Настоящее изобретение относится к новым носителям для вакцин, а также для других молекул, созданным на основе ядерного антигена вируса гепатита В. В ядерный антиген вируса гепатита встраивают чужеродные аминокислотные последовательности, которые могут быть выведены из возбудителей болезни (патогенов). Вакцина предназначена для того, чтобы обеспечивать возможность продуцирования антител к указанным белковым последовательностям, при этом предпочтение отдается обладающим защитным действием или нейтрализующим антителам. Частицы, предлагаемые в изобретении, могут также стимулировать клеточный иммунный ответ (Т-клетки). Ядерный белок вируса гепатита В характеризуется тем, что большое количество его копий обладают способностью образовывать капсидоподобные частицы. Эти капсидоподобные частицы (CLP) наиболее пригодны для создания вакцин, поскольку они стимулируют иммунную систему и тем самым усиливают производство антител. Понятие «вакцина» в контексте настоящего описания предпочтительно обозначает систему-носитель, которая может вызывать как гуморальный, так и клеточный иммунный ответ.
Капсиды вирусов гепатита В представляют собой обладающие икосаэдральной симметрией наночастицы (диаметром примерно 30 нм), которые состоят из 180 или 240 копий вирусного ядерного белка. Ядерный белок обозначают как HBcAg. Капсиды могут служить в качестве представляющих собой частицы носителей для чужеродных молекул и их предпочтительно применяют в качестве усиливающих иммунную реакцию носителей антигена для вакцин. С помощью соответствующих модификаций ядерного белка можно презентовать чужеродные молекулы на поверхности частиц (а также при необходимости внутри них). Для презентации на внешней поверхности оптимальным является сцепление чужеродной молекулы с аминокислотными (ак) остатками, расположенными в центральной области аминокислотной последовательности ядерного белка (область примерно от ак 73 до ак 94), которые включают иммунодоминантный В-клеточный эпитоп «с/е1» и которые в наибольшей степени «выставлены» на поверхности 3-мерной структуры (3D-структуры) частицы (ак обозначает аминокислоту).
В WO 01/77158 описаны слитые белки ядерного антигена вируса гепатита В, в которых гетерологичные эпитопы встроены предпочтительно в область между положениями 61 и 90. В US 2003/0198649 описаны частицы ядерного антигена вируса гепатита В, в которых иммуногены соединены посредством лигандных структур с ядерным белком вируса гепатита В (HBV). У Kratz и др., PNAS, 1999, cc.1915-1920 описана презентация GFP (зеленый флуоресцентный белок) на поверхности ядерного белка HBV. В данном случае аминокислоты 79 и 80 ядерного белка были заменены на состоящую из 238 аминокислот последовательность GFP.
У Nassal и др., Eur. J. Immunol., 2005, cc.655-665 описан продукт слияния полноразмерного белка OspA Borrelia burgdorferi и капсидного белка вируса гепатита В. И в этом случае также аминокислоты 79 и 80 были заменены аминокислотами с 18 по 273 белка OspA, однако при этом между ядерным белком и OspA были встроены линкерные последовательности. У Skamel и др. (Journal of Biological Chemistry, 2006, cc.17474-17481) описана презентация полноразмерного белка OspC Borrelia burgdorferi с помощью капсидоподобных частиц вируса гепатита В. Однако было установлено, что известные из существующего уровня техники модифицированные ядерные белки, предлагавшиеся для решения проблемы, не сохраняют тенденцию к образованию капсидоподобных частиц (CLP) требуемым образом и, следовательно, не обладают удовлетворительной способностью стимулировать иммунный ответ.
Это означает, что при генетическом встраивании чужеродных молекул на основе пептидов или белков в ядерный белок, чужеродную последовательность связывают с ядерным белком как на ее N-конце, так и на ее С-конце. При создании настоящего изобретения было установлено, что двухсторонняя связь резко ограничивает тип и набор пригодных чужеродных последовательностей.
Представленная в настоящем описании система с расщепленным ядром устраняет указанные ограничения за счет того, что ядерный белок создают в виде двух раздельных частей, которые, как неожиданно было установлено при создании изобретения, спонтанно взаимодействуют и образуют капсидные частицы подобно тому, как это происходит в случае непрерывной белковой цепи. Чужеродные молекулы можно сливать либо с N-концевым фрагментом («coreN»), либо с С-концевым фрагментом («coreC») и в результате этого они оказываются сцепленными с белком-носителем только через один конец. За счет этого в значительной степени устраняются обусловленные двухсторонней связью структурные ограничения на встраивание чужеродных последовательностей в непрерывную пептидную цепь ядерного белка.
Таким образом, система с расщепленным ядром, предлагаемая в изобретении, позволяет осуществлять:
(I) презентацию чужеродных молекул, которые вследствие своего размера и/или структуры могут презентоваться в контексте обычного непрерывного ядерного белка лишь в ограниченной степени или не могут презентоваться вообще;
(II) презентацию гетеродимерных чужеродных белков;
(III) презентацию обладающих способностью к взаимодействию чужеродных молекул в гибкой хорошо доступной форме, что существенно облегчает взаимодействие с требуемыми молекулами-партнерами;
(IV) презентацию дополнительных чужеродных молекул через посредство других «выставленных наружу» N- и С-концов, которые присутствуют в системе с расщепленным ядром, но не присутствуют в обычной системе с непрерывным ядром.
Вирусы гепатита В (HBV) представляют собой оболочечные вирусы. Внутренний нуклеокапсид называется также коровой (ядерной) частицей («ядром») и серологически его определяют как ядерный антиген вируса гепатита В (HBcAg). Ядра состоят из 180 или 240 копий ядерного белка длиной 183-185 ак (в зависимости от подтипа HBV). Ядерный белок может экспрессироваться гетерологично (в бактериях, дрожжах, эукариотических клетках); при этом спонтанно образуются «капсидоподобные частицы» (т.е. CLP). Эти частицы являются неинфекционными, поскольку они не содержат ни вирусный геном, ни внешнюю оболочку. Такие CLP можно получать также из HBV более близкородственных млекопитающих (например, сурка; суслика и др.) и из HBV находящихся в более дальнем родстве птиц (например, утки; цапли и др.). В принципе, согласно изобретению можно применять любые HBV-последовательности, однако предпочтительными являются последовательности HBV, которые способны заражать человека.
Аминокислотная последовательность, соответственно нуклеотидная последовательность ядерного белка HBV являются известными (см. Galibert и др., Nature, 1979, cc.646-650; Nassal, Gene, 1988, cc.279-294 или WO 01/77158; указанные публикации включены в настоящее описание в качестве ссылки). Предпочтительно применяют последовательность ядерного белка подтипа ayw, но можно применять также варианты, модификации HBV-последовательности, а также последовательность HBV других млекопитающих или HBV птиц. Последовательности хранятся в доступных для научной общественности банках генов.
Помимо патогенного для человека вируса гепатита В (HBV в узком смысле) существует целый ряд родственных специфических в отношении конкретного вида животных вирусов гепатита В, например, HBV североамериканского лесного сурка (английское название: лесной сурок; зоологическое название: Marmota monax; вирус гепатита В сурка, т.е. WHV), калифорнийская земляная белка (английское название: калифорнийская земляная белка, зоологическое название: Spermophilus beecheyi, вирус гепатита В земляной белки, т.е. GSHV) и др.
Эти вирусы обладают генетической структурой, сходной со структурой человеческого HBV, но имеют выраженные отличия в своей нуклеотидной последовательности и, как следствие, в аминокислотной последовательности своих белков.
Применение ядра человеческого HBV в качестве носителя для вакцины имеет два возможных ограничения.
1. Часть индивидуумов, страдающих хронической HBV-инфекцией, обладает толерантностью к антигенам HBV (это отсутствие иммунного ответа обусловливает, в частности, персистентность инфекции). Ядро действует также в качестве зависимого от Т-клетки антигена (Т-зависимый антиген). Поэтому Т-клеточные эпитопы в последовательности ядерного белка вносят вклад в сильную иммуногенность вакцин на основе CLP ядерных HBV. Это, по-видимому, не оказывает воздействия на индивидуумов, хронически инфицированных HBV, поскольку их Т-клетки толерантны к ядру HBV. Однако не имеется такой толерантности к ядру WHV и HBV других животных (см. Billaud J.N. и др., Advantages to the use of rodent hepadnavirus core proteins as vaccine platforms. Vaccine, 25(9), 19 февраля 2007 г., cc.1593-606). Для этого специфического круга индивидуумов, которые потенциально могут быть подвергнуты вакцинации, применение ядерного белка HBV животных в качестве носителя может иметь преимущество.
2. Благодаря своей очень сильной иммуногенности ядро вызывает сильный В-клеточный ответ к ядру (антитела к HBcAg) у всех индивидуумов, остро или хронически инфицированных HBV; кроме того, характерным является то, что в организме излеченных индивидуумов образуются антитела к оболочечным белкам HBV (антитела к HBsAg). Применяемые в настоящее время профилактические вакцины против HBV-инфекции основаны исключительно на оболочечных белках HBV (HBsAg). Таким образом, успешная вакцинация приводит, также как и перенесенная инфекция, к образованию антител к HBsAg. В определенных случаях с точки зрения диагностики представляет интерес выяснение того, приобрел ли индивидуум антитело к HBsAg в результате инфекции или после HbsAg-вакцинации, это можно определить путем обнаружения антитела к HbcAg, которое образуется только после инфекции.
Вакцина на основе ядра HBV человека может вызывать помимо требуемого ответа на внедрившийся чужеродный белок также определенный ответ к ядру (антитело к HBcAg). В том случае, когда одновременно присутствует антитело к HbsAg, это может затруднять выявление различия между результатами HBV-инфекции и HbsAg-вакцинации.
В случае системы с расщепленным ядром производство антитела к HBcAg уже является минимальным, поскольку основной эпитоп, а именно с/е1, физически разделен, и поэтому не может более распознаваться большинством антител к HBcAg. Однако сохраняется остаточный ответ в виде антитела к HBcAg на другие области последовательности ядерного белка. Этот ответ можно предупреждать или дополнительно снижать путем применения ядерного белка нечеловеческого HBV, предпочтительно ядерного белка WHV, в качестве основы носителя.
Наиболее важными компонентами системы-носителя с расщепленным ядром, предлагаемой в изобретении, являются, во-первых, ядерный белок вируса гепатита В и, во-вторых, чужеродная молекула, к которой требуется вызвать иммунный ответ.
В наиболее широком варианте осуществления изобретения ядерный белок получают из любого вируса гепатита В. Известны различные типы вируса гепатита В. Согласно изобретению предпочтительно применяют те последовательности, которые выведены из вирусов гепатита В, выделенных из млекопитающих и обладающих специфичностью к ним. Наиболее предпочтительно применяют HBV человека. Однако наряду с этим можно применять также такие вирусы гепатита В, которые имеют последовательность ядерного белка, происходящую из других животных, например, птиц, таких как утки или цапли.
Другой компонент, а именно, чужеродная молекула, к которой требуется вызвать иммунный ответ, в принципе может представлять собой любую молекулу. В данном случае предпочтительными являются белковые последовательности и наиболее предпочтительно те белковые последовательности, которые присутствуют на поверхности патогенов и которые вступают в контакт с иммунной системой. Если иммунная система способна продуцировать антитела к таким поверхностным структурам патогенов, то последние инактивируются после контакта с отдельными компонентами иммунной системы вакцинированного пациента, при этом проникнувший патоген, как правило, разрушается.
На основе сравнения имеющихся в базах данных последовательностей вируса гепатита В сурка (WHV) с аминокислотной последовательностью ядерного белка HBV (подтипа ayw) было установлено, что они идентичны друг другу примерно на 60%. Наибольшие различия в последовательностях выявлены в области, находящейся между аминокислотными положениями 66 и 94, причем эта последовательность включает эпитоп с/е1 и в системе с расщепленным ядром HBV, предлагаемой в изобретении, сайт рестрикции между N-концевой областью и С-концевой областью ядерного белка. В наиболее предпочтительном варианте осуществления изобретения этот сайт рестрикции расположен между аминокислотами 79 и 81.
Для создания систем носителей с расщепленным ядром WHV, предлагаемых в изобретении, последовательности ядерного белка WHV расщепляли на уровне нуклеиновой кислоты между положением 79 (кодирует аминокислоту Glu) и положением 80 (кодирует аминокислоту глутамин). На нуклеотидном уровне последовательность, кодирующую сайт расщепления BamHI, добавляли на карбокси-конец N-концевого сегмента ядерного белка WHV (сокращенно обозначен как WcoreN). За ним вплотную расположен второй рибосомальный сайт связывания, а также сайт, расщепляемый эндонуклеазой Nde1, который, в свою очередь, перекрывается стартовым кодоном сегмента coreC WHV (WcoreC). Это расположение идентично предпочтительному расположению в соответствующих конструкциях с расщепленным ядром HBV. Это позволяет осуществлять перенос вставок непосредственно из одной системы в другую с помощью расщепления в BamHI-Nde1-сайтах. В представленном в данном описании случае WcoreN содержит благодаря встроенному сайту рестрикции BamHI консервативную замену, а именно, E79D, и, кроме того, Р в качестве C-концевой аминокислоты. Это не оказывает вредного воздействия на способность образовывать частицы, и C-концевой пролин, благодаря его устойчивости к действию протеазы, повышает стабильность расщепленного белка. Аналогично расщепляемому ядру HBV WcoreC содержит дополнительный стартовый кодон метионина, в данном случае расположенные перед Q80 (перед S81 в случае HBV). Эта конструкция с расщепленным ядром WHV так же эффективно образует частицы, как и соответствующая конструкция с расщепленным ядром HBV.
В другом варианте осуществления настоящего изобретения С-концевая область и N-концевая область ядерного белка могут происходить из различных вирусов гепатита В. В этом случае гибридная система-носитель с расщепленным ядром, предлагаемая в изобретении, имеет три главных компонента, а именно, N-концевую область ядерного белка из вируса гепатита В, например, вируса гепатита В человека, чужеродную молекулу, к которой иммунная система может вырабатывать ответ, и С-концевую область ядерного белка, происходящую из другого вируса гепатита В, например, из WHV. Такая гибридная конструкция с расщепленным ядром также без каких-либо проблем образует частицы. Поскольку аминокислотные последовательности ядерных белков из различных вирусов гепатита В отличаются друг от друга, то В- и Т-клеточные эпитопы также являются различными. Следовательно, с помощью гибридной системы с расщепленным ядром можно оказывать специфическое воздействие на иммунный ответ на конкретный носитель.
