Область техники, к которой относится изобретение
Изобретение относится к области медицинской фармакологии, более конкретно к поиску соединений, снижающих функциональную активность протеазы вируса иммунодефицита человека (протеазы ВИЧ) путем ингибирования ассоциации ее субъединиц. Изобретение ориентировано на разработку новых лекарственных средств.
Предшествующий уровень техники настоящего изобретения
Терапия, основанная на применении ингибиторов протеазы ВИЧ, продляет жизнь пациентов с синдромом приобретенного иммунодефицита (Sepkowitz К.A. AIDS - the first 20 years. N. Engl. J. Med. 2001, 344, 1764-1772). Протеаза ВИЧ - фермент, расщепляющий вирусный полипротеин на функционально активные зрелые белки, необходимые для воспроизводства и сборки ВИЧ. Обычно используют конкурентные ингибиторы протеазы, непосредственно связывающиеся активным центром фермента. Эффективность применения таких ингибиторов в ходе лечения быстро уменьшается из-за точечных мутаций вирусного генома, подавляющих связывание протеазой ингибитора в большей степени, чем ее ферментативную активность. В результате организм поражается возникшими штаммами ВИЧ, которые приобрели резистентность к используемому ингибитору.
Альтернативой служит ингибирование протеазы путем блокирования образования функционально-активной формы фермента (Bannwarth L., Reboud-Ravanx М. An alternative strategy for inhibiting multidrug-resistant mutants of the dimeric HIV-1 protease by targeting the subunit interface. Biochem. Soc. Trans. 2007 Jun; 35(Pt 3):551-4), которая представляет собой димер гомологичных субъединиц. Главный участок их контакта представляет собой бета-структуру, тяжи которой являются С- и N-концевыми фрагментами субъединиц. Показано, что пептиды и пептидомиметики, имитирующие эти фрагменты, препятствуют сборке субъединиц, действуя в качестве неконкурентных ингибиторов протеазы ВИЧ (Zutshi R., Brickner М., Chmielewski J. Inhibiting the assembly of protein-protein interfaces. Current Opinion in Chemical Biology, 1998, 2(1): 62-66(5)). Такого рода ингибиторы предпочтительнее в качестве лекарственных средств, поскольку для возникновения резистентности необходимы сочетанные мутации в заведомо консервативных терминальных участках полипептидной цепи протеазы (Archakov A.I., Govorun V.M., Dubanov A.V., Ivanov Y.D., Veselovsky A.V., Lewi P., Janssen P. Protein-protein interactions as a target for drugs in proteomics. Proteomics. 2003 Apr; 3(4):380-91).
Изученные в настоящее время ингибиторы димеризации представляют собой модифицированные пептиды или пептидомиметики, а также вещества синтетического и естественного происхождения (Camarasa M.J., Velázquez S., San-Félix A., Pérez-Pérez M.J., Gago F. Dimerization inhibitors of HIV-1 reverse transcriptase, protease and integrase: a single mode of inhibition for the three HIV enzymes? Antiviral Res. 2006 Sep; 71(2-3):260-7). Co структурной точки зрения указанные вещества имитируют протяженные участки бета-структуры.
Раскрытие настоящего изобретения
Изобретение относится к способу поиска новых химических соединений - ингибиторов димеризации (ИД) - способных воздействовать на сборку субъединиц протеазы ВИЧ путем конкурентного взаимодействия с контактной площадкой субъединиц претеазы а также влияющих на конформацию субъединиц, снижая их сродство друг к другу.
