ПЕРЕКРЕСТНАЯ ССЫЛКА НА СВЯЗАННЫЕ ЗАЯВКИ
По этой заявке испрашивается приоритет по Предварительной Заявке США No. 62/737,155, поданной 27 сентября 2018 г., все содержание которой включено в настоящий документ посредством ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение направлено на терапию для лечения рака, включающую введение антагониста SFRP2 либо в качестве монотерапии, либо в комбинации с антагонистом PD-1 одновременно или последовательно пациенту, нуждающемуся в этом.
Все публикации, патенты, патентные заявки и другие ссылки, приведенные в этой заявке, включены в настоящий документ посредством ссылки в их полном объеме для всех целей и в той же степени, как если бы каждая отдельная публикация, патент, патентная заявка или другая ссылка были специально и индивидуально указаны для включения посредством ссылки в их полном объеме для всех целей. Цитирование ссылки в настоящем документе не должно толковаться как признание того, что таковое является предшествующим уровнем техники для настоящего изобретения.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Лиганды Wnt представляют собой секретируемые гликопротеины, которые активируют нижестоящие эффекторы путем связывания с трансмембранными рецепторами, связанными с G-белком клеточной поверхности, известными как frizzled-рецепторы. Активация сигнализации Wnt участвует в нормальном эмбриональном развитии, но дисрегуляция этого пути вовлекалась в прогрессирование опухоли при различных видах рака (1, 2). Относящиеся к frizzled секретируемые белки (SFRP) ранее рассматривались как ингибиторы канонического Wnt-бета (β)-катенинового пути (1) с предположением того, что SFRP2 может быть супрессором опухоли. Однако несколько дополнительных исследований показали, что SFRP2 может действовать как агонист β-катенина, а не антагонист (3-7), что предполагает его роль в промотировании опухоли.
В настоящее время имеются убедительные доказательства вклада SFRP2 в промотирование роста опухоли при раке молочной железы (5, 8-11), ангиосаркоме (9, 10), остеосаркоме (12), рабдомиосаркоме (13), саркоме альвеолярной мягкой части (14), злокачественной глиоме (15), множественной миеломе (16), почечно-клеточном раке (2), раке предстательной железы (17), раке легкого (18) и меланоме (19). Кроме того, молекулярная визуализация in vivo SFRP2 показывает, что экспрессия SFRP2 увеличивается пропорционально размеру опухоли (20), и авторы изобретения показали, что мышиное моноклональное антитело к SFRP2 ингибирует рост ангиосаркомы и рака молочной железы in vivo (21). Кроме того, Techavichit et al показали, что SFRP2 высоко сверхэкспрессируется в метастатической остеосаркоме, а сверхэкспрессия в клетках низкометастатической остеосаркомы увеличивает метастазы in vivo, в то время как нокдаун SFRP2 в высокометастатической остеосаркоме уменьшает миграцию клеток и инвазию in vitro (12). Помимо прямого воздействия SFRP2 на опухолевые клетки SFRP2 участвует в опухолевом ангиогенезе (9, 10, 19, 22-24). Таким образом, SFRP2 играет двойную роль в прямой активации опухолевого роста и вторичном влиянии на активацию ангиогенеза.
В эндотелиальных клетках SFRP2 активирует неканонический путь Wnt/Ca2, а не канонический путь β-катенина, для стимулирования ангиогенеза (22, 24). Путь Wnt/Ca2+ опосредуется активированными G-белками и фосфолипазами. Это приводит к транзиторным повышениям свободного кальция в цитоплазме и активации фосфатазы, кальциневрина, которая дефосфорилирует ядерный фактор активированных Т-клеток (NFAT), который затем транслоцируется из цитоплазмы в ядро. Все больше данных подтверждают критическую роль NFAT в опосредовании роста опухоли, включая рост клеток, выживаемость, инвазию и ангиогенез (25). Белки NFAT также играют решающую роль в развитии и функционировании иммунной системы, включая активацию Т-клеток. В частности, ядерный NFAT взаимодействует с другими транскрипционными факторами для регуляции множества генов, участвующих в функциях иммунной системы (26), включая IL2 и циклооксигеназу 2 (27).
Комбинированная Терапия
Введение двух лекарственных средств для лечения определенного состояния, такого как рак, вызывает ряд потенциальных проблем. In vivo взаимодействия между двумя лекарственными средствами сложны. Эффекты любого отдельного лекарственного средства связаны с его абсорбцией, распределением и элиминацией. Когда в организм вводятся два лекарственных средства, каждое из них может влиять на всасывание, распределение и элиминацию другого и, следовательно, изменять эффекты другого. Например, одно лекарственное средство может ингибировать, активировать или индуцировать выработку ферментов, участвующих в метаболическом пути элиминации другого лекарственного средства (44). В одном из примеров было экспериментально показано, что комбинированное введение глатирамера ацетата (GA) и интерферона (IFN) отменяет клиническую эффективность любой терапии (49). В другом эксперименте сообщалось, что добавление преднизона в комбинированную терапию с IFN-β антагонизировало его ап-регуляторный эффект (48). Таким образом, когда два лекарственных средства вводятся для лечения одного и того же состояния, невозможно предсказать, будет ли каждый из них дополнять, не оказывать влияния или препятствовать терапевтической активности другого у субъекта.
Взаимодействие между двумя лекарственными средствами может не только влиять на предполагаемую терапевтическую активность каждого лекарственного средства, но и повышать уровень токсических метаболитов (44). Взаимодействие может также усиливать или ослаблять побочные эффекты каждого лекарственного средства. Следовательно, при введении двух лекарственных средств для лечения заболевания невозможно предсказать, какие изменения произойдут в отрицательном боковом профиле каждого лекарственного средства. В одном из примеров было отмечено, что комбинация натализумаба и интерферона β-1а увеличивает риск непредвиденных побочных эффектов (47, 45, 46).
Кроме того, трудно точно предсказать, когда проявятся эффекты взаимодействия между этими двумя лекарственными средствами. Например, метаболические взаимодействия между лекарственными средствами могут проявляться при первоначальном введении второго лекарственного средства, после того как оба лекарственных средства достигли стационарной концентрации или после прекращения приема одного из лекарственных средств (44).
Таким образом, состояние уровня техники на момент подачи заявки таково, что эффекты дополнительной или комбинированной терапии двумя лекарственными средствами, в частности антагонистом SFRP2 совместно с антагонистом PD-1, не могут быть предсказаны с какой-либо разумной уверенностью до тех пор, пока не будут доступны результаты комбинированного исследования.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение направлено на фармацевтическую комбинацию, содержащую терапевтически эффективное количество антагониста SFRP2, CD38 и/или PD-1 и терапевтически эффективное количество антагониста PD-1. Изобретение также направлено на способ лечения рака, включающий одновременное или последовательное введение терапевтически эффективного количества антагониста SFRP2, CD38 и/или PD-1 и терапевтически эффективного количества антагониста PD-1 пациенту, нуждающемуся в этом. Изобретение также направлено на способ лечения некоторых видов рака, включающий введение терапевтически эффективного количества антагониста SFRP2, CD38 и/или PD-1 пациенту, нуждающемуся в этом.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Чертежи, описанные ниже, носят исключительно иллюстративный характер и не предназначены для ограничения объема изобретения.
Фигура 1: Реактивные Т-клетки селезенки мыши gp100 культивировали в течение 3 дней отдельно или в присутствии клеток Hs578T (верхний ряд) или RF420 (нижний ряд) и обрабатывали в течение 3 дней. Интенсивность измеряли для каждого состояния анализом FACS. В качестве положительного контроля в этом эксперименте были использованы антитела к CD3 и к CD28 (TCR стим). Процент супрессии вычисляли на основе способа определения индекса деления. Индекс деления рассчитывают путем умножения индекса пролиферации на процент делящихся клеток и, таким образом, представляет состояние деления всей популяции. Эксперименты повторяли трижды. Репрезентативное наложение представлено слева, в то время как совокупные данные всех повторов представлены на гистограмме (*р<0,01).
Фигура 2: А) Белок FZD5 присутствует в Т-клетках. В-С) Т-клетки обрабатывали SFRP2 (30 нМ) в течение 1 ч и выделяли (В) ядерную и (С) цитоплазматическую фракции. Образцы зондировали антителами к указанным белковым маркерам. D) Т-клетки обрабатывали антигеном gp100 (0,87 мкМ) или mAb hSFRP2 (10 мкМ) отдельно или в комбинации в течение 60 мин и выделяли ядерную фракцию. Уровни белка NFATc3 в обработанных SFRP2 клетках сравнивали с таковыми в необработанных клетках. А-С, Е). Актин: контроль загрузки для цитоплазматической фракции; Гистон Н3 и TATA: контроль загрузки для ядерных фракций. B-D) Денситометрию проводили с помощью knageJ и плотности вычисляли путем умножения средней интенсивности на поверхность каждой полосы. Контроль загрузки использовался для устранения межпробовой изменчивости. Окончательные результаты были получены путем нормализации каждого значения к необработанным контрольным (B-D) или обработанным антигеном образцам (D).
Фигура 3. А) Т-клетки селезенки обрабатывали либо IL2, либо IL2 + антигеном TCR, либо IL2 + антигеном TCR + TGFb. Или IL2 + антигеном TCR + TGFb и mAb hSFRP2. Белковые лизаты экстрагировали и подвергали Вестерн-блот-зондированию на SFRP2. Это показывает, что SFRP2 увеличивается с TCR и TGF b, который уменьшается с mAb hSFRP2. В) Концентрацию NAD+ спленоцитов мыши обрабатывали IL-2 (6000 ед/лунка), TCR/TGFp (5 нг/мл) или без него и mAb hSFRP2 (10 нМ) или без него в течение 3 дней (n=3 на группу). mAb hSFRP2 увеличивал концентрацию NAD+ по сравнению с необработанным контролем (*р=0,02). С) Количество клеток CD38+ (ось Z). Клетки обрабатывали так же, как и выше, за исключением 36 часов. Наблюдалось увеличение количества клеток CD38 + с добавлением TCR/TGFp, которое было достоверно ингибировано mAb hSFRP2 (n=3, ** р<0,001).
Фигура 4. mAb SFRP2 ингибирует PD-1 в Т-клетках. Т-клетки селезенки обрабатывают только IL2, или IL2 с антигеном TCR и TGFB, или IL2 с антигеном TCR и TGFB и mAb hSFRP2. Клетки анализировали с помощью FACS. TCR и TGFB увеличивают Гистограмму PD-1, которая инвертируется с mAb hSFRP2.
Фигура 5. А) Клетки RF420 остеосаркомы вводили внутривенно мышам C57BL6. Обработки контрольным IgG1 или mAb hSFRP2 (4 мг/кг каждые 3 дня), начиная спустя 10 дней после инъекции опухолевых клеток. Через три недели животных усыпляли, иссекали легкие и подсчитывали поверхностные узелки *: р≤0,0001; n=12). В) Репрезентативные легкие с метастазами опухоли. С) Т-клетки, выделенные из селезенок мышей C57BL/6, которым вводили клетки RF420 и обрабатывали контрольным IgG1 или mAb hSFRP2. Клетки окрашивали CD38, флуорохромом и анализировали среднюю интенсивность флуоресценции (MFI) с помощью FACS. Гистограмма, показывающая измерения флуоресценции, полученные из Т-клеток, выделенных из 4 различных селезенок для каждой обработки (n=4). CD38 статистически отличался от mAb hSFRP2 как в спленоцитах, так и в TIL* р≤0,001.
Фигура 6. Клетки RF420 остеосаркомы мыши вводили в хвостовую вену мышей C57BL/6. Начиная с 7-го дня мышей лечили либо контрольным IgG1, mAb hSFRP2, мышиным mAb PD-1, либо комбинацией обоих антител в течение 21 дня. Мышей усыпляли и забирали легкие. В каждой группе подсчитывали количество поверхностных метастазов и микрометастазов по Н&Е. При обработке mAb PD-1 не наблюдалось снижения числа мет. Наблюдалось достоверное снижение количества мет.при hSFRP2 в качестве монотерапии (р<0,001), которое в дальнейшем увеличивалось при комбинации (р<0,001).
Фигура 7: Активность in vitro гуманизированного mAb SFRP2. (А) Кривая концентрация-отклик ЕС50: половинная максимальная эффективная концентрация; Kd: равновесная константа диссоциации; Хилл: коэффициент Хилла. (В) Гистограмма (С-Н) Гистограммы, показывающие влияние увеличения концентрации mAb hSFRP2 (от 0 до 10 мкМ) на апоптоз (С, F; n=8) и некроз (D, G; n=8) пролиферацию (Е, Н; n=12) в клетках Hs578T рака молочной железы (С-Е) и клетках SVR ангиосаркомы (F-H). *: р≤0,05; **: р≤0,001. Пролиферацию измеряли с помощью Cyquant®, а апоптоз и некроз - с помощью Аннексина V и йодида пропидия. Результаты апоптоза и некроза представляют собой компиляцию 2 независимых экспериментов, содержащих по 4 лунки в каждом, n=8).
Результаты, представленные для пролиферации, представляют собой компиляцию 3 экспериментов, каждый из которых содержит 4 повтора (n=12).
Фигура 8: Влияние Гуманизированного mAb SFRP2 на рост опухоли при ангиосаркоме и раке молочной железы. A) AUC: Площадь Под Кривой; Т 1/2: Период Полувыведения; CL: клиренс; Vd: объем распределения; Стах: максимальная концентрация в сыворотке. Каждая точка данных представляет собой среднее ± SEM измерений для по меньшей мере 3 независимых образцов (n=3 в каждый момент времени). А-С) День отсчитывается от исходной даты, которая составляет 30 дней с момента инокуляции опухоли.
Фигура 9: Обработка гуманизированным mAb SFRP2 способствует апоптозу в опухолях. Верхняя Гистограмма показывает увеличение количества апоптотических клеток в опухолях, обработанных mAb hSFRP2 (белые полосы), по сравнению с опухолями, обработанными контрольным IgG1 (черные полосы). *:р≤0,05. Нижние изображения: Залитые парафином SVR (верхние панели) и Hs578T (нижние панели) опухоли были срезаны и обработаны для окрашивания TUNEL. Для каждой опухоли было сфотографировано в общей сложности 5 полей, подсчитано количество апоптотических клеток (коричневых) в каждом поле и усреднено для каждой опухоли. Для анализа использовали в общей сложности 10 опухолей на одну обработку (n=10).
Фигура 10: Гуманизированное mAb SFRP2 уменьшает рост метастатической остеосаркомы. А) Количество поверхностных узелков легкого после обработок. В) Спленоциты и TIL были собраны у мышей, обработанных контрольным IgG1 и mAb hSFRP2, и подвергнуты проточной цитометрии.
Фигура 11: Комбинация Гуманизированного mAb SFRP2 и ниволумаба ингибирует рост метастатической остеосаркомы. А) Количество поверхностных узелков легкого после различных обработок. В) График, показывающий измерения флуоресценции, полученные из Т-клеток, выделенных из 4 различных селезенок для каждой обработки (n=4), *** р≤0,001. Средняя интенсивность флуоресценции (MFI).
Фигура 12: Конкурентный анализ ELISA SFRP2 с использованием вариантных антител.
Фигура 13: SDS Page. 1 мкг очищенного лидера mAb hSFRP2 на 4-12% геле NuPAGE-SDS.
Фигура 14: Реакции пролиферации здоровых донорских Т-клеток на тестовые антитела.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Следует понимать, что терминология, используемая в настоящем документе, предназначена для описания конкретных вариантов выполнения изобретения и не предназначена для ограничения. Кроме того, хотя в практике или тестировании изобретения могут быть использованы любые способы, устройства и материалы, подобные или эквивалентные описанным в настоящем документе, далее описываются конкретные способы, устройства и материалы.
Изобретение относится к способу лечения рака, включающему введение количества антагониста SFRP2 и количества антагониста PD-1 субъекту, нуждающемуся в этом, причем количества, взятые вместе, эффективны для лечения субъекта. Изобретение также относится к фармацевтической комбинации, содержащей количество антагониста SFRP2, такого как mAb SFRP2, и количество антагониста PD-1, такого как антитело к PD-1. В одном варианте выполнения изобретение относится к новому гуманизированному моноклональному антителу к SFRP2 (mAb hSFRP2), которое уменьшает CD38 в спленоцитах и опухолевых инфильтрирующих лимфоцитах (TIL) in vivo и обладает превосходным сопутствующим эффектом с антителом PD-1 при ингибировании роста опухоли in vivo. В другом варианте выполнения изобретения гуманизированное моноклональное антитело к SFRP2 снижает PD-1 в лимфоцитах in vitro. Таким образом, mAb hSFRP2 по изобретению влияет на клеточные функции, ингибируя неканонический путь WNT в нескольких типах клеток.
В другом варианте выполнения изобретение относится к способу лечения рака, включающему введение терапевтически эффективного количества антагониста SFRP2, антагониста CD38 и/или антагониста PD-1 и терапевтически эффективного количества антагониста PD-1 субъекту, нуждающемуся в этом.
В другом варианте выполнения изобретение относится к способу лечения рака, включающему введение терапевтически эффективного количества антагониста SFRP2, антагониста CD38 и антагониста PD-1 и терапевтически эффективного количества антагониста PD-1 субъекту, нуждающемуся в этом.
В другом варианте выполнения изобретение относится к способу лечения рака, включающему введение терапевтически эффективного количества антагониста SFRP2 и/или антагониста CD38 и терапевтически эффективного количества антагониста PD-1 субъекту, нуждающемуся в этом.
В другом варианте выполнения изобретение относится к способу лечения рака, включающему введение терапевтически эффективного количества антагониста SFRP2 и антагониста CD38 и терапевтически эффективного количества антагониста PD-1 субъекту, нуждающемуся в этом.
В другом варианте выполнения изобретение относится к способу лечения рака, включающему введение терапевтически эффективного количества антагониста SFRP2 или антагониста CD38 и терапевтически эффективного количества антагониста PD-1 субъекту, нуждающемуся в этом.
В другом варианте выполнения изобретение относится к способу лечения рака, включающему введение терапевтически эффективного количества антагониста SFRP2 и терапевтически эффективного количества антагониста PD-1 субъекту, нуждающемуся в этом.
В одном варианте выполнения изобретения антагонист SFRP2 представляет собой: (а) антитело или антигенсвязывающий фрагмент антитела, который специфически связывается с рецептором SFRP2 и ингибирует его активацию, или (b) растворимую форму рецептора SFRP2, которая специфически связывается с лигандом SFRP2 и ингибирует связывание лиганда SFRP2 с рецептором SFRP2.
В одном варианте выполнения изобретения антагонист PD-1 представляет собой: (а) антитело или антигенсвязывающий фрагмент антитела, который специфически связывается с рецептором PD-1 и ингибирует его активацию, или (b) растворимую форму рецептора PD-1, которая специфически связывается с лигандом PD-1 и ингибирует связывание лиганда PD-1 с рецептором PD-1.
Саркомы представляют собой гетерогенную группу злокачественных новообразований, включающую >50 различных подтипов, каждый из которых обладает уникальными клиническими и патологическими свойствами. В целом, смертность составляет 50%, и большинство излечений достигается при полной хирургической резекции с лучевой терапией или без нее. Результаты применения химиотерапевтических агентов при нерезектабельном или метастатическом заболевании были неутешительными с минимальной долгосрочной пользой и 5-летней выживаемостью пациентов с метастатическим заболеванием всего 15% (34). Доксорубицин вызывает ответную реакцию от 20% до 25%. Для лечения сарком в последнее время изучаются ингибиторы PD-1. В ретроспективном исследовании 28 пациентов с метастатическими саркомами мягких тканей, получавших ниволумаб, 50% пациентов имели частичный ответ или стабильное заболевание (35). Несмотря на некоторую активность таргетных агентов при саркоме, для улучшения ответа и результата необходимы улучшенные терапевтические агенты и новые комбинации терапевтических средств. В этом исследовании авторы изобретения сообщают о разработке гуманизированного mAb SFRP2, которое не является иммуногенным и связывается с SFRP2 с высокой аффинностью. mAb hSFPR2 не только подавляет рост опухоли в качестве единственного агента в трех линиях опухолевых клеток (ангиосаркома, остеосаркома и карцинома-саркома молочной железы), но и при остеосаркоме этот эффект значительно превосходит действие только ингибитора PD-1.
