Описание
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к генному кластеру, отвечающему за биосинтез сафрацина, к его применению в генной инженерии и к новым сафрацинам, получаемым путем коррекции механизмов биосинтеза.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
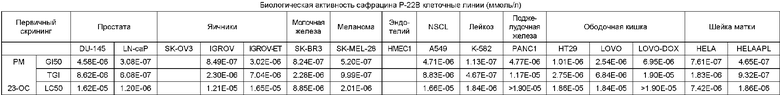
Сафрацины, представляющие собой семейство новых соединений с широким спектром мощной антибактериальной активности, были обнаружены в культуральном бульоне Pseudomonas sp. Сафрацины встречаются в двух штаммах Pseudomonas sp., Pseudomonas fluorescens A2-2, выделенного из образца почвы, собранного в Тагавагуне, Фукуока, Япония (Ikeda et al., J. Antibiotics 1983, 36, 1279-1283; WO 82 00146 и JP 58113192), и в штамме Pseudomonas fluorescens SC 12695, выделенного из образцов воды, взятых из канала Раритан-Делаваре возле Нью-Джерси (Meyers et al., J. Antibiot. 1983, 36(2), 190-193). Сафрацины A и В, продуцируемые Pseudomonas fluorescens А2-2, были исследованы в отношении их воздействия на различные линии опухолевых клеток и было обнаружено, что они кроме антибактериальной активности обладают противоопухолевой активностью:
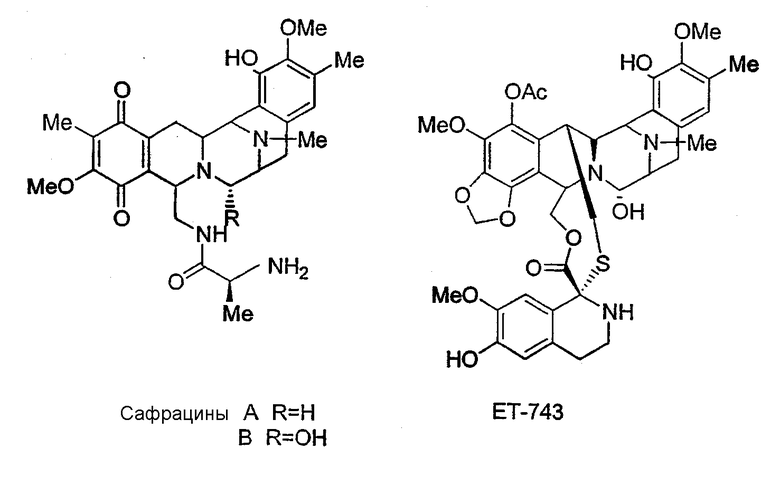
В результате структурного сходства между сафрацином В и ЕТ-743 с помощью сафрацина возможно осуществлять полусинтез высокоперспективного мощного нового противоопухолевого средства ЕТ-743, выделенного из оболочечного морского организма Еcteinascidia turbinata, который в настоящее время проходит II фазу клинических испытаний в Европе и США. Полусинтез ЕТ-743 был осуществлен с использованием сафрацина В в качестве исходного соединения (Cuevas et al., Organic Lett. 2000, 10, 2545-2548; WO 00 69862 и WO 01 87895).
Альтернативный способ получения сафрацинов или их структурных аналогов химическим синтезом посредством воздействия на гены, осуществляющие контроль вторичного метаболизма, является весьма перспективными и с помощью него можно получать эти соединения биосинтетическим путем. Кроме того, благодаря структуре сафрацина можно осуществлять комбинаторный биосинтез.
В связи с тем, что сафрацины имеют структуру комплекса и его получение из Pseudomonas fluorescens А2-2 ограничено, было бы весьма желательно изучить генетическую основу их синтеза с тем, чтобы создать способы воздействия на них заданным образом. Это могло бы увеличить количество продуцируемых сафрацинов, поскольку природные штаммы, продуцирующие сафрацин, обычно дают лишь небольшие концентрации вторичных метаболитов, представляющих интерес. Такие способы также могут обеспечить продукцию сафрацинов у тех хозяинов, которые не продуцируют такие соединения. Кроме того, генетическое воздействие может использоваться для комбинаторного создания новых аналогов сафрацина, которые могут обладать улучшенными свойствами и которые могут использоваться в полусинтезе новых эктеинасцидиновых соединений.
Однако успех биосинтетического подхода главным образом зависит от доступности новых генетических систем и от генов, кодирующих новые ферменты с соответствующей активностью. Изучение генного кластера сафрацина вносит вклад в основные положения комбинаторного биосинтеза за счет расширения спектра генов, которые уникальным образом ассоциированы с биосинтезом сафрацина, что приведет к возможному получению новых предшественников и сафрацинов путем комбинаторного биосинтеза.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящее время можено идентифицировать и клонировать гены биосинтеза сафрацина, создав генетическую базу для улучшения, а также воздействия заданным образом на продуктивность Pseudomonas sp. и используя генетические методы синтеза аналогов сафрацина. Кроме того, эти гены кодируют ферменты, которые вовлечены в процессы биосинтеза, приводя к образованию структур, таких как предшественники сафрацина, которые могут сформировать базу комбинаторной химии для получения большого разнообразия соединений. Эти соединения могут быть подвергнуты скринингу на наличие большого диапазона биологической активности, включая противораковую активность.
В этой связи, в первом аспекте настоящее изобретение относится к нуклеиновой кислоте, соответственно к выделенной нуклеиновой кислоте, которая включает последовательность ДНК (включая ее мутации или варианты), которая кодирует нерибосомальные пептидсинтетазы, ответственные за биосинтез сафрацинов. Настоящее изобретение относится к генному кластеру, соответственно к выделенному генному кластеру с открытыми рамками считывания, кодирующими полипептиды, которые относятся к сборке молекулы сафрацина.
В одном из аспектов настоящее изобретение относится к композиции, включающей по меньшей мере одну последовательность нуклеиновой кислоты, соответственно к выделенной молекуле нуклеиновой кислоты, которая кодирует по меньшей мере один полипептид, катализирующий по меньшей мере одну стадию биосинтеза сафрацинов. В композиции могут присутствовать две или более такие последовательности нуклеиновой кислоты. Изобретение также относится к ДНК или соответствующей молекуле РНК.
В частности, настоящее изобретение относится к последовательности нуклеиновой кислоты, соответственно к выделенной последовательности нуклеиновой кислоты, из генного кластера сафрацина, включающего указанную последовательность нуклеиновой кислоты, часть или части указанной последовательности нуклеиновой кислоты, где указанные часть или части кодируют полипептид или полипептиды, или биологически активный фрагмент полипептида или полипептидов, к последовательности одноцепочечной нуклеиновой кислоты, полученной из указанной последовательности нуклеиновой кислоты, или к последовательности одноцепочечной нуклеиновой кислоты, выделенной из части или частей указанной последовательности нуклеиновой кислоты, или к последовательности двуцепочечной нуклеиновой кислоты, полученной из последовательности одноцепочечной нуклеиновой кислоты (такой как кДНК из мРНК). Последовательность нуклеиновой кислоты может представлять ДНК или РНК.
Более конкретно настоящее изобретение относится к последовательности нуклеиновой кислоты, соответственно к выделенной последовательности нуклеиновой кислоты, которая включает или содержит по меньшей мере SEQ ID 1, ее варианты или части, или по меньшей мере один из генов sacA, sacB, sacC, sacC, sacD, sacE, sacF, sacG, sacH, sacH, sacI, sacJ, orf1, orf2, orf3 или orf4, включая их варианты или части. Длина частей может составлять по меньшей мере 10, 15, 20, 25, 50, 100, 1000, 2500, 5000, 10000, 20000, 25000 или более нуклеотидов. Обычно длина этих частей находится в диапазоне от 100 до 5000 или от 100 до 2500 нуклеотидов и обладает биологической активностью.
Мутанты или варианты включают полинуклеотидные молекулы, в которых по меньшей мере один нуклеотидный остаток изменен, замещен, удален или введен. Также возможны множественные изменения различных нуклеотидов в 1, 2, 3, 4, 5, 10, 15, 25, 50, 100, 200, 500 или более положениях. Также возможны вырожденные варианты, которые кодируют один и тот же полипептид, а также невырожденные варианты, которые кодируют разные полипептиды. Часть, мутант или вариант последовательности нуклеиновой кислоты соответственно кодирует полипептид, который сохраняет биологическую активность соответствующего полипептида, кодируемого любой из открытых рамок считывания генного кластера сафрацина. Также возможны аллельные формы и полиморфизмы.
Настоящее изобретение также относится к последовательности выделенной нуклеиновой кислоты, способной гибридизоваться в жестких условиях с последовательностью нуклеиновой кислоты по настоящему изобретению. Особенно предпочтительной является гибридизация с последовательностью нуклеиновой кислоты по настоящему изобретению, имеющей транслируемую длину.
Настоящее изобретение также относится к нуклеиновой кислоте, кодирующей полипептид, который по меньшей мере на 30%, предпочтительно на 50%, предпочтительно на 60%, более предпочтительно на 70%, в частности на 80%, 90%, 95% или более идентичен по аминокислотной последовательности полипептиду, кодируемому любой из открытых рамок считывания генного кластера сафрацина sacA - sacJ и оrf1 - orf4 (SEQ ID 1 и гены, кодируемые в SEQ ID 1) или кодируемому их вариантом или частью. Полипептид при этом сохраняет биологическую активность соответствующего полипептида, кодируемого любой из открытых рамок считывания генного кластера сафрацина.
В частности, настоящее изобретение относится к последовательности выделенной нуклеиновой кислоты, кодирующей любой белок SacA, SacB, SacC, SacD, SacE, SacF, SacG, SacH, SacI, SacJ, Orf1, Orf2, Orf3 или Orf4 (SEQ ID 2-15) и к ее вариантам, мутантам или частям.
В одном из аспектов последовательность выделенной нуклеиновой кислоты по настоящему изобретению кодирует пептидсинтетазу, L-Tyr производное гидроксилазы, L-Tyr производное метилазы, L-Tyr О-метилазу, метилтрансферазу или монооксигеназу, или белок резистентности к сафрацину.
Настоящее изобретение также относится к зонду для гибридизации, который представляет собой последовательность нуклеиновой кислоты, определенную выше, или ее часть. Зонды соответствующим образом включают последовательность из 5, 10, 15, 20, 25, 30, 40, 50, 60 или более нуклеотидных остатков. Предпочтительными являются последовательности с длиной от 25 до 60. Настоящее изобретение также относится к применению определенного выше зонда для обнаружения гена сафрацина или эктеинасцидина. В частности, данный ген используется для обнаружения генов в Ecteinascidia turbinata.
В родственном аспекте настоящее изобретение относится к полипептиду, кодируемому последовательностью нуклеиновой кислоты, определенной выше. Предусматривается полная последовательность, вариант, мутант или фрагмент полипептидов.
В другом аспекте настоящее изобретение относится к вектору, предпочтительно к вектору экспрессии, предпочтительно к космиде, содержащему последовательность нуклеиновой кислоты, кодирующую белок или биологически активный фрагмент белка, где указанная нуклеиновая кислота определена выше.
В другом аспекте настоящее изобретение относится к клетке-хозяину, трансформированной одной или более последовательностями нуклеиновой кислоты, определенными выше, или вектором, вектором экспрессии или космидой, определенной выше. Предпочтительную клетку-хозяина трансформируют экзогенной нуклеиновой кислотой, содержащей генный кластер, который кодирует полипептиды, достаточные для обеспечения сборки сафрацина или аналога сафрацина. Предпочтительно клетка-хозяин представляет собой микроорганизм, более предпочтительно бактерию.
Настоящее изобретение также относится к рекомбинантной бактериальной клетке-хозяину, в которой по меньшей мере часть последовательности нуклеиновой кислоты, определенной выше, разрушается с образованием рекомбинантной клетки-хозяина, которая продуцирует измененные уровни сафрацинового соединения или аналога сафрацина, по сравнению с соответствующей нерекомбинантной бактериальной клеткой-хозяином.
Настоящее изобретение также относится к способу получения сафрацинового соединения или аналога сафрацина, который заключается в ферментации в условиях и в среде, подходящих для продукции такого соединения или аналога, организма, такого как Pseudomonas sp., в котором повышается количество генов/кластера сафрацина, кодирующих полипептиды, достаточные для осуществления сборки сафрацина или аналогов сафрацина.
Настоящее изобретение также относится к способу получения сафрацинового соединения или аналога сафрацина, который заключается в ферментации в условиях или в среде, подходящих для получения такого соединения или аналога, организма, такого как Pseudomonas sp., в котором экспрессия генов, кодирующих полипептиды, достаточная для осуществления сборки сафрацина или аналогов сафрацина, модулируется путем воздействия или замещения одного или более генов или последовательностей, ответственных за регуляцию такой экспрессии. Предпочтительно экспрессия генов при этом повышается.
Настоящее изобретение также относится к применению композиции, содержащей по меньшей мере одну выделенную последовательность нуклеиновой кислоты, определенную выше, или ее модификацию, для комбинаторного биосинтеза нерибосомальных пептидов, дикетопиперазиновых колец и сафрацинов.
В частности, настоящий способ включает взаимодействие соединения, которое является субстратом полипептида, кодируемого одной или несколькими открытыми рамками считывания генного кластера биосинтеза сафрацина, определенного выше, с полипептидом, кодируемым одной или несколькими открытыми рамками считывания генного кластера биосинтеза сафрацина, при этом полипептид химически модифицирует это соединение.
В еще одном варианте настоящее изобретение также относится к способу получения сафрацина или аналога сафрацина. Этот способ включает получение микроорганизма, трансформированного экзогенной нуклеиновой кислотой, содержащей генный кластер сафрацина, который кодирует полипептиды, достаточные для осуществления сборки указанного сафрацина или аналога сафрацина; культивирование бактерий в условиях, при которых возможен биосинтез сафрацина или аналога сафрацина; и к выделению указанного сафрацина или аналога сафрацина из указанной клетки.
Настоящее изобретение также относится к любому соединению-предшественнику P2, P14, их аналогам и производным и к их применению в комбинаторном биосинтезе нерибосомальных белков, дикетопиперазиновых колец и сафрацинов.
Кроме того, настоящее изобретение также относится к новым сафрацинам, полученным путем выключения генов сафрацина P19B, P22A, P22В, D и Е, и к их применению в качестве антимикробных или противоопухолевых средств, а также к их применению в синтезе эктеинасцидиновых соединений.
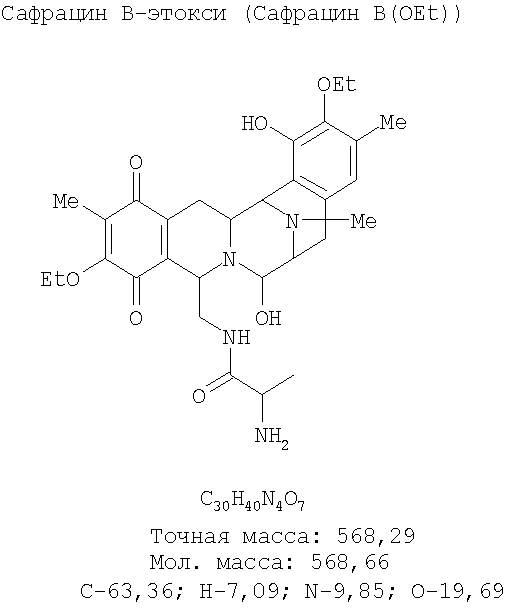
Настоящее изобретение также относится к новым сафрацинам, полученным путем направленного биосинтеза, определенного выше, и к их применению в качестве антимикробных или противоопухолевых средств, а также к их применению в синтезе эктеинасцидиновых соединений. В частности, изобретение также относится к сафрацин-B-этокси или сафрацин-А-этокси и к их применению.
