Область техники, к которой относится изобретение
Настоящее изобретение в целом относится к способу измерения биологической активности диацилглицерин-ацилтрансферазы (DGAT). В частности, настоящее изобретение относится к способу быстрого массового скрининга соединений, которые способны модулировать биологическую активность DGAT. Более конкретно - настоящее изобретение относится к системе анализа для измерения активности DGAT, основанной на применении специальных мицелл по способу FlashPlate™.
Уровень техники
Триглицериды представляют собой основную форму запасания энергии у эукариот. Расстройства или нарушения равновесия в метаболизме триглицеридов вовлечены в патогенeз и повышенный риск ожирения, синдрома инсулинорезистентности и диабета II типа неалкогольной жировой болезни печени и коронарной болезни сердца (см., Lewis, et al., Endocrine Reviews (2002) 23:201 и Malloy and Kane, Adv. Intern. Med. (2001) 47:111). Кроме того, гипертриглицеридемия часто является нежелательным последствием терапии рака (см., Bast, et al. Cancer Medicine, 5th Ed., (2000) B.C. Decker, Hamilton, Ontario, CA).
Ключевым ферментом в синтезе триглицеридов является ацил-CoA:диацилглицерин-ацилтрансфераза или DGAT. DGAT представляет собой микросомальный фермент, который широко экспрессируют ткани млекопитающих и который катализирует присоединение 1,2-диацилглицерина (DAG) и жирного ацил-CoA для образования триглицеридов (TG) в эндоплазматическом ретикулуме (обзор в Chen and Farese, Trends Cardiovasc Med. (2000) 10:188 и Farese, et al., Curr. Opin. Lipidol (2000) 11:229). Первоначально полагали, что DGAT единственным образом контролирует катализ последней стадии ацилирования диацилглицерина до триглицерида в двух основных путях синтеза триглицеридов, глицеринфосфатного и моноацилглицеринового путей. Вследствие того, что триглицериды считаются необходимыми для жизни и их синтез, как полагали, осуществляется посредством единственного механизма, ингибирование синтеза триглицеридов посредством ингибирования активности DGAT в значительной степени не исследовано.
Гены, кодирующие DGAT1 мыши и родственные гомологи у человека ARGP1 и ARGP2, сейчас клонированы и охарактеризованы (Cases, et al., Proc. Natl. Acad. Sci. (1998) 95:13018; Oelkers, et al., J. Biol. Chem. (1998) 273:26765). Ген DGAT1 мыши использовали для получения мышей с нокаутом DGAT, чтобы лучше прояснить функцию гена DGAT.
Неожиданно, мыши, не способные экспрессировать функциональный фермент DGAT (мыши Dgat-/-), являются жизнеспособными и даже способными синтезировать триглицериды, что указывает на множественные каталитические механизмы, вносящие вклад в синтез триглицеридов (Smith, et al., Nature Genetics (2000) 25:87). Также идентифицировали другие ферменты, которые катализируют синтез триглицеридов, например, DGAT2 и диацилглицерин-трансацилазу (Buhman, J. Biol. Chem., выше и Cases, et al., J. Biol. Chem. (2001) 276:38870). Исследования нокаута гена у мышей выявили, что DGAT2 играет фундаментальную роль в синтезе триглицеридов у млекопитающих и необходим для жизни. Мыши с нарушенным DGAT2 обладают липопенией и умирают вскоре после рождения, очевидно, из-за значительного снижения количества субстратов для энергетического метаболизма и из-за нарушенной проницаемости в барьерной функции кожи (Farese et al. JBC (2004) 279:11767).
Существенно, мыши Dgat-/- являются устойчивыми к ожирению, вызванному питанием, и остаются худыми. Даже при кормлении пищей с высоким содержанием жира (21% жира) мыши Dgat-/- сохраняют массы тела, сравнимые с мышами, которых кормят обычной пищей (4% жира), и обладают более низкими уровнями общих триглицеридов тела. Устойчивость к ожирению у мышей Dgat-/- существует не вследствие сниженной калорийности питания, но как результат повышенного потребления энергии и пониженной устойчивости к инсулину и лептину (Smith, et al., Nature Genetics, выше; Chen and Farese, Trends Cardiovasc Med., выше; и Chen, et al., J. Clin. Invest. (2002) 109:1049). Кроме того, у мышей Dgat-/- снижались интенсивности всасывания триглицеридов (Buhman, et al., J. Biol. Chem. (2002) 277:25474). В дополнение к ускоренному метаболизму триглицеридов, мыши Dgat-/- также обладают ускоренным метаболизмом глюкозы, со сниженными уровнями глюкозы и инсулина после глюкозной нагрузки, по сравнению с мышами дикого типа (Chen and Farese, Trends Cardiovasc Med., выше).
Открытие того, что различные ферменты участвуют в катализе синтеза триглицерида из диацилглицерина, является важным, потому что это дает возможность модулировать один каталитический механизм данной биохимической реакции, чтобы достичь терапевтических результатов у индивидуума с минимальными нежелательными побочными эффектами. Соединения, которые ингибируют превращение диацилглицерина в триглицерид, например, специфическим ингибированием активности гомолога DGAT1 у человека, найдут применение в снижении концентраций в организме и всасывания триглицеридов, чтобы терапевтически противодействовать патогенным эффектам, вызванным патологическим метаболизмом триглицеридов при ожирении, синдроме инсулинорезистентности и выраженном диабете II типа, застойной сердечной недостаточности и атеросклерозе и, как последствия, терапии рака.
Вследствие все возрастающей распространенности ожирения, диабета II типа, заболевания сердца и рака во всем мире существует насущная необходимость в развитии новых терапевтических способов для эффективного лечения и профилактики данных заболеваний. Таким образом, существует заинтересованность в разработке соединений, которые могут эффективно и специфически ингибировать каталитическую активность DGAT. Однако ранее не наладили массовый скрининг для выделения специфических ингибиторов DGAT из-за технических трудностей, связанных с наладкой такого анализа.
Общепринятые анализы DGAT обладают низкими активностями порядков пмоль TG/мин/мг микросомального белка и загрязнены продуктами нескольких других ферментативных реакций. Более того, продукт реакции, катализируемой DGAT, обычно определяют анализом TLC (Cases S., et al., PNAS (1998) 95:13018; Cheng D., et al., Biochem J. (2001) 359:707; Erickson S.K., et al., J. Lipid Res. (1980) 21:930) или с использованием громоздких способов экстракции органическим растворителем (Coleman R. A., et al., Meth. Enzymology (1992) 209:98). Вследствие многочисленных стадий, предусматриваемых способами экстракции, и низкой производительности анализа TLC ни один из доступных в настоящее время анализов DGAT не пригоден в формате высокопроизводительного скрининга.
В первой попытке улучшить доступные анализы DGAT Ramharack R.R. и Spahr M.A. (EP 1219716 и US 2002/0127627) изменили способ посредством использования системы растворителей, содержащей сочетание ацетона и хлороформа. С использованием такой системы растворителей обычный способ экстракции можно упростить до способа экстракции в 1 стадию. Однако целью настоящего изобретения является дальнейшее упрощение анализа, чтобы выйти на способ, который более пригоден для высокопроизводительного скрининга посредством устранения необходимости в длительных стадиях экстракции, и предоставление анализа, который можно осуществить в формате одной лунки.
Сущность изобретения
Как отмечено выше, настоящее изобретение относится к анализу DGAT, специально приспособленному, чтобы позволить быстрый, массовый скрининг соединений, основанный на применении специальных мицелл по способу FlashPlate™.
Таким образом, в первом аспекте настоящее изобретение относится к способу измерения активности DGAT; указанный способ включает в себя: контактирование мицелл, содержащих, по меньшей мере, один субстрат DGAT, с содержащими DGAT микросомами, и определение образования триглицерида в реакционной смеси, полученной таким образом.
