Изобретение относится к биотехнологии, в частности к клеточной биоинженерии, а именно к клеточным технологиям скрининга молекулярных мишеней. Может быть использовано при разработке и фармакологическом контроле новых лекарственных средств.
Мембранные белки представляют собой обширный класс клеточных белков; наиболее биологически значимыми мембранными белками являются белки-рецепторы (например, GPCR, тирозин-киназные рецепторы), ионные каналы, белки-транспортеры, белки дыхательной цепи и др. Мембранные белки участвуют в регуляции клеточной активности путем связывания различных лигандов на поверхности клетки и проведения сигнала внутрь клетки. Возникновение множества заболеваний и патологий, таких как рак, диабет, сердечная аритмия, гипертония, иммунодефицитные состояния, связано с нарушением уровня экспрессии и/или нормальной функциональной активности мембранных белков. Различные мембранные белки, в первую очередь рецепторы и ионные каналы, являются молекулярными мишенями многих лекарств.
Поиск и отбор лигандов, способных корректировать аномальную активность мембранных белков, является ключевым этапом при разработке лекарственных препаратов нового поколения. Создание новых экспериментальных методов для доклинического скрининга прототипов лекарственных препаратов по их способности взаимодействовать с белками-мишенями направлено на повышение эффективности выявления наиболее перспективных лигандов с потенциально высоким лекарственным действием. В основе этих методов лежит определение лигандсвязывающей активности мембранных белков, позволяющее оценивать характер взаимодействия мембранного белка с лигандом (специфичность, константа диссоциации комплекса и др.), исследовать влияние различных модуляторов на взаимодействие с лигандом и пр.
Основная трудность тестирования лигандсвязывающей активности мембранных белков в растворах заключается в необходимости применения различных детергентов для выделения белка и связанной с этим высокой вероятности денатурации белка в процессе выделения. Кроме того, проведение биохимических тестов в присутствии детергентов затруднено или вообще невозможно.
Для определения тестирования связывания лигандов с мембранными белками, такими как ионные каналы, белки-рецепторы, в том числе GPCR, получение которых в функционально-активном состоянии представляет определенные трудности, используются клеточные тест-системы на основе эукариотических клеточных линий, экспрессирующих заданный рецептор. Методы тестирования основаны на функциональной экспрессии рекомбинантных белков в составе клеточной мембраны и на возможности проводить связывание мембранного белка с лигандом на поверхности клеток. Клеточные технологии позволяют исключить процедуру выделения мембранного белка, следствием чего является: упрощение, ускорение и удешевление технологии связывания; сохранение мембранным белком конформации, определяемой липидным окружением бактериальной мембраны. Связывание мембранного белка в составе клеточной мембраны дает возможность проводить тестирование в физиологических условиях (солевой состав, рН буфера) в отсутствие детергентов, используемых традиционно при выделении мембранных белков из состава мембран.
Известен способ тестирования связывания с лигандом одного из GPCR-рецепторов - hNMUR (рецептор нейромедина U человека), экспрессированного в эукариотических линиях клеток (СНО, НЕК 293), путем измерения уровня биосинтеза репортерного гена люциферазы. [Li X. et.al. Functional characterization of cell lines for high-throughput screening of human neuromedin U receptor subtype 2 specific agonists using a luciferase reporter gene assay. Eur. J. Pharm. Biopharm., 2007, doi:10.1016/j.ejpb.2007.01.004]. Репортерные клеточные линии получают путем со-трансфекции клеток плазмидами, содержащими а) ген целевого белка hNMUR - рецептора нейромедина U и б) ген репортерного белка люциферазы. Активация рецептора hNMUR пептидом природного происхождения нейромедином U инициирует каскад внутриклеточных реакций, связанных с активностью аденилатциклазы и метаболизмом кальция. Изменения уровня внутриклеточного кальция и/или цАТФ, в свою очередь, влияют на уровень транскрипции репортерного белка. Тестирование проводят по определению уровня экспрессии репортерного белка люциферазы, являющегося конечным звеном в цепи клеточных реакций.
Недостатком этого способа, основанного на непрямом определении связывания целевого белка с лигандом, является необходимость тщательной стандартизации условий с целью исключения влияния множества неспецифических факторов на уровень биосинтеза репортерного белка и связанная с этим сложность оценки полученных данных.
Известен наиболее близкий к заявленному способ тестирования связывания мембранного белка с лигандом, основанный на флуоресцентной детекции образования комплекса лиганда с мембранным белком-рецептором (GPCR), экспрессирующимся в эукариотической линии клеток [Mellentin-Michelotti J. ef al. Determination of ligand-binding affinities for endogenous seven-transmembrane receptors using flourometric microvolume assay technology. Analytical Biochemistry, 1999, 272:182-190]. В анализе используют клеточные линии астроцитомы UC11 и нейробластомы, продуцирующие соответственно человеческие рецепторы NK1 и GalR1, а также рекомбинантную клеточную линию NPY-Y2 с высоким уровнем экспрессии рецептора NPY-Y2. Эту линию получают путем трансфекции клеток СНО-К1 плазмидой sr-alpha-NPY-Y2R, содержащей ген рецептора NPY-Y2.
Клетки для анализа готовят двумя способами: в первом способе клетки культивируют в монослое, затем для отделения клеток от подложки проводят трипсинолиз и готовят клеточную суспензию для анализа связывания с лигандом. Во втором варианте клетки засевают в планшеты за сутки до проведения измерений и анализируют связывание лиганда с прикрепленными клетками в монослое.
Лиганды - нейропептиды (SP, нейрокинин А, галанин и др.) - метят флуоресцентным красителем Су5, при этом способность связываться с рецептором у меченых аналогов сравнима с исходными лигандами.
Процедуру связывания флуоресцентно-меченого лиганда с мембранным белком выполняют двумя способами. В первом способе суспензию, содержащую 10000 клеток, смешивают с меченым лигандом в объеме 100 мкл, выдерживают в течение часа, затем вносят в лунки планшета и через 15 мин после оседания клеток на дно проводят измерение флуоресценции связавшегося на клеточной мембране лиганда с помощью лазерного сканирующего микроскопа.
Во втором варианте проводят анализ клеток, выращенных в монослое в лунках планшета. Для этого из лунок отбирают культуральную жидкость, а к адсорбированным на дне клеткам прибавляют раствор меченого лиганда, инкубируют в течение часа и проводят измерения.