Для выявления новых ИД использовался способ, основанный на вычислительной методике подбора соединений по компьютерной модели структуры протеазы. С использованием подхода, известного как «аланиновый скрининг», выявлялись аминокислотные остатки, определяющие силу взаимодействия между субъединицами протеазы. Было установлено, что замена на остаток аланииа в структуре протеазы аминокислотных остатков, перечисленных в таблице 1 (далее - контактные точки), приводит к резкому снижению энергии взаимодействия субъединиц. Для каждой контактной точки методом молекулярного докинга проводился скрининг базы данных низкомолекулярных соединений и ряда сконструированных пептидомиметиков на предмет выявления кандидатов, обладающих высокими значениями энергии связи с субъединицей. С использованием известных ИД было установлено пороговое значение для функции оценки эффективности взаимодействия соединения с контактной точкой. В результате были отобраны соединения, характеризующиеся значениями энергии связи выше установленного порога, а также те из них, которые характеризовались высоким сродством к участкам поверхности молекулы протеазы ВИЧ. Последнее оценивалось методом молекулярной динамики, в ходе которой регистрировалось влияние связывания соединения с поверхностными элементами белковой молекулы на конформационные свойства контактных участков. В таблице 1 приведены номера аминокислотных остатков в составе субъединицы протеазы, которые являются определяющими для молекулярного механизма действия ИД. Таким образом, примененная вычислительная методика позволила отобрать структуры как избирательно связывающиеся с участками взаимодействия субъединиц протеазы ВИЧ, так и влияющие на пространственную конформацию субъединицы. Отобранные указанным способом ИД приведены в таблице 3.
Для подтверждения механизма действия отобранные соединения выборочно тестировались согласно измерительной схеме, основанной на высокоточной регистрации межмолекулярных взаимодействий на оптическом биосенсоре модели Biacore-3000. В измерительной и контрольной кювете прибора были ковалентно иммобилизованы молекулы протеаза ВИЧ в димерной форме. Затем в контрольной кювете субъединицы димеров протеаза ВИЧ необратимо связывались друг с другом путем дополнительных ковалентных связей, тогда как в измерительной кювете осуществлялась диссоциация димеров до мономеров путем промывки потоком буферной смеси.
Указанная экспериментальная схема позволяла регистрировать взаимодействие ингибитора с отдельными субъединицами (мономерами) протеазы, помещавшимися в измерительном канале, за вычетом взаимодействия с комплексом из двух субъединиц (димерами) в контрольном канале. Обнаружено, что найденные ингибиторы влияют на характеристики взаимодействия субъединиц и при этом не затрагивают характеристики целого (собравшегося) димера протеазы.
Для проверки влияния найденных ИД на ферментативную активность протеазы использовалась стандартная методика, в основе которой лежит спектрофотометрическое измерение процесса ферментативного гидролиза тестового субстрата. В измерительной кювете находился объем буферного раствора, содержащего протеазу, в который вносили ИД в различных концентрациях и в течение 20 мин измеряли поглощение при длине волны 300 нм. На основе полученных данных рассчитывали значения константы ингибирования. Полученные значения для 4-х из 15 выборочно протестированных соединений варьировали с диапазоне от 1,5 до 0,01 мкМ, что сравнимо с эффективностью известных на настоящее время ингибиторов димеризации протеазы ВИЧ-1.
Таким образом, настоящее изобретение также относится к способу ингибирования димеризации субъединиц протеазы ВИЧ, предусматривающему контактирование, по меньшей мере, одного соединения-ингибитора протеазы ВИЧ, выявленного в соответствии с описанным выше способом, со средой, содержащей протеазу ВИЧ. Эффективность ингибирования димеризации субъединиц протеазы ВИЧ указанными соединениями наглядно подтверждена представленными ниже экспериментальными данными.
Краткое описание чертежей
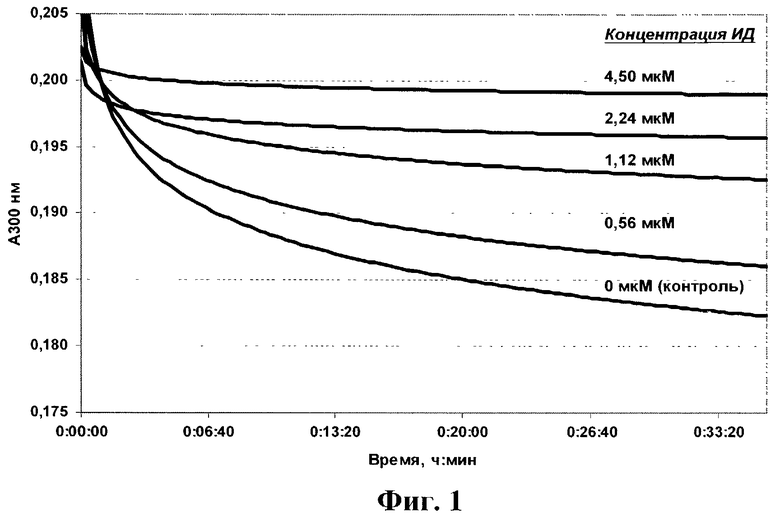
На фиг.1 в качестве примера приведены кинетические записи изменения поглощения при 300 нм во времени в присутствии ИД (препарат №10).
