Изобретение относится к биотехнологии, может быть использовано в тканевой инженерии и клеточной трансплантологии с целью получения тканевой структуры сетчатки для исследования клеточных и молекулярных механизмов регенерации сетчатки и тестирования лекарств in vitro, а также для восстановления сетчатки при различных нейродегенеративных заболеваниях.
На сегодняшний день возрастная макулярная дегенерация (ВМД) - одно из самых распространенных и малоизученных заболеваний глаз, угрожающих значительным снижением зрения, слепотой и инвалидностью. ВМД характеризуется постепенным снижением центрального зрения у лиц старше 50-60 лет, приводящего к значительной потере зрения в пожилом возрасте. Существует две основные формы ВМД - «сухая» и «влажная». В 9 из 10 случаев ВМД встречается «сухая» форма. Она, как правило, характеризуется медленным прогрессирующим снижением зрения. «Влажная» форма ВМД развивается примерно в 1 из 10 случаев и может вызвать серьезную потерю зрения в довольно короткий промежуток времени - иногда за несколько месяцев. Лечение ВМД (макулодистрофии) остается актуальной проблемой современной офтальмологии.
Известен способ лечения «влажной» формы возрастной макулярной дегенерации, включающий интравитреальное введение препарата Авастин [Э.В.Бойко, О.В.Филохина, С.В.Сосновский и др. Первый опыт применения препарата Авастин в лечении влажной формы возрастной макулярной дегенерации, Современные технологии лечения витреоретинальной патологии - 2007. Сборник научных статей по материалам научно-практической конференции (Москва, 23 марта 2007 г.) под редакцией Х.П.Тахчиди. Москва, 2007 г., С.54-58]. Препарат Авастин представляет собой рекомбинантное моноклональное антитело, которое селективно связывается с биологически активным фактором роста эндотелия сосудов (VEGF) и нейтрализует его, ингибируя взаимодействие VEGF с его рецепторами на поверхности эндотелиальных клеток, что приводит к снижению роста новообразованных сосудов. Недостатком способа является непродолжительность действия, что требует повторных инъекций препарата.
Известен способ лечения возрастной макулярной дегенерации сетчатки, по которому выполняют частичную витрэктомию премакулярного отдела стекловидного тела, индуцируют отслойку задней гиалоидной мембраны без ее удаления и в сформировавшееся ретрогиалоидное пространство вводят препарат Авастин [патент RU №2368359, МПК A61F 9/00].
Известен способ лечения возрастной макулярной дегенерации сетчатки [патент RU №2408335, МПК A61F 9/00]. Способ включает лазерное воздействие и введение ретиналамина, представляющего собой лиофилизат, полученный путем уксуснокислой экстракции из сетчатки глаза крупного рогатого скота или свиней, содержащий комплекс низкомолекулярных пептидов.
Однако все эти методы лечения направлены только на поддержание зрения и предотвращение его ухудшения.
Достижения клеточной биологии в конце XX века стали основой для разработки биомедицинских технологий. Они позволили использовать культивируемые клетки человека в медицине для восстановления структурной и функциональной активности поврежденных тканей в ситуациях, где традиционные методы лечения малоэффективны или бессильны.
Известен способ лечения «сухой» формы возрастной макулярной дегенерации сетчатки, по которому макулярную область сетчатки облучают низкоинтенсивным лазерным излучением и через 3 часа проводят внутривенное введение аутологичных культивированных мезенхимальных стволовых клеток костного мозга пациента [патент RU 2333737, МПК A61F 9/00].
Известен способ лечения «сухой» формы возрастной макулярной дегенерации сетчатки [патент RU 2375016, МПК A61F 9/00]. Способ заключается в интравитреальном введении взвеси аутологичных культивированных мезенхимальных стволовых клеток собственного костного мозга пациента.
Считается, что клетки предшественники, выделенные вне сетчатки и трансплантированные путем системного введения или инъекции субретинально или в полость стекловидного тела взрослого глаза, могут интегрироваться в сетчатку и тем самым улучшать ее функцию.
Однако недостатком известных способов является то, что полученные результаты в основном неоднозначны, маргинальны или негативны и указывают, что простые трансплантации взрослых не ретинальных предшественников в патологически измененную сетчатку не способны приводить к существенным долговременным клиническим улучшениям.
В качестве лечебной терапии в последнее десятилетие интенсивно исследуется идея трансплантирования здоровых клеток ретинального пигментного эпителия (РПЭ) [Sommer F. Hyalocytes in Tissue Engineering First Steps Towards a Cell-based Vitreous Substitute, Dissertation, 2006, p.14-15]. Попытки использовать слои клеток РПЭ без какого-нибудь поддерживающего матрикса связаны с некоторыми недостатками. Во-первых, более трудно управлять клетками в течение трансплантации [Lu L., Nyalakonda К., Kam L., Bizios R., Goepferich A., Mikos A.G. Retinal pigment epithelial cell adhesion on novel micropatterned surfaces fabricated from synthetic biodegradable polymers // Biomaterials. - 2001. - Vol.22. - P.291-297]. В дополнении неспецифическая ферментативная обработка (например, использование трипсина/ЭДТА) культивированных слоев РПЭ приводит к существенному снижению метаболизма ретиноидов [Recum Н.А., Okano Т., Kim S.W., Bernstein P.S. Maintenance of retinoid metabolism in human retinal pigment epithelium cell culture // Exp. Eye Res. - 1999. - Vol.69. - Р.97-107].
Одним из направлений биотехнологии, которое занимается созданием биологических заместителей тканей и органов, является тканевая инженерия. Тканевая инженерия - создание новых тканей и органов для терапевтической реконструкции поврежденного органа посредством доставки в нужную область опорных структур, клеток, молекулярных и механических сигналов для регенерации. На первом этапе создания тканеинженерной конструкции отбирают собственный или донорский клеточный материал (биопсия или аутопсия), выделяют тканеспецифичные клетки и культивируют их.
Одной из центральных задач по лечению патологически измененной сетчатки является идентификация оптимального типа клеток, чтобы активно управлять репаративной функцией сетчатки. Идеальным типом клеток должны быть клетки, полностью детерминированные к судьбе клеток сетчатки и еще сохраняющие способность к экспансии in vivo или in vitro. Кроме того, важна для интересующего типа клеток способность дифференцироваться в функциональную специализированную ткань. Так как эпителиальные клетки обычно не выживают в суспензии, клетки РПЭ должны прикрепляться к поверхности для того чтобы избежать апоптоз [Tezel Т.Н., Del Priore L.V. Reattachment to a substrate prevents apoptosis of human retinal pigment epithelium // Graefes. Arch. Clin. Exp. Ophthalmol. - 1997. - Vol.235. - Р.41-47].
Поэтому в состав тканеинженерной конструкции, или трансплантата, кроме культуры клеток входит специальный носитель (матрица). Матрицы могут быть выполнены из различных биосовместимых материалов. Клетки полученной культуры наносятся на матрицу, после чего такая трехмерная структура переносится в биореактор с питательной средой, где инкубируется в течение определенного времени. При формировании сложных пространственных структур необходим специальный матрикс, повторяющий форму восстанавливаемой ткани. Биоматериалы, используемые для получения матриц, должны быть биологически инертными и после перенесения в организм обеспечивать локализацию нанесенного на них клеточного материала в определенном месте. Для создания тканей и органов применяются синтетические материалы, материалы на основе природных полимеров (хитозан, альгинат, коллаген), а также биокомпозитные материалы. Одними из первых в тканевой инженерии стали применяться биодеградируемые синтетические биоматериалы на основе полимеров органических кислот, например молочной (PLA, полилактат) и гликолевой (PGA, полигликолид). Матрицы на основе органических кислот легли в основу создания таких органов и тканей, как кожа, кость, хрящ, сухожилие, мышцы (поперечно-полосатая, гладкая и сердечная), тонкая кишка и др.
Группа исследователей под руководством A.G.Mikos предложила использовать биодеградируемые полимерные пленки как временные субстраты для культивируемых клеток РПЭ и последующую трансплантацию этих полимер-клеточных комплексов в субретинальное пространство. Однако культивированные клетки РПЭ на этих пленках, сделанных из комбинации молочной и гликолевой кислоты (PLGA), теряли свою характерную кубическую морфологию в течение 7 дней культивирования [Thomson R.C., Giordano G. G., Collier J.H., Ishaug S.L., Mikos A.G., Lahiri-Munir D., Garcia C.A. Manufacture and characterization of poly(alpha-hydroxy ester) thin films as temporary substrates for retinal pigment epithelium cells // Biomaterials. - 1996. - Vol.17. - P.321-327].
Для того чтобы сохранить нормальную морфологию и функцию клеток РПЭ in vitro исследователи разработали новый деградируемый микросубстрат из PLGA и блокатор кополимеров из полиэтиленгликоля (PEG) и PLA, используя микроконтактную печать. Поверхности этой пленки, состоящие из адгезивного (PLGA) и неадгезивного (PEG/PLGA) доменов, позволяли клеткам прикрепляться и распространяться, и разрешали поддерживать дифференцированный клеточный фенотип в течение 8-часового периода исследования. Полимерный субстрат должен был облегчить обращение с клетками в течение трансплантации и обеспечить корректную ориентацию трансплантата в субретинальном пространстве [Lu L., Kam L., Hasenbein М., Nyalakonda К., Bizios R., Goepferich A., Young J.F., Mikos A.G. Retinal pigment epithelial cell function on substrates with chemically micropatterned surfaces // Biomaterials. - 1999. - Vol.20. - Р.2351-2361]. Хотя PLA и PLGA являются биодеградируемыми, у этих материалов имеются недостатки: изменение pH окружающих тканей при расщеплении в организме и недостаточная механическая прочность, что не позволяет использовать их как универсальный материал для матриц и подложек.
Известно использование амниотических мембран, которые модулируют пролиферацию и дифференцировку клеток РПЭ в культуре [Ohno-Matsui К., Ichinose S., Nakahama К., Yoshida Т., Kojima A., Mochizuki М., Morita I. The effects of amniotic membrane on retinal pigment epithelial cell differentiation // Mol. Vis. - 2005. - Vol.11. - P.1-10]. Амниотические мембраны при трансплантации действуют как соответствующий субстрат для эпителизации и получили широкое использование в офтальмологии для лечения персистирующих эпителиальных дефектов [Dua H.S. Amniotic membrane transplantation // Br. J. Ophthalmol. - 1999. - Vol.83. - Р.748-752]. В работе [Stanzel B.V., Espana E.M., Grueterich M., Kawakita T., Parel J.M., Tseng S.C., Binder S. Amniotic membrane maintains the phenotype of rabbit retinal pigment epithelial cells in culture // Exp. Eye Res. - 2005. - Vol.80. - P.103-112.] было показано, что лишенные эпителия амниотические мембраны повышают образование монослоя из РПЭ с плотными контактами. Их рекомендовали использовать как матрикс, содержащий базальную мембрану, для того чтобы облегчить трансплантацию РПЭ при лечении ВМД.
Изоляция клеток РПЭ с использованием ферментов (таких как коллагеназа 3 типа/гиалуронидаза) и последующее пассирование на термически чувствительной поверхности является подходящим методом для того, чтобы сохранить метаболическую активность культивированных клеток РПЭ для трансплантации [von Recum Н.А., Okano Т., Kim S.W., Bernstein P.S. Maintenance of retinoid metabolism in human retinal pigment epithelium cell culture // Exp. Eye Res. - 1999. - Vol.69. - Р.97-107].
Белки внеклеточного матрикса, в том числе коллагеновые структуры успешно используются в качестве подложки для культивирования. Однако создать искусственную объемную конструкцию из коллагена, повторяющую структуру любого органа, в том числе и сетчатки глаза, не представляется возможным.
Это обстоятельство инициировало исследования, направленные на разработку способа, позволяющего получать децеллюлированные матриксы органов с сохранением их пространственной организации. Технология, предусматривающая девитализацию ткани аллографта длительной мягкой обработкой растворами детергентов, приводит к разрушению всех клеточных элементов донора, оставляя неизменным соединительнотканный остов органа. Ацеллюлярность полученного каркаса обеспечивает его неиммуногенность, а остутствие поперечных сшивок не препятствует заселению его клетками реципиента. Кроме того, пространственные особенности подложки и химический состав матрикса служат дополнительными стимулами для их поддержания пролиферации, самоорганизации и выполнения дифференцировочных функций. Трехмерные модели (3D матрикс), представляющие собой тканеинженерные конструкции, на сегодняшний день являются наилучшими моделями для оценки поведения и дифференцировки предетерминированных клеток.
Известен способ получения соединительнотканного каркаса кровеносного сосуда. Способ позволяет эффективно отмывать клетки сосуда без повреждения коллагеновых и эластических волокон [Егорова М.В., Роговская Ю.В., Иванов А.В., Андреев С.Л., Ахмедов Ш.Д., Афанасьев С.А., Экономичная технология получения бесклеточной матрицы артериального сосуда животных и человека, Клеточные технологии в биологии и медицине, №2, 2011, с.110-113].
Известен метод экстракции клеточных компонентов из тканей тела и структур, которые должны быть использованы как материал для имплантации. Децеллюляризация улучшает биосовместимость и уменьшает иммунологическую реакцию против тканевого трансплантата [заявка GB №2375771, МПК A61L 27/26].
Известен способ совмещения культивированных остеогенных клеток и трехмерного материала-носителя путем иммобилизации клеток на поверхности материала-носителя (деминерализованный костный матрикс). Изобретение обеспечивает получение тканеинженерного эквивалента кости, включающего полноценное совмещение предифференцированных в условиях in vitro остеогенных клеток и трехмерного материала-носителя, изготовленного из деминерализованного костного матрикса [патент RU №2414916, МПК A61K 35/32].
Известна гелевая модель внеклеточного матрикса для изучения развития и дифференцировки эмбриональных СК (ЭСК) in vitro и метод для изучения механизмов развития ЭСК [заявка CN №101711890, МПК A61L 27/38]. В заявке описана трехмерная модель дифференцировки и развития ЭСК, внедрение ЭСК в эту модель в различной клеточной плотности, обнаружение феномена образования эмбриоидных телец.
Учитывая ключевую роль РПЭ в сохранности и жизнеспособности всей сетчатки, пересадка РПЭ может решить проблему ВМД.
Чтобы выяснить биологическую сложность органогенеза сетчатки и ее все увеличивающееся пересечение с регенеративной функцией сетчатки, а также для решения целесообразности внедрения в клиническую практику клеточных трансплантатов на основе клеток РПЭ, необходимы новые модельные системы, в которых могут быть проанализированы индивидуальные вариации типов клеток предшественников, в том числе культивированных клеток РПЭ, сигналы внеклеточного матрикса, двух- и трехмерные структуры и тканевая функция в отношении их эффектов на глобальную физиологическую функцию интактного органа. В частности необходимо разработать способ моделирования репаративной функции сетчатки для оценки поведения и дифференцировки культивируемых клеток РПЭ при проведении доклинических исследований.
Задачей, на решение которой направлено предлагаемое изобретение, является разработка способа моделирования тканевой структуры сетчатки глаза человека.
Техническим результатом изобретения является создание ex vivo трехмерной модели культуры клеток РПЭ с заданными параметрами количества, плотности и тканевой дифференцировки на биологически инертном матриксе, не ухудшающем рост и дифференцировку клеточной культуры.
