Изобретение относится к области микробиологии и паразитологии микроорганизмов и может быть использовано для определения простейших Blastocysts spp. с различной степенью вирулентности у пациентов с нарушениями качественного и/или количественного состава микробиоценоза кишечника, инвазированных бластоцистами.
В последние годы в России отмечается чрезвычайно напряженная эпидемиологическая обстановка по паразитозам. В стране ежегодно официально регистрируется более 1,3 миллионов больных паразитарными заболеваниями, среди которых отмечается значительный рост заболеваемости кишечными протозоозами. В связи с этим, особую актуальность приобретает широко распространенная «новая» протозойная инвазия - бластоцистоз, обусловленная паразитированием преимущественно в толстой кишке простейших Blastocysts spp.
Данный возбудитель длительное время не привлекал внимание специалистов как энтеропатоген, однако в настоящее время имеется достаточное количество работ, подтверждающих роль Blastocysts spp. в патологии человека. Мишенью данных простейших является желудочно-кишечный тракт человека.
С целью аргументации этиологической значимости бластоцист в развитии патологического процесса, как и всех условно-патогенных микроорганизмов, необходимо подтверждение их вирулентности. Несмотря на то, что до сих пор отсутствуют экспериментальные модели, позволяющие воспроизвести полный цикл паразита, существует большое количество экспериментальных и клинико-биологических работ, подтверждающих значение Blastocysts spp. как патогена.
Патогенность Blastocysts spp. была убедительно доказана в опытах на безмикробных морских свинках, которых заражали per os (Мое К.Т. et al. Experimental Blastocysts hominis infection in laboratory mice // Parasitol. Res. -1998. - V.83. - P.319-325.) и непосредственным введением паразита в слепую кишку (Markell Е.К. et al. В.hominis Pathogen or fellow traveller // J.Trop. Med. Hyg. - 1986. - V.35. - P. 1023-1026). У всех животных развивалась не только колонизация, но и инвазия кишечника бластоцистами, сопровождающаяся диареей. В результате все животные теряли в весе и проявляли повышенную сонливость. Гистологическое исследование толстого кишечника выявило интенсивное воспаление слизистой оболочки с явлениями некроза.
Общепринятым методом определения вирулентности микроорганизмов является тест внутрибрюшинного заражения мышей. Однако этот способ является длительным и малоэффективным, так как не дает полного представления о наличии генетических маркеров вирулентности в геноме микроорганизмов.
Задачей изобретения является создание метода определения простейших Blastocysts spp. с различной степенью вирулентности, обеспечивающего получение технического результата, состоящего в повышении эффективности определения вирулентности простейших за счет проведения молекулярно-генетического анализа расщепления хромосомной ДНК бластоцист эндонуклеазами рестрикции.
Предлагаемый способ определения вирулентности простейших бластоцист методом полиморфизма длин фрагментов рестрикции с использованием определенного набора эндонуклеаз не имеет идентичных аналогов при решении поставленной задачи.
Используемый метод ПДФР широко применяется в настоящее время в генетических исследованиях популяций, поскольку наличие в геноме исследуемого организма рестрикционного фрагмента ДНК определенной длины является прекрасным генетическим маркером и одновременно фенотипическим признаком, тесно связанным с генотипом организма. Однако большинство исследований направлены, например, на изучение распространения такого маркера в популяциях, передачей его от родителей к потомству при скрещиваниях и направлены в дальнейшем для построения генетических карт исследуемых организмов, поскольку ПДФР-маркеры благодаря их четкой принадлежности определенным генетическим локусам не уступают по информативности распространенным биохимическим маркерам и во многих случаях оказываются удобнее сложных стенотипических признаков (таких, как цвет глаз, окраска волос или шерсти, форма цветков и листьев), определяемых многими генными локусами. Это широко представлено в работах Лыоина Б., 1987, Lindblom В. et al., 1988, Roberts R. et al., 2005.
Также этот довольно простой метод в последнее время применяется с целью изучения геномов млекопитающих для поиска сайтов узнавания эндонуклеаз рестрикции, позволяющий производить расчет длин и количества образуемых при расщеплении фрагментов ДНК с последующим построением диаграмм их распределения (Абдурашитов М.А. с соавт., 2006).