В особом варианте осуществления настоящего изобретения образование триглицерида определяют с использованием системы сцинтиллирующей твердой подложки, такой как, например, flashplate.
Настоящее изобретение также относится к способу определения того, способно ли тестируемое соединение модулировать активность DGAT; указанный способ включает в себя: контактирование мицелл, содержащих, по меньшей мере, один субстрат DGAT, с микросомами, содержащими DGAT, в присутствии и отсутствие тестируемого соединения и определение образования триглицерида в реакционной смеси, полученной таким образом и в которой изменение в образовании TG в присутствии тестируемого соединения указывает, что указанное соединение способно модулировать активность DGAT.
В альтернативном варианте осуществления образование триглицерида в вышеупомянутом скрининговом анализе определяют с использованием системы сцинтиллирующей твердой подложки, такой как, например, flashplate.
В особом варианте осуществления настоящего изобретения вышеупомянутые скрининговые анализы применяют для определения способности тестируемого соединения ингибировать активность DGAT, в которых уменьшение образования TG в присутствии тестируемого соединения указывает, что указанное соединение является ингибитором DGAT.
Также целью настоящего изобретения является предоставление применения мицелл, содержащих субстрат DGAT, по способу согласно изобретению.
Настоящее изобретение также относится к способам лечения или профилактики состояний и расстройств, ассоциированных с DGAT, включающим введение нуждающемуся в нем субъекту терапевтически эффективного количества соединения, выявленного способом скрининга согласно изобретению.
Описание последовательностей
SEQ ID NO:1 представляет собой нуклеотидную последовательность DGAT1 человека.
SEQ ID NO:2 представляет собой аминокислотную последовательность DGAT1 человека.
SEQ ID NO:3 представляет собой нуклеотидную последовательность DGAT2 человека.
SEQ ID NO:4 представляет собой аминокислотную последовательность DGAT2 человека.
Краткое описание чертежей
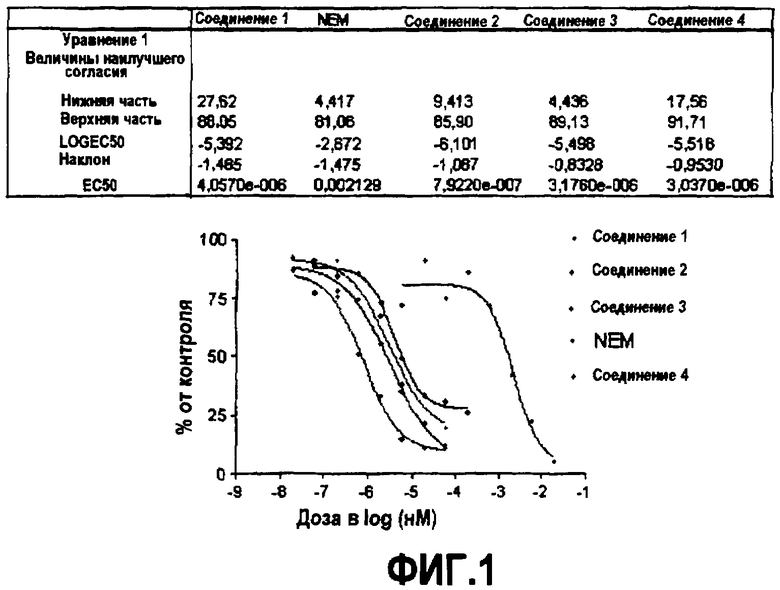
Фиг.1. Эффекты ингибиторов на активность DGAT с использованием скринингового анализа с FlashPlate™ с 384 лунками.
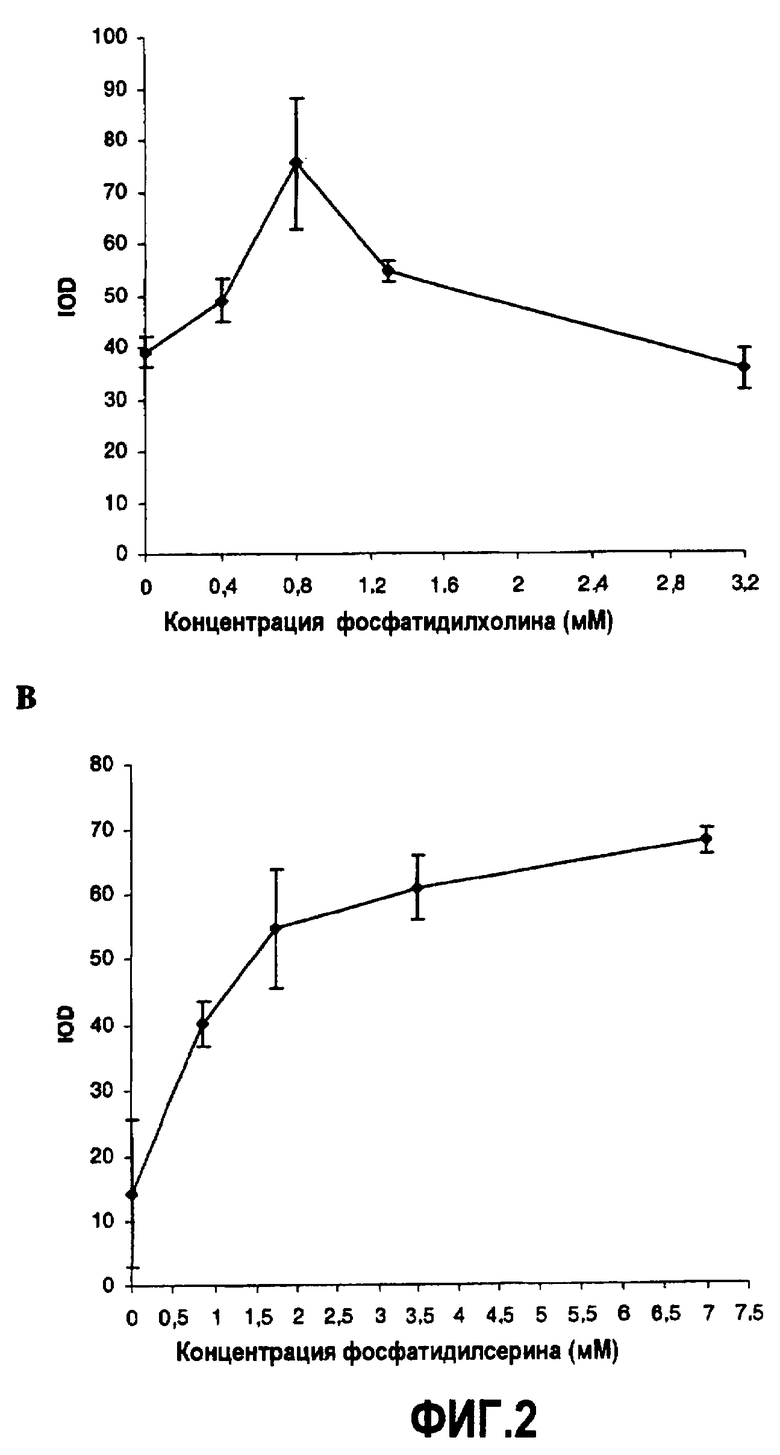
Фиг.2. Эффекты фосфатидилсерина (PS) и фосфатидилхолина (PC) в мицеллах, содержащих субстрат DGAT, на активность DGAT в скрининговом анализе с FlashPlate™. При фиксированной концентрации PS (3,5 мМ) и разных концентрациях PC (фиг.2A) и при фиксированной концентрации PC (1,3 мМ) и разных концентрациях PS (фиг.2B).
Подробное описание изобретения
Настоящее изобретение относится к способу измерения биологической активности диацилглицерин-ацетилтрансферазы (DGAT) в анализе, который позволяет быстрый и массовый скрининг способности соединений модулировать активность DGAT.
Под активностью «DGAT» подразумевают перенос жирных кислот, активированных коферментом A, в 3-положение 1,2-диацилглицеринов, формируя молекулу триглицерида.
Как применяют здесь, термин «триглицерид» (TG, триацилглицерин или нейтральный жир) относится к сложному триэфиру глицерина и жирных кислот. Триглицериды являются, как правило, неполярными и нерастворимыми в воде. Фосфоглицериды (или глицерофосфолипиды) являются основными липидными компонентами биологических мембран. Жиры и масла у животных в значительной степени содержат смеси триглицеридов.
Как применяют здесь, термин «модулировать» означает повышение или понижение функции. Предпочтительно, соединение, которое модулирует активность DGAT, модулирует ее, по меньшей мере, на 10%, более предпочтительно, по меньшей мере, на 25% и наиболее предпочтительно, по меньшей мере, на 50%, и его можно определить как «модулятор» активности DGAT.
Способ в основном включает стадии сочетания мицелл, содержащих, по меньшей мере, один субстрат DGAT, с микросомами, содержащими DGAT, инкубации таким образом полученной реакционной смеси в течение заранее установленного времени, остановку реакции и определение количества образованного TG как индикатора активности DGAT.
Мицеллы, содержащие субстрат DGAT, состоят из фосфолипидных липосом, как правило, содержащих фосфатидилсерин или фосфатидилхолин, более конкретно - содержащих фосфатидилсерин и фосфатидилхолин, предпочтительно с фосфатидилхолином в концентрации, меньшей или равной концентрации фосфатидилсерина, еще более конкретно - содержащих фосфатидилсерин и фосфатидилхолин в молярном соотношении 3:1, наиболее конкретно, содержащих фосфатидилсерин и фосфатидилхолин в молярном соотношении 3,5:1,3. Субстратами DGAT, в основном применяемыми в способах по настоящему изобретению, являются 1,2-диацилглицерин (DAG), такой как, например, 1-стеароил-2-арахидонил-sn-глицерин или 1,2-диолеоил-sn-глицерин, и жирная кислота, активированная коферментом A, такая как, например, пальмитоил-CoA или олеоил-CoA. В особом варианте осуществления настоящего изобретения мицеллы, содержащие субстрат DGAT, содержат фосфатидилсерин и фосфатидилхолин в отношении по массе 1:1 и 1,2-диолеоил-sn-глицерин в качестве субстрата DGAT. В предпочтительном варианте осуществления мицеллы состоят из фосфатидилхолина и фосфатидилсерина в концентрации 1,3 мМ и 3,5 мМ соответственно, с 1,6 мМ DAG в качестве субстрата. Указанные мицеллы, содержащие субстрат DGAT, можно получить, как, например, описано в примере 3 ниже, и хранить как запас мицелл при -20°C для дальнейшего использования.
Содержащие DGAT микросомы при применении в способах по настоящему изобретению можно получать или из систем сверхэкспрессии из клеток насекомых, или из тканевых препаратов микросом, предпочтительно источником фермента для измерений активности являются системы сверхэкспрессии из клеток насекомых.
Тканевые препараты микросом, как правило, получают из печени и кишечника, как, например, описано Coleman R. (Coleman R., Diacylglycerol acyltransferase and monoacylglycerol acyltransferase from liver and intestine. Methods in Enzymology 1992; 209:98-104).
В системах сверхэкспрессии из клеток насекомых применяют препараты мембран клеток насекомых (sf9, sf21 или клеток High Five), трансфицированных подходящим экспрессирующим вектором, таким как, например, коммерчески доступной бакуловирусной системой экспрессии Bac-to-Bac, содержащей последовательность нуклеиновой кислоты, кодирующей фермент DGAT. Препараты мембран получают с использованием способов, известных в данной области и, как правило, включающих лизирование и гомогенизирование клеток с использованием устройств для гомогенизации и собирание суммарных клеточных мембран ультрацентрифугированием. Таким образом полученные препараты мембран можно разделить на аликвоты и хранить в 10% глицерине при -80°C для дальнейшего использования.
Реакцию DGAT с ее субстратами, как правило, инициируют контактированием микросом, содержащих DGAT, с мицеллами, как определено выше, в присутствии жирной кислоты, активированной коферментом A, в частности в присутствии олеоил-CoA, где, необязательно, часть указанной жирной кислоты, активированной коферментом A, помечена для детектирования. Детектируемая метка, как применяют здесь, означает включение радиоизотопов, таких как 14C или 3H, или флуоресцентных меток, таких как, например, пирендекановая кислота. В согласии с целью изобретения является предоставление применения радиоактивно меченной или флуоресцентно меченной жирной кислоты, активированной коферментом A, в способах согласно изобретению, в частности, применение [14C]-олеоил-CoA или (1-пирен-1-ил)деканоил-CoA. В более конкретном варианте осуществления настоящего изобретения применяют [14C]-олеоил-CoA.
Реакционную смесь, как правило, инкубируют при температуре, варьирующей от комнатной температуры до 37°C в течение заранее установленного времени, такого как, например, от 5 мин до 180 мин, более конкретно - при, по меньшей мере, 23°C в течение, по меньшей мере, 15 мин, еще более конкретно -при 37°C в течение 120 мин.
Остановку реакции DGAT с ее субстратами можно осуществить добавлением ингибитора DGAT, такого как, например, N-этилмалеимид, N-(7,10-диметил-11-оксо-10,11-дигидро-дибензо[b,f][1,4]оксазепин-2-ил)-4-гидрокси-бензамид или OT-13540 (Masahiko Ikeda, Chinatsu Suzuki, Yasuhide Inoue: Effects of OT-13540, a potential antiobesity compound, on plasma triglyceride levels in experimental hypertriglyceridemia; XIIIth International Symposium on Atherosclerosis (Kyoto, Japan) Sep-Oct, 2003). Альтернативно, реакцию останавливают с использованием денатурирующего вещества, такого как щелочной раствор для остановки, содержащий этанол, т.е. 12,5% абсолютного этанола, приблизительно 10% деионизованной воды, приблизительно 2,5% 1н NaOH и приблизительно 75% раствора, содержащего приблизительно 78,4% изопропанола, приблизительно 19,6% н-гептана и приблизительно 2,0% деионизованной воды или хлороформ-метанола. В особом варианте осуществления настоящего изобретения реакцию останавливают с использованием N-этилмалеимида, N-(7,10-диметил-11-оксо-10,11-дигидро-дибензо[b,f][1,4]оксазепин-2-ил)-4-гидрокси-бензамида или OT-13540, более конкретно - с использованием N-этилмалеимида.
Нуклеиновые кислоты
Как применяют в способах по настоящему изобретению, последовательность нуклеиновой кислоты, кодирующая фермент DGAT, как подразумевают, включает в себя последовательности нуклеиновых кислот, кодирующие или DGAT1 человека (SEQ ID NO:2), или DGAT2 человека (SEQ ID NO:4), а также последовательности нуклеиновых кислот, кодирующие у другого животного, в частности у другого млекопитающего, более конкретно - у другого примата, гомологи DGAT1 и DGAT2 человека. Указанные гомологи DGAT будут, как правило, обладать, по меньшей мере, 50%, например 60%, 70%, 80%, 90%, 95% или 98% идентичности последовательности с SEQ ID NO:2 или SEQ ID NO:4. Последовательность нуклеиновой кислоты, как применяют здесь, включает ДНК (включая как геномную ДНК, так и кДНК) и РНК. Там, где нуклеиновая кислота согласно изобретению включает РНК, ссылку на последовательности, показанные в сопроводительных списках, следует интерпретировать как ссылку на эквивалентную РНК, с заменой T на U.
Нуклеиновая кислота по изобретению может являться одно- или двухцепочечной. Одноцепочечные нуклеиновые кислоты по изобретению включают в себя антисмысловые нуклеиновые кислоты. Таким образом, будет понятно, что ссылка на SEQ ID NO:1 или ее гомологи включает комплементарные последовательности, если из контекста явно не следует обратное.
Последовательность кДНК DGAT по изобретению можно клонировать с использованием способов клонирования стандартным PCR (полимеразной цепной реакцией). Это включает создание пары праймеров к 5'- и 3'-концам противоположных цепей SEQ ID NO:1 или SEQ ID NO:3, приведение праймеров в контакт с мРНК или кДНК, полученной из библиотеки кДНК млекопитающих, осуществление полимеразной цепной реакции в условиях, в которых происходит амплификация желаемой области, выделение амплифицированного фрагмента (например, очисткой реакционной смеси в агарозном геле) и получение амплифицированной ДНК. Праймеры можно сконструировать содержащим участки узнавания подходящим ферментом рестрикции так, что амплифицированную ДНК можно клонировать в подходящий вектор для клонирования.
Полинуклеотиды, которые не на 100% гомологичны последовательности SEQ ID NO:1 или SEQ ID NO:3, но которые кодируют SEQ ID NO:2 или SEQ ID NO:4, или другие полипептиды по изобретению, можно получить рядом способов.
Например, можно осуществить сайт-специфический мутагенез последовательности SEQ ID NO:1 или SEQ ID NO:3. Это пригодно там, где, например, молчащие изменения кодона необходимы для последовательностей, чтобы оптимизировать предпочтения кодонов для конкретной клетки-хозяина, в которой экспрессируются полинуклеотидные последовательности. Другие изменения последовательности могут являться желательными для того, чтобы ввести участки узнавания ферментом рестрикции или изменить свойства или функции полипептидов, кодируемых полинуклеотидами. Дополнительные изменения могут являться желательными для представления конкретных изменений кодирования, которые необходимы для обеспечения, например, консервативных замен.
Нуклеиновые кислоты по изобретению могут содержать дополнительные последовательности на 5'- или 3'-конце. Например, синтетические или природные 5'-лидерные последовательности можно присоединить к нуклеиновой кислоте, кодирующей полипептиды по изобретению. Дополнительные последовательности могут также включать 5'- или 3'-нетранслируемые области, необходимые для транскрипции нуклеиновых кислот по изобретению в конкретных клетках-хозяевах.
Кроме того, гомологи DGAT, из другого животного, в частности млекопитающего (например, крыс или кроликов), более конкретно - примата, включая мышь, можно получить и применять в способах по настоящему изобретению. Такие последовательности можно получить созданием или получением библиотек кДНК, полученных из делящихся клеток или тканей, или библиотек геномной ДНК из других видов животных и исследовать такие библиотеки посредством зондов, содержащих всю или часть SEQ ID NO:1 или SEQ ID NO:3, в условиях от средней до высокой строгости (например, 0,03 M хлорид натрия и 0,03 M цитрат натрия от, приблизительно, 50°C до, приблизительно, 60°C).
Идентичность последовательности
Процентное значение идентичности последовательностей нуклеиновых кислот и полипептидов можно вычислить с использованием коммерчески доступных алгоритмов, которые сравнивают контрольную последовательность с запрашиваемой последовательностью. Следующие программы (предоставленные National Center for Biotechnology Information) можно использовать для определения гомологий/идентичностей: BLAST, gapped BLAST, BLASTN и PSI-BLAST, которые можно использовать с параметрами по умолчанию.
Алгоритм GAP (Genetics Computer Group, Madison, WI) использует алгоритм Needleman и Wunsch для выравнивания двух полных последовательностей, который максимизирует число совпадений и минимизирует число разрывов. Как правило, используют параметры по умолчанию, со штрафом за создание разрыва = 12 и штрафом за продолжение разрыва = 4.
Другим способом определения наилучшего полного совпадения между последовательностью нуклеиновой кислоты или ее частью и запрашиваемой последовательностью является использование компьютерной программы FASTDB, основанной на алгоритме Brutlag et al. (Comp. App. Biosci., 6; 237-245 (1990)). Программа предоставляет глобальное выравнивание последовательности. Результатом указанного глобального выравнивания последовательности является процентное значение идентичности. Подходящими параметрами, используемыми в поиске последовательности ДНК посредством FASTDB для вычисления процентного значения идентичности, являются: матрица=унитарная, k-группа=4, штраф за несовпадение=1, штраф за присоединение=30, длина рандомизированной группы=0, пороговый показатель=1, штраф за разрыв=5, штраф за размер разрыва=0,05 и размер окна=500 или длина в нуклеотидных основаниях запрашиваемой последовательности, что из них является более коротким. Подходящими параметрами для вычисления процентного значения идентичности и сходства аминокислотного выравнивания являются: матрица=PAM 150, k-группа=2, штраф за несовпадение=1, штраф за присоединение=20, длина рандомизированной группы=0, пороговый показатель=1, штраф за разрыв=5, штраф за размер разрыва=0,05 и размер окна=500 или длина в нуклеотидных основаниях запрашиваемой последовательности, что из них является более коротким.
Векторы
Последовательности нуклеиновых кислот по настоящему изобретению можно встраивать в векторы, в частности экспрессирующие векторы. Вектор можно использовать для репликации нуклеиновой кислоты в совместимой клетке-хозяине. Таким образом, в дополнительном варианте осуществления изобретение относится к способу создания полинуклеотидов по изобретению посредством введения полинуклеотида по изобретению в реплицирующийся вектор, вводя вектор в совместимую клетку-хозяина и выращивая клетки-хозяева в условиях, в которых осуществима репликация вектора. Вектор можно выделить из клетки-хозяина. Подходящие клетки-хозяева описаны ниже в связи с экспрессирующими векторами.
Предпочтительно, полинуклеотид по изобретению в векторе функционально связан с контрольной последовательностью, которая способна обеспечивать экспрессию кодирующей последовательности в клетке-хозяине, т.е. вектор является экспрессирующим вектором.
Термин «функционально связанный» относится к смежному расположению, в котором описанные компоненты находятся во взаимоотношении, позволяющем им функционировать предназначенным способом. Контрольную последовательность, «функционально связанную» с кодирующей последовательностью, лигируют таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с контрольными последовательностями.
Подходящие векторы можно выбрать или сконструировать содержащими подходящие регуляторные последовательности, включая промоторные последовательности, терминаторные фрагменты, последовательности полиаденилирования, энхансерные последовательности, маркерные гены и другие последовательности в зависимости от ситуации. Векторами в соответствующих случаях могут являться плазмиды, вирусные, например, фаговые, фагемидные или бакуловирусные космиды, YAC, BAC или PAC. Векторы включают векторы для генной терапии, например, векторы, основанные на векторах из аденовируса, аденоассоциированного вируса, ретровируса (такого как HIV или MLV) или альфавируса.
Векторы можно снабдить точкой начала репликации, необязательно, промотором для экспрессии указанного полинуклеотида и, необязательно, регулятором промотора. Векторы могут содержать один или более маркерных генов для селекции, например, ген устойчивости к ампициллину в случае бактериальной плазмиды или ген устойчивости к неомицину для вектора млекопитающего. Векторы можно использовать in vitro, например, для получения РНК или использовать для трансфекции или трансформации клетки-хозяина. Вектор можно также адаптировать для использования in vivo, например, в способах генной терапии. Хорошо известны системы для клонирования и экспрессии полипептида во множестве разных клеток-хозяев. Подходящие клетки-хозяева включают бактерии, эукариотические клетки, такие как из млекопитающих и дрожжевые, и бакуловирусные системы. Линии клеток млекопитающих, доступные в данной области для экспрессии гетерологичного полипептида, включают клетки яичника китайского хомячка, клетки HeLa, эмбриональные клетки, почки хомячка, клетки COS и многие другие.
Промоторы и другие сигналы регуляции экспрессии можно выбрать совместимыми с клеткой-хозяином, для которой сконструирован экспрессирующий вектор. Например, промоторы дрожжей включают промоторы GAL4 и ADH S. cerevisiae, промоторы nmt1 и adh S. pombe. Промоторы млекопитающих включают промотор металлотионеина, который можно индуцировать в ответ на тяжелые металлы, такие как кадмий. Также можно использовать вирусные промоторы, такие как промотор большого T-антигена SV40 или аденовирусные промоторы. Все эти промоторы являются легко доступными в данной области.
Векторы могут включать другие последовательности, такие как промоторы или энхансеры для управления экспрессией вставленной нуклеиновой кислоты, последовательности нуклеиновых кислот такие, чтобы продуцировать полипептид в качестве слитого белка и/или нуклеиновую кислоту, кодирующую сигналы секреции так, чтобы продуцируемый белок в клетке-хозяине секретировать клеткой.
Векторы для продукции полипептидов по изобретению, применяемые в генной терапии, включают векторы, которые несут последовательность мини-гена по изобретению.
Дополнительные подробности см., например, в Molecular Cloning: a Laboratory Manual: 2nd edition, Sambrook et al., 1989, Cold Spring Harbor Laboratory Press. Многие известные способы и протоколы для манипуляции нуклеиновой кислотой, например, получения конструкций нуклеиновых кислот, мутагенеза, секвенирования, введения ДНК в клетки и экспрессии генов, и анализа белков подробно описаны в Current Protocols in Molecular Biology, Ausubel et al. eds., John Wiley & Sons, 1992.
Векторы можно трансформировать в подходящую клетку-хозяина, как описано выше, для обеспечения экспрессии полипептида по изобретению. Таким образом, в дополнительном аспекте изобретение относится к способу получения полипептидов согласно изобретению, который включает культивирование клетки-хозяина, трансформированной или трансфицированной экспрессирующим вектором, как описано выше, в условиях, обеспечивающих экспрессию вектором кодирующей последовательности, кодирующей полипептиды и получение экспрессируемых полипептидов. Полипептиды можно также экспрессировать в системах in vitro, таких как лизат ретикулоцитов.
Дополнительный вариант осуществления по изобретению относится к клеткам-хозяевам, трансформированным или трансфицированным векторами для репликации и экспрессии полинуклеотидов по изобретению. Клетки будут выбраны, чтобы являться совместимыми с указанным вектором, и могут, например, являться бактериальными, дрожжевыми, из насекомых или из млекопитающих. Клетки-хозяева можно культивировать в условиях для экспрессии гена так, чтобы продуцировать кодируемый полипептид. Если полипептид экспрессируется соединенным с подходящим сигнальным лидерным пептидом, он может секретироваться клеткой в культуральную среду. После продукции экспрессией полипептид можно выделить и/или очистить из клетки-хозяина и/или культуральной среды, в зависимости от случая, и далее применять по желанию, например, в составлении композиции, которая может содержать один или более дополнительных компонентов, такой как фармацевтическая композиция, которая содержит один или более фармацевтически приемлемых наполнителей, связующих веществ или носителей.
Полинуклеотиды согласно изобретению можно также вставлять в векторы, описанные выше, в антисмысловой ориентации для того, чтобы обеспечить продукцию антисмысловой РНК или рибозимов.
Препараты мембран
Специфика получения таких клеточных мембран, как применяют в способах по настоящему изобретению, может в некоторых случаях определяться природой последующего анализа, но, как правило, включает сбор целых клеток и разрушение клетки, например, обработкой ультразвуком в охлажденном во льду буфере (например, 20 мМ Трис-HCl, 1 мМ EDTA, pH 7,4 при 4°C). Полученный грубый лизат клеток затем осветляют от клеточного дебриса низкоскоростным центрифугированием, например, при 200×g в течение 5 мин при 4°C. Дальнейшее осветление и насыщение мембранами осуществляют в конце с использованием стадии высокоскоростного центрифугирования, такого как, например, 40000×g, в течение 20 мин при 4°C и полученный осадок мембран промывают суспендированием в охлажденном во льду буфере и повторяют стадию высокоскоростного центрифугирования. После заключительной промывки осадок мембран ресуспендируют в буфере для анализа. Концентрации белка определяют способом по Bradford (1976) с использованием бычьего сывороточного альбумина в качестве стандарта. Мембраны можно использовать сразу или заморозить для дальнейшего использования.
В способах по настоящему изобретению мембраны инкубируют с субстратами DGAT, как здесь описано выше, в присутствии или в отсутствие соединений, тестируемых на их способность модулировать активность DGAT. Активность DGAT определяют измерением образования TG, где указанное образование TG, как правило, определяют измерением включения радиоактивно меченного TG в мицеллы по изобретению с использованием сцинтиллирующей твердой подложки, такой как, например, по коммерчески доступному способу FlashPlate™. Данные сглаживают нелинейными кривыми с использованием GraphPad prism.
Таким образом, можно идентифицировать соединения-агонисты или антагонисты, которые модулируют активность DGAT. Особой целью настоящего изобретения является применение препаратов мембран в способах идентификации соединений, которые способны ингибировать активность DGAT, т.е. идентификации антагонистов DGAT.
Терапевтические составы
Таким образом, изобретение далее относится к новым веществам-модуляторам, в частности антагонистам, полученным в анализе согласно настоящему изобретению, и композициям, содержащим такие вещества. Вещества, которые связываются с рецептором и которые обладают активностью агониста или антагониста, можно применять в способах лечения заболеваний, патология которых характеризуется действием фермента DGAT, в частности ожирения и заболеваний, связанных с высоким уровнем триацилглицерина, и такое применение оформляет дополнительный аспект изобретения. Расстройства или нарушения равновесия метаболизма триглицеридов связаны с патогенeзом и повышенным риском ожирения, синдрома инсулинорезистентности и диабета II типа, неалкогольной жировой болезни печени и коронарной болезни сердца (см. Lewis, et al., Endocrine Reviews (2002) 23:201 и Malloy and Kane, Adv. Intern. Med. (2001) 47:111). Кроме того, гипертриглицеридемия часто является нежелательным следствием терапии рака (см. Bast, et al. Cancer Medicine, 5th Ed., (2000) B. C. Decker, Hamilton, Ontario, CA).
Настоящее изобретение также относится к способам лечения или профилактики состояния или расстройства, выбранного из группы, состоящей из ожирения, диабета, нервной анорексии, булимии, кахексии, синдрома X, метаболического синдрома, инсулинорезистентности, гипергликемии, гиперурикемии, гиперинсулинемии, гиперхолестеринемии, гиперлипидемии, дислипидемии, смешанной дислипидемии, гипертриглицеридемии, неалкогольной жировой болезни печени, атеросклероза, артериосклероза, острой сердечной недостаточности, застойной сердечной недостаточности, заболевания коронарных артерий, кардиомиопатии, инфаркта миокарда, стенокардии, гипертензии, гипотензии, инсульта, ишемии, повреждения при ишемии-реперфузии, аневризмы, рестеноза, стеноза сосудов, солидных опухолей, рака кожи, меланомы, лимфомы, рака молочной железы, рака легкого, рака ободочной и прямой кишки, рака желудка, рака пищевода, рака поджелудочной железы, рака предстательной железы, рака почки, рака печени, рака мочевого пузыря, рака шейки матки, рака матки, рака яичка и рака яичника, включая введение субъекту, нуждающемуся в нем, терапевтически эффективного количества соединения по изобретению. Для данного способа и способов, представленных ниже, соединение по изобретению, в некоторых вариантах осуществления, будут вводить в сочетании со вторым терапевтическим средством.
Средства по изобретению можно вводить в эффективном количестве. Так как многие из вышеупомянутых состояний являются хроническими и часто неизлечимыми, будет понятно, что «лечение», как подразумевают, включает достижение купирования симптомов на период времени, такой как несколько часов, дней или недель, и включает замедление прогрессирования течения заболевания.
Такие средства можно составить в композиции, содержащие средство вместе с фармацевтически приемлемым носителем или разбавителем. Средство может находиться в форме физиологически функционального производного, такого как сложный эфир или соль, такая как аддитивная соль кислоты или основная соль металла, или N- или S-оксид. Композиции можно составить для любого подходящего пути и способов введения. Фармацевтически приемлемые носители или разбавители включают те, которые используют в составах, пригодных для перорального, ректального, назального, ингаляционного, местного (включая буккальное и подъязычное), вагинального или парентерального (включая подкожное, внутримышечное, внутривенное, внутрикожное, интратекальное и эпидуральное) введения. Выбор носителя или разбавителя, конечно, будет зависеть от предполагаемого пути введения, которое может зависеть от средства и его терапевтического назначения. Составы можно удобно представлять в единичной дозированной форме и можно получить любым из способов, хорошо известных в данной области фармации. Такие способы включают стадию приведения в сочетание активного ингредиента с носителем, который состоит из одного или более вспомогательных ингредиентов. Как правило, составы получают единообразно и непосредственно приводят в сочетание активный ингредиент с жидкими носителями или мелкоизмельченными твердыми носителями или с обоими и, затем, при необходимости, формуют продукт.
Для твердых композиций можно использовать общепринятые нетоксические твердые носители, включающие, например, маннит, лактозу, целлюлозу, производные целлюлозы, крахмал, стеарат магния, сахарин натрия, тальк, глюкозу, сахарозу, карбонат магния и т.п. фармацевтической чистоты. Активное соединение, как определено выше, можно составлять в виде суппозиториев с использованием, например, полиалкиленгликолей, ацетилированных триглицеридов и т.п. в качестве носителя. Жидкие фармацевтически вводимые композиции можно, например, получить растворением, диспергированием и т.д. активного соединения, как определено выше, и, необязательно, фармацевтических адъювантов в носителе, таком как, например, вода, водно-солевой раствор декстрозы, глицерин, этанол и т.п., таким образом, формируя раствор или суспензию. По желанию, фармацевтическая композиция для введения может также содержать незначительные количества нетоксичных вспомогательных веществ, таких как увлажняющие или эмульгирующие вещества, вещества, поддерживающие pH, и т.п., например, ацетат натрия, монолаурат сорбитана, триэтаноламин, олеат триэтаноламина и т.д. Фактические способы получения таких лекарственных форм известны или будут очевидными для специалистов в данной области; например, см. Remington's Pharmaceutical Sciences, Mack Publishing Company, Easton, Pennsylvania, 15th Edition, 1975.
Композиция или состав для введения, в любом случае, будет содержать количество активного(ых) соединения(й) в количестве, эффективном для облегчения симптомов у субъекта, которого лечат.
Можно получить лекарственные формы или композиции, содержащие активный ингредиент в диапазоне от 0,25 до 95%, составленном в равновесии с нетоксичным носителем.
Для перорального введения фармацевтически приемлемую нетоксичную композицию формируют включением любого из обычно используемых наполнителей, таких как, например, маннита, лактозы, целлюлозы, производных целлюлозы, кроскармеллозы натрия, крахмала, стеарата магния, сахарина натрия, талька, глюкозы, сахарозы, карбоната магния и т.п. фармацевтической чистоты. Такие композиции принимают форму растворов, суспензий, таблеток, пилюль, капсул, порошков, составов с пролонгированным высвобождением и т.п. Такие композиции могут содержать 1-95% активного ингредиента, более предпочтительно 2-50%, наиболее предпочтительно 5-8%.
Парентеральное введение, как правило, характеризуют как инъекцию, или подкожную, внутримышечную или внутривенную. Средства для инъекций можно получить в общепринятых формах, или как жидкие растворы или суспензии, твердые формы, пригодные для растворения или суспендирования в жидкости перед инъекцией, или как эмульсии. Подходящими эксципиентами являются, например, вода, солевой раствор, декстроза, глицерин, этанол или т.п. Кроме того, по желанию, фармацевтические композиции для введения могут также содержать незначительные количества нетоксичных вспомогательных веществ, таких как увлажняющие или эмульгирующие вещества, вещества, поддерживающие pH, и т.п., такие как, например, ацетат натрия, монолаурат сорбитана, олеат триэтаноламина, триэтаноламин и т.д.
Процентное содержание активного соединения, содержащегося в таких парентеральных композициях, в высокой степени зависит от его конкретной природы так же, как активность соединения и потребности субъекта. Однако применимы процентные содержания активного ингредиента от 0,1% до 10% в растворе и будут больше, если композиция является твердым веществом, которое затем разведут до вышеупомянутых процентных содержаний. Предпочтительно, композиция будет содержать 0,2-2% активного средства в растворе.
Данное изобретение будет лучше понято посредством ссылки на экспериментальные подробности, которые следуют, но специалисты в данной области легко примут во внимание, что эти подробности только иллюстрируют изобретение, как описано более подробно в формуле изобретения, которая следует после этого. Кроме того, на всем протяжении данной заявки цитировались разные публикации. Описание данных публикаций включено таким образом в качестве ссылки в данную заявку, чтобы более подробно описать состояние данной области, которой принадлежит это изобретение.
ПРИМЕРЫ
Следующие примеры иллюстрируют изобретение. Другие варианты осуществления будут понятны специалисту в данной области в свете данных примеров.
ПРИМЕР 1
Экспрессия DGAT
DGAT, ацил-CoA:диацилглицерин-ацилтрансфераза, является ключевым ферментом в биосинтезе триглицеридов. DGAT катализирует реакцию переноса ацильного остатка с жирного ацил-CoA на диацилглицерин, формируя TAG с использованием диацилглицерина (DAG) и жирного ацил-CoA в качестве субстратов.
DGAT1 человека (SEQ ID NO:1) клонировали в вектор pFastBac, содержащий начало трансляции, маркер FLAG на N-конце, как описано в литературе, и вирусную последовательность Kozak (AAX), предшествующая ATG, для улучшения экспрессии в клетках насекомых. Так как DGAT является мембранным белком, экспрессию осуществляли, как описано в литературе (Cases, S., Smith, S.J., Zheng, Y., Myers H.M., Lear, S.R., Sande, E., Novak, S., Collins, C, Welch, C.B., Lusis, A.J., Erickson, S.K. and Farese, R.V. (1998) Proc. Natl. Acad. Sci USA 95, 13018-13023.) с использованием клеток SF9.
ПРИМЕР 2
Получение мембран с DGAT
Клетки SF9, трансфицированные 72 час, собирали центрифугированием (13000 об/мин-15 мин-4°C) и лизировали в 2x500 мл буфера для лизиса (0,1 М сахароза, 50 мМ KCl, 40 мМ KH2PO4, 30 мМ EDTA, pH 7,2). Клетки гомогенизировали в клеточном дезинтеграторе. После центрифугирования при 1380 об/мин-15 мин-4°C (SN удаляли) осадок ресуспендировали в 500 мл буфера для лизиса и собирали суммарные клеточные мембраны ультрацентрифугированием при 34000 об/мин (100000×g) в течение 60 мин (4°C). Собранные мембраны ресуспендировали в буфере для лизиса, разделяли на аликвоты и хранили в 10% глицерине при -80°C до использования.
ПРИМЕР 3
Получение мицелл
Материалы
a) 1,2-диолеоил-sn-глицерин, 10 мг/мл (DAG)
Выпаривают раствор в ацетонитриле в атмосфере азота и заново растворяют в хлороформе в конечной концентрации 10 мг/мл.
b) L-α-фосфатидилхолин, 1 мг/мл (PC)
Растворяют в хлороформе в конечной концентрации 1 мг/мл и хранят при 4°C.
c) L-α-фосфатидил-L-серин, 1 мг/мл (PS)
Растворяют в хлороформе в конечной концентрации 1 мг/мл и хранят при 4°C.
Способ
Добавляют 1 мл DGA к 10 мл PC и 10 мл PS в резервуаре из толстого стекла. Выпаривают в атмосфере азота и помещают в лед на 15 минут. Таким образом полученную суспензию заново растворяют в 10 мл Трис-HCl (10 мМ, pH 7,4) с обработкой ультразвуком во льду. Способ обработки ультразвуком состоит из циклов обработки ультразвуком по 10 секунд в бане для обработки ультразвуком с последующим охлаждением во льду и повторением этого цикла обработки ультразвуком до получения гомогенного раствора (занимает приблизительно 15 минут). Таким образом полученные мицеллы хранят при -20°C до дальнейшего использования, и они содержат DAG в конечной концентрации 1,61 мМ.
Для подтверждения оптимального соотношения по массе 1:1 фосфатидилсерина и фосфатидилхолина в мицеллах, содержащих субстрат DGAT, авторы настоящего изобретения анализировали эффект разных соотношений в анализе DGAT с FlashPlate™. Для разных сочетаний фосфатидилхолина и фосфатидилсерина готовили отдельные смеси. Аликвоты маточных растворов диолеоил-sn-глицерина (10 мг/мл), L-α-фосфатидилхолина (1 мг/мл) и L-α-фосфатидил-L-серина (1 мг/мл) в хлороформе объединяли в стеклянных флаконах и выпаривали в атмосфере азота и помещали в лед на 15'. Повторное растворение осуществляли в 10 мл Трис-HCl (10 мМ, pH 7,4) обработкой ультразвуком во льду. Аликвоты хранили при -20°C.
В первой серии экспериментов концентрацию PC изменяли, чтобы изменить соотношение PC:PS. Оптимальная концентрация в мицеллах фосфатидилхолина для активности DGAT являлась 0,8 мМ (фиг.2A) с 3,5 мМ фосфатидилсерина в мицеллах. К сожалению, данная концентрация давала нестабильные, невоспроизводимые мицеллы, указывая на то, что критическая концентрация мицелл оказалась недостигнутой. Липиды, как правило, определяют на основе свойств растворимости. Они легко растворимы в неполярных растворителях и практически нерастворимы в воде. Мерой растворимости амфипатических молекул в воде является их критическая концентрация мицелл (CMC). Ее определяют как концентрацию свободных молекул в растворе в равновесии с молекулами в агрегированной форме. Обычная моющая жидкость содержит детергенты с CMC в мМ диапазоне концентраций (3). С другой стороны, использование концентраций, больших, чем 0,8 мМ, снижало активность DGAT. Авторы настоящего изобретения сделали вывод, что в данных условиях 1,6 мМ фосфатидилхолин является оптимальным для воспроизводимого образования мицелл с приемлемой активностью DGAT.
Это подтвердили во второй серии экспериментов, в которых изменяли концентрацию PS, чтобы изменить соотношение PS:PC. Тестирование разных концентраций мицеллярного фосфатидилсерина в мицеллах, содержащих 1,6 мМ диацилглицерина, выявило хороший дозовый эффект активности DGAT вплоть до 3,5 мМ, после чего достигалась почти максимальная активность DGAT (фиг.2B). Использование меньшего количества фосфатидилсерина не только снижало активность, но также приводило, аналогично фосфатидилхолину, к менее стабильным и невоспроизводимым мицеллам. В отсутствие фосфатидилсерина почти вся активность DGAT исчезала, что указывает на то, что фосфатидилсерин важен для активности. Авторы настоящего изобретения сделали вывод, что в данных условиях 3,5 мМ фосфатидилсерин является оптимальным для воспроизводимого формирования мицелл с приемлемой активностью DGAT. Беря в расчет не только максимальную активность, но также стабильность и воспроизводимость формирования мицелл, оптимальные концентрации достигаются для фосфатидилхолина и фосфатидилсерина при 1,3 мМ и 3,5 мМ соответственно. Представляется, что в данной постановке фосфатидилсерин важен для активности DGAT и фосфатидилхолин для стабилизации и воспроизводимости мицелл.
ПРИМЕР 4
Анализ DGAT с использованием FlashPlate™
Материалы
a) Буфер для анализа
50 мМ Трис-HCl (pH 7,4), 150 мМ MgCl2, 1 мМ EDTA, 0,2% BSA.
b) 5 М N-этилмалеимид
Растворяют 5 г в конечном объеме 8 мл 100% DMSO и хранят при -20°C в аликвотах до дальнейшего использования.
c) Смесь с субстратом (для планшета на 1384 лунки = 3840 мкл)
612 мкл маточной смеси мицелл (конечная концентрация 51 мкМ)
16,6 мкл олеоил-CoA, 9,7 мМ
23 мкл [3H]-олеоил-CoA (49 Ки/ммоль, 500 мкКи/мл)
3188,4 мкл Трис, pH 7,4, 10 мМ
d) Смесь с ферментом (для планшета на 1384 лунки = 3520 мкл) (5 мкг/мл)
Добавляют 11,73 мкл маточной смеси мембран с DGAT (маточная смесь 1500 мкг/мл) к 3508 мкл буфера для анализа.
e) Смесь для остановки (для планшета на 1384 лунки = 7,68 мл) (250 мМ)
Добавляют 384 мкл N-этилмалеимида (5M) к 3,456 мл 100% DMSO и далее разводят 3,84 мл указанного раствора 3,84 мл 10% DMSO.
Способ
Активность DGAT в препаратах мембран анализировали в 50 мМ Трис-HCl (pH 7,4), 150 мМ MgCl2, 1 мМ EDTA и 0,2% BSA, содержащем 50 мкМ DAG, 32 мкг/мл PC/PS и 8,4 мкМ [3H]-олеоил-CoA (при удельной активности 30 нКи/лунка) в конечном объеме 50 мкл в формате для 384 лунок с использованием Basic Image FlashPlate™ со сдвигом в красную область (Perkin Elmer Кат. № SMP400).
Подробно, 10 мкл смеси с ферментом и 10 мкл смеси с субстратом добавляли к 30 мкл буфера для анализа, необязательно, в присутствии 1 мкл DMSO (холостая проба и контроли) или 1 мкл тестируемых соединений. Данную реакционную смесь инкубировали 120 минут при 37°C и ферментативную реакцию останавливали добавлением 20 мкл смеси для остановки. Планшеты запечатывали и позволяли везикулам осесть в течение ночи при комнатной температуре. Планшеты центрифугировали 5 минут при 1500 об/мин и измеряли в Leadseeker.
Обсуждение
На данный момент фактически нет коммерчески доступного высокопроизводительного совместимого анализа, вероятно, вследствие того, что в традиционных ферментативных анализах используют препараты везикул, чтобы имитировать естественное окружение фермента, где он встроен в мембрану.
Традиционно необходимы разделение посредством TLC или экстракция растворителем для отделения радиоактивно меченного DAG или ацил-COA от образованного радиоактивно меченного TG. Данная дополнительная стадия обработки перед измерением образованного радиоактивно меченного TG делает данные традиционные способы менее пригодными для высокопроизводительного скрининга, где каждая стадия не только увеличивает продолжительность цикла, но может также нарушать воспроизводимость и последовательный вывод данных анализа.
