ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к некоторым неразветвленным карбаматным производным некоторых производных Wnt5-α, а также к лечению меланомы с использованием этих производных Wnt5-α.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Меланома является злокачественной опухолью меланоцитов, которые встречаются преимущественно в коже, но также в кишечнике и глазу. Она представляет собой один из наиболее редко встречающихся типов рака кожи, но является причиной большинства связанных с раком кожи смертельных исходов. Злокачественная меланома является потенциально опасным типом рака кожи. Это обусловлено неконтролируемым ростом пигментных клеток, называемых меланоцитами. Несмотря на многие годы интенсивного лабораторного и клинического исследования единственным эффективным методом лечения является хирургическое удаление первичной опухоли до достижения ею толщины, большей, чем 1 мм.
Приблизительно 160000 новых случаев меланомы диагностируется во всем мире каждый год, и она наиболее часто встречается у мужчин и представителей европеоидной расы. Она более распространена в европейских популяциях, проживающих в солнечном климате, чем в других группах. В соответствии с сообщением ВОЗ примерно 48000 связанных с меланомой смертельных исходов наблюдается во всем мире в год. Злокачественная меланома является причиной 75 процентов всех смертельных исходов, связанных с раком кожи.
Лечение включает хирургическое удаление опухоли; адъювантное лечение; химио- и иммунотерапию или лучевую терапию.
Риск развития меланомы зависит от двух групп факторов: внутренних и обусловленных окружающей средой. «Внутренние» факторы обычно представляют собой семейный анамнез индивидуумов и унаследованный генотип, тогда как наиболее важным фактором окружающей среды является солнечная экспозиция. Эпидемиологические исследования указывают на то, что экспозиция ультрафиолетового излучения (УФА и УФВ) является одним из основных факторов, способствующих развитию меланомы. УФ-излучение вызывает повреждение в ДНК клеток, в основном димеризацию тимина, которая в отсутствие репарации может приводить к возникновению мутаций. Если клетки делятся, эти мутации передаются в новые поколения клеток. Если мутации происходят в онкогенах или в генах опухолевых супрессоров, скорость митоза в несущих мутацию клетках может становиться неконтролируемой, приводя к образованию опухоли. Случайная чрезмерная солнечная экспозиция (приводящая к «солнечному ожогу») причинно связана с меланомой.
Возможные значимые элементы в определении риска включают интенсивность и продолжительность солнечной экспозиции, возраст, при котором происходит солнечная экспозиция, и степень пигментации кожи. Экспозиция в период детства является более важным фактором риска, чем экспозиция в период зрелого возраста. Это видно из миграционных исследований в Австралии, где жители проявляют тенденцию к сохранению профиля риска своей страны рождения, если они мигрировали в Австралию в зрелом возрасте. Индивиды с солнечными ожогами в виде волдырей или шелушением кожи (особенно в первые двадцать лет жизни) имеют значительно больший риск развития меланомы. Это не означает, что солнечный ожог является причиной меланомы. Наоборот, он просто статистически коррелирует. Причиной является чрезмерная УФ-экспозиция. Было показано, что солнцезащитный крем несмотря на то что предотвращает солнечный ожог, не защищает от меланомы. Многие исследователи утверждают, что солнцезащитный крем может даже увеличить риск развития меланомы. Русые и рыжеволосые жители, персоны с множеством атипичных невусов или с диспластическими невусами и персоны, родившиеся с гигантскими врожденными меланоцитарными невусами, имеют повышенный риск развития меланомы.
Заболеваемость меланомой увеличилась в последние годы, но не ясно, какой объем изменений в поведении, в окружающей среде или в раннем выявлении вовлечены в этот процесс.
Для понимания того, как солнцезащитный крем может уменьшить солнечный ожог и в то же время вызвать развитие меланомы, необходимо различать прямое повреждение ДНК и непрямое повреждение ДНК. Генетический анализ показывает, что 92% всех меланом вызвано непрямым повреждением ДНК. Семейная меланома является генетически гетерогенной, и локусы семейной меланомы были идентифицированы на хромосомных плечах 1p, 9p и 12q.
Признаками и симптомами меланомы являются:
асимметричное повреждение кожи;
граница повреждения неоднородная;
меланомы обычно являются многоцветными;
родинки, большие чем 5 мм, более вероятно являются меланомами, чем родинки меньшего размера.
Эволюция (т.е. изменение) родинки или повреждения может являться указанием на то, что повреждение становится злокачественным.
Наиболее распространенные типы меланомы в коже представляют собой:
поверхностно генерализованную меланому (SSM);
узловатую меланому;
акральную лентигинозную меланому;
злокачественное лентиго (меланома) [меланому типа злокачественного лентиго].
Любая из вышеупомянутых типов может вырабатывать меланин (и являться темной по цвету) или нет (и является амеланотической - нетемной). Аналогично любой подтип может проявлять десмоплазию (реакция плотных волокон с нейротропизмом), которая является маркером агрессивного поведения и тенденции к местному рецидиву.
Другие:
светлоклеточная саркома (меланома мягких тканей);
меланома слизистой;
увеальная меланома.
Особенности, которые влияют на прогноз, представляют собой толщину опухоли в миллиметрах (толщина по Бреслоу), глубину, связанную со структурой кожи (уровень Кларка), тип меланомы, наличие изъязвлений, наличие лимфатической/перинейральной инвазии, наличие опухоль-инфильтрующих лимфоцитов (если имеются, прогноз улучшается), место поражения, наличие сопутствующих поражений и наличие региональных или отдаленных метастазов.
Некоторые типы меланомы имеют плохие прогнозы, но это объясняется их толщиной. Интересно, что менее инвазивные меланомы даже с метастазами в лимфатический узел влекут за собой более лучший прогноз, чем глубокие меланомы без региональных метастазов во время стадии распространения опухоли. Локальные рецидивы имеют тенденцию к поведению, сходному с первичной опухолью, если только они не находятся в областях широкого локального иссечения (в отличие от многоэтапного иссечения или иссечения путем вырубания/срезания), так как такие рецидивы проявляют тенденцию к лимфатической инвазии.
Когда меланомы распространяются в лимфатические узлы, один из наиболее важных факторов представляет собой число узлов со злокачественностью. Степень злокачественности в узле также важна; микрометастазы, у которых злокачественность носит лишь микроскопический характер, имеют более благоприятный прогноз, чем в случае макрометастазов. В некоторых случаях микрометастазы могут быть определены только посредством специального окрашивания, и, если злокачественность определяется только посредством более редко используемого теста, известного как полимеразная цепная реакция (ПЦР), прогноз становится лучше. Макрометастазы, для которых злокачественность клинически очевидна (в некоторых случаях рак полностью замещает узел), имеют значительно худший прогноз, и, если узлы спаяны, или если они имеют внекапсульное протяжение, прогноз становится еще более плохим.
Когда они представляют собой отдаленные метастазы, рак обычно считается неизлечимым. Пятилетняя частота выживания составляет менее чем 10%. Средняя выживаемость 6-12 месяцев. Лечение является паллиативным, сфокусированным на продлении жизни и качестве жизни. В некоторых случаях пациенты могут жить многие месяцы или даже годы с метастазирующей меланомой (в зависимости от агрессивности лечения). Метастазы в кожу и легкие имеют более лучший прогноз. Метастазы в головной мозг, кость и печень ассоциированы с худшим прогнозом.
Меланома проявляется на различных стадиях, которые обозначаются как Стадия 0, которая представляет собой меланому in situ, имеющая 100% выживаемость, Стадия I/II, которая представляет собой инвазивную меланому, имеющая 85-95% выживаемость, Стадия II, которая представляет собой меланому высокого риска, имеющая 40-85% выживаемость, Стадия III, которая представляет собой региональный метастаз, имеющая 25-60% выживаемость, Стадия IV, которая представляет собой отдаленный метастаз, имеющая 9-15% выживаемость, основанную на AJCC 5-летней выживаемости при надлежащем лечении.
Хирургия является способом лечения первого выбора для локализованной кожной меланомы. Также имеется зависимость от стадии выполненной биопсии сторожевого лимфатического узла, хотя существует разногласие по опытным данным этой процедуры. Лечение прогрессирующей злокачественной меланомы проводится на основе многопрофильного подхода.
Меланомы высокого риска могут потребовать адъювантного лечения. В Соединенных Штатах большинство пациентов в остальном с хорошим здоровьем будут начинать с года лечения высокой дозой интерферона, которая имеет тяжелые побочные эффекты, но может улучшить прогноз для пациента. Это требование не поддерживается всей научной работой на данный момент, и в Европе интерферон обычно не используется вне объема клинических испытаний.
Используются разные химиотерапевтические агенты, включая дакарбазин (также названный как DTIC), иммунотерапия (с использованием интерлейкина-2 (IL-2) или интерферона (IFN)), а также локальная перфузия используется различными центрами. Они могут иногда показывать значительный успех, но в целом благоприятный исход при метастазирующей меланоме довольно ограничен. IL-2 (Пролейкин®) является первой новой терапией, одобренной для лечения метастазирующей меланомы в течение 20 лет. Исследования показывают, что IL-2 дает возможность полной и продолжительной ремиссии этого заболевания, хотя только у небольшого процента пациентов. Ряд новых агентов и новых подходов находятся на экспертизе и являются перспективными.
Лучевая терапия часто используется после хирургической резекции для пациентов с локально или регионально прогрессирующей меланомой, или для пациентов с нерезецируемыми отдаленными метастазами. Она может снижать частоту локальных рецидивов, но не увеличивает выживаемость.
Была интенсивно исследована молекулярная основа меланомной прогрессии, и анализ экспрессии генов идентифицировал несколько генов, дифференциально экспрессируемых в инвазивных формах меланомы по сравнению с менее инвазивной меланомой или доброкачественным невусом, один такой ген представляет собой Wnt-5a (Bittner et al., 2000). Wnt-5a является секретируемым, обогащенным цистеином белком, который подвергается посттрансляционному гликозилированию и липидным модификациям (Kurayoshi et al., 2007). После своей секреции Wnt-5a действует ауто- или паракринным образом путем связывания со своим рецептором, было показано, что в злокачественной меланоме Wnt-5a связывается с сопряженным с G-белком рецептором Frizzled-5 (Weeraratna, 2002). Он считается как не канонический белок Wnt, указывая на то, что он не действует первоначально через сигнальный путь β-катенина. Значение Wnt-5a в прогрессии рака было исследовано на различных типах рака в течение последних лет. Было показано, что Wnt-5a проявляет активность опухолевого супрессора при раке молочной железы, раке щитовидной железы, лимфоме, нейробластоме, раке толстой кишки и раке печени (Jönsson 2002; Kremenevskaja 2005; Liang 2003; Blanc 2005; Dejmek 2005; Liu 2008). Однако в других типах рака, подобного злокачественной меланоме и раку желудка, было показано, что увеличенная экспрессия Wnt-5a стимулирует опухолевую прогрессию (Bittner et al., 2000, Weeraratna, 2002; Lewis et al., 2005; Kurauoshi et al., 2006). На основании этих результатов можно заключить, что в некоторых раках вещество, имитирующее эффекты Wnt-5a, может служить ингибитором опухолевой прогрессии (Säfholm, 2006), тогда как в других раках, подобных злокачественной меланоме, ингибитор опосредованной Wnt-5a опухолевой прогрессии будет необходим.
В отношении нижележащих функциональных эффектов белка Wnt-5a в злокачественной меланоме доступны лишь ограниченные данные (Weeraratna, et al., 2002; Dissanayake et al., 2007). В клетках, полученных из образцов меланомной ткани, было показано, что увеличенная экспрессия Wnt-5a индуцирует усиление клеточной адгезии, миграции и инвазии. В том же исследовании авторы также показали, что эффекты Wnt-5a опосредовались через рецептор Frizzled-5 и сигнал нижележащей протеинкиназы C (PKC) (Weeraratna, 2002). В более поздней статье авторы далее показывают, что Wnt-5a индуцирует эпителиально-мезенхимный переход (EMT) при посредстве PKC-индуцированной экспрессии Snail, которая приводит к снижению уровня E-кадгерина, но увеличению уровня виментина (Dissanayake et al., 2007). Однако остается еще вопрос в отношении подлинной причины увеличенной экспрессии Wnt-5a в злокачественных меланомах.
В недавнем исследовании Hoek и соавторы на основании анализа ДНК-микрочипов предположили, что трансформирующий фактор роста-β (TGF-β) играет решающую роль в регуляции экспрессии гена Wnt-5a (Hoek et al., 2006). Весьма интересно, что члены суперсемейства TGF-β (Van Belle et al., 1996) и костного морфогенетического белка (BMP, Rothhammer et al., 2005) проявляют увеличенную экспрессию в злокачественной меланоме. Кроме того, по меньшей мере некоторые функциональные эффекты TGF-β также перекрываются с таковыми Wnt-5a. Конкретнее, как упоминалось выше для Wnt-5a, TGF-β1 индуцирует EMT и увеличивает в меланомной клетке миграцию и метастатический потенциал (Janji et al., 1999; Gouon et al., 1996). Наконец, Wnt-5a и TGF-β1 опосредуют изменения в клеточном белке уровней E-кадгерина, некоторых интегринов и матриксных металлопротеиназ (Dissanayake et al., 2007; Janji et al., 1999). Существуют публикации по нераковым системам, которые демонстрируют прямую связь между сигнальной системой TGF-β и экспрессией Wnt-5a. Например, было показано, что в мезенхимных клетках зачатка крыла курицы TGF-β3 увеличивает экспрессию Wnt-5a, приводя к активации PKCα и хондрогенной дифференцировке (Jin et al., 2006). В более недавней публикации на мышах показали, что TGF-β1 увеличивает экспрессию Wnt-5a в эпителиальных клетках молочной железы, приводя к ингибированию протокового вытяжения и бокового ветвления в развивающейся молочной железе (Roarty и Serra, 2007). Вследствие этого ингибирование сигнальной системы TGF-β1 может потенциально являться привлекательным механизмом, посредством которого Wnt-5a опосредует миграцию опухолевой клетки, и метастазирование может быть замедлено.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к производному Wnt-5a, используемому для лечения меланомы и рака желудка, а также к способу лечения меланомы и рака желудка.
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
В частности, изобретение относится к неразветвленным карбаматным производным, особенно N-бутоксикарбонильным производным некоторых пептидов Wnt-5a, и конкретнее к неразветвленному карбаматному производному, в частности N-бутоксикарбонильному производному одного или нескольких пептидов

Другой аспект по изобретению относится к неразветвленным карбаматным производным, в частности N-бутоксикарбонильному производному вышеупомянутых пептидов для применения в лечении меланомы и рака желудка.
Другой аспект по изобретению относится к фармацевтической композиции, содержащей по меньшей мере одно из неразветвленных карбаматных производных, в частности N-бутоксикарбонильное производное вышеупомянутых пептидов для применения в лечении меланомы и рака желудка.
В предпочтительном варианте осуществления изобретения фармацевтическая композиция представляет собой топическую композицию.
Другой аспект по изобретению относится к способу лечения меланомы путем введения терапевтически эффективного количества неразветвленного карбаматного производного, в частности N-бутоксикарбонильного производного вышеупомянутых пептидов пациенту, страдающему меланомой и раком желудка.
Другой аспект по изобретению относится к способу профилактического лечения меланомы путем введения терапевтически эффективного количества неразветвленного карбаматного производного, в частности N-бутилоксикарбонильного производного вышеупомянутых пептидов пациенту, находящемуся в зоне риска развития меланомы и рака желудка.
Термин “неразветвленное карбаматное производное” в настоящем описании означает одно из производных группы N-метилоксикарбонильного, N-этилоксикарбонильного, N-n-пропилоксикарбонильного или N-бутилоксикарбонильного производного, при этом последнее может быть предпочтительным.
Настоящее изобретение будет описано ниже путем ссылки на некоторые выполненные эксперименты.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
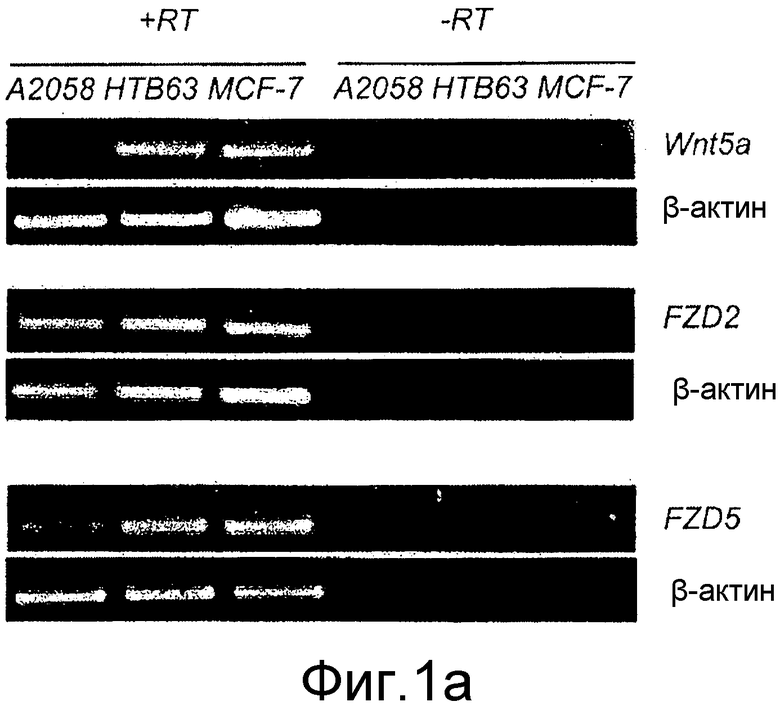
Фиг. 1 иллюстрирует характеристику меланомных клеточных линий A2058 и HTB63.
A) Анализ отсутствия или присутствия мРНК Wnt-5a, Frizzled-2 и Frizzled-5 в меланомных клетках A2058 и HTB63. Клеточная линия MCF-7 (M) рака молочной железы человека служила в качестве положительного контроля для всех этих транскриптов и β-актин - в качестве нагрузочного контроля. Плюс (+RT) и минус (-RT) указывают реакции, выполненные с и без обратной транскриптазы. Для Fzd2 и Fzd5 ПЦР-реакции выполняются с использованием в 3,5 раза большего количества кДНК, чем для β-актинового контроля.
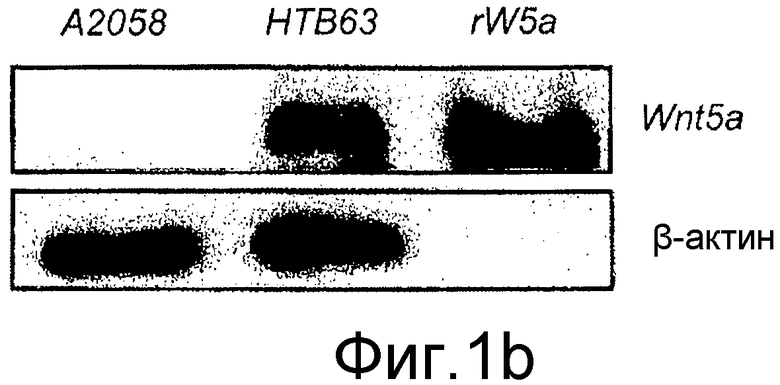
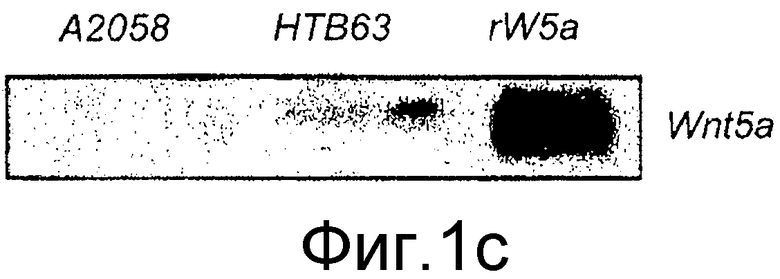
B) Для дальнейшей характеристики присутствия транскрипта Wnt-5a в клетках HTB63 изобретатель также определял с помощью Вестерн-блота клеточные уровни белка Wnt-5a в клетках A2058 и HTB63 с использованием рекомбинантного Wnt-5a (rW5a) в качестве положительного контроля и β-актина в качестве нагрузочного контроля. Изобретатель также выполнял Вестерн-блот анализ в бессывороточной культуральной среде, собранной из клеток A2058 и клеток HTB63 через 48 часов для выявления отсутствия или присутствия секретируемого из этих клеток белка Wnt-5a. Рекомбинантный белок Wnt-5a служил в качестве положительного контроля. Каждый из представленных результатов повторяли как независимые эксперименты по меньшей мере три раза.
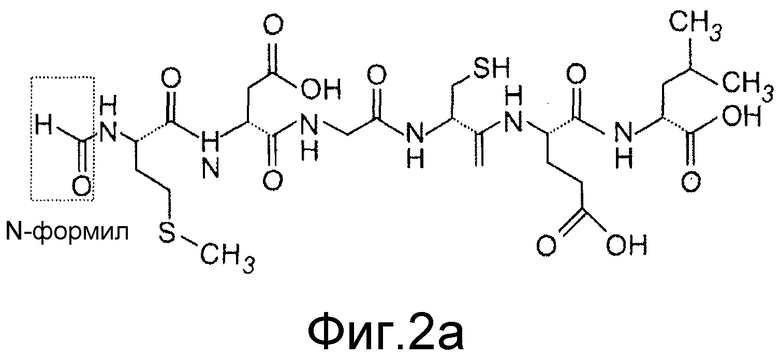
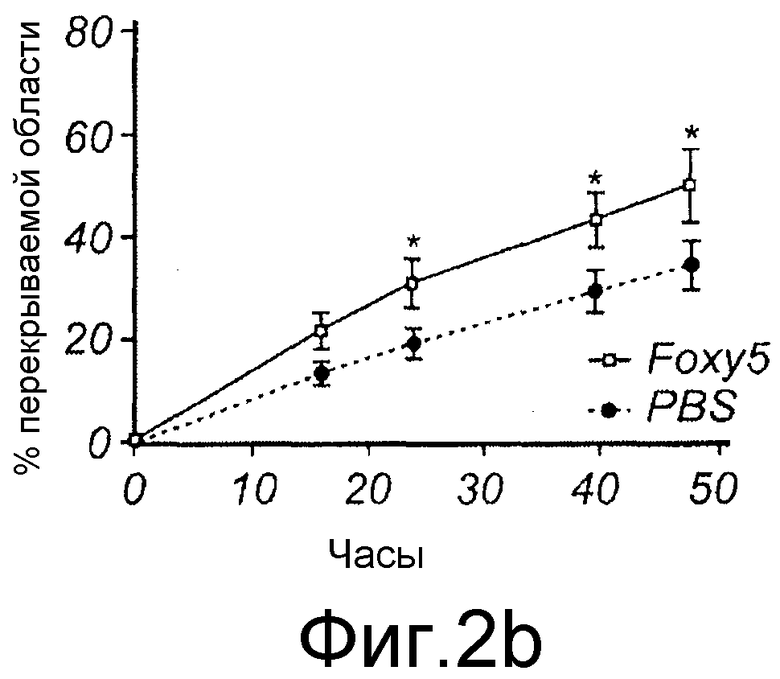
Фиг. 2 иллюстрирует Foxy5, который является Wnt-5a агонистом в меланомных клетках. A показывает структуру Foxy5 (формильная группа выделена). B показывает, что Foxy5 (50 мкМ) стимулирует миграцию клеток A2058 (анализ ранения-заживления) в пределах периода, составляющего 0, 16, 24, 40 и 48 часов. Планка погрешностей представлена s.e.m. Двусторонний критерий Стьюдента; *p<0,05.
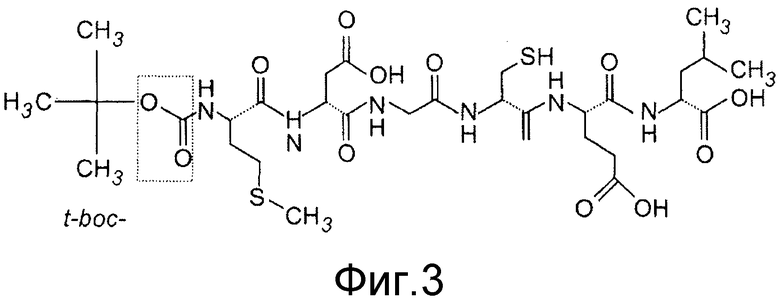
Фиг. 3 показывает структуру Box5, который является модифицированным аналогом Foxy5.
Фиг. 4 иллюстрирует эффекты Wnt-5a и нового N-бутилоксикарбонильного гексапептида, Box5, на адгезию и миграцию меланомных клеток.
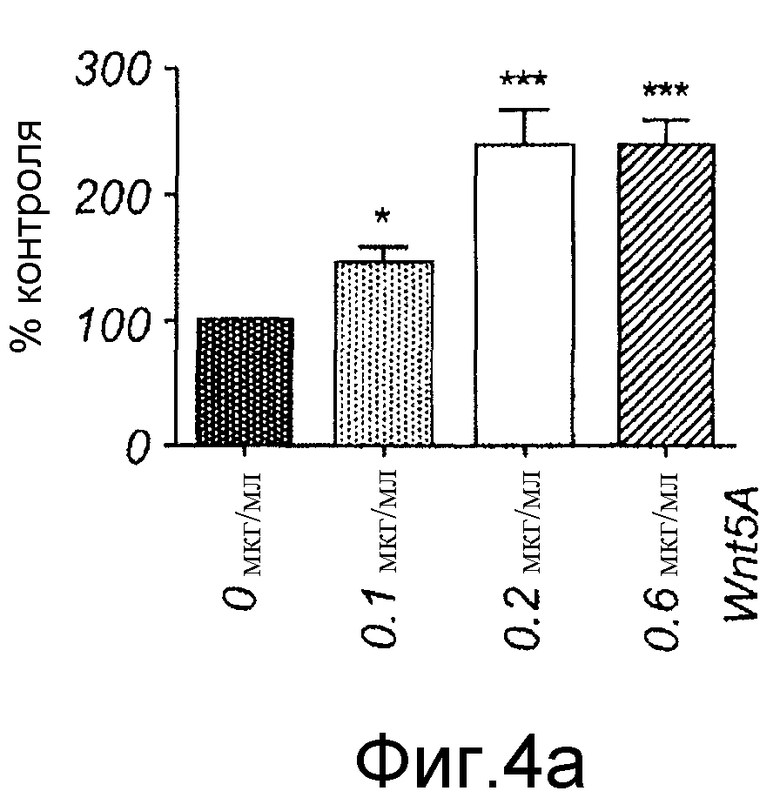
A) Меланомные клетки A2058 стимулировали указанными концентрациями Wnt-5a, разъединяли Версеном и ресуспендировали как одиночные клетки в бессывороточной среде в присутствии или в отсутствие рекомбинантного Wnt-5a в указанных концентрациях. Клеткам затем обеспечивали прилипание в 96-луночном планшете. По истечении периода 60 минут неприлипшие клетки смывали, в то время как прилипшие клетки окрашивали и определяли их число. Это число представлено как процент контрольной (без Wnt-5a) стимуляции.
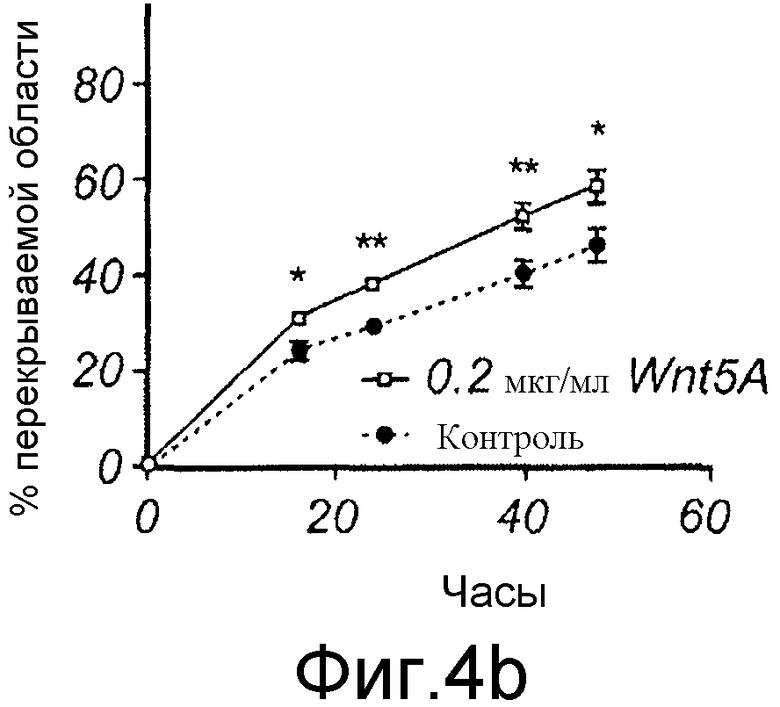
B) Меланомные клетки A2058 культивировали до слияния в 12-луночном планшете, после чего росчерком вносили в каждую лунку, среду заменяли на свежую бессывороточную среду, не содержащую (закрашенные кружки) или содержащую 0,2 мкг/мл Wnt-5a (незакрашенные квадраты).
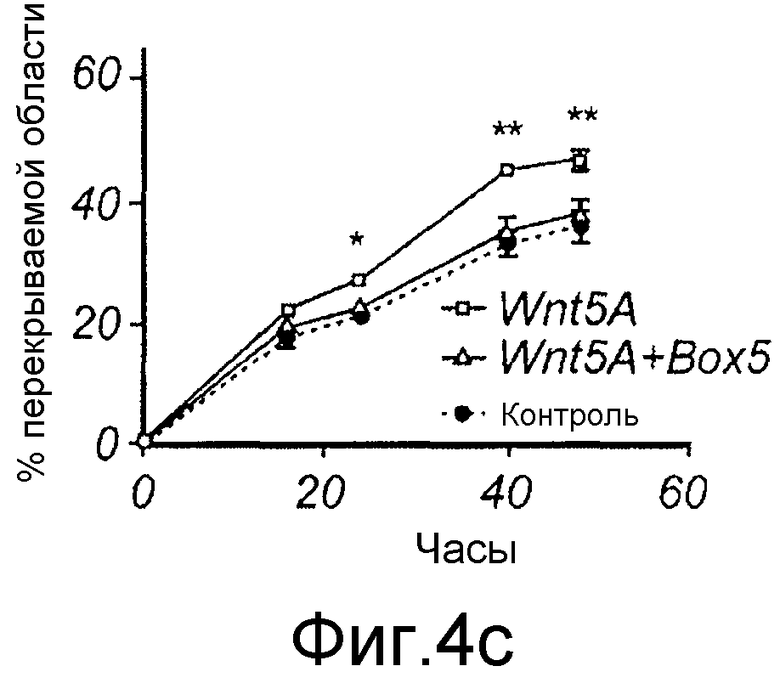
C) Меланомные клетки A2058 культивировали до слияния в 12-луночном планшете, после чего росчерком вносили в каждую лунку, среду заменяли на свежую бессывороточную среду в отсутствие любой добавки (закрашенный кружок), присутствии 0,2 мкг/мл Wnt-5a одного (незакрашенные квадраты) или в присутствии 0,2 мкг/мл Wnt-5a со 100 мкМ Box5 (незакрашенный треугольник).