В одном из аспектов настоящее изобретение относится к получению структур, близких к сафрацинам и эктеинасцидинам, которые не могут быть получены химическим путем или это получение затруднено. Еще один аспект относится к использованию знаний, позволяющих осуществить биосинтез эктеинасцидинов в Ecteinascidia turbinata, например, c использованием этих последовательностей или их частей в качестве зондов в указанном организме или предполагаемом симбионте.
В основном настоящее изобретение открывает широкие возможности и позволяет получать эктеинасцидины с помощью генной инженерии.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1: Структурная организация хромосомного участка ДНК, клонированного в космиду pL30p. Показан участок ДНК P. fluorescens A2-2, содержащий генный кластер сафрацина. На чертеже показаны и sacABCDEFGH, и sacIJ, генные опероны и модулярная организация пептидсинтетаз, полученная на основе sacA, sacB и sacC. Указываются следующие домены: С - конденсация; Т - тиолирование; А - аденилирование и Re - редуктаза. Показано также расположение других генов, имеющихся в космиде pL30p (orf1 - orf4), а также их предполагаемые функции.
Фиг.2: Консервативные ядерные мотивы среди NRPS. Консервативные аминокислотные последовательности в белках SacA, SacB и SacC и их сравнение с гомологичными последовательностями из Myxococcus xanthus DM50415.
Фиг.3: Механизм биосинтеза NRPS, предлагаемый для образования дипептида Ala-Gly. Стадия a*: аденилирование Ala; b*: перенос на 4'-фосфопантетеиниловое плечо; c*: перенос к сайту ожидания/элонгации; d*: аденилирование Gly; e*: перенос на 4'-фосфопантетеиниловое плечо; f*: конденсация элонгируемой цепи на 4'-фосфопантетеиниловом плече со стартерной цепью на сайте ожидания/элонгации; g*: дипептид Ala-Gly, присоединенный к фосфопантетеиниловому плечу SacA и h*: перенос удлиненной цепи к следующему сайту ожидания/элонгации.
Фиг.4: Перекрестный эксперимент: А. Схема фрагментов ДНК А2-2, клонированных в вектор pBBR1-MCS2, и продукты, полученные в гетерологичном хозяине. B. Параметры ВЭЖХ сафрацина, продуцируемого в штамме дикого типа, в сравнении с sacF мутантом. Добавление предшественника Р2 к мутанту sacF при условии, что оба находятся в транс-положении и был использован синтетический метод, приводит к образованию сафрацина B. SfcA - сафрацин A и SfcB - сафрацин B.
Фиг.5: Схема механизма биосинтеза сафрацина и промежуточных продуктов биосинтеза. Отдельные ферментативные стадии указаны сплошными стрелками, а реакции с множественными стадиями указаны прерывистыми стрелками.
Фиг.6: Разрушение гена сафрацина и полученные соединения. А. Разрушение гена и молекул предшественника, синтезированных с использованием сконструированных мутантов. Гены, помеченные звездочкой, не принадлежат к сафрациновому кластеру. Было показано, что инактивация генов orf1, orf2, orf3 и orf4 не влияет на продукцию сафрацина. B. Параметры ВЭЖХ сафрацина, продуцируемого в штамме дикого типа, и в мутантах sacA, sacI и sacJ. Показаны структуры различных полученных молекул.
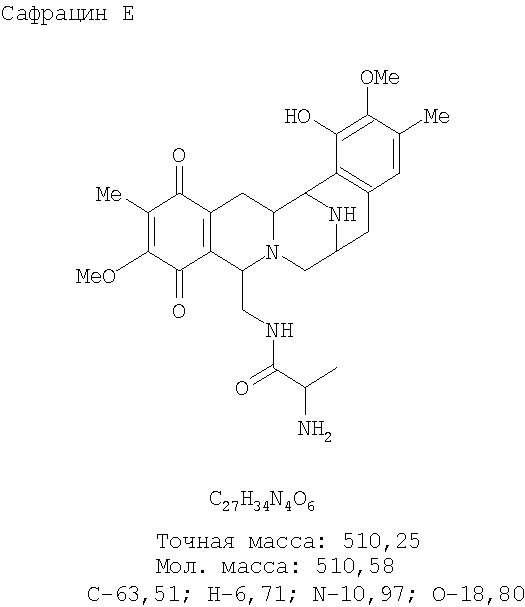
Фиг.7: Структура различных молекул, полученных при разрушении гена. Инактивация белка SacJ (a) приводит к образованию молекул P22B, P22A и Р19, тогда как разрушение гена sacI (b) дает только соединение P19. Разрушение sacI в сочетании с восстановленной экспрессией sacJ дает два новых сафрацина: сафрацин D (возможный предшественник в полусинтезе ET-729) и сафрацин Е (с).
Фиг.8: Добавление специфически разработанных "неприродных" предшественников (Р3). Химическая структура двух молекул, полученных при добавлении Р3 соединения к мутанту sacF.
Фиг.9: Схема процесса разрушения гена путем простой рекомбинации с использованием фрагмента гомологичной ДНК, клонированного в pK18:MOB (плазмида, интегрированная в Pseudomonas).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Нерибосомальные пептидсинтетазы (NRPS) представляют собой ферменты, отвечающие за биосинтез семейства соединений, которое охватывает большое количество структурно и функционально различных природных продуктов. Так, например, пептидов с биологической активностью, которая обеспечивает структурный каркас соединений с разнообразной биологической активностью, таких как антибиотики, противовирусные, противоопухолевые и иммуносупрессорные соединения (Zuber et al., Biotechnology of Antibiotics 1997 (W. Strohl, ed.), 187-216 Marcel Dekker, Inc., N.Y; Мarahiel et al., Chem. Rev. 1997, 97, 2651-2673).
Несмотря на структурное различие, большая часть этих биологически активных пептидов имеет общий механизм биосинтеза. В соответствии с этой моделью образование пептидной связи происходит на мультиферментных комплексах, разработанных пептидсинтетазах, на которых аминокислотные субстраты активируются за счет гидролиза АТФ до соответствующего аденилата. Полученный нестабильный промежуточный продукт затем переносится на другой сайт мультиферментного комплекса, где он связывается в виде сложного тиоэфира с цистеаминовой группой фермент-связанного 4'-фосфопантетенинилового (4'-РР) кофактора. На этой стадии тиол-активированные субстраты могут модифицироваться, например эпимеризироваться или N-метилироваться. Тиоэтерифицированные аминокислотные субстраты затем объединяются с образованием пептидного продукта через постадийную элонгацию в серии реакций транспептидации. При таком временном размещении пептидсинтетаз модули, по-видимому, действуют независимо друг от друга, но они действуют сообща и катализируют образование соответствующих пептидных связей (Stachelhaus et al., Science 1995, 269, 69-72; Stachelhaus et al., Сhem. Biol, 1996, 3, 913-921). Общая схема нерибосомального биосинтеза пептидов подробно описана в литературе (Marahiel et al., Chem. Rev. 1997, 97, 2651-2673; Konz and Marahiel, Chem. and Biol. 1999, 6, R39-R48; Moffit and Neilan, FEMS Microbiol. Letters 2000, 191, 159-167).
Было клонировано большое количество бактериальных оперонов и генов грибов, кодирующих пептидсинтетазы, которые были недавно клонированы, секвенированы и частично охарактеризованы, что дало ценную информацию об архитектуре молекулы (Marahiel et al., Chem. and Biol. 1997, 4, 561-567). Использовали различные подходы клонирования, включая исследование библиотек экспрессии антителами, возникающими в ответ на пептидсинтетазы, комплементацию дефицитных мутантов и использование олигонуклеотидов, сконструированных на основе аминокислотных последовательностей фрагментов пептидсинтетазы.
Анализ первичной структуры этих генов показал наличие явных гомологичных доменов размером примерно 600 аминокислот. Эти специфические функциональные домены состоят по меньшей мере из шести высококонсервативных последовательностей с длиной от трех до восьми аминокислот, порядок следования которых и расположение во всех известных доменах очень сходен (Kusard and Marahiel, Peptide Research, 1994, 7, 238-241). Использование вырожденных олигонуклеотидов, полученных из консервативных участков, дает возможность идентификации и клонирования пептидсинтетаз из геномных ДНК путем использования методики полимеразно-цепьевой реакции (ПЦР) (Kusard and Marahiel, Peptide Research, 1994, 7, 238-241; Borchert et al., FEMS Microbiol Letters 1992, 92, 175-180).
Структура сафрацина предполагает, что это соединение синтезируется по механизму NRPS. Клонирование и экспрессия нерибосомальных пептидсинтетаз и ассоцированных «хвостовых» ферментов из кластера сафрацина Pseudomonas fluorescens A2-2 могло бы позволить продукцию неограниченного количества сафрацина. Кроме того, клонированные гены могли бы использоваться для создания комбинаторными методами новых аналогов сарфацина, которые могут обладать улучшенными свойствами и которые могут использоваться в полусинтезе новых эктеинасцидинов. Кроме того, клонирование и экспрессия генного кластера сафрацина в гетерологичных системах или сочетание генного кластера сафрацина с другими генами в системе NRPS может привести к созданию новых лекарственных средств с улучшенной активностью.
Настоящее изобретение относится, в частности, к последовательности ДНК, кодирующей NRPS, которая ответственна за биосинтез сафрацина, например синтетаз сафрацина. Авторы описали участок длиной 26705 пн (SEQ ID No. 1) из генома Pseudomonas fluorescens A2-2, клонированного в космиде pL30P, и показали путем экспериментов с выключением генов и гетерологичной экспрессии, что этот участок ответственен за биосинтез сафрацина. Авторы экспрессировали космиду pL30P в двух штаммах Pseudomonas sp., которые не продуцируют сафрацин, и в результате осуществили продукцию сафрацина A и B на уровнях до 22% в случае Pseudomonas fluorescens (CECT 378), и 2% в случае Pseudomonas aerugenosa (CECT 110) по сравнению с продукцией P. fluorescens A2-2. Предсказанные аминокислотные последовательности различных белков, кодируемых этой последовательностью ДНК, показаны в последовательностях SEQ ID No. 2 - SEQ ID No. 15 соответственно.
Выделенный из P. fluorescens A2-2 генный кластер биосинтеза сафрацина характеризуется наличием нескольких открытых рамок считывания (OРС), которые организованы в два различных оперона (фиг.1), в оперон с восемью генами (sacABCDEFGH) и оперон с двумя генами (sacIJ), которым предшествуют достаточно консервативные участки предположительных перекрывающихся промоторов. Генный кластер биосинтеза сафрацина присутствует лишь в одной копии генома P. fluorescens A2-2.
Полученные авторами результаты указывают на то, что оперон из восьми генов возможно отвечает за биосинтез каркаса сафрацина, а оперон из двух генов возможно ответственен за создание на финальном этапе «хвостового» участка в молекулах сафрацина.
В опероне sacABCDEFGH установленные аминокислотные последовательности, кодируемые sacA, sacB и sacC, очень сходны с генными продуктами NRPS. В пределах установленных аминокислотных последовательностей SacA, SacB и SacC на каждой из ОРС идентифицирован один пептидсинтетазный модуль.
Первым удивительным свойством белков сафрациновых NRPS является то, что среди известных активных сайтов и коровых участков пептидсинтетаз (Konz and Marahiel, Chem. and Biol. 1999, 6, R39-R48) первый коровый участок является слабо консервативным во всех трех пептидсинтетазах: SacA, SacB и SacC (фиг.2). Другие пять коровых участков являются достаточно консервативными в трех генах сафрациновых NRPS. Биологическое значение первого корового участка (LKAGA) не известно, но ядерные последовательности SGT(ST)TGxPKG (Gocht and Marahiel, J. Bacteriol. 1994, 176, 2654-266; Konz and Marahiel, Chem. and Biol. 1999, 6, R39-R48), TGD (Gocht and Marahiel, J. Bacteriol. 1994, 176, 2654-2662; Konz and Marahiel, 1999) и KIRGxRIEL (Pavela-Vrancic et al., J. Biol Chem. 1994, 269, 14962-14966; Konz and Marahiel, Chem. and Biol. 1999, 6, R39-R48) могут быть предназначены для связывания АТФ и гидролиза. Остаток серина в коровой последовательности LGGxS может быть представлен как сайт образования сложного тиоэфира (D'Souza et al., J. Bacteriol. 1993, 175, 3502-3510; Vollenbroich et al., FEBS Lett. 1993, 325(3), 220-4; Konz and Marahiel, Chem. and Biol. 1999, 6, R39-R48) и связывания 4'-фосфопантетеина (Stein et al., FEBS Lett. 1994, 340, 39-44; Konz and Marahiel, Chem. and Biol. 1999, 6, R39-R48). Эти результаты наряду с тем фактом, что сафрацин, по всей видимости, синтезируется из аминокислот, поддерживает гипотезу о том, что образование нерибосомальной пептидной связи через механизм участия тио-содержащей матрицы происходит в процессе пути биосинтеза сафрацина и что sacA, sacB и sacC кодируют соответствующие пептидсинтетазы. В соответствии с этим механизмом аминокислоты активируются как аминоациладенилаты в результате гидролиза АТФ и затем ковалентно связываются с ферментом через карбоксил-тиоэфирные связи. Затем на последующих стадиях происходит транспептидация и образование пептидной связи.
Во-вторых, удивительно, что полученные авторами данные четко указывают на то, что в системе синтетаз сафрацина не сохраняется правило колинеарности, в соответствии с которым порядок модулей аминокислотного связывания по хромосоме соответствует порядку аминокислот в пептиде. В соответствии с информацией, имеющейся в базе данных по гомологии последовательностей, а также по гомологии сафрациновых и сафрамициновых структур, SacA отвечает за распознавание и активацию остатка Gly, а SacВ и SacС отвечают за распознавание и активацию двух производных L-Tyr, которые включаются в сафрациновый каркас, тогда как предполагаемый ген Ala-NRPS может отсутствовать в генном кластере сафрацина. В нескольких нерибосомальных генных кластерах пептидсинтетаз, например, пути биосинтеза пристамицина (Crecy-Lagard et al., J. of Вacteriol. 1997, 179(3), 705-713) и фосфинотрицинового трипептида (Schwartz et al,. Appl Environ Microbiol 1996, 62, 570-577), первый NRPS расположен не рядом со вторым геном NRPS. Конкретно в пути биосинтеза пристамицина первый структурный ген (snbA) и второй структурный ген (snbC) находятся на расстоянии 130 т.п.н. Такая ситуация не наблюдается в генном кластере сафрацина, для которого по результатам гетерологичной экспрессии с использованием космиды pL30P четко видно отсутствие NRPS гена, поскольку происходит гетерологичная продукция сафрацина.
В-третьих, даже если вопрос о механизме, посредством которого образуется дипептид Ala-Gly, остается открытым, присутствие sacA за пределами С домена на аминоконце первого гена NRPS позволяет предположить, что данный ген будет действовать в системе бифункциональной активации аденилирования. Авторы полагают, что Ala сначала может быть внесен на фосфопантетеиниловое «плечо» SacA (фиг.3a* и b*) перед переносом в положение ожидания, домен конденсации, расположенный на N-конце SacA (фиг.3с*). Затем Gly аденилат вносится на то же самое фосфопантетеиниловое плечо (фиг.3d* и e*), расположенное в сайте элонгации, и происходит элонгация (фиг.3f*). Плечо первого модуля будет на данном этапе нести дипептид Ala-Gly (фиг.3g*). Авторы полагают, что этот/такой дипептид будет затем перенесен в положение ожидания второго фосфопантетеинилового плеча (фиг.3h*), расположенное в SacB, для продолжения синтеза на основном каркасе сафрацинового тетрапептида. В альтернативном механизме биосинтеза дипептид Ala-Gly может непосредственно вводиться в SacA. В этом случае дипептид может был образован в результате активности представителей семейства высокоактивных пептидилтрансферазных рибозимов (Sun et al., Chem. and Biol. 2002, 9, 619-626) или в результате активности ферментов бактериального протеолиза.