Блокада либо рецептора PD-1, либо его лиганда PD-L1 улучшила общую выживаемость в исследованиях Фазы III у пациентов с меланомой, немелкоклеточным раком легкого и раком почки. Ранние исследования показывают, что блокада пути PD-1 может принести пользу подгруппе пациентов со многими другими типами рака. Тем не менее, большинство пациентов не реагируют на блокаду пути PD-1, и крайне необходимо понимание улучшения скорости ответа (36).
В то время как авторы изобретения и другие ранее показали роль SFRP2 на ангиогенез и опухолевые клетки (9, 10, 19, 20, 22, 24), настоящее исследование изобретателей раскрывает новый механизм: SFRP2 стимулирует NFAT не только в эндотелиальных клетках и опухолевых клетках, но и в Т-клетках. Учитывая, что индукция PD-1 после стимуляции TCR Т-клеток CD4 и CD8 требует NFAT, поскольку ингибитор пути кальциневрин/NFAT циклоспорин А способен блокировать PD-1 (37, 38), авторы изобретения предположили, что блокирование SFRP2 уменьшит истощение эффекторных Т-клеток и приведет к лучшему контролю опухоли. Данные авторов изобретения показывают, что в то время как экспрессия маркеров истощения, CD5 и CD103, не изменялась, наблюдалось снижение экспрессии неканонической эктонуклеотидазы CD38, экспрессия которой на Т-клетках также недавно была показана как обратно коррелирующая с контролем опухоли (39). CD38 регулирует противоопухолевый Т-клеточный ответ, а генетическая абляция или опосредованное антителами нацеливание CD38 на Т-клетки улучшает контроль опухоли. Кроме того, было также показано, что Т-клетки со сниженной экспрессией CD38 сохраняют высокую способность к секреции эффекторных цитокинов и не являются дисфункциональными, несмотря на экспрессию PD-1. Было также показано, что экспрессия CD38 высоко экспрессируется на неперепрограммируемых дисфункциональных Т-клетках PD1hi в фиксированном хроматином состоянии (40). Кроме того, комбинированная блокада PD-1 и CTLA-4 устраняет дефицитные по CD38 опухоли у мышей, а несущие опухоль мыши, получавшие комбинированные блокирующие PD-1 и CTLA-4 антитела, развивают резистентность через повышенную регуляцию CD38 (41). Таким образом, снижение экспрессии CD38 может спасти Т-клетки от истощения, вызванного опухолью.
Поскольку было показано, что кальциевая и NFAT сигнализация регулируют экспрессию CD38 в различных типах клеток (42), вполне вероятно, что его ингибирование SFRP2 привело к снижению сигнализации Ca2+/NFAT в Т-клетках, что привело к снижению экспрессии CD38. В В-клетках NFATc1, как сообщалось, имеет решающее значение для экспрессии CD38 (43), что привело авторов изобретения к гипотезе о том, что mAb hSFRP2 может снижать CD38 через его ингибирующее действие на NFATc3 в Т-клетках. Однако данные авторов изобретения подтверждают, что ингибирование SFRP2 наряду с PD-1 приводит к лучшему контролю опухоли, вероятно, благодаря нацеливанию как на сниженную иммуносупрессию из-за CD38 в Т-клетках, так и на сигнальный путь Wnt в опухолях. Без данных авторов изобретения не было никаких оснований ожидать, что такая комбинация будет лучше контролировать опухоль.
В одном варианте выполнения изобретения субъект имеет повышенную экспрессию CD38 и/или PD-1, если какие-либо клетки в организме субъекта, например, Т-клетки, имеют большую экспрессию CD38 и/или PD-1, чем соответствующий здоровый субъект или субъект с раком, который не имеет такой повышенной экспрессии.
Определения
Артикли «а» и «an» используются в этом раскрытии для обозначения одного или более чем одного (т.е. по меньшей мере одного) грамматического объекта артикля.
«Субъект» представляет собой человека, и термины «субъект» и «пациент» в настоящем документе используются взаимозаменяемо.
Термин «лечение» в отношении субъекта охватывает, напр., индуцирование ингибирования, регрессии или стазиса заболевания или расстройства; или лечение, улучшение или по меньшей мере частичное улучшение расстройства; или облегчение, уменьшение, супрессию, ингибирование, снижение тяжести, устранение или существенное устранение или улучшение симптома заболевания или расстройства. «Ингибирование» прогрессирования заболевания или осложнения заболевания у субъекта означает предотвращение или уменьшение прогрессирования заболевания и/или осложнения заболевания у субъекта.
«Симптом», связанный с раком, включает любое клиническое или лабораторное проявление, связанное с раком, и не ограничивается тем, что субъект может чувствовать или наблюдать.
«Введение субъекту» или «введение пациенту (человеку)» означает предоставление, выдачу или применение медицинских препаратов, лекарственных средств или лекарств субъекту/пациенту для облегчения, лечения или уменьшения симптомов, связанных с состоянием, напр., патологическим состоянием. Введение может быть периодическим введением.
В настоящем документе термин «периодическое введение» означает повторное/периодическое введение, разделенное определенным периодом времени. Период времени между введениями предпочтительно является последовательным от времени к времени. Периодическое введение может включать в себя введение, напр., один раз в день, два раза в день, три раза в день, четыре раза в день, еженедельно, два раза в неделю, три раза в неделю, четыре раза в неделю и т.д.
Как используется в настоящем документе, «разовая доза», «разовые дозы» и «единичная лекарственная форма(ы)» означают количество/количества (или форму/формы с количеством) лекарственного средства на одно введение.
Как используется в настоящем документе, «эффективный» или «терапевтически эффективный», когда речь идет о количестве антагониста PD-1 и/или антагониста SFRP2, относится к количеству антагониста PD-1 и/или антагониста SFRP2, которое является достаточным для получения желаемого терапевтического ответа. В некоторых вариантах выполнения изобретения эффективное количество относится к количеству, эффективному в дозировках и в течение периодов времени, необходимых для достижения желаемого терапевтического или профилактического результата. Терапевтически эффективное количество антагониста SFRP2, и/или антагониста PD-1/PD-L1, или ингибитора по изобретению может варьироваться в зависимости от таких факторов, как состояние заболевания, возраст, пол и вес индивидуума, а также способность антитела или антител вызывать желаемый ответ у индивидуума. Терапевтически эффективное количество включает количество, при котором любые токсические или вредные эффекты антитела или антител перевешиваются терапевтически полезными эффектами. В одном варианте выполнения изобретения количество антагониста SFRP2 и количество антагониста PD-1 при введении в комбинации являются эффективными для лечения субъекта. Согласно некоторым вариантам выполнения изобретения антитело по изобретению вводят в количестве от 0,1 мг/кг массы тела до 100 мг/кг массы тела. Согласно другим вариантам выполнения изобретения антитело по изобретению вводят в количестве от 0,5 мг/кг массы тела до 20 мг/кг массы тела. Согласно дополнительным вариантам выполнения изобретения антитело по изобретению вводят в количестве от 1,0 мг/кг массы тела до 10 мг/кг массы тела.
Нижеследующие сокращения общих терминов, используемых в настоящем описании, применяются независимо от того, появляются ли данные термины отдельно или в сочетании с другими группами:
Площадь под кривой (AUC); Бычий сывороточный альбумин (BSA); Кальций (Са2+); Карбоксифлуоресцеин сукцинимидиловый эфир (CFSE); Клиренс (CL); Константа диссоциации (Kd); Фермент-связанный иммуносорбентный анализ (ELISA); Фетальная Бычья Система (FBS); Флуоресцентно-активированная сортировка клеток (FACS); Frizzled 5 (FZD5); Гуманизированное моноклональное антитело к SFRP2 (mAb hSFRP2); Человеческий рекомбинантный секретируемый относящийся к frizzled белок 2 (hrSFRP2); Пероксидаза хрена (HRP); Половинная максимальная эффективная концентрация (ЕС50); Внутривенное (i.v.); Внутрибрюшинное (i.p.); Модификация Базальной Среды Игла (DMEM); Средняя Интенсивность флуоресценции (MIF); Некомпартментный анализ (NCA); Ядерный фактор активированных Т-клеток (NFAT); Фармакокинетика (PK); Белок 1 программируемой клеточной гибели (PD-1); относящийся к frizzled секретируемый белок 2 (SFRP2); Т-клеточный рецептор (TCR); Терминальный период полувыведения (Т1/2); и Объем распределения (Vd).
Комбинация по изобретению может быть составлена для ее одновременного, раздельного или последовательного введения по меньшей мере с фармацевтически приемлемым носителем, добавкой, адъювантом или транспортным средством, как описано в настоящем документе. Таким образом, комбинация двух активных соединений может быть введена:
• как комбинация, которая является частью одного и того же состава лекарственного средства, два активных соединения таким образом вводят одновременно, или
• как комбинация двух единиц, каждая с одной из активных субстанций, дающих возможность одновременного, последовательного или раздельного введения.
Как используется в настоящем документе, «комбинация» означает совокупность реагентов для применения в терапии либо одновременным, либо единовременным введением. Одновременное введение относится к введению смеси (будь то истинная смесь, суспензия, эмульсия или другая физическая комбинация) антагониста PD-1 и антагониста SFRP2, CD38 и/или PD-1. В этом случае комбинация может быть смесью или отдельными контейнерами антагониста PD-1, SFRP2, CD38 и/или антагониста PD-1, которые объединяются непосредственно перед введением. Единовременное введение или сопутствующее введение относится к раздельному введению антагониста PD-1 и антагониста SFRP2, CD38 и/или PD-1 в одно и то же время, или достаточно близко друг к другу, так, чтобы наблюдалась синергическая активность относительно активности либо одного антагониста PD-1, либо одного антагониста SFRP2, CD38 и/или PD-1, достаточно близко во времени, чтобы индивидуальные терапевтические эффекты каждого агента перекрывались.
Как используется в настоящем документе, «дополнение» или «дополнительная терапия» означает совокупность реагентов для применения в терапии, в которой субъект, получающий терапию, начинает первую схему лечения одним или несколькими реагентами до начала второй схемы лечения одним или несколькими отличающимися реагентами в дополнение к первой схеме лечения, так что не все реагенты, используемые в терапии, начинают принимать одновременно. Например, добавление терапии антагонистом PD-1 пациенту, уже получающему терапию антагонистом SFRP2, CD38 и/или PD-1.
В практике изобретения может быть использован любой известный антагонист PD-1, широкий спектр которого известен и раскрыт в данной области техники. Антагонист PD-1 предпочтительно нейтрализует биологическую функцию после связывания. Антагонист PD-1 предпочтительно представляет собой антагонист PD-1 человека. Необязательно антагонистом PD-1 может быть антитело, такое как моноклональное антитело или его фрагмент; химерное моноклональное антитело (такое как химерное моноклональное антитело чело века-мыши); полностью человеческое моноклональное антитело; рекомбинантное моноклональное антитело человека; фрагмент гуманизированного антитела; растворимый антагонист PD-1, включая низкомолекулярные блокирующие PD-1 агенты. Необязательно, антагонист PD-1 представляет собой функциональный фрагмент или слитый белок, содержащий функциональный фрагмент моноклонального антитела, такой как Fab, F(ab')2, Fv и предпочтительно Fab. Предпочтительно, фрагмент является пегилированным или инкапсулированным (напр., для стабильности и/или длительного высвобождения). Антагонистом PD-1 может быть также верблюжье антитело. Используемые в настоящем документе антагонисты PD-1 включают, но без ограничения, ингибиторы рецепторов PD-1.
Антагонист PD-1 может быть выбран, например, из одного или из комбинации ниволумаба, пембролизумаба, авелумаба, дурвалумаба, цемиплимаба или атезолизумаба или их функционального фрагмента.
Любой известный антагонист SFRP2 и/или CD38 может быть использован в практике изобретения, широкий спектр которых известен и раскрыт в данной области техники. Антагонист SFRP2 и/или CD38 предпочтительно нейтрализует биологическую функцию после связывания. Антагонист SFRP2 и/или CD38 предпочтительно является антагонистом SFRP2 и/или CD38 человека. Необязательно, антагонистом SFRP2 и/или CD38 может быть антитело, такое как моноклональное антитело или его фрагмент; химерное моноклональное антитело (такое как химерное моноклональное антитело человека-мыши); полностью человеческое моноклональное антитело; рекомбинантное моноклональное антитело человека; фрагмент гуманизированного антитела; растворимый антагонист SFRP2 и/или CD38, включая низкомолекулярные блокирующие SFRP2 и/или CD38 агенты. Необязательно, антагонист SFRP2 и/или CD38 представляет собой функциональный фрагмент или слитый белок, содержащий функциональный фрагмент моноклонального антитела, такой как Fab, F(ab')2, Fv и предпочтительно Fab.
Предпочтительно, фрагмент является пегилированным или инкапсулированным (напр., для стабильности и/или длительного высвобождения). Антагонистом SFRP2 и/или CD38 также может быть верблюжье антитело. Используемые в настоящем документе антагонисты SFRP2 и/или CD38 включают, но без ограничения, ингибиторы рецепторов SFRP2 и/или CD38. Например, антагонисты SFRP2 раскрыты в Патентах США No. 8,734,789 и 9,073,982, содержание которых настоящим включено посредством ссылки.
Далее изобретение будет дополнительно описано в приведенных ниже примерах, которые предназначены только для иллюстрации и не ограничивают объем изобретения.
ПРИМЕРЫ
Раскрытие далее иллюстрируется следующими примерами, которые не должны толковаться как ограничивающие это раскрытие по объему или духу конкретными процедурами, описанными в настоящем документе. Следует понимать, что примеры приведены для иллюстрации конкретных вариантов выполнения изобретения и что таким образом не предполагается никакого ограничения объема раскрытия. Следует также понимать, что можно прибегнуть к различным другим вариантам выполнения изобретения, модификациям и их эквивалентам, которые могут быть предложены специалистами в данной области техники, не отступая от духа настоящего раскрытия и/или объема прилагаемой формулы изобретения.
Пример 1
Гуманизированное mAb SFRP2 препятствует ингибированию пролиферации Т-клеток опухолевыми клетками. Поскольку SFRP2 активирует NFATc3 и белки NFAT регулируют пролиферацию Т-клеток (28), авторы изобретения исследовали, влияет ли mAb hSFRP2 на пролиферацию Т-клеток после активации со стимуляцией TCR (антитело к CD3/к CD28) (Фиг. 1). Т-клетки инкубировали отдельно, со стимуляцией TCR, со стимуляцией TCR + опухолевые клетки, со стимуляцией TCR + опухолевые клетки+контрольный IgG1 или со стимуляцией TCR + опухолевые клетки + mAb hSFRP2. По сравнению с пролиферацией, наблюдаемой только в популяции Т-клеток, антиген TCR (положительный контроль) увеличивал пролиферацию. Пролиферация была снижена в присутствии Hs578T, линии клеток метапластического рака молочной железы человека (Фигура 1). Добавление контрольного IgG1 к со-культуре не имело никакого эффекта. Для сравнения, добавление mAb hSFRP2 в со-культуры частично препятствовало пролиферации Т-клеток. Этот эффект также наблюдался при со-культивировании Т-клеток в присутствии клеток RF420 остеосаркомы мыши, где присутствие mAb hSFRP2 существенно препятствовало супрессии пролиферации, опосредованной опухолевой клеткой (Фиг. 1). Опять же, добавление контрольного IgG1 не влияло на пролиферацию по сравнению с Т-клетками, обработанными TCR в присутствии клеток RF420.
SFRP2 индуцирует сигнализацию Wnt в Т-клетках Рецептор FZD5 связывает SFRP2 в эндотелиальных клетках для стимуляции активации NFATc3 и ангиогенеза (23). Однако роль SFRP2 в активации Т-клеток и передаче сигналов Wnt ранее не оценивалась. Анализ Вестерн-блот лизатов Т-клеток показал, что белок FZD5 присутствует в Т-клетках (Фиг. 2А). Т-клетки селезенки мыши стимулировали SFRP2 (30 нМ) в течение 1 часа и выделяли ядерную и цитоплазматическую фракции. В цитоплазматической фракции наблюдалось увеличение CD38 при обработке SFRP2 (Фиг. 2В). В ядерной фракции наблюдалось увеличение NFATc3 при обработке SFRP2 (Фиг. 2С). Затем Т-клетки обрабатывали родственным антигеном в течение трех дней с mAb hSFRP2 или без него и собирали ядерные фракции. Наблюдалось увеличение NFATc3 в ядерной фракции при стимуляции родственным антигеном, a NFATc3 снижался в ядерной фракции при обработке mAb hSFRP2 (Фиг. 2D).
mAb hSFRP2 ингибирует PD-1 и CD38 в Т-клетках и восстанавливает NAD. Затем была проведена оценка того, ингибирует ли CD38 и восстанавливает ли уровень NAD+ в TGFβ-экспонированных Т-клетках обработка mAb hSFRP2 Т-клеток in vitro. TGFβ представляет собой цитокин, присутствующий в микроокружении опухоли, который повышает CD38 из Т-клеток. На Фигуре 3А показано, что обработка Т-клеток селезенки с помощью IL2, антигена TCR и TGFβ приводит к увеличению SFRP2 по вестерн-блот.Анализ FACS показал статистически значимое увеличение клеток CD38 + с добавлением TCR/TGFβ, которое было значительно ингибировано mAb hSFRP2 (Фигура 3c, n=3, р<0,001). Наряду с этим наблюдалось обратное увеличение концентрации NAD+ при обработке hSFRP2 (Фигура 3b, n=3, р=0,02). Кроме того, было рассмотрено, непосредственно ингибирует ли mAb SFRP2 PD-1 в Т-клетках селезенки. Т-клетки селезенки CD8+ и CD4+ обрабатывали с помощью IL2, антигена TCR и TGFβ, которые увеличивают PD-1. Это было обращено вспять с добавлением mAb SFRP2 (Фиг. 4).
Пример 2
Прогноз Остеосаркомы и Варианты Лечения. Остеосаркома (OS) представляет собой наиболее распространенную первичную злокачественную опухоль кости, обычно поражающую подростков и молодых взрослых. Если это возможно, первичная опухоль резецируется хирургическим путем с применением как неоадъювантной, так и адъювантной химиотерапии. Однако даже при химиотерапии излечиваются только две трети пациентов с изначально резектабельным заболеванием, при этом долгосрочная выживаемость наступает у <30% пациентов с метастатической или рецидивирующей опухолями. Легкие вовлечены в около 80% случаев метастатического заболевания, и последующий респираторный дистресс является причиной большинства летальных исходов (29). Хотя иммунотерапия показала эффективность при некоторых типах опухолей, введение пембролизумаба привело к недостаточной эффективности лечения остеосаркомы в исследовании фазы II (SARC028), в котором только 5% пациентов с метастатической остеосаркомой имели объективный ответ на пембролизумаб (30). Отсутствие новых активных агентов блокировало любой прогресс в повышении выживаемости пациентов с остеосаркомой на протяжении более трех десятилетий, и крайне необходимы новые подходы к лечению (31).
Все больше данных убедительно подтверждают вклад секретируемого SFRP2 в метастазы остеосаркомы. SFRP2 сверхэкспрессируется при метастатической остеосаркоме по сравнению с неметастатической остеосаркомой (32). Высокая экспрессия SFRP2 в образцах пациентов с OS коррелирует с плохой выживаемостью, а сверхэкспрессия SFRP2 подавляет нормальную дифференцировку остеобластов, способствует развитию особенностей OS и облегчает ангиогенез (33). Функциональные исследования показали, что стабильная сверхэкспрессия SFRP2 в локализованных клетках OS человека и мыши значительно увеличивает миграцию клеток и инвазивную способность in vitro, а также повышает метастатический потенциал in vivo. Дополнительные исследования по нокауту SFRP2 в метастатических клетках OS показали снижение способности клеток к миграции и инвазии in vitro, что подтверждает критический биологический фенотип, выполняемый SFRP2 (12). Таким образом, SFRP2 стал потенциальной терапевтической мишенью для остеосаркомы. Было также показано, что SFRP2 способствует росту опухоли при раке молочной железы (5, 8-11), ангиосаркоме (9, 10), рабдомиосаркоме (13), саркоме альвеолярной мягкой части (14), злокачественной глиоме (15), множественной миеломе (16), почечно-клеточном раке (2), раке предстательной железы (17), раке легкого (18) и меланоме (19). Учитывая недостаточную эффективность иммунотерапии при остеосаркоме и данные авторов изобретения, подробно описанные в другом месте этой заявки, авторы изобретения исследовали, будет ли комбинация гуманизированного моноклонального антитела к SFRP2 (mAb hSFRP2) усиливать активность ингибитора PD-1.