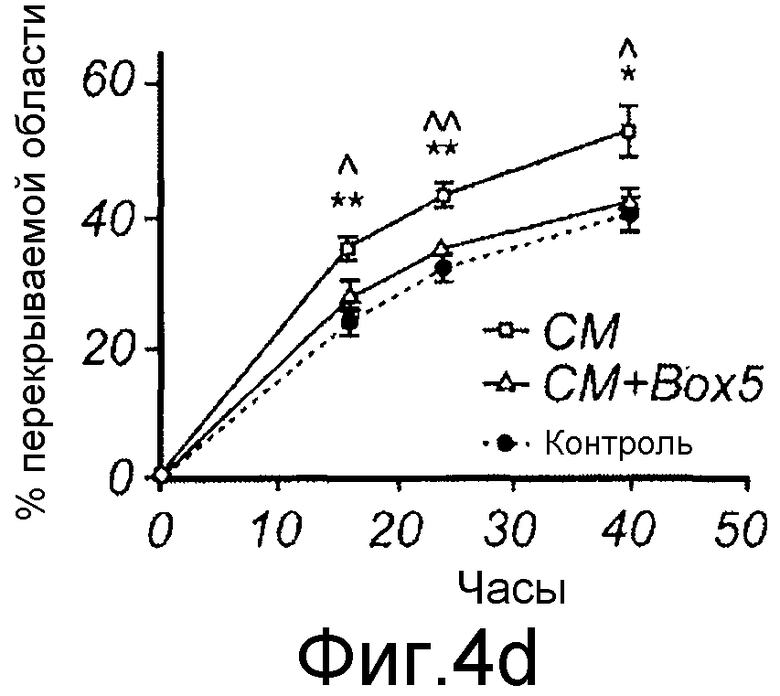
D) Меланомные клетки HTB63 культивировали до слияния в 12-луночной чашке, после чего росчерком вносили в каждую лунку, среду заменяли на свежую бессывороточную среду (закрашенный кружок), кондиционированную среду (незакрашенные квадраты) или кондиционированную среду с добавлением 100 мкМ Box5 (незакрашенный треугольник). Для регистрации изменений в миграции на панелях B-D картину учитывали с каждого росчерка/лунки из одной и той же области клеток спустя 0, 16, 24, 40 или 48 часов, и ранение-заживление выявляли как процент закрытия раневой поверхности.
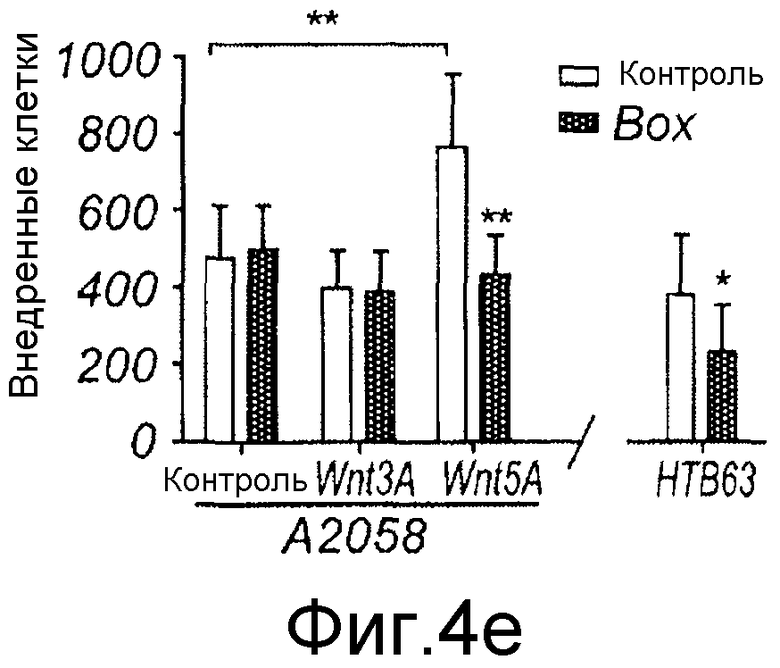
E) До начала каждого эксперимента клетки A2058 (шесть столбцов слева) и клетки HTB63 (два столбца справа) разъединяли Версеном и ресуспендировали как одиночные клетки в бессывороточной среде. Клетки предварительно инкубировали в течение 40 минут с использованием непрерывного встряхивания в отсутствие (незакрашенные столбцы) или присутствии (залитые краской столбцы) 100 мкМ Box5. Аликвоту клеточной суспензии, содержащую 25000 клеток, затем добавляли в верхнюю Transwell камеру и нижнюю камеру заполняли содержащей сыворотку (10%) средой. Как указывалось, 0,1 мкг/мл Wnt-3a, 0,2 мкг/мл Wnt-5a и/или 100 мкМ Box5 добавляли в верхнюю камеру. Клеткам затем давали возможность внедриться в течение 24 часов, после чего прикрепленные клетки на нижней стороне мембраны подсчитывали. Результаты даны как среднее ± SEM (n=5-7). *=p<0,05, **=p<0,01 и ***=p<0,001, где значения сравниваются с контролем (Фиг. 5D, CM против CM + Box5, ^=p<0,05, ^^=p<0,01 и ^^^=p<0,001).
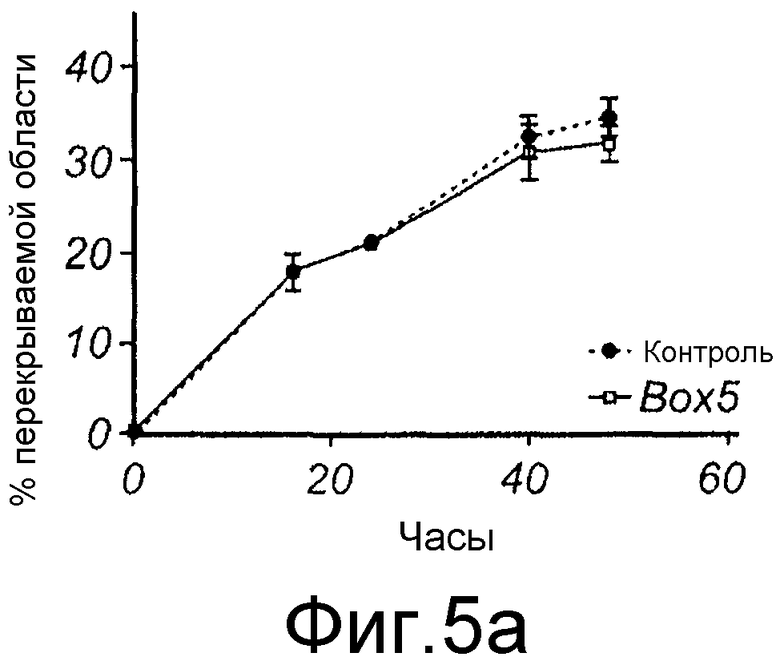
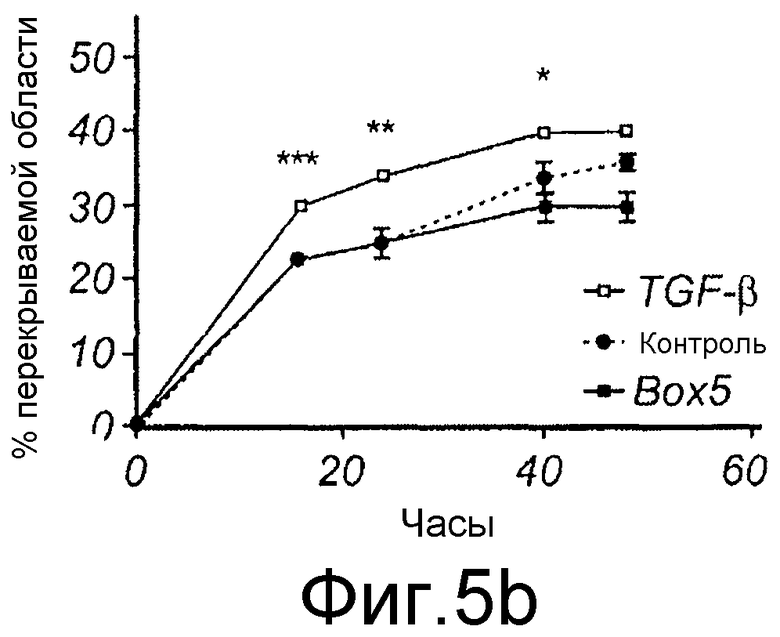
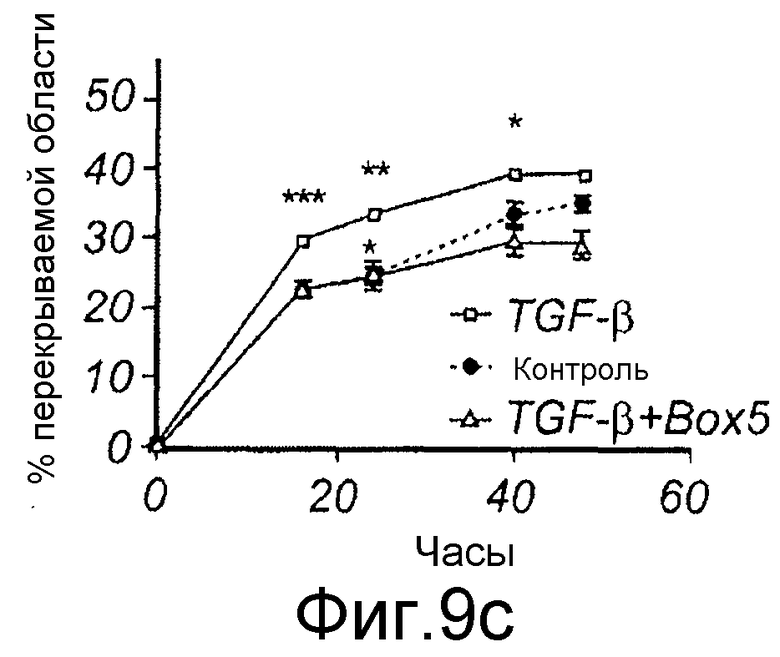
Фиг. 5 иллюстрирует, что Box5 не оказывает влияния на спонтанную миграцию меланомных клеток A2058, но может ингибировать индуцированную TGFβ1 миграцию. A) Анализ заживления раны клеток A2058 в присутствии (□) или в отсутствие (●) 100 мкМ Box5. B) Анализ заживления раны клеток A2058, предварительно инкубированных с или без 100 мкМ Box5 в течение 40 минут и затем далее стимулированных с или без 5 нг/мл TGFβ1, как указывалось. Все данные по заживлению раны выражаются как процент закрытия раневой поверхности через 0, 16, 24, 40 и 48 часов.
Фиг. 6 иллюстрирует, что сигнальный путь Wnt-5a/Ca2+ является необходимым для опосредованной Wnt-5a инвазии меланомных клеток. A) rWnt-5a (0,1 мкг/л; добавление указывается стрелкой) запускает быстрый цитозольный сигнал Ca2+ в клетках A2058. B) Предварительная инкубация клеток A2058 с 10 мкМ MAPT/AM в течение 30 минут отменяет rWnt-5a (0,1 мкг/мл) стимуляцию (показано стрелкой) цитозольного Ca2+. C) MAPT/AM отменяет индуцированную Wnt-5a клеточную инвазию A2058. Клетки предварительно инкубировали с 10 мкМ MAPT/AM в течение 30 минут, затем стимулировали с/без rWnt-5a (0,2 мкг/л), и затем с 1 мкМ MAPT/AM на всем протяжении эксперимента по инвазии (24 часа), где последнее условие обработки обладало таким же хелатирующим эффектом на Ca2+, как 10 мкМ MAPT/AM в течение 30 минут, показано на Фиг. 6A. Планка погрешностей представляет s.e.m. Двусторонний критерий Стьюдента; *p<0,05, ***p<0,001.
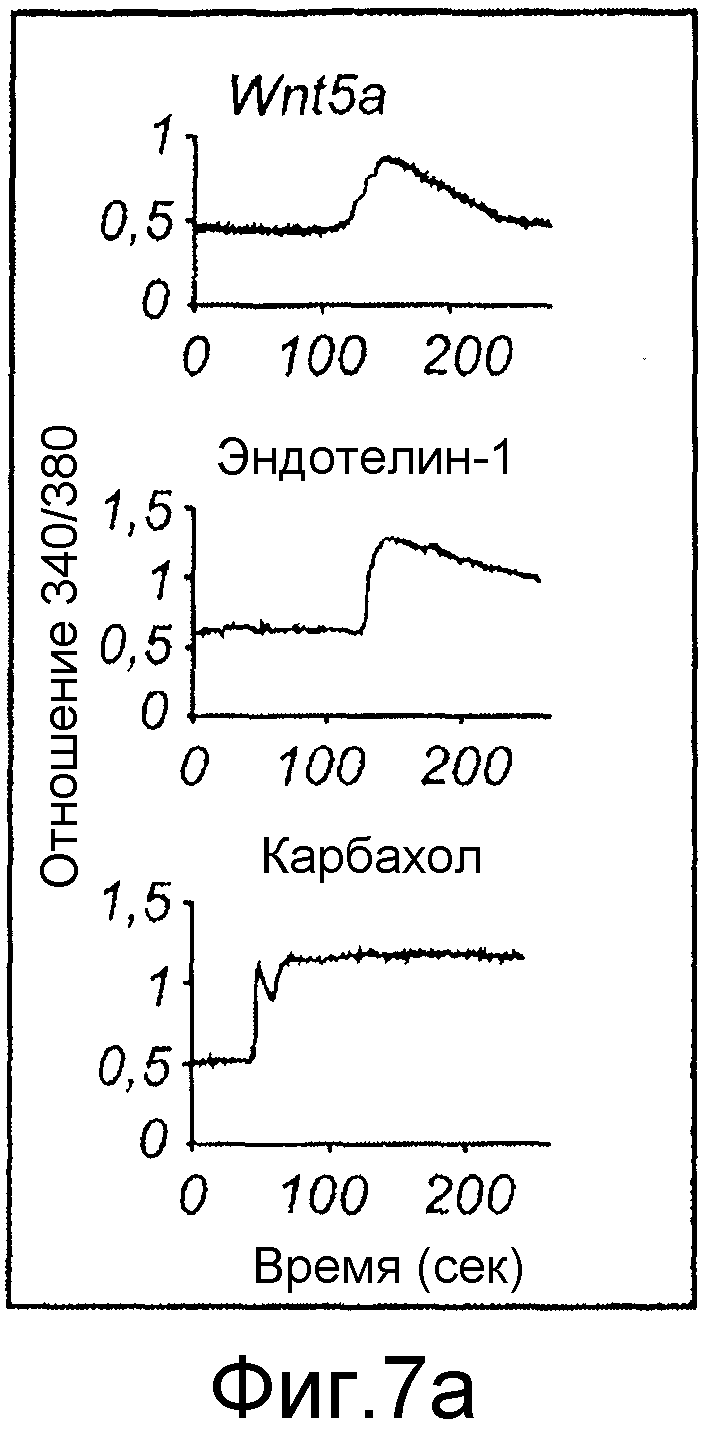
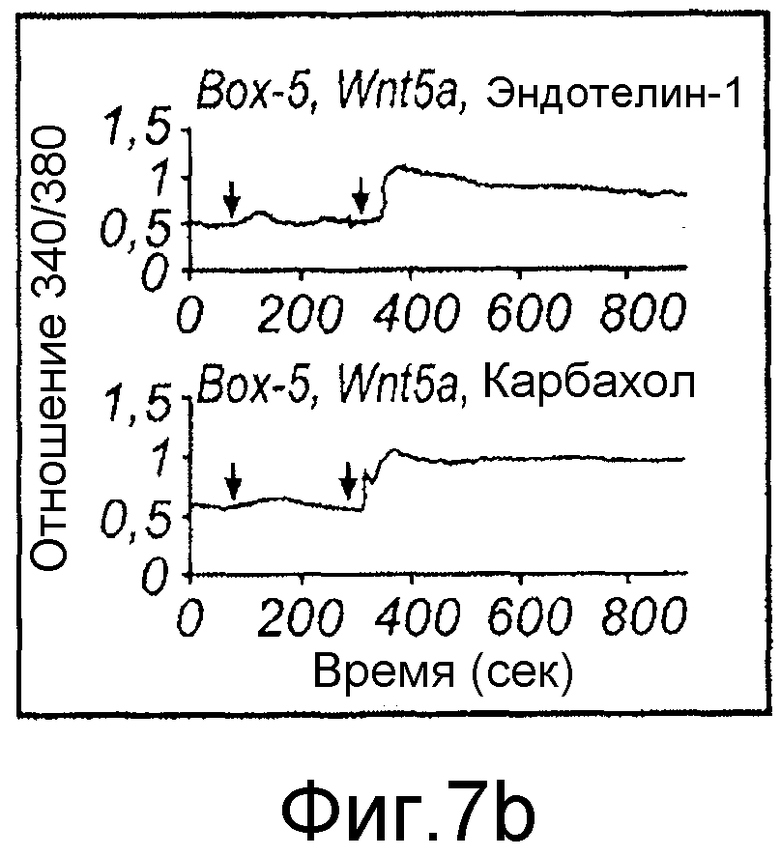
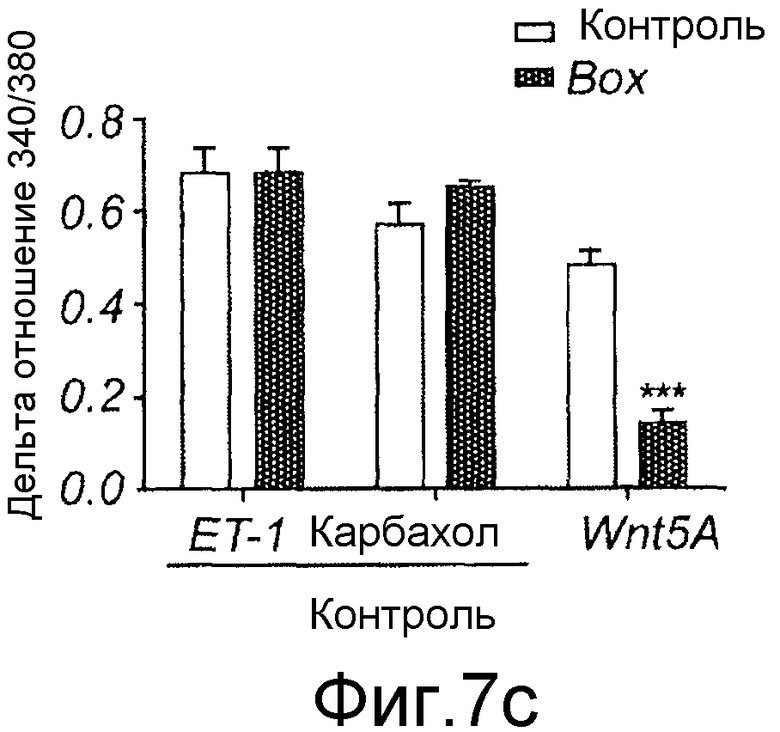
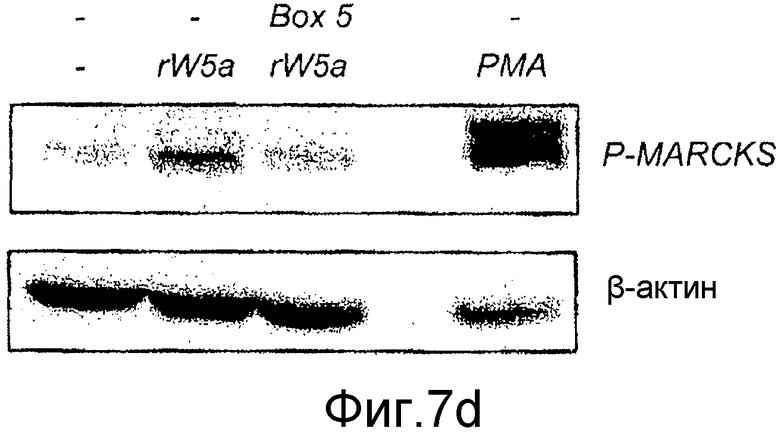
Фиг. 7 иллюстрирует влияние Box5 на индуцированную Wnt-5a сигнальную систему Ca2+ и активацию PKC.