Для создания предпочтительных гибридных систем-носителей с расщепленным ядром, предлагаемых в изобретении, последовательность ядерного белка WHV расщепляли на уровне нуклеиновой кислоты между положением 79 (кодирует аминокислоту Glu) и положением 80 (кодирует аминокислоту глутамин). На нуклеотидном уровне последовательность, кодирующую сайт рестрикции BamH1, добавляли на карбокси-конец N-концевого сегмента ядерного белка WHV (сокращенно обозначен как WcoreN). За ним расположен вплотную второй рибосомальный сайт связывания, а также сайт, расщепляемый эндонуклеазой Nde1, который, в свою очередь, перекрывается стартовым кодоном сегмента coreC WHV (WcoreC). Это расположение идентично предпочтительному расположению в соответствующих конструкциях с расщепленным ядром HBV. Это позволяет осуществлять перенос вставок непосредственно из одной системы в другую с помощью расщепления в BamH1-Nde1-сайтах. В представленном в данном описании случае WcoreN содержит благодаря встроенному сайту рестрикции BamH1 консервативную замену, а именно, E79D, и, кроме того, Р в качестве C-концевой аминокислоты. Это не оказывает вредного воздействия на способность образовывать частицы, и C-концевой пролин, благодаря его устойчивости к действию протеазы, повышает стабильность расщепленного белка. Аналогично расщепленному ядру HBV WcoreC содержит дополнительный стартовый кодон метионина, в данном случае перед Q80 (перед S81 в случае HBV). Оказалось, что эта конструкция может без проблем образовывать частицы.
CLP встречающегося в естественных условиях HbcAg, а также рекомбинантного HBV, обладают высокой иммуногенностью (независимый от Т-клетки и зависимый от Т-клетки антиген). Существенное значение для этого имеет симметричная мультимерная состоящая из частиц структура ядерной частицы. Эту структуру можно выявлять с помощью обычной электронной микроскопии с «негативным окрашиванием», а также с помощью электронной криомикроскопии. Структуру, состоящую из отдельных частиц, можно выявлять биохимическим методом с помощью седиментации в градиентах плотности сахарозы (коэффициент седиментации в зависимости от варианта составляет 60-80 S; для сравнения: коэффициент седиментации растворимых мономерных белков: примерно 1-5 S; эукариотических рибосомальных субъединиц: 40 S + 60 S). Кроме того, частицы отличаются специфическими миграционными характеристиками при нативном гель-электрофорезе в агарозных гелях, которые отличны от миграционных характеристик форм, не состоящих из частиц. Это позволяет определять, могут ли образовываться CLP или присутствуют белки, не образующие CLP-структуру.
Ядерный белок HBV содержит 183 или 185 аминокислот (ядерный белок WHV содержит 187 аминокислот) и состоит из двух доменов: первые примерно 140 ак необходимы и достаточны для сборки CLP (домен сборки); C-концевая область богата основными ак и связывается с нуклеиновыми кислотами. Укороченный на С-конце вариант ядерного белка (ак 1-149) экспрессировался в исходных экспрессионных системах Е.coli существенно более эффективно, чем полноразмерный белок. Поэтому CLP ядерного белка 1-149, т.е. без домена, связывающего нуклеиновую кислоту, использовали для исследований структур с помощью электронной микроскопии (ЭМ) умеренного и высокого разрешения. Это позволило определить кристаллическую структуру CLP и белка. Как и было предсказано на основе биохимических данных, ядерный белок образует очень стабильные симметричные димеры, которые собираются в ансамбль с образованием CLP, состоящих из 90 или 120 димеров.
Ядерные частицы имеют выраженные «шипы»; каждый димер образует «шип», структура которого представляет собой состоящий из 4 спиралей пучок, в котором каждый мономер имеет две длинные α-спирали, расположенные в центре аминокислотной последовательности. Кончики «шипов» содержат аминокислоты (ак), соответствующие области с ак 74 по ак 85. Эта область несет также иммунодоминантный B-клеточный эпитоп HBcAg (эпитоп с/е1), и ее очень высокая иммуногенность может объясняться тем, что указанные остатки «выставлены» на поверхность.
На фиг.1 схематично изображена структура ядерного белка. Первичная последовательность ядерного белка HBV изображена в левой части фиг.1. Полноразмерный белок состоит из 183 ак (некоторые подтипы HBV имеют 185 ак); для образования CLP необходимы только примерно 140 N-концевых ак (домен сборки); предпочтительно C-концевая ак находится в положении 149; His-метку можно сливать с этим положением и с любым другим последующим положением по ходу транскрипции вплоть до ак 183, не нарушая CLP. Эпитоп с/е1 расположен в области ак 78-83 и образует часть петли между центральными α-спиралями (внизу слева). Спирали сами по себе могут быть укороченными, по-видимому, минимально необходимыми являются аминокислотные остатки вплоть до положения Gly73 в coreN и ак-остатки, начиная с Gly94 в coreC. В правой части фиг.1 представлено схематическое изображение димера ядерного белка. Показаны только две центральные спирали мономера. Эпитоп с/е1 расположен на поверхности частицы.
Для успешного образования частиц аминокислотная последовательность ядра должна включать область ак по меньшей мере вплоть до положения 140, предпочтительно по меньшей мере до положения 149, но она может простираться дальше вплоть до C-концевой ак 183 или ак 185, в зависимости от подтипа HBV. Часть аминокислотной последовательности, расположенная по ходу транскрипции за положением 140, а лучше за положением 149, можно заменять чужеродной последовательностью, которая после этого, как правило, попадает внутрь CLP. Такие чужеродные аминокислоты не приводят к образованию основных антител, однако они могут выполнять другие функции.
Благодаря чрезвычайно высокой иммуногенности HbcAg рекомбинантные CLP HBV представляют интерес с точки зрения применения в качестве повышающего иммуногенность носителя чужеродных антигенов.
Альтернативным путем решения является химическое сочетание чужеродных молекул с заранее сформированными CLP. В настоящем изобретении предложено генетическое слияние чужеродных аминокислотных последовательностей с ядерным белком. На основе эмпирических данных даже до определения кристаллографической структуры с помощью рентгеновских лучей было установлено, что область эпитопа с/е1 наилучшим образом приспособлена для индуцирования сильных B-клеточных ответов. В настоящее время, принимая во внимание указанную структуру, стало очевидным, что такие слияния представляют собой вставки в область петли, находящуюся между центральными спиралями. Поскольку указанная петля сама по себе не несет никакой структурной функции, то 3D-структура ядерного белка может, по крайней мере в принципе, сохраняться после инсерции чужеродных последовательностей. В зависимости от встроенной чужеродной последовательности некоторые из слитых белков могут сохранять способность образовывать регулярные CLP, хотя это не всегда гарантировано, поскольку чужеродная аминокислотная последовательность может оказывать негативное влияние на формирование структуры слитого белка. С другой стороны, состоящая из отдельных частиц структура является важной для обеспечения высокой иммуногенности.
Ранее уже встраивали многочисленные чужеродные последовательности в ядерный белок и в некоторых случаях экспериментально было выявлено повышение иммуногенности (Ulrich и др., Adv. Virus Res., 1998, cc.141-182). Однако, поскольку многие попытки встраивания относительно длинных чужеродных последовательностей, состоящих более чем из примерно 40 ак, оказались неудачными (частицы переставали образовываться), то было сделано предположение о том, что существует обусловленная естественными факторами ограниченная способность к включению чужеродных последовательностей, т.е. их длина может составлять вплоть до 40 ак или в особом случае вплоть до 120 ак.
Было установлено, что чужеродные аминокислотные последовательности, слитые на С-конце и на N-конце с ядерным белком, образуют частицы только в том случае, когда они удовлетворяют по меньшей мере двум необходимым критериям:
(I) 3D-структура чужеродного белка должна быть устроена таким образом, чтобы его N-конец и С-конец соответствовали пространственному положению сайтов связывания в ядерном белке (С-концу N-концевого ядерного фрагмента, т.е. ядерным ак от примерно 1 до примерно 78 [coreN]; N-концу C-концевого ядерного фрагмента, т.е. ядерным ак от примерно 80 до 149 или 183 [coreC]);
(II) если чужеродный белок сам по себе способен к гомомерным взаимодействиям (с образованием димеров, тримеров и т.д.), то структура указанных гомоолигомеров, по-видимому, должна также соответствовать структуре обоих фрагментов ядерного белка.
Таким образом, чужеродные белки, N- и С-концы которых расположены близко друг к другу в нативной 3D-структуре, хорошо приспособлены для презентации в описанных ранее слитых белках. GFP удовлетворяет обоим этим требованиям, но многие другие белки не удовлетворяют им; одним из примеров таких белков является белок А наружной оболочки (поверхностный белок А, OspA) возбудителя болезни Лайма, т.е. Borrelia burgdorferi, N- и С-концы которого расположены на противоположных сторонах длинной 3D-структуры.
Встраивание OspA или других белков, имеющих аналогичную неблагоприятную структуру, приводит к возникновению напряжений в слитом белке. Могут иметь место следующие ситуации: либо чужеродный белок сохраняет правильную укладку и препятствует сближению двух фрагментов ядерного белка друг с другом, препятствуя тем самым их укладке и последующей димеризации и образованию частиц; либо укладка фрагмента ядерного белка мешает укладке чужеродного белка, так что она либо более не является нативной (т.е. изменяется антигенность), либо, если укладка в значительной степени отсутствует, то происходит агрегация.
В случае OspA эту проблему оказалось возможным преодолеть лишь частично за счет применения очень длинных линкерных последовательностей. Эти линкерные последовательности, прежде всего при их применении в вакцинах, могут вызывать проблему, заключающуюся в том, что они сами могут обладать нежелательной антигенностью, которая может приводить к вредным перекрестным реакциям, например, с эндогенными антигенами. Кроме того, соответствующие белковые препараты обладали лишь очень ограниченной способностью к образованию регулярных CLP.
Вследствие димерной структуры ядерного белка-носителя и геометрически ограниченной поверхности частиц-носителей в случае очень больших чужеродных белков или чужеродных белков, имеющих структуру, сильно отличающуюся от сферической, может дополнительно возникать проблема, обусловленная общей стерической помехой (пространство, доступное на «сферической поверхности» CLP, ограничено). Максимальный размер чужеродного белка зависит от нескольких факторов. С помощью способа, предлагаемого в изобретении, уже удалось презентовать чужеродный белок, состоящий примерно из 320 ак (CSP).
Таким образом, объектом настоящего изобретения является вакцина с расщепленным ядром, которая содержит в виде отдельных полипептидов coreN-домен и coreC-домен ядерного белка вируса гепатита В и по меньшей мере одну чужеродную аминокислотную последовательность, к которой требуется вызвать гуморальный и при необходимости клеточный иммунный ответ, где указанная чужеродная аминокислотная последовательность слита с С-концом coreN-домена или с N-концом coreC-домена и ядерный белок обладает способностью образовывать капсидоподобные частицы. Предпочтительно система-носитель обладает способностью вызывать ответ в виде нейтрализующих антител.
Очень важно, чтобы ядерный белок в вакцине на основе расщепленного ядра, предлагаемой в изобретении, был расщеплен, т.е. разделен в области эпитопа с/е1, т.е. примерно между аминокислотами 73 и 94. Затем чужеродную аминокислотную последовательность можно сшивать с С-концом coreN-области. В этом случае N-конец coreC-фрагмента начинается с аминокислоты, на которой был расщеплен coreN-белок. Можно также изымать путем делеции часть эпитопа с/е1, т.е. удалять одну или несколько аминокислот из области между аминокислотами 73 и 94. В альтернативном варианте можно чужеродную аминокислотную последовательность, к которой требуется вырабатывать антитела, сливать с N-концом coreC-области. В этом случае C-концевая область coreN-фрагмента оканчивается аминокислотой, в которой был расщеплен ядерный белок. Основное свойство, которое должна сохранить вакцина на основе расщепленного ядра, заключается в том, что частицы вакцины на основе расщепленного ядра, несущие слитые с ними чужеродные аминокислотные последовательности, все еще должны обладать способностью образовывать ядерные капсидоподобные частицы. Это проверяют либо с использованием градиента сахарозы, либо с помощью нативного электрофореза в агарозном геле. Можно применять также анализ с использованием электронного микроскопа.
Чужеродная аминокислотная последовательность, к которой требуется вырабатывать антитела, предпочтительно представляет собой последовательность, происходящую из поверхностных структур микроорганизмов. Такие микроорганизмы, как правило, вызывают заболевание у человека. Если могут образовываться нейтрализующие антитела к таким структурам, то это приводит к тому, что иммунная защита может очень быстро удалять проникшие микроорганизмы. Примером бактерии, разработка вакцины против которой продвинулась уже довольно далеко, является Borrelia burgdorferi, возбудитель болезни Лайма. Чужеродные аминокислотные последовательности, которые предпочтительно применяют согласно изобретению, представляют собой последовательности поверхностных белков OspA и OspC Borrelia burgdorferi. Однако чужеродные аминокислотные последовательности могут представлять собой также другие последовательности, происходящие из патогенных организмов. Их примером может служить возбудитель малярии Plasmodium falciparum.
Другими чужеродными аминокислотными последовательностями, к которым могут вырабатываться антитела, являются последовательности, происходящие из вирусов. Предпочтительно применяют аминокислотные последовательности, происходящие из белков вирусов, которые вступают в контакт с иммунной системой организма-хозяина. Такими белками являются, прежде всего, поверхностные белки, поскольку, как правило, они первыми вступают в контакт с иммунной системой организма-хозяина. Однако они могут представлять собой также белки, которые высвобождаются в процессе жизненного цикла вируса. В зависимости от типа вируса они могут представлять собой, например, вирусные ядерные белки или нуклеокапсидные белки.
Однако предлагаемую в настоящем изобретении вакцину с расщепленным ядром можно применять для создания не только антител к экзогенным аминокислотным последовательностям, но также и антител к нежелательным эндогенным структурам, таким, например, как маркеры опухоли. Опухолевые клетки часто экспрессируют поверхностные маркеры, отличные от маркеров, экспрессируемых здоровыми клетками. Таким образом, если в вакцине на основе расщепленного ядра презентуются эндогенные аминокислотные последовательности, то вызывается иммунный ответ, заставляющий организм вырабатывать в увеличенном количестве антитела к указанным опухолевым клеткам. С помощью таких антител иммунная защита может легко распознавать и в конце концов уничтожать опухолевые клетки. Однако система-носитель, предлагаемая в изобретении, может индуцировать также клеточный иммунный ответ, что может повышать эффективность борьбы.
Настоящее изобретение относится также к способу получения вакцины против гетерологичной белковой последовательности. Вакцина с расщепленным ядром содержит две части ядерного антигена вируса гепатита В, причем либо гетерологичная белковая последовательность слита с coreN-областью белка, либо coreC-область слита с С-концом гетерологичной белковой последовательности. Положение в аминокислотной цепи, в котором два домена отделяют друг от друга, находится между положением 73 и положением 94. Индивидуальные аминокислоты могут быть удалены из этой области, представляющей собой эпитоп с/е1.
Способ, предлагаемый в изобретении можно осуществлять в принципе двумя различными путями. Если две части антигена с расщепленным ядром экспрессируются в виде только одной полипептидной цепи, то последовательность, распознаваемую протеазой, встраивают в требуемое положение согласно изобретению. Это позволяет расщеплять полипептид протеазой на две определенные части. В этом варианте осуществления изобретения можно также осуществлять коэкспрессию протеазы, которая расщепляет последовательность в определенном сайте. Последовательность гена, кодирующая ее, может располагаться либо на том же самом векторе, либо на отдельном векторе.
В другом предпочтительном варианте осуществления изобретения два указанных выше полипептида экспрессируют с использованием специального вектора. Предпочтительно применяют «бицистронный» вектор. Это означает, что одну часть вакцины с расщепленным ядром экспрессируют в виде полипептидной цепи и что стоп-кодон расположен на С-конце. Однако вскоре после этого можно снова применять рибосомы и осуществлять экспрессию второго полипептида. Это гарантирует то, что обе части вакцины с расщепленным ядром продуцируются примерно в равных количествах и не требуется принимать меры для того, чтобы организм-хозяин не потерял один из векторов, что может привести к тому, что будет присутствовать только одна часть вакцины с расщепленным ядром.
Понятие «слитый белок» или «слитый с» означает, что два различных белка или полипептида связаны друг с другом с помощью пептидной связи. Такие слитые белки образуются в результате экспрессии кодирующих их нуклеотидных последовательностей, которые присоединяют друг за другом.
При создании настоящего изобретения было установлено, что основная связанная с химией белка проблема с точки зрения широкой применимости системы-носителя HBV-CLP (конкретно в качестве носителя вакцины, как правило, для презентации чужеродных молекул) возникает при двухстороннем сцеплении встроенного чужеродного белка на его N- и С-концах. Количество и тип чужеродных белков, которые можно встраивать, резко возрастает в том случае, если оказывается возможным освободить любую одну из двух ковалентных связей (на N- или С-конце вставки) и, если, тем не менее, при этом происходит образование частиц, поскольку разделенные фрагменты ядерного белка («расщепленного ядра») обнаруживают друг друга и правильно укладываются. Это является важным объектом настоящего изобретения.
На фиг.2 схематично показано, что встраивание чужеродного белка, имеющего неблагоприятную 3D-структуру (например, OspA), может препятствовать формированию компетентной в отношении образования частиц структуры ядерного белка-носителя (верхняя часть схемы); в другом варианте образование правильной 3D-структуры носителя ядерного белка может препятствовать образованию нативной 3D-структуры чужеродного белка (нижняя часть схемы). Это может оказывать негативное воздействие на связывание требуемых антител. Эту стерическую проблему можно устранять, если освободить (обозначено стрелкой) одну из двух ковалентных связей между вставкой, представляющей собой чужеродный белок, и носителем (на С-конце или на N-конце вставки). Одним из возможных путей достижения этого является последующее расщепление непрерывного слитого белка, для чего необходимо дополнительно встраивать сайт, расщепляемый специфической протеазой. Поэтому предпочтительно с самого начала осуществлять экспрессию двух фрагментов в виде отдельных единиц.
Решение, предлагаемое в изобретении, открывает также и другие возможности:
(I) поскольку эпитоп с/е1 расположен на поверхности частицы, то на этой поверхности образуются новые N- и С-концы, которые можно дополнительно дериватизировать. Например, чужеродную молекулу Х можно сливать с coreN-фрагментом и чужеродную молекулу Y можно сливать с coreC-фрагментом. Возможным применением является презентация гетеродимерных чужеродных молекул. Однако следует иметь ввиду, что в этом случае не должно быть стерической помехи для обеих частей, присоединяемых путем слияния.
(II) специфические подобласти встроенной чужеродной последовательности, например, особенно важный эпитоп, могут быть ориентированы по направлению к поверхности частицы или от нее, в зависимости от сцепления через N-конец или С-конец. Одним из примеров является так называемый «LA2-эпитоп» OspA В. burgdorferi, который, как было установлено, находится в C-концевой области. Антитела, которые распознают этот эпитоп, обладают нейтрализующим действием.
Это важно также для встраивания чужеродных последовательностей, которые, в свою очередь, презентуют дополнительные поверхности для взаимодействия (для присоединения третьих молекул, см. примеры). Поскольку обладающая способностью к взаимодействию чужеродная последовательность сцеплена только на одной стороне, то ее не ограничивает никакая 3D-структура; напротив, в зависимости от типа, она может обладать гибкостью или может без помех образовывать правильную 3D-структуру. Это существенно облегчает взаимодействия с партнером по взаимодействию.
Настоящее изобретение относится также к лекарственным средствам, прежде всего вакцинам, которые содержат капсидоподобные частицы на основе системы с расщепленным ядром.
Лекарственные средства, предлагаемые в изобретении, содержат капсидоподобные частицы, предлагаемые в изобретении, и они предпочтительно оказывают положительное воздействие на иммунную систему. Как правило, система-носитель с расщепленным ядром, предлагаемая в изобретении, презентует иммунной системе чужеродную аминокислотную последовательность, к которой иммунная система вырабатывает антитела, предпочтительно нейтрализующие антитела. Затем эти антитела принимают решающее участие в борьбе с микроорганизмом или вирусом, проникшим в организм, или они способствуют специфическому разрушению нежелательных опухолевых клеток.
В другом варианте осуществления изобретения систему-носитель с расщепленным ядром, предлагаемую в изобретении, можно применять в качестве диагностического и/или аналитического средства. В примерах 12 и 13 такие применения проиллюстрированы более подробно. С помощью таких средств можно, например, сделать доступными обладающие собственной флуоресценцией и связывающие антитела частицы, которые, со своей стороны, связываются с антигеном и тем самым позволяют обнаруживать его с помощью флуоресценции. В другом варианте осуществления изобретения система-носитель с расщепленным ядром, предлагаемая в изобретении, может содержать пептидные последовательности, которые специфически связываются с определенным ионами металлов/лантанидами. Если такие пептиды встраивают в систему с расщепленным ядром, то получают частицы, которые содержат много репортерных атомов/ионов такого рода на одну частицу и поэтому их легче обнаруживать.
Пример 1
Ядерный белок дикого типа с встроенной последовательностью, распознаваемой TEV-протеазой
Последовательность, распознаваемую протеазой вируса гравировки табака (TEV), встраивали в эпитоп с/е1 ядерного белка дикого типа (wt) 1-149 (замена ак Р79+А80 на GGGGT-ENLYFQGT-GGGG; применение G-остатков в качестве линкеров гарантировало доступность распознаваемой последовательности для протеазы). Рекомбинантный экспрессируемый белок образовывал частицы. Их инкубировали в присутствии протеазы TEV и успех реакции расщепления проверяли с помощью ДСН-ПААГ. Затем смесь, полученную в результате расщепления протеазой, осаждали в градиенте сахарозы. Происходило практически полное расщепление в ожидаемом положении, поскольку образовывались два фрагмента почти одинаковой длины. Несмотря на расщепление, белок осаждался в градиенте практически в том же положении, что и нерасщепленный ядерный wt-белок 1-149. Сопоставимые результаты получали с использованием слитого белка на основе полноразмерного ядерного белка 1-183.
Пример 2
Варианты ядерного белка с более крупными встроенными чужеродными последовательностями
В качестве примера для получения слитого белка с несколько большей вставкой последовательность искусственного пептида «ACID» (фланкированную богатыми Gly линкерами; всего 65 ак), которая может взаимодействовать с комплементарным пептидом «BASE», встраивали в эпитоп с/е1 (O'Shea и др., Curr. Biol., 1993, cc.656-667); после чего за ней по ходу транскрипции встраивали последовательность, распознаваемую TEV-протеазой GGGGSGGGVEDGGGGSGGGGT-AQLEKELQALEKENAQLEWELQALEKELAQTG-ENLYFQGTGGGG.
Слитый белок образовывал CLP. Их выделяли и инкубировали, как описано выше, с TEV-протеазой. И в этом случае также происходило специфическое расщепление, но частицы оставались интактными. В этом случае фрагменты, полученные в результате расщепления с помощью TEV-протеазы, имели разные размеры, и поэтому их можно было различать непосредственно в ДСН-геле. Несмотря на расщепление, происходило совместное осаждение фрагментов в специфических для CLP градиентных фракциях.
Этот пример демонстрирует, что CLP, состоящие из ядерного белка с встроенным пептидом длиной 65 ак (линкер + ACID + сайт, расщепляемый TEV-протеазой), также можно специфически расщеплять, сохраняя тем не менее структуру в виде частиц.
Пример 3
В качестве другого примера описан соответствующий эксперимент с использованием ядерного белка, содержащего аминокислотную последовательность 18-273 белка OspA Borrelia burgdorferi. Соответствующий белок без сайта, расщепляемого протеазой TEV, описан у Nassal и др., 2005; лишь некоторая часть его образовывала регулярные частицы, что было видно также по широкому распределению в градиенте сахарозы. Таким образом, вакцину на основе нерасщепленного ядра можно применять лишь в ограниченной мере. Кроме того, эту вакцину нельзя применять на людях/в медицинских целях, поскольку для получения по меньшей мере небольшой части препарата в форме CLP требуются очень длинные внешние линкерные последовательности, которые обладают собственным нежелательным антигенным потенциалом.
После встраивания в сайт, расщепляемый протеазой TEV, слитый белок обрабатывали протеазой TEV, как описано в примере 2, и затем осаждали в градиенте сахарозы. В этом случае также происходило специфическое расщепление, однако оно не было полным (предположительно стерическая помеха, вызываемая OspA, приводила к плохой доступности сайта, расщепляемого протеазой TEV); осуществляли классификацию фрагментов с помощью моноклональных антител, которые специфически распознают N-концевой или C-концевой ядерный фрагмент; следовательно, антитело к coreN реагирует с нерасщепленным слитым белком, а также с N-концевым отщепленным фрагментом. Тем не менее, частично расщепленный продукт обладал типичными для частиц миграционными характеристиками. Эти данные свидетельствуют о том, что образование частиц можно существенно улучшать путем размыкания одной из двух связей между встроенным чужеродным белком и ядерным носителем.
Этот пример показывает, что слитый ядерный белок, несущий вставку в виде большого чужеродного белка (состоящий из 255 ак OspA + линкер + сайт, расщепляемый протеазой TEV), можно специфически расщеплять (хотя и менее эффективно). В то время как описанный выше слитый белок непрерывное ядро-OspA характеризовался широким распределением в градиенте плотности (Nassal и др., 2005), (частично) расщепленный слитый белок выражено концентрируется в градиентных фракциях, типичных для частиц. Это свидетельствует о том, что эффективность образования частиц повышается в результате расщепления.
В другом эксперименте продемонстрировано, что коэкспрессируемая (из совместимой плазмиды) протеаза TEV расщепляет слитые ядерные белки, несущие сайты, расщепляемые протеазой TEV, иногда более эффективно, чем это возможно при последующем расщеплении in vitro. С помощью осаждения в градиенте сахарозы можно обнаружить, что и слитые белки, которые уже были расщеплены в бактериях, также образуют частицы.
В примерах 1-3 продемонстрировано, что предварительно сформированные CLP остаются интактными после расщепления в области эпитопа с/е1. С точки зрения структуры это можно объяснить тем фактом, что соединительная петля между двумя образующими «шипы» спиралями сама по себе не играет структурообразующую роль, хотя этого нельзя было ожидать. Это приводит к более предпочтительному варианту осуществления изобретения, в котором вместо последующего расщепления непрерывной белковой цепи coreN- и coreC-фрагменты экспрессируют непосредственно в виде отдельных фрагментов белка, очевидно, что при этом они спонтанно встречаются друг с другом и образуют частицы. Такой подход имеет по меньшей мере три существенных преимущества:
(I) упрощение: не требуется дополнительной стадии расщепления с помощью протеазы TEV (или другой протеазы);
(II) не требуется дополнительной пептидной последовательности в слитом белке для специфического распознавания протеазой;
(III) образование частиц, даже если оно происходит только частично, с использованием стерически неблагоприятного чужеродного белка, такого как OspA, требует применения очень длинных линкерных последовательностей; от них можно отказаться, если устранить стерические помехи, поскольку в этом случае не требуется присоединение на обоих концах к носителю-ядерному белку.
Пункты (II) и (III) являются особенно важными прежде всего для применений в вакцинах, поскольку любая дополнительная последовательность может приводить к непредсказуемым иммунологическим последствиям (новые эпитопы, присутствующие в дополнительных последовательностях, возможные перекрестные реакции с эндогенными эпитопами).
Пример 4
Экспрессионные конструкции для экспрессии фрагментов coreN и coreC в примерно эквимолярных количествах
Непрерывная пептидная цепь содержит продукты, которые в дальнейшем должны быть отщеплены, безусловно в эквимолярных количествах. Следовательно, при раздельной экспрессии две части должны продуцироваться аналогичным образом примерно в эквимолярных количествах для обеспечения эффективной сборки. В первых экспериментах для экспрессии двух частей белка-носителя применяли две отдельные совместимые плазмиды, однако лишь с ограниченным успехом.
В предпочтительном варианте осуществления изобретения применяют бицистронные векторы, которые обладают, кроме того, тем преимуществом, что в этом случае можно избежать отбора с помощью другого антибиотика (в ином случае для получения необходима вторая плазмида).
Конструкции типа 1 содержат расположенные за общим промотором (в данном случае за промотором РНК-полимеразы фага Т7) две кассеты экспрессии, перед каждой из которых расположен сайт связывания рибосом (RBS; последовательность Шайна-Дальгарно). В векторах, предложенных в настоящем изобретении, трансляция фрагмента coreN прекращается искусственным стоп-кодоном, расположенным за ак пролином 79 (Р79), и искусственный стартовый кодон, расположенный перед ак серином 81 (S81), позволяет инициировать трансляцию второго фрагмента. Такая структура оперона часто встречается в бактериях («полицистронная мРНК»; рибосомы могут связываться с каждым RBS на мРНК относительно независимо и инициировать трансляцию гена, локализованного на 3'-конце). В предложенных конструкциях второй RBS является точной копией первого RBS, который выведен из исходных векторов рЕТ; безусловно можно аналогичным образом применять другие последовательности RBS. Стартовый кодон ATG второго цистрона является частью Nde-сайта рестрикции (CATATG), что упрощает клонирование чужеродных последовательностей.