Для решения поставленной задачи способ моделирования тканевой структуры сетчатки глаза человека путем колонизирования бесклеточного каркаса сетчатки глаза клетками ретинального пигментного эпителия заключается в том, что бесклеточный каркас сетчатки глаза получают девитализирующей обработкой сетчатки из энуклиированного при аутопсии глазного яблока путем последовательного промывания его в спиртовом растворе при концентрации 70% и в холодном растворе Хэнкса с антибиотиками, удаления по зубчатой линии переднего сегмента глазного яблока, отделения стекловидного тела и нейральной сетчатки от ретинального пигментного эпителия и выделения сетчатки, подрезая ее в области зрительного нерва, выделенную сетчатку для девитализирующей обработки помещают в 10-15 мл 10 мМ ЭДТА в среде DMEM на 20 часов при температуре 4°C, затем обрабатывают 3% раствором Тритона X-100 в течение 2 часов при температуре 4°C и отмывают в дистиллированной воде, далее сетчатку последовательно помещают в 0,025% раствор ДНазы I в 1 М растворе NaCl на 1 час при температуре 4°C и в 4% дезоксихолат натрия на 1-2 часа при температуре 22°C и затем отмывают в дистиллированной воде для удаления детергента и клеточного дебриса, глазную чашу наполняют холодным раствором Хэнкса с ЭДТА и через 15-30 мин собирают клетки нативного ретинального пигментного эпителия, часть собранных клеток переносят в лизирующий раствор TRI®-Reagent из расчета 1 мл раствора на 50-100 мг ткани, пипетируют до гомогенного состояния раствора и лизат клеток нативного ретинального пигментного эпителия инкубируют 5-7 мин при температуре 22°C, другую часть собранных клеток переносят в стерильную среду для выделения, состоящую из питательной среды DMEM/F12 (1:1), дополненной 15% эмбриональной телячьей сывороткой, 2 мМ L-глутамина и антибиотиками и центрифугируют в течение 5-7 мин при 600-800 об/мин, затем клетки ресуспендируют в полной культуральной среде, состоящей из питательной среды DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ного раствора заменимых аминокислот, 1%-ного раствора нейральной добавки N2, 20 нг/мл основного фактора роста фибробластов и антибиотиков, и после пипетирования распределяют в культуральных флаконах без покрытия и культивируют при температуре 37°C и 5% CO2, через 5-7 суток проводят первую смену среды и продолжают культивировать со сменой среды через 2-3 суток до достижения клетками конфлюэнтного монослоя и образования первичной культуры, затем проводят пассирование клеток первичной культуры, используя раствор трипсина-версена (1:3), далее клетки помещают в культуральные флаконы и в слайд-камеры в плотности 1 - 2×104 кл./см2 и культивируют, культивированные в культуральных флаконах клетки 1-5 пассажа в виде адгезивных монослойных культур, коммитированные в нейральном направлении, снимают с культуральной поверхности раствором трипсина-версена (1:3) и в виде суспензии культивированных клеток ретинального пигментного эпителия направляют на колонизирование бесклеточного каркаса сетчатки, причем бесклеточный каркас сетчатки перед колонизированием промывают в питательной культуральной среде DMEM/F12 (1:1) с антибиотиками, переносят в слайд-камеру и заливают полной культуральной средой, а колонизирование бесклеточного каркаса сетчатки глаза осуществляют посредством диффузионного осаждения суспензии культивированных клеток ретинального пигментного эпителия в плотности 4 - 5,5×104 кл./см2 в полной культуральной среде того же состава и культивирования при температуре 37°C и 5% CO2, со сменой среды через 2-3 суток до колонизирования культивированными клетками бесклеточного каркаса сетчатки и формирования монослоя клеток на его поверхности с получением тканевой структуры сетчатки глаза, которую направляют на изучение локализации культивируемых клеток и на иммуногистохимическую (ИГХ) оценку потенциала дифференцировки культивируемых клеток в бесклеточном каркасе сетчатки глаза.
В частном варианте для изучения локализации культивируемых клеток проводят окрашивание ядер клеток витальным красителем Hoechst 33342 для люминесцентной микроскопии.
В другом частном варианте для изучения локализации культивируемых клеток колонизированный клетками бесклеточный каркас сетчатки через 12-25 суток фиксируют 4% параформальдегидом, промывают его в фосфатно-солевом буфере (PBS, pH 7.4), проводят по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт, заливают в парафин, изготавливают гистологические срезы и окрашивают гематоксилином и эозином для световой микроскопии.
В другом частном варианте ИГХ оценку потенциала дифференцировки культивируемых клеток в бесклеточном каркасе сетчатки проводят методом иммунопероксидазного окрашивания, используя мышиные моноклональные антитела к CD31, CD34, тирозиназе, СК7, СК8, СК18, СК19, виментину, GFAP, S100 и NSE.
В другом частном варианте используют антибиотики пенициллин-стрептомицин в стандартной концентрации - пенициллин 100 Ед./мл и стрептомицин 100 мкг/мл.
В другом частном варианте бесклеточный каркас сетчатки глаза после удаления детергента и клеточного дебриса помещают в питательную культуральную среду DMEM/F12 с антибиотиками при температуре 4-8°C на хранение в течение 2-20 суток до фиксации 4% параформальдегидом или колонизирования культвированными клетками.
В другом частном варианте фиксированный бесклеточный каркас сетчатки промывают в фосфатно-солевом буфере (PBS, pH 7.4), проводят по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт, заливают в парафин, изготавливают гистологические срезы, которые монтируют на предметные стекла и окрашивают гематоксилином и эозином для контроля адекватности девитализирующей обработки с помощью световой и/или люминесцентной микроскопии.
В другом частном варианте после инкубирования лизат клеток нативного ретинального пигментного эпителия хранят при температуре минус 20°C до проведения полимеразной цепной реакции (ПЦР) - ПЦР оценки.
В другом частном варианте клетки первичной культуры и культивированные клетки 1-5 пассажа направляют для проведения ПЦР оценки.
В другом частном варианте для проведения ПЦР оценки определяют уровень экспрессии генов, кодирующих транскрипционные факторы Oct4 (POU5F1), Nanog, Рах6 и маркеры дифференцировки Musashi 1, β tubulin III, RPE65 выделением тотальной РНК и синтеза кДНК, при этом уровень экспрессии генов оценивают по интенсивности свечения полос, полученных при электрофоретическом разделении ПЦР-продуктов в 1%-ном агарозном геле на анализаторе гелей.
В другом частном варианте культивирование клеток в слайд-камерах проводят в 1-, 2-, 4- или 8-луночных слайд-камерах, через 2-3 суток, при достижении клетками субконфлюэнтного слоя, их фиксируют охлажденным ацетоном и направляют на ИГХ оценку потенциала дифференцировки клеток.
В другом частном варианте ИГХ оценку потенциала дифференцировки культивированных клеток осуществляют методами иммунопероксидазного и иммунофлуоресцентного окрашивания, используя первичные мышиные моноклональные и кроличьи поликлональные антитела, Рах6, Sox2, виментин, Nestin, β tubulin III, nNOS, тирозин гидроксилаза, NF 68 and 200 kDa, GFAP, CNPase, Iba, N-cadherin, Коннексин-43, RPE65, CRALBP, Recoverin, PanCK, Macrophage antigen, Фибронектин, Factor Von Willebrand, Ki67, а также полимер из набора EnVision™, конъюгированный с пероксидазой хрена, (Dako, Дания) или вторичные антитела: набор SFX с антителами козы против мыши, меченные Alexa Fluor® 488, антитела против иммуноглобулинов кролика, конъюгированные с флуорохромом Texas Red, разведенные в 0,1%-ном растворе бычьего сывороточного альбумина.
В другом частном варианте бесклеточный каркас сетчатки глаза, перенесенный в слайд-камеру, заливают полной культуральной средой, состоящей из питательной среды DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2, 20 нг/мл основного фактора роста фибробластов и антибиотиками.
В другом частном варианте бесклеточный каркас сетчатки глаза, перенесенный в слайд-камеру, заливают полной культуральной средой, состоящей из питательной среды DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина и антибиотиками, и помещают в CO2-инкубатор, имеющий температуру 37°C, 5% CO2 в воздухе для уравновешивания pH среды.
Предлагаемое изобретение иллюстрируется на следующих фигурах.
На фиг.1 представлена схема моделирования тканевой структуры сетчатки глаза человека.
На фиг.2 представлен гистологический препарат нейральной части сетчатки глаза взрослого человека в тесной связи со стекловидным телом, фиксация 4% параформальдегидом, парафиновый срез (контроль). Окраска гематоксилином и эозином, масштабное увеличение Х200.
А - поперечный срез,
Б - продольный срез.
На фиг.3 представлена бесклеточная сетчатка глаза взрослого человека непосредственно после девитализирующей обработки, фазовый контраст.
А - масштабное увеличение Х40,
Б - масштабное увеличение Х200.
На фиг.4 представлена бесклеточная сетчатка глаза взрослого человека на границе с бесклеточным каркасом стекловидного тела, парафиновый срез. Окраска гематоксилином и эозином, масштабное увеличение Х400.
А - через 2 суток после девитализирующей обработки,
Б-Г - через 20 суток после девитализирующей обработки.
На фиг.5 представлена культура клеток ретинального пигментного эпителия глаза взрослого человека 2-го пассажа. Иммунопероксидазное окрашивание, ядра докрашены гематоксилином. Масштабное увеличение Х200.
А - маркер Nestin - коричневое окрашивание,
Б - маркер β tubulin III - коричневое окрашивание,
В - маркер NF 68 and 200 kDa - коричневое окрашивание.
На фиг.6 представлена культура клеток ретинального пигментного эпителия глаза взрослого человека 2-го пассажа. Иммунофлуоресцентное исследование, ядра окрашены DAPI (синее свечение). Масштабная линейка 200 µm.
А - маркер nNOS - красное свечение,
Б - маркер GFAP - красное свечение,
В - маркер CNPase - зеленое свечение.
На фиг.7 представлены результаты ПЦР-анализа, иллюстрирующие характер экспрессии генов RPE65, Oct4, Nanog, Musashi 1, Рах6, β Tubulin III в первичных (ПК) и пассируемых (Р1-Р4) адгезивных клеточных культурах ретинального пигментного эпителия глаза взрослого человека, а также в контролях - в клеточной культуре постнатальных фибробластов (К) и нативном ретинальном пигментном эпителии (Н). В качестве гена для нормировки использован RPL19.
На фиг.8-10 представлена колонизация бесклеточной сетчатки и бесклеточного каркаса стекловидного тела глаза взрослого человека аутологичными клетками ретинального пигментного эпителия 2-го пассажа.
На фиг.8 на 7-е сутки, фазовый контраст.
А - скопления округлых клеток на поверхности бесклеточного каркаса сетчатки, масштабное увеличение X100,
Б - округлые образования, соединенные между собой клетками вытянутой формы, масштабное увеличение Х200.
На фиг.9 - на 7-е сутки, фазовый контраст.
А - округлые образования, соединенные между собой клетками вытянутой формы, масштабное увеличение X100,
Б - две субпопуляции, масштабное увеличение X100.
На фиг.10 - на 12-е сутки, монослой клеток ретинального пигментного эпителия на поверхности бесклеточного каркаса сетчатки глаза, фазовый контраст, масштабное увеличение X100.
На фиг.11 - на 12-е сутки, ядра клеток ретинального пигментного эпителия, образующих монослой на поверхности бесклеточного каркаса сетчатки глаза, окрашены витальным ядерным красителем Hoechst 33342, люминесцентная микроскопия, масштабное увеличение X100.
На фиг.12-16 представлены гистологические срезы бесклеточного каркаса сетчатки глаза взрослого человека, интимно связанного с бесклеточным каркасом стекловидного тела; бесклеточные каркасы колонизированы аутологичными клетками ретинального пигментного эпителия 2-го пассажа, окраска гематоксилином и эозином.
На фиг.12 представлено
А - конгломераты клеток ретинального пигментного эпителия округлой формы на каркасе стекловидного тела, квадратом выделен фрагмент с одним из клеточных конгломератов, масштабное увеличение Х40,
Б - увеличение фрагмента, масштабное увеличение Х400.
На фиг.13 - клетки ретинального пигментного эпителия заселили всю внутреннюю пограничную мембрану сетчатки глаза, граничащую с задней гиалоидной мембраной стекловидного тела, масштабное увеличение X100.
На фиг.14 - монослой кубовидной формы клеток ретинального пигментного эпителия, плотно прилежащих друг к другу; видна миграция клеток в каркас стекловидного тела, масштабное увеличение Х400.
На фиг.15 - клетки ретинального пигментного эпителия уплощенной формы, заселивших внутреннюю пограничную мембрану сетчатки глаза, прилежащей к задней гиалоидной мембране стекловидного тела, масштабное увеличение Х400.
На фиг.16 - конгломерат клеток ретинального пигментного эпителия на поверхности каркаса стекловидного тела, масштабное увеличение Х200.
На фиг.17-28 представлен бесклеточный каркас сетчатки глаза взрослого человека, интимно связанный с бесклеточным каркасом стекловидного тела; бесклеточные каркасы колонизированы аутологичными клетками ретинального пигментного эпителия 2-го пассажа. Иммунопероксидазное окрашивание на серийных парафиновых срезах.
На фиг.17 - без первичных антител (контроль), масштабное увеличение Х200.
На фиг.18 - негативное окрашивание на маркер эндотелиальных клеток CD31.
А - на поверхности внутренней пограничной мембраны бесклеточной сетчатки глаза, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение Х200,
Б - в клеточных конгломератах, располагающихся на бесклеточном каркасе стекловидного тела, масштабное увеличение Х200.
На фиг.19 - негативное окрашивание на маркер эндотелиальных клеток CD34.
А - на поверхности внутренней пограничной мембраны бесклеточной сетчатки глаза, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение Х400,
Б - на поверхности бесклеточной сетчатки глаза и в клеточных конгломератах, масштабное увеличение X100.
На фиг.20 - негативное окрашивание на маркер клеток, синтезирующих меланин, тирозиназу.
А - на поверхности бесклеточной сетчатки, масштабное увеличение Х200,
Б - в клеточных конгломератах, располагающихся на бесклеточном каркасе стекловидного тела, масштабное увеличение Х200.
На фиг.21 - коричневое (позитивное) окрашивание дедифференцированных клеток ретинального пигментного эпителия на маркер эпителия СК7.
А - на поверхности внутренней пограничной мембраны бесклеточной сетчатки глаза, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение Х200,
Б - на поверхности бесклеточной сетчатки глаза и в клеточных конгломератах, масштабное увеличение Х200.
На фиг.22 - коричневое (позитивное) окрашивание мигрирующих клеток ретинального пигментного эпителия на маркер эпителия СК19.
А - на поверхности внутренней пограничной мембраны бесклеточной сетчатки, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение X100,
Б - на поверхности бесклеточной сетчатки глаза в клеточных конгломератах, масштабное увеличение Х200.
На фиг.23 - коричневое (позитивное) окрашивание дифференцированных клеток ретинального пигментного эпителия на маркер эпителия СК8.
А - на поверхности внутренней пограничной мембраны бесклеточной сетчатки глаза, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение Х200,
Б - на поверхности бесклеточной сетчатки глаза в клеточных конгломератах, масштабное увеличение Х400.
На фиг.24 - коричневое (позитивное) окрашивание дифференцированных клеток ретинального пигментного эпителия на маркер эпителия СК18.
