Изобретения относятся к медицине (диагностика вирусных заболеваний человека), генетической инженерии и биотехнологии и представляют собой рекомбинантную плазмидную ДНК pVR6-II3, несущую фрагмент dVR6-II3 для специфического выявления ротавирусов II субгруппы, и штамм E. coli - продуцент зонда.
Ротавирусы, являющиеся этиологическим агентом острого гастроэнтерита, характеризуются вариабельностью генома и, как следствие, существованием как минимум двух серологических подгрупп вируса (c/гр). Субгрупповая дифференциация ротавирусов имеет важное значение для изучения особенностей эпидемиологии ротавирусной инфекции и клинических проявлений гастроэнтерита, вызванного ротавирусами различных серологических подгрупп.
Субгрупповая дифференциация штаммов ротавирусов может быть проведена методом молекулярной гибридизации с использованием зонда - ДНК последовательности, кодирующей субгрупповой антиген известной специфичности. Субгрупповую принадлежность штамма ротавируса определяет шестой сегмент двунитевой (дн) геномной РНК.
Известны клонированные в плазмидах комплементарные ДНК (кДНК) копии шестого сегмента генома ротавируса обезьян - шт. SAII, ротавируса крупного рогатого скота - шт. RF, ротавирусов свиньи - шт. Готфрид; лошади - шт. Н-2, F 1-14 и ротавирусов человека различных субгрупп - шт. 1076, S 2 (I-я с/гр), шт. Wa (II-я с/гр). Определены и сравнены между собой их первичные последовательности. Показаны диагностические и дифференцирующие возможности ДНК-копий шестого гена различных штаммов ротавирусов в реакции молекулярной гибридизации. Установлено, что степень гомологии полноразмерных нуклеотидных последовательностей шестого гена ротавирусов различных субгрупп достигает 77-95%, что ставит под сомнение возможность использования полноразмерных кДНК для субгрупповой дифференциации ротавирусов методом молекулярной гибридизации. В связи с этим для создания специфичного дифференцирующего зонда необходим выбор участка последовательности кДНК, наиболее вариабельного у ротавирусов различных серологических подгрупп. Известны штаммы E. coli, содержащие рекомбинантные плазмиды с кДНК шестого гена ротавирусов различных субгрупп. Использование этих клеток в качестве продуцентов гибридизационного зонда для субгрупповой дифференциации ротавирусов нецелесообразно в связи с тем, что содержащийся в них генетический материал недостаточно специфично гибридизуется с РНК ротавирусов различных субгрупп.
Целью изобретения является создание штамма Escherichia сoli, содержащего плазмиду - продуцент ДНК(РНК) зонда для специфического выявления ротавирусов II-й субгруппы.
Поставленная цель достигается тем, что:
1. Выделяют днРНК ротавирусов человека, характеризующуюся длинным электрофоретипом (ЭФ-типом).
2. Синтезируют ДНК фрагмент dVR6, комплементарный шестому сегменту РНК. Проводят гибридизационный анализ.
3. Определяют первичную структуру dVR6 фрагмента.
4. Проводят анализ последовательности dVR6 фрагмента с использованием программы "Microgenic" фирмы Beckman.
5. Выделяют фрагмент dVR6-II3. Проверяют его специфичность методом молекулярной гибридизации.
6. Конструируют плазмиду pVR6-II3, в которой dVR6-II3 встроен в полилинкер плазмиды pGEM4Z.
7. Создают штамм E. coli, содержащий плазмиду pVR6-II3 - продуцент ДНК (РНК) зонда для специфического выявления ротавирусов II-й субгруппы.
Сущность предлагаемых объектов изобретения состоит в следующем:
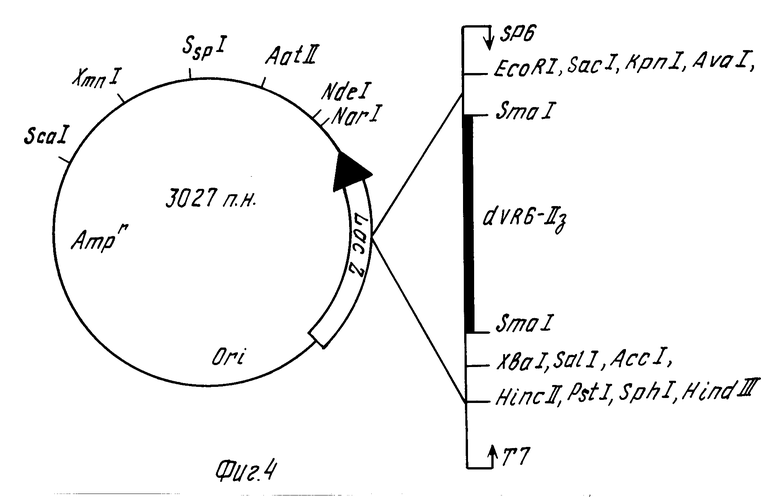
A. Плазмида pVR6-II3 состоит из:
- Sma I-Sma I фрагмента ДНК (dVR6-II3) размером 281 п.н. с определенной последовательностью;
- Sma I-Sma I фрагмента ДНК векторной плазмиды pGEM4Z размером 2746 Т. п.н. и имеет:
- уникальные сайты рестрикции для эндонуклеаз EcoR I, Sac I, Kpn I, Ava I, Xba I, Sal I, Acc I, Hinc II, Pst I, Sph I, Hind III, Sca I, Xmn I, Sep I, Aat II; Nde I;
два слайда рестрикции для эндонуклеазы Sma I;
- промоторы для SP6 РНК полимеразы и Т7 РНК полимеразы;
- ori репликации плазмиды pGEM4Z;
- селективный маркер Amp;
- область, кодирующую α-пептид β -галактозидазы, синтез которой инактивирован встройкой dVR6-II3 фрагмента кДНК.
Б. Штамм - продуцент ДНК-зонда для выявления ротавирусов II-й субгруппы, полученный трансформацией клеток E. coli TG1 с помощью рекомбинантной плазмиды pVR6-II3.
Штамм бактерий Escherichia coli, содержащий плазмиду pVR6-II3, депонирован во Всесоюзной коллекции микроорганизмов под N ВКМ CR-354D, характеризуется следующими признаками:
Морфологические признаки.
Клетки прямые, палочковидные (1,2-1,6 х 2,0 х 6,0 мкм), подвижные, имеют перитрихиальные жгутики, грамотрицательные, неспороносные.
Культуральные признаки.
Клетки хорошо растут на обычно используемых питательных средах. На 1,5% -ном питательном агаре Дифко колонии гладкие, серые блестящие, края ровные, мутные; при выращивании в жидких средах - мясопептонном бульоне и L-бульоне, образуют ровную интенсивную муть;
Физиолого-биохимические признаки.
Оптимальная температура жизнедеятельности 37оС, рН 6,8-7,5. В качестве источника углерода используют многие углеводы, органические кислоты, спирты. В качестве источника азота используют минеральные соли (в аммонийной или нитратной форме), органические соединения (в виде пептона, триптона, аминокислот).
Устойчивость к антибиотикам.
Проявляют устойчивость к ампициллину (100-150 мг/л).
Генетические признаки: Δ (lac pro) Hi, str A, sup E, end A, sbe B, hst R-, F', tra D 36, pro AB, lac IV, 2Δ M15.
Присутствие в штаммах плазмидных ДНК, содержащих фрагменты - зонды, комплементарные шестому сегменту РНК ротавируса человека II c/гр., подтверждается путем проверки их устойчивости к ампициллину, а также путем выделения и анализа плазмидной ДНК одним из известных методов, а также методом блот-гибридизации.
П р и м е р 1. Выделение очищенной РНК ротавирусов человека.
РНК ротавируса человека (РВЧ) выделяли из инфекционного материала больного ротавирусным гастроэнтеритом (изолят N 3480). Ротавирусы данного изолята имеют длинный тип миграции РНК (фиг.1).
500 мг инфекционного материала гомогенизируют в 3 мл буфера, содержащего 0,01 М СН3COONa, 0,3 M NaCl, 4 мМ ЭДТА (рН 5,2), 1% ДСН и инкубируют 1 ч при 37оС. Затем добавляют равный объем прогретой до 65оС фенол:хлороформенной (1: 1) смеси, перемешивают и инкубируют 3-5 мин при 65оС. Центрифугируют при 5 тыс. об/мин в течение 10 мин и отбирают супернатант. Экстракцию повторяют один раз при 65оС и затем при комнатной температуре аналогично описанному выше. К отобранному супернатанту добавляют 0,1 объема 3 М ацетата натрия (рН 5,2) и 2 объема этанола, выдерживают 18-20 ч при -20оС и центрифугируют при 5 тыс. об/мин в течение 1 ч. Осадок растворяют в 1 мл буфера, содержащего 0,01 М Трис-HCl, рН 8,0, 0,001 М MgCl2, 0,3 M NaCl, 20 мкг/мл панкреатической ДНКазы, инкубируют 30 мин при 25оС, добавляют равный объем фенола (рН 8,0), интенсивно перемешивают и центрифугируют при 5 тыс. об/мин в течение 10 мин. Водную фазу отбирают. РНК преципитируют этанолом как описано выше и растворяют в буфере, содержащем 10 мМ Трис-HCl, pH 8,0, 1 мМ ЭДТА (ТЕ-буфер). Осадки хранят при -20оС.
На следующем этапе работы проводят электрофорез РНК в 10% ПААГ в течение 18 ч с использованием буферной системы Лэммли. Гель окрашивают этидиумбромидом и под ультрафиолетом вырезают шестой сегмент РНК. Кусочек геля измельчают, заливают буфером, содержащим 0,5 М ацетата аммония (рН 8,0), 0,1% ДСН и 1 мМ ЭДТА, выдерживают при 37оС в течение ночи и проводят фенольную экстракцию. РНК преципитируют этанолом, высушивают, растворяют в ТЕ-буфере и используют для получения полноразмерной ДНК-копии.
П р и м е р 2. Получение полноразмерной копии шестого гена.
А. Синтез двухцепочечной кДНК
Синтез кДНК осуществляют по известному методу (McCrae M.A., McCorquodale J.G. J. Virol. 1982, 44, N 3, р. 1076-1079).
днРНК (5-10 мкг) в ТЕ-буфере денатурируют при 100оС 5 мин и охлаждают при 0оС. Реакцию полиаденилирования 3'-концов РНК проводят в объеме 200 мкл при 37оС в буфере, содержащем 50 мМ Трис-HCl pH 7,9, 10 мМ MgCl2, 2,5 мМ MnCl2, 200 мМ NaCl, 50 мкг/мл БСА, 0,1 мМ АТФ (включая 20 μ Сi H3-АТФ) и 1/10 объема поли-аполимеразы (0,5 мкг) в течение 10 мин, за которые присоединяется 15-20 остатков АМФ. Степень аденилирования определяют по известной методике. Полученный препарат экстрагируют фенолом и осаждают 2,5 объемами этанола.
К препарату полиаденилированной РНК в буфере ТЕ добавляют 5 мкг олиго-dT12-18, денатурируют 3 мин при 100оС и охлаждают во льду. Синтез кДНК проводят в буфере, содержащем 50 мМ Трис-HCl pH 8,3, 50 мМ KCl, 10 мМ MgCl2, 10 М ДТТ, 4 мМ пирофосфата натрия, 0,5 мМ dАТФ, d ТФ, d ГТФ, 0,5 мМ d ТТФ, 10μ Ci α-P32-АТФ в объеме реакционной смеси 100 мкл. Добавляют 25 ед. обратной транскриптазы из вируса миелобластоза птиц (Омутнинский химический завод) и инкубируют 2 ч при 42оС.
Цепи РНК удаляют щелочным гидролизом (0,25 М NaOH 20 мин при 70оС). Препарат нейтрализуют HCl, депротеинизируют равным объемом фенола, нуклеиновые кислоты осаждают этанолом и растворяют в 50 мкл ТЕ-буфера. Препарат очищают от невключившихся d НТФ на колонке с сефадексом G-50 (medium), уравновешенной ТЕ-буфером. Отбирают фракции, содержащие кДНК.
Для получения двунитевой (дн) кДНК препарат прогревают 2 мин при 100оС, добавляют NaCl до 0,2 М и отжигают комплементарные молекулы ДНК 30 мин при 65оС, 60 мин при 50оС, 30 мин при 40оС. Двунитевую кДНК осаждают этанолом.
Достройку дн кДНК до образования тупых концов проводят с помощью фермента Кленова в буфере, содержащем 0,1 М Hepes pH 6,9, 10 мМ MgCl2, 5 мМ ДТТ, 0,14 М KCl, 1 мМ каждого g НТФ, 50 ед. фрагмента Кленова, при 15оС в течение 20 ч. Реакцию останавливают добавлением 2 мкл 0,5 М ЭДТА, проводят экстракцию фенолом и хроматографию на колонке с сефадексом G-50. Фракции, содержащие кДНК, объединяют, дн кДНК осаждают этанолом.
Реакцию достройки полиС-оннекторов проводят в 25 мкл инкубационной смеси следующего состава: 0,2 M какодилат натрия рН 7,9, 8 мМ MgCl2, 1 мМ β-меркаптоэтанола, 2 мМ g ЦТФ, 20 ед. концевой нуклеотидилтрансферазы и 0,5 мкг дн кДНК. Реакцию проводят при 37оС в течение 30 мин, экстрагируют фенолом и осаждают этанолом как описано выше.
Б. Клонирование кДНК в векторной плазмиде рИС9
5 мкг векторной плазмиды рИС9 гидролизуют рестрикционной эндонуклеазой Pst I в 50 мкл буфера, содержащего 20 мМ KCl, 10 мМ Трис-HCl, pH 8,0, 10 мМ MgCl2, 60 мМ МЭ (У2) и 10 ед. Pst I. Реакцию проводят при 37оС 1 ч, затем депротеинизируют 2-кратной экстракцией фенолом (рН 8,0), фенол удаляют экстракцией равным объемом хлороформа и плазмидную ДНК осаждают спиртом. Достройку поли-G-коннекторов проводят при 37оС 30 мин, как указано выше (за исключением того, что вместо gЦТФ берут gГТФ, а вместо MgCl2 - CoCl2).
Плазмидную ДНК и дн кДНК смешивают в эквимолярных соотношениях в 20 мкл буфера, содержащего 10 мМ Трис-HCl pH 7,5, 1 мМ ЭДТА, 0,1 М NaCl, инкубируют 5 мин при 65оС, затем 2 ч при 42оС и охлаждают до 0оС.
Полученным препаратом ДНК трансформируют компетентные клетки E. coli. C этой целью кл. E. coli Z 85 выращивают при 37оС в 10 мл бульона 2хУТ (Bacto-триптон 16 г/л, дрожжевой экстракт 10 г/л, NaCl 5 г/л) до титра 3˙ 108 кл/мл. После охлаждения во льду клетки собирают центрифугированием (5 тыс. об/мин, 10 мин, 0оС), суспендируют в 10 мл раствора, содержащего 10 мМ Трис-HCl pH 8,0, 50 мМ CaCl2, выдерживают 10 мин во льду. Клетки повторно центрифугируют при 5 тыс об/мин 10 мин при 0оС, суспендируют в 0,5 мл того же раствора и используют для трансформации.
К 50 мкл смеси гибридных молекул ДНК добавляют 100 мкл клеток, инкубируют 1 ч при 0оС, затем 2 мин при 42оС. После 10-кратного разбавления бульоном 2хУТ клетки подращивают 2 ч с интенсивной аэрацией при 37оС и высеивают на агаризованную среду 2хУТ, содержащую 0,004% X-gal, 40 мкМ IPTG и 100 мкг/мл ампициллина.
