ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Аспекты по изобретению относятся к способам получения дифункциональных алканов в клетках-хозяевах. В частности, аспекты по изобретению описывают компоненты генов, ассоциированных с продукцией дифункционального алкана из исходного материала углеводов в клетках-хозяевах. Более конкретно, аспекты по изобретению описывают метаболические пути для продукции адипиновой кислоты, аминокапроновой кислоты, капролактама, гексаметилендиамина через 2-кетопимелиновую кислоту.
УРОВЕНЬ ТЕХНИКИ
Сырая нефть является основным исходным материалом для синтеза ключевых химических веществ и полимеров. Поскольку нефть становится все более дефицитной и дорогой, биологическая обработка возобновляемого сырья в производстве химических веществ с использованием живых микроорганизмов или их очищенных ферментов вызывает возрастающий интерес. Биологическую обработку, в частности брожение, использовали в течение многих столетий для приготовления напитков. За последние 50 лет микроорганизмы коммерчески использовали для получения соединений, таких как антибиотики, витамины и аминокислоты. Тем не менее использование микроорганизмов для производства промышленных химических веществ было гораздо менее распространено. Только недавно стало понятно, что микроорганизмы могут обладать способностью обеспечивать экономически выгодное получение определенных соединений, которые сложны или дорогостоящи при получении обычными химическими способами.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Аспекты по изобретению относятся к метаболически сконструированной клетке-хозяину для продукции α,ω-дифункциональных Cn-алканов из α-кетокислоты, где концевые функциональные группы α и ω выбраны из группы -OH, -COOH и -NH3 и где n представляет собой целое число в диапазоне от 4 до 8, причем метаболически сконструированная клетка-хозяин генетически модифицирована при помощи нуклеиновой кислоты, содержащей, по меньшей мере, одну нуклеотидную последовательность, кодирующую, по меньшей мере, один фермент биосинтетического пути. В некоторых вариантах осуществления нуклеиновая кислота содержит нуклеотидные последовательности, кодирующие два или более продуктов генов. Метаболически сконструированная клетка-хозяин может представлять собой прокариотическую клетку. Например, в некоторых вариантах осуществления метаболически сконструированная клетка-хозяин является анаэробной прокариотической клеткой. Метаболически сконструированную клетку-хозяина можно выбрать из группы, состоящей из E. Coli, C. glutanicum, B. flavum и B. lactofermentum.
В предпочтительных вариантах осуществления α-кетокислота представляет собой α-кетоглутарат и превращается в α-кетоадипат, α-кетопимелат или α-кетосуберат. В предпочтительных вариантах осуществления клетка-хозяин содержит последовательности нуклеиновой кислоты, кодирующие гомоцитратсинтазу, гомоаконитазу и гомоизоцитратдегидрогеназу. Гомоцитратсинтазу можно выбрать из группы, состоящей из AksA, NifV, hcs и Lys20/21. Гомоаконитазу можно выбрать из группы, состоящей из AksD/E, LysT/U, Lys4, большой/малой субъединиц 3-изопропилмалатдегидратазы и их гомологов. Гомоизоцитратдегидрогеназу можно выбрать из группы, состоящей из AksF, Hicdh, Lysl2, 2-оксосубератсинтазы, 3-изопропилмалатдегидрогеназы и их гомологов.
В предпочтительных вариантах осуществления α,ω-дифункциональный алкан содержит шесть атомов углерода и выбран из группы, состоящей из аминокапроновой кислоты, адипата, гексаметилендиамина, 6-гидроксигексамина, 1,6-гександиола, 6-аминогексанала, 6-аминогексанола и 6-гидроксигексаноата.
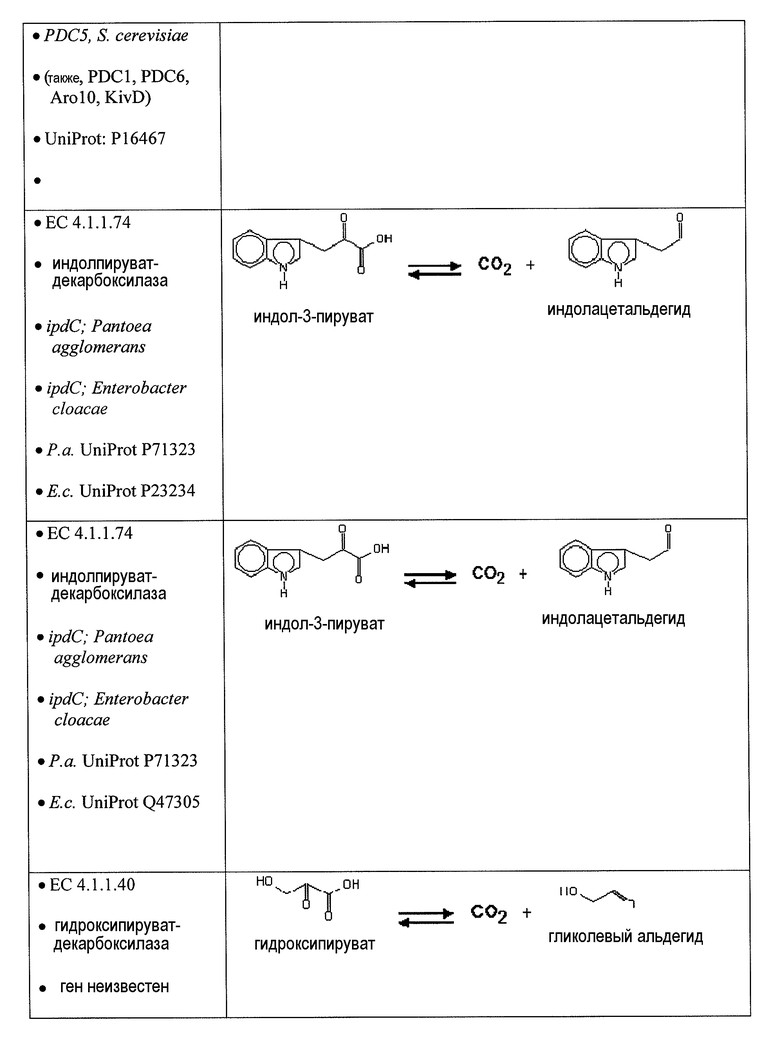
Аспекты по изобретению относятся к метаболически сконструированным клеткам-хозяевам для продукции адипиновой кислоты из α-кетопимелата, причем клетка-хозяин дополнительно содержит нуклеиновую кислоту, кодирующую фермент декарбоксилазу и фермент альдегиддегидрогеназу. В некоторых вариантах осуществления фермент декарбоксилаза представляет собой 2-кетодекарбоксилазу и катализирует превращение α-кетопимелата в полуальдегид адипата, и альдегиддегидрогеназа катализирует превращение полуальдегида адипата в адипиновую кислоту. 2-Кетодекарбоксилазу можно выбрать из группы 2-кетоглутаратдекарбоксилазы (kgd), 2-кетоизовалератдекарбоксилазы (kivD), декарбоксилазы трансаминированной аминокислоты (ARO10), бензоилформиатдекарбоксилазы (mdlC), 2-кетоаргининдекарбоксилазы (aruI), фосфонопируватдекарбоксилазы (fom2), изофермента пируватдекарбоксилазы (PDC6, PDC1), изофермента 2 пируватдекарбоксилазы (PDC5, PDC1, PDC6, Aro10, KivD), индолпируватдекарбоксилазы (ipdC) и их гомологов. В некоторых вариантах осуществления декарбоксилаза обладает, по меньшей мере, 30% идентичности при сравнении с 2-кетоглутаратдекарбоксилазой (kgd), 2-кетоизовалератдекарбоксилазой (kivD), декарбоксилазой трансаминированной аминокислоты (ARO10), бензоилформиатдекарбоксилазой (mdlC), 2-кетоаргининдекарбоксилазой (aruI), фосфонопируватдекарбоксилазой (fom2), изоферментом пируватдекарбоксилазы (PDC6, PDC1), изоферментом 2 пируватдекарбоксилазы (PDC5, PDC1, PDC6, Aro10, KivD), индолпируватдекарбоксилазой (ipdC) и их гомологами. В предпочтительных вариантах осуществления альдегиддегидрогеназа представляет собой 6-оксогексаноатдегидрогеназу (ChnE) и ее гомологи. В других вариантах осуществления альдегиддегидрогеназа обладает, по меньшей мере, 30% идентичностью с 6-оксогексаноатдегидрогеназой (ChnE).
Аспекты по изобретению относятся к метаболически сконструированной клетке-хозяину для продукции аминокапроновой кислоты из α-кетопимелата, причем клетка-хозяин дополнительно содержит нуклеиновую кислоту, кодирующую фермент аминотрансферазу и декарбоксилазу. В некоторых вариантах осуществления 2-аминотрансфераза катализирует превращение α-кетопимелата в 2-аминопимелат, и декарбоксилаза катализирует превращение 2-аминопимелата в аминокапроновую кислоту. Аминотрансферазу можно выбрать из группы, состоящей из α-аминоадипатаминотрансферазы-1 (AADAT), аминоадипатаминотрансферазы (LysN), диаминопимелатдегидрогеназы (ddb и dapdh) и их гомологов. В некоторых вариантах осуществления декарбоксилаза представляет собой глутаматдекарбоксилазу. В некоторых вариантах осуществления глутаматдекарбоксилаза кодируется геном или фрагментом гена, выбранным из группы, состоящей из Gad6/7, GadA, GadB и lysA. В некоторых вариантах осуществления декарбоксилаза катализирует превращение α-кетопимелата в полуальдегид адипата, и аминотрансфераза катализирует превращение полуальдегида адипата в аминокапроновую кислоту. Кетодекарбоксилазу можно выбрать из группы 2-кетоглутаратдекарбоксилазы (kgd), 2-кетоизовалератдекарбоксилазы (kivD), декарбоксилазы трансаминированной аминокислоты (ARO10), бензоилформиатдекарбоксилазы (mdlC), 2-кетоаргининдекарбоксилазы (aruI), фосфонопируватдекарбоксилазы (fom2), изофермента пируватдекарбоксилазы (PDC6, PDC1), изофермента 2 пируватдекарбоксилазы (PDC5, PDC1, PDC6, Aro10, KivD), индолпируватдекарбоксилазы (ipdC) и их гомологов. Аминотрансферазу можно выбрать из группы, состоящей из трансаминазы ГАМК, Lys6-дегидрогеназы, трансаминазы орнитиноксокислоты, лизинаминотрансферазы, 4-аминобутиратаминотрансферазы, 4-аминобутиратаминотрансферазы, 4-аминобутиратаминотрансферазы, сахаропиндегидрогеназы (LYS9 и LYS1) или любых гомологичных им белков.
Аспекты по изобретению относятся к продукции гексаметилендиамина из аминокапроновой кислоты, причем клетка-хозяин содержит нуклеиновую кислоту, кодирующую альдегиддегидрогеназу и аминотрансферазу. В некоторых вариантах осуществления альдегиддегидрогеназа катализирует превращение аминокапроновой кислоты в 6-аминогексанал, и аминотрансфераза катализирует превращение 6-аминогексанала в 6-гексаметилендиамин. В предпочтительных вариантах осуществления альдегиддегидрогеназа представляет собой фермент ALDH (EC 1.2.1-). В некоторых вариантах осуществления аминотрансферазу можно выбрать из группы, состоящей из α-аминоадипатаминотрансферазы-1 (AADAT), аминоадипатаминотрансферазы (LysN), диаминопимелатдегидрогеназы (ddb, dapdh) и гомологичных им белков.
Аспекты изобретения относятся к метаболически сконструированной клетке-хозяину для продукции гексаметилендиамина из α-кетопимелата, причем клетка-хозяин содержит нуклеиновую кислоту, кодирующую аминотрансферазу, редуктазу, дегидрогеназу и декарбоксилазу. В некоторых вариантах осуществления аминотрансфераза катализирует превращение α-кетопимелата в 2-аминопимелат, редуктаза катализирует превращение 2-аминопимелата в 2-амино-7-оксогептаноат, дегидрогеназа катализирует превращение 2-амино-7-оксогептаноата в 2,7-диаминогептаноат, и декарбоксилаза катализирует превращение 2,7-диаминогептаноата в гексаметилендиамин. Аминотрансферазу можно выбрать из группы, состоящей из α-аминоадипатаминотрансферазы-1 (AADAT), аминоадипатаминотрансферазы (LysN), диаминопимелатдегидрогеназы (ddb, dapdh) и их вариантов. В некоторых вариантах осуществления редуктаза представляет собой аминоадипатредуктазу или ее гомолог. В некоторых вариантах осуществления аминоадипатредуктаза кодируется Sc-Lys2. В некоторых вариантах осуществления дегидрогеназа представляет собой сахаропиндегидрогеназу или ее гомолог и кодируется Sc-Lys9 или Sc-Lysl или их вариантами. Декарбоксилазу можно выбрать из группы, состоящей из лизиндекарбоксилазы, орнитиндекарбоксилазы и их вариантов.
Некоторые аспекты по изобретению относятся к метаболически сконструированной клетке-хозяину для продукции 6-гидроксигексаноата из α-кетопимелата, причем клетка-хозяин содержит нуклеиновую кислоту, кодирующую алкогольдегидрогеназу. Другие аспекты по изобретению относятся к метаболически сконструированной клетке-хозяину для продукции 1,6-гександиола из 6-гидроксигексаноата, причем клетка-хозяин содержит нуклеиновую кислоту, кодирующую алкогольдегидрогеназу или альдегиддегидрогеназу. Алкогольдегидрогеназу можно выбрать из 6-гидроксигексаноатдегидрогеназы, бутанолдегидрогеназы, ADHIV-дегидрогеназы, пропандиолоксидоредуктазы, ADH6 и их гомологов.
Аспекты по изобретению относятся к способам получения α,ω-дифункциональных Cn-алканов из α-кетоглутарата, где концевые функциональные группы α и ω выбраны из группы -OH, -COOH и -NH3 и где n представляет собой целое число в диапазоне от 4 до 7, включающим культивирование клетки-хозяина в условиях, подходящих для получения α,ω-дифункциональных Cn-алканов; и выделение α,ω-дифункциональных Cn-алканов.
В некоторых аспектах изобретение относится к клетке-хозяину, которая содержит одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из гомоцитратсинтазы (EC 2.3.3.-), гомоаконитазы (EC 2.3.3.14), гомоизоцитратдегидрогеназы (EC 1.1.1.-) и их сочетания. В некоторых вариантах осуществления метаболически сконструированные клетки-хозяева продуцируют α-кетоадипат, α-кетопимелат, α-кетосуберат или их сочетание из α-кетоглутарата. В некоторых вариантах осуществления сконструированная клетка-хозяин содержит одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из декарбоксилазы α-кетокислоты (EC 4.1.1.-) и дегидрогеназы (EC 1.1.1.-) и их сочетания. В некоторых вариантах осуществления сконструированная клетка-хозяин продуцирует Cn-дикарбоновую кислоту, где n представляет собой целое число в диапазоне от 4 до 7. Например, сконструированная клетка-хозяин продуцирует адипат. В других вариантах осуществления сконструированная клетка-хозяин продуцирует Cn-гидроксикарбоновую кислоту, где n представляет собой целое число в диапазоне от 4 до 7. Например, сконструированные клетки-хозяева продуцируют 6-гидроксигексаноат. Еще в других вариантах осуществления сконструированная клетка-хозяин продуцирует Cn-алкандиол, где n представляет собой целое число в диапазоне от 4 до 7, например, 1,6-гександиол.
В некоторых аспектах по изобретению сконструированная клетка-хозяин, продуцирующая α-кетоадипат, α-кетопимелат, α-кетосуберат или их сочетание из α-кетоглутарата, содержит одну или несколько экзогенных последовательностей нуклеиновых кислот, кодирующих, по меньшей мере, один полипептид, выбранный из декарбоксилазы α-кетокислоты (EC 4.1.1.-), аминотрансферазы (EC 1.4.1.-), аминотрансферазы (EC 2.6.1.-) и их сочетания. В предпочтительных вариантах осуществления сконструированная клетка-хозяин продуцирует аминокапроновую кислоту. В некоторых вариантах осуществления сконструированная клетка-хозяин содержит, кроме того, одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из альдегиддегидрогеназы (EC 1.2.1.3), и продуцирует Cn-аминоальдегид или Cn-диаминоалкан, где n представляет собой целое число в диапазоне от 4 до 7. Например, сконструированная клетка-хозяин продуцирует гексаметилендиамин или 6-гидроксигексамин.
В некоторых вариантах осуществления сконструированная клетка-хозяин содержит одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из аминоадипаттрансферазы (EC 2.6.1.39), диаминопимелатдегидрогеназы (EC 1.4.1.16), глутаматдекарбоксилазы (EC 4.1.1.-) и их сочетания. В некоторых вариантах осуществления клетка-хозяин продуцирует Cn-аминокарбоновую кислоту, где n представляет собой целое число в диапазоне от 4 до 7. Например, сконструированная клетка-хозяин по пункту продуцирует аминокапроновую кислоту.
Некоторые аспекты по изобретению относятся к сконструированной клетке-хозяину для продукции α,ω-дифункциональных Cn-алканов из α-кетокислоты, содержащей одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из α-аминоадипатаминотрансферазы (EC 2.6.1.39), диаминопимелатдегидрогеназы (EC 1.4.1.16), аминоадипатредуктазы (EC 1.2.1.31), сахаропиндегидрогеназы (EC 1.5.1.-), лизиндекарбоксилазы (EC 4.1.1.18), орнитиндекарбоксилазы (EC4.1.1.17) и их сочетания. В некоторых вариантах осуществления сконструированная клетка-хозяин продуцирует Cn-аминоальдегид или Cn-диаминоалкан, где n представляет собой целое число в диапазоне от 4 до 7. Например, сконструированная клетка-хозяин продуцирует гексаметилендиамин. В некоторых аспектах по изобретению сконструированная клетка-хозяин дополнительно содержит одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из 3-оксоацил-[ацилпереносящий белок]-редуктазы (EC 1.1.1.100), синтазы жирных кислот (EC 2.3.1.-), дегидратазы (EC 4.2.1.59), 3-гидроксиоктаноил-[ацилпереносящий белок]-дегидратазы (EC 4.2.1.59), еноил-[ацилпереносящий белок]-редуктазы (EC 1.3.1.9) и их сочетания. В некоторых предпочтительных вариантах осуществления сконструированная клетка-хозяин продуцирует Cn-дикарбоновую кислоту, где n представляет собой целое число в диапазоне от 5 до 8. Например, сконструированная клетка-хозяин продуцирует адипат. В некоторых вариантах осуществления сконструированная клетка-хозяин дополнительно содержит одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из альдегиддегидрогеназы (EC 1.2.1.3), и продуцирует Cn-гидроксикарбоновую кислоту, где n представляет собой целое число в диапазоне от 5 до 8, например, 6-гидроксигексаноат. В некоторых вариантах осуществления сконструированная клетка-хозяин продуцирует Cn-алкандиол, где n представляет собой целое число в диапазоне от 5 до 8, например 1,6-гександиол.