А - на поверхности внутренней пограничной мембраны бесклеточной сетчатки глаза, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение X100,
Б, В, Г - на поверхности бесклеточной сетчатки глаза и в клеточных конгломератах, масштабное увеличение Х200.
На фиг.25 - коричневое (позитивное) окрашивание на маркер глиальных клеток виментин.
А - на поверхности внутренней пограничной мембраны бесклеточной сетчатки глаза, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение Х200,
Б - на поверхности бесклеточной сетчатки глаза, масштабное увеличение Х200.
На фиг.26 - коричневое (позитивное) окрашивание на маркер дифференцированных глиальных клеток S100.
А - на поверхности внутренней пограничной мембраны бесклеточной сетчатки глаза, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение Х200,
Б - на поверхности бесклеточной сетчатки глаза и в клеточных конгломератах, масштабное увеличение Х40.
На фиг.27 - коричневое (позитивное) окрашивание на нейрональный маркер NSE.
А - на поверхности внутренней пограничной мембраны бесклеточной сетчатки глаза, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение Х200,
Б - на поверхности бесклеточной сетчатки глаза и в клеточных конгломератах, масштабное увеличение Х40.
На фиг.28 - окрашивание на маркер глиальных клеток GFAP.
A, Б - негативное окрашивание в клетках ретинального пигментного эпителия, располагающихся на поверхности внутренней пограничной мембраны бесклеточной сетчатки, граничащей с бесклеточным каркасом стекловидного тела, масштабное увеличение Х200,
B, Г - негативное окрашивание в клетках ретинального пигментного эпителия, располагающихся на поверхности внутренней пограничной мембраны бесклеточной сетчатки глаза, граничащей с бесклеточным каркасом стекловидного тела, и коричневое (позитивное) окрашивание в бесклеточном каркасе сетчатки (в виде сети), масштабное увеличение Х200,
Д - коричневое (позитивное) окрашивание в отдельных клеточных конгломератах, масштабное увеличение X100,
Е - коричневое (позитивное) окрашивание в отдельных клеточных конгломератах, масштабное увеличение Х400.
На фигурах показано:
1 - гиалоидная строма стекловидного тела;
2 - нейральная сетчатка;
3 - соединение внутренней пограничной мембраны сетчатки с задней гиалоидной мембраной стекловидного тела;
4 - сосудистый каркас;
5 - каркас нейральной части сетчатки, состоящий из базальных мембран клеток Мюллера;
6 - каркас стекловидного тела;
7 - внутренняя пограничная мембрана сетчатки;
8 - субпопуляция клеток ретинального пигментного эпителия, прикрепленная к поверхности бесклеточного каркаса сетчатки;
9 - субпопуляция клеток ретинального пигментного эпителия, распластанная на поверхности культурального пластика и образовавшая монослой;
10 - монослой кубовидной формы клеток, плотно прилежащих друг к другу;
11 - слой клеток уплощенной формы.
Моделирование тканевой структуры сетчатки глаза человека (трехмерной модели) осуществляли по схеме, представленной на фиг.1.
Получение бесклеточного каркаса сетчатки из трупного материала осуществляли путем очищения ее от функционирующих клеток (девитализирующей обработкой).
Глазные яблоки взрослого человека энуклиировали при аутопсии, освобождали от окружающих тканей и промывали один раз в 70%-ном спирте и 3 раза в холодном растворе Хэнкса с антибиотиками в стандартной концентрации - пенициллин 100 Ед./мл и стрептомицин 100 мкг/мл. Передний сегмент глазного яблока удаляли круговым разрезом позади зубчатой линии, которая хорошо определялась при обычном освещении, как граница между темным и светлым участками. Стекловидное тело и нейральную сетчатку отделяли от ретинального пигментного эпителия (РПЭ) и выделяли, подрезая нейральную сетчатку в области зрительного нерва.
Выделенную сетчатку обрабатывали по методике Carlson [Carlson Е.С. 1989. Fenestrated subendothelial basement membranes in human retinal capillaries. Invest. Ophthalmol. Vis. Sci. 30 (9): 1923-1932]. Сетчатку целиком помещали в 10-15 мл 10 мМ ЭДТА в среде DMEM на 20 часов при температуре 4°C. Затем сетчатку обрабатывали 3% раствором Тритона Х-100 в течение 2 часов при температуре 4°C, несколько (3-4) раз отмывали в дистиллированной воде. Далее ее последовательно помещали в 0,025% раствор ДНазы I в 1 М растворе NaCl на 1 час при 4°C, затем в 4% дезоксихолат натрия на 1-2 часа при температуре 22°C и затем помещали несколько (3-5) раз в дистиллированную воду для того, чтобы удалить детергент и клеточный дебрис.
Все растворы были предварительно стерилизованы через 0,22 мкм фильтр.
Наблюдение за сетчаткой проводили с помощью инвертированного микроскопа Olympus СКХ31 (Япония), а фотографирование с помощью камеры Olympus DP70 (Япония).
После последовательной обработки детергентами и ДНазой сетчатка уменьшалась в размере и при исследовании с помощью фазово-контрастной микроскопии представляла собой бесклеточный каркас, состоящий из волокон внеклеточного матрикса, компонентов внутренней пограничной мембраны сетчатки (коллагеновых фибрилл и базальных мембран Мюллеровских клеток) и базальных мембран капилляров.
Бесклеточный каркас сетчатки после отмывания в дистиллированной воде для удаления детергента и клеточного дебриса помещали в питательную культуральную среду DMEM/F12 с антибиотиками при температуре 4-8°C на хранение в течение 2-20 суток до фиксации 4% параформальдегидом или колонизирования культивированными клетками.
Фиксированный бесклеточный каркас сетчатки промывали в фосфатно-солевом буфере (PBS, pH 7.4), проводили по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт, заливали в парафин, изготавливали гистологические срезы, которые монтировали на предметные стекла и окрашивали гематоксилином и эозином для контроля адекватности девитализирующей обработки с помощью световой и/или люминесцентной микроскопии.
Получение нативного РПЭ.
Для получения нативного РПЭ глазную чашу глазного яблока после извлечения нейральной сетчатки наполняли холодным раствором Хэнкса с ЭДТА и спустя 15-30 мин начинали процесс сбора клеток нативного РПЭ. В течение всей процедуры глазные чаши находились на льду в растворе Хэнкса. Контролируя процесс под бинокулярной лупой, наконечником 200 мкл проводили по слою и одновременно засасывали клетки, отделившиеся от мембраны Бруха.
Часть клеток РПЭ переносили в стерильные пробирки, убирали надосадочную жидкость и заливали лизирующим раствором TRI®-Reagent (Sigma, США) из расчета 1 мл раствора TRI®-Reagent на 50-100 мг ткани, пипетировали до гомогенного состояния раствора, лизат инкубировали 5 мин при температуре 22°C, а затем переносили в стерильные пробирки и хранили при температуре минус 20°C до проведения молекулярного анализа выделения тотальной РНК и синтеза кДНК (ПЦР оценка).
Другую часть клеток РПЭ направляли на получение культур клеток.
Получение культур клеток РПЭ.
Для культивирования клетки РПЭ переносили в стерильную среду для выделения следующего состава: питательная среда DMEM/F12 (1:1), дополненная 15% эмбриональной телячьей сыворотки, 2 мМ L-глутамина и антибиотиками пенициллином-стрептомицином в стандартной концентрации, и центрифугировали в течение 5-7 мин при 600-800 об/мин. Далее клетки РПЭ ресуспендировали в полной культуральной среде, следующего состава: питательная среда DMEM/F12 (1:1), дополненная 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2 (Gibco, Invitrogen, США), 20 нг/мл основного фактора роста фибробластов (оФРФ) и антибиотиками пенициллином-стрептомицином в стандартной концентрации, затем после мягкого пипетирования равномерно распределяли в культуральных флаконах площадью 25 см2 без покрытия (Greiner, Германия) и культивировали при температуре 37°C и 5% CO2. Спустя 5-7 суток проводили первую смену среды и продолжали культивировать со сменой среды через 2-3 суток до достижения клетками конфлюэнтного монослоя и образования первичной культуры.
Далее проводили пассирование конфлюэнтных первичных культур клеток, используя раствор трипсина - версена (1:3). Клетки культивировали в плотности 1 - 2×104 кл./см2 в полной культуральной среде, следующего состава: питательная среда DMEM/F12 (1:1), дополненная 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2 (Gibco, Invitrogen, США), 20 нг/мл основного фактора роста фибробластов (оФРФ) и антибиотиками пенициллином-стрептомицином в стандартной концентрации. Культивирование проводили во флаконах и в 2 или 4-луночных слайд-камерах для культур клеток (BD Falcon™, BD Biosciences, США). Слайд-камера представляла собой предметное стекло для роста клеток, к которому прикреплена разделительная решетка из пластика, образующая камеру с определенным числом лунок (существуют 1-, 2-, 4- и 8-луночные слайд-камеры), прикрываемая сверху крышкой для поддержания стерильных условий.
Клетки РПЭ, культивированные на слайд-камерах, через 2-3 суток при достижении ими субконфлюэнтного слоя фиксировали охлажденным ацетоном. Далее предметные стекла с клетками освобождали от разделительных решеток, высушивали при температуре 20°C, заворачивали в фольгу и хранили в низкотемпературном холодильнике при температуре минус 70-80°C до проведения ИГХ оценки дифференцировки полученных культур.
Клетки РПЭ, культивированные в культуральных флаконах, росли в виде адгезивных монослойных культур. Наблюдение за клеточными культурами РПЭ проводили с помощью инвертированного микроскопа Olympus СКХ31 (Япония).
С одних культуральных флаконов клеточные культуры РПЭ 1-5 пассажа, коммитированные в нейральном направлении, снимали с культуральной поверхности раствором трипсина-версена (1:3) и в виде суспензии использовали для колонизирования бесклеточного каркаса сетчатки в плотности 4 - 5,5×104 кл./см2.
С других культуральных флаконов клетки первичной культуры и культуры клеток РПЭ 1-5 пассажа использовали для проведения ПЦР оценки дифференцировки клеток. Для этого монослой клеток промывали 1 раз DPBS (ПанЭко, Россия), заливали лизирующим раствором TRI®-Reagent (Sigma, США) из расчета 1 мл на 25 см2 культурального флакона, клеточный лизат пипетировали до гомогенного прозрачного состояния раствора, гомогенат инкубировали 5 мин при температуре 22°C, а затем переносили в стерильные пробирки и хранили при температуре минус 20°C до проведения выделения тотальной РНК и синтеза кДНК (ПЦР оценки дифференцировки культивируемых клеток РПЭ).
ИГХ и ПЦР оценку дифференцировки (коммитирования в нейральном направлении) культивируемых клеток РПЭ осуществляли следующим образом.
Проведение ИГХ оценки дифференцировки культивируемых клеток РПЭ.
Для ИГХ оценки дифференцировки (коммитирования в нейральном направлении) клеток в первичной и пассируемых культурах применяли методы иммунопероксидазного и иммунофлуоресцентного окрашивания, используя первичные мышиные моноклональные и кроличьи поликлональные антитела, Рах6, Sox2, виментин, Nestin, β tubulin III, nNOS, тирозин гидроксилаза, NF 68 and 200 kDa, GFAP, CNPase, Iba, N-cadherin, Коннексин-43, RPE65, CRALBP, Recoverin, PanCK, Macrophage antigen, Фибронектин, Factor Von Willebrand, Ki67, а также полимер из набора EnVision™, конъюгированный с пероксидазой хрена (Dako, Дания), или вторичные антитела: набор SFX с антителами козы против мыши, меченные Alexa Fluor® 488 (Molecular Probes, США), антитела против иммуноглобулинов кролика, конъюгированные с флуорохромом Texas Red (Jackson ImmunoResearch, США), разведенные в 0,1%-ном растворе бычьего сывороточного альбумина (BSA, Sigma, США).
В таблице 1 приведена характеристика использованных антител.
Предметные стекла с фиксированными на них клетками РПЭ извлекали из низкотемпературного холодильника, освобождали от фольги и после испарения конденсата помещали в 0,01 М фосфатно-солевой буфер (PBS, pH 7.4) (ООО «БиолоТ», Россия) для отмывки от ацетона.
Иммунопероксидазное окрашивание препаратов проводили по следующей схеме: блокировка эндогенной пероксидазы с Dual Endogenous Enzyme Block из набора EnVision™ (Dako, Дания) в течение 10 мин, блокировка неспецифического связывания с 1%-ным раствором BSA (Sigma, США) в течение 20 мин, инкубация с первичными антителами в течение 17 часов при температуре 4°C, инкубация с полимером из набора EnVision™, конъюгированным с пероксидазой хрена в течение 15 мин, инкубация с раствором DAB 5-10 мин. Далее препараты докрашивали гематоксилином, покрывали водной нефлуоресцентной средой (Shandon Immu-Mount™, Thermo Electron Corporation, США) и заключали под покровное стекло.
Проведение иммунофлуоресцентного исследования препаратов включало: блокировку неспецифического связывания реагентом Image-iT™ FX signal enhancer из SFX набора (Molecular Probes, США) в течение 30 мин при температуре 22°C, инкубацию с первичными антителами в течение 17 часов при температуре 4°C, связывание первичных антител с соответствующими вторичными, меченными флуорохромами, антителами в течение 1 часа при температуре 22°C, покрытие средой с ядерным красителем DAPI (Vectashield®, Vector Laboratories, США) либо докрашивание ядерным люминесцентным красителем Hoechst 33342 в концентрации 5 мкг/мл (Sigma, США) с последующим покрытием глицерином, заключение препаратов под покровное стекло.
ИГХ оценку и фотографирование проводили с помощью светового микроскопа Olympus АНЗ (Япония).
Проведение ПЦР оценки дифференцировки культивируемых клеток РПЭ.
Для молекулярного анализа (ПЦР оценки) выделение тотальной РНК проводили согласно инструкции производителя TRT®-Reagent (Sigma, США), синтез кДНК проводили с помощью RevertAid Н Minus Kit (Fermentas), используя 1 мкг тотальной РНК, обработанной ДНКазой (Fermentas). Для проведения ПЦР оценки определяли уровень экспрессии генов, кодирующих транскрипционные факторы Oct4 (POU5F1), Nanog, Рах6, и маркеры дифференцировки Musashi 1, β tubulin III, RPE65.
В таблице 2 представлена характеристика функции генов и последовательность праймеров для ПЦР оценки.
Условия амплификации: температура 94°C - 45 с, температура 56°C - 45 с, температура 72°C - 1 с, 30-40 циклов. кДНК библиотеки были нормированы по большому рибосомальному белку человека с молекулярной массой 19 кДа (RPL19). Уровень экспрессии генов оценивали по интенсивности свечения полос, полученных при электрофоретическом разделении ПЦР-продуктов в 1%-ном агарозном геле, на анализаторе гелей (BIO-RAD™XR, США).
Колонизирование бесклеточного каркаса сетчатки глаза взрослого человека культивируемыми клетками РПЭ осуществляли следующим образом.