Флуоресцентные сигналы от нагруженных fura-2 меланомных клеток A2058, предварительно инкубированных (в течение ночи) и инкубированных в отсутствие или присутствии Box5 (100 мкМ,) регистрировали после стимуляции с использованием Wnt-5a (0,1 мкг/мл), эндотелина-1 (ET-1)(10 нМ) или карбакола (5 мкМ). A) Репрезентативные следы Ca2+ от клеток A2058, стимулированных Wnt-5a, эндотелином-1 или карбаколом, последние два представляют собой контроли лиганда рецептора G-белка. B) Два следа Ca2+ от меланомных клеток A2058, предварительно инкубированных и инкубированных с Box5 и затем стимулированных с использованием Wnt-5a (первая стрелка), и затем снова с использованием эндотелина-1 (вторая стрелка верхний след) или карбакола (вторая стрелка в нижнем следе). Все показанные следы являются репрезентативными для по меньшей мере пяти отдельных экспериментов. C) Показаны накопленные результаты изменений ∆Ca2+ в отношении значений (спонтанный уровень к уровню пика), зарегистрированных от клеток A2058, стимулированных Wnt-5a, эндотелином-1 или карбаколом в отсутствие (незакрашенные столбцы) или в присутствии Box5 (залитые краской столбцы). D) Предварительная инкубация с Box5 (100 мкМ) в течение ночи ингибирует фосфорилирование MARCKS через 45 минут или стимуляцию с использованием rWnt-5a (0,2 мкг/мл). 1 нМ PMA использовали как положительный индикатор фосфорилирования MARCKS. Результаты даны как среднее ±SEM, ***p<0,001.
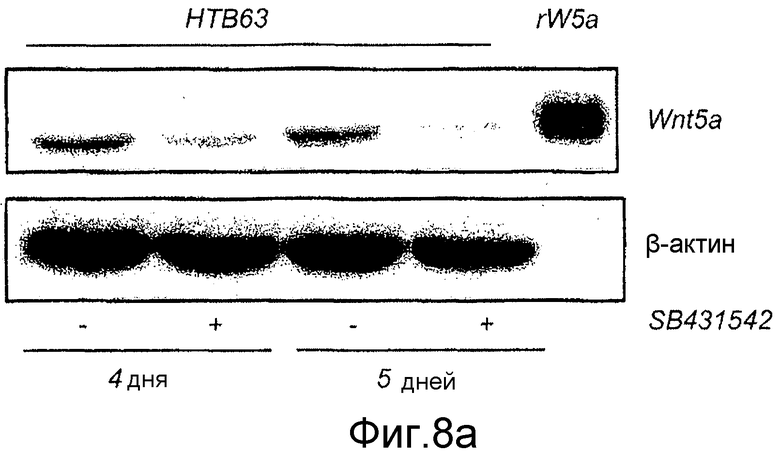
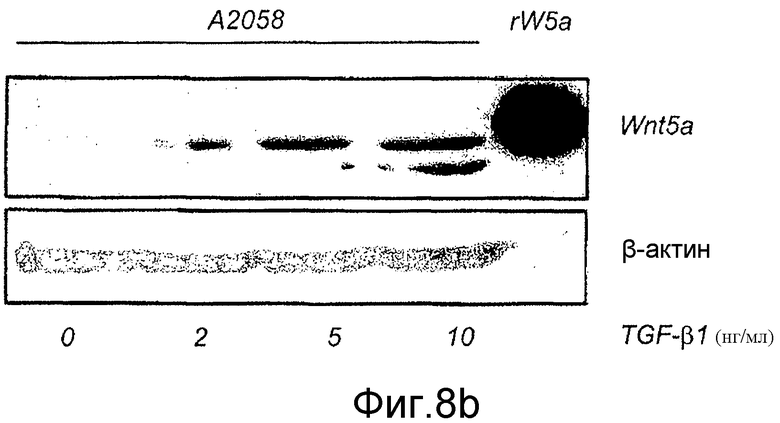
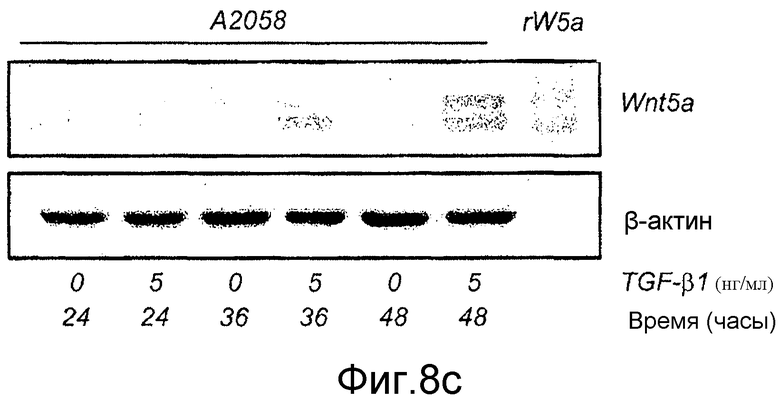
Фиг. 8 иллюстрирует эффекты сигнальной системы TGF-β1 на экспрессию белка Wnt-5a в меланомных клетках A2058 и HTB63.
A) Репрезентативный Вестерн-блот показывает эффекты инкубированных меланомных клеток HTB63 в отсутствие или присутствии селективного TGF-β1 рецепторного антагониста SB431542 (10 мкМ) в течение 4 или 5 дней на эндогенную экспрессию Wnt-5a. Рекомбинантный Wnt-5a служил в качестве положительного контроля и β-актин в качестве нагрузочного контроля.
B) Вестерн-блоты показывают эффекты 24-часовой стимуляции с использованием увеличенных концентраций TGF-β1 на экспрессию Wnt-5a в клетках A2058. Рекомбинантный Wnt-5a служил в качестве положительного контроля и β-актин в качестве нагрузочного контроля. C) Вестерн-блоты показывают эффекты стимуляции с использованием 5 нг/мл TGF-β1 для увеличенных периодов времени на экспрессию Wnt-5a в клетках A2058. Рекомбинантный Wnt-5a служил в качестве положительного контроля и β-актин в качестве нагрузочного контроля. Каждый из представленных результатов повторяли как независимые эксперименты по меньшей мере четыре раза.
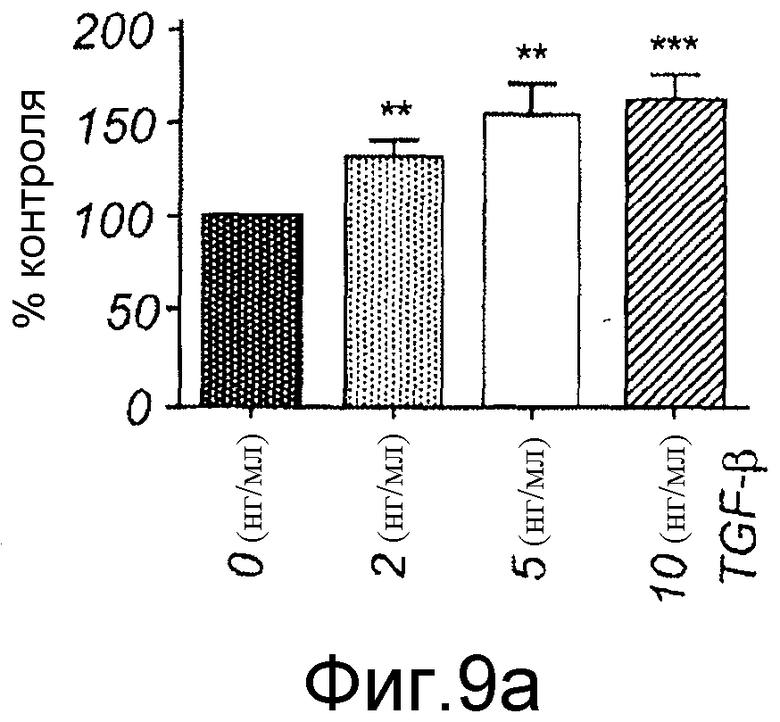
Фиг. 9 иллюстрирует эффекты TGF-β1 и Box5 на адгезию и миграцию меланомных клеток.
A) Меланомные клетки A2058 стимулировали указанными концентрациями TGF-β1, разъединяли Версеном и ресуспендировали как одиночные клетки в бессывороточной среде. Клеткам затем обеспечивали прилипание в 96-луночном планшете и после периода 60 минут не прилипшие клетки смывали, в то время как прилипшие клетки окрашивали и определяли их число. Это число представлено как процент контрольной (без Wnt-5a) стимуляции.
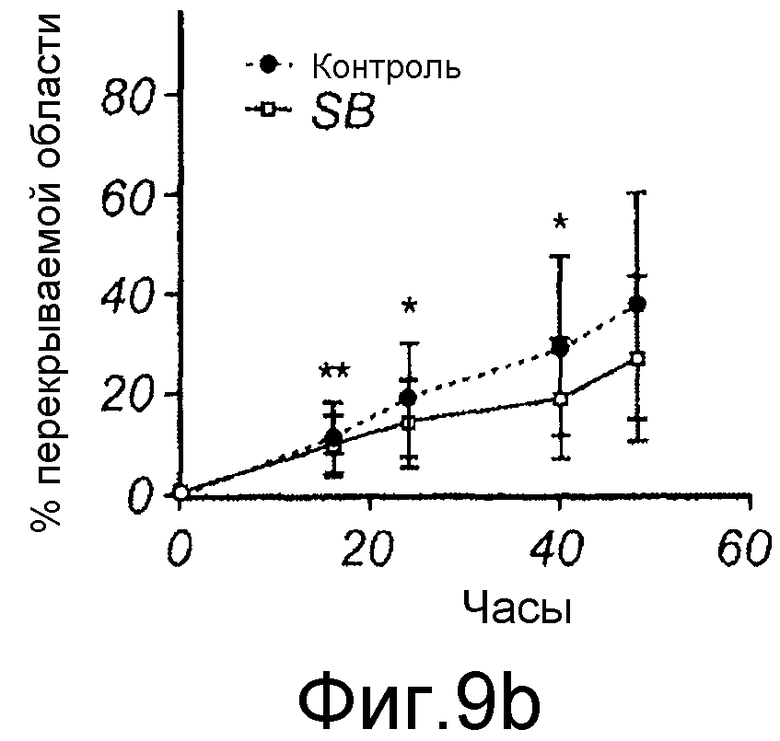
B) Меланомные клетки HTB63 культивировали до слияния в 12-луночной чашке, после чего росчерком вносили в каждую лунку, среду заменяли на свежую бессывороточную среду в отсутствие (закрашенный кружок) или в присутствии 10 мкМ SB431542 (незакрашенный квадрат).
C) Меланомные клетки A2058 культивировали до слияния в 12-луночной чашке, после чего росчерком вносили в каждую лунку, среду заменяли на свежую бессывороточную среду в отсутствие любой добавки (закрашенный кружок) или в присутствии 5 нг/мл TGF-β1 индивидуально (незакрашенный квадрат), или в присутствии 5 нг/мл TGF-β1 со 100 мкМ Box5 (незакрашенный треугольник). В приведенных экспериментах на панелях B и C картину получали от каждого росчерка/лунки из одной и той же области клеток через 0, 16, 24, 40 или 48 часов, и заживление раны выражали как процент закрытия раневой поверхности.