И, в-четвертых, хотя в случае большинства прокариотических пептидсинтетаз фосфоэстеразный фрагмент, который, скорее всего, ответственен за высвобождение зрелой пептидной цепи из ферментного комплекса, сливается с С-терминальным концом последнего модуля связывания аминокислоты (Marahiel et al, Chem. Rev. 1997, 97, 2651-2673), в случае сафрациновых синтетаз ТЕ домен отсутствует. Вероятно, в пути синтеза сафрацина после прохождения последней стадии элонгации тетрапептид может высвобождаться по альтернативному подходу осуществления терминации синтеза пептидной цепи, что также происходит при синтезе сафрамицина (Pospiech et al., Microbiol. 1996, 142, 741-746). В рамках данного конкретного подхода процесс терминации катализируется доменом редуктазы на карбокситерминальном конце SacC пептидсинтетазы, которая катализирует восстановительное расщепление связанной ацильной группы, присоединенной к Т-домену, с высвобождением линейного альдегида.
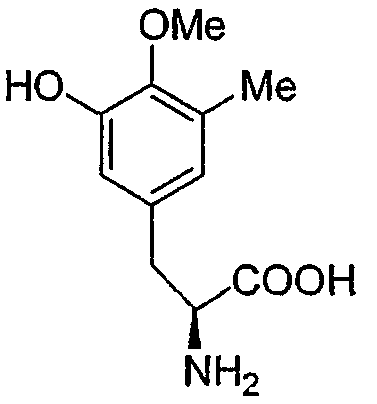
Перекрестные эксперименты авторов показали, что последние две аминокислоты, введенные в сафрациновую молекулу, представляют собой два производных L-Tyr, называемых Р2 (3-гидрокси-5-метил-О-метилтирозин) (фиг.4, 5), вместо двух L-Tyr, как предполагалось в случае синтеза сафрамицина. Во-первых, продукты двух генов (sacF и sacG), аналогичные бактериальным метилтрансферазам, как было показано, участвуют в O-, C-метилировании L-Tyr с образованием P14 (3-метил-О-метилтирозин), предшественника Р2. Возможный механизм может быть основан на том, что сначала происходит О-метилирование, а затем осуществляется С-метилирование аминокислотного производного. Во-вторых, Р2, субстрат для пептидсинтетаз SacB и SacC образуется при гидроксилировании P14 под действием SacD (фиг.4, 5):
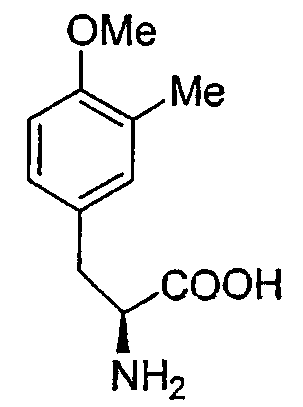
Точная масса: 209,11
Мол. масса: 209,24,
С - 63,14; H - 7,23; N - 6,69;
O - 22,94
Точная масса: 225,10
Мол. масса: 225,24,
С - 58,66; H - 6,71; N - 6,22;
O - 28,41
Кроме генов, участвующих в биосинтезе сафрацина, в опероне sacABCDЕFGH были также обнаружены два гена sacE и sacH с неизвестной функцией и вовлеченные в механизм устойчивости к сафрацину соответственно. Авторы показали, что sacH ген кодирует белок, который в случае гетерологичной экспрессии в других штаммах Pseudomonas генерирует выраженное повышение резистентности к сафрацину B. Предполагают, что SacH является трансмембранным доменом, который трансформирует C21-OH группу сафрацина B в C21-H группу с образованием сафрацина A, соединения с меньшей антибиотической и противоопухолевой активностью. И наконец, даже несмотря на то, что все еще неизвестны возможные функции SacE, было обнаружено, что гомологи этого гена близки к различным вторичным метаболитам в биосинтетической цепи генного кластера в геномах некоторых микроорганизмов, что позволяет предположить наличие консервативной функции данных генов в образовании или регуляции вторичных метаболитов.
В опероне sacIJ установленные аминокислотные последовательности, кодируемые sacI и sacJ, в высокой степени сходны с генными продуктами метилтрансферазы и гидроксилазы/монооксигеназы соответственно. Данные, полученные авторами, показывают, что SacI представляет собой фермент, ответственный за N-метилирование, присутствующее в сафрациновой структуре, и что SacJ представляет собой белок, который дополнительно гидроксилирует на одном из производных L-Tyr в тетрапептиде, с образованием хиноновой структуры, присутствующей во всех сафрациновых молекулах. N-метилирование представляет собой одну из модификаций пептидов, синтезируемых по нерибосомальному механизму, которые вносят значительный вклад в их биологическую активность. За исключением сафрамицина (Pospiech et al., Microbiol. 1996, 142, 741-746), который продуцируют бактерии и который N-метилируется, все известные N-метилированные нерибосомальные белки продуцируются грибами или актиномицетами и в большинстве случаев за такое N-метилирование отвечает домен, который находится в нерибосомальной пептидсинтетазе (см. табл.I).
лоты
На фиг.5 показан вероятный путь синтеза сафрацина, где указаны специфические аминокислотные субстраты, использующиеся в каждой реакции конденсации, и различные ферменты, активность которых необходима после конденсации.
Для дальнейшей оценки роли генов биосинтеза сафрацина авторами были сконструированы мутанты с выключением каждого из генов в кластере сафрацина (фиг.6). Разрушение генов NRPS (sacA, sacB и sacC), а также sacD, sacF и sacG, приводит к получению мутантов, не продуцирующих сафрацин и Р2. Полученные авторами результаты указывают на то, что гены из sacА-sacH являются частью одного и того же генетического оперона. В результате разрушения генов sacI и sacJ образуются три новые молекулы: P19B, Р22A и P22B (фиг.6):
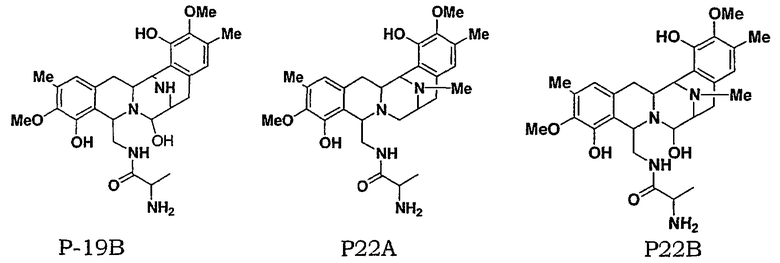
Продукция P22A и P22B (фиг.7а*) sacJ мутантом указывает на то, что роль SacJ заключается в осуществлении дополнительного гидроксилирования в левых производных L-Tyr аминокислоты сафрацина, вовлеченных в хиноновое кольцо. Образование sacI мутантом Р19В (фиг.7 b*) молекулы, подобной сафрацину, в которой отсутствует N-метилирование и хиноновое кольцо, подтверждает тот факт, что SacI является N-метилтрансферазой и дает основание полагать, что sacIJ является опероном, участвующим в транскрипции. Образование Р19В мутантом sacJ (фиг.7а*) также дает основание предположить, что N-метилирование, вероятно, происходит после образования хинонового кольца. Даже несмотря на то, что указанные новые структуры не обладают интересующей антимикробной активностью в отношении B. subtilis или не оказывают высокого цитотоксического воздействия на раковые клетки, они могут служить в качестве новых предшественников для полусинтеза новых активных молекул. Что касается структурной активности, наблюдение что Р19В, Р22А и Р22В теряют свою активность, позволяет полагать, что отсутствие хинонового кольца в сафрациновой структуре непосредственно связано с потерей активности молекул семейства сафрацинов.
Разрушение sacl гена с последующим восстановлением экспрессии гена sacJ приводит к образованию двух новых сафрацинов. Два полученные антибиотика с уровнями продукции такими же высокими, как и в случае продуцирования сафрацина А/сафрацина В в штамме дикого типа, были названы сафрацин D и сафрацин Е (фиг.7 с*):
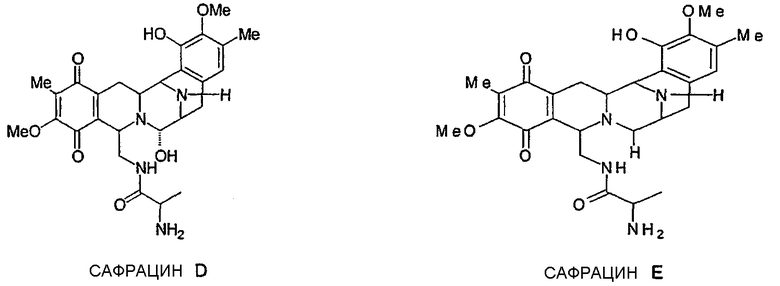
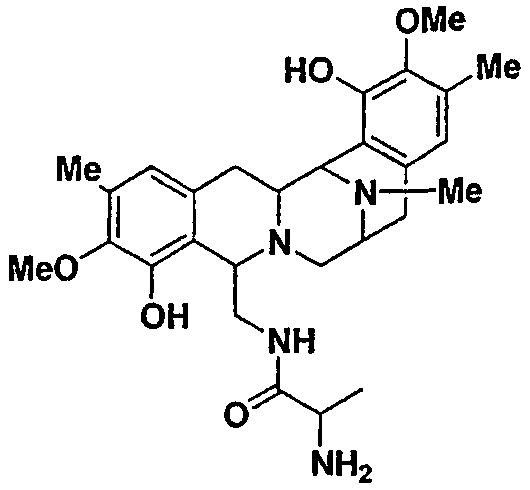
Сафрацин D и сафрацин Е представляют собой молекулы, подобные сафрацину В и сафрацину А соответственно, в которых отсутствует N-метилирование. Было показано, что и сафрацин D и сафрацин Е обладают такой же антибактериальной и противоопухолевой активностью, что и сафрацин В и сафрацин А соответственно. Сафрацин D, кроме высокой антибактериальной и противоопухолевой активности, обладает свойствами, позволяющими использовать его в полусинтезе эктеинасцидина ЕТ-789, мощного противоопухолевого средства, а также в полусинтезе новых эктеинасцидинов.
Возникает вопрос, касающийся роли белка, подобного аминопептидазе, кодируемого геном, расположенным в 3-сайте сафрацинового оперона. Инактивация путем встраивания orfl (PM-S1-14) не влияет на продукцию сафрацина А/сафрацина В. Что касается функциональных свойств, остается неясным вопрос, может ли данный белок выполнять какую-либо роль в метаболизме сафрацина. Должны быть исследованы более детально другие гены, присутствующие в космиде pL30P (orf2-orf4).
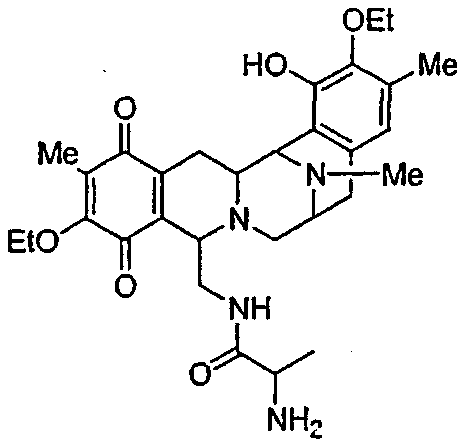
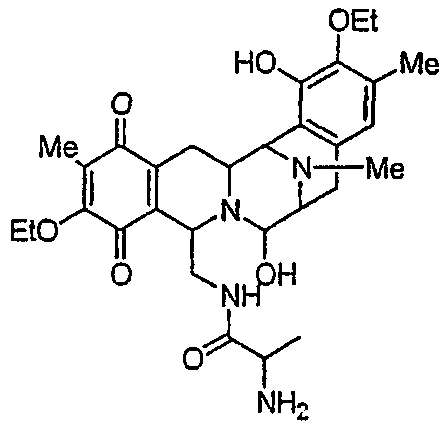
В другом аспекте настоящее изобретение относится к необходимым инструментам для получения новых специфически сконструированных "неприродных" молекул. Добавление специфически модифицированного предшественника производного Р2, обозначенного как Р3, 3-гидрокси-5-метил-O-этилтирозина, к мутанту sacF приводит к образованию двух "неприродных" сафрацинов, которые включают данный специфически модифицированный предшественник, сафрацин A(OEt) и сафрацин B(OEt) (фиг.8):
Точная масса: 552,29
Мол. масса: 552,66, С - 65,20; H - 7,30; N - 10,14; O - 17,37
Точная масса: 568,29
Мол. масса: 568, 66; С - 63,36; H - 7,09; N - 9,85; O - 19,69
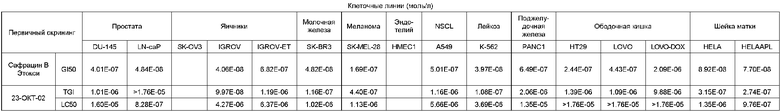
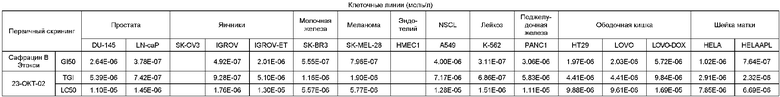
Два новых сафрацина являются мощными антибиотическими и противоопухолевыми соединениями. Биологическая активность сафрацина A(OEt) и сафрацина B(OЕt) характеризуются такой же эффективностью, что и активность сафрацина А и сафрацина В соответственно. Указанные новые сафрацины могут стать источником новых мощных противоопухолевых средств, а также источником молекул для полусинтеза новых эктеинасцидинов.
Кроме того, гены, участвующие в синтезе сафрацина, могут быть объединены с другими генами нерибосомальных пептидсинтетаз с получением в результате новых "неприродных" средств и аналогов с улучшенной активностью.
ПРИМЕРЫ
Пример 1: Экстракция молекул нуклеиновой кислоты из Pseudomonas fluorescens A2-2
Бактериальные штаммы
Штаммы Pseudomonas sp. выращивают при 27°С в бульоне Луриа-Бертани (Luria-Bertani (LB)) (Ausubel et al., 1995, J. Wiley and Sons, New York, N.Y). Штаммы E. coli выращивают при 37°С в среде LB. Антибиотики используют в следующих концентрациях: ампициллин (50 мкг/мл), тетрациклин (20 мкг/мл) и канамицин (50 мкг/мл) (см. табл.II).
Манипуляции с ДНК
Если не указано другого, то используют стандартные методики молекулярной биологии для манипуляций ДНК in vitro и клонирования (Sambrook et al., 1989, Cold Spring Harbor, NY: Cold Spring Harbor Laboratory).
Экстракция ДНК
Общую ДНК из культур Pseudomonas fluorescens A2-2 получают по опубликованным методикам (Sambrook et al., 1989, Cold Spring Harbor, NY: Cold Spring Harbor Laboratory).
Компьютерный анализ
Данные последовательностей объединяют и анализируют их с помощью пакета программ DNA-Star.
Пример 2: Идентификация генов NRPS, ответственных за образование сафрацина в Pseudomonas fluorescens A2-2
Создание праймера
Ранее Марахил с соавт. (Marahiel et al., Chem. Rev. 1997, 97, 2651-2673) опубликовали данные о высококонсервативных коровых мотивах в каталитических доменах циклических и разветвленных пептидсинтетаз. На основании результатов множественных сравнений последовательностей для ряда описанных пептидсинтетаз были использованы консервативные участки пептидсинтетаз А2, А3, А5, А6, А7 и А8 модулей аденилирования и модулей Т тиолирования в качестве мишени для создания вырожденного праймера (Turgay and А2, Мarahiel, Рeptide Res. 1994, 7, 238-241). В выбранные модулы были введены предпочтительные в соответствии с вырожденностью нуклеотиды и получен Pseudomonas sp. с ожидаемым высоким содержанием G/C. Все олигонуклеотиды получают от компании ISOGEN (Bioscience BV). При использовании вырожденных олигонуклеотидов получен ПЦР фрагмент, созданный на основе последовательностей YGPTЕ (A5 ядро) и LGGXS (T ядро). Указанные олигонуклеотиды были обозначены как PS34-YG и PS6-FF соответственно.
