Изобретение относится к медицине, конкретно к онкологии, и касается веществ, обладающих способностью предотвращать злокачественное перерождение нормальных клеток млекопитающих в опухолевые.
Проблема опухолевых заболеваний является одной из острейших проблем современного здравоохранения. Рак (злокачественные опухолевые заболевания) является одной из основных причин смерти в мире. По данным ВОЗ в настоящее время на онкологию приходится 13% от общего числа смертей. За последние 25 лет заболеваемость раком выросла в 1,5-2,0 раза, а к 2030 году по прогнозам вырастет еще втрое, причем особенно тревожно положение в развитых странах. Для различных разновидностей рака характерна одна общая черта - эти болезни чрезвычайно трудно излечить. Следует признать, что лечение онкологических заболеваний в настоящее время высокозатратно и сравнительно малоэффективно. В то же время считается, что до 40% случаев заболевания раком можно предотвратить с помощью здорового рациона питания, физической активности и воздержания от употребления табака. Профилактика может включать употребление различных продуктов и биопрепаратов, содержащих вещества, предотвращающие перерождение нормальных клеток в раковые.
Противоопухолевые хемопревентивные вещества - это обычно природные вторичные метаболиты или их синтетические аналоги, которые ингибируют трансформацию нормальных клеток в прораковые или тормозят прогрессию прораковых клеток в раковые [Hong W.K., Sporn M.B. (1997) Recent advances in chemoprevention of cancer. Science 278:1073-1077; Sporn M.B. (1976) Approaches to prevention of epithelial cancer during the preneoplastic period. Cancer Res 36:2699-2702; Umar A., Viner J.L., Hawk E.T. (2001) The future of colon cancer prevention. Ann NY Acad Sci 952:88-108].
Следовательно, эффективное канцерпревентивное вещество должно вклиниваться в процесс канцерогенеза с тем, чтобы уничтожить прораковые клетки до того, как они трансформируются в раковые [Wattenberg L.W. (1995) What are the critical attributes for cancer chemopreventive agents? Ann NY Acad Sci 768:73-81; Smith T.J., Hong J-Y, Wang Z-Y, Yang C.S. (1995) How can carcinogenesis be inhibited? Ann NY Acad Sci 768:82-90; Kelloff G.J., Crowell J.A., Steele V.E., Lubet R.A., Boone C.W., Malone W.A., et al. (1999) Progress in cancer chemoprevention. Ann NY Acad Sci 889:1-13].
Некоторые противораковые хемопревентивные вещества, к примеру, ретиноиды и антиэстрогены оказывают цито статическое действие на трансформированные клетки путем воздействия на клеточную пролиферацию или дифференциацию [Hong W.K., Sporn M.B. (1997) Recent advances in chemoprevention of cancer. Science 278:1073-1077; Wattenberg LW (1995) What are the critical attributes for cancer chemopreventive agents? Ann NY Acad Sci 768:73-81; Kelloff G.J., Crowell J.A., Steele V.E., Lubet R.A., Boone C.W., Malone W.A., et al. (1999) Progress in cancer chemoprevention. Ann NY Acad Sci 889:1-13]. Такие вещества должны в течение долгого времени вводиться в организм людей, подверженных повышенному риску раковых заболеваний. В этом случае токсичность веществ, в течение долгого времени вводимых в организм, а также возможность развития резистентности к ним, могут ограничить возможность использования данных хемопревентивных веществ [Wattenberg L.W. (1995) What are the critical attributes for cancer chemopreventive agents? Ann NY Acad Sci 768:73-81]. Альтернативный подход заключается в поиске веществ, которые быстро удаляют прораковые клетки путем индукции в них апоптоза.
Публикуются данные о все увеличивающемся числе хемопревентивных противораковых веществ, которые стимулируют апоптоз прораковых и раковых клеток in vitro и in vivo [Gupta S., Hastak K, Ahmad N, Lewin J.S., Mukhtar H. (2001) Inhibition of prostate carcinogenesis in TRAMP mice by oral infusion of green tea polyphenols. Proc Natl Acad Sci USA 98:10350-10355; Lu Y-P, Lou Y-R, Li X-H, Xie J-G, Brash D., Huang M-T, et al. (2000) Stimulatory effect of oral administration of green tea or caffeine on ultraviolet light-induced increases in epidermal wild-type p53, p21 (WAFI/CIPI), and apoptotic sunburn cells in SKH-1 mice. Cancer Res 60:4785-4791; Lu Y-P, Lou Y-R, Li X-H, Xie J-G, Yen P., Huang M-T, Conney A.H. (1997) Inhibitory effect of black tea on the growth of established skin tumors in mice: effects on tumor size, apoptosis, mitosis and bromodeoxyuridine incorporation into DNA. Carcinogenesis 18:2163-2169; Samaha H.S., Kelloff G.J., Steele V., Rao C.V., Reddy B.S. (1997) Modulation of apoptosis by sulindac, curcumin, phenylethyl-3-methylcaffeate and 6-phenylhexyl isothiocyanate apoptotic index as a biomarker in colon cancer chemoprevention and promotion. Cancer Res 57:1301-1305; Yang K, Lamprecht SA, Liu Y, Shinozaki H, Fan K, Leung D, et al. (2000) Chemoprevention studies on the flavonoids quercetin and rutin in normal and azoxymethane-treated mouse colon. Carcinogenesis 21:1655-1660; Tanaka Т., Kohno H., Sakata K., Yamada Y., Hirose Y., Sugie S., et al. (2002) Modifying effects of dietary capsaicin and rotenone on 4-nitroquinoline-1-oxide-induced rat tongue carcinogenesis. Carcinogenesis 23:1361-1367]. При использовании такого подхода долговременный прием хемопревентивных веществ становится ненужным, что снижает риск проявления токсического эффекта данных препаратов или развития резистентности по отношению к ним [Wattenberg L.W. (1995) What are the critical attributes for cancer chemopreventive agents? Ann NY Acad Sci 768:73-81; Smith T.J., Hong J-Y, Wang Z-Y, Yang C-S (1995) How can carcinogenesis be inhibited? Ann NY Acad Sci 768:82-90; Kelloff GJ, Crowell J.A., Steele V.E., Lubet R.A., Boone C.W., Malone W.A., et al. (1999) Progress in cancer chemoprevention. Ann NY Acad Sci 889:1-13].
