и практически исключительно в месте заражения (Gurr & Rushton, 2005). В этой связи задача настоящего изобретения заключается в создании таких новых цис-регуляторных элементов, обеспечивающих индуцируемость патогеном, и их сочетаний.
Ниже приводится более подробное описание некоторых терминов, используемых в данной заявке.
Под "элиситором" в данном изобретении понимается индуктор или медиатор, способствующий мерам защиты от растительных патогенов, например, синтезу фитоалексинов. Элиситоры могут быть либо эндогенного или экзогенного происхождения. Предпочтительно, чтобы (экзогенный) элиситор происходил из патогена и распознавался растением. К ним также относятся ПАМП (патоген-ассоциированные молекулярные паттерны), например, флагеллин, РЕР25 и хитин. Элиситоры можно использовать, чтобы имитировать заражение или контакт с патогеном, при котором элиситор вносится искусственно, в отсутствие патогена. В связи с данным изобретением элиситоры используются, в частности, для проверки индуцируемости промоторов.
"Одиночная последовательность" - последовательность нуклеотидов или оснований (пар оснований), причем каждая позиция в одиночной последовательности определяется только одним четко заданным основанием (а, с, g или t). Одиночная последовательность изолируется из природного промотора и является результатом биоинформационного анализа. Она состоит из сердцевинной последовательности и боковых частей последовательности. Понятие "одиночная последовательность" означает также молекулу нуклеиновой кислоты, последовательность нуклеотидов или (пар) оснований соответствует одиночной последовательности.
"Сердцевинная последовательность" - это последовательность нуклеотидов или (пар) оснований в определенным участке цис-регуляторного элемента, причем данный участок отвечает за функциональность цис-регуляторного элемента. Сердцевинная последовательность является частью одиночной. Понятие "сердцевинная последовательность" означает также молекулу нуклеиновой кислоты, последовательность нуклеотидов или (пар) оснований соответствует сердцевинной последовательности.
"Промотор" - не транслированная последовательность ДНК, обычно ориентированная против направления кодирующего участка, содержащая точку связи с РНК-полимеразой и инициирующая транскрипцию ДНК. Кроме того, в промоторе часто есть другие элементы, являющиеся регулятора экспрессии гена (например, цис-регуляторные элементы).
"Минимальный промотор" - промотор, имеющий только базовые элементы, необходимые для инициации транскрипции (например, ТАТА-бокс и (или) инициатор). "Химерным промотором" называется промотор, не встречающийся в природе и состоящий из нескольких элементов. Он состоит из минимального промотора и по направлению вверх по меньшей мере один цис-регуляторный элемент, который представляет собой точку связи для транс-активных факторов (trans-acting factors, например, факторов транскрипции). Химерный промотор создается в соответствии с требованиями и индуцируется или тормозится различными факторами. Выбор цис-регуляторного элемента или сочетаний таких элементов имеет решающее значение, например, для специфичности или уровня активности промотора. Цис-регуляторный элемент в химерном промоторе либо гетерологичен используемому минимальному промотору, т.е. элемент и используемый минимальный промотор происходят из разных организмов или видов (например, как показано на рис. 15А-С), либо гомологичен ему, т.е. элемент и используемый минимальный промотор встречаются в одном природном промоторе, однако цис-регуляторный элемент сам по себе или как дополнительный элемент в химерном промоторе и природный промотор локализованы в разном генетическом окружении. Под химерным промотором также понимается (природный) промотор, который был изменен мультимеризацией по меньшей мере одного цис-регуляторного элемента (например, как показано на рис. 15D).
"Комплементарная" последовательность нуклеотидов относительно двуспиральной ДНК означает, что во второй цепочке ДНК, комплементарной первой цепочке, в соответствии с правилами сочетания оснований и с учетом ориентации есть нуклеотидные основания, соответствующие основаниям первой цепочки (пример: 5'-gcat-3' комплементарен 5'-atgc-3').
"Патоген" - организм, который при взаимодействии с растением приводит к возникновению симптомов заболевания одного или нескольких органов этого растения. К таким патогенам относятся например, животные, грибковые, бактериальные или вирусные организмы и оомицеты.
Под "заражением патогеном" понимается самый ранний момент времени, в который обмен веществ патогена подготавливается к проникновению в ткани растения-хозяина. Например, у грибов или оомицетов к этому относится прорастание гифы или образование специальных структур, например, гифы для пенетрации и апрессории.
Под "патоген-/элиситор-индуцируемостью" в контексте изобретения понимается особое свойство промотора, который после заражения патогеном или применения элиситора приводит к усиленной по меньшей мере в два раза транскрипции оперативно связанного гена. Кроме того, под "патоген-/элиситор-индуцируемостью" в контексте изобретения понимается свойство генов, которые после заражения патогеном или применения элиситора транскрибируются с по меньшей мере двойным усилением.
Согласно изобретению поставленная задача решается путем создания новых цис-регуляторных элементов, способствующих индуцируемости патогеном и (или) элиситором. Они значительно отличаются от уже известных элементов, в частности, в пределах сердцевинной последовательности, и не представляют собой их вариации. Таким образом, цис-регуляторные элементы согласно изобретению должны представлять собой точки распознавания и (или) связи для новых факторов транскрипции, следовательно, индуцируемость патогеном и (или) элиситором является новой характеристикой. Цис-регуляторные элементы согласно изобретению были идентифицированы биоинформационными методами в промоторах патоген- или РАМР- (элиситор)-индуцированных генов Arabidopsis thaliana. В результате различных этапов анализа изолированные одиночные последовательности цис-регуляторных элементов удалось соотнести с восемью группами мотивов (группы 1, 5, 11, 12, 18, 21, 27 и 32). Кроме того, несколько изолированных одиночных последовательностей удалось соотнести с группой мотивов 21n. К одной группе мотивов относятся те одиночные последовательности, которые при сравнении идентифицированных мотивов обнаруживают высокую степень консервативности. Таким образом, все одиночные последовательности одной группы мотивов совпадают в одном характерном мотиве сердцевинной последовательности. Следовательно, поставленная задача решается с помощью изолированного цис-регуляторного элемента, включающего молекулу нуклеиновой кислоты, нуклеотидная последовательность которой соответствует одному из мотивов сердцевинной последовательности из:
a) vaaagtm,
b) aaacca,
c) scaaam,
d) acrcg,
e) sktgkact,
f) mrtsack,
g) ccaccaa,
h) tcgtctcttc или
i) wwkgwc
Менее консервативные позиции основании в характерных мотивах сердцевинной последовательности а) - i): Υ' означает гуанин (g) или аденин (а), то есть пуриновое основание, 'k' означает гуанин (g) или тимин (t)урацил (u), 's' означает гуанин (g) или цитозин (с), 'm' означает аденин (а) или цитозин (с) и 'w' означает аденин (а) или тимин (t)/урацил (u). Определенный мотив сердцевинной последовательности воспроизводит по меньшей мере одну частичную последовательность сердцевинной последовательности каждой одиночной последовательности группы мотивов, относящейся к мотиву сердцевинной последовательности, причем эта частичная последовательность может составлять минимум 30% общей сердцевинной последовательности одной одиночной последовательности. В группах мотивов сердцевинной последовательности g) и h) мотив сердцевинной последовательности соответствует всей сердцевинной последовательности одиночных последовательностей. Изобретение также включает изолированный цис-регуляторный элемент, включающий молекулу нуклеиновой кислоты, нуклеотидная последовательность которой соответствует одному мотиву сердцевинной последовательности, комплементарному а) - i). Характерный мотив сердцевинной последовательности определенной группы мотивов может также неоднократно встречаться в сердцевинной последовательности определенной одиночной последовательности, причем мотивы сердцевинной последовательности могут перекрываться в ней и (или) обладать разной ориентацией. Например, сердцевинная последовательность Cis09 из группы мотивов 1, с одной стороны, имеет частичную последовательность, соответствующую мотиву сердцевинной последовательности acrcg, с другой стороны, - перекрывающуюся на два основания частичную последовательность, которая соответствует комплементарному мотиву сердцевинной последовательности acrcg, и именно cgygt. Другим примером является сердцевинная последовательность 21G-2_M1_S2 из группы мотивов 11, где сердцевинная последовательность имеет две частичные последовательности, которые перекрываются одним основанием и соответствуют комплементарному мотиву сердцевинной последовательности ааасса, а именно tggttt.
Для групп мотивов 1, 5, 11, 12, 21, 21 η и 27 на основании экспериментальных данных о функциональности удалось определить семейный мотив, включающий характерный мотив сердцевинной последовательности. Семейный мотив является производным отличительным признаком фактора транскрипции или семейства факторов транскрипции. Преимущество семейного мотива состоит в том, что он объединяет возможные варианты области распознавания/связывания. Предпочтительно, чтобы семейный мотив определенной группы мотивов объединял все сердцевинные последовательности одиночных последовательностей этой группы. С этой целью для части одиночных последовательностей учитывалась комплементарная цепочка цис-регуляторного элемента; в дальнейшем такие комплементарные последовательности идентифицированных цис-регуляторных последовательностей обозначаются '(inv) ', если это необходимо для понимания. Определенный таким образом семейный мотив имеет длину минимум 15 нуклеотидов, предпочтительно минимум 13 нуклеотидов, особенно предпочтительно минимум 11 нуклеотидов. Кроме соответственного мотива сердцевинной последовательности семейный мотив имеет боковые части, которые при использовании изобретенного цис-регуляторного элемента в химерном промоторе оказывают значительное количественное воздействие на его свойства, например, фоновую активность и силу экспрессии. При увеличении расстояния между определенным основанием боковых частей и сердцевинной последовательностью в одиночной последовательности количественное воздействие ослабевает. Для определения семейного мотива можно дополнительно учитывать и другие высококонсервативные отдельные основания, выходящие за пределы сердцевинных последовательностей одиночных последовательностей и расширяющиеся семейный мотив. Семейные мотивы групп мотивов 1, 5,11, 12, 21, 21 η и 27 перечислены в столбце 2 таблицы 1. Таким образом, изобретение включает и изолированный цис-регуляторный элемент, включающий молекулу нуклеиновой кислоты, нуклеотидная последовательность которой
a) соответствует семейному мотиву согласно SEQ ID NO: 1, включающему мотив сердцевинной последовательности vaaagtm,
b) соответствует семейному мотиву согласно SEQ ID NO: 2, включающему мотив сердцевинной последовательности ааасса,
c) соответствует семейному мотиву согласно SEQ ID NO: 3, включающему мотив сердцевинной последовательности scaaam,
d) соответствует семейному мотиву согласно SEQ ID NO: 4, включающему мотив сердцевинной последовательности acrcg,
e) соответствует семейному мотиву согласно SEQ ID NO: 5, включающему мотив сердцевинной последовательности sktgkact,
f) соответствует семейному мотиву согласно SEQ ID NO: 6, включающему мотив сердцевинной последовательности mrtsack,
g) соответствует семейному мотиву согласно SEQ ID NO: 41, включающему мотив сердцевинной последовательности wwkgwc,
или нуклеотидная последовательность которой соответствует определенному семейному мотиву, комплементарному а) - д).
Кроме того, семейный мотив может быть и короче. В самой короткой форме он определяется как минимальный семейный мотив, который после упорядочения одиночных последовательностей в соответствии с общим мотивом сердцевинной последовательности включает только те позиции оснований, который встречаются в сердцевинных последовательностях всех одиночных последовательностей одной группы мотивов. В некоторых случаях минимальный семейный мотив соответствует мотиву сердцевинной последовательности. В столбце 2 таблицы 1 (заголовок "Семейный мотив") подчеркнуты минимальные семейные мотивы групп мотивов 1, 5, 11, 12, 21, 21n и 27.
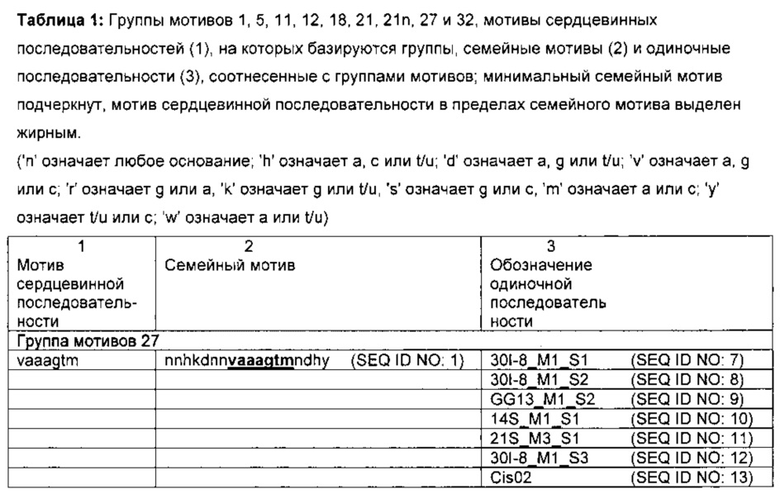
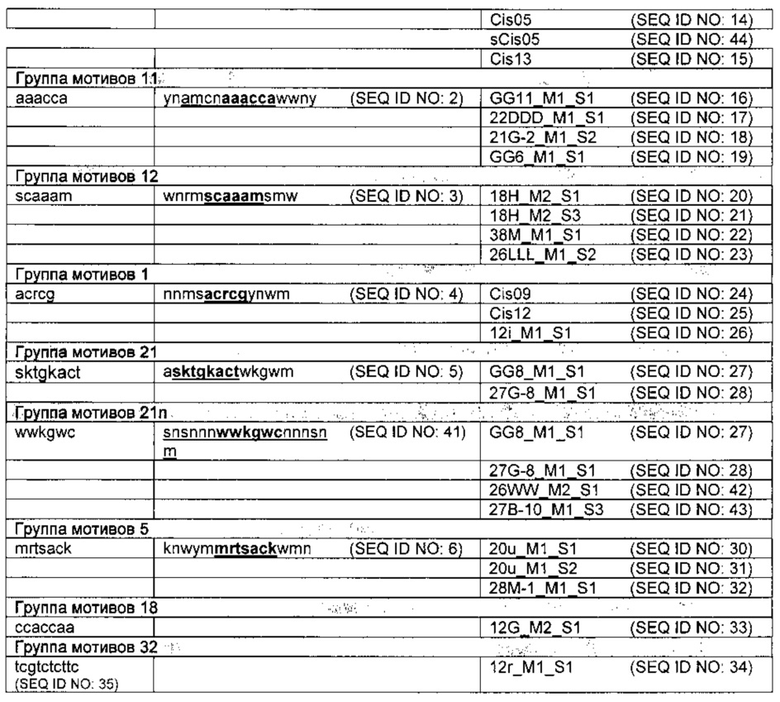
Изобретение также касается всех идентифицированных и изолированных одиночных последовательностей цис-регуляторных элементов согласно изобретению и их сердцевинных последовательностей (таблицы 1 и 2). Таким образом, задача решается за счет изолированного цис-регуляторного элемента, включающего
a) молекулу нуклеиновой кислоты с последовательностью нуклеотидов согласно SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29, SEQ ID NO: 30, SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34, SEQ ID NO: 42, SEQ ID NO: 43 или SEQ ID NO: 44,
b) молекулу нуклеиновой кислоты с последовательностью нуклеотидов согласно сердцевинной последовательности из SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29, SEQ ID NO: 30, SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34, SEQ ID NO: 42, SEQ ID NO: 43 или SEQ ID NO: 44, или
с) молекулу нуклеиновой кислоты с последовательностью нуклеотидов, комплементарной одной из последовательностей нуклеотидов из а) или b). Цис-регуляторный элемент, включающий молекулу нуклеиновой кислоты с последовательностью нуклеотидов согласно SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26 относится к группе мотивов 1, элемент с последовательностью нуклеотидов согласно SEQ ID NO: 30, SEQ ID NO: 31 или SEQ ID NO: 32 к группе мотивов 5, элемент с последовательностью нуклеотидов согласно SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18 или SEQ ID NO: 19 к группе мотивов 11, элемент с последовательностью нуклеотидов согласно SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22 или SEQ ID NO: 23 к группе мотивов 12, элемент с последовательностью нуклеотидов согласно SEQ ID NO: 33 к группе мотивов 18, элемент с последовательностью нуклеотидов согласно SEQ ID NO: 27 или SEQ ID NO: 28 к группе мотивов 21, элемент с последовательностью нуклеотидов согласно SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 42 или SEQ ID NO: 43 к группе мотивов 21n, элемент с последовательностью нуклеотидов согласно SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 44 или SEQ ID NO: 15 к группе мотивов 27 и элемент с последовательностью нуклеотидов согласно SEQ ID NO: 34 - к группе мотивов 32.
Изобретенный цис-регуляторный элемент может иметь длину менее 50 нуклеотидов, предпочтительно менее 40 нуклеотидов и особенно предпочтительно менее 30 нуклеотидов. Сердцевинная последовательность изобретенного цис-регуляторного элемента может иметь длину менее 20 нуклеотидов, предпочтительно менее 15 нуклеотидов, особенно предпочтительно менее 10 нуклеотидов, однако она не должна быть короче 6 нуклеотидов.
Некоторые одиночные последовательности цис-регуляторных элементов согласно изобретению могут помимо идентифицированной новой сердцевинной последовательности содержать и сердцевинную последовательность известного цис-регуляторного элемента, способствующего патоген-/элиситор-индуцируемости, что при определенных обстоятельствах может влиять и на специфичность новой идентифицированной сердцевинной последовательности и (или) идентифицированной одиночной последовательности. Одним из примеров является цис-регуляторный элемент Cis05 (SEQ ID NO: 14), которые помимо идентифицированной сердцевинной последовательности между позициями нуклеотидов 14 и 20 содержит также сердцевинную последовательность W-бокса между позициями нуклеотидов 29 и 35. Для этой цели были проведены подробные анализы мутаций (см. рис. 5А, 5В и 11). Цис-регуляторный элемент согласно изобретению можно использовать в химерном промоторе, причем данный элемент придает химерному промотору специфическую патоген- и (или) элиситор-индуцируемость. Таким образом, данное изобретение включает химерный промотор, который подходит для того, чтобы под действием заражения патогеном или обработки патогенным элиситором воздействовать на экспрессию оперативно связанной молекулы нужной нуклеиновой кислоты, например, гетерологической последовательности ДНК, в растительной клетке, и который состоит из минимального промотора и по меньшей мере одного цис-регуляторного элемента согласно изобретению. Один изобретенный цис-регуляторный элемент в таком химерном промоторе уже может содействовать значительной патоген- и (или) элиситор- индуцируемости. Таким образом, этого цис-регуляторного элемента уже достаточно для конструирования патоген-/элиситор-реактивного химерного промотора в сочетании с минимальным промотором.
Предпочтительно, чтобы это был такой химерный промотор, содержащий только один или несколько цис-регуляторных элементов согласно изобретению, который реагировал бы только на патоген и (или) элиситор, то есть этот промотор не должен индуцироваться (или должен индуцироваться лишь в небольшой степени) другими стимулами, например, абиотическим стрессом. Индукция такого химерного промотора, включающего один или несколько цис-регуляторных элементов согласно изобретению, после контакта с патогеном/элиситором минимум в 2 раза, предпочтительно в 10 раз, особенно предпочтительно минимум в 25 раз сильнее индукции без контакта с патогеном/элиситором (фоновая активность).
В предпочтительном варианте индуцированная экспрессия ограничивается местом заражения, то есть в аналогичной или меньшей степени по сравнению с контролируемой экспрессией природных PR-генов. Особенно предпочтительно, чтобы контролируемая активация транскрипции химерным промотором согласно изобретению происходила только в тех клетках, которые контактируют с патогеном или патогенным элиситором. Однако в результате взаимодействия клеток транскрипция может активироваться и в тех клетках, которые окружают место или места заражения. Однако химерные промоторы согласно данному изобретению не ограничиваются исключительно патоген-реактивными. При сочетании с другими регуляторными элементами индуцированную экспрессию можно задать более точно, например, путем комбинирования с цис-регуляторным элементом, который способствует, например, определенной специфичности ткани, индуцируемости при хранении, холоде или высокой температуре, а также специфической активности на определенных этапах развития. Химерные промоторы согласно изобретению могут включать и по меньшей мере одну комбинацию из минимум двух цис-регуляторных элементов, причем данная (минимум одна) комбинация включает по меньшей мере один цис-регуляторный элемент согласно изобретению. Другими цис-регуляторными элементами в комбинации для создания химерного промотора могут быть и известные элементы, способствующие патоген-/элиситор-индукции, например, W-бокс, S-бокс или D-бокс (см. WO 00/29592).
Кроме того, изобретение включает химерный промотор, который содержит один или несколько мономеров и (или) один или несколько мультимеров цис-регуляторных элементов согласно изобретению. Предпочтительными формами являются димеры и тетрамеры Мономеры сами по себе или отдельные мономеры в мультимере могут иметь разные ориентации, т.е. они могут быть расположены, например, комплементарно. Мультимерные цис-регуляторные элементы согласно изобретению могут быть функционально связаны, то есть в мультимерной форме они демонстрируют синергетическое или антагонистическое действие, например, на связывающую способность фактора транскрипции, который, в том числе, распознает характерный мотив сердцевинной последовательности определенной группы мотивов. Изобретение также включает химерный промотор, который подходит для того, чтобы под действием заражения патогеном или обработки патогенным элиситором воздействовать на экспрессию оперативно связанной молекулы нужной нуклеиновой кислоты в растительной клетке, и который состоит из минимального промотора и по меньшей мере двух цис-регуляторных элементов согласно изобретению, причем минимум два цис-регуляторных элемента согласно изобретению могут быть функционально связаны в гомо- и (или) гетеромерной форме. Индукция химерного промотора, включающего минимум один мультимер цис-регуляторных элементов согласно изобретению, после контакта с патогеном/элиситором минимум в 2 раза, предпочтительно в 10 раз, особенно предпочтительно минимум в 25 раз сильнее индукции без контакта с патогеном/элиситором (фоновая активность).