Условия ПЦР для амплификации ДНК из Pseudomonas fluorescens A2-2
Фрагмент внутреннего участка нерибосомальных пептидсинтетаз (NRPS) амплифицируют с использованием олигонуклеотидов PS-34-YG и PS6-FF и хромосомной ДНК из P. fluorescens A2-2 в качестве матрицы. Используют реакционный буфер и полимеразу Taq от компании Promega. Циклы осуществляют в Personal thermocycler (Eppendorf), которая включает 30 циклов по 1 минуте при 95°С, 1 минуту при 50°C и 2 минуты при 72°C. ПЦР продукты имеют ожидаемый размер (примерно 750 п.н.), который был рассчитан на основе информации о расположении праймеров внутри NRPS доменов других генов синтетазы.
Клонирование ДНК
Фрагменты, амплифицированные ПЦР, клонируют в векторе pGEM-Teasy по методике производителя (Qiagen, Inc., Valencia, CA). Клонированные таким способом фрагменты фланкируют двумя сайтами рестрикции EcoRI, с тем чтобы облегчить последующее клонирование в других плазмидах (см. ниже). Поскольку NRPS ферменты являются модулярными, клоны из вырожденных ПЦР праймеров отражают пул фрагментов из разных доменов.
Секвенирование ДНК
Все анализы секвенирования проводят с помощью праймеров, направленных против вектора клонирования, с использованием автоматизированного секвенатора ABI (Perkin-Elmer). Клонированные последовательности ДНК идентифицируют с использованием сервера BLAST Национального Центра биотехнологической информации (National Center for Biotechnology Information), доступного через интернет (Altsсhul et al., Nucleic Acids Res. 1997, 25, 3389-3521). Все последовательности имеют сигнальные участки для NRPS и демонстрируют высокое сходство по результатам поисков в системе BLAST с бактериальными NRPS, указывая на то, что они фактически имеют пептидное происхождение. Кроме того, поиски возможного сходства по доменам проводят с использованием вэб-сервера PROSITE (European Molecular Biology Laboratory, Heidelberg, Germany).
Разрушение гена в Pseudomonas fluorescens A2-2
Для анализа функции клонированных генов данные гены разрушают путем гомологичной рекомбинации (фиг.9). Для данной цели рекомбинантные плазмиды (производные pG-PS), несущие генный фрагмент NRPS, расщепляют рестрикционным ферментом EcoRI. Полученные фрагменты, принадлежащие гену, который будет мутирован, клонируют в мобилизуемой плазмиде pK18mob (Schafer et al., Gene 1994, 145, 69-73), которая представляет собой интегрируемую в хромосому плазмиду, способную реплицироваться в штаммах E. coli, и не может реплицироваться в Pseudomonas. Рекомбинантную плазмиду вводят сначала в штамм E. coli S17-λPIR путем трансформации, а затем в P. fluorescens A2-2 путем конъюгации двух родительских вариантов (Herrero et al., J. Bacteriol 1990, 172, 6557-6567). Различные разведения конъюгата помещают на твердую среду LB, содержащую ампициллин и канамицин, и инкубируют в течение ночи при 27°C. Отбирают канамицин-резистентные трансконъюгаты, содержащие плазмиды, интегрированные в геном путем гомологичной рекомбинации.
Биологический анализ (биотест) образования сафрацина
Штаммы P. fluorescens A2-2 и его производные инкубируют в эрленмейровских колбах с дефлектором на 50 мл, содержащих ферментационную среду с соответствующими антибиотиками. Вначале используют ферментационную среду SA3 (Ikeda Y.J. Ferment. Technol. 1985, 63, 283-286). Для повышения продуктивности процесса ферментации используют статистико-математические методы, такие как метод Плакетта-Бурмана (Plackett-Burman) для отбора питательных компонентов, а также методы оптимизации поверхностной ответной реакции (Hendrix C. Chemtech 1980, 10, 488-497) для определения оптимального уровня каждой ключевой независимой переменной. Проводят также эксперименты, направленные на улучшение условий культивирования, таких как температуре инкубации и скорость перемешивания. И наконец, выбирают высокопродуктивную по сафрацину В среду, названную 16В (152 г/л маннита, 35 г/л G20-25 дрожжей, 26 г/л CaCO3, 14 г/л сульфата аммония, 0,18 г/л хлорида железа, pH 6,5).
Продукцию сафрацина анализируют путем определения способности к ингибированию твердой культуры Bacillus subtilis 10 мкл супернатанта 3-дневной культуры Pseudomonas sp., инкубированной при 27°С (Alijah et al., Appl Microbiol Biotechnol 1991, 34, 749-755). Культуры P. fluorescens A2-2 образуют зоны ингибирования диаметром 10-14 мм, тогда как непродуцирующие мутанты не ингибируют рост B. subtilis. Три выделенных клона содержат поврежденные пути биосинтеза сафрацина. Для подтверждения полученных результатов был проведен анализ уровня образованных сафрацинов методом ВЭЖХ.
Анализ образования сафрацина методом ВЭЖХ
Супернатант анализируют с использованием ВЭЖХ Symmetry C-18, 300Å, колонка 5 мкм, 250 х 4,6 мм (Waters) с защитной рубашкой (Symmetry C-18, колонка 5 мкм, 3,9 х 20 мм, Waters). В качестве подвижной фазы используют градиент аммоний-ацетатного буфера (10 мМ, 1% диэтаноламин, pH 4,0) - ацетонитрила. Сафрацин выявляют по поглощению при длине волны 268 нм. На фиг.6 показаны ВЭЖХ-профили сафрацина и предшественников сафрацина, продуцируемых штаммом Pseudomonas fluorescens A2-2, и различных сафрацин-подобных структур, продуцируемых мутантами P. fluorescens.
Пример 3. Клонирование и анализ последовательности сафрацинового кластера
Обратная ПЦР и гибридизация с фаговой библиотекой
Саузерн-гибридизация с использованием хромосомных ДНК мутантов подтвердила корректность разрушения гена и показала, что фрагмент пептидсинтетазы, клонированный в плазмиде pK18mob, необходим для продукции сафрацина. Анализ полученных не продуцирующих сафрацин мутантов показал, что все они включают разрушение гена в одном и том же гене sacA.
Обратная ПЦР на основе геномной ДНК и скрининг фаговой библиотеки с использованием геномной ДНК P. fluorescens A2-2 выявил наличие дополнительных генов, фланкирующих ген sacA, которые, по всей вероятности, участвуют в биосинтезе сафрацина.
Номер доступа в базе данных GenBank к информации по нуклеотидной последовательности кластера, связанного с биосинтезом сафрацина в P. fluorescens A2-2, AY061859.
Конструирование космидной библиотеки и гетерологичная эксперссия
Для выявления, может ли сафрациновый кластер придавать способность к биосинтезу сафрацина непродуцирующему штамму, указанный кластер был клонирован в широком диапазоне космидных векторов (pLAFR3, Staskawicz B, et al., J. Bacteriol 1987, 169, 5789-5794) и конъюгирован с разными штаммами из коллекции Pseudomonas sp.
Для получения клона, содержащего целый кластер, конструируют и подвергают скринингу космидную библиотеку. Для данной цели хромосомную ДНК частично разрушают рестрикционным ферментом PstI, фрагменты подвергают дефосфорилированию и лигируют в PstI сайте космидного вектора pLAFR3. Космиды упаковывают с использованием экстрактов, способствующих упаковке (Gigapack III gold) (Stratagene), в соответствии с рекомендациями производителя. Инфицированные клетки штамма XL1-Blue помещают на LB агар, содержащий в качестве добавки 50 мкг/мл тетрациклина. Отбирают положительные клоны по методу гидридизации колоний с DIG-меченым фрагментом ДНК, принадлежащим к 3'-концу сафрацинового кластера. Для гарантии клонирования всего кластера проводят новую гибридизацию колоний с 5'-концевым фрагментом ДНК. Только космида pL30р демонстрирует множественные гибридизации с зондами ДНК. Для подтверждения точности клонирования проводят ПЦР-амплификацию и секвенирование ДНК с использованием олигонуклеотидов ДНК, принадлежащих к сафрациновой последовательности. Размер вставки в pL30Р составляет 26705 п.н. pL30p клон ДНК трансформируют в Е. coli S17λPIR и полученный штамм конъюгируют с гетерологичными штаммами Pseudomonas sp. Космиду pL30p вводят в P. fluorescens CECT378 и P. aeruginosa СЕСТ110 путем конъюгации двух родительских вариантов, как было описано выше. После идентификации клона, кодирующего целый кластер, определяют, способен ли данный потенциальный штамм продуцировать сафрацин. Продукцию сафрацина в конъюгированных штаммах оценивают анализом метода ВЭЖХ и с помощью биологического теста, в котором используют супернатанты бульонных культур, как было описано ранее.
Штамм P. fluorescens CECT378, экспрессирующий космиду pL30p (PM-19-002), способен продуцировать сафрацин в значительных количествах, тогда как образование сафрацина в штамме P. aeruginosa CECT110, экспрессирующем pL30Р (РМ-16-002), в 10 раз меньше, чем в CECT378. Продукция сафрацина в указанных штаммам составляет примерно 22% и 2% от общей продукции в сравнении с природным штаммом-продуцентом.
Гены, участвующие в образовании сафрацина. Анализ последовательностей оперонов sacABCDEFGH и sacIJ
Компьютерный анализ последовательности ДНК pL30Р выявил 14 ОРС (фиг.1). Сайт возможного связывания с рибосомом предшествует каждому из ATG старт-кодонов.
В опероне sacABCDEFGH три очень больших ОРС: sacA, sacB и sacC (положения 3052-6063, 6080-9268, и 9275-13570 в последовательности сафрацина P. fluorescens A2-2 SEQ ID No. 1 соответственно) могут быть считаны в том же направлении и кодируют предположительные NRPS сафрацина: SacA (1004 аминокислот, Mr 110452), sacB (1063 аминокислот, Mr 117539) и sacC (1432 аминокислот, Mr 157331). Три гена NRPS содержат домены, соответствующие активирующим аминокислоту доменам известных пептидсинтетаз. Конкретно домен, активирующий аминокислоту, из указанных генов NRPS очень близок к трем из четырех доменов, активирующих аминокислоту (Gly, Tyr и Tyr), найденных в NRPS сафрацина в Myхococcus xanthus (Pospiеch et al., Microbiology 1995, 141, 1793-803; Pospiеch et al., Microbiol. 1996, 142, 741-746). В частности, SacA (SEQ ID No: 2) демонстрирует 33% идентичность с белком синтетазы В сафрамицина Mx1 B (SafB) из M. xanthus (номер доступа в NCBI U24657), тогда как SacB (SEQ ID No: 3) и SacC (SEQ ID No: 4) характеризуются 39% и 41% идентичности соответственно, с синтетазой А сафрамицина Mx1 (SafA) из M. хanthus (номер доступа в NCBI U24657). На фиг.2 показаны в сравнительном аспекте SacA, SacB и SacC, а также различные домены, активирующих аминокислоту, и NRPS сафрамицина.
Имеет место считывание пяти малых ОРС по направлению транскрипции sacC, как и генов NRPS (фиг.1). Первая из них sacD (положение 13602-14651 в сафрациновой последовательности P. fluorescens A2-2) предположительно кодирует белок SacD (350 аминокислот, Mr 39187, SEQ ID No:5), при этом не выявляется сходных вариантов в GeneBank DB. Следующая sacЕ (положение 14719-14901 в сафрациновой последовательности P. fluorescens A2-2) кодирует предположительно малый белок SacЕ (61 аминокислота, Mr 6729, SEQ ID No: 6), который демонстрирует некоторое сходство с имеющими в базах данных белками неизвестной функции (ОРС 1 из Streptomyces viridоchromоgenes (номер доступа в NCBI Y17268; 44% идентичности) и MbtH из Mycobacretium tuberculosis (номер доступа в NCBI Z95208; 36% идентичности). Третья ОРС sacF (положение 14962-16026 в сафрациновой последовательности P. fluorescens A2-2) кодирует белок из 355 остатков с расчетной молекулярной массой 39834 (SEQ ID No: 7). Данный белок наиболее близок к метилтрансферазе гидроксинейроспорина (CrtF) из Chloroflexus aurantiacus (номер доступа в NCBI AF288602; 25% идентичности). Нуклеотидная последовательность четвертой ОРС sacG (положение 16115-17155 в сафрациновой последовательности P. fluorescens A2-2) соответствует предсказанному генному продукту из 347 аминокислот с молекулярной массой 3822 кДА (SEQ ID No: 8). Белок, названный SacG, близок к бактериальным О-метилтрансферазам, включая О-диметилпуромицин-О-метилтрансферазу (DmpM) из Streptomyces anulatus (номер доступа в NCBI Р42712; 31% идентичности). Компьютерный поиск также показывает, что данный белок содержит три мотива последовательности, обнаруженные в различных S-аденозилметионин-зависимых метилтрансферазах (Kagan and Clarke, Arch. Biochem. Biophys. 1994, 310, 417-427). Пятый ген, SacH (положение 17244-17783 в сафрациновой последовательности P. fluorescens A2-2) кодирует предположительный белок SacH (180 аминокислот; Mr 19632; SEQ ID No: 9). Компьютерный поиск сходства между расчетной аминокислотной последовательностью SacH и другими белковыми последовательностями выявляет идентичность с некоторыми консервативными гипотетическими белками неизвестной функции, которые содержат достаточно консервативный трансмембранный мотив и сайт активности типа дигидрофолятредуктазы (консервативный гипотетический белок Pseudomonas aeruginosa PAO1, номер доступа в NCBI Р3469; 35% идентичности).
Против хода считывания оперона sacABCDEFGH расположены два гена sacIJ в антисмысловом направлении. Ген sacI (положение 2513-1854) кодирует белок из 220 аминокислот (Mr 24219; SEQ ID No: 10), который ближе всего к метилтрансферазе убихинона/манехинона из Thermotoga maritime (номер доступа в NCBI AE001745; 32% идентичности). Ген sacJ (положение 1861-335) кодирует белок из 509 аминокислот (SEQ ID No: 11) c молекулярной массой 55341 Да, который аналогичен бактериальным монооксигеназам/гидроксилазам, включая предположительно монооксигеназу из Bacillus subtilis (номер доступа в NCBI Y14081; 33% идентичности) и Streptomyces coelicolor (номер доступа в NCBI AL109972; 29% идентичности).
Опероны sacABCDEFGH и sacIJ транскрибируются по-разному и разделены участком примерно 450 п.н. Оба оперона фланкированы оставшимися транспортируемыми фрагментами.
Гены, родственные генам сафрацинового кластера
Была обнаружена предположительная ОРС (orf1; положение 18322-19365 в сафрациновой последовательности P. fluorescens A2-2), расположенная на 3'-конце сафрациновой последовательности (фиг.1). Белок ORF1 (SEQ ID No: 12) демонстрирует сходство с аминопептидазами из базы данных Gene Bank (пептидаза семейства М20/M25/M40 из Сaulobacter crescentus CB15; номер доступа в NCBI NP422131; 30% идентичности). С использованием стратегии, описанной в примере 2, показано, что разрушение гена orf1 не влияет на продукцию сафрацина в P. fluorescens А2-2.
Сафрациновую последовательность клонируют по 3'-концу в космиде pL30p и выявляют три предположительные ОРС (orf2, orf3 и orf4). Считывание гена orf2 в направлении, противоположном направлению считывания оперона sacABCDEFGH (положение 22885 - 21169, SEQ ID No: 1), дает белок ORF2 (SEQ ID No: 13), имеющий сходство с сенсорным белком HoxX в Aquifex aeolicus (номер доступа в NCBI NC000918.1; 35% идентичности), тогда как ген orf3 (положение 23730-23041 в SEQ ID No: 1) кодирует белок ORF3 (SEQ ID No: 14), который имеет 44% идентичности с белком, близким к гликозилтрансферазе Xanthomonas axonopodis pv. Citri str. 306 (номер доступа в NCBI NP642442).