Однако до настоящего времени нет работ, позволяющих выявить генетические маркеры патогенных свойств бластоцист и на основе рестрикционного анализа ДНК определить зависимость между полиморфизмом длин фрагментов нуклеотидных последовательностей и вирулентностью простейших.
Указанный технический результат в методе определения Blastocystis spp. с различной степенью вирулентности достигается тем, что применяется молекулярно-генетический метод рестрикционного анализа бактериальной ДНК или анализ полиморфизма длин фрагментов рестрикции (ПДФР) ДНК.
Для фрагментирования ДНК бластоцист использовали рестриктазы (ферменты, расщепляющие ДНК) или рестрикционные эндонуклеазы, выделенные из бактериальных клеток. Эти ферменты in vivo участвуют в узнавании и разрушении чужеродных для микроорганизмов ДНК, расщепляя внутренние участки молекулы на сравнительно небольшие фрагменты. Рестриктазы узнают специфические последовательности из 4-6, реже 8-12 нуклеотидов в двухцепочечной молекуле ДНК (сайты рестрикции) и "разрезают" ее в местах локализации этих последовательностей. Количество образующихся рестрикционных фрагментов ДНК при использовании одной рестриктазы зависит от количества сайтов рестрикции, а размер фрагментов определяется положением этих сайтов по всей длине исходной молекулы ДНК.
В работе были использованы рестриктирующие эндонуклеазы (рестриктазы): EcoR 1, BamH 1, Нае III, Hind III, Pst I, производство НПО «СибЭнзим», г.Новосибирск (infos@ibenzyme.ru), в соответствующем буфере.
Сущность изобретения поясняется рисунками, на которых изображены:
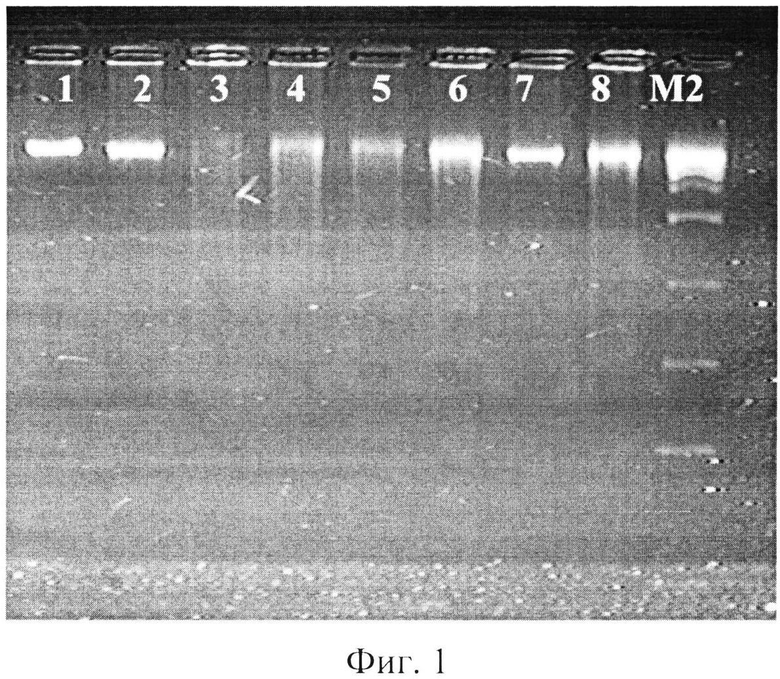
фиг.1 - электрофореграмма рестрикционных фрагментов ДНК штаммов бластоцист с использованием рестриктазы BamH I;
где 1, 2 - авирулентные штаммы бластоцист,
3, 4 - слабовирулентные штаммы бластоцист,
5, 6- умеренновирулентные штаммы бластоцист,
7, 8 - высоковирулентные штаммы бластоцист,
М2 - маркер молекулярного веса - 300-10000 п.н.