В предпочтительном варианте химерных промоторов согласно изобретению расстояние от минимального промотора до первого в направлении вверх цис-регуляторного элемента согласно изобретению составляет от 0 до 300 пар оснований, предпочтительно от 0 до 70 пар, особенно предпочтительно менее 10 пар. Дополнительно или альтернативно расстояние между двумя одинаковыми мономерами цис-регуляторных элементов согласно изобретению в мультимерной форме составляет от 0 до 10 пар оснований. Предпочтительно, чтобы два отдельных мультимера в химерном промоторе согласно изобретению разделялись примерно 0-50 парами оснований.
Специфические комбинации цис-регуляторных элементов согласно изобретению с другими цис-регуляторными элементами согласно изобретению или с другими известными регуляторными элементами или фрагментами, например, S-боксом, D-боксом или Gst1 по результатам экспериментальных анализов неожиданно продемонстрировали воздействие на свойства промоторов, например, на невысокую фоновую активность или особо специфическую либо особо сильную индуцируемость (до 183-кратной 2xCis13-2xCis05). При этом некоторые комбинации демонстрируют синергетическое действие на определенное свойство промотора, например, на фактор индукции (см. например 2xCis13-2xC¡s05 в петрушке, рис. 12 и 13), а другие - антагонистическое действие на фактор индукции, но особенно низкую фоновую активность (например, 4xCis05-2xD в сахарной свекле). Кроме того, изобретение включает все возможные сочетания цис-регуляторных элементов согласно изобретению между собой и с известными цис-регуляторными элементами, оказывающие предпочтительное синергетическое или антагонистическое действие на индуцируемость промотора. Предпочтительными комбинациями являются те, которые можно выбрать из следующей группы: 4х sCis05, 4х 20u_M1_S1, 4х 27G-8_M1_S1, 4х 38M_M1_S1, 4х 18H_M2_S3, 4х 18H_M2_S1, 4х GG13_M1_S2, 4х 21S_M3_S1, 4х 30I-8_M1_S2, 2х Cis02 - 2х Cis02, 2х Cis02 - 2х Cis05, 2х Cis02 - 2х Cis12, 2х Cis02 - 2х C¡s13, 2х Cis02 - 2х D, 2х Cis02 - 2х S, 2х Cis02 - Gst1, 2х Cis02 - 2х 30I-8_M1_S2, 2х Cis05 - 2х Cis02, 2х Cis05 - 2х Cis05, 2х Cis05 - 2х Cis12, 2х C¡s05 - 2х Cis13, 2х Cis05 -2х D, 2х Cis05 - 2х S, 2х C¡s05 - Gst1, 2x Cis05 - 2x 30I-8_M1_S2, 2x Cis12 - 2x Cis02, 2x C¡s12-2x C¡s05, 2x Cis12-2x Cis12, 2x C¡s12-2x Cis13, 2x Cis12-2x D, 2x Cis12-2x S, 2x Cis12 - Gst1, 2x Cis12 - 2x 30I-8_M1_S2, 2x Cis13 - 2x Cis02, 2x Cis13 - 2x Cis05, 2x Cis13-2x Cis12, 2x Cis13-2x Cis13, 2x Cis13-2x D, 2x Cis13-2xS, 2x C¡s13-Gst1, 2x C¡s13 - 2x 30I-8_M1_S2, 2x D - 2x Cis02, 2x D - 2x C¡s05, 2x D - 2x Cis12, 2x D -2x Cis13, 2x D - 2x 30I-8_M1_S2, 2x S - 2x Cis02, 2x S - 2x Cis05, 2x S - 2x Cis12, 2x S -2x Cis13, 2x S - 2x 30I-8_M1_S2, Gst1 - 2x Cis02, Gst1 - 2x Cis05, Gst1 - 2x Cis12, Gst1 -2x Cis13, Gst1 - 2x 30I-8_M1_S2, 2x 30I-8_M1_S2 - 2x Cis02, 2x 30I-8_M1_S2 - 2x C¡s05, 2x 30I-8_M1_S2 - 2x Cis12, 2x 30I-8_M1_S2 - 2x Cis13, 2x 30I-8_M1_S2 - 2x D, 2x 30I-8_M1_S2 - 2x S, 2x 30I-8_M1_S2 - Gst1 и 2x 30I-8_M1_S2 - 2x 30I-8_M1_S2. Особенно предпочтительные комбинации с неожиданным воздействием на фактор индукции и активность приведены в таблице 3.
Данное изобретение также относится к химерным промоторам, которые включают по меньшей мере одну из вышеназванных комбинаций цис-регуляторных элементов.
В предпочтительном варианте химерных промоторов согласно изобретению минимальный промотор происходит, например, из промотора CaMV35S, для однодольных растений, например, из промотора пшеницы TaPal (SEQ ID NO: 39), промотора кукурузы ZmUbiquitin (SEQ ID NO: 40) или промотора риса OsGnsI (SEQ ID NO: 38), для двудольных растений из известных минимальных промоторов (WO 07/147395). Кроме того, для конструирования химерного промотора согласно изобретению можно использовать и минимальные промоторы из других источников.
В любом случае, химерный промотор согласно изобретению соответствует основным требованиям, которые предъявляются к строгому регулированию экспрессии трансгена в генной инженерии, например, при создании растения, резистентного к определенному патогену/заболеванию. Трансген - это оперативно связанная с химерным промотором молекула нуклеиновой кислоты, например, гетерологическая последовательность ДНК, кодирующая для гена резистентности (R-гена) автоактивируемый ген резистентности, ген авирулентности, другой эффектор, протеин, токсичный по отношению к по меньшей мере одному патогену, компонент передачи сигнала, протеин, участвующий в синтезе фитоалексинов, двуцепочечную РНК для образования направленной против патогена siPHK или антимикробный пептид. Кроме того, многочисленные опыты с петрушкой (Petroselinum erispum), Arabidopsis thaliana, пшеницей (Triticum sp.) и сахарной свеклой (Beta vulgaris) показали, что химерный промотор согласно изобретению функционирует с разными видами растений (см., например, 4xCis05 в петрушке, сахарной свекле и пшенице).
Определение семейного мотива для каждой группы мотивов 1, 5, 11, 12, 21, 21n и 27 открывает перед специалистом новые возможности конструирования химерных промоторов согласно данному изобретению. Наблюдаемая активность разных членов групп мотивов 27 и 12 (рис. 8) показывает, что боковые участки последовательности можно использовать для точной настройки нужных уровней экспрессии. Это относится и к настройке уровней экспрессии в зависимости от вида растения. Семейный мотив воспроизводит возможности варьирования отдельных оснований в боковых областях, показывая специалисту, насколько можно модифицировать эти области. Кроме того, на основании семейного мотива специалист получает информацию о том, насколько консервативны отдельные позиции оснований в пределах этого мотива. При этом следует исходить из того, что модификация сильно консервативного основания влияет на конечные свойства химерного промотора сильнее, чем слабо консервативное основание.
Кроме того, изобретение касается рекомбинантного гена, включающего химерный промотор согласно изобретению. Предпочтительно, чтобы рекомбинантный ген был таким, чтобы химерный промотор был оперативно связан с молекулой нуклеиновой кислоты, например, с гетерологической последовательностью ДНК. Такая гетерологическая последовательность ДНК кодирует, в частности, (поли)пептид, цитотоксичный протеин (например, Bt-токсин, авирулентный протеин или такие энзимы, как глюкозооксидазы, вырабатывающие реактивные формы кислорода) антитело, антисмысловую РНК, смысловую РНК, фактор транскрипции, протеазу, нуклеазу, липазу, ингибитор энзимов или измеримый маркер (например, люциферазу, GFP или β-галактосидазу). Последние маркеры, а также маркеры, известные на данный момент можно использовать в тестах, чтобы определить патогенную специфичность химерного промотора согласно изобретению или идентифицировать эффекторы, способствующие индукции химерного промотора или тормозящие ее. Химерные промоторы согласно изобретению можно также использовать в технологиях подавления экспрессии генов на базе PHKi, причем оперативно связанная молекула нуклеиновой кислоты кодирует, например, антисмысловую РНК, смысловую РНК или двуцепочечную РНК (dsPHK). Молекула РНК может представлять собой короткую нуклеотидную последовательность (минимум 10 нуклеотидов, предпочтительно минимум 14 нуклеотидов, в некоторых вариантах до 100 нуклеотидов и более), в основном комплементарную специфической последовательности тРНК и (или) последовательности ДНК соответствующего гена. Стандартные методы технологии PHKi описаны в известных работах.
В принципе оперативно связанную кодирующую последовательность можно модифицировать так, чтобы продукт трансляции был локализован в нужной части клетки, например ядре, эндоплазматической сети, митохондрии, цитоплазме или вакуоли либо вне клетки (апопластически). Специалистам известны подходящие методы модификации (Gorlich, Science 271 (1996), 1513-1518; Hicks, Plant Physiol. 107 (1995), 1055-1058; Rachubinski, Cell 83 (1995), 525-528; Schatz, Science 271 (1996), 1519-1526; Schnell, Cell 83 (1995), 521-524; Verner, Science 241 (1988), 1307-1313; Vitale, BioEssays 14 (1992), 151-160).
Рекомбинантный ген согласно изобретению можно использовать как сам по себе, так и как часть вектора. Таким образом, данное изобретение касается и вектора, включающего химерный промотор согласно изобретению или рекомбинантный ген согласно изобретению. Предпочтительно, чтобы это был растительный вектор экспрессии, который должен предпочтительно включать и маркер селекции для растений. Примеры подходящих маркеров были приведены выше. Методы конструирования таких векторов известны специалистам и описаны, например, в Sambrook, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (1989) N.Y. and Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.V. (1989).
Настоящее изобретение также касается прокариотической или эукариотической клетки-хозяина, включающей химерный промотор, рекомбинантный ген или вектор согласно изобретению, причем химерный промотор сам по себе или как часть рекомбинантного гена либо вектора, а также часть химерного промотора как цис-регуляторный элемент является гетерологичной по отношению к прокариотической или эукариотической клетке-хозяину, например, происходит из клетки или организма с другим генетическим фоном, либо гомологической по отношению к прокариотической или эукариотической клетке-хозяину, однако в этом случае он должен быть локализован в другом генетическом окружении, отличаясь таким образом от природного химерного промотора или его части.
Химерный промотор, рекомбинантный ген или вектор согласно изобретению могут быть либо интегрированы в геном прокариотической или эукариотической клетки-хозяина (предпочтительно стабильно интегрированы), либо находиться в клетке в экстрахромосомной форме, например, как плазмид.
Кроме того, изобретение касается метода создания трансгенного растения, включающего внесение химерного промотора, рекомбинантного гена или вектора согласно изобретению в по меньшей мере одну клетку растения или внесение химерного промотора, рекомбинантного гена или вектора согласно изобретению в по меньшей мере одну растительную клетку клеточной культуры, из которой затем восстанавливается трансформированное или трансгенное растение. Предпочтительно, чтобы химерный промотор, рекомбинантный ген или вектор были интегрированы в геном растения, особенно предпочтительно - стабильно интегрированы. Для экспрессии требуемой молекулы нуклеиновой кислоты в растительных клетках под контролем химерного промотора согласно изобретению молекулу можно соединять с другими регуляторными последовательностями, например, с полиаденильным хвостом на 3'-конце. Методы внесения генов или генетического материала в растение или растительную клетку, а также методы восстановления трансформированной растительной клетки известны, например, трансформация растительных клеток или тканей с участием Agrobacteríum tumefaciens или Agrobacterium rhizogenes с помощью Т-ДНК, слияние протопластов, инъекция, электропорация, вакуумная инфильтрация или биолистические методы. Известны также методы подготовки подходящих векторов для внесения генов или генетического материала в растение или растительную клетку (Sambrook, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (1989) N.Y. and Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.V. (1989)).
В альтернативном варианте растительную клетку можно модифицировать так, чтобы она вырабатывала эндогенный ген под контролем химерного промотора согласно изобретению или под контролем нативного, модифицированного цис-регуляторными элементами согласно изобретению промотора эндогенного гена. Внесение подобного химерного промотора, который в естественном состоянии не регулирует экспрессию определенного гена или определенной геномной последовательности, в нужном месте генома растения или внесение цис-регуляторных элементов согласно изобретению в нативный промотор, может осуществляться стандартными методами, например, путем целенаправленной интеграции ('gene targeting') с помощью цинк-пальцевых нуклеаз (Urnov et al., Nature Reviews 2010_Genome editing with engineered zinc finger nucleases; Townsend et al., Nature 2009_High-frequency modification of plant genes using engineered zinc-finger nucleases) или TAL-эффекторных нуклеаз (WO 2010/079430; WO 2011/072246). При этом модификация нативного промотора эндогенного гена подразумевает и дополнительное внесение цис-регуляторного элемента согласно изобретению в нативный промотор, который изначально содержит цис-регуляторный элемент согласно изобретению, то есть мультимеризацию имеющихся цис-регуляторных элементов. Подобный модифицированный промотор может обладать измененными свойствами по сравнению с нативным вариантом, например, иной специфичностью, уровнем экспрессии или фоновой активностью. Из модифицированных растительных клеток можно восстанавливать модифицированные растения с помощью известных методов.
Таким образом, изобретение касается описанных трансгенных (трансформированных) растений, которые были трансформированы с помощью химерного промотора, рекомбинантного гена или вектора согласно изобретению, и описанных растений, модифицированных путем внесения минимум одного цис-регуляторного элемента согласно изобретению или одного химерного промотора согласно изобретению. Трансгенные или модифицированные растения могут принадлежать любому требуемому виду. Это могут быть однодольные, двудольные или покрытосемянные растения, предпочтительно сельскохозяйственные или садоводческие культуры, например, кукуруза, рис, пшеница, рожь, ячмень, овес, сорго, картофель, рапс, подсолнечник, соя, хлопок или сахарная свекла.
В предпочтительном варианте изобретения трансгенное или модифицированное растение является полностью устойчивым или обладает повышенной устойчивостью к одному или нескольким патогенам по сравнению с нетрансгенным или немодифицированным растением того же вида (диким типом).
Данное изобретение также включает часть, ткань клетку или семя трансгенного или модифицированного растения, причем, эта часть, ткань, клетка или семя также имеют внесенный в растение трансген или модификацию.
Варианты данного изобретения описываются на примерах с использованием прилагаемых иллюстраций и последовательностей:
Рис. 1: В качестве примера клонирования одиночных последовательностей как химерных промоторов показаны плазмиды с элементом 2xCis05 и мультимеризованным элементом 4xCis05. Плазмиды выведены из pΒΤΊΟ-GUS (Sprenger and Weisshaar (2000): The Plant Journal 22, 1-8). Структура других плазмидов является соответствующей. Amp: устойчивость к ампициллину; WRKY30-Cis05: двойная одиночная последовательность Cis05. 35S-minimal: минимальный промотор 35S; Luc-m3: репортерный ген люциферазы; pAnos: терминатор nos.
Рис. 2: патоген-индуцируемые одиночные последовательности (А-X) для различных видов. В верхних строках приведены названия одиночной последовательности, группа мотивов и сами одиночные последовательности. Сердцевинные последовательности мотива, который привел к выбору одиночной последовательности, выделены жирным и подчеркнуты. Ниже приводятся знак базового, идентифицированного биоинформационным способом мотива и его матрица. Если было протестировано несколько одиночных последовательностей одного мотива, они приводятся вместе.
Рис. 3: Все одиночные последовательности одной группы мотивов, давшие положительный результат, а также выведенные из них мотивы последовательностей и семейные мотивы (рис. 3А-G). В самой верхней строке названа соответствующая группа мотивов. Ниже показано распределение всех одиночных последовательностей, давших положительный результат и включающих как минимум сердцевинные последовательности и - при наличии - другие консервативные основания. Основания, из которых выводится мотив сердцевинной последовательности, приведены в рамке.
Рис. 4: Филогенетическое древо идентифицированных групп мотивов. Древо было создано в результате кластерного анализа с использованием веб-сервера STAMP. После номера группы мотивов в скобках указано количество мотивов в данной группе.
Рис. 5: А) Мутагенез одиночной последовательности Cis05. В используемой последовательности помимо мотивов Cis05 (жирный, подчеркнутый) содержится W-бокс (жирный, в рамке). В оба мотива были внесены мутации. С тетрамеризированными мутировавшими дериватами были проведены описанные анализы петрушки на индукцию с РАМР РЕР25. РАМР-индуцированная активность химерных промоторов была измерена через 4, 8 и 24 ч (правая сторона). Мутации в мотиве Cis05 (Cis05mut1) и в W-боксе (Cis05mut2) ведут к значительному снижению индуцированной активности. Индуцируемость утрачивается полностью только, если мутировали оба мотива (Cis05mut1+2). В) sCis05 - это укороченный вариант Cis05, который содержит только мотив Cis05, но не W-бокс. РЕР25-индуцируемость sCis05 и мутированных дериватов проверялась так, как описано в 5А. Два столбца означают два биологических репликата (независимые трансформации). Для ориентации под дериватами sCis05 показан консенсус W-бокса.
Рис. 6А: Элиситор-чувствительная экспрессия репортерных генов химерных промоторов 4x30l-8_M1_S2 и 4x18H_M2_S1 с мутациями в одиночных последовательностях. Мутировавшие основания подчеркнуты. Активация проводилась РЕР25 в протопластах петрушки. Нуклеотидные последовательности мутировавших дериватов одиночных последовательностей приведены под диаграммами. +, с элиситором РЕР25; -, без элиситора. 2S2D: положительный контроль; MS23GUS: отрицательный контроль (пустой вектор).
Рис. 6В: Элиситор-чувствительная экспрессия репортерных генов химерных промоторов 4x20u_M1_S1 и 4x27G-8_M1_S1 с мутациями в одиночных последовательностях. Мутировавшие основания подчеркнуты. Активация проводилась РЕР25 в протопластах петрушки. Нуклеотидные последовательности мутировавших дериватов одиночных последовательностей приведены под диаграммами.
Рис. 7: Двоичный вектор для трансформации репортерного гена люциферазы под контролем химерных промоторов в свекле. В качестве примера показан вектор с химерным промотором 4xCis05. nptll: устойчивость к канамицину; WRKY30-Cis05: двойная одиночная последовательность Cis05. 35S-minimal: минимальный промотор 35S; Iuc-m3: репортерный ген люциферазы; Anos: терминатор nos.
Рис. 8А: Индуцированная Cercospora beticola активность промотора в стабильно трансформированной сахарной свекле. У указанных конструктов для нескольких независимых трансформантов была определена активность люциферазы после заражения С.beticola в растениях in vitro (по 4 репликата на трансформант и момент времени). На основании измеренных значений была рассчитана медиана. На верхней диаграмме приведены результаты для одиночных последовательностей из группы мотивов 12, на нижней - результаты для элементов из группы мотивов 27. Ко: контроль (имитация заражения); inf; заражение С.beticola; 1d -4d: дней после инокуляции (d.p.i.). Контроль: не трансгенные растения.
Рис. 8В: Индуцированная Cercospora beticola активность промотора 4xCis05 и его дериватов в стабильно трансформированной сахарной свекле. Для 4xCis05 и его дериватов была определена активность люциферазы после заражения С.beticola в нескольких независимых трансформантах (по 4 репликата на трансформант и момент времени). На основании измеренных значений была рассчитана медиана. Последовательность разных дериватов приведена на рис. 5А и 5В. Под диаграммой дополнительно для каждого из различных дериватов указано, содержит ли он мотив Cis05 или W-бокс. Ко: контроль (имитация заражения); inf; заражение С.beticola; 1d-4d: дней после инокуляции (d.p.i.). non transgenic: не трансгенный контроль.
Рис. 9: Индуцированная Cercospora beticola активность химерного промотора 4xGG6_M1_S1 в стабильно трансформированной сахарной свекле. Для 10 независимых трансформантов с конструктом 4xGG6_M1_S1-luc была определена активность люциферазы после заражения С.beticola в растениях in vitro (по 4 репликата на трансформант и момент времени). Ко: контроль (имитация заражения); inf; заражение С.beticola; 2d, 3d, 4d и 7d: дней после инокуляции (d.p.i.). 3DC4156: не трансгенные контрольные растения.
Рис. 10: Карта плазмида, используемого для переходных тестов в пшенице. В качестве примера приведен плазмид химерного промотора 4xCis05. Rue: репортерный ген люциферазы Renilla. AMP: устойчивость к ампициллину. WRKY30-Cis05: двойная одиночная последовательность Cis05.
Рис.11: Тест на индукцию химерного промотора 4xCis05 и его мутировавших дериватов с вызванными фузариозом мутациями в мотиве Cis05-Motiv (Cis05mut1), в W-боксе (Cis05mut2) или в обоих мотивах (Cis05mut1+2). Соответствующие конструкты были временно преобразованы в пшенице, активность люциферазы измерялась через 20 часов после инкубации с возбудителем фузариоза. 4xCis05-dam/dcm означает тест, в котором плазмидная ДНК неметилирующего штамма E.coli использовалась для исключения индукции плотины dam/dcm-метилированной ДНК (также являющейся потенциальным РАМР). При мутации сердцевинной последовательности Cis05 индуцируемость полностью утрачивается. Напротив, мутация в W-боксе не приводит ни к каким изменениям. Последовательности Cis05 и его мутировавших дериватов приведены справа. Мутировавшие основания выделены красным.
Рис. 12: Индуцированная и не индуцированная активность химерных комбинаторных промоторов после РЕР25-индукции в петрушке. Тесты проводились на трех биологических репликатах, синяя линия означает фактор индукции. Под диаграммой в нижнем ряду приведены элементы в позиции 5', в верхнем ряду - в позиции 3'. 3018b - другое обозначение одиночной последовательности 30I-8_M1_S2.
Рис. 13: Синергетическое и антагонистическое взаимодействие одиночных последовательностей в химерных комбинаторных промоторах после РЕР25-индукции в петрушке. Фиолетовым цветом обозначен фактически измеренный фактор индукции, синим - фактор индукции, ожидаемый на основании факторов индукции отдельных элементов. Соотношение этих значений обозначено желтой линией. Если точки желтой линии находятся выше значения 1, значит, отдельные элементы демонстрируют синергетическое взаимодействие.
Рис. 14: Трансгенная сахарная свекла с конструктом 4xCis05-RFP была заражена Cercospora beticola. Заражение ведет к активации промотора 4xCis05, что, в свою очередь, приводит к образованию красного флуоресцентного протеина RFP. Протеин под микроскопом светится красным цветом. Как видно, индукция и, следовательно, свечение ограничены областью проникновения или местом заражения.