Третий ген расположен на 3'-конце SEQ ID No: 1 (положение 25037-26095). Данный ген, названный orf4 (положение 2513-1854), кодирует белок ОRF4 (SEQ ID No: 15), который больше всего напоминает гипотетический белок Escherichia coli из семейства изохоризматазы YcdL (номер доступа в NCBI P75897; 32% идентичности).
Указанные три гена, скорее всего, не участвуют в пути биосинтеза сафрацина, однако только дальнейшее разрушение указанных генов сможет подтвердить данное предположение.
Различные найденные последовательности ДНК перечислены в конце описания.
Пример 4. Функциональный анализ локуса сафрацина и поиски возможных предшественников
Поскольку в настоящее время неизвестен путь синтеза сафрацина в P. fluorescens А2-2, инактивация каждого из генов, описанных в примере 3, будет представлять собой фундаментальные исследования механизма биосинтеза сафрацина в данном штамме.
Для анализа функциональных свойств каждого конкретного белка в пути продукции сафрацина проводят разрушение каждого конкретного гена в кластере, за исключением sacЕ. Все генетические мутанты, полученные по данной стратегии разрушения, были ранее описаны.
Фиг.6 представляет собой краткое описание различных мутантов, сконструированных в рамках настоящего изобретения, а также краткое описание соединений, получаемых с использованием мутантов, содержащих разрушенный ген. В штамме дикого типа методами ВЭЖХ были четко обнаружены и сафрацин А, и сафрацин В, а также другие соединения, Р2 и Р14 (см. фиг.6, WT). Разрушение гена sacA (PM-S1-002), sacB (PM-S1-003), sacC (PM-S1-004), sacD (PM-S1-010), sacF (PM-S1-008) и sacG (PM-S1-009) приводит к получению генных мутантов, которые не способны продуцировать ни сафрацин А, ни сафрацин В, ни соединения-предшественники, и имеют время удержания менее 15 минут, Р2 и Р4 соответственно. Изучение структуры Р2 и Р14 показывает, что Р14 представляет собой 3-метил-О-метилтирозин, тогда как Р2 представляет собой 3-гидрокси-5-метил-О-метилтирозин. В связи с малым размером гена sacЕ такой sacЕ мутант невозможно получить путем генного разрушения, но в настоящее время продолжаются попытки получения делеции по данному гену. Сверхэкспрессия белка SacЕ в транс-положении не оказала эффекта на продукцию сафрацина В/А. Мутанты sacI (PM-S1-006) продуцируют Р2, Р14 и значительные количества соединения, названного Р19В (фиг.6, фиг.7b*). Изучение структуры Р19В показывает, что данное соединение является сафрацин-подобной молекулой, в которой N-Met и одна из ОН групп хинонового кольца отсутствуют. В мутантах sacJ (PM-S1-005) были получены P2, P14, P19B и два новых соединения, названных Р22А и Р22В (фиг.6, фиг.7а*).
Изучение структуры Р22А и Р22В показало, что они представляют собой молекулы, близкие к сафрацину А и сафрацину В соответственно, но без одной -ОН группы в хиноновом кольце. Биологическое тестирование экстрактов мутантов sacI и sacJ выявляет очень низкую активность против Bacillus subtilis.
Разрушение sacI гена путем восстановления экспрессии гена sacJ приводит к получению нового мутанта, продуцирующего сафрацины, PM-S1-007. Оба антибиотика, образуемых на таком же высоком уровне, как и сафрацин А и сафрацин В в штамме дикого типа, были обозначены как сафрацин D и сафрацин Е (фиг.7с*). Cафрацин D и сафрацин E представляют собой молекулы сафрацина B и сафрацина A соответственно, в которых отсутствует N-метилирование.
Указанные результаты позволяют в значительной мере полагать, что i) гены sacA, sacB и sacC кодируют NRPS сафрацина; ii) гены sacD, sacF и sacG ответственны за трансформацию L-Tyr в L-Tyr-производное Р2 и iii) sacI и sacJ ответственны за «хвостовые» модификации, которые позволяют превратить Р19 и Р22 в сафрацин.
Характеристика природных предшественников
Р-14
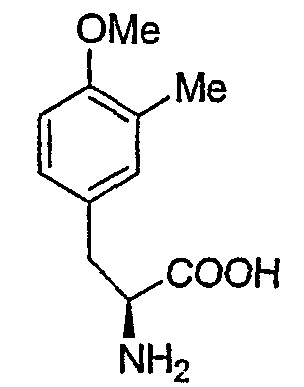
Точная масса: 209,11, Мол. масса: 209,24
С - 63,14; H - 7,23; N - 6,69; O - 22,94
Штамм:
Pseudomonas fluorescens А2-2 (дикий тип) (PM-S1-001)
Условия ферментации:
В посевную среду YMP3, содержащую 1% глюкозы, 0,25% мясного экстракта, 0,5% бактопептона, 0,25% NaCl, 0,8% CaCO3, инокулируют 0,1% замороженную вегетативную культуру микроорганизмов и инкубируют на роторной качалке (250 об/мин) при 27°C. После 30 часов инкубации 2% об/об посевной культуры переносят в эрленмейеровские колбы на 2000 мл, содержащие 250 мл продуцирующей среды М-16В, которая состоит из 15,2% маннита, 3,5% сухих пекарских дрожжей, 1,4% (NH4)2SO4, 0,001% FeCl3, 2,6% СаCO3. Температура инкубации составляет 27°С в момент инокуляции до окончания 40 часов инкубации и затем 24°С до окончания процесса (71 час). Значение pH не контролируют. Скорость перемешивания на роторной качалке составляет 220 об/мин с эксцентриситетом 5 см.
Выделение:
После 71 часов инкубации объединяют содержимое двух эрленмейеровских колб и 500 мл ферментационного бульона осветляют центрифугированием при 7500 об/мин в течение 15 минут. К супернатанту добавляют 50 г смолы XAD-16 (Аmberlite) и перемешивают в течение 30 минут при комнатной температуре. Затем смолу восстанавливают из осветленного бульона фильтрованием. Далее смолу промывают два раза дистиллированной водой и экстрагируют 250 мл изопропанола (2-PrOH). Спиртовой экстракт сушат в высоком вакууме с получением 500 мг неочищенного экстракта. Данный неочищенный экстракт растворяют в метаноле и очищают на хроматографической колонке с использованием сефадекса LH-20 и метанола в качестве подвижной фазы. Соединение Р-14 элюируют и сушат, получая 15 мг желтоватого твердого вещества. Чистоту определяют с использованием метода аналитической ВЭЖХ и метода 1H-ЯМР.
Р-14 также выделяют аналогичным образом из культур мутанта sacJ (PM-S1-005) с использованием полупрепаративной ВЭЖХ на последней стадии процесса очистки.
Биологическая активность:
АКТИВНОСТЬ ОТСУТСТВУЕТ.
Спектроскопические данные:
ESMS m/z 254 (C11H14NO3Na2 +), 232 (C11H15NO3Na+), 210 (M+H+), 1Н ЯМР (300 МГц, CD3OD): 7,07 (д, J=8,1 Гц, H-9), 7,06 (с, H-5), 6,84 (д, J=8,1 Гц, H-8), 3,79 (с, H-11), 3,72 (дд, J=8,7, 3,9 Гц, H-2), 3,20 (дд, J=14,4, 3,9 Гц, H-3a), 2,91 (дд, J=14,4, 8,9 Гц, H-3b), 2,16 (с, H-10). 13С ЯМР (75 МГц, CD3OD): 174,1 (C-1), 158,6 (C-7), 132,5 (C-5), 128,9 (C-9), 128,5 (C-4), 128,0 (C-6), 111,4 (C-8), 57,6 (C-2), 55,8 (C-11), 37,4 (C-3), 16,3 (C-10).
Р-2
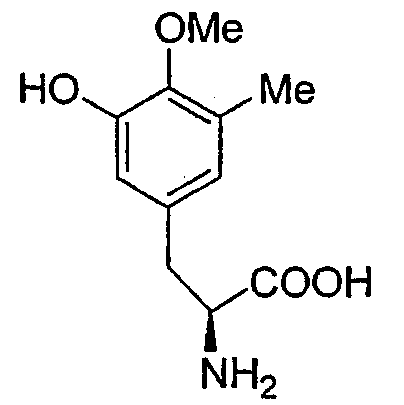
Точная масса: 225,10, Мол. масса: 225,24
С - 58,66; H - 6,71; N - 6,22; O - 28,41
Штамм:
Pseudomonas fluorescens А2-2 (дикий тип) (PM-S1-001).
Условия ферментации:
Тот же процесс, что и в случае Р-14.
Выделение:
Проводят процедуру, аналогичную использованной в случае Р-14, за исключением хроматографии на сефадексе, при которой позже элюируются фракции, содержащие Р-2. Для очистки Р-2 обязательна стадия полупрепаративной ВЭЖХ (колонка Symmetry Preр C-18, 7,8 x 150 мм, 10 мМ AcONH4 pH 3/СH3CN (95:5), в течение 5 минут и затем градиент 5-6,8% CH3CN в течение 3 минут).
Также выделяют соединения из ферментационного бульона Pseudomonas putida ATCC12633+pB5H83 (PM-17-004), полученного в результате гетерологичной экспрессии.
Биологическая активность:
АКТИВНОСТЬ ОТСУТСТВУЕТ.
Спектроскопические данные:
ESMS m/z 226 [M+H]+; 1Н ЯМР (CD3OD, 300 МГц): 6,65 (д, J=1,8 Гц, H-5), 6,59 (д, J=1,8 Гц, H-9), 3,72 (с, H-11), 3,71 (дд, J=9,0, 4,2 Гц, H-2), 3,16 (дд, J=14,4, 4,2 Гц, H-3a), 2,83 (дд, J=14,4, 9,0 Гц, H-3b), 2,22 (с, H-10); 13C ЯМР (ДMСO, 75 МГц): 170,88 (с, C-1), 150,025 (с, C-7), 144,56 (с, C-8), 132,28 (с, C-4), 130,36 (с, C-6), 121,73 (д, C-5), 115,55 (д, C-9), 59,06 (кв, 7-OMe), 55,40 (д, C-2), 36,21 (т, C-3), 15,86 (кв, 6-Me).
Характеристика сафрацин-подобных соединений, полученных путем выключения функции гена
Соединение Р-22В
Точная масса: 526,28
Мол. масса: 526,62
С - 63,86; H - 7,27; N - 10,64; O - 18,23
Штамм:
sacJ мутант Pseudomonas fluorescens А2-2 (PM-S1-005).
Условия ферментации:
В биореактор для выращивания культуры (Bioеngineering LP-351) с суммарной вместимостью 75 литров выливают 50 литров среды SAM-7 (50 л), состоящей из декстрозы (3,2%), маннита (9,6%), сухих пекарских дрожжей (2%), сульфата аммония (1,4%), вторичного фосфата калия (0,03%), хлорида калия (0,8%), 6-гидрата хлорида железа (III) (0,001%), L-тирозина (0,1%), карбоната кальция (0,8%), поли-(пропиленгликоль) 2000 (0,05%) и пеногасителя ASSAF 1000 (0,2%), и после стерилизации добавляют стерильные антибиотики (ампициллин 0,05 г/л и канамицин 0,05 г/л). Затем в среду инокулируют посевную культуру (2%) мутантного штамма PM-S1-005. Ферментацию проводят в течение 71 часа в условиях аэрации и перемешивания (1,0 л/л/мин и 500 об/мин). Температуру поддерживают в диапазоне от 27°С (с момента инокуляции в течение 24 часов) до 25°С (начиная с 24 часа до окончания процесса). Значение pH поддерживают на уровне 6,0 путем автоматического добавления разбавленной серной кислоты, начиная с 22 часа до окончания процесса.
Выделение:
Весь бульон осветляют (центрифуга Шарплс (Sharples)). pH осветленного бульона доводят до pH 9,0 путем добавления 10% NaOH и экстрагируют 25 литрами этилацетата. После 20 минут перемешивания обе фазы разделяют. Органическую фазу замораживают в течение ночи, затем фильтруют для удаления льда и выпаривают с получением жирного темно-зеленого экстракта (65,8 г).
Полученный экстракт смешивают с 500 мл гексана (добавляют два раза по 250 мл) и фильтруют для удаления растворимых в гексане примесей. Оставшееся твердое вещество после сушки дает 27,4 г сухого зелено-бежевого экстракта.
Новый полученный экстракт растворяют в метаноле и очищают хромотографией на сефадексе (Sephadex LH-20) (с использованием метанола в качестве подвижной фазы растворителя) и сафрацин-подобные вещества элюируют в центральных фракциях (Условия анализа в ТСХ: силикагель, нормальная фаза, подвижная фаза: EtОAc: MeOH 5:3. Прим. значение Rf: 0,3 для P-22B, 0,25 для P-22A and 0,1 для P-19).
Объединенные фракции (7,6 г), содержащие три сафрацин-подобных соединения, очищают на колонке с силикагелем с использованием смеси EtОAc: MeOH в градиенте от 50:1 до 0:1 и других хроматографических систем (изократическая система CHCl3:МеОН:H2O:АсОН, 50:45:5:0,1). Соединения Р22-А, Р22-В и Р19-В очищают методом ВЭЖХ с обращением фазы (колонка SummetryPrep С-18, 150×7,8 мм, 4 мл/мин, подвижная фаза: 5 мин, МеОН:H2O (0,02% ТФУ), 5:95 и градиент от МеОН:H2O (0,02% ТФУ) 5:95 до МеОН 100% в течение 30 минут).
Противомикробная активность: на твердой среде.
Bacillus subtilis: 10 мкг/диск (диаметр 6 мм): зона ингибирования 10 мм.
Спектроскопические данные:
HRFABMS m/z 509,275351 [М-H2O+Н]+(вычислено для: C28H37N4O5 509,276396 Δ 1,0 mmu); LRFABMS с использованием m-NBA в качестве матрицы m/z (относительная интенсивность) 509 [М-H2O+Н]+(5), 460 (2,7), 391 (3).
1H NMR (CD3OD, 500 МГц): 6,70 (с, Н-15), 6,52 (с, Н-5), 4,72 (ушир.с, Н-11), 4,66 (д, J=2,0 Гц, Н-21), 4,62 (дд, J=8,4, 3,7 Гц, Н-1), 3,98 (ушир.д, J=7,6 Гц, Н-13), 3,74 (с, 7-ОМе), 3,71 (с, 17-ОМе), 3,63 (м, перекрывающиеся сигналы, Н-25), 3,62 (м, перекрывающиеся сигналы, Н-3), 3,30 (м, Н-22а), 3,29 (м, Н-14а), 3,18 (д, J=18,6 Гц, Н-14b), 2,90 (м, Н-4а), 2,88 (м, Н-22b), 2,76 (с, 12-NMe), 2,30 (с, 16-Ме), 2,22 (м, Н-4b), 1,16 (д, J=7,4 Гц, Н-26);
13С ЯМР (CD3OD, 125 МГц): 170,75 (с, C-24), 149,24 (с, C-18), 147,54 (с, C-8), 145,95 (с, C-7), 145,82 (с, С17), 133,93 (с, C-16), 132,31 (с, C-9), 131,30 (с, C-6), 128,95 (с, C-20), 121,93 (д, C-15), 121,76 (д, C-5), 121,44 (с, C-10), 112,45 (с, C-19), 92,87 (д, C-21), 60,86 (кв, 7-OMe), 60,76 (кв, 17-OMe), 59,39 (д, C-11), 57,96 (д, C-13), 55,51 (д, C-1), 54,29 (д, C-3), 50,08 (д, C-25), 45,55 (т, C-22), 40,43 (кв, 12-NMe), 32,56 (т, C-4), 25,84 (т, C-14), 17,20 (кв, C-26), 16,00 (кв, 16-Me), 15,81 (кв, 6-Me).