Характеризуя исследуемое соединение как возможный лиганд целевого мембранного белка, тестируют способность этого соединения конкурировать с меченым лигандом за образование комплексов с мембранным белком. Способность конкурировать за связывание характеризуют константой IC50, которая численно равна концентрации исследуемого соединения, вытесняющего 50% флуоресцентно-меченого лиганда из комплексов с мембранным белком.
Для этого составляют смеси, содержащие фиксированную концентрацию меченого лиганда и возрастающие концентрации немеченого исследуемого соединения, тестируемого в качестве возможного лиганда. После инкубации клеток со смесями в каждой лунке планшета измеряют интегральную интенсивность флуоресценции по сканируемой области. Для каждой концентрации исследуемого соединения вычисляют среднее значение интенсивности флуоресценции по 16-ти лункам и данные концентрационной зависимости связывания меченого лиганда в условиях конкуренции с исследуемым соединением анализируют в программе Sigmaplot. Измеряемая интенсивность флуоресценции соответствует флуоресценции лиганда, связавшегося на поверхности клеток (флуоресценция меченого лиганда в растворе не детектируется), а интенсивность флуоресценции прямо пропорциональна количеству связанного лиганда. В результате сканирования получают также флуоресцентные изображения клеток и определяют, таким образом, локализацию лиганда на клеточной поверхности.
С помощью описанного метода тестирования определяют, например, значение 1С50 для нейропептида SP (проводят конкурентное вытеснение меченого пептида Cy5-SP немеченым SP пептидом), связывающегося с рецептором NK1 на клетках UC11, которое составляет 0.7 нМ. Это значение соответствует данным, получаемым с помощью радиоизотопного метода анализа. В случае конкурентного вытеснения меченого пептида Cy5-SP пептидом NKA, имеющим слабое сродство к рецепторам NK1 на UC11 клетках, константа IС50 составляет 75 нМ.
В описанных выше клеточных тест-системах используются для анализа клеточные линии с относительно высоким уровнем экспрессии природного мембранного рецептора или стабильные эукариотические клеточные линии, полученные путем трансфекции клеток плазмидой, содержащей ген целевого рекомбинантного белка. Процесс создания таких клеточных линий является длительным, трудозатратным и дорогим; не всегда уровень экспрессии целевого белка оказывается достаточен для надежной регистрации.
Изобретение решает задачу создания простого и эффективного способа качественного и количественного определения связывания мембранного белка с исследуемым соединением - предполагаемым лигандом.
Поставленная задача решается за счет того, что способ тестирования связывания мембранного белка с лигандом включает экспрессию рекомбинантного мембранного белка в клетках E.coli в составе цитоплазматической мембраны, приготовление препарата сферопластов из клеток E.coli путем пермеабилизации клеточной стенки, приготовление меченого лиганда, внесение сферопластов в лунки планшета, инкубацию сферопластов с меченым лигандом, детекцию на поверхности сферопластов комплекса мембранного белка с лигандом и измерение уровня флуоресценции на поверхности сферопластов методом флуоресцентной оптической микроскопии, определение константы диссоциации Kd для комплекса мембранного белка с лигандом, проведение конкурентного вытеснения флуоресцентно-меченного лиганда немеченым лигандом из комплекса с мембранным белком, определение константы IС50 для немеченого лиганда и расчет константы диссоциации Ki для немеченого лиганда.
Способ осуществляют следующим образом.
В составе цитоплазматической мембраны Е. coli осуществляют функциональную экспрессию мембранного белка KcsA-Kv1.3. Этот белок представляет собой гибрид, в котором участок поровой петли калиевого канала KcsA из бактерии Streptomyces lividans заменен на гомологичный участок эукариотического вольтзависимого калиевого канала Kv1.3. Ген KcsA получают путем амплификации на матрице хромосомной ДНК Streptomyces lividans и клонируют в плазмиду рЕТ28а фирмы Novagen, США. Полученную плазмиду pETKcsA используют в ПЦР для синтеза гибридного гена KcsA-Kv1.3. Клетки E.coli BL21(DE3) трансформируют полученной плазмидой pETKcsA-Kv1.3 и проводят культивирование. Биосинтез целевого мембранного белка KcsA-Kv1.3 в клетках E.coli определяют с помощью электрофоретического анализа суммарного клеточного белка.
Аналогичным образом проводят конструирование гена гибридного белка KcsA-Kν1.1 в составе плазмиды pETKcsA-Kν1.1 и последующее культивирование клеток E.coli BL21(DE3) для наработки белка KcsA-Kv1.1. Этот белок содержит участок поровой петли эукариотического вольтзависимого калиевого канала Kv1.1.
Подготовка клеток для анализа заключается в приготовлении сферопластов из клеток E.coli, экспрессирующих мембранный белок KcsA-Kv1.3 (или KcsA-Kv1.1) в составе цитоплазматической мембраны. Пермеабилизацию и последующее удаление наружной клеточной стенки осуществляют путем обработки клеток E.coli раствором, содержащим лизоцим, Трис-НСl и ЭДТА, и последующей стабилизации цитоплазматической мембраны добавлением MgCl2. В результате такой обработки флуоресцентно-меченый лиганд связывается с мембранным белком на поверхности сферопластов E.coli.
Флуоресцентно-меченый лиганд, обладающий сродством к участку поровой петли эукариотического калиевого канала Kv1.3 (а также канала Kv1.1), получают путем мечения пептидного токсина (агитоксина, AgTx2) флуоресцентным красителем TAMRA.
Процедуру связывания флуоресцентно-меченого лиганда AgTx2-TAMRA с мембранным белком KcsA-Kv1.3 (или KcsA-Kv1.1), локализованным в цитоплазматической мембране клеток E.coli, проводят одним из двух вариантов. В обоих случаях используют лунки планшета, дно которых предварительно обрабатывают раствором полилизина для иммобилизации клеток. В первом варианте суспензию сферопластов инкубируют в лунках планшета в солевом буфере с флуоресцентно-меченым лигандом заданной концентрации, а затем проводят регистрацию флуоресцирующих клеток, осевших на дно лунки, или планшет подвергают центрифугированию для более полного осаждения клеток и проводят регистрацию флуоресцирующих клеток. Во втором варианте суспензию сферопластов вносят в лунки планшета, выдерживают в течение 30 мин для оседания сферопластов на дно лунки или подвергают центрифугированию для более полного осаждения клеток, затем неосевшие сферопласты удаляют, проводят инкубацию иммобилизованных сферопластов в солевом буфере с флуоресцентно-меченым лигандом и регистрируют флуоресцирующие клетки.