Гуманизированное mAb SFRP2 ингибирует метастазы in vivo. Для оценки противоопухолевой активности mAb hSFRP2 у иммунокомпетентной мыши mAb hSFRP2 протестировали на модели опухолевых метастазов, мышиной остеосаркоме RF420, у мышей C57BL/6. Клетки RF420 вводили в хвостовую вену мышей C57BL/6. Наличие метастазов в легких верифицировали через 7 дней после первичной инъекции опухолевых клеток. В первом эксперименте лечение mAb hSFRP2 (4 мг/кг вводили i.v. каждые 3 дня) начинали на 10-й день после инъекции опухоли и сравнивали с лечением контрольным IGg1. В конечной точке наблюдалось достоверное снижение количества легочных поверхностных узелков при лечении mAb hSFRP2 по сравнению с контролем (n=7, р<0,01, Фиг. 5). При оценке маркеров истощения клеточной поверхности мы заметили, что CD38, который, как было показано, тесно соэкспрессируется с PD-1 (41), был значительно снижен в спленоцитах (n=4, р<0,01) у мышей, получавших mAb hSFRP2, по сравнению с теми, кто получал контрольный IgG1 (Фиг. 5).
Введение mAb hSFRP2 с ингибитором PD-1 мыши эффективно ингибирует рост метастатической остеосаркомы in vivo. Клетки RF420 остеосаркомы мыши вводили в хвостовую вену мышей C57BL/6. Через 7 дней мышей лечили либо контрольным IgG1 4 мг/кг i.v. еженедельно, mAb hSFRP2 4 мг/кг i.v. каждые 3 дня, mAb PD-1 мыши (200 мкг/мышь) каждые 3 дня, либо комбинацией обоих антител. После 21 дня лечения мышей усыпляли и забирали легкие. В каждой группе подсчитывали количество поверхностных метастазов. Комбинация mAb hSFRP2 уменьшала количество поверхностных узелков по сравнению с контрольным IgG1 на 75% (Фиг. 6).
Способы Примеров 1-2
Антитела и Белки. Контрольный IgG1, омализумаб, был приобретен у компании Novartis (Basel, Switzerland). Рекомбинантный белок SFRP2 человека (SFRP2) был получен, как описано ранее (23), и предоставлен Protein Expression and Purification Core Lab Университета Северной Каролины в Чапел-Хилле. Гуманизированное моноклональное антитело к SFRP2 (mAb hSFRP2) было получено, как описано ранее и как описано в Примере 4, и очищено от эндотоксина.
В вестерн-блотах использовались следующие первичные антитела: антитела кролика к CD38 (#14637) и антитела кролика к гистону Н3 (#2650) были получены от Cell Signaling (Danvers, MA, USA), кролика к FZD5 (#H00007855-D01P, Abnova, Taipei city, Taiwan), мыши к PD1 (#66220-1, Proteintech, Rosemont, IL, USA), кролика к NFATc3 (#SAB2101578) и кролика к актину (#А2103, Sigma-Aldrich, St Louis, MO, USA). Вторичными антителами были: пероксидаза хрена (HRP)-конъюгированная к мыши (#7076, Cell Signaling); HRP-конъюгированная к кролику (#403005, Southern Biotech, Birmingham, AL, USA). Для анализа FACS антитело к CD38-PE крысы (#102707) было получено от BioLegend (San Diego, СА, USA). Анти-мышиный CD3 (#ВЕ00011) и анти-мышиный CD28 (#ВЕ0015-1) были получены от BioXCell (West Lebanon, NH, USA). Следующие антитела были приобретены в Biolegend, San Diego, СА и использованы для проточной цитометрии: к CD103 (клон 2Е7 cat # 121435), к CD5: Фрагмент антигена gp100 был получен от AnaSpec (#AS-62589).
Культура Клеток. Были получены клетки RF420 и мышиной остеосаркомы, полученные из генетически модифицированной модели остеосаркомы мыши (32). Клетки культивировали при 37°С в увлажненной атмосфере 5% CCh-95% комнатного воздуха. Клеточные линии были аутентифицированы АТСС® и мышиные клетки тестировались Charles River Research Animal (Wilmington, MA, USA) на наличие патогенов грызунов, включая микоплазму, всякий раз, когда они использовались in vivo.
Флуоресцентно-Активированная Сортировка Клеток (FACS) анализ клеточной пролиферации по измерению интенсивности сигнала Сукцинимидилового Эфира Карбоксифлуоресцеина (CFSE). Разбавление сигнала CFSE тесно коррелирует с увеличением пролиферации клеток. Т-клетки селезенки предварительно помечали красителем CFSE в соответствии с инструкциями CellTrace™ CFSE Cell Proliferation Kit (Thermo Fisher Scientific, Waltham, MA, USA). Затем клетки оставляли необработанными или активировали растворимым антителом к CD3 (#ВЕ0001-1, BioXCell, West Lebanon, NH, USA; 2 мкг/мл)/к CD28 (#BE0015-1, BioXCell; 2 мкг/мл) либо отдельно, либо в присутствии опухолевых клеток (RF420 или Hs578T карциномы-саркомы молочной железы) в соотношении 2:1 в течение 3 дней. Кроме того, некоторые со-культуры обрабатывали контрольным IgG1 (10 мкМ) или mAb hSFRP2 (10 мкМ). Через 3 дня Т-клетки из со-культур использовали для измерения интенсивности CFSE. Среднюю интенсивность флуоресценции (MFI) измеряли с помощью FACS, и анализ проводился с помощью программного обеспечения FlowJo.
Вестерн-блоты. Т-клетки селезенки обрабатывали в течение 1 часа SFRP2 (30 нМ) или без него или mAb hSFRP2 (10 мкМ). Контрольные клетки для SFRP2 получали только среду, а для экспериментов с mAb hSFRP2 получали 10 мкМ IgG1. Затем клетки центрифугировали при 1000 об/мин в течение 10 мин. Среду удаляли и клетки хранили замороженными при температуре -80°С перед обработкой. Экстракты ядер готовили с использованием реагента ядерной и цитоплазматической экстракции NE-PER, как описано в руководстве производителя (Pierce Biotechnology, Rockford, IL). Т-клетки селезенки, полученные от трансгенных мышей Pmell (The Jackson laboratory, Bar Harbor, ME, USA), обрабатывали в течение 1 часа rhSFRP2 (30 нМ) или без него или mAb hSFRP2 (10 мкМ). Контрольные клетки для rhSFRP2 получали только среду, а для экспериментов с mAb hSFRP2 получали 10 мкМ IgG1. Затем клетки центрифугировали при 1000 об/мин в течение 10 мин. Среду удаляли и клетки хранили замороженными при температуре -80°С перед обработкой. Экстракты ядер готовили с использованием реагента ядерной и цитоплазматической экстракции NE-PER, как описано в руководстве производителя (Pierce Biotechnology, Rockford, IL). Концентрацию белка измеряли с помощью Bio-Rad Protein Assay (Bio-Rad Laboratories, Hercules, CA, USA). Равные количества белка были загружены в гели SDS-PAGE. Белки переносили на поливинилидендифторидную мембрану, а вестерн-блоттинг проводили с использованием следующих первичных антител: антител кролика к CD38 и кролика к Гистону Н3, кролика к FZD5, мыши к PD1, кролика к NFATc3 и кролика к актину. Использовались следующие вторичные антитела: HRP-конъюгированное антимышиное и HRP-конъюгированное анти-кроличье. Для визуализации использовался субстрат ECL Advance (GE Healthcare Bio-Sciences, Piscataway, NJ, USA).
Затем мы оценили, ингибирует ли CD38 и восстанавливает ли уровень NAD+ в TGFβ-экспонированных Т-клетках обработка mAb hSFRP2 in vitro. Селезенки получали от мышей C57/BL6 и создавали суспензию из отдельных клеток и ресуспендировали в лизирующем буфере ACK в течение 1 минуты с PBS. Добавляли 1% FCS, чтобы остановить реакцию. Попав в суспензию отдельных клеток, клетки CD4+ и CD8+ выделяли отрицательным вычитанием с использованием смеси следующих антител: TCR119, CD25, GR1, NK1.1, CD11C, CD11B, CD19 и инкубировали на льду в течение 15 минут.Клетки инкубировали с 200 мкл раствора шариков, связанных со стрептавидином. После выделения клетки подсчитывали, и 400000 клеток помещали в пластины, предварительно покрытые aHTH-CD3 (2 мкг/мл) и анти-CD28 (5 мкг/мл). Отрицательный контроль содержал только выделенные клетки в среде, обогащенной IL-2, и не имел покрытия aHTH-CD3/Cd28 (TCR). Каждая экспериментальная лунка с клетками содержала TCR и IL-2 ((6000 Ед/мл) и одно из следующих экспериментальных условий: mAb hSFRP2 (10 мкМ) с TGFβ (5нг/мл) или без него. Все условия были выполнены в трех экземплярах и культивировались в течение трех дней. После эксперимента клетки подсчитывали и либо окрашивали для FACS, либо обрабатывали для анализа NAD с помощью набора для анализа на основе клеток NAD/NADH. Для анализа NAD требовалось не менее 250000 клеток, которые обрабатывались непосредственно в соответствии с протоколом NAD. Клетки центрифугировали и инкубировали при перемешивании с пермеабилизирующим буфером. После дополнительного центрифугирования образцы и стандарты инкубировали с реакционным буфером в течение 1 ч 30 мин при перемешивании. Оптические Плотности были окончательно считаны при 450 нм с помощью планшетного считывателя. Для анализа FACS 300000 клеток ресуспендировали в PBS и инкубировали в Живом-мертвом пятне по протоколу производителя, затем промывали PBS, закручивали с удалением супернатанта. Затем клетки суспендировали в мастер-смеси антител и окрашивающего буфера (50 мкл/образец), содержащей анти-CD38 РЕ/Су5 (1/200), анти-CD4 FITC (1/100), анти-CD8 АРС (1/200), анти-PD1 РЕ (1/200), буферизовали в течение 20 мин при комнатной температуре и защищали от света. Клетки окончательно фиксировали в 4% параформальдегиде в течение 10-15 минут, а затем ресуспендировали в 250 мкл окрашивающего буфера.
Рост метастатической остеосаркомы in vivo. В первом эксперименте клетки RF420 остеосаркомы (5×105), суспендированные в стерильном PBS, вводили i.v. через хвостовую вену 6-8-недельным мышам C57BL/6 (10 самок и 13 самцов). На 7-й день 2 мыши были умерщвлены, их легкие были удалены, зафиксированы в 10% формалине, залиты парафином и окрашены гематоксилином и эозином. Срезы рассматривали под микроскопом на наличие метастазов. Как только наличие метастазов было подтверждено, на 10-й день мышей лечили 4 мг/кг контрольного IgG1 или 4 мг/кг mAb hSFRP2 (n=10). После 3 недель лечения животных умерщвляли и удаляли легкие и селезенки. Затем подсчитывали поверхностные узелки в легких и сравнивали между группами лечения. Селезенки собирали свежими для выделения Т-клеток для проточной цитометрии.
Затем клетки RF420 (5×105), повторно суспендированные в стерильном PBS, вводили i.v. в хвостовую вену 6-8-недельных мышей-самцов и самок С57В1/6, купленных у Envigo (Indianapolis, IN, USA) (код штамма 044). Мыши были случайным образом распределены на 4 группы: контрольная (омализумаб, n=13); mAb hSFRP2 (n=11); мышиное mAb PD-1 (n=12); mAb PD-1 + mAb hSFRP2 (n=12). Лечение начинали через 10 дней после инокуляции опухолевых клеток. Дозировка, путь доставки и частота были следующими: контроль (омализумаб) - 4 мг/кг i.v. один раз в неделю; mAb hSFRP2 - 4 мг/кг i.v. каждые 3 дня; mab pd-1 - 8 мг/кг внутрибрюшинно (i.p.) каждые 3 дня. После 23 дней лечения животных умерщвляли, резецировали их легкие и подсчитывали поверхностные узелки. Поверхностные узелки подсчитывали по снимкам полных легких, сделанным сразу после резекции. Легкие фиксировали в формалине и заливали парафином. Они были секционированы и окрашены Н&Е.
Проточная цитометрия. Окрашивание для CD38 проводили путем инкубации спленоцитов из эксперимента с инъекциями остеосаркомы RF420 в хвостовую вену с первичными антителами к CD38 в буфере FACS (0,1% Бычий Сывороточный Альбумин (BSA) в PBS) в течение 30 мин при 4°С. Образцы были изучены на средние уровни интенсивности флуоресценции (MFI) на LSRFortessa и проанализированы с помощью программного обеспечения FlowJo (Tree Star, OR).
Статистика. Все расчеты мощности и размера выборки были выполнены с использованием PASS версии 08.0.13. Эксперименты in vitro проводились в трех экземплярах и повторялись трижды. Количественные показатели были собраны техником, слепым к экспериментальным условиям, чтобы смягчить потенциальное смещение. Групповые сравнения непрерывных показателей проводили с использованием двух выборочных Г-тестов или ANOVA для двух- или мультигрупповых сравнений соответственно.
Пример 3
Гуманизация mAb SFRP2. Химерные антитела и комбинации композитных тяжелых и легких цепей (всего 16 антител) были протестированы на связывание с SFRP2 в конкурентном анализе ELISA. В частности, серия разведений очищенных полностью человеческих композитных вариантов IgG конкурировала с фиксированной концентрацией биотинилированного антитела мыши для связывания с SFRP2 Пептидом В. Затем связанное биотинилированное mAb 80.8.6 (mAb SFRP2 мыши) было обнаружено с использованием субстрата стрептавидина HRP и ТМВ. Это продемонстрировало, что эффективность связывания всех композитных антител с SFRP2 была в целом сопоставима с эффективностью химерных антител, причем все варианты показали улучшение по сравнению с антителом мыши (Фиг. 12). Химерные антитела и композитные варианты анти-SFRP2 очищали от супернатантов клеточных культур на колонке с Протеин А-сефарозой, буфером, обмененным на PBS рН 7,4, и количественно определяли с помощью OD280нм с использованием коэффициента экстинкции (Ес (0,1%) = 1,76) на основе предсказанной аминокислотной последовательности. Тестирование эндотоксина лидирующего mAb hSFRP2 показало эндотоксин <0,5 EU/m. SDS-PAGE лидирующего mAb hSFRP2 показала две полосы, соответствующие тяжелой и легкой цепям (Фиг. 13). На Фиг. 13, SDS Page. 1 мкг очищенного лидирующего mAb hSFRP2 загружали на 4-12% гель NuPAGE-SDS. Предварительно окрашенный PageRuler Plus маркер был загружен, чтобы позволить калибровку полос.Полоса 1 была восстановлена β-меркаптоэтанолом; для образца присутствовали две полосы, соответствующие тяжелой цепи и легкой цепи. Полоса 2 была неукороченной.
Тестирование иммуногенности mAb hSFRP2. Лидирующие полностью гуманизированные и химерные антитела к SFRP2 были протестированы в отношении когорты из 22 здоровых доноров с использованием Т-клеточного анализа временной динамики EpiScreen™ для определения относительного риска иммуногенности. Полностью гуманизированное антитело к SFRP2 не индуцировало положительных реакций при использовании порога SI ≥ 2,0, р<0,05 ни у одного из доноров в анализе пролиферации, тогда как химерное антитело к SFRP2 индуцировало положительные реакции пролиферации Т-клеток у 23% доноров. Результаты с контрольным антигеном KLH показывают, что имелась хорошая корреляция (<10% вариабельности между анализами) между положительными и отрицательными результатами в повторных исследованиях, что свидетельствует о высоком уровне воспроизводимости в анализе (Фиг. 14). На Фиг. 14 были отобраны образцы РВМС из объемных культур и оценены на пролиферацию на 5, 6, 7 и 8-е сутки после инкубации с тремя тестовыми образцами. Ответы пролиферации с SI ≥ 2,0 (р<0,05), обозначенные пунктирной линией, которые были значимыми (р<0,05) при использовании непарного, двухвыборочного t-критерия Стьюдента считались положительными на этой Фигуре.
Гуманизированное mAb SFRP2 связывает SFRP2 с высокой аффинностью. Для того чтобы определить аффинность связывания лидирующего mAb hSFRP2 с SFRP2, rhSFRP2 (1 мкМ) инкубировали с возрастающими концентрациями mAb hSFRP2 в микропланшетном твердофазном анализе связывания белка ELISA. mAb hSFRP2 связало rhSFRP2 с ЕС50 8,72 нМ nKd 74,1 нМ. На Фиг. 1А показано, что гуманизированное mAb SFRP2 связывает rhSFRP2 с высокой аффинностью и дает кривую концентрация-реакция, показывающую поглощение 480 нм, измеренное после связывания возрастающих концентраций mAb hSFRP2 с заданной концентрацией 1 мкМ rhSRP2 в анализе ELISA (n=16).
Гуманизированное mAb SFRP2 ингибирует образование эндотелиальной трубки, пролиферацию опухолевых клеток и способствует апоптозу опухоли. В соответствии с предыдущими отчетами rhSFRP2 индуцировал увеличение числа точек ветвления по сравнению с контрольными клетками (n=4, р≤0,05). На Фиг. 7В представлена гистограмма, показывающая влияние rhSFRP2 и mAb hSFRP2 на формирование эндотелиальной трубки 2Н11. Для получения этих данных клетки 2Н11 инкубировали и обрабатывали либо только контрольным IgG1 (5 мкМ), либо IgG1 (5 мкМ) + белком rhSFRP2 (30 нМ), либо комбинацией rhSFRP2 (30 нМ) и mAb hSFRP2 (от 0,5 до 10 мкМ). n=4 *: р≤0,05; **: р≤0,001. И наоборот, увеличение концентрации mAb hSFRP2 значительно противодействовало влиянию rhSFRP2 на образование трубок (n=4, р≤0,05). IC50 для ингибирования посредством mAb hSFRP2 стимулированного SFRP2 образования трубок составляла 4,9±2 мкМ.
Влияние mAb rhSFRP2 на пролиферацию опухолевых клеток, апоптоз и некроз в клетках Hs578T карциномы/саркомы молочной железы человека и клетках SVR ангиосаркомы мыши оценивали in vitro. Лечение mAb hSFRP2 значительно увеличивало апоптоз опухоли как в клетках рака молочной железы Hs578T (Фиг. 7С; р≤0,05 и р≤0,001 для 5 мкМ и 10 мкМ mAb hSFRP2 соответственно), так и в клетках SVR ангиосаркомы (Фиг. 7F; р≤0,001 как для 5 мкМ, так и для 10 мкМ mAb hSFRP2) без изменения некроза. Лечение mAb hSFRP2 не оказывало влияния на пролиферацию SVR (Фиг. 7Н), но значительно снижало пролиферацию опухолевых клеток Hs578T рака молочной железы (Фиг. 7Е, 5 мкМ р≤0,05, 10 мкМ р≤0,001).
Определение эффективности и токсичности mAb hSFRP2 in vivo. Мышей, инокулированных клетками SVR ангиосаркомы, лечили mAb hSFRP2 в дозах 2, 4, 10 и 20 мг/кг i.v. каждые три дня; или контрольным IgG1 в течение 21 дня. Ни у одной из мышей, обработанных антителами, не было потери веса или летаргии. Патологических изменений в печени или легких не наблюдалось даже при дозе 20 мг/кг.В конце эксперимента массы оставались одинаковыми между группами (32,2±1,4 г для контроля; 31,3±1,1 г для 2 мг/кг; 32,1±0,5 г для 4 мг/кг; 31,8±0,9 г для 10 мг/кг и 32,7±1,0 г для 20 мг/кг.Доза с максимальным эффектом составила 4 мг/кг, где наблюдалось уменьшение объема опухоли на 69% (n=5 в группе, р=0,05).
Для изучения фармакокинетических свойств антитела голым мышам вводили однократную дозу mAb hSFRP2 4 мг/кг через хвостовую вену и собирали образцы крови в разные моменты времени (Фиг. 8). Лечение рекомбинантным hSFRP2 приводило к увеличению мембранного CD38 и ядерного белка NFATc3, в то время как mAb hSFRP2 ингибирует накопление ядерного NFATc3 в Т-клетках. Данные на Фиг. 8А демонстрируют, что белок FZD5 присутствует в Т-клетках. На Фиг. 8А представлен фармакокинетический график, показывающий снижение концентрации mAb hSFRP2 в сыворотке крови мышей с течением времени после однократной i.v. инъекции 4 мг/кг.Период полувыведения антитела в сыворотке крови животных составил 4,1±0,5 суток при максимальной концентрации в сыворотке крови (Стах) 7,8±1,0 мг/л и клиренсе (CL) 13,0±0,6 мл/час.