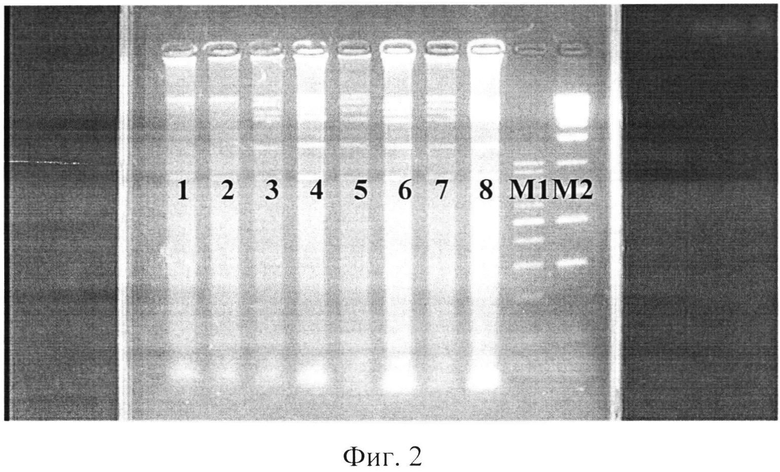
фиг.2 - электрофореграмма рестрикционных фрагментов ДНК штаммов бластоцист с использованием рестриктазы Нае III;
где 1, 2 - авирулентные штаммы бластоцист,
3, 4 - слабовирулентные штаммы бластоцист,
5, 6 - умеренновирулентные штаммы бластоцист,
7, 8 - высоковирулентные штаммы бластоцист,
Ml - маркер молекулярного веса - 100-1000 п.н.,
М2 - маркер молекулярного веса - 300-10000 п.н.
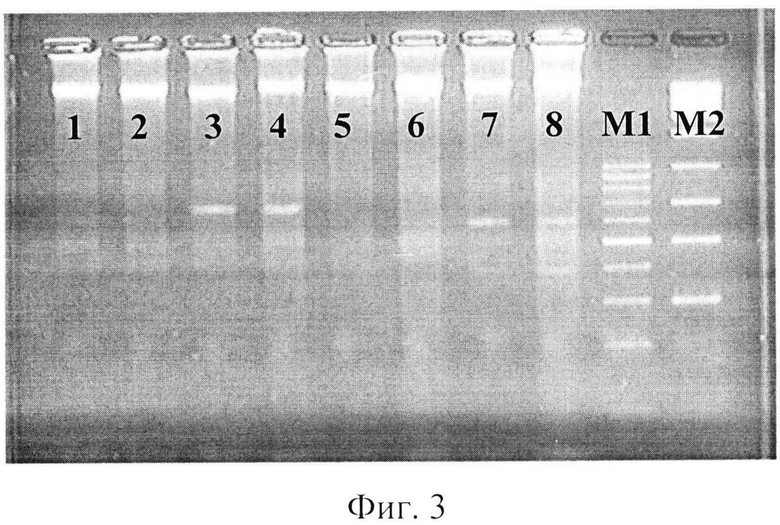
фиг.3 - электрофореграмма рестрикционных фрагментов ДНК штаммов бластоцист с использованием рестриктазы Hind III;
где 1,2 - авирулептные штаммы бластоцист,
3, 4 - слабовирулентные штаммы бластоцист,
5, 6 - умеренновирулентные штаммы бластоцист,
7, 8 - высоковирулентные штаммы бластоцист,
Ml - маркер молекулярного веса - 100-1000 п.н.,
М2 - маркер молекулярного веса - 300-10000 п.н.