Конструкции типа 2 («стоп/старт-конструкции») не содержат отдельный второй RBS; вместо этого инициирующий кодон расположенного относительно него по ходу транскрипции гена (NNN TGA) перекрывается стоп-кодоном расположенного относительно него против хода транскрипции гена (NNA TGA) в смещенной на 1 рамке считывания. В этом случае рибосомы могут после считывания первого гена сразу реинициироваться на ATG второго гена; такой порядок расположения позволяет осуществлять совместную экспрессию (коэкспрессию) двух генов; аналогичные порядки расположения стоп-/старт-конструкций можно обнаружить в некоторых бактериофагах. Они присутствуют также в некоторых эукариотических не содержащих LTR-ретротранспозонах (стык открытых рамок считывания ОРС1/ОРС2). На основе известных последовательностей, например, в опероне Trp E.coli, можно применять другие комбинации смежных или перекрывающихся стоп-/старт-кодонов (например, ТАА TG, TGA TG и т.д.).
Предпочтительные экспрессионные векторы схематично изображены на фиг.3.
Вверху: Конструкции типа 1. Промотор, например, промотор РНК-полимеразы фага Т7, контролирует транскрипцию бицистронной мРНК (для эффективной терминации можно встраивать терминирующую последовательность на его 3'-конец). Первый цистрон кодирует аминокислотную последовательность 1-79 ядерного белка HBV. Расположенный против хода транскрипции сайт связывания рибосом (RBS1) обеспечивает инициацию трансляции (малая и большая субъединицы рибосомы изображены овалами). Стоп-кодон расположен далее по ходу транскрипции на 3'-конце кодона Р79. На 5'-конце второго цистрона распложен другой RBS (RBS2), необходимый для инициации трансляции coreC-фрагмента. Этот фрагмент дополнительно содержит собственный кодон инициации (ATG). В данном примере последовательность coreC начинается с А1а80 и она может простираться до ак в положении 140, более предпочтительно до положения 149 или до действительного конца, находящегося в положении 183. Участок аминокислотной последовательности от положения 140, предпочтительно от положения 149, и далее можно также заменять чужеродными последовательностями.
Внизу: Конструкции типа 2. Эти конструкции содержат только один RBS, расположенный против хода транскрипции относительно первого цистрона. Трансляция coreC-фрагмента происходит в результате реинициации, поскольку, как проиллюстрировано, искусственный стартовый кодон фрагмента coreC (ATG) перекрывается стоп-кодоном фрагмента coreN (TGA). В данном примере последовательность coreC начинается с Ser81.
Аминокислоты 77, 78 и 79 ядерного белка представляют собой глутамат (Е; кодоны: GAA, GAG), аспартат (D; кодоны: GAC или GAT), ак 79 представляет собой пролин (Р; кодоны: CCN). В созданных конструкциях типа 1 в качестве аминокислотной последовательности EDP была выбрана нуклеотидная последовательность GAG GAT CCN, что позволило создать сайт рестрикции BamHI (GGATCC); этот уникальный сайт рестрикции BamHI позволяет осуществлять простое клонирование различных ДНК, кодирующих coreN- и coreC-фрагменты. Нуклеотид N в кодоне пролина CCN не является строго определенным; стоп-кодон может представлять собой ТАА, TAG или TGA.
Последовательность, простирающаяся до конца первого гена в конструкциях типа 2, представляет собой G GAT CCa TGA. Эта последовательность также содержит сайт рестрикции BamHI (подчеркнут) для клонирования различных ДНК, кодирующих coreN- или coreC-фрагменты. Р79 в созданных конструкциях типа 2 должен иметь кодон Сса, а стоп-кодон должен представлять собой TGA (>CCa TGa) для того, чтобы мог быть создан перекрывающий инициирующий кодон ATG для второго гена. Кодон для первой ак второго гена после стартового кодона ATG должен начинаться с А. Альтернативные стоп-/старт-нуклеотидные последовательности могут представлять собой TGA TG или ТАА TG, как это имеет место в опероне Trp Е.coli между TrpE и TrpD или также между TrpB и TrpA (Das и Yanofsky, Nucl. Acids Res., 1989, cc.9333-9340; Oppenheim и Yanofsky, Genetics, 1960, cc.785-795).
Каркасом созданных векторов (рЕТ28а2-ххх) служил имеющийся на рынке вектор рЕТ-28, в котором, однако, ген, обусловливающий устойчивость к канамицину, был заменен на ген, обусловливающий устойчивость к ампициллину. Однако, безусловно можно применять также другие каркасные векторы (имеющие различные сайты инициации репликации, различные гены, обусловливающие устойчивость, различные промоторы, различные RBS). Важным является бицистронный порядок расположения генов coreN и coreC, реализуемый либо с помощью двух раздельных RBS, либо посредством определенного порядка расположения стоп-/старт-кодонов. В экспериментах, проведенных при создании изобретения, оказалось возможным осуществлять экспрессию нескольких белков с расщепленным ядром, имеющих конструкцию как типа 1, так и типа 2. Однако в данном случае количество конструкций типа 2 (стоп-/старт-конструкции) было меньше. Кроме того, для конструкций типа 2 возможность вариации последовательности была более ограниченной (см. выше). Таким образом, конструкции типа 1 могут найти более широкое применение.
В качестве C-концевой ак coreN-фрагмента был выбран Р79, поскольку пролин наиболее устойчив к воздействию протеазы и тем самым минимизируется расщепление фрагмента. На основе известной 3D-структуры и описанных вариантов делеций в указанной центральной области можно предположить, что
(I) конец coreN-фрагмента может располагаться в области ак, находящихся примерно между положениями 72 и 74, прежде всего на остатке Gly73,
(II) начало coreC-фрагмента может располагаться в области ак, находящихся примерно между положениями 78 и 86, возможно вплоть до Gly94, причем это не приводит к существенному изменению результатов по сравнению с теми, которые получают при использовании предельных выбранных для них положений (конец coreN на Р79, начало coreC на S81, с расположенным против хода транскрипции относительно него инициирующим кодоном ATG).
Вследствие различных механизмов транскрипции и трансляции описанные конструкции нельзя применять в такой форме для экспрессии в эукариотических клетках; вместо прокариотического промотора необходимо использовать эукариотический промотор (например, можно применять немедленно-ранний промотор цитомегаловируса (CMV-IE) и многие другие), а также необходимо встраивать сигнал полиаденилирования (например, из вируса SV40 или многие другие сигналы) по ходу транскрипции после конца открытой(ых) рамки(ок) считывания.
Примерно одинаковый уровень экспрессии фрагментов coreN и coreC в эукариотических организмах можно обеспечивать путем:
(I) котрансфекции двумя плазмидами (одной для coreN, одной для coreC);
(II) трансфекции плазмидой с двумя независимыми кассетами экспрессии;
(III) трансфекции плазмидой с функциональной бицистронной мРНК;
например, второй ген можно экспрессировать с помощью расположенного выше против хода транскрипции внутреннего рибосомального сайта проникновения (IRES);
(IV) порядок расположения стоп-/старт-конструкций, аналогичный тому, который имеет место в векторе типа 2, может функционировать также и в эукариотических организмах.
Пример 5
Раздельная экспрессия wt-coreN- и -coreC-фрагментов приводит к образованию интактных CLP
Ядерный белок дикого типа экспрессировали в виде двух частей с использованием экспрессионных векторов на основе конструкции типа 1 (фиг.3) в клетках Е.coli BL21, в одном случае последовательность включала ак 1-149, в другом случае последовательность включала ак 1-183. Продуцировались оба ожидаемые фрагменты и происходила их сборка с образованием интактных CLP. Это было установлено с помощью седиментации в градиентах сахарозы и нативного гель-электрофореза в агарозе. Образование CLP выявляли непосредственно, осуществляя сравнение полученных с помощью электронного микроскопа изображений (негативное окрашивание) частиц, которые содержали непрерывную цепь из ак 1-183, несущую C-концевую His6-метку, и частиц, содержащих систему с расщепленным ядром. При таком уровне разрешения различий не было выявлено.
Wt-HBV-CLP были очень робастными и обладали устойчивостью к мочевине в молярных концентрациях; CLP, состоящие из укороченного на С-конце ядерного белка 1-149, можно было подвергать диссоциации с помощью 3-5М мочевины с образованием димеров и осуществлять повторную ассоциацию in vitro с образованием CLP путем изменения буферных условий (удаление мочевины, обеспечение нейтрального значения рН, повышение концентрации соли). Сравнительное исследование стабильности CLP, содержащих расщепленное ядро, продемонстрировало сопоставимую высокую стабильность CLP с расщепленным ядром. Это свидетельствует о высокой взаимной аффинности coreN- и coreC-фрагментов, что является очень важным с точки зрения применения в качестве системы носителя для чужеродных молекул.
Пример 6
Раздельная экспрессия слитого с чужеродным белком coreN- и wt-coreC-фрагментов и наоборот позволяет получать интактные CLP
Для доказательства того, что система с расщепленным ядром в принципе пригодна для образования CLP и презентации чужеродных белков, применяли в качестве модели GFP, слитый либо с coreN, либо с coreC; для создания конструкции coreN использовали либо вектор типа 1 (несущий второй RBS), либо вектор типа 2 (несущий стоп-/старт-конструкцию). Все конструкции позволяли осуществлять экспрессию белков, обладающих зеленой флуоресценцией, которые образовывали частицы согласно результатам анализа методом седиментации в градиенте сахарозы.
Для того чтобы получить дополнительное подтверждение физической ассоциации соответствующих coreN- и coreC-фрагментов, аликвоты градиентных фракций подвергали электрофорезу в нативных агарозных гелях. В этом случае частицы сохраняются интактными так же, как и хромофор GFP. Фракции, содержащие как GFP, связанный с coreN, так и GFP, связанный с coreC, давали отчетливую зеленую флуоресцентную полосу; слитый белок, не содержащий GFP, оставался в верхних градиентных фракциях и имел другие миграционные характеристики и отличался более диффузным распределением (более быстрая диффузия в геле вследствие существенно меньшего размера структуры по сравнению с частицами, состоящими из 180 или 240 субъединиц). CoreC обнаруживали также с помощью антитела к α-coreC в зеленой флуоресцентной полосе (т.е. содержащей GFP) как в случае GFP, связанного с coreN, так и GFP, связанного с coreC.
В этом примере продемонстрированы два момента.
(I) В системе с расщепленным ядром чужеродный белок, состоящий примерно из 240 ак, в данном случае GFP, можно сливать как с coreN, так и с coreC, не нарушая образование CLP.
(II) Векторы с расщепленным ядром как типа 1, так и типа 2, можно применять для коэкспрессии coreN и coreC, содержащих слитую с ними чужеродную последовательность.
Пример 7
Презентация с помощью CLP с использованием системы с расщепленным ядром имеющих медицинское значение чужеродных белков, которые нельзя или можно лишь в ограниченной степени презентовать с помощью обычных средств
OspA Borrelia burgdorferi
Как указано выше, сцепление чужеродного белка с белком-носителем на обоих концах, которое необходимо при использовании известной в настоящее время системы, приводит к значительным топологическим ограничениям. В то время как GFP обладает естественной «подходящей формой», OspA не обладает таковой. Таким образом, OspA использовали в качестве примера чужеродного белка, имеющего неблагоприятную структуру, который можно применять в системе с расщепленным ядром, и его сливали либо с coreN, либо с coreC. В отличие от применявшихся ранее непрерывных конструкций (описанных у Nassal и др., 2005), которые отличались широким распределением в градиенте, оба слитых белка выражено концентрировались во фракциях, типичных для частиц. Анализ с помощью электронной микроскопии продемонстрировал существенно более высокую эффективность образования CLP как для слияния OspA с coreN, так и для слияния OspA с coreC, по сравнению с применявшейся ранее непрерывной конструкцией.
Этот пример доказывает, что
(I) система с расщепленным ядром позволяет осуществлять презентацию с помощью CLP чужеродных белков, структура которых мешает образованию CLP в обычной системе с непрерывным ядром,
(II) система с расщепленным ядром позволяет осуществлять эффективное формирование CLP путем слияния чужеродного белка как с coreN, так и с coreC.
Аналогично этому оказалось возможным также успешно презентовать OspC Borrelia burgdorferi с использованием системы с расщепленным ядром, предлагаемой в изобретении.
Пример 8
Вокругспорозоитный белок возбудителя малярии Plasmodium falciparum
Другим патогенным белком, имеющим очень большую важность с точки зрения создания вакцин, является вокругспорозоитный белок (CSP) возбудителя малярии Plasmodium falciparum. CSP (в данном случае использовали форму, имеющую общую длину 319 ак) содержит состоящий примерно из 110 ак повтор тетрапептидного мотива NANP/NVDP. Структура CSP до настоящего времени не выяснена; состоящий примерно из 50 ак C-концевой домен, с 4 остатками Cys, по-видимому (на основе гомологии последовательности), укладывается аналогично повторам тромбоспондина типа 1. Хотя иммуногенной является последовательность повтора, однако другие области CSP также могут содержать важные эпитопы. С использованием обычной системы с непрерывным ядром оказалось невозможным включить полноразмерный CSP в CLP-форму. В то же время это легко удалось сделать с использованием системы с расщепленным ядром. В настоящее время получены данные, однозначно свидетельствующие о слиянии CSP с coreN; коэкспрессия с coreC (как в случае конструкции, содержащей 149 ак, так и конструкции, содержащей 183 ак) позволяла эффективно получать состоящие из частиц структуры.
Получали серию конструкций (как на основе ядра 1-149, так и на основе ядра 1-183), которые содержали только одну последовательность повтора или укороченный CSP без богатого Cys домена, в каждом случае в системе с непрерывным или расщепленным ядром; кроме того, полный CSP сливали с coreN. CLP-препараты применяли при проведении сравнительных исследований иммуногенности на мышах, которые, однако, к настоящему времени еще не завершены. Полученные к настоящему моменту времени данные свидетельствуют о том, что вакцина, созданная на основе системы с расщепленным ядром, обладает более высокими характеристиками по сравнению с существующими вакцинами. Это объясняется тем, что полный CSP может образовывать CLP только при использовании системы-носителя, предлагаемой в изобретении. В отличие от этого при использовании других систем CLP не образуются. В результате этого CSP-вакцины, полученные с использованием системы-носителя, предлагаемой в изобретении, могут обладать более высокой иммуногенностью и индуцировать образование прежде всего нейтрализующих антител.
Пример 9
CLP с расщепленным ядром в качестве носителей для чужеродных белков усиливают В-клеточный ответ на чужеродный белок
В исследовании иммуногенности на мышах проводили сравнение 5 конструкций ядро-OspA, созданных на основе системы с расщепленным ядром, с липидированным OspA (LipOspA), который не был слит с ядром; последний является основой имеющихся на рынке вакцин Lymerix, которые в настоящее время считаются «золотым стандартом» для вакцин против болезни Лайма. Липидный фрагмент в LipOspA (трис-пальмитоилцистеин-(Pam3-Cys)) имеет важное значение для его относительно высокой иммуногенности, нелипидированный OspA обладает лишь очень слабой иммуногенностью (см. у Nassal и др., 2005). Три конструкции ядро-OspA содержали полноразмерный OspA (ак 18-273), а другие две содержали укороченный OspA (начинающийся на ак 185, простирающийся до ак 273).
Для данной цели 6 групп мышей, каждая из которых состояла из 5 мышей линии BALB/c, иммунизировали 4 раза, вводя каждый раз по 10 мкг антигена (в дни 0, 14, 29, 49):
Группа 1: LipOspA;
Группа 2: полноразмерный OspA (ак 18-273), слитый с coreN, coreC содержал ядерные ак с 81 по 183;
Группа 3: полноразмерный OspA (ак 18-273), слитый с coreN, coreC содержал ядерные ак с 81 по 149;
Группа 4: укороченный OspA (ак 185-273), слитый с coreN, coreC содержал ядерные ак с 81 по 183;
Группа 5: укороченный OspA (ак 185-273), слитый с coreN, coreC содержал ядерные ак с 81 по 149;
Группа 6: полноразмерный OspA (ак 18-273), слитый с coreC на ядерной ак 183, coreN содержал ак 1-79.
Для выявления индуцированных антител брали образцы крови в день -1 (т.е. за 1 день до первой иммунизации; неиммунная сыворотка) и в дни 8, 26, 36 и 57. С помощью ELISA оценивали следующие параметры: кинетику продуцирования специфических в отношении OspA антител; соответствующую пропорцию LA2-эквивалентных антител, LA2 представляет собой моноклональное антитело, в отношении которого известно, что оно обладает нейтрализующей способностью и распознает комплексный конформационный эпитоп в прерывистых аминокислотных последовательностях ниже по ходу транскрипции относительно ак 185 OspA.
Результаты этого эксперимента подробно представлены на фиг.4.
На фиг.4А представлены данные о кинетике индукции общих антител к OspA (в мкг специфического антитела (Ат) на мл сыворотки). Уже после второй иммунизации сыворотка мышей, иммунизированных конструкциями на основе расщепленного ядра, содержащими полноразмерный OspA, имела обнаруживаемые титры антител к OspA, которые были сопоставимы с титрами, вызываемыми LipOspA. После 3-й и 4-й иммунизации титры антител к OspA у мышей, иммунизированных конструкциями на основе расщепленного ядра, содержащими полноразмерный OspA, выражено превышали титры у мышей, иммунизированных LipOspA; особенно высокие титры достигались после иммунизации конструкцией, содержащей OspA18-273, слитый с coreC (группа 6). Укороченный OspA185-273 в контексте соге149 (группа 5) и core183 (группа 4) вызывал лишь существенно более слабый специфический в отношении OspA ответ.
На фиг.4Б представлено прямое сравнение общих титров Ат к OspA с титрами антител к эпитопу нейтрализующего LA2 в C-концевой области OspA. Полноразмерный OspA, слитый с coreN, в контексте как core149, так и core183 (группы 2 и 3 соответственно), индуцировал более высокие титры Ат, эквивалентного антителу LA2 по сравнению с LipOspA, а полноразмерный OspA, слитый с coreC, вызывал существенно меньшие титры. Исследование соответствующего содержания (титров) нейтрализующих Ат (обеспечение защиты от заражения возбудителем В. burgdorferi) в настоящее время продолжается.
Этот пример демонстрирует, что CLP с расщепленными ядрами индуцируют специфические антитела к презентируемому полноразмерному чужеродному белку OspA, причем их титры выше, чем в случае применения используемого в настоящее время липидированного LipOspA, в котором наличие липидного фрагмента является важным для иммуногенности. Различные пропорции Ат, эквивалентных LA2, которые вырабатываются после иммунизации с использованием OspA, слитого с coreN (примерно 30%), по сравнению с иммунизацией с использованием OspA, слитого с coreC (<5%) свидетельствуют о том, что тип связи влияет на тип вырабатываемых Ат (см. пример 10).
Пример 10
Направленная индукция специфических в отношении определенной области антител к чужеродному белку в результате альтернативного слияния либо с coreN, либо с coreC
В случае белков, N- и С-концы которых не находятся в непосредственной близости друг к другу, как это имеет место в GFP, слияние с coreN приводит к ориентации по отношению к поверхности частицы, отличной от той, которая возникает при слиянии с coreC (coreN: С-конец чужеродного белка «снаружи»; coreC: N-конец чужеродного белка «снаружи»). Следует ожидать, что при применении в качестве В-клеточных вакцин те фрагменты белка, которые простираются в каждом случае наиболее далеко в растворитель, будут вызывать наиболее сильный гуморальный иммунный ответ. Таким образом, можно индуцировать различные специфические в отношении определенной области антитела путем слияния чужеродного белка либо с coreN (С-конец чужеродного белка «снаружи») или с coreC (N-конец чужеродного белка «снаружи»).
На фиг.5 для чужеродного белка с вытянутой структурой, такого как представленный в настоящем описании OspA, схематично изображен возможный путь определения посредством слияния либо с coreN, либо с coreC, какая часть молекулы наиболее далеко отходит от поверхности CLP. Хорошо картированным эпитопом в OspA является LA2-эпитоп, расположенный в C-концевой области, который распознается нейтрализующим моноклональным антителом LA2. Как ожидалось, содержащие OspA CLP с расщепленным ядром, в которых OspA слит с coreN (следовательно, выставлен наружу С-конец OspA) индуцировали сильный LA2-эквивалентный ответ, в то время как CLP, содержащие OspA, слитый с coreC, индуцировали лишь слабый LA2-эквивалентный ответ. Вместо этого усиливался ответ к другим областям OspA.
Ожидалось, что в случае использования OspA в качестве чужеродного белка слияние с coreN должно обеспечивать хороший доступ к LA2-эпитопу в C-концевой области OspA, в то время как слияние с coreC должно обеспечивать хороший доступ к N-концевой области OspA, которая до настоящего времени недостаточно полно охарактеризована с иммунологической точки зрения. Представленные выше данные убедительно подтверждают эту концепцию (очень высокие титры антител к полному OspA, но лишь незначительные титры эквивалентных LA2 антител в случае OspA, слитого с coreC).
Таким образом, система с расщепленным ядром обладает высокой активностью в отношении усиления иммунного ответа как в случае слияния чужеродного белка с coreN, так и в случае слияния чужеродного белка с coreC.
Тип/область специфичности индуцируемых антител можно контролировать путем выбора сайта связывания (coreN или coreC). Для достижения оптимальных результатов вакцинации можно применять смеси вакцин на основе расщепленного ядра, в которых одна часть гетерологичной чужеродной аминокислотной последовательности слита с фрагментом coreN и другая часть слита с фрагментом coreC, что обеспечивает оптимальную презентацию различных эпитопов.
Пример 11
Слияния, представляющие интерес с точки зрения их применения в других областях помимо применения в вакцинах
Помимо прямого применения в качестве усиливающих иммунный ответ состоящих из частиц носителей для антигенных вакцин, которые содержат пептид и в данном случае прежде всего белок, можно рассматривать многочисленные другие применения такой платформы-носителя. В каждом случае большое количество (180 или 240) презентуемых молекул располагают симметрично в тесной близости друг к другу. Если указанные молекулы представляют собой способные к взаимодействию чужеродные молекулы (вакцины представляют собой специальный случай: взаимодействие с антителами), то частица, содержащая множество копий чужеродной молекулы, обладает существенно более высокой авидностью по сравнению с мономерной чужеродной молекулой (ср. пентамеры существующих в естественных условиях IgM; а также тетрамеры МНС, применяемые в иммунодиагностике). Для того чтобы иметь возможность вступать в реакцию со своими партнерами по взаимодействию, такие способные к взаимодействию чужеродные молекулы должны быть доступными на поверхности CLP. Система на основе расщепленного ядра существенно облегчает эту задачу по сравнению с обычной непрерывной системой.
Целый ряд служащих в качестве моделей чужеродных последовательностей экспрессировали с использованием системе на основе расщепленного ядра, при этом при использовании всех протестированных вставок происходило образование CLP. Для некоторых из них осуществляли непосредственную оценку способности к взаимодействию.
Схема на основе расщепленного ядра позволяет сделать доступными для дальнейшей дериватизации новые концы на поверхности CLP. На фиг.5 изображен димер с расщепленным ядром, в котором ACID-пептид слит с coreN, указанный пептид может взаимодействовать с комплементарным BASE-пептидом. Если молекула Х слита с BASE-пептидом, то Х связывается с ACID-CLP. Благодаря схеме на основе расщепленного ядра ACID приобретает гибкость; ACID, встроенный в систему с непрерывным ядром, вступает во взаимодействие с BASE-пептидом, однако, поскольку ACID- и BASE-пептиды образуют жесткую скрученную двойную спираль (O'Shea и др., 1993), то структура ядерного носителя становится нестабильной и CLP распадаются. В отличие от этого при использовании схемы на основе расщепленного ядра CLP должны сохранять стабильность. Такое сочетание можно расширить: вместо ACID-пептида можно сливать с coreN или coreC другие способные осуществлять взаимодействие молекулы. Затем можно подвергать взаимодействию соответствующие CLP с В, специфическим партнером А по взаимодействию. Такими парами партнеров по взаимодействию являются His6-Ni-NTa, являющийся акцептором биотина пептид - стрептавидин; Z33 - иммуноглобулин.
Ниже описаны более подробно другие возможные сочетания:
а) His6-метка:
Сливали с coreN через Gly2-линкер; получали частицы, которые связывались с Ni2 +ТА-агарозой. В данном случае His-метка являлась легко доступной в отличие от непрерывной системы. Использование сопоставимой вставки (His7) в эпитоп с/е1 в системе с непрерывным ядром не приводило к получению белка в значительных количествах, поскольку в этом случае остаток His6-метки не являлся стерически доступным.
б) Пептид, являющийся акцептором биотина (ВА):
Акцептор биотина (ВА23) представляет собой искусственный состоящий из 13 ак пептид (GLNDIFEAQKIEWH), который биотинилировали с помощью биотин-лигазы BirA из Е.coli. Для эффективного биотинилирования требуется свободный доступ, который обеспечивается в системе на основе расщепленного ядра, но не обеспечивается или обеспечивается лишь в незначительной степени в системе на основе непрерывного ядра. Слитый белок расщепленное ядро-ВА образовывал CLP и биотинилировался в Е.coli. С презентованным на CLP биотином могут связываться авидин/стрептавидин или их конъюгаты.
в) Домен Z33 белка А
Белок А из S. aureus связывается с иммуноглобулинами с высокой аффинностью («белок А-сефароза» для иммунопреципитации). Белок А состоит из 5 структурно сходных Ig-связывающих доменов. Z33 представляет собой один из таких доменов с модифицированной аминокислотной последовательностью; общая длина: 33 ак). Z33 все еще обладает высокой аффинностью к Ig (Kd 40 нМ). Для связывания с различными Ig требуется структурная гибкость и, кроме того, доступность. Последовательность Z33 сливали с coreN с использованием длинной линкерной последовательности: GGGGSGGGVEDGGGGSGGGGT-FNMQQQRRFYEALHDPNLNEEQRNAKIKSIREDP.
При использовании системы на основе расщепленного ядра происходило образование CLP. После добавления меченного с помощью ФИТЦ иммуноглобулина и повторного осаждения можно было обнаружить часть флуоресценции в типичных для частиц градиентных фракциях. Это не имело места в том случае, когда вместо Z33-слияния применяли CLP с ядерным wt-белком.
г) Вставка, представляющая собой ACID-пептид
ACID-пептид, встроенный в систему на основе непрерывного ядра, взаимодействует с комплементарным BASE-пептидом («пептид Velcro», т.е. пептид типа «застежка-липучка») посредством формирования пептидов в виде скрученной спирали (O'Shea и др., 1993), при этом происходят очень большие изменения ядерной структуры вплоть до ее расщепления; это обусловлено тем, что ACID-пептид сам по себе принимает гибкую структуру, которая не вызывает никакого «стресса» в ядерной структуре. Добавление BASE-пептида приводит к взаимодействию с последовательностью ACID, в результате чего обе пептидные последовательности принимают жесткую конформацию в виде скрученной спирали. Напряжение возникает вследствие наличия связей на обеих сторонах. ACID-пептид, фиксированный только на одной стороне, должен обладать способностью взаимодействовать без напряжения с добавленным BASE-пептидом или BASE-пептидом, слитым с другими партнерами. В любом случае состоящая из частиц структура слитого ACID-белка в обычной системе с непрерывным ядром остается интактной после расщепления TEV-протеазой связи С-конца с ядерным носителем.
Расщепленные ядра с «выставленной» последовательностью ACID можно загружать чужеродными белками, с которыми связывают комплементарный BASE-пептид. Это является одной из нескольких возможностей осуществления последующей загрузки CLP с расщепленным ядром выбранными чужеродными молекулами.
Подробно описанные выше механизмы сочетания позволяют создавать CLP, несущие один из пригодных для сочетания компонентов. К указанным CLP легко можно присоединять белок, слитый с другим пригодным для сочетания компонентом. Таким путем можно, например, презентовать иммунной системе структуры, не сходные с пептидными.
д) Одновременное слияние различных чужеродных молекул с coreN и coreC
В предыдущих примерах в каждом случае одну чужеродную последовательность сливали либо с coreN, либо с coreC. В принципе можно также сливать две различные чужеродные молекулы одновременно и с coreN и coreC; примером могут служить две субъединицы гетеродимерного чужеродного белка. В качестве модели применяли состоящий из двух частей GFP, первый сегмент которого, содержащий первые 10 из 11 β-нитей GFP, сливали с coreN, а второй сегмент, содержащий 11-ю β-нить GFP, сливали с coreC. В результате коэкспрессии с использованием вектора типа 1 получали зеленые флуоресцентные CLP. Таким образом, происходила сборка доменов coreN и coreC с образованием способной к сборке структуры CLP и две части GFP дополняли друг друга, в результате чего получалась правильная 3D-структура с образованием GFP-хромофора.
Этот пример демонстрирует, что можно осуществлять одновременное слияние различных чужеродных последовательностей с coreN и coreC с образованием функциональных структур как носителя с расщепленным ядром, так и двух различных чужеродных последовательностей, слитых с coreN и coreC.
Пример 12
CLP с расщепленным ядром, презентующие GB1-домен белка G на своей поверхности
В другом конкретном примере осуществляли встраивание так называемого GB1-домена иммуноглобулинсвязывающего белка, «белка G», из Streptococcus spp., который является функционально родственным белку А, в систему на основе расщепленного ядра HBV. Белок G также состоит из нескольких сходных доменов. Подобно белку А белок G связывается с иммуноглобулинами, но обладает различной специфичностью в отношении видов-источников и подтипов IgG. Благодаря этой комплементарности белка G и белка А они оба поступают на рынок в качестве реагентов для иммунологических анализов, основанных на связывании с различными иммуноглобулинами. Подобно белку А белок G и выведенный из него GB1-домен также связываются с фрагментом константной области (Fc-фрагментом) антитела, при этом вариабельная, распознающая антиген область антитела остается доступной для связывания с антигеном. Таким образом, существует возможность связывания с несколькими различными антителами.
