Настоящее изобретение относится к мономерному домену VHH, полученному из анти-VP6-антител верблюдовых, димерному домену, способу иммунизации, способу выявления ротавируса, композициям, способам профилактики и лечения ротавирусных инфекций. Более конкретно изобретение относится к мономерному домену (VHH), полученному из антител верблюдовых, при этом указанный домен может иметь любые аминокислотные последовательности, показанные в SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3 или SEQ ID No. 4, и при этом указанные домены связываются с белком VP6 ротавируса группы A.
УРОВЕНЬ ТЕХНИКИ
Ротавирус группы A (RV) является главной причиной тяжелой диареи у детей и у молодняка многих видов животных, представляющих хозяйственный интерес (коров, свиней, лошадей, южноамериканских верблюдовых и т.д.). В качестве проблемы здравоохранения RV является третьей наиболее распространенной причиной смерти, ассоциированной с тяжелой диареей, в развивающихся странах (2 миллиона смертей в год). С другой стороны, RV-индуцированная диарея у животных, предназначенных для употребления в пищу, например у молодых телят, приводит к высоким затратам, связанным с профилактикой или лечением.
RV группы A представляют собой частицы, состоящие из тройного белкового капсида. Наружная поверхность капсида состоит из белков VP4 и VP7, которые являются высоковариабельными антигенами; до настоящего времени описано, по меньшей мере, 27 вариантов VP4 (P-типы) и 16 вариантов VP7 (G-типы). Каждое сочетание G-P-типов индуцирует образование нейтрализующих антител, которые имеют низкую перекрестную реактивность с другими G-P-типами; по этой причине в вакцины необходимо включать разные штаммы, которые циркулируют в организме вида-мишени.
Промежуточный капсид состоит из тримерного белка VP6, масса которого составляет 51% от массы вириона. В зависимости от наличия или отсутствия двух разных эпитопов в белке VP6 (узнаваемых моноклональными антителами мАт 255/60 и 631/9), штаммы RV группы A дополнительно делят на подгруппы (Sb) I, II, I+II и подгруппа, в которой нет ни I, ни II. RV человека обычно относятся к подгруппе Sb II, тогда как RV животных главным образом относятся к Sb I. Белок VP6 является высокоиммуногенным; у инфицированных естественным образом людей и животных развивается сильный гуморальный ответ, направленный против эпитопов VP6. Независимо от указанных выше подгрупп VP6 является высококонсервативным белком среди всех RV группы A (гомология аминокислотной последовательности >90%), и общие антигены могут быть выявлены с помощью поликлональных антисывороток с широкой реакционной способностью или с помощью моноклональных антител. Поэтому VP6 является антигеном-мишенью в большинстве иммунодиагностических тестов, предназначенных для выявления RV группы A. Антитела, направленные против указанного белка, не обладают нейтрализующей активностью in vitro. Однако моноклональные IgA-антитела могут блокировать репликацию вирусов у мышей внутриклеточно.
В настоящее время профилактика RV-индуцированной диареи у животных основана на пассивной иммунизации, тогда как для людей используют активную иммунизацию. У животных парентеральные вакцины на основе инактивированных вирусов применяют для беременных самок, чтобы защитить новорожденных посредством переноса материнских антител через молозиво и молоко. Указанная методика является высокоэффективной для профилактики симптомов тяжелой диареи и снижения заболеваемости и смертности у пораженного болезнью поголовья скота, но не способна предотвращать RV-инфекцию, так как методика существенно не снижает количество вируса, выделяемого инфицированными животными (Parreno, V.C. et al., Vet. Immunol. Immunopathol. 100: 7-24, 2004). Только непрерывное присутствие высоких титров пассивных анти-RV-антител в просвете кишечника (продуцированных естественным путем или искусственно добавленных в молоко) полностью защищает от диареи и существенно снижает экскрецию вирусов (Fernandez, F.M. et al., Vaccine 16: 507-16, 1998; Saif, LJ. et al., Infect. Immun. 41: 1118-31, 1983, и Saif, L.J. et al., Adv. Exp. Med. Biol. 216B: 1815-23, 987).
Для детей одобрены две вакцины на основе живых вирусов, аттенуированных в результате генетической реассоциации. В случае обоих продуктов доказана высокая эффективность против тяжелой RV-индуцированной диареи. Однако, учитывая историю интуссусцепции, ассоциированной с вакциной, ранее используемой на человеке (Murphy, T. V. et al., J. Infect. Dis. 187: 1309-13, 2003), и недавнее открытие RV-виремии у естественным образом инфицированных детей (Ray, P. et al., J. Infect. Dis. 194: 588-93, 2006, и Blutt, S.E. et al., Lancet 362: 1445-9, 2003), безвредность указанных вакцин была подвергнута сомнению, особенно в случае недоношенных детей, детей с ослабленным иммунитетом и плохо питающихся детей. Поэтому необходима альтернативная дополнительная методика профилактики и лечения RV-индуцированной диареи.
В случае методики, основанной на пассивном иммунитете, как в случае грудного вскармливания, показано, что введение анти-RV-антител, очищенных из молозива коров или яиц (анти-RV IgG человека и коров и IgY из желтка куриных яиц), снижает заболевание диареей как у людей, так и у животных. Но возможность получения экономичным способом больших количеств антител, обладающих воспроизводимыми свойствами, является низкой. Поэтому необходимо создание антител, предназначенных для пассивной анти-RV-иммунизации животных и человека, в частности антител, которые могут быть получены в промышленном масштабе, которые не вызывают иммунологических реакций, которые являются достаточно маленькими, чтобы эффективно достигать эпитопов консервативных внутренних белков, и которые способны распознавать и ингибировать репликацию штаммов с разными генотипами (полиреактивные).
Домен VHH тяжелой цепи антитела верблюдовых массой 15 кД является наименьшим известным природным доменом, обладающим полной антигенсвязывающей способностью, и является идеальным доменом для создания библиотеки кодирующих ДНК для одноцепочечных фрагментов, обладающих способностью узнавать природные антигены. Кроме того, можно использовать методику иммунизации лам, чтобы обогатить библиотеку VHH, направленными против представляющего интерес антигена. Благодаря своим особым свойствам домены VHH, полученные из тяжелых цепей антител лам, являются весьма универсальными средствами для разработки диагностических реагентов и продуктов, предназначенных для профилактики или лечения RV-индуцированной диареи. Например, недавно сообщалось, что VHH, направленный против RV-штамма G-типа G3, полученный в дрожжах, проявляет нейтрализующую активность in vitro, и очищенный VHH способен снижать частоту встречаемости и продолжительность RV-индуцированной диареи у лактирующих мышей (Pant, N. et al., J. Infect. Dis. 194: 1580-8, 2006, и van der Vaart J. M. et al., Vaccine, May 8, 24(19): 4130-7, 2006). Однако указанные авторы не смогли выяснить, против какого вирусного белка направлены полученные VHH, и они предполагали, что VHH могли быть направлены против конформационных эпитопов наружных белков.
В патентном документе WO 2006/056306 описано получение и применение доменов VHH или их фрагментов для лечения инфекций, вызванных энтеропатогенными микроорганизмами, например RV. Показано получение указанного VHH или его применение в системе высвобождения в конкретном месте. Например, раскрыто высвобождение специфичного VHH в желудочно-кишечном тракте при инкапсулировании в альгинате. Кроме того, в качестве способа высвобождения предлагается применение трансгенных пробиотических микроорганизмов, которые высвобождают специфичные VHH-антитела, и при этом указанные микроорганизмы могут заселять кишечник человека. Предлагаются различные методики получения лекарственных средств и пищевых продуктов с использованием VHH-антител, которые инкапсулированы или экспрессируются пробиотическими микроорганизмами. Полученный VHH не связывается с VP6, не может быть нейтрализующим, а также используется не как таковой, а в системе контролируемого высвобождения.
В патентном документе US 20050054001 Muyldermans Serge описывает состоящие из тяжелых цепей антитела, функциональные домены состоящих из тяжелых цепей антител, функциональные VH-домены или их фрагменты, которые содержат некоторые модифицированные или мутантные аминокислотные последовательности. При этом не писаны последовательности, которые соответствуют доменам VHH, которые связывают VP6 RV.
В патентном документе WO 00/65057 описаны моновалентные белки, которые содержат один вариабельный домен, который связывается с вирусными антигенами, в частности бактериофага P2 Lactococcus. Описаны только последовательности VHH, которые ингибируют указанный бактериофаг.
В патентном документе US 2007/0009512, Hamers с соавторами, и в предыдущих документах тех же авторов, описаны фрагменты тяжелой цепи иммуноглобулинов и их применение для ветеринарного лечения, например пассивной иммунотерапии или серотерапии. Описанный VHH узнает только столбнячный токсин. Способ, применяемый для получения VHH, основан на получении мРНК иммунизированных верблюдовых.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Одной из целей настоящего изобретения является получение мономерного домена (VHH), происходящего из антител верблюдовых, при этом указанный домен может иметь одну из аминокислотных последовательностей, показанных в SEQ ID No. 1 , SEQ ID No. 2, SEQ ID No. 3 или SEQ ID No. 4, и при этом указанные домены связываются с белком VP6 RV группы A.
Другой целью настоящего изобретения является получение димерного домена, который связывается с белком VP6 RV группы A, при этом указанный слитый белок содержит, по меньшей мере, одну последовательность мономера, показанную в SEQ ID No. 1 , SEQ ID No. 2, SEQ ID No. 3 или SEQ ID No. 4. В предпочтительном варианте слитый белок содержит аминокислотную последовательность, показанную в SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11 и SEQ ID No. 12.
Другой целью настоящего изобретения является способ иммунологического выявления ротавируса, который включает в себя осуществление контакта RV-содержащего образца с одной из аминокислотных последовательностей, показанных в SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 6, SEQ ID No. 7, SEQ ID No. 8, SEQ ID No. 9, или их сочетанием; и выявление.
Указанный способ иммунологического выявления можно осуществлять с использованием любой из методик, известных в данной области, например, основанных на иммунологическом улавливании тестов ELISA, ELISPOT, конкурентного ELISA, магнитных шариков или теста быстрого выявления в свежих образцах «pen-side».
Другой целью настоящего изобретения является получение композиции, которая придает пассивный иммунитет млекопитающему, которая содержит любую из последовательностей, показанных в SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 6, SEQ ID No. 7, SEQ ID No. 8 или SEQ ID No. 9, эксципиент и иммуномодуляторы.
Другой целью настоящего изобретения является способ профилактики инфекций, вызываемых RV, который включает в себя введение эффективного количества любой из последовательностей, показанных в SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12, или их сочетания млекопитающему, которое нуждается в таком введении, при этом указанные последовательности вводят как таковые или в сочетании с веществами, которые инкапсулируют и защищают их от разрушения в желудочно-кишечном тракте.
Другой целью настоящего изобретения является способ лечения инфекций, вызываемых RV, при этом указанный способ включает в себя введение эффективного количества любой из аминокислотных последовательностей, показанных в SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12, и их сочетания млекопитающему, которое нуждается в таком введении, либо как таковых, либо в сочетании с веществами, которые инкапсулируют и защищают антитела от разрушения в желудочно-кишечном тракте.
ОПИСАНИЕ ФИГУР
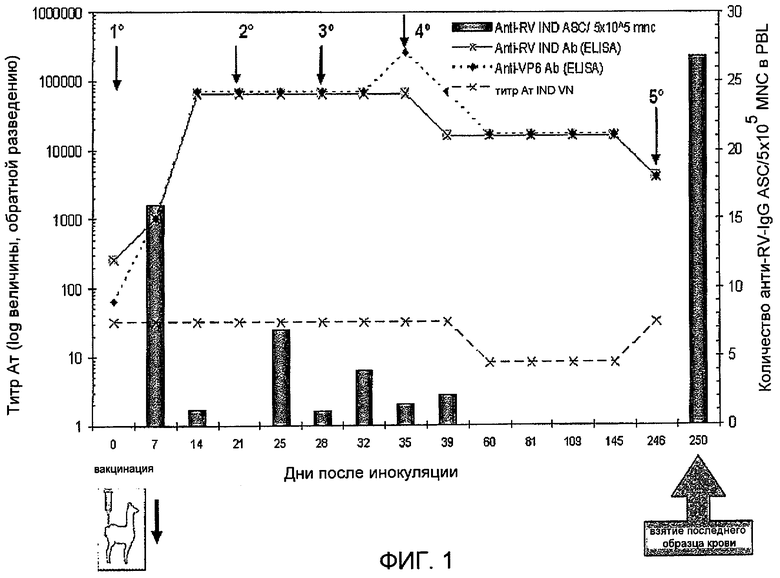
Фиг 1. Иммунизация ламы. Схема иммунизации, сбор образцов, конечное взятие крови и оценка гуморального ответа в виде антител против ротавируса в сыворотке в ходе иммунизации: титры антител, измеряемые (i) в ELISA с использованием рекомбинантного VP6, (ii) в ELISA с использованием ротавируса (IND; Sb I P[5]G6), (iii) с помощью нейтрализации вируса и (iv) в ELISPOT с использованием такого же ротавируса (IND; SbI P[5]G6). Временные точки вакцинации указаны стрелками.
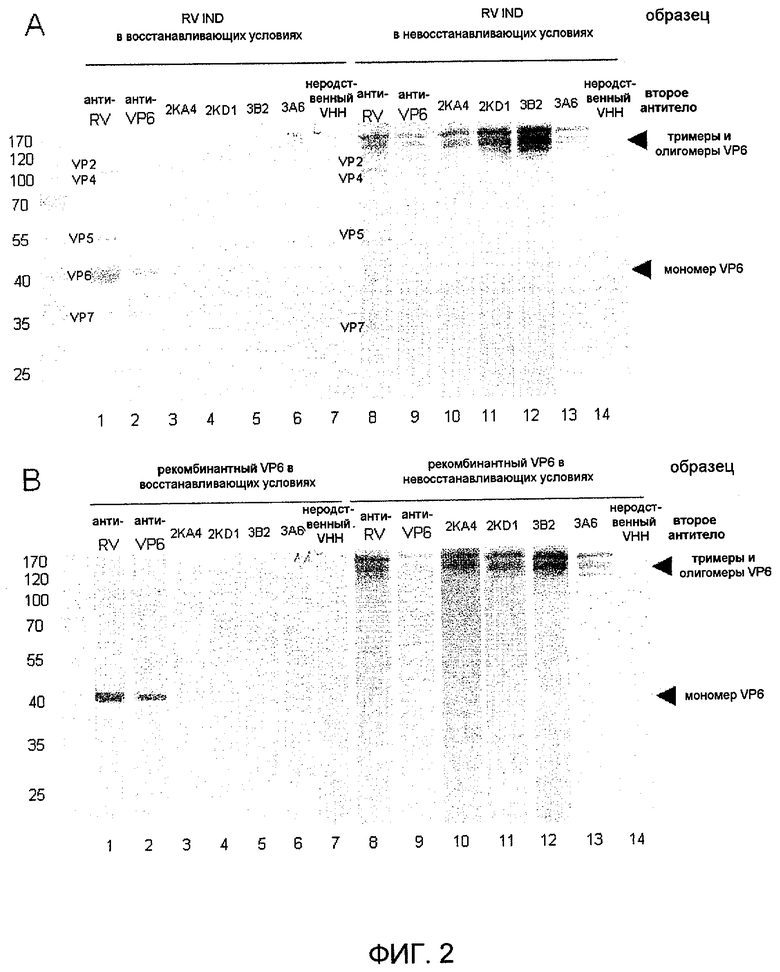
Фиг 2. Выявление нативного и рекомбинантного белка VP6 в Вестерн-блот-анализе. BRV IND (A) или рекомбинантный VP6 (B), разгоняемые в восстанавливающих условиях или невосстанавливающих условиях и выявляемые с использованием: дорожки 1 и 8 - поликлональной бычьей сыворотки против RV группы A; 2 и 9 - анти-VP6-мАт (RG25A10); 3 и 10 - VHH 2KA4 анти-VP6; 4 и 11 - VHH 2KD1 анти-VP6; 5 и 12 - VHH 3B2 анти-VP6; 6 и 13 - VHH 3A6 анти-VP6; 7 и 14 - не родственные VHH.
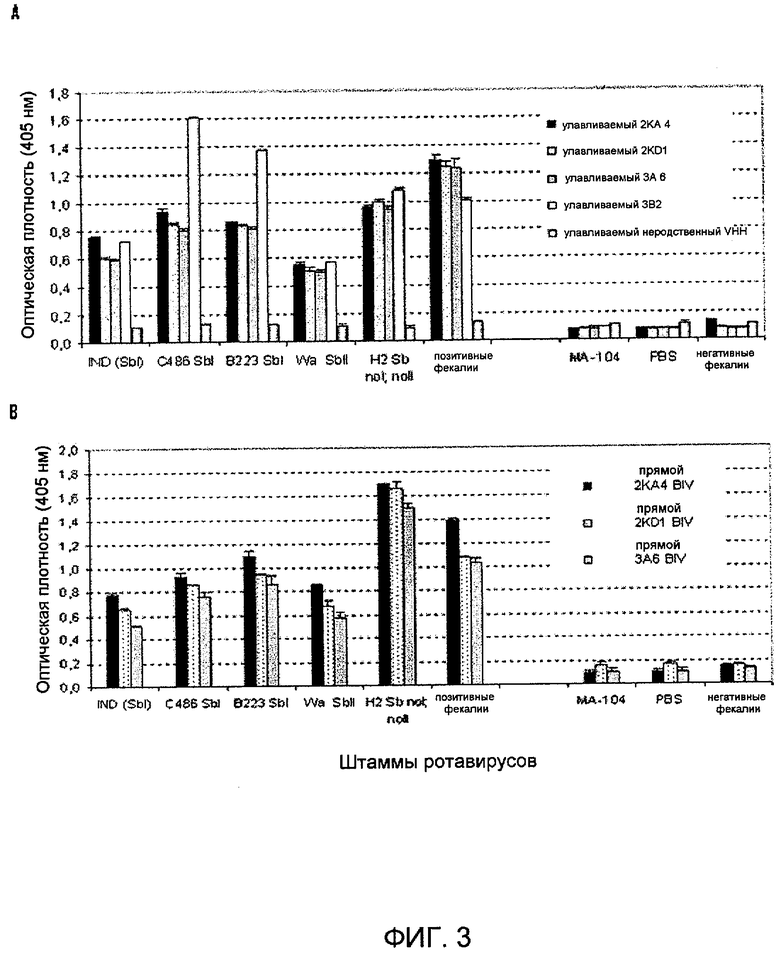
Фиг. 3. VHH-ELISA: Выявление ротавирусных штаммов с реактивностью разных подгрупп и разными специфичностями G/P-типов из разных видов животных.
A) Улавливаемый антителом мономерный VHH 2KA4, 2KD1, 3A6 (2 мкг/лунку).
B) Прямое покрывание бивалентным VHH biv2KA4, biv2KD1, biv3A6 (1 мкг/лунку). Надосадок культуры ткани бычьего ротавируса IND (SbI; P[5]G6), C486 (SbI; P[1]G6) и B223 (SbI; P[11]G10); ротавируса Wa человека (SbII; P[8]G1) и ротавируса H2 лошадей (Sb нет I, нет II; P[12]G3); позитивный образец кала: образец кала, соответствующий пику выделения вируса в среду у теленка, экспериментально инфицированного бычьим ротавирусом IND; MA-104: надосадок ложно инфицированных клеток. PBS: (пустой контроль реакции), негативный образец кала: образец кала теленка, отрицательный в отношении наличия ротавируса.
Величина ошибки, отмеченная на графике, показывает стандартное отклонение для двух независимых измерений.