Рис. 15: Примерное схематическое изображение химерного промотора согласно изобретению. Химерный промотор оперативно связан с соответствующей молекулой нуклеиновой кислоты и включает (А) кроме минимального промотора гетерологичный цис-регуляторный элемент, (В) кроме минимального промотора димер/мультимер гетерологичного цис-регуляторного элемента или (С) кроме минимального промотора два димера/мультимера с различными гетерологичными цис-регуляторными элементами. Кроме того, на (D) показан природный химерный промотор, включающий эндогенный минимальный промотор и эндогенный цис-регуляторный элемент, причем этот промотор был модифицирован путем интеграции дополнительного гомологичного цис-регуляторного элемента.
Рис. 16: Трансгенные растения арабидопсис с тетрамером цис-регуляторного элемента Cis05 в химерном промоторе, контролирующем экспрессию репортерного гена GUS, были исследованы на индуцируемую патогеном и повреждением, а также специфическую тканевую активность элемента. Для данного промотора были исследованы 10 независимых трансформантов. В каждом случае показана репрезентативная линия. На левом изображении (5 d.p.i. с H. arabidopsidis) показана активность промотора после заражения совместимым патогеном Hyaloperonospora arabidopsidis, на правом (mock control) - соответствующий контрольный образец. Промотор демонстрирует четкую индукцию, вызванную Hyaloperonospora arabidopsidis (потемнение растительной ткани).
Кроме того, на правом изображении показан лист с надрезом. Синеватый оттенок в месте надреза должен свидетельствовать о вызванной повреждением индукции промотора с тетрамером цис-регуляторного элемента Cis05.
Биоинформационная идентификация цис-регуляторных элементов согласно изобретению:
Биоинформационная идентификация новых цис-регуляторных элементов базируется на общедоступных данных об экспрессии, полученных в результате микроматричного анализа. Эти сведения хранятся в таких базах данных, как TAIR, NASCArrays, Geo или ArrayExpress, а также могут быть получены непосредственно из публикаций об экспериментах с использованием микроматричного анализа (например, Rhee et al., 2003; Craigon et al., 2004; Barret und Edgar, 2006; Brazma et al., 2006, Zipfel et al., 2004, 2006; Bülow et al., 2007; Wan et al., 2008). Для биоинформационной идентификации новых цис-регуляторных элементов сначала на основании общедоступных данных были определены группы генов растения Arabidopsis thaliana, экспрессия которых индуцируется такими патогенами, как P. syringae, В. cinérea или такими РАМР, как flg22 или хитин. Затем последовательности промоторов этих групп генов были извлечены из последовательности генома Arabidopsis thaliana (TAIR; http://www.arabidopsis.org). При помощи различных известных алгоритмов (MEME, Bioprospector, Alignace, BEST и т.п.) эти последовательности были исследованы на обогащенные мотивы.
Запросы в базы данных
Для отправки запросов в базы данных с целью идентификации корегулируемых генов была написана программа, позволяющая идентифицировать гены, которые активируются или индуцируются при возникновении максимум шести различных стимулов. Для идентификации корегулируемых генов в базы данных было отправлено более 700 запросов. В результате было получено более 400 групп совместно индуцируемых генов, подходящих для идентификации общих цис-регуляторных мотивов с использованием программного пакета BEST (Che et al., 2005). У 77 групп количество совместно регулируемых генов было сокращено до 120 путем увеличения необходимого фактора индукции. После этого общее количество групп корегулируемых генов (2-120) составляло 510.
Из этих 510 групп генов были выделены последовательности промотора длиной от 500 до 1000 пар оснований в направлении вверх от сайта инициации транскрипции корегулируемых генов, сайт инициации транскрипции которых был известен. Последовательности промоторов групп корегулируемых генов были исследованы на наличие консервативных мотивов последовательностей с использованием программ BEST, Cismodule, MD-Scan, BloProspector или MEME. При этом были выбраны мотивы длиной 5-10 нуклеотидов, 10-15 нуклеотидов и 15-20 нуклеотидов. Таким образом, было выполнено примерно 500×3 (длины мотивов)=1500 запросов. В большинстве случаев увеличение длины мотивов не привело к идентификации новых мотивов, которые отсутствовали бы при меньшей длине. В рамках анализов BEST мотив классифицировался в том случае, если он был найден по меньшей мере двумя из четырех программ BEST.
При определенных различных стимулах имелись одинаковые корегулируемые гены и, следовательно, одинаковые мотивы. Затем идентичные мотивы избыточных групп генов были объединены в новые мотивы (таблица групп генов), чтобы в рамках систематизирующих сравнительных анализов всех мотивов между собой сравнивались только уникальные мотивы.
Был составлен каталог всех идентифицированных мотивов плюс файл анализа и обозначения последовательностей отдельных мотивов. Обозначения последовательностей (Crooks et al., 2004), созданные на сайте http://weblogo.berkeley.edu/, отображают консервативность нуклеотидов на отдельных позициях мотива. Матрица была создана из образующих мотив последовательностей, последовательности которых, а также соответствующие гены также воспроизводятся. С помощью программы STAMP (Mahony and Beños, 2007), которая находится по адресу http://www.benoslab.pitt.edu/stamp, идентифицированные мотивы были сравнены с уже известными цис-регуляторными элементами (PLACE, Agris, Athamap). Кроме того, на сервере STAMP удалось объединить схожие мотивы в группу. Программа выдает филогенетическое древо, отражающее сходства групп мотивов (рис. 4).
Подтверждение патоген-индуцируемости идентифицированных иис-регуляторных элементов
Так как в результате использования биоинформационных методов возможно определение ложно-положительных последовательностей, патоген-индуцируемость следует подтвердить экспериментальным путем. Для экспериментального подтверждения последовательности, идентифицированные биоинформационным способом, были клонированы с использованием стандартных технологий клонирования ДНК как тетрамеры перед репортерным геном люциферазы и проверены на индуцируемость PAMP РЕР25 в переходных системах экспрессии в петрушке. Для экспериментального исследования были выбраны элементы, являющиеся новыми и не имеющими сходства с известными цис-регуляторными элементами вызванной патогеном индукции. Одиночные последовательности, приведенные в таблице 2, были синтезированы in vitro с линкерами Spel и Xbal или Spel и Sail и клонированы в плазмид MS23. При этом MS23 имеет либо репортерный ген β-глюкуронидазы (GUS), либо репортерный ген люциферазы с минимальным промотором 35S. Все элементы были тетрамеризованы и секвенированы для проверки. В качестве примера клонирования одиночных последовательностей как химерных промоторов на рис. й показаны плазмиды с элементом 2xCis05 и мультимеризованным элементом 4xCis05.


В дальнейшем тетрамеризованные одиночные последовательности были проверены на индуцируемость под действием PAMP РЕР25 во временно трансформированных клетках петрушки из одной клеточной культуры. Помимо общей валидации это было необходимо для подтверждения того, что эти элементы индуцируются в разных видах растений и разными элиситорами.
Для теста на петрушке из клеточной культуры возрастом 5 дней были изолированы протопласты. Для этого в центрифуге были отделены 35 мл клеточной культуры, ресуспендированы в 90 мл стерильного отфильтрованного раствора с 0,5% целлюлазы, 0,2% мацерозима R-10 и 0,24 Μ CaCl2 и инкубированы в течение ночи при 26°C с раскачиванием. После этого выделенные протопласты были подвергнуты гранулированию в центрифуге, промыты 40 мл 0,24 Μ CaCl2 и затем ресуспендированы в 50 мл среды Р5 (1 пакет готовой среды Gamborgs В-5, 1 мг 2,4 D, 96,9 г сахарозы, рН 5,7 с 1Μ КОН, стерильная фильтрация). После центрифугирования протопласты плавают на поверхности среды Р5, и их можно снять. Очистка средой Р5 была повторена 2 раза.
Для трансформации в пробирке с навинчивающейся крышкой объемом 10 мл находилось 5 мкг тестируемого промоторного конструкта, 2,5 мкг экспрессирующего основную люциферазу Renilla вектора нормализации и 200 мкл PEG. В среду было добавлено 200 мкл протопластов, затем было проведено осторожное смешивание и инкубация в темноте при комнатной температуре в течение 20 минут. Затем реакция была остановлена путем добавки 5 мл 0.275 М раствора CaNO3. Трансформированные протопласты были отделены в центрифуге, добавлены в 6 мл среды Р5 и разделены на 2 аликвоты. Одна аликвота была активирована с помощью Рер25 (конечная концентрация: 300 нг/мл; последовательность: VTAGAEVWNQPVRGFKVYEQTEMT), вторая использовалась как контрольная. После инкубации в течение ночи протопласты были собраны в центрифуге. Активность люциферазы была установлена с помощью набора "Dual Luziferase Kit" (Promega, Маннгейм, Германия) в люминометре "Sirius" (Berthold Detection System GmbH, Пфорцхайм, Германия). Для этого клетки петрушки были подвергнуты гранулированию в центрифуге и лизированию в течение 20 минут при 4°C в пассивном лизирующем буфере PLB на 150 мкл (Passive Lysis Buffer; Promega, Маннгейм, Германия). Остатки клеток в течение 20 минут отделялись в настольной центрифуге при 13000 об/мин и 4°C.
В зависимости от того, использовался репортерный ген люциферазы или GUS, лизат обрабатывался по-разному. При использовании репортерного гена люциферазы из супернатанта с выделенной люциферазой 5 мкл пробы в пробирках по 5 мл (Sarstedt, арт.55.476) смешивались с 50 мкл буфера LARII (Promega, Маннгейм, Германия). Буфер содержит субстрат люциферазы, благодаря чему люминометром можно измерить активность энзима и, следовательно, промотора. Измерение делится на предварительный этап в течение 2 секунд и измерение люциферазы в течение 10 с. Затем добавляется 50 мкл буфера Stop & Glo (Promega, Маннгейм, Германия) и осторожно смешивается путем нагрева. Буфер останавливает активность люциферазы и позволяет измерить основную активность люциферазы Renilla вектора нормализации. Измеренное значение используется для нормализации различных коэффициентов трансформации. Это измерение также делится на предварительный этап в течение 2 секунд и измерение люциферазы в течение 10 с. Если использовался репортерный ген β-глюкуронидазы (GUS) из Е. coli (Jefferson et al., 1987), подтверждение проводилось в энзимной реакции, в которой субстрат MUG был гидролизован в 4-MU. Затем на основании флуоресценции проводилось подтверждение и определение количества 4-MU.
В соответствии с этими методами была измерена активность с РАМР РЕР25 и без него для всех исследованных одиночных последовательностей.
В таблице 2 приведены все протестированные одиночные последовательности. Кроме того, было указано, индуцировались ли они РАМР РЕР25 в петрушке. Цис-регуляторные элементы (одиночные последовательности) с идентифицированной патоген-индуцируемостью и их мотивы приведены на рис. 2.
Анализы мутаций идентифицированных сердцевинных последовательностей
Чтобы доказать, что РАМР - и патоген-индуцируемость отдельных последовательностей вызвана идентифицированными сердцевинными последовательностями, для некоторых одиночных последовательностей были проведены анализы мутаций. Для этого были созданы мутировавшие дериваты одиночных последовательностей. Мутировавшие одиночные последовательности синтезировались как олигонуклеотиды. Клонирование плазмидов проводилось в соответствии с конструктами с химерными промоторами без мутаций. Затем конструкты были проверены на РЕР25-индуцируемость в петрушке в соответствии с описанием выше. Результаты анализов мутаций приведены на рис. 5А, рис. 5В, рис. 6А и рис. 6В. Для всех пяти исследованных элементов удалось показать, что индуцируемость вызывается идентифицированной сердцевинной последовательностью.
Особое значение это имеет для одиночных последовательностей Cis05 и 30I-8 М1 S2 группы 27. Сердцевинная последовательность 30I-8_M1_S2 частично перекрывается с комплементарной W-боксу (TTGAC; Rushton et al., 1996) последовательностью (GTCAA). Перекрывающаяся последовательность W-бокса была найдена также в одиночных последовательностях 30I-8_M1_S3, Cis02 и Cis13. Однако в результате анализов мутаций, в частности, варианта 30l-8_M1_S2_mut2, удалось показать, что основания за пределами W-бокса имеют решающее значение для РАМР-индуцируемости (рис. 6А). Кроме того, у других членов группы 27 нет перекрывающейся последовательности W-бокса. Соответственно, обязательно необходимая для W-бокса сердцевинная последовательность TGAC не является частью мотива последовательности или семейного мотива группы 27 (рис. 3F). Таким образом, мотивы или одиночные последовательности группы 27 не являются вариантами W-бокса.
В одиночной последовательности Cis05 последовательность W-бокса находится за пределами сердцевинной. В этом случае анализы мутаций показали, что этот W-бокс все же способствует РАМР-индуцируемости. Однако мутация только сердцевинной последовательности Cis05 или только W-бокса ведет к снижению индуцируемости, а полная потеря индуцируемости возможна только при мутации обоих элементов. Таким образом, речь идет об одиночной последовательности с двумя функциональными, РАМР - и патоген-индуцируемыми цис-регуляторными элементами: известным W-боксом и новым мотивом Cis05 (рис. 2Т). При этом сочетание обоих элементов имеет значительно более высокую активность, чем элементы по отдельности. Чтобы ограничить способствующую индуцируемости область последовательности мотива Cis05 (рис. 2Т), он был тетрамеризован отдельно от W-бокса (sC¡s05), также были созданы еще четыре мутированных деривата мотива Cis05. Дериваты были проверены на индуцируемость РАМР РЕР25 в петрушке в соответствии с описанием выше (рис. 5 В). При этом помимо решающего значения сердцевинной последовательности для индуцируемости удалось показать, что оба основания, находящихся перед сердцевинной последовательностью в направлении 5', важны для индуцируемости Cis05, что также подтверждает, что речь идет не о варианте W-бокса.
На основании результатов других анализов мутаций удалось также подтвердить сердцевинные последовательности элементов 20u_M1_S1 и 27G-8_M1_S1. Тесты проводились в соответствии с описанием выше. Результаты этих анализов приведены на рис. 6В (вверху: 20u_M1_S1; внизу: 27G-8_M1_S1).
Выведение семейных мотивов
Благодаря тому, что тестировались цис-регуляторные элементы в петрушке, идентифицированные в промоторах Arabidopsis, было подтверждено наличие биологической функциональности идентифицированных цис-регуляторных элементов, которая является общей для разных видов растений. Как и ожидалось, эксперименты доказали, что не все последовательности, идентифицированные биоинформационными способами, функциональны. Примерно треть протестированных последовательностей ДНК, индуцировалась в петрушке под действием РЕР25. Другие последовательности, идентифицированные в рамках биоинформационного анализа, были ложно-положительными или не продемонстрировали нужную, общую для разных видов растений биологическую функциональность. Эти важные результаты можно было использовать, чтобы на основании функционально действенных одиночных последовательностей идентифицировать семейные мотивы групп мотивов 1,5, 11, 12, 21, 21n и 27, а также соответствующие сильно консервативные характерные сердцевинные последовательности (рис. 3). С этой целью все функционально действенные одиночные последовательности, относящиеся к одной группе мотивов, были объединены, и на их базе был выведен один консенсус и один мотив. Сначала в качестве характерного мотива сердцевинной последовательности семейных мотивов была определена область, которая во всех базовых одиночных последовательностях является частью сердцевинной последовательности. Сердцевинная последовательность группы мотивов 27 имеется также в последовательности LS10 (Lebel et al., 1998). Там в нативном промоторе PR-1 в этой области последовательности была создана мутация, которая включала 10 оснований и привела к сильному снижению SA- или INA-индуцируемости нативного промотора. Однако полученный там результаты не позволяют вывести мотив или сердцевинную последовательность. Кроме того, не продемонстрирована возможность использования LS10 для химерных промоторов. Кроме того, семейный мотив группы 27 отделяет эту группу от последовательности LS10. Этот семейный мотив исключает наличие С на позиции 5, в то время как в LS10 на этой позиции С имеется. Кроме того, он исключает наличие G на позиции 17, в то время как в LS10 на этой позиции G имеется. Наконец, на позиции 18 требуется Τ или С, в то время как в LS10 на этой позиции имеется А.
Подтверждение индуцируемости. вызываемой патогенным грибком Cercospora beticola в стабильно трансформируемой сахарной свекле
Новые цис-регуляторные элементы также должны отличаться способностью к индукции в разных видах растений под действием разных РАМР и патогенов. Чтобы показать индуцируемость в важном с агрономической точки зрения растении под действием важного с агрономической точки зрения патогена, одиночные последовательности, давшие положительные результаты в петрушке, были устойчиво трансформированы в сахарной свекле. С этой целью химерные промоторы с тетрамеризованными отдельными последовательностями, включая ген luc, были переклонированы с помощью участков контакта Ascl и Sad в двоичный вектор 1xW1-luc-kan, базирующийся на двоичном векторе pGPTV плазмид. В качестве примера на рис. 7 показан вектор 4xCis05-luc-kan. Соответствующие векторы были составлены для всех проверенных элементов. ДНК плазмида двоичных векторов была изолирована из Е. Coli и с помощью системы Gene Puiser® II. Electroporation System при настройке 25 мФ и 2,5 кВ трансформирована в Agrobacterium tumefaciens, штамм GV3101. Рекомбинантные клоны A. tumefaciens отбирались с использованием антибиотика канамицина (50 мг/л). Трансформация сахарной свеклы проводилась согласно работе Lindsey et al. (1991) с использованием канамицина.
Трансгенность растения была проверена с помощью ПЦР. Использование праймеров GTGGAGAGGCTATTCGGTA (SEQ ID NO: 36) и CCACCATGATATTCGGCAAG (SEQ ID NO: 37) привело к амплификации участка ДНК длиной 553 пар оснований из гена nptll. ПЦР проводилась с использованием 10 нг геномной ДНК, праймера концентрацией 0,2 мкМ при температуре отжига 55°C в аппарате Multlcycler РТС-200 (MJ Research, Уотертаун, США).
10-20 независимых трансгенных линий были вегетативно размножены в культуре in vitro и инфицированы Cercospora beticola (Isolât Ahlburg). Через 1, 2, 3 и 4 дня собиралось по 4 растения из линии, и их активность люциферазы измерялась с помощью Promega Luciferase Assay System, 100 образцов, кат. Е1500 (LAR). Для этого образцы были помещены в 4 емкости с буфером CCLR (Cell Culture Lysis Reagent - лизирующий реагент клеточных культур, 5х) и обработаны средством Heidolph (RZR 2020). Остатки растений обрабатывались в центрифуге 10 минут при 4°C и 14000 об/мин, супернатант использовался для измерения люциферазы. Для измерения 10 мкл образца были пипетками добавлены в пробирку объемом 5 мл (Sarstedt, арт. 55.476), в которую затем было добавлено 100 мкл реагента для анализа люциферазы (LAR; Promega, Маннгейм, Германия). Затем было проведено осторожное смешивание и определение активности люциферазы люминометром "Sirius" (Berthold Detection System GmbH, Пфорцхайм, Германия).
В том числе, тестировались новые элементы из групп 12 и 27 (классификацию групп см. рис. 3D и F). На основании результатов измерения активности люциферазы 10-20 независимых трансгенных линий с химерным промотором была рассчитана медиана. Тесты разных промоторов разделены на группы, из которых взяты элементы. Результаты приведены на рис. 8А. Прямое сравнение показывает, что одиночные последовательности одной группы мотивов демонстрируют явно различающуюся активность несмотря на высокую степень гомологичности содержащихся в них мотивов. Таким образом, нуклеотиды семейного мотива, примыкающие к сердцевинной последовательности, могут значительно влиять на силу промотора и фоновую активность. Следовательно, разные последовательности одной группы мотивов не обладают идентичными функциями, то есть позволяют развитие химерных промоторов с количественными различиями регулирования экспрессии генов. Для тестирования следующей группы мотивов элемент GG6_M1 (одиночная последовательность GG6_M1_S1) был устойчиво трансформирован в сахарной свекле в соответствии с описанием выше и протестирован на индуцируемость под действием Cercospora. С этой целью 10 независимых трансгенных растений (свекла) были вегетативно размножены в культуре in vitro и инфицированы Cercospora beticola (Isolât Ahlburg). Через 2, 3, 4 и 7 дней собиралось по 4 растения из линии, и их активность люциферазы измерялась с помощью Promega Luciferase Assay System, 100 образцов, кат. Е1500 (LAR). Результаты приведены на рис. 9; они показывают явную индуцируемость одиночной последовательности GG6_M1_S1 в сахарной свекле под действием Cercospora beticola. Кроме того, гистохимический анализ показал, что индукция происходит практически только в проводящей ткани.
Для исследования влияния последовательности W-бокса в одиночной последовательности Cis05 в стабильно трансформированных растения мутировавшие дериваты Cis05mut1 и Cis05mut2, а также укороченная одиночная последовательность sCis05 из тестов с петрушкой были стабильно трансформированы в сахарной свекле. Создание конструктов и трансформация проводились в соответствии с описанием выше. 18 независимых трансгенных линий были вегетативно размножены in vitro и инфицированы Cercospora beticola (Isolât Ahlburg). Через 1, 2, 3 и 4 дня собиралось по 4 растения из линии, и их активность люциферазы измерялась в соответствии с описанием выше (рис. 8 В).
Анализ мутаций в стабильно трансформированных растениях показывает, что данный W-бокс сам по себе, т.е. с мутировавшим мотивом Cis05, обеспечивает лишь слабую патоген-индуцируемость под действием Cercospora. Мотив Cis05 сам по себе (мутировавший W-бокс или укороченный элемент без W-бокса), например, явно обеспечивает индуцируемость. Вся одиночная последовательность Cis05 с W-боксом также обеспечивает индуцируемость, но по сравнению с дериватами без W-бокса обладает повышенной фоновой активностью. Таким образом, в стабильно трансформированной сахарной свекле мотив Cis05 сам по себе равнозначен комбинации с W-боксом или даже превосходит ее.