В данном примере GB1-домен встраивали в систему на основе расщепленного ядра, при этом было продемонстрировано, что (I) происходит образование CLP и (II) указанные CLP (но не контрольные CLP, которые не несут GB1) обладают способностью связываться с различными антителами.
Аналогично тому, как это осуществляли в описанных выше примерах, GB1-домен генетически сливали с coreN или с coreC и коэкспрессировали с соответствующими не несущими указанны домен coreC- или coreN-фрагментами.
Специфическая аминокислотная последовательность соответствует ак с 229 по 284 белка-предшественника белка G (регистрационный номер в Swiss Prot; Р06654); SD-структура является известной (регистрационный номер в PDB: 1PGA; ссылка: Gallagher Т., Alexander P., Bryan P., Gililand G.L., Two crystal structures of the В 1 immunoglobulin-binding domain of streptococcal protein G and comparison with NMR Biochemistry 33, 1994, cc.4721-4729).
coreDJS-GGGGSGGGGTQ-YKLILNGKTLKGETTTEAVDAATAEKVFKQYANDNGVDGEWTYDDATKTFTVTEdP
Связывание с coreN осуществляли с помощью линкера G4SG4TQ; предпоследнюю ак, а именно K, заменяли на D (строчная буква в последовательности); предыдущий остаток Е представляет собой последнюю ак, видимую на рентгенограмме; замена K>D позволяли встраивать на нуклеотидном уровне сайт рестрикции BamHI (Gag gat cct TAG); таким образом, он находился в положении, гомологичном положению в описанных выше конструкциях.
Как и в других примерах, происходило эффективное образование частиц, что обнаруживали с помощью седиментации в градиенте сахарозы, анализа миграционных характеристик в нативном агарозном геле и электронной микроскопии.
Для выявления способности GB1-доменов, присутствующих в CLP, вступать во взаимодействие осуществляли инкубацию частиц с иммуноглобулинами и затем повторную седиментацию в градиенте сахарозы; при этом наблюдалась выраженная седиментация антител с частицами. Кроме того, связывание с IgG подтверждалось также тем фактом, что coreN-GB1-фрагмент непосредственно вступал в реакцию с конъюгатами вторичного антитела после ДСН-гель-электрофореза по данным анализа методом Вестерн-блоттинга. Следовательно, модифицированный GB1 обладал способностью, даже в контексте слияния с coreN, эффективно ренатурироваться и связываться со своим лигандом IgG.
Для доказательства того, что связанный с частицей GB1 действительно связывается с иммуноглобулином, аликвоту соответствующей фракции градиента сахарозы инкубировали с 5нМ меченным с помощью золота антителом, несвязанное антитело удаляли путем повторной седиментации в градиенте сахарозы и содержащую частицы фракцию анализировали с помощью электронной микроскопии. Присутствие обладающих электронной плотностью комплексов золото-частицы свидетельствует о связывании антитела с интактными CLP.
Пример 13
Содержащие GFP и варианты GFP CLP с расщепленным ядром, которые обладают собственной флуоресценцией и презентуют обладающие способностью к взаимодействию чужеродные молекулы
В примере 11 д) проиллюстрированы слияния, имеющие важное значение для применений помимо применения в вакцинах, где осуществляли ассоциацию двух частей молекулы GFP (GFP β1-10 и GFP β11) с образованием нативной GFP-структуры (зеленая флуоресценция) и формированием CLP, в которых GFPβ1-10 слита с coreN и GFPβ11 слита с coreC.
В настоящем примере дополнительные чужеродные последовательности сливали с GFPβ11, а именно с GB1-доменом белка G или доменами preS поверхностных белков HBV человека, HBV утки и HBV цапли. Это оказалось возможным только потому, что система с расщепленным ядром приводит к образованию нового N-конца на GFPβ11, который, кроме того, вследствие специфической ориентации на CLP с расщепленным ядром выходит наружу из поверхности CLP. Конструирование соответствующих экспрессионных плазмид осуществляли аналогично описанным выше методам, на основе векторов с расщепленным ядром, кодирующих по отдельности фрагменты coreN и согеС, в этом случае уже GFPβ1-10 была слита с coreN и GFPβ11 с coreC.
На основе полученных ранее данных следовало ожидать, что N-конец GFPβ11-сегмента должен выступать наружу с поверхности CLP и, следовательно, присутствие другого чужеродного домена, в принципе, не должно влиять ни на способность coreN- и coreC-фрагментов образовывать структуру в виде частицы, ни на способность GFPβ1-10- и GFPβ11-фрагментов образовывать нативную структуру GFP. Результаты, полученные в этом примере, схематично представлены на фиг.7. На панелях А-Г этого чертежа представлено следующее.
А. Основная структура обеих частей слитого белка. GFPβ1-10 слита с coreN, GFPβ11 слита с coreC. Поскольку GFPβ11 несет новый N-конец, то с ним можно сливать другой домен, в данном случае GB1. Б. Обладающие собственной флуоресценцией CLP получали, как уже продемонстрировано ранее, путем взаимодействия coreN с coreC (образование CLP) и GFPβ1-10 с GFPβ11 (образование GFP-хромофора). Однако, кроме того, они презентуют GB1 в доступной форме на своей поверхности. В. Различные антитела могут связываться посредством своего Fc-фрагмента с GB1-доменами. Г. В свою очередь, антитела могут взаимодействовать посредством своих различных вариабельных областей со специфическим для них антигеном. Таким образом, CLP с расщепленным ядром, несущие GFP-GB1, могут заменять обычные флуоресцентно меченные вторичные антитела при проведении непрямого иммуннофлуоресцентного анализа. На фиг.7 проиллюстрировано применение для флуоресцентного мечения тубулина в двух различных клетках. Аналогично этому можно рассматривать другие применения, основанные на связывании антител с антигеном.
Действительно, оказалось возможным осуществлять эффективную коэкспрессию других модифицированных фрагментов в Е.coli, где они образовывали CLP согласно данным, полученным методом седиментации в градиенте сахарозы и на основе анализа миграционных характеристик в нативном агарозном геле. Кроме того, такие CLP обладали типичной для GFP флуоресценцией и, следовательно, указанные фрагменты GFP связывались друг с другом правильным образом. Этот пример показывает, что GB1-фрагменты также имели правильную укладку и были доступными на поверхности CLP.
Способность обладающих собственной флуоресценцией CLP, презентующих GB1, связываться со связанными с антигеном антителами обнаруживали путем повышения проницаемости клеток линии HeLa (человеческих) или LMH (куриных), которые затем инкубировали с мышиным первичным антителом к тубулину. В одном случае клетки инкубировали общепринятым образом с меченным с помощью химического флуоресцентного агента (Су2) вторичным антителом (козье антимышиное антитело) или в альтернативном варианте с обладающими собственной флуоресценцией CLP с расщепленным ядром, презентующими GB1, и затем отмывали и, наконец, обследовали клетки с помощью флуоресцентной микроскопии.
В обоих случаях получали сопоставимые флуоресцентные изображения (как и ожидалось, происходило цитоплазматическое окрашивание, исключая ядра клеток). Таким образом, GB1-домены должны обладать способностью связываться с первичными антителами к тубулину, связанными с тубулином.
Таким образом, обладающие собственной флуоресценцией CLP с расщепленным ядром, презентующие GB1, можно применять в качестве альтернативы обычным конъюгатам, содержащим вторичное антитело с флуоресцентной меткой, для обнаружения связанного с антигеном первичного антитела.
Это доказывает, что GB1-домены все еще обладают способностью взаимодействовать с IgG и, следовательно, должны иметь правильную укладку. Кроме того, это доказывает, что обладающие собственной флуоресценцией CLP, презентующие GB 1, можно применять в качестве общего реагента для обнаружения (сопоставимого с флуоресцентно меченным вторичным антителом).
Аналогичные конструкции получали с использованием вариантов GFP, обладающих флуоресценцией другого цвета, причем в этом случае в имеющуюся конструкцию GFP вводили путем направленного мутагенеза известные мутации, которые изменяли спектры поглощения и испускания. Конкретно, создавали варианты желтого флуоресцентного белка (YFP) и цианового флуоресцентного белка (CFP). Оба варианта образовывали CLP, которые в каждом случае обладали типичными спектрами поглощения. Таким образом, можно создавать несущие GFP CLP с расщепленным ядром, презентующие GB1, которые обладают различными хромофорными свойствами и которые пригодны для применения в иммунологии в тех случаях, когда требуется мультифлуоресценция.
Система с расщепленным ядром позволяет создавать обладающие собственной флуоресценцией CLP, которые презентуют на своей поверхности обладающую способностью к взаимодействию другую чужеродную молекулу. Если эта чужеродная молекула представляет собой GB1, то она все еще обладает способностью специфически взаимодействовать с иммуноглобулинами, даже если они связаны со своим антигеном-мишенью. В результате этого появляется новое возможное применение помимо применения в вакцине, а именно, основанное на флуоресценции непосредственное обнаружение различных антигенов с использованием обладающих собственной флуоресценцией CLP. Такие обладающие собственной флуоресценцией способные к взаимодействию CLP с расщепленным ядром особенно пригодны для диагностических и аналитических применений.
Обладающие собственной флуоресценцией CLP с расщепленным ядром, которые презентуют чужеродные домены, отличные от GB1 белка G.
Представленные выше результаты показывают, что на CLP с расщепленным ядром, которые содержат состоящий из двух частей GFP, можно презентовать другой чужеродный домен, а именно GB1, в функциональной форме, т.е. способный взаимодействовать со специфическим лигандом, таким как IgG. Однако, GB1, состоящий менее чем из 60 ак, является сравнительно небольшим доменом. Для проверки общего подхода создавали гомологичные конструкции, в которых GB1 заменяли на preS-домены больших оболочечных белков HBV человека (HBV), утки (DHBV) и цапли (HHBV). Эти домены опосредуют специфическое связывание с соответствующими вирусами в их естественных клетках-мишенях, т.е. гепатоцитах, соответствующих видов-хозяев. Поскольку HBV состоит из 108 ак, а DHBV и HHBV примерно из 160 ак, то preS-домены существенно превышают GB1, и, кроме того, их последовательности отличаются от GB1, а также друг от друга.
Все три домена встраивали в слитые белки, содержащие расщепленное ядро-GFP, аналогично методу, который применяли для GB1 (см. фиг.7; замена GB1 на соответствующие preS-домены). Все три эффективно образовывали CLP, что обнаруживали с помощью седиментации в градиенте сахарозы, анализа миграционных характеристик и электрофореза в нативном агарозном геле и с помощью электронной микроскопии; CLP обладали типичной для GFP флуоресценцией (т.е. происходила ассоциация GFP-фрагментов с образованием правильной 3D-структуры GFP); кроме того, содержащие preS HBV и preS DHBV CLP вступали в реакцию с моноклональными антителами к preS HBV и preS DHBV; соответствующее антитело к preS вируса цапли в настоящее время не имеется в распоряжении.
Еще одним вариантом применения является идентификация клеточных рецепторов для конкретных вирусов. Можно предположить, что благодаря большому количеству (240) preS-доменов на частицу, CLP должны обладать очень большой авидностью в отношении указанных рецепторов, обеспечивая тем самым стабильное связывание с ними и также их идентификацию. Несущие рецептор клетки можно идентифицировать благодаря обусловливаемой GFP флуоресценции CLP с помощью флуоресцентной микроскопии или FACS; солюбилизированные рецепторные молекулы из таких клеток можно инкубировать с CLP и затем после седиментации в градиентах сахарозы они должны концентрироваться в типичных для частиц градиентных фракциях.
Пример 14
Несущие OspA CLP с расщепленным ядром, в которых OspA слит с coreN, индуцируют более высокие титры защитных антител к OspA, чем LipOspA, несущие OspA CLP с расщепленным ядром, в которых OspA слит с coreC, индуцируют меньшие титры
Как проиллюстрировано на фиг.5, на ориентацию презентуемого чужеродного белка можно направленно воздействовать путем сцепления либо с coreN, либо с coreC. В случае OspA это приводит к наиболее выгодной экспозиции либо C-концевого эпитопа моноклонального антитела LA-2, которое, как известно, является нейтрализующим (слияние с coreN), либо N-концевой области OspA, которая не содержит никаких известных эпитопов нейтрализующих антител (слияние с coreC). Как продемонстрировано на фиг.4, обе несущие OspA CLP с расщепленным ядром (в данном случае: группа 2 соответствует слиянию с coreN и группа 6 - слиянию с coreC) индуцировали титры общих антител к OspA, но пропорция антител, эквивалентных LA-2, была существенно выше в случае слияния с coreN, как это и следовало ожидать.
Однако представленные в данном случае данные представляют собой данные, полученные in vitro. Для того чтобы оценить непосредственно защитный потенциал индуцируемых антител in vivo, изучали способность конкретных вариантов иммунной сыворотки нейтрализовать искусственно вызванную инфекцию В. burgdorferi с использованием принятой для этой цели модели заболевания на мышах (см. Nassal и др., European. J. Immunol. 2005, cc.655-665). Мышам линии SCID вводили i.p. соответствующие варианты иммунной сыворотки, после чего через 2 ч осуществляли контрольное заражение путем введения s.c. в каждом случае по 103 спирохет на мышь. У невакцинированных мышей это, как правило, приводит к инфекции, которая проявляется в виде характерных для артрита симптомов, которые можно оценивать полуколичественно в левом и правом бедренно-предплюсневом суставах. Группа из 3 мышей, которой вводили только сыворотку здоровой мыши, служила в качестве отрицательного контроля. Группы (в каждом случае по 6 мышей), которым вводили либо очищенное моноклональное антитело LA2, которое, как известно, является нейтрализующим, либо индуцированные LipOspA антитела, служили в качестве положительного контроля. Группам подопытных животных (тест-группы, также включающие по 6 животных каждая) вводили иммунную сыворотку, которую получали с использованием OspA в системе с расщепленным ядром, где OspA слит с coreN, или OspA в системе с расщепленным ядром, где OspA слит с coreC. Для оценки количественных различий осуществляли пассивную иммунизацию сначала с использованием 5 мкг, а затем 1 мкг LA2, или взятой в объемах, эквивалентных указанным объемам LA2, иммунной сывороткой, полученной от животных, вакцинированных LipOspA или OspA в системе с расщепленным ядром, где OspA слит с coreN. Результаты представлены на фиг.8.
Вследствие низкой пропорции антител, эквивалентных LA2, в иммунной сыворотке, индуцируемых при использовании OspA в системе с расщепленным ядром, где OspA слит с coreC, ее применяли в объемах, эквивалентных тем, которые использовали в случае OspA в системе с расщепленным ядром, где OspA слит с coreN, безотносительно к титру антител, эквивалентных LA2. Оценку артрита в баллах проводили в дни 13, 17, 21, 28, 35, 45 и 52. Результаты, полученные в день 52, обобщены на фиг.8.