Некоторые аспекты по изобретению относятся к сконструированной клетке-хозяину для продукции α,ω-дифункциональных Cn-алканов из α-кетокислоты, содержащей одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из α-аминоадипатаминотрансферазы (EC 2.6.1.39), диаминопимелатдегидрогеназы (EC l.4.1.16), аминоадипатредуктазы (EC 1.2.1.31), сахаропиндегидрогеназы (EC 1.5.1.-), лизиндекарбоксилазы (EC 4.1.1.18), орнитиндекарбоксилазы (EC 4.1.1.17) и их сочетания, выбранный из 3-оксоацил-[ацилпереносящий белок]редуктазы (EC 1.1.1.100), синтазы жирных кислот (EC 2.3.1.-), дегидратазы (EC 4.2.1.59), 3-гидроксиоктаноил-[ацилпереносящий белок]дегидратазы (EC 4.2.1.59), еноил-[ацилпереносящий белок]редуктазы (EC 1.3.1.9) и их сочетания, по меньшей мере, один полипептид, выбранный из альдегиддегидрогеназы, и одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из аминотрансферазы (EC 1.4.1.-) и аминотрансферазы (EC 2.6.1.-). В некоторых вариантах осуществления клетка-хозяин продуцирует Cn-аминокарбоновую кислоту, где n представляет собой целое число в диапазоне от 5 до 8, например аминокапроновую кислоту. В некоторых вариантах осуществления сконструированная клетка-хозяин дополнительно содержит одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из альдегиддегидрогеназы (EC 1.2.1.3), аминотрансферазы (EC 1.4.1.-) и аминотрансферазы (EC 2.6.1.-) и их сочетания. В некоторых вариантах осуществления сконструированная клетка-хозяин продуцирует 1,n-диаминоалкан, где n представляет собой целое число в диапазоне от 5 до 8, например гексаметилендиамин. В других вариантах осуществления сконструированная клетка-хозяин продуцирует n-аминоспирт, где n представляет собой целое число в диапазоне от 5 до 8, например 6-аминогексанол.
Некоторые аспекты по изобретению относятся к сконструированной клетке-хозяину, содержащей одну или несколько экзогенных последовательностей нуклеиновой кислоты, кодирующих, по меньшей мере, один полипептид, выбранный из аминотрансферазы (EC 2.6.1.7), альдегиддегидрогеназы (EC 1.2.1.3), глутаматполуальдегидальдегидамутазы (EC 5.4.3.8), 3-оксоацил-[ацилпереносящий белок]редуктазы (EC 1.1.1.100), синтазы жирных кислот (EC 2.3.1.-), дегидратазы (EC 4.2.1.59), 3-гидроксиоктаноил-[ацилпереносящий белок]дегидратазы (EC 4.2.1.59), еноил-[ацилпереносящий белок]редуктазы (EC 1.3.1.9) и их сочетания. В некоторых вариантах осуществления сконструированная клетка-хозяин продуцирует n-аминокарбоновую кислоту, где n представляет собой целое число в диапазоне от 5 до 8, например аминокапроновую кислоту.
Некоторые аспекты по изобретению относятся к способу получения α,ω-дифункционального Cn-алкана из α-кетоглутарата, где концевые функциональные группы α и ω выбраны из группы -OH, -COOH и -NH3 и где n представляет собой целое число в диапазоне от 5 до 8, причем способ включает культивирование сконструированной клетки-хозяина в условиях, достаточных для получения α,ω-дифункционального Cn-алкана; и выделение α,ω-дифункционального Cn-алкана.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Изобретение можно более полно понять, исходя из следующего ниже подробного описания и фигур, которые являются частью заявки.
На фигуре 1 показан путь биосинтеза адипиновой кислоты. Стадии от a до 1 описывают путь 2-кетоудлинения из пути биосинтеза кофермента B из Methanococcus jannaschii. Отмеченные стадии a, b, c, d, e, f, g, h, m и n представляют собой превращение субстрата в продукт, описанное ниже.
На фигуре 2 представлена схема последовательности операций для получения с помощью микроорганизмов дифункциональных C(n-1)-алканов, начиная с Cn-α-кетокислоты.
На фигуре 3 представлена схема последовательности операций для получения с помощью микроорганизмов дифункциональных гексанов, начиная с 2-кетопимелата.
На фигуре 4 представлена схема последовательности операций для получения с помощью микроорганизмов дифункциональных Cn-алканов, начиная с Cn-α-кетокислоты.
На фигуре 5 представлена схема последовательности операций для получения с помощью микроорганизмов дифункциональных Cn-гексанов, начиная с 2-кетоадипата.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем документе следующие ниже термины и фразы будут иметь значения, установленные ниже. Если не определено иначе, все технические и научные термины, используемые в настоящем документе, имеют то же самое значение, как обычно понятно специалисту в данной области.
Формы единственного числа включают указание на множественное число, если контекст явным образом не предписывает иначе.
Термины «содержит» и «содержащий» применяют во всеохватывающем общедоступном смысле, означая, что могут быть включены дополнительные элементы.
Термин «включая» применяют в значении «включая в качестве неограничивающих примеров». «Включая» и «включая в качестве неограничивающих примеров» используют взаимозаменяемо.
Все публикации, упомянутые в настоящем документе, включены в настоящий документ в качестве ссылки. Публикации, обсуждаемые в настоящем документе, приведены исключительно для их описания до даты подачи заявки по настоящему изобретению. Ничто в настоящем документе не следует рассматривать в качестве признания, что настоящее изобретение не имеет право противопоставлять факт создания изобретения с более ранним приоритетом такой публикации на основании предшествующего изобретения.
В аспектах по изобретению предлагаются способы и материалы для получения органических алифатических, представляющих интерес соединений быстрым, недорогим и экологически безопасным способом. В связи с этим настоящее изобретение удовлетворяет большому количеству коммерческих и промышленных потребностей. Термин «органическая молекула» относится, например, к любой молекуле, которая преимущественно состоит из углерода и водорода, такой как, например, алканы. Представляющие интерес органические соединения, такие как дифункциональные алканы, диолы, дикарбоновые кислоты и т.д., можно использовать для синтеза пластмассы, нейлона и других продуктов, обычно получаемых из нефти и углеводородов. Аспекты по изобретению относятся к синтезу дифункциональных n-алканов, в которых углеводородные цепи Cn получают из углеводородной цепи Cn или Cn+1, в которой n представляет собой число от приблизительно 1 до приблизительно 8, такое как от приблизительно 2 до приблизительно 5 или от приблизительно 3 до приблизительно 4. В предпочтительном варианте осуществления дифункциональные n-алканы получают из α-(n+1) или n-кетокислоты.
Аспекты по изобретению относятся к получению представляющих интерес дифункциональных алканов в микроорганизмах и предлагают способы получения дифункциональных алканов из источника углеводов в микроорганизме. В настоящем документе «дифункциональные алканы» относится к алканам, содержащим две функциональных группы. Термин «функциональная группа» относится, например, к группе атомов, расположенных некоторым способом, который определяет химические свойства группы и молекулы, к которой она присоединена. Примеры функциональных групп включают атомы галогена, гидроксильные группы (-OH), группы карбоновой кислоты (-COOH) и аминогруппы (-NH2) и т.п. «Спирт» относится, например, к алкильной группе, в которой один или несколько атомов водорода заменено на группу -OH. Термин «первичный спирт» относится, например, к спиртам, в которых группа -OH связана с концевым атомом углерода или атомом углерода на обрыве цепи, таким как в 1-бутаноле, 1-гексаноле и т.п. Термин «вторичный спирт» относится, например, к спиртам, в которых группа -OH связана с атомом углерода, который связан с одним атомом водорода и двумя другими атомами углерода, таким как в 2-бутаноле, 2-гексаноле и т.п. Термин «третичный спирт» относится, например, к спиртам, в которых группа -OH связана с атомом углерода, который связан с тремя другими атомами углерода, таким как в метилпропаноле (трет-бутаноле) и т.п. «Амин» относится, например, к алкильной группе, в которой один или несколько атомов водорода заменены на группу -NH2. «Карбонильное соединение» относится, например, к органическому соединению, содержащему карбонильную группу, C=O, такому как, например, альдегиды, которые имеют общую формулу RCOH; кетоны, которые имеют общую формулу RCOR'; карбоновые кислоты, которые имеют общую формулу RCOOH; и сложные эфиры, которые имеют общую формулу RCOOR'.
Способ предусматривает использование микроорганизмов, способных продуцировать один из следующих ниже, представляющих интерес дифункциональных алканов, в частности, адипиновую кислоту, аминокапроновую кислоту, HMD, 6-гидроксигексаноат. Другие представляющие интерес дифункциональные алканы включают 1,3-пропандиол, глицерин, акриловую кислоту, кадаверин, 3-гидроксипропионовую кислоту, пентаметилендиамин, малеиновую кислоту, янтарную кислоту, адипиновую кислоту, себациновую кислоту, глутаровую кислоту и субериновую кислоту и т.д. Описано несколько способов химического синтеза, например, для адипиновой кислоты и ее промежуточных продуктов, таких как муконовая кислота и полуальдегид адипата; для капролактама и его промежуточных продуктов, таких как 6-аминокапроновая кислота; для гексан-1,6-диаминогексана или гексанметилендиамина; для 3-гидроксипропионовой кислоты и ее промежуточных продуктов, таких как полуальдегид малоната, но только малое количество биологических способов было описано для некоторых из данных органических химических веществ. Поэтому в аспектах по изобретению предлагаются метаболически сконструированные способы, выделенные нуклеиновые кислоты или сконструированные нуклеиновые кислоты, полипептиды или сконструированные полипептиды, клетки-хозяева или генетически сконструированные клетки-хозяева, способы и материалы для получения дифункциональных алканов из неистощимого исходного материала. Источники углерода, подходящие для стартовой точки, включают углеводы и синтетические промежуточные продукты. Примеры углеводов, которые клетки способны метаболизировать, включают сахара, декстрозы, триглицериды и жирные кислоты. Промежуточные продукты из метаболического пути, такие как пируват, оксалоацетат, 2-кетоглутарат, также можно использовать в качестве стартовой точки. Аспекты по изобретению относятся к сконструированным полипептидам и полинуклеотидам, кодирующим ферменты, обладающие активностью или повышенной активностью в отношении природных или неприродных субстратов или обладающие широкой субстратной специфичностью (например, каталитическая разнородность, такая как субстратная разнородность). Термин «полипептид» и термины «белок» и «пептид», которые в настоящем документе используют взаимозаменяемо, относится к полимеру аминокислот, включая, например, продукты генов, встречающиеся в природе белки, гомологи, ортологи, паралоги, фрагменты и другие эквиваленты, варианты и аналоги упомянутого выше. Термин «полипептид, обладающий ферментативной активностью», относится к любому полипептиду, который катализирует химическую реакцию других веществ без собственного разрушения или изменения после завершения реакции. Типично, полипептид, обладающий ферментативной активностью, катализирует образование одного или нескольких продуктов из одного или нескольких субстратов. В некоторых аспектах по изобретению свойства каталитической разнородности некоторых ферментов можно комбинировать с белковым конструированием и можно использовать в новых метаболических путях и биосинтетических применениях. В некоторых вариантах осуществления существующие ферменты модифицируют для применения в органическом биосинтезе. В некоторых предпочтительных вариантах осуществления ферменты, вовлеченные в продукцию представляющих интерес дифункциональных n-алканов, включают в качестве неограничивающих примеров 2-аминодекарбоксилазы, 2-кетодекарбоксилазы, концевые аминотрансферазы, 2-аминотрансферазы, алкогольдегидрогеназы, альдегиддегидрогеназы, аминоальдегиддегидрогеназы, дегидрогеназы и дегидратазы. В некоторых вариантах осуществления механизм реакции фермента можно изменить для катализа новых реакций, для изменения, увеличения или повышения субстратной специфичности. Следует принимать во внимание, что если известна структура фермента (например, кристаллическая структура), свойства ферментов можно модифицировать при помощи рационального реконструирования (см. патентную заявку США US20060160138, US20080064610 и US20080287320, которые в полном объеме включены в качестве ссылки). Модификации или улучшения свойств фермента могут являться результатом встраивания модификаций в полипептидную цепь, которые могут, фактически, исказить структуру-функцию фермента и/или взаимодействие с другой молекулой (например, субстрат в сравнении с неприродным субстратом). В данной области хорошо известно, что некоторые участки полипептида могут являться крайне необходимыми для ферментативной активности. Например, небольшие искажения состава аминокислот, вовлеченных в катализ и/или входящих в состав связывающих субстраты домены, будут оказывать достоверное влияние на функцию фермента. Некоторые аминокислотные остатки могут находиться в важных положениях для сохранения вторичной или третичной структуры фермента, и, таким образом, также приводить к заметным изменениям свойств фермента, если их модифицировать. В некоторых вариантах осуществления возможные компоненты пути являются вариантами любых из упомянутых выше. Такие варианты можно получить случайным мутагенезом или можно получить при рациональном конструировании для появления ферментативной активности, обладающие, например, измененной субстратной специфичностью, повышенной ферментативной активностью, большей устойчивостью и т.д. Таким образом, в некоторых вариантах осуществления количество модификаций упомянутого исходного фермента, которые приводят к появлению фермента, обладающего необходимым свойством, может включать одну или несколько аминокислот, 2 или более аминокислот, 5 или более аминокислот, 10 или более аминокислот или 20 или более аминокислот, вплоть до 10% от общего количества аминокислот, вплоть до 20% от общего количества аминокислот, вплоть до 30% от общего количества аминокислот, вплоть до 40% от общего количества аминокислот, составляющих упомянутый фермент, или вплоть до 50% от общего количества аминокислот, составляющих упомянутый фермент.
Специалистам в данной области будет понятно, что сконструированные пути, представленные в качестве примера в настоящем документе, описаны в отношении, но не ограничены, разновидности определенных генов и охватывают гомологи или ортологи нуклеиновой кислоты или аминокислотных последовательностей. Гомологи и ортологи последовательностей обладают относительно высокой степенью идентичности/схожести последовательности при выравнивании с использованием известных в данной области способов.
Аспекты по изобретению относятся к новым микроорганизмам или «генетически модифицированным» микроорганизмам или клеткам-хозяевам, которые были сконструированы для того, чтобы приобрести новые метаболические возможности или новые метаболические пути. В настоящем документе термин «генетически модифицированные» микроорганизмы относится к микроорганизмам, имеющим, по меньшей мере, одно генетическое изменение, обычно не обнаруживаемое в штамме дикого типа вида сравнения. В некоторых вариантах осуществления генетически сконструированные микроорганизмы создают для экспрессии или повышенной экспрессии, по меньшей мере, одного определенного фермента в важной точке метаболического пути и/или блокируют синтез других ферментов, чтобы преодолеть или обойти метаболические «узкие места». Термин «метаболический путь» относится к серии двух или более ферментативных реакций, в которых продукт одной ферментативной реакции становится субстратом для другой ферментативной реакции. На каждой стадии метаболического пути образуются промежуточные соединения, и они используются в качестве субстратов для последующей стадии. Такие соединения можно назвать «метаболическими промежуточными продуктами». Продукты на каждой стадии также называют «метаболитами».
В аспектах по изобретению предлагаются способы создания и формирования сконструированных метаболических путей. В некоторых аспектах по изобретению альтернативные пути для получения представляющего интерес продукта из одного или нескольких доступных и неистощимых субстратов можно сформировать в одной или нескольких клетках-хозяевах или представляющих интерес микроорганизмах. Следует принимать во внимание, что конструирование пути для получения представляющих интерес дифункциональных алканов может предполагать использование множества ферментов и поэтому продвижение через путь может быть неоптимальным для получения представляющего интерес продукта. Таким образом, в некоторых аспектах по изобретению продвижение оптимально сочетают посредством модуляции уровня активности ферментов пути относительно друг друга. Примеры такой модуляции предлагаются на всем протяжении заявки.
В аспектах по изобретению предлагаются генетически модифицированные клетки-хозяева или микроорганизмы и способы их применения для продукции дифункциональных n-алканов из α-кетокислот. Клетка-хозяин в настоящем документе относится к эукариотической клетке in vivo или in vitro, прокариотической клетке или к клетке из многоклеточного организма (например, клеточной линии), культивируемой в виде одноклеточного организма. Клетка-хозяин может являться прокариотической (например, бактериальной, такой как E. coli или B. subtilis) или эукариотической (например, дрожжевой, клеткой млекопитающих или насекомых). Например, клетки-хозяева могут представлять собой бактериальные клетки (например, Escherichia coli, Bacillus subtilis, Mycobacterium spp., M. tuberculosis или другие подходящие бактериальные клетки), археи (например, Methanococcus Jannaschii или Methanococcus Maripaludis или другие подходящие клетки архей), дрожжевые клетки (например, виды Saccharomyces, такие как S. cerevisiae, S. pombe, виды Picchia, виды Candida, такие как C. albicans, или другие подходящие виды дрожжей). Эукариотические или прокариотические клетки-хозяева могут быть, или были, генетически модифицированы (также обозначаемые как «рекомбинантная клетка-хозяин», «метаболически сконструированные клетки» или «генетически сконструированные клетки») и их используют в качестве реципиентов нуклеиновой кислоты, например, экспрессирующего вектора, который содержит нуклеотидную последовательность, кодирующую один или несколько продуктов генов биосинтетического или сконструированного пути. Эукариотические и прокариотические клетки-хозяева также обозначают потомство исходных клеток, которые были созданы генно-инженерным способом при помощи нуклеиновой кислоты. В некоторых вариантах осуществления клетку-хозяина можно выбрать вследствие ее метаболических свойств. Например, если выбор или скрининг относится к определенному метаболическому пути, возможно, будет полезным применять клетку-хозяина, в которой имеет место соответствующий путь. Такая клетка-хозяин может обладать определенной физиологической адаптацией, которая позволит ей подвергать процессу или импортировать, или экспортировать один или несколько промежуточных продуктов или продуктов пути. Однако в других вариантах осуществления клетку-хозяина, которая не экспрессирует никаких ферментов, ассоциированных с определенным представляющим интерес путем, можно выбрать для того, чтобы иметь возможность идентифицировать все компоненты, необходимые для такого пути, используя соответствующие наборы генетических элементов и не полагаясь на клетку-хозяина, чтобы обеспечить прохождение одной или нескольких отсутствующих стадий.