Регистрацию флуоресцирующих клеток (то есть комплексов флуоресцентно-меченого лиганда с мембранным белком на поверхности сферопластов) проводят с помощью флуоресцентной оптической микроскопии с применением одного из следующих методов: метода лазерной сканирующей конфокальной микроскопии, эпифлуоресцентной микроскопии или многофотонной сканирующей микроскопии. Результатом применения любого из этих методов являются цифровые микрофотографии, описывающие распределение флуоресцентно-меченого лиганда на мембране сферопластов.
Среднее количество связавшегося флуоресцентно-меченого лиганда в расчете на клетку оценивается по цифровой фотографии путем измерения средней яркости свечения меченого лиганда на поверхности отдельных клеток и усреднения этих данных по выборке клеток.
Для определения константы Kd диссоциации комплекса меченого лиганда и мембранного белка проводят: инкубацию клеток в лунках планшета с возрастающей концентрацией меченого лиганда; регистрацию флуоресцирующих клеток для каждой концентрации меченого лиганда; оценку среднего количества связавшегося флуоресцентно-меченого лиганда в расчете на клетку; построение зависимости среднего количества связавшегося лиганда от концентрации внесенного в среду лиганда; аппроксимацию построенной зависимости функцией
I=IмC/(Kd+C),
где Iм - максимальное количество связавшегося лиганда, соответствующее выходу функции на плато, С - концентрация внесенного в среду лиганда, Кd - искомая константа диссоциации комплекса меченого лиганда и мембранного белка.
Проведение конкурентного вытеснения меченого лиганда исследуемым соединением (немеченым потенциальным лигандом) из комплексов с целевым мембранным белком проводят путем инкубации клеток со смесью, содержащей фиксированную концентрацию меченого лиганда и возрастающую концентрацию исследуемого соединения. Если соединение вытесняет меченый лиганд, то оно само является лигандом. В этом случае для исследуемого соединения определяют константу IС50. Проводят: регистрацию флуоресцирующих клеток для каждой концентрации исследуемого соединения; оценку среднего количества связавшегося флуоресцентно-меченого лиганда в расчете на клетку; построение зависимости среднего количества связавшегося меченого лиганда от концентрации внесенного в среду исследуемого соединения; определение по построенной зависимости константы IC50, которая численно равна концентрации исследуемого соединения, вытесняющего 50% флуоресцентно-меченого лиганда из комплексов с мембранным белком.
Расчет константы диссоциации Кi для исследуемого соединения, являющегося лигандом, проводят в соответствии с уравнением Ченга-Прусоффа [Cheng Y., Prusoff W.H. Relationship between the inhibition constant (K.1) and the concentration of inhibitor which causes 50 per cent inhibition (150) of an enzymatic reaction Biochem. Pharmacol. 1973, 22:3099-3108] по формуле:
IC50=Ki(1+[L*]/Kd),
где [L*] - концентрация свободного меченого лиганда AgTx2-TAMRA (принимается равной общему количеству меченого лиганда).
Заявляемый способ позволяет осуществлять эффективный и простой способ качественного и количественного определения связывания мембранного белка с лигандом, расширяет возможности тестирования связывания мембранного белка с лигандом, повышает чувствительность и точность измерений, облегчает детекцию и обсчет данных, полученных с помощью флуоресцентных методов анализа, позволяет получать точные количественные оценки всех параметров связывания.
Эффективность и простота способа обеспечивается тем, что для анализа связывания мембранного белка с лигандом используют клетки E.coli, экспрессирующие целевой мембранный белок в составе цитоплазматической мембраны. Все стадии получения этих клеток (клонирование генов целевых белков в прокариотические плазмидные векторы, трансформация этими векторами клеток E.coli, поддержание и культивирование штаммов, содержащих плазмидные векторы, проведение индукции целевого мембранного белка) отличаются простотой и эффективностью технологических операций, дешевизной питательных сред и реактивов, не требуют дорогостоящего оборудования для культивирования.
Эффективность метода определяется также тем, что его можно отнести к технологии флуориметрического тестирования в микрообъеме (FMAT): анализ проводят в микрообъемах (50 мкл) с использованием планшетов; процедура анализа представляет собой «гомогенный» способ тестирования, т.е. не требует удаления несвязавшегося меченого лиганда и исследуемого соединения из анализируемого образца; регистрацию лиганда, связавшегося с клетками, осуществляют одним из методов флуоресцентной микроскопии.
Эффективность и простота этого способа как клеточной технологии тестирования обеспечивается тем, что исключается процедура выделения мембранного белка из состава бактериальной мембраны.
Детекция связывания флуоресцентно-меченного лиганда с мембранным белком на поверхности бактериальной цитоплазматической мембраны представляет собой новую систему тестирования связывания мембранного белка с лигандом и, таким образом, расширяет возможности тестирования лигандсвязывающей активности мембранных белков.
Получение качественных и количественных параметров связывания обеспечивается тем, что в заявленном способе для детекции комплекса мембранного белка с лигандом используют лазерный сканирующий микроскоп, в результате чего исследователь получает флуоресцентные изображения клеток. Эти изображения позволяют, во-первых, локализовать комплекс мембранного белка с лигандом в составе клеточной мембраны и, во-вторых, произвести измерения средней яркости свечения флуоресцентного лиганда на поверхности отдельных клеток (для последующих расчетов констант связывания).
Повышение чувствительности и точности измерений в значительной мере определяется высоким уровнем флуоресцентного сигнала, детектируемого на поверхности единичной клетки. Высокий уровень флуоресцентного сигнала, в свою очередь, является следствием, во-первых, высокого уровня экспрессии ряда мембранных белков в составе бактериальной мембраны (до 1000000 молекул рецептора на клетку и выше), что значительно превосходит уровень экспрессии мембранных белков в эукариотических клетках (10-100000 молекул на клетку), и, во-вторых, высокой плотности молекул мембранного белка в составе бактериальной мембраны (площадь поверхности бактериальной мембраны в 2000 раз меньше поверхности эукариотической клетки). Кроме того, неспецифическое (фоновое) связывание флуоресценто-меченного лиганда на поверхности бактериальной цитоплазматической мембраны практически отсутствует. Таким образом, высокий уровень флуоресцентного сигнала и значительное превышение уровня этого сигнала по отношению к фоновой флуоресценции позволяет повысить чувствительность метода, способствует упрощению обработки данных и получению более достоверных количественных характеристик.
Облегчение детекции и обсчета данных обеспечивается, во-первых, применением нерадиоактивного, а именно флуоресцентного метода анализа; во-вторых, однородностью популяции клеток бактерий, несущих мембранный белок (в отличие от эукариотеских клеток); в-третьих, специальными процедурами приготовления сферопластов и тестирования. Процедура приготовления сферопластов, разработанная специально для визуализации флуоресцентно-окрашенных клеток в лазерном сканирующем микроскопе, основана на простых приемах (измерении оптической плотности клеточной суспензии, низкоскоростном центрифугировании, смешивании реагентов) и не требует для выполнения больших временных затрат (не более одного часа). Процедура приводит к получению четких изображений отдельных сферопластов, равномерному распределению флуоресцентного окрашивания на мембране отдельной клетки, а также соседних клеток, что является необходимым условием анализа связывания и проведения точных количественных расчетов. В результате этой процедуры происходит полное удаление внешней клеточной стенки, отсутствует слипание сферопластов, а также происходит количественное образование сферопластов из исходных клеток. Процедура позволяет хранить полученные сферопласты в течение, по крайней мере, 10 дней с сохранением их способности к связыванию с флуоресцентным токсином. Указанные эффекты достигаются соблюдением соотношений концентрации исходных клеток в суспензии и концентраций реагентов, таких как лизоцим, ЭДТА, MgCl2, а также последовательности смешивания реагентов и соблюдения температурного режима. Получению четких изображений отдельных сферопластов способствует и разработанная процедура тестирования, в которой для иммобилизации сферопластов на дне лунки планшета используют обработку лунок раствором полилизина.
Получение точных количественных оценок всех параметров связывания обеспечивается возможностью определения константы диссоциации комплекса мембранного белка с меченым лигандом (Kd), проведения теста по конкурентному ингибированию и определения величины IС50, а также расчета константы диссоциации комплекса мембранного белка с немеченым лигандом (Ki). Расчет констант диссоциации Kd и Ki позволяет количественно сопоставить аффинность используемых лигандов к мембранному белку.
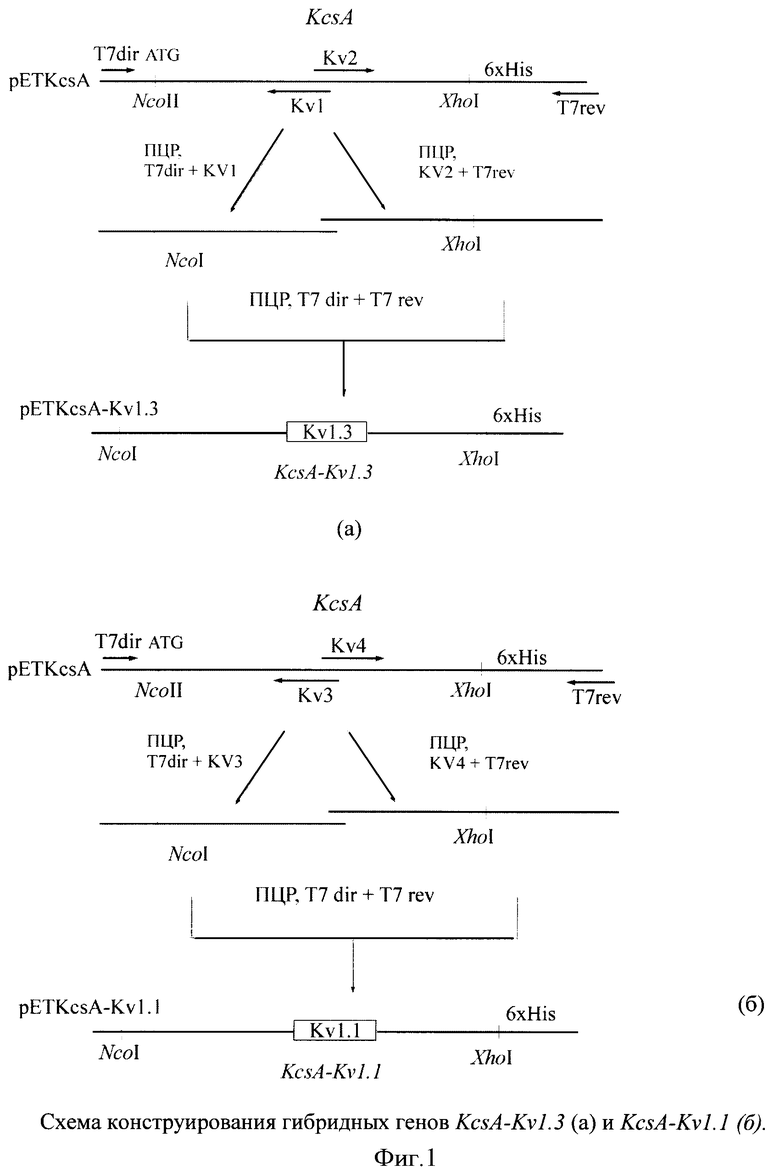
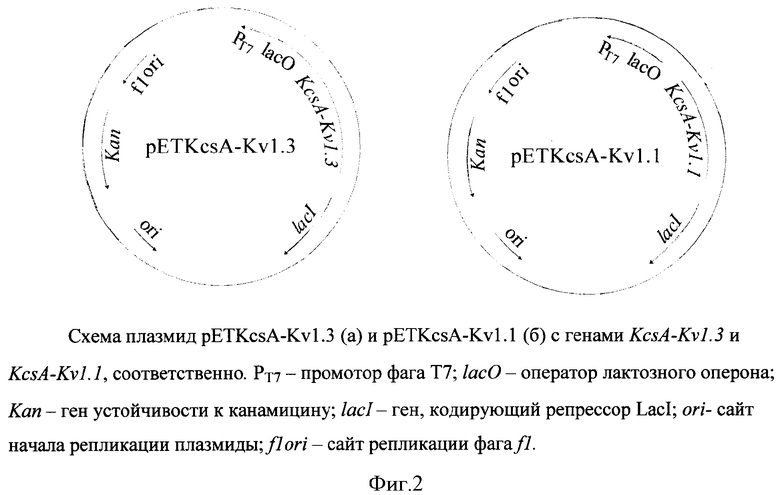
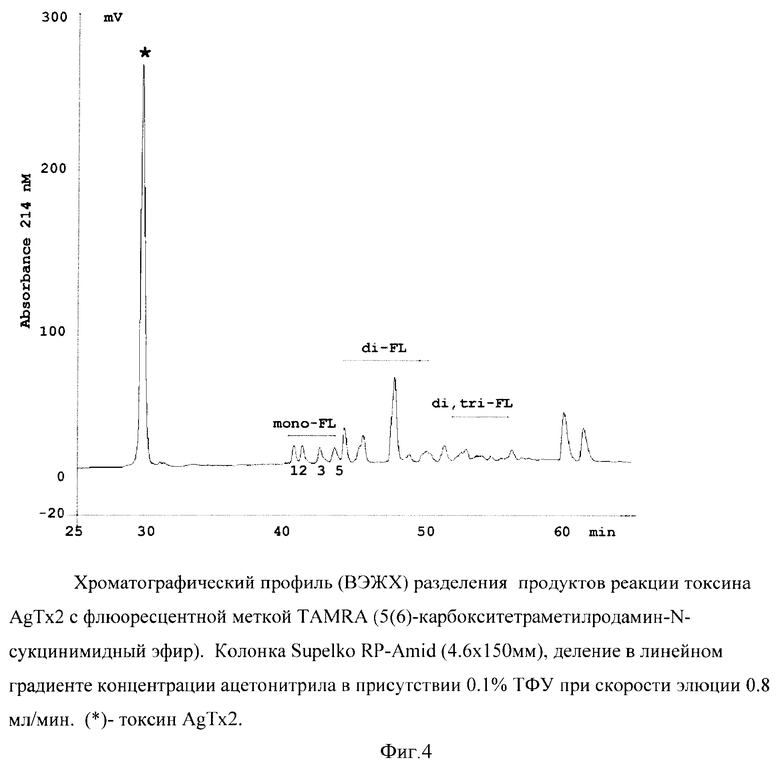
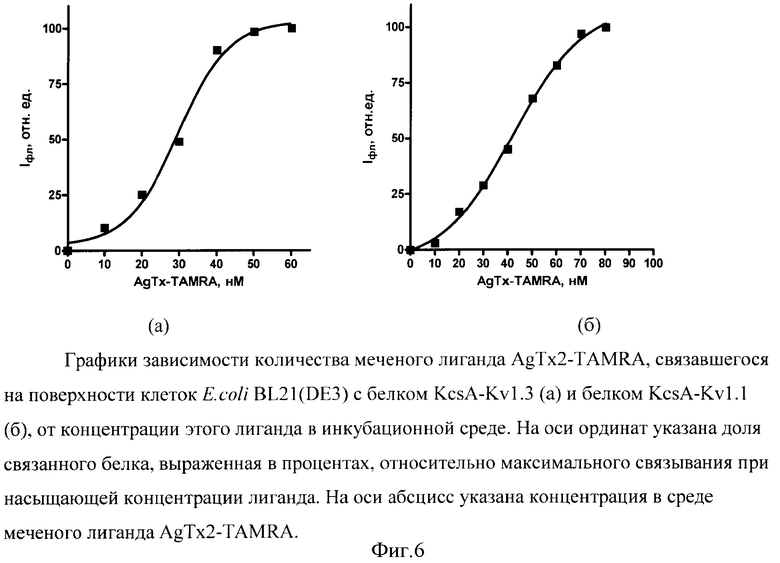
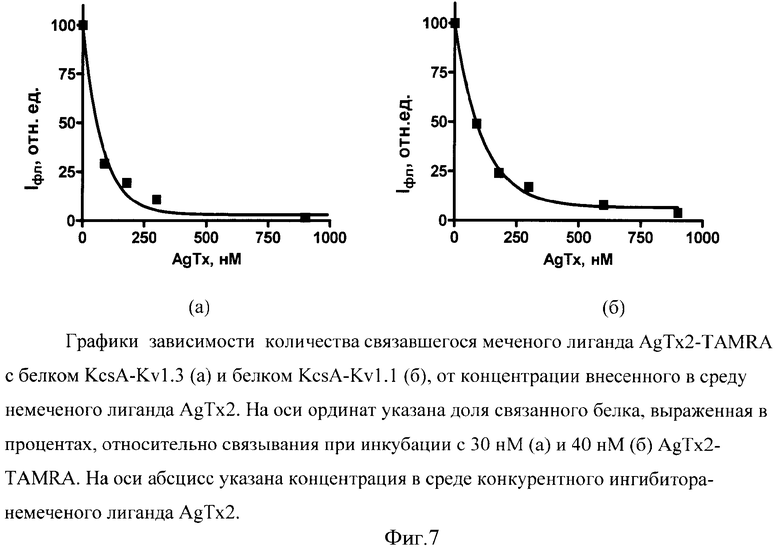
Для иллюстрации изобретения представлены следующие графические материалы: схема конструирования гибридых генов KcsA-Kν1.3 и KcsA-Kν1.1 (фиг.1, а,б); схемы плазмид pETKcsA-Kv1.3 и pETKcsA-Kv1.1 с генами KcsA-Kv1.3 и KcsA-Kv1.1, соответственно (фиг.2, а,б); фотография электрофоретического разделения суммарного белка клеток BL21(DE3) с плазмидами pETKcsA-Kv1.3 и pETK.csA-Kv1.1 в 13.5%-ном SDS-ПААГ (фиг.3); хроматографический профиль (ВЭЖХ) разделения продуктов реакции токсина AgTx2 с флюоресцентной меткой TAMRA (5(6)-карбокситетраметилродамин-N-сукцинимидный эфир) (фиг.4); цифровые микрофотографии клеток E.coli BL21(DE3) с белками KcsA-Kv1.3 и KcsA-Kv1.1, связывающих на своей поверхности флуоресцентно-меченый лиганд AgTx2-TAMRA (фиг.5); графики зависимости количества меченого лиганда AgTx2-TAMRA, связавшегося на поверхности клеток E.coli BL21(DE3), от концентрации этого лиганда в инкубационной среде для белков KcsA-Kv1.3 и KcsA-Kv1.1 (фиг.6); графики зависимости количества связавшегося меченого лиганда AgTx2-TAMRA от концентрации внесенного в среду немеченого лиганда AgTx2 для белков KcsA-Kv1.3 и KcsA-Kv1.1 (фиг.7).
Изобретение иллюстрируют примеры.
Пример 1.
Получение клеток E.coli, экспрессирующих мембранный белок KcsA-Kv1.3: Хромосомную ДНК Streptomyces lividans выделяют из мицеллия Streptomyces lividans 66 [Rascher, A. et al. 2003. Cloning and characterization of a gene cluster for geldanamycin production in Streptomyces hygroscopicus. FEMS Microbiology Lett.218:223-230]. Ген KcsA (EMBL GenBank, accession number Z37969) амплифицируют методом полимеразной цепной реакции (ПЦР) на матрице хромосомной ДНК Streptomyces lividans [Legros, С.et al. 2000. Generating a High Affinity Scorpion Toxin Receptor in KcsA-Kv1.3 Chimeric Potassium Channels. J. Biol. Chem.275: 16918-16924], а затем клонируют в плазмиду рЕТ28а (фирмы Novagen, США) по сайтам рестриктаз Ncol и Xhol. Полученная плазмида pETKcsA кодирует рекомбинантный белок KcsA-6xHis, содержащий гексагистидиновый таг на С-конце.
Для конструирования гена KcsA-Kvl.3, кодирующего гибридный белок, в котором последовательность аминокислотных остатков (а.о.) 52-64 белка KcsA (EMBL accession Р0А334) заменена на последовательность а.о. 371-383 белка Kv1.3 (EMBL accession P2201), проводят ПЦР, в которой в качестве матрицы используют плазмиду pETKcsA. Схема конструирования гибридного гена представлена на фиг.1а. Для получения фрагментов ДНК, содержащих нуклеотидную последовательность соответствующих участков гена Kv1.3, используют синтетические олигонуклеотидные праймеры KV1 и KV2. Амплификацию 5'-концевой части гена KcsA проводят в присутствии праймеров Т7 direct (прямого) и KV1; 3'-концевую часть - KV2 и Т7 reverse (обратного):
Т7 direct: 5' - TAATACGACTCACTATAGGG
Т7 reverse: 5'-GCTAGTTATTGCTCAGCGG
KV1: 5'-GCTGAAACCGCTGGTCGGATCATCTGCCTCAGCCAGGACGGC
KV2: 5'- ACCAGCGGTTTCAGCAGCATCCCGGATGCGCTGTGGTGGTCC
После выделения из реакционной смеси полученные фрагменты ДНК, содержащие перекрывающиеся последовательности в области праймеров KV1 и KV2, используют для конструирования полноразмерного гена KcsA-Kv1.3 с помощью пары праймеров Т7 direct и Т7 reverse. Полученный фрагмент ДНК гидролизуют рестриктазами Ncol и Xhol и клонируют по соответсвующим сайтам в вектор рЕТ28а. Структуру полученной плазмиды pETKcsA-Kv1.3 подтверждают секвенированием. Схема плазмиды pETKcsA-Kv 1.3 с геном KcsA-Kv1.3 представлена на фиг.2а. Гибридный ген KcsA-Kv1.3 имеет следующую нуклеотидную последовательность и кодирует соответствующую аминокислотную последовательность (последовательность гена Ку1.3 подчеркнута):

Клетки E.coli BL21(DE3) трансформируют плазмидой pETKcsA-Kv1.3; ночную культуру засевают в жидкую питательную среду LB (среда Luria Bertani), содержащую 40 мкг/мл канамицина, до начальной мутности OD560=0.2 и культивируют при 37°С и качании 200 об/мин. При OD560=0.8 проводят индукцию культуры добавлением ИПТГ до концентрации 50 мкМ и продолжают выращивание в течение 18 час.
Экспрессию белка KcsA-Kv1.3 оценивают с помощью разделения суммарного клетотчного лизата электрофорезом в SDS-ПААГ [Laemmli, U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685]. Биосинтез белка KcsA-Kv1.3 в клетках E.coli показан на фиг.3а.
Приготовление сферопластов:
Суспензию клеток E.coli BL21(DE3) объемом 0.1 мл с оптической плотностью OD560=3.0 о.е., содержащих мембранный белок KcsA-Kv1.3, охлаждают до 4°С, и клетки осаждают центрифугированием при 4000 g в течение 3 мин. Клетки промывают охлажденным буфером, содержащим 20 мМ Трис-HCl, рН 7.5, 50 мМ NaCl, центрифугируют и суспендируют в 1 мл буфера А (50 мМ Трис-HCl, рН 7.5, 0.3 М сахароза, 0.3 мМ ЭДТА). К клеточной суспензии добавляют раствор лизоцима до конечной концентрации 20 мкг/мл, смесь инкубируют при слабом качании в течение 30 мин при 4°С, а затем добавляют раствор MgCl2 до конечной концентрации 10 мМ и через 10 мин растворы NaCl и КСl до конечной концентрации соответственно 100 и 20 мМ. Полученную суспензию сферопластов (около 10(6) клеток/мл) хранят при 4°С в течение 10 дней.
Получение флуоресцентно-меченого лиганда:
AgTx2 из яда скорпиона Leirus quinquestriatus - один из наиболее изученных классических ингибиторов калиевых каналов. AgTx2 широко применяется в изучении бактериального канала KcsA и его гибридов с потенциал-зависимыми каналами млекопитающих. Токсин AgTx2 представляет собой полипептид длиной 38 а.о. со следующей первичной структурой:
GVPINVSCTGSPQCIKPCKDAGMRFGKCMNRKCHCTPK
Кодирующую последовательность гена AgTx2 клонируют в вектор рЕТ32b по сайтам Kpn1/BamH1. При индукции 0.1-0.4 мМ IPTG и инкубации при 25°С в линиях клеток BL21(DE3) и Origami В, содержащих плазмиду pETAgTx2, образуются растворимые продукты экспрессии. Эффективную очистку гибридных белков проводят из суммарного растворимого клеточного лизата в одну стадию с использованием металлоаффинной хроматографии на сорбенте Talon (Clontech). Выход гибридного белка после металлоаффинной хроматографии составляет 80 мг/л для Trx-AgTx2 на богатой среде.
Для выделения целевых пептидов из состава гибридных белков используют гидролиз энтерокиназой по сайту DDDDK, предварительно введенному при создании экспрессирующей конструкции, с последующей очисткой пептидов методом ВЭЖХ на обращенной фазе. Состав рекомбинантных пептидов подтверждают MALDI- масс-спектрометрией. Общий выход продуктов после расщепления энтерокиназой составляет 50% для AgTx2.
Для получения необходимого количества флуоресцентно-меченого токсина AgTx2 к 10 наномолям токсина AgTx2, растворенного в 100 мкл 50 мМ фосфата натрия рН 8.5, добавляют 30 наномолей метки TAMRA (5(6)-карбокситетраметилродамин-N-сукцинимидный эфир) в ДМСО (3 мкл), реакционную смесь инкубируют при 25°С 4 часа, продукты реакции разделяют ВЭЖХ в следующих условиях: колонка Supelko RP-Amid (4.6×150 мм), элюция в линейном градиенте концентрации ацетонитрила в присутствии 0.1% ТФУ при скорости 0.8 мл/мин. Моно-меченые производные определяют масс-спектрометрически.
Наибольшей активностью обладает моно-производное 3 (фиг.4), которое содержит флуоресцентную метку, связанную с С-концевым остатком Lys38.
Проведение процедуры связывания мембранного белка с лигандом и детекции связавшегося лиганда:
В лунки планшета вносят по 50 мкл раствора 0,1%-ного полилизина, выдерживают около часа при 37°С для адсорбции полимера на дно планшета, затем остатки раствора удаляют, планшет высушивают и используют для проведения анализа.
В лунки планшета вносят по 50 мкл суспензии сферопластов, затем добавляют 2 мкл флуоресцентно-меченого AgTx2 до конечной концентрации 30 нМ и выдерживают в течение 40 мин при 20°С.
Детекцию образования комплекса мембранного белка KcsA-Kv1.3 с флуоресцентно-меченым лигандом AgTx2 осуществляют методом лазерной сканирующей конфокальной микроскопии с помощью установки LSM510 МЕТА. Цифровая микрофотография, описывающая распределение флуоресцентно-меченого лиганда на мембране сферопластов, представлена на фиг.5а и 5б.
Параллельно проводят процедуру связывания мембранного белка KcsA-Kv1.3 с флуоресцентно-меченым лигандом следующим образом: в лунки планшета вносят по 50 мкл суспензии сферопластов, выдерживают 30 мин при 4°С, затем суспензию неосевших клеток отбирают, а в лунки, содержащие адсорбированные на дне сферопласты, вносят по 50 мкл буфера В (50 мМ Трис-HCl, 0.3 М сахароза, 0.3 мМ ЭДТА, 10 мМ MgCl2, 20 мМ КСl, 100 мМ NaCl, 0.1 % BSA), содержащего 30 нМ лиганда. Смесь выдерживают в течение 1 час при 20°С. Детекцию связавшегося лиганда проводят, как описано выше.
Определение константы Kd диссоциации комплекса флуоресцентно-меченого лиганда и мембранного белка KcsA-Kv1.3:
Константа Kd характеризует аффинность меченого токсина к мембранному белку KcsA-Kv1.3, экспонированному на поверхности сферопластов. Для определения Kd сферопласты инкубируют с увеличивающимися концентрациями меченого токсина AgTx2-TAMRA. Для этого в лунки планшета вносят по 50 мкл суспензии сферопластов, выдерживают 30 мин при 4°С, затем суспензию неосевших клеток отбирают, а в лунки вносят по 50 мкл буфера В, содержащего последовательно увеличивающиеся концентрации меченого токсина AgTx2-TAMRA. Смесь выдерживают в течение 1 ч при 20°С, затем проводят детекцию комплексов, как описано выше.
Для каждой концентрации AgTx2-TAMRA получают по 5 цифровых микрофотографий размером 120×120 мкм, на каждой из которых присутствует 100-150 клеток. Все изображения получают в идентичных условиях при одинаковом напряжении на детекторе и размере зоны сканирования.
Полученные цифровые микрофотографии обрабатывают при помощи программы Image J: убирают шумы, используя функцию Порог (Threshold), применяют функцию Анализ частиц (Analyze Particles) для определения измерения средней яркости свечения AgTx2-TAMRA на поверхности отдельных клеток и общего количества клеток. Средняя яркость свечения AgTx2-TAMRA на клетке пропорциональна количеству токсина, связавшегося на поверхности сферопласта. С использованием полученного массива цифровых данных для каждой концентрации AgTx2-TAMRA рассчитывают усредненное по выборке клеток количество связавшегося AgTx2-TAMRA в расчете на клетку (в единицах средней яркости свечения) и стандартное отклонение этого значения. Строят график зависимости среднего количества связавшегося лиганда от концентрации внесенного в среду лиганда (фиг.6а) и аппроксимируют эту зависимость функцией
I=IмC/(Kd+C),
определяя методом наименьших квадратов величину Кd. Для приведенной на фиг.6а зависимости значение Kd составляет Kd=29,6±1,6 нМ.
Проведение конкурентного вытеснения меченого лиганда исследуемым соединением из комплексов с целевым мембранным белком; определение константы IC50 и расчет константы диссоциации Кi для немеченого токсина AgTx2.
Для определения константы IC50 для немеченого AgTx2 сферопласты инкубируют в присутствии фиксированной концентрации меченого токсина AgTx2-TAMRA и увеличивающихся концентраций немеченого токсина AgTx2. Для этого в лунки планшета вносят по 50 мкл суспензии сферопластов, выдерживают 30 мин при 4°С, затем суспензию неосевших клеток отбирают, а в лунки вносят по 50 мкл буфера В, содержащего AgTx2-TAMRA в концентрации 30 нМ, и последовательно увеличивающиеся концентрации немеченого токсина AgTx2 (60, 90, 180, 300 и 900 нМ). Смесь выдерживают в течение 1 ч при 20°С, затем проводят регистрацию флуоресцирующих клеток для каждой концентрации AgTx2, как описано выше. Оценку среднего количества связавшегося флуоресцентно-меченого лиганда в расчете на клетку проводят, как описано выше. На фиг.7а представлена зависимость среднего количества связавшегося AgTx2-TAMRA от концентрации внесенного в среду немеченого лиганда AgTx2. Значение константы IC50, соответствующей концентрации AgTx2, при которой происходит вытеснение 50% AgTx2-TAMRA из комплексов с KcsA-Kv1.3, равно IC50=55,8±1,0 нМ, а расчетная константа диссоциации для немеченого токсина AgTx2 равна Ki=55,8/(1+29,6/30)=28,1±1,0 нМ.
Пример 2.
Пример 2 проводят, как пример 1, но конструирование гена KcsA-Kv1.1, кодирующего гибридный белок, в котором последовательность аминокислотных остатков (а.о.) 52-64 белка KcsA (EMBL accession POA334) заменена на последовательность а.о. 347-363 белка Kv1.1 (EMBL accession NP000208), проводят следующим образом: для получения в реакции ПЦР фрагментов ДНК, содержащих нуклеотидную последовательность соответствующих участков гена Kv1.1, используют синтетические олигонуклеотидные праймеры Т7 direct и Т7 reverse, а также KV3 и KV4:
KV3: 5'- GCTGAAACCGCTGGTCGGATCATCTGCCTCAGCCAGGACGGC
KV4: 5'- ACCAGCGGTTTCAGCAGCATCCCGGATGCGCTGTGGTGGTCC
Схема конструирования гибридного гена KcsA-Kv1.1 представлена на фиг.1б, а схема плазмиды pETKcsA-Kv1.1 с геном KcsA-Kv1.1 представлена на фиг.2б. Гибридный ген KcsA-Kv1.1 имеет следующую нуклеотидную последовательность и кодирует соответствующую аминокислотную последовательность (последовательность гена Kv1.1 подчеркнута):

Трансформацию клеток BL21(DE3) плазмидой pETKcsA-Kv1.1, индукцию культуры и разделение суммарного клетотчного лизата электрофорезом в SDS-ПААГ проводят, как в примере 1; биосинтез белка KcsA-Kv1.1 в клетках E.coli показан на фиг.3б.
Проведение процедуры связывания мембранного белка KcsA-Kv1.1 с лигандом и детекции связавшегося лиганда проводят, как в примере 1; цифровая микрофотография, описывающая распределение флуоресцентно-меченого лиганда на мембране сферопластов, содержащих KcsA-Kv1.1, представлена на фиг.5в и 5г.
Определение константы Kd мембранного белка KcsA-Kv1.1, экспонированному на поверхности сферопластов, в отношении меченого токсина проводили, как в примере 1. Зависимость количества меченого лиганда AgTx2-TAMRA, связавшегося на поверхности клеток E.coli BL21(DE3) с белком KcsA-Kv1.1, от концентрации лиганда в инкубационной среде представлена на фиг.6б. Для приведенной на фиг.6б зависимости значение Kd составляет Kd=42±2 нМ.
Проведение конкурентного вытеснения меченого лиганда исследуемым соединением из комплексов с целевым мембранным белком KcsA-Kv1.1; определение константы IС50 и расчет константы диссоциации Кi для немеченого токсина AgTx2 проводили, как в примере 1. На фиг.7б представлена зависимость среднего количества связавшегося AgTx2-TAMRA от концентрации внесенного в среду немеченого лиганда AgTx2. Значение константы IC50, соответствующей концентрации AgTx2, при которой происходит вытеснение 50% AgTx2-TAMRA из комплексов с KcsA-Kv1.1, равно IC50=80±2 нМ нМ, а расчетная константа диссоциации для немеченого токсина AgTx2 равна Ki=80/(1+40/42)=41±2 нМ.


Способ относится к области биотехнологии, а именно к способу качественной и количественной оценки связывания мембранного белка с лигандом. Способ включает экспрессию рекомбинантного мембранного белка в клетках E.coli в составе цитоплазматической мембраны. Готовят препарат сферопластов из клеток E.coli путем пермеабилизации клеточной стенки с использованием лизоцима, ЭДТА, MgCl2. Готовят меченый лиганд, вносят сферопласты в лунки планшета, инкубируют сферопласты с меченым лигандом. Детектируют комплекс мембранного белка с лигандом на поверхности сферопластов. Измеряют уровень флуоресценции на поверхности сферопластов методом флуоресцентной микроскопии. Определяют константы диссоциации Kd для комплекса мембранного белка с лигандом. Проводят конкурентное вытеснение флуоресцентно-меченого лиганда немеченым лигандом из комплекса с мембранным белком. Определяют константы IC50 для немеченого лиганда и рассчитывают константы диссоциации Кi для немеченого лиганда. Изобретение позволяет упростить способ и повысить эффективность качественной и количественной оценки связывания мембранного белка с исследуемыми соединениями. 7 ил.
Способ качественной и количественной оценки связывания мембранного белка с лигандом, включающий экспрессию рекомбинантного мембранного белка KcsA-Kvl.3 в клетках E.coli в составе цитоплазматической мембраны, приготовление препарата сферопластов из клеток E.coli путем пермеабилизации клеточной стенки с использованием лизоцима, ЭДТА, MgCl2, приготовление меченого лиганда, внесение сферопластов в лунки планшета, обработанного раствором полилизина, инкубацию сферопластов с меченым лигандом, детекцию на поверхности сферопластов комплекса мембранного белка с лигандом и измерение уровня флуоресценции на поверхности сферопластов методом флуоресцентной микроскопии, определение константы диссоциации Kd для комплекса мембранного белка с лигандом, проведение конкурентного вытеснения флуоресцентно-меченого лиганда с мембранным белком немеченым лигандом из комплекса с мембранным белком, определение константы IС50 для немеченого лиганда и расчет константы диссоциации Ki для немеченого лиганда, на основе полученных данных делают вывод об аффинности связывания мембранного белка с лигандом.
| MELLENTIN-MICHELOTTI J et al., Determination of ligand binding affinities for endogenous seven-transmembrane receptors using fluorometric microvolume assay technology, Anal Biochem | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| US 2004058403, 25.03.2004 | |||
| MAGGIONI A et al., Targeting the expression of functional murine CMP-sialic acid transporter to the E.coli inner | |||