Широко известными хемопревентивными веществами, предотвращающими перерождение нормальных клеток в опухолевые, являются полифенолы из зеленого чая, флавоноиды из ягод, ресвератрол из красного винограда, капсаицин из перца, куркумин из тропического растения куркума и многие другие [Lu Y-P, Lou Y-R, Li XH, Xie J-G, Brash D., Huang M-T, et al. (2000) Stimulatory effect of oral administration of green tea or caffeine on ultraviolet light-induced increases in epidermal wild-type p53, p21 (WAFI/CIPI), and apoptotic sunburn cells in SKH-1 mice. Cancer Res 60:4785-4791; Samaha H.S., Kelloff G.J., Steele V., Rao C.V., Reddy B.S. (1997) Modulation of apoptosis by sulindac, curcumin, phenylethyl-3-methylcaffeate and 6-phenylhexyl isothiocyanate apoptotic index as a biomarker in colon cancer chemoprevention and promotion. Cancer Res 57:1301-1305; Yang K., Lamprecht S.A., Liu Y., Shinozald H., Fan K., Leung D., et al. (2000) Chemoprevention studies on the flavonoids quercetin and rutin in normal and azoxymethane-treated mouse colon. Carcinogenesis 21:1655-1660; Tanaka Т., Kohno H., Sakata K., Yamada Y., Hirose Y, Sugie S., et al. (2002) Modifying effects of dietary capsaicin and rotenone on 4-nitroqumoline-1-oxide-induced rat tongue carcinogenesis. Carcinogenesis 23:1361-1367].
В качестве веществ, предотвращающих злокачественное перерождение нормальных клеток, в последнее время все большее внимание привлекают морские природные соединения и их синтетические аналоги и производные, которые способны вызывать апоптоз трансформированных и раковых клеток.
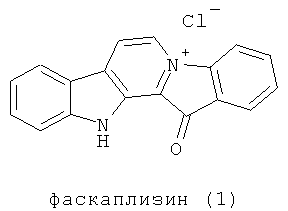
Впервые фаскаплизин (1) был выделен в 1988 году из морской губки Fascaplysinopsis Bergquist sp., собранной в южной части Тихого Океана близ островов Фиджи. Было показано, что этот красный пигмент является пентациклическим алкалоидом (1). Его структура, представляющая собой плоскую сопряженную систему, была установлена с помощью спектральных методов и рентгеноструктурного анализа [Hörmann A., Chaudhuri В., Fretz H. DNA binding of the marine sponge pigment fascaplysin // Bioorg. Med. Chem. 2001. Vol.9. P.917; Roll D.M., Ireland C.M., Lu H.S. M., Clardy J. Fascaplysin, an unusual antimicrobial pigment from the marine sponge Fascaplysinopsis sp. // J. Org. Chem. 1988. Vol.53. P.3276-3278].
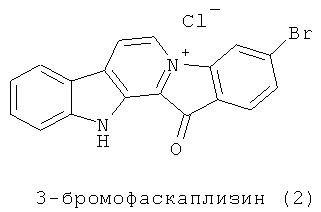
3-Бромофаскаплизин (2) впервые был выделен в 2003 году из асцидии Didemnum sp. [Segraves N.L., Lopez S., Johnson T.A., Said S.A., Fu X., Schmitz F.J., Pietraszkiewicz H., Valeriote F.A., Crews P. Structures and cytotoxicities of fascaplysin and related alkaloids from two marine phyla - Fascaplysinopsis sponges and Didemnum tunicates // Tetrahedron Lett. 2003. Vol.44. P.3471-3475]. Подобные соединения, как оказалось позже, встречаются довольно часто среди метаболитов морских губок. Так, в 2004 году было сообщено о выделении 12 новых и 7 ранее известных алкалоидов фаскаплизинового типа.
Алкалоиды фаскаплизинового типа обладают разносторонней биологической активностью и, в частности, привлекают внимание ученых своими противоопухолевыми свойствами. Лучше всего изучена биологическая активность фаскаплизина. Выяснилось, что он обладает очень широким спектром действия, включающим противобактериальную (против Staphylococcus aureus, Escherichia coli, Candida albicans, Saccharomyces cerevisiae и т.д.), противомалярийную, противогрибковую, противовирусную и противоопухолевую активности [Segraves N.L., Lopez S., Johnson T.A., Said S.A., Fu X., Schmitz F., Pietraszkiewicz H., Valeriote F.A., Crews P. Structures and cytotoxicities of fascaplysin and related alkaloids from two marine phyla - Fascaplysinopsis sponges and Didemnum tunicates // Tetrahedron Lett. 2003. Vol.44. P.3471-3475]. Именно противоопухолевая активность и вызывает наибольший интерес. Так, фаскаплизин проявляет цитотоксические свойства против таких опухолевых клеток, как: MALME-3M (меланома, IC50 0,03 мкг/мл), МКФ 7 (рак молочной железы, IC50 0,14 мкг/мл), OVCAR-3 (рак яичников, IC50 0,16 мкг/мл), А549 (рак легких, IC50 0,38 мкг/мл), L-1210 (лейкемия, IC50 0.2 мкг/мл) [Charan R.D., МсКее Т.С., Gustafson К.R., Pannell L.К., Boyd M.R. Thorectandramine, a novel β-carboline alkaloid from the marine sponge Thorectandra sp. // Tetrahedron Lett. 2002. Vol.43. P.5201-5204].
Механизм действия фаскаплизина на процессы, происходящие в клетке, достаточно хорошо изучен. Было показано, что фаскаплизин является селективным ингибитором циклинзависимой киназы 4 (CDK 4), которая является ключевым ферментом в регуляции перехода между фазами G0 и G1 в ходе клеточного цикла [Aubry С., Jenkins P.R., Mahale S., Chaudhuri В., Sutcliffe M.J. New fascaplysin-based CDK4-specific inhibitors: design, synthesis, and biological activity // Chem. Commun. 2004. P.1696-1697]. Для успешного прохождения этой точки необходимо наличие особых факторов транскрипции, относимых к группе E2F. Их непрерывному синтезу препятствует белок ретинобластома (pRB). Именно циклинзависимая киназа 4 в комплексе с циклином D1 проводит фосфорилирование этого белка, что вызывает его дезактивацию и в свою очередь приводит к высвобождению факторов E2F, прохождению точки G0/G1 и в дальнейшем ведет к клеточному делению. В нормальной клетке предусмотрена система регуляции активности CDK 4 природными ингибиторами этого фермента, такими как р16. При мутациях иногда возникает сбой в работе данной системы, клетка начинает бесконтрольно делиться, что приводит к возникновению злокачественной опухоли. Использование низкомолекулярных ингибиторов CDK4 существенно тормозит и даже останавливает рост опухоли. В этой связи фаскаплизин представляет огромный интерес как потенциальное лекарственное средство при лечении злокачественных новообразований.
Изучение действия фаскаплизина на асцитный вариант карциномы Эрлиха в условиях in vivo показало его малую активность против этой разновидности раковых опухолей [Попов A.M., Макарьева Т.Н., Федореев С.А., Стоник В.А. Противоопухолевая и цитостатическая активности низкомолекулярных метаболитов из морских тропических губок // Химиотерапия опухолей в СССР. 1991. Т.56. С.61-66].