Как и ожидалось, у всех животных, которым вводили сыворотку здоровых мышей, развивался артрит. Уровень защиты, обусловленной моноклональным антителом LA-2, снижался от 4/6 до 1/6 животных при уменьшении дозы от 5 до 1 мкг, использование иммунной сыворотки LipOspA обеспечивало защиту 5/6 животных. Пассивная иммунизация иммунной сывороткой, полученной при использовании OspA в системе с расщепленным ядром, где OspA слит с coreN, обеспечивала защиту всех (6/6) животных от инфекции, даже при введении низкой дозы. OspA, слитый с coreC, обладал заметно более низкой защитной активностью (защищено 4/6 животных при введении высокой дозы и 1/6 животных при введении низкой дозы).
CLP с расщепленным ядром, несущие OspA, где OspA слит с coreN, вызывали высокие титры антител, эквивалентных LA2, которые обладали защитным действием in vivo. Защитный потенциал превосходил потенциал антител, индуцированных LipOspA. CLP с расщепленным ядром, несущие OspA, где OspA слит с согеС индуцировали высокие общие титры антител к OspA и лишь небольшая их пропорция представляла собой антитела, эквивалентные LA-2, и они обладали лишь небольшим нейтрализующим потенциалом. Таким образом, при использовании слияния чужеродного антигена с coreN индуцировался иммунный ответ, который количественно отличался от ответа, вызываемого слиянием с coreC.
Пример 15
Вокругспорозоитный белок (CSP) возбудителя малярии Plasmodium falciparum
Полноразмерный CSP в системе с расщепленным ядром индуцировала более сильный и широкий иммунный ответ, чем иммунодоминантная последовательность CSP-повтора в обычной системе с непрерывным ядром.
Ранее было установлено, что полный CSP, состоящий из 319 ак, может экспрессироваться только в системе с расщепленным ядром с образованием CLP; это было невозможно осуществлять в обычной непрерывной системе. Хотя укороченную версию без богатого цистеином C-концевого домена (ниже обозначена как CSPshort) оказалось возможным получать также и в обычной непрерывной системе, однако белок имел выраженную тенденцию к осаждению через несколько дней, этого не происходило в системе с расщепленным ядром, что свидетельствует о ее более высоких химических свойствах касательно белка.
Характерной особенностью CSP является присутствие множественного повтора тетрапептидных мотивов NANP и NVDP (24 мотива NANP и 3 мотива NVDP в полноразмерной версии, применяемой согласно настоящему изобретению); известно, что эти повторы являются иммуногенами, и ранее была описана экспериментальная вакцина на основе системы с непрерывным ядром. Поэтому для сравнительного исследования иммуногенности была создана соответствующая обычная конструкция, в которой последовательность повтора NANPNVDP(NANP)3NVDP была встроена между ядерными ак D78 и S81 и которая образовывала CLP, как это и ожидалось, поскольку вставка имела небольшой размер. Указанную конструкцию применяли в исследовании иммуногенности для сравнения с полноразмерным CSP и укороченным на С-конце CSPshort. Мышей иммунизировали в каждом случае одинаковыми количествами соответствующих CLP (20 мкг/мышь); количества продуцируемых антител определяли с помощью ELISA через 2, 4, 6 и 8 недель; затем проводили повторную иммунизацию (аналогичную первой) и снова определяли титры через 2, 4 и 6 недель после повторной иммунизации. В качестве тест-антигенов для оценки B-клеточного ответа на носитель использовали рекомбинантный CSP (в виде слитого белка из Е.coli), NANP- и NVDP-пептиды и рекомбинантный HBcAg. Антигены иммобилизовывали на планшетах для ELISA и затем анализировали с использованием 2-кратных серийных разведении иммунных сывороток. Наибольшее возможное разведение, которое вызывало сигнал, превышающий фоновый, указано на фиг.9.
Данные результаты позволяют сделать следующие выводы.
а) B-клеточный ответ на рекомбинантный CSP
Все три иммуногена вызывали очень сильный ответ на CSP; однако ответ на полноразмерный CSP был еще в 4 раза более сильным (титр: 1:12×106 по сравнению с 1:3×105 для повтора и укороченного CSP соответственно), это означает, что после разбавления иммунной сыворотки в 12 миллионов раз (!) она еще обладала способностью распознавать белок CSP.
б) B-клеточные ответы на присутствующие в повторе пептиды NANP и NVDP
Ответы на NANP-пептид, как и следовало ожидать, были сходными для всех конструкций. Ответы на NVDP-пептид, который присутствует в повторе в меньшем количестве, были сходными, но выражено более высокими для полноразмерной CSP-конструкции даже после первой иммунизации (без повторной иммунизации) (титр 1:2,5×104 по сравнению с 1:5×103).
в) B-клеточные ответы на CSP-последовательности, не присутствующие в повторе
Только CSPshort и полноразмерный CSP индуцировали ответ на пептиды CSP12 (EEPSDKHIEQYLKKIQNSLS; положения 246-264 в полноразмерной конструкции с расщепленным ядром) и CSP8 (GNGIQVRIKPGSANKPKDELD; положения 274-294 в полноразмерной конструкции с расщепленным ядром). Следовало ожидать отсутствие соответствующих ответов после иммунизации с использованием входящих в повтор пептидов в системе с непрерывным ядром, поскольку последовательности CSP8 и CSP12 в нем не присутствуют. Важность результата заключается в том, что именно полноразмерный CSP можно включать только в систему с расщепленным ядром, с использованием системы с расщепленным ядром можно также презентовать другие эпитопы, которые могут вносить вклад в защитное действие вакцины.
г) B-клеточные ответы на ядерный носитель
Как CSPshort, так и полноразмерный CSP в системе с расщепленным ядром вызывали примерно в 25 раз более слабый ответ на ядерный носитель по сравнению с входящими в повтор пептидами в непрерывной системе (титр 1:1,25×105 сравнению с 3×106). Поскольку требуемый ответ на чужеродный белок являлся по меньшей мере таким же (CSPshort) или даже превышал его (полноразмерный CSP) при использовании входящих в повтор пептидов в системе с непрерывным ядром, то специфическая иммуногенность по отношению к встроенной чужеродной последовательности существенно повышалась при использовании укороченного и прежде всего полноразмерного CSP в системе с расщепленным ядром. Кроме того, сильный ответ на ядерный носитель затрудняет дифференциальную диагностику пациентов с положительной реакцией на антитела к HBsAg (выявление различия между инфекцией и вакцинацией с использованием антител к HBsAg); таким образом, благоприятным является низкий ответ на ядерный носитель.
д) Полноразмерный CSP в системе с расщепленным ядром активирует Т-клетки
Для выявления индукции Т-клеточного ответа осуществляли стимуляцию клеток селезенки мышей, специфически иммунизированных рекомбинантным HbcAg или соответственно известным Т-клеточным пептидом (НВс 120-140), или рекомбинантным CSP или соответственно CSP-пептидами NANP и CSP8, и затем оценивали производство IL-2. Все конструкции вызывали образование Т-клеток, которые активировались HBcAg, HBc120-140, рекомбинантным CSP и NANP. Однако стимулировать пептидом CSP8 производство IL-2 оказалось возможным только в клетках селезенки мышей, иммунизированных полноразмерным CSP в системе с расщепленным ядром.
Таким образом, и на Т-клеточном уровне конструкция полноразмерного CSP также вызывала более широкий ответ по сравнению с непрерывной конструкцией, содержащей только входящие в повтор пептиды.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЕКТОР ДЛЯ ПЕРЕНОСА И ВАКЦИНА ПРОТИВ ТУБЕРКУЛЕЗА | 2007 |
|
RU2453603C2 |
| ВАКЦИНЫ ПРОТИВ ВИРУСОВ ГЕПАТИТА | 1995 |
|
RU2189254C2 |