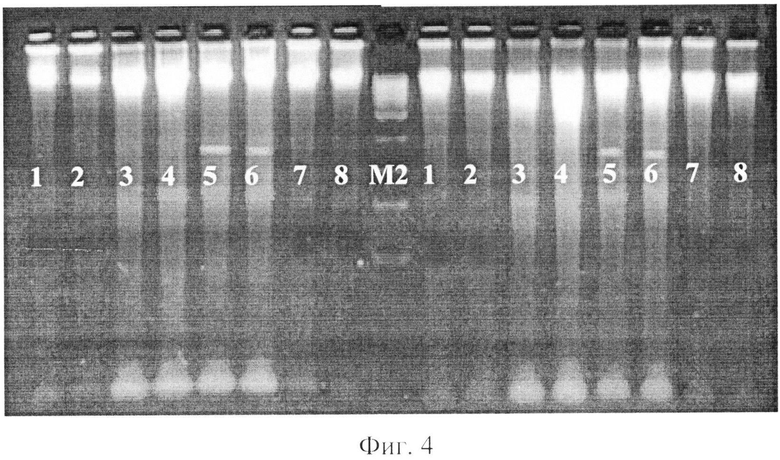
фиг.4 - электрофореграмма рестрикционных фрагментов ДНК штаммов бластоцист с использованием рестриктазы Pst I;
где 1, 2 - авирулентные штаммы бластоцист,
3, 4 - слабовирулентные штаммы бластоцист,
5, 6 - умеренновирулентные штаммы бластоцист,
7, 8 - высоковирулентные штаммы бластоцист,
М2 - маркер молекулярного веса - 300-10000 п.н.
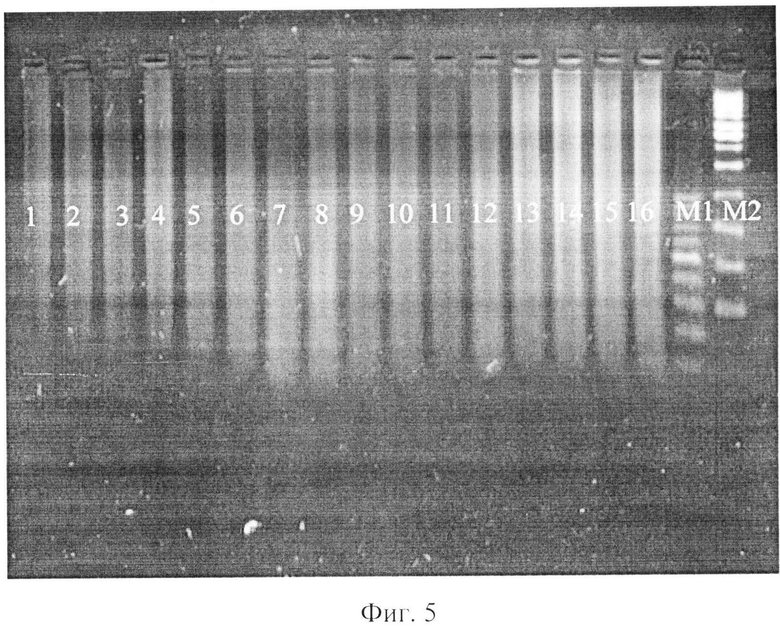
фиг.5 - электрофореграмма рестрикционных фрагментов ДНК штаммов бластоцист с использованием рестриктазы EcoR I;
где 1-4 - авирулентные штаммы бластоцист,
5-8 - слабовирулентные штаммы бластоцист,
9-12 - умеренновирулентные штаммы бластоцист,
13-16 - высоковирулентные штаммы бластоцист,
Ml - маркер молекулярного веса - 100-1000 п.н.,
М2 - маркер молекулярного веса - 300-10000 п.н.
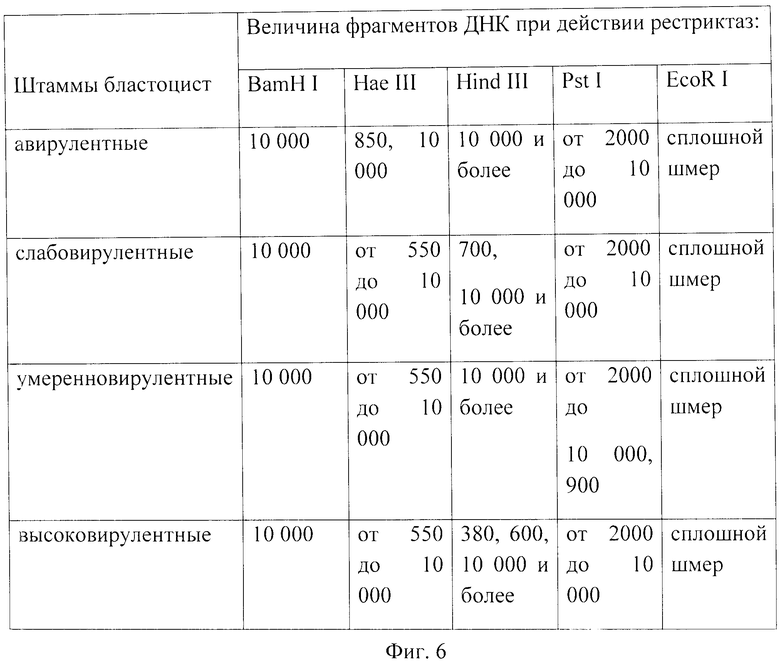
фиг.6 - величина фрагментов ДНК бластоцист с различной степенью проявления вирулентности (п.н.).