Бесклеточный каркас сетчатки промывали несколько (5-7) раз в питательной среде DMEM/F12 (1:1) с антибиотиками пенициллином-стрептомицином в стандартной концентрации, переносили в стерильную 1-луночную слайд-камеру (BD Falcon™, BD Biosciences, США), заливали 2 мл одного из двух следующих составов полной культуральной среды: питательной средой DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2 (Gibco, Invitrogen, США), 20 нг/мл основного фактора роста фибробластов (оФРФ) и антибиотиками пенициллином-стрептомицином в стандартной концентрации или питательной средой DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина и антибиотиками пенициллином-стрептомицином в стандартной концентрации, и помещали в CO2-инкубатор, имеющий температуру 37°C, 5% CO2 в воздухе, для уравновешивания pH среды.
На бесклеточный каркас сетчатки человека высаживали посредством диффузионного осаждения суспензии культивированных клеток РПЭ как аутологичных, так и аллогенных, выращенных в монослойных культурах. Клетки РПЭ высаживали в плотности 4 - 5,5×104 кл./см2 в 2-3 мл полной культуральной среды того же состава, в котором находился бесклеточный каркас. Клетки культивировали в присутствии бесклеточного каркаса при температуре 37°C и 5% CO2, со сменой среды через 2-3 суток до колонизирования клетками бесклеточного каркаса сетчатки и формирования монослоя клеток на его поверхности с получением тканевой структуры сетчатки глаза, которую направляли на изучение локализации культивируемых клеток и на ИГХ оценку потенциала дифференцировки культивируемых клеток в бесклеточном каркасе сетчатки глаза.
Наблюдение за поведением клеточных культур РПЭ на бесклеточной сетчатке проводили с помощью инвертированного микроскопа Olympus СКХ31 (Япония).
При определении на поверхности бесклеточного каркаса сетчатки монослоя клеток РПЭ проводили окрашивание ядер последних витальным красителем Hoechst 33342 для люминисцентной микроскопии.
Фиксацию каркасов сетчаток, колонизированных культивированными клетками РПЭ человека, проводили через 12-25 суток 4% параформальдегидом. Фиксированные препараты промывали в фосфатно-солевом буфере (PBS, pH 7.4), проводили по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт и заливали в парафин.
Из залитых в парафин блоков изготавливали на микротоме гистологические срезы толщиной 5 мкм, которые монтировали на предметные стекла, покрытые адгезивом. Изучение локализации культивированных клеток РПЭ в бесклеточном каркасе сетчатки проводили на серийных парафиновых срезах, окрашенных гематоксилином и эозином, с помощью светового микроскопа Olympus АН3 (Япония) для световой микроскопии.
Проведение ИГХ оценки потенциала дифференцировки культивированных клеток РПЭ в бесклеточном каркасе сетчатки на серийных парафиновых срезах проводили методом иммунопероксидазного окрашивания с реактивами набора LSAP (Dako, Дания), используя готовые к использованию мышиные моноклональные антитела к CD31, CD34, тирозиназе, СК7, СК8, СК18, СК19, виментину, GFAP, S100 и NSE (Dako, Дания), по следующей схеме: тепловая обработка депарафинированных срезов в СВЧ-печи (мощность 650-750 Вт) в течение 5 мин с соответствующими растворами для восстановления антигена (Dako, Дания), блокировка эндогенной пероксидазы раствором из набора LSAP в течение 10 мин, блокировка неспецифического связывания 1%-ным раствором BSA в течение 20 мин, инкубация с первичными антителами в течение 17 часов при температуре 4°C, инкубация с вторичными биотинилированными антителами из набора LSAP в течение 20 мин, инкубация с стрептавидином, конъюгированным пероксидазой хрена, из набора LSAP в течение 15 мин, инкубация с раствором DAB 5-10 мин. Далее препараты докрашивали гематоксилином, покрывали водной средой (Faramount, Dako, Дания) и заключали под покровное стекло.
Предлагаемое изобретение поясняется следующими примерами.
Пример 1.
В качестве контроля при оценке адекватности девитализирующих обработок других сетчаток использовали сетчатку без девитализирующей обработки, которую получали следующим образом. Глазное яблоко освобождали от окружающих тканей и промывали один раз в 70%-ном спирте и 3 раза в холодном растворе Хэнкса с антибиотиками в стандартной концентрации - пенициллин 100 Ед./мл и стрептомицин 100 мкг/мл. Передний сегмент глазного яблока удаляли круговым разрезом позади зубчатой линии, которая хорошо определяется при обычном освещении, как граница между темным и светлым участками. Стекловидное тело и нейральную сетчатку отделяли от РПЭ и выделяли, подрезая нейральную сетчатку в области зрительного нерва. Сетчатку после извлечения целиком помещали в 4% параформальдегид для фиксирования. Фиксированную сетчатку промывали в фосфатно-солевом буфере (PBS, pH 7.4), проводили по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт и заливали в парафин. Из залитого в парафин блока изготавливали на микротоме гистологические срезы толщиной 5 мкм, которые монтировали на предметные стекла и окрашивали гематоксилином и эозином (фиг.2А, Б).
Бесклеточный каркас сетчатки глаза получали девитализирующей обработкой энуклиированного глазного яблока при аутопсии следующим образом.
Глазные яблоки взрослого человека (ж, 50 лет) освобождали от окружающих тканей и промывали один раз в 70%-ном спирте и 3 раза в холодном растворе Хэнкса с антибиотиками пенициллином-стрептомицином в стандартной концентрации. Передний сегмент глаза удаляли круговым разрезом позади зубчатой линии. Стекловидное тело и нейральную сетчатку отделяли от РПЭ и выделяли, подрезая нейральную сетчатку в области зрительного нерва.
Сетчатка из одного глазного яблока после извлечения была целиком помещена в 10 мМ ЭДТА (в среде DMEM) объемом 15 мл на 20 час при температуре 4°C. Затем сетчатка была обработана 3% раствором Тритона Х-100 в течение 2 часов при 4°C, 3 раза отмыта дистиллированной водой. Далее она была последовательно помещена в 0,025% раствор ДНазы I в 1 М растворе NaCl на 1 час при температуре 4°C, затем перенесена в 4% дезоксихолат натрия на 1 час при 22°C и 3 раза промыта дистиллированной водой для удаления детергента и клеточного дебриса. Все растворы были предварительно стерилизованы через 0,22 мкм фильтр. После такой последовательной обработки сетчатка была исследована в фазовом контрасте с помощью инвертированного микроскопа Olympus СКХ31 (Япония) (фиг.3А, Б). Далее она хранилась в течение 20 суток при температуре 4°C в среде DMEM/F12 с антибиотиками пенициллином-стрептомицином в стандартной концентрации до фиксации 4% параформальдегидом. Фиксированную сетчатку промывали в фосфатно-солевом буфере (PBS, pH 7.4), проводили по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт и заливали в парафин.
Из залитого в парафин блока изготавливали на микротоме гистологические срезы толщиной 5 мкм, которые монтировали на предметные стекла и окрашивали гематоксилином и эозином (фиг.4Б, В, Г).
Данная сетчатка использовалась для контроля адекватности девитализирующей обработки и возможности длительного хранения. Как показало микроскопическое исследование гистологических срезов, видно, что сетчатка после последовательной обработки детергентами и ДНазой представляла собой бесклеточный каркас, состоящий из волокон внеклеточного матрикса внутренней пограничной мембраны сетчатки и вплетенных в нее фибрилл стекловидного тела, что свидетельствует об адекватности девитализирующей обработки. Отсутствие микробной контаминации культуральной среды при длительном хранении бесклеточного каркаса сетчатки, свидетельствующее о сохранении стерильности биоматериала, позволяет использовать его после длительного периода ожидания, необходимого для наращивания клеточной массы.
Сетчатка из второго глазного яблока после извлечения была целиком помещена в 10 мМ ЭДТА (в среде DMEM) объемом 10 мл на 20 час при температуре 4°C. Затем сетчатка была обработана 3% раствором Тритона Х-100 в течение 2 часов при температуре 4°C, 3 раза отмыта дистиллированной водой. Далее она была последовательно помещена в 0,025% раствор ДНазы I в 1 М растворе NaCl на 1 час при температуре 4°C, затем перенесена в 4% дезоксихолат натрия на 1 час при температуре 22°C и 3 раза промыта дистиллированной водой для удаления детергента и клеточного дебриса. После такой последовательной обработки детергентами и ДНазой сетчатка хранилась в течение 20 суток при температуре 4°C в среде DMEM/F12 с антибиотиками пенициллином-стрептомицином в стандартной концентрации до колонизирования ее культивированными клетками РПЭ.
Две глазные чаши после извлечения нейральных сетчаток наполняли холодным раствором Хэнкса с ЭДТА и спустя 15 мин начинали процесс сбора клеток нативного РПЭ. В течение всей процедуры глазные чаши находились на льду в растворе Хэнкса. Контролируя процесс под бинокулярной лупой, наконечником 200 мкл проводили по слою и одновременно засасывали клетки, отделившиеся от мембраны Бруха.
Клетки нативного РПЭ, собранные из одного глазного яблока, переносили в стерильные пробирки, убирали надосадочную жидкость и заливали лизирующим раствором TRI®-Reagent (Sigma, США) из расчета 1 мл раствора TRI®-Reagent на 100 мг ткани, пипетировали до гомогенного состояния раствора, лизат клеток инкубировали 5 мин при температуре 22°C, переносили в стерильные пробирки и хранили при температуре минус 20°C до проведения выделения тотальной РНК и синтеза кДНК (ПРЦ оценки).
В клетках нативного РПЭ при проведении ПЦР оценки выявлялась экспрессия на мажорном уровне гена RPE65, являющегося маркером клеток РПЭ (фиг.7). Белковый продукт гена участвует в цикле фототрансдукции при взаимодействии РПЭ с фоторецепторами. Кроме того, в нативном РПЭ наблюдалась экспрессия на минорном уровне гена плюрипотеного статуса Oct4 (POU5F1) и маркеров нейральных стволовых клеток Musashi 1 и Рах6, что позволяет предположить наличие в нативной ткани немногочисленной популяции клеток со свойствами нейральных стволовых-прогениторных клеток.
Клетки нативного РПЭ, собранные из второго глазного яблока, переносили в стерильную среду для выделения следующего состава: питательная среда DMEM/F12 (1:1), дополненная 15% эмбриональной телячьей сыворотки, 2 мМ L-глутамина и антибиотиками пенициллина-стрептомицина в стандартной концентрации, и центрифугировали в течение 5 мин при 800 об/мин. Клетки РПЭ ресуспендировали в полной культуральной среде, следующего состава: питательная среда DMEM/F12 (1:1), дополненная 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2 (Gibco, Invitrogen, США), 20 нг/мл основного фактора роста фибробластов (оФРФ) и антибиотиками пенициллина-стрептомицина в стандартной концентрации, затем после мягкого пипетирования равномерно распределяли в стерильных пластиковых флаконах площадью 25 см2 без покрытия (Greiner, Германия) и культивировали при температуре 37°C и 5% CO2. Спустя 5-7 суток проводили первую смену среды и продолжали культивировать со сменой среды через 2-3 суток до достижения клетками конфлюэнтного монослоя и образования первичной культуры.
Пассирование конфлюэнтной первичной культуры проводили, используя раствор трипсина-версена (1:3). Клетки культивировали в плотности 1 - 2×104 кл./см2 в полной культуральной среде, следующего состава: питательная среда DMEM/F12 (1:1), дополненная 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2 (Gibco, Invitrogen, США), 20 нг/мл основного фактора роста фибробластов (оФРФ) и антибиотиками пенициллина-стрептомицина в стандартной концентрации. Культивирование проводили во флаконах и в 2- или 4-луночных слайд-камерах для культур клеток (BD Falcon™, BD Biosciences, США). Клетки РПЭ на слайд-камерах через 2-3 суток при достижении ими субконфлюэнтного слоя фиксировали охлажденным ацетоном. Далее предметные стекла с клетками освобождали от разделительных решеток, предметные стекла высушивали при 20°C, заворачивали в фольгу и хранили в низкотемпературном холодильнике при температуре минус 70-80°C до проведения ИГХ оценки полученных культур.
Клеточные культуры при ИГХ оценке были положительны на пронейральные (Рах6), нейральные (Nestin (фиг.5А), β тубулин III (фиг.5Б), nNOS (фиг.6А), ТН, NF 68 and 200kDa (фиг.5В), N-cadherin, Recoverin) и глиальные (виментин, GFAP (фиг.6Б), CNPase (фиг.6В)) маркеры, а также маркер эпителиальных клеток PanCK. ИГХ оценку и фотографирование проводили с помощью светового микроскопа Olympus АН3 (Япония).
Клетки РПЭ во флаконах росли в виде адгезивных монослойных культур. Наблюдение за клеточными культурами РПЭ проводили с помощью инвертированного микроскопа Olympus СКХ31 (Япония).
С одних культуральных флаконов клетки первичной культуры и культуры клеток РПЭ 1-5 пассажа использовали для проведения ПЦР оценки. Для этого монослой клеток промывали 1 раз DPBS (ПанЭко, Россия), заливали лизирующим раствором TRI®-Reagent (Sigma, США) из расчета 1 мл на 25 см2 культурального флакона, клеточный лизат пипетировали до гомогенного прозрачного состояния раствора, гомогенат инкубировали 5 мин при температуре 22°C, а затем переносили в стерильные пробирки и хранили при температуре минус 20°C до проведения выделения тотальной РНК и синтеза кДНК.
ПЦР оценка культивируемых клеток РПЭ взрослого человека по сравнению с нативным РПЭ показала резкое падение уровня экспрессии гена RPE65 (фиг.7), маркера клеток РПЭ, участвующего в цикле фототрансдукции при взаимодействии их с фоторецепторами, что свидетельствует о начале дедифференцировки. В клетках первичной и пассируемых культур одновременно детектировалась экспрессия генов-маркеров стволовых-прогениторных клеток: Oct4 (POU5F1), Nanog, Musashi 1 и Рах6.
Наличие экспрессии нейральных маркеров β tubulin III, Рах6 и Musashi 1 свидетельствовало о нейральном потенциале клеточных культур РПЭ.
Таким образом, результаты проведенного ИГХ оценки и молекулярно-генетического анализа показали, что в условиях культивирования клетки РПЭ глаза взрослого человека претерпевают сильные изменения, они способны к дедифференцировке и развитию по нейральному пути.
Еще с одного культурального флакона клеточные культуры РПЭ 2-го пассажа (20 суток культивирования), коммутированные в нейральном направлении, снимали с культуральной поверхности раствором трипсина-версена (1:3) и в виде суспензии в плотности 5,5×104 кл./см2 использовали для колонизирования аутологичного бесклеточного каркаса сетчатки, хранившегося в течение 20 суток при температуре 4°C в среде DMEM/F12.
Перед нанесением культивированных клеток РПЭ бесклеточная сетчатка была исследована под инвертированным микроскопом на предмет микробной контаминации. Отсутствие микробной контаминации культуральной среды при длительном хранении бесклеточного каркаса сетчатки, свидетельствующее о сохранении стерильности биоматериала, позволило использовать его после длительного периода ожидания, необходимого для наращивания аутологичной клеточной массы. Бесклеточная сетчатка была промыта 5 раз в питательной среде DMEM/F12 (1:1) с антибиотиками пенициллином-стрептомицином в стандартной концентрации и перенесена в стерильную 1-луночную слайд-камеру (BD Falcon™, BD Biosciences, США), залита 2 мл полной культуральной среды, состоящей из питательной среды DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2 (Gibco, Invitrogen, США), 20 нг/мл основного фактора роста фибробластов (оФРФ) и антибиотиками пенициллина-стрептомицина в стандартной концентрации, и помещена в CO2-инкубатор, имеющий температуру 37°C, 5% CO2 в воздухе, для уравновешивания pH среды.