| БАКУЛОВИРУСНЫЙ ВЕКТОР И ЕГО ПРИМЕНЕНИЕ | 2007 |
|
RU2588464C2 |
| СЛИТЫЙ БЕЛОК | 2016 |
|
RU2748643C2 |
| ИММУНОГЕННЫЕ ГИБРИДНЫЕ ПОЛИПЕПТИДЫ ПРОТИВ ОЖИРЕНИЯ И КОМПОЗИЦИЯ ВАКЦИН ПРОТИВ ОЖИРЕНИЯ, СОДЕРЖАЩАЯ ЭТИ ПОЛИПЕПТИДЫ | 2007 |
|
RU2418005C2 |
| СЛИТЫЕ БЕЛКИ ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ ИММУНОГЕННЫХ УСИЛИВАЮЩИХ АГЕНТОВ ДЛЯ ИНДУЦИРОВАНИЯ АНТИГЕНСПЕЦИФИЧЕСКОГО Т-КЛЕТОЧНОГО ОТВЕТА | 2013 |
|
RU2619187C2 |
| РЕКОМБИНАНТНЫЙ ВИРУСНЫЙ ВЕКТОР И СИСТЕМА ЭКСПРЕССИИ В КЛЕТКАХ РАСТЕНИЯ ГИБРИДНОГО БЕЛКА, ВКЛЮЧАЮЩЕГО ВНЕКЛЕТОЧНЫЙ ДОМЕН БЕЛКА М2 ВИРУСА ГРИППА, ПРИСОЕДИНЕННЫЙ К ЯДЕРНОМУ АНТИГЕНУ ВИРУСА ГЕПАТИТА В | 2010 |
|
RU2460796C2 |
| БАКУЛОВИРУСНЫЕ ВЕКТОРЫ С ДВОЙНЫМ ПРОМОТОРОМ, ВКЛЮЧАЮЩИМ В СЕБЯ ПРОМОТОР ПОЗВОНОЧНОГО И ПРОМОТОР БАКУЛОВИРУСА, КОНТРОЛИРУЮЩИМ ИММУНОГЕННЫЙ СЛИТЫЙ ГЕН | 2008 |
|
RU2491093C2 |
| ВСПОМОГАТЕЛЬНЫЙ ПЛАЗМИДНЫЙ ЛЕНТИВИРУСНЫЙ ЭКСПРЕССИОННЫЙ ВЕКТОР ДЛЯ ПОЛУЧЕНИЯ ВЫСОКИХ ТИТРОВ VPX-СОДЕРЖАЩИХ ЛЕНТИВИРУСНЫХ ЧАСТИЦ, ОБЕСПЕЧИВАЮЩИЙ ЭФФЕКТИВНОЕ ЗАРАЖЕНИЕ МОНОЦИТОВ И ДЕНДРИТНЫХ КЛЕТОК ЧЕЛОВЕКА | 2017 |
|
RU2697781C2 |
| СРЕДСТВА И СПОСОБЫ ЛЕЧЕНИЯ ВИРУСА ГЕПАТИТА B | 2017 |
|
RU2740802C2 |







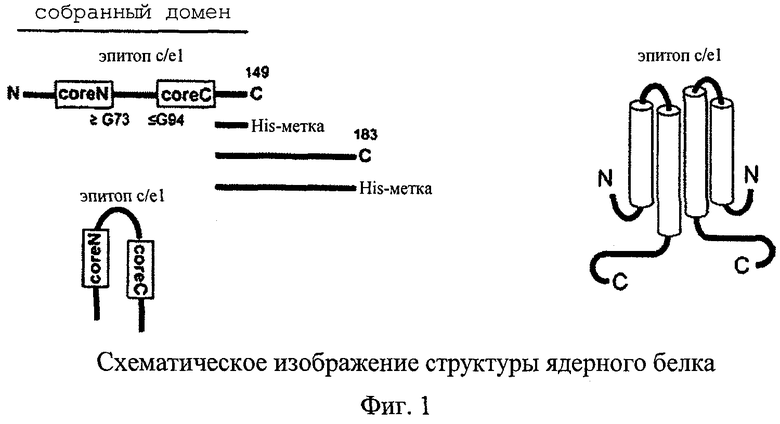
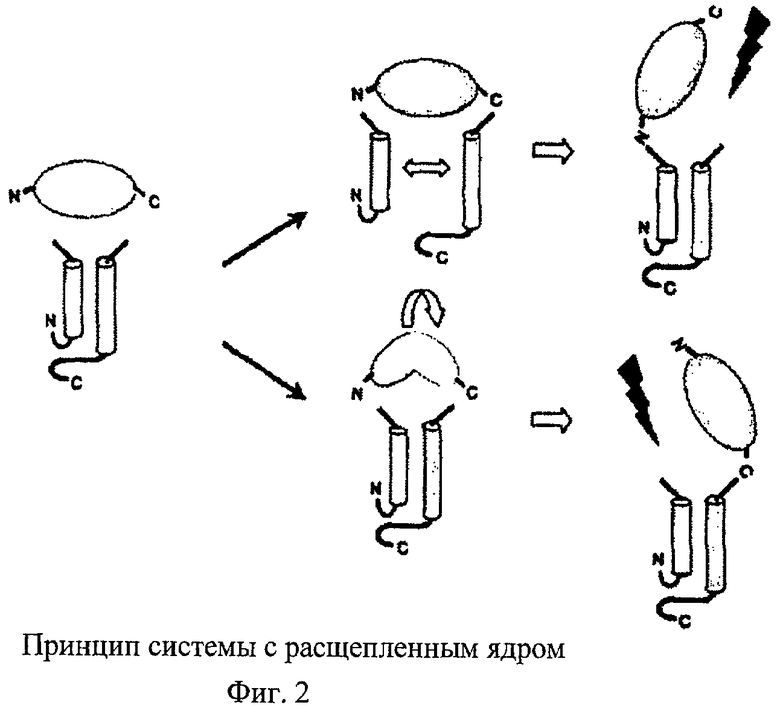
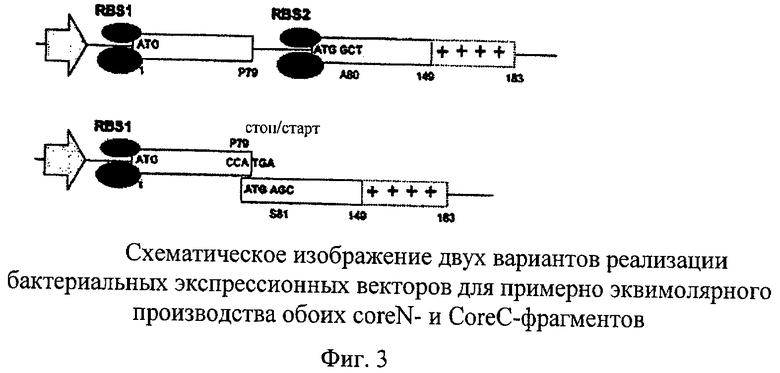
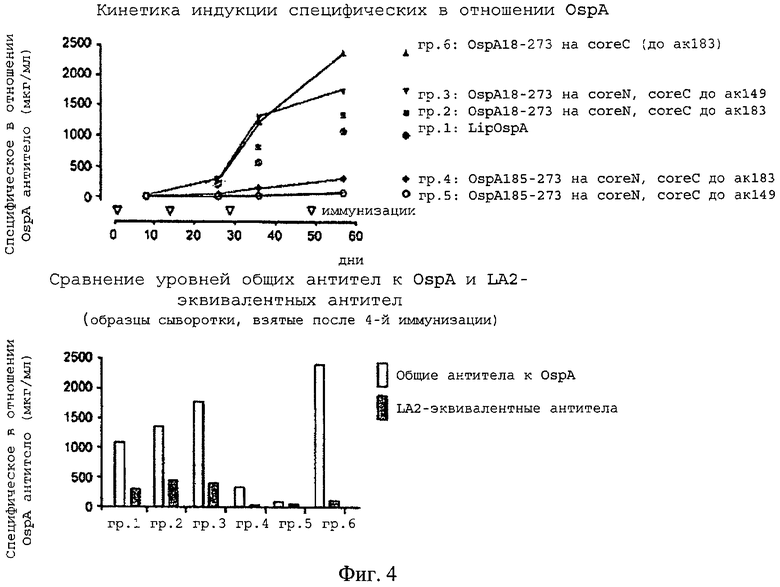
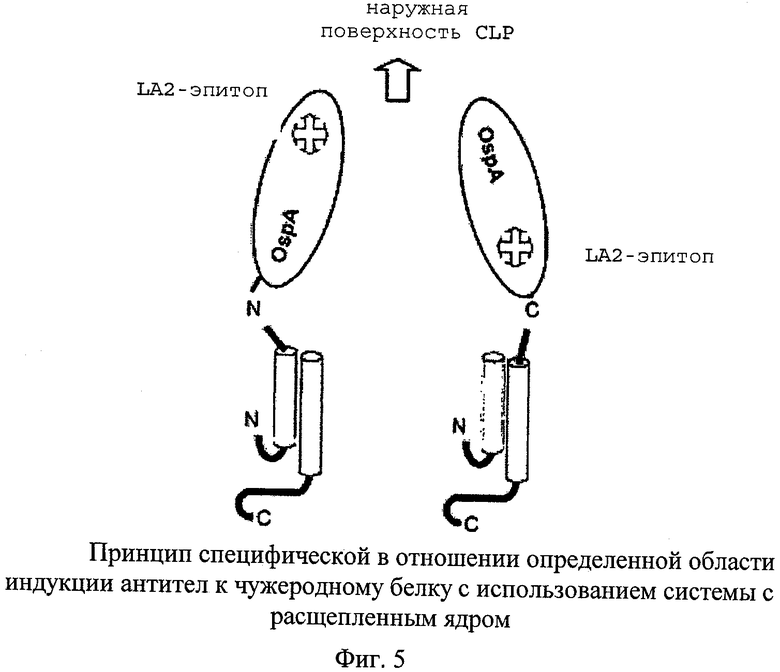
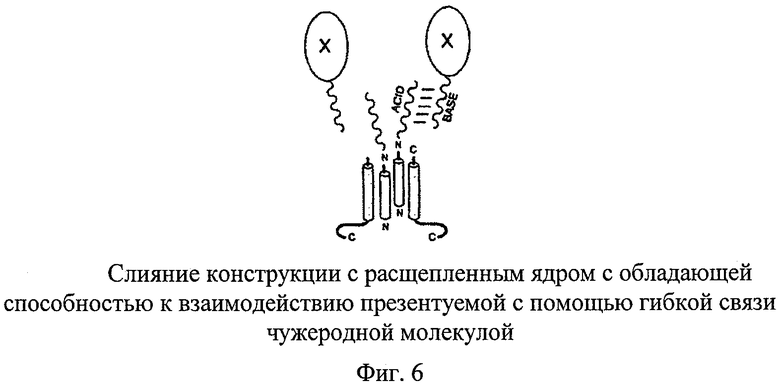
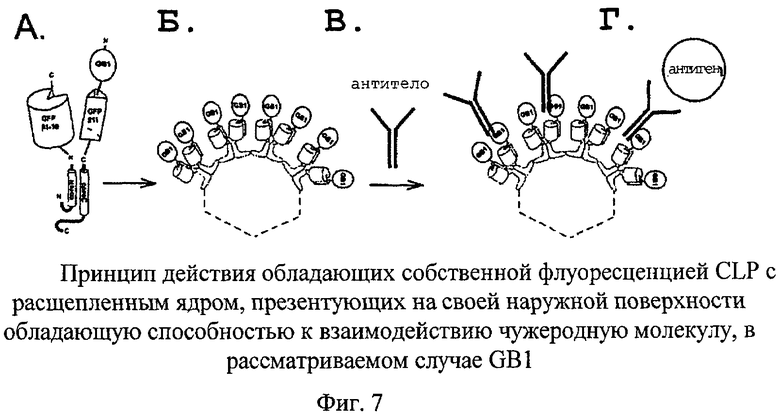
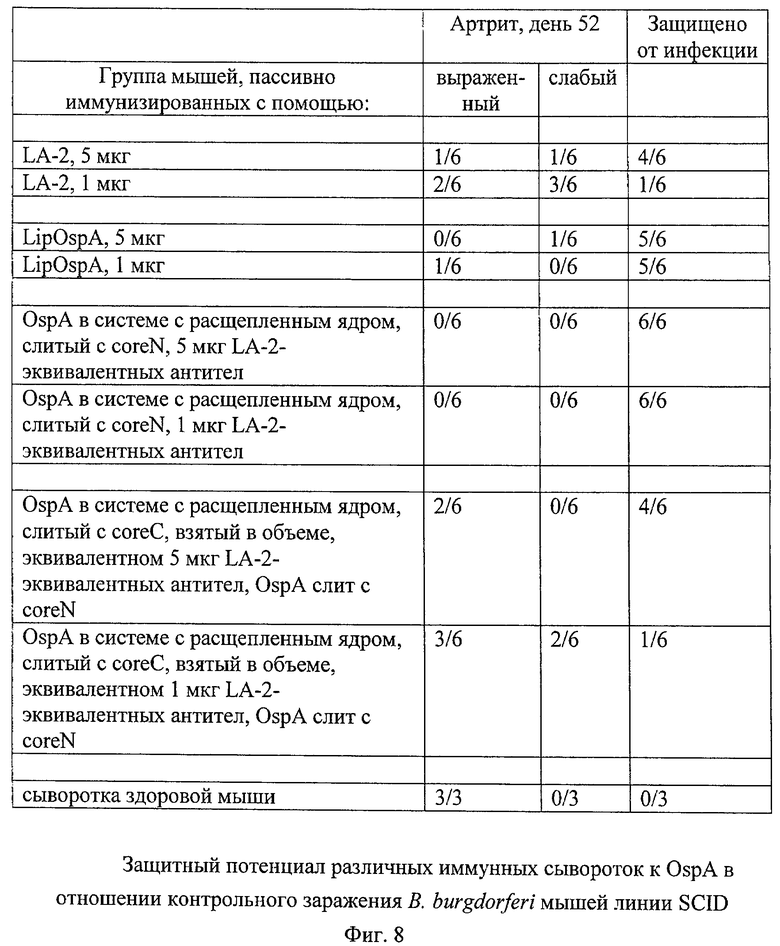
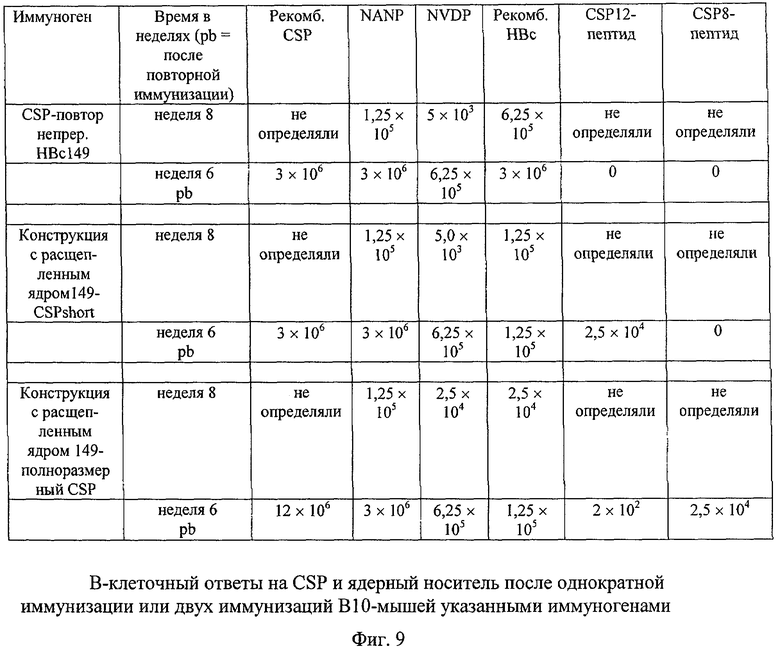
Изобретение относится к области биотехнологии, генной инженерии и вирусологии. Раскрыты субстанции, представляющие собой носитель с расщепленным ядром, которые содержат в виде отдельных полипептидов coreN-и соrеС-доменов ядерного белка вируса гепатита В и по меньшей мере одну чужеродную молекулу, к которой требуется вызвать иммунный ответ. Согласно изобретению чужеродную молекулу, прежде всего гетерологичную чужеродную аминокислотную последовательность, сливают либо с С-концом coreN-домена, либо с N-концом соrеС-домена, и ядерный белок обладает способностью образовывать капсидоподобные частицы. Также описаны соответствующий способ получения и диагностическое и лекарственное средство. Изобретение может быть использовано в медицине. 4 н. 12 з.п. ф-лы, 9 ил., 15 пр.
1. Система-носитель с расщепленным ядром, которая служит носителем чужеродных молекул на основе пептидов и белков, имеющих в виде отдельных полипептидов copeN- и соrеС-домены ядерного белка вируса гепатита В и по меньшей мере одну чужеродную молекулу, к которой требуется вызвать иммунный ответ, где чужеродная молекула слита с С-концом coreN-домена или с N-концом соrеС-домена и ядерный белок обладает способностью образовывать капсидоподобные частицы, и где чужеродная молекула слита с аминокислотой ядерного белка, находящейся между положениями 73 и 94 ядерного белка.
2. Система-носитель с расщепленным ядром по п.1, отличающаяся тем, что вирус гепатита В является специфическим для млекопитающих.
3. Система-носитель с расщепленным ядром по п.2, отличающаяся тем, что ядерный белок имеет последовательность вируса гепатита В человека.
4. Система-носитель с расщепленным ядром по п.2, отличающаяся тем, что ядерный белок имеет аминокислотную последовательность WHV.
5. Система-носитель с расщепленным ядром по п.1, отличающаяся тем, что чужеродная молекула имеет аминокислотную последовательность белка патогенной бактерии.
6. Система-носитель с расщепленным ядром по п.1, отличающаяся тем, что чужеродная молекула имеет аминокислотную последовательность белка патогенного для эукариотического организма, прежде всего возбудителя малярии Plasmodium falciparum.
7. Система-носитель с расщепленным ядром по п.1, отличающаяся тем, что чужеродная молекула имеет аминокислотную последовательность белка вируса.
8. Система-носитель с расщепленным ядром по п.1, отличающаяся тем, что чужеродная молекула имеет аминокислотную последовательность белка, обладающего собственным патогенным потенциалом, прежде всего белка-маркера опухоли.
9. Система-носитель с расщепленным ядром по п.1, отличающаяся тем, что чужеродная молекула выбрана из группы, включающей
а) последовательность пептида-акцептора биотина;
б) последовательность ACID-пептида;
в) последовательность Z33-домена, происходящего из белка А;
г) последовательность GB1-домена белка G.
10. Способ получения системы-носителя для чужеродных молекул, прежде всего чужеродных белков, к которым требуется индуцировать иммунный ответ, где система-носитель, во-первых, имеет белковую последовательность ядерного антигена вируса гепатита В и, во-вторых, несет чужеродную молекулу, прежде всего гетерологичную белковую последовательность,
при этом чужеродную молекулу, прежде всего гетерологичную белковую последовательность, сливают с coreN- или соrеС-доменом за положением 73 или перед положение 94 в ядерной области вируса гепатита В, и при этом полипептидную цепь носителя расщепляют либо после домена соrеN, либо перед доменом соrеС,
отличающийся тем, что либо домен coreN и чужеродная молекула, прежде всего гетерологичная белковая последовательность, либо домен соrеС и чужеродная молекула, прежде всего гетерологичная белковая последовательность, экспрессируются по отдельности в виде слитых белков,
либо встраивают последовательность, распознаваемую протеазой, между coreN и чужеродной молекулой, прежде всего гетерологичной белковой последовательностью, или между чужеродной молекулой, прежде всего гетерологичной белковой последовательностью, и соrеС, и после экспрессии, но до применения вакцину с расщепленным ядром расщепляют с помощью протеазы на два полипептида, и
получают капсидоподобные частицы в качестве системы-носителя для чужеродных молекул на основе пептидов или белков.
11. Способ по п.10, отличающийся тем, что гетерологичную белковую последовательность присоединяют путем слияния после аминокислоты 74 или перед аминокислотой 85 ядерного антигена вируса гепатита В.
12. Способ по одному из пп.10-11, отличающийся тем, что гетерологичная последовательность чужеродного белка состоит по меньшей мере из 40 аминокислот.
13. Способ по п.10, отличающийся тем, что гетерологичная последовательность чужеродного белка состоит по меньшей мере из 120 аминокислот.
14. Способ по п.10, отличающийся тем, что два отдельных полипептида экспрессируют с помощью бицистронного вектора, при этом первый полипептид содержит coreN-домен и второй полипептид содержит соrеС-домен, и гетерологичную белковую последовательность сливают либо с С-концом coreN-домена, либо с N-концом соrеС-домена.
15. Лекарственное средство, которое является вакциной, содержащее капсидоподобные частицы, отличающееся тем, что указанные частицы содержат систему-носитель с расщепленным ядром по одному из пп.1-8.
16. Диагностическое средство, содержащее капсидоподобные частицы, отличающееся тем, что указанные частицы содержат систему-носитель с расщепленным ядром по одному из пп.1-8.
| US 2003175863 A1, 18.09.2003 | |||
| VOGEL MAREN ET AL: "Quaternary structure is critical for protein display on capsid-like particles (CLPs): Efficient generation of hepatitis В virus CLPs presenting monomeric but not dimeric and tetrameric fluorescent proteins" | |||
| PROTEINS STRUCTURE FUNCTION AND BIOINFORMATICS, Bd | |||
| Способ окисления боковых цепей ароматических углеводородов и их производных в кислоты и альдегиды | 1921 |
|
SU58A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |