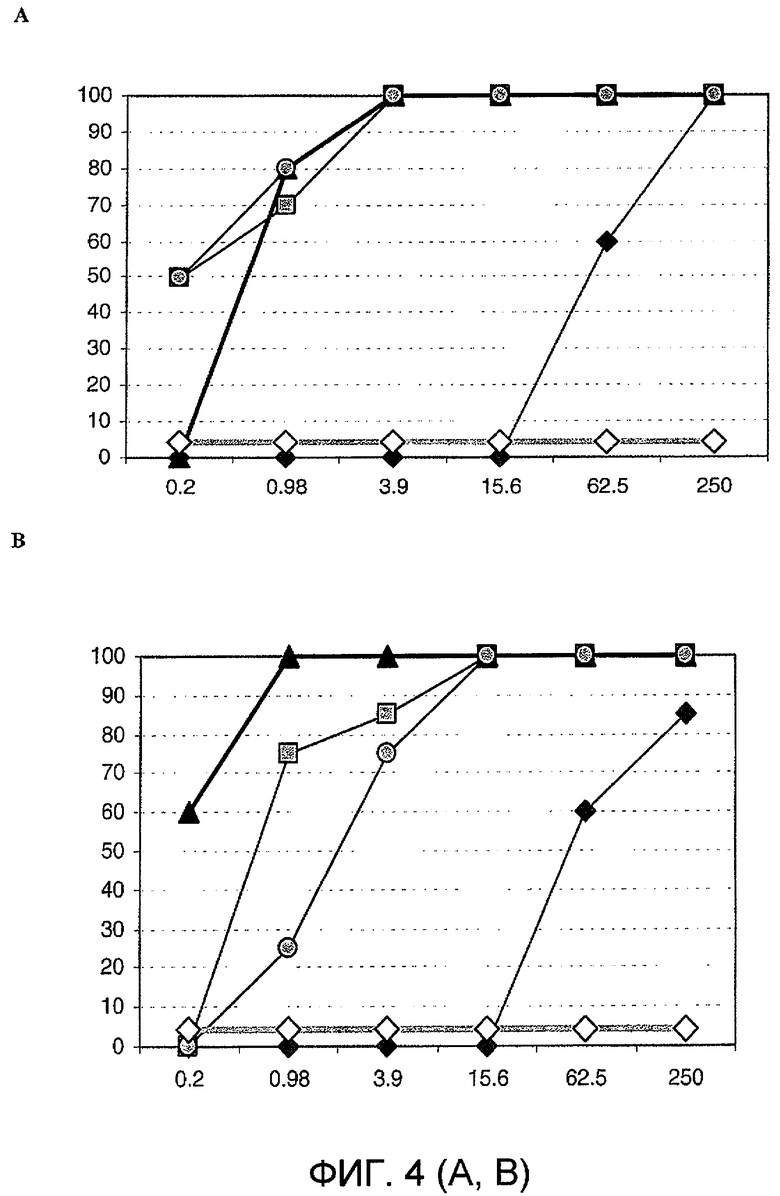
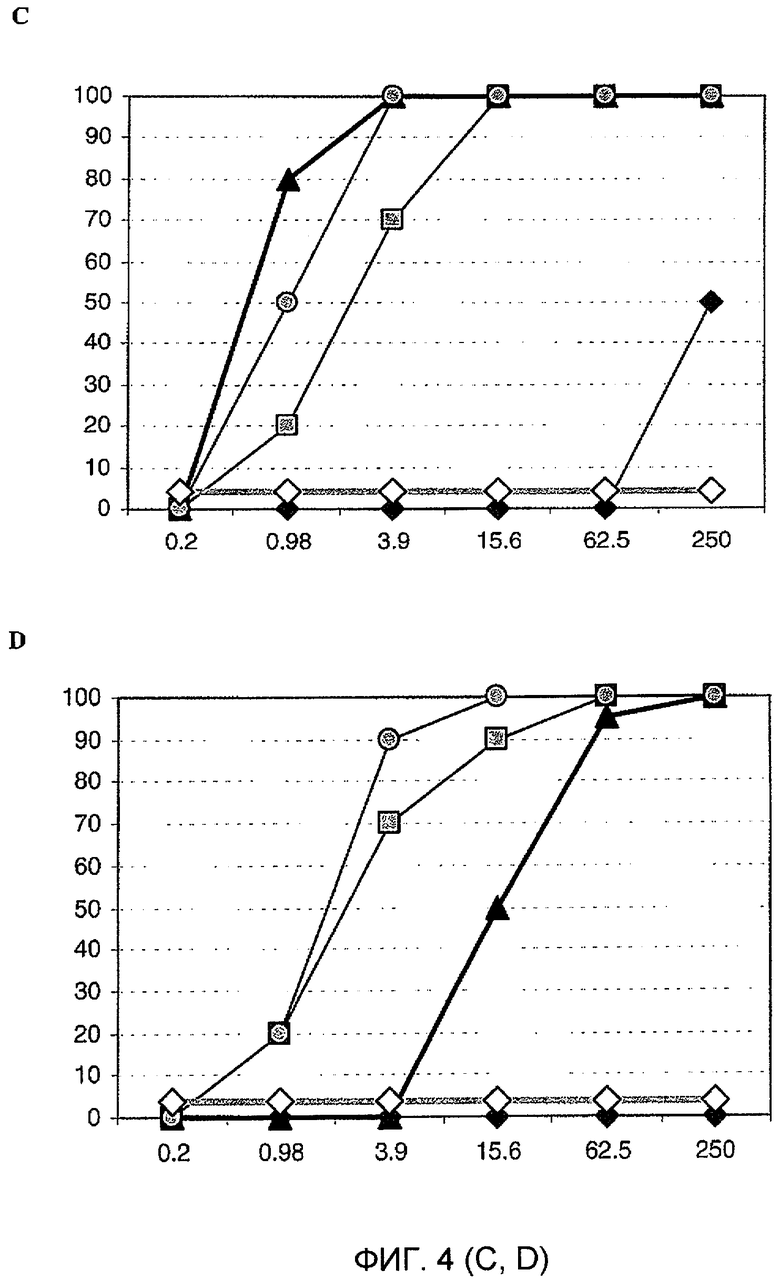
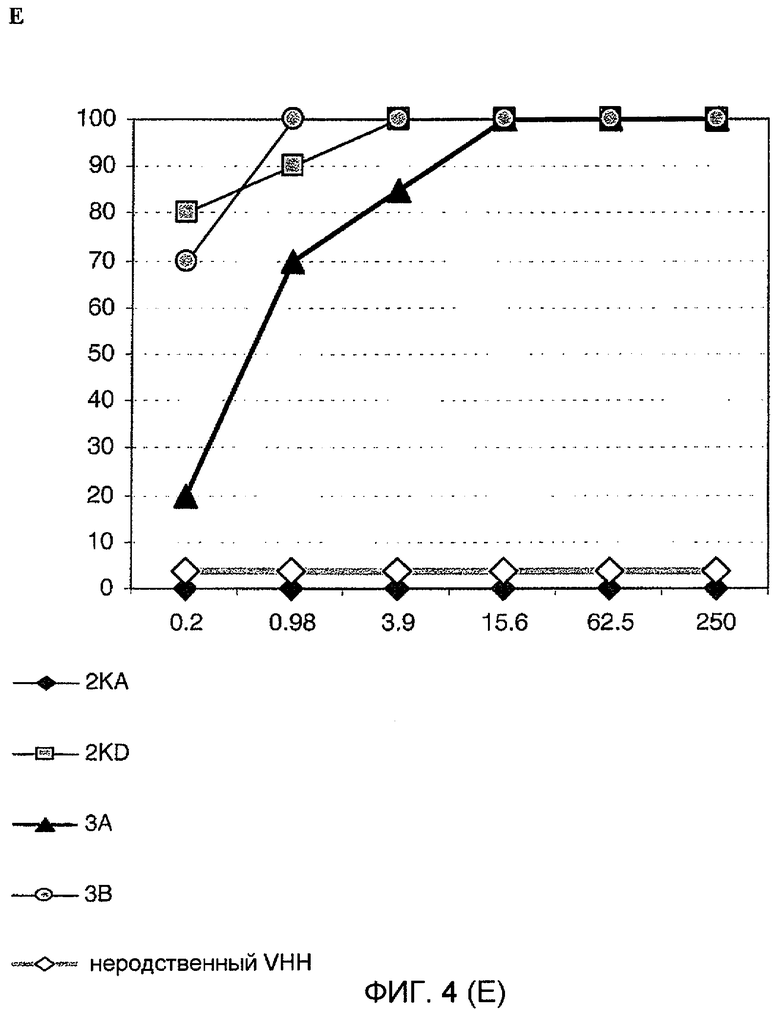
Фиг. 4. Анализ уменьшения очага флуоресценции вируса in vitro. Четырехкратное разведение каждого моновалентного VHH 2KA4, 2KD1, 3A6 и 3B2 смешивали с таким же объемом ротавируса, содержащего 100 ФОЕ. Концентрацию VHH, которая приводила к >80% уменьшению зараженности, считали защитной.
A. Бычий ротавирус C486 (гомолог антигена, используемого для вакцинации и заражения мышей);
B. Бычий ротавирус IND (гомолог антигена, используемого при отборе связывающего агента);
C. Бычий ротавирус B223;
D. Ротавирус Wa человека;
E. Ротавирус H2 лошадей.
На графиках представлены суммарные результаты двух независимых анализов.
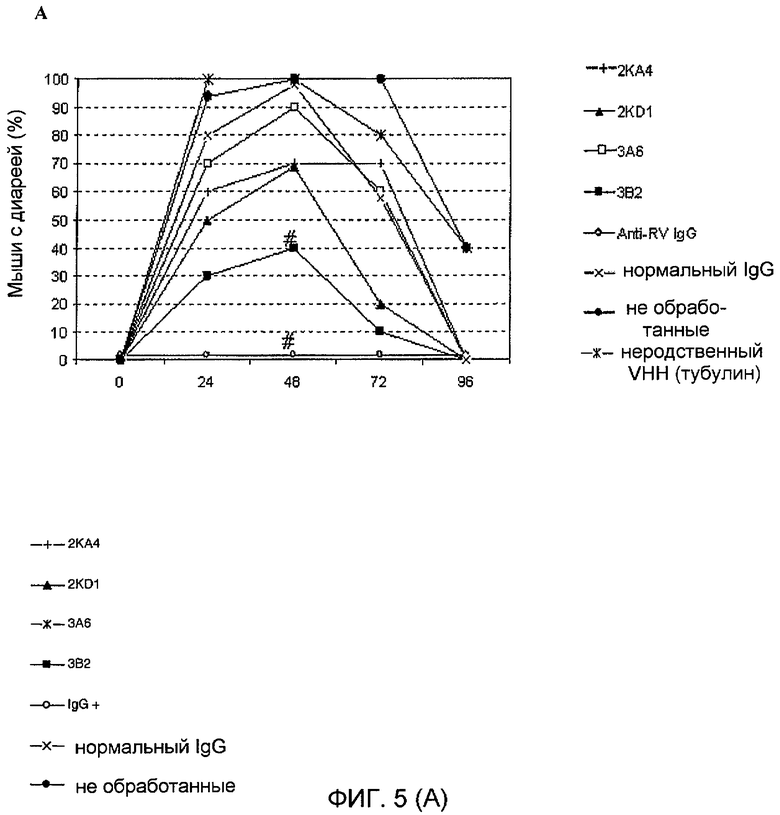
Фиг. 5. Степень защиты от диареи, достигаемая при использовании моновалентного VHH 2KA4, 2KD1, 3A6 и 3B2 в новорожденных мышей, зараженных ротавирусом. Мышатам давали 100 мкг (100 мкл) каждого VHH с 0 по 5 день один раз в сутки внутрижелудочным путем. В 1 день мышат заражали внутрижелудочно RV через 2 часа после обычного кормления. За диареей наблюдали ежедневно вплоть до 96 часов после заражения.
A. Заражение: 30 DD50 (6×105 ФОЕ) бычьего ротавируса C486. Эксперимент осуществляли в трех независимых анализах, используя по 5 мышей в группе.
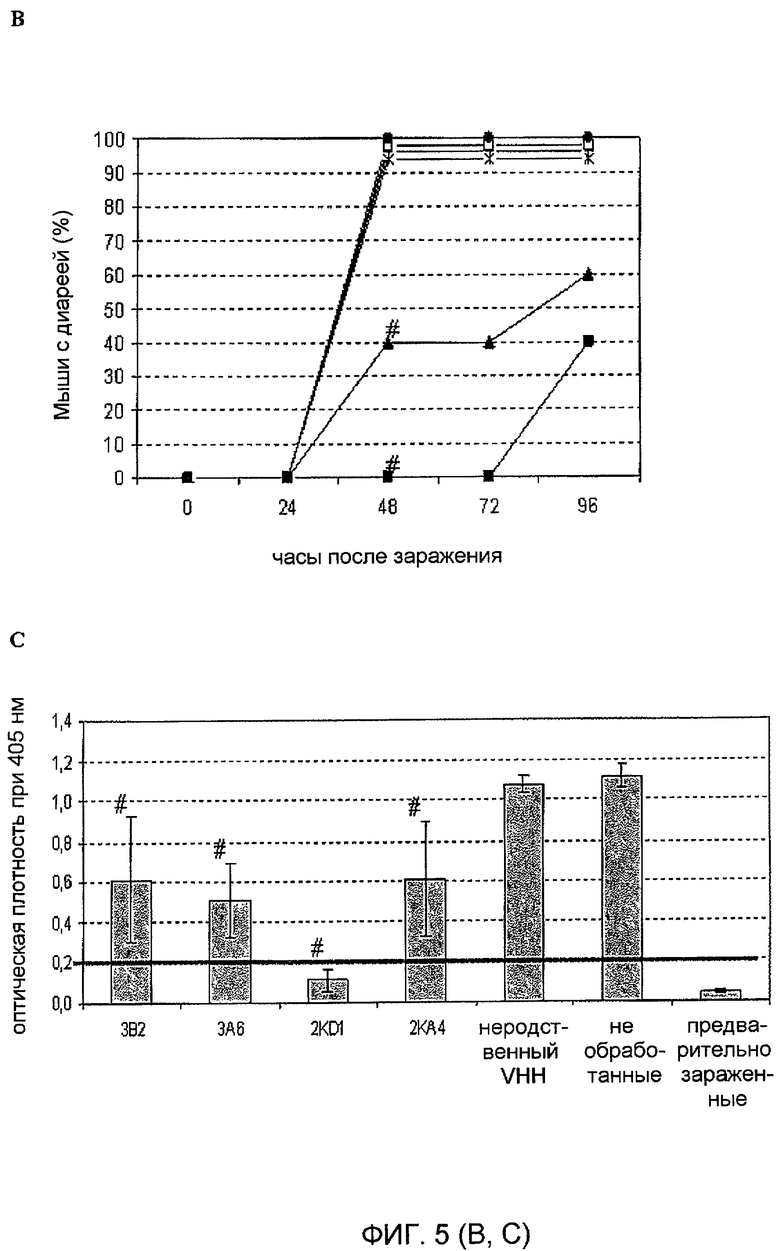
B. Заражение; 316 DD50 ротавируса ECw мышей. Эксперимент осуществляли в трех независимых анализах, используя по 5 мышей в группе.
C. Количественная оценка выделения вируса с помощью ELISA в 10% масс./об. гомогенатах тонкого кишечника.
Символ # означает процент пораженных животных, который значимо отличается от необработанной/зараженной группы, точный критерий Фишера, p<0,05.
Столбики представляют среднее значение оптической плотности в ELISA при 405 нм на группу. Величина ошибка, отмеченная на графике, показывает ± стандартное отклонение. Значение отсечения в ELISA: 0,200.
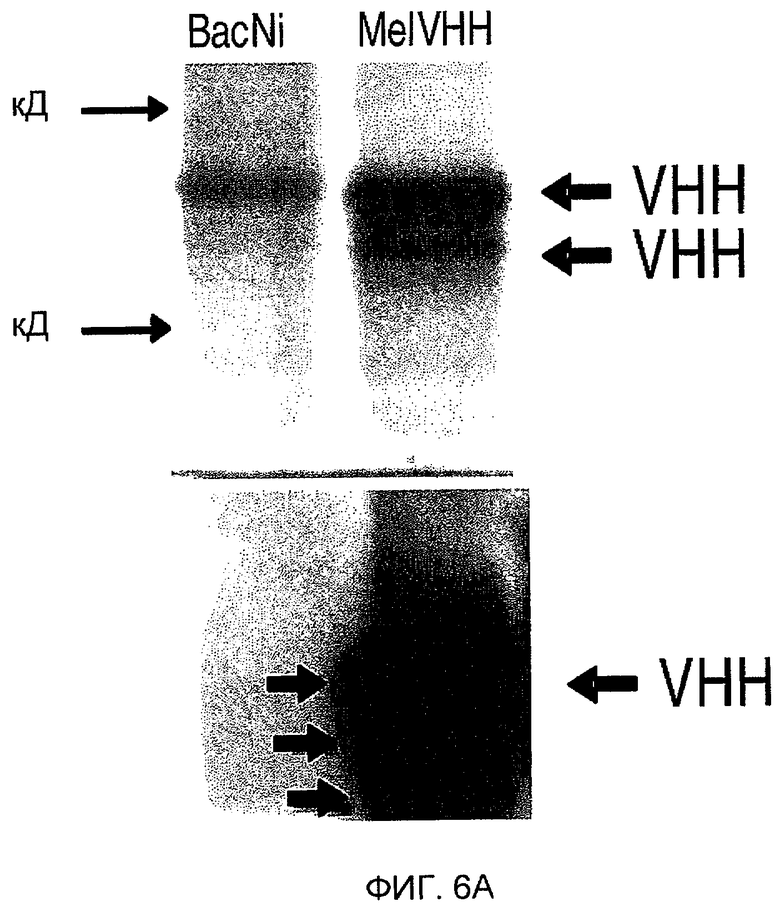
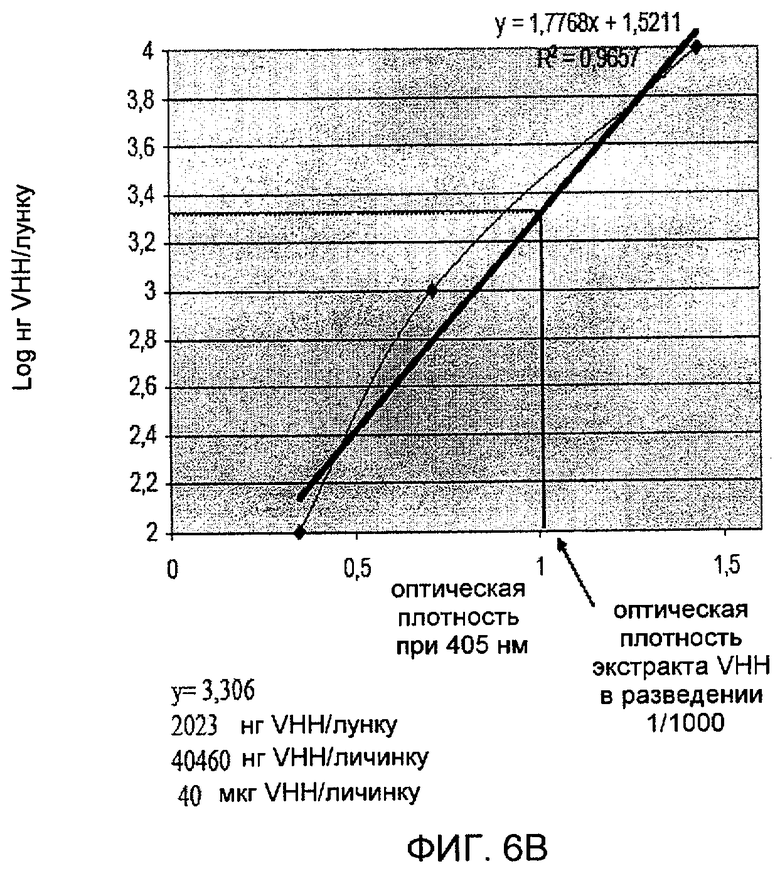
На фиг. 6A показана экспрессия VHH в личинках T.ni. Уровни экспрессии являются достаточно высокими для того, чтобы выявить две полосы в области ожидаемой молекулярной массы при окрашивании Кумасси. На фиг. 6B показана количественная оценка VHH, экспрессированного в системе личинок.
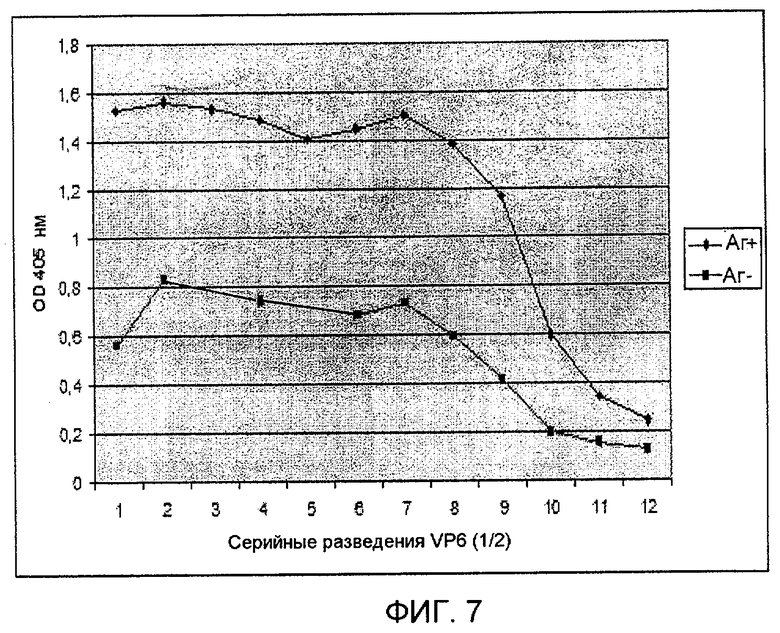
На фиг. 7 показан способ ELISA с использованием VHH из личинок. Суммарные экстракты растворимых белков (TSP) из личинок, экспрессирующих VP6 [Аг(+)], или из инфицированных личинок, имеющих не встроенный рекомбинантный бакуловирус [Аг(-)], использовали для покрывания микропланшетов для ELISA (Polysorp, Nunc, Denmark) с использованием серийных разведений, начиная с 40 мкг/лунку в 50 мМ карбонатном/бикарбонатном буфере, pH 9,6 и инкубировали в течение ночи при 4°C.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В целях описания настоящей заявки термин «домен» означает часть последовательности белка (антитела) и структуру, которая может эволюционировать, функционировать и существовать независимо от остальной части цепи белка. Каждый домен образует компактную трехмерную структуру и часто может быть независимо стабильным и может независимо подвергаться фолдингу. Многие белки состоят из нескольких структурных доменов. Один домен, по-видимому, может встречаться в нескольких эволюционно родственных белках. Длина доменов варьируется примерно от 25 аминокислот до 500 аминокислот. Домены способны связывать эпитопы. Домены согласно настоящему изобретению связываются с белком VP6 RV группы A.
В целях описания настоящей заявки подразумевается, что термины «VHH», «домен VHH», «мономерный VHH» и «мономер VHH» имеют одно и то же значение и являются взаимозаменяемыми.
В целях описания настоящей заявки подразумевается, что термины «белок VP6», «антиген VP6» и «VP6» имеют одно и то же значение и являются взаимозаменяемыми.
В целях описания настоящей заявки подразумевается, что термины «димерный домен», «димеры VHH» и «димерный VHH» имеют одно и то же значение и являются взаимозаменяемыми.
Термин «гомодимер» определяют как белок или полипептид, образованный двумя идентичными мономерными доменами при наличии или в отсутствие связывающей последовательности между ними.
Термин «гетеродимер» определяют как белок или полипептид, образованный двумя разными мономерными доменами при наличии или в отсутствие связывающей последовательности между ними.
Выражение «подходящие условия роста» относится к подходящим условиям среды, созданным для улучшения роста трансгенных клеток (трансформированных, трансфицированных или инфицированных вектором, определенным в настоящем изобретении). Например, инфицированных личинок поддерживают в камерах для роста при 28°C и собирают в указанное время.
В целях описания настоящего изобретения термин «вакцина» означает препарат, который применяют для повышения иммунитета к конкретному заболеванию. Композиция, предназначенная для придания пассивного иммунитета млекопитающему, отличается тем, что она содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No.1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12 и их сочетаний; эксципиент и иммуномодуляторы.
Выражение «эффективное количество аминокислотной последовательности (полипептида)», вводимое млекопитающему для профилактики инфекции, вызываемой RV, относится к предварительно определяемому количеству аминокислотной последовательности (полипептида), необходимого для связывания эпитопа и блокирования действия патогена (вируса), который в соответствующих условиях может инфицировать организм хозяина.
Выражение «эффективное количество аминокислотной последовательности (полипептида)», вводимое млекопитающему для лечения инфекции, вызываемой RV, относится к предварительно определяемому количеству аминокислотной последовательности (полипептида), необходимого для связывания эпитопа и блокирования действия патогена, который уже инфицировал организм хозяина.
В целях описания настоящего изобретения термин «млекопитающее», в общем, относится к классу млекопитающих, например человеку, коровам, козам, овцам, свиньям и лошадям.
Связывающую последовательность определяют как аминокислотную последовательность, которая связывает два мономерных домена.
Описан отбор, получение и характеристика VHH-антител, направленных к белку промежуточного капсида RV группы A, VP6. Показано, что VHH согласно изобретению являются белками с широкой реакционной способностью, которые могут быть обработаны так, чтобы использовать их в универсальном иммунодиагностическом анализе для выявления RV группы A. Кроме того, показано, что некоторые из отобранных анти-VP6-VHH обладают широкой нейтрализующей активностью in vitro, и что некоторые из указанных VHH защищают от вирусного заражения in vivo. На основе полученных сведений авторы изобретения полагают, что VHH согласно изобретению являются первыми описанными молекулами, которые связываются с белком VP6 и обладают нейтрализующей активностью.
Чтобы получить домены VHH согласно изобретению, осуществляли иммунизацию животных согласно схеме, показанной на фигуре 1. Чтобы оценить иммунный ответ у ламы, анализировали титры анти-RV- и анти-VP6-антител с помощью ELISA, анализов нейтрализации вирусов (VN) и специфичные секретирующие антитела клетки, циркулирующие в периферической крови, анализировали с помощью ELISPOT. Как и предполагалось, в 0 день после инокуляции (DPI) у лам получен позитивный результат в отношении анти-RV-антител, что свидетельствовало о предшествующем контакте с антигеном. Однако не выявлено секретирующих антитела клеток, циркулирующих в крови. На 7 день после инъекции титр анти-IND-RV или анти-VP6-антител, определенный в ELISA, был высоким, и в периферической крови определяли пик клеток, секретирующих RV-специфичные антитела (16 клеток, продуцирующих анти-RV-IgG/5 x 105 мононуклеарных клеток). Гуморальный ответ достигал плато, начиная с 14 дня после инъекции, с высокими титрами антител как к целому вирусу, так и к белку VP6. Напротив, титры нейтрализующих антител оставались сходными и очень низкими в случае всех антител к разным исследованным RV (IND, B223, Wa и H2). Хотя в сыворотке получали очень высокие титры антител, количество клеток, секретирующих анти-RV-антитела, выявленных в крови, уменьшалось с каждой бустер-иммунизацией (фиг. 1). По этой причине и чтобы обеспечить достаточный период времени, обеспечивающий созревание аффинности антител, лама получала конечную дозу VP6 намного позже на 246 день после инъекции (примерно через 7 месяцев после 4-ой дозы). Наконец, у ламы брали кровь через 4 дня после последней бустер-иммунизации, при этом значение количества клеток, секретирующий анти-RV-IgG, достигало 26,8/5×105 мононуклеарных клеток. 6×108 мононуклеарных клеток извлекали из 900 мл крови (которые содержали, по меньшей мере, 32160 клеток, секретирующих RV-специфичные IgG-антитела согласно результатам ELISPOT). На основе процессированной РНК (210 мкг) создавали фаговую библиотеку VHH, которая содержала 6×107 клонов. Используемая схема вакцинации показала, что для того, чтобы получить VHH согласно изобретению, важнее достичь высоких значений количества клеток, секретирующих специфичные антитела, чем высоких титров антител, направленных против представляющих интерес антигенов. В случае используемой схемы вакцинации можно получить достаточные количества клеток, секретирующих специфичные антитела. Результаты, полученные в ходе иммунизации, позволили авторам особое внимание обратить на то, что наилучшим способом регистрации иммунного ответа у иммунизированных лам для создания библиотеки VHH является выбор такого способа, который позволяет оценивать количество секретирующих специфичные антитела клеток, циркулирующих в периферической крови, вместо определения в сыворотке титров антител против представляющего интерес антигена. Согласно с полученной картиной клеток, секретирующих антитела, показано, что схема вакцинации с длительным интервалом между последними двумя дозами стимулирует циркуляцию большего количества клеток, секретирующих специфичные антитела, в периферической крови. Также было показано, что на 4 день после инокуляции присутствует большее количество циркулирующих клеток, секретирующих антитела против представляющего интерес антигена, чем на 7 день после инокуляции. Схему иммунизации следует осуществлять таким образом, чтобы вводить ламам бустер-дозу, по меньшей мере, через 5 месяцев после последней дозы, и колонии фагов создают, когда количество клеток, секретирующих IgG против антигена-мишени в периферической крови, достигает примерно 20. Предпочтительно в случае анти-VP6-VHH количество клеток, секретирующих IgG-антитела, в периферической крови должно составлять примерно 30 клеток, секретирующих IgG-антитела против полного RV. Специалистам в данной области известно, что можно использовать другие схемы иммунизации, чтобы получить подходящий VHH для применения в способах и композициях согласно настоящему изобретению.
Чтобы отобрать фаги, которые экспрессировали анти-RV-VHH, осуществляли три раунда селекции in vitro, используя RV IND в качестве антигена. Отобрали 192 клона. Рестрикционные анализы осуществляли для всех клонов, которые сильно отличались в отношении связывания VP6 в библиотеке VHH, что определяли с помощью ELISA фагов. Клоны также анализировали в ELISA, чтобы определить их способность связываться с RV и с VP6. Из 14 клонов с разными последовательностями отобрали 10 клонов, для которых показано более сильное специфичное связывание с RV и VP6, и такие клоны субклонировали в экспрессирующем векторе, который обеспечивает гексагистидиновую метку на карбоксильном конце для облегчения очистки (таблица 1).
Сводные результаты количественной оценки для отбора доменов VHH
ELISA фага
ELISA фага
Отобрали четыре клона, которые более сильно связывались со штаммами RV, которые соответствовали разным подгруппам; указанные клоны названы 2KA4 (SEQ ID No. 1), 2KD1 (SEQ ID No. 2), 3A6 (SEQ ID No. 3) и 3B2 (SEQ ID No. 4), которые узнавали рекомбинантный VP6 и его нативный аналог RV IND, судя по оценке с помощью Вестерн-блота, который показывает, что указанный VHH связывается с конформационными эпитопами такого белка VP6 (фиг. 2). Ниже VHH, называемые 2KA4, 2KD1, 3A6 и 3B2, представляют собой домены VHH согласно изобретению.
Последовательности ДНК, которые кодировали каждый из доменов VHH, указаны ниже:
SEQ ID No. 5 кодирует домен SEQ ID No. 1;
SEQ ID No. 6 кодирует домен SEQ ID No. 2;
SEQ ID No. 7 кодирует домен SEQ ID No. 3;
SEQ ID No. 8 кодирует домен SEQ ID No. 4.
Специалистам в данной области понятно, что любая последовательность ДНК, которая кодирует указанные домены, входит в объем настоящего изобретения. Например, аминокислотная последовательность SEQ ID No. 1 может кодироваться последовательностью ДНК SEQ ID No. 5 или другой последовательностью ДНК, которая отличается от последовательности SEQ ID No. 5 вследствие вырожденности генетического кода. Такой же пример применим для каждой из аминокислотных последовательностей (SEQ ID No. 2, 3 и 4), кодируемых соответствующими последовательностями ДНК (SEQ ID No. 6, 7 и 8).
Специфичные анти-VP6-VHH согласно изобретению были очень эффективны в качестве реагентов для иммунодиагностики RV. Мономерные формы VHH согласно изобретению анализировали в ELISA в качестве улавливающих антител, в качестве вторых антител или иммобилизованы с помощью анти-His-антител. VHH согласно изобретению способны выявлять штаммы RV из организма человека или животного разных специфичных подгрупп и разных типов G и P (фиг. 3A).
С другой стороны, конструировали экспрессирующие векторы, чтобы получить димерные VHH согласно изобретению, которые специфично связываются с VP6, которые содержали идентичные гены VHH, соединенные связывающей последовательностью, сходной с последовательностью шарнира IgA человека. Димерные VHH согласно изобретению также оценивали в ELISA в качестве прямого улавливающего антитела и выявили отчетливые воспроизводимые сигналы в случае всех исследованных штаммов RV (фиг. 3B). Димерные VHH согласно изобретению можно использовать для сенсибилизации планшета для ELISA, таким образом исключая необходимость в использовании антител, улавливающих VHH, и полученные при этом результаты по выявлению RV были лучше, чем в случае мономерных аналогов.
Стоит подчеркнуть, что выходы мономерных VHH, экспрессированных в периплазме E. coli, были сравнимы с выходами, о которых сообщалось другими авторами для E. coli и дрожжей.
Доказано, что мономерные и димерные VHH согласно настоящему изобретению весьма полезны для диагностики RV и поэтому могут быть использованы в качестве рекомбинантных моноклональных антител в любых иммунодиагностических анализах, известных специалистам в данной области. Специалистам в данной области понятно, что мономерные и димерные домены VHH согласно изобретению можно применять для любого типа иммунодиагностических анализов, чтобы выявить RV, и что указанные анализы входят в объем настоящего изобретения.
Анти-VP6-домены согласно изобретению можно использовать для конструирования димеров, таких как димеры, описанные в настоящей публикации, например гомодимеры, или можно сливать с образованием гетеродимеров, например, в результате слияния домена 3B2 и домена 3A6, или слияния любых двух мономеров VHH согласно изобретению. Кроме того, три или более мономеров VHH согласно изобретению также можно сливать или сочетать с образованием гомотримеров или гетеротримеров. Все мультимерные формы, которые возникают в результате сочетания мономеров VHH согласно изобретению, независимо от того, содержат ли они или не содержат связывающую последовательность, входят в объем настоящего изобретения. В предпочтительном варианте димеры VHH согласно изобретению имеют последовательности, указанные в SEQ ID No. 9, или SEQ ID No. 10, или SEQ ID No. 11, или SEQ ID No. 12. Например, димеры могут содержать связывающую аминокислотную последовательность, такую как последовательность, показанная в SEQ ID No. 13, или любая другая последовательность, которая действует в качестве шарнирной последовательности.
В следующей ниже таблице показаны некоторые характеристики димерных и мономерных VHH согласно изобретению.
Характеристики
2 на основании группы из 6 культур по 0,5 л каждой, очищенный с помощью колонки с антителами против гистидиновой метки;
3 выявление RV-штаммов Sb I, Sb II, Sb без I; без II.
Анти-VP6-VHH-домены согласно изобретению способны нейтрализовать разные штаммы RV in vitro. Три из четырех мономеров согласно изобретению (2KD1, 3A6 и 3B2) проявляли широкую нейтрализующую активность in vitro. Способность к нейтрализации каждого VHH была однородной по отношению ко всем оцениваемым штаммам RV. Концентрации мономеров от 3,9 мкг/мл были способны полностью нейтрализовать инфекционность, вызываемую 100 ФОЕ штаммов RV C486 (P[1]G6), IND (P[5]G6), B223 (P[11]G10), Wa P[8]G1 и H2 (P[12]G3) in vitro. В таблице 3 указан титр нейтрализующих антител против разных штаммов RV при использовании раствора с концентрацией 2 мг/мл каждого мономера.
Титр разных нейтрализующих мономерных и димерных доменов VHH
Sbl
P[5]G6
Sbl
P[1]G6
Sbl
P[11]G10
Sbll
P[8]G1
Sb nol
P[12]G3
Следовательно, мономеры VHH способны нейтрализовать штаммы RV, относящиеся к разным сочетаниям G/P-типов, которые обычно не индуцируют перекрестную нейтрализацию. Мономер VHH с наибольшей нейтрализующей способностью представлял собой мономер 2KD1. С другой стороны, хотя мономер 2KA4 способен соответственно узнавать все RV в ELISA, он не нейтрализовал никакие исследованные штаммы. Способность VHH согласно изобретению нейтрализовать высокие титры штаммы RV с разными антигенами имеет большое значение и может сделать их полинейтрализующими молекулами; такое свойство делает их возможными средствами для профилактики или лечения RV-индуцированной диареи, независимо от серотипа (27 P-типов и 16 G-типов).
Димерный VHH проявлял более слабую нейтрализующую активность, чем его мономерные аналоги.
Оценивали способность мономеров VHH согласно изобретению оказывать лечебное и профилактическое действие в случае диареи, вызванной инфекцией RV. С этой целью новорожденным мышам ежедневно внутрижелудочно вводили дозу VHH в течение 5 дней (с 0 по 4 день). Мышей заражали в 1 день штаммом вируса C486 также внутрижелудочным путем (фиг. 5). 60% мышей, обработанных мономерным VHH согласно изобретению, 3B2, были защищены от RV-индуцированной диареи. Такая защита была значимо выше, когда проводили сравнение между обработанными мышами и необработанными мышами или по сравнению с мышами, обработанными неродственным VHH (p=0,0108), когда все мыши страдали от диареи (таблица 4 и фигура 5). Кроме того, тяжесть и продолжительность диареи у животных, обработанных VHH согласно изобретению, были значимо снижены по сравнению с контрольными группами.
Защита от заражения ротавирусом у лактирующих мышей
Средние значения в некоторых колонках с разными буквенными обозначениями отличаются значимо (критерий Крускала-Уоллиса, p<0,05).
Проценты пораженных животных с разными буквенными обозначениями отличаются значимо (точный критерий Фишера, p<0,05).
Следует отметить, что самый высокий титр нейтрализации был получен против гетерологичного штамма RV человека (Wa, SbII, P[8]G1), который считают штаммом, в большинстве случаев ассоциированным с гастроэнтеритом у детей во всем мире.
Получение и очистку указанных фрагментов антител можно осуществить с высокими выходами, что приводит к более низкой себестоимости продукции. Это особенно существенно для развивающихся стран, в которых распространение инфекции и заболеваемость/смертность от RV огромны, и затраты на лечение и профилактику являются решающими ограничивающими факторами.
Любому специалисту в данной области понятно, что на основе приведенного в данной публикации описания домены VHH согласно изобретению могут быть получены в результате синтеза соответствующей нуклеотидной последовательности и ее экспрессии в любой клетке-хозяине без необходимости в создании фаговой библиотеки.
Любому специалисту в данной области понятно, что различные сочетания и смеси мономеров VHH, димеров VHH или мультимеров VHH согласно изобретению можно использовать для иммунодиагностики, профилактики RV-инфекций и лечения млекопитающих, инфицированных RV, без изменения сущности настоящего изобретения, и при этом все возможные сочетания и смеси входят в объем настоящего изобретения.
Как указано выше, отобрали четыре клона, которые сильнее связываются со штаммами RV, которые соответствовали разным подгруппам. Такие клоны названы 2KA4 (SEQ ID NO: 1), 2KD1 (SEQ ID NO: 2), 3A6 (SEQ ID NO: 3) и 3B2 (SEQ ID NO: 4), и они узнавали рекомбинантный VP6 и его нативный аналог RV IND, судя по оценке с помощью Вестерн-блота, в котором показано, что указанный VHH связывается с линейными эпитопами такого белка VP6.
Указанные домены согласно изобретению (SEQ ID NO: 1-4) или кодирующие их нуклеотидные последовательности (SEQ ID NO: 5-8) могут быть встроены в соответствующий рекомбинантный вектор, такой как экспрессирующий вектор. Следовательно, настоящее изобретение, кроме того, относится к экспрессирующему вектору, который содержит указанные последовательности. Выбор вектора зависит от типа клетки-хозяина, в которую собираются вводить вектор. В качестве иллюстрации вектор может представлять собой плазмиду или вектор, который после внедрения в клетку-хозяина интегрируется или не интегрируется в геном указанной клетки-хозяина. Указанный вектор может быть получен с использованием любого известного способа, имеющегося в данной области [Sambrook et al., 1989]. В предпочтительном варианте может быть использован вектор согласно изобретению, встраиваемый в геном клеток растений или животных. Таким образом, вектор согласно изобретению может представлять собой, например, Agrobacterium tumefaciens или вирусный вектор, способный экспрессироваться в клетках растений или животных. В конкретном варианте вирусным вектором, используемом в настоящем изобретении, является бакуловирус (см. пример 5).
Вектор может быть использован для трансформации, трансфекции или инфекции клеток растений, водорослей или животных, предпочтительно клеток насекомых или личинок. Поэтому настоящее изобретение, кроме того, относится к клеткам, трансформированным, трансфицированным или инфицированным вектором согласно изобретению.
В предпочтительном варианте согласно изобретению трансгенная клетка представляет собой клетку животного, предпочтительно клетку насекомого и более предпочтительно личинки указанного насекомого. Следовательно, изобретение, кроме того, относится к трансгенному животному, отличному от человека, такому как трансгенное насекомое или трансгенная личинка, с высоким выходом экспрессирующему пептид, характеризуемый последовательностями SEQ ID: 1-4.
Следовательно, вектор согласно изобретению можно использовать для получения и/или хранения доменов согласно изобретению, характеризуемых последовательностями SEQ ID NO: 1- 4 и/или 9-12. Поэтому настоящее изобретение, кроме того, относится к способу получения доменов согласно изобретению, который включает в себя выращивание клетки или организма, трансфицированного, трансформированного или инфицированного вектором согласно изобретению в условиях, которые обеспечивают получение указанных доменов. Условия оптимизации культивирования трансгенной клетки или организма будут зависеть от типа используемой клетки или организма. При необходимости способ получения доменов согласно изобретению дополнительно включает в себя выделение и очистку любым способом, известным в данной области.
В предпочтительном варианте настоящее изобретение относится к антителам, отличающимся тем, что они содержат любой из доменов согласно изобретению:
- мономерный домен VHH, полученный из антител верблюдовых, отличающийся тем, что он содержит последовательности, выбранные из группы, состоящей из последовательностей SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3 и SEQ ID No. 4, при этом указанные домены связываются с белком VP6 ротавируса (RV) группы A;
- димерный домен VHH, который связывается с белком VP6 RV группы A, отличающийся тем, что он содержит, по меньшей мере, одну мономерную последовательность, выбранную из группы, состоящей из последовательностей, показанных в SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3 и SEQ ID No. 4.
Другой аспект настоящего изобретения относится к набору для иммунодетекции RV, содержащему аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12 и их сочетаний.
Изобретение, кроме того, относится к аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12 и их сочетаний, для применения в способе профилактики инфекций, вызванных RV. Другими словами, изобретение относится к применению аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12 и их сочетаний, для производства композиция для профилактики инфекций, вызванных RV.
Кроме того, изобретение относится к аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12 и их сочетаний, для применения в способе лечения инфекций, вызванных RV. Другими словами, изобретение относится к применению аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12 и их сочетаний, для производства композиции для лечения инфекций, вызванных RV.
Наконец, настоящее изобретение относится к способу пассивной иммунизации, который включает в себя инокуляцию эффективного количества определенных выше антител в организм человека или животного.
ДЕПОЗИТ МИКРООРГАНИЗМОВ СОГЛАСНО БУДАПЕШТСКОМУ ДОГОВОРУ
Плазмида pFBMelVHH (см. пример 5) депонировали в Испанской коллекции типов культур (CECT); University of Valencia, Spain, с номером доступа CECT7431, 02.07.08.
ПРИМЕРЫ
Настоящее изобретение лучше иллюстрировано следующими примерами, которые не следует интерпретировать как ограничение объема изобретения. Напротив, необходимо ясно понимать, что могут быть использованы другие варианты, модификации и эквиваленты изобретения, которые могут предполагать специалисты в данной области на основании приведенного описания, не отходя от сути настоящего изобретения и/или не выходя за рамки объема прилагаемой формулы изобретения.
Пример 1: Получение и очистка мономерного и димерного VHH согласно изобретению
Получение библиотеки VHH согласно изобретению:
Эталонный штамм бычьего IND RV (SbI; P[5]G6) использовали в качестве антигена в способе биопэннинга, чтобы отобрать VHH. Чтобы получить панель RV, на которой представлены реактивности разных подгрупп с разными сочетаниями G- и P-типов из разных видов животных и человека, эталонные штаммы RV, перечисленные в таблице 5, включали в разные анализы, осуществляемые при получении VHH. Вирусы размножали в клетках почки обезьяны (MA-104). Также включали фекальный образец от лишенного молозива новорожденного теленка, инфицированного штаммом IND во временной точке до инокуляции и в точке пика размножения вируса.
Эталонные штаммы ротавирусов, используемые в разных способах,
осуществляемых во время получения и характеристики VHH
согласно изобретению
Гуморальный ответ у лам (VN)
Характеристика связывающего агента (VN, ELISA)
Заражение мышей
Характеристика связывающего агента (VN, ELISA)
Гуморальный ответ у лам (VN, ELISA, ELISPOT)
Характеристика связывающего агента (VN, ELISA)
Характеристика связывающего агента (VN, ELISA)
Нет II
Характеристика связывающего агента (VN, ELISA)
Иммунизация лам: белок VP6, полученный из штамма бычьего RV
C486 (SbIP[1]G6) получали в клетках Sf9, инфицированных рекомбинантным бакуловирусом. Самцы лам в возрасте одного года получали пять доз неочищенного клеточного экстракта, содержащего 500 мкг VP6 в смеси с масляным адъювантом INTA (Marcol:Arcel:Span:Tween) в 0, 21 , 28, 35 и 246 дни. Образцы сыворотки и крови брали в 0, 4 и 7 день после каждой инокуляции. Гуморальный ответ оценивали с помощью ELISA и анализа нейтрализации вируса (VN) (см. ниже). Чтобы оценить ответ эффекторных B-клеток, адаптировали анализ ELISPOT, в котором определяли количество клеток, секретирующих RV-специфичные антитела, в периферической крови инокулированной ламы, на основе описанных ранее анализов ELISPOT на свиньях и телятах (Parreno, V. C. et. al., Vet. Immunol. Immunopathol. 100: 7-24, 2004, и Parreno, V.V. et. al., J. Vet. Med. B infect. Dis. Vet. Public Health 48: 713-20, 2001, включенные в настоящее описание исключительно в виде ссылки). Коротко, клетки MA-104, инфицированные BRV IND (при этом более чем 80% инфекции выявляли с помощью иммунофлуоресценции), выращенные в 96-луночном планшете, фиксировали 70% ацетоном, сушили на воздухе и хранили при -20°C вплоть до использования. Суспензии мононуклеарных клеток (MNC), полученных из периферической крови (PB) ламы, которой проводили инокуляцию, добавляли в лунки (1×106; 5×105; 2,5×105 и 1,25×105 клеток/лунку). После центрифугирования при 500 g в течение 5 минут планшеты инкубировали в течение 12-14 часов при 37°C в 5% CO2. Планшеты промывали PBS с 0,05% твина-20, чтобы удалить слипшиеся клетки, и получали пятна, добавляя такой же конъюгат, который использовали в ELISA, в разведении 1/1500 на 2 часа при 37°C, затем добавляли 50 мкл системы субстрата пероксидазы TMB (KLP, Maryland, USA).
Приготовление, инокуляцию и сбор образцов из организма ламы осуществлялись обученным персоналом под наблюдением ветеринара в соответствии с протоколами, одобренными комитетом по этике в отношении животных и благополучию животных INTA.
Получение библиотеки VHH и отбор VP6-связывающих VHH согласно изобретению: Из общего объема 900 мл крови, собранной на 4 день после последней инъекции, 6×108 мононуклеарных клеток выделяли с помощью центрифугирования в градиенте фикол-пак; затем их центрифугировали, замораживали в жидком азоте и хранили при -80°C. Суммарную РНК экстрагировали, используя оборудование для экстракции РНК (Macherey Nagel; Nucleospin RNA II), получая 250 мкг РНК. Затем синтезировали первую цепь кДНК, используя обратную транскриптазу Superscript III (Invitrogen) и олиго-dT-праймеры (12-18) (Invitrogen) или случайные праймеры (Invitrogen). В 20 мкл реакционной смеси использовали 0,2, 1 или 5 мкг суммарной РНК. кДНК, кодирующие VHH и VH, специфично амплифицировали в ПЦР, используя праймеры CALL01 (SEQ ID No. 14) и CALL02 (SEQ ID No. 15), которые отжигались с лидером и последовательностью CH2. Фрагмент длиной 600 п.н. (экзоны VHH-CH2 без экзона CH1) элюировали из 1,6% агарозного геля после отделения его от фрагмента длиной 900 п.н. (экзона VH-CH1-CH2). Затем VHH амплифицировали в дополнительной гнездовой ПЦР с праймерами, которые отжигались с областью каркаса 1 (SEQ ID No. 16) и областью каркаса 4 (SEQ ID No. 17), и с праймерами, содержащими сайты рестрикции для последующих стадий клонирования: VHHfor2: (SEQ ID No. 18) с сайтами рестрикции для NcoI и PstI, и VHHrev2 (SEQ ID No. 19) с сайтом рестрикции для NotI. Конечные ПЦР-фрагменты лигировали, используя расположенные выше сайты рестрикции NcoI или PstI и расположенный ниже сайт рестрикции NotI, в фагмидный вектор pAO-Lib, модифицированный вариант pHEN4 (Arbabi Ghahroudi M, Desmyter A, Wyns L, Hamers R, Muyldermans S. Selection and identification of single domain antibody fragments from camel heavy-chain antibodies. FEBS Lett. 1997 Sep. 15; 414(3): 521-6), который содержит нерелевантную длинную последовательность, которая удаляется после инсерции VHH, для того, чтобы задержать размножение вектора без вставки VHH. Клетки Escherichia coli (TG1) трансформировали лигированным материалом и клетки высевали. Колонии снимали с планшетов, промывали и хранили при -80°C в среде LB с добавлением глицерина (конечная концентрация 50%).
Специфичные VHH собирали из библиотеки, используя методику фагового дисплея. Библиотеку VHH инфицировали хелперным фагом M13 (Invitrogen), и фаговые частицы, которые экспрессируют репертуар VHH, спасали и преципитировали ПЭГ, как описано в Marks, J. D., Hoogenboom, H. R., Bonnert, T.P., McCafferty, J., Griffiths, A.D., Winter, G. JMB, 1991. Обогащение специфичными VHH осуществляли, используя два-три раунда селекции in vitro, то есть используя методику биопэннинга. Иммунологические пробирки покрывали разведением 1/50 полуочищенного BRV IND (SbI; P[5]G6) или разведением 1/5000 поликлональной антисыворотки против RV морской свинки в карбонатном буфере, pH 9,6, и оставляли в течение ночи при 4°C, затем осуществляли стадию блокирования и улавливали разведение 1/50 такого же BRV IND. Спасенные фаги инкубировали с BRV IND либо непосредственно, либо с предварительно улавливаемыми и промытыми BRV IND, и связанные фаговые частицы элюировали 100 мМ триэтиламином при pH 10,0 и сразу же нейтрализовали, используя трис, pH 7,4. Элюированные фаги использовали для инфекции клеток TG1 в экспоненциальной фазе роста. После второго или третьего раунда биопэннинга выращивали отдельные колонии и соответствующие клоны VHH анализировали в ELISA фагов.
Экспрессия и очистка мономерных и димерных VHH согласно изобретению: кДНК VHH клонов, которые были позитивными в ELISA, повторно клонировали, используя ферменты рестрикции NcoI и NotI, в экспрессирующем векторе pHEN6 (публикации Con rath, K. E. M. et. al. Antimicrob. Agents Chemother. 45: 2807-12, 2001, включенная в настоящее описание исключительно в виде ссылки), который обеспечивает направляющую в периплазму последовательность pelB и шестигистидиновую метку на карбоксильном конце. Бивалентный VHH конструировали с помощью ПЦР-амплификации последовательности VHH, используя праймеры Bivfor2 (SEQ ID No. 20) и Bivrev2 (SEQ ID No. 21) (SEQ ID No. 22), которые кодируют линкер, родственный шарниру IgA человека. ПЦР-продукт и вектор pHEN6, содержащий матрицу VHH, расщепляли NcoI и PstI и лигировали, получая вектор pAO-biv, который содержит бивалентный VHH. Чтобы получить моновалентный или бивалентный VHH, клетки E. coli XL1 Blue трансформировали разными плазмидными конструкциями. Экспрессию VHH индуцировали 1 мМ изопропил-D-тиогалактопиранозидом в течение 16 часов при 27°C (Sambrook, J., and Russell, D.W., 2001, Molecular Cloning). После центрифугирования клеток периплазматические белки экстрагировали осмотическим шоком. VHH очищали из периплазматического экстракта, используя хелатирующую колонку N-High-Trap HP (Amersham Biosciences).
Пример 2: Характеристика VHH согласно изобретению
Вестерн-блот: Концентраты VP6, экспрессированного в бакуловирусной системе, и концентраты BRV IND ресуспендировали в буфере для образцов Лэммли, который кипятили в течение 10 минут. Затем их пропускали через колонку с 12% SDS-ПААГ и переносили на мембрану Immobilon P (Millipore, Berdford, MA). Мембрану блокировали в течение 45 минут, используя PBS/твин (0,05%), содержащий 10% обезжиренного молока, и каждый из VHH (4 мкг/мл) инкубировали в течение 2 часов при температуре окружающей среды. Затем мембрану промывали PBS/твин (0,05%) и инкубировали в течение ночи при 4°C с антителом против пентагистидина (разведение 1/500 в PBS/твин (0,05%), БСА (3%). Наконец их инкубировали с конъюгированным с HRP антителом козы против IgG мыши (разведение 1/5000) (Amersham, Pharmacia, Biotech) в течение 40 минут при температуре окружающей среды. Результаты анализов получали, используя ECL (Amersham Biosciences).
Секвенирование VHH согласно изобретению: Чтобы секвенировать VHH, использовали олигонуклеотиды «прямой M13» и «обратный M13», следуя указанному способу: готовый набор для реакции циклического секвенирования с использованием меченных красителем терминаторов (Applied Biosystem) в автоматическом секвенаторе ДНК ABI-Prism 377 (Perkin Elmer, Applied Biosystems).
Пример 3: Иммунологические анализы RV с использованием VHH согласно изобретению
Иммуноферментный анализ (ELISA) и Вестерн-блот: Эксперименты на основе ELISA осуществляли в 96-лучноных планшетах Maxisorp (Nunc) посредством прямой сенсибилизации RV или посредством улавливания RV или рекомбинантного VP6 поликлональным антителом, полученным у гнотобиологической свиньи. В качестве антигенов для негативного контроля использовали псевдоинфицированные клетки MA-104 и неродственный белок, экспрессированный в бакуловирусах (белок E2 вируса бычьей диареи). PBS использовали в качестве пустого контроля, и сыворотку неиммунизированной морской свинки использовали в качестве контроля негативного улавливания.
Присутствие анти-RV-антител в сыворотке лам анализировали, как описано в Parreno, V.C. et. al., Vet. Immunol. Immunopathol. 100: 7-24, 2004, и присутствие анти-VP6-антител анализировали, используя адаптированный протокол, описанный Fernandez et al. (Fernandez, F.M. et al., Vaccine 16: 507-16, 1998). IgG ламы выявляли, используя меченное пероксидазой антитело козы против IgG (H+L) ламы (Bethyl, lab inc, Montgomery, CA, USA) в разведении 1/2000. Фаги, полученные из отдельных клонов, полученных в результате биопэннинга, анализировали в ELISA фагов. Коротко, отдельные клоны E. coli TG1 в экспоненциальной фазе роста, содержащие разные гены VHH в векторе phen4, инфицировали хелперными фагами M13, чтобы получить фаговые частицы, которые экспрессируют VHH, слитый с поверхностным белком, и анализировали надосадок культуры, содержащий фаговое потомство, в планшетах для ELISA, сенсибилизированных BRV IND или VP6. Связанные фаги выявляли, используя конъюгированное с HRP анти-M13p8-антитело (Amersham, Pharmacia, Biotech) в разведении 1/5000, в течение 40 минут при температуре окружающей среды. Анализы регистрировали, используя H2O2/ABTS (Zymed).
Во-первых, моновалентные или бивалентные VHH, очищенные с использованием 6-His-метки на карбоксильном конце, исследовали в качестве реагентов для выявления RV или VP6 в ELISA, как описано выше, регистрируя результаты с использованием моноклонального антитела против пентагистидина (Qiagen, 1/5000) и конъюгированное с HRP антитело козы против Ig мыши. Во-вторых, их анализировали в качестве улавливающего RV реагента как посредством прямой сенсибилизации планшета для ELISA с использованием 10 мкг/мл VHH, так и посредством улавливания с помощью 10 мкг/мл моноклонального антитела против гистидина и затем 20 мкг/мл VHH. Результаты анализов регистрировали, используя поликлональную антисыворотку против RV, полученную от не получавшего молозива теленка, гипериммунизированного BRV IND (разведение 1/2000) и меченного пероксидазой антитела против IgG (H+L) быка (KPL, Gaitherburg, Maryland, USA) в разведении 1/5000.
Димерный VHH тестировали в ELISA в качестве реагента, улавливающего RV, в концентрации 10 мкг/мл.
Мономерный VHH также анализировали в качестве вторых антител, и результаты ELISA регистрировали, используя моноклональные антитела против пентагистидина и конъюгированные с HRP антимышиные антитела козы (разведение 1:1000) (Amersham, Pharmacia, Biotech).
Анализы нейтрализации вирусов: Титры антител, нейтрализующих вирусы IND, C486, B223, Wa и H2, в образцах сыворотки лам, и очищенные VHH определяли, используя нейтрализацию фокусов флуоресценции (FFN), как описано в To, T.L. et al. (J. Gen. Virol. 79 (Pt 11): 2661-72, 1998). Коротко, 100 мкл серийных разведений сыворотки ламы, выбранных очищенных мономеров или димеров VHH смешивали с равными объемами вируса, чтобы получить 100 фокус-образующих единиц (ФОЕ)/100 мкл смеси, и инкубировали в течение одного часа при 37°C. 100 мкл смеси антитело-вирус высевали на монослой MA-104 (4 повтора) и инкубировали в течение 48 часов при 37°C. Планшеты фиксировали 70% ацетоном и результаты анализа регистрировали, используя ФИТЦ-меченое анти-RV-антитело не получавшего молозива теленка, гипериммунизированного RV. Титр VN выражали в виде обратной величины наибольшего разведения образца, которое приводит к >80% уменьшению фокусов флуоресценции.
Пример 4: Применение мономерных и димерных VHH согласно изобретению для профилактики и/или лечения млекопитающих
Анализ защиты от RV у новорожденных мышей:
100 мкг каждого мономера анти-VP6-VHH в 100 мкл вводили четырехдневным мышам Balb/c, используя внутрижелудочный зонд, один раз в сутки, начиная с 0 дня и в течение 5 суток. Лактирующих мышей заражали, используя 100 мкл BRV C486 (SbI; P[1]G6), содержащих 2×106 ФОЕ/мл, в 1 день через 2 часа после обычной дозы VHH и затем вводили 20 мкл 5% раствора бикарбоната также внутрижелудочным путем. Инокулят был способен вызывать диарею у 100% необработанных контрольных мышей. В эксперименте использовали следующие контрольные группы: (i) мыши, которым инокулировали RV и которых не обрабатывали антителами; ii) мыши, обработанные таким же количеством неродственного VHH, направленного против клеточного белка; iii) мыши, обработанные 450 мкг аффинно очищенного IgG, полученного из поликлональной антисыворотки морской свинки с титром VN 2048 против гомологичного RV; iv) мыши, обработанные таким же количеством IgG серонегативной контрольной морской свинки; v) мыши, которых не инфицировали и не обрабатывали. RV-индуцированную диарею клинически оценивали прямой пальпацией живота мышей в течение 5 дней исследования. Тяжесть диареи анализировали ежедневно, используя количественное значение, основанное на окраске и консистенции стула, как описано в VanCott, J. L. et al., J. Virol. 80: 4949-61, 2006. Точный критерий Фишера использовали для сравнения доли мышей с диареей в группах. Непараметрический критерий Крускала-Уоллиса использовали для сравнения среднего значения начала, продолжительности и тяжести диареи в обработанных группах.
Пример 5: Создание рекомбинантного бакуловируса
Рекомбинантный бакуловирус BacMeIVHH создавали на основе плазмиды phen 6, содержащей полную последовательность VHH 3B2. Белок амплифицировали в ПЦР с плазмиды phen 6, используя следующие праймеры: SEQ ID NO: 23 и SEQ ID NO: 24. Затем указанный ампликон клонировали в векторе pFastMeIB2 в рамке с сигнальной последовательностью насекомых, полученной из мелитина медоносной пчелы, используя сайты рестрикции BamHI и XbaI, включенные в соответствующие праймеры. Полученную плазмиду pFBMeIVHH характеризовали с помощью автоматизированного секвенирования и использовали для создания рекомбинантного бакуловируса BacMeIVHH с применением бакуловирсной системы Bac-to-Bac® (Invitrogen, USA), следуя инструкциям производителя. Рекомбинантный бакуловирус размножали и амплифицировали в клетках насекомых sf21, чтобы достичь титров инфекционности от 107 до 109 БОЕ/мл, и исходные культуры хранили при 4°C в случае ежедневного применения и при -80°C в случае долговременного хранения.
Условия роста насекомых и инокуляция
Для экспериментов по исследованию экспрессии личинкам пятой возрастной стадии (личинки последней возрастной стадии перед окукливанием) массой примерно 250 мг инъецировали рекомбинантные бакуловирусы вблизи ложной ножки (по направлению к полости тела), используя известные дозы БОЕ/личинку. Инфицированных личинок содержали в камерах для роста при 28°C и собирали в указанное время. Затем личинок сразу замораживали и хранили при -20°C вплоть до обработки.
Культуры клеток насекомых также инфицировали, используя известные дозы, инфицированные клетки выдерживали при 28°C в течение 72 часов. Наконец инфицированные культуры собирали и осадки клеток также немедленно замораживали и хранили при -20°C вплоть до обработки.
Получение экстрактов белка
Суммарные растворимые белки (TSP) из личинок T. ni получали, измельчая замороженные личинки в буфере для экстракции, содержащем тритон 0,01%; DTT 25 мМ и смесь ингибиторов белка (полная, Roche, Germany) в 1X PBS.
Анализ экстрактов белка
Окрашивание Кумасси синим и анализ с помощью иммуноблоттинга осуществляли для количественной оценки и выявления специфичного белка VHH, входящего в состав TSP. Таким образом, 20 мкг TSP наносили на дорожку 12% SDS-полиакриламидного геля. После электрофореза гели красили раствором Кумасси синего или переносили на нитроцеллюлозные мембраны (Schleicher and Schuell) для осуществления Вестерн-блот-анализа.
Для Вестерн-блот-анализа SDS-ПААГ (12%) переносили на нитроцеллюлозную мембрану (Bio-Rad, USA). Мембрану блокировали в течение ночи при 4°C в PBS-0,05% твин-20 (PBST) с 4% обезжиренного молока (блокирующий буфер, BF) и затем инкубировали при комнатной температуре (КТ) в течение 1 часа, используя анти-VHH-сыворотку кролика (1:100 в BF). Мембрану 3 раза промывали PBST и, наконец, добавляли конъюгированное с HPR антитело против IgG кролика (1:2000 в BF, Sigma, USA) на 1 час в качестве второго антитела. После тщательной промывки PBST полосы белка выявляли, используя систему регистрации для Вестерн-блоттинга ECL на пленках Hyperfilm ECL (Amersham, USA) (смотри фигуру 6A).
Функциональный анализ
Экстракты TSP из личинок, экспрессирующих VP6 (белок бычьего ротавируса) [Аг(+)], или из инфицированных личинок без встроенного рекомбинантного бакуловируса [Аг(-)] использовали для покрывания микропланшетов для ELISA (Polysorp, Nunc, Denmark) серийными разведениями по 40 мкг/лунку в 50 мМ карбонатном/бикарбонатном буфере, pH 9,6 и инкубировали в течение ночи при 4°C. На следующий день планшеты четыре раза промывали PBST. Планшеты последовательно инкубировали в течение 1 часа при 37°C при постоянном встряхивании с блокирующим раствором (PBST-2% БСА, 100 мкл/лунку) в течение 30 минут, затем с экстрактами TSP из личинок, экспрессирующих VHH, по 2,5 мкг/лунку в блокирующем буфере в течение 1 часа. Затем планшеты 4 раза промывали PBST и снова блокировали в течение 30 минут. Затем добавляли 100 мкл/лунку поликлонального антитела (в разведении 1:100) против VHH, полученного у кролика, и инкубировали в течение 1 часа при 37°C. Планшеты 4 раза промывали PBST. Наконец добавляли 100 мкл/лунку конъюгированного с HRP антитела против IgG кролика в разведении 1:2000 в блокирующем растворе. Для реакции с субстратом планшеты четыре раза промывали и в планшеты добавляли 100 мкл/лунку 1 мМ 2,2'-азино-бис(3-этилбензотиазолин-6-сульфоновой кислоты) ABTS, (KPL, USA). Пероксидазной реакции давали возможность протекать в течение 5-10 минут при комнатной температуре, и реакции регистрировали при 405 нм в считывающем устройстве для микропланшетов для ELISA (Multiskan EX, Thermo Electron Corp, USA) (см. фиг. 7).
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения аминокислотных последовательностей, кодирующих наноантитела, нейтрализующие токсин А Clostridioides difficile | 2023 |
|
RU2833930C1 |
| ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ РОТАВИРУСНОЙ ИНФЕКЦИИ, СОДЕРЖАЩАЯ ГИБРИДНЫЙ БЕЛОК В КАЧЕСТВЕ АКТИВНОГО АГЕНТА (ВАРИАНТЫ) | 2013 |
|
RU2539913C2 |
| Вирусоподобные химерные частицы для индукции специфического иммунитета против вируса тяжелого острого респираторного синдрома SARS-CoV-2, содержащие белки коронавируса и ротавируса | 2022 |
|
RU2779810C1 |
| ИММУНОГЛОБУЛИН С ОДНИМ ВАРИАБЕЛЬНЫМ ДОМЕНОМ ПРОТИВ F-БЕЛКА PCB | 2016 |
|
RU2730671C2 |
| ТЕРАПЕВТИЧЕСКИЕ ПОЛИПЕПТИДЫ, ИХ ГОМОЛОГИ, ИХ ФРАГМЕНТЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ МОДУЛЯЦИИ АГРЕГАЦИИ, ОПОСРЕДОВАННОЙ ТРОМБОЦИТАМИ | 2004 |
|
RU2524129C2 |
| РОТАВИРУСНЫЕ ЧАСТИЦЫ С ХИМЕРНЫМИ ПОВЕРХНОСТНЫМИ БЕЛКАМИ | 2014 |
|
RU2698049C2 |
| Однодоменное антитело ламы Н5 и его производное H5-Fc, специфически связывающие RBD-домен S-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2022 |
|
RU2793967C1 |
| РЕКОМБИНАНТНОЕ ОДНОДОМЕННОЕ АНТИТЕЛО, СПОСОБНОЕ СПЕЦИФИЧЕСКИ СВЯЗЫВАТЬ ФАКТОР НЕКРОЗА ОПУХОЛЕЙ ЧЕЛОВЕКА, И ЕГО ПРОИЗВОДНЫЕ | 2012 |
|
RU2530553C2 |
| Иммунобиологическое средство на основе вирусоподобных частиц для индукции специфического иммунитета против инфекции, вызываемой ротавирусом А человека | 2022 |
|
RU2795055C1 |
| Рекомбинантное антитело, специфичное к фактору некроза опухоли и маркеру иммунных клеток миелоидного ряда | 2016 |
|
RU2652876C1 |








Настоящее изобретение относится к области иммунологии. Предложены мономерные и димерные домены VHH, полученные из антител верблюдовых, способные связывать антиген VP6 ротавируса группы А, а также способ выявления ротавируса группы А, композиция для придания пассивного иммунитета в отношении ротавируса группы А, вектор, клетки и способ получения домена VHH по изобретению и набор для иммунологического выявления ротавируса группы А. Данное изобретение может найти дальнейшее применение в профилактике и лечении ротавирусных инфекций. 11 н. и 3 з.п. ф-лы, 7 ил., 5 табл., 5 пр.
1. Мономерный домен VHH, способный связывать антиген VP6 ротавируса группы А, отличающийся тем, что он содержит последовательность, выбранную из группы, состоящей из последовательностей SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3 и SEQ ID No. 4.
2. Димерный домен VHH, способный связывать антиген VP6 ротавируса группы А, отличающийся тем, что он содержит
(а) две последовательности, либо идентичные, либо разные, выбранные из группы, состоящей из последовательностей SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3 и SEQ ID No. 4; или
(b) последовательность, выбранную из группы, состоящей из SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11 и SEQ ID No. 12.
3. Димерный домен VHH по п.2, отличающийся тем, что он содержит связывающую последовательность SEQ ID No. 13.
4. Способ иммунологического выявления ротавируса А, отличающийся тем, что он включает: a) осуществление контакта содержащего ротавирус образца с мономерным или димерным доменом VHH по п.1 или 2 соответственно; и b) обработку для выявления результатов.
5. Способ по п.4, отличающийся тем, что он выбран из группы, состоящей из основанных на иммунологическом улавливании тестов ELISA, ELISPOT, конкурентного ELISA, теста с использованием магнитных шариков или любого другого хроматографического способа иммуноанализа.
6. Композиция, предназначенная для придания пассивного иммунитета в отношении ротавируса группы А млекопитающему, отличающаяся тем, что она содержит эффективное количество мономерного или димерного домена VHH по п.1 или 2 соответственно; эксципиент и иммуномодуляторы.
7. Клетка для экспрессирования домена VHH, содержащая плазмиду pFBMelVHH, кодирующую VHH 3B2 (SEQ ID No. 4), депонированная в Испанской Коллекции Типов Культур (СЕСТ) с номером доступа СЕСТ7431.
8. Экспрессионный вектор, отличающийся тем, что он содержит нуклеотидную последовательность, которая кодирует любой из мономерных или димерных доменов по пп.1-3.
9. Вектор по п.8, отличающийся тем, что указанный вектор представляет собой вирус, в частности бакуловирус.
10. Трансгенная клетка для экспрессии мономерного или димерного домена VHH по любому из пп.1-3, трансформированная, трансфицированная или инфицированная вектором по п.8 или 9.
11. Способ получения доменов по пп.1-3, который включает следующие стадии:
a) трансформацию, или трансфекцию, или инфекцию клеток-хозяев в культуре или в личинке насекомого рекомбинантным вектором по п.8 или 9;
b) поддержание роста клеток в соответствующих условиях;
c) выделение и очистку доменов.
12. Набор для иммунологического выявления ротавируса группы А, содержащий мономерный или димерный домен VHH по п.1 или 2 соответственно.
13. Мономерный домен VHH, содержащий последовательность, выбранную из группы: SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3, SEQ ID No. 4, для применения в способе профилактики или лечения инфекций, вызываемых ротавирусом группы А.
14. Димерный домен VHH, содержащий
(а) две последовательности, либо идентичные, либо разные, выбранные из группы: SEQ ID No. 1, SEQ ID No. 2, SEQ ID No. 3 и SEQ ID No. 4; или
(b) последовательность, выбранную из группы: SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11 и SEQ ID No. 12,
для применения в способе профилактики или лечения инфекций, вызываемых ротавирусом группы А.
| CORTHESY B | |||
| et al | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| LOPEZ S | |||
| et al | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИРОТАВИРУСНОГО ИММУНОГЛОБУЛИНА | 1998 |
|
RU2129017C1 |