СОЕДИНЕНИЕ Р-22А
Штамм:
Тот же, что и в случае Р-22В.
Условия ферментации:
Те же, что и в случае Р-22В.
Выделение:
То же, что и в случае Р-22В.
Биологическая активность сафрацина Р-22А
Противоопухолевая активность
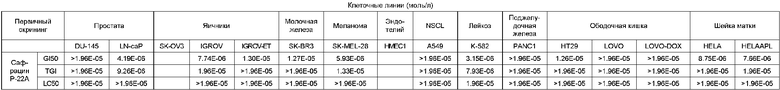
Противомикробная активность: на твердой среде.
Bacillus subtilis: 10 мкг/диск (диаметр 6 мм):
АКТИВНОСТЬ ОТСУТСТВУЕТ.
Спектроскопические данные:
HRFABMS m/z 511,290345 [М+Н]+ (вычислено для C28H39N4O5 511,292046 Δ 1,7 mmu); LRFABMS с использованием m-NBA в качестве матрицы m/z (относительная интенсивность) 511 [М+Н]+ (61), 409 (25), 391 (4); 1Н ЯМР (CD3OD, 500 МГц): 6,68 (с, Н-15), 6,44 (с, Н-5), 3,71 (с, 7-ОМе), 3,67 (с, 17-ОМе), 2,72 (с, 12-NMe), 2,28 (с, 16-Ме), 2,20 (с, 6-Ме), 0,87 (д, J=7,1 Гц, Н-26).
Соединение Р-19В
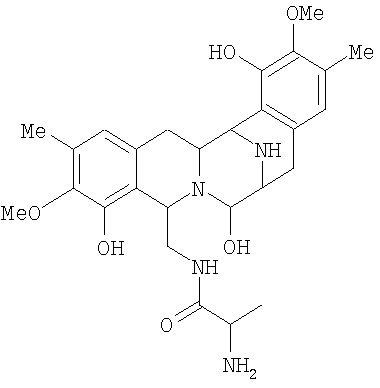
Штамм:
Тот же, что и в случае Р-22В.
Условия ферментации:
Те же, что и в случае Р-22В.
Выделение
То же, что и в случае Р-22В.
Биологическая активность сафрацина Р-19В
Противоопухолевая активность
Клеточные линии (моль/л)
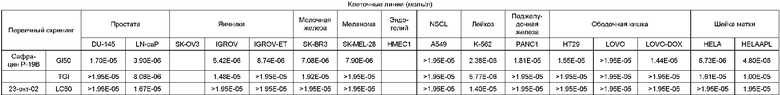
Противомикробная активность: на твердой среде.
Bacillus subtilis: 10 мкг/диск (диаметр 6 мм):
АКТИВНОСТЬ ОТСУТСТВУЕТ.
Спектроскопические данные:
HRFABMS m/z 495,260410 [М-H2O+Н]+ (вычислено для: C27H35N4O5 495,260746 Δ 0,3 mmu); LRFABMS с использованием m-NBA в качестве матрицы m/z (относительная интенсивность) 495 [М-Н2О+Н]+ (13), 460 (3), 391 (2); 1Н ЯМР (CD3OD, 500 МГц): 6,67 (с, Н-15), 6,5 (с, Н-5), 3,73 (с, 7-ОМе), 3,71 (с, 17-ОМе), 2,29 (с, 16-Ме), 2,24 (с, 6-Ме), 1,13 (д, J=7,1 Гц, Н-26).
Новые сафрациновые соединения, полученные путем выключения функции гена
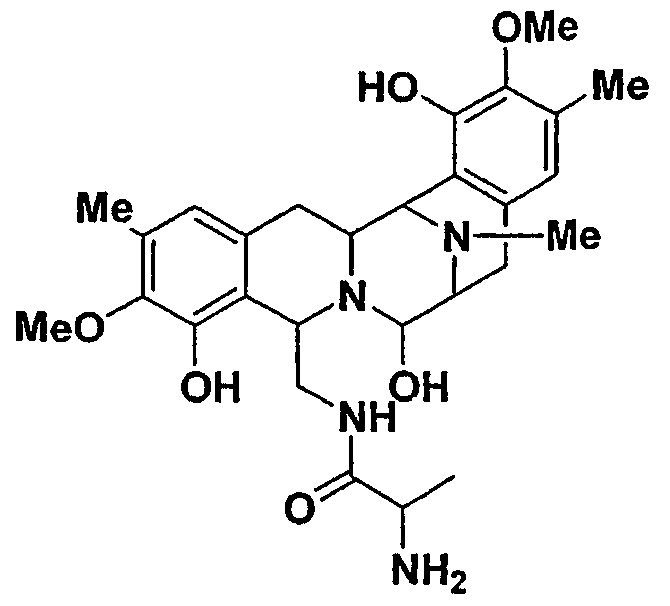
Сафрацин D
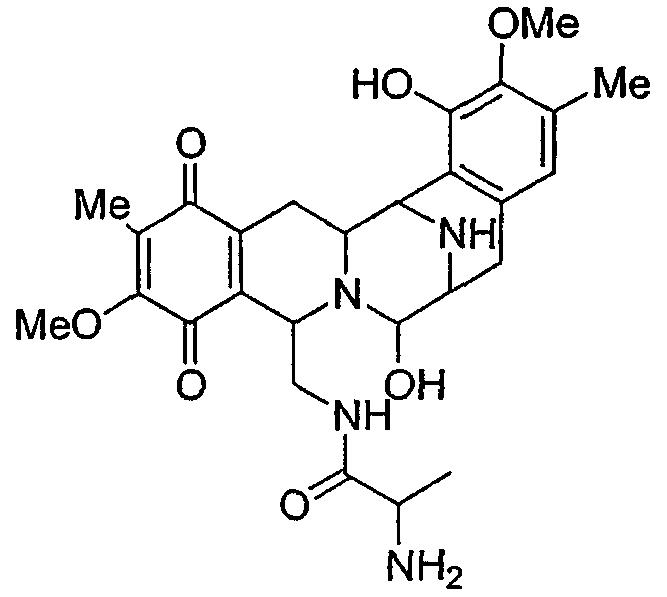
Точная масса: 526,24
Мол. масса: 526,58
С - 61,58; H - 6,51; N - 10,64; O - 21,27
Штамм:
sacI с восстановленной экспрессией sacJ из P. fluorescens А2-2 (PM-S1-007)
Условия ферментации:
В биореактор для выращивания культуры (Bioеngineering LP-351) с суммарной вместимостью 75 литров выливают 50 литров среды SAM-7 (50 л), состоящей из декстрозы (3,2%), маннита (9,6%), сухих пекарских дрожжей (2%), сульфата аммония (1,4%), вторичного фосфата калия (0,03%), хлорида калия (0,8%), 6-гидрата хлорида железа (III) (0,001%), L-тирозина (0,1%), карбоната кальция (0,8%), поли-(пропиленгликоль) 2000 (0,05%) и пеногасителя ASSAF 1000 (0,2%) и после стерилизации добавляют стерильные антибиотики (ампициллин 0,05 г/л и канамицин 0,05 г/л). Затем в среду инокулируют посевную культуру (2%) мутантного штамма PM-S1-007. Ферментацию проводят в течение 89 часа в условиях аэрации и перемешивания (1,0 л/л/мин и 500 об/мин). Температуру поддерживают в диапазоне от 27°С (с момента инокуляции в течение 24 часов) до 25°С (начиная с 24 часа до окончания процесса). Значение pH поддерживают на уровне 6,0 путем автоматического добавления разбавленной серной кислоты, начиная с 27 часа до окончания процесса.
Выделение:
Полученную таким образом культуральную среду (45 л) после удаления клеток центрифугированием доводят до pH 9,5 разбавленным гидроксидом натрия и экстрагируют дважды 25 литрами этилацетата. Смесь вводят в резервуар с перемешиванием при комнатной температуре на 20 минут. Две фазы разделяют путем центрифугирования по типу жидкость-жидкость. Органические фазы замораживают при -20°С, фильтруют для удаления льда и выпаривают до получения 35 г маслянистого темного неочищенного экстракта. После растирания с 5 л гексана экстракт (12,6 г) очищают флэш-хроматографией на колонке (диаметр 5,5 см, длина 20 см) с использованием в качестве нормальной фазы силикагеля, мобильная фаза: этилацетат: MeOH: по 1 л каждой из систем: 1:0; 20:1; 10:1; 5:1 и 7:3. Элюируют фракции по 250 мл и объединяют в связи с условиями ТСХ (силикагель, нормальная фаза, EtOAc:MeOH, 5:2, сафрацин D: Rf - 0,2; сафрацин E - 0,05). Фракцию, содержащую неочищенные сафрацин D и E, выпаривают в высоком вакууме (2,2 г). Необходима дополнительная стадия очистки для разделения сафрацина D и E в аналогичных условиях (EtOAc:MeOH от 1:0 до 5:1), и после этого фракции, содержащие сафрацин D и E, разделяют и выпаривают, и дальше очищают хроматографией на колонке с сефадексом LH-20, которую элюируют метанолом.
Полученные сафрацины D и E представляют собой формы, осажденные из CH2Cl2 (80 мл) и гексана (1500 мл), имеющие вид сухого зеленого/желтоватого сухого твердого вещества (800 мг сафрацина D) и (250 мг сафрацина Е).
Биологическая активность сафрацина D
Противоопухолевый скрининг:
Клеточные линии (моль/л)
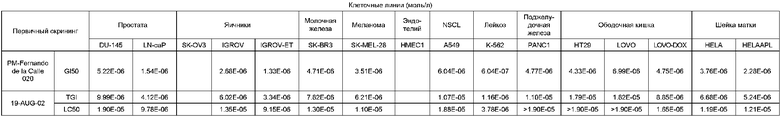
Противомикробная активность: на твердой среде.
Bacillus subtilis: 10 мкг/диск (диаметр 6 мм): Зона ингибирования: 15 мм в диаметре.
Спектроскопические данные:
ESMS: m/z 509 [М-H2O+Н]+; 1Н ЯМР (CDCl3, 300 МГц): 6,50 (с, С-15), 4,02 (с, ОМе), 3,73 (с, ОМе), 2,22 (с, Me), 1,85 (с, Me), 0,80 (д, J=7,2 Гц); 13С ЯМР (CDCl3, 75 МГц): 186,51, 181,15, 175,83, 156,59, 145,09, 142,59, 140,78, 137,84, 131,20, 129,01, 126,88, 121,57 (2 × С), 82,59, 60,92, 60,69, 53,12, 21,40, 50,68, 50,22, 48,68, 40,57, 29,60, 25,01, 21,46, 15,64, 8,44.
Штамм:
Тот же, что и в случае сафрацина D.
Условия ферментации:
Та же партия, что и в случае сафрацина D.
Выделение
См. условия получения сафрацина D.
Биологическая активность сафрацина Е
Противоопухолевый скрининг:
Клеточные линии (моль/л)
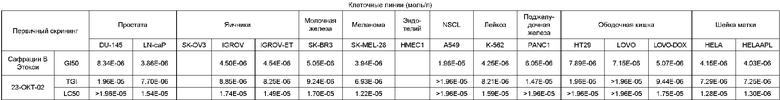
Противомикробная активность: на твердой среде.
Bacillus subtilis: 10 мкг/диск (диаметр 6 мм): Зона ингибирования: 9,5 мм.
Спектроскопические данные:
ESMS: m/z 511 [M+H]+; 1H ЯМР (CDCl3, 300 МГц): 6,51 (c, C-15), 4,04 (c, OMe), 3,75 (c, OMe), 2,23 (c, Me), 1,89 (c, Me), 0,84 (д, J=6,6 Гц); 13C ЯМР (CDCl3, 75 МГц): 186,32, 181,28, 175,83, 156,43, 145,27, 142,75, 141,05, 137,00, 132,63, 128,67, 126,64, 122,00, 120,69, 60,69, 60,21, 59,12, 58,04, 57,89, 50,12, 49,20, 46,72, 39,88, 32,22, 25,33, 21,29, 15,44, 8,23.
Пример 5. Перекрестные эксперименты по исследованию добавляемых компонентов
Гетерологичная экспрессия генов биосинтеза предшественников сафрацина для продукции Р2 и Р14
С целью пролить свет на механизм биосинтеза Р2 и Р14 авторы проводят клонирование и экспрессию по направлению считывания генов NRPS для определения их биохимической активности.
Для достижения сверхпродукции Р14 клонируют гены sасEFGH (pB7983) (фиг.4). Для достижения сверхпродукции Р2 в гетерологичной системе клонируют гены sacD - sасH (pB5H83) (фиг.4). Для достижения данной цели используют ПЦР-амплифицированные фрагменты, несущие интересующие гены, с использованием олигонуклеотидов, которые содержат сайт рестрикции XbaI на 5'-конце. Используют олигонуклеотиды PFSC79 (5'-CGTCTAGACACCGGCTTCATGG-3') и PFSC83 (5'-GGTCTAGATAACAGCCAACAAACATA-3') для амплификации генов sacE - sacH; и используют олигонуклеотиды 5HPT1-XB (5'-CATCTAGACCGGACTGATATTCG-3') и PFSC83 (5'-GGTCTAGATAACAGCCAACAAACATA-3') для амплицикации генов sacD - sacH. ПЦР-фрагменты, расщепленные XbaI, клонируют в сайте рестрикции XbaI в pBBR1-MCS2 плазмиде (Kovach et al., Gene 1994, 166, 175-176). Обе плазмиды pB7983 и pB5H83 вводят по отдельности в три гетерологичные бактерии P. fluorescens (CECT 378), P. putida (ATCC12633) P.stutzeri (ATCC 17588) путем конъюгации (см таблицу II). По данным ВЭЖХ анализа, в ферментированном культуральном бульоне с штаммами-трансконъюгатами наблюдается большое количество Р14 соединения во всех трех штаммах, содержащих плазмиду pB7983, тогда как большие количества Р2 и Р14 продуктов наблюдаются в том случае, если проводить экспрессию pB5H83 плазмиды в гетерологичных бактериях.
Перекрестный вариант культивирования
Как видно из данных примера 4, мутанты sacF - (PM-S1-008) и sacG - (PM-S1-009) не способны продуцировать ни сафрацин, ни P2 и P14 соединения. Добавление химически синтезированного P2 к указанным мутантам в ходе ферментации приводит к получению сафрацина.
Кроме того, совместное культивирование гетерологичного штамма P. stutzeri (ATCC 17588), несущего плазмиду pB5H83 (PM-18-004), при экспрессии которой образуются P2 и P14, с одним из двух штаммов sacF - и sacG -, приводит к образованию сафрацина. Совместное культивирование гетерологичного штамма P. stutzeri (ATCC 17588), несущего плазмиду pB7983 (PM-18-005), экспрессия которого приводит только к получению Р14, с одним из двух мутантов P. fluorescens А2-2, указанных ранее, вообще не приводит к образованию сафрацина. Полученные результаты позволяют полагать, что Р14 трансформируется в Р2, молекулу, которая может легко транспортироваться внутрь и наружу через клеточную стенку Pseudomonas sp., и присутствие которого абсолютно необходимо для биосинтеза сафрацина.
Пример 6. Биологическая продукция новых "неприродных" молекул
Добавление в количестве 2 г/л специфически модифицированного производного предшественника Р2, Р3 3-гидрокси-5-метил-O-этилтирозина к sacF мутанту (PM-S1-008) в ходе ферментации, дает два «неприродных» сафрацина, которые включают модифицированный предшественник Р3 в свою структуру, сафрацин A(OEt) и сафрацин В(OEt).
Штамм:
Мутант sacF на основе P.fluorescens А2-2 (PM-S1-008).
Условия ферментации:
Посевную среду, содержащую 1% глюкозы, 0,25% мясного экстракта, 0,5% бактопептона, 0,25% NaCl, 0,8% СаСО3, инокулируют 0,1% замороженной вегетативной культуры микроорганизмов и инкубируют на роторной качалке (250 об/мин) при 27°С. После 30 часов инкубации 2% об/об посевной культуры мутанта PM-S1-008 переносят в эрленмейеровские колбы на 2000 мл, содержащие 250 мл среды М-16 В для продукции, состоящей из 15,2% маннита, 3,5% сухих пекарских дрожжей, 1,4% (NH4)2, 0,001% FeCl3, 2,6% СаCO3 и 0,2% Р3 (3-гидрокси-5-метил-О-метилтирозин). Температура в ходе инкубации составляет 27°С, начиная с момента инокуляции до 40 часов, и затем 24°С до окончания процесса (71 час). Значение pH не контролируют. Скорость перемешивания в роторной качалке составляет 220 об/мин с эксцентриситетом 5 см.
Выделение:
Объединяют содержимое 4 х 2000/250 мл эрленмейеровских колб (970 мл), центрифугируют (12000 об/мин, 4°С, 10', J2-21 центрифуга BECKMAN) для удаления клеток. pH в очищенном бульоне (765 мл) доводят до значения 9,0, добавляя 10% NaOH. Затем осветленный щелочью бульон экстрагируют в пропорции 1:1 (объем/объем) EtOAc (x2). Органическую фазу выпаривают в высоком вакууме и получают жирный темный экстракт (302 мг).
Экстракт промывают путем растирания с гексаном для удаления примесей и твердые компоненты очищают колоночной хроматографией с использованием нормальной фазы силикагеля и смеси этилацетат/метанол (от 12:1 до 1:1). Фракции анализируют в УФ при использовании метода ТСХ (силикагель 60, подвижная фаза EtOAc:MeOH, 5:4, Rf - 0,3 для Сафрацина B-OЕt и 0,15 для сафрацина A-OЕt). В результате данной процедуры получают сафрацин B OЕt (25 мг) и сафрацин A OЕt (20 мг).
Биологическая активность сафрацина В(OEt)
Противоопухолевая активность
Клеточные линии (моль/л)
Противомикробная активность: на твердой среде.
Bacillus subtilis: 10 мкг/диск (диаметр 6 мм): зона ингибирования 17,5 мм.
Спектроскопические данные:
ESMS: m/z 551 [М-H2O+Н]+; 1H ЯМР (CDCl3, 300 МГц): 6,48 (с, Н-15), 2,31 (с, 16-Ме), 2,22 (с, 12-NMe), 1,88 (с, 6-Ме), 1,43 (т, J=6,9 Гц, Ме-этокси), 1,35 (т, J=6,9 Гц, Ме-этокси), 0,81 (д, J=7,2 Гц, Н-26).
Штамм:
Тот же, что и в случае сафрацина В(OEt).
Условия ферментации:
Те же, что и в случае сафрацина В(OEt).
Выделение:
Объединяют содержимое 4 × 2000/250 мл эрленмейеровских колб (970 мл), центрифугируют (12000 об/мин, 4°С, 10', J2-21 центрифуга BECKMAN) для удаления клеток. В осветленном бульоне (765 мл) доводят значение рН до 9,0, добавляя 10% NaOH. Затем осветленный щелочью бульон экстрагируют в пропорции 1:1 (объем/объем) EtOAc (х2). Органическую фазу выпаривают в высоком вакууме и получают жирный темный экстракт (302 мг).
Экстракт промывают путем растирания с гексаном для удаления примесей и твердые компоненты очищают колоночной хроматографией с использованием нормальной фазы силикагеля и смеси этилацетат/метанол (от 12:1 до 1:1). Фракции анализируют в УФ при использовании метода ТСХ (силикагель 60, подвижная фаза EtOAc:MeOH, 5:4, Rf - 0,3 для сафрацина B-OEt и 0,15 для сафрацина A-OEt). В результате данной процедуры получают сафрацин В OEt (25 мг) и сафрацин A OEt (20 мг).
Биологическая активность сафрацина A(OEt)
Противоопухолевая активность
Противомикробная активность: на твердой среде.
Bacillus subtilis: 10 мкг/диск (диаметр 6 мм): зона ингибирования 10 мм.
Спектроскопические данные:
ESMS: m/z 553 [M+H]+; 1Н ЯМР (CDCl3, 300 МГц): 6,48 (c, H-15), 2,33 (c, 16-Me), 2,21 (c, 12-NMe), 1,88 (c, 6-Me), 1,42 (т, J=6,9 Гц, Ме-этокси), 1,34 (т, J=6,9 Гц, Ме-этокси), 0,8 (д, J=6,9 Гц, H-26).
Пример 7. Ферментативная трансформация сафрацина В в сафрацин А
Для оценки ферментативной активности, направленной на превращение сафрацина В в сафрацин А, культуры различных штаммов после 120 часов ферментации (см. условия в примере 2: Биологический анализ (биотест) образования сафрацина) собирают и центрифугируют (9000 об/мин х 20 минут). Анализируемые штаммы включают P. fluorescens A2-2, а также штамм дикого типа и P. fluorescens CECT378 + pBHPT3 (PM-19-006) в качестве клетки-хозяина для гетерологичной экспрессии. Супернатант отбрасывают и клетки промывают (NаCl 0,9%) дважды и ресуспендируют в 60 мл 100 мМ фосфатного буфера, pH 7,2. 20 мл клеточной суспензии распределяют по трем эрленмейеровским колбам:
а) клеточная суспензия + сафрацин В (400 мг/л);
b) клеточная суспензия, нагретая до 100°С в течение 10 минут, + сафрацин В (400 мг/л) (отрицательный контроль);
с) клеточная суспензия без сафрацина В (отрицательный контроль).
Биохимическую реакцию проводят в процессе инкубирования при 27°С и скорости 220 об/мин, и каждые 10 минут отбирают образцы. Трансформацию сафрацина В в сафрацин А отслеживают методом ВЭЖХ. Полученные результаты четко показывают, что ген sacH, клонированный в pBHPT3, кодирует белок, ответственный за трансформацию сафрацина В в сафрацин А.
На основании данных результатов авторы провели анализ, с тем чтобы выяснить, способен ли данный тот же самый фермент распознавать другой субстрат, такой как эктеинасцидин 743 (ET-743), и трансформировать данное соединение в Et-745 (в котором гидроксигруппа в положении С-21 отсутствует). Повторяют указанные выше эксперименты с получением эрленмейеровских колб, содержащих:
а) клеточную суспензию + ЕТ-743 (приблизительно 567 мг/л);
b) клеточную суспензию, нагретую до 100°С в течение 10 минут, + ЕТ-743 (567 мг/л) (отрицательный контроль);
с) клеточную суспензию без ЕТ-743 (отрицательный контроль).
Биохимическую реакцию проводят в процессе инкубации при 27°С и скорости 220 об/мин, и образцы отбирают в точке 0, 10 минут, 1 час, 2 часа, 3 часа, 4 часа, 20 часов, 40 часов, 44 часа, 48 часов. Трансформацию ЕТ-743 в ЕТ-745 отслеживают методом ВЭЖХ. Полученные результаты четко показывают, что ген sacH, клонированный в pBHPT3, кодирует белок, ответственный за трансформацию ЕТ-743 в ЕТ-745. Полученный результат показывает, что данные ферменты распознают эктеинасцидин как субстрат и что он может использоваться при биологической трансформации широкого перечня структур.
ПОСЛЕДОВАТЕЛЬНОСТИ



























































| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЫ-МИШЕНИ ДЛЯ НАПРАВЛЕННОГО ВОЗДЕЙСТВИЯ НА АЗОТФИКСАЦИЮ ДЛЯ УЛУЧШЕНИЯ КАЧЕСТВ РАСТЕНИЙ | 2018 |
|
RU2805085C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ 1-БУТАНОЛА | 2006 |
|
RU2429295C2 |
| СПОСОБ КОНСТРУИРОВАНИЯ РЕКОМБИНАНТНЫХ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ PANTOEA, И СПОСОБ ПРОДУКЦИИ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ PANTOEA | 2006 |
|
RU2418069C2 |
| ГЕНЫ БУТИРОБЕТАИН/КРОТОНОБЕТАИН-L-КАРНИТИН-МЕТАБОЛИЗМА И ИХ ПРИМЕНЕНИЕ ДЛЯ МИКРОБИОЛОГИЧЕСКОГО ПОЛУЧЕНИЯ L-КАРНИТИНА | 1994 |
|
RU2133773C1 |
| СПОСОБ ПОЛУЧЕНИЯ ГИАЛУРОНАНА В РЕКОМБИНАНТНОЙ КЛЕТКЕ-ХОЗЯИНЕ | 2002 |
|
RU2346049C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОВЫШЕНИЯ ПРОДУКЦИИ ЭНДУРАЦИДИНА В ГЕННО-ИНЖЕНЕРНОМ ШТАММЕ Streptomyces fungicidicus | 2017 |
|
RU2790405C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ЧЕТЫРЕХУГЛЕРОДНЫХ СПИРТОВ | 2006 |
|
RU2394913C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ АМИНОКИСЛОТ СЕМЕЙСТВА АСПАРТАТА С ИСПОЛЬЗОВАНИЕМ МИКРООРГАНИЗМОВ | 2010 |
|
RU2535973C2 |
| Средство редактирования генома на основе белка LigD из бактерии Pseudomonas putida и Cas9 комплекса | 2022 |
|
RU2797049C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПИРРОЛОХИНОЛИНОХИНОНА (PQQ) С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ РОДА Methylobacterium ИЛИ Hyphomicrobium | 2011 |
|
RU2504584C2 |
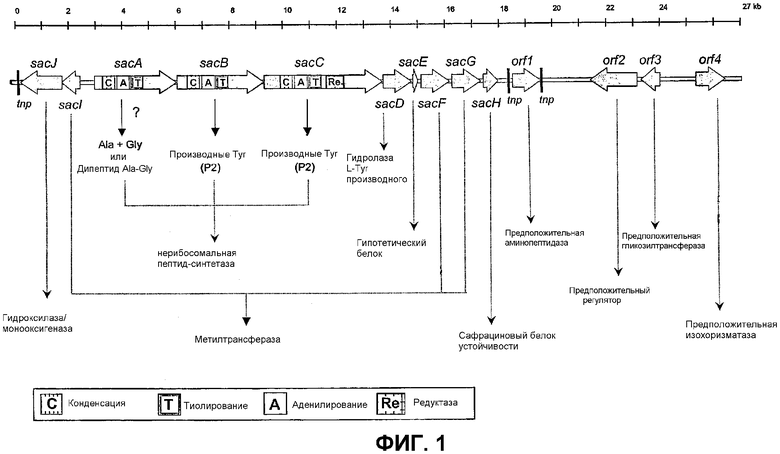
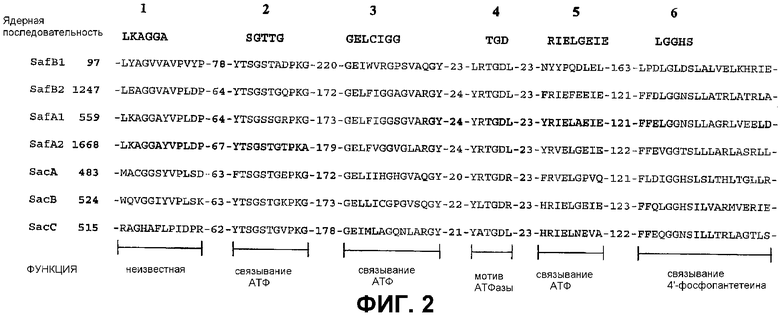
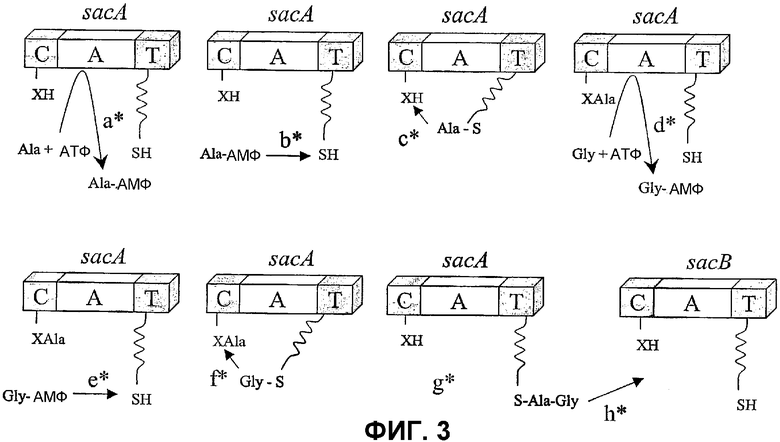
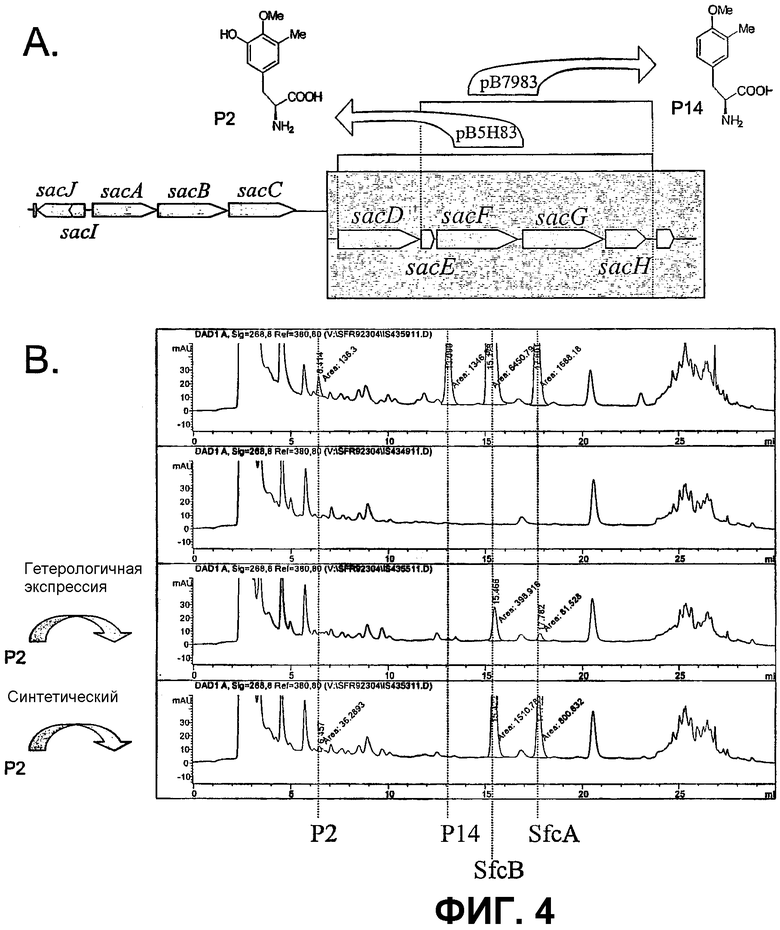
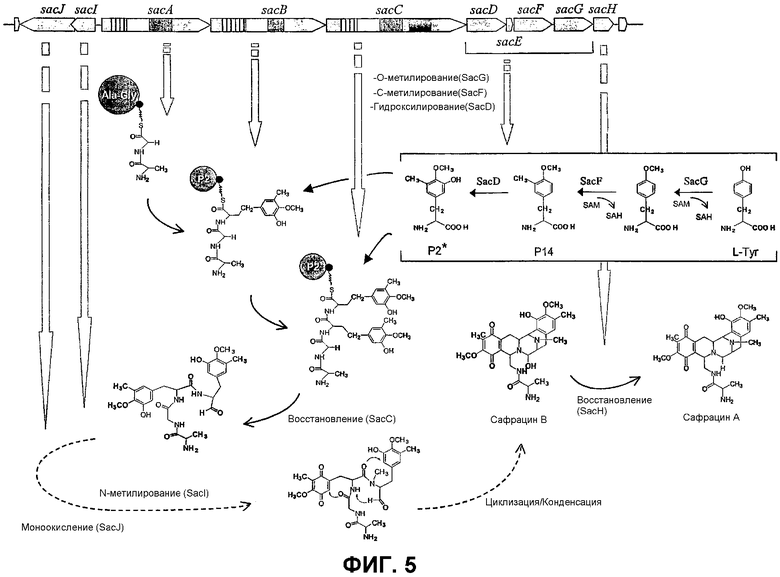
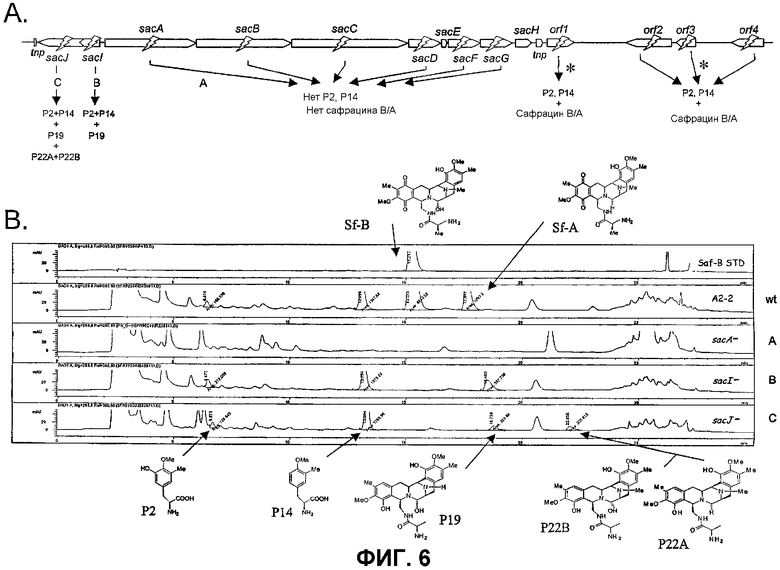
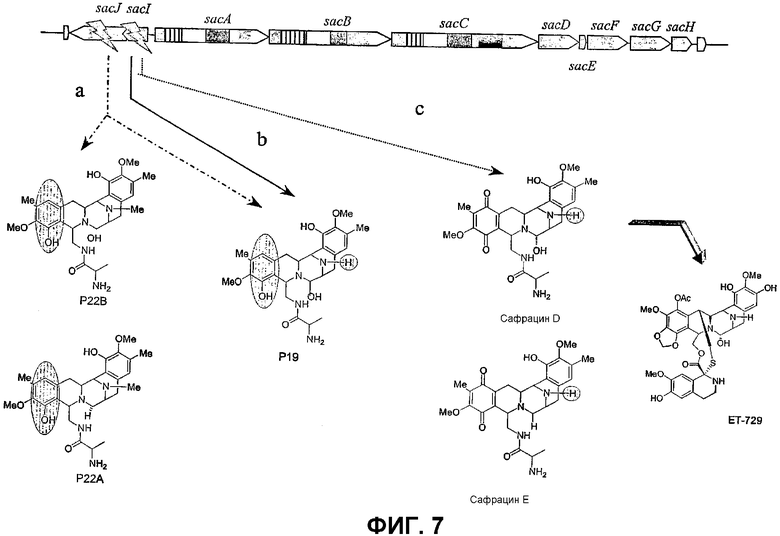
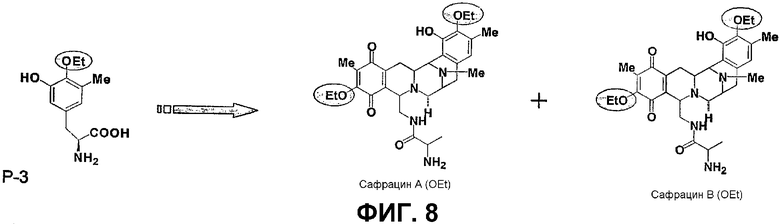
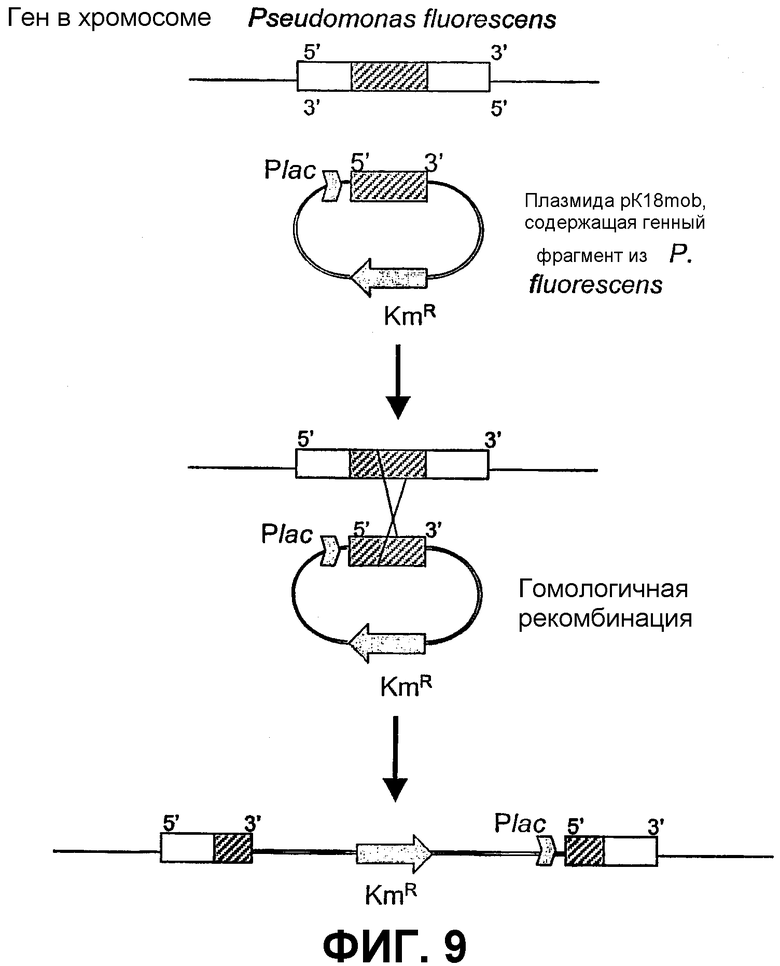
Изобретение относится к области молекулярной биологии и биотехнологии и может быть использовано в медико-биологической промышленности при производстве антимикробных и противоопухолевых препаратов. Выделен фрагмент геномной ДНК Pseudomonas fluorescens А2-2, включающий полноразмерный генный кластер биосинтеза сафрацина (А и В), при исследовании которого установлено наличие нескольких ОРС, организованных в два оперона: sacABCDEFGH и sacIJ. Путем экспрессии нуклеиновой кислоты, включающей полный генный кластер сафрацина, в гетерологичной системе получены рекомбинантные формы природных сафрацинов А и В. В результате удаления или нарушения функционирования отдельных генов, входящих в состав оперонов, и применения полученных модифицированных форм нуклеиновой кислоты в технологии рекомбинантных ДНК синтезированы аналоги сафрацина, которые могут быть использованы в качестве антимикробных или противоопухолевых средств, а также в синтезе эктеинасцидиновых соединений. 13 н. и 7 з.п. ф-лы, 9 ил.
1. Нуклеиновая кислота, обеспечивающая биосинтез сафрацина А и В, которая выбрана из группы, состоящей из:
a) нуклеиновой кислоты, содержащей полноразмерный генный кластер сафрацина Pseudomonas fluorescens А2-2 и характеризующейся последовательностью SEQ ID NO:1;
b) нуклеиновой кислоты, по существу, соответствующей оперонам sacABCDEFGH и sacIJ генного кластера сафрацина Pseudomonas fluorescens А2-2 и характеризующейся структурой и расположением в последовательности SEQ ID NO:1, как показано на фиг.1;
c) нуклеиновой кислоты, представляющей собой вариант а) или b), который включает замены в кодирующих участках, определенные вырожденностью генетического кода.
2. Нуклеиновая кислота по п.1, содержащая гены sacA, sacB, scC, sacD, sacE, sacG и sacH.
3. Нуклеиновая кислота, обеспечивающая биосинтез структурного аналога сафрацина, которая представляет собой генетически модифицированную форму нуклеиновой кислоты по п.1 и характеризуется отсутствием или нарушением функционирования гена sacI и/или гена sacJ оперона sacIJ.
4. Нуклеиновая кислота по п.3, в которой ген sacI отсутствует или разрушен.
5. Нуклеиновая кислота по п.3, в которой ген sacJ отсутствует или разрушен.
6. Нуклеиновая кислота по п.3, в которой ген sacI отсутствует или разрушен, а экспрессия гена sacJ восстановлена.
7. Нуклеиновая кислота, используемая для получения сафрацина или его структурного аналога, которая представляет собой генетически модифицированную форму нуклеиновой кислоты по п.1 и характеризуется отсутствием или нарушением функционирования гена sacF или гена sacG.
8. Вектор экспрессии, содержащий нуклеиновую кислоту по п.1.
9. Клетка-хозяин Pseudomonas sp., трансформированная вектором по п.8, обеспечивающая биосинтез сафрацинов А и В.
10. Способ получения сафрацинов А и В, включающий:
a) культивирование клетки-хозяина, содержащей нуклеиновую кислоту по п.1 в подходящей культурной среде и в условиях для пролиферации клеток;
b) восстановление культуральной среды и
c) очистку сафрацинов из культуральной среды.
11. Способ получения структурного аналога сафрацина, включающий:
a) культивирование клетки-хозяина, содержащей нуклеиновую кислоту по п.3 в подходящей культурной среде и в условиях для пролиферации клеток;
b) восстановление культуральной среды и
c) очистку соединения сафрацина из культуральной среды.
12. Способ по п.11, где указанная клетка-хозяин содержит нуклеиновую кислоту по п.4.
13. Способ по п.11, где указанная клетка-хозяин содержит нуклеиновую кислоту по п.5.
14. Способ по п.11, где указанная клетка-хозяин содержит нуклеиновую кислоту по п.6.
15. Способ получения сафрацинов А и В, включающий:
a) культивирование клетки-хозяина, содержащей нуклеиновую кислоту по п.7, в подходящей культуральной среде и в условиях для пролиферации клеток;
b) восстановление культуральной среды и
c) очистку сафрацинов из культуральной среды,
где указанная культуральная среда содержит L-Tyr производное Р2 со следующей структурой:
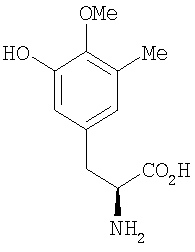 .
.
16. Способ получения структурного аналога сафрацина, включающий:
a) культивирование клетки-хозяина, содержащей нуклеиновую кислоту по п.7, в подходящей культуральной среде и в условиях для пролиферации клеток;
b) восстановление культуральной среды и
c) очистку сафрацинов из культуральной среды, где указанная культуральная среда содержит L-Tyr производное Р3 со следующей структурой:
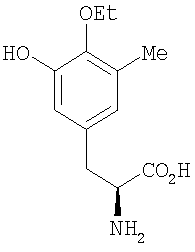 .
.
17. Структурный аналог сафрацина формулы:
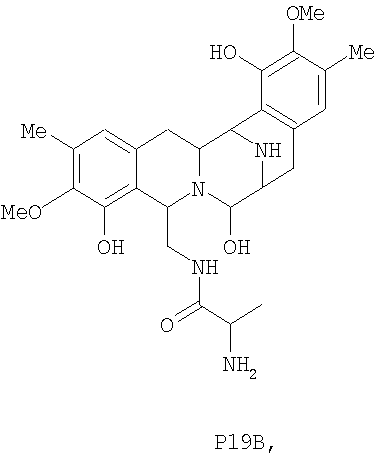
полученный способом по п.12 или 13.
18. Структурный аналог сафрацина формулы:
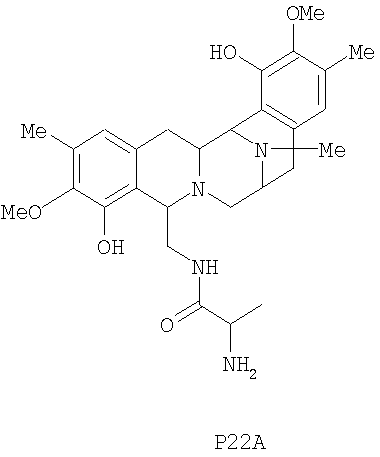 или
или 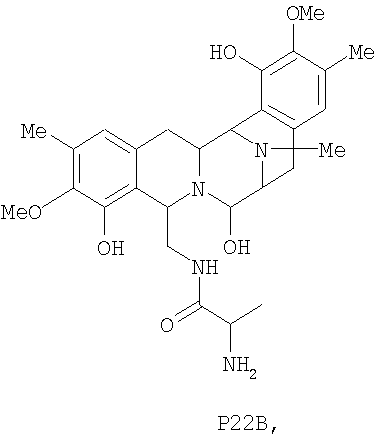
полученный способом по п.13.
19. Структурный аналог сафрацина формулы:
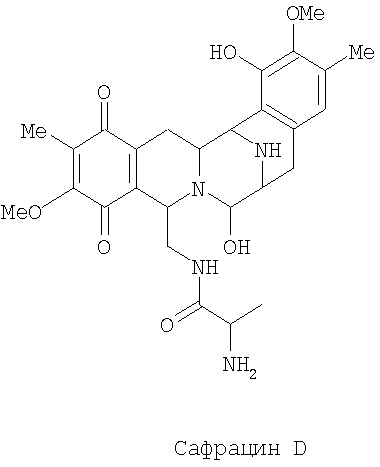 или
или 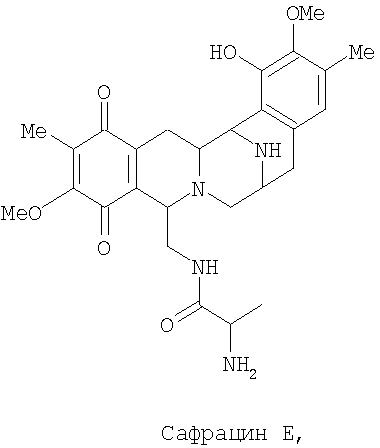
полученный способом по п.14.
20. Структурный аналог сафрацина формулы:
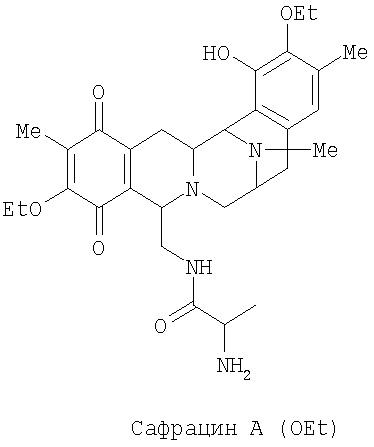 или
или 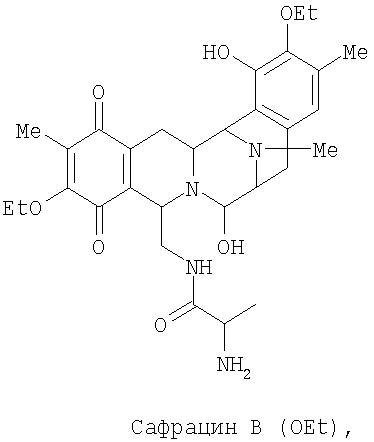
полученный способом по п.16.
| IKEDA et al | |||
| J | |||
| Antibiotics, 36, 1279-1283, 1983 | |||
| Устройство для очистки хлопка-сырца | 1946 |
|
SU69862A1 |
| POSPIESCH А | |||
| et al | |||
| Microbiology, 142, 741-746, 1996. | |||