Для подтверждения эффективности дозы, выявленной в эксперименте MTD, авторы изобретения повторили эксперимент с опухолями SVR ангиосаркомы на большем количестве животных (n=10 животных/группа) и начали лечить их 4 мг/кг mAb hSFRP2. Т-клетки обрабатывали rhSFRP2 (30 нМ) в течение 1 ч и процессировали с помощью набора NE-PER для разделения цитоплазматической и ядерной фракций (Фиг. 8В). Образцы зондировали антителами к указанным белковым маркерам, и уровни белков в обработанных клетках сравнивали с таковыми в необработанных клетках. Через 3 недели опухоли, обработанные mAb hSFRP2, были на 43% меньше, чем опухоли, обработанные контрольным IgG1 (1631,3±283 мм3 для контроля, 928,5±148 мм3 для mAb hSFRP2; р≤0,05).
Затем авторы изобретения рассмотрели, может ли mAb hSFRP2 влиять на рост других типов опухолей. Мышей с ксенотрансплантатами Hs578T карциномы-саркомы молочной железы лечили mAb hSFRP2 или контрольным IgG1. Т-клетки обрабатывали антигеном gp100 (0,87 мкМ) или mAb hSFRP2 (10 мкМ) отдельно или в комбинации в течение 60 мин и выделяли ядерные фракции (Фиг. 8С). Уровни белка NFATc3 в обработанных rhSFRP2 клетках сравнивали с таковыми в необработанных клетках. Сравнение между контрольной и каждой обработанной группой в каждый момент времени показало, что 22-й, 25-й дни лечения и все временные точки с 31-го дня от исходного уровня являются статистически значимыми (р=0,05). На самом деле у мышей, получавших mAb hSFRP2, наблюдалось уменьшение объема опухоли на 61% (n=11, *Р<0,05). Кроме того, не было никакой потери веса или летаргии ни у одной из получавших лечение мышей.
Гуманизированное mAb SFRP2 индуцирует апоптоз в опухолях in vivo. mAb hSFRP2 индуцирует апоптоз in vitro и ингибирует пролиферацию в клетках рака молочной железы, и авторы изобретения исследовали, сохраняются ли эти фенотипы in vivo. В то время как доля пролиферативных (положительных по Ki67) клеток не была затронута лечением mAb hSFRP2 по сравнению с контрольными IgG1 опухолями (23±1,6% против 29±4,2% для опухолей SVR; 18±2,7% против 18±2,8% для опухолей Hs578T, p=NS), доля апоптотических клеток увеличилась на 188% в опухолях SVR (8,4±0,9 в контрольном IgG1, 24,2±3,5 в опухолях mAb hSFRP2; n=10, р≤0,05) и на 181% в опухолях Hs578T (15,1±4,9 в контрольном IgG1, 42,4±3,9 в опухолях mAb hSFRP2; n=10, р≤0,05)(Фиг. 9).
Для того чтобы оценить противоопухолевую активность mAb hSFRP2 у иммунокомпетентной мыши, авторы изобретения протестировали mAb hSFRP2 в мышиной остеосаркоме RF420 у мышей C57BL/6 на модели опухолевых метастазов. Клетки RF420 остеосаркомы вводили в хвостовую вену мышей C57BL/6. На 10-й день было начато лечение контрольным IgG1 или mAb hSFRP2. Мышей усыпляли на 21-й день лечения и подсчитывали поверхностные узелки. У мышей, получавших mAb hSFRP2, наблюдалось достоверное снижение количества поверхностных узелков по сравнению с контролем (n=7, р<0,01, Фиг. 10А). При оценке маркеров клеточной поверхности на истощение авторы изобретения заметили, что CD38, который, как было показано, тесно соэкспрессируется с PD-1, был значительно снижен на спленоцитах (n=4, р<0,01) и TIL (n=4, р<0,01) у мышей, получавших mAb hSFRP2, по сравнению с контрольным IgG, без достоверной разницы в PD-1, CD103 и CD5 (n=3) (Фиг. 6В). Экспрессия других маркеров истощения, таких как PD-1, CD103, TNFα или CD5, не была значимой в спленоцитах или TIL (n=4, p=NS).
Во втором эксперименте с остеосаркомой клетки RF420 остеосаркомы вводили внутривенно иммунокомпетентным мышам. Исследование было разделено на четыре группы. Первую группу лечили mAb hSFRP2 4 мг/кг i.v. каждые 3 дня. Была также контрольная группа IGgl, группа, которой вводили ниволумаб, антитело к PD-1, каждые 3 дня в дозе 8 мг/кг i.v., и группа, которая получала как mAb hSFRP2, так и антитело к PD-1. Лечение начинали на 10-й день после инъекции, а через три недели животных усыпляли, резецировали их легкие и подсчитывали поверхностные узелки *: р≤0,0001; **: р≤0,01, n=12). Эти группы сравнивали для измерения развития метастазов в легких. Каждая индивидуальная обработка уменьшала количество поверхностных узелков по сравнению с контрольным IgG1 (43,6±6,8 для контрольного IgG1, 18,3±3,4 для mAb hSFRP2, 16,3±1,1 для ниволумаба; р≤0,0001, Фиг. 11А). Наблюдалось снижение частоты метастатических поражений на 80% по сравнению с мышами, получавшими комбинацию mAb hSFRP2 и ниволумаба, и мышами, получавшими контрольный IgG1 (IRR=0,20, 95% CI=0,13-0,32; р<0,0001). Наблюдается снижение частоты метастатических поражений на 51% по сравнению с мышами, получавшими комбинацию mAb hSFRP2 и ниволумаба, и мышами, получавшими один агент mAb hSFRP2 (IRR=0,49, 95% CI=0,31-0,77; p=0,0021). Наблюдается снижение частоты метастатических поражений на 45% по сравнению с мышами, получавшими комбинацию mAb hSFRP2 и ниволумаба, и мышами, получавшими единственный агент ниволумаб (IRR=0,55, 95% CI=0,35-0,86; р=0,0084) (Фиг. 11А).
Авторы изобретения измерили влияние ниволумаба и mAb hSFRP2, назначаемых в качестве индивидуальных методов лечения или в комбинации, на уровень CD38 в Т-клетках мыши. В частности, Т-клетки выделяли из селезенки мышей C57BL/6, которым вводили клетки RF420 и обрабатывали контрольным IgG1, mAb hSFRP2, ниволумабом или комбинацией mAb hSFRP2 и ниволумаба. Затем клетки окрашивали CD38, меченным флуорохромом, и анализировали среднюю интенсивность флуоресценции (MFI) посредством FACS. Лечение ниволумабом само по себе не оказывало влияния на уровень CD38. Однако mAb hSFRP2 снижал поверхностную экспрессию CD38 в Т-клетках по сравнению с Т-клетками, полученными из группы, обработанной контрольным антителом IgG (р<0,001, Фиг. 11В), что указывает на то, что нацеливание на SFRP2 является достаточным для снижения экспрессии CD38 на Т-клетках. Эти результаты подтверждают предположение о том, что введение hSFPR2 mAb может восстановить Т-клеточный иммунный ответ и предотвратить рост опухоли. Следует отметить, что поскольку ниволумаб является человеческим антителом, он не лучше всего подходит для лечения на мышиной модели.
Гуманизация моноклонального антитела к SFRP2. Гены V-области, кодирующие мышиное моноклональное антитело 80.8.6 к SFRP2 (21), были первоначально клонированы и использованы для создания химерных антител, содержащих мышиные V-области в сочетании с константными областями тяжелой цепи и константными областями легкой цепи κ IgG1 человека. Химерные антитела и комбинации композитных тяжелых и легких цепей (всего 16 антител) экспрессировали в клетках NS0 или HEK293, очищали и тестировали на связывание с пептидом SFRP2 в конкурентном анализе ELISA.
Тестирование на иммуногенностъ. Лидирующее полностью гуманизированное антитело к SFRP2 (VH2/VK5) и эталонное химерное антитело к SFRP2 оценивали на иммуногенный потенциал с помощью Т-клеточных анализов временной динамики EpiScreen™, где объемные культуры получали с использованием РВМС, обедненных CD8+, а пролиферацию Т-клеток измеряли в различные моменты времени путем включения [3Н]-Тимидина после добавления образцов. Лидирующее полностью гуманизированное и химерное антитела к SFRP2 протестировали в отношении когорты из 22 здоровых доноров с использованием Т-клеточного анализа временной динамики EpiScreen™ для определения относительного риска неспецифической иммуногенности. Образцы были протестированы в конечной концентрации 50 мкг/мл на основе предыдущих исследований Antitope, показывающих, что эта насыщающая концентрация достаточна для стимуляции обнаруживаемых антителоспецифических Т-клеточных реакций. Для оценки иммуногенного потенциала каждого образца был использован Т-клеточный анализ временной динамики EpiScreen™ с анализом пролиферации для измерения активации Т-клеток. Поскольку образцы ранее не оценивались в анализе на основе РВМС, была определена первоначальная оценка любого грубого токсического воздействия образцов на жизнеспособность РВМС. Жизнеспособность клеток рассчитывали с использованием красителя трипанового синего, исключающего РВМС, через 7 дней после культивирования с тестируемыми образцами.
Антитела и Белки. В вестерн-блотах использовались следующие первичные антитела: антитела кролика к CD38 (#14637) и кролика к гистону Н3 (#2650) были получены от Cell Signaling (Danvers, MA, USA), кролика к FZD5 (#H00007855-D01P, Abnova, Taipei city, Taiwan), мыши к PD1 (#66220-1, Proteintech, Rosemont, IL, USA), кролика к NFATc3 (#SAB2101578) и кролика к актину (#А2103, Sigma-Aldrich, St Louis, MO, USA). Вторичными антителами были: HRP-конъюгированное анти-мышиное (#7076, Cell Signaling); HRP-конъюгированное анти-кроличье (#403005, Southern Biotech, Birmingham, AL, USA). Для ELISA: HRP-конъюгированный козий анти-человеческий IgG от Abeam, Cambridge, MA, USA. Для анализа FACS антитело к CD38-PE крысы (#102707) было получено от BioLegend (San Diego, СА, USA). Анти-мышиный CD3 (#ВЕ00011) и анти-мышиный CD28 (#ВЕ0015-1) были получены от BioXCell (West Lebanon, NH, USA). Контрольный IgG1, омализумаб, был приобретен у компании Novartis (Basel, Switzerland). Белок SFRP2 человека (rhSFRP2) был получен, как описано ранее. Фрагмент антигена gp100 был получен от AnaSpec (#AS-62589).
Микропланшетный Твердофазный Анализ Связывания Белка (ELISA) для Определения Аффинности Связывания rhSFRP2 с mAb hSFRP2. Для определения ЕС50 для rhSFRP2 и mAb hSFRP2 использовали микропланшетный твердофазный анализ связывания белка. Плоскодонные 96-луночные микропланшеты с покрытием Ni2+ (#15442, Thermo Fisher Scientific, Waltham, MA, USA) блокировали 0,05% бычьим сывороточным альбумином (BSA, #001-000-162, Jackson ImmunoResearch, West Grove, PA, USA) в фосфатном буферном физиологическом растворе (PBS, #ВР399-1, Fisher Scientific, Waltham, MA, USA) в течение ночи при температуре 4°С. 1 мкМ меченого гис rhSFRP2, разведенного в PBS (рН 7,4), инкубировали на блокированном планшете в течение ночи при температуре 37°С. Пластины промывали 3 раза 250 мкл/лунку PBS. Увеличивающиеся дозы mAb hSFRP2 в PBS (0 пМ, 100 пМ, 200 пМ, 400 пМ, 800 пМ, 1,6 нМ, 3,15 нМ, 6,3 нМ, 12,5 нМ, 25 нМ, 50 нМ, 100 нМ) инкубировали на пластине с rhSFRP2 при 37°С в течение ночи. Пластины промывали 3 раза, блокировали в течение 1 часа при комнатной температуре в 0,1% BSA в PBS и затем инкубировали со 100 мкл/лунку вторичного антитела (HRP-конъюгированного козьего анти-человеческого IgG), разведенного 1:40 000 в PBS, в течение 1 часа при 37°С. После того как пластины были промыты 5 раз, каждую лунку инкубировали со 100 мкл субстрата K-Blue ТМВ (#308176, Neogen, Lexington, KY, USA) в течение 5 минут в темноте. Реакцию останавливали 100 мкл 2н. H2SO4. Поглощение считывали при 450 нм. Расчеты ЕС50 были определены с помощью нелинейного регрессионного анализа с переменным наклоном с использованием GraphPad Prism log (ингибитор) по сравнению с нормализованной функцией отклика-переменного наклона с вершиной, ограниченной до 100%. ЕС50 был преобразован в Kd с использованием уравнения Ченга-Прусоффа, где концентрация агониста и ЕС50 были равны (40). Результаты выражаются в виде среднего значения ± стандартная ошибка среднего. Каждая точка данных является результатом 8 независимых измерений (n=8).
Культура Клеток. Эндотелиальные клетки 2Н11 мыши (#CRL-2163, АТСС®, Manassas, VA, USA) культивировали в Opti-MEM (#22600134, Thermo Fisher Scientific, Waltham, MA, USA) с 5% - ной инактивированной теплом фетальной бычьей сывороткой (FBS, #FB-12, Omega Scientific, Biel/Bienne, Switzerland) и 1%-ным пенициллином/стрептомицином (v/v). Тройные отрицательные клетки Hs578T карциномы-саркомы молочной железы человека (#30-202, АТСС®, Manassas, VA, USA) культивировали в DMEM (АТСС®) с 10% FBS, 0,01 мг/мл бычьего инсулина (#10516,Sigma-Aldrich, St. Louis, MO, USA) и 1% пенициллина/стрептомицина (#MT30009C, Thermo Fisher Scientific). Клетки SVR ангиосаркомы были получены из Американской Коллекции Типов Культур (#CRL-2280, АТСС®) и культивированы в Opti-MEM (Thermo Fisher Scientific) с 8% FBS и 1% пенициллином/стрептомицином (об./об.). Клетки RF420 остеосаркомы мыши, полученные из генетически модифицированной модели остеосаркомы мыши (41), были получены от доктора Джейсона Т. Юстейна (Texas Children's Cancer and Hematology Centers, Department of Pediatrics, Baylor College of Medicine, Houston, TX, USA) и культивированы в DMEM (АТСС®) с 10% термоинактивированным FBS и 1% пенициллином/стрептомицином (об./об.). Все клеточные линии культивировали при 37°С в увлажненной атмосфере 5% СО2-95% комнатного воздуха. Все клеточные линии были аутентифицированы АТСС® и мышиные клетки тестировались Charles River Research Animal (Wilmington, MA, USA) на наличие патогенов грызунов, включая микоплазму, всякий раз, когда они использовались in vivo. Мышиные Т-клетки были выделены от мышей C57BL/6 и gp100 реактивных TCR, несущих Pmel трансгенных мышей на фоне C57BL/6, полученных из Лаборатории Джексона (Bar Harbor, ME, USA).
Анализ образования эндотелиальной трубки. Эндотелиальные клетки 2Н11 покрывали в Opti-MEM 5% FBS и оставляли оседать в течение 24 часов. Покой индуцировался поддержанием клеток в Opti-MEM с 2,5% FBS в течение ночи. Matrigel™ (#ЕСМ625, Millipore, Bedford, MA, USA) полимеризовали в лунках 96-луночной пластины в соответствии с протоколом Анализа Ангиогенеза In vitro. В этом анализе было подготовлено девять условий обработки: только IgG1 (5 мкМ; омализумаб); белок rhSFRP2 (30 нМ) с IgG1 (5 мкМ); или rhSFRP2 (30 нМ) в сочетании с увеличением концентрации mAb hSFRP2 (0,5, 1, 5, 10 или 20 мкМ). Обработки, ресуспендированные в Opti-MEM с 2,5% FBS, предварительно инкубировали на качалке при 37°С, 5% СО2, в течение 90 минут перед добавлением их в клетки. 1,9×104 клеток ресуспендировали в 150 мкл предварительно инкубированных обработок, затем инкубировали еще 30 мин на качалке при 37°С, 5% СО2. Наконец, суспензию клеток добавляли в каждую лунку, уже покрытую полимеризованным Matrigel™. Каждый эксперимент повторяли 4 независимых раза, с n=4 в каждом условии. Контрольным клеткам давали свежий Opti-MEM с 2,5% FBS и 5 мкМ IgG1. Для каждого условия обработки после 4 ч инкубации при 37°С, 5% СО2 изображения получали с помощью 4-кратного объектива EVOS FL Digital Imaging System (Thermo Fisher Scientific, Waltham, MA, USA). Точки ветвления подсчитывали с помощью программного обеспечения Image J Angiogenesis Analysis (National Institutes of Health, Bethesda, MD, USA). В программном обеспечении GraphPad Prism данные были проанализированы для определения IC50 с использованием нелинейной регрессии и семейства уравнений «Доза-реакция - Ингибирование».
Анализ Пролиферации. Клетки Hs578T карциномы-саркомы молочной железы и SVR ангиосаркомы покрывали в 96-луночной пластине при 3000 клеток/лунку. Через 4 часа в питательную среду добавляли mAb hSFRP2 (1,5 или 10 мкМ) в указанных концентрациях. Клетки инкубировали в течение 72 часов при 37°С, 5% СО2. Пролиферацию оценивали с помощью набора Cyquant Direct Cell Proliferation Assay (#C35011, Thermo Fisher Scientific, Waltham, MA, USA). Изображения были получены с помощью EVOS FLc Digital hnaging System (Thermo Fisher Scientific). Клетки подсчитывали с помощью программного обеспечения FIJI cell counting.
Апоптоз/Некроз. Клетки Hs578T карциномы-саркомы молочной железы и SVR ангиосаркомы покрывали в 16-луночных предметных слайдах-камерах (#178599, Thermo Fisher Scientific, Waltham, MA, USA) при 2×104, 3×104 и 7,5×103 клеток/лунка соответственно. На следующий день клетки инкубировали при 37°С, 5% СО2 с 1, 5 или 10 мкМ mAb hSFRP2 или 5 мкМ контрольного IgG1 в суспензии с питательной средой в течение 2 часов. Некроз и апоптоз определяли в соответствии с протоколом для набора для Обнаружения Апоптоза/Некроза (#PK-СА707-30017, PromoCell, GmbH, Heidelberg, Germany). Изображения получали с помощью Цифровой Системы Визуализации EVOS FLc (Thermo Fisher Scientific, Waltham, MA, USA). Клетки подсчитывали с помощью программного обеспечения knageJ cell counting. Каждая точка данных была результатом 2 независимых повторов экспериментов, каждый из которых содержал 4 отдельных лунки (всего n=8).
Вестерп-блоты. Т-клетки селезенки, полученные от трансгенных мышей Pmell (The Jackson laboratory, Bar Harbor, ME, USA), обрабатывали в течение 1 часа rhSFRP2 (30 нМ) или без него или mAb hSFRP2 (10 мкм). Контрольные клетки для rhSFRP2 получали только среду, а для экспериментов с mAb hSFRP2 давали 10 мкМ IgG1. Затем клетки центрифугировали при 1000 об/мин в течение 10 мин. Среду удаляли и клетки хранили замороженными при температуре -80°С перед обработкой. Ядерные экстракты готовили с использованием реагента ядерной и цитоплазматической экстракции NE-PER, как описано в руководстве производителя (Pierce Biotechnology, Rockford, IL). Концентрацию белка измеряли с помощью Bio-Rad Protein Assay (Bio-Rad Laboratories, Hercules, CA, USA). Равные количества белка были загружены в гели SDS-PAGE. Белки переносили на поливинилидендифторидную мембрану, а вестерн-блоттинг проводили с использованием следующих первичных антител: антител кролика к CD38 и кролика к Гистону Н3, кролика к FZD5, мыши к PD1, кролика к NFATc3 и кролика к актину. Использовались следующие вторичные антитела: HRP-конъюгированное анти-мышиное и HRP-конъюгированное антикроличье. Для визуализации использовался субстрат ECL Advance (GE Healthcare Bio-Sciences, Piscataway, NJ, USA).
