Изобретение касается новых белков, обладающих эстеразной активностью, в особенности бутинол I эстеразной активностью, получаемых из Pseudomonas glumae, а также кодирующих их нуклеотидных последовательностей, экспрессионных блоков, векторов и рекомбинантных микроорганизмов; изобретение также касается технологии производства этих белков и их использования для ферментативного, в особенности энантио-селективного ферментативного гидролиза эфиров или переэтерификации органических эфиров.
Эстеразы и липазы принадлежат к классу гидролаз, которые могут использоваться при синтезе оптически активных, органических соединений в технических процессах и отличаются высокой специфичностью к субстрату. Они могут переносить ацильные группы на нуклеофилы, например на карбонильные группы, по механизму, аналогичному действию сериновой протеазы, или гидролитически расщеплять эфирные связи. Для каталитической триады эстераз, липаз и сериновых протеаз характерен общий участок последовательности из аминокислот Ser, His и Asp, причем активный Ser атакует карбонильный атом углерода, а две другие аминокислоты участвуют в распределении заряда. Эстеразы и липазы могут также переносить ацильные группы на другие нуклеофилы, например, на тио-группы тио-эфиров или на активированные амины.
Липазы осуществляют гидролиз длинноцепочечных глицериновых эфиров и для них характерна активизация на границе раздела фаз, т.е. активный центр становится доступным только в присутствии липидного субстрата. Липазы сохраняют стабильность в неводных органических растворителях и используются в многочисленных технических процессах для кинетического расщепления рацемата, т.е. когда один из энантиомеров эфира преобразовывается значительно быстрее, чем другие. Это позволяет в последующем выделять его из реакционного раствора, благодаря различию в физических и химических свойствах.
Накамура (Nakamura К. et al., Tetrahedron; Asymmetry 9 (1999), 4429-4439) описывает разделение рацематов 1-алкин-3-ола путем переэтерификации с помощью коммерчески выпускаемых липаз (Amano АК, АН и PS; Amano Pharmaceuticals Co. Ltd.) в гидрофобных растворителях, причем энантио-селективность повышается с увеличением длины цепи донора ацильных групп, а стерически крупные остатки (хлорацетат, винил-бензоат) негативно влияют на реакцию. Янг (Yang Н. et al., J. Org. Chem. 64 (1999), 1709-1712) описывает энантио-селективную технологию производства оптически активных кислот путем переэтерификации с виниловыми эфирами с использованием в качестве катализатора липазы В из Candida antarctica. Этиловые эфиры приводят при этом к заметному уменьшению скорости и селективности реакции. Липаза, выделенная из Burkholderia plantarii (Pseudomonas plantarii или glumae) DSM 6535 используется для энантио-селективного ацилирования рацемических аминов с помощью этилметок-сиацетата (Balkenhohl F. et al., J. Prakt. Chem. 339 (1997), 381-384).
Эстеразы осуществляют катализ энантио-селективного образования и разрыва эфирных связей (прямая и обратная реакция). При переэтерификации с целью получения оптически активных спиртов преимущественно используются виниловые эфиры, поскольку при этом после превращения в результате таутомеризации в альдегид или кетон пропадает спиртовая функция эфира, что препятствует протеканию обратной реакции. Для эстераз, в отличие от липаз, активизация на границе раздела фаз не характерна, и, кроме того, они способны осуществлять преобразование органических соединений с более короткой длиной цепи. Из различных организмов выделены эстеразы, характеризующиеся специфичностью к различным субстратам.
Так, выделенная из Pseudocardia thermophila эстераза FERM-BP-6275 используется при гидролизе оптически активных эфиров уксусной кислоты (европейская заявка на патент ЕР-А-0892044).
Эстераза, выделенная из Bacillus acidocaldarius, обладает узким спектром субстратов при гидролизе эфиров и характеризуется более низкой энантио-селективностью (Manco G. et al., Biochem. J. 332 (1998), 203-212).
Ацилаза 1, выделенная из Aspergillus, используется для получения вторичных спиртов путем переэтерификации с виниловыми эфирами в органических неполярных растворителях, причем преимущественно преобразованию подвергаются вторичные спирты с более короткими боковыми цепями (Faraldos J. et al., Synlett 4 (1997), 367-370). Из Pseudomonas fluorescens DSM 50 106 была клонирована связанная с мембраной лактонспецифическая эстераза (Khalameyzer V. et al., Appl. and Environ. Microbiol. 65(2) (1999), 477-482), а из Q-мутанта E.coli ацетил-эстераза (Peist R. et al., J. Bacteriol. 179 (1997), 7679-7686). Однако детальное исследование энантио-селективности и субстратной специфичности обоих этих эстераз не проводилось. В Rhodococcus sp. NCBM 11216 экспрессируются 4 эстеразы, с RR1 по RR4, отличающиеся различной специфичностью. При синтезе эфиров из нафтола и кислоты RR1 и RR2 преимущественно преобразуют кислоты с более короткими углеродными цепочками, в то время как RR3 и RR4 обладают специфичностью к кислотам с более длинными углеродными цепочками и стерически крупными остатками (Gudelj М. et al., J. Mol. Cat. В, Enzymatic 5 (1998), 261-266).
Пока, однако, не получены эстеразы, которые обладали бы широким спектром субстратов, характеризовались высокой энантиоселективностью и могли бы использоваться в технических процессах для получения небольших органических молекул, таких как оптически активные спирты, кислоты или эфиры с короткими углеродными цепочками. Задачей настоящего изобретения является разработка эстераз, обладающих, по меньшей мере, одним из вышеуказанных свойств.
Оказалось, что эта задача может быть успешно решена с помощью белка, обладающего бутинол I эстеразной активностью и содержащего, по меньшей мере, одну частичную аминокислотную последовательность, соответствующую последовательности номер 3, 4, 5 или 6:
a) FIETLGLERPVLVGHSLGGAIALAVGLDYPER (последовательность номер 3),
b) IALIAPLTHTETEP (последовательность номер 4),
c) GGGMMGLRPEAFYAASSDLV (последовательность номер 5)
d) AIDAIFAPEPV (последовательность номер 6)
(здесь для обозначения аминокислот используется однобуквенный код, причем первая аминокислота соответствует N-концу аминокислотной последовательности),
а также его функциональных эквивалентов, обладающих бутинол I эстеразной активностью.
Особенно перспективной при решении этой задачи является разработка бутинол I эстеразы, содержащей аминокислотную последовательность, соответствующую последовательности номер 2 или кодируемую нуклеотидной последовательностью, соответствующей последовательности номер 1, а также функциональных эквивалентов этого белка.
Вышеуказанные белки в дальнейшем для простоты будут именоваться как бутинол I эстеразы.
"Функциональными эквивалентами" или аналогами конкретно описанных полипептидов или белков в рамках настоящего изобретения называются отличные от них полипептиды или белки, обладающие при этом нужной биологической активностью, в особенности ферментативной активностью.
В рамках данного изобретения под "функциональными эквивалентами", в первую очередь, понимают мутантные белки, содержащие, по меньшей мере, одну из вышеуказанных частичных последовательностей, в которых заменена одна или несколько аминокислот, но обладающие, несмотря на это, по меньшей мере, одной из заявленных в данном изобретении биологической активностью. "Функциональные эквиваленты" включают в себя, таким образом, мутантные белки, полученные путем добавления, замены, делеции и/или вставки одной или нескольких аминокислот, причем указанные изменения могут затрагивать любой участок последовательности при условии, что мутантный белок сохраняет заявленные в данном изобретении свойства. Функциональная эквивалентность также имеет место при качественном соответствии ферментативных свойств мутантного и неизмененного полипептида, т.е. если, например, одни и те же субстраты преобразовываются этими белками с различной скоростью.