Скрининг активности DGAT по настоящему изобретению по-прежнему имитирует естественное окружение фермента, так как в нем применяют как препараты мембран, содержащие DGAT, так и мицеллы, содержащие субстрат DGAT, но особенно приспособлен к массовому скринингу активности DGAT, так как он является способом на основе одной лунки, что устраняет необходимость отделения образованного радиоактивно меченного TG от радиоактивно меченного ацил-COA. Данный формат скрининга на основе одной лунки достижим, так как наблюдаемая радиолюминесценция происходит только от образованного радиоактивного триацилглицерина, который вступает в непосредственную близость с поверхностью flashplate, в отличие от радиоактивно меченного ацил-CoA, который остается в растворе.
В заключение настоящее изобретение относится к способу, который более пригоден для высокопроизводительного скрининга посредством устранения необходимости в продолжительных стадиях TLC и экстракции и обеспечивает более воспроизводимые и надежные результаты.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ЛИПИДОВ | 2012 |
|
RU2804627C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЛИПИДОВ | 2012 |
|
RU2656996C2 |
| Способы получения липидов | 2011 |
|
RU2636344C2 |
| ФЕРМЕНТАТИВНАЯ ОБРАБОТКА МАСЕЛ | 2004 |
|
RU2377307C2 |
| НОВАЯ ЛИЗОФОСФОЛИПИД-АЦИЛТРАНСФЕРАЗА | 2010 |
|
RU2534559C2 |
| ГЕНЫ ДИАЦИЛГЛИЦЕРОЛ-АЦИЛТРАНСФЕРАЗЫ И ИХ ИСПОЛЬЗОВАНИЕ | 2010 |
|
RU2514655C2 |
| СПОСОБЫ РАФИНИРОВАНИЯ МАСЛА | 2010 |
|
RU2573916C2 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИНЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ В ТРАНСГЕННЫХ РАСТЕНИЯХ | 2005 |
|
RU2449007C2 |
| ВАРИАНТЫ ИЗОПРЕНСИНТАЗЫ, ПРИМЕНЯЕМЫЕ ДЛЯ УЛУЧШЕНИЯ ПРОДУЦИРОВАНИЯ ИЗОПРЕНА МИКРООРГАНИЗМАМИ | 2009 |
|
RU2516343C2 |








Изобретение относится к биотехнологии и представляет собой способ измерения активности DGAT. Способ включает контактирование стабильных и воспроизводимых мицелл, содержащих, по меньшей мере, один субстрат DGAT, с микросомами, содержащими DGAT, и определение образования триглицерида в таким образом полученной реакционной смеси, при условии, что мицеллы содержат фосфатидилсерин и фосфатидилхолин. Изобретение относится также к способу идентификации того, способно ли тестируемое соединение модулировать активность DGAT, причем указанный способ включает контактирование стабильных и воспроизводимых мицелл, содержащих, по меньшей мере, один субстрат DGAT, с микросомами, содержащими DGAT, в присутствии и отсутствии тестируемого соединения, и определение образования триглицерида в таким образом полученных реакционных смесях, и в котором изменение в образовании триглицерида в присутствии тестируемого соединения указывает на то, что указанное соединение способно модулировать активность DGAT, при условии, что мицеллы содержат фосфатидилсерин и фосфатидилхолин. Изобретение позволяет быстро, просто и эффективно определить активность DGAT. 2 н. и 24 з.п. ф-лы, 2 ил.
1. Способ измерения активности DGAT, где указанный способ включает: контактирование стабильных и воспроизводимых мицелл, содержащих, по меньшей мере, один субстрат DGAT, с микросомами, содержащими DGAT, и определение образования триглицерида в таким образом полученной реакционной смеси, при условии, что мицеллы содержат фосфатидилсерин и фосфатидилхолин.
2. Способ по п.1, в котором мицеллы содержат фосфатидилхолин в концентрации, которая меньше или равна концентрации фосфатидилсерина.
3. Способ по п.1, в котором мицеллы содержат фосфатидилсерин и фосфатидилхолин в молярном соотношении 3:1.
4. Способ по п.1, в котором мицеллы содержат фосфатидилсерин и фосфатидилхолин в молярном соотношении 3,5:1,3.
5. Способ по п.1, в котором мицеллы содержат фосфатидилсерин и фосфатидилхолин в весовом соотношении 1:1.
6. Способ по п.1, в котором реакционная смесь дополнительно содержит жирную кислоту, активированную коферментом А.
7. Способ по п.6, в котором жирная кислота, активированная коферментом А, выбрана из пальмитоил-СоА или олеоил-СоА.
8. Способ по п.6, в котором часть указанной жирной кислоты, активированной коферментом А, помечена для детектирования.
9. Способ по п.8, в котором часть указанной жирной кислоты, активированной коферментом А, радиоактивно мечена.
10. Способ по любому из пп.6-9, в котором жирная кислота, активированная коферментом А, является олеоил-СоА, и часть указанного олеоил-СоА является [3H]-олеоил-СоА.
11. Способ по п.1, в котором субстрат DGAT состоит из стеароил-2-арахидонил-sn-глицерина или 1,2-диолеоил-sn-глицерина.
12. Способ по п.1, в котором микросомы, содержащие DGAT, являются препаратом мембран клеток насекомых, экспрессирующих белок DGAT1 человека (SEQ ID NO:2).
13. Способ по п.1, в котором образование триглицерида определяют с применением сцинтиллирующей твердой подложки.
14. Способ идентификации того, способно ли тестируемое соединение модулировать активность DGAT, где указанный способ включает:
контактирование стабильных и воспроизводимых мицелл, содержащих, по меньшей мере, один субстрат DGAT, с микросомами, содержащими DGAT, в присутствии и отсутствии тестируемого соединения, и определение образования триглицерида в таким образом полученных реакционных смесях, и в котором изменение в образовании триглицерида в присутствии тестируемого соединения указывает на то, что указанное соединение способно модулировать активность DGAT, при условии, что мицеллы содержат фосфатидилсерин и фосфатидилхолин.
15. Способ по п.14, в котором мицеллы содержат фосфатидилхолин в концентрации, которая меньше или равна концентрации фосфатидилсерина.
16. Способ по п.14, в котором мицеллы содержат фосфатидилсерин и фосфатидилхолин в молярном соотношении 3:1.
17. Способ по п.14, в котором мицеллы содержат фосфатидилсерин и фосфатидилхолин в молярном соотношении 3,5:1,3.
18. Способ по п.14, в котором мицеллы содержат фосфатидилсерин и фосфатидилхолин в весовом соотношении 1:1.
19. Способ по п.14, в котором реакционная смесь дополнительно содержит жирную кислоту, активированную коферментом А.
20. Способ по п.19, в котором жирная кислота, активированная коферментом А, выбрана из пальмитоил-СоА или олеоил-СоА.
21. Способ по п.19, в котором часть указанной жирной кислоты, активированной коферментом А, помечена для детектирования.
22. Способ по п.21, в котором часть указанной жирной кислоты, активированной коферментом А, радиоактивно мечена.
23. Способ по любому из пп.19-22, в котором жирная кислота, активированная коферментом А, является олеоил-СоА, и часть указанного олеоил-СоА является [3H]-олеоил-СоА.
24. Способ по п.14, в котором субстрат DGAT состоит из стеароил-2-арахидонил-sn-глицерина или 1,2-диолеоил-sn-глицерина.
25. Способ по п.14, в котором микросомы, содержащие DGAT, являются препаратом мембран клеток насекомых, экспрессирующих белок DGAT1 человека (SEQ ID NO:2).
26. Способ по п.14, в котором образование триглицерида определяют с применением сцинтиллирующей твердой подложки.
| Аванкамера насосной станции | 1984 |
|
SU1219716A1 |
| ZHU D et.al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| abstract | |||
| WATERMAN I.J | |||
| et.al | |||
| Distinct ontogenic patterns of overt and latent DGAT activities of rat liver microsomes | |||
| J Lipid Res. | |||