В некоторых вариантах осуществления анаэробные бактериальные организмы являются метаболически сконструированными. В настоящем документе анаэробный организм представляет собой любой организм, которому для роста не требуется кислород (т.е. анаэробные условия). Преимущественно, бактериальная клетка может представлять собой клетку E. coli, C. glutanicum, B. flavum или B. lactofermentum; данные штаммы в настоящее время применяются в промышленном масштабе для получения аминосоединениий, используя процессы бактериальной ферментации. Например, C. glutanicum широко используют для продукции аминокислоты (например, L-глутамата, L-лизина, см. Eggleging L et al., 2005, Handbook for Corynebacterium glutanicum. Boca Raton, USA: CRC Press).
Метаболически сконструированную клетку по изобретению получают, трансформируя клетку-хозяина при помощи, по меньшей мере, одной нуклеотидной последовательности, кодирующей ферменты, вовлеченные в конструируемые метаболические пути. В настоящем документе термин «нуклеотидная последовательность», «последовательность нуклеиновой кислоты» и «генетическая конструкция» используют взаимозаменяемо, и он обозначает полимер РНК или ДНК, одно- или двухцепочечный, необязательно содержащий синтетические, неприродные или измененные нуклеотидные основания. Нуклеотидная последовательность может содержать один или несколько сегментов кДНК, геномной ДНК, синтетической ДНК или РНК. В предпочтительном варианте осуществления нуклеотидная последовательность оптимизирована в отношении кодона, чтобы отражать типичную частоту использования кодона клетки-хозяина, без изменения полипептида, кодируемого нуклеотидной последовательностью. В определенных вариантах осуществления термин «оптимизация кодона» или «оптимизированный в отношении кодона» относится к модификации состава кодона последовательности нуклеиновой кислоты, не модифицируя последовательность полипептида, кодируемого нуклеиновой кислотой, чтобы увеличить экспрессию в определенной клетке-хозяине. В определенных вариантах осуществления термин предназначен для того, чтобы охватить модификацию состава кодона последовательности нуклеиновой кислоты в качестве средства контроля уровня экспрессии полипептида (например, либо увеличивать, либо снижать уровень экспрессии). Таким образом, аспекты по изобретению включают нуклеиновые последовательности, кодирующие ферменты, вовлеченные в конструируемые метаболические пути. В некоторых вариантах осуществления метаболически сконструированная клетка может экспрессировать один или несколько полипептидов, обладающих ферментативной активностью, необходимой для выполнения стадий, описанных ниже. Например, определенная клетка может содержать одну, две, три, четыре, пять или больше пяти последовательностей нуклеиновой кислоты, причем каждая кодирует полипептид(ы), необходимые для осуществления превращения α-кетокислоты в дифункциональный алкан. Альтернативно одна молекула нуклеиновой кислоты может кодировать один или более одного полипептида. Например, одна молекула нуклеиновой кислоты может содержать последовательности нуклеиновой кислоты, которые кодируют два, три, четыре или даже пять различных полипептида. Последовательности нуклеиновой кислоты, применимые в изобретении, описываемые в настоящем документе, можно получать из различных источников, таких как, например, амплификация последовательности кДНК, библиотеки ДНК, синтез de novo, исключение геномного сегмента. Последовательности, получаемые из таких источников, можно затем модифицировать, применяя стандартную технологию молекулярной биологии и/или технологию рекомбинантных ДНК, чтобы получить нуклеотидные последовательности, имеющие требуемые модификации. Иллюстративные способы модификации последовательностей нуклеиновой кислоты включают, например, сайт-направленный мутагенез, ПЦР-мутагенез, делецию, вставку, замену, перестановку части последовательности с использованием ферментов рестрикции, необязательно в сочетании с лигированием, гомологичной рекомбинацией, сайт-специфической рекомбинацией или различным их сочетанием. В других вариантах осуществления последовательности нуклеиновой кислоты могут представлять собой синтетическую последовательность нуклеиновой кислоты. Синтетические полинуклеотидные последовательности можно получить, применяя целый ряд способов, описанных в патенте США 7323320, в одной из заявок, находящихся одновременно на рассмотрении патентного ведомства, с регистрационным номером 11/804996 и в патентных публикациях США №№ 1006/0160138 и 2007/0269870, которые включены в настоящий документ в качестве ссылки в полном объеме.
Способы трансформации клеток бактерии, растения и животного хорошо известны в данной области. Общеизвестные способы бактериальной трансформации включают электропорацию и химическую модификацию.
В некоторых вариантах осуществления генетически модифицированная клетка-хозяин является генетически модифицированной таким образом, чтобы она продуцировала, когда ее культивируют in vitro в подходящей среде, представляющий интерес продукт или промежуточный продукт на уровне, по меньшей мере, 0,1 г/л, по меньшей мере, 1 г/л или, по меньшей мере, 10 г/л. Следует принять во внимание, что уровень представляющего интерес продукта или его метаболических промежуточных продуктов, продуцируемый генетически модифицированной клеткой-хозяином, можно контролировать различными способами. В некоторых вариантах осуществления уровень экспрессии контролируют количеством копий последовательности нуклеиновой кислоты, кодирующей один или несколько ферментов, вовлеченных в конструируемый путь (например, высококопийный экспрессирующий вектор по сравнению со средне- или низкокопийными экспрессирующими векторами). Предпочтительно последовательности нуклеиновой кислоты встраивают внутрь клетки, используя вектор. Низкокопийные экспрессирующие векторы, как правило, обеспечивают получение менее чем 20 копий векторов на клетку (например, от 1 до приблизительно 5, от 5 до приблизительно 10, от 10 до приблизительно 15, от 15 до приблизительно 20 копий экспрессирующего вектора на клетку. Подходящие низкокопийные экспрессирующие векторы для прокариотических клеток (например, E. Coli) включают в качестве неограничивающих примеров pAYC184, pBeloBac11, pBR332, pBAD33, pBBRlMCS и их производные, pSC101, SuperCos (космида) и pWE15 (космида). Среднее количество копий экспрессирующих векторов, как правило, обеспечивает получение от приблизительно 20 до приблизительно 50 копий экспрессирующих векторов на клетку или приблизительно от 20 до 80 копий экспрессирующих векторов на клетку). Подходящие среднекопийные экспрессирующие векторы для прокариотических клеток (например, E. Coli) включают в качестве неограничивающих примеров pTrc99A, pBAD24 и векторы, содержащие участок начала репликации ColE1, и их производные. Большое количество копий экспрессирующих векторов, как правило, обеспечивает получение приблизительно от 80 до приблизительно 200 или более копий экспрессирующих векторов на клетку. Подходящие высококопийные экспрессирующие векторы для прокариотических клеток (например, E. Coli) включают в качестве неограничивающих примеров векторы pUC, PCV1, pBluescript, pGEM и pTZ.
В аспектах по изобретению предлагаются экспрессирующие кассеты, содержащие нуклеиновую кислоту или ее субпоследовательность, кодирующую полипептид, вовлеченный в конструируемый путь. В некоторых вариантах осуществления экспрессирующая кассета может содержать нуклеиновую кислоту, которая функционально связана с транскрипционным элементом (например, промотором) и с терминатором. В настоящем документе термин «кассета» относится к нуклеотидной последовательности, способной экспрессировать определенный ген, если указанный ген встроен так, чтобы оказаться функционально связанным с одной или несколькими регуляторными последовательностями, присутствующими в нуклеотидной последовательности. Таким образом, например, экспрессирующая кассета может содержать необходимый гетерологичный ген, который будет экспрессироваться в клетке-хозяине. В некоторых вариантах осуществления одну или несколько экспрессирующих кассет можно встраивать в вектор известными рекомбинантными способами. Промотор представляет собой последовательность нуклеотидов, которая инициирует и контролирует транскрипцию требуемой последовательности нуклеиновой кислоты при помощи фермента РНК-полимераза. В некоторых вариантах осуществления промоторы могут быть индуцируемыми. В других вариантах осуществления промоторы могут быть конститутивными. Неограничивающие примеры подходящих промоторов для применения в прокариотических клетках-хозяевах включают промотор РНК-полимеразы бактериофага T7, промотор trp, промотор lac-оперона и т.п. Неограничивающий пример подходящего сильного промотор для применения в прокариотических клетках-хозяевах включает промотор lacUV5, T5, T7, Trc, Tac и т.п. Неограничивающие примеры подходящих промоторов для применения в эукариотических клетках включают предранний промотор CMV, ранний или поздний промотор SV40, промотор тимидинкиназы HSV и т.п. Участки контроля над терминацией могут также происходить из различных генов, присущих предпочтительным хозяевам.
В некоторых вариантах осуществления первый фермент сконструированного пути может находиться под контролем первого промотора, и второй фермент сконструированного пути может находиться под контролем второго промотора, где первый и второй промоторы обладают различной силой. Например, первый промотор может быть сильнее, чем второй промотор, или второй промотор может быть сильнее, чем первый промотор. Таким образом, уровень первого фермента можно повысить относительно уровня второго фермента в сконструированном пути, увеличивая число копий первого фермента и/или увеличивая силу промотора, с которым первый фермент функционально связан, относительно силы промотора, с которым функционально связан второй фермент. В некоторых других вариантах осуществления множество ферментов сконструированного пути может находиться под контролем одного и того же промотора. В других вариантах осуществления изменение участка связывания рибосомы влияет на трансляцию и экспрессию различных ферментов в пути. Изменение участка связывания рибосомы можно использовать исключительно для контроля относительной экспрессии ферментов в пути или можно использовать совместно с указанными выше модификациями промоторов и оптимизацией кодона, которые также влияют на уровни экспрессии.
В иллюстративном варианте осуществления экспрессия ферментов возможного пути может зависеть от наличия субстрата, с которым будет взаимодействовать фермент пути в реакционной смеси. Например, экспрессию фермента, который катализирует превращение A в B, можно индуцировать в присутствии A в среде. Экспрессию таких ферментов пути можно индуцировать, либо добавляя соединение, которое вызывает индукцию, либо посредством естественного накопления соединения во время прохождения пути биосинтеза (например, индуктор может представлять собой промежуточный продукт, получаемый в ходе процесса биосинтеза с выходом требуемого продукта).
В некоторых вариантах осуществления реализуемые при помощи компьютера методики конструирования можно применять для создания альтернативных путей для представляющей интерес органической молекулы. В некоторых вариантах осуществления базы данных содержат информацию о геноме, и ее связанные части можно использовать для конструирования новых метаболических путей. Примеры баз данных представляют собой MetaCyc (база данных метаболических путей и ферментов, база данных по биокатализу/биодеградации Миннесотского университета (база данных каталитических реакций микроорганизмов и путей биодеградации органических химических соединений), LGAND (состоящая из нескольких частей база данных, в которой предлагается информация о метаболитах и других химических соединениях, взаимосвязи субстрат-продукт, представляющие метаболические и другие реакции, и информацию о молекулах ферментов). База данных компонентов пути может также содержать компоненты с предсказанными, вероятными или неизвестными функциями. Базы также могут содержать псевдокомпоненты с определенными функциями, которые могут обладать неопределенным составом. В некоторых вариантах осуществления программа может конструировать регуляторные и/или функциональные элементы, которые находятся в общественной собственности (например, которые не защищены патентными правами и/или не облагаются лицензионным сбором). Базы данных свободно доступных генетических элементов можно создать и/или использовать в качестве источника последовательностей нуклеиновой кислоты, которые можно комбинировать для того, чтобы разработать альтернативные пути. Альтернативные пути, содержащие различные комбинации известных функциональных и/или регуляторных элементов (например, из различных организмов), можно конструировать, компоновать и/или тестировать. Библиотеки, включающие вариации в участках элементов ферментов, можно использовать для выявления взаимосвязанных эффектов различных типов ферментов или различных вариантов одного и того же фермента. Библиотеки, включающие вариации в участках регуляторных элементов, можно использовать для определения оптимального уровня экспрессии или для контроля регуляции среди ряда генов.
Нуклеиновые кислоты, кодирующие различные пути, можно объединять. В некоторых вариантах осуществления функциональные свойства различных сконструированных путей можно проверять in vivo, трансформируя клетки-хозяева или организмы с использованием подходящих объединенных нуклеиновых кислот и исследуя свойства сконструированных организмов. В некоторых вариантах осуществления функциональные свойства различных сконструированных путей можно проверять in vitro, выделяя компоненты, экспрессируемые объединенными нуклеиновыми кислотами, и тестируя соответствующие комбинации компонентов в системе in vitro.
I. Сконструированный путь синтеза кофермента B для получения 2-кетокислот (от C5 до C8)
Аспекты изобретения относятся к новым сконструированным способам получения дифункциональных n-алканов посредством реакций удлинения цепи α-кетокислоты, вовлеченных в биосинтез кофермента B (см. фигуру 1). В настоящем документе α-кетокислоты или 2-оксокислоты или 2-кетокислоты используют взаимозаменяемо и конструируют органические кислоты, содержащие кетоновую функциональную группу в непосредственной близости от группы карбоновой кислоты. Реакции удлинения цепи α-кетокислоты (также называемые удлинением 2-оксокислоты), вовлеченные в биосинтез кофермента B, применяют в настоящем документе для обозначения пути биосинтеза, в котором α-кетоглутарат (C5-цепь) и ацетилCoA преобразуются в α-кетосуберат (C8-цепь), предшественник кофермента B (7-меркаптогептаноилтреонинфосфат) и, возможно, биотин. Множество организмов могут синтезировать α-кетоглутарат через оксалоацетат, получаемый либо из PEP посредством фермента РЕР-карбоксилазы, либо из пирувата посредством зависимого от биотина фермента пируваткарбоксилазы. α-Кетоглутарат является ключевым промежуточным продуктом в цикле Кребса и служит исходным компонентом для пути удлинения α-кетокислот. Путь кофермента B с удлинением α-кетокислоты включает ферменты, которые катализируют следующие ниже стадии:
(1) конденсация α-кетоглутарата и ацетилCoA с образованием гомоцитрата (например, под действием гомоцитратсинтазы, такой как, например, AksA, NifV, Hcs, Lys 20/21);
(2) дегидратация и гидратация до (2R,3S) гомоизоцитрата с цис-гомоаконитатом, служащим в качестве промежуточного продукта (например, под действием гомоаконитазы, такой как, например, AskD/E, LysT/U, Lys4, 3-изопропилмалатдегидратаза);
(3) окислительное декарбоксилирование (2R,3S) гомоизоцитрата до α-кетоадипата (например, под действием гомоизоцитратдегидрогеназы, такой как, например, AksF, Hicdh, Lys12, 2-оксосубератсинтаза, 3-изопропилмалатдегидрогеназа).
Получившийся α-кетоадипат (C6-цепь) затем подвергается двум последовательным сериям реакций удлинения цепи α-кетокислот, чтобы произвести α-кетопимелат в качестве промежуточного продукта и α-кетосуберат. α-Кетосуберат, получающийся из данного ряда реакций, затем подвергается неокислительному декарбоксилированию с образованием 7-оксогептановой кислоты, предшественника кофермента B.
Один хорошо описанный пример пути удлинения α-кетокислоты представляет собой путь биосинтеза кофермента B у Methanococcus jannaschii. В данном пути имеют место три фермента, катализирующие стадии a-l на фигуре 1. AksA (катализирующий стадии a, e и i) представляет собой гомоцитратсинтазу; AksD/E (катализирующий стадии b, c, f, g, j, k) является гетеротетрамером AksD и AksE и представляет собой гомоаконитазу; AksF (катализирующий стадии d, h и l) представляет собой гомоизоцитратдегидрогеназу. Данные ферменты, как было показано, все катализируют стадии реакции, и AksD/E M. jannaschii представляет собой единственную до настоящего времени гомоаконитазу, которая, как было показано, катализирует обе гидролазные реакции (Howell et al., Biochem., 1998; Howell et al., J. Bacteriol., 2000; Drevland et al., JBC, 2008). Следует отметить, что M. jannaschii является термофильной метанопродуцирующей бактерией, и все ферменты Aks были охарактеризованы при 50-60°C. В некоторых вариантах осуществления можно использовать альтернативные гомологи Aks из других метанопродуцирующих бактерий, которые размножаются при 37°C. Такие гомологи Aks были идентифицированы и они включают гены Methanococcus maripaludis S2 aksA (MMP0153), MMP1480, MMP0381 и aksF (MMP0880).
Путь, начинающийся с α-кетоглутарата (C5), включает различные, следующие ниже промежуточные продукты в зависимости от цикла удлинения: α-кетоадипат (C6), α-кетопимелат (C7) и α-кетосуберат (C8). Следовательно, в некоторых аспектах по изобретению в зависимости от длины углеводородной цепи (например, C4, C5, C6, C7) требуемого продукта никакого удлинения (для получения дифункционального бутана) может не потребоваться, может потребоваться только один цикл удлинения (например, стадии с a до d для получения дифункционального пентана), два удлинения (например, стадии с a до h для получения дифункционального гексана) или три удлинения (например, стадии с a по l для получения дифункционального гептана). Следует поэтому принять во внимание, что в зависимости от длины углеводородной цепи может оказаться желательным максимально увеличить доступность либо промежуточного продукта 2-кетоадипата, либо промежуточного продукта кетопимелата. В некоторых вариантах осуществления желательно сохранить стадии от a до d и удалить стадии от e до l, чтобы максимально увеличить доступность промежуточного продукта 2-кетоадипата. В других вариантах осуществления может оказаться желательным сохранить стадии от a до h и удалить стадии от i до l, чтобы максимально увеличить доступность промежуточного продукта кетопимелата.