На бесклеточный каркас сетчатки высаживали посредством диффузионного осаждения суспензию аутологичных клеток РПЭ 2-го пассажа в количестве 470 тыс (плотностью 5,5×104 кл./см2) в 3 мл полной культуральной среды того же состава, в котором находился бесклеточный каркас. Клетки культивировали при температуре 37°C и 5% CO2, со сменой среды через 2-3 суток.
На 2-е сутки при исследовании с помощью фазово-контрастного микроскопа отмечалось разделение клеток на 2 субпопуляции: одни клетки распластывались на поверхности слайд-камеры, а другие прикреплялись к поверхности бесклеточного матрикса, но оставались округлой формы.
На 7-е сутки на поверхности бесклеточного каркаса сетчатки определялись скопления клеток округлой формы (фиг.8А; фиг.9Б), а также округлые образования, соединенные между собой вытянутой формы клетками (фиг.8Б; фиг.9А).
На 12-е сутки на поверхности бесклеточного каркаса сетчатки определялся монослой (фиг.10), ядра в котором окрашивались витальным красителем Hoechst 33342 (фиг.11).
Фиксацию колонизированного культивированными клетками бесклеточного каркаса сетчатки 4% параформальдегидом проводили через 12 суток. Фиксированные препараты промывали в фосфатно-солевом буфере (PBS, pH 7.4), проводили по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт и заливали в парафин.
Из залитых в парафин блоков изготавливали на микротоме гистологические срезы толщиной 5 мкм, которые монтировали на предметные стекла, покрытые адгезивом.
Изучение локализации культивируемых клеток РПЭ в бесклеточной сетчатке проводили на серийных парафиновых срезах, окрашенных гематоксилином и эозином, с помощью светового микроскопа Olympus АН3 (Япония). Клетки РПЭ заселили всю внутреннюю пограничную мембрану сетчатки, граничащую с задней гиалоидной мембраной стекловидного тела. При этом они образовывали и монослой кубовидной формы клеток, плотно прилежащих друг к другу (фиг.13, фиг.14), и слой клеток уплощенной формы (фиг.13; фиг.15).
Кроме того, клетки РПЭ образовывали конгломераты клеток РПЭ округлой формы и на поверхности бесклеточного матрикса сетчатки и на поверхности стекловидного тела (в местах отсутствия сетчатки) (фиг.12А, Б, фиг.16).
ИГХ оценку дифференцировки культивируемых клеток РПЭ, заселивших бесклеточную сетчатку, проводили иммунопероксидазным методом, используя готовые к использованию мышиные моноклональные антитела к CD31, CD34, тирозиназе, СК7, СК8, СК18, СК19, виментину, GFAP, S100 и NSE (Dako, Дания).
Клетки РПЭ, заселившие сетчатку, были негативны на маркеры эндотелиальных клеток (CD31 (фиг.18А, Б), CD34 (фиг.19А, Б)) и маркер меланоцитов (тирозиназу (фиг.20А, Б)). Позитивное окрашивание отмечалось на маркеры дифференцированных (СК8 (фиг.23А, Б) и СК18 (фиг.24А, Б, В, Г)) и культивированных (СК7 (фиг.21А, Б) и СК19 (фиг.22А, Б)) клеток РПЭ, а также на глиальные (GFAP (фиг.28А, Б, В, Г, Д, Е), S100 (фиг.26А, Б), виментин (фиг.25А, Б)) и нейрональный (NSE) (фиг.27А, Б) маркеры. GFAP-позитивное окрашивание определялось в виде сети в округлых образованиях (фиг.28В, Е).
Негативное окрашивание на маркеры эндотелиальных клеток (CD31 и CD34) свидетельствует как об отсутствии новообразованных сосудов, так и об адекватности девитализирующей обработки (гибели эндотелиальных клеток сосудов сетчатки после последовательной обработки детергентами и ДНазой).
Негативное окрашивание на тирозиназу свидетельствует об отсутствии клеток, синтезирующих меланин, в т.ч. меланоцитов. Эти данные еще раз подтверждают данные, полученные при ИГХ анализе культивируемых клеток РПЭ, об отсутствии контаминирования культур клеток РПЭ клетками из подлежащей хориоидеи (фибробластами, меланоцитами, эндотелиальными клетками и перицитами).
В нативном РПЭ в норме всех видов млекопитающих, в том числе и человека, главными промежуточными филаментами являются цитокератины СК8 и СК18, маркеры простого эпителия, причем доминантным типом является СК18. Маркеры протокового эпителия СК7 и СК19 в клетках нативного РПЭ человека не встречаются. Эта специфичность делает СК18 эффективным маркером для идентификации клеток РПЭ.
В культуре клетки РПЭ экспрессируют СК 7 и СК19, а экспрессия СК8 и СК18 либо не изменяется, либо исчезает.
Как видно из характера окрашивания на цитокератины клеток РПЭ взрослого человека, заселившего бесклеточный каркас сетчатки, доминантным типом является СК18. В тех же местах, где локализуется СК18, обнаруживаются в убывающем порядке СК8, СК7 и СК19. СК19 экспрессируется в небольшом числе клеток. Наличие цитокератин-позитивного окрашивания во всех клеточных образованиях, обнаруженных в бесклеточном матриксе, свидетельствует о получении клеточных культур из РПЭ. Наличие СК18- и СК8-позитивньгх клеток свидетельствует о дифференцировке культивированных клеток в зрелые клетки РПЭ. Кроме того, учитывая существующие доказательства того, что СК19 экспрессируется в мигрирующих клетках РПЭ, а также то, что с клеточной миграцией может быть связан и СК18, можно предположить, что окончательной дифференцировки все же не произошло и можно ожидать дальнейшего развития при более длительном культивировании. Эту же мысль подтверждает ИГХ окрашивание бесклеточного каркаса сетчатки, колонизированного клетками РПЭ взрослого человека, на другие маркеры.
В нативной сетчатке взрослого человека в клетках Мюллера и астроцитах экспрессируется GFAP, маркер астроцитов и нейральных стволовых клеток, и виментин. Тогда как внутриклеточный Са2+-связывающий регуляторный белок S100 экспрессируется в синапсах между нейронами (фоторецепторными и биполярными клетками) сетчатки взрослого человека.
Клетки РПЭ человека in vitro приобретают выраженное окрашивание на виментин. Выявление одновременной экспрессии глиальных маркеров (виментина, S100 и GFAP) в культивируемых клетках РПЭ связано с дифференцировкой клеток в глиальном направлении. Астроглиальные клетки - это наиболее многочисленные клетки в нервной ткани. Они образуют трехмерную сеть, которая является опорным каркасом для нейронов.
В исследовании развития астроцитов в субвентрикулярной зоне постнатального головного мозга взрослого человека было показано, что S100 является маркером зрелых глиальных клеток (астроцитов), который еще долго экспрессируется после окончания экспрессии GFAP. S100 колокализуется с промежуточными филаментами GFAP и виментином и участвует в их фосфорилировании.
NSE или γ (нейрон-специфическая енолаза, гамма-энолаза) - гликолитический фермент, участвующий в метаболизме глюкозы. Фермент NSE присутствует в клетках нейроэктодермального происхождения, в том числе в дифференцированных клетках РПЭ, нейронах головного мозга и периферической нервной ткани. В культивируемых клетках РПЭ NSE исчезает.
Как видно из примера, ИГХ окрашивание клеток РПЭ взрослого человека, заселивших бесклеточный каркас сетчатки, показало наличие реакции на цитокератины (СК7, 8, 18, 19), нейральный (NSE) и глиальные (виментина, S100 и GFAP) маркеры. В клетках, располагающихся на поверхности внутренней пограничной мембраны, выявляется окрашивание на цитокератины, что свидетельствует об их сходстве с нативными клетками РПЭ, однако одновременное выявление с цитокератинами экспрессии NSE, виментина и S100 указывает на незавершенность дифференцировки клеток. GFAP в этих клетках не обнаруживается. Экспрессия этого белка наблюдается в волокнистых структурах, а также в клеточных образованиях, собранных в округлые конгломераты. Это указывает на то, что дедифференцированные клетки РПЭ человека в условиях 3D культивирования на девитализированном матриксе сетчатки детерминируются в нейральном направлении.
В таблице 3 представлены данные сравнения локализации ИГХ маркеров в нейральной сетчатке и в клетках РПЭ in vivo и in vitro.
Клетки РПЭ заселяли внутреннюю пограничную мембрану сетчатки, граничащую с задней гиалоидной мембраной стекловидного тела, где они, во-первых, образовывали монослой кубовидной формы клеток, плотно прилежащих друг к другу, по морфологии и ИГХ маркерам соответствующий дифференцированным клеткам РПЭ нативной сетчатки. Во-вторых, клетки РПЭ образовывали слой клеток, уплощенной формы, располагающихся более разряжено, по форме напоминающих глиальные клетки нейральной сетчатки. В-третьих, клетки РПЭ образовывали конгломераты, которые обнаруживались и на поверхности бесклеточного каркаса сетчатки и на поверхности стекловидного тела (в местах отсутствия сетчатки). При микроскопическом и ИГХ исследованиях эти конгломераты представляли собой матрикс нейральной части сетчатки, заселенный клетками РПЭ.
Как видно из примера, бесклеточный каркас сетчатки через 20 суток после девитализирующей обработки успешно колонизировался аутологичными культивированными клетками РПЭ. Также исследование показало, что культивированные клетки РПЭ 2-ого пассажа обладают выраженными потенциями, они могут редифференцироваться (возвращаться в дифференцированное состояние РПЭ) и трансдифференцироваться в клетки нейральной сетчатки.
Таким образом, колонизирование бесклеточного матрикса сетчатки жизнеспособными аутологичными культивированными клетками РПЭ, показавшими специфическую миграцию и дифференцировку, свидетельствует, что осуществлено моделирование тканевой структуры сетчатки глаза человека.
Пример 2.
Глазные яблоки взрослого человека (м, 63 года) освобождали от окружающих тканей и промывали один раз в 70%-ном спирте и 3 раза в холодном растворе Хэнкса с антибиотиками в стандартной концентрации - пенициллин 100 Ед./мл и стрептомицин 100 мкг/мл. Передний сегмент глазных яблок удаляли круговым разрезом позади зубчатой линии, которая хорошо определяется при обычном освещении, как граница между темным и светлым участками. Стекловидное тело и нейральную сетчатку отделяли от РПЭ и выделяли, подрезая нейральную сетчатку в области зрительного нерва.
Сетчатки глазных яблок после извлечения были целиком помещены в 10 мМ ЭДТА (в среде DMEM) объемом 10 мл на 20 час при температуре 4°C. Затем сетчатки были обработаны 3% раствором Тритона Х-100 в течение 2 час при температуре 4°C, 4 раза отмыты дистиллированной водой. Далее они были последовательно помещены в 0,025% раствор ДНазы I в 1 М растворе NaCl на 1 час при температуре 4°C, затем перенесены в 4% дезоксихолат натрия на 1 час при температуре 22°C и 3 раза отмыты дистиллированной водой для удаления детергента и клеточного дебриса. Все растворы были предварительно стерилизованы через 0,22 мкм фильтр. Далее они хранились в течение 2 суток при температуре 4°C в среде DMEM/F12 с антибиотиками пенициллином-стрептомицином в стандартной концентрации до фиксации 4% параформальдегидом одной сетчатки и колонизирования второй сетчатки.
Фиксированную сетчатку промывали в фосфатно-солевом буфере (PBS, pH 7.4), проводили по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт и заливали в парафин. Из залитого в парафин блока изготавливали на микротоме гистологические срезы толщиной 5 мкм, которые монтировали на предметные стекла и окрашивали гематоксилином и эозином. Данная сетчатка использовалась для контроля адекватности девитализирующей обработки (фиг.4А).
После обработки сетчатки были исследованы в фазовом контрасте с помощью инвертированного микроскопа Olympus СКХ31 (Япония) и сфотографированы.
Как показало исследование с помощью фазово-контрастной микроскопии, сетчатки после последовательной обработки представляли собой бесклеточный каркас, состоящий из базальных мембран капилляров, волокон внеклеточного матрикса, компонентов внутренней пограничной мембраны сетчатки (коллагеновых фибрилл и базальных мембран Мюллеровских клеток) и вплетенных в нее фибрилл задней гиалоидной мембраны стекловидного тела, что свидетельствует об адекватности девитализирующей обработки.
Две глазные чаши после извлечения нейральных сетчаток наполняли холодным раствором Хэнкса с ЭДТА и спустя 30 мин начинали процесс сбора клеток нативного РПЭ. В течение всей процедуры глазные чаши находились на льду в растворе Хэнкса.
Контролируя процесс под бинокулярной лупой, наконечником 200 мкл проводили по слою и одновременно засасывали клетки, отделившиеся от мембраны Бруха.
Клетки нативного РПЭ, собранные из одного глазного яблока, переносили в стерильные пробирки, убирали надосадочную жидкость и заливали лизирующим раствором TRI®-Reagent (Sigma, США) из расчета 1 мл раствора TRI®-Reagent на 50 мг ткани, пипетировали до гомогенного состояния раствора, лизат клеток инкубировали 7 мин при температуре 22°C, переносили в стерильные пробирки и хранили при температуре минус 20°C до проведения ПЦР оценки дифференцировки клеток.
В клетках нативного РПЭ при проведении ПЦР оценки выявлялась экспрессия на мажорном уровне гена RPE65, являющегося маркером клеток РПЭ. Кроме того, в нативном РПЭ наблюдалась экспрессия на минорном уровне гена плюрипотеного статуса Oct4 (POU5F1) и маркеров нейральных стволовых клеток Musashi 1 и РахG, что позволяет предположить наличие в нативной ткани немногочисленной популяции клеток со свойствами нейральных стволовых-прогениторных клеток.
Клетки нативного РПЭ, собранные из второго глазного яблока, переносили в стерильную среду для выделения следующего состава: питательная среда DMEM/F12 (1:1), дополненная 15% эмбриональной телячьей сыворотки, 2 мМ L-глутамина и антибиотиками пенициллина-стрептомицина в стандартной концентрации, и центрифугировали в течение 7 мин при 600 об/мин. Клетки РПЭ ресуспендировали в полной культуральной среде, следующего состава: питательная среда DMEM/F12 (1:1), дополненная 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2 (Gibco, Invitrogen, США), 20 нг/мл основного фактора роста фибробластов (оФРФ) и антибиотиками пенициллина-стрептомицина в стандартной концентрации, затем после мягкого пипетирования равномерно распределяли в стерильных пластиковых флаконах площадью 25 см2 без покрытия (Greiner, Германия) и культивировали при температуре 37°C и 5% CO2. Спустя 5-7 суток проводили первую смену среды и продолжали культивировать со сменой среды через 2-3 суток до достижения клетками конфлюэнтного монослоя и образования первичной культуры.
Затем проводили пассирование конфлюэнтной первичной культуры, используя раствор трипсина - версена (1:3). Клетки культивировали в плотности 1 - 2×104 кл./см2 в полной культуральной среде, следующего состава: питательная среда DMEM/F12 (1:1), дополненная 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2 (Gibco, Invitrogen, США), 20 нг/мл основного фактора роста фибробластов (оФРФ) и антибиотиками пенициллина-стрептомицина в стандартной концентрации. Культивирование проводили во флаконах и в 2- или 4-луночных слайд-камерах для культур клеток (BD Falcon™, BD Biosciences, США).