Анализ FACS клеточной пролиферации по измерению интенсивности сигнала CFSE. Разбавление сигнала CFSE тесно коррелирует с увеличением пролиферации клеток. Т-клетки селезенки трансгенных мышей Pmell предварительно помечали красителем CFSE в соответствии с инструкциями Набора CellTrace™ CFSE Cell Proliferation (Thermo Fisher Scientific, Waltham, MA, USA). Затем клетки оставляли необработанными или активировали растворимым антителом к CD3 (#ВЕ0001-1, BioXCell, West Lebanon, NH, USA; 2 мкг/мл)/к CD28 (#BE0015-1, BioXCell; 2 мкг/мл) либо отдельно, либо в присутствии опухолевых клеток (SVR ангиосаркомы или Hs578T карциномы-саркомы молочной железы) в соотношении 2:1 в течение 3 дней. Кроме того, некоторые со-культуры обрабатывали контрольным IgG1 (10 мкМ) или mAb hSFRP2 (10 мкМ). Через 3 дня Т-клетки из со-культур использовали для измерения интенсивности CFSE. Средняя интенсивность флуоресценции (MFI) измерялась с помощью FACS, и анализ проводился с помощью программного обеспечения FlowJo.
Максимальная Переносимая Доза (MTD) mAb hSFRP2 in vivo. Протоколы экспериментов на животных соответствовали руководящим принципам NIH по уходу и использованию лабораторных животных. 106 клеток SVR ангиосаркомы вводили подкожно в правый бок 6-недельным голым мышам самцам и самкам, полученным из Charles River (Wilmington, MA, USA). На следующий день мышей (n=5 в группе) обрабатывали i.v. контрольным PBS с различными концентрациями очищенного mAb hSFRP2 (2, 4, 10 и 20 мг/кг), вводимого через хвостовую вену каждые 3 дня. Животных лечили и измеряли объемы опухоли каждые три дня, пока контрольные опухоли не достигали среднего диаметра 2 см, который определяли как конечную точку. После эвтаназии опухоли, легкие и печени собирали и фиксировали в 10% формалине.
Фармакокинетическое исследование. Самцам и самкам мышей C57BL/6 вводили 4 мг/кг mAb hSFRP2 в разные моменты времени (0, 5 мин, 1, 2, 7, 14, 21, 28, 35 и 42 дня). Для каждой временной точки использовали по три мыши (n=3). В конечной точке образцы крови брали через воротную вену и помещали в разделительные трубки (#367981, Becton Dickinson, Franklin Lakes, NJ, USA). Образцы центрифугировали при 1300xg в течение 15 мин.
Микропланшетный Твердофазный Анализ Связывания Белка (ELISA) для Фармакокинетики (PK) mAb hSFRP2. Плоскодонные 96-луночные микропланшеты с покрытием Ni2+ блокировали 0,05% BSA в PBS в течение ночи при 4°С. 1 мкМ меченого гис rhSFRP2, разведенного в PBS (рН 7,4), инкубировали в течение ночи при 37°С. Пластины промывали 3 раза 250 мкл/лунку PBS. Затем в тарелку добавляли разведенную 1:50 мышиную сыворотку и инкубировали, слегка встряхивая при 37°С в течение ночи.
Пластины промывали 3 раза, блокировали в течение 1 часа при комнатной температуре в 0,1% BSA в PBS и затем инкубировали со 100 мкл/лунку вторичного антитела (HRP конъюгированного козьего анти-человеческого IgG), разведенного 1:40000 в PBS в течение 1 часа при 37°С. После того как пластины были промыты 5 раз, каждую лунку инкубировали со 100 мкл субстрата K-Blue ТМВ в течение 5 минут в темноте. Реакцию останавливали с помощью 100 мкл 2н. H2SO4. Поглощение считывали при 450 нм. Для оценки РК определяли AUC, t1/2, CL, Vd, Tmax и Cmax с помощью некомпартментного анализа (NCA) в EXCEL и (42). В NCA используется линейное трапециевидное правило для определения площади под кривой зависимости концентрации в плазмы от времени (AUC). Т1/2 представляет собой терминальный период полураспада. Для расчета AUC концентрации в нМ из ELISA были преобразованы в мг/л.
Ажотрансплантаты ангиосаркомы in vivo. 106 клеток SVR ангиосаркомы вводили подкожно в правый бок 6-недельных голых мышей самцов и самок, полученных из Charles River (Wilmington, MA, USA). На следующий день мышам (n=10 животных/группа) вводили i.v. mAb hSFRP2 (4 мг/кг) или контрольный IgG1 (омализумаб 4 мг/кг) через хвостовую вену и обрабатывали каждые 3 дня. Последовательные измерения штангенциркулем перпендикулярных диаметров, выполняемые два раза в неделю, использовались для расчета объема опухоли по следующей формуле: [(L (мм) х W (мм) × Н (мм)) × 0,5]. Мышей ежедневно контролировали на предмет оценки состояния организма и веса. Мышей умерщвляли, когда контроль достигал 2 см в диаметре, и опухоли резецировали, фиксировали в формалине и заливали парафином.
Ксенотрансплантаты Hs578T карциномы-саркомы молочной железы in vivo. Ксенотрансплантаты Hs578T помещали в 5-6-недельные самки голых мышей из Charles River (Wilmington, MA, USA). Мыши были инокулированы в жировое тело молочной железы 106 клетками, и лечение начинали, когда средний размер опухоли составлял приблизительно 100 мм3 (30-й день). Животным вводили 4 мг/кг mAb hSFRP2 (n=11) i.v. каждые 3 дня или 4 мг/кг контрольного IgG1 (n=11) до конечной точки, когда контрольные опухоли достигали 2 см в диаметре. Опухоли измеряли два раза в неделю с помощью штангенциркуля, а затем рассчитывали объем опухоли, как описано выше. Опухоли резецировали, фиксировали в формалине и заливали парафином.
Метастатическая RF420 остеосаркома in vivo. В первом эксперименте клетки RF420 остеосаркомы (5×105), суспендированные в стерильном PBS, вводили i.v. через хвостовую вену 6-8-недельным мышам C57BL/6 (10 самок и 13 самцов). На 7-й день 2 мыши были умерщвлены, их легкие были удалены, зафиксированы в 10% формалине, залиты парафином и окрашены гематоксилином и эозином. Срезы рассматривали под микроскопом на наличие метастазов. Как только наличие метастазов было подтверждено (на 10-й день мыши получали контрольный IgG1 4 мг/кг или mAb hSFRP2 4 мг/кг (n=10). После 3 недель лечения животных умерщвляли и удаляли легкие. Подсчитывали поверхностные узелки. Во втором эксперименте клетки RF420 (5×105), повторно суспендированные в стерильном PBS, вводили i.v. в хвостовую вену 6-8-недельных мышей самцов и самок С57В1/6, приобретенных у Envigo (Indianapolis, IN, USA) (код штамма 044). Мыши были случайным образом распределены на 4 группы: контрольная (омализумаб, n=13); mAb hSFRP2 (n=11); ниволумаб (NDC# 0003-3772-11, Bristol-Meyers Squibb, Princeton, NJ) (n=12); ниволумаб + mAb hSFRP2 (n=12). Лечение начинали через 10 дней после инокуляции опухолевых клеток. Дозировка, способ доставки и частота были следующими: контроль (омализумаб) 4 мг/кг внутривенно один раз в неделю; mAb hSFRP2 4 мг/кг внутривенно каждые 3 дня; ниволумаб 8 мг/кг внутривенно каждые 3 дня. После 23 дней лечения животных умерщвляли, резецировали их легкие и подсчитывали поверхностные узелки. Поверхностные узелки подсчитывали по снимкам полных легких, сделанным сразу после резекции. Селезенки собирали свежими для выделения Т-клеток, иммуногистохимии и туннельного анализа.
Иммуногистохимия. Фиксированные формалином, залитые парафином срезы опухоли дважды депарафинизировали в течение десяти минут в ксилоле и дважды гидратировали в абсолютном этаноле, дважды в 95%-ном этаноле, а затем в водопроводной воде. Срезы инкубировали в 3%-ной перекиси водорода в течение десяти минут при комнатной температуре с последующим двумя промывками в PBS IX. Стадию извлечения антигена из цитратного буфера проводили в овощной пароварке с использованием набора Vector Antigen Retrieval Citrate Buffer pH6 (H-3300) в течение 40 минут с 10 минутами для охлаждения. Срезы инкубировали в блокирующей сыворотке, входящей в Набор Vector Rabbit IMPRESS HRP (MP-4100), в увлажненной камере для срезов при комнатной температуре в течение 1 часа. Затем блокирующую сыворотку сливали, и предметные стекла инкубировали втечение ночи при 4°С сразведением антитела Ki67 1:40 (РА1-21520). На следующий день срезы промывали 3 раза в PBS в течение 5 мин/промывка. Добавляли вторичное антитело из набора Vector Rabbit IMPRESS HRP и инкубировали срезы в течение 30 мин RT, а затем 3 раза промывали в PBS в течение 5 мин/промывка. Раствор DAB готовили и добавляли к предметным стеклам в соответствии с инструкциями набора Vector DAB (SK-4100) в течение 5 мин, промывали в PBS и окрашивали гематоксилином в течение 30 секунд. Затем предметные стекла промывали дистиллированной водой, затем аммиачным спиртом, дважды обезвоживали в 95%-ном этаноле, дважды в 100%-ном этаноле, дважды в ксилоле, а затем устанавливали с помощью покровного стекла. Пролиферацию опухоли определяли количественно как количество положительно окрашенных клеток/единица площади, используя в среднем 3 поля на срез.
Анализ TUNEL. Срезы опухолей Hs578T и SVR окрашивали на наличие апоптотических клеток в соответствии с протоколом производителя набора для обнаружения апоптоза Apoptag® Peroxidase In Situ (#S7100). Все срезы были депарафинизированы с помощью Histoclear (#HS-200, National Diagnostics, Atlanta, GA, USA). Следующие материалы не поставлялись вместе с набором TUNEL и приобретались отдельно: 30% перекись водорода (#5155-01, J. Т. Baker, Phillipsburg, NJ, USA), Протеиназа К (#21627, Millipore, Burlington, MA, USA), Усиленный металлами набор субстратов DAB (#34065, Thermoscientific, Waltham, MA, USA), стабильный буфер субстратов пероксидазы IX (#1855910, Thermoscientific, Waltham, MA, USA) и 1-бутанол (#B7908, Sigma-Aldrich, St. Louis, MO, USA). Пять полей были случайным образом отобраны в каждом образце и сфотографированы с помощью микроскопа EVOS FLc (Life Technologies Inc., Waltham, MA, USA). В каждом поле апоптоз опухоли определялся количественно как количество апоптотических ядер/HPF.
Проточная цитометрия. Окрашивание на поверхностную экспрессию CD38 проводили путем инкубации спленоцитов из эксперимента с инъекциями остеосаркомы RF420 в хвостовую вену антителом к CD38-PE крысы (1:200; #102707, Biolegend, San Diego, СА, USA) в буфере FACS (0,1% BSA в PBS) в течение 30 мин при 4°С. Образцы подвергали скринингу на уровень средней интенсивности флуоресценции CD38 (MFI) на LSRFortessa и анализировали с помощью программного обеспечения Flow Jo (Tree Star, OR).
Статистика. Для анализов in vitro статистические различия между обработками IgG1 и mAb hSFRP2 были рассчитаны с использованием двустороннего t-критерия Стьюдента, при этом р≤0,05 считался значимым. Для исследования опухолей in vivo при ангиосаркоме (где лечение начинали на следующий день после инокуляции опухоли) использовали двусторонний t-критерий Стьюдента. Для Hs578T, где лечение было начато на 30-й день, когда опухоли были ощутимы, данные были нормализованы, чтобы скорректировать различия в исходных объемах опухоли, разделив объем опухоли с 34-го по 82-й день на исходный объем опухоли каждой группы (30-й день). Использовали двухпробный t-критерий для каждой временной точки и сравнивали объем опухоли между обработанными и контрольными животными. Для того чтобы удовлетворять предположению о нормальности для t-критерия, нормализованный объем опухоли логарифмически преобразуется. Для множественных сравнений в исследовании остеосаркомы авторы изобретения моделировали количество макрометастатических поражений в зависимости от группы лечения с использованием отрицательной биномиальной обобщенной линейной модели (NBGLM). Сравнение групп лечения проводили с использованием модельных линейных контрастов. Все анализы проводились с использованием R версии 3.2.3. Авторы изобретения суммировали коэффициенты частоты встречаемости (IRR) и соответствующие 95% доверительные интервалы (CI) при сравнении IgG1, mAb hSFRP2 и ниволумаба с комбинацией mAb hSFRP2 и ниволумаба. Авторы изобретения предполагали, что комбинированная терапия снизит частоту макрометастатических поражений по сравнению с одноагентной терапией, и поэтому построили IRR с лечением (IgG1, mAb hSFRP2 или ниволумаб), представленным в знаменателе, облегчая интерпретацию влияния комбинированной терапии по отношению к одному агенту.
СПИСОК ЛИТЕРАТУРЫ
1. Kawano Y, Kypta R. Secreted antagonists of the Wnt signalling pathway
1. J Cell Sci. 2003;116(Pt 13):2627-34.
2. Yamamura S, et al. Oncogenic functions of secreted Frizzled-related protein 2 in human renal cancer. MolCancer Ther. 2010;9(6):1680-7.
3. Esteve P, et al..Secreted frizzled-related proteins are required for Wnt/beta-catenin signalling activation in the vertebrate optic cup
1. Development. 2011;138(19):4179-84.
4. Gehmert S, Sadat S, Song YH, Yan Y, Alt E. The anti-apoptotic effect of IGF-1 on tissue resident stem cells is mediated via PI3-kinase dependent secreted frizzled related protein 2 (Sfrp2) release. BiochemBiophysRes Commun. 2008;371(4):752-5.
5. Lee JL, Chang CJ, Wu SY, Sargan DR, Lin CT. Secreted frizzled-related protein 2 (SFRP2) is highly expressed in canine mammary gland tumors but not in normal mammary glands. Breast Cancer Res Treat. 2004;84(2): 139-49.
6. Melkonyan HS, Chang WC, Shapiro JP, Mahadevappa M, Fitzpatrick PA, Kiefer MC, Tomei LD, Umansky SR. SARPs: a family of secreted apoptosis-related proteins. Proc NatlAcadSciUSA. 1997;94(25): 13636-41.
7. Mirotsou M, Zhang Z, Deb A, Zhang L, Gnecchi M, Noiseux N, Mu H, Pachori A, Dzau V. Secreted frizzled related protein 2 (Sfrp2) is the key Akt-mesenchymal stem cell-released paracrine factor mediating myocardial survival and repair. Proc NatlAcadSciUSA. 2007;104(5): 1643-8.
8. Gene Expression Omnibus [www.ncbi.nlm.nih.gov/geo/].
9. Bhati R, et al. Molecular characterization of human breast tumor vascular cells. Am J Pathol. 2008; 172(5): 1381-90.
10. Fontenot E, et al. A Novel Monoclonal Antibody to Secreted Frizzled Related Protein 2 Inhibits Tumor Growth. Molecular Cancer Therapeutics. 2013.
11. Lee JL, Chang CJ, Chueh LL, Lin CT. Secreted Frizzled Related Protein 2 (sFRP2) Decreases Susceptibility to UV-Induced Apoptosis in Primary Culture of Canine Mammary Gland Tumors by NF-kappaB Activation or JNK Suppression. Breast Cancer Res Treat. 2006;100(1):49-58.
12. Techavichit P, Gao Y, Kurenbekova L, Shuck R, Donehower LA, Yustein JT. Secreted Frizzled-Related Protein 2 (sFRP2) promotes osteosarcoma invasion and metastatic potential. BMC Cancer. 2016;16(1):869. doi: 10.1186/s12885-016-2909-6. PubMed PMID: 27821163; PMCID: PMC5100268.
13. Singh S, et al. Impaired Wnt signaling in embryonal rhabdomyosarcoma cells from p53/c-fos double mutant mice. The American journal of pathology. 2010;177(4):2055-66. doi: 10.2353/ajpath.2010.091195. PubMed PMID: 20829439; PMCID: PMC2947299.
14. Tanaka M, Homme M, Yamazaki Y, Shimizu R, Takazawa Y, Nakamura T. Modeling Alveolar Soft Part Sarcoma Unveils Novel Mechanisms of Metastasis. Cancer Res. 2017;77(4):897-907. doi: 10.1158/0008-5472.CAN-16-2486. PubMed PMID: 27979841.
15. Roth W, Wild-Bode C, Platten M, Grimmel C, Melkonyan HS, Dichgans J, Weller M. Secreted Frizzled-related proteins inhibit motility and promote growth of human malignant glioma cells. Oncogene. 2000; 19(37):4210-20.
16. Oshima T, Abe M, Asano J, Нага T, Kitazoe K, Sekimoto E, Tanaka Y, Shibata H, Hashimoto T, Ozaki S, Kido S, hioue D, Matsumoto T. Myeloma cells suppress bone formation by secreting a soluble Wnt inhibitor, sFRP-2. Blood. 2005; 106(9):3160-5. doi: 10.1182/blood-2004-12-4940. PubMed PMID: 16030194.
17. Sun Y, Zhu D, Chen F, Qian M, Wei H, Chen W, Xu J. SFRP2 augments WNT16B signaling to promote therapeutic resistance in the damaged tumor micro environment. Oncogene. 2016;35(33):4321-34. doi: 10.1038/onc.2015.494. PubMed PMID: 26751775; PMCID: 4994019.
18. Xiao X, Xiao Y, Wen R, Zhang Y, Li X, Wang H, Huang J, Liu J, Long T, Tang J. Promoting roles of the secreted frizzled-related protein 2 as a Wnt agonist in lung cancer cells. Oncol Rep.2015;34(5):2259-66. doi: 10.3892/or.2015.4221. PubMed PMID: 26323494; PMCID: 4583535.
19. Kaur A, et al. sFRP2 in the aged microenvironment drives melanoma metastasis and therapy resistance. Nature. 2016;532(7598):250-4. doi: 10.1038/naturel7392. PubMed PMID: 27042933; PMCID: PMC4833579.
20. Tsuruta JK, Klauber-DeMore N, Streeter J, Samples J, Patterson C, Mumper RJ, Ketelsen D, Dayton P. Ultrasound molecular imaging of secreted frizzled related protein-2 expression in murine angiosarcoma. PloS one. 2014;9(1):e86642. doi: 10.1371/journal.pone.0086642. PubMed PMID: 24489757; PMCID: 3906081.
21. Fontenot E, et al. A novel monoclonal antibody to secreted frizzled-related protein 2 inhibits tumor growth. MolCancer Ther. 2013;12(5):685-95. doi: 1535-7163.MCT-12-1066 [pii];10.1158/1535-7163.MCT-12-1066 [doi].
22. Siamakpour-Reihani S, Caster J, Bandhu ND, Courtwright A, Hilliard E, Usary J, Ketelsen D, Darr D, Shen XJ, Patterson C, Klauber-DeMore N. The Role of Calcineurin/NFAT in SFRP2 Induced Angiogenesis-A Rationale for Breast Cancer Treatment with the Calcineurin Inhibitor Tacrolimus. PLoSOne. 2011;6(6):e20412.
23. Peterson YK, Nasarre P, Bonilla IV, Hilliard E, Samples J, Morinelli ТА, Hill EG, Klauber-DeMore N. Frizzled-5: a high affinity receptor for secreted frizzled-related protein-2 activation of nuclear factor of activated T-cells c3 signaling to promote angiogenesis. Angiogenesis. 2017;20(4):615-28. doi: 10.1007/s10456-017-9574-5. PubMed PMID: 28840375; PMCID: PMC5675522.
24. Courtwright A, Siamakpour-Reihani S, Arbiser JL, Banet N, Hilliard E, Fried L, Livasy C, Ketelsen D, Nepal DB, Perou CM, Patterson C, Klauber-DeMore N. Secreted Frizzle-Related Protein 2 Stimulates Angiogenesis via a Calcineurin/NFAT Signaling Pathway. Cancer Res. 2009.
25. Mancini M, Toker A. NFAT proteins: emerging roles in cancer progression. Nat Rev Cancer. 2009;9(11):810-20. doi: 10.1038/nrc2735. PubMed PMID: 19851316; PMCID: PMC2866640.
26. Macian F. NFAT proteins: key regulators of T-cell development and function. Nat Rev Immunol. 2005;5(6):472-84. doi: 10.1038/nril632. PubMed PMID: 15928679.
27. Urso K, Alfranca A, Martinez-Martinez S, Escolano A, Ortega I, Rodriguez A, Redondo JM. NFATc3 regulates the transcription of genes involved in T-cell activation and angiogenesis. Blood. 2011;118(3):795-803. doi: 10.1182/blood-2010-12-322701. PubMed PMID: 21642596.
28. Dutta D, Barr VA, Akpan I, Mittelstadt PR, Singha LI, Samelson LE, Ashwell JD. Recruitment of calcineurin to the TCR positively regulates T cell activation. Nat Immunol. 2017;18(2):196-204. doi: 10.1038/ni.3640. PubMed PMID: 27941787.
29. Lindsey BA, Markel JE, Kleinerman ES. Osteosarcoma Overview. Rheumatol Ther. 2017;4(l):25-43. doi: 10.1007/s40744-016-0050-2. PubMed PMID: 27933467; PMCID: PMC5443719.
30. Tawbi HA, et al. Pembrolizumab in advanced soft-tissue sarcoma and bone sarcoma (SARC028): a multicentre, two-cohort, single-arm, open-label, phase 2 trial. Lancet Oncol. 2017;18(11):1493-501. doi: 10.1016/S1470-2045(17)30624-1. PubMed PMID: 28988646.
31. Wedekind MF, Wagner LM, Cripe TP. Immunotherapy for osteosarcoma: Where do we go from here? Pediatr Blood Cancer. 2018;65(9):e27227. doi: 10.1002/pbc.27227. PubMed PMID: 29923370.
32. Zhao S, et al. NKD2, a negative regulator of Wnt signaling, suppresses tumor growth and metastasis in osteosarcoma. Oncogene. 2015;34(39):5069-79. doi: 10.1038/onc.2014.429. PubMed PMID: 25579177; PMCID: PMC4802362.
33. Kim H, Yoo S, Zhou R, Xu A, Bernitz JM, Yuan Y, Gomes AM, Daniel MG, Su J, Demicco EG, Zhu J, Moore KA, Lee DF, Lemischka IR, Schaniel C. Oncogenic role of SFRP2 in p53-mutant osteosarcoma development via autocrine and paracrine mechanism. Proc Natl Acad Sci U S A. 2018;115(47):E11128-E37. doi: 10.1073/pnas.1814044115. PubMed PMID: 30385632; PMCID: PMC6255152.
34. Schoffski P, Cornillie J, Wozniak A, Li H, Hompes D. Soft tissue sarcoma: an update on systemic treatment options for patients with advanced disease. Oncol Res Treat. 2014;37(6):355-62. doi: 10.1159/000362631. PubMed PMID: 24903768.
35. Paoluzzi L, Cacavio A, Ghesani M, Karambelkar A, Rapkiewicz A, Weber J, Rosen G. Response to anti-PD1 therapy with nivolumab in metastatic sarcomas. Clin Sarcoma Res. 2016;6:24. doi: 10.1186/sl3569-016-0064-0. PubMed PMID: 28042471; PMCID: PMC5200964.
36. Zhang J, et al. Cyclin D-CDK4 kinase destabilizes PD-L1 via cullin 3-SPOP to control cancer immune surveillance. Nature. 2018;553(7686):91-5. doi: 10.1038/nature25015. PubMed PMID: 29160310; PMCID: PMC5754234.
37. Bally AP, Lu P, Tang Y, Austin JW, Scharer CD, Ahmed R, Boss JM. NF-kappaB regulates PD-1 expression in macrophages. J Immunol. 2015;194(9):4545-54. doi: 10.4049/jimmunol. 1402550. PubMed PMID: 25810391; PMCID: PMC4402259.
38. Oestreich KJ, Yoon H, Ahmed R, Boss JM. NFATcl regulates PD-1 expression upon T cell activation. J Immunol. 2008;181(7):4832-9. PubMed PMID: 18802087; PMCID: PMC2645436.
39. Chatterjee S, et al. CD38-NAD(+)Axis Regulates Immunotherapeutic Anti-Tumor T Cell Response. Cell Metab. 2018;27(1):85-100 e8. doi: 10.1016/j.cmet.2017.10.006. PubMed PMID: 29129787; PMCID: PMC5837048.
40. Philip M, et al.. Chromatin states define tumour-specific T cell dysfunction and reprogramming. Nature. 2017;545(7655):452-6. doi: 10.1038/nature22367. PubMed PMID: 28514453; PMCID: PMC5693219.
41. Chen L, et al. CD38-Mediated Immunosuppression as a Mechanism of Tumor Cell Escape from PD-1/PD-L1 Blockade. Cancer Discov. 2018. doi: 10.1158/2159-8290.CD-17-1033. PubMed PMID: 30012853.
42. Vukcevic M, Zorzato F, Spagnoli G, Treves S. Frequent calcium oscillations lead to NFAT activation in human immature dendritic cells. J Biol Chem. 2010;285(21):16003-11. doi: 10.1074/jbc. Ml 09.066704. PubMed PMID: 20348098; PMCID: PMC2871469.
43. Marklin M, Bugl, S., Bechtel, M., Poljak, A., Kopp, H., Kanz, L., Rao, A., Wirths, S., Muller, M. Ca2+/NFAT Signaling REgulates the Expression of CD38 and ZAP70 in Murine В Cells and Controls Bl a Cell Homeostasis. Blood. 2011; 118(21): 183.
44. Guidance for Industry. In vivo drug metabolism/drug interaction studies-study design, data analysis, and recommendations for dosing and labeling, U.S. Dept. Health and Human Svcs., FDA, Ctr. for Drug Eval. and Res., Ctr. For Biologies Eval. and Res., Clin./Pharm., November 1999 <http://www.fda.gov/cber/gdlns/metabol.pdf>.
45. Kleinschmidt-DeMasters et al. (2005) New England Journal of Medicine, 353:369-379.
46. Langer-Gould et al. (2005) New England Journal of Medicine, 353:369-379.
47. Rudick et al. (2006) New England Journal of Medicine, 354:911-923
48. Salama, Hassan H., et al. "Effects of Combination Therapy of Beta-Interferon la and Prednisone on Serum Immunologic Markers in Patients with Multiple Sclerosis." Multiple Sclerosis, vol. 9, no. 1, Feb. 2003, pp. 28-31. PubMed.
49. Brod et al. (2000) Annals of Neurology, 47:127-131.
50. Патенты США No. 8,734,789 и 9,073,982
* * *
Следует понимать, что изобретение не ограничивается конкретными вариантами выполнения изобретения, описанными выше, поскольку могут быть сделаны вариации конкретных вариантов выполнения изобретения и все же подпадающие под сферу применения прилагаемой формулы изобретения.
Изобретение будет далее описано, без ограничения, следующими пронумерованными параграфами:
1. Фармацевтическая комбинация, содержащая терапевтически эффективное количество антагониста SFRP2, антагониста CD38 и/или антагониста PD-1 и терапевтически эффективное количество антагониста PD-1.
2. Фармацевтическая комбинация в соответствии с параграфом 1, в которой антагонист SFRP2, CD38 и/или PD-1 представляет собой:
а. антитело или антигенсвязывающий фрагмент антитела, который специфически связывается с рецептором SFRP2, CD38 и/или PD-1 и ингибирует его активацию, или
b. растворимую форму рецептора SFRP2, CD38 и/или PD-1, которая специфически связывается с лигандом SFRP2, CD38 и/или PD-1 и ингибирует связывание лиганда SFRP2, CD38 и/или PD-1 с рецептором SFRP2, CD38 и/или PD-1.
3. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-2, где антагонист SFRP2, CD38 и/или PD-1 представляет собой моноклональное антитело (mAb) SFRP2, CD38 и/или PD-1.
4. Фармацевтическая комбинация в соответствии с параграфом 3, где моноклональное антитело к SFRP2 является человеческим или гуманизированным.
5. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-4, где антагонист PD-1 представляет собой:
a. антитело или антигенсвязывающий фрагмент антитела, который специфически связывается с рецептором PD-1 и ингибирует его активацию, или
b. растворимую форму рецептора PD-1, которая специфически связывается с лигандом PD-1 и ингибирует связывание лиганда PD-1 с рецептором PD-1.
6. Фармацевтическая комбинация в соответствии с параграфом 5, где антагонист PD-1 представляет собой растворимую форму рецептора PD-1 и лиганд PD-1 представляет собой PD-L1 или PD-L2.
7. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-5, где антагонист PD-1 представляет собой моноклональное антитело PD-1.
8. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-5, где антагонист PD-1 представляет собой ниволумаб.
9. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-5, где антагонист PD-1 представляет собой пембролизумаб, авелумаб, дурвалумаб, цемиплимаб или атезолизумаб.
10. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-9, где терапевтически эффективное количество антагониста SFRP2, CD38 и/или PD-1 составляет от 0,1 мг/кг массы тела до 100 мг/кг массы тела.
11. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-9, где терапевтически эффективное количество антагониста SFRP2, CD38 и/или PD-1 составляет 0,2-3, 0,27-2,70, 0,27, 0,54, 1,35 или 2,70 мг на кг массы тела.
12. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-11, где терапевтически эффективное количество антагониста SFRP2, CD38 и/или PD-1 составляет 10 мг - 200 мг, 17 мг, 33 мг, 84 мг или 167 мг
13. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-12, где терапевтически эффективное количество антагониста PD-1 составляет от 0,1 мг/кг массы тела до 100 мг/кг массы тела.
14. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-12, где терапевтически эффективное количество антагониста PD-1 составляет 0,02-1,2, 0,027-1,08, 0,027 или 1,08 мг/кг массы тела.
15. Фармацевтическая комбинация в соответствии с любым одним из параграфов 1-14, где терапевтически эффективное количество антагониста PD-1 составляет 1-80, 1,6-67, 1,6 или 67 мг/кг массы тела.
16. Способ лечения рака, включающий введение терапевтически эффективного количества антагониста SFRP2, CD38 и/или PD-1 и терапевтически эффективного количества антагониста PD-1 субъекту, нуждающемуся в этом.
17. Способ в соответствии с параграфом 16, где введение осуществляется одновременно или последовательно.
18. Способ в соответствии с параграфом 16 или 17, где антагонист SFRP2, CD38 и/или PD-1 представляет собой:
a. антитело или антигенсвязывающий фрагмент антитела, который специфически связывается с рецептором SFRP2, CD38 и/или PD-1 и ингибирует его активацию, или
b. растворимую форму рецептора SFRP2, CD38 и/или PD-1, которая специфически связывается с лигандом SFRP2, CD38 и/или PD-1 и ингибирует связывание лиганда SFRP2, CD38 и/или PD-1 с рецептором SFRP2, CD38 и/или PD-1.
19. Способ в соответствии с любым одним из параграфов 16-18, где антагонист SFRP2, CD38 и/или PD-1 представляет собой моноклональное антитело (mAb) SFRP2, CD38u/или PD-1.
20. Способ в соответствии с параграфом 19, где моноклональное антитело к SFRP2 является человеческим или гуманизированным.
21. Способ в соответствии с любым одним из параграфов 16-20, где антагонист PD-1 представляет собой:
a. антитело или антигенсвязывающий фрагмент антитела, который специфически связывается с рецептором PD-1 и ингибирует его активацию, или
b. растворимую форму рецептора PD-1, которая специфически связывается с лигандом PD-1 и ингибирует связывание лиганда PD-1 с рецептором PD-1.
22. Способ в соответствии с параграфом 21, где антагонист PD-1 представляет собой растворимую форму рецептора PD-1, и лиганд PD-1 представляет собой PD-L1 или PD-L2.
23. Способ в соответствии с любым одним из параграфов 16-22, где антагонист PD-1 представляет собой моноклональное антитело PD-1.
24. Способ в соответствии с любым одним из параграфов 16-23, где антагонист PD-1 представляет собой ниволумаб.
25. Способ в соответствии с любым одним из параграфов 16-23, где антагонист PD-1 представляет собой пембролизумаб, авелумаб, дурвалумаб, цемиплимаб или атезолизумаб.
26. Способ в соответствии с любым одним из параграфов 16-25, где указанный рак представляет собой рак молочной железы.
27. Способ в соответствии с любым одним из параграфов 16-26, где указанный рак представляет собой ангиосаркому, рак легкого, остеосаркому, меланому, немелкоклеточный рак легкого или рак почки.
28. Способ в соответствии с любым одним из параграфов 16-27, где введение антагониста SFRP2, CD38 и/или PD-1 предшествует введению антагониста PD-1.
29. Способ в соответствии с любым одним из параграфов 16-27, где введение антагониста PD-1 предшествует введению антагониста SFRP2, CD38 и/или PD-1.
30. Способ в соответствии с любым одним из параграфов 16-29, где антагонист SFRP2, CD38 и/или PD-1 вводят в дополнение к антагонисту PD-1.
31. Способ в соответствии с любым одним из параграфов 16-29, где антагонист PD-1 вводят в дополнение к антагонисту SFRP2, CD38 и/или PD-1.
32. Способ в соответствии с любым одним из параграфов 16-31, где антагонист SFRP2, CD38 и/или PD-1 вводят ежедневно, чаще одного раза в день или реже одного раза в день.
33. Способ в соответствии с любым одним из параграфов 16-31, где антагонист SFRP2, CD38 и/или PD-1 вводят один раз в 3 дня, один раз в неделю, один раз в 2 недели, один раз в 3 недели или один раз в 4 недели.
34. Способ по любому одному из параграфов 16-33, где антагонист PD-1 вводят ежедневно, чаще одного раза в день или реже одного раза в день.
35. Способ в соответствии с любым одним из параграфов 16-33, где антагонист PD-1 вводят один раз в 3 дня, один раз в неделю, один раз в 2 недели, один раз в 3 недели или один раз в 4 недели.
36. Способ в соответствии с любым одним из параграфов 16-35, где антагонист PD-1 представляет собой ниволумаб и количество ниволумаба, вводимого субъекту, составляет 3 мг/кг массы тела каждые 3 недели, 240 мг каждые 2 недели или 480 мг каждые 4 недели.
37. Способ в соответствии с любым одним из параграфов 16-35, где антагонист PD-1 представляет собой пембролизумаб и количество пембролизумаба, вводимого субъекту, составляет 200 мг каждые 3 недели.
38. Способ в соответствии с любым одним из параграфов 16-35, где антагонист PD-1 представляет собой авелумаб и количество авелумаба, вводимого субъекту, составляет 800 мг каждые 2 недели.
39. Способ в соответствии с любым одним из параграфов 16-35, где антагонист PD-1 представляет собой дурвалумаб и количество дурвалумаба, вводимого субъекту, составляет 10 мг/кг массы тела каждые 2 недели.
40. Способ в соответствии с любым одним из параграфов 16-35, где антагонист PD-1 представляет собой цемиплимаб и количество цемиплимаба, вводимого субъекту, составляет 250 мг каждые 3 недели.
41. Способ в соответствии с любым одним из параграфов 16-35, где антагонист PD-1 представляет собой атезолизумаб и количество атезолизумаба, вводимого субъекту, составляет 840 мг каждые 2 недели, 1200 мг каждые 3 недели или 1680 мг каждые 4 недели.
42. Способ в соответствии с любым одним из параграфов 16-41, где субъект получает терапию антагонистом PD-1 до начала терапии антагонистом SFRP2, CD38 и/или PD-1.
43. Способ в соответствии с любым одним из параграфов 16-41, где субъект получает терапию антагонистами SFRP2, CD38 и/или PD-1 до начала терапии антагонистами PD-1.
44. Способ в соответствии с параграфом 42 или 43, где субъект получает первую терапию в течение по меньшей мере 8 недель, по меньшей мере 10 недель, по меньшей мере 24 недель, по меньшей мере 28 недель, по меньшей мере 48 недель или по меньшей мере 52 недель до начала второй терапии.
45. Способ в соответствии с любым одним из параграфов 16-44, где периодическое введение антагониста SFRP2, CD38 и/или PD-1 и/или антагониста PD-1 продолжается в течение по меньшей мере 3 дней, по меньшей мере 30 дней, по меньшей мере 42 дней, по меньшей мере 8 недель, по меньшей мере 12 недель, по меньшей мере 24 недель или по меньшей мере 6 месяцев.
46. Способ в соответствии с любым одним из параграфов 16-45, где каждое из количества антагониста SFRP2, CD38 и/или PD-1 при приеме в отдельности и количество антагониста PD-1 при приеме в отдельности эффективно для лечения субъекта.
47. Способ в соответствии с любым одним из параграфов 16-45, где либо количество антагониста SFRP2, CD38 и/или PD-1 при приеме в отдельности, либо количество антагониста PD-1 при приеме в отдельности, либо каждое такое количество при приеме в отдельности не является эффективным для лечения субъекта.
48. Способ в соответствии с любым одним из параграфов 16-45, где либо количество антагониста SFRP2, CD38 и/или PD-1 при приеме в отдельности, либо количество антагониста PD-1 при приеме в отдельности, либо каждое такое количество при приеме в отдельности менее эффективно для лечения субъекта.
49. Способ в соответствии с любым одним из параграфов 16-48, где субъект представляет собой пациента-человека.
50. Способ в соответствии с любым одним из параграфов 16-49, где пациент ранее получал терапию антагонистом PD-1 и прекратил получать терапию антагонистом PD-1 до получения комбинированной терапии.
51. Способ в соответствии с любым одним из параграфов 16-50, где пациент ранее не отвечал на терапию антагонистом PD-1 или введение монотерапии антагониста PD-1 не лечило субъекта.
52. Набор для лечения пациента, страдающего раком, содержащий терапевтически эффективное количество антагониста SFRP2, CD38 и/или PD-1, терапевтически эффективное количество антагониста PD-1 и вкладыш, содержащий инструкции по применению набора.
53. Фармацевтическая композиция, содержащая количество антагониста PD-1 и количество антагониста SFRP2, CD38 и/или PD-1.
54. Фармацевтическая композиция в соответствии с параграфом 53, содержащая по существу количество антагониста PD-1 и количество антагониста SFRP2, CD38 и/или PD-1.
55. Фармацевтическая композиция в соответствии с параграфом 53 или 54 для применения в лечении субъекта, страдающего раком, в которой количество антагониста PD-1 и количество антагониста SFRP2, CD38 и/или PD-1 предназначены для введения одновременно, в одно время или сопутствующим образом.
56. Терапевтическая упаковка для дозирования субъекту, страдающему раком, или для применения при дозировании субъекту, страдающему раком, которая содержит: а) одну или более разовых доз, каждая из которых содержит: i) количество антагониста PD-1 и ii) количество антагониста SFRP2, CD38 и/или PD-1, где соответствующие количества указанного антагониста PD-1 и указанного антагониста SFRP2, CD38 и/или PD-1 в указанной разовое дозе эффективны при сопутствущем введении указанному субъекту для лечения субъекта, и b) готовый фармацевтический контейнер, где указанный контейнер включает указанную разовую дозу или разовые дозы, где указанный контейнер дополнительно включает или содержит маркировку, направленную на применение указанной упаковки при лечении указанного субъекта.
57. Антагонист SFRP2, CD38 и/или PD-1 для применения в качестве дополнительной терапии или в комбинации с антагонистом PD-1 в лечении субъекта, страдающего раком.
58. Антагонист PD-1 для применения в качестве дополнительной терапии или в комбинации с SFRP2, CD38 и/или антагонистом PD-1 в лечении субъекта, страдающего раком.
59. Применение количества антагониста SFRP2, CD38 и/или PD-1 и количества антагониста PD-1 в получении комбинации для лечения субъекта, страдающего раком, где антагонист SFRP2, CD38 и/или PD-1 и антагонист PD-1 получены для введения одновременно, в одно время или сопутствующим образом.
60. Комбинация антагониста SFRP2, CD38 и/или PD-1 и антагониста PD-1 для применения в получении лекарственного средства.
61. Комбинация в соответствии с параграфом 60, где лекарственное средство предназначено для лечения, предотвращения или облегчения симптома рака.
62. Способ лечения рака, включающий введение терапевтически эффективного количества моноклонального антитела (mAb) SFRP2 субъекту, нуждающемуся в этом, где субъект имеет повышенную экспрессию CD38 и/или PD-1.
63. Способ по параграфу 62, где Т-клетки субъекта имеют повышенную экспрессию CD38 и/или PD-1.
64. Способ в соответствии с параграфом 62 или 63, где моноклональное антитело к SFRP2 является человеческим или гуманизированным.
65. Способ в соответствии с любым одним из параграфов 62-64, где указанный рак представляет собой рак молочной железы.
66. Способ в соответствии с любым одним из параграфов 62-64, где указанный рак представляет собой ангиосаркому, рак легкого, остеосаркому, меланому, немелкоклеточный рак легкого или рак почки.
67. Способ в соответствии с любым одним из параграфов 62-66, где моноклональное антитело к SFRP2 вводят ежедневно, чаще одного раза в день или реже одного раза в день.
68. Способ в соответствии с любым одним из параграфов 62-67, где моноклональное антитело к SFRP2 вводят один раз в 3 дня, один раз в неделю, один раз в 2 недели, один раз в 3 недели или один раз в 4 недели.
69. Способ в соответствии с любым одним из параграфов 62-67, где периодическое введение моноклонального антитела к SFRP2 продолжается в течение по меньшей мере 3 дней, по меньшей мере 30 дней, по меньшей мере 42 дней, по меньшей мере 8 недель, по меньшей мере 12 недель, по меньшей мере 24 недель или по меньшей мере 6 месяцев.
70. Способ в соответствии с любым одним из параграфов 62-69, где субъектом является пациент-человек.
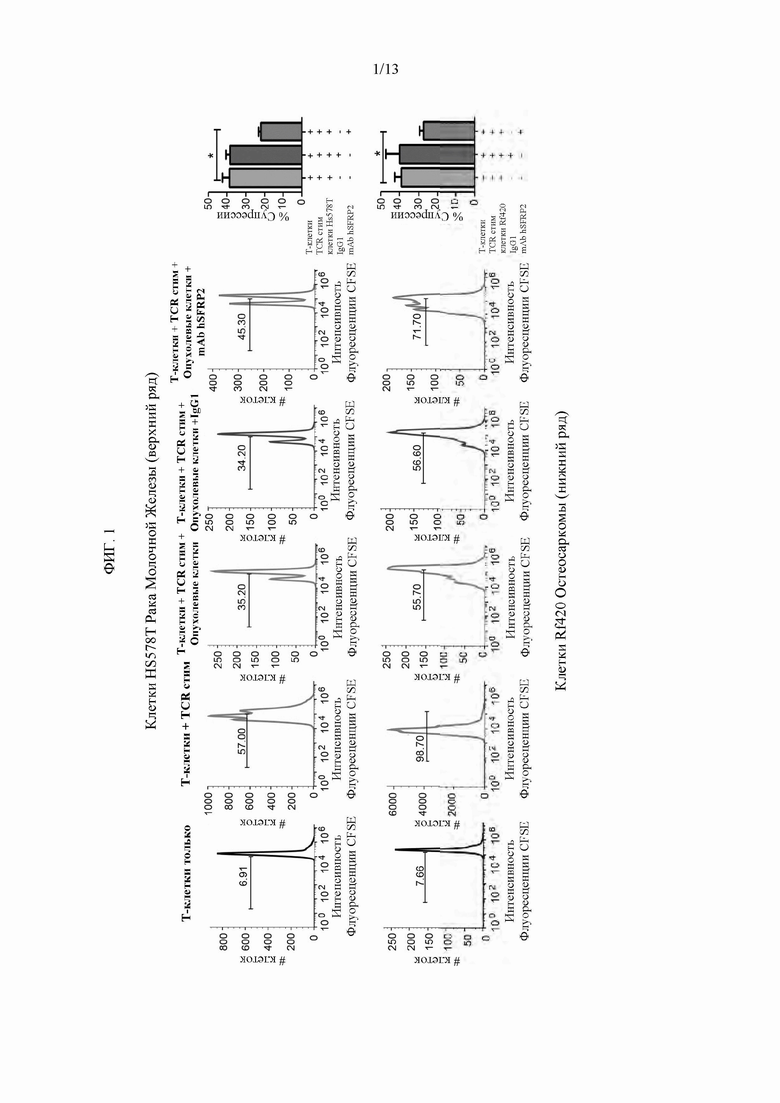
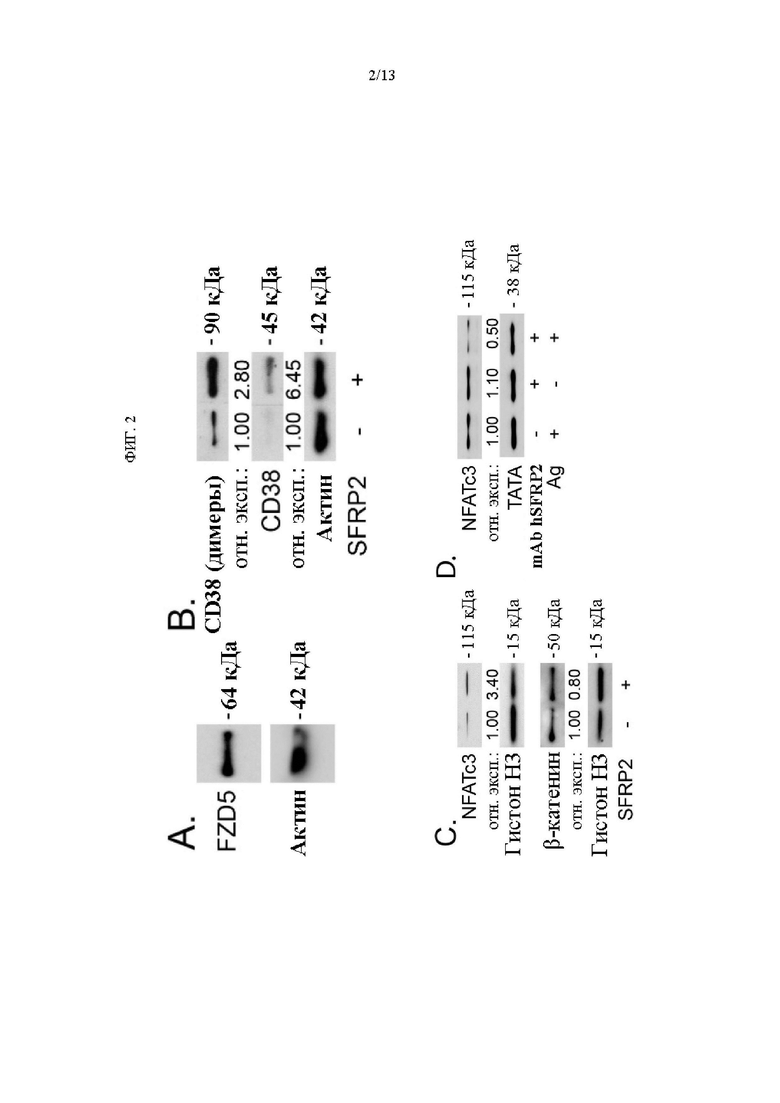
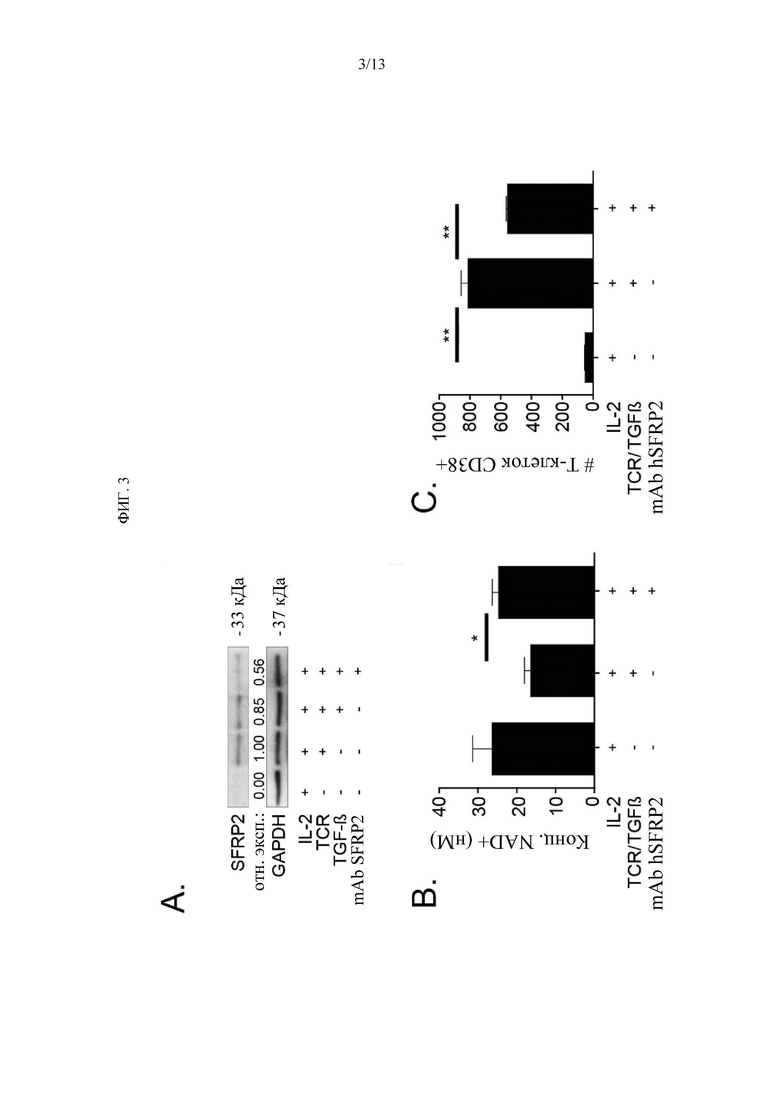
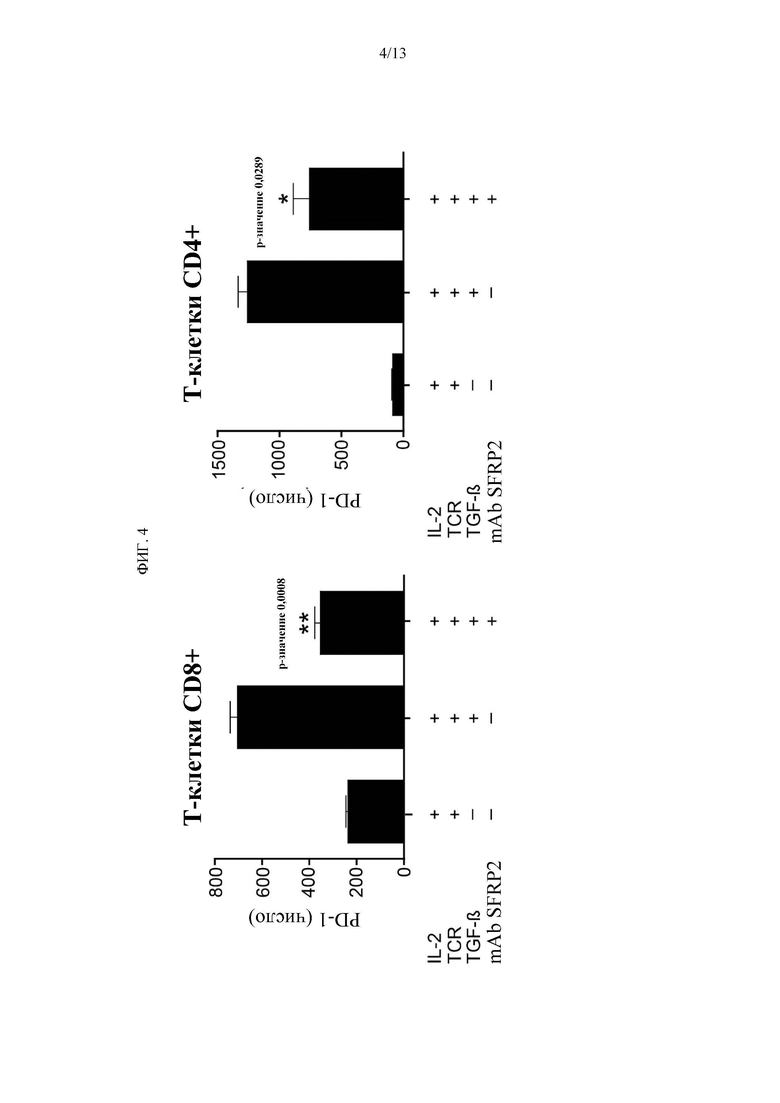
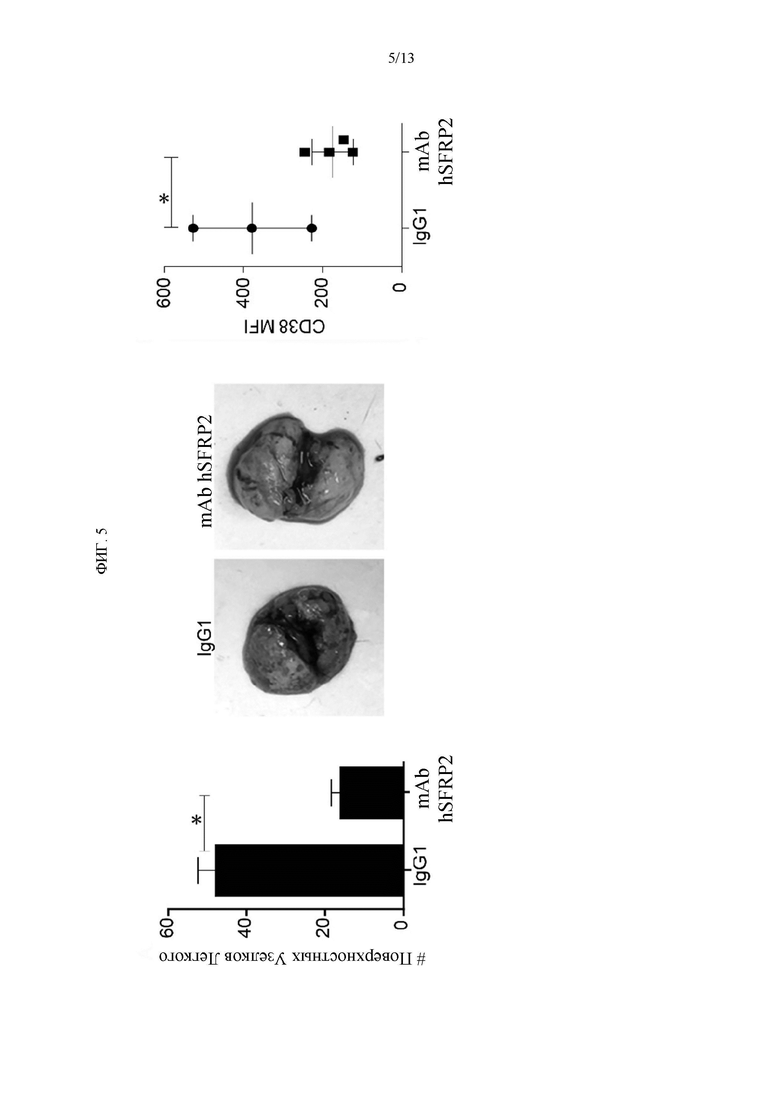
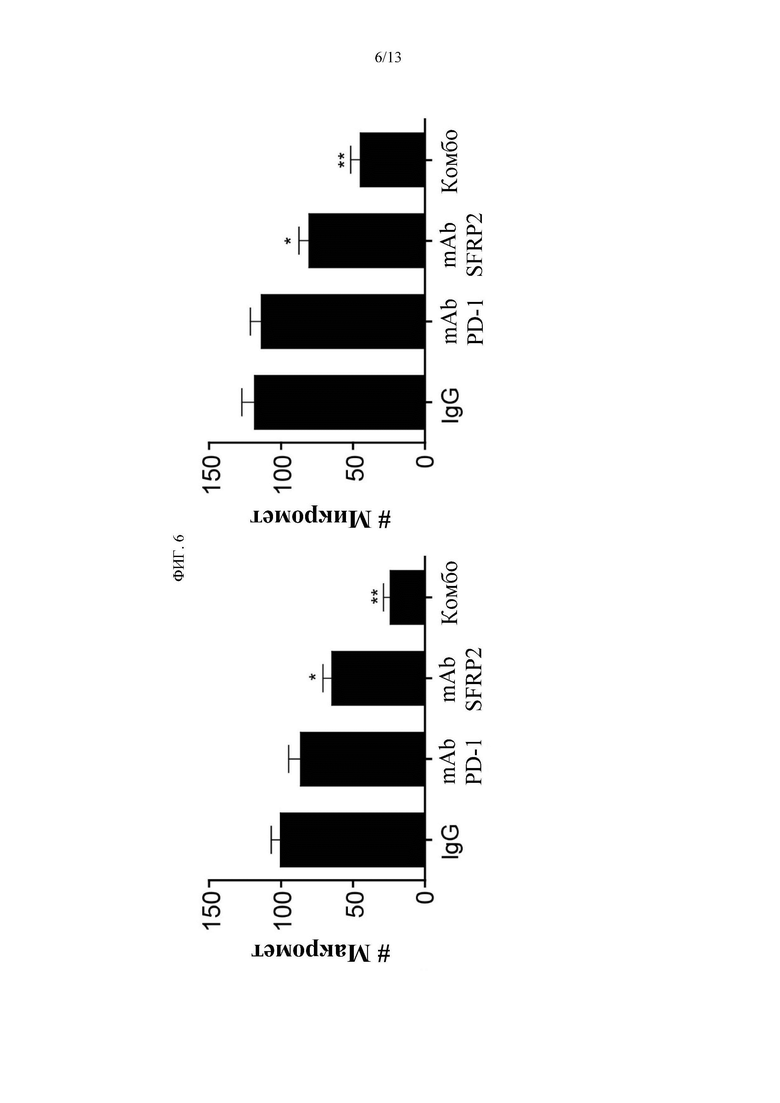
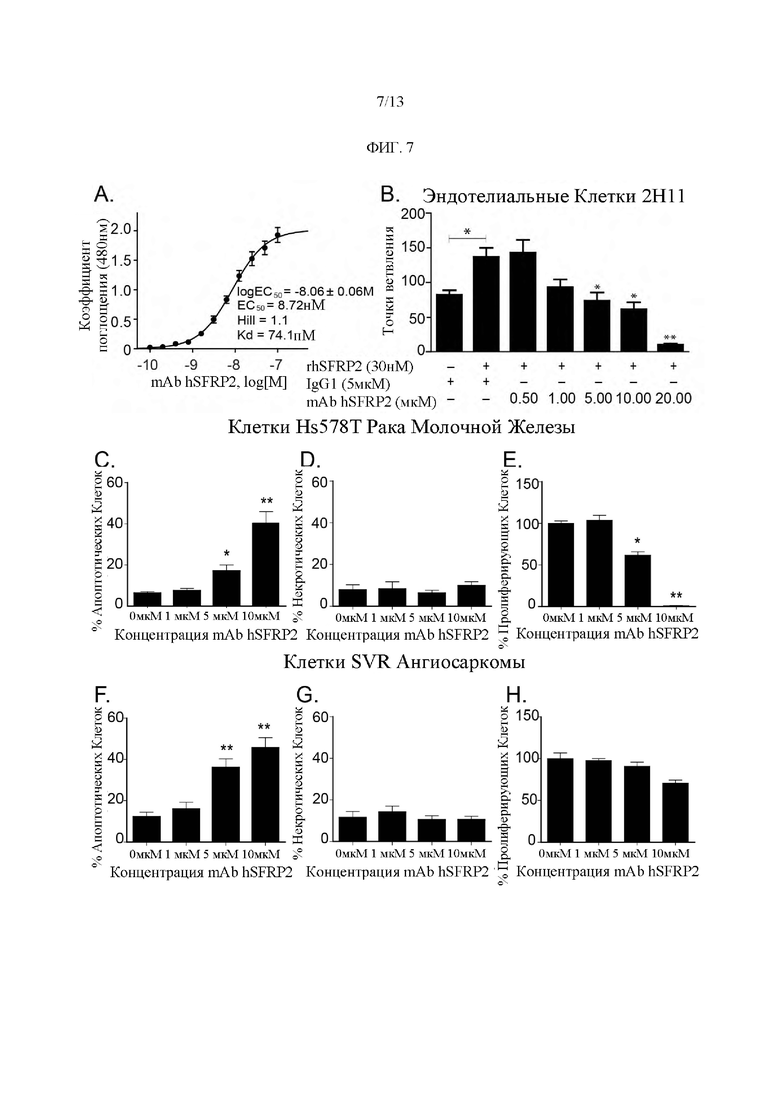
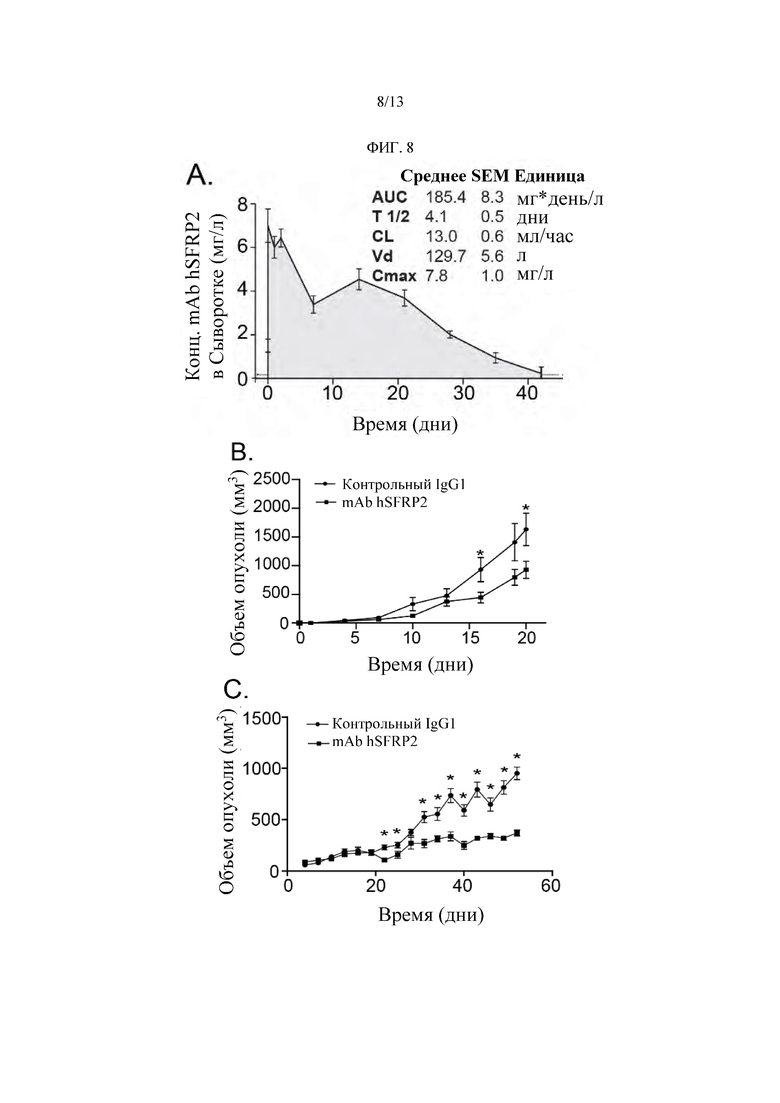
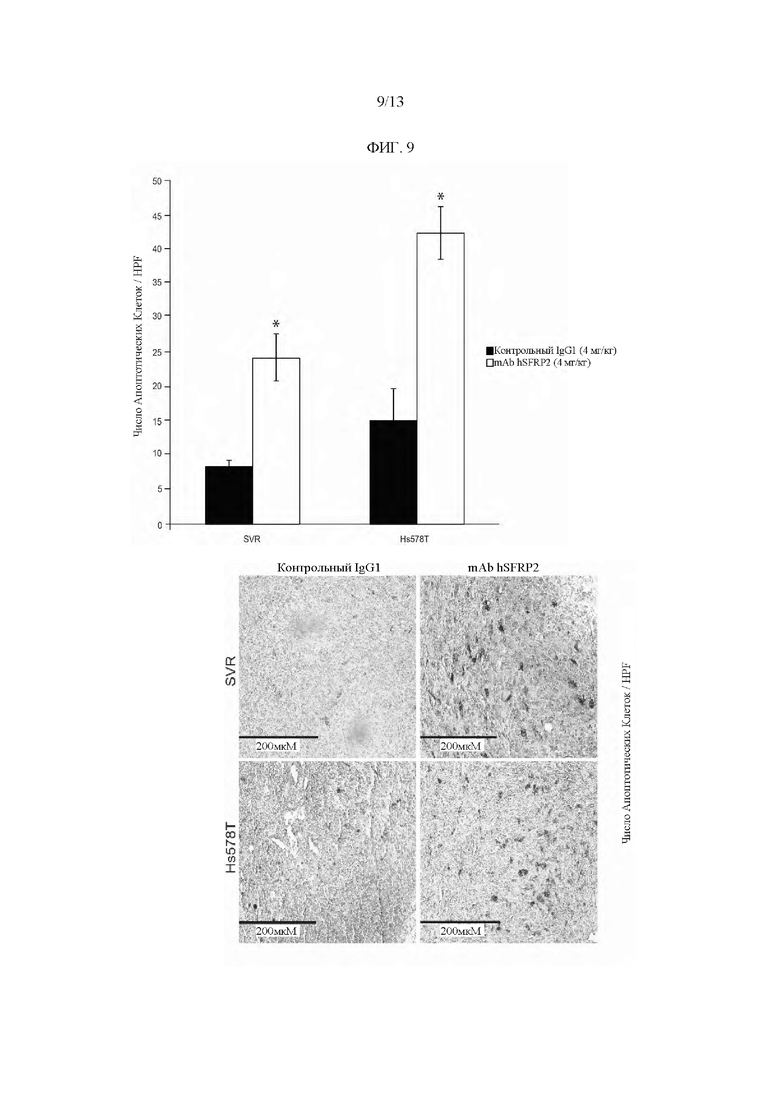
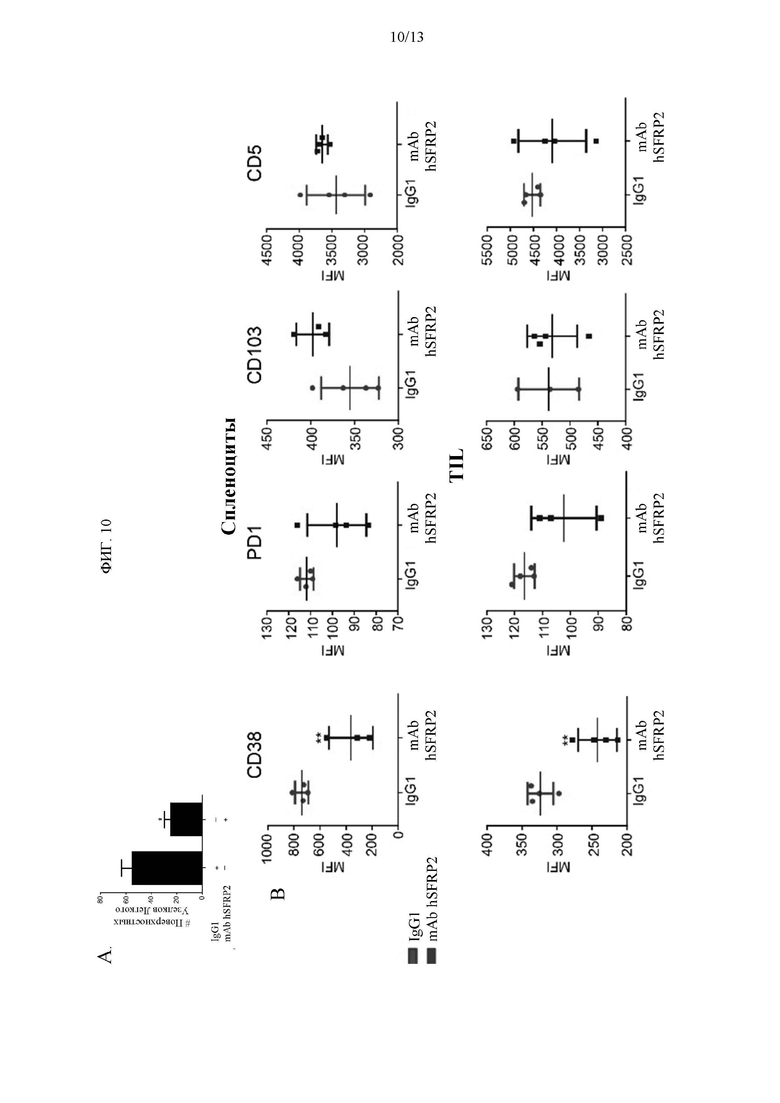
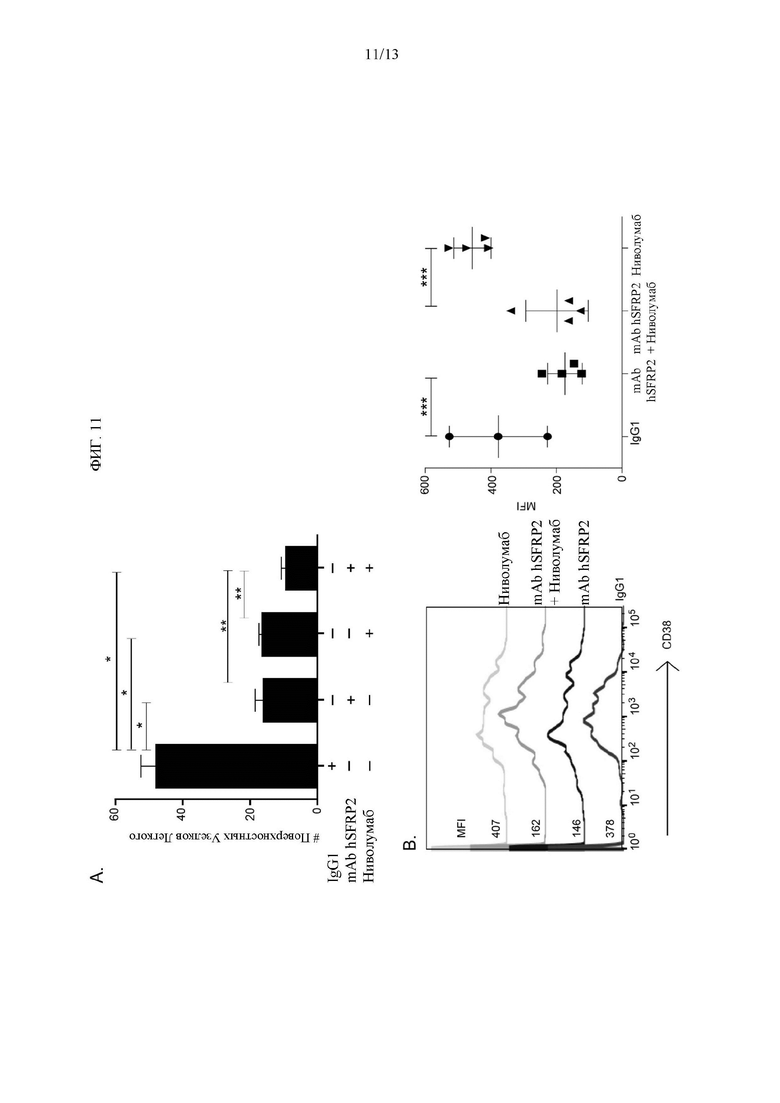
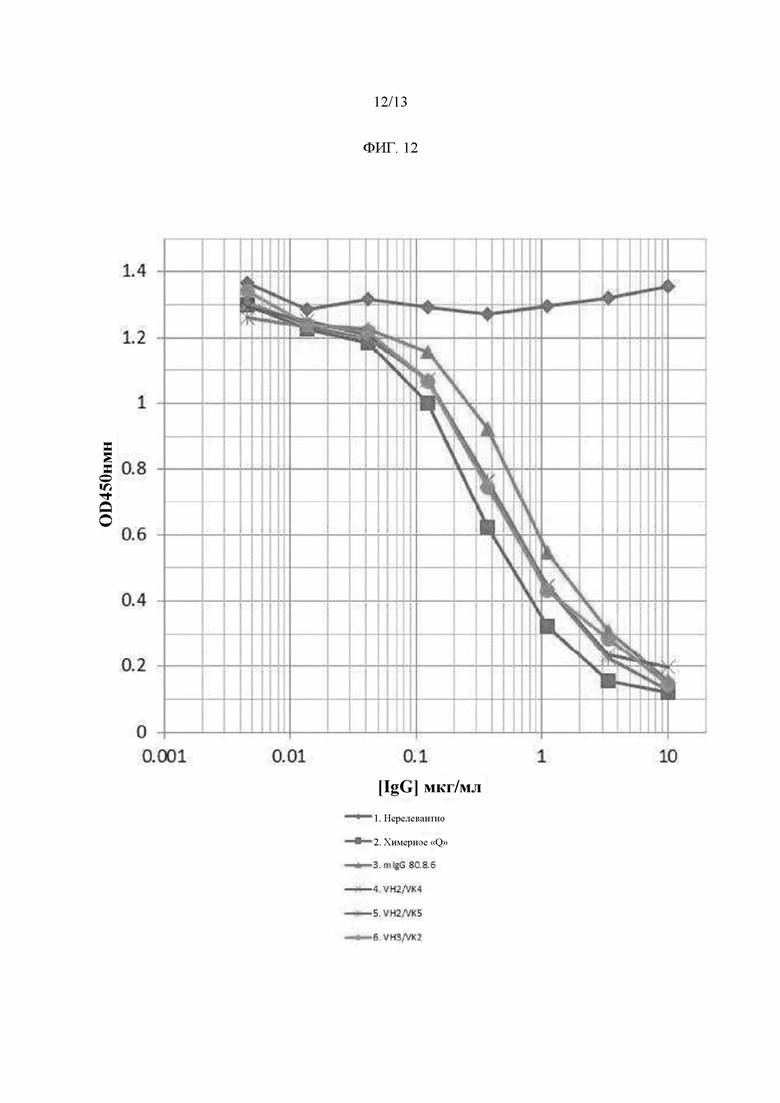
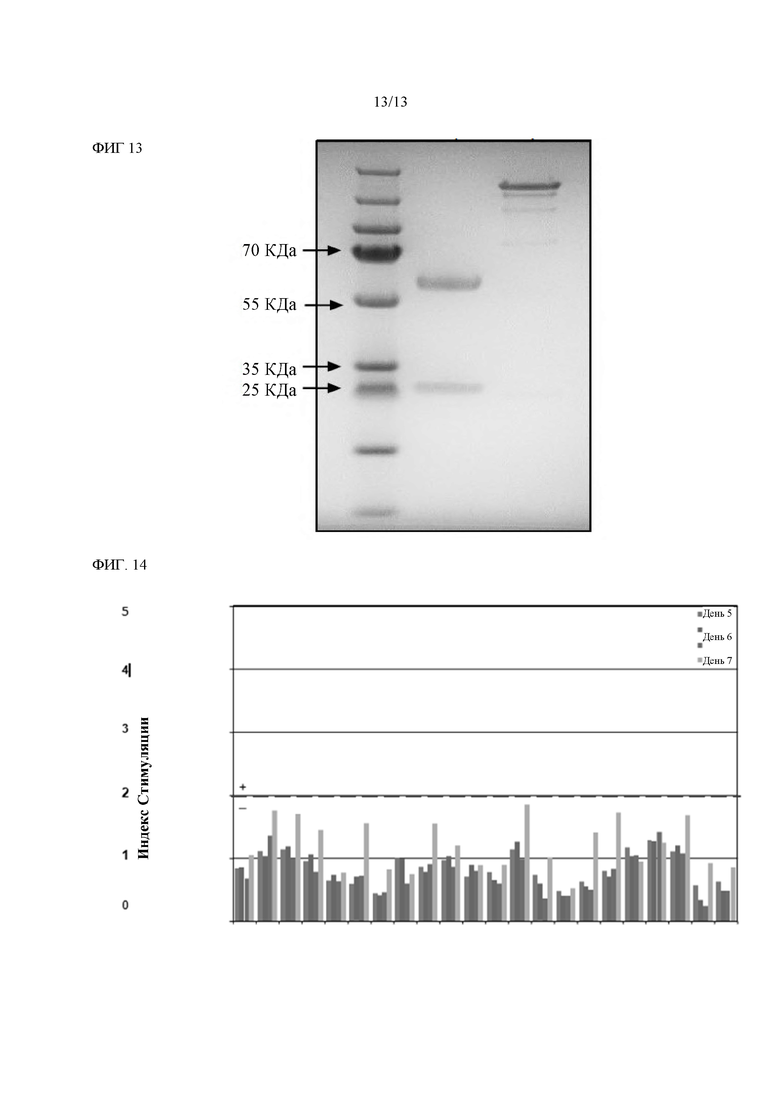
Группа изобретений относится к фармацевтической комбинации, включающей антагонист SFRP2 и антагонист антитела PD-1. Предложены способ, набор, фармацевтическая композиция для лечения остеосаркомы, основанные на применении терапевтически эффективного количества гуманизированного моноклонального антитела к SFRP2 и терапевтически эффективного количества ниволумаба. Гуманизированное моноклональное антитело к SFRP2 содержит V-области мышиного моноклонального антитела 80.8.6, константные области тяжелой цепи IgG1 человека и константные области легкой цепи κ. Изобретения обеспечивают эффективное ингибирование роста метастатической остеосаркомы in vivo. 4 н. и 11 з.п. ф-лы, 14 ил., 3 пр.
1. Способ лечения остеосаркомы, включающий введение терапевтически эффективного количества гуманизированного моноклонального антитела к SFRP2 и терапевтически эффективного количества ниволумаба субъекту, нуждающемуся в этом, причем гуманизированное моноклональное антитело к SFRP2 содержит i) V-области мышиного моноклонального антитела 80.8.6 к SFRP2, ii) константные области тяжелой цепи IgG1 человека и iii) константные области легкой цепи κ.
2. Способ по п. 1, где гуманизированное моноклональное антитело к SFRP2 вводят ежедневно, один раз в 3 дня, один раз в неделю, один раз в 2 недели, один раз в 3 недели или один раз в 4 недели.
3. Способ по п. 1, где ниволумаб вводят ежедневно, один раз в 3 дня, один раз в неделю, один раз в 2 недели, один раз в 3 недели или один раз в 4 недели.
4. Способ по п. 1, где количество ниволумаба, вводимого субъекту, составляет 3 мг/кг массы тела каждые 3 недели, 240 мг каждые 2 недели или 480 мг каждые 4 недели.
5. Способ по п. 1, где субъект получает терапию ниволумабом до начала терапии гуманизированным моноклональным антителом к SFRP2.
6. Способ по п. 5, где субъект получает терапию ниволумабом в течение по меньшей мере 8 недель, по меньшей мере 10 недель, по меньшей мере 24 недель, по меньшей мере 28 недель, по меньшей мере 48 недель или по меньшей мере 52 недель до начала терапии гуманизированным моноклональным антителом к SFRP2.
7. Способ по п. 1, где субъект получает терапию гуманизированным моноклональным антителом к SFRP2 до начала терапии ниволумабом.
8. Способ по п. 7, где субъект получает терапию гуманизированным моноклональным антителом к SFRP2 в течение по меньшей мере 8 недель, по меньшей мере 10 недель, по меньшей мере 24 недель, по меньшей мере 28 недель, по меньшей мере 48 недель или по меньшей мере 52 недель до начала терапии ниволумабом.
9. Способ по п. 1, где введение гуманизированного моноклонального антитела к SFRP2 и/или ниволумаба продолжается в течение по меньшей мере 3 дней, по меньшей мере 30 дней, по меньшей мере 42 дней, по меньшей мере 8 недель, по меньшей мере 12 недель, по меньшей мере 24 недель или по меньшей мере 6 месяцев.
10. Набор для лечения пациента, страдающего остеосаркомой, содержащий терапевтически эффективное количество гуманизированного моноклонального антитела к SFRP2, терапевтически эффективное количество ниволумаба и вкладыш, содержащий инструкции по применению набора, причем гуманизированное моноклональное антитело к SFRP2 содержит i) V-области мышиного моноклонального антитела 80.8.6 к SFRP2, ii) константные области тяжелой цепи IgG1 человека и iii) константные области легкой цепи κ.
11. Фармацевтическая композиция для лечения остеосаркомы, содержащая терапевтически эффективное количество гуманизированного моноклонального антитела к SFRP2 и терапевтически эффективное количество ниволумаба, причем гуманизированное моноклональное антитело к SFRP2 содержит i) V-области мышиного моноклонального антитела 80.8.6 к SFRP2, ii) константные области тяжелой цепи IgG1 человека и iii) константные области легкой цепи κ.
12. Фармацевтическая композиция по п. 11, где терапевтически эффективное количество гуманизированного моноклонального антитела к SFRP2 составляет от 0,1 до 100 мг/кг массы тела.
13. Фармацевтическая композиция по п. 11, где терапевтически эффективное количество гуманизированного моноклонального антитела к SFRP2 составляет 10-200 мг.
14. Фармацевтическая композиция по п. 11, где терапевтически эффективное количество ниволумаба составляет от 0,1 до 100 мг/кг массы тела.
15. Применение гуманизированного моноклонального антитела SFRP2 для лечения остеосаркомы в комбинации с ниволумабом, причем гуманизированное моноклональное антитело к SFRP2 содержит i) V-области мышиного моноклонального антитела 80.8.6 к SFRP2, ii) константные области тяжелой цепи IgG1 человека и iii) константные области легкой цепи κ.
| Токарный резец | 1924 |
|
SU2016A1 |
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| FONTENOT E | |||
| ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2013 |
|
RU2599417C2 |