В настоящее время большое внимание уделяется аналогам фаскаплизина. Однако относительно их биологической активности известно гораздо меньше, чем о биологической активности фаскаплизина.
О биологической активности 3-бромофаскаплизина до сих пор было известно немногое. Сообщалось лишь, что наряду с другими производными фаскаплизина 3-бромофаскаплизин был протестирован на цитотоксическую активность методом STS (Solid Tumor Selectivity). В этом исследовании он оказался менее активным, чем фаскаплизин. В экспериментах использовались мышиные опухолевые С38 клетки и опухолевые клетки человека H116 [Segraves N.L., Lopez S., Johnson T.A., Said S.A., Fu X., Schmitz F.J., Pietraszkiewicz H., Valeriote F.A., Crews P. Structures and cytotoxicities of fascaplysin and related alkaloids from two marine phyla - Fascaplysinopsis sponges and Didemnum tunicates // Tetrahedron Lett. 2003. Vol.44. P.3471-3475]. О молекулярном механизме цитотоксического действия 3-бромофаскаплизина (2) в литературе сведений нет.
Опубликована также международная заявка, в которой описано получение, установление структуры и применение некоторых производных фаскаплизина в качестве терапевтических противоопухолевых средств [Chaudhuri В., Mahale S.G. Fascaplysin derivatives and their use in the treatment of cancer. PCT Int. App.; WO 2009/022104 A1]. Однако использование 3-бромфаскаплизина (2) в качестве средства, предотвращающего трансформацию нормальных клеток в опухолевые, не описано.
Авторами в доступной патентной и другой научно-технической литературе не найдено указание на использование экстрактов, содержащих 3-бромофаскаплизин или индивидуального 3-бромофаскаплизина в качестве средства, предотвращающего трансформацию нормальных клеток в опухолевые.
Задача изобретения - расширение арсенала средств и фармацевтических композиций, способных избирательно предотвращать злокачественное перерождение нормальных клеток млекопитающих в опухолевые.
Задача решена применением 3-бромофаскаплизина (2) в качестве средства, предотвращающего трансформацию нормальных клеток млекопитающих в опухолевые.
Новое назначение 3-бромофаскаплизина в качестве средства, обладающего способностью предотвращать злокачественное перерождение нормальных клеток млекопитающих в опухолевые, не вытекает с очевидностью из его известных свойств и обнаружено авторами впервые.
3-Бромофаскаплизин (2), использованный для изучения его активности по предотвращению трансформации нормальных клеток млекопитающих в опухолевые, был получен нами синтетическим путем [Zhidkov M.E., Baranova O.V., Balaneva N.N., Fedorov S.N., Radchenko O.S., and Dubovitskii S.V. The first syntheses of 3-bromofascaplysin, 10-bromofascaplysin and 3,10-dibromofascaplysin - marine alkaloids from Fascaplysinopsis reticulata and Didemnum sp. by application of a simple and effective approach to the pyrido[1,2-a:3,4-b']diindole system // Tetrahedron Lett. 2007. V.48. P.7998-8000]. Синтез 3-бромофаскаплизина (2) состоит из 4-х стадий и основан на формировании исходя из соответствующим образом замещенных фенилуксусной кислоты и триптамина, пиридо[1,2-а:3,4-b']дииндольной системы, которая является скелетной основой алкалоида. Общий выход 3-бромофаскаплизина (2) составляет 43%.
Задача решена также применением 3-бромофаскаплизина (2) для приготовления фармацевтической композиции, предотвращающей трансформацию нормальных клеток млекопитающих в опухолевые.
Фармацевтические композиции, предотвращающие трансформацию нормальных клеток млекопитающих в опухолевые, получают, смешивая эффективное количество 3-бромофаскаплизина (2) с традиционными фармацевтически или фармакологически приемлемыми наполнителями. Выражение «фармацевтически или фармакологически приемлемый» обозначает молекулярные компоненты и композиции, которые не вызывают негативных, аллергических или иных нежелательных реакций при введении их животному или человеку. Природа наполнителя зависит от способа введения. Например, если желательным является пероральное введение, то может быть выбран твердый наполнитель, тогда как для внутривенного введения может быть использован жидкий солевой раствор.
Технический результат, обеспечиваемый изобретением, заключается как в способности 3-бромофаскаплизина (2) предотвращать трансформацию нормальных клеток млекопитающих в опухолевые, так и в возможности использования этого вещества для приготовления фармацевтических композиций, предотвращающих трансформацию нормальных клеток млекопитающих в опухолевые.
Изобретение расширяет арсенал средств и фармацевтических композиций, предотвращающих трансформацию нормальных клеток млекопитающих в опухолевые.
На фиг.1 представлено дозозависимое ингибирование 3-бромофаскаплизином (2) EGF-индуцированной опухолевой трансформации мышиных эпителиальных клеток JB6 Р+ Cl41.
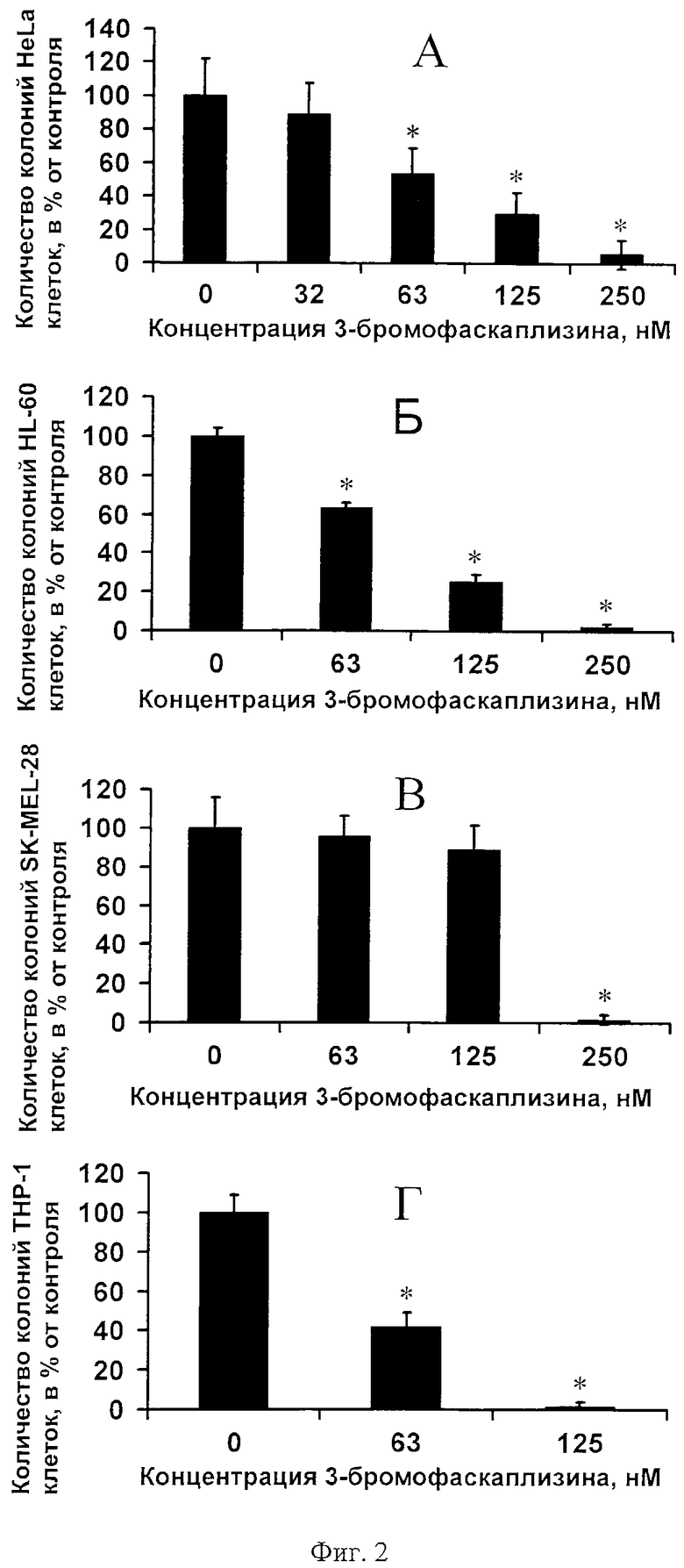
На фиг.2 представлено дозозависимое ингибирование 3-бромофаскаплизином (2) колонеобразования опухолевых клеток человека: HeLa (A), HL-60 (Б), SK-MEL-28 (В), ТНР-1 (Г).