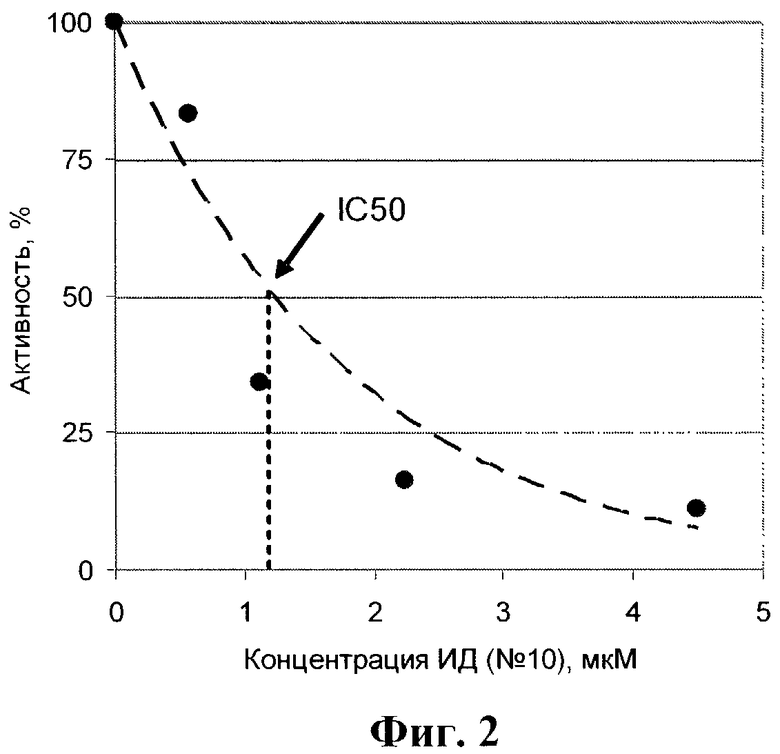
На фиг.2 приведена зависимость активности пВИЧ в присутствии различных концентраций ИД (препарат №10). Из зависимости активности пВИЧ в присутствии различных концентраций ИД было рассчитаны значения IC50.
Пример осуществления настоящего изобретения
Пример 1. Подбор соединений (а) и тестирование с использованием виртуальной протеазы ВИЧ (б)
(а)
1. Материалы
1.1. Структура протеазы ВИЧ
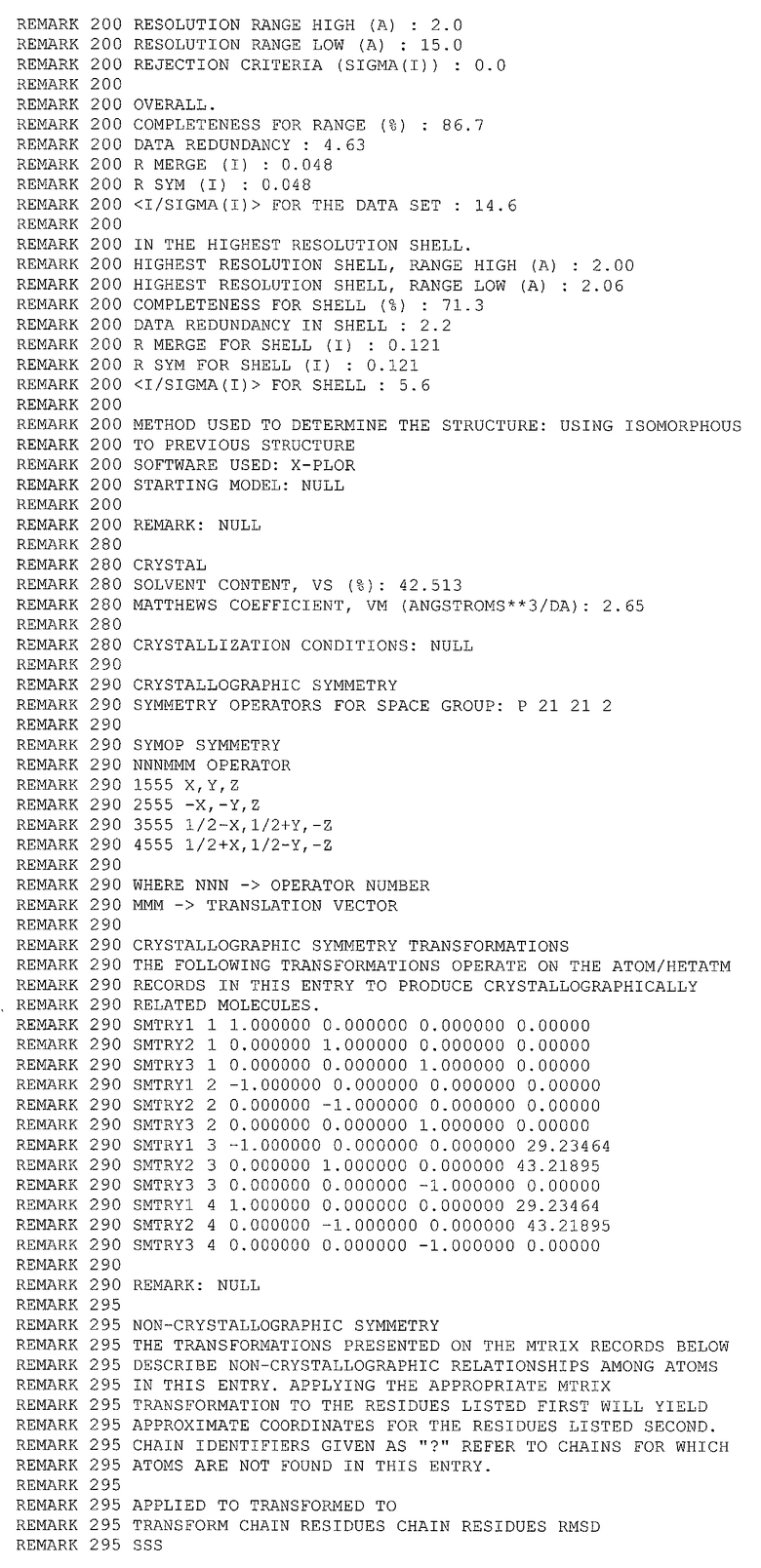
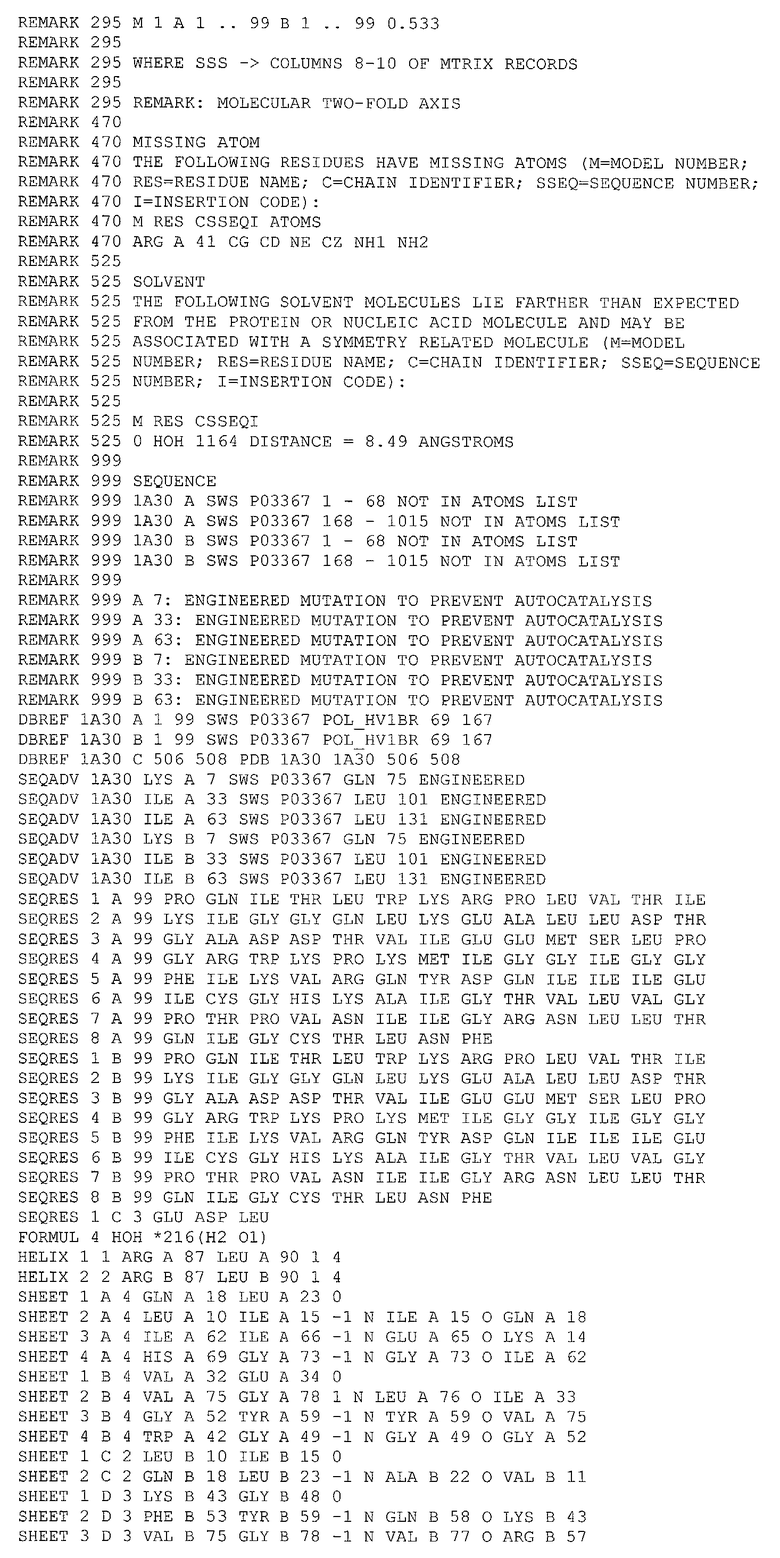
В качестве координат исходной структуры протеазы ВИЧ-1 использовались координаты, полученные рентгеноструктурным способом, депонированные в Protein Data Bank под кодом 1А30. Дата депонирования 27/12/1998. Код фермента по ЕС: 3.4.23.16. Разрешение РСМ 2,0 ангстрем.
Состав структуры:
1. Симметричный димер следующего аминокислотного состава (по 99 аминокислотных остатков в каждой цепи)
2. Трипептидный конкурентный ингибитор
GLU ASP LEU
3. 216 молекул воды.
1.2. Базы данных низкомолекулярных соединений
1. База данных NCI Diversity Set, включающая информацию о 1990 химических соединениях, отобранных из базы данных NCI (140000 соединений доступных в Национальном институте рака США), с учетом исключения структурно подобных соединений.
2. База данных NCI, включающая информацию о более чем 250000 химических соединений.
2. Методы
2.1. Оптимизация структуры
Из исходной структуры были удалены: молекулы ингибитора, молекулы воды. Координаты атомов водорода восстановлены с использование программы Reduce (Word, J.M.; Lovell, S.C.; LaBean, Т.Н.; Taylor, H.C.; Zalis, M.E.; Presley, B.K.; Richardson, J.S.; Richardson, D.C. J. Mol. Biol. 1999, 285, 1711-1733). Структура димера была подвергнута процедуре симуляции молекулярной динамики в вакууме. Использовалась программа CHARMM (Brooks, B.R.; Bruccoleri, R.; Olafson, В.; States, D.; Swaninathan, S.; Karplus, M. J. Comp. Chem. 1983, 4, 187-217), выбранное молекулярно-механическое силовое поле Merck (Halgren, T.A. J. Сотр. Chem. 1996, 17, 490-519). Время уравновешивания системы 50 нс с шагом 1 фс.
2.2. Аланиновый скрининг
Симуляция процедуры аланинового сканирования был выполнена также средствами программы CHARMM с использованием процедуры молекулярной динамики. Собственное время симуляции после уравновешивания составляет 10 пс (шаг 1 фс) с записью промежуточных состояний через каждые 10 шагов. Для каждого промежуточного состояния проводилась последовательная виртуальная замена каждого аминокислотного остатка субъединицы протеазы на аланин, в ходе каждой из них проводилась оценка изменений внутренней энергии комплекса субъединиц (в ккал/моль) по формуле
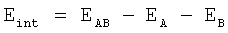 ,
,
где Eint - энергия взаимодействия между субъединицами, ЕАВ - энергия димера EA, EB - энергия каждой из цепей.
2.3. Молекулярный докинг
Отбор структур низкомолекулярных соединений из баз данных для тестирования производился методом молекулярного докинга с использование программы Autodock (Morris, G.M.; Goodsell, D.S.; Halliday, R.S,; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. J. Comp. Chem. 1998, 19, 1639-1662). В качестве посадочного места в процедуре докинга использовались зоны на поверхности одной из субъединиц димера, определенные в результате аланинового сканирования. Вторая субъединица была удалена, конформация рабочей субъединицы соответствовала полученной после процедуры оптимизации структуры. Процедура локирования выполнялась в две стадии. Первая стадия включала в себя локирование всех молекул базы данных NCI Diversity Set no 10 попыток на каждую. Использовалась процедура поиска с использованием генетического алгоритма, параметры поиска по умолчанию. Вторая стадия - уточнение положения отобранных на первом шаге молекул. Использовались тот же алгоритм и набор параметров, однако число попыток было увеличено до 50 и изменена точка входа в генератор псевдослучайных чисел. Для каждого полученного в результате последней процедуры варианта комплекса проводилась независимая от оценочной функции Autodock оценка энергии связывания соединений с субъединицей протеазы ВИЧ, с использованием программы DS. Для тестирования предлагались соединения с энергией связывания ниже 6 ккал/моль.
2.4. Дополнительный поиск структур по подобию
Для отобранных по результатам локирования молекул был проведен поиск структурно подобных соединений по полной базе данных NCI. Для найденных подобных соединений процедура молекулярного докинга повторялась полностью и отбирались соединения с энергией связывания ниже 6 ккал/моль.
3. Полученные результаты
3.1. Аланиновый скрининг
Результаты процедуры симуляции аланинового сканирования представлены в таблице 2.
Из результатов аланинового скрининга отобраны те, в которых разность между стандартным отклонением и энергией субъединиц (D=S-Eint) отрицательна. В таблице 2 приведены первые 15 строк, полученных после сортировки по параметру D. Из отобранных аминокислотных остатков были выбраны только те, которые находятся на поверхности белка. Ниже приведены номера и названия аминокислотных остатков, которые по результатам аланинового скринига определяют контакты ИД с субъединицей протеазы ВИЧ.
1) Asn98, Phe99. Декартовы координаты прямоугольной области локирования относительно приложенного файла (см. приложение I):
- минимальные значения по осям X, Y, Z - 15,290, 30,630, -4,210;
- максимальные значения по осям X, Y, Z - 37,790, 53,130, 18,290.
2) Asp29, Asp30 (взаимодействие с областью 3 на противоположной субъединице).
3) Arg8 (взаимодействие с областью 2 на противоположной субъединице).
4) Gly49, Gly51, Gly52.
3.2. Молекулярный докинг
По результатам первой стадии локирования было отобрано 300 соединений с лучшей энергией связывания.
По результатам второго локирования и независимой от Autodock оценки энергии взаимодействия лигандов с субъединицей протеазы ВИЧ (пороговое значение -6 ккал/моль) было отобрано 20 соединений.
3.3. Дополнительный поиск структур по подобию
По результатам поиска по подобию, прохождению полного цикла локирования и отбора по энергии (пороговое значение -7 ккал/моль) было отобрано 14 соединений (см. таблицу 3).
(6)
Измерение ферментативной активности протеазы ВИЧ в присутствии ингибиторов.
1. Материалы
1.1. Препарат протеазы ВИЧ-1
В работе была использована рекомбинантная протеаза ВИЧ-1 фирмы Bachem AG (Швейцария) (препарат Н-9040).
1.2. Хромогенный субстрат для протеазы ВИЧ
Для анализа ферментативной активности протеазы ВИЧ был использован специальный хромогенный субстрат VII: H-Lys-Ala-Arg-Val-Tyr-п-нитро-Phe-Glu-Ala-Nle-NH2 производства фирмы Bachem AG (Швейцария) (препарат Н-1286).
1.3. Контрольный ингибитор димеризации (ИД) протеазы ВИЧ
Для позитивного контроля в работе был использован известный пептидных ИД (ингибитор Шрама) (пальмитоил-TVSYEL).
1.4. Образцы низкомолекулярных соединений
Для экспериментальной проверки действия потенциальны ингибиторов димеризации на ферментативную активность протеазы ВИЧ были взяты 3 образца соединений, отобранных в ходе in vitro скрининга с помощью ОБ.
1.5. Другие реактивы и препараты
Другие реактивы и препараты, использованные в работе, были аналитической чистоты.
2. Методы.
2.1. Кинетические спектральные измерения
Для регистрации кинетики ферментативной реакции был использован спектрофотометр Libra S32PC (Biochrom, Англия) в режиме «Кинетика». Измерение производили при 25°С по снижению поглощения при 300 нм (пик поглощения субстрата VII).
2.2. Анализ ферментативной активности протеазы ВИЧ
Для измерения ферментативной активности протеазы ВИЧ был использован хромогенный субстрат VII, имеющий максимум поглощения в УФ при 300 нм.
Препарат протеазы ВИЧ (100 нг) инкубировали 35 мин при 25°С в 240 мкл буфера (1 М NaCl, 1 мМ ЭДТА, 0,1% CHAPS, 8% ДМСО, 0,1 М ацетат натрия, рН 5,0) в присутствии (опыт) или отсутствии (контроль) разных концентраций потенциального ИД. Конечная концентрация протеазы ВИЧ была примерно 18 нМ. Затем смесь переносили в кварцевую полумикрокювету и запускали реакцию добавлением 10 мкл раствора субстрата VII до конечной концентрации 25 мкМ.
Смесь быстро перемешивали и активность протеазы ВИЧ регистрировали при 25°С по снижению поглощения при 300 нм в течение 35 мин. Активность протеазы ВИЧ (мкмоль субстрата/(мин·мг фермента)) вычисляли по формуле
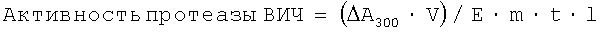
где ΔА300 - изменение оптической плотности,
V - общий объем смеси (л),
Е - коэффициент молярной экстинкции субстрата (М-1см-1),
m - масса протеазы (мг),
t - время измерения (мин),
l - толщина измерительной кюветы (см).
Ингибирующее действие потенциальных ИД и КИД выражалось величиной IC50 (концентрация ингибитора, подавляющая 50% активности фермента).
3. Результаты.
На фиг.1 в качестве примера приведены кинетические записи изменения поглощения при 300 нм во времени в присутствии ИД. Можно видеть, что ИД тормозит ферментативную реакцию. Из зависимостей активности протеазы ВИЧ в присутствии различных концентраций ИД (в качестве примера см. фиг.2) были рассчитаны значения IC50. Полученные результаты приведены в таблице 4.
Таким образом, все три потенциальных ИД, отобранных в эксперименте in vitro с помощью тест-системы на ОБ, показали достаточную ингибирующую активность, сравнимую с контрольным пептидным ИД (ингибитор Шрама).
| название | год | авторы | номер документа |
|---|---|---|---|
| Мутантная АТФ-зависимая протеаза и способ получения L-аминокислоты с ее применением | 2021 |
|
RU2831447C1 |
| СПОСОБЫ ПРИМЕНЕНИЯ ТЕРМОСТАБИЛЬНЫХ СЕРИНОВЫХ ПРОТЕАЗ | 2017 |
|
RU2771261C2 |
| ВАРИАНТЫ ПРОТЕАЗ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2790443C2 |
| АКТИВИРУЕМЫЕ ПРОТЕАЗОЙ СВЯЗЫВАЮЩИЕ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИЕ МОЛЕКУЛЫ | 2017 |
|
RU2830288C2 |
| ВАРИАНТЫ ПРОТЕАЗЫ КЛАДЫ APRL И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2733987C2 |
| СПОСОБ ИНГИБИРОВАНИЯ РЕТРОВИРУСНОЙ ИНФЕКЦИИ (ВАРИАНТЫ), ИНГИБИТОР ПРОТЕАЗЫ, КОДИРУЮЩАЯ ЕГО НУКЛЕИНОВАЯ КИСЛОТА И СПОСОБ РЕКОМБИНАНТНОГО ПРОДУЦИРОВАНИЯ СЕРИНОВОГО ИНГИБИТОРА | 1993 |
|
RU2126266C1 |
| МУЛЬТИПРОТЕАЗНЫЙ СПОСОБ | 2017 |
|
RU2774157C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2721274C2 |
| СПОСОБЫ ПРОИЗВОДСТВА ПРОТЕОЛИТИЧЕСКИ ПРОЦЕССИРОВАННЫХ ПОЛИПЕПТИДОВ | 2019 |
|
RU2727402C1 |
| СПОСОБЫ ПРОИЗВОДСТВА ПРОТЕОЛИТИЧЕСКИ ПРОЦЕССИРОВАННЫХ ПОЛИПЕПТИДОВ | 2018 |
|
RU2719164C1 |


























Изобретение относится к способу выявления соединения, являющегося неконкурентным ингибитором протеазы вируса иммунодефицита человека (ВИЧ) (SEQ ID No: 1), предусматривающему выявление методами аланинового скинирования и молекулярного докинга соединения, по меньшей мере, с одним атомом аминокислотного остатка, выбранного из группы, состоящей из Asn98, Phe99, Asp29, Asp30, Arg8, Gly49, Gly51 и Gly52 SEQ ID NO: 1, на расстоянии не более 5 ангстрем. 3 з.п. ф-лы, 2 ил., 4 табл.
1. Способ выявления соединения, являющегося неконкурентным ингибитором протеазы вируса иммунодефицита человека (ВИЧ) (SEQ ID NO: 1), предусматривающий выявление методами аланинового скинирования и молекулярного докинга соединения, по меньшей мере, один атом которого контактирует, по меньшей мере, с одним атомом аминокислотного остатка, выбранного из группы, состоящей из Asn98, Phe99, Asp29, Asp30, Arg8, Gly49, Gly51 и Gly52 SEQ ID NO: 1, на расстоянии не более 5 Å.
2. Способ по п.1, в соответствии с которым аланиновый скрининг проводится средствами программы CHARMM с использованием процедуры молекулярной динамики.
3. Способ по п.1, в соответствии с которым собственное время симуляции после уравновешивания составляет 10 пс (шаг 1 фс) с записью промежуточных состояний через каждые 10 шагов, а для каждого промежуточного состояния проводится последовательная виртуальная замена каждого аминокислотного остатка субъединицы протеазы на аланин, в ходе каждой из которых проводится оценка изменений внутренней энергии комплекса субъединиц (в ккал/моль) по формуле:
,
где Eint - энергия взаимодействия между субъединицами, EAB - энергия димера ЕА, ЕВ - энергия каждой из цепей.
4. Способ по п.1, в соответствии с которым в процедуре докинга в качестве посадочного места используются зоны на поверхности одной из субъединиц димера, определенные в результате аланинового сканирования, а именно: Asn98, Phe99, Asp29, Asp30, Arg8, Gly49, Gly51 и Gly52 SEQ ID NO: 1.
| WO 2002014271 A1, 21.02.2002 | |||
| WO 2005097800 A1, 20.01.2005 | |||
| RU 2005139393 A, 27.07.2006 | |||
| Fakhfakh MA, Fournet A, Prina E, Mouscadet JF, Franck X, Hocqueniiller R, Figadere B | |||
| Synthesis and biological evaluation of substituted quinolines: potential treatment of protozoal and retroviral co-infections | |||
| Bioorganic and Medicinal Chemistry | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |