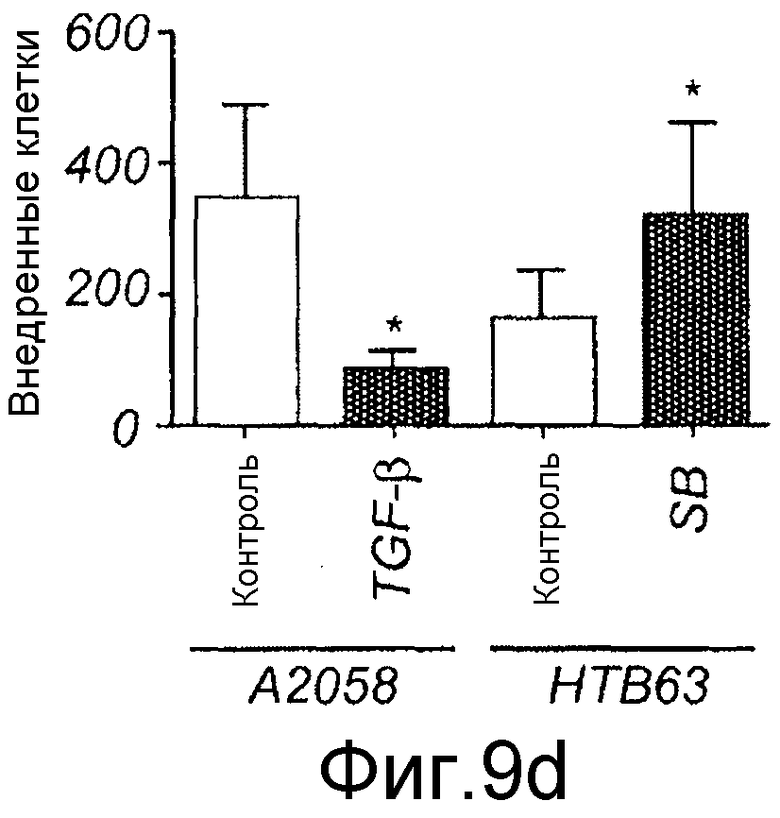
D) До начала каждого эксперимента клетки A2058 (два столбца слева) и клетки HTB63 (два столбца справа) разъединяли Версеном и ресуспендировали как одиночные клетки в бессывороточной среде RPMI. Аликвоту клеточной суспензии, содержащую 25000 клеток, затем добавляли в верхнюю Transwell камеру и нижнюю камеру заполняли содержащей сыворотку (10%) средой. Как указывалось, клеткам давали возможность внедриться в отсутствие (незакрашенные столбцы) или в присутствии 5 нг/мл TGF-β1 (залитый краской столбец), или 10 мкМ SB431542 (залитый краской столбец) в верхней камере. Клеткам затем давали возможность внедриться в течение 24 часов, после чего прикрепленные клетки на нижней стороне мембраны подсчитывали. Результаты даны как среднее ± SEM (n=5-10). *=p<0,05, **=p <0,01 и ***=p<0,001.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Антитела и пептиды
Использовали следующие первичные антитела: β-актин моноклональное AC-15 Ab (Sigma Aldrich, St. Louis, MO); TGF-β1 куриное поликлональное Ab (R&D Systems Europe Ltd., Абингдон, Великобритания). Поликлональное антитело к Wnt-5a получили в лаборатории изобретателя против аминокислот 275-290 зрелой молекулы Wnt-5a, как ранее описано (Jonsson et al., 2002). Вторичная конъюгированная с пероксидазой анти-куриная IgY (IgG) полная молекула изготовлена Sigma Aldrich (St. Louis, MO); все другие конъюгированные с пероксидазой IgG получили от Dakopatts (Глоструп, Дания). Inbiolabs Ltd (Таллинн, Эстония) синтезировал новый производный от Wnt-5a N-бутилоксикарбонильный гексапептид (Met-Asp-Gly-Cys-Glu-Leu; Box5) для двух различных событий. Две партии пептида Box5 имели сходные результаты в анализах исследователя. Синтезированные партии пептида Box5 (>95% чистоты) проверяли на качество с помощью обращенно-фазовой высокоэффективной жидкостной хроматографии (RP-HPLC) и масс-спектрометрии. Использованный формилированный контрольный пептид: формил-Nle-Leu-Phe-Nle-Tyr-Lys изготовлен Sigma-Aldrich (St. Louis, MO).
Химические реактивы - бензамидин, бычья сыворотка, все типы среды для тканевой культуры изготовлены Sigma-Aldrich (St. Louis, MO). Человеческие рекомбинантные белки Wnt-5a, Wnt-3a и TGF-β1 приобретены у R&D Systems Europe Ltd. (Абингдон, Великобритания). Человеческий и свиной эндотелин-1 и карбакол приобретены у Sigma Aldrich (St. Louis, MO). Протеазные ингибиторы пефаблок, лейпептин и апротинин изготовлены Roche Molecular Biochemicals (Мангейм, Германия). Селективный ингибитор рецептора типа 1 TGF-β1, подобной рецептору активина киназы ALK5 и ее родственных форм ALK4 и 7, SB431542 (Inman et al., 2002) приобретен у Tocris Bioscience (Tocris Cookson Ltd., Бристоль, Великобритания). Реагенты для определения усиленной хемилюминесценции (ECL) приобретены у Santa Cruz Biotechnology, Inc. (Стокгольм, Швеция), тогда как все другие реагенты для электрофореза поступили от BioRad (Ричмонд, Калифорния). Все другие химические реактивы были химически чистыми и приобретены у Sigma-Aldrich (St. Louis, MO).
Клеточная культура
Человеческая злокачественная меланомная клеточная линия A2058 была любезно предоставлена László Kopper из Department of Pathology and Experimental Cancer Research [Отдел патологии и экспериментального исследования рака], Университет Земмельвейс, Будапешт, Венгрия. Клетки A2058 поддерживались в RPMI 1640 с добавлением 10% FBS, 5 ед/мл пенициллина, 0,5 ед/мл стрептомицина и 2 мМ глутамина.
HTB63 (также названная как HT144) человеческая злокачественная меланомная клеточная линия приобретена из Американской коллекции типовых культур (ATCC; LGC Promochem AB, Boras, Швеция) и поддерживалась в среде Мак-Коя 5A с добавлением 10% FBS, 5 ед/мл пенициллина, 0,5 ед/мл стрептомицина и 2 мМ глутамина.
Человеческие клетки карциномы молочной железы, MCF7 (положительный контроль экспрессии Wnt-5a) выращивали в DMEM с добавлением 10% FBS, 5 ед/мл пенициллина, 0,5 ед/мл стрептомицина и 2 мМ глутамина. Все клеточные культуры поддерживались при 37°C в увлажненной атмосфере 5% двуокиси углерода.
Вестерн-блот
Клетки прямо лизировали в 1X буфере Лэммли, содержащем DTT, и кипятили в течение 10 минут, или лизировали в буфере, содержащем 50 мМ Трис-HCl (pH 7,5), 1% Тритон X-100, 100 мМ NaCl, 10 мМ MgCl2, 20% глицерин, 1 мМ Na3VO4 и протеазные ингибиторы (20 мкг/мл апротинина, 1 мкг/мл лейпептина, 2,5 мМ бензамидина и 2 мМ пефаблока). Клетки, обработанные лизирующим буфером, центрифугировали при 15000 об/мин в течение 5 минут при 4°C. Содержание белка в каждом образце определяли и регулировали обеспечением равной нагрузки белка на каждой дорожке. Затем добавляли 50 мМ DTT и 5x концентрированный буфер Лэммли и образцы кипятили в течение 5 минут. Образцы разделяли с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS) и впоследствии переносили на мембраны PVDF. Для иммуноблоттинга мембраны блокировали в PBS с добавлением 0,2% Твин 20 и 1% обезжиренного молока для Wnt-5a или 3% обезжиренного молока в случае всех других антител в течение 1 часа. Затем мембраны инкубировали в течение 1 часа при комнатной температуре или в течение ночи при 4°C с индикаторным первичным Ab (1:25000 для β-актина; 1:1000 для Wnt-5a и 1:1000 для TGF-β1) в 2% обезжиренном молоке или 1,5% BSA. После экстенсивного промывания в PBS с 0,2% Твина мембраны инкубировали в течение 1 часа с конъюгированным с пероксидазой хрена вторичным Ab в 2% обезжиренном молоке или 1,5% BSA и снова экстенсивно промывали. Наконец, Ab-антигенные комплексы определяли с использованием усиленной хемилюминесценции. Для повторного зондирования мембраны освобождали с использованием раствора для стриппинга Reblot Strong solution от Chemicon International (Темекула, Калифорния). Показано, что Вестерн-блоты являлись репрезентативными в по меньшей мере трех независимых экспериментах.
ОТ-ПЦР
Экстракцию РНК выполняли с использованием TRIzol® от Invitrogen в соответствии с инструкциями изготовителя. Концентрацию РНК измеряли с использованием нанокапельного спектрофотометра ND-1000 (Bio-Rad (Геркулес, Калифорния). Перед обратной транскрипцией РНК обрабатывали 1 ед/мл ДНКазыI (Invitrogen). кДНК синтезировали с использованием случайных гексамеров от Fermentas (Хельсингборг, Швеция), 1-2 мкг общей РНК с использованием обратной транскриптазы (RT) M-MuLV [вирус лейкоза мышей Молони] (Fermentas, Хельсингборг, Швеция). Для ПЦР-реакций в объеме 50 мкл использовали 5 мкл RT реакции в 1X Taq полимеразном буфере (75 мМ Трис-HCl, 20 мМ [NH4]2SO4, 0,01% Твин 20) с добавлением 2,5 мМ MgCl2, 200 мМ дНТФ, 1 мкМ каждого праймера и 1 единицу ДНК-полимеразы Taq (Fermentas, Хельсингборг, Швеция). ПЦР-праймеры были следующими: Wnt-5a прямой: 5'-GGATTGTTAAACTCAACTCTC-3' (SEQ. ID. NO: 16);
Wnt-5a обратный: 5'-ACACCTCTTTCCAAACAGGCC-3' (SEQ. ID. NO: 17); β-актин прямой: 5'-TTCAACACCCCAGCCATGTA-3' (SEQ. ID. NO: 18);
β-актин обратный: 5'-TTGCCAATGGTGATGACCTG-3' (SEQ. ID. NO: 19); Frizzled-2 прямой: 5'-ACATCGCCTACAACCAGACC-3' (SEQ. ID. NO: 20); и
Frizzled-2 обратный: 5'-CTCGCCCAGAAACTTGTAGC-3' (SEQ. ID. NO: 21);
Frizzled-5 прямой: 5'-ACACCCGCTCTACAACAAGG-3' (SEQ. ID. NO: 22); и
Frizzled-5 обратный: 5'-CGTAGTGGATGTGGTTGTGC-3' (SEQ. ID. NO: 23). Реакции ОТ-ПЦР, показанные на Фиг. 1, являются репрезентативными в по меньшей мере трех независимых экспериментах.
Клеточная адгезия
Клетки, предварительно обработанные и стимулированные, как описано ниже, разъединяли Версеном и ресуспендировали в бессывороточной среде RPMI и образцы, содержащие 30000 клеток из каждой обработанной группы, добавляли в каждую лунку 96-луночного планшета. Клеткам затем обеспечивали прилипание в течение 60 минут при 37°C в увлажненной атмосфере 5% двуокиси углерода, после чего неприлипшие клетки смывали с использованием PBS. Прилипшие [адгезивные] клетки фиксировали в 1% глутаральдегиде в течение 10 минут при комнатной температуре и затем окрашивали с использованием 0,5% кристаллвиолета в 20% метаноле в течение 10 минут. Наконец краситель из каждой группы клеток растворяли в 50% уксусной кислоте. Количество растворенного красителя из каждой лунки затем измеряли в планшете-ридере Fluostar [спектрофотометр для прочтения планшетов] (BMG Lab Technologies GmbH, Оффенберг, Германия) при 544 нм. Индивидуальные образцы от каждого отдельного эксперимента анализировали четырехкратно и накопленные данные основывались на 5 отдельных экспериментах. Результаты иллюстрированы на Фиг. 4.
A) Меланомные клетки A2058 стимулировали указанными концентрациями Wnt-5a, разъединяли Версеном и ресуспендировали как одиночные клетки в бессывороточной среде в присутствии или в отсутствие рекомбинантного Wnt-5a в указанных концентрациях. Клеткам затем обеспечивали прилипание в 96-луночном планшете. По истечении периода 60 минут неприлипшие клетки смывали, в то время как прилипшие клетки окрашивали и определяли их число. Это число представлено как процент контрольной (без Wnt-5a) стимуляции (Фиг. 4A).
B) Меланомные клетки A2058 культивировали до слияния в 12-луночном планшете, после чего росчерком вносили в каждую лунку, среду заменяли на свежую бессывороточную среду, не содержащую (закрашенные кружки) или содержащую 0,2 мкг/мл Wnt-5a (незакрашенные квадраты) (Фиг. 4B).
C) Меланомные клетки A2058 культивировали до слияния в 12-луночном планшете, после чего росчерком вносили в каждую лунку, среду заменяли на свежую бессывороточную среду в отсутствие любой добавки (закрашенный кружок), присутствии 0,2 мкг/мл Wnt-5a одного (незакрашенные квадраты) или в присутствии 0,2 мкг/мл Wnt-5a со 100 мкМ Box5 (незакрашенный треугольник) (Фиг. 4C).
D) Меланомные клетки HTB63 культивировали до слияния в 12-луночной чашке, после чего росчерком вносили в каждую лунку, среду заменяли на свежую бессывороточную среду (закрашенный кружок), кондиционированную среду (незакрашенные квадраты) или кондиционированную среду с добавлением 100 мкМ Box5 (незакрашенный треугольник). Для регистрации изменений в миграции на панелях B-D картину учитывали с каждого росчерка/лунки из одной и той же области клеток спустя 0, 16, 24, 40 или 48 часов, и ранение-заживление выявляли как процент закрытия раневой поверхности (Фиг. 4D).
E) До начала каждого эксперимента клетки A2058 (шесть столбцов слева) и клетки HTB63 (два столбца справа) разъединяли Версеном и ресуспендировали как одиночные клетки в бессывороточной среде. Клетки предварительно инкубировали в течение 40 минут с использованием непрерывного встряхивания в отсутствие (незакрашенные столбцы) или в присутствии (залитые краской столбцы) 100 мкМ Box5. Аликвоту клеточной суспензии, содержащую 25000 клеток, затем добавляли в верхнюю Transwell камеру и нижнюю камеру заполняли содержащей сыворотку (10%) средой. Как указывалось, 0,1 мкг/мл Wnt-3a, 0,2 мкг/мл Wnt-5a и/или 100 мкМ Box5 добавляли в верхнюю камеру. Клеткам затем давали возможность внедриться в течение 24 часов, после чего прикрепленные клетки на нижней стороне мембраны подсчитывали. Результаты даны как среднее ± SEM (n=5-7). *=p<0,05, **=p<0,01 и ***=p<0,001, где значения сравниваются с контролем (Фиг. 4D, CM против CM + Box5, ^=p<0,05, ^^=p<0,01 и ^^^=p<0,001).
Анализ заживления раны
Клетки помещали в 12-луночный планшет и обеспечивали рост в сливном слое в полной [среде] RPMI (для клеток A2058) или в полной среде Мак-Коя 5A (для клеток HTB63). Как указывалось, клетки предварительно инкубировали с пептидом Box5 (100 мкМ) или контрольным растворителем в течение 40 минут при непрерывном встряхивании. На рану затем наносили путем росчерка через сливной слой клетки с использованием микродозатора. В продолжение анализа периода миграции клетки инкубировали в бессывороточной среде или в случае клеток HTB63 в их собственной бессывороточной кондиционированной среде, собранной из клеток, культивированных в течение 48 часов, и использованной в течение 2 дней после сбора. Точные условия каждого эксперимента описаны выше. Для избежания потери активности клеточную среду заменяли через 24 часа. Картину для каждого росчерка получали из одной и той же области клеток через 0, 16, 24, 40 или 48 часов, и заживление раны измеряли как процент закрытия раневой поверхности. Для каждого экспериментального условия заживление раны анализировали трижды. Все данные основаны на 3-8 отдельных экспериментах, как указано.
Клеточная инвазия
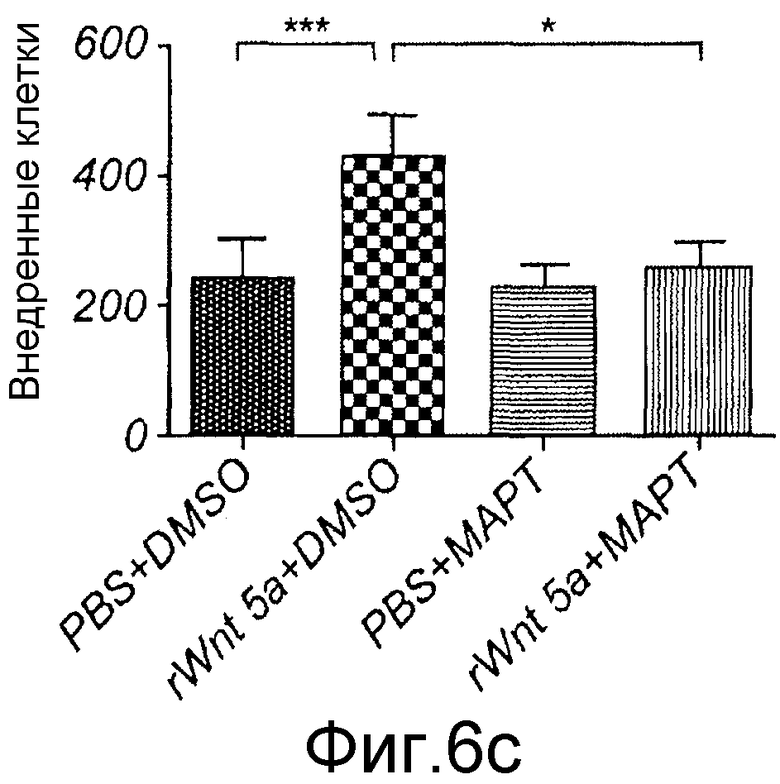
Клеточную инвазию анализировали с использованием анализа в инвазионной камере BD MatrigelTM (BD Biosciences, Бедфорд, Массачусетс). Перед началом каждого эксперимента клетки разъединяли Версеном и ресуспендировали как одиночные клетки в бессывороточной среде RPMI. Как указывалось, клетки предварительно инкубировали с пептидом Box5 (100 мкМ) или контрольным растворителем в течение 40 минут при непрерывном встряхивании. Аликвоту клеточной суспензии, содержащую 25000 клеток, и 100 мкМ Box5 или контрольный растворитель затем добавляли в верхнюю transwell камеру и нижнюю камеру заполняли содержащей сыворотку (10%) средой. Одновременно Wnt-5a, Wnt-3a или TGF-β1 ингибитор SB431542 добавляли в верхнюю камеру, как указано на Фиг. 7. Инвазионные камеры встряхивали горизонтально в течение 5 минут для обеспечения равномерного распределения клеток в суспензии и впоследствии через мембранную поверхность. Клеткам давали возможность внедриться в течение указанного периода времени при 37°C в увлажненной атмосфере 5% CO2. Среду удаляли и клетки фиксировали в 4% параформальдегиде в течение 10 минут. Клетки окрашивали с использованием 0,5% кристаллвиолета в 20% метаноле в течение 10 минут и не внедренные клетки на внутренней стороне мембраны удаляли с использованием аппликатора с хлопковым наконечником. Мембрану вырезали из камеры с использованием лезвия скальпеля и окрашенные клетки на нижней стороне камеры подсчитывали.
Определение уровней цитозольного свободного кальция
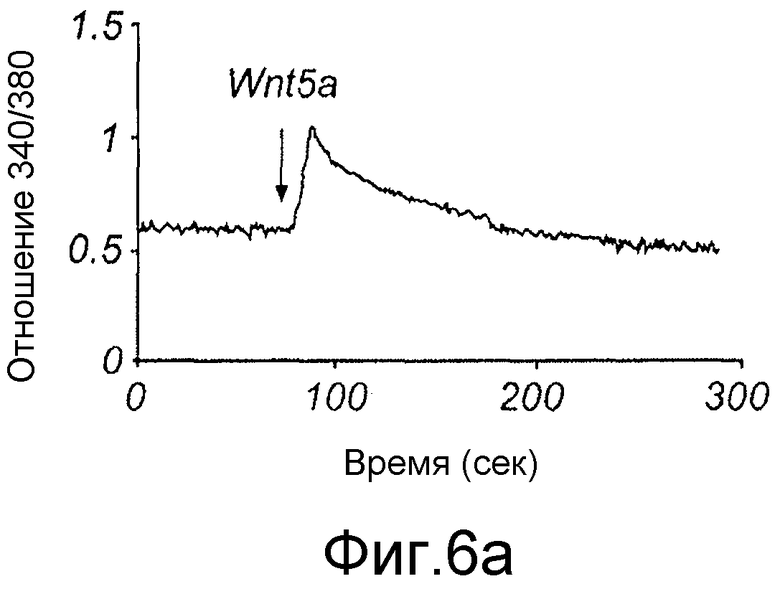
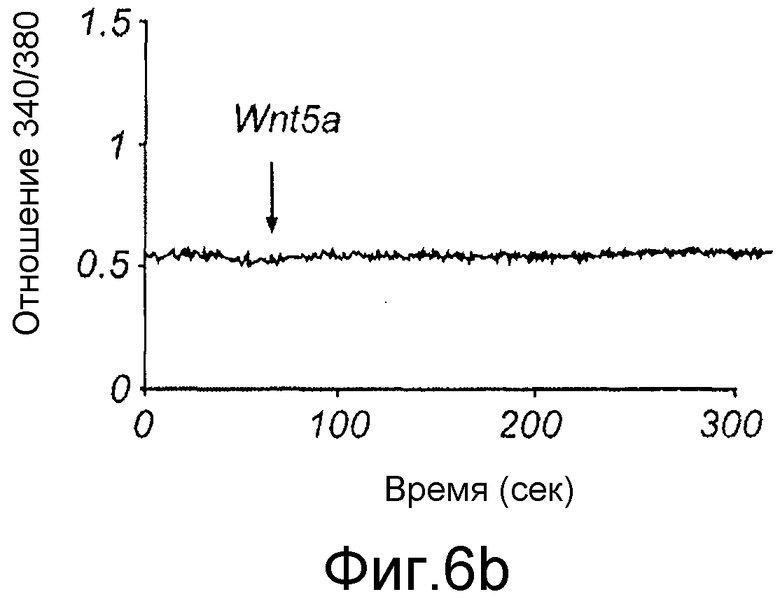
Клетки, выращенные на стеклянном покровном стекле, инкубировали с 4 мкМ fura-2/AM в культуральной среде в течение 30 минут при 37°C (Dejmek et al., 2006). После загрузки fura-2 клеток покровные стекла промывали и устанавливали в специально сконструированную камеру, в которую добавляли содержащую кальций среду (136 мМ NaCl, 4,7 мМ KCl, 1,2 мМ MgSO4, 1,1 мМ CaCl2, 1,2 мМ KH2PO4, 5 мМ NaHCO3, 5,5 мМ глюкозы и 20 мМ Hepes, pH 7,4). Камеру затем помещали в систему, состоящую из микроскопа NIKON Diaphot, соединенного с системой визуализации Photon Technology International (PTI). Клеткам сначала обеспечивали состояние покоя в течение 10 минут перед проведением любой стимуляции. Флуоресценцию fura-2 затем регистрировали непрерывно от клеток до и после стимуляции с использованием Wnt-5a, эндотелина-1 или карбакола (как указывалось в условных обозначениях к Фиг. 8) с использованием возбуждающей длины волны, быстро изменяющейся между 340 и 380 нм, тогда как эмиссионная длина волны устанавливалась при 510 нм. Отношения интенсивности флуоресценции (340/380 нм) в дальнейшем рассчитывали и анализировали с использованием программы PTI Image Master.
Статистический анализ - критерий Стьюдента для непарных образцов использовали для анализа различий в экспериментах, где *=p<0,05, **=p<0,01 и ***=p<0,001.
Характеристика меланомных клеточных линий A2058 и HTB63
Для дальнейшего исследования эффектов экспрессии и передачи сигнала Wnt-5a в меланомных клетках решили использовать две различные человеческие меланомные клеточные линии, A2058 и HTB63. И та, и другая меланомные клетки A2058 и HTB63 экспрессировали рецепторы Frizzled-5 (Фиг. 1A), предполагаемый Wnt-5a рецептор в меланомных клетках (Weeraratna, 2002). С другой стороны наблюдали значительно более слабую экспрессию рецепторов Frizzled-2 в клеточных линиях. Дальнейшее определение характеристик показало, что клетки A2058 лишены эндогенной экспрессии мРНК и белка Wnt-5a, тогда как клетки HTB63, напротив, проявляют постоянную эндогенную экспрессию мРНК (Фиг. 1A) и белка (Фиг. 1B) Wnt-5a.
Для того чтобы убедиться в том, что эндогенный Wnt-5a экспрессируемый в клетках HTB63, действительно секретируется, среду, в которой выращивали клетки, анализировали с помощью Вестерн-блота на присутствие Wnt-5a. Нижний блот на фиг. 1B четко показывает, что клетки HTB63 не только экспрессируют, но также секретируют Wnt-5a. Экспрессирующие Wnt-5a и Frizzled-5 клетки MCF-7 рака молочной железы и рекомбинантный Wnt-5a использовали в качестве положительных контролей в этих экспериментах. Эти данные показывают, что экспрессия Wnt-5a в этих клеточных линиях регулируется на транскрипционном уровне, что отличается от ткани и клеток рака молочной железы, где она регулируется на трансляционном уровне (Dejmek, Leandersson). В настоящее время факторы, которые регулируют транскрипцию Wnt-5a в меланомных клетках, не известны.
Создание пептида антагониста Wnt-5a
Ранее изобретатель идентифицировал Wnt-5a производное, N-формилированный гексапептид (Foxy5; Фиг. 2A), который функционирует как агонист сигнальной системы Wnt-5a. Этот пептид имитирует эффекты Wnt-5a в клеточных линиях рака молочной железы in vivo. Обнаружено, что Foxy5 может также имитировать промигрирующие эффекты Wnt-5a в меланомных клетках A2058 (Фиг. 2B), показывая, что этот пептид функционирует как Wnt-5a агонист в различных клеточных типах. Интересно, что ранее было показано, что специфическая модификация формилированного, полученного из бактерий хемотактического пептида (формил-Met-Leu-Phe), превращает молекулу из агониста в аналог антагониста. В настоящее время показано, что такая модификация Foxy5 может также изменять ее функцию Wnt-5a агониста на функцию антагониста. Этот пептид t-boc-Met-Asp-Gly-Cys-Glu-Leu был назван как Box5 (Фиг. 3).
Эффекты Wnt-5a и N-бутилоксикарбонильного гексапептида Box5 на клеточную адгезию и миграцию
Эффекты Wnt-5a тестировали на адгезию меланомных клеток A2058. Wnt-5a увеличивал адгезивную способность меланомных клеток A2058, и максимальный эффект получали после стимуляции 0,2 мкг/мл (Фиг. 4A). На основании этих данных затем исследовали, как эта концентрация рекомбинантного Wnt-5a влияет на миграцию клеток A2058 в анализе заживления раны. Результаты, приведенные на фиг. 4B, четко показывают, что добавление Wnt-5a (0,2 мкг/мл) в начале экспериментов увеличивает миграцию меланомных клеток A2058. Для специфического ингибирования этой, индуцированной Wnt-5a миграции меланомных клеток впоследствии разработали и тестировали новый N-бутилоксикарбонильный модифицированный пептид. Ранее было показано, что полученный из Wnt-5a формилированный гексапептид может имитировать ингибирующее действие Wnt-5a на миграцию клеток рака молочной железы in vitro (Säfholm, 2006) и что этот пептид также ингибирует метастаз рака молочной железы на мышиной модели (Säfholm, 2008). В данном описании тестировали возможность добавления бутилоксикарбонильной группы к N-концевому метиониновому остатку гексапептида для получения ингибитора сигнальной системы Wnt-5a. Основой для этой манипуляции являются данные о том, что такая модификация полученного из бактерии хемотактического пептида (формил-Met-Leu-Phe), как было сообщено, изменяет действия пептида от агонистического на антагонистическое (Derian, 1996). Способность этого бутилоксикарбонил-модифицированного гексапептида, далее называемого как Box5, тестировали на ингибирование миграции меланомных клеток A2058 и HTB63 в анализе заживления раны. Box5 отменяет индуцированную Wnt-5a миграцию клеток A2058 (Фиг. 4C), но не влияет на естественную миграцию этих клеток, которые лишены эндогенной экспрессии (Фиг. 5A). Также показали, что опосредованная TGF-β1 миграция клеток A2058 может быть блокирована предварительной инкубацией с Box5 (Фиг. 5B). Эти данные также поддерживались данными о том, что Box5, но не формилированный контрольный гексапептид (данные не показаны), ингибирует миграцию клеток HTB63 в том же размере, что и при изменении кондиционированной среды (содержащей секретированный Wnt-5a) на свежую бессывороточную среду (лишенную Wnt-5a) (Фиг. 4D). С помощью анализа заживления раны исследуют миграцию клеток, присутствующих в монослое, которая характеризуется многочисленными клетка-клеточными взаимодействиями. Однако это не отражает ситуацию in vivo, когда опухолевые клетки усиливают внедрение во внеклеточный матрикс и поэтому далее выполняли аналогичные эксперименты по анализу инвазии.
Во время процесса метастазирования опухолевые клетки должны внедряться во внеклеточный матрикс, поэтому тестировали эффективность Box5 в блокировании клеточной инвазии на 3-мерной модели клеточной культуры. Добавление Box5 отменяло индуцированную Wnt-5a инвазию клеток A2058, эффект не наблюдали, когда клетки стимулировали каноническим Wnt лигандом, Wnt-3a (Фиг. 4E). Box5 также проявлял способность ингибировать инвазию клеток HTB63 путем противодействия эффектам эндогенного Wnt-5a (Фиг. 4E). В совокупности эти данные показывают, что Box5 является потенциальным антагонистом опосредованной Wnt-5a миграции и инвазии меланомных клеток, и те, и другие являются необходимыми компонентами процесса метастазирования.
Результаты, полученные в анализе заживления раны, подтвердились при анализе инвазии в отношении способности Wnt-5a стимулировать инвазию меланомных клеток A2058. К тому же, этот эффект Wnt-5a на инвазию меланомных клеток не наблюдали, когда клетки стимулировали каноническим лигандом Wnt-3a. Кроме того, добавление пептида Box5 отменяло индуцированную Wnt-5a инвазию меланомных клеток A2058, но эффект не наблюдали при спонтанной инвазии. При этом Box5 мог также ингибировать инвазию меланомных клеток HTB63, которые проявляли эндогенную экспрессию и секрецию Wnt-5a.
Сигнальный путь Wnt/Ca 2+ необходим для опосредованной Wnt-5a инвазии меланомных клеток
Для идентификации молекулярной основы антагонистических функций Box5 исследовали сигнальные пути, которые необходимы для индуцированной Wnt-5a инвазии меланомных клеток. Обнаружено, что Wnt-5a стимулирует быстрый цитозольный сигнал Ca2+ в клетках A2058 (Фиг. 6A), который может быть ингибирован путем использования внутриклеточного Ca2+ хелатора MAPT/AM (Фиг. 6B).
Ca2+ хелирование посредством MAPT/AM использовали для оценки инвазивной способности меланомных клеток в отсутствие индуцированного Ca2+ сигнального пути Wnt-5a. Проинвазивный эффект Wnt-5a на клетки A2058 полностью отменялся посредством MAPT/AM (Фиг. 6C). Это демонстрирует, что Ca2+ сигнальный компонент Wnt-5a стимуляции необходим для опосредования инвазии меланомных клеток.
Эффекты N-бутилоксикарбонильного гексапептида Box5 на индуцированную Wnt-5a сигнальную систему
Для дальнейшего исследования свойств гексапептида Box5 и его селективного взаимодействия с Wnt-5a рецепторами анализировали его эффекты непосредственно на индуцированную Wnt-5a рецепторную сигнальную систему. Ранее показано, что Wnt-5a запускает быстрое увеличение цитозольного свободного Ca2+ в клетках щитовидной железы (Kremenevskaja, 2005) и в клетках рака молочной железы (Dejmek, 2006). Данные показывают, что Wnt-5a также запускает быстрый цитозольный Ca2+ сигнал в меланомных клетках A2058 (Фиг. 7A), сходный с быстрыми ответами, индуцированными двумя другими сопряженными с G-белком контрольными рецепторными лигандами, эндотелином-1 и карбахолом (Фиг. 7A). Необходимо отметить, что для того чтобы получить приблизительно сходные Ca2+ ответы для всех трех лигандов, концентрацию Wnt-5a снижали от 0,2 до 0,1 мкг/мл в этих сериях экспериментов.
Эффекты Box5 затем тестировали на клетках, которые сначала стимулировали Wnt-5a и впоследствии эндотелином-1 или карбахолом (Фиг. 7B). Эти эксперименты показали, что Box5 селективно ингибирует Wnt-5a-, но не эндотелин-1 или карбахол-индуцированный внутриклеточный сигнал Ca2+ в меланомных клетках A2058. Накопленные результаты по влиянию Box5 на индуцированный Wnt-5a сигнал Ca2+ показали, что имеется более чем 70% ингибирование по сравнению с пиком значений сигнала Ca2+ в отсутствие Box5 (Фиг. 7C). Сходные эксперименты по Ca2+, в которых клетки стимулировали эндотелином-1 или карбахолом в отсутствие Box5, не выявили значимого эффекта этого пептида на сигнальную систему Ca2+ (Фиг. 7C).
Один из нижележащих эффектов сигнальной системы Wnt-5a представляет собой активацию PKC. Стимуляция Wnt-5a клеток A2058 приводит к увеличению фосфорилирования MARCKS и эндогенного PKC субстрата, который ингибируется в присутствии пептида Box5 (Фиг. 7D). Эти данные показывают, что функции Box5 по блокированию инвазии меланомных клеток путем прямого противодействия стимулированной Wnt-5a сигнальной системе Ca2+ и PKC приводят к нижележащему последующему ингибированию опосредованной Wnt-5a клеточной инвазии.
Хотя регистрируемые изменения внутриклеточного Ca2+ являются очень чувствительным анализом исследования модуляции сопряженной с G-белком сигнальной системы, она не связана прямо с подвижностью меланомных клеток. Однако ранние исследования показали нижележащее влияние индуцированной Wnt-5a активации PKC на регуляцию миграции меланомных клеток (Weeraratna 2002, Dissanayake et al., 2007).
В настоящем исследовании уровень аутофосфорилирования PKC (Weeraratna 2002, Dissanayake et al., 2007) не оценивали, так как его связь с киназной активностью PKC не ясна. Вместо этого эффекты Wnt-5a и Box5 на фосфорилирование эндогенного PKC субстрата MARCKS анализировали в качестве прямого средства оценки уровня активности PKC в меланомных клетках. Стимуляция посредством Wnt-5a меланомных клеток A2058 приводит к прямому увеличению фосфорилирования MARCKS с пиками от 9 до 15 мин (Фиг. 7A). Этот эффект Wnt-5a отменялся посредством пептида Box5 (Фиг. 7B). Эти результаты снова подтверждают селективный эффект Box5 на миграцию и инвазию меланомных клеток (Фиг. 4C-D) и далее поддерживают гипотезу о том, что Box5 является селективным пептидным Wnt-5a антагонистом.
Эффекты SB431542 и TGF-β1 на экспрессию белка Wnt-5a в клетках HTB63 и A2058
В настоящее время регуляция транскрипции Wnt-5a не ясна. В других клеточных типах, совсем недавно показанных на эпителиальных клетках протоков молочной железы в процессе развития (Roarty and Serra, 2007), обнаружили, что TGF-β1 отвечает за регуляцию экспрессии Wnt-5a на транскрипционном уровне. Для того чтобы исследовать возможность того, что сходный механизм существует в меланомных клетках, непосредственно тестировали способность селективного ингибитора рецептора TGF-β1 типа I, SB431542, и рекомбинантного TGF-β1 влиять на экспрессию Wnt-5a в клетках. Приведенные данные на фиг. 7C показывают, что когда клетки HTB63 (которые проявляют эндогенную экспрессию Wnt-5a) содержали на полной среде Мак-Коя 5A с добавлением 10 мкМ SB431542 в течение 4-5 дней, эндогенная экспрессия белка Wnt-5a значимо снижалась через 4 дня инкубации и почти исчезала через 5 дней инкубации. Стимуляция клеток A2058 (которые лишены эндогенной экспрессии Wnt-5a) в течение 36 ч различными концентрациями рекомбинантного TGF-β1 приводила к увеличению экспрессии белка Wnt-5a (Фиг. 8B). Эти данные показывают, что приблизительно максимальная экспрессия Wnt-5a достигается при стимуляции 5 нг/мл TGF-β1 (Фиг. 8B) и что стимуляция клеток A2058 5 нг/мл TGF-β1 требует 36 ч стимуляции для получения четко определяемого увеличения экспрессии белка Wnt-5a (Фиг. 8C). Эти результаты подтверждают, что TGF-β1 регулирует экспрессию в по меньшей мере двух клеточных линиях злокачественной меланомы, используемых в этом исследовании. Это повышает возможность того, что опосредованной Wnt-5a миграции меланомных клеток можно противодействовать косвенно путем блокирования сигнальной системы TGF-β1 в этих клетках.
Эффекты TGF-β1, SB431542 и Box5 на адгезию, миграцию и инвазию меланомных клеток
Для исследования того, действительно ли Wnt-5a является нижележащим регулятором индуцированной TGF-β1 клеточной миграции, эффекты TGF-β1 сначала тестировали на адгезию меланомных клеток A2058. Обнаружили, что TGF-β1 стимулировал адгезивную способность меланомных клеток A2058 и что максимальный эффект получили после стимуляции 5 нг/мл (Фиг. 9A). На основе этих данных затем исследовали, как эта концентрация рекомбинантного TGF-β1 влияла на миграцию клеток A2058 в анализе заживления раны. Результаты, приведенные на фиг. 9B, четко показывают, что добавление TGF-β1 (5 нг/мл) в начале экспериментов увеличивает миграцию меланомных клеток A2058. В хорошем соответствии с этими результатами обнаружили, что 10 мкМ SB431542, ингибитора рецептора TGF-β типа I, ингибировали миграцию клеток HTB63 (Фиг. 9C). Далее тестировали способность Box5 ингибировать миграцию A2058 в анализе заживления раны. Box5 отменяет индуцированную TGF-β1 миграцию в двумерном анализе заживления раны (Фиг. 9D). Box5 не влиял на спонтанную миграцию меланомных клеток A2058 (данные не показаны). Однако когда тестировали влияние TGF-β1 (5 нг/мл) на миграцию в более комплексном анализе миграции, получили противоречивые результаты. В инвазионном анализе TGF-β1 ингибировал миграцию клеток A2058, и SB431542 стимулировал миграцию клеток HTB63 (Фиг. 9E). Поэтому сделали вывод, что TGF-β1 является непрогнозируемой мишенью блокирования зависимой от Wnt-5a миграции меланомных клеток вследствие его множественных нижележащих эффектов.
В настоящем исследовании было показано, что модифицированный полученный из Wnt-5a гексапептид Box5 селективно ингибирует индуцированную Wnt-5a сигнальную систему в меланомных клетках и блокирует опосредованную Wnt-5a миграцию этих клеток в анализе заживления раны и инвазионном анализе. Основа для конструирования Box5 появилась из предыдущей работы, в которой провели предсказание вторичной структуры/доступной для растворителя поверхности по методу PHD (Rost, 1996), и затем скринировали полученные из Wnt-5a малые низкомолекулярные пептиды на способность реконструировать эффекты Wnt-5a на клетки рака молочной железы, в которых отсутствует эндогенная экспрессия Wnt-5a (Särndahl, 2006). В этом исследовании охарактеризовали гексапептид Met-Asp-Gly-Cys-Glu-Leu, который после формилирования N-концевого Met, названный как Foxy5, оказался способен имитировать эффекты Wnt-5a на сигнальную систему и ингибировать миграцию клеток рака молочной железы (Särndahl, 2006). Эти эффекты Foxy5 утрачивались, если клетки инкубировали с ранее описанным (Sen, 2001; Weeraratna 2002) блокирующим анти-Frizzled-5 антителом, что указывает на то, что Foxy5 опосредует свое влияние на клетку рака молочной железы через сопряженный с G-белком рецептор Frizzled-5 (Säfholm, 2006). Было показано, что такой же рецептор Frizzled отвечает за сигнальную систему и функциональные эффекты Wnt-5a на меланомные клетки (Weeraratna 2002 и Dissanayake, 2007).
Имеется несколько примеров пептидных лигандов, которые могут специфически активировать отдельный рецептор; они включают трипептид Arg-Gly-Asp, который функционирует как лиганд рецептора интегрина (Pierschbacher и Rouslahti, 1984), два гексапептида, которые специфически активируют сопряженные с G-белком активируемые протеазой рецепторы 1 и 4 (Andersen, 1999) и противодействуют септапептиду, который связывается с сопряженным с G-белком рецептором тромбина (Pakala, 2000). Однако для настоящего исследования наиболее интересный пептидный лиганд представляет собой полученный из бактерии формилированный-Met-Leu-Phe трипептид, который активирует лейкоциты путем связывания с высокой аффинностью с сопряженными с G-белком формилпептидными рецепторами на этих клетках (Le, 2002). Если формильная группа этого пептида заменяется на бутилоксикарбонильную группу, этот трипептид еще связывается с таким же рецептором, но вместо действия как агонист этот бутилоксикарбонилированный трипептид теперь действует как антагонист (Derian, 1996). Такая же модификация Met-Asp-Gly-Cys- Glu-Leu гексапептида очевидно, как показано в данном описании, превращает его в селективный для Wnt-5a пептид антагонист в меланомных клетках.
Данные четко показывают, что сигнальная система Wnt-5a согласованно стимулирует миграцию в меланомных клетках A2058 и HTB63 при использовании анализа заживления раны или инвазионого анализа, и что Box5 во всех этих ситуациях блокирует зависимую от Wnt-5a миграцию меланомных клеток. Несмотря на имеющееся доказательство того, что TGF-β1 регулирует транскрипцию Wnt-5a в меланомных клеточных линиях A2058 и HTB63 и что добавление Wnt-5a всегда стимулирует миграцию меланомных клеток, очень противоречивые эффекты отметили, когда стимулировали меланомные клетки посредством TGF-β1. Полученные эффекты оказались сопоставимыми с типом анализа, использованным для исследования клеточной миграции. По всей вероятности эти различные эффекты TGF-β1 на миграцию меланомных клеток сопоставимы с убедительно подтвержденными документальными доказательствами множественных эффектов на опухолевые клетки. Необходимо отметить, что в ситуациях, в которых TGF-β1 стимулировал миграцию меланомных клеток, Box5 эффективно ингибировал это влияние на миграцию. Следовательно, данные, по-видимому, служат доказательством того, что прямое вмешательство в сигнальную систему Wnt-5a посредством соединения, такого как Box5, может представлять собой эффективный новый терапевтический подход к селективному ингибированию метастаза злокачественной меланомы.
Использованные сокращения:
BMP - костный морфогенетический белок;
EMT - эпителиально-мезенхимный переход;
PKC - протеинкиназа C; и
TGF-β - трансформирующий фактор роста-β.
Фармацевтические составы
При их использовании в качестве лекарственных препаратов, соединения по этому изобретению обычно вводили в форме фармацевтических композиций. Эти соединения могут быть введены различными путями, включая пероральный и ректальный. Эти соединения являются эффективными в форме пероральных композиций. Такие композиции получают способом, хорошо известным в фармацевтической области, и они включают по меньшей мере одно активное соединение.
Это изобретение также включает фармацевтические композиции, которые содержат в качестве активного ингредиента одно или несколько соединений, описанных в настоящем описании, в комбинации с фармацевтически приемлемыми носителями. Для получения композиций по этому изобретению активный ингредиент обычно смешивают с эксципиентом, разводят эксципиентом или включают в такой носитель, который может быть в форме капсулы, пакета-саше, бумажном или другом контейнере. Когда эксципиент служит растворителем, он может быть твердым, полутвердым или жидким материалом, который действует как основа, носитель или среда для активного ингредиента. Таким образом, композиции могут быть в форме таблеток, пилюль, порошков, пастилок, пакетов-саше, капсул, эликсиров, суспензий, эмульсий, растворов, сиропов, мягких и твердых желатиновых капсул, суппозиториев и расфасованных порошков.
Для получения состава может быть необходимо измельчить соединение с получением соответствующего размера частицы до комбинирования с другими ингредиентами. Если соединение является практически нерастворимым, его обычно измельчают до размера частицы менее чем 200 меш. Если соединение является в значительной степени водорастворимым, размер частицы обычно регулируется путем измельчения с получением существенно однородного распределения в составе, например примерно 40 меш.
Некоторые примеры приемлемых эксципиентов включают лактозу, декстрозу, сахарозу, сорбит, маннит, крахмалы, аравийскую камедь, кальция фосфат, альгинаты, трагакант, желатин, кальция силикат, микрокристаллическую целлюлозу, поливинилпирролидон, целлюлозу, стерильную воду, сироп и метилцеллюлозу. Составы могут дополнительно включать: смазывающие агенты, такие как тальк, магния стеарат и минеральное масло; смачивающие агенты; эмульгирующие и суспендирующие агенты; консервирующие агенты, такие как метил- и пропил гидрокси-бензоаты; подслащивающие агенты; и ароматизирующие агенты. Композиции по изобретению могут быть получены так, чтобы обеспечить быстрое, непрерывное или замедленное высвобождение активного ингредиента после введения пациенту посредством используемых процедур, известных в данной области.
Композиции предпочтительно получают в стандартной лекарственной форме. Термин «стандартные лекарственные формы» относится к физически дискретным единицам, приемлемым в качестве однократных доз для человека и других млекопитающих, каждая единица содержит заранее определенное количество активного материала, рассчитанного для получения желаемого терапевтического эффекта в сочетании с приемлемыми фармацевтическими эксципиентами. Предпочтительно вышеприведенное соединение Формулы (I) применяется в не более чем примерно 20 весовых процентах фармацевтической композиции, более предпочтительно в не более чем примерно 15 весовых процентах с нахождением баланса с фармацевтически инертным носителем (носителями).
Активное соединение является эффективным в пределах широкого диапазона доз и обычно вводится в фармацевтически эффективном количестве. Например, когда лекарственное средство вводится через оральный путь, каждая доза содержит от примерно 1 мг до примерно 1000 мг, предпочтительно от примерно 2 мг до примерно 500 мг, более предпочтительно от примерно 5 мг до примерно 100 мг, еще более предпочтительно от примерно 5 мг до примерно 60 мг активного ингредиента. Следует понимать, однако, что количество фактически введенного соединения будет определяться врачом в свете соответствующих обстоятельств, включая патологическое состояние, которое нужно лечить, выбранный путь введения, конкретное введенное соединение и его относительную активность, возраст, массу и ответ индивидуального пациента, тяжесть симптомов пациента и тому подобное. С принципиальной точки зрения состав должен вводиться одновременно с приемом пищи и должен затем вводиться в количестве, обеспечивающем достаточное ингибирование липидов. Таким образом, организм может иметь потребность в некотором количестве липидов с диетологической точки зрения, и это может затем оказывать влияние на количество введенных ингибирующих соединений по изобретению. Действие соединений по изобретению происходит в тонкой кишке, и поэтому отсутствует дальнейшее влияние как таковое, за исключением возможных метаболитов соединений.
Для получения твердых композиций, таких как таблетки, основной активный ингредиент смешивают с фармацевтическим эксципиентом с формированием твердой предварительно полученной композиции, содержащей гомогенную смесь соединения по настоящему изобретению. Если относиться к этим предварительно полученным композициям как к гомогенным, это означает, что активный ингредиент диспергирован равномерно по всей композиции, так что композиция может быть легко подразделена на равные эффективные стандартные лекарственные формы, такие как таблетки, пилюли и капсулы. Этот твердый предварительный состав затем подразделяют на стандартные лекарственные формы типа, описанного выше, содержащие активный ингредиент по настоящему изобретению.
Таблетки, пилюли или гранулы по настоящему изобретению могут быть покрыты или же смешаны с получением лекарственной формы, обеспечивающей преимущество пролонгированного действия. Например, таблетка или пилюля может включать внутренний или внешний лекарственный компонент, последний может находиться в виде оболочки формы. Два компонента могут быть разделены энтеросолюбильным слоем, который препятствует разложению в желудке и обеспечивает прохождение внутреннего компонента в неизменном виде в двенадцатиперстную кишку или замедленное высвобождение. Различные материалы могут быть использованы для таких энтеросолюбильных слоев или покрытий, такие материалы включают ряд полимерных кислот и смесей полимерных кислот с такими материалами, как шеллак, цетиловый спирт и целлюлозы ацетат.
Таблетки, пилюли или гранулы по настоящему изобретению могут быть покрыты покрытием с непрерывным высвобождением, способствующим высвобождению в поджелудочной железе, поскольку липаза поджелудочной железы поставляется свободно в кишечник. Такое непрерывно высвобождаемое покрытие будет, таким образом, допускать небольшое высвобождение, при необходимости, в желудке, но обеспечивать полное высвобождение в верхней части тонкой кишки.
Например, таблетка может быть получена путем прессования или формования. Прессованные таблетки могут быть получены путем прессования на приемлемой машине. Композиция по настоящему изобретению в сыпучей форме, такой как порошок или гранулы, при необходимости смешивается со связующим лубрикантом, инертным растворителем и/или поверхностно активным, или диспергирующим агентом. Формованные таблетки могут быть получены путем формования на приемлемой машине смеси порошкообразного соединения, увлажненного инертным жидким растворителем.
В предпочтительном варианте осуществления изобретения по меньшей мере один фармацевтически приемлемый эксципиент представляет собой лубрикант, наполнитель или их смесь. Предпочтительные эксципиенты включают, но не ограничиваясь ими, лактозу, кроскармелозу, микрокристаллическую целлюлозу, предварительно желатинизированный крахмал и магния стеарат.
Лубриканты для получения дозированных составов фармацевтических композиций по изобретению включают, но не ограничиваясь ими, кукурузный крахмал, картофельный крахмал или другие крахмалы, желатин, природные и синтетические камеди, такие как аравийская камедь, натрия альгинат, альгиновую кислоту, другие альгинаты, порошкообразный трагакант, гуаровую камедь, целлюлозу и ее производные (например, этилцеллюлоза, целлюлозы ацетат, карбоксиметилцеллюлозы кальций, натрия карбоксиметилцеллюлоза), поливинилпирролидон, метилцеллюлозу, предварительно желатинизированный крахмал, гидроксипропил метилцеллюлозу (например, № 2208, 2906, 2910), микрокристаллическую целлюлозу и их смеси.
Приемлемые формы микрокристаллической целлюлозы включают, например, материалы, проданные как AVICEL-PH-101, AVICEL-PH-103 и AVICEL-PH-105 (поставляемый от FMC Corporation, American Viscose Division, Маркус Хук, Пенсильвания). Особенно приемлемое связующее вещество представляет собой смесь микрокристаллической целлюлозы и натрия карбоксиметилцеллюлозы, проданной как AVICEL-RC-581 FMC Corporation.
Примеры приемлемых наполнителей для использования в лекарственных формах соединений по изобретению включают, но не ограничиваясь ими, тальк, кальция карбонат (например, гранулы или порошок), микрокристаллическую целлюлозу, порошкообразную целлюлозу, декстраны, каолин, маннит, салициловую кислоту, сорбит, крахмал, предварительно желатинизированный крахмал и их смеси.
Как правило, от примерно 50 до примерно 99 весовых процентов твердой лекарственной формы по изобретению представляет связующее вещество и/или наполнитель.
Разрыхлители используются, чтобы вызвать разрушение таблетки при экспозиции в водной окружающей среде. Слишком большое количество разрыхлителя будет приводить к получению таблеток, которые могут разрушаться во флаконе вследствие атмосферной влажности; слишком маленькое количество разрыхлителя может быть недостаточно для того, чтобы произошло разрушение, и может, таким образом, изменять скорость и объем высвобождения соединения по изобретению из лекарственной формы. Таким образом, достаточное количество разрыхлителя, которое является не слишком маленьким и не слишком большим, отражается на изменении высвобождения лекарственного средства, которое должно быть использовано для формирования твердых лекарственных форм по изобретению. Количество используемого разрыхлителя варьирует исходя из типа состава и способа введения и легко прослеживается специалистом в данной области. Как правило, от примерно 0,5 до примерно 15 весовых процентов разрыхлителя, предпочтительно от примерно 1 до примерно 5 весовых процентов разрыхлителя может быть использовано в фармацевтической композиции.
Приемлемые разрыхлители, которые могут быть использованы для формирования твердых лекарственных форм, включают, но не ограничиваясь ими, агар-агар, альгиновую кислоту, кальция карбонат, микрокристаллическую целлюлозу, кроскармелозы натрия, кросповидон, полакрилина калий, натрия гликолат крахмала, картофельный или маниоковый крахмал, другие крахмалы, предварительно желатинизированный крахмал, другие крахмалы, глины, другие альгины, другие целлюлозы, камеди и их смеси.
Приемлемые лубриканты для использования в твердых лекарственных формах включают, но не ограничиваясь ими, кальция стеарат, магния стеарат, минеральное масло, легкое минеральное масло, глицерин, сорбит, маннит, полиэтиленгликоль, другие гликоли, стеариновую кислоту, натрия лаурил сульфат, тальк, гидрированное растительное масло (например, арахисовое масло, хлопковое масло, подсолнечное масло, сезамовое масло, оливковое масло, кукурузное масло и соевое масло), цинка стеарат, этил олеат, этил лаурат, агар и их смеси. Дополнительные лубриканты включают, например, силоидный силикагель (AEROSIL 200, произведенный W. R. Grace Co. Балтимор, Мэриленд), коагулированный аэрозоль синтетического кремния (проданный Degussa Co. Плано, Техас), CAB-O-SIL (пирогенный продукт кремния диоксида, проданный Cabot Co. Бостон, Массачусетс) и их смеси. Лубрикант может быть при необходимости добавлен, как правило, в количестве менее чем примерно 1 весовой процент фармацевтической композиции.
Предпочтительно каждая твердая лекарственная форма содержит от примерно 5 мг до примерно 3000 мг соединения по изобретению. Предпочтительно каждая твердая лекарственная форма содержит от примерно 5 мг до примерно 25 мг, примерно 100 мг, примерно 200 мг, примерно 250 мг или примерно 500 мг соединения по изобретению. Твердые лекарственные формы, приемлемые для орального введения, предпочтительно содержат от примерно 5 мг до примерно 200 мг соединения по изобретению.
Жидкие формы, в которые новые композиции по настоящему изобретению могут быть включены для перорального введения, включают водные растворы, соответственно ароматизированные сиропы, водные и ароматизированные эмульсии с пищевыми маслами, такими как кукурузное масло, хлопковое масло, сезамовое масло, кокосовое масло или арахисовое масло, а также эликсиры и подобные фармацевтические основы. Жидкие составы могут быть также использованы для ингаляционного введения, где активный компонент суспендируется в жидкости для введения с использованием носового распылителя. Тем самым активное соединение может быть поглощено слизистыми мембранами в носовом ходе или поглощаться легкими.
Другие композиции для носового введения представлены как сухие композиции, использующие газ-пропеллент для введения сухой композиции в носовой ход и/или легкие.
Кроме того, фармацевтические композиции, содержащие одно или несколько соединений по изобретению, могут быть введены в комбинации с любым другим приемлемым лекарственным средством, например для лечения желудочно-кишечных нарушений. Когда применяется комбинированная терапия, фармацевтическая композиция, содержащая соединение(я) по изобретению и вторичное лекарственное средство, может быть введена одновременно, последовательно или раздельно. Каждый компонент, используемый в комбинированной терапии, применяется в количестве, достаточном для его предусмотренного назначения. Например, вторичное лекарственное средство применяется в достаточных количествах для обеспечения уменьшения рассматриваемого симптома in vivo.
Предпочтительно диапазон доз для соединений по изобретению составляет от примерно 1 мг до примерно 1000 мг на дозу, более предпочтительно от примерно 2 мг до примерно 500 мг, даже более предпочтительно от примерно 5 мг до примерно 100 мг и еще более предпочтительно от примерно 5 мг до примерно 60 мг. К тому же конкретная используемая доза будет зависеть от пациента (возраст, масса и т.п.) и тяжести заболевания (легкое, умеренное, тяжелое). Наконец, может быть также получена фармацевтическая композиция, содержащая два активных ингредиента для введения лекарственных средств одновременно.
Введение представленного лекарственного средства (лекарственных средств) будет нормально происходить в связи с приемом пищи, когда липаза-колипаза выделяется в свободном виде вследствие переваривания пищи, и оптимальное ингибирование будет происходить ниже двенадцатиперстной кишки.
ПРИМЕРЫ
Следующие препараты и примеры предоставлены, чтобы дать возможность специалистам в данной области более четко понимать и применять на практике настоящее изобретение. Они не должны считаться как ограничивающие объем изобретения, но всего лишь иллюстрируют и представляют его.
Примеры состава
Пример 1
Получение твердых желатиновых капсул, содержащих следующие ингредиенты:
(мг/капсула)
Вышеупомянутые ингредиенты смешивают и заполняют твердые желатиновые капсулы в количестве 340 мг.
Пример 2
Формула таблеток, полученных с использованием ингредиентов, представленных ниже:
(мг/таблетка)
Компоненты смешивают и прессуют с формированием таблеток, каждая массой 240 мг.
Пример 3
Таблетки, каждая из которых содержит 30 мг активного ингредиента, получены следующим образом:
(мг/таблетка)
(в виде 10% раствора в стерильной воде)
Активный ингредиент, крахмал и целлюлозу пропускают через NO: 20 меш сито США и тщательно перемешивают. Раствор поливинилпирролидона смешивают с полученными порошками, который затем пропускают через 16 меш сито США. Полученные таким образом гранулы сушат при 50-60°C и пропускают через 16 меш сито США. Натрия карбоксиметил крахмал, магния стеарат и тальк предварительно пропускали через NO: 30 меш сито США, затем добавляли в гранулы, которые после перемешивания прессуют на таблеточной машине с получением таблеток, каждая массой 120 мг.
Пример 4
Капсулы, каждая из которых содержит 40 мг лекарственного средства, получены следующим образом:
(мг/капсула)
Активный ингредиент, крахмал и магния стеарат смешивают, пропускают через NO: 20 меш сито США и заполняют твердые желатиновые капсулы в количестве 150 мг.
Пример 5
Суппозитории, каждый из которых содержит 25 мг активного ингредиента, получены следующим образом:
Активный ингредиент пропускают через NO: 60 меш сито США и суспендируют в насыщенных глицеридах жирной кислоты, предварительно расплавленных с использованием минимально необходимого тепла. Смесь затем наливают в формочку для суппозитория номинальной емкостью 2,0 г и охлаждают.
Пример 6
Суспензии, каждая из которых содержит 50 мг лекарственного средства на дозу 5,0 мл, получены следующим образом:
Активный ингредиент, сахарозу и ксантановую камедь смешивают, пропускают через NO: 10 меш сито США и затем перемешивают с полученным ранее раствором микрокристаллической целлюлозы и натрия карбоксиметил целлюлозы в воде. Натрия бензоат, ароматизатор и краситель разводят в той же воде и добавляют с перемешиванием. Достаточное количество воды затем добавляют для получения необходимого объема.
Пример 7
Состав может быть получен следующим образом:
(мг/капсула)
Активный ингредиент, крахмал и магния стеарат смешивают, пропускают через NO: 20 меш сито США и наполняют твердые желатиновые капсулы в количестве 425,0 мг.
Другие приемлемые составы для использования в настоящем изобретении могут быть найдены в Remington's Pharmaceutical Sciences, под редакцией E.W. Martin (Mack Publishing Company, 18th ed., 1990).





Изобретение относится к новым неразветвленным карбаматным производным некоторых пептидов Wnt-5a, в частности к N-бутилоксикарбонильному производному, их фармацевтическим композициям и их использованию для лечения меланомы или рака желудка. 5 н. и 2 з.п. ф-лы, 9 ил., 7 пр.
1. Неразветвленное карбаматное производное пептида Wnt5-α, состоящее из пептида с SEQ ID. NO: 1, причем производное выбрано из группы, состоящей из N-метилоксикарбонильного производного, N-этилоксикарбонильного производного, N-n-пропилоксикарбонильного производного и N-бутилоксикарбонильного производного.
2. Применение неразветвленного карбаматного производного пептида Wnt5-α, состоящего из пептида с SEQ ID. NO: 1, для лечения меланомы и рака желудка, причем производное выбрано из группы, состоящей из N-метилоксикарбонильного производного, N-этилоксикарбонильного производного, N-n-пропилоксикарбонильного производного и N-бутилоксикарбонильного производного.
3. Способ лечения человека, страдающего меланомой или раком желудка, где терапевтически эффективное количество неразветвленного карбаматного производного пептида Wnt5-α, состоящего из пептида с SEQ ID. NO: 1, вводят указанному человеку, причем производное выбрано из группы, состоящей из N-метилоксикарбонильного производного, N-этилоксикарбонильного производного, N-n-пропилоксикарбонильного производного и N-бутилоксикарбонильного производного.
4. Способ профилактического лечения меланомы или рака желудка, где терапевтически эффективное количество неразветвленного карбаматного производного пептида Wnt5-α, состоящего из пептида с SEQ ID. NO: 1, вводят пациенту, находящемуся в зоне риска развития меланомы или рака желудка, причем указанное производное выбрано из группы, состоящей из N- метилоксикарбонильного производного, N-этилоксикарбонильного производного, N-n-пропилоксикарбонильного производного и N-бутилоксикарбонильного производного.
5. Фармацевтическая композиция для лечения меланомы и рака желудка, содержащая терапевтически эффективное количество неразветвленного карбаматного производного пептида Wnt5-α, состоящего из пептида с SEQ ID. NO: 1, в комбинации с одним или несколькими фармацевтически приемлемыми инертными эксципиентами и/или адъювантами, причем неразветвленным карбаматным производным является одно из группы N-метилоксикарбонильного, N-этилоксикарбонильного, N-n-пропилоксикарбонильного или N-бутилоксикарбонильного производного.
6. Фармацевтическая композиция по п.5, где композиция составлена как топическая композиция.
7. Фармацевтическая композиция по п.5, где композиция составлена как инъецируемая композиция.
| WO 2006130082 A1, 12.07.2006 | |||
| WO 1998023730 A1, 04.06.1998 | |||
| US 7247426 B2, 24.07.2007 | |||
| WO 2004030615 A2, 15.04.2004 | |||
| WO 1998023730 A1, 04.06.1998 | |||
| RU 2010147806 A, 10.06.2012 |