Метод осуществляют поэтапно следующим образом.
1. Выделение ДНК бластоцист
Выделение ДНК простейших проводили с использованием фенольно-хлороформной методики выделения согласно инструкции по применению комплекта реагентов для выделения ДНК, утвержденным приказом Росздравнадзора от 30 июня 2008 №5008-Пр/08, регистрационное удостоверение МЗ CP РФ №ФСР 2008/02938.
Фенольно-хлороформная экстракция для выделения ДНК бластоцист осуществляли с применением наборов «ВекторДНК-экстракция», ЗАО «Вектор-Бест» (пос. Кольцево, Новосибирской области).
В эксперименте использовали чистые культуры Blastocysts spp., выращенные на среде Surech. Для установления оптимального срока обработки исследуемого материала было проведено выделение ДНК через 3 суток после хранения культур при t=-20°С.
Этапы выделения ДНК:
1) полученные культуры бластоцист, объемом до 2 мл, отливали в пробирки типа Эппендорф;
2) центрифугировали пробирки при 13000 об/мин в течение 15 минут при комнатной температуре;
3) не задевая осадок, полностью удаляли надосадочную жидкость из каждой пробирки отдельным наконечником;
4) проводили электрофорез в 1% агарозовом геле. Это стандартный метод использовался для разделения, идентификации и очистки фрагментов ДНК. Окрашивание проводили флуоресцирующим и интеркалирующим в ДНК красителем - бромистым этидием в низкой концентрации, связывающегося с фрагментами молекулы и дающего специфическое розовое окрашивание в ультрафиолетовой области спектра;
5) в агарозовый гель помещали выделенные фрагменты ДНК и заливали буферным раствором ТВЕ (трис-борат) при концентрации 50 мМ, рН 7,5-7,8 и разгоняли в течение 3 часов;
6) гели просматривали с помощью трансиллюминатора.
II. Проведение расщепления ДНК бластоцист рестриктазами
Для каждого фермента рестрикции существуют оптимальные условия реакции, которые приводились в описании, прилагаемой фирмой-изготовителем. Основные переменные параметры - это температура инкубации и состав буфера.
Реакцию останавливали добавлением 5 мкл стоп-раствора, содержащего 0,1 М ЭДТА, 0,05% бромфенолового синего и 40% сахарозы. Электрофоретическое разделение продуктов рестрикции амплифицированной ДНК проводили в 2% агарозе (Sigma) в трис-ацетатном буфере с этидий-ор-бромидом (0,5 мг/л) при 120 V в течение 3 ч. Идентификация нужных фрагментов ДНК в таком геле возможна только путем гибридизации с мечеными ДНК-зондами. Синтез ДНК-зондов осуществлялся в автоматизированных машинах, позволяющих синтезировать фрагменты однонитевой ДНК длиной свыше 100 нуклеотидных звеньев со строго определенной первичной структурой. Такие молекулы можно использовать для специфического связывания с исследуемыми участками гена.
Для определения длины фрагментов ДНК использовали маркеры молекулярного веса ДНК (Ml- маркер молекулярного веса - 100-1000 п.н., М2-маркер молекулярного веса - 300-10000 п.н., НПО "СибЭнзим"). Определение длин полученных рестриктов осуществляли с помощью компьютерной программы Gel Pro Analyzer, версия 4.0.00.001. Процент идентичности длин фрагментов рассчитывали для каждой пары микроорганизмов, сравнивая картины рестрикции отдельно по каждой рестриктазе. При сравнении длин рестриктов идентичными считали фрагменты ДНК, длина которых различалась не более чем на 5%.
Далее был проведен ряд экспериментов с использованием рестриктаз на экстрагированной ДНК авирулентных, слабовирулентных, умеренновирулентных и высоковирулентных штаммах бластоцист, выделенных из клинического материала больных.
При использовании рестриктазы BamH 1 - G▼GATCC/CCTAG▲G. на дорожках 1% агарозного геля наблюдались полосы окрашенной бромистым этидием ДНК размером около 10000 п.н. Коротких фрагментов хромосомной ДНК бластоцист размером 100-10000 п.н. в ходе эксперимента обнаружено не было (фиг.1).
При проведении реакции с экстрагированной тотальной ДНК бластоцист с различной степенью выраженности вирулентности были получены следующие результаты: на дорожках агарозного геля после электрофореза у авирулентных бластоцист ясно выражены фрагменты ДНК размером около 850 п.н., в остальных дорожках ДНК бластоцист с различной степенью выраженности вирулентности, кроме этих фрагментов, наблюдались полосы с размерами от 1500 до 10000 п.н. (фиг.2).
Следовательно, рестриктаза Нае III позволяет дифференцировать авирулентные и вирулентные штаммы бластоцист без выявления выраженности вирулентных свойств.
При использовании рестриктазы Hind III - A▼AGCTT/TTCGA▼А на дорожках 1% агарозного геля наблюдались полосы размером от 10000 и более п.н., окрашенные бромистым этидием. Кроме этого, на дорожках ДНК слабовирулентных штаммов бластоцист наблюдалась полоса размером 700 п.н.; на дорожках агарозы ДНК высоковирулентных штаммов бластоцист выявлялись две полосы 380 и 600 п.н. Следовательно, рестриктаза Hind III позволяет выявить слабо- и высоковирулентных простейших (фиг.3).
При проведении экспериментов рестрикцирования (расщепления с помощью эндонуклеаз) выделенной ДНК штаммов бластоцист рестриктазы Pst I - CTGCA▼G/G▲ACGTC были получены следующие результаты: на дорожках агарозного геля штаммов бластоцист с различной степенью выраженности вирулентности наблюдались яркие размытые полосы (шмеры) размером от 2000 до 10000 (на отдельных дорожках и более) п.н. (фиг.4).
Однако на дорожках хромосомной ДНК умеренновирулентных бластоцист наблюдали фрагменты размером около 900 п.н. У других штаммов бластоцист таких полос не обнаруживалось. Таким образом, рестриктаза Pst I позволила выявить только умеренновирулентные штаммы Blastocystis spp.
При использовании рестриктазы EcoR I с сайтом узнавания G G▼AATTC/CTTAA▲G на дорожках 1% агарозного геля изолятов бластоцист с различной степнью выраженности вирулентности наблюдали полосы шмер по всей длине дорожек, свидетельствующих о множественных фрагментах ДНК различной длины, перекрывающих друг друга (фиг.5).
Таким образом, можно сделать заключение о наличии в структуре ДНК всех штаммов бластоцист множественных сайтов GAATTC/CTTAAG, что свидетельствует о невозможности выявления степени вирулентности бластоцист при помощи рестриктазы EcoR I.
На фиг.6 представлены результаты определения вирулентности штаммов бластоцист, полученных методом рестрикционного анализа ДНК простейших с использованием рестриктирующих эндонуклеаз (рестриктаз): EcoR I, BamH I, Нае III, Hind III, Pst I.
Таким образом, в работе предложен метод определения вирулентности Blastocystis spp., основанный на анализе полиморфизма длин фрагментов рестрикции (ПДФР) ДНК бластоцист. В эксперименте представлены результаты анализа ПДФР ПЦР-продукта длинной 10000 пар нуклеотидов, полученного при выделении ДНК простейших с применением пяти эндонуклеаз рестрикции EcoR I, BamH I, Нае III, Hind III, Pst 1.
Проведенный рестрикционный анализ показал, что использование рестриктаз BamH I, EcoR I не позволяет типировать бластоцисты с различной степенью выраженности вирулентности.
Применение рестриктазы Нае III позволяет проводить дифференциацию авирулентных штаммов бластоцист на основе наличия фрагментов размером 850 п.н. и вирулентных на основе наличия фрагментов ДНК размером от 550 до 10000 п.н.
При помощи рестриктазы Pst I произведено типирование бластоцист с умеренно выраженной вирулентностью на основе наличия фрагментов ДНК размером около 900 п.н.
В эксперименте с применением рестриктазы Hind III было произведено типирование бластоцист со слабовыраженной вирулентностью на основе фрагментов ДНК, размером 700 п.н., а также высоковирулентных штаммов бластоцист на основе наличия фрагментов ДНК, размеров 380 и 600 п.н.
Таким образом, разработанный способ определения Blastocystis spp. с различной степенью вирулентности позволил достичь значительно высоких результатов при определении генетических маркеров вирулентности в геноме бластоцист.
Предлагаемый метод обеспечивает:
1. Достаточно простой и универсальный способ идентификации простейших Blastocystis spp. в зависимости от степени выраженности вирулентных свойств на основе предложенных комбинаций эндонуклеаз рестрикции (Нае III, Pst I и Hind III).
2. Повышение эффективности определения вирулентности бластоцист за счет использования метода рестрикционного анализа ДНК простейших, основанного на выявлении не только потенциально патогенных штаммов бластоцист, которые выявляются традиционно биологическим методом исследования, но и позволяет выявить весь спектр генов в геноме бластоцист, обладающих свойствами патогенности.
Изобретение относится к области микробиологии и паразитологии микроорганизмов. Способ предусматривает рестрикционный анализ бактериальной ДНК бластоцист с использованием комбинации эндонуклеаз рестрикции - Нае III, Pst I и Hind III, позволяющих идентифицировать простейших с различной степенью вирулентности. При этом применение рестриктазы Нае III позволяет проводить дифференциацию авирулентных штаммов бластоцист на основе наличия фрагментов размером 850 п.н. и вирулентных на основе наличия фрагментов ДНК размером от 550 до 10000 п.н. При помощи рестриктазы Pst I выявляют бластоцисты с умеренно выраженной вирулентностью на основе наличия фрагментов ДНК размером около 900 п.н. Применение рестриктазы Hind III позволяет типировать бластоцисты со слабовыраженной вирулентностью на основе фрагментов ДНК, размером 700 п.н., а также высоковирулентные штаммы бластоцист на основе наличия фрагментов ДНК, размеров 380 и 600 п.н. Способ позволяет повысить эффективность определения вирулентности простейших. 6 ил.
Способ определения простейших Blastocysts spp. с различной степенью вирулентности, отличающийся тем, что осуществляют рестрикционный анализ бактериальной ДНК бластоцист с использованием комбинации эндонуклеаз рестрикции - Нае III, Pst I и Hind III, позволяющих идентифицировать простейших с различной степенью вирулентности, при этом применение рестриктазы Нае III позволяет проводить дифференциацию авирулентных штаммов бластоцист на основе наличия фрагментов размером 850 п.н. и вирулентных на основе наличия фрагментов ДНК размером от 550 до 10000 п.н., при помощи рестриктазы Pst I возможно определение бластоцист с умеренно выраженной вирулентностью на основе наличия фрагментов ДНК размером около 900 п.н., а применение рестриктазы Hind III позволяет типировать бластоцисты со слабовыраженной вирулентностью на основе фрагментов ДНК размером 700 п.н., а также высоковирулентные штаммы бластоцист на основе наличия фрагментов ДНК размером 380 и 600 п.н.
| INIT I et al, Restriction enzyme digestion analysis of PCR-amplified DNA of Blastocystis hominis isolates, Southeast Asian J Trop Med Public Health, 2007 Nov., abstract | |||
| ЕМЕЛЬЯНЕНКО П.А | |||
| и др., Ветеринарная микробиология, изд | |||
| "Колос", 1982, с.84-85 | |||
| MOE K.T | |||
| et al, Experimental Blastocystis hominis infection in laboratory mice, Parasitol.Res, 1998, v.83, p.319-325 |