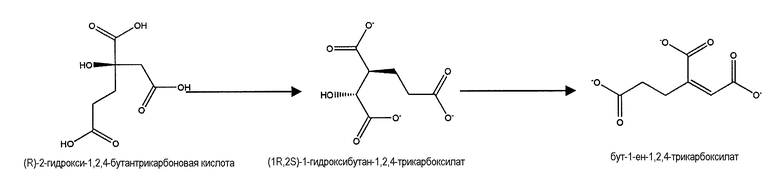
Как описано выше, каждый цикл удлинения содержит группу трех ферментов: ацилтрансферазу или гомолог ацилтрансферазы, гомоаконитазу или гомолог гомоаконитазы и гомоизоцитратдегидрогеназу или гомолог гомоизоцитратдегидрогеназы. Первая реакция каждой стадии удлинения катализируется ферментом ацетилтрансфераза, который при переносе преобразует ацильные группы в алкильные группы. В некоторых вариантах осуществления фермент ацилтрансфераза представляет собой гомоцитратсинтазу (EC 2.3.3.14). Ферменты гомоцитратсинтазы, которые катализируют химическую реакцию ацетил-CoA + H2O + 2-оксоглутарат ↔ (R)-2-гидроксибутан-1,2,4-трикарбоксилат + CoA.
Продукт (R)-2-гидроксибутан-1,2,4-трикарбоксилат также известен как гомоцитрат. Было показано, что некоторые гомоцитратсинтазы, такие как AksA, обладают широким субстратным диапазоном и катализируют конденсацию оксоадипата и оксопимелата с ацетил-CoA (Howell et al., 1998, Biochemistry, Vol. 37, pp. l0108-10117). В некоторых аспектах по изобретению предлагается гомоцитратсинтаза, обладающая субстратной специфичностью в отношении оксоглутарата или в отношении оксоглутарата и в отношении оксоадипата. Предпочтительная гомоцитратсинтаза известна под номером EC 2.3.3.14. В основном, процесс выбора подходящих ферментов может включать поиск ферментов среди естественного разнообразия при помощи поиска гомологов из других организмов и/или создание и поиск искусственной вариабельности и выбор вариантов с выбранной специфичностью и активностью фермента. Подходящие гомоцитратсинтазы, подходящие гомоаконитазы и подходящая гомоизоцитратдегидрогеназа перечислены в таблице 1.
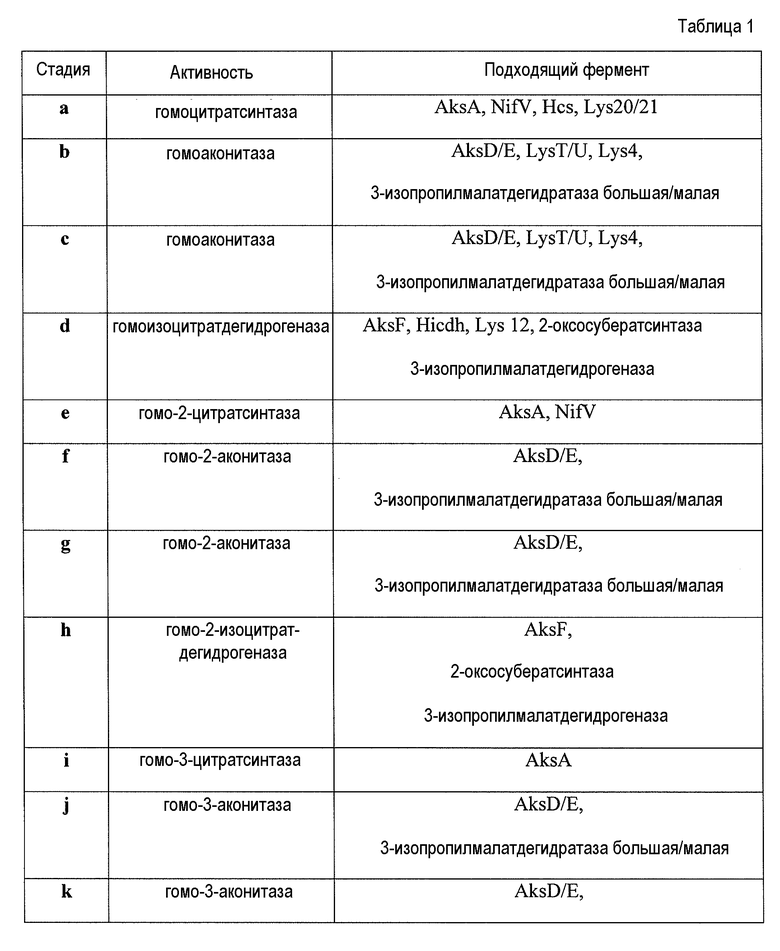
В некоторых вариантах осуществления первую стадию пути конструируют так, чтобы она катализировалась гомоцитратсинтазой NifV или гомологами NifV. Гомологи NifV обнаруживают у множества организмов, включая в качестве неограничивающих примеров Azotobacter vinelandii, Klebsiella pneumoniae, Azotobacter chroococcum, Frankia sp. (штамм FaC1), Anabaena sp. (штамм PCC 7120), Azospirillum brasilense, Clostridium pasteurianum, Rhodobacter sphaeroides, Rhodobacter capsulatus, Frankia alni, Carboxydothermus hydrogenoformans (штамм Z-2901/DSM 6008), Anabaena sp. (штамм PCC 7120), Frankia alni, Enterobacter agglomerans, Erwinia carotovora subsp. atroseptica (Pectobacterium atrosepticum), Chlorobium tepidum, Azoarcus sp. (штамм BH72), Magnetospirillum gryphiswaldense, Bradyrhizobium sp. (штамм ORS278), Bradyrhizobium sp. (штамм BTAi1/ATCC BAA-1182), Clostridium kluyveri (штамм ATCC 8527/DSM 555/NCIMB 10680), Clostridium kluyveri (штамм ATCC 8527/DSM 555/NCIMB 10680), Clostridium butyricum 5521, Cupriavidus taiwanensis (штамм R1/LMG 19424), Ralstonia taiwanensis (штамм LMG 19424), Clostridium botulinum (штамм Eklund 17B/тип B), Clostridium botulinum (штамм Alaska E43/тип E3), Synechococcus sp. (штамм JA-2-3B'a(2-13)) (йеллоустонская цианобактерия B-Prime), Synechococcus sp. (штамм JA-3-3Ab) (йеллоустонская цианобактерия A-Prime), Geobacter sulfurreducens и Zymomonas mobilis. NifV, как было показано, использует оксоглутарат (ферментативная стадия a) и оксоадипат (ферментативная стадия e) в качестве субстрата, но, как было продемонстрировано, не использует оксопимелат в качестве субстрата (см. Zheng et al., (1997) J. Bacteriol. Vol. 179, pp. 5963-5966). Таким образом, сконструированный путь 2-кетоудлинения, содержащий гомоцитратсинтазу NifV, исключает стадии от i до l в пути 2-кетоудлинения и максимально увеличивает доступность промежуточного продукта 2-кетопимелата.
В некоторых вариантах осуществления первая стадия пути, как конструируют, катализируется гомоцитратсинтазой Lys20 или Lys21. Lys20 и Lys21 представляют собой два изофермента гомоцитратсинтазы, вовлеченных в первую стадию пути биосинтеза лизина в дрожжах Saccharomyces cerevisiae. Гомологи Lys20 или Lys21 обнаруживают у множества организмов, таких как Pichia stipitis и Thermus thermophilus. Ферменты Lys20 и Lys21, как было показано, используют оксоглутарат в качестве субстрата, но не используют оксоадипат или оксопимелат. Таким образом, сконструированный путь 2-кетоудлинения, содержащий Lys20/21, исключает стадии от e до l в пути 2-кетоудлинения и максимально увеличивает доступность 2-оксоадипата. В некоторых вариантах осуществления ферменты, катализирующие реакцию с участием ацетилкофермента А и α-кетокислот в качестве субстратов, используют для превращения а-кетокислоты в гомоцитрат (например, EC 2.3.3.-). Methanogenic archaea содержит три близкородственных гомолога AksA: 2-изопропилмалатсинтазу (LeuA) и цитрамалат(2-метилмалат)синтазу (GimA), которые конденсируют ацетил-CoA с пируватом. Данный фермент, как полагают, принимает участие в биосинтезе изолейцина в метанопродуцирующих бактериях и, возможно, других видах, не имеющих треониндегидратазу. В некоторых вариантах осуществления фермент ацилтрансфераза является изопропилмалатсинтазой (например, LeuA, EC 2.3.3.13) или цитрамалатсинтазой (например, CimA, EC 2.3.1.182).
Вторая стадия пути кетоудлинения катализируется ферментом гомоаконитаза. Фермент гомоаконитаза катализирует реакцию гидратации и дегидратации:
гомоцитрат ↔ цис-гомоаконитат + H2O
В некоторых вариантах осуществления гомоаконитаза предствляет собой AksD/E, lysT/U или lys4 или их гомологи или варианты. Гомоаконитазы AksD/E и lysT/U, как было показано, состоят из двух полипептидов AksD и AksE, lysT и lysU, соответственно.
Последняя стадия каждого цикла кетоудлинения катализируется гомоизоцитратдегидрогеназой. Гомоизоцитратдегидрогеназа (например, EC 1.1.1.87) представляет собой фермент, который катализирует химическую реакцию:
(1R,2S)-1-гидроксибутан-1,2,4-трикарбоксилат + NAD+ ↔ 2-оксоадипат + CO2 + NADH + H+.
В некоторых вариантах осуществления гомоцитратдегидрогеназа включает в качестве неограничивающих примеров AksF, Hicdh, lys12 и LeuB (EC 1.1.1.85). LeuB представляет собой 3-изопропилмалатдегидрогеназу (EC 1.1.1.85) (IMDH) и катализирует третью стадию биосинтеза лейцина в бактериях и грибах, окислительное декарбоксилирование 3-изопропилмалата в 2-оксо-4-метилвалерат. Было показано, что 2-кетоизовалерат превращается в 2-кетоизокапроат на 3 стадии цикла удлинения при помощи LeuA (2-изопропилмалатсинтазы), LeuC, LeuD (комплекса 3-изопропилмалатизомеразы) и LeuB (3-изопропилмалатдегидрогеназы) в пути биосинтеза лейцина. Следует принимать во внимание, что данные ферменты обладают широкой субстратной специфичностью (см. Zhang et al., (2008), P.N.A.S) и могут катализировать реакции удлинения α-кетокислоты. В некоторых вариантах осуществления LeuA, LeuC, LeuD и LeuB катализируют удлинение α-кетоглутарата до α-кетоадипата и удлинение α-кетоадипата до α-кетопимелата.
II. Сконструированные пути для получения дифункциональных алканов (от C4 до C7) из α-кетокислот (от C5 до C8)
Существует несколько возможных путей получения дифункциональных алканов из источников α-кетокислот с использованием рекомбинантных микроорганизмов, как показано на фигуре 2. Аспекты по изобретению относятся к превращению с помощью микроорганизмов α-кето-промежуточных продуктов в молекулы дифункционального бутана, дифункционального пентана, дифункционального гексана, дифункционального гептана. Молекулы представляющего интерес дифункционального бутана включают в качестве неограничивающих примеров 1,4-бутандиол, 1-гидроксибутаноат, янтарную кислоту, 1,4-диаминобутан, 4-аминобутанал и 4-аминобутанол. Молекулы представляющего интерес дифункционального пентана включают в качестве неограничивающих примеров 1-гидроксипентаноат, 1,5-пентандиол, глутарат, кадаверин (пентан-1,5-диамин), 5-аминопентанал и 5-аминопентанол. Молекулы представляющего интерес дифункционального гексана включают в качестве неограничивающих примеров 1-гидроксигексаноат, 1,6-гександиол, адипат, гексаметилендиамин, аминокапроновую кислоту, 6-аминогексанал и 6-аминогексанол. Молекулы представляющего интерес дифункционального гептана включают в качестве неограничивающих примеров 1-гидроксигептаноат, 1,7-гептандиол, пимелиновую кислоту, 1,7-диаминогептан, 7-аминогептанал и 7-аминогептанол.
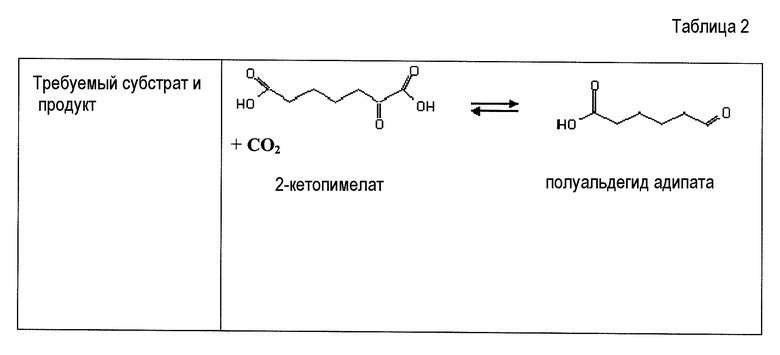
В некоторых вариантах осуществления первый возможный путь получения дифункциональных алканов включает, прежде всего, декарбоксилирование α-кетокислоты (ферментативная стадия I). В некоторых аспектах по настоящему изобретению описан способ удаления карбоксильной группы без окислительного декарбоксилирования. Такой способ осуществляют, экспрессируя в клетке-хозяине белок, обладающий биологической активностью, по существу схожей с декарбоксилазой α-кетокислоты, для того чтобы получить полуальдегид карбоновой кислоты. Термин «декарбоксилаза α-кетокислоты» (KDC) относится к ферменту, который превращает α-кетокислоты в полуальдегид карбоновой кислоты и диоксид углерода. Некоторые особенно представляющие интерес KDC известны под следующими ниже номерами: EC 4.1.1.1; EC 4.1.1.80, EC 4.1.1.72, 4.1.1.71, 4.1.1.7, 4.1.1.75, 4.1.1.82, 4.1.1.74 (см. ниже таблицу 2). Некоторые из KDC обладают широким субстратным диапазоном, тогда как другие KDC являются более субстратспецифичными. KDC доступны из большого количества источников, включая в качестве неограничивающих примеров S. cerevisiae и бактерии. В некоторых иллюстративных вариантах осуществления используемые KDC включают в качестве неограничивающих примеров KivD из Lactococcus lactis (UniProt Q684J7), ARO10 (UniProt Q06408) из S. cerevisiae, PDC1 (UniProt P06169), PDC5 (UniProt P16467), PDC6 (UniProt P26263), Thi3 из S. cerevisiae, kgd из M. tuberculosis (UniProt 50463), mdlc из P. putida (UniProt P20906), arul из P. aeruginosa (UniProt AAG08362), fom2 из S. wedmorensis (UniProt Q56190), Pdc из Clostridium acetobutyculum, ipdC из E. coacae (UniProt P23234) или любые гомологичные белки из тех же самых или других видов микроорганизмов. В некоторых вариантах осуществления декарбоксилаза кетокислоты представляет собой пируватдекарбоксилазу, известную под номером по классификации EC EC 4.1.1.1. Пируватдекарбоксилаза представляет собой ферменты, которые катализируют декарбоксилирование пировиноградной кислоты до ацетальдегида и диоксида углерода. Пируватдекарбоксилазы доступны из большого количества источников, включая в качестве неограничивающих примеров S. cerevisiae и бактерии (см. патент США 20080009609, который включен в настоящий документ в качестве ссылки). В некоторых вариантах осуществления декарбоксилаза α-кетокислоты представляет собой декарбоксилазу α-кетоизовалерата KivD, которая в природе катализирует превращение α-кетоизовалерата в изомасляный альдегид и диоксид углерода. В некоторых вариантах осуществления декарбоксилаза α-кетокислоты представляет собой декарбоксилазы α-кетокислот с разветвленными цепями (номер EC 4.1.1.72). Следует принимать во внимание, что поскольку некоторые KDC, как было показано, обладают широким субстратным диапазоном, субстратная специфичность может являться важным фактором для рассмотрения при выборе источников генов. Следовательно, в некоторых вариантах осуществления ферменты пируватдекарбоксилазы конструируют для того, чтобы проявить преимущество α-кетосуберата, α-кетопимелата, α-кетоадипата или α-кетоглутарата над пируватом. Предпочтительно сконструированный фермент проявляет, по меньшей мере, в 2 раза, по меньшей мере, в 5 раз, по меньшей мере, в 10 раз, по меньшей мере, в 20 раз, по меньшей мере, в 50 раз, по меньшей мере, в 100 раз увеличенное предпочтение по сравнению с пируватом.
В предпочтительных вариантах осуществления KDC, экспрессирующаяся в рекомбинантных клетках-хозяевах, катализирует превращение α-кетосуберата в полуальдегид пимелиновой кислоты или превращение α-кетопимелата в полуальдегид адипиновой кислоты, или превращение α-кетоадипата в полуальдегид глутаровой кислоты, или превращение α-кетоглутарата в полуальдегид янтарной кислоты.
В некоторых вариантах осуществления полуальдегид карбоновой кислоты (например, полуальдегид янтарной кислоты, полуальдегид глутаровой кислоты, полуальдегид адипиновой кислоты и/или полуальдегид пимелиновой кислоты) превращается в гидроксикарбоновую кислоту (гидроксилбутаноат, гидроксипентаноат, гидроксигексаноат, гидроксигептаноат) при помощи алкогольдегидрогеназы, которая преобразует альдегидную функциональную группу в спиртовую функциональную группу (ферментативная стадия 1, фиг. 2). Алкогольдегидрогеназы (ADH) (EC 1.1.1.1 и EC 1.1.1.2) катализируют обратимое восстановление кетонов и альдегидов до спиртов с восстановлением NAD+ до NADH. В некоторых вариантах осуществления алкогольдегидрогеназа включает в качестве неограничивающих примеров adhA или adhB (из Z. mobilis), бутанолдегидрогеназу (из Clostridium acetobutylicum), пропандиолоксидоредуктазу (из E. coli) и ADHIV алкогольдегидрогеназу (из Saccharomyces) или ADH6 (из S. cerevisiae).
В некоторых вариантах осуществления гидрокарбоновую кислоту подвергают дегидрированию с использованием алкогольдегидрогеназы или альдегиддегидрогеназы (ферментативная стадия 2, фиг. 2 и фиг. 3), чтобы получить алкандиол, такой как 1,4-бутандиол, 1,5-пентандиол, 1,6-гександиол и/или 1,7-гептандиол.
Альдегид-NAD(+)-дегидрогеназная активность и алкоголь-NAD(+)-дегидрогеназные активности могут осуществляться при помощи двух различных полипептидов или осуществляться посредством одного полипептида, такого как многофункциональная альдегид-алкогольдегидрогеназа (EC 1.2.1.10) из E. coli (Goodlove et al. Gene 85:209-14, 1989; инвентарный номер в GenBank M33504). Полипептиды, обладающие (NAD(P)+)-альдегиддегидрогеназной (EC 1.2.1.-) или (NAD(+))-альдегиддегидрогеназной (EC 1.2.1.3) активностью, можно использовать в сочетании с алкогольдегидрогеназой, чтобы восстановить остающуюся карбоновую кислоту до спирта, получая на выходе алкандиол. Нуклеиновую кислоту, кодирующую такие полипептиды, можно получать из различных видов, включая без ограничения S. cerevisiae.
В других вариантах осуществления полуальдегид карбоновой кислоты (например, полуальдегид янтарной кислоты, полуальдегид глутаровой кислоты, полуальдегид адипиновой кислоты, полуальдегид пимелиновой кислоты) превращается в дикарбоновую кислоту (ферментативная стадия 3, фиг. 2 и фиг. 3), такую как адипиновая кислота, янтарная кислота, глутаровая кислота и/или пимелиновая кислота, посредством использования альдегиддегидрогеназы.
В одном аспекте изобретения стадия первоначального декарбоксилирования α-кетокислоты сопровождается ферментативной стадией с использованием 1-аминотрансферазы (ферментативная стадия 4, фиг. 2 и фиг.3) для получения аминокарбоновой кислоты. В частности, представляющий интерес продукт можно синтезировать, используя данный сконструированный путь, включающий аминокапроновую кислоту, аминобутановую кислоту, аминогептановую кислоту и/или аминопентановую кислоту. В некоторых вариантах осуществления ферментативная стадия 4 включает аминотрансферазу полуальдегида карбоновой кислоты. Ферменты, обладающие активностью аминотрансферазы полуальдегида карбоновой кислоты, доступны из большого количества источников, включающих в качестве неограничивающих примеров B. subtilis, S. cerevisiae. H. sapiens, F. lutescens, S. clavuligerus. Иллюстративные примеры, способные катализировать аминотрансферазные реакции, представляют собой трансаминазу орнитиноксокислоты (rodD из B. subtilis, OAT из H. sapiens, EC 2.6.1.13), орнитинтрансаминазу аргинин-деструктивного фермента (EC 2.6.1.13), лизинаминотрансферазу (EC 2.6.1.36) из F. lutescens или S. clavuligerus, 4-аминобутиратаминотрансферазу Mus musculus (EC 2.6.1.19), 4-аминобутиратаминотрансферазу Sus scrofa (EC 2.6.1.19), 4-аминобутиратаминотрансферазу S. cerevisiae (EC 2.6.1.19), сахаропиндегидрогеназу LYS9 и LYSl S. cerevisiae (EC 1.5.1.10 и 1.5.1.7, соответственно) или любые гомологичные белки из тех же самых или других видов микроорганизмов.
Два возможных пути для получения представляющих интерес диаминоалканов или аминоспирта включают превращение аминокарбоновой кислоты в аминоальдегид с использованием альдегиддегидрогеназы (ферментативная стадия 5, фиг.2 и фиг.3), после чего следует ферментативная стадия 6 или ферментативная стадия 7. В другом варианте осуществления 2-аминодикарбоновая кислота может подвергаться действию 2-аминодекарбоксилазы (ферментативная стадия 8) с получением аминокарбоновой кислоты. В некоторых вариантах осуществления ферментативная стадия 8 сопровождается стадией дегидрирования, катализируемой и альдегиддегидрогеназой (ферментативная стадия 5). Полученный в результате аминоальдегидный метаболит можно использовать в качестве субстрата для двух различных ферментативных стадий. Ферментативная стадия 6 включает 1-аминотрансферазу и катализирует превращение аминоальдегида в диаминоалкан. Альтернативно, ферментативная стадия 7, включающая алкогольдегидрогеназу, катализирует превращение аминоальдегида в аминоспирт. ферментативная стадия 6 включает фермент аминотрансферазу и позволяет получить диаминоалканы, такие как гексаметилендиамин (HDD), кадаверин (или пентаметилендиамин), диаминобутан и/или димаминогептан. Ферментативная стадия 7 включает алкогольдегидрогеназу и позволяет получить аминобутанол, аминопентанол, аминогексанол (или 6-гидроксигексамин 6HH) и аминогептанол.
Второй возможный путь включает, прежде всего, 2-аминотрансферазу (ферментативная стадия II, фиг. 2 и фиг. 3) с образованием 2-аминодикарбоновой кислоты. Предпочтительные возможные представляющие интерес гены, которые экспрессируют белки с аминотрансферазной активностью, включают в качестве неограничивающих примеров lysN (кодирующий α-аминоадипатаминотрансферазу из T. thermophilus EC 2.6.1.7) и гомологи (например kat2); aadat (кодирующий аминоадипатаминотрансферазу в R. norvegicus, EC 2.6.1.39), AADAT (кодирующий аминоадипатаминотрансферазу у H. sapiens). Альтернативные ферменты, обладающие аминотрансферазной активностью, включают в качестве неограничивающих примеров ddh (из Corynebacterium glutanicum, кодирующий мезо-диаминопимелат-D-дегидрогеназу, EC 1.4.1.16) и dapdh (из Lysinibacillus sphaericus, кодирующий мезо-диаминопимелатдегидрогеназу, EC 1.4.1.16). Возможные гены, экспрессирующие белки с аминопимелатдекарбоксилазной активностью, включают в качестве неограничивающих примеров глутаматдекарбоксилазы (EC 4.1.1.15), например, gadA/B, кодирующий изоформу A или изоформу B в E. coli, GAD1 (из S. cerevisiae), GAD1/2 (из A. thaliana), GAD1/2 (из H. sapiens) и их гомологи, или диаминопимелатдекарбоксилазу, такую как, например, LysA (EC 1.1.1.20, из E. Coli или Bacillus subtilis) или AT5G11880 (из A. thaliana), или AT3G14390 (из A. thaliana).
III. Сконструированный путь для получения C6-дифункциональных алканов из α-кетопимелата
Аспекты по изобретению относятся к сконструированным путям для получения представляющих интерес C6-дифункциональных алканов. В частности, аспекты по изобретению относятся к получению адипиновой кислоты, аминокапроновой кислоты (устойчивому предшественнику кислоты капролактама), гексаметилендиамина и 6-гидроксигексаноата (фиг. 3). Следует принимать во внимание, что для получения с помощью микроорганизмов C6-дифункциональных алканов необходимо удалить стадии с i до l, для того чтобы максимально увеличить доступность 2-кетопимелата для превращения с помощью микроорганизмов в полуальдегид адипата (стадия m). В некоторых вариантах осуществления AksA заменяют на NifV, фермент из A.vinelandii, который, как было показано, действует и на 2-кетоглутарат, и на 2-кетоадипат, но не на 2-кетопимелат (Howell et al., Biochem., 1998; Howell et al., J. Bacteriol., 2000; Drevland et al., JBC, 2008). Такая замена позволила бы устранить стадии с i до l в пути удлинения 2-кетокислоты. Следует принимать во внимание, что в зависимости от длины углеводородной цепи необходимого продукта может потребоваться только один цикл удлинения (например, стадии a-d). В некоторых иллюстративных вариантах осуществления глутаровую кислоту, общеизвестный пластификатор и предшественник сложных полиэфиров с формулой HO2C(CH2)3CO2H, (также называемая пентандиовая кислота), можно получить с помощью микроорганизмов из 2-кетоглутарата, который подвергали одному циклу удлинения, чтобы получить кетоадипат. Глутаровую кислоту применяют для производства полимеров, таких как полиэфирполиолы, и полиамидов. В некоторых вариантах осуществления, чтобы осуществить один цикл удлинения, ферменты Hcs и Lys20/21 (из путей биосинтеза лизина в S. cerevisiae и T. thermophilus) можно предоставить возможность для удаления стадий e-l, для того чтобы максимально увеличить жизнеспособность 2-кетоадипата для получения с помощью микроорганизмов глутаровой кислоты.
A. Сконструированные пути для получения адипиновой кислоты
1. Общие сведения об адипиновой кислоте:
В 2005 году мировой спрос на адипиновую кислоту составлял 2,7 миллиона метрических тонн. Исторически спрос на адипиновую кислоту вырастает на 2% ежегодно, и до 2009 года ожидается увеличение на 2-3%. Адипиновая кислота раз за разом занимает одно из ведущих пяти мест среди химических веществ, производимых в США. Почти 90% получаемой внутри страны адипиновой кислоты используется для производства нейлона-6,6. Другие применения адипиновой кислоты включают производство смазочных веществ, смол, полиэфирполиолов и пластификаторов, и ее используют в качестве пищевого подкисляющего средства.
Существует три основных способа коммерческого производства: циклогексановый способ, циклогексаноловый способ, способ бутадиенкарбонилированием. В основном промышленном способе синтеза адипиновой кислоты применяется первоначальное окисление кислородом воздуха циклогексана, чтобы получить на выходе смесь циклогексанона (кетона) и циклогексанола (спирта), которую обозначают KA (см., например, патент США № 5221800). Также для получения KA коммерчески используют гидрирование фенола, хотя на данный способ приходится только 2% от всего производства адипиновой кислоты. KA, производимую посредством двух способов, окисляют с помощью азотной кислоты, чтобы получить адипиновую кислоту. Восстановленные оксиды азота, включающие NO2, NO и N2O, получают в качестве побочных продуктов и перерабатывают их обратно до азотной кислоты на различных уровнях. Становится все более и более интересным для промышленности и благоприятным для окружающей среды конструировать несинтетические, биологические пути доступа к адипиновой кислоте. Было описано большое количество микробиологических способов. Организмы дикого типа и мутантные организмы, как было показано, превращают возобновляемое сырье, такое как глюкоза и другие углеводороды, в адипиновую кислоту (см., например, WO9507996 и патенты США №№ 5272073, 5487987 и 5616496). Схожим образом, организмы, обладающие нитрилазной активностью, как было показано, превращают нитрилы в карбоновые кислоты, включая адипиновую кислоту (см., например, патент США № 5629190). Дополнительно, организмы дикого типа были использованы для превращения циклогексана и циклогексанола и других спиртов в адипиновую кислоту (см., например, патент США № 6794165; и патентные заявки США №№ 2003087403 и 20020127666). Например, в одном ферментативном пути циклогексанол преобразуется в адипиновую кислоту, причем ферментативный путь включает гены, выделенные из Acinetobacter, кодирующие гидроксилацил-CoA-дегидрогеназу; еноил-CoA-гидратазу, ацил-CoA-дегидрогеназу, убихиноноксидоредуктазу, монооксигеназу, альдегиддегидрогеназу. В качестве другого ферментативного пути для превращения циклогексанола в адипиновую кислоту предложили путь, включающий промежуточные продукты циклогексанол, циклогексанон, 2-гидроксициклогексанон, ε-капролактон, 6-гидроксикапроновую кислоту. Некоторые определенные ферментативные активности были продемонстрированы в данном пути, включая циклогексанолдегидрогеназу, связанную с NADPH циклогексаноноксигеназу, ε-капролактонгидролазу и связанную с NAD(NADP) дегидрогеназу 6-гидроксикапроновой кислоты (Tanaka et al., Hakko Kogaku Kaishi (1977), 55(2), 62-7). Альтернативный ферментативный путь, как требуются, включает циклогексанол, циклогексанон, 1-окса-2-оксоциклогептан, 6-гидроксигексаноат, 6-оксогексаноат и адипат (Donoghue et al., Eur. J. Biochem., 1975, 60(1), 1-7).
Следовательно, проблема, которую необходимо решить, заключается в том, чтобы предоставить способ синтеза адипиновой кислоты, который не только позволяет избежать зависимости от относительно экологически чувствительных исходных веществ, таких как нефть, но также и делает эффективным использование не являющихся нефтехимическими недорогих возобновляемых ресурсов. Далее было бы желательно предоставить способ синтеза адипиновой кислоты, который позволяет избежать необходимости в существенном потреблении энергии и который сводит до минимума образование токсичных побочных продуктов.
2. Получение с помощью микроорганизмов адипиновой кислоты по α-кетопимелатному пути
Некоторые аспекты по изобретению относятся к декарбоксилированию 2-кетопимелата до полуальдегида адипата (или 6-оксогексаноат) и дегидрированию полуальдегида адипата с получением адипиновой кислоты. В некоторых вариантах осуществления все опубликованные в MetaCyc реакции декарбоксилазы проверяли в отношении возможных декарбоксилаз 2-кетопимелата. Перечень ферментов и активностей получили на основе критериев (i) обнаруженной активности в отношении 2-кетокарбоксилата и (ii) доступности информации о последовательности белка (см. таблицу 2). В некоторых вариантах осуществления ферменты, перечисленные в таблице 2, проверяют в отношении декарбоксилазной активности для всех 2-кетокислот в предлагаемых сконструированных путях (т.е. для 2-кетопимелата, 2-кетоадипата и 2-кетоглутарата).
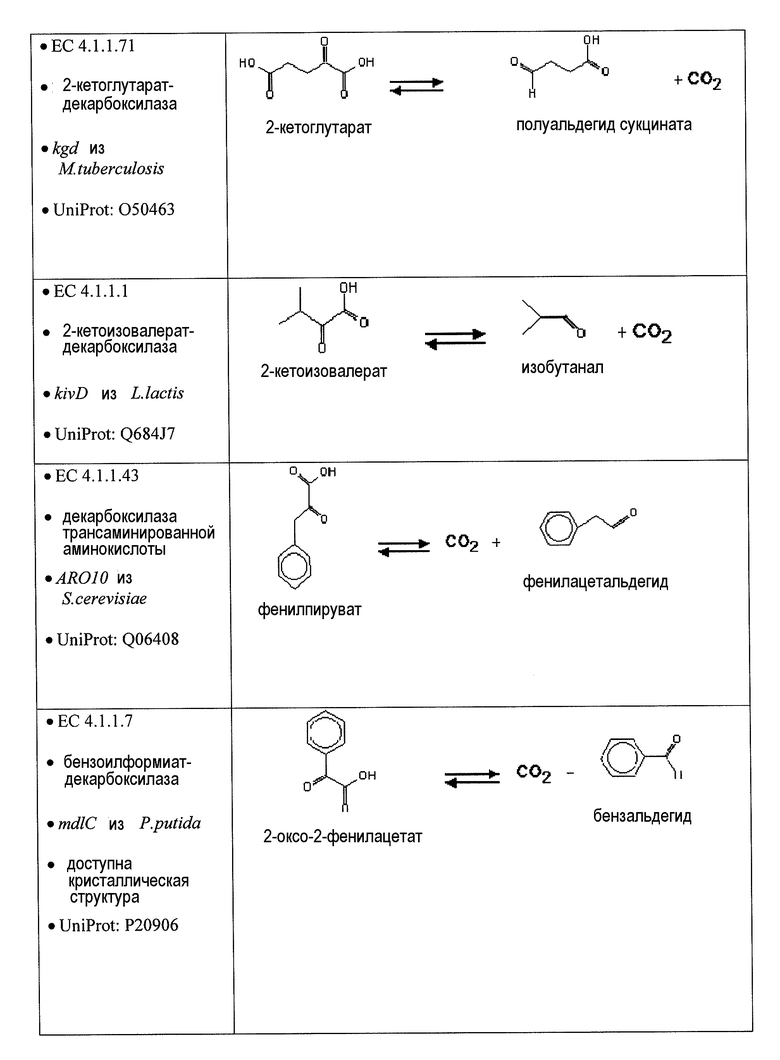

В предпочтительном варианте осуществления превращение полуальдегида адипата в адипат (стадия n) катализируют с использованием фермента ChnE или гомолога фермента ChnE. ChNE представляет собой фермент-связанную с NADP+ 6-оксогексаноатдегидрогеназу и, как было показано, он катализирует дегидрирование 6-оксогексаноата до адипата в пути деградации циклогексанола у Acinetobacter sp. (см. Iwaki et al., Appl. Environ. Microbiol. 1999, 65(11): 5158-5162). В другом варианте осуществления α-кетоглутаровая полуальдегиддегидрогеназа (EC 1.2.1.26, например, AraE) преобразует полуальдегид адипата в адипат.
B. Сконструированные пути для получения капролактама
1. Общие сведения о капролактаме
Капролактам, прежде всего, применяется при изготовлении синтетических волокон, особенно нейлона 6, который также применяется в щетинных кистях, текстильных жестких прокладках, пленочных покрытиях, синтетической коже, пластмассах, пластификаторах, носителях, перекрестных сшивках для полиуретанов и в синтезе лизина. Приблизительно 2,5 миллиарда тонн нейлона 6 производится ежегодно во всем мире. Производство нейлона достигается полимеризацией с раскрытием цикла мономера ε-капролактама. Исходным химическим соединением для получения ε-капролактама является бензол, который превращают либо в циклогексан, либо в фенол и каждое из двух химических веществ превращают через циклогексанон в циклогексаноноксим, и затем данный промежуточный продукт нагревают в серной кислоте. Следовательно, проблема, которую необходимо решить, состоит в том, чтобы предоставить способ синтеза капроновой кислоты, который не только позволяет избежать зависимости от относительно экологически чувствительных исходных веществ, таких как нефть, но также и делает эффективным применение не являющихся нефтехимическими недорогих возобновляемых ресурсов. Далее было бы желательно предоставить способ синтеза капроновой кислоты, который позволяет избежать необходимости в значительном потреблении энергии и который сводит до минимума образование токсичных побочных продуктов.
2. Сконструированные пути для получения капролактама
Аспекты по изобретению относятся к двум сконструированным путям для получения с помощью микроорганизмов аминокапроновой кислоты из α-кетопимелата. Первый возможный путь включает, прежде всего, кетопимелатаминотрансферазу (ферментативная стадия II, фиг. 2 и фиг. 3), после чего следует аминопимелатдекарбоксилаза (ферментативная стадия 8). Возможные представляющие интерес гены, которые экспрессируют белки с кетопимелатаминотрансферазной активностью, включают в качестве неограничивающих примеров lysN (кодирующий α-аминоадипатаминотрансферазу из T. thermophilus EC 2.6.1.7) и гомологи (например, kat2); aadat (кодирующий аминоадипатаминотрансферазу у R. norvegicus) и AADAT (кодирующий аминоадипатаминотрансферазу у H. sapiens). Перечень возможных ферментов и активностей создавали на основе критериев (i) продемонстрированного или возможного превращения 2-оксопимелата в 2-аминопимелиновую кислоту в присутствии глутарата и (ii) доступности информации о последовательности белка (см. таблицу 3).
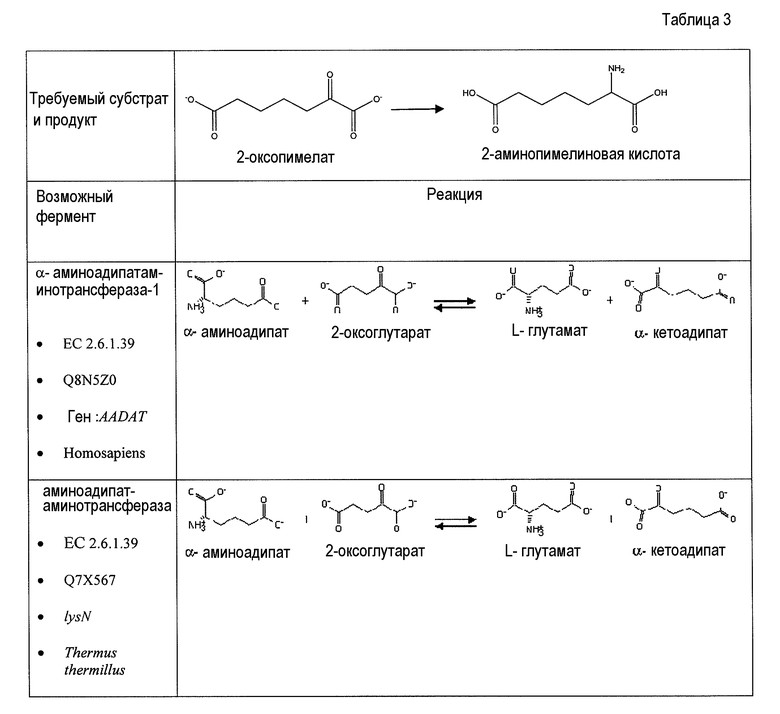
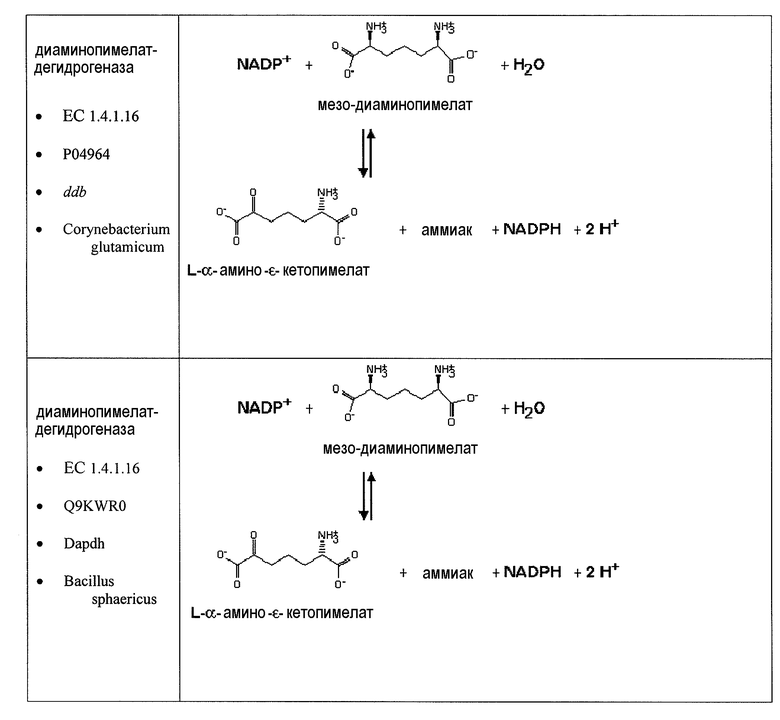
Возможные гены, экспрессирующие белки с аминопимелатдекарбоксилазной активностью, включают в качестве неограничивающих примеров глутаматдекарбоксилазы (gadA/B, кодирующий изоформу A или изоформу B в E. coli, GAD1 в S. cerevisiae, GAD1/2 в A. thaliana, GAD1/2 у H. sapiens) и их гомологи. Перечень возможных ферментов и активностей создавали на основе критериев (i) продемонстрированного или возможного превращения 2-аминопимелиновой кислоты в 6-аминогексановую кислоту в присутствии кетоглутарата и (ii) доступности информации о последовательности белка (см. таблицу 4). 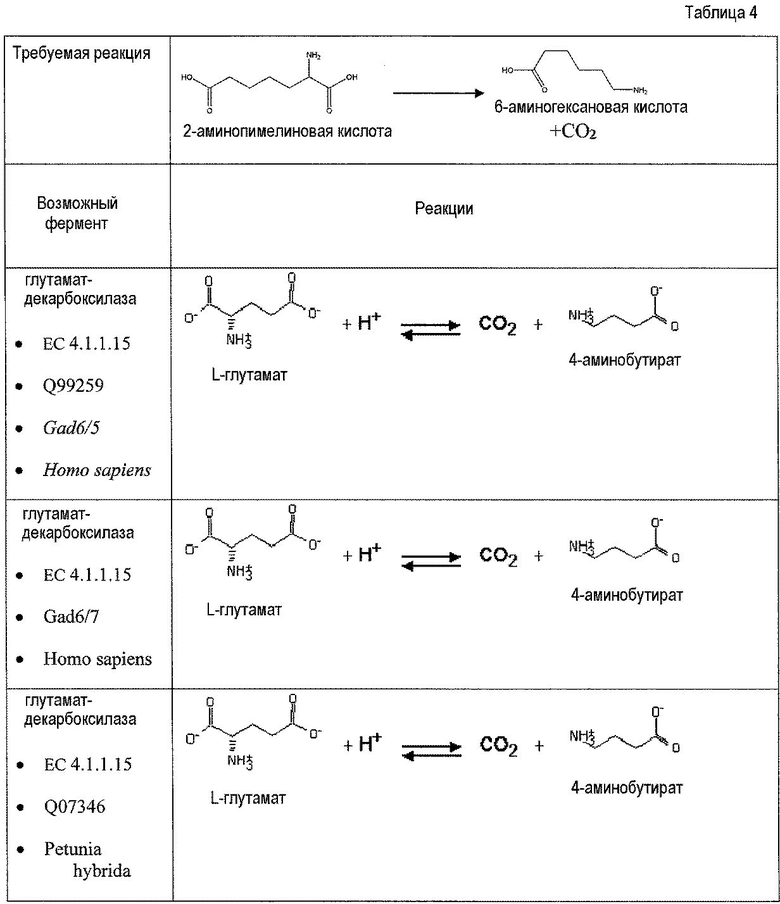
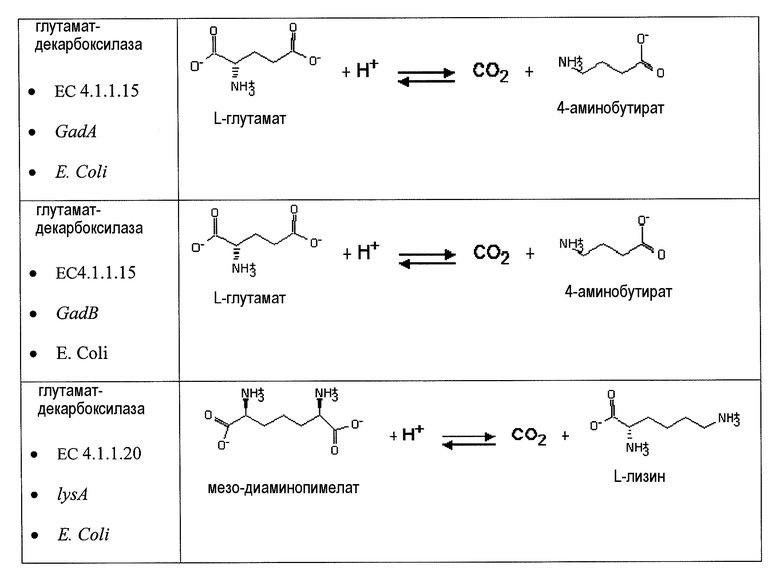
Второй возможный путь включает, прежде всего, кетопимелатдекарбоксилазу (ферментативная стадия I), после которой следует аминотрансфераза полуальдегида адипата (ферментативная стадия 4).
В некоторых вариантах осуществления фермент, который катализирует декарбоксилирование, представляет собой фермент KivD (как описано выше) или гомолог фермента KivD. В некоторых вариантах осуществления аминотрансфераза полуальдегида адипата, используемая во втором сконструированном пути, включает в качестве неограничивающих примеров трансаминазу орнитиноксокислоты (rodD из B. subtilis, OAT из H. sapiens, EC 2.6.1.13), орнитинтрансаминазу аргинин-деструктивного фермента (EC 2.6.1.13), лизинаминотрансферазу (EC 2.6.1.36) из F. lutescens или S. clavuligerus, 4-аминобутиратаминотрансферазу Mus musculus (EC 2.6.1.19), 4-аминобутиратаминотрансферазу Sus scrofa (EC 2.6.1.19), 4-аминобутиратаминотрансферазу S. cerevisiae (EC 2.6.1.19), сахаропиндегидрогеназу LYS9 и LYS1 S. cerevisiae (EC 1.5.1.10 и 1.5.1.7, соответственно) или любые гомологичные белки из тех же самых или других видов микроорганизмов.
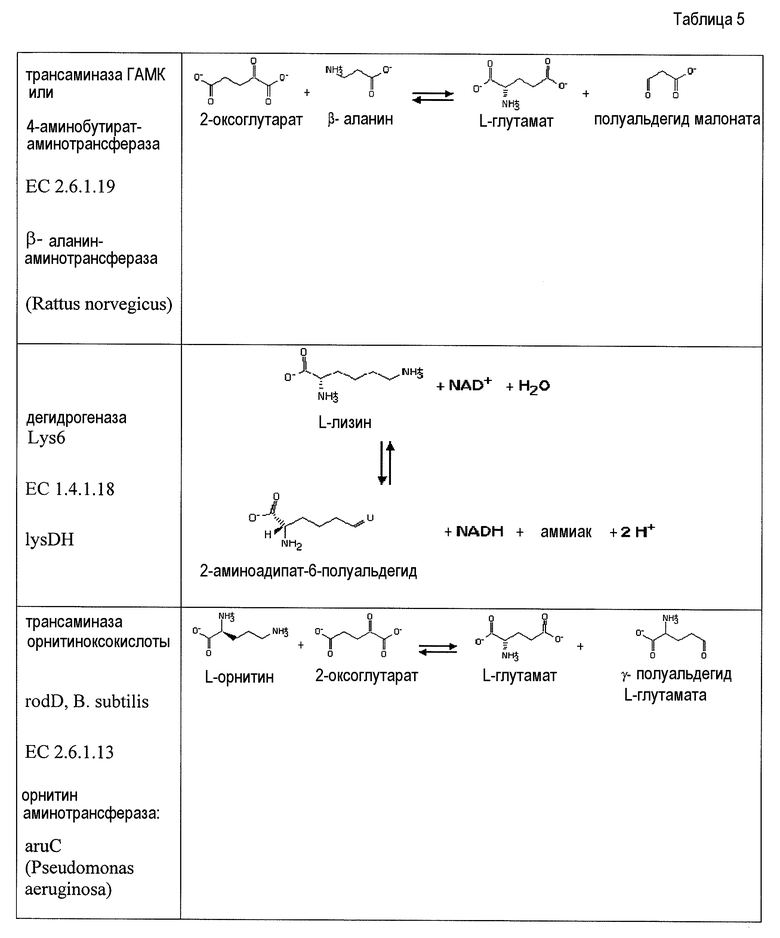
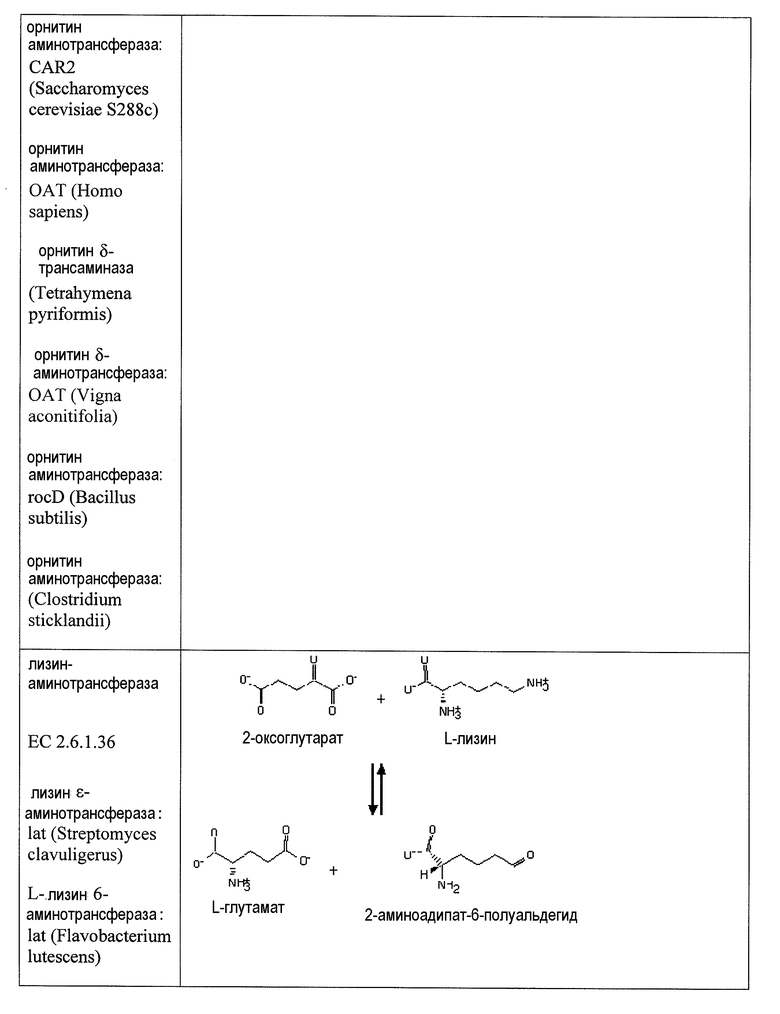
C. Сконструированные пути для получения гексаметилендиамина (HMD)
Аспекты по изобретению относятся к сконструированным путям для получения с помощью микроорганизмов гексаметилендиамина из α-кетопимелата. Гексаметилендиамин, главным образом, применяют для производства нейлона 6,6, нейлона 6,10, нейлона 6,66. Нейлон 6,6 и нейлон 6,10 можно изготовить в виде различных типов нейлоновых смол и нейлоновых волокон.
Первый сконструированные путь, как показано на фиг. 2 и 3, включает, прежде всего, декарбоксилирование 2-кетопимелата до аминокапроновой кислоты, как описано выше в аминокапроновом сконструированном пути (ферментативные стадии I, после чего следует ферментативная стадия 4 или ферментативная стадия II, после чего следует ферментативная стадия 8). В некоторых вариантах осуществления фермент альдегиддегидрогеназа экспрессируется и катализирует превращение аминокапроновой кислоты в промежуточный продукт 6-аминогексанал (ферментативная стадия 5), и фермент 1-аминотрансфераза экспрессируется и катализирует превращение 6-аминогексанала в HMD (ферментативная стадия 6). Альтернативно сконструированный путь включает стадию фосфорилирования (с использованием киназы) перед дегидрированием и ферментативные реакции аминопереноса, описанные выше. Следует принимать во внимание, что стадия фосфорилирования и стадии дегидрирования аналогичны стадии фосфорилирования (катализируемой аспартокиназой EC 2.7.2.4) и стадии дегидрирования (катализируемой ферментом EC 1.2.1.11), вовлеченных в синтез полуальдегида аспартата в лизиновом пути. Согласно второму сконструированному пути превращение 2-аминопимелата, получаемого из α-кетопимелата (ферментативная стадия II, фиг. 3), в гексаметилендиамин состоит из ферментативных стадий 9, 10 или 11 и 12, которые объединяют ферменты или гомологичные ферменты, охарактеризованные в пути биосинтеза лизин IV. Путь включает следующие ниже превращения субстратов в продукты:
• ферментативная стадия 9: 2-аминопимелат в 2-амино-7-оксогептаноат (или 2-аминопимелат-7-полуальдегид), которая катализируется, например, аминоадипатредуктазой или гомологичным ферментом (например, Sc-Lys2, EC 1.2.1.31);
• ферментативная стадия 10 или 11: амино-7-оксогептаноат в 2,7-диаминогептаноат, которая катализируется, например, сахаропиндегидрогеназой (например, Sc-Lys9, EC 1.5.1.10 или Sc-LyS1, EC 1.5.1.7);
• ферментативная стадия 12: 1,7-диаминогептаноат в гексаметилендиамин, которая катализируется, например, лизиндекарбоксилазой или орнитиндекарбоксилазой.
Следует принимать во внимание, что начиная с α-кетоадипата, схожий путь приведет к получению кадаверина.
D. Сконструированный путь для получения 6-гидроксигексаноата (6HH)
Один аспект по изобретению относится к сконструированному пути для получения с помощью микроорганизмов 6-гидроксигексаноата (6HH) из полуальдегида адипата, промежуточного продукта в сконструированном пути для адипиновой кислоты, описанном выше. 6HH представляет собой 6-углеродный гидроксиалканоат, который можно сформировать в капролактон или непосредственно полимеризовать с получением пластмасс на основе сложных полиэфиров (полигидроксиалканоата PHA). В некоторых вариантах осуществления полуальдегид адипата превращают в 6HH простым гидрированием и реакцию катализируют алкогольдегидрогеназой (EC 1.1.1.1). Данный фермент принадлежит семейству оксидоредуктаз, которые специфически действуют на группу CH-OH донора с использованием NAD+ или NADP+ в качестве акцептора. В некоторых вариантах осуществления используют 6-гидроксигексаноатдегидрогеназу (EC 1.1.1.258), которая катализирует следующую ниже химическую реакцию.
6-гидроксигексаноат + NAD+ ↔ 6-оксогексаноат + NADH + H+
Другие алкогольдегидрогеназы включают в качестве неограничивающих примеров adhA или adhB (из Z. mobilis), бутанолдегидрогеназу (из Clostridium acetobutylicum), пропандиолоксидоредуктазу (из E. coli) и ADHIV-алкогольдегидрогеназу (из Saccharomyces).
E. Сконструированный путь для получения 1,6-гександиола
Один аспект по изобретению относится к сконструированному пути для получения с помощью микроорганизмов 1,6-гександиола из полуальдегида адипата, промежуточного продукта в сконструированном пути для адипиновой кислоты, описанном выше. 1,6-Гександиол представляет собой ценный промежуточный продукт в химической промышленности. Он применяется в целом ряде синтезов полимеров, таких как получение сложных полиэфиров для полиуретановых эластомеров и полимерных пластификаторов, и его также используют для улучшения качества бензина.
В некоторых вариантах осуществления полуальдегид адипата превращают в 6-гидроксигексаноат при помощи алкогольдегидрогеназы (ферментативная стадия 1, фиг. 2) и затем в 1,6-гександиол под действием алкогольдегидрогеназы или альдегиддегидрогеназы (ферментативная стадия 2, фиг. 2).
Алкогольдегидрогеназы (ADH) (EC 1.1.1.1 и 1.1.1.2) катализируют обратимое восстановление кетонов и альдегидов до спиртов с восстановлением NAD+ до NADH. В некоторых вариантах осуществления алкогольдегидрогеназа включает в качестве неограничивающих примеров adhA или adhB (из Z. mobilis), бутанолдегидрогеназу (из Clostridium acetobutylicum), пропандиолоксидоредуктазу (из E. coli) и ADHIV-алкогольдегидрогеназу (из Saccharomyces), ADH6 (из S. cerevisiae).
Альдегид-NAD(+)-дегидрогеназная активность и алкоголь-NAD(+)-дегидрогеназные активности можно осуществлять при помощи двух различных полипептидов или осуществлять посредством одного полипептида, такого как многофункциональная альдегидалкогольдегидрогеназа (EC 1.2.1.10) из E. coli (Goodlove et al. Gene 85:209-14, 1989; инвентарный номер в GenBank M33504). Полипептиды, обладающие (NAD(P)+)-альдегиддегидрогеназной (EC 1.2.1.-) или (NAD(+))-альдегиддегидрогеназной (EC 1.2.1.3) активностью, а также нуклеиновую кислоту, кодирующую такие полипептиды, можно получать из различных видов, включая без ограничения S. cerevisiae.
F. Сконструированный путь для получения 6-аминогексанола
В некоторых вариантах осуществления Cn-омега-аминоспирт можно получить из Cn-концевой аминокислоты, где количество атомов углерода находится в диапазоне от 4 до 7, сочетая гамма-аминобутиральдегиддегидрогеназу (EC 1.2.1.3 или 1.2.1.19 или 1.2.1.47) и алкогольдегидрогеназу. В предпочтительном варианте осуществления 6-аминогексанол получают из аминокапроновой кислоты, сочетая гамма-аминобутиральдегиддегидрогеназу (EC 1.2.1.3 или 1.2.1.19 или 1.2.1.47) и указанную выше алкогольдегидрогеназу с использованием метаболического пути для аминокапроновой кислоты.
IV. Сконструированные пути для получения дифункциональных алканов (от C5 до C8) из α-кетокислот (от C5 до C8)
Существует насколько возможных путей для получения дифункциональных алканов (Cn) из источников α-кетокислот (Cn) при помощи рекомбинантных микроорганизмов, как показано на фиг. 5. Аспекты по изобретению относятся к превращению с помощью микроорганизмов α-кето-промежуточных продуктов в молекулы дифункциональных пентанов, дифункциональных гексанов, дифункциональных гептанов, дифункциональных октанов. Молекулы представляющих интерес дифункциональных пентанов включают в качестве неограничивающих примеров 1-гидроксипентаноат, 1,5-пентандиол, глутарат, кадаверин (пентан-1,5-диамин), 5-аминопентанал и 5-аминопентанол. Молекулы представляющих интерес дифункциональных гексанов включают в качестве неограничивающих примеров 1-гидроксигексаноат, 1,6-гександиол, адипат, гексаметилендиамин, аминокапроновую кислоту, 6-аминогексанал и 6-аминогексанол. Молекулы представляющих интерес дифункциональных гептанов включают в качестве неограничивающих примеров 1-гидроксигептаноат, 1,7-гептандиол, пимелиновую кислоту, 1,7-диаминогептан, 7-аминогептанал и 7-аминогептанол. Молекулы представляющих интерес дифункциональных октанов включают в качестве неограничивающих примеров 1,4-октандиол, 1-гидроксиоктаноат, каприловую кислоту, 1,4-диаминооктан, 4-аминооктанал и 4-аминооктанол.
Один аспект по изобретению относится к первому сконструированному пути для получения дифункциональных алканов, который включает 2-аминотрансферазу (ферментативная стадия II), как описано выше. Согласно первому сконструированному пути превращение Cn-α-кетокислоты в дифункциональные Cn-алканы состоит из ферментативных стадий II, 21, 22, 23 и описанных выше стадий 5, 6 и 7, включающих следующие ниже ферменты и превращения субстратов в продукты:
• Ферментативная стадия II: Cn-α-кетокислота в 2-амино-1,n-дикарбоновую кислоту, которая катализируется, например, 2-аминотрансферазой или гомологичным ферментом (например, lysN (EC 2.6.1.7), kat2; aadat и AADAT).
• Ферментативная стадия 21: 2-амино-1,n-дикарбоновая кислота в (n-l)-амино-n-оксокарбоновую кислоту, которая катализируется, например, аминоальдегиддегидрогеназой.
• Ферментативная стадия 22: (n-1)-амино-n-оксокарбоновая кислота в (n-1)-оксо-n-аминокарбоновую кислоту, которая катализируется 2-аминоальдегидмутазой. В иллюстративном варианте осуществления аминоальдегидмутаза представляет собой глутамат-1-полуальдегид-2,1-аминомутазу (EC 5.4.3.8). Глутамат-1-полуальдегид-2,1-аминомутаза (GSAM) описана в пути биосинтеза порфирина и катализирует реакцию превращения (S)-4-амино-5-оксопентаноата в 5-аминолевулинат.
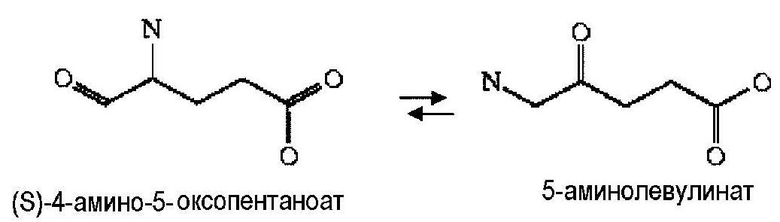
В некоторых вариантах осуществления аминоальдегидмутаза включает глутамат-1-полуальдегид-2,1-аминомутазу из thermosynechococcus elongates, из thermus thermophilics и из aeropyrum pernix (ген heml) и полипептиды, кодируемые геном gsa, например, из Synechococcus sp.
• Ферментативная стадия 23: (n-1)-оксо-n-аминокарбоновая кислота в n-аминокарбоновую кислоту, которая катализируется тремя ферментами: дегидрогеназой (стадия 23a), превращающей кетоновую группу в группу вторичного спирта. За дегидрированием следует дегидратация, которая катализируется дегидратазой (стадия 23b), катализирующей превращение группы вторичного спирта в алкен, и дегидрогеназа, катализирующая превращение алкена в алкан (стадия 23c).
• Ферментативная стадия 5: n-аминокарбоновая кислота в n-аминоальдегид, катализируемая альдегиддегидрогеназой (например, EC 1.2.1.3), после чего следует либо ферментативная стадия 6, либо ферментативная стадия 7.
• Ферментативная стадия 6: n-аминокарбоновая кислота в n-аминоспирт, катализируемая алкогольдегидрогеназой, например, алкогольдегидрогеназами (EC 1.1.1.1 и EC 1.1.1.2).
• Ферментативная стадия 7: n-аминоальдегид в 1,n-диаминоалкан, катализируемая 1-аминотрансферазой.
Другой аспект изобретения относится ко второму пути для получения с помощью микроорганизмов представляющих интерес дифункциональных алканов (от C5 до C8) из α-кетокислоты такой же длины (от C5 до C8). Второй сконструированный путь включает, прежде всего, стадию удаления α-кетона (ферментативная стадия III). В некоторых вариантах осуществления реакция удаления кетона составляет три стадии ферментативных реакций, включающих дегидрогеназу, катализирующую превращение кетоновой группы во вторичный спирт; дегидратазу, катализирующую превращение вторичного спирта в алкен, и дегидрогеназу, катализирующую превращение алкена в алкан. В иллюстративном варианте осуществления адипиновую кислоту синтезируют из α-кетоадипата, объединяя активности первой дегидрогеназы, дегидратазы и второй дегидрогеназы. В некоторых вариантах осуществления первая и вторая дегидрогеназы одинаковы.
В некоторых вариантах осуществления дегидрогеназа представляет собой 3-оксоацил-[ацилпереносящий белок]редуктазу (EC 1.1.1.100), которая, как показано, катализирует следующую ниже реакцию в суперпути биосинтеза жирных кислот:
В некоторых вариантах осуществления дегидрогеназа представляет собой β-кетоацил-[ацилпереносящий белок]редуктазу (ген FabG из E. Coli), синтазу жирных кислот (ген Fas2 из Saccharomyces cerevisiae) или синтазу жирных кислот (ген FASN из Homo sapiens). В некоторых вариантах осуществления дегидратаза представляет собой 3-гидроксиоктаноил-[ацилпереносящий белок]дегидратазу (EC 4.2.1.59), которая катализирует следующую ниже реакцию:
S-Гидроксиоктаноил-[ацилпереносящий белок]дегидратаза включает в качестве неограничивающих примеров 3-гидроксиацил-АПБ-дегидразу из Spinacia oleracea, β-гидроксиацил-АПБ-дегидразу (ген FabA из E. Coli), β-гидроксиацил-АПБ-дегидратазу (ген FabZ из E. Coli). В некоторых вариантах осуществления вторая дегидрогеназа представляет собой еноил-ацилпереносящий белок-редуктазу (EC 1.3.1.9), такую как еноил-АПБ-редуктаза (ген fabI из E. Coli), которая, как показано, катализирует следующую ниже реакцию:

В некоторых вариантах осуществления первая и вторая дегидрогеназы идентичны (синтаза жирных кислот (ген Fas2 из Saccharomyces cerevisiae или ген FASN из Homo sapiens)).
Получение дифункционального алкана включает ферментативную стадию 30, после которой следуют различные ферментативные стадии в зависимости от метаболита или конечного представляющего интерес продукта. Биологический сконструированный путь для получения n-гидроксикарбоновой кислоты и 1,n-алкандиолов включает одну или несколько алкоголь- и/или альдегиддегидрогеназ. В одном из вариантов осуществления сконструированный путь включает ферментативную стадию 30, после которой следуют ферментативные стадии 1 и 2, или ферментативную стадию 30, после которой следует ферментативная стадия 31, как описано ниже и проиллюстрировано на фиг. 4 и фиг. 5. В некоторых вариантах осуществления биологический сконструированный путь для получения n-аминокарбоновой кислоты, n-аминоальдегида, n-аминоспирта или 1,n-диаминоалкана включает одну или несколько аминотрансфераз (ферментативные стадии 4 и 6), альдегиддегидрогеназу (ферментативная стадия 5) и алкогольдегидрогеназу (ферментативная стадия 7). В иллюстративном варианте осуществления аминокапроновую кислоту получают из 6-оксогексаноата посредством сконструированного пути, включающего альдегиддегидрогеназу (ферментативная стадия 30), после которой следует аминотрансфераза (ферментативная стадия 4). Аминокапроновую кислоту можно впоследствии превращать в гексаметилендиамин при помощи альдегиддегидрогеназы (ферментативная стадия 5), за которой следует аминотрансфераза (ферментативная стадия 6). Ферментативные стадии и превращения субстратов в продукты перечислены ниже и проиллюстрированы на фиг. 4 и фиг. 5:
• Ферментативная стадия 30: дикарбоновая кислота в полуальдегид карбоновой кислоты, которая катализируется альдегиддегидрогеназой.
• Ферментативная стадия 1: полуальдегид карбоновой кислоты в гидроксилкарбоновую кислоту, которая катализируется алкогольдегидрогеназой.
• Ферментативная стадия 2: гидроксикарбоновая кислота в 1,n-алкандиол, которая катализируется альдегиддегидрогеназой или алкогольдегидрогеназой.
• Ферментативная стадия 31: полуальдегид карбоновой кислоты в 1,n-алкандиол, которая катализируется альдегиддегидрогеназой или алкогольдегидрогеназой.
• Ферментативная стадия 4: полуальдегид карбоновой кислоты в n-аминокарбоновую кислоту в n-аминоальдегид, которая катализируется альдегиддегидрогеназой.
• Ферментативная стадия 6: n-аминоальдегид в 1,n-диаминоальдегид, которая катализируется 1-аминотрансферазой.
• Ферментативная стадия 7: n-аминоальдегид в n-аминоспирт, которая катализируется алкогольдегидрогеназой.
Методики скрининга
В соответствии со способами, описываемыми в настоящем документе, реакционные смеси для создания пути можно готовить в любом сосуде, который позволяет осуществлять рост и/или инкубацию клеток. Например, реакционная смесь может находиться в биореакторе, в колбе или планшете для клеточной культуры, в многолуночном планшете (например, 96-, 384-, 1056-луночных планшетах для микротитрования и т.д.), колбе для культур, ферментере или других сосудах для роста или инкубации клеток.
Скрининг можно проводить, определяя экспрессию селективного маркера, который, при некоторых генетических обстоятельствах, позволяет клеткам, экспрессирующим маркер, выжить, в то время как другие клетки погибают (или наоборот). Эффективные методики скрининга необходимы для того, чтобы обеспечить эффективную разработку новых путей, используя способы, описываемые в настоящем документе. Предпочтительно подходящие методики скрининга для соединений, получаемых ферментативными путями, позволяют осуществлять быстрый и чувствительный отбор в отношении представляющих интерес свойств. Визуальные (калориметрические) анализы оптимальны в этом отношении и легко применимы для соединений с подходящими свойствами поглощения света. Более совершенные методики скрининга включают, например, высокопроизводительный анализ ВЭЖХ-МС, SPME (твердофазную микроэкстракцию) и ГХ-МС (газовую хроматографию-масс-спектрометрию) (см. Handbook of analytical derivatization reaction, D. R. Knapp; John Wiley & Sons, 1979). В некоторых случаях робот для скрининга подсоединяют к системе ВЭЖХ-МС для автоматической подачи и быстрого анализа образца. Такие методики позволяют проводить высокопроизводительное определение и количественный анализ практически любого необходимого соединения.
Биологически получаемые представляющие интерес продукты можно выделять из ферментационной среды или клеточного экстракта, применяя известные в данной области способы. Например, твердые вещества или клеточный дебрис можно удалять посредством центрифугирования или фильтрации. Представляющие интерес биопродукты можно выделять дистилляцией, экстракцией жидкости жидкостью, мембранным выпариванием, адсорбцией или применяя любые известные в данной области способы.
В некоторых вариантах осуществления идентификацию представляющего интерес продукта можно провести с использованием ВЭЖХ. Например, готовят стандартные образцы с известным количеством органического продукта в среде (например, адипиновая кислота, аминокапроновая кислота). Время удержания продуцируемой адипиновой кислоты можно затем сравнить со временем удержания аутентичного стандарта. В некоторых вариантах осуществления идентификацию представляющего интерес продукта можно провести с использованием ГХ-МС. Разделенные образцы затем анализируют на масс-селективном детекторе и сравнивают с предыдущими масс-спектрами и временами удержания аутентичных стандартов.
В некоторых вариантах осуществления клеточный экстракт можно проверять при помощи скрининга в отношении ферментативной активности. Например, оксогексаноатдегидрогеназную активность можно определить, измеряя степень увеличения поглощения при 340 нм, как описано у Donoghue и Trudgill (Eur. J. Biochem., 1975, 60:1-7).