Еще один важный признак химерных промоторов согласно изобретению состоит в том, что патоген-индуцированная активность ограничена областью заражения. Это показано на рис. 14 на примере промотора 4xCis05 согласно изобретению. Этот промотор был слит с красным флуоресцирующим репортерным геном RFP, а полученный таким способом конструкт стабильной трансформирован в сахарной свекле. Под лазерным сканирующим микроскопом активность наблюдается как красное свечение. На рис. 14 показана локальная индукция промотора 4xCis05 вокруг места проникновения гифы Cercospora.
Расширенная патоген-индуиируемость. общая для разных видов
В качестве еще одного примера широких возможностей использования данных цис-регуляторных элементов в разных видах растений для Cis05 приводятся результаты переходных тестов на индуцируемость в однодольном растении (пшенице) под действием грибка Fusarium culmorum. Так как минимальный промотор 35S в пшенице вызывает недостаточную активность, элементы Cis05 потребовалось переклонировать. Для этого с помощью энзимов Есо311 и Xbal они были вырезаны из плазмидов, использовавшихся в тестах с петрушкой, и клонированы в вектор pubiTATARucll, открытый с помощью Есо311 и Bcul (рис. 10). Соответствующие конструкты были созданы для Cis05, а также для мутировавших одиночных последовательностей Cis05 - C¡s05mut1 и Cis05mut2, - в которых мутировал либо мотив Cis05, либо W-бокс. Эти конструкты были биолистически трансформированы в инфицированные Fusarium graminearum первичные листья пшеницы сорта "Тайфун", а также в не инфицированные контрольные листья.
Для инфицирования мицелий Fusarium graminearum был соскоблен предметной пластиной с пластины с грибницей и быстро измельчен и суспендирован в 200 мл воды с помощью устройства Ultraturrax. Затем было добавлено 200 мкл 2-процентного средства "тритон", первичные листья пшеницы опущены в суспензию на 1 минуту. После этого инфицированные кусочки листьев и кусочки не инфицированных контрольных листьев были положены на агаровые пластинки с Н2O и подвергнуты переходной трансформации с помощью генной пушки Biorad в соответствии с инструкциями производителя и при использовании мембран на 1100 фунтов на кв. дюйм. В качестве фактора нормализации использовался основной вектор экспрессии люциферазы.
Затем листья пшеницы проходили инкубацию в течение ночи при 25°C. Для определения активности люциферазы листья были растерты каждый в 1 мл буфера PLB с морским песком. После обработки в центрифуге в течение 20 минут при 4°C из супернатанта с выделенной люциферазой 5 мкл пробы в пробирках по 5 мл (Sarstedt, арт. 55.476) смешивались с 50 мкл буфера LARII (Promega, Маннгейм, Германия). Буфер содержит субстрат люциферазы, благодаря чему можно измерить активность вектора нормализации. Измеренное значение используется для нормализации различных коэффициентов трансформации. Измерение делится на предварительный этап в течение 2 секунд и измерение люциферазы в течение 10 с. Затем добавляется 50 мкл буфера Stop & Glo (Promega, Маннгейм, Германия) и осторожно смешивается путем нагрева. Буфер останавливает активность люциферазы и позволяет измерить активность люциферазы Renilla, которая соответствует активности промоторов Cis05. Это измерение также делится на предварительный этап в течение 2 секунд и измерение люциферазы в течение 10 с.
Индуцированная и неиндуцированная активность промотора 4xCis05, а также его мутировавших дериватов была измерена в рамках 5 повторяющихся биологических тестов (рис. 11). Затем весь тест был повторен еще раз. Оба повтора показывают, что в пшенице патоген-индуцированная активность вызывается мотивом Cis05, в то время как мотив W-бокса не способствует активности, индуцируемой Fusarium graminearum.
Анализ индуцируемой патогеном и повреждением активности и специфической тканевой активности элементов Cis02. Cis05. Cis09, Cis12 и Cis13 в Arabidopsis
Новые цис-регуляторные элементы также должны отличаться способностью к индукции в разных видах растений под действием разных РАМР и патогенов. Для дополнительного контроля активности химерных промоторов 6 промоторов с тетрамезированными одиночными последовательностями были стабильно трансформированы в Arabidopsis. С этой целью тетрамеризованные элементы Cis02, Cis05, Cis09, Cis12 и Cis13 с минимальным промотором 35S перед репортерным геном GUS были клонированы в вектор трансформации pBIN-GUS. Готовый конструкт был трансформирован в агробактерии. Затем была проведена трансформация методом погружения цветочных почек Arabidopsis (Clough and Bent, 1998) с целью стабильной интеграции конструктов промотора GUS в геном растения. Селекция трансгенных растений проводилась с использованием канамицина. Для каждого элемента были исследованы 10 независимых трансформатов. Активность репортерного гена GUS и, следовательно, промоторов, видна благодаря окрашиванию GUS (Jefferson et al., 1987). Активация соответствующего химерного промотора приводит к экспрессии энзима (GUS), образующего синий краситель. Таким образом, цвет отображает активность соответствующего химерного промотора. Для тестирования патоген-индуцируемости промоторов трансгенные растения Arabidopsis были инфицированы совместимым патогеном Hyaloperonospora arabidopsidis. Через 5 дней после заражения активность промоторов была подтверждена окрашиванием GUS. Кроме того, отдельные листья были надрезаны лезвием для тестирования индуцируемости промоторов, вызываемой повреждением (рис. 16).
Элемент Cis02 показал хорошую патоген-индуцируемость и невысокую индуцируемость при повреждении. Элемент Cis05 показал в целом сильную активность; индукция под действием И. Arabidopsidis также сильна. Элемент Cis09 показал хорошую индукцию промотора после инфекции и почти нулевую активность после повреждения. Элемент Cis12, как и элемент Cis09 во всех проверенных линиях показал почти нулевую активность после повреждения и явную индукцию под действием патогена Н. arabidopsidis. Элементы Cis12 и Cis09 имеют общий семейный мотив. Элемент Cis13 также сильно индуцируется при инфекции Н. arabidopsidis. Однако явной индукции при повреждении не наблюдается. В качестве примера на рис. 16 показан окрас GUS трансгенных растений A. thaliana, которые экспрессируют репортерный ген GUS под контролем химерного промотора с 4 Cis05.
Комбинаторика цис-регуляторных элементов
Благодаря синергетическому взаимодействию или интеграции различных путей передачи сигнала (можно представить себе промотор, который может воспринимать и интегрировать сигналы „EF-Tu" И „flg22", чтобы активироваться) химерные комбинированные промоторы, состоящие из разных цис-регуляторных элементов, могут обладать более высокой специфичностью и (или) активностью, чем содержащиеся в них отдельные элементы (Rushton et al., 2002). Для выявления оптимальных комбинаций новых цис-регуляторных элементов для химерных комбинаторных промоторов при использовании стандартных методов клонирования ДНК были созданы химерные комбинированные промоторы со всеми возможными сочетаниями "2×2" описанных здесь цис-регуляторных элементов Cis02, Cis05, Cis12, Cis13 и 30I-8_M1_S2, a также названных здесь элементов D-бокс (Rushton et al., 2002), S-бокс (Kirsch et al., 2000) и Gstl-бокс (Strittmatter et al., 1996). При этом метод был тем же, что и при тетрамеризации одиночных последовательностей, только клонировались не два идентичных димера (например, 2x30l-8_M1_S2 и 2x30l-8_M1_S2), а два разных (например, 2xCis05 и 2x30l-8_M1_S2). Созданные таким способом химерные комбинаторные промоторы тестировались в переходных системах экспрессии в петрушке на индуцируемость под действием РАМР РЕР25. Результаты (абсолютная не индуцированная и индуцированная активность химерных комбинаторных промоторов и фактор индукции) приведены на рис. 12. Удалось идентифицировать химерные комбинаторные промоторы с особенно сильной и специфической индуцируемостью, фактор индукции и активность которых выше, чем у химерных промоторов, состоящих только из повторов отдельных цис-регуляторных элементов (рис. 13). Химерные комбинаторные промоторы с самыми высокими факторами индукции и сильнейшей активностью приведены в таблице 3.
Таблица 3: Сочетания патоген-индуцируемых одиночных последовательностей с максимальными коэффициентами индукции и индуцированными действиями.
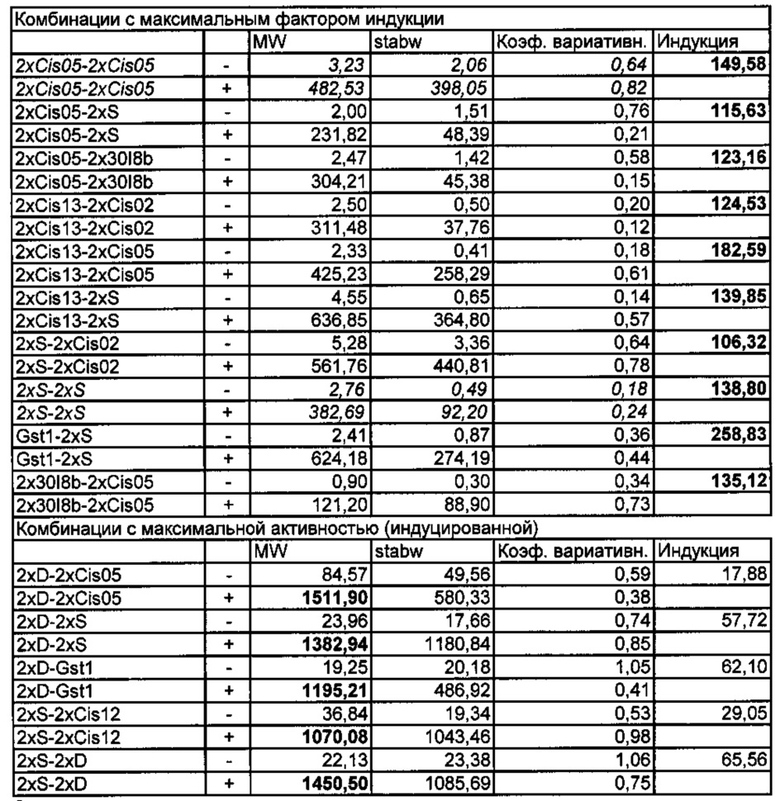
Ссылки
Bailey, T.L., С.Elkan (1994). "Fitting a mixture model by expectation maximization to discover motifs in biopolymers." Proc Int Conf Intell Svst Mol Biol. 2:28-36.
Bulow, L, M. Schindler, et al. (2007). "PathoPlant: a platform for microarray expression data to analyze co-regulated genes involved in plant defense responses." Nucleic Acids Res 35 (Database issue): D841-845.
Che, D., S. Jensen, et al. (2005). "BEST: binding-site estimation suite of tools." Bioinformatics 21 (12): 2909-2911.
Clough, S.J. and Bent, A.F. (1998)." Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana." Plant J. 16(6):735-43.
Eulgem, T., P. J. Rushton, et al. (2000). "The WRKY superfamily of plant transcription factors." Trends Plant Sci 5(5):199-206.
Gorlich, D. and I. W. Mattaj (1996). "Nucleocvtoplasmic transport." Science 271(5255):1513-1518.
Gurr, S. J. and P. J. Rushton (2005). "Engineering plants with increased disease resistance: how are we going to express it?" Trends Biotechnol 23(6):283-290.
Hahlbrock, K., D. Scheel, et al. (1995). "Oligopeptide elicitor-mediated defense gene activation in cultured parsley cells." Proc Natl Acad Sci USA 92(10):4150-4157.
Hicks, G.R., H. M. Smith, et al. (1995). "Three classes of nuclear import signals bind to plant nuclei." Plant Phvsiol 107(4):1055-1058.
Himmelbach, Α., L. Liu et al. (2010). "Promoters of the Barley Germin-Like GER4 Gene Cluster Enable Strong Transgene Expression in Response to Pathogen Attack" The Plant Cell 22(3):937-952.
Howies, P., javascript:popRef('al') G. Lawrence, et al. (2005). "Autoactive Alleles of the Flax L6 Rust Resistance Gene Induce Non-Race-Specific Rust Resistance Associated with the Hypersensitive Response." Molecular Plant-Microbe Interactions 18(6):570-582.
Humphry, M., P. Bednarek, et al. (2010). "A regulon conserved in monocot and dicot plants defines a functional module in antifungal plant immunity." Proc Natl Acad Sci U S A 107(50):21896-21901.
Jefferson, R.Α., Kavanagh, T. A & Bevan, M.W. (1987). "GUS fusions, 8-glucuronidase as a sensitive and versatile gene fusion marker in higher plants." EMBO J 6(13):3901-3907.
Kirsch, С., M. Takamiya-Wik, et al. (2000) "A novel regulatory element involved in rapid activation of parsley ELI7 gene family members by fungal elicitor or pathogen infection." Mol Plant Pathol. 1(4):243-51.
Kirsch, С., Ε. Logemann, et al. (2001) "A highly specific pathogen-responsive promoter element from the immediate-early activated CMPG1 gene in Petroselinum crispum." Plant J. 26(2):217-27.
Kovalchuk, N., M. Li, et al. (2010). "Defensin promoters as potential tools for engineering disease resistance in cereal grains." Plant Biotechnol J 8(1):47-64.
Lebel, E., P. Heifetz, et al. (1998). "Functional analysis of regulatory sequences controlling PR-1 gene expression in Arabidopsis." Plant J 16(2):223-233.
Liu, X., D. L. Brutlag, et al. (2001). "BioProspector: discovering conserved DNA motifs in upstream regulatory regions of co-expressed genes." Рас Svrnp Biocomput: 127-138.
Morris, K., S. A. MacKerness, et al. (2000). "Salicylic acid has a role in regulating gene expression during leaf senescence." Plant J 23(5):677-685.
Rushton, P.J., A. Reinstadler, et al. (2002). "Synthetic plant promoters containing defined regulatory elements provide novel insights into pathogen- and wound-induced signaling." Plant Cell 14(4):749-762.
Rushton, P. J., J. T. Torres, et al. (1996). "Interaction of elicitor-induced DNA-binding proteins with elicitor response elements in the promoters of parsley PR1 genes." EMBO J 15(20):5690-5700.
Schatz, G. and B. Dobberstein (1996). "Common principles of protein translocation across membranes." Science 271 (5255):1519-1526.
Schumacher, S.В., О. Van den Hauwe, et al. (2003). "Development of a dual luciferase reporter screening assay for the detection of synthetic glucocorticoids in animal tissues." Analyst 128(12):1406-1412.
Strittmatter, G., G. Gheysen, et al. (1996). "Infections with various types of organisms stimulate transcription from a short promoter fragment of the potato gst1 gene." Mol Plant Microbe Interact 9(1):68-73.
Venter, M. (2007). "Synthetic promoters: genetic control through cis engineering." Trends Plant Sci 12(3):118-124.
Verner, K. and G. Schatz (1988). "Protein translocation across membranes." Science 241(4871):1307-1313.
Wan, J., X. C. Zhang, et al. (2008). "A LysM receptor-like kinase plays a critical role in chitin signaling and fungal resistance in Arabidopsis." Plant Cell 20(2):471-481.
Zipfel, C., G. Kunze, et al. (2006). "Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation." ÇeH 125(4):749-760.
Zipfel, C., S. Robatzek, et al. (2004). "Bacterial disease resistance in Arabidopsis through flagellin perception." Nature 428(6984):764-767.
WO 00/29592 (Max-Planck-Gesellschaft zur Forderung der Wissenschaften e.V.). Chimeric promoters capable of mediating gene expression in plants upon pathogen infection and uses thereof.
WO 02/50293 (Max-Planck-Gesellschaft zur Forderung der Wissenschaften e.V.). Pflanzen mit verbesserter Widerstandskraft.
WO 03/00898 (Syngenta Participations AG). Plant genes involved in defense against pathogens.
WO 07/147395 (KWS SAAT AG). Pathogen induzierbarer synthetischer Promotor
WO 10/079430 (U. Bonas et al.). Modular DNA-binding domains and methods of use.
WO 11/072246 (Regents of the University of Minnesota; Iowa State University Research Foundation Inc.). TAL effector-mediated DNA modification.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПАТОГЕН-ИНДУЦИРУЕМЫЙ СИНТЕТИЧЕСКИЙ ПРОМОТОР | 2007 |
|
RU2406761C2 |
| АУТОАКТИВИРУЮЩИЙСЯ БЕЛОК УСТОЙЧИВОСТИ | 2006 |
|
RU2375453C2 |
| БОЛЕЗНЕУСТОЙЧИВЫЕ РАСТЕНИЯ | 2006 |
|
RU2393227C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2675524C2 |
| СПОСОБ ПОВЫШЕНИЯ РЕЗИСТЕНТНОСТИ ТРАНСГЕННЫХ РАСТЕНИЙ К ПАТОГЕННЫМ ВОЗДЕЙСТВИЯМ | 2007 |
|
RU2428480C9 |
| ПЕСТИЦИДНЫЕ КОМПОЗИЦИИ И ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ | 2013 |
|
RU2662672C2 |
| ИСКУССТВЕННЫЙ ПРОМОТОР ДЛЯ ЭКСПРЕССИИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2003 |
|
RU2326167C2 |
| НОВЫЙ ИММУНОГЕННЫЙ ЭКСТРАКТ ГРИБОВ И ПАТТЕРН-РАСПОЗНАЮЩИЙ РЕЦЕПТОР РАСТЕНИЙ | 2013 |
|
RU2700621C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| КОНСТИТУТИВНЫЕ ПРОМОТОРЫ СОИ | 2014 |
|
RU2714724C2 |
























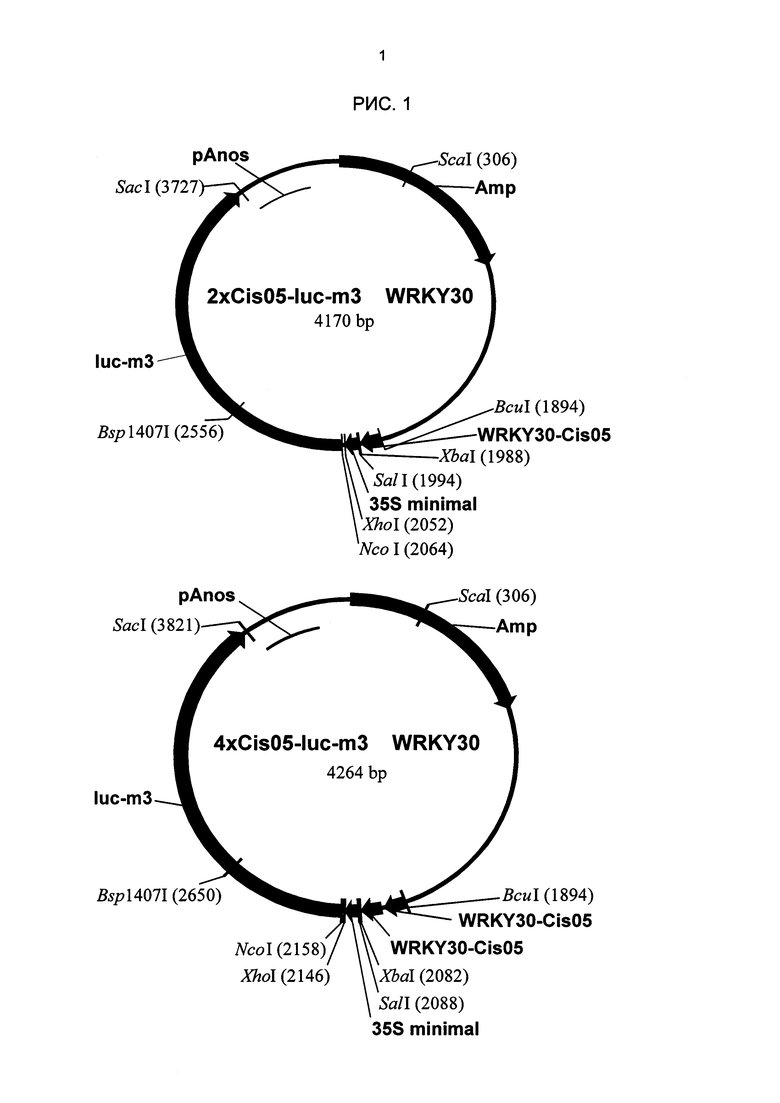
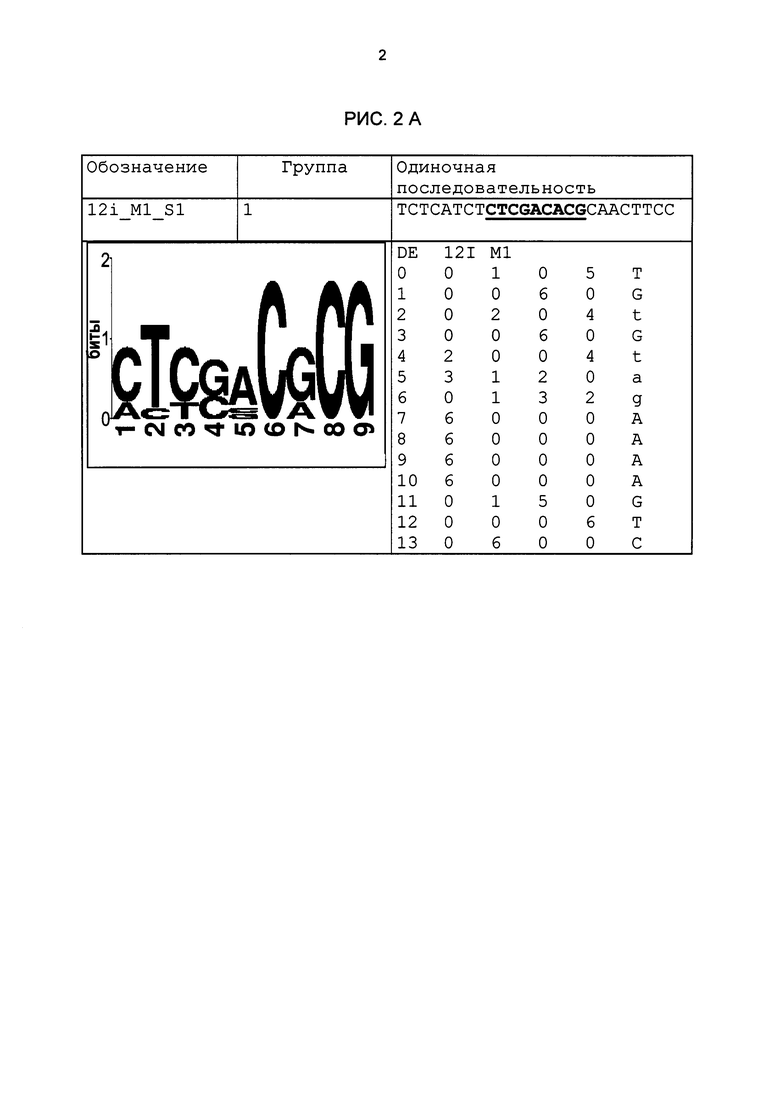

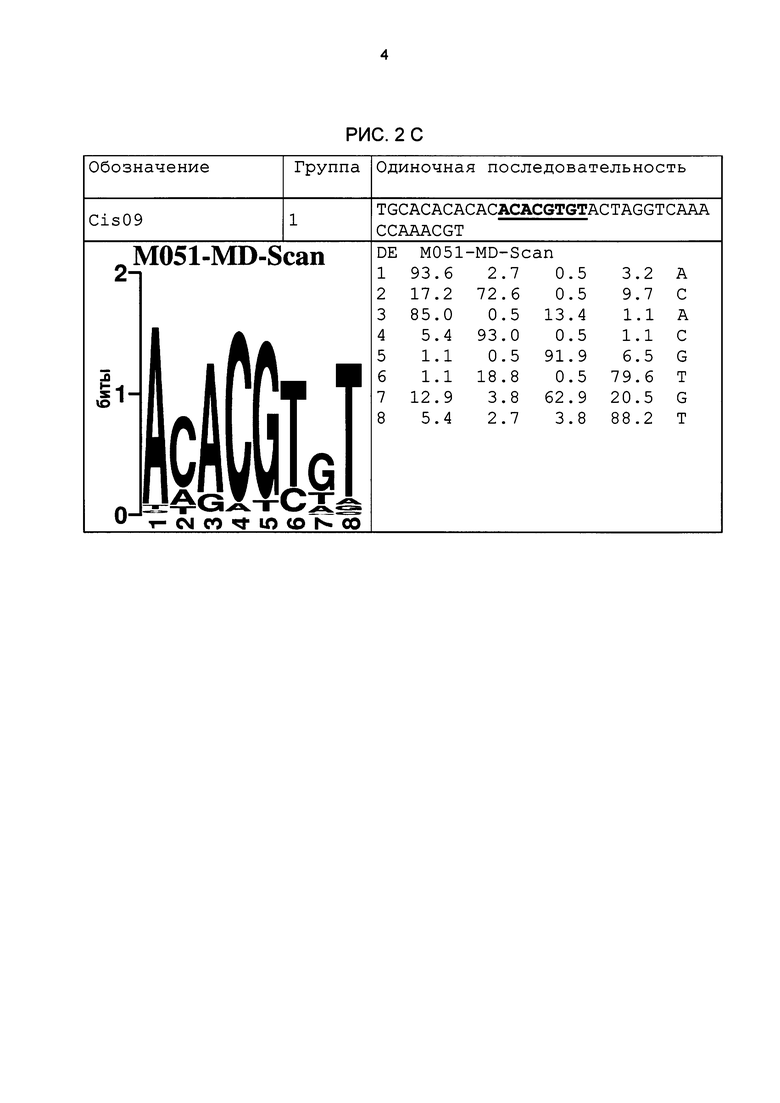


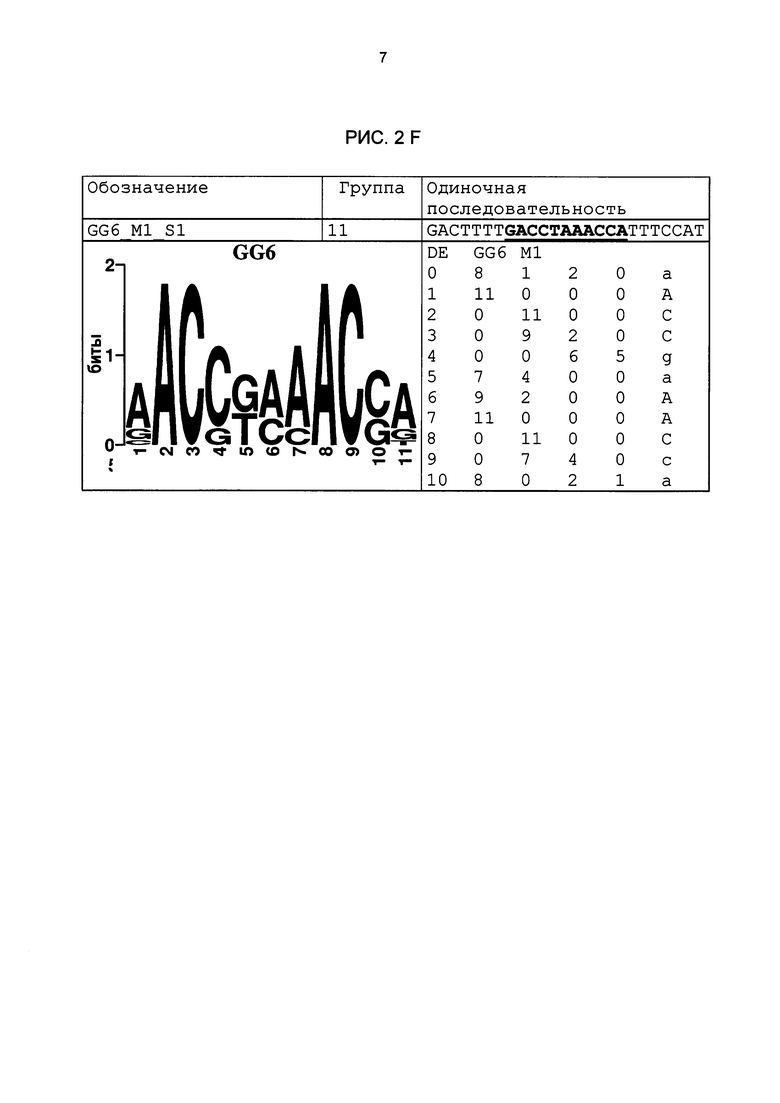
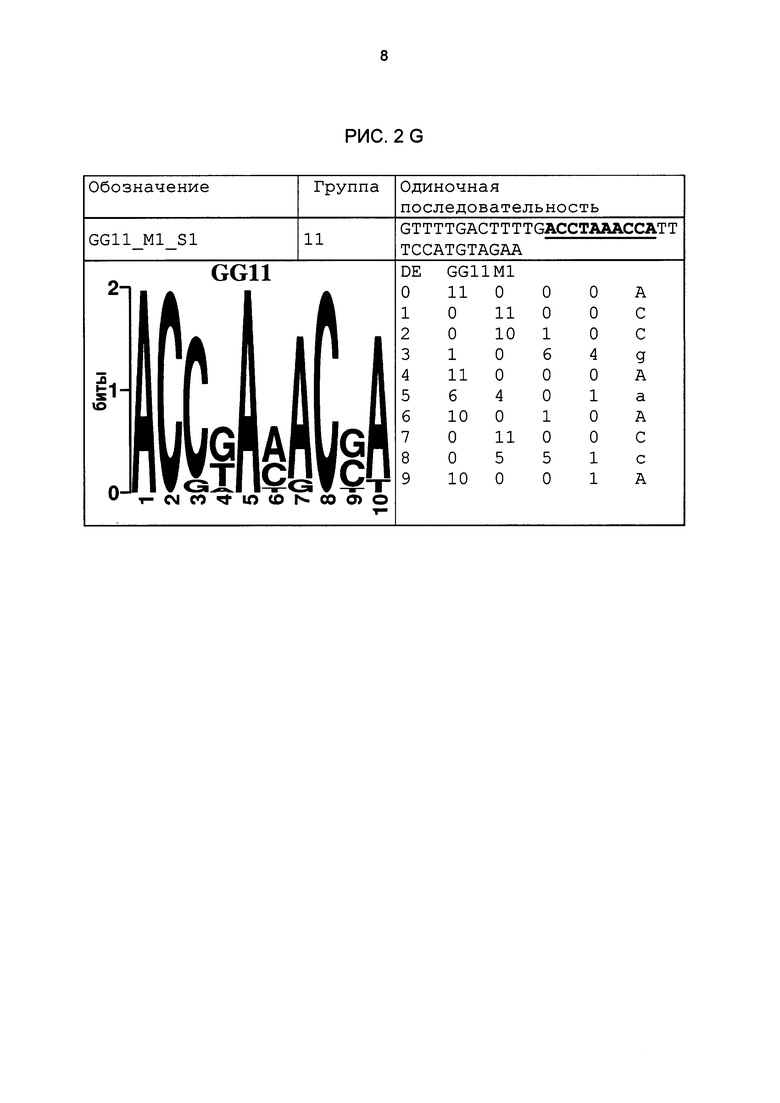
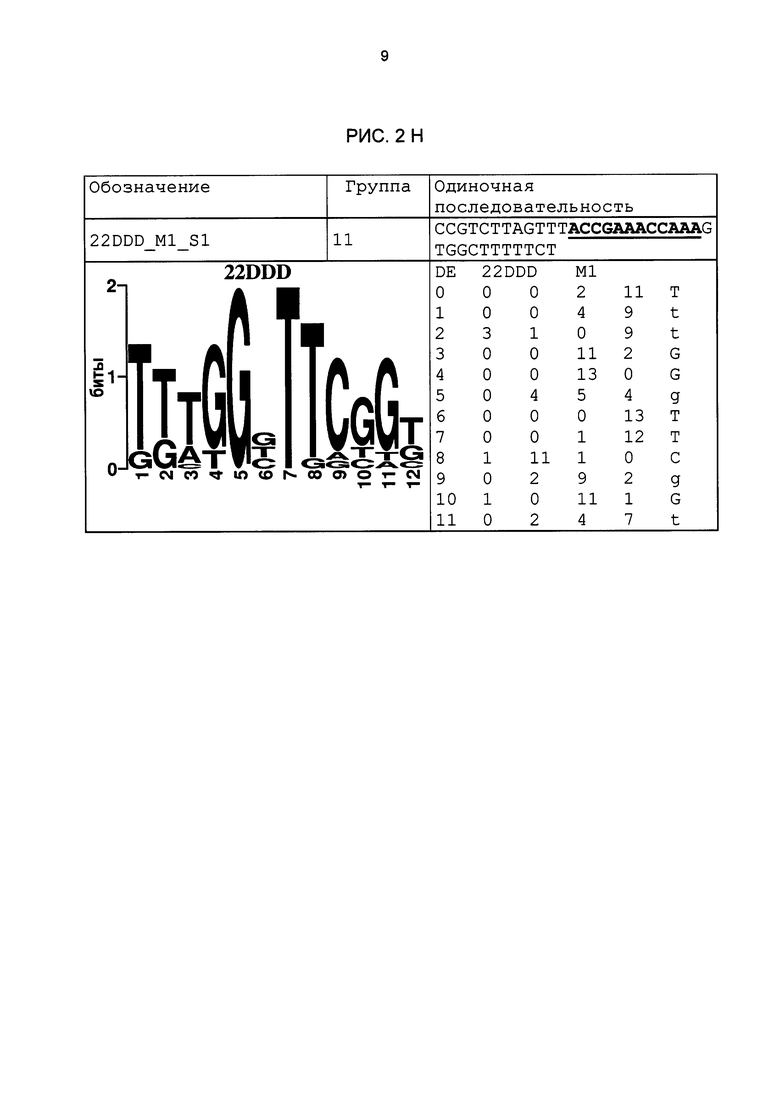



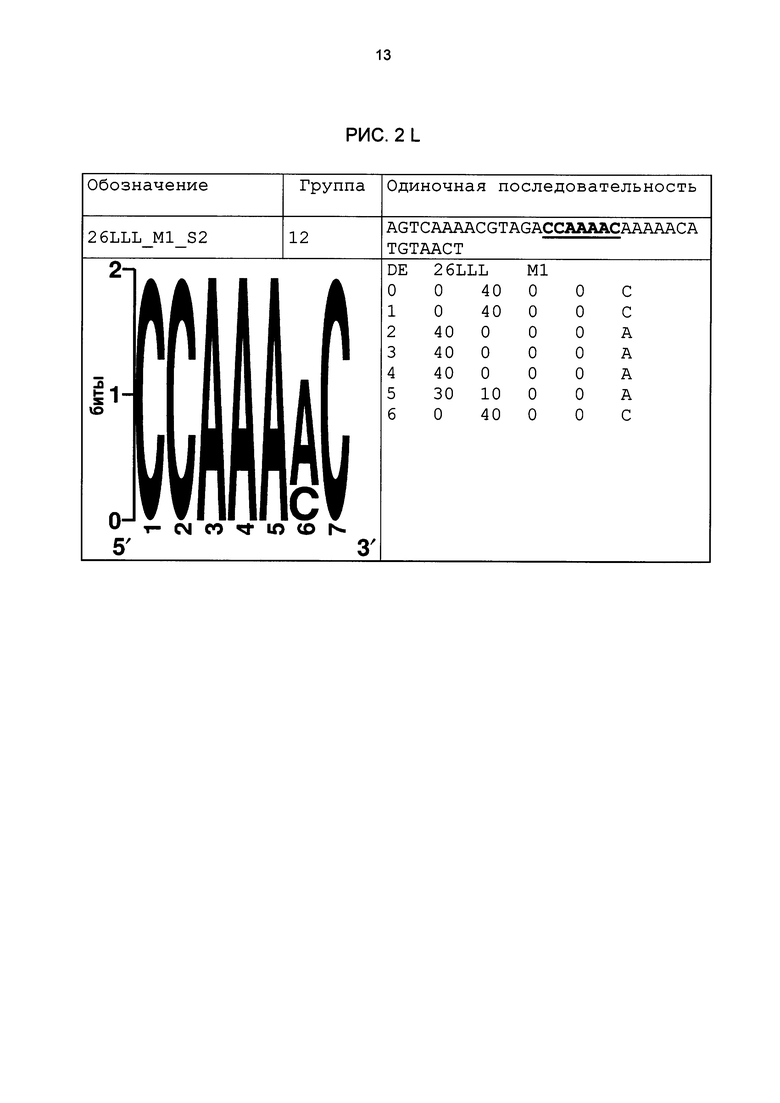


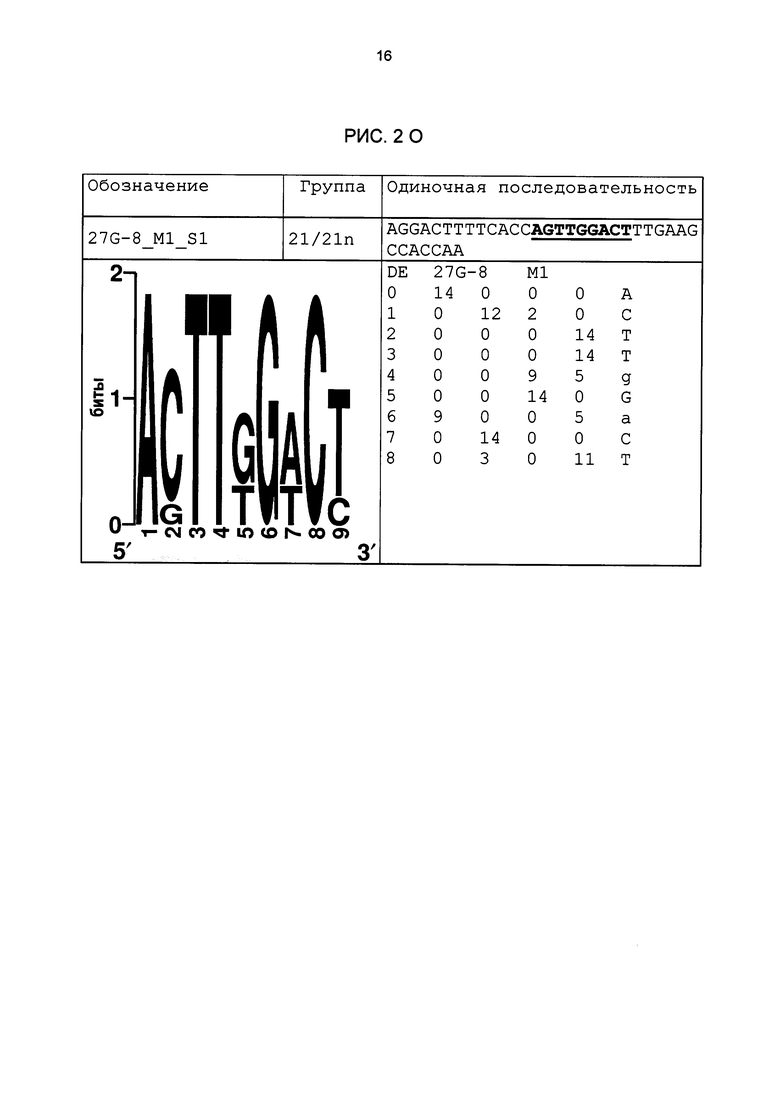

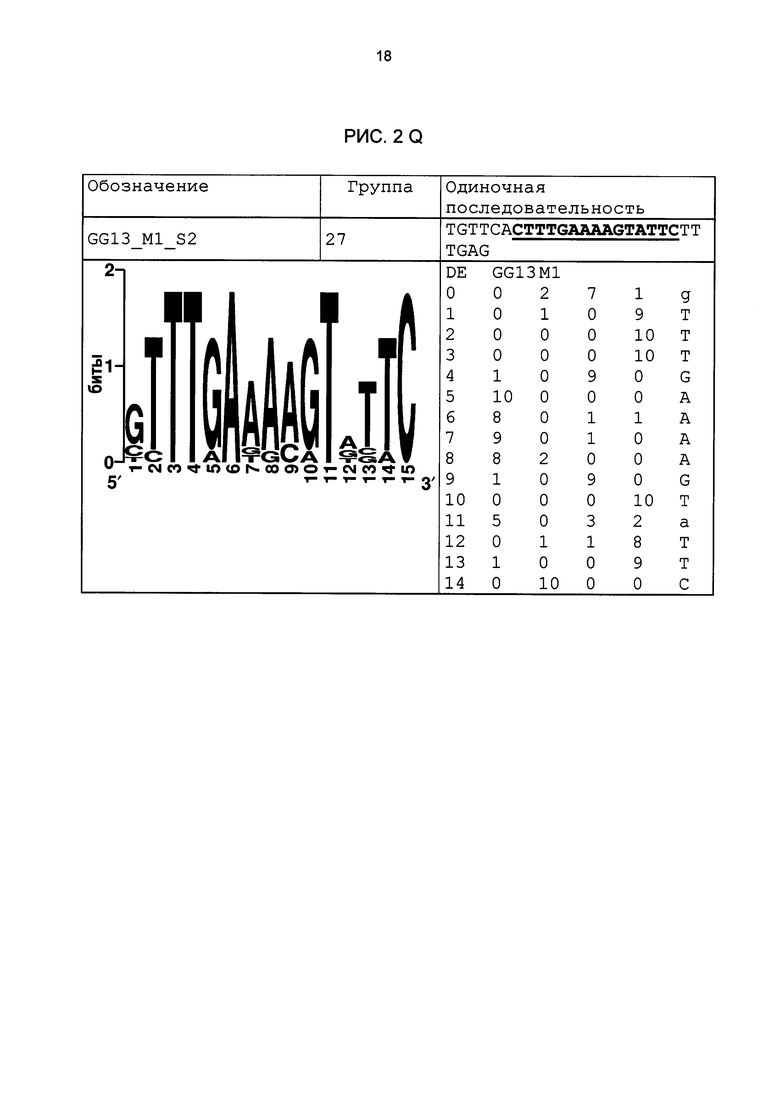
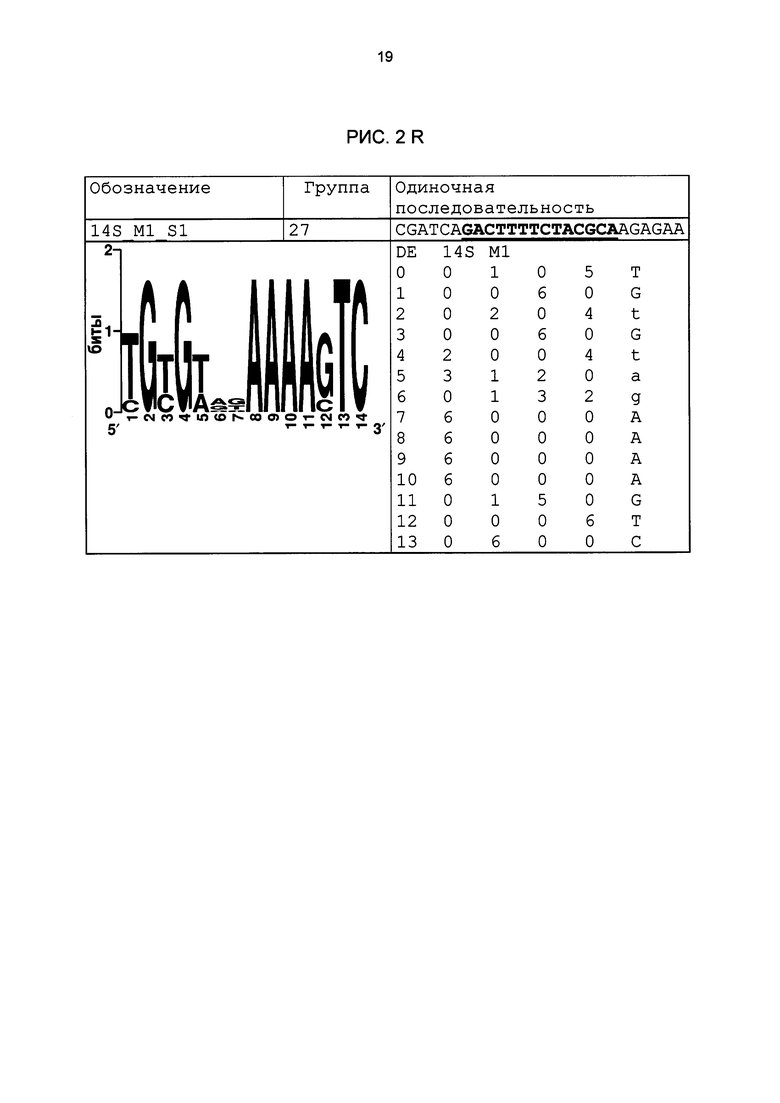

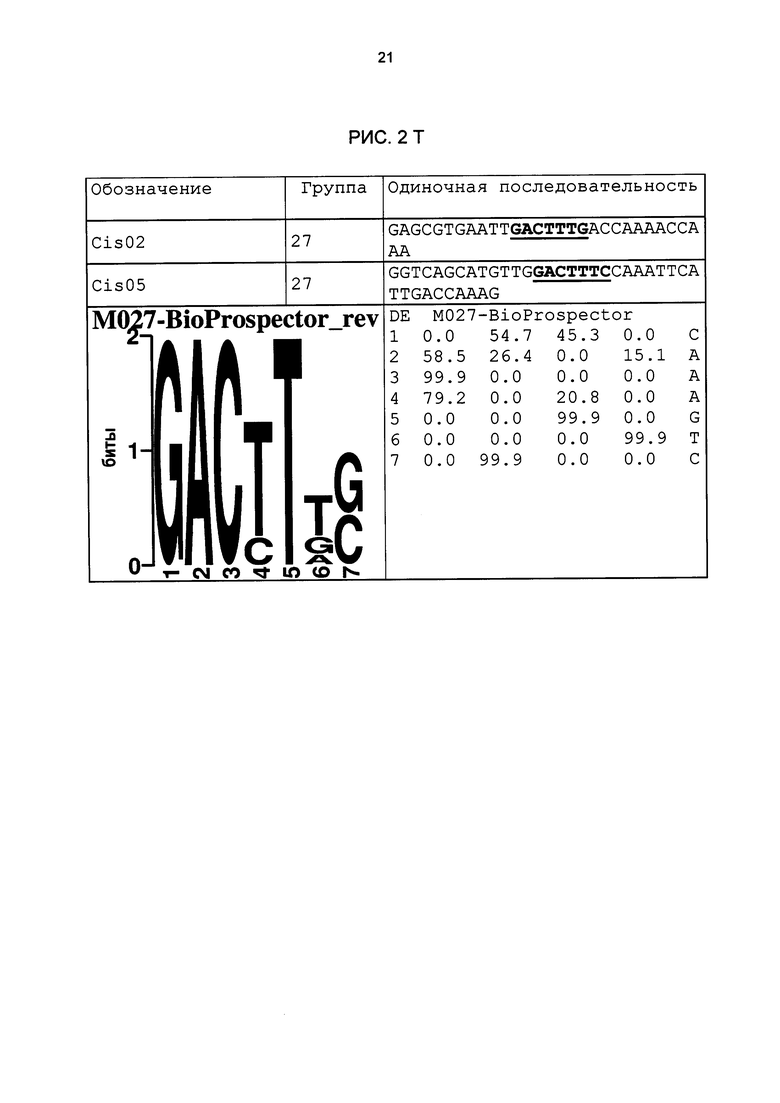
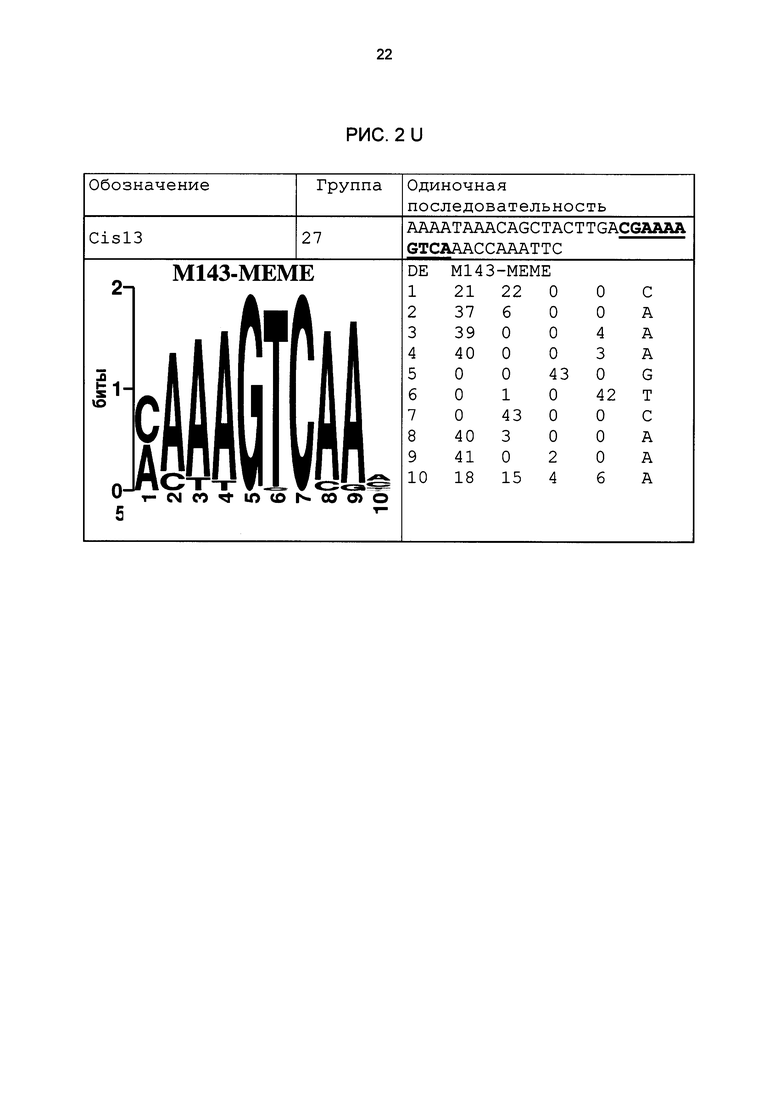

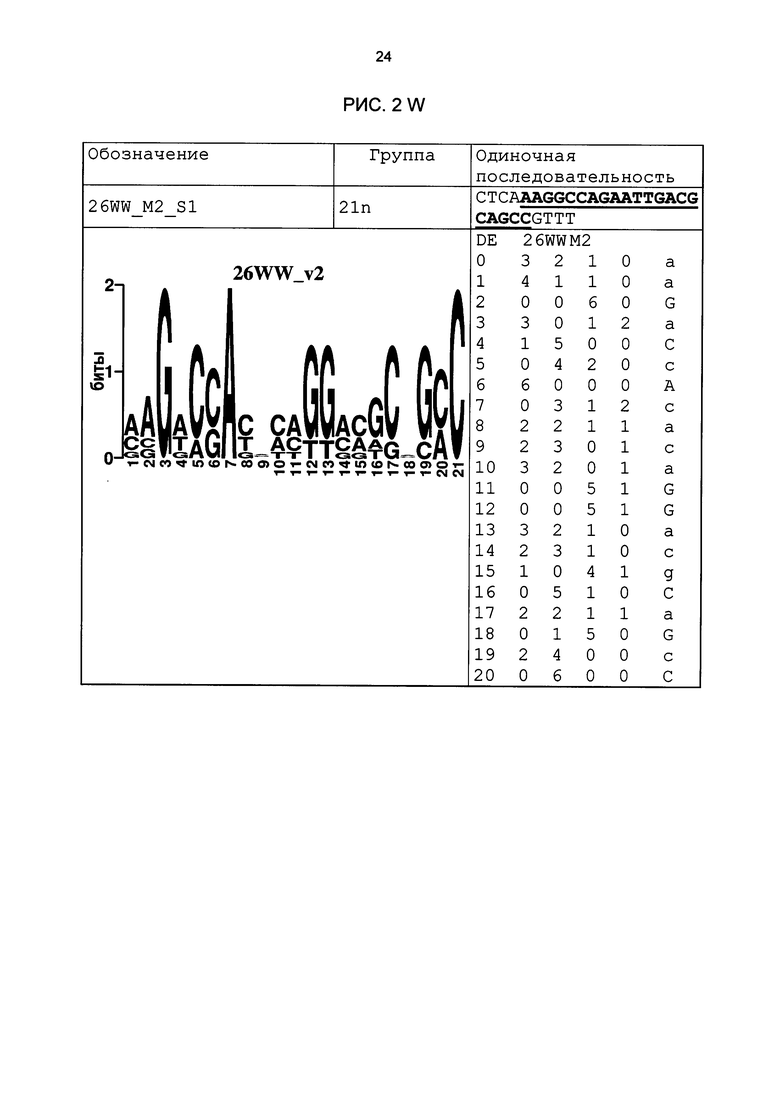
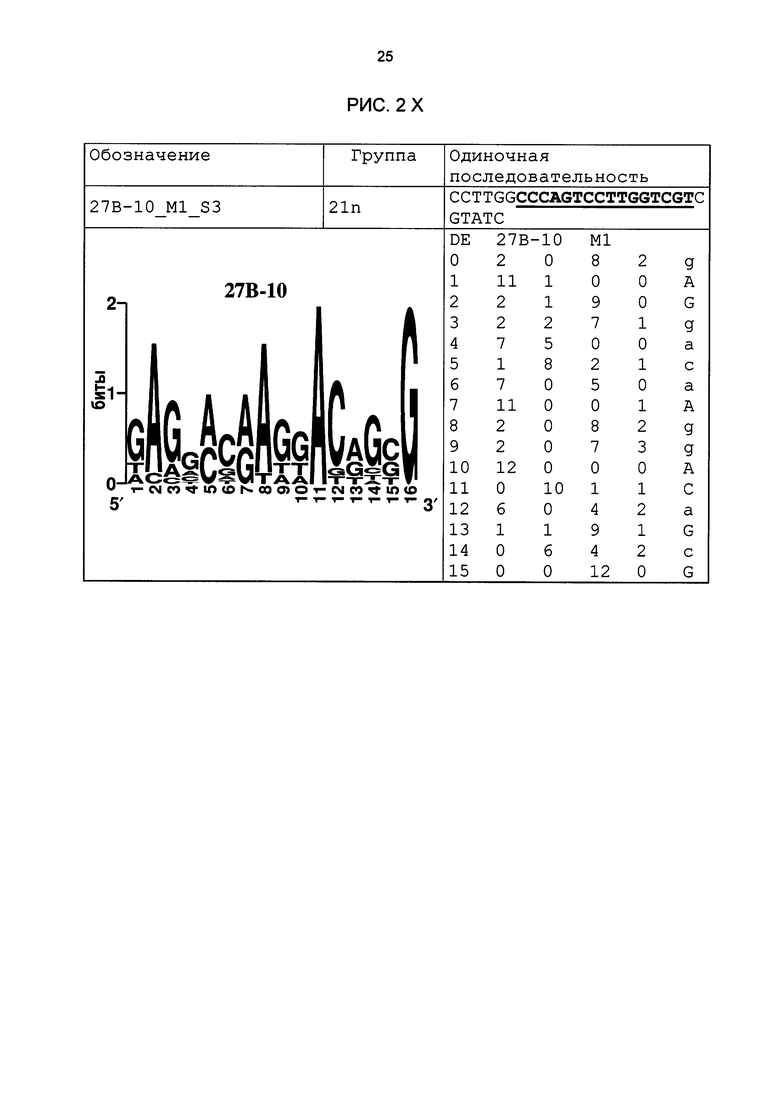
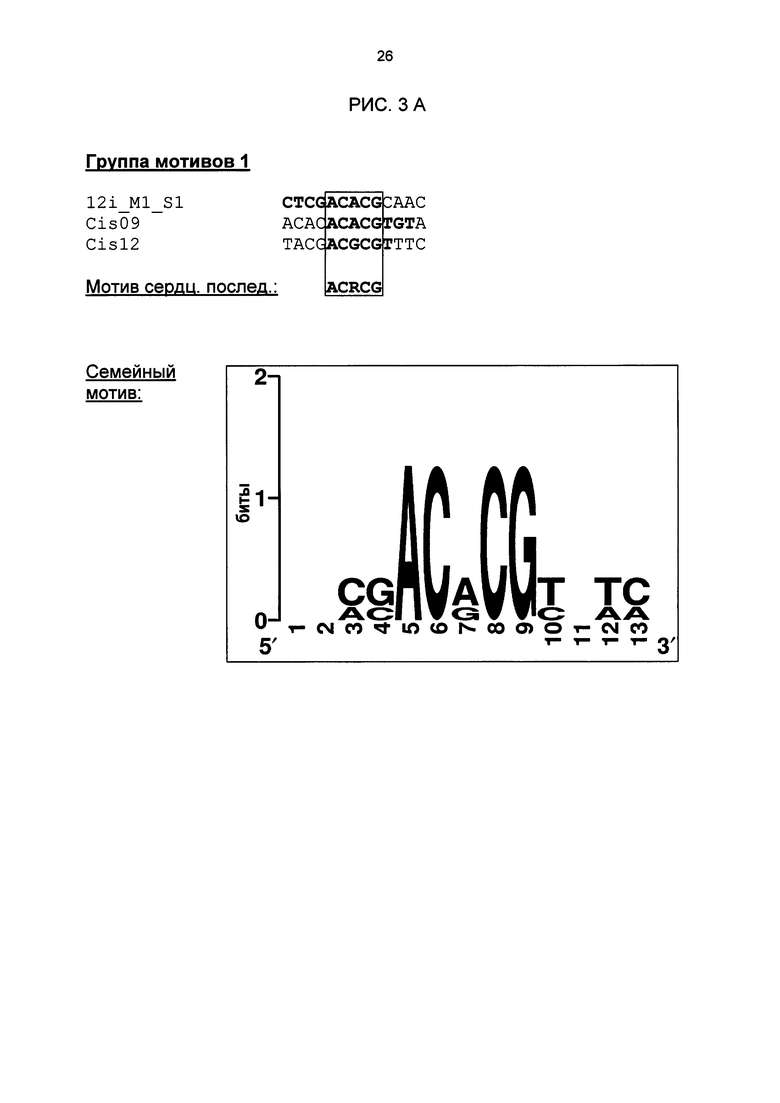
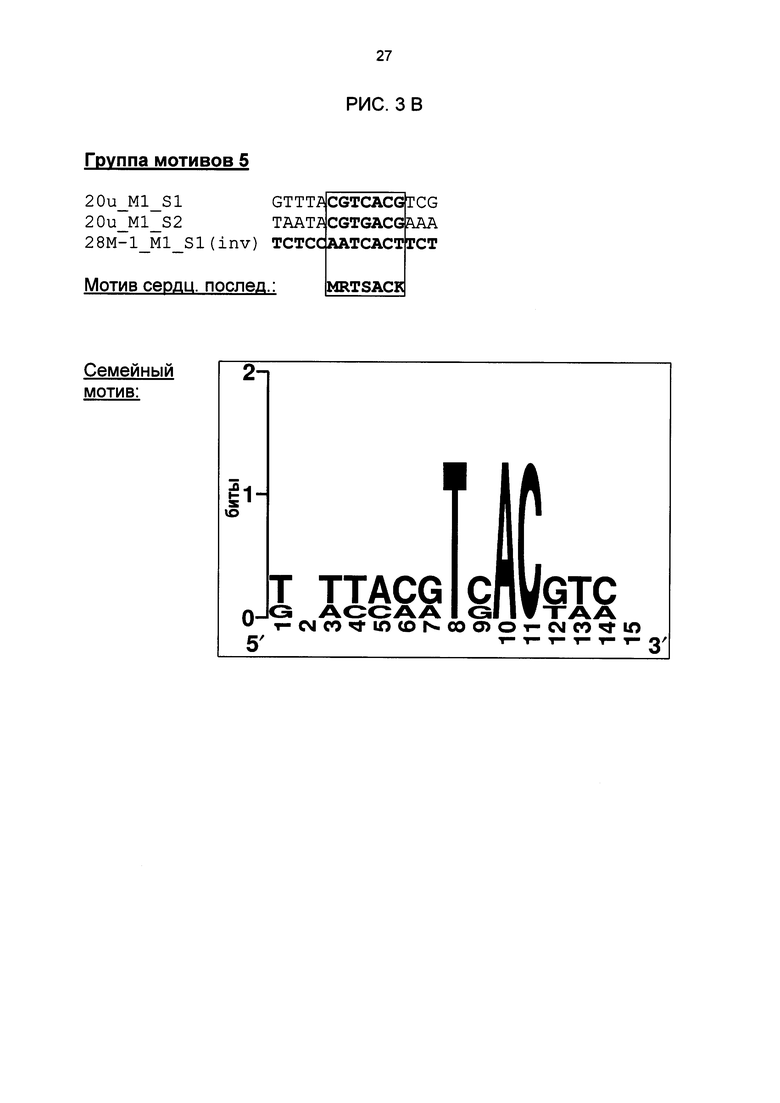

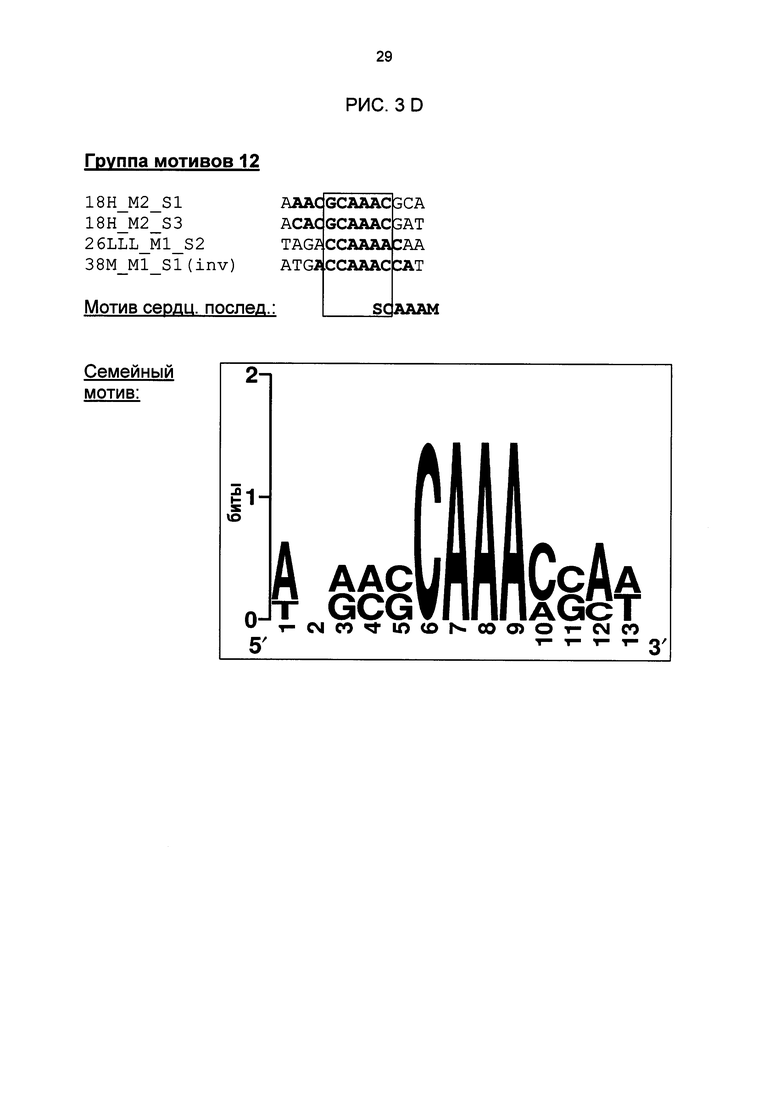

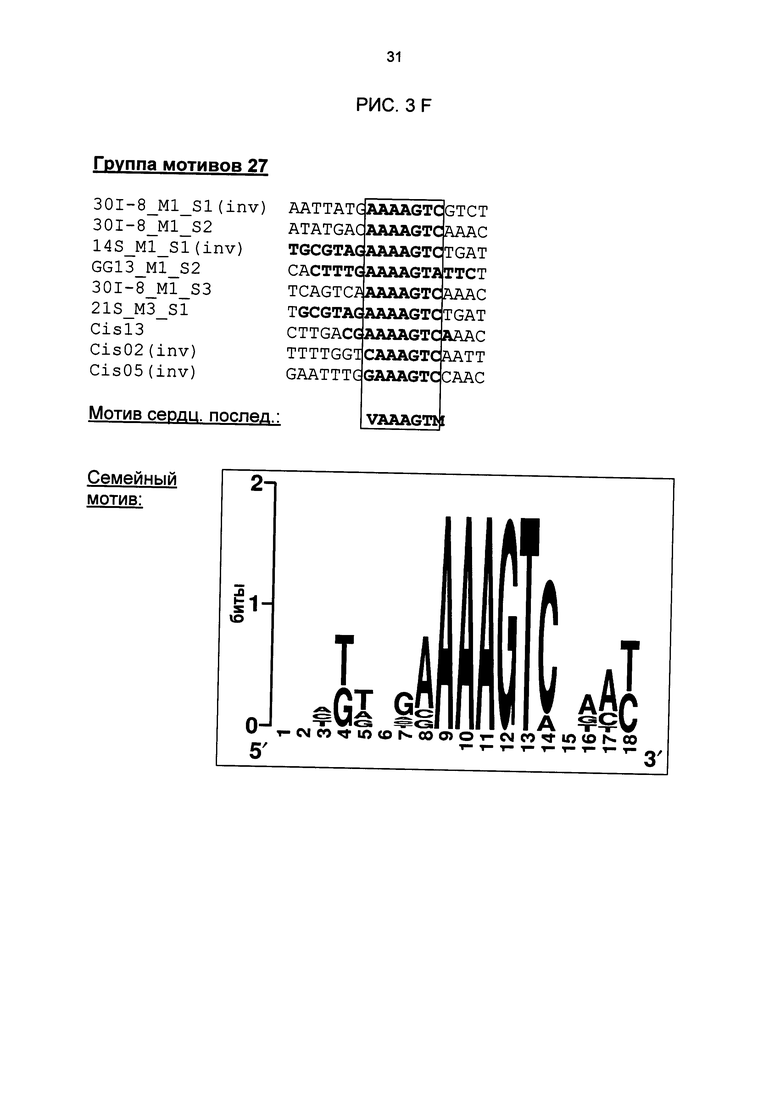
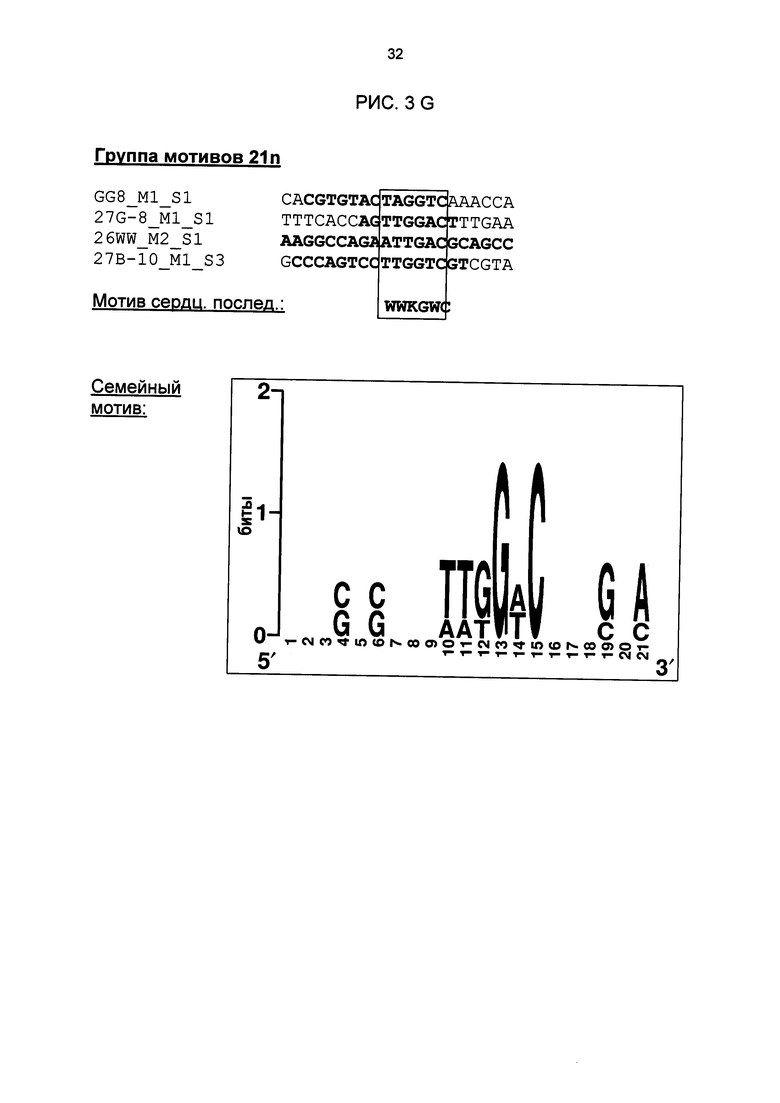
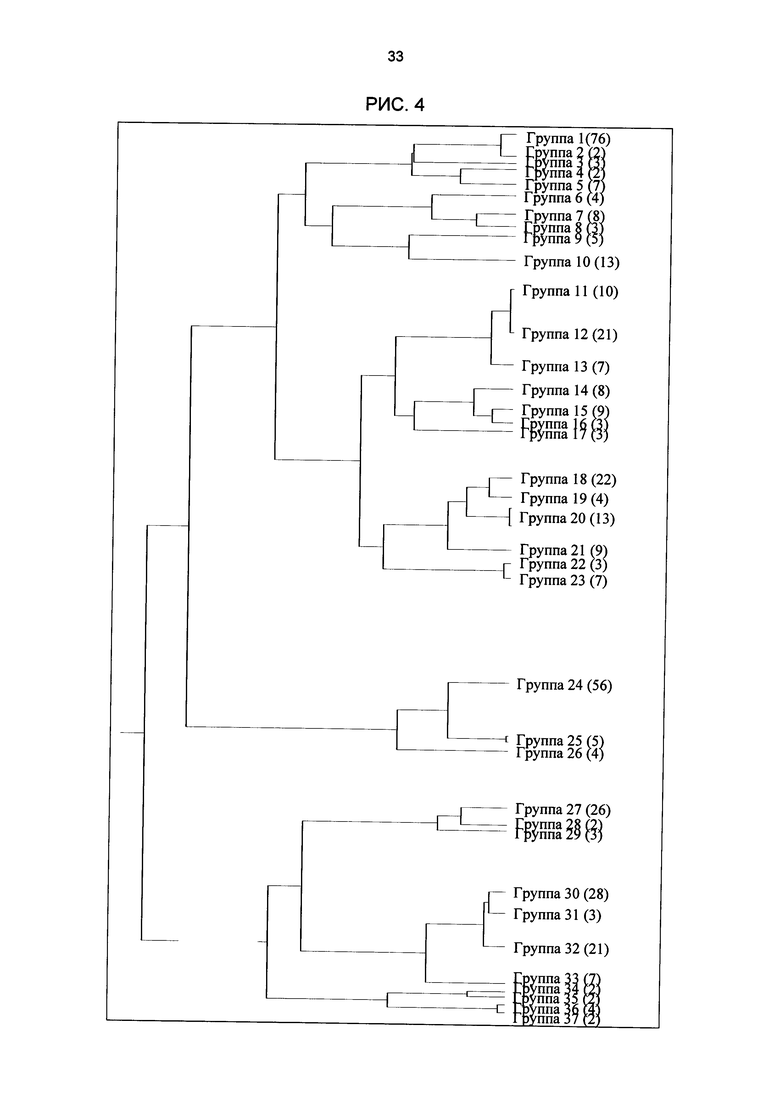
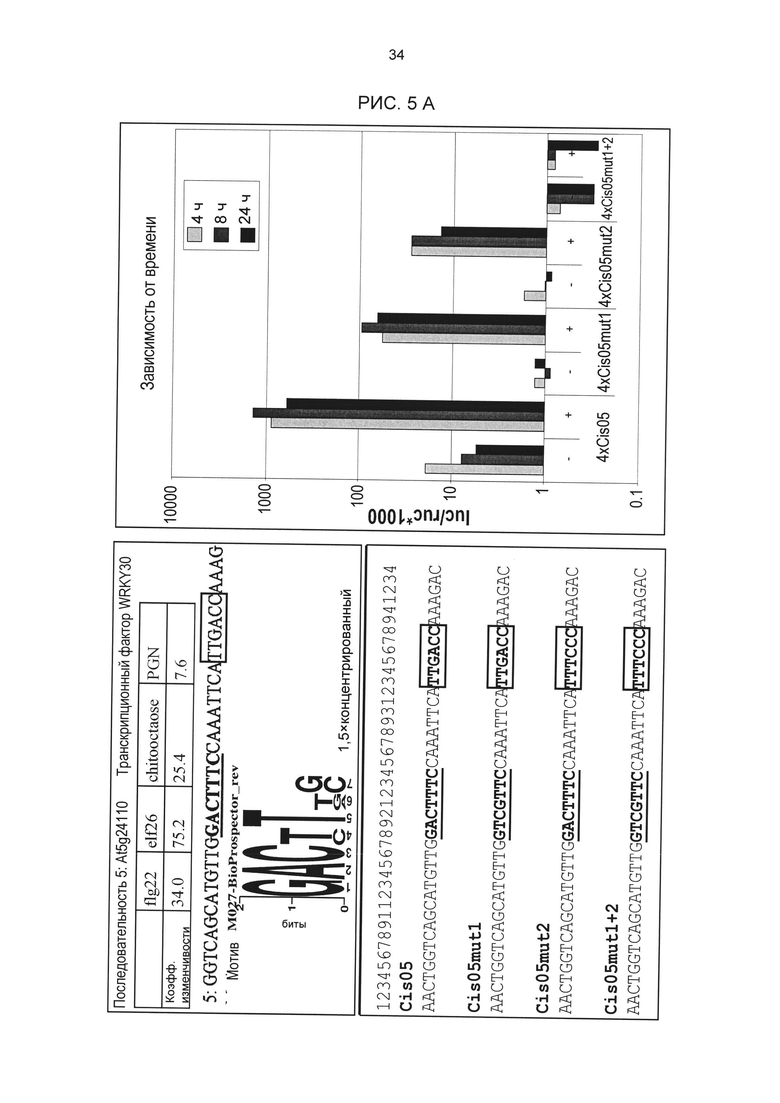
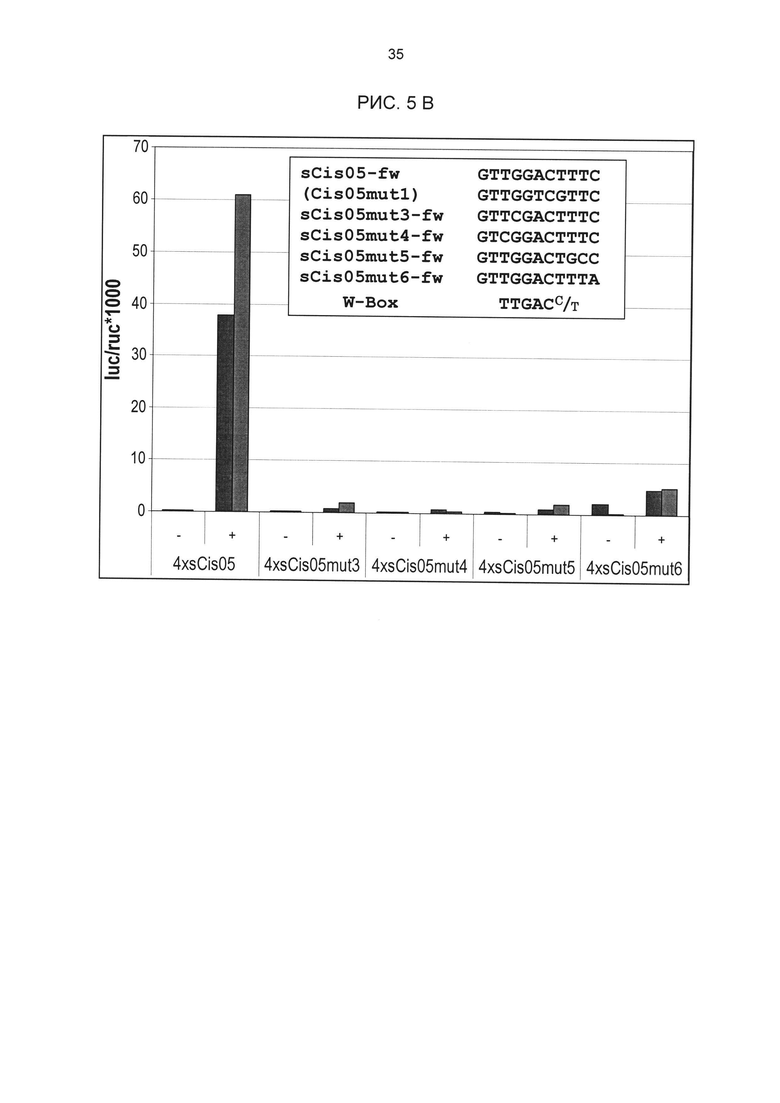
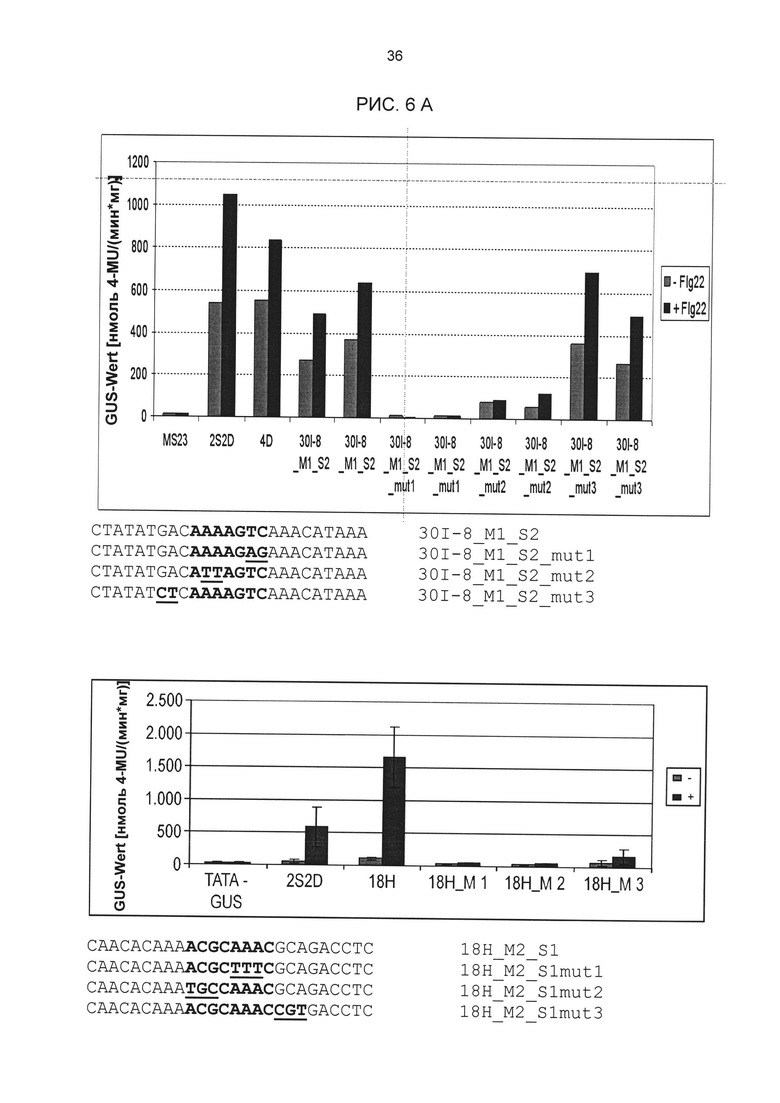
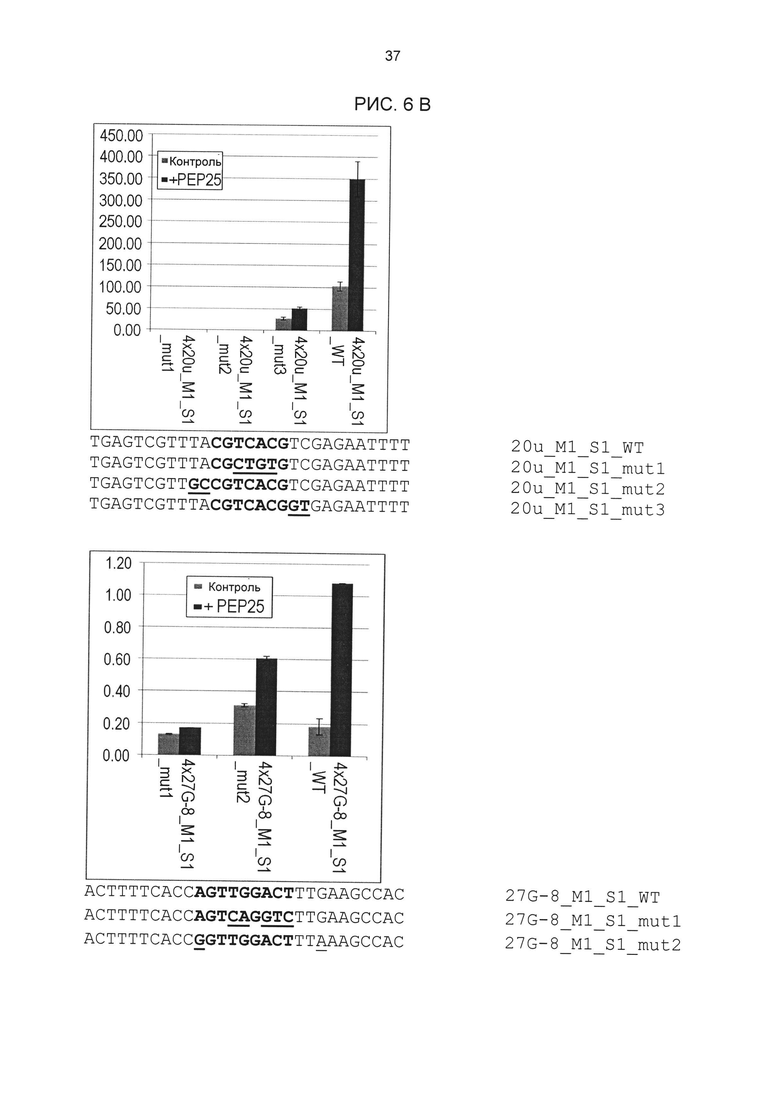
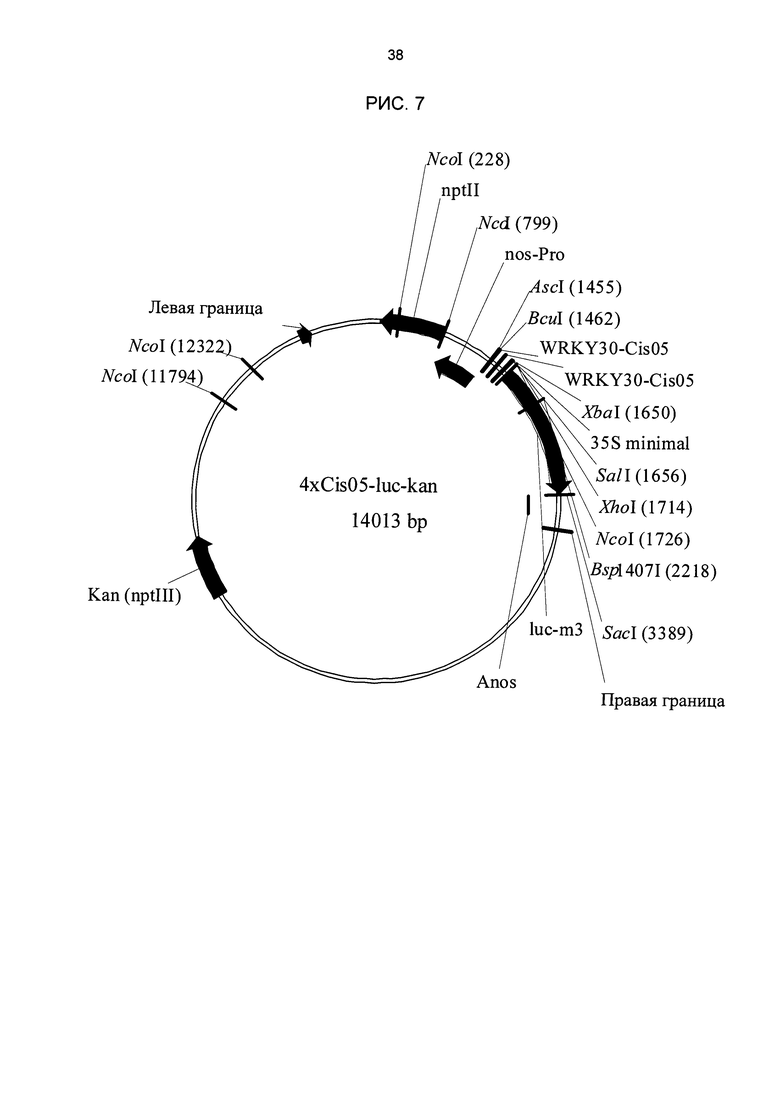