Клетки РПЭ на слайд-камерах через 2-3 суток при достижении ими субконфлюэнтного слоя фиксировали охлажденным ацетоном. Далее предметные стекла с клетками освобождали от разделительных решеток, предметные стекла высушивали при температуре 20°C, заворачивали в фольгу и хранили в низкотемпературном холодильнике при температуре минус 70-80°C до проведения ИГХ оценки полученных культур.
Клеточные культуры при ИГХ оценке были положительны на пронейральные (Рах6), нейральные (Nestin, β-tubulin III, nNOS, TH, NF 70 and 200kd, N-cadherin, Recoverin) и глиальные (виментин, GFAP, CNPase) маркеры, а также маркер эпителиальных клеток PanCK. ИГХ оценку и фотографирование проводили с помощью светового микроскопа Olympus АН3 (Япония). Поскольку такие клетки, которые могут контаминировать клеточные культуры РПЭ из подлежащей хориоидеи, как фибробласты, меланоциты, эндотелиальные клетки и перициты, являются цитокератин-негативны, то наличие окрашивания на PanCK подтверждало получение клеточных культур из РПЭ.
Окрашивание на пронейральные, нейральные и глиальные маркеры демонстрирует, что клетки РПЭ человека в условиях культивирования были коммитированы в нейральном направлении.
Клетки РПЭ во флаконах росли в виде адгезивных монослойных культур. Наблюдение за клеточными культурами РПЭ проводили с помощью инвертированного микроскопа Olympus СКХ31 (Япония).
С культуральных флаконов клетки первичной культуры и культуры клеток РПЭ 1-5 пассажа использовали для проведения ПЦР оценки. Для этого монослой клеток промывали 1 раз DPBS (ПанЭко, Россия), заливали лизирующим раствором TRI®-Reagent (Sigma, США) из расчета 1 мл на 25 см2 культурального флакона, клеточный лизат пипетировали до гомогенного прозрачного состояния раствора, гомогенат инкубировали 5 мин при температуре 22°C, а затем переносили в стерильные пробирки и хранили при температуре минус 20°C до проведения выделения тотальной РНК и синтеза кДНК.
ПЦР оценка культивируемых клеток РПЭ взрослого человека по сравнению с нативным РПЭ показала резкое падение уровня экспрессии гена RPE65, маркера клеток РПЭ, участвующего в цикле фототрансдукции при взаимодействии их с фоторецепторами, что свидетельствует о начале дедифференцировки. В клетках первичной и пассируемых культур одновременно детектировалась экспрессия генов-маркеров стволовых-прогениторных клеток: Oct4 (POU5F1), Nanog, Musashi 1 и Рах6. Наличие экспрессии нейральных маркеров β tubulin III, Рах6 и Musashi 1 свидетельствовало о нейральном потенциале клеточных культур РПЭ.
Результаты проведенной ИГХ оценки и молекулярно-генетического анализа показали, что в условиях культивирования клетки РПЭ глаза взрослого человека претерпевают сильные изменения, они способны к дедифференцировке и развитию по нейральному пути, т.е. комитируются в направлении ретинальных нейральных клеток.
Далее проводили колонизирование бесклеточного каркаса сетчатки, хранившегося в течение 2 суток при температуре 4°C в среде DMEM/F12, суспензией аллогенных клеток РПЭ 5-го пассажа в плотности 4×104 кл./см2. В качестве аллогенных клеток РПЭ для колонизирования бесклеточного каркаса сетчатки использовали клеточные культуры РПЭ 5-го пассажа, полученные из глазного яблока взрослого человека (ж, 50 лет). Способ получения нативных клеток РПЭ, способ получения первичной культуры и культур клеток РПЭ 1-5 пассажа, коммитирования клеток РПЭ в нейральном направлении и снятия их с культуральной поверхности раствором трипсина-версена (1:3) приведены в примере 1.
Перед нанесением культивированных клеток РПЭ бесклеточный каркас сетчатки был промыт 7 раз в питательной среде DMEM/F12 (1:1) с антибиотиками пенициллином-стрептомицином в стандартной концентрации и перенесен в стерильную 1-луночную слайд-камеру (BD Falcon™, BD Biosciences, США), залит 2 мл полной культуральной среды, состоящей из питательной среды DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина и антибиотиками, и помещен в CO2-инкубатор, имеющий температуру 37°C, 5% CO2 в воздухе, для уравновешивания pH среды.
На бесклеточный каркас сетчатки человека высаживали посредством диффузионного осаждения суспензию аллогенных клеток РПЭ 5-го пассажа в количестве 350 тыс (плотностью 4×104 кл./см2) в 2 мл полной культуральной среды того же состава, в котором находился бесклеточный каркас. Клетки культивировали при температуре 37°C и 5% CO2, со сменой среды через 2-3 суток.
На 2-е сутки при исследовании с помощью фазово-контрастного микроскопа отмечалось разделение клеток на 2 субпопуляции: одни клетки распластывались на поверхности культурального пластика, а другие прикреплялись к поверхности бесклеточного матрикса, но оставались округлой формы.
На 14-е сутки на поверхности бесклеточного матрикса определялись округлые клетки.
Фиксацию колонизированного культивированными клетками бесклеточного каркаса сетчатки 4% параформальдегидом проводили через 25 суток. Фиксированные препараты промывали в фосфатно-солевом буфере (PBS, pH 7.4), проводили по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт и заливали в парафин.
Из залитых в парафин блоков изготавливали на микротоме гистологические срезы толщиной 5 мкм, которые монтировали на предметные стекла, покрытые адгезивом.
Изучение локализации культивируемых клеток РПЭ в бесклеточной сетчатки проводили на серийных парафиновых срезах, окрашенных гематоксилином и эозином, с помощью светового микроскопа Olympus АН3 (Япония). Клетки РПЭ образовывали конгломераты округлой формы и на поверхности бесклеточного матрикса сетчатки и на поверхности стекловидного тела (в местах отсутствия сетчатки).
Оценку дифференцировки культивируемых клеток РПЭ, заселивших бесклеточную сетчатку, проводили методом иммунопероксидазного окрашивания, используя готовые к использованию мышиные моноклональные антитела к CD31, CD34, тирозиназе, СК7, СК8, СК18, СК19, виментину, GFAP, S100 и NSE (Dako, Дания).
Клетки РПЭ, заселившие сетчатку, были негативны на маркеры эндотелиальных клеток (CD31, CD34) и маркер меланоцитов (тирозиназу). Позитивное окрашивание отмечалось на маркеры дифференцированных (СК8 и СК18) и культивированных (СК7 и СК19) клеток РПЭ, а также на глиальные (виментин, S100 и GFAP) и нейрональный (NSE) маркеры. GFAP-позитивное окрашивание определялось в виде сети в округлых образованиях.
Как видно из примера, бесклеточный каркас сетчатки через 2 суток после девитализирующей обработки успешно колонизировался аллогенными культивированными клетками РПЭ. В присутствии 3D-матрикса аллогенные клетки РПЭ 5-го пассажа колонизировали бесклеточный каркас сетчатки, образуя конгломераты округлой формы и на поверхности бесклеточного матрикса сетчатки и на поверхности стекловидного тела (в местах отсутствия сетчатки).
Культивированные клетки РПЭ, иммобилизованные на носителе из бесклеточного матрикса сетчатки, сохраняют свою жизнеспособность, пролиферативную и миграционную активность и заданное направление гистоспецифической дифференцировки в течение минимум 25 суток.
Таким образом, колонизирование бесклеточного каркаса сетчатки жизнеспособными аллогенными культивированными клетками РПЭ, показавшими специфическую миграцию и дифференцировку, свидетельствует, что осуществлено моделирование тканевой структуры сетчатки глаза человека.
Предлагаемое изобретение позволяет создать способ моделирования тканевой структуры и/или репаративной функции сетчатки путем создания ex vivo не имеющей мировых аналогов трехмерной модели, состоящей из бесклеточного матрикса сетчатки человека, биологически инертного, не ухудшающего рост и дифференцировку клеточной культуры ретинального пигментного эпителия, с заданными параметрами количества, плотности и тканевой дифференцировки.
Изобретение может быть использовано для изучения дифференцировки культивированных клеток ретинального пигментного эпителия, для исследования действия на пигментный эпителий при добавлении в среду культивирования разных факторов, моделирования процессов, происходящих в пигментном эпителии при повреждении и патологических состояниях сетчатки, а также изучения регенерационных ответов клеток пигментного эпителия сетчатки.
Изобретение позволит определить направление дальнейших исследований, которые в будущем помогут создать жизнеспособный заместитель сетчатки глаза человека и тканевые аналоги нервной ткани для замещения пораженных тканей сетчатки, позволяющих лечить и/или предотвращать глазные болезни, такие как возрастная макулярная дегенерация как влажная, так и сухая, глаукома, диабетическая ретинопатия, включая диабетический отек желтого пятна, хориоидальная неоваскулярная мембрана, увеит, миопическая дегенерация, глазные опухоли, окклюзия центральной вены сетчатки, покраснение радужки, неоваскуляризация глаз, центральная серозная ретинопатия, болезни поверхности глаза, такие как синдром сухого глаза, окклюзия центральной артерии сетчатки, кистозный макулярный отек и любое другое дегенеративное заболевание глаз.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ДЕДИФФЕРЕНЦИРОВАННЫХ КЛЕТОК РЕТИНАЛЬНОГО ПИГМЕНТНОГО ЭПИТЕЛИЯ ГЛАЗА ВЗРОСЛОГО ЧЕЛОВЕКА | 2009 |
|
RU2409663C1 |
| СПОСОБ ВЫДЕЛЕНИЯ И ФИКСАЦИИ КЛЕТОК РЕТИНАЛЬНОГО ПИГМЕНТНОГО ЭПИТЕЛИЯ ТРУПНЫХ ГЛАЗ ЧЕЛОВЕКА | 2011 |
|
RU2464783C1 |
| СПОСОБ ПОЛУЧЕНИЯ КУЛЬТУРЫ КЛЕТОК РЕТИНАЛЬНОГО ПИГМЕНТНОГО ЭПИТЕЛИЯ ИЗ ГЛАЗА ВЗРОСЛОГО ДОНОРА-ТРУПА | 2014 |
|
RU2569481C1 |
| Способ получения образца фрагмента ретинального пигментного эпителия сетчатки | 2024 |
|
RU2836977C1 |
| Способ получения фрагмента нативного пигментного эпителия сетчатки в виде универсального и стандартизированного образца для дальнейшего изучения РПЭ глаза взрослого донора-трупа | 2024 |
|
RU2835179C1 |
| СПОСОБ ПОЛУЧЕНИЯ ОРГАННОЙ КУЛЬТУРЫ СОБСТВЕННО СОСУДИСТОЙ ОБОЛОЧКИ ИЗ ГЛАЗА ВЗРОСЛОГО ДОНОРА-ТРУПА | 2014 |
|
RU2576573C1 |
| СПОСОБ ПОЛУЧЕНИЯ ОРГАННОЙ КУЛЬТУРЫ ХОРОИДАЛЬНО-ПИГМЕНТНОГО КОМПЛЕКСА ИЗ ГЛАЗА ВЗРОСЛОГО ДОНОРА-ТРУПА | 2014 |
|
RU2578526C1 |
| СПОСОБ УЛУЧШЕНИЯ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ СЕТЧАТКИ ПРИ ЕЕ ПАТОЛОГИИ РАЗЛИЧНОГО ГЕНЕЗА | 2004 |
|
RU2279886C2 |
| Способ субретинальной трансплантации клеток ретинального пигментного эпителия (РПЭ), дифференцированных из индуцированных плюрипотентных стволовых клеток человека, при атрофии ретинального пигментного эпителия в эксперименте | 2019 |
|
RU2729937C1 |
| СПОСОБЫ ВЫДЕЛЕНИЯ И КУЛЬТИВИРОВАНИЯ ЧЕЛОВЕЧЕСКИХ РЕТИНАЛЬНЫХ КЛЕТОК-ПРЕДШЕСТВЕННИКОВ | 2019 |
|
RU2809003C2 |
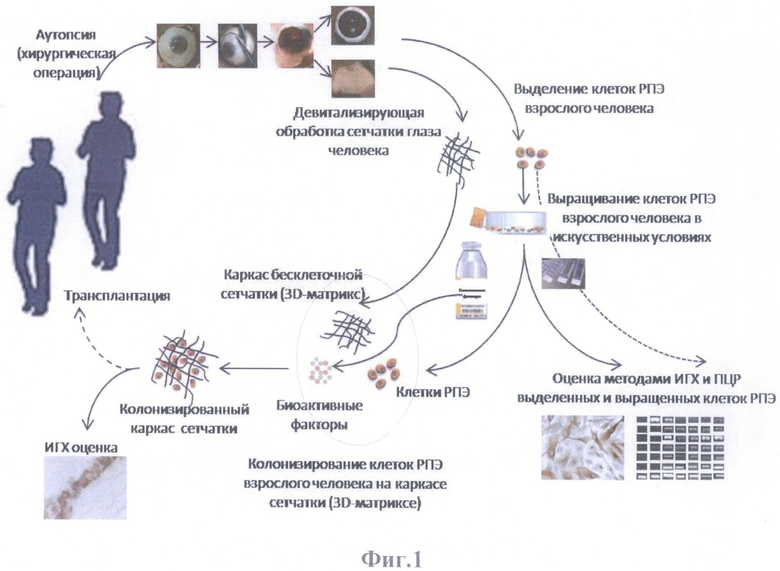
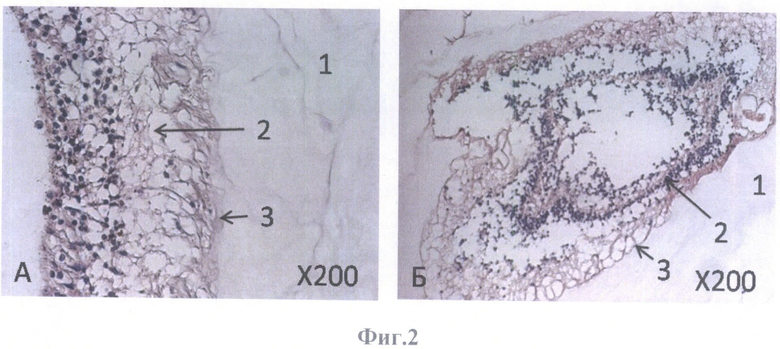

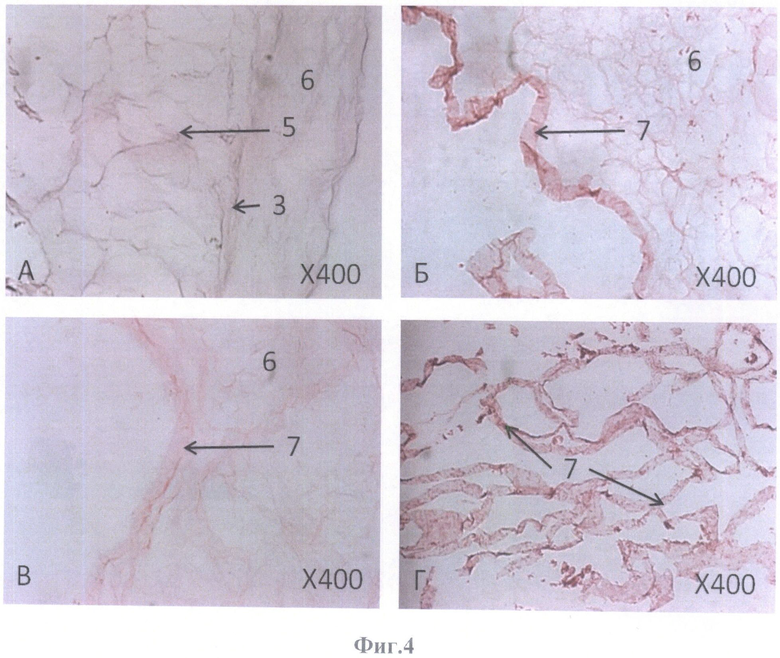
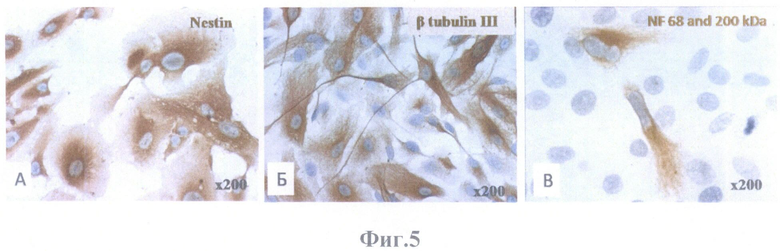

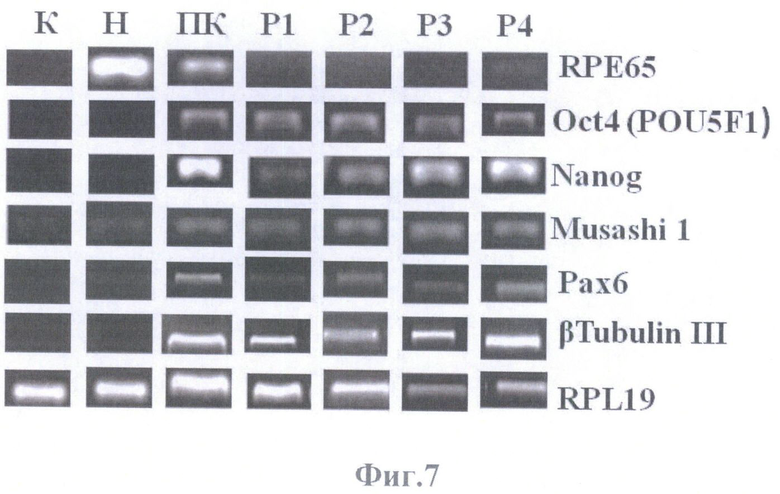
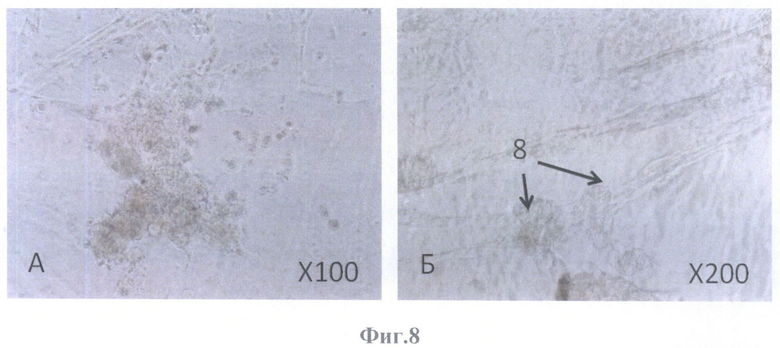
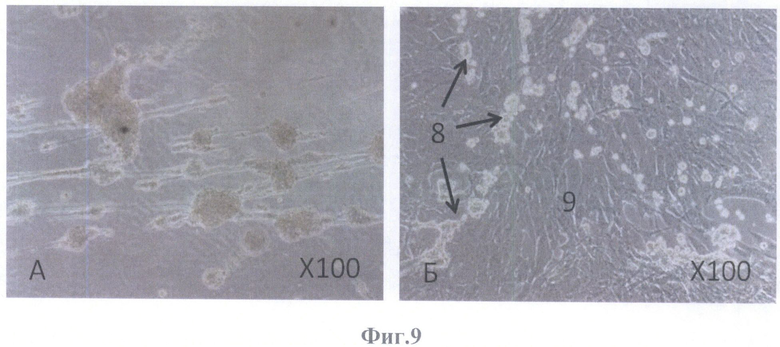
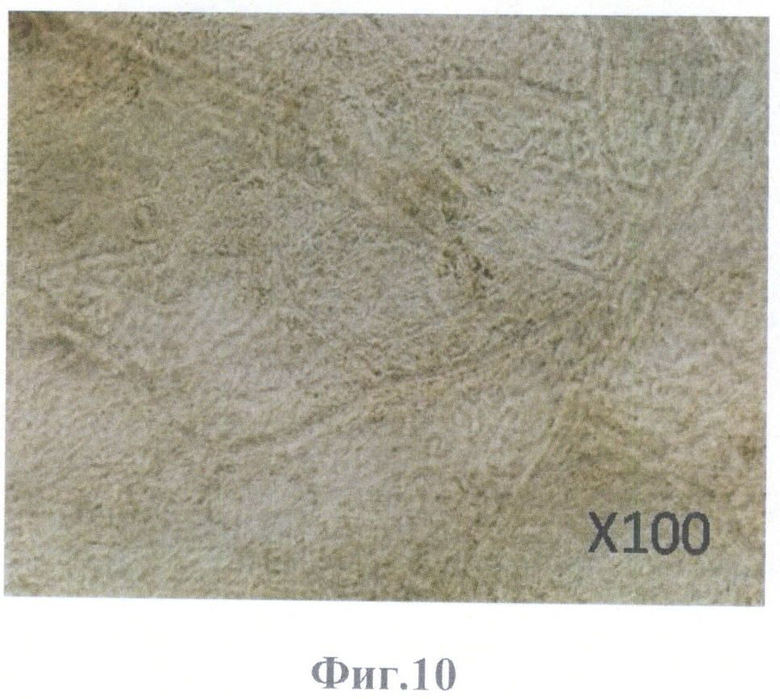

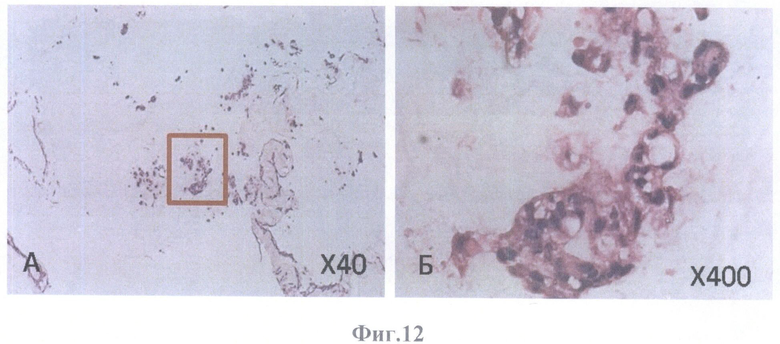
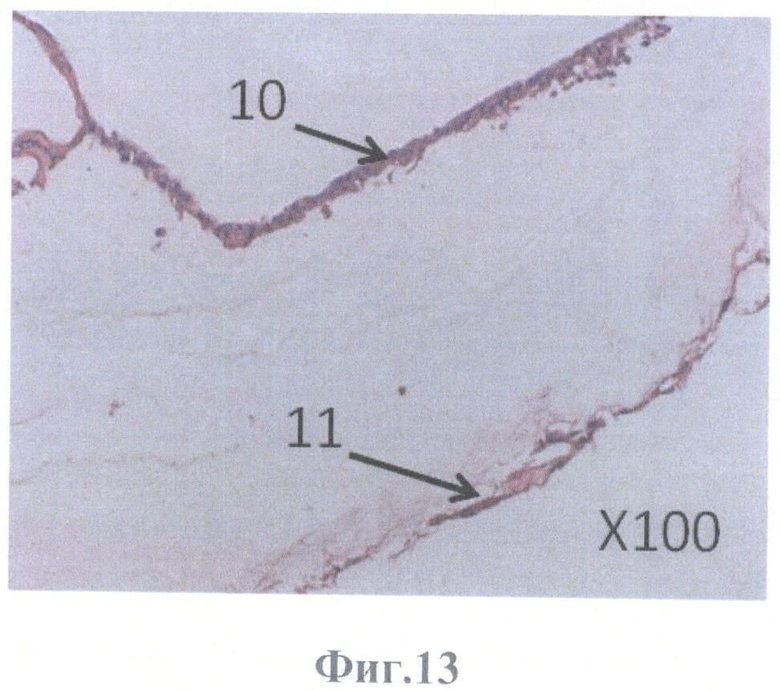
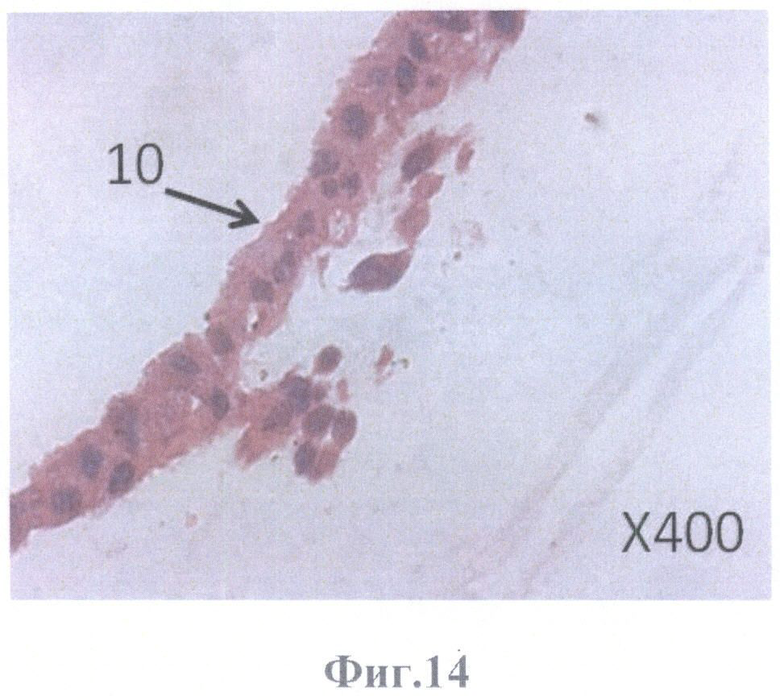

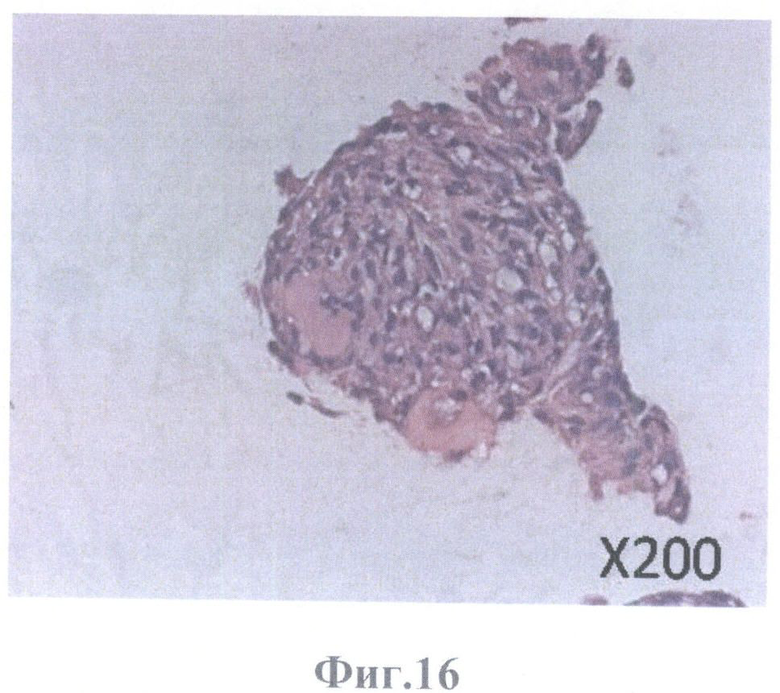
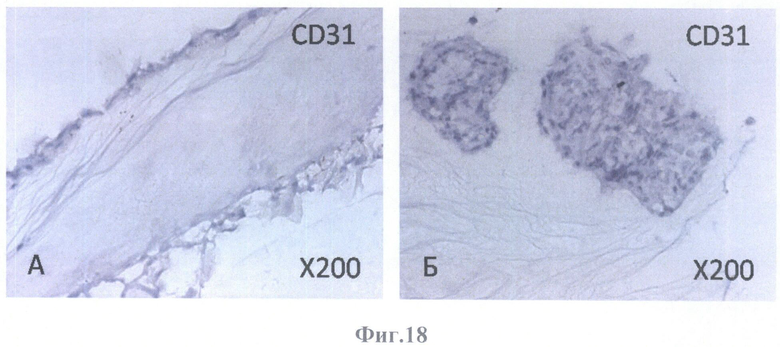
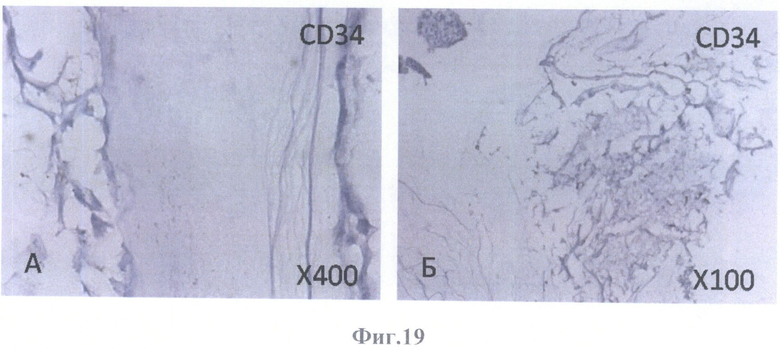
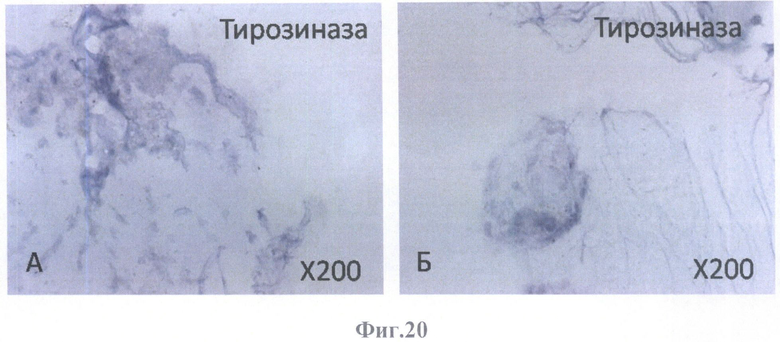
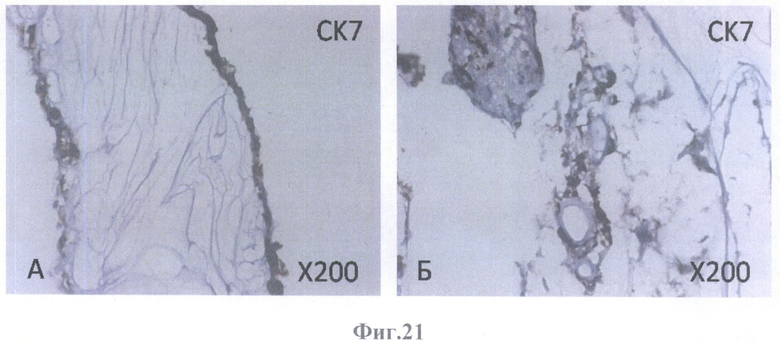
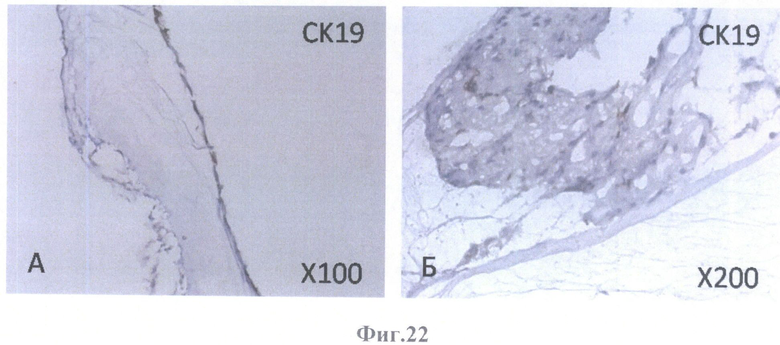
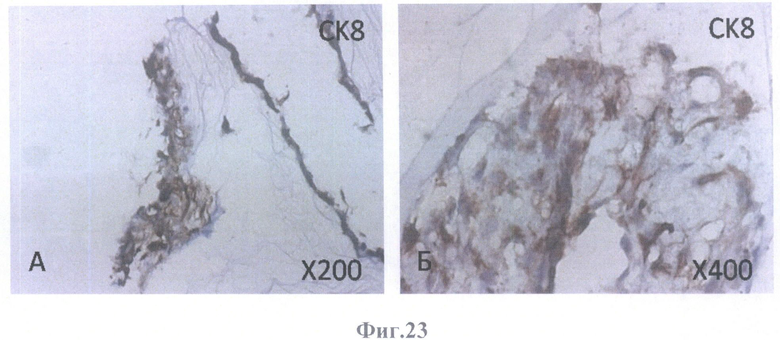

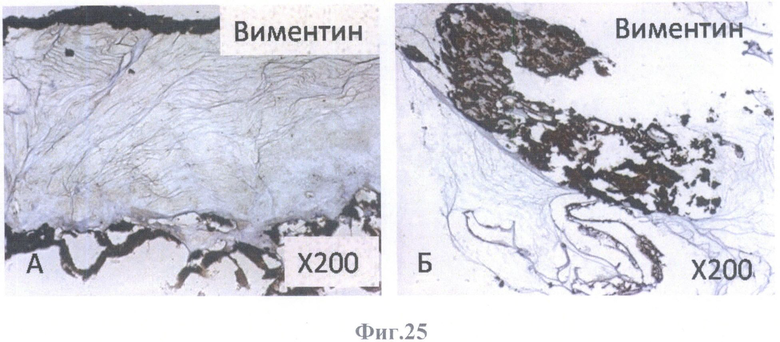

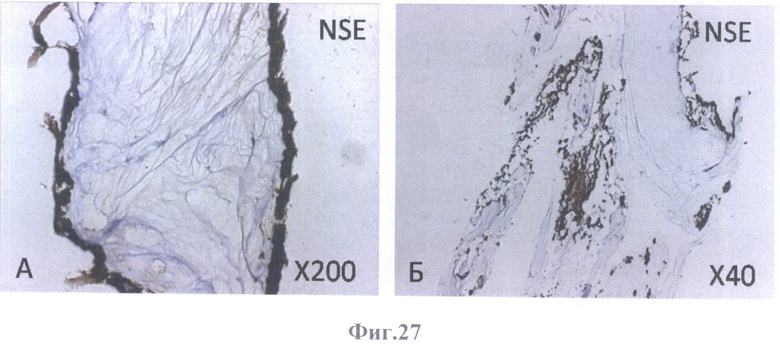

Изобретение относится к биотехнологии и медицине, и может быть использовано в тканевой инженерии и клеточной трансплантологии с целью моделирования тканевой структуры сетчатки. Для этого энуклиируют глазное яблоко при аутопсии, промывают его в спиртовом растворе и в растворе Хэнкса с антибиотиками. Затем удаляют по зубчатой линии передний сегмент глазного яблока. Отделяют стекловидное тело и нейтральную сетчатку от ретинального пигментного эпителия (РПЭ). Выделенную сетчатку подвергают девитализирующей обработке для создания бесклеточного каркаса сетчатки глаза. Этот каркас промывают в питательной среде с антибиотиками. Затем каркас переносят в слайд-камеру и колонизируют клетками РПЭ. Для получения культуры клеток РПЭ глазную чашу глазного яблока заполняют раствором Хэнкса с ЭДТА. Затем собирают клетки нативного РПЭ. Часть клеток РПЭ переносят в стерильную среду, для выделения, центрифугируют. Затем выделенные клетки ресуспендируют в ростовой среде, пипетируют, распределяют во флаконы и культивируют со сменой среды до образования первичной культуры. Далее проводят пассирование клеток первичной культуры. Затем эти клетки культивируют в культуральных флаконах. Культивированные клетки 1-5 пассажа в виде монослойных культур снимают с культуральной поверхности и в виде суспензии культивированных клеток РПЭ направляют на колонизирование бесклеточного каркаса сетчатки. При этом, наблюдение за сетчаткой и контроль девитализирующей обработки осуществляли методом микроскопии. Для наблюдения за клеточными культурами проводят ИГХ опенку, ПЦР опенку, микроскопию. Способ обеспечивает создание трехмерной модели культуры клеток РПЭ с заданными параметрами количества, плотности и тканевой дифференцировки, благодаря поэтапному контролированию процесса моделирования. 13 з.п. ф-лы, 2 пр., 3 табл., 28 ил.
1. Способ моделирования тканевой структуры сетчатки глаза человека путем колонизирования бесклеточного каркаса сетчатки глаза клетками ретинального пигментного эпителия, заключающийся в том, что бесклеточный каркас сетчатки глаза получают девитализирующей обработкой сетчатки из энуклиированного при аутопсии глазного яблока путем последовательного промывания его в спиртовом растворе при концентрации 70% и в холодном растворе Хэнкса с антибиотиками, удаления по зубчатой линии переднего сегмента глазного яблока, отделения стекловидного тела и нейтральной сетчатки от ретинального пигментного эпителия и выделения сетчатки, подрезая ее в области зрительного нерва, выделенную сетчатку для девитализирующсй обработки помещают в 10-15 мл 10 мМ ЭДТЛ в среде DMEM на 20 ч при температуре 4°С, затем обрабатывают 3%-ным раствором Тритона Х-100 в течение 2 ч при температуре 4°С и отмывают в дистиллированной воде, далее сетчатку последовательно помещают в 0,025%-ный раствор ДНазы 1 в 1 М растворе NaCl на 1 ч при температуре 4°С и в 4%-ный дезоксихолат натрия на 1-2 ч при температуре 22°С и затем отмывают в дистиллированной воде для удаления детергента и клеточного дебриса, глазную чащу наполняют холодным раствором Хэнкса с ЭДТА и через 15-30 мин. собирают клетки нативного ретинального пигментного эпителия, часть собранных клеток переносят в лизирующий раствор TRI®-Reagent из расчета 1 мл раствора на 50-100 мг ткани, пипетируют до гомогенного состояния раствора и лизат клеток нативного ретинального пигментного эпителия инкубируют 5-7 мин при температуре 22°С, другую часть