В практике настоящих способов будут применять, если не указано иначе, общепринятые способы клеточной биологии, культивирования клеток, молекулярной биологии, трансгенной биологии, микробиологии, рекомбинантной ДНК и иммунологии, конструирования, роботизации, оптики, компьютерного программного обеспечения и интеграции. Методики и способы, как правило, выполняют согласно общепринятым способам в данной области и различным общим ссылкам, которые известны специалистам в данной области. Такие методики полностью описаны в литературе. См., например, Molecular Cloning A Laboratory Manual, 2nd Ed., ed. by Sambrook, Fritsch и Maniatis (Cold Spring Harbor Laboratory Press: 1989); DNA Cloning, Volumes I and II (D. N. Glover ed., 1985); Oligonucleotide Synthesis (M. J. Gait ed., 1984); Mullis et al. U.S. Patent № 4683195; Nucleic Acid Hybridization (B. D. Hames & S. J. Higgins eds. 1984); Transcription And Translation (B. D. Hames & S. J. Higgins eds. 1984); Culture Of Animal Cells (R. I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); монографию Methods In Enzymology (Academic Press, Inc., N. Y.); Gene Transfer Vectors For Mammalian Cells (J. H. Miller and M. P. Calos eds., 1987, Cold Spring Harbor Laboratory); Methods In Enzymology, Vols. 154 and 155 (Wu et al. eds.), Immunochemical Methods In Cell And Molecular Biology (Mayer and Walker, eds., Academic Press, London, 1987); Handbook Of Experimental Immunology, Volumes HV (D. M. Weir and C. C. Blackwell, eds., 1986); Manipulating the Mouse Embryo, (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y., 1986); Lakowicz, J. R. Principles of Fluorescence Spectroscopy, New York:Plenum Press (1983) и Lakowicz, J. R. Emerging Applications of Fluorescence Spectroscopy to Cellular Imaging: Lifetime Imaging, Metal-ligand Probes, Multi-photon Excitation and Light Quenching, Scanning Microsc. Suppl VOL. 10 (1996) pages 213-24, в отношении методик флуоресценции, Optics Guide 5 Melles Griot.RTM. Irvine Calif, в отношении общих оптических способов, Optical Waveguide Theory, Snyder & Love, опубликованную Chapman & Hall, и Fiber Optics Devices and Systems Peter Cheo, опубликованную в Prentice-Hall в отношении теории волоконной оптики и материалов.
ЭКВИВАЛЕНТЫ
Настоящее изобретение относится, в частности, к композициям и способам метаболического конструирования. В то время как обсуждались определенные варианты осуществления заявленного изобретения, упомянутое выше описание является иллюстративным и неограничивающим. Множество изменений по изобретению станут очевидны специалистам в данной области при просмотре данного описания. Полный объем изобретения следует определить на основе формулы изобретения, наряду с их полным объемом эквивалентов и спецификации, наряду с такими изменениями.
ВКЛЮЧЕНИЕ В ОПИСАНИЕ ИЗОБРЕТЕНИЯ СВЕДЕНИЙ В КАЧЕСТВЕ ССЫЛКИ
Все публикации и патенты, упомянутые в настоящем документе, включая пункты, перечисленные ниже, включены, таким образом, в полном объеме в качестве ссылки, как если бы каждая отдельная публикация или патент были определенно и индивидуально обозначены так, чтобы быть включенными в качестве ссылки. В случае конфликта настоящая заявка, включая любые определения в настоящем документе, будет проверяться.
| название | год | авторы | номер документа |
|---|---|---|---|
| Новые трансаминазы и способ дезаминирования аминосоединения с их применением | 2016 |
|
RU2718778C2 |
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ОСНОВАННОГО НА РЕКОМБИНАЦИИ ПОЛУЧЕНИЯ СОЕДИНЕНИЙ ШАФРАНА | 2012 |
|
RU2676730C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ЧЕТЫРЕХУГЛЕРОДНЫХ СПИРТОВ | 2006 |
|
RU2394913C2 |
| СПОСОБ ПОЛУЧЕНИЯ 2,4-ДИГИДРОКСИБУТИРАТА | 2013 |
|
RU2645260C2 |
| МОДИФИЦИРОВАННЫЕ МИКРООРГАНИЗМЫ И СПОСОБЫ ПОЛУЧЕНИЯ БУТАДИЕНА С ИХ ПРИМЕНЕНИЕМ | 2012 |
|
RU2639564C2 |
| ДРОЖЖИ, ПРОДУЦИРУЮЩИЕ ЭКТОИН | 2018 |
|
RU2745157C1 |
| ПОЛУЧЕНИЕ 1,4 ДИАМИНОБУТАНА | 2010 |
|
RU2577967C2 |
| НОВЫЕ ФЕРМЕНТЫ МЕТАНОЛДЕГИДРОГЕНАЗЫ ИЗ Bacillus | 2013 |
|
RU2624017C2 |
| БЕСКЛЕТОЧНОЕ ПОЛУЧЕНИЕ САХАРОВ | 2018 |
|
RU2776637C2 |
| Способ получения 2,4-дигидроксимасляной кислоты | 2011 |
|
RU2626531C2 |
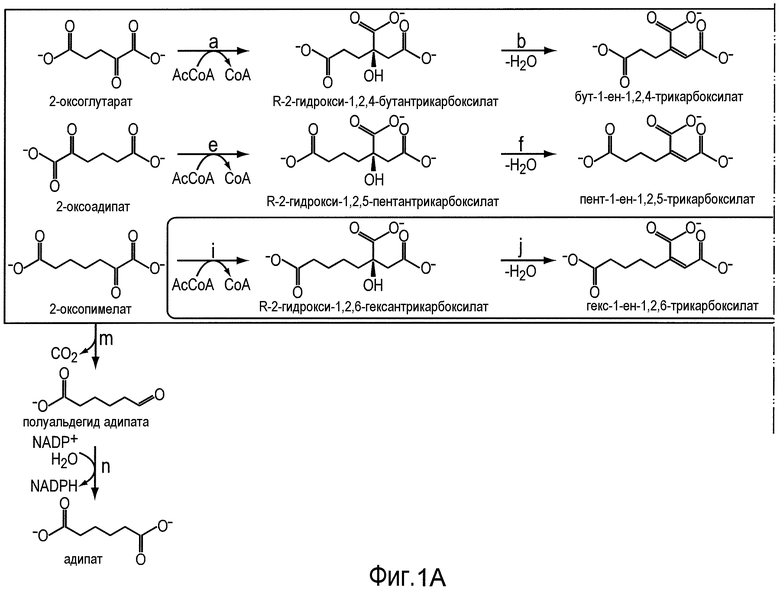
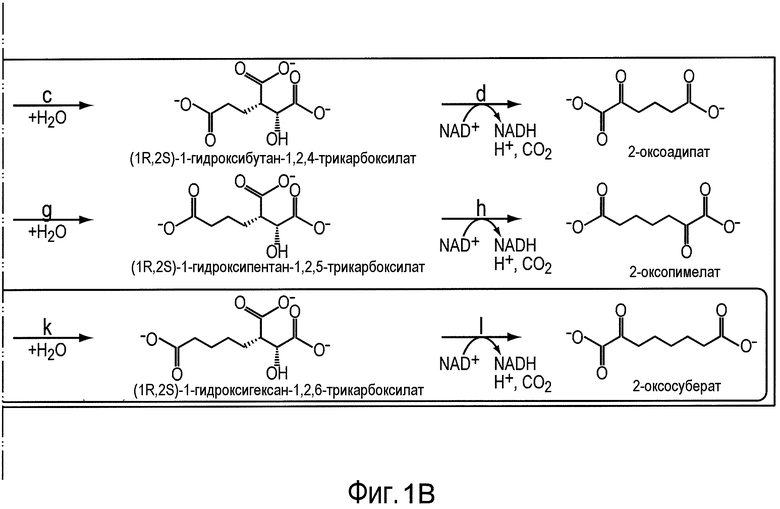
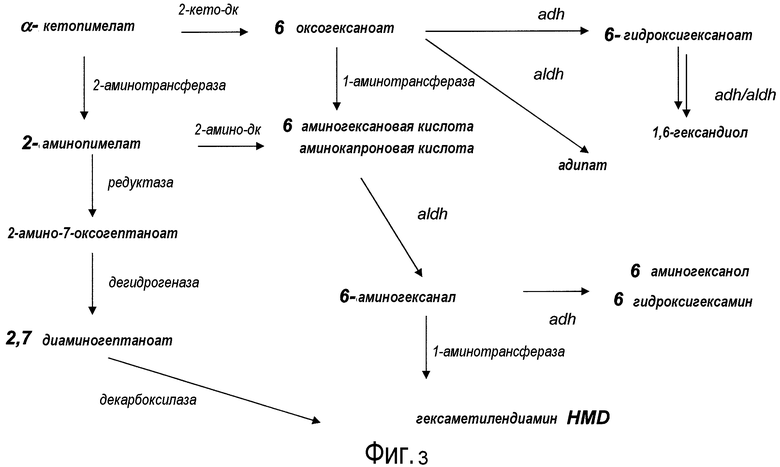
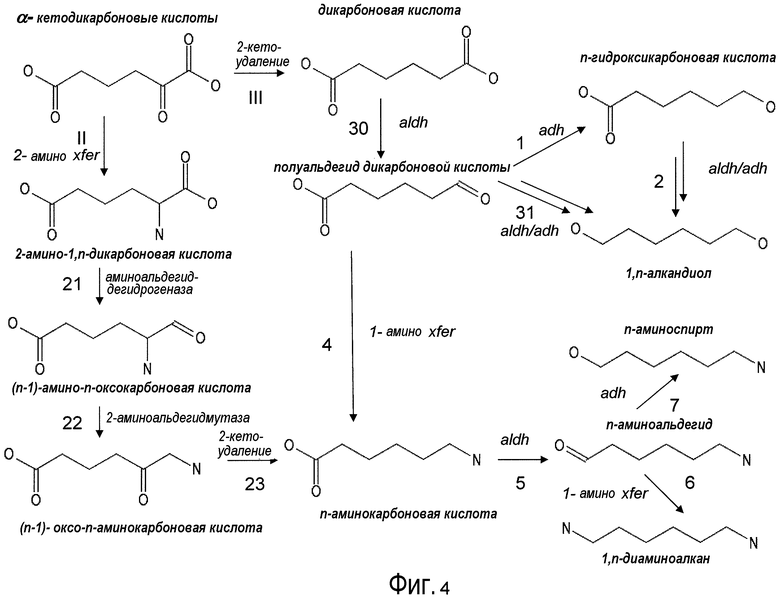
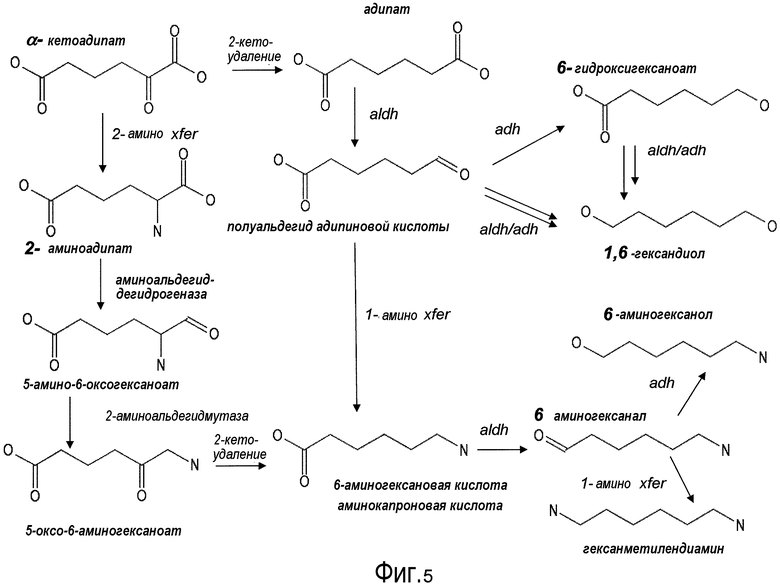
Изобретение относится к области биохимии и молекулярной биологии. Предложен способ получения дифункциональных алканов в рекомбинантной клетке-хозяине из исходного материала углеводов в клетках-хозяевах. Изобретение может быть использовано для получения соединений с использованием микроорганизмов для производства химических веществ, производство которых обычными методами является сложным и дорогостоящим. 2 н. и 5 з.п. ф-лы, 5 ил., 5 табл.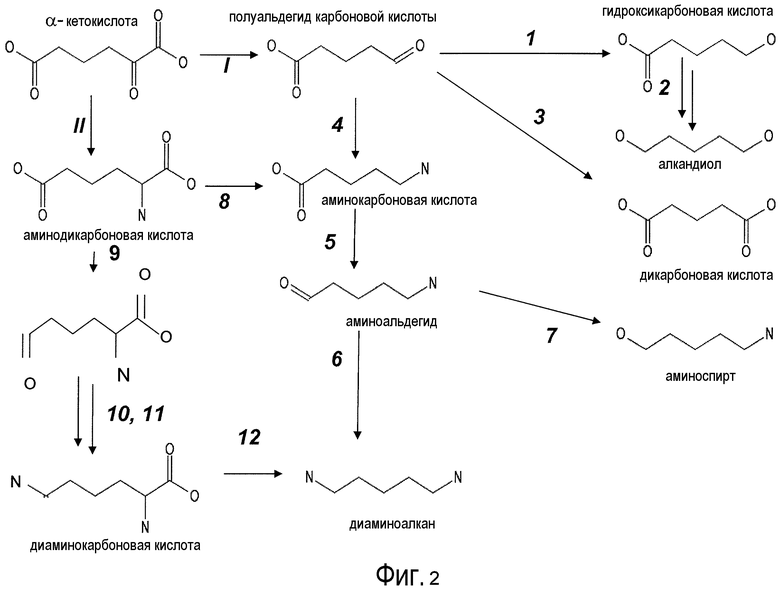
1. Способ получения α,ω-дифункционального Cn-алкана, включающий:
(a) получение рекомбинантной клетки-хозяина, где клетка-хозяин содержит нуклеиновую кислоту, содержащую по меньшей мере одну нуклеотидную последовательность, кодирующую по меньшей мере один фермент пути биосинтеза, для продукции α,ω-дифункционального Сn-алкана из α-кетоглутарата, где концевые функциональные группы α и ω выбраны из группы -ОН, -СООН и -NH3, и где n представляет собой целое число в диапазоне от 4 до 8, где клеткой-хозяином является бактериальная клетка или дрожжевая клетка;
(b) культивирование рекомбинантной клетки-хозяина в условиях, подходящих для продукции α,ω-дифункционального Cn-алкана; и
(c) выделение α,ω-дифункционального Сn-алкана.
2. Способ получения α,ω-дифункционального Cn-алкана, включающий:
(а) получение рекомбинантной клетки-хозяина, где клетка-хозяин содержит нуклеиновую кислоту, содержащую по меньшей мере одну нуклеотидную последовательность, кодирующую гомоцитратсинтазу, гомоаконитазу и гомоизоцитратдегидрогеназу для продукции α-кетоадипата, α-кетопимелата или α-кетосуберата, и по меньшей мере одну нуклеотидную последовательность, кодирующую фермент 2-кетодекарбоксилазу, где клеткой-хозяином является бактериальная клетка или дрожжевая клетка;
(b) культивирование рекомбинантной клетки-хозяина в условиях, подходящих для продукции α,ω-дифункционального Cn-алкана, где концевые функциональные группы α и ω выбраны из группы -ОН, -СООН и -NH3, и где n представляет собой целое число в диапазоне от 4 до 8; и
(c) выделение α,ω-дифункционального Сn-алкана.
3. Способ по п.2, в котором:
(a) гомоцитратсинтаза выбрана из группы, состоящей из AksA, NifV, hcs, Lys20/21 и их гомологов;
(b) гомоаконитаза выбрана из группы, состоящей из AksD/E, LysT/U, Lys4, большой/малой субъединиц 3-изопропилмалатдегидратазы и их гомологов; и
(c) гомоизоцитратдегидрогеназа выбрана из группы, состоящей из AksF, Hicdh, Lysl2, 2-оксосубератсинтазы, 3-изопропилмалатдегидрогеназы и их гомологов.
4. Способ по п.2, в котором 2-кетодекарбоксилаза выбрана из группы 2-кетоглутаратдекарбоксилазы (kgd), 2-кетоизовалератдекарбоксилазы (kivD), декарбоксилазы трансаминированной аминокислоты (ARO10), бензоилформиатдекарбоксилазы (mdlC), 2-кетоаргининдекарбоксилазы (aruI), фосфонопируватдекарбоксилазы (fom2), изофермента пируватдекарбоксилазы (PDC6, PDC1), изофермента 2 пируватдекарбоксилазы (PDC5, PDC1, PDC6, Aro10, KivD), индолпируватдекарбоксилазы (ipdC) и их гомологов.
5. Способ по п.2, в котором клетка-хозяин дополнительно содержит по меньшей мере одну нуклеотидную последовательность, кодирующую:
(а) фермент альдегиддегидрогеназу для продукции дифункционалыюго Cn-алкана, где по меньшей мере одна из концевых функциональных групп представляет собой карбоновую кислоту (-СООН), где альдегиддегидрогеназа необязательно представляет собой 6-оксогексаноатдегидрогеназу (ChnE) и ее гомологи, или
(b) фермент аминотрансферазу для продукции дифункционального Сn-алкана, где по меньшей мере одна из концевых функциональных групп представляет собой амин (-NH3), где аминотрансфераза необязательно выбрана из группы, состоящей из α-аминоадипатаминотрансферазы-1 (AADAT), аминоадипатаминотрансферазы (LysN), диаминопимелатдегидрогеназы (ddb и dapdh), трансаминазы ГАМК, дегидрогеназы Lys6, трансаминазы орнитиноксокислоты, лизинаминотрансферазы, 4-аминобутиратаминотрансферазы, сахаропиндегидрогеназы (LYS9 и LYS1) и их гомологов, или
(c) алкогольдегидрогеназу для продукции дифункционального Сn-алкана, где по меньшей мере одна из концевых функциональных групп представляет собой спирт (-ОН), и где алкогольдегидрогеназа необязательно выбрана из 6-гидроксигексаноатдегидрогеназы, бутанолдегидрогеназы, дегидрогеназы ADHIV, пропандиолоксидоредуктазы, ADH6 и их гомологов.
6. Способ по любому из пп.1-5, в котором α,ω-дифункциональный алкан имеет шесть атомов углерода и выбран из группы, состоящей из адипиновой кислоты, аминокапроновой кислоты, гексаметилендиамина, 6-гидроксигексамина, 1,6-гександиола, 6-аминогексанала, 6-аминогексанола и 6-гидроксигексаноата.
7. Способ по п.1, в котором α-кетокислота представляет собой α-кетоглутарат, α-кетоадипат, α-кетопимелат или α-кетосуберат.
| Способ получения производных алканов или алкенов | 1982 |
|
SU1329615A3 |
| WO 2009046375 А2, 09.04.2009. | |||