Приготовление фармацевтических композиций иллюстрируется следующими примерами:
Пример 1. 3-бромофаскаплизин (2) в эффективном количестве смешивают с соответствующим количеством смеси крахмала и гипса и прессуют в таблетку.
Пример 2. Фармацевтическую композицию готовят, как описано в примере 1, но в качестве наполнителя используют смесь сорбита и стеарата кальция.
Пример 3. 3-бромофаскаплизин (2) в эффективных концентрациях растворяют в соответствующем количестве физраствора.
Исследование биологической активности 3-бромофаскаплизина (2).
I. Материалы и методы.
1. Принятые сокращения.
ДМСО - Диметилсульфоксид
EGF - Epidermal growth factor (эпидермальный фактор роста)
FBS - Fetal bovine serum (сыворотка бычьих эмбрионов)
HPLC - High performance liquid chromatography (высокоэффективная жидкостная хроматография)
IC50 - Inhibition concentration 50% (концентрация, вызывающая гибель 50% клеток)
INCC50 - Inhibition of number of colonies concentration 50% (концентрация, ингибирующая злокачественную трансформацию 50% клеток)
PBS - Phosphate-buffered saline (фосфатно-солевой буфер)
ТРА - 12-O-Tetradecanoylphorbol-13-acetate (12-O-тетрадеканоилфорбол-13-ацетат)
нМ - нонамоль/литр
мкМ - микромоль/литр
мМ - миллимоль/литр
М - моль/литр
мкл - микролитр
мл - милилитр
SD - стандартное отклонение от среднего
PBS - фосфатно-солевой буферный раствор.
ВМЕ - basal medium Eagle (питательная среда Игла для культивирования клеток млекопитающих)
RPMI, DMEM, MEM - питательные среды для культивирования клеток млекопитающих, изготовленные на основе среды Игла ВМЕ.
МТТ - 3-(4,5-диметилтиазолил)-2,5-дифенилтетразолиум бромид (реагент для определения цитотоксичности)
MTS - 5-(3-Карбоксиметоксифенил)-2-(4,5-диметилтиазолил)-3-(4-сульфофенил) тетразолиум, внутренняя соль (реагент для определения цитотоксичности).
2. Клеточные культуры
Мышиные эпителиальные клетки JB6 Р+ Cl41, а также опухолевые клетки человека, HeLa (рак шейки матки), ТНР-1 (лейкемия, моноциты), HL-60 (лейкемия, промиелоциты), SK-MEL-28 (меланома), MDA-MB-231 (рак молочной железы), SNU-C4 и DLD-1 (рак кишечника) из коллекции АТСС, Rockville, MD (США) выращивались в инкубаторе Sanyo MCO-15AC в монослое для прикрепленных (линии JB6 Р+ Cl41, SNU-C4, DLD-1, SK-MEL-28, HeLa, MDA-MB-231) или в суспензии для неприкрепленных клеток (линии ТНР-1 и HL-60) при 37°С и в атмосфере 5% СО2.
Для клеток линий JB6 Р+ Cl41 и SK-MEL-28 использовалась среда MEM, содержащая 5% FBS (10% FBS для SK-MEL-28), 2 mM раствора L-глутамина и 15 мкг/мл гентамицина.
Для клеток линий HeLa, ТНР-1, HL-60, SNU-C4 и DLD-1 использовалась среда RPMI-1640, содержащая 10% FBS, 2 mM раствора L-глутамина и 15 мкг/мл гентамицина.
Для клеток линии MDA-MB-231 использовалась среда DMEM, содержащая 10% FBS, 2 mM раствора L-глутамина и 15 мкг/мл гентамицина.
3. Приготовление растворов веществ.
Базовый (стоковый) раствор 3-бромофаскаплизина (2) с концентрацией вещества 20 мМ готовили в ДМСО, из которого получали растворы нужной концентрации разбавлением в культуральной среде. Содержание ДМСО в разбавленных растворах не превышало 0,5% во всех опытах.
4. Метод определения канцерпревентивной (предотвращающей злокачественное перерождение клеток) активности веществ (метод мягкого агара).
Эксперименты по изучению противоопухолевого профилактического эффекта исследуемых веществ проводили методом мягкого агара, в шестилуночных планшетах [Nakamura Y., Colburn N.Н., Cindhart T.D. Role of reactive oxygen in tumor promotion: implication of superoxide anion in promotion of neoplastic transformations in JB-6 cells by TPA // Carcinogenesis. 1985. V.6, №2. P.229-235]. Метод основан на способности мышиных эпидермальных JB6 Р+ Cl41 клеток (8×103 кл./мл) перерождаться в опухолевые под действием активирующего их эпидермального фактора роста (EGF), взятого в концентрации 10 нг/мл и, как следствие, образовывать колонии. Клетки, не обработанные эпидермальным фактором роста, колоний в мягком агаре не образуют. Методика рассчитана на приготовление 1 контрольного и 5 экспериментальных 6-луночных планшетов с мягким агаром. Всего за один эксперимент можно исследовать 10 различных концентраций (каждая в трипликате) одного или нескольких веществ:
А. Приготовление Agar Mix: В стерильной бутыле объемом 250 мл смешивали 18 мл PBS, 18 мл FBS, 100 мкл раствора гентамицин-сульфата с концентрацией 10 мг/мл (раствор в PBS), 2 мл 0,2 М раствора L-глутамина в PBS и 70 мл среды 2×ВМЕ. Смесь перемешивали и помещали в водяную баню (45°С) на 20 мин. Затем к смеси добавляли 72 мл разогретого до 50°С 1,25% раствора агара в воде (специально очищена для работы с клеточными культурами) и получали таким образом 180 мл смеси Agar Mix.
Б. Приготовление Agar Bottom: 180 мл смеси Agar Mix поделили на 2 части: 60 мл и 120 мл, каждая в стерильной бутыле объемом 250 мл. К 120 мл смеси Agar Mix добавляли 60 мкл раствора EGF с концентрацией 20 мкг/мл в PBS и получили таким образом 120 мл смеси Agar Bottom. Обе смеси, 120 мл Agar Bottom и 60 мл Agar Mix помещали в водяную баню (45°С).
В. Подготовка контрольного планшета (Bottom): В первые 3 лунки контрольного 6-луночного планшета добавляли по 3 мл Agar Mix (без EGF), в оставшиеся 3 лунки - по 3 мл Agar Bottom (с EGF). Для застывания растворов в лунках оставляли планшет на 30 мин при комнатной температуре
Г. Подготовка экспериментальных планшетов с веществами: В стерильные пробирки объемом 15 мл помещали рассчитанные объемы растворов исследуемого вещества так, чтобы получить требуемые концентрации в расчете на 10 мл конечного раствора, по одной пробирке на одну концентрацию одного вещества. Затем в каждую пробирку добавляли по 10 мл Agar Bottom (с EGF), перемешивали с помощью пипетки. Из каждой пробирки добавляли по 3 мл полученных растворов вещества с одной и той же концентрацией в 3 лунки одного из приготовленных 6-луночных планшетов. Для застывания растворов в лунках оставили планшеты на 30 мин при комнатной температуре.
Д. Подготовка JB6 P+ Cl41 клеток: Из бутыли, в которой выращивались JB6 P+ Cl41 клетки, с помощью пипетки Пастера удаляли клеточную среду, затем клетки промывали 5 мл PBS и добавляли 2 мл 0,25% раствор трипсина в PBS. Затем клетки с раствором трипсина инкубировались в течение 5 минут при 37°С в атмосфере 5% CO2, полученную суспензию клеток осторожно перемешивали с помощью пипетки, после чего в бутыль с клетками добавляли 8 мл соответствующей клеточной среды и суспензию снова перемешивали. Открепившиеся в процессе трипсинизации клетки переносили в стерильную пробирку и центрифугировали при 1000 об/мин в течение 10 мин. Далее с помощью пипетки Пастера удаляли супернатант и добавляли 5 мл среды 1×BME-10% FBS. После перемешивания считали концентрацию клеток в получившейся суспензии с помощью камеры Горяева.
Далее, путем смешивания необходимых объемов полученной суспензии клеток и среды 1×BME-10% FBS готовили 20 мл клеточной суспензии с концентрацией клеток 2,4×104 кл./мл.
Е. Подготовка контрольного планшета (Тор): Из оставшихся 51 мл Agar Mix отбирали в стерильный бутыль 30 мл раствора и добавляли в них 23 мкл 20 мкг/мл раствора EGF в PBS, таким образом получили 30 мл Agar Mix + EGF. Далее в первые 3 лунки 6-луночного контрольного планшета поверх застывшего Agar Mix (без EGF) добавляли по 1 мл предварительно приготовленной смеси: 1,2 мл клеточной суспензии JB6 Р+ Cl41 клеток и 2,4 мл Agar Mix (без EGF), получая, таким образом, 3 готовых лунки с нулевым контролем. Во вторые 3 лунки (с Agar Bottom) 6-луночного контрольного планшета поверх застывшего Agar Bottom добавляли по 1 мл предварительно приготовленной смеси 2,4 мл Agar Mix + EGF и 1,2 мл клеточной суспензии JB6 P+ Cl41 клеток, получая таким образом 3 готовых лунки со 100% контролем. После этого планшет помещали в инкубатор и инкубировали в течение 7 суток при 37°С в атмосфере 5% СО2.
Ж. Подготовка экспериментальных планшетов: В стерильные пробирки помещали необходимые объемы раствора исследуемого вещества (исходя из исследуемого диапазона концентраций) в расчете на 3,6 мл конечного раствора, по одной пробирке на одну концентрацию одного вещества. Затем в каждую пробирку добавляли по 2,4 мл Agar Mix + EGF и 1,2 мл клеточной суспензии JB6 P+ Cl41 клеток, перемешивали и добавляли по 1 мл полученной смеси в соответствующие лунки 6-луночных планшетов поверх застывшего раствора веществ в Agar Bottom. После этого планшет помещали в инкубатор и инкубировали в течение 7 суток при 37°С в атмосфере 5% CO2.
Полученная концентрация клеток в верхнем слое лунок всех планшетов равна 2,4×104×1,2/(1,2+2,4)=8×103 кл./мл. Концентрация агара в лунках в нижнем слое равна 1,25%×72/180=0,5%. Концентрация агара в лунках в верхнем слое равна (1,25%×72/180)×1,2/(1,2+2,4)=0,33%.
З. Получение результатов: После инкубации в течение 7 дней колонии живых клеток в лунках контрольного и экспериментальных планшетов были подсчитаны с помощью микроскопа Olympus (Япония). Количество колоний на рисунках показано в процентном отношении к количеству колоний в лунках со 100% контролем. Были проведены два независимых эксперимента.
5. Определение цитотоксической активности MTS-методом.
Для определения цитотоксической активности исследуемого вещества использовали стандартный MTS-метод (усовершенствованная модификация МТТ-метода) [Barltrop J.A., Owen Т.С., Cory А.H., Cory J.G. 5-(3-Carboxymethoxyphenyl)-2-(4,5-dimethylthiazolyl)-3-(4-sulfophenyl)tetrazolium, inner sault (MTS) and related analogs of 3-(4,5-dimethylthiazolyl)-2,5-diphenyltetrazolium bromide (MTT) reducing to purple water-soluble formazans as cell-viability indicators // Bioorg. Med. Chem Lett. 1991. №11. P.611-614]. Метод основан на способности живых клеток перерабатывать MTS-реагент (желтая окраска, λmax=382 нм) в формазан (красная окраска, λmax=492 нм).
Схема переработки MTS-реагента живыми (метаболитически активными) клетками в формазан.
Описание метода:
А. Приготовление планшета с клетками: Для прикрепленных клеток из бутыли, в которой выращивали клетки, с помощью пипетки Пастера удаляли клеточную среду, затем клетки промывали 5 мл PBS и добавляли 2 мл 0,25% раствора трипсина в PBS. Затем клетки с раствором трипсина инкубировали в течение 5 минут при 37°С в атмосфере 5% СО2, после этого осторожно перемешивали с помощью пипетки и к полученной суспензии клеток добавляли 8 мл соответствующей клеточной среды. Открепившиеся в процессе трипсинизации клетки переносили в пробирку и центрифугировали при 1000 об/мин в течение 10 мин. Далее с помощью пипетки Пастера удаляли супернатант и добавляли 5 мл соответствующей среды. После перемешивания считали концентрацию клеток в получившейся суспензии с помощью камеры Горяева.
Для неприкрепленных клеток клеточную суспензию (без предварительной обработки трипсином) центрифугировали при 1000 об/мин в течение 10 мин. Далее с помощью пипетки Пастера удаляли супернатант и добавляли 5 мл соответствующей среды. После перемешивания считали концентрацию клеток в получившейся суспензии с помощью камеры Горяева. Далее, путем смешивания необходимых объемов полученной суспензии клеток и соответствующей среды готовили клеточную суспензию с концентрацией - 6×105 кл/мл для прикрепленных и 12×105 кл/мл для неприкрепленных клеток - для загрузки в планшет.
Далее клетки высевали в 96-луночный планшет в лунки В1-H12, по 50 мкл клеточной суспензии на 1 лунку для неприкрепленных клеток и по 100 мкл клеточной суспензии на 1 лунку для прикрепленных клеток. Таким образом, количество клеток на 1 лунку в обоих случаях составляло 6000 клеток. В лунки А1-А12 добавляли соответствующую среду без клеток - по 50 мкл при приготовлении планшета с неприкрепленными клетками и по 100 мкл в случае прикрепленных клеток.
Планшеты инкубировали при 37°С в атмосфере 5% СО2 в течение 1 суток.
Б. Приготовление растворов веществ: На аналитических весах брали навеску исследуемого вещества и растворяли ее в необходимом объеме ДМСО, так, чтобы концентрация вещества в полученном растворе была 20 мМ. Далее приготавливали растворы вещества соответствующих концентраций в соответствующей питательной среде.
В. Загрузка веществ в планшет: В случае прикрепленных клеток из всех лунок с помощью пипетки Пастера удаляли клеточную среду и в лунки С1-H12 помещали приготовленные ранее растворы с исследуемым веществом по 100 мкл в каждую лунку, по 3 лунки с одной и той же концентрацией вещества. В лунки В1-В12 и А1-А12 добавляли по 100 мкл соответствующей среды без вещества (эти лунки служат в качестве контрольных).
Для неприкрепленных клеток к 50 мкл уже имеющейся в каждой лунке клеточной суспензии добавляли еще 50 мкл раствора исследуемого вещества в соответствующей среде. Таким образом, концентрация вещества в клеточной среде уменьшалась в 2 раза по сравнению с исходной, что необходимо учитывать при приготовлении растворов вещества в среде до загрузки их на планшет. В лунки В1-В12 и А1-А12 добавляли по 50 мкл соответствующей среды без вещества. После этого планшеты инкубировали при 37°С в атмосфере 5% CO2 в течение 1 суток.
Г. Получение результатов: По завершении инкубирования в каждую лунку добавляли по 20 мкл MTS-реагента. Затем планшеты инкубировали при 37°С в атмосфере 5% СО2 еще в течение 2 часов. После чего оптическую плотность среды в каждой лунке регистрировали с помощью спектрофотометрического планшетного ридера при 492 нм (интенсивность поглощения, обусловленного наличием формазана) и 690 нм (результат использовали в качестве фонового показателя). Интенсивность окраски формазана при 492 нм прямо пропорциональна количеству оставшихся живых (метаболически активных) клеток [Barltrop J.A., Owen Т.С., Cory А.Н., Cory J.G. 5-(3-Carboxymethoxyphenyl)-2-(4,5-dimethylthiazolyl)-3 -(4-sulfophenyl)tetrazolium, inner sault (MTS) and related analogs of 3-(4,5-dimethylthiazolyl)-2,5-diphenyltetrazolium bromide (MTT) reducing to purple water-soluble formazans as cell-viability indicators // Bioorg. Med. Chem. Lett. 1991. №11. P.611-614]. Для определения цитотоксической активности вещества использовали также соответствующие спектрофотометрические показатели контрольных лунок на планшете: лунок с нулевым контролем (А1-А12), в которые не высевали клетки и не добавляли вещества, но добавляли MTS-реагент, и лунок со 100%-ным контролем (В1-В12), в которые высевали клетки в том же количестве, что и в экспериментальные, не добавляли вещества, но также добавляли MTS-реагент.
Д. Обработка результатов: Для вычисления количества живых клеток, оставшихся в экспериментальных лунках:
1. Из значения интенсивности поглощения среды при 492 нм в каждой лунке вычитали значение интенсивности поглощения среды при 690 нм в соответствующей лунке.
2. Находили среднее значение полученных в пункте 1 результатов для лунок с нулевым контролем и вычитали его из значений, полученных в пункте 1 для всех остальных лунок.
3. Вычисляли среднее значение полученных в пункте 2 результатов для лунок со 100%-ным контролем, IK.
4. Вычисляли количество живых клеток в каждой экспериментальной лунке (N), в процентах по сравнению с контрольными лунками, по формуле:
N=(IЭ/IK)×100%,
где IЭ - это интенсивность поглощения среды в каждой экспериментальной лунке, полученное в пункте 2;
IK - среднее значение полученных в пункте 3 результатов для лунок со 100%-ным контролем;
Для каждой клеточной линии было проведено два независимых эксперимента.
II. Результаты исследования биологической активности 3-бромофаскаплизина и их обсуждение.
Противоопухолевая профилактическая (предотвращающая злокачественное перерождение клеток) активность.
К канцерпревентивным веществам относятся природные метаболиты или их синтетические аналоги, которые ингибируют трансформацию нормальных клеток в прораковые или прогрессию прораковых клеток в опухолевые. Следовательно, эффективное канцерпревентивное вещество должно вмешиваться в процесс канцерогенеза и тем или иным образом нейтрализовывать прораковые клетки, не давая им превратиться в опухолевые. Для исследования синтезированного 3-бромофаскаплизина (2) на канцерпревентивную активность мы использовали широко применяемый метод мягкого агара, мышиные эпителиальные JB6 P+ Cl41 клетки и опухолевые клетки человека HeLa (рак шейки матки), SK-MEL-28 (меланома), HL-60 и ТНР-1 (лейкемия), а также эпидермальный фактор роста (EGF) в качестве промотора опухолевой трансформации JB6 Р+ Cl41 клеток [Colburn N.H. Former B.F., Nelson K.A., Yuspa S.H. Tumour promoter induces anchorage independence irreversibly // Nature. 1979. V.281. P.589-591; Dong Z., Birrer M.J., Watts R.G., Matrisian L.M., Colburn N.H. Blocking of tumor promoter-induced AP-1 activity inhibits induced transformation in JB6 mouse epidermal cells // Proc. Natl. Acad. Sci. USA. 1994. V.91, №2. P.609-613; Dong Z., Watts S.G., Sun Y., Colburn N.H. Progressive elevation of AP-1 activity during preneoplastic-to-neoplastic progression as modeled in mouse JB6 cell variants // Int. J. Oncol. 1995. V.7. P.359-364; Strickland J., Sun Y., Dong Z., Colburn N.H. Grafting assay distinguishes promotion sensitive from promotion resistant JB6 cells // Carcinogenesis. 1997. V.18. P.1135-1138)]. Система клональных генетических вариантов JB6 клеток, которая включает в себя чувствительные к трансформации (Р+) и нечувствительные (Р-), а также трансформированные JB6 Cl41 клетки, широко используется в поиске канцерпревентивных веществ и изучении их свойства на молекулярном уровне. Различные типы JB6 клеток находятся на различных стадиях процесса их преобразования от пренеопластического до неопластического состояния и от ранней до поздней стадий такого преобразования [Colburn N.H., Wendel Е., Srinivas L. Responses of preneoplastic epidermal cells to tumor promoters and growth factors: Use of promoter-resistant variants for mechanism studies. // J. Cell. Biochem. 1982. V.18, №3. P.261-270; Huang С., Ma W., Young M.R., Colburn N., Dong Z. Shortage of mitogen-activated protein kinase is responsible for resistance to AP-1 transactivation and transformation in mouse JB6 cells // Proc. Natl. Acad. Sci. USA. 1998. V.95, №1. P.156-161; Bernstein L.R., Colburn N.H. AP1/jun function is differentially induced in promotion-sensitive and resistant JB6 cells // Science. 1989. V.244, №4904. P.566-569].
JB6 P+ Cl41 клетки трансформируются после их обработки промоторами злокачественной трансформации, такими как EGF или ТРА. В процессе такой трансформации происходит активирование ядерного фактора транскрипции АР-1, который регулирует транскрипцию различных генов, ответственных за процессы воспаления, пролиферации и метастазирования [Dong Z., Birrer M.J., Watts R.G., Matrisian L.M., Colburn N.H. Blocking of tumor promoter-induced AP-1 activity inhibits induced transformation in JB6 mouse epidermal cells // Proc. Natl. Acad. Sci. USA. 1994. V.91, №2. P.609-613; Huang С., Ma Y, Dawson M.I., Rincon M., Flavell R.A., Dong Z. Blocking activator protein - 1 activity, but not activating retinoic acid response element, is required for the antitumor promotion effect of retinoic acid // Proc. Natl. Acad. Sci. USA. 1997. V.94, №11. P.5826-5830].
Полученный результат дозозависимого ингибирования 3-бромофаскаплизином (2) опухолевой трансформации JB6 Р+ Cl41 клеток отражен на фиг.1 как количество колоний трансформированных JB6 Р+ Cl41 клеток в процентах от контроля в зависимости от концентрации исследуемого вещества (диаграмма получена с помощью программы Microsoft Excel). Каждый «столбик» на фиг.1 представляет собой усредненный результат двух независимых экспериментов, по три образца в каждом ± SD (стандартное отклонение от среднего). Астериск (*) указывает на результат, статистически достоверно отличающийся от контроля, р<0.05.
На основании этих данных, с помощью компьютерной программы Statistica 6.0 методом регрессии, были произведены расчеты концентрации (INCC50), при которой исследуемое вещество ингибирует опухолевую трансформацию 50% клеток JB6 Р+ Cl41. В результате было вычислено, что INCC50 3-бромофаскаплизина (2) по отношению к опухолевой трансформации JB6 Р+ Cl41 клеток = 106,4 нМ, что почти в три раза меньше цитотоксической концентрации (IC50=284 нМ) 3-бромофаскаплизина (2) для той же клеточной линии. Результаты представлены в таблице.
Результаты дозозависимого ингибирования 3-бромофаскаплизином (2) фенотипической экспрессии (колонеобразования) опухолевых клеток человека HeLa, HL-60, ТНР-1, SK-MEL-28 в мягком агаре отражен на фиг.2 как количество колоний HeLa (A), HL-60 (Б), SK-MEL-28 (В) и ТНР-1 (Г), клеток в процентах от контроля в зависимости от концентрации исследуемого вещества (диаграмма получена с помощью программы Microsoft Excel). Каждый «столбик» на фиг.2 представляет собой усредненный результат двух независимых экспериментов, по три образца в каждом ± SD (стандартное отклонение от среднего). Астериск (*) указывает на результат, статистически достоверно отличающийся от контроля, р<0.05.
На основании этих данных, с помощью компьютерной программы Statistica 6.0 методом регрессии, были произведены расчеты концентрации (INCC50), при которой 3-бромофаскаплизин (2) ингибирует на 50% колонеобразование HeLa, HL-60, ТНР-1 и SK-MEL-28 клеток в мягком агаре. В результате было вычислено, что INCC50 3-бромофаскаплизина (2) по отношению к опухолевой трансформации HeLa, HL-60, SK-MEL-28 и ТНР-1 клеток = 108,1; 103,5; 162,9; и 59,7 нМ, соответственно, что в несколько раз меньше соответствующих цитотоксических концентраций (IC50 = 337; 549; 785; 521 нМ) 3-бромофаскаплизина (2) для тех же клеточных линий. Результаты представлены в таблице.
Таким образом, проведенные эксперименты показали, что 3-бромофаскаплизин (2) способен ингибировать опухолевую трансформацию JB6 Р+ Cl41 клеток, а также колонеобразование HeLa, HL-60, ТНР-1, SK-MEL-28 опухолевых клеток человека в концентрациях меньше цитотоксических (IC50) для той же клеточной линии в 3, 3, 5, 5 и 8 раз соответственно (фиг.1, 2; таблица). Это говорит о возможности использования 3-бромофаскаплизина (2) в качестве средства профилактики раковых заболеваний.
Противоопухолевая цитотоксическая активность.
Цитотоксическая активность 3-бромофаскаплизина (2) по отношению к восьми линиям опухолевых и нормальных клеток человека и мыши была изучена MTS-методом. Результаты представлены в таблице в виде количества живых клеток определенной линии в зависимости от концентрации исследуемого вещества в среде.
Для каждого значения, отражающего процент живых клеток относительно контроля, указано стандартное отклонение от среднего. Астериск (*) указывает на результат, статистически достоверно отличающийся от контроля, р<0.05 (Манн-Уитни  -тест).
-тест).
На основании данных, приведенных в таблице, и с помощью компьютерных программ Microsoft Excel и Statistica 6.0 были произведены расчеты IC50 (концентрация исследуемого вещества, при которой гибнет 50% исследуемых клеток) для 3-бромофаскаплизина (2) по отношению к каждой из восьми исследованных клеточных линий. Полученные IC50 также представлены в таблице.
Из результатов, представленных в таблице, можно сделать вывод о том, что 3-бромофаскаплизин (2) более активен по отношению к клеткам рака кишечника (DLD-1 и SNU-C4) по сравнению с другими линиями опухолевых клеток человека.
Необходимо отметить, что цитотоксическая активность 3-бромофаскаплизина (2) по отношению к приведенным в таблице клеточным линиям ранее не изучалась и приводится впервые.
| название | год | авторы | номер документа |
|---|---|---|---|
| СРЕДСТВО, ПРЕДОТВРАЩАЮЩЕЕ ТРАНСФОРМАЦИЮ НОРМАЛЬНЫХ КЛЕТОК МЛЕКОПИТАЮЩИХ В ОПУХОЛЕВЫЕ | 2010 |
|
RU2429840C1 |
| СРЕДСТВО, ВОЗДЕЙСТВУЮЩЕЕ НА АКТИВНОСТЬ НЕКОТОРЫХ ЯДЕРНЫХ ФАКТОРОВ ТРАНСКРИПЦИИ | 2011 |
|
RU2457840C1 |
| ГЛИКОЗИДНЫЕ ПРОИЗВОДНЫЕ 1,2-ДИТИОЛ-3-ТИОНА ИЛИ 1,2-ДИТИОЛ-3-ОНА И ЛЕКАРСТВЕННЫЕ СРЕДСТВА НА ИХ ОСНОВЕ | 2013 |
|
RU2534525C9 |
| ТЕРАПЕВТИЧЕСКИЕ ХИНОНЫ | 2005 |
|
RU2411229C2 |
| ИНГИБИТОР ПРОТЕИНКИНАЗЫ ТОРК С НАПРАВЛЕННЫМ ХЕМОПРЕВЕНТИВНЫМ ДЕЙСТВИЕМ IN VITRO | 2014 |
|
RU2566268C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТЬЮ | 2009 |
|
RU2414920C1 |
| МОНОЯДЕРНЫЕ ДИНИТРОЗИЛЬНЫЕ КОМПЛЕКСЫ ЖЕЛЕЗА, СПОСОБ ПОЛУЧЕНИЯ МОНОЯДЕРНЫХ ДИНИТРОЗИЛЬНЫХ КОМПЛЕКСОВ ЖЕЛЕЗА, ДОНОР МОНООКСИДА АЗОТА, ПРИМЕНЕНИЕ МОНОЯДЕРНОГО ДИНИТРОЗИЛЬНОГО КОМПЛЕКСА ЖЕЛЕЗА В КАЧЕСТВЕ ПРОТИВООПУХОЛЕВОГО ЛЕКАРСТВЕННОГО СРЕДСТВА | 2011 |
|
RU2494104C2 |
| Клеточная линия рака молочной железы человека BrCCh4e | 2019 |
|
RU2717654C1 |
| Способ ингибирования метастазирования опухолей путем подавления дедифференцировки опухолевых клеток | 2023 |
|
RU2840966C1 |
| ЦИТОТОКСИЧЕСКАЯ И ПРОТИВОВИРУСНАЯ АКТИВНОСТЬ 3-АЦИЛОКСИМЕТИЛ-3-ОКСО-1-ЦИАНО-2,3-СЕКО-2-НОР-ТРИТЕРПЕНОИДОВ | 2018 |
|
RU2686100C1 |
Предложено применение 3-бромофаскаплизина в качестве средства, предотвращающего трансформацию нормальных клеток млекопитающих в опухолевые, а также для приготовления фармацевтических композиций с данным (канцерпревентивным) действием. Показано, что 3-бромофаскаплизин оказывает раково-превентивное действие в нецитотоксических для нормальных клеток концентрациях. Изобретение может найти применение в медицине, конкретно в онкологии, а также в научных исследованиях в области клеточной молекулярной биологии. 2 н.п. ф-лы, 2 ил., 1 табл.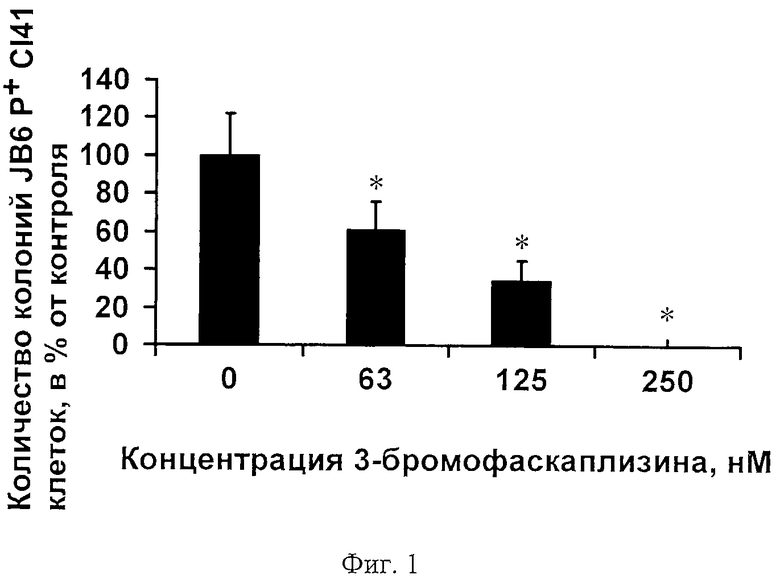
1. Применение 3-бромофаскаплизина в качестве средства, предотвращающего трансформацию нормальных клеток млекопитающих в опухолевые.
2. Применение 3-бромофаскаплизина для приготовления фармацевтической композиции, предотвращающей трансформацию нормальных клеток млекопитающих в опухолевые.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| КОНДЕНСИРОВАННЫЕ ГЕТЕРОЦИКЛИЧЕСКИЕ СУКЦИНИМИДНЫЕ СОЕДИНЕНИЯ | 2001 |
|
RU2298554C2 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| KR 20090017928 A, 19.02.2009 Реферат [он-лайн] [найдено 2010-12-16] (Найдено из базы данных Esp@cenet) | |||
| Жидков M.E | |||
| и др | |||
| Синтез бромофаскаплизинов - цитотоксичных алкалоидов из морских беспозвоночных | |||
| Исследования в области физико-химической биологии и | |||