собранных клеток переносят в стерильную среду для выделения, состоящую из питательной среды DMEM/F12 (1:1), дополненной 15%-ной эмбриональной телячьей сывороткой, 2 мМ L-глутамина и антибиотиками и центрифугируют в течение 5-7 мин при 600-800 об/мин, затем клетки ресуснендируют в полной культуральной среде, состоящей из питательной среды DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ного раствора заменимых аминокислот, 1%-ного раствора нейральной добавки N2, 20 нг/мл основного фактора роста фибробластов и антибиотиков, и после пипетирования распределяют в культуральпых флаконах без покрытия и культивируют при температуре 37°С и 5% СО2, через 5-7 суток проводят первую смену среды и продолжают культивировать со сменой среды через 2-3 суток до достижения клетками конфлюэнтного монослоя и образования первичной культуры, затем проводят пассирование клеток первичной культуры, используя раствор трипсина - версена (1:3), далее клетки помещают в культуральные флаконы и в слайд-камеры в плотности 1 - 2·104 кл./см2 и культивируют, культивированные в культуральных флаконах клетки 1-5 пассажа в виде адгезивных монослойных культур, коммитированные в нейральном направлении, снимают с культуральной поверхности раствором трипсина-версена (1:3) и в виде суспензии культивированных клеток ретинального пигментного эпителия направляют на колонизирование бесклеточного каркаса сетчатки, причем бесклеточный каркас сетчатки перед колонизированием промывают в питательной культуральной среде DMEM/F12 (1:1) с антибиотиками, переносят в слайд-камеру и заливают полной культуральной средой, а колонизирование бесклеточного каркаса сетчатки глаза осуществляют посредством диффузионного осаждения суспензии культивированных клеток ретинального пигментного эпителия в плотности 4-5,5·104 кл./см2 в полной культуральной среде того же состава и культивирования при температуре 37°С и 5% СО2, со сменой среды через 2-3 суток до колонизирования культивированными клетками бесклеточного каркаса сетчатки и формирования монослоя клеток на его поверхности с получением тканевой структуры сетчатки глаза, которую направляют на изучение локализации культивируемых клеток и на ИГХ оценку потенциала дифференцировки культивируемых клеток в бесклеточном каркасе сетчатки глаза.
2. Способ по п.1, отличающийся тем, что для изучения локализации культивируемых клеток проводят окрашивание ядер клеток витальным красителем Hoechst 33342 для люминесцентной микроскопии.
3. Способ по п.1, отличающийся тем, что для изучения локализации культивируемых клеток колонизированный клетками бесклеточный каркас сетчатки через 12-25 суток фиксируют 4%-ным параформальдегидом, промывают его в фосфатно-солевом буфере (PBS, pH 7.4), проводят по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт, заливают в парафин, изготавливают гистологические срезы и окрашивают гематоксилином и эозином для световой микроскопии.
4. Способ по п.1, отличающийся тем, что ИГХ оценку потенциала дифференцировки культивированных клеток в бесклеточном каркасе сетчатки проводят методом иммунопероксидазного окрашивания, используя мышиные моноклональные антитела к CD31, CD34, тирозиназе, СК7, CK8, СК18, СК19, виментину, GFAP, SIOO и NSH.
5. Способ по п.1, отличающийся тем, что используют антибиотики пенициллин-стрептомицин в стандартной концентрации - пенициллин 100 Ед./мл и стрептомицин 100 мкг/мл.
6. Способ по п.1, отличающийся тем, что бссклеточный каркас сетчатки глаза после удаления детергента и клеточного дебриса помещают в питательную культуральную среду DMEM/F12 с антибиотиками при температуре 4-8°С на хранение в течение 2-20 суток до фиксации 4% параформальдегидом или колонизирования культивированными клетками.
7. Способ по п.6, отличающийся тем, что фиксированный бесклеточный каркас сетчатки промывают в фосфатно-солевом буфере (PBS, pH 7.4), проводят по спиртам с восходящей концентрацией 70, 96 и 100%-ный спирт, заливают и парафин, изготавливают гистологические срезы, которые монтируют на предметные стекла и окрашивают гематоксилином и эозином для контроля адекватности девитализирующей обработки с помощью световой и/или люминесцентной микроскопии.
8. Способ по п.1, отличающийся тем, что после инкубирования лизат клеток нативного ратинального пигментного эпителия хранят при температуре минус 20°С до проведения ПЦР оценки.
9. Способ по п.1, отличающийся тем, что клетки первичной культуры и культивированные клетки 1-5 пассажа направляют для проведения ПЦР оценки.
10. Способ по п.9, отличающийся тем, что для проведения ПЦР оценки определяют уровень экспрессии генов, кодирующих транскрипционные факторы Oct4 (POU5F1), Nanog, Рах6 и маркеры дифференцировки Musashi 1, β tubulin III, RPE65 выделением тотальной РНК и синтеза кДНК, при этом уровень экспрессии генов оценивают но интенсивности свечения полос, полученных при электрофоретическом разделении ПЦР-продуктов в 1%-ном агарозном геле на анализаторе гелей.
11. Способ по п.1, отличающийся тем, что культивирование клеток в слайд-камерах проводят в 1-, 2-, 4- или 8-луночных слайд-камерах, через 2-3 суток, при достижении клетками субконфлюэнтного слоя, их фиксируют охлажденным ацетоном и направляют на ИГХ оценку потенциала дифференцировки клеток.
12. Способ по п.11, отличающийся тем, что ИГХ оценку потенциала дифференцировки культивированных клеток осуществляют методами иммунопероксидазного и иммунофлуоресцентного окрашивания, используя первичные мышиные моноклональные и кроличьи поликлональные антитела, Рах6, Sox2, вимннтин, Nestin, β tubulin III, nNOS, тирозин гидроксилаза, NF 68 and 200 кДа, GFAP, CNPasc, Iba, N-cadherin, Коннексин-43, RPE65, CRALBP, Rccoverin, PanCK, Macrophage antigen, Фибронектин, Factor Von Willcbrand, Ki67, а также полимер из набора EnVision™, конъюгированный с пероксидазой хрена, (Dako, Дания) или вторичные антитела: набор SFX с антителами козы против мыши, меченные Alexa Fluor® 488, антитела против иммуноглобулинов кролика, коньюгированные с флуорохромом Texas Red, разведенные в 0,1%-ном растворе бычьего сывороточного альбумина.
13. Способ по п.1, отличающийся тем, что бесклеточный каркас сетчатки глаза, перенесенный в слайд-камеру, заливают полной культуральной средой, состоящей из питательной среды DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина, 1%-ным раствором заменимых аминокислот, 1%-ным раствором нейральной добавки N2, 20 нг/мл основного фактора роста фибробластов и антибиотиками.
14. Способ по п.1, отличающийся тем, что бесклеточный каркас сетчатки глаза, перенесенный в слайд-камеру, заливают полной культуральной средой, состоящей из питательной среды DMEM/F12 (1:1), дополненной 10% эмбриональной телячьей сыворотки, 2 мМ L-глутамина и антибиотиками и помещают в СО2-инкубатор, имеющий температуру 37°С, 5% СО2 в воздухе для уравновешивания pH среды.
| СПОСОБ ПОЛУЧЕНИЯ ДЕДИФФЕРЕНЦИРОВАННЫХ КЛЕТОК РЕТИНАЛЬНОГО ПИГМЕНТНОГО ЭПИТЕЛИЯ ГЛАЗА ВЗРОСЛОГО ЧЕЛОВЕКА | 2009 |
|
RU2409663C1 |
| WO 2008129554 A1, 30.10.2008 | |||
| ТАХЧИДИ X.П | |||
| и др | |||
| Сравнительная характеристика влияния фактора пигментного эпителия (PEDF) и авастина на органотипические культуры сетчатки | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| SHOZO SONODA et al | |||
| A protocol for the culture and diflerentiation of highly polarized human retinal pigment | |||