Естественно, "функциональные эквиваленты" включают в себя также полипептиды, выделенные из других организмов, а также природные варианты белка. Например, возможно обнаружение гомологичных участков при сравнении последовательностей и выявление эквивалентных ферментов в соответствие с конкретными данными, приводимыми в данном изобретении.
К "функциональным эквивалентам" относятся также фрагменты, преимущественно отдельные домены или участки последовательности, заявленных в данном изобретении полипептидов, обладающие, например, нужной биологической функцией.
К "функциональным эквивалентам" относятся, кроме того, фьюжн-белки, содержащие одну из вышеуказанных полипептидных последовательностей или ее функциональный аналог, которая соединена, по меньшей мере, с одной, функционально отличной от нее гетерологической последовательностью на функциональном N- или С-конце (т.е. так, что различные части фьюжн-белков не оказывают существенного функционального влияния друг на друга). Без ограничения общности к гетерологическим последовательностям такого рода относятся, например, сигнальные пептиды, ферменты, иммуноглобулин, поверхностные антигены, рецепторы или их лиганды.
В рамках данного изобретения к "функциональным эквивалентам" относятся также гомологи конкретно описанных полипептидов или белков. Они должны обладать, по меньшей мере, 60%, преимущественно не менее чем 75% и в особенности более чем 85% так, например, 90%, 95% или 99%, гомологией с конкретно описанными полипептидными последовательностями, рассчитанной по алгоритму Пирсона и Липмана, Proc. Natl. Acad, Sei. (USA) 85(8), 1988, 2444-2448.
Гомологи заявленных в данном изобретении белков или полипептидов могут быть получены путем мутагенеза, например с помощью точечных мутаций или укорочения белка. Понятие "гомолог", как оно используется здесь, касается различных форм белка, действующих в качестве агонистов или антагонистов белковой активности.
Гомологи заявленных в данном изобретении белков могут быть идентифицированы путем скрининга комбинаторных банков мутантов, например укороченных мутантов. Так, банк различных вариантов белков может быть получен путем комбинаторного мутагенеза на уровне нуклеиновой кислоты, например, путем ферментативного лигирования смеси синтетических олигонуклеотидов. Имеется множество методов получения банков потенциальных гомологов из вырожденных олигонуклеотидных последовательностей. Химический синтез вырожденных олигонуклеотидных последовательностей может быть проведен на аппарате для синтеза ДНК, и синтетический ген может затем лигироваться в подходящий экспрессионный вектор. Использование вырожденных генных комбинаций позволяет получать смесь всех последовательностей, кодирующих нужную комбинацию потенциальных белковых последовательностей. Методы синтеза вырожденных олигонуклеотидов хорошо известны специалистам (Z.B.Narang, S.A. (1983) Tetrahedron 39:3; Itakura et al. (1984) Annu. Rev. Biochem. 53:323; Itakura et al., (1984) Science 198:1056; Ike et al. (1983) Nucleic Acids Res. 11:477).
Кроме того, можно использовать банки фрагментов белковых кодонов с целью получения вариационной популяции белковых фрагментов для скрининга и последующей селекции гомологов заявленного в данном изобретении белка. В одном из вариантов исполнения банк фрагментов кодирующей последовательности может быть получен путем обработки двухцепочечных ПЦР-фрагментов кодирующей последовательности ДНК с помощью нуклеазы в условиях, при которых в среднем на одну молекулу приходится только один разрыв, последующего денатурирования двухцепочечной ДНК, ренатурирования ДНК с образованием двухцепочечной ДНК, которая может включать в себя смысловые/антисмысловые пары от различным образом порезанных продуктов, удаления одноцепочечных участков из новообразованных дуплексов путем обработки с помощью нуклеазы S1 и лигирования полученного банка фрагментов в экспрессионный вектор. С помощью этой методики может быть получен экспрессионный банк, содержащий кодирующие последовательности N-концевых, С-концевых и внутренних фрагментов различных размеров заявленного в данном изобретении белка.
В настоящее время существует множество методов скрининга комбинаторных банков генных продуктов, полученных посредством точечных мутаций или укорочения, а также методов скрининга банков кДНК на предмет поиска генных продуктов, обладающих заданными свойствами. Эти методики позволяют осуществлять быстрый скрининг геномных банков, полученных посредством комбинаторного мутагенеза гомологов заявленного в данном изобретении белка. Наиболее часто используемые методики скрининга больших геномных банков, позволяющие выполнять анализ с высокой производительностью, включают в себя операции по клонированию геномного банка в реплицируемых экспрессионных векторах, трансформации подходящих клеток полученным банком векторов и экспрессии комбинаторных генов в условиях, при которых выявление нужной активности облегчает выделение вектора, кодирующего ген, продукт которого был обнаружен. Технология рекурсивного совокупного мутагенеза (Recursive-Ensemble-Mutagenese, REM), позволяющая повысить частоту возникновения функциональных мутантов в банках, может использоваться в сочетании со скрининг-тестами, для идентификации гомологов (Arkin и Yourvan (1992) PNAS 89:7811-7815; Delgrave et al. (1993) Protein Engineering 6(3):327-331).
Предпочтительные функциональные эквиваленты, заявленные в данном изобретении, имеют последовательность, отличающуюся от последовательности номер 2, по крайней мере, в одном положении, причем эти изменения в аминокислотной последовательности преимущественно лишь незначительно, т.е. не более чем на ±90%, в особенности на ±50% или не более чем на ±30%, изменяют степень активности эстеразы. Эти изменения можно определить с использованием стандартных субстратов, таких, например, как бутинол-бутират в стандартизированных условиях (например, 20 мМ субстрата, 10 мМ фосфатный буфер, рН 7,4 Т=20°С).
Объектом настоящего изобретения являются в особенности такие функциональные эквиваленты, которые включают в себя, по меньшей мере, одну частичную последовательность из, по меньшей мере, 10 следующих друг за другом аминокислотных остатков из последовательности, соответствующей последовательности номер 2, и обладают вышеуказанной активностью по отношению к стандартному субстрату.
Без ограничения общности, примером такого рода частичных последовательностей являются частичные последовательности, полученные из вышеуказанных последовательностей номер 3, 4, 5 и 6.
Другие предпочтительные функциональные эквиваленты заявленных в данном изобретении эстераз включают в себя, таким образом, по меньшей мере, одну из частичных последовательностей, полученную из последовательностей номер 3, 4, 5 или 6, причем по сравнению с конкретно указанной частичной последовательностью в аминокислотной последовательности функционального эквивалента одна или несколько аминокислот заменены, удалены, вставлены или добавлены, и причем активность эстеразы отличается от активности эстеразы нативного белка (последовательность номер 2) не более чем на±90% или±50%, преимущественно не более чем на ±30%.
Заявленные в данном изобретении бутинол I эстеразы имеют преимущественно молекулярный вес в пределах от 40 до 42 кДа, преимущественно 41,3 кДа, определяемый путем SDS-электрофореза в геле. Они получаются преимущественно из Pseudomonas glumae Lu 2023 номер DSM 13176. Другие варианты штамма могут быть получены, например, из Pseudomonas glumae Lu 8093 путем селекции, в частности, в культуре, разводимой на плашках с минимальной питательной средой, где единственным доступным источником углерода служит этилфенилацетат.
Объектом настоящего изобретения являются также полинуклеотиды, кодирующие бутинол I эстеразу и нуклеотидные последовательности, соответствующие последовательности номер 1 или ее производным.
Объектом настоящего изобретения являются в особенности нуклеотидные последовательности (последовательности одно- и двухцепочечной ДНК и РНК, такие как кДНК и мРНК), кодирующие один из вышеуказанных полипептидов или белков и их функциональные эквиваленты, которые можно получить, например, с помощью искусственных аналогов нуклеотидов.
Объектом настоящего изобретения являются также выделенные молекулы нуклеиновой кислоты, кодирующие заявленные в данном изобретении полипептиды или, соответственно белки, либо их биологически активные участки, а также фрагменты нуклеотидной последовательности, которые могут быть использованы, например, в качестве зондов для гибридизации или праймеров для идентификации или амплификации заявленных в данном изобретении кодирующих нуклеотидных последовательностей.
Заявленные в данном изобретении молекулы нуклеиновой кислоты могут также содержать не транслируемые последовательности на 3'- и/или 5'-конце кодирующей области гена.
"Выделенная" молекула нуклеиновой кислоты должна быть отделена от других нуклеиновых кислот, присутствующих в природном источнике нуклеиновых кислот, а также очищена от прочего клеточного материала или культуральной питательной среды, если ДНК получают рекомбинантными методами, или быть очищена от химических реагентов и промежуточных продуктов реакции при синтезе ДНК химическим способом.
Заявленная в данном изобретении молекула нуклеиновой кислоты может быть выделена с использованием стандартных молекулярно-биологических методов и информации о ее нуклеотидной последовательности, указанной в данном изобретении. Например, кДНК может быть выделена из подходящего банка кДНК, при этом конкретно описанные полные последовательности или их участки могут использоваться в качестве зондов для гибридизации по стандартным методикам (которые описаны, в частности, в следующей литературе Sambrook J., Fritsch E.F. и Maniatis Т. Molecular Cloning: A Laboratory Manual. 2. Aufl., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989). Кроме того, молекулы нуклеиновой кислоты, в частности описанные последовательности или их участки, могут быть получены посредством полимеразной цепной реакции, причем при этом используются олигонуклеотидные праймеры, созданные на базе этой последовательности. Амплифицированные таким образом молекулы ДНК могут быть клонированы в подходящий вектор и охарактеризованы посредством анализа последовательности ДНК. Заявленные в данном изобретении олигонуклеотиды в дальнейшем могут быть получены с помощью стандартных методов синтеза, например, с помощью автоматического аппарата для синтеза ДНК.
Объектом настоящего изобретения являются, кроме того, молекулы нуклеиновой кислоты, комплементарные конкретно описанным нуклеотидным последовательностям или их отдельным участкам.
Заявленные в данном изобретении нуклеотидные последовательности позволяют получать зонды и праймеры, используемые для идентификации и/или клонирования гомологичных последовательностей в других типах клеток и организмах. Подобные зонды или, соответственно, праймеры обычно включают в себя область нуклеотидной последовательности, которая при заданных условиях способна гибридизоваться по крайней мере с последовательностью из 12, преимущественно не менее чем из 25, например из 40, 50 или 75 следующих друг за другом нуклеотидов смысловой цепи заявленной в данном изобретении нуклеотидной последовательности или соответствующей антисмысловой цепи.
Другие заявленные в данном изобретении нуклеотидные последовательности являются производными последовательности номер 1 и отличаются от нее в результате добавления, замены, вставки или делеции одного или нескольких нуклеотидов, кроме того, они кодируют полипептиды, обладающие требуемыми свойствами, в особенности, заявленной в данном изобретении эстеразной активностью в указанном выше диапазоне колебаний ферментативной активности.
Объектом настоящего изобретения являются также нуклеотидные последовательности, отличающиеся от конкретно указанных последовательностей наличием так называемых молчащих мутаций или, соответственно, мутаций не меняющих значение кодона, если, как это имеет место у некоторых организмов, таблица генетического кода в исходном организме или организме хозяина имеет свои особенности, а также встречающиеся в природе варианты этих последовательностей, такие как сплайсинговые или аллельные варианты. Объектом настоящего изобретения являются также нуклеотидные последовательности, полученные из исходной в результате консервативной замены нуклеотидов (т.е. когда в последовательности белка одна аминокислота заменяется другой аминокислотой, имеющей такой же заряд, размер, полярность, и/или растворимость).
Объектом настоящего изобретения являются также нуклеотидные последовательности, произошедшие из конкретно указанных последовательностей в результате генетического полиморфизма молекулы нуклеиновые кислоты. Эти генетические полиморфизмы между отдельными организмами могут существовать внутри популяции из-за естественной вариабельности. Эти естественные вариации обычно затрагивают от 1 до 5% нуклеотидной последовательности гена.
Кроме того, объектом настоящего изобретения являются также нуклеотидные последовательности, гибридизирующиеся с вышеуказанными кодирующими последовательностями или комплементарные им. Эти полинуклеотиды можно обнаружить, путем просмотра банков геномной или кДНК, и, при необходимости, размножить методом ПЦР с помощью подходящих праймеров и затем выделить, например, с помощью подходящих зондов. Другая возможность заключается в трансформации подходящих микроорганизмов заявленными в данном изобретении полинуклеотидами или векторами, размножении этих микроорганизмов и вместе с ними соответствующих полинуклеотидов и последующем их выделении. Кроме того, заявленные в данном изобретении полинуклеотиды также могут быть синтезированы химическим способом.
Под способностью полинуклеотидов "гибридизоваться" понимают способность поли- или олигонуклеотида при заданных условиях связываться с ближайшей комплементарной последовательностью, в то время как при этих же условиях неспецифического связывания между не комплементарными последовательностями не происходит. При этом степень комплементарности последовательностей должна составлять от 70 до 100%, преимущественно от 90 до 100%. Свойство комплементарных последовательностей специфически связываться друг с другом используется, в частности, в методике Нозерн- или Саузерн-блот анализа или при связывании праймеров в ПЦР. Обычно при этом используются олигонуклеотиды длиной 30 пар оснований. Под заданными условиями, например, в методе Нозерн-блот анализа понимают использование для элюирования неспецифически гибридизированных кДНК-зондов или олигонуклеотидов теплого, с температурой 50-70°С, преимущественно 60-65°С, промывочного раствора, например, следующего состава: 0,1×SSC-буфер плюс 0,1% SDS (20×SSC: 3М NaCl, 0,3М цитрат натрия, рН 7,0). При этом, как было указано выше, связанными друг с другом остаются только те нуклеиновые кислоты, которые обладают высокой степенью комплементарности. Условия, которые требуется задавать при использовании соответствующих методов, хорошо известны специалистам и описаны, в частности, в следующей литературе Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6.
Объектом настоящего изобретения являются также экспрессионные блоки, содержащие, по меньшей мере, один заявленный в данном изобретении полинуклеотид, оперативно связанный с регуляторными нуклеотидными последовательностями. Предпочтительными являются экспрессионные блоки, в которых с 5'-конца от заявленного в данном изобретении полинуклеотида находится промоторная последовательность, которая, таким образом, позволяет контролировать экспрессию бутинол I эстеразы. Особенно предпочтительными являются экспрессионные блоки, в которых с 3'-конца от заявленного в данном изобретении полинуклеотида находится терминаторная последовательность, а также, при необходимости, другие обычные регуляторные элементы, оперативно связанные с кодирующей последовательностью бутинол I эстеразы. Под оперативным соединением понимают последовательное расположение промотора, кодирующей последовательности, терминатора и, при необходимости, других регуляторных элементов таким образом, что каждый из регуляторных элементов может соответствующим образом выполнять свою функцию до, во время или после экспрессии кодирующей последовательности. Среди оперативно связанных последовательностей могут также присутствовать последовательности-мишени, а также усилители трансляции, энхансеры, сигналы полиаденилирования и другие регуляторные элементы. К часто используемым регуляторным элементам относятся также селекционные маркеры, репортерные гены, сигналы амплификации, точки начала репликации и другие подобные последовательности.
Помимо искусственных регуляторных последовательностей экспрессионные блоки могут также содержать природную регуляторную последовательность, расположенную перед собственным структурным геном. Посредством генетических изменений этот природный регулятор может быть при необходимости выключен, а экспрессия генов повышена или снижена. Впрочем, экспрессионный блок может иметь и более простое строение, т.е. перед структурным геном могут отсутствовать вставки дополнительных регуляторных сигналов, а природная промоторная последовательность со своими регуляторными элементами может оставаться на своем месте. Вместо удаления природной регуляторной последовательности ее можно заменить таким образом, что регуляция более не будет осуществляться, а экспрессия гена будет усилена или наоборот снижена. Нуклеотидные последовательности могут содержаться в экспрессионном блоке в одной или нескольких копиях.
В качестве промоторов для этих целей могут быть использованы: промоторы cos-, tac-, trp-, tet-, trp-tet-, lpp-, lac-, lpp-lac-, laclq-, T7-, T5-, T3-, gal-, trc-, ara-, SP6-, 1-PR- или 1-PL-, преимущественно в грамотрицательных бактериях; а также промоторы грамположительных бактерий amy и SPO2, дрожжевые промоторы ADC1, MFa, АС, Р-60, CYC1, GAPDH или промоторы растений CaMV/35S, SSU, OCS, lib4, usp, STLS1, В33, nos, а также убикитиновый или фазеолиновый промоторы. Особенно предпочтительным является использование индуцируемых промоторов, например, свето- и, в особенности, температурно-индуцируемых промоторов, таких как промотор PrPl.
В принципе могут использоваться все природные промоторы со своими регуляторными последовательностями. Кроме того, возможно также использование синтетических промоторов.
Указанные регуляторные последовательности должны обеспечивать требуемую экспрессию нуклеотидной последовательности и кодируемого ей белка. В зависимости от организма хозяина это может означать, например, что ген начинает экспрессироваться или повышает экспрессию только после индукции, либо что он сразу начинает экспрессироваться и/или повышает свою экспрессию.
Регуляторные последовательности или, соответственно, факторы влияют на экспрессию гена преимущественно позитивным образом и, тем самым, позволяют повышать или снижать ее интенсивность. Таким образом, возможно усиление регуляторных элементов преимущественно на уровне транскрипции при помощи сильных сигналов транскрипции, таких как промоторы и/или "энхансеры". Кроме того, возможно также усиление уровня трансляции, например, за счет повышения стабильности мРНК.
Получение заявленных в данном изобретении экспрессионных блоков осуществляется посредством соединения подходящего промотора с подходящей кодирующей полинуклеотидной последовательностью бутинол I эстеразы, а также терминатором или сигналом полиаденилирования. При этом используются стандартные методы получения рекомбинантной ДНК и технология клонирования, описанные в литературе, в частности, в Т. Maniatis, Е.F. Fritsch и J. Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989), а также в T.J.Silhavy, M.L.Berman и L.W.Enquist, Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1984) и в Ausubel, F.M. et al., Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley Interscience (1987).
Объектом настоящего изобретения являются также рекомбинантные векторы для трансформации эукариотического и прокариотического организма хозяина, несущие заявленный в данном изобретении полинуклеотид или заявленный в данном изобретении экспрессионный блок. Эти векторы позволяют осуществлять экспрессию бутинол I эстеразы в подходящем организме хозяина. Эти векторы хорошо известны специалистам и описаны в литературе, в частности, в "Cloning Vectors" (Pouwels P.H. et al., Hrsg, Eisevier, Amsterdam-New York-Oxford, 1985). Под векторами понимают помимо плазмид также все остальные векторы, хорошо известные специалистам, такие как фаги, вирусы, например, SV40, CMV, бакуловирусы и аденовирусы, транспозоны, IS-элементы, фазмиды, космиды, а также линейные или кольцевые ДНК. Эти векторы могут реплицироваться в организме хозяина автономно или в составе хромосомы.
С помощью заявленных в данном изобретении векторов можно получать рекомбинантные микроорганизмы, например, трансформированные, по меньшей мере, одним из заявленных в данном изобретении векторов и могущие использоваться для получения рекомбинантной эстеразы. Преимущественно вышеописанные рекомбинантные экспрессионные блоки, заявленные в данном изобретении вводятся и экспрессируются в подходящей системе организма хозяина в составе экспрессионного вектора. При этом преимущественно используются хорошо известные специалистам стандартные методы клонирования и трансфекции, позволяющие добиться экспрессии указанных нуклеиновых кислот в соответствующей экспрессионной системе. Подходящие системы описаны в литературе, в частности, в Current Protocols in Molecular Biology, F.Ausubel et al. Hrsg., Wiley Interscience, New York 1997.
В качестве организма хозяина для трансформации заявленными в данном изобретении векторами в принципе могут использоваться любые организмы, способные осуществлять экспрессию заявленных в данном изобретении полинуклеотидов, их аллельные варианты, их функциональные эквиваленты или производные. В качестве организма хозяина могут использоваться, в частности, бактерии, грибы, дрожжи, растительные или животные клетки. Предпочтительными организмами являются бактерии, такие как Gattungen Escherichia, например, Escherichia coli, Streptomyces, Bacillus или Pseudomonas, эукариотические микроорганизмы, такие как Saccharomyces cerevisiae, Aspergillus, высшие эукариотические клетки животных или растений, например, клетки Sf9 или СНО. Селекция успешно трансформированных организмов может осуществляться при помощи генов-маркеров, также содержащихся в векторе или экспрессионном блоке. В качестве подобных генов-маркеров могут использоваться гены устойчивости к антибиотикам и гены, кодирующие ферменты, способные катализировать реакции окрашивания трансформированных клеток. Эти клетки затем могут быть отобраны с помощью системы автоматической сортировки клеток. Организмы, успешно трансформированные вектором, несущим соответствующий ген устойчивости к антибиотикам, можно отобрать, выращивая их на питательной среде, содержащей соответствующий антибиотик. При селекции с помощью афинной хроматографии могут использоваться белки-маркеры, расположенные на поверхности клетки.
Таким образом, объектом настоящего изобретения являются также микроорганизмы, несущие заявленные в данном изобретении векторы, а также мутант Pseudomonas glumae, Lu 2023, под номером DSM 13176, эндогенно экспрессирующий бутинол I эстеразу.
Заявленные в данном изобретении бутинол I эстеразы характеризуются, прежде всего, тем, что они катализируют, по меньшей мере, одну из следующих реакций:
a) энантио-селективный гидролиз оптически активных эфиров формулы I
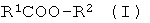
где R1 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, a R2 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, С7-С15-аралкил или содержащий одно или несколько ядер, при необходимости, однократно или многократно замещенный ароматический остаток,
R1 и/или R2 включают в себя, по меньшей мере, один асимметрический атом углерода, причем особенно предпочтительной является формула, где асимметрическим является атом углерода из R1 или, соответственно R2, соединенный с атомом углерода или с атомом кислорода сложноэфирной связью; и
b) энантио-селективная переэтерификация эфиров формулы I с помощью оптически активных спиртов формулы II
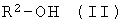
где R2 имеет указанное выше значение и, при необходимости, содержит один или несколько асимметрических атомов углерода, причем особенно предпочтительной является формула, где асимметрическим является атом углерода, имеющий группу ОН-.
Объектом настоящего изобретения являются также способ энантио-селективного гидролиза эфиров при помощи бутинол I эстеразы, причем бутинол I эстеразы приводят в контакт со смесью стереоизомеров оптически активных эфиров формулы I, и получающееся в результате стереоселективного гидролиза одного из двух стереоизомеров оптически активное соединение и/или не гидролизированный энантиомер эфира выделяют из реакционного раствора. Бутинол I эстераза может также осуществлять гидролиз подобных эфиров формулы I, которые не обладают оптической активностью.
Объектом настоящего изобретения являются также способ энантио-селективной переэтерификации, причем смесь стереоизомеров оптически активного спирта формулы II приводят в контакт с эфиром формулы I в присутствии бутинол I эстеразы, и не прореагировавший стереоизомер спирта выделяют из реакционного раствора, или смесь стереоизомеров оптически активного эфира формулы I приводят в контакт со спиртом формулы II в присутствии бутинол I эстеразы, и стереоизомер содержащегося в эфире оптически активного спирта выделяют из реакционного раствора. При переэтерификации в качестве ацилирующего средства для оптически активного спирта преимущественно используется виниловый эфир. Это позволяет предотвратить протекание обратной реакции, поскольку при этом после взаимодействия в результате таутомеризации пропадает спиртовая функция винилового эфира. Бутинол I эстераза катализирует также процессы переэтерификации, при которых ни эфир, ни спирт не обладают оптической активностью.
Предпочтительными субстратами для гидролиза эфиров являются эфиры этанола, пропанола, бутанола и особенно предпочтительны бутиниловые эфиры (бутиноловый эфир, эфир 1-метил-проп-2-инола) с карбоновыми кислотами, такими как уксусная кислота, пропионовая кислота, масляная кислота, пентановая кислота, гексановая кислота, гептановая кислота, октановая кислота, молочная кислота, 2-этилгексановая кислота, 3-метилмасляная кислота, метоксиуксусная кислота, 2-метилпропионовая кислота, 2-бутеновая кислота, 3-хлорпропионовая кислота и 2-метилпентановая кислота. Особенно предпочтительными являются бутиниловые эфиры масляной кислоты (бутинилбутират) и метилмасляной кислоты.
Предпочтительными спиртами для переэтерификации являются этанол, пропанол и бутанол, особенно предпочтительным является бутинол.
Предпочтительными эфирами при переэтерификации являются виниловые эфиры уксусной, пропионовой и масляной кислоты.
В качестве реакционного раствора в вышеуказанной методике используются такие органические растворители, как алканы, эфир, толуол, диоксан, метилизобутилкетон, метил-трет-бутиловый эфир (МТВЕ) и тому подобные вещества. При гидролизе эфиров могут также использоваться смеси из используемого буферного раствора и органических растворителей, таких как МТВЕ и гептан или толуол.
Объектом настоящего изобретения являются также полученные по указанной выше методике с использованием бутинол I эстеразы оптически активные спирты, карбоновые кислоты или эфиры.
Разделение рацематов, т.е. энантио-селективность, и скорость реакции зависят от размера и гидрофобности кислотного остатка. Реакция протекает преимущественно при комнатной температуре при значении рН от 6 до 9, особенно предпочтительным является значение рН от 7,0 до 7,4. При этом эстераза может использоваться в форме выделенного или очищенного фермента, экспрессирующих эстеразу клеток микроорганизма, надосадочной жидкости культуры этих клеток, клеточного лизата или экстракта, а также в форме иммобилизированного фермента. Продукты реакции могут быть выделены из реакционного раствора посредством химических или физических методов разделения химических веществ, которые хорошо известны специалистам. Бутинол I эстераза может быть выделена из реакционной смеси путем фильтрации через мембранный фильтр.
Иммобилизация эстеразы может осуществляться при помощи полиакриламида, альгиновой кислоты или каррагенов. Эстераза также может по известной методике ковалентно или посредством абсорбции быть связана с подходящим носителем. Иммобилизация бутинол I эстеразы преимущественно осуществляется посредством лиофилизации на кизельгуре или путем осаждения сульфатом аммония.
Как отмечалось выше, бутинол I эстераза получается из Pseudomonas glumae Lu 2023. Она, впрочем, может быть также получена по известной методике пептидного синтеза.
Кроме того, бутинол I эстераза может быть также получена из эукариотических или прокариотических организмов, экспрессирующих бутинол I эстеразы, например, из микроорганизмов, несущих заявленный в данном изобретении вектор.
Таким образом, объектом настоящего изобретения является также методика получения бутинол I эстеразы, заключающаяся в культивировании микроорганизмов, продуцирующих бутинол I эстеразу, или трансформированных одним из заявленных в данном изобретении векторов, индуцировании, при необходимости, экспрессии бутинол I эстеразы и выделении бутинол I эстеразы из клеточной культуры. Культивирование и ферментация микроорганизмов можно осуществлять по известной методике. Выращивание бактерий, например, может осуществляться на среде ТВ или LB при температуре от 20 до 40°С и значении рН от 6 до 9. Подробнее условия культивирования описаны в соответствующей литературе, например: Т. Maniatis, E.F.Fritsch and J. Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989).
После культивирования клетки вскрываются, и бутинол I эстеразу получают из лизата по методике выделения белка. Вскрытие клеток может осуществляться с помощью высокочастотного ультразвука, под высоким давлением, например в соответствующих ячейках (электролизерах), путем осмолиза, с помощью детергентов, литических ферментов или органических растворителей, в гомогенезаторах, предпочтительно в измельчителях со стеклянными шариками, или с использованием комбинации нескольких перечисленных методов. После центрифугирования белки и другие растворимые молекулы остаются в надосадочной жидкости. После высаждения ДНК хлоридом марганца получается раствор со значительно меньшей вязкостью. Белки можно селективно осаждать путем "высаливания", например, при помощи сульфата аммония или фосфата калия. Преципитация может также осуществляться путем изменения значения рН или температуры, либо с помощью органических растворителей, таких как метанол, этанол или ацетон. После преципитации в солевом растворе их можно удалить путем диализа.
Дальнейшая очистка бутинол I эстеразы может быть выполнена по известной методике хроматографической очистки, например, с помощью хроматографии через молекулярное сито (гель-фильтрация), хроматографии в Q-сефарозе, ионнообменной хроматографии и гидрофобной хроматографии, а также другими принятыми методами, такими как ультрафильтрация, кристаллизация и нативный электрофорез в геле.
Без ограничения сущности настоящее изобретение далее более подробно разъясняется на представленных примерах со ссылками на прилагаемый чертеж. При этом на чертеже для сравнения приводится заявленная в данном изобретении частичная аминокислотная последовательность бутинол I эстеразы вместе с частичной последовательностью лактонспецифической эстеразы из Pseudomonas fluorescens. Query: Частичная последовательность заявленного в данном изобретении клона LU2898. Sbjct: Частичная последовательность фермента из Pseudomonas fluorescens (Accession No.: 087637).
Пример 1
Селекция мутанта Pseudomonas glumae, экспрессирующего бутинол I эстеразу
Исходным штаммом скрининга являлся продуцирующий липазу штамм Pseudomonas (Burkholderia) glumae Lu 8093. С помощью липазы, продуцируемой этим штаммом, можно проводить целый ряд интересных реакций (Balkenhohl F. et al., J. Prakt. Chem. 339 (1997), 381-384). Однако эфиры молочной кислоты, эфиры пировинной кислоты и эфиры фенилуксусной кислоты не могут служить субстратом для липазы и не могут гидролизироваться с помощью этого штамма.
Продукты гидролиза могут, однако, служить источником углерода. Таким образом можно производить поиск мутантов Lu 8093, способных осуществлять гидролиз этих эфиров и расти, используя продукты гидролиза в качестве источника углерода. Мутанты, обладающие новой эстеразной активностью, можно, таким образом, выявлять по их способности расти на этих эфирах.
Условия селекции: клетки штамма Lu 8093 культивировались в питательной среде в течение 16 часов и затем осаждались центрифугированием. Клетки дважды промывали в солевом растворе и высевали по 106 клеток на плашках с минимальной питательной средой, содержащей 0,5 или соответственно 1,0 г/л этилфенилацетата в качестве единственного источника углерода. Сначала, однако, никакого роста не наблюдалось. Только через 4-6 дней появлялись отдельные колонии. Их количество в последующие дни увеличивалось.
Мутант Lu 2023 отбирался из таких содержащих эстеразу положительных мутантов. Оказалось, что новая эстеразная активность также может использоваться для селективного гидролиза небольших органических молекул. В примере показан селективный гидролиз бутинолового эфира.
Пример 2
Ферментация Pseudomonas glumae Lu 2023
Для получения бутинол I эстеразы клетки Pseudomonas glumae Lu 2023 культивировали в 14 л среды и высевали активную биомассу.
Клетки Pseudomonas glumae Lu 2023 наносились на плашки с агаром, с минеральной средой М12 и содержащие 1 г/л ЕРА, и инкубировались при температуре 28°С от 36 до 48 часов. Плашки затем могли храниться при температуре 4°С в течение четырех недель.
Ферментация штамма проводилась в 14-литровом ферментере Infors хху. Для получения предварительной культуры 250 мл среды заражали 2-3 каплями штамма и инкубировали, перемешивая со скоростью 200 об/мин при температуре 28°С в течение 24 часов. Основная культура получалась при следующих условиях:
Температура 28°С
Приток воздуха 7 л/мин
Скорость перемешивания 600 об/мин
Время ферментации порядка 24 ч
Встроенные датчики рН и pO2
Среда для предварительной и основной культуры
15 г/л 65%
дрожжевой автолизат Springer
1,6 г/л сульфат магния семиводный
0/02 г/л хлорид кальция двуводный
3,5 г/л дигидрофосфат калия
3,5 г/л дикалиевая соль ортофосфорной кислоты
5 г/л диаммониевая соль ортофосфорной кислоты
6 мл ингибитор пенообразования Pluriol Р2000
Указанные компоненты растворялись в воде VE и рН полученного раствора доводилось 25% раствором аммиака до значения 6,5. Следовые элементы 5 мл/л и глюкоза 2 г/л отдельно стерилизовались посредством фильтрования.
После стерилизации и дополнения среды в ферментер добавляли 0,5 г/л этилфенилацетата. Возникающая при ферментации пена подавлялась добавлением ингибитора пенообразования Pluriol Р2000. Ферментация прерывалась после того, как значение pO2 в ферментере поднималось выше 85%. Содержимое ферментере затем центрифугировали при температуре не выше 15°С и на скорости порядка 9000-10000 g, и надосадочная жидкость удалялась. Клеточная масса замораживалась при температуре -16°С.
Пример 3
Очистка бутинол I эстеразы из Pseudomonas glumae Lu 2023
Клетки (100 мл, 50 г влажной массы) Pseudomonas glumae (Lu 2023) вскрывались на измельчителе со стеклянными шариками (100 мл стеклянных шариков, диаметром 0,5 мм) при температуре 4°С и на скорости 3000 об/мин. После центрифугирования (30 мин, 10000 об/мин) и удаления стеклянных шариков к надосадочной жидкости (300 мл) добавлялся хлорид марганца (рН 7-7,5; конечной концентрации 50 мМ) и производилось осаждение. После повторного центрифугирования значение рН надосадочной жидкости доводилось до 8,0 и к ней добавлялся EDTA до концентрации 50 мМ. Эти объемы очищались посредством хроматографии на Q-сефарозе (300 мл). После загрузки образцов колонки промывались 50 мМ Tris/HCI. Нужная фракция собиралась и концентрировалась путем ультрафильтрации (100 кДа). Путем хроматографии через молекулярное сито (колонка диаметром 5 см, высотой 90 см; материал S-300) бутинол I эстераза отделялась от неспецифической эстеразы. Полученная активная фракция была мутной и повторно концентрировалась. Эстераза, очевидно, была связана с клеточной мембраной. Мембранная фракция затем обрабатывалась протеазами (трипсин, весовое соотношение от 1:50 до 1:100). При этом из мембранной фракции исчезали все белки за исключением небольшого числа, дающих несколько полос при электрофорезе в полиакриламидном геле SDS. Эстеразная активность при этом сохранялась. Полученные полосы разделялись посредством нативного электрофореза в геле (0,04% SDS) и активность в нативном геле по-прежнему указывала на присутствие эстеразы. Она элюировалась из геля и давала чистые полоски при денатурирующем электрофорезе в полиакриламидном геле SDS.
Очищенный таким образом белок переносился методом блотинга на мембрану PVDF и секвенировался, либо полученные после расщепления трипсином пептиды разделялись методом обратной фазы HPLC и затем секвенировались. Поскольку аминоконцевой конец белка был заблокирован, получались только расщепленные трипсином пептиды. Различные аминокислотные последовательности обнаруживают слабую гомологию с муконат-циклоизомеразой, ЕС 5.5.1.1, из Acinetobacter Iwoffii и Pseudomonas putida, а также с лактон-эстеразой из Pseudomonas fluorescens. Пептид с аминокислотной последовательностью AIDAIFAPEGV обнаруживает гомологию с пептин-эстеразой (ЕС 3.1.1.11).
На чертеже для сравнения приводится заявленная в данном изобретении частичная аминокислотная последовательность бутинол I эстеразы вместе с частичной последовательностью лактонспецифической эстеразы из Pseudomonas fluorescens
Пример 4
Иммобилизация бутинол I эстеразы
Для иммобилизации бутинол I эстеразы используются различные методы.
1. Бутинол I эстераза инактивируется путем осаждения ацетоном в присутствии кизельгура. 25 мг белка смешивалось с 3,5 г кизельгура (Merck) и в течение 10 минут к смеси добавлялось 1,4 л ацетона (-20°С). Загруженный носитель затем отделялся на стеклянном нутч-фильтре G3, остаток на фильтре промывался холодным ацетоном и высушивался.
2. Бутинол I эстераза не связывается с Accurel (Akzo).
3. Иммобилизация бутинол I эстеразы (2,3 ед./г, тест-ЕРА) может осуществляться посредством лиофилизации на кизельгуре. Для этого ферментативный раствор смешивался с кизельгуром и замораживался при температуре -80°С. Затем твердая фаза высушивалась посредством лиофилизации.
4. Иммобилизация бутинол I эстеразы (454 мед/г, тест-ЕРА) может осуществляться путем осаждения сульфатом аммония. Для этого фермент высаждается 80% раствором сульфата аммония в присутствии кизельгура.
Пример 5
Разделение рацематов при помощи бутинол I эстеразы из Pseudomonas glumae Lu 2023
Выполнение (стандартная исходная смесь)
В реакционный раствор, содержащий 20 ммоль бутинол-бутирата (1-метил-проп-2-иниловый эфир масляной кислоты) в фосфатном буфере (200 мл, 10 ммол, рН 7,4), при перемешивании добавляли 100 единиц бутинол I эстеразы. Значение рН непрерывно измерялось и посредством добавления раствора едкого натра поддерживалось на уровне рН 7,4. Через временные интервалы, приведенные в Таблице 1, производился отбор проб, которые дважды обрабатывались метил-трет-бутиловым эфиром (МТВЕ), после чего органический фаза анализировалась посредством GC (Chiraldex GTA). Бутинол I эстераза может быть выделена из реакционной смеси путем фильтрации через мембранный фильтр.
С увеличением процентного содержания менее предпочтительного энантиомера эфира, интенсивность его преобразования возрастала. Это приводило к тому, что примерно через 45 минут ее-значение S-бутинола в реакционной смеси начинало снижаться. Своего максимума ее-значение продукта достигало примерно через 30-40 минут, и оно составляло 84% (83-97,9%). При этом ее-значение эдукта достигало за 90 минут значения 99%. ее-значение (избыток энантиомера) определяется как процентное соотношение содержания предпочтительно преобразуемого энантиомера к содержанию менее предпочтительно преобразуемого энантиомера. Это значение в большинстве случаев характеризует оптическую чистоту раствора. Снижение значения рН в течение первых 30 минут протекало линейно. Примерно через 100 минут изменение значения рН становилось несущественным.
Остаточная активность эстеразы в водной фазе после обработки метил-трет-бутиловым эфиром составляла еще около 50%.
В Таблице 1 показана зависимость от времени соотношения энантиомеров при превращении бутинол-бутирата с помощью бутинол I эстеразы. R- и S-конфигурация определены согласно R/S-номенклатуре Кан, Прелог и Инголд, оба являются хиральными молекулами. Степень превращения определяется как процентное содержание преобразованного эфира в реакционной смеси.
Пример 6
Зависимость специфичности бутинол I эстеразы от размера и гидрофобности/заряда кислотной части эфира
Стандартная исходная смесь
В реакционный раствор, содержащий 20 ммоль бутинолового эфира в фосфатном буфере (200 мл, 10 ммол, рН 7,4), при перемешивании добавляли 100 единиц бутинол I эстеразы. Значение рН непрерывно измерялось и посредством непрерывного титрования поддерживалось на уровне рН 7,0. Отобранные пробы дважды обрабатывались метил-третичным-бутиловым эфиром (МТВЕ), после чего органический фаза анализировалась посредством GC (Chiraldex GTA).
Результат
Эффективность разделения рацематов и скорость реакции зависят от размера и гидрофобности кислотной части. Наилучшими субстратами для бутинол эстеразы являлись бутиноловые эфиры масляной и метилмасляной кислоты. Липазы не проявляют активности в отношении этих субстратов. Это относится также к длинноцепочечным эфирам, таким как n-бутиниловый эфир декановой кислоты.
В Таблице 2 показана зависимость от кислотной компоненты преобразованного эфира соотношения энантиомеров при превращении эфира с помощью бутинол I эстеразы.
Пример 7
Переэтерификация в органической среда с использованием бутинол I эстеразы
В реакционный раствор, содержащий 10 ммоль рацемической смеси бутинола и 5 ммоль винилового эфира масляной кислоты, растворенных в 50 мл метил-третичного-бутилового эфира (МТВЕ), добавляли 9 единиц нанесенной на кизельгур бутинол I эстеразы (3,3 г) и в течение 24 часов перемешивали при комнатной температуре. После фильтрации растворитель удаляли, и смесь продуктов характеризовали посредством GC.
При 47% превращении оставался (R)-бутинол (18% ее) и бутират (S)-бутинола (45% ее).
В метилизобутилкетоне при 43% превращении содержались (R)-бутинол с ее-значением 16% и бутират (в)-бутинола с ее-значением 52%.
В Таблице 3 показана зависимость от кислотной компоненты преобразованного эфира соотношения энантиомеров при превращении эфира с помощью бутинол I эстеразы.
| название | год | авторы | номер документа |
|---|---|---|---|
| БУТИНОЛ I ЭСТЕРАЗА | 2001 |
|
RU2333958C2 |
| СПОСОБЫ УСИЛЕНИЯ ДЕГРАДАЦИИ ИЛИ ПРЕВРАЩЕНИЯ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА | 2006 |
|
RU2441912C2 |
| НИТРИЛГИДРАТАЗА ИЗ Rhodococcus | 2005 |
|
RU2385932C2 |
| МИКРООРГАНИЗМ, ОБЛАДАЮЩИЙ УЛУЧШЕННОЙ СПОСОБНОСТЬЮ К ПРОДУЦИРОВАНИЮ L-ЛИЗИНА, И СПОСОБ ПРОИЗВОДСТВА L-ЛИЗИНА С ИСПОЛЬЗОВАНИЕМ ДАННОГО МИКРООРГАНИЗМА | 2015 |
|
RU2663135C2 |
| УСТОЙЧИВЫЕ К ЦИАНИДАМ НИТРИЛГИДРАТАЗЫ | 2005 |
|
RU2385876C2 |
| НОВАЯ АЛЬДОЛАЗА И СПОСОБ ПОЛУЧЕНИЯ ЗАМЕЩЕННЫХ α-КЕТОКИСЛОТ | 2003 |
|
RU2307871C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pEst877, ДЕТЕРМИНИРУЮЩАЯ ЭКСПРЕССИЮ ПОЛИПЕПТИДА С АКТИВНОСТЬЮ ЭСТЕРАЗЫ Psychrobacter cryohalolentis К5 НА ПОВЕРХНОСТИ КЛЕТОК ESCHERICHIA COLI, И ШТАММ БАКТЕРИЙ Escherichia coli BL21(DE3)pLysS/pEst877-ПРОДУЦЕНТ ПОЛИПЕПТИДА С АКТИВНОСТЬЮ ЭСТЕРАЗЫ Psychrobacter cryohalolentis К5 НА ПОВЕРХНОСТИ КЛЕТОК | 2013 |
|
RU2526213C1 |
| СИГНАЛЬНЫЕ ПЕПТИДЫ ТАТ ДЛЯ ПРОДУКЦИИ БЕЛКОВ В ПРОКАРИОТАХ | 2008 |
|
RU2487937C2 |
| НОВАЯ ЛИЗИНДЕКАРБОКСИЛАЗА И СПОСОБ ПОЛУЧЕНИЯ КАДАВЕРИНА С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2016 |
|
RU2699516C2 |
| КОМПОЗИЦИЯ И СПОСОБ ДЛЯ ПОЛУЧЕНИЯ ДУШИСТОГО СЛОЖНОГО ЭФИРА | 2008 |
|
RU2511409C2 |

Изобретение относится к биотехнологии и касается способа получения белка, обладающего бутинол I эстеразной активностью, содержащего, по меньшей мере, одну частичную аминокислотную последовательность, соответствующую последовательности номер 3, 4, 5 или 6. Согласно предложенному способу культивируют микроорганизм, эндогенно продуцирующий данный белок или трансформированный рекомбинантным вектором для трансформации эукариотического или прокариотического организма хозяина, содержащим полинуклеотид, кодирующий вышеупомянутый белок, а также комплементарные ему полинуклеотиды, и гибридизируемые с ним нуклеотидные последовательности, или экспрессионную кассету, содержащую, по меньшей мере, один вышеупомянутый полинуклеотид, оперативно связанный с, по меньшей мере, одной регуляторной нуклеотидной последовательностью, и выделяют белок из клеточной культуры. Изобретение также касается способа энантио-селективного ферментативного гидролиза эфиров и способа энантио-селективной переэтерификации с использованием полученного белка. Изобретение позволяет получить и эффективно использовать бутинол I эстеразы с высокой энантио-селективностью. 6 н. и 20 з.п. ф-лы, 1 ил., 3 табл.
1. Способ получения белка, обладающего бутинол I эстеразной активностью, содержащего, по меньшей мере, одну частичную аминокислотную последовательность, соответствующую последовательности номер 3, 4, 5 или 6, в котором культивируют микроорганизм, эндогенно продуцирующий данный белок или трансформированный рекомбинантным вектором для трансформации эукариотического или прокариотического организма-хозяина, содержащим полинуклеотид, кодирующий вышеупомянутый белок, а также комплементарные ему полинуклеотиды, и гибридизируемые с ним нуклеотидные последовательности, или экспрессионную кассету, содержащую, по меньшей мере, один вышеупомянутый полинуклеотид, оперативно связанный с, по меньшей мере, одной регуляторной нуклеотидной последовательностью, и выделяют белок из клеточной культуры.
2. Способ по п.1, отличающийся тем, что в качестве продуцирующего микроорганизма используют штамм Pseudomonas glumae Lu 2023 под номером DSM 13176.
3. Способ по п.1, отличающийся тем, что белок содержит аминокислотную последовательность, соответствующую последовательности номер 2.
4. Способ по п.1, отличающийся тем, что белок содержит полипептидную цепочку с молекулярным весом порядка 41300 Да.
5. Способ по п.1, отличающийся тем, что белок является получаемым штаммом Pseudomonas glumae Lu 2023 под номером DSM 13176.
6. Способ по п.1, отличающийся тем, что белок катализирует, по меньшей мере, одну из следующих реакций:
a) энантиоселективный гидролиз оптически активных эфиров формулы I
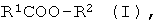
где R1 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, а R2 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, С7-С15-аралкил или содержащий одно или несколько ядер, при необходимости, однократно или многократно замещенный ароматический остаток,
R1 и/или R2 включают в себя, по меньшей мере, один асимметрический атом углерода; и
b) энантиоселективная переэтерификация эфиров формулы I с помощью оптически активных спиртов формулы II
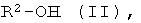
где R2 имеет указанное выше значение, и при необходимости, содержит один или несколько асимметрических атомов углерода.
7. Белок, получаемый способом по одному из пп.1 - 6.
8. Штамм Pseudomonas glumae Lu 2023 под номером DSM 13176 - продуцент белка по п.7.
9. Микроорганизм, отличающийся тем, что он несет рекомбинантный вектор для трансформации эукариотического или прокариотического организма-хозяина, содержащий полинуклеотид, кодирующий белок, обладающий бутинол I эстеразной активностью, содержащий, по меньшей мере, одну частичную аминокислотную последовательность, соответствующую последовательности номер 3, 4, 5 или 6, а также комплементарные ему полинуклеотиды, и гибридизируемые с ним нуклеотидные последовательности, или экспрессионную кассету, содержащую, по меньшей мере, один вышеупомянутый полинуклеотид оперативно связанный с, по меньшей мере, одной регуляторной нуклеотидной последовательностью.
10. Микроорганизм по п.9, отличающийся тем, что белок содержит аминокислотную последовательность, соответствующую последовательности номер 2.
11. Микроорганизм по п.9, отличающийся тем, что белок содержит полипептидную цепочку с молекулярным весом порядка 41300 Да.
12. Микроорганизм по п.9, отличающийся тем, что белок является получаемым штаммом Pseudomonas glumae Lu 2023 под номером DSM 13176.
13. Микроорганизм по п.9, отличающийся тем, что белок катализирует, по меньшей мере, одну из следующих реакций:
a) энантио-селективный гидролиз оптически активных эфиров формулы I
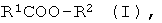
где R1 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, а R2 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, С7-С15-аралкил или содержащий одно или несколько ядер, при необходимости, однократно или многократно замещенный ароматический остаток,
R1 и/или R2 включают в себя, по меньшей мере, один асимметрический атом углерода; и
b) энантио-селективная переэтерификация эфиров формулы I с помощью оптически активных спиртов формулы II
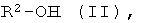
где R2 имеет указанное выше значение, и при необходимости, содержит один или несколько асимметрических атомов углерода.
14. Способ энантио-селективного гидролиза эфиров с использованием белка по п.7, обладающего бутинол I эстеразной активностью, содержащего, по меньшей мере, одну частичную аминокислотную последовательность, соответствующую последовательности номер 3, 4, 5 или 6, в котором
a) белок приводят в контакт со смесью стереоизомеров оптически активного эфира формулы I; и
b) получающиеся в результате стерео-селективного гидролиза одного из стереоизомеров оптически активные соединения и/или не гидролизированные энантиомеры эфира выделяют из реакционного раствора.
15. Способ по п.14, отличающийся тем, что в качестве реакционного раствора используют органический растворитель.
16. Способ по п.14, отличающийся тем, что белок содержит аминокислотную последовательность, соответствующую последовательности номер 2.
17. Способ по п.14, отличающийся тем, что белок содержит полипептидную цепочку с молекулярным весом порядка 41300 Да.
18. Способ по п.14, отличающийся тем, что белок производится штаммом Pseudomonas glumae Lu 2023 под номером DSM 13176.
19. Способ по любому из п.п.14-18, отличающийся тем, что белок катализирует, по меньшей мере, одну из следующих реакций:
а) энантио-селективный гидролиз оптически активных эфиров формулы I
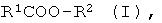
где R1 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, а R2 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, С7-С15-аралкил или содержащий одно или несколько ядер, при необходимости, однократно или многократно замещенный ароматический остаток,
R1 и/или R2 включают в себя, по меньшей мере, один асимметрический атом углерода; и
b) энантио-селективная переэтерификация эфиров формулы I с помощью оптически активных спиртов формулы II
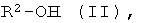
где R2 имеет указанное выше значение, и при необходимости, содержит один или несколько асимметрических атомов углерода.
20. Способ энантиоселективной переэтерификации, в котором
a) смесь стереоизомеров оптически активного спирта формулы II приводят в контакт с эфиром формулы I в присутствии белка по п.7, обладающего бутинол I эстеразной активностью, содержащего, по меньшей мере, одну частичную аминокислотную последовательность, соответствующую последовательности номер 3, 4, 5 или 6, и не прореагировавший стереоизомер спирта выделяют из реакционного раствора; или
b) смесь стереоизомеров оптически активного эфира формулы I приводят в контакт со спиртом формулы II в присутствии белка по п.7, обладающего бутинол I эстеразной активностью, содержащего, по меньшей мере, одну частичную аминокислотную последовательность, соответствующую последовательности номер 3, 4, 5 или 6, и один из стереоизомеров содержащегося в эфире оптически активного спирта выделяют из реакционного раствора.
21 Способ по п.20, отличающийся тем, что при переэтерификации в качестве ацилирующего средства для оптически активного спирта используют виниловый эфир.
22. Способ по п.20, отличающийся тем, что в качестве реакционного раствора используют органический растворитель.
23. Способ по п.20, отличающийся тем, что белок содержит аминокислотную последовательность, соответствующую последовательности номер 2.
24. Способ по п.20, отличающийся тем, что белок содержит полипептидную цепочку с молекулярным весом порядка 41300 Да.
25. Способ по п.20, отличающийся тем, что белок является получаемым штаммом Pseudomonas glumae Lu 2023 под номером DSM 13176.
26. Способ по любому из пп.20-25, отличающийся тем, что белок катализирует, по меньшей мере, одну из следующих реакций:
a) энантиоселективный гидролиз оптически активных эфиров формулы I
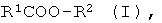
где R1 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, а R2 означает нормальный или разветвленный, при необходимости, однократно или многократно замещенный алкил с 1-10 атомами углерода, алкенил с 2-10 атомами углерода, алкинил с 2-10 атомами углерода, С7-С15-аралкил или содержащий одно или несколько ядер, при необходимости, однократно или многократно замещенный ароматический остаток,
R1 и/или R2 включают в себя, по меньшей мере, один асимметрический атом углерода; и
b) энантиоселективная переэтерификация эфиров формулы I с помощью оптически активных спиртов формулы II
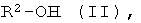
где R2 имеет указанное выше значение, и при необходимости, содержит один или несколько асимметрических атомов углерода.
| Шатун для двигателя внутреннего сгорания | 1980 |
|
SU892044A1 |
| MANCO G et | |||
| al | |||
| Overexpression and properties of a new thermophilic and thermostable esterase from Bacillus acidocaldarius with sequence similarity to hormone-sensitive lipase subfamily | |||
| Biochem | |||
| J | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| ROGALSKA E | |||
| et | |||
| al | |||
| Stereoselective hydrolysis of triglycerides by animal and microbial lipases | |||
| Chirality | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
| Способ получения бактериальной холинэстеразы | 1986 |
|
SU1472502A1 |