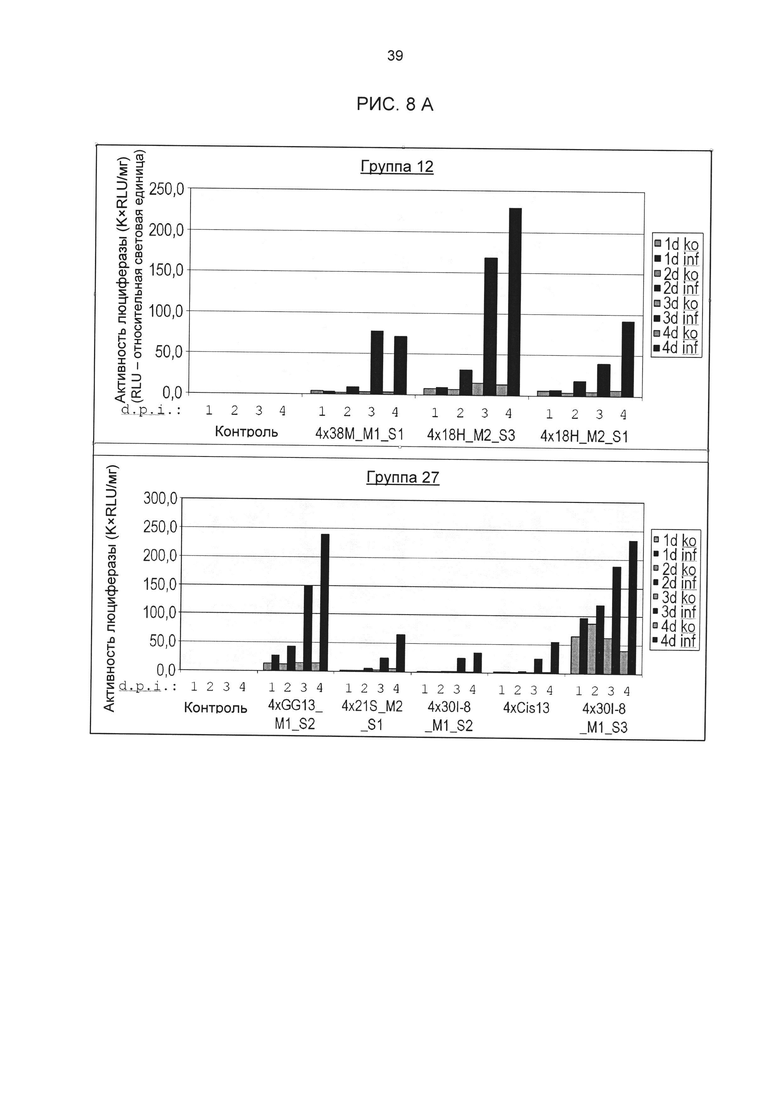
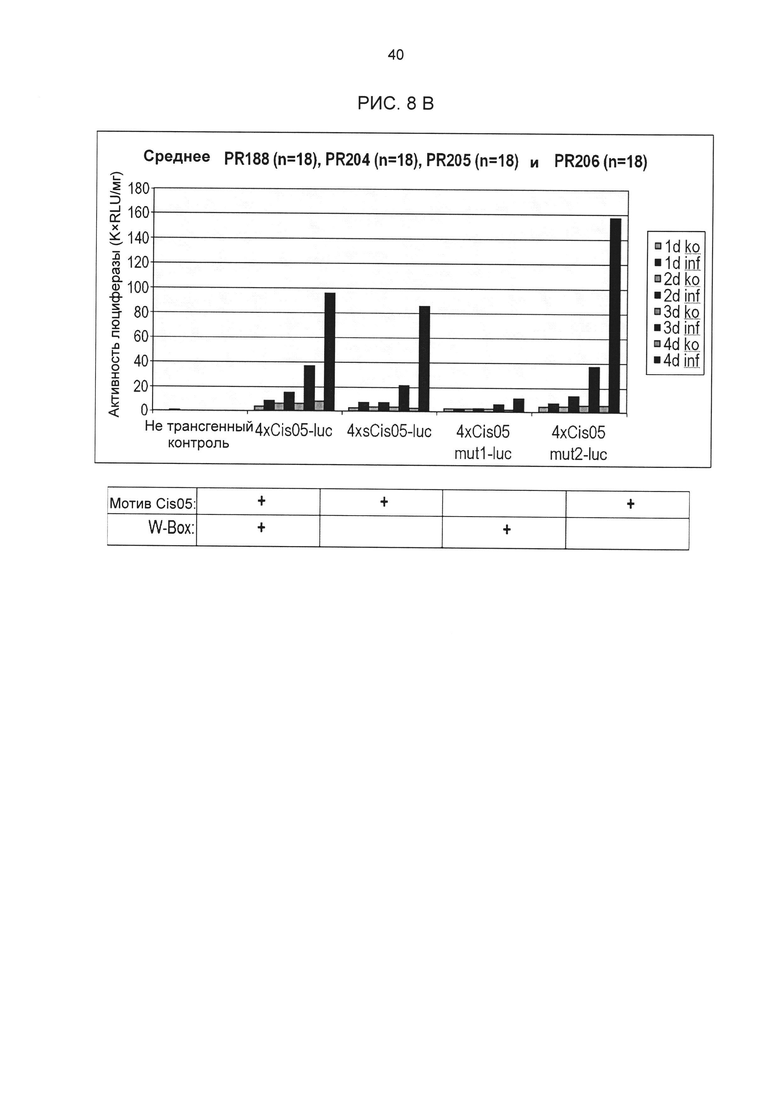
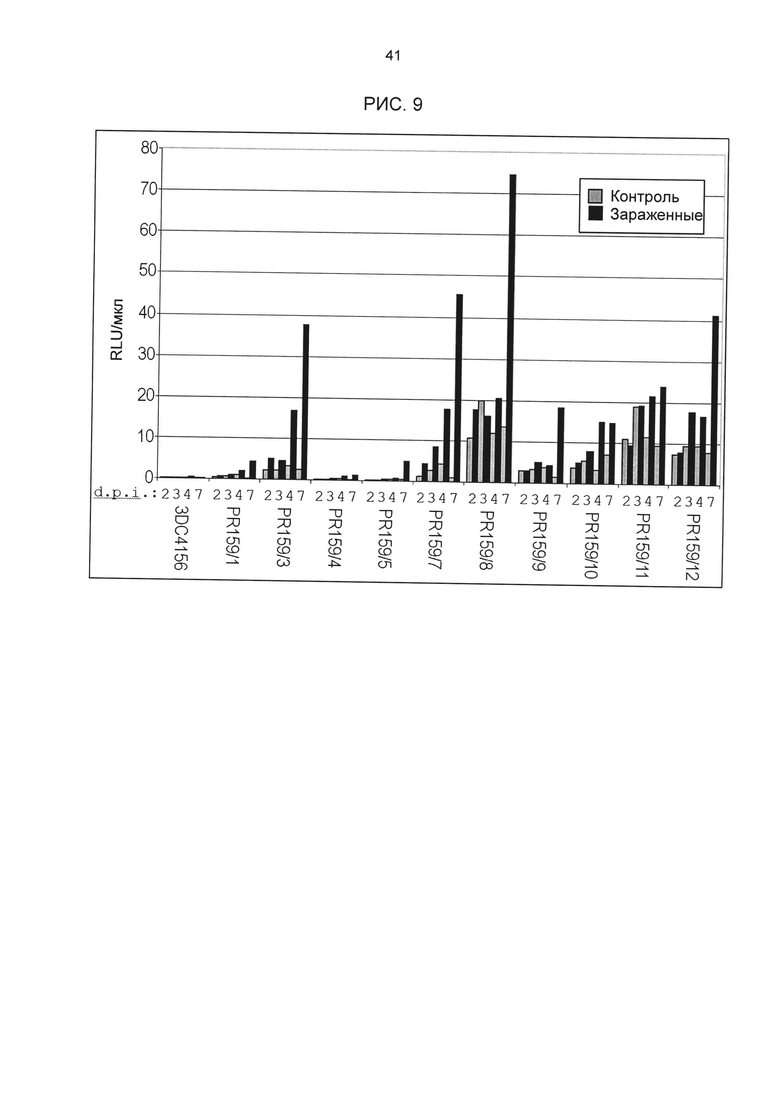
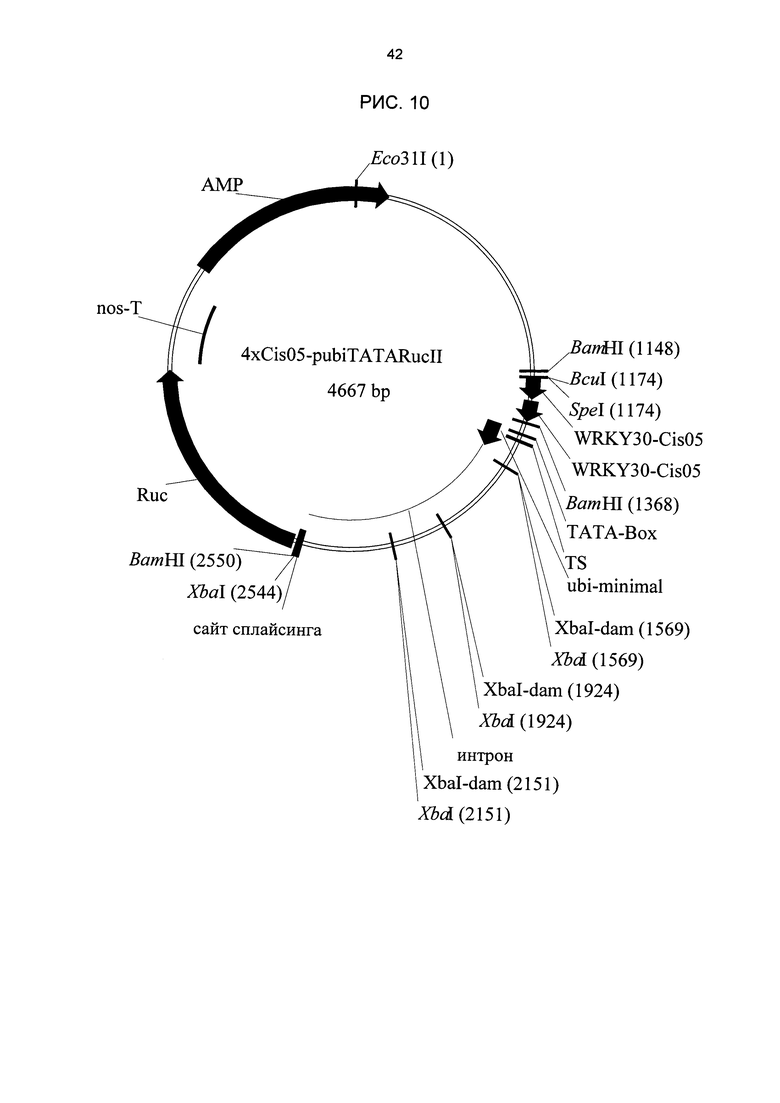
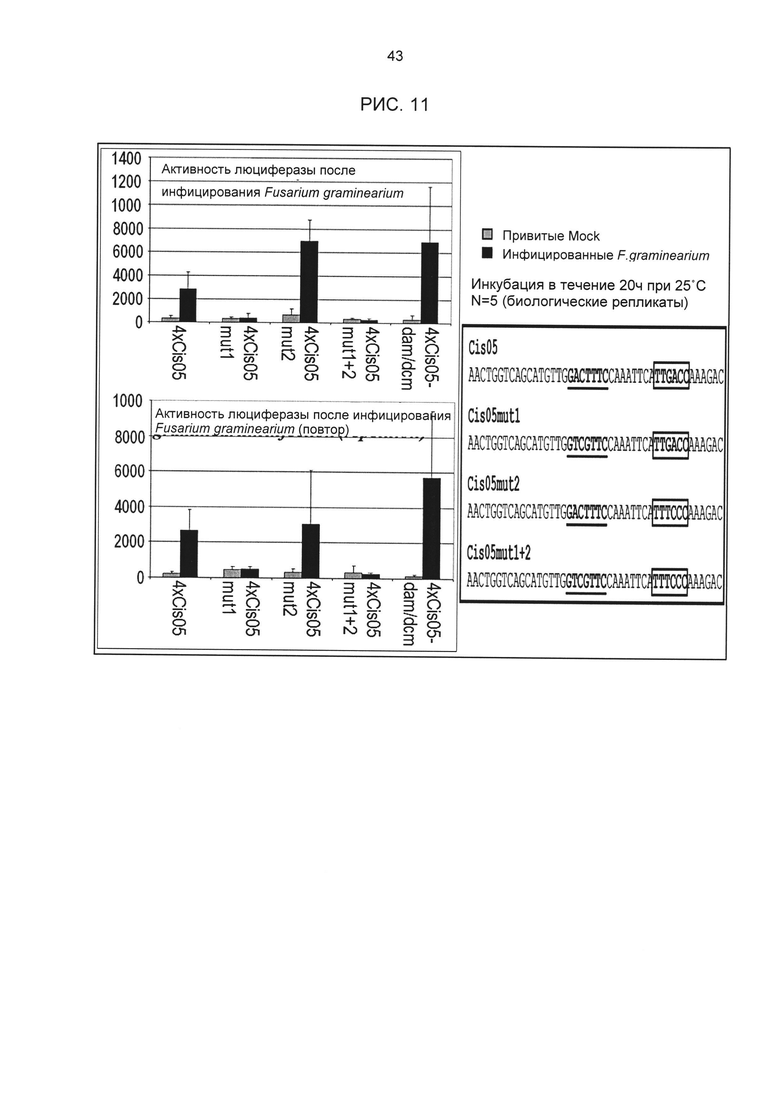
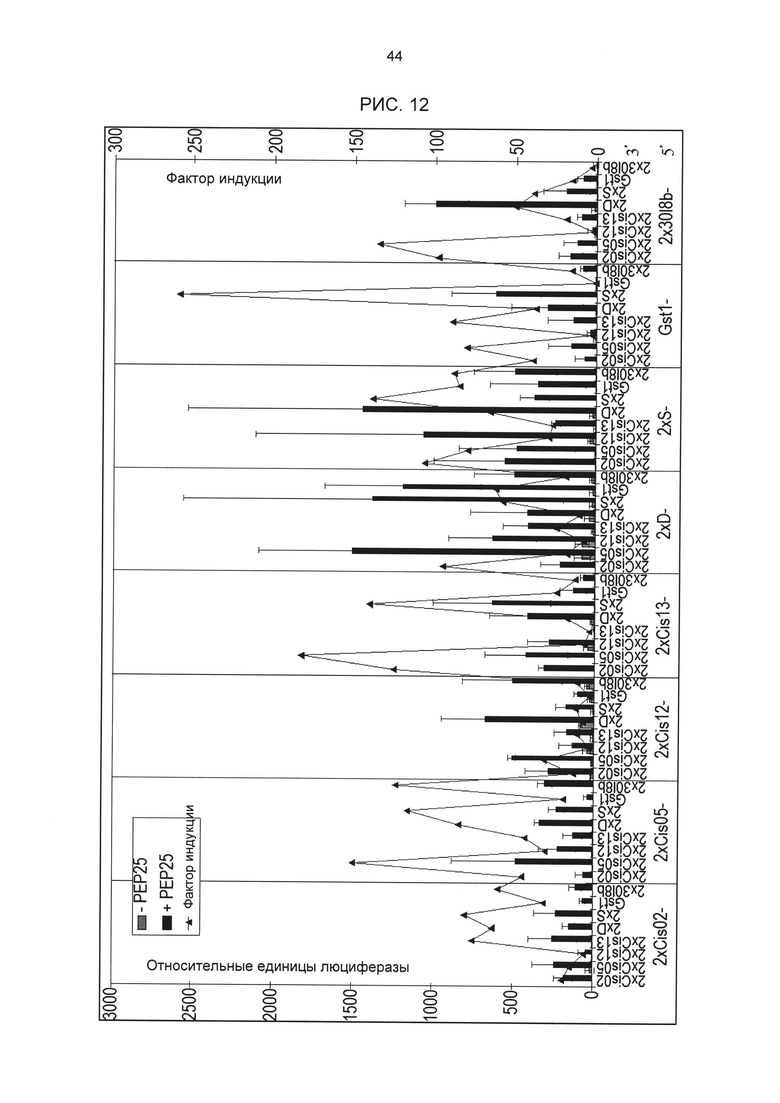
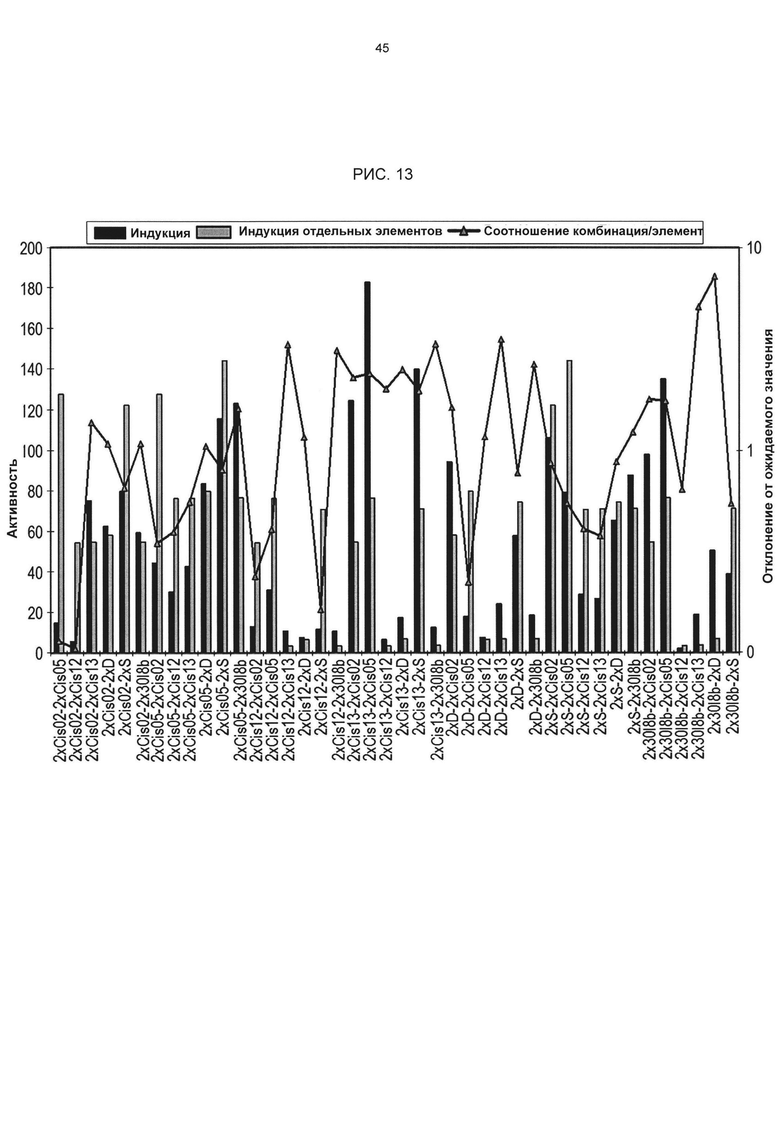



Изобретение относится к области биотехнологии, а именно к изолированным цис-регуляторным элементам для придания промотору индуцируемости патогеном, химерному промотору, обладающему индуцируемостью патогеном, рекомбинантному гену для экспрессии в растительной клетке после контакта с патогеном, экспрессирующим вектору для повышения устойчивости растения к патогенам, где патоген представляет собой грибок или оомицет. Также изобретение представляет собой прокариотическую или эукариотическую клетку для получения растения с повышенной устойчивостью к патогенам, трансгенное растение для снижения риска заражения патогеном, семя растения для культивирования растения со сниженным риском заражения, ткань растения для in vitro культивирования, часть растения для снижения риска заражения патогеном, способ создания трансгенного растения, способ патоген-индуцируемой экспрессии эндогенного гена в растительной клетке под контролем нативного промотора, растительную клетку и растение, способные к патоген-индуцируемой экспрессии эндогенного гена. Изобретение обеспечивает технологию создания трансгенного растения, в частности, стойкого к действию патогена, где патоген представляет собой грибок или оомицет. 14 н. и 3 з.п. ф-лы, 16 ил., 3 табл.
1. Изолированный цис-регуляторный элемент для придания промотору индуцируемости патогеном, представляющий собой молекулу нуклеиновой кислоты, последовательность нуклеотидов которой соответствует одному из мотивов:
a) nnhkdnnvaaagtmndhy,
b) ааасса,
c) scaaam,
d) acrcg,
e) asktgkactwkgwm,
f) knwymmrtsackwmn,
g) ccaccaa,
h) tcgtctcttc или
i) snsnnnwwkgwcnnnsnm,
где n означает любое основание; h означает а, с или t/u; d означает a, g или t/u; v означает a, g или с; r означает g или а; k означает g или t/u; s означает g или с; m означает а или с; y означает t/u или с; w означает а или t/u;
причем патоген представляет собой грибок или оомицет.
2. Изолированный цис-регуляторный элемент по п. 1, отличающийся тем, что мотив включает сердцевинную последовательность цис-регуляторного элемента.
3. Изолированный цис-регуляторный элемент для придания промотору индуцируемости патогеном, включающий молекулу нуклеиновой кислоты с последовательностью нуклеотидов согласно SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 30, SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34, SEQ ID NO: 42, SEQ ID NO: 43 или SEQ ID NO: 44, причем патоген представляет собой грибок или оомицет.
4. Химерный промотор, обладающий индуцируемостью патогеном, состоящий из минимального промотора и по меньшей мере одного цис-регуляторного элемента по любому из пп. 1-3, причем патоген представляет собой грибок или оомицет.
5. Химерный промотор по п. 4, отличающийся тем, что он включает по меньшей мере один мультимер цис-регуляторных элементов, причем по меньшей мере один из этих цис-регуляторных элементов является элементом по любому из пп. 1-3.
6. Химерный промотор по п. 5, отличающийся тем, что мультимер представляет собой димер или тетрамер.
7. Рекомбинантный ген для экспрессии в растительной клетке после контакта с патогеном, включающий химерный промотор по любому из пп. 4-6, причем патоген представляет собой грибок или оомицет.
8. Экспрессионный вектор для повышения устойчивости растения к патогенам, включающий химерный промотор по любому из пп. 4-6 или рекомбинантный ген по п. 7, причем патоген представляет собой грибок или оомицет.
9. Прокариотическая или эукариотическая клетка для получения растения с повышенной устойчивостью к патогенам, включающая химерный промотор по любому из пп. 4-6, рекомбинантный ген по п. 7 или вектор по п. 8, причем патоген представляет собой грибок или оомицет.
10. Трансгенное растение для снижения риска заражения патогеном, трансформированное химерным промотором по любому из пп. 4-6, рекомбинантным геном по п. 7 или вектором по п. 8, причем патоген представляет собой грибок или оомицет.
11. Семя растения для культивирования растения со сниженным риском заражения, трансформированного химерным промотором по любому из пп. 4-6, рекомбинантным геном по п. 7 или вектором по п. 8.
12. Ткань растения для in vitro культивирования, трансформированного химерным промотором по любому из пп. 4-6, рекомбинантным геном по п. 7 или вектором по п. 8.
13. Часть растения для снижения риска заражения патогеном, трансформированного химерным промотором по любому из пп. 4-6, рекомбинантным геном по п. 7 или вектором по п. 8, причем патоген представляет собой грибок или оомицет.
14. Способ создания трансгенного растения по п. 10, включающий
a) внесение химерного промотора согласно одному из пп. 4-6, рекомбинантного гена по п. 7 или вектора по п. 8 в по меньшей мере одну клетку растения и
b) регенерацию растения.
15. Способ патоген-индуцируемой экспрессии эндогенного гена в растительной клетке под контролем нативного промотора, модифицированного цис-регуляторным элементом, включающий внесение цис-регуляторного элемента по любому из пп. 1-3 в нативный промотор эндогенного гена, причем патоген представляет собой грибок или оомицет.
16. Растительная клетка для экспрессии эндогенного гена под контролем химерного или нативного промотора, которая содержит указанный эндогенный ген, включающий химерный промотор по любому из пп. 4-6 или нативный промотор эндогенного гена, модифицированный внесением цис-регуляторного элемента по любому из пп. 1-3.
17. Растение для патоген-индуцируемой экспрессии эндогенного гена, которое содержит растительную клетку по п. 16, причем патоген представляет собой грибок или оомицет.
| EP1637607 A2, 22.03.2006 | |||
| WO9503690 A1, 9.02.1995 | |||
| ФРАГМЕНТ ДНК, ПОЛУЧАЕМЫЙ ИЗ Arabidopsis thaliana, ЕГО СУБФРАГМЕНТ ИЛИ КОМБИНАЦИЯ СУБФРАГМЕНТОВ, ПОСЛЕДОВАТЕЛЬНОСТЬ ХИМЕРНОЙ ДНК И ЕЕ ИСПОЛЬЗОВАНИЕ, РЕПЛИКОН (ВАРИАНТЫ) | 1996 |
|
RU2198219C2 |
| Сверлильный станок для пуговиц | 1930 |
|
SU29592A1 |
| ПАТОГЕН-ИНДУЦИРУЕМЫЙ СИНТЕТИЧЕСКИЙ ПРОМОТОР | 2007 |
|
RU2406761C2 |