Из выращенных при 37оС клонов трансформантов белого цвета выделяют плазмидную ДНК щелочным методом. Осадок плазмидной ДНК собирают центрифугированием, промывают 70%-ным спиртом и суспендируют в 100 мкл буфера ТЕ.
Выделенные плазмидные ДНК расщепляют эндонуклеазой рестрикции Pst I в 20 мкл буфера У2, содержащего 1 мкг ДНК и 2 ед. Pst I при 37оС в течение 1 ч. Полученные препараты анализируют в 1%-ном агарозном геле. Электрофорез проводят в трис-ацетатном буфере (0,04 М Трис-HCl pH 7,6, 0,03 М CH3COONa, 1 мМ ЭДТА) при постоянном токе (20 мА) в течение 3-4 ч. Гель окрашивают бромистым этидием (0,5 мкг/мл) и фотографируют в ультрафиолетовом свете.
По данным рестрикционного анализа отбирают плазмидные ДНК, содержащие вставки, по молекулярной массе соответствующие полноразмерной ДНК-копии. Плазмидную ДНК, содержащую полноразмерную копию шестого гена d VR6, обозначают p VR6.
В. Проверка специфичности dVR6-фрагмента
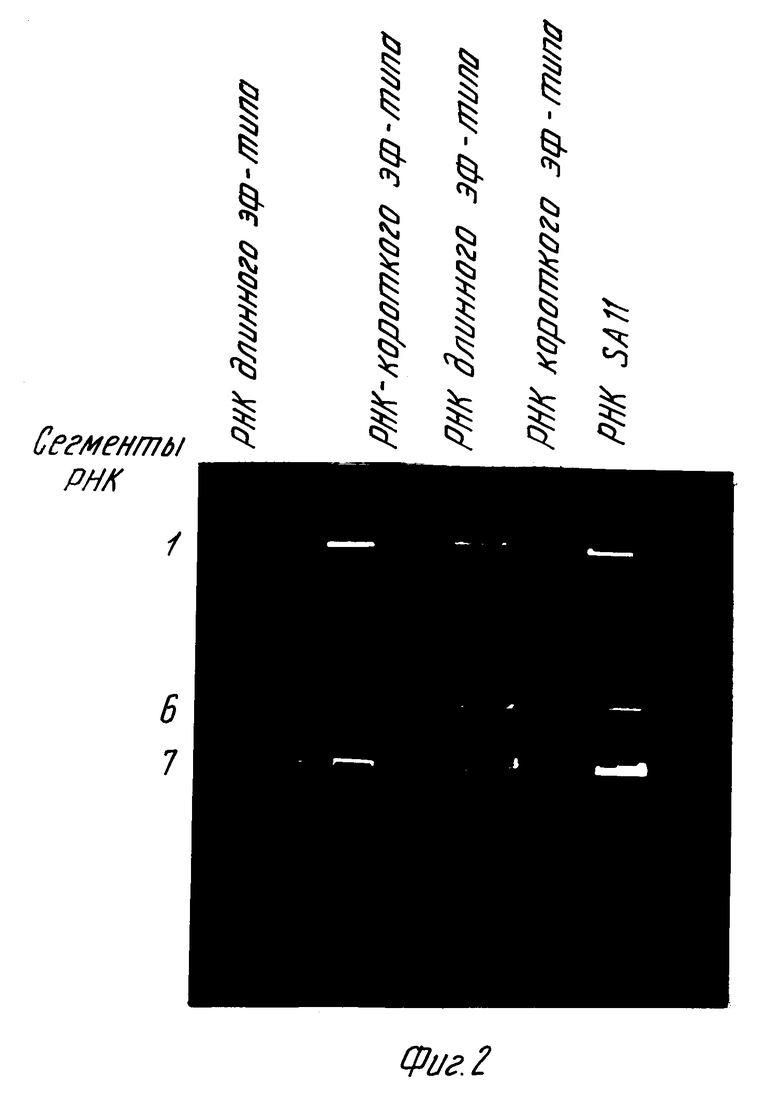
Фрагмент dVR6 нарабатывают в препаративном количестве, как описано выше, метят Р32-дАТФ в процессе ник-трансляции и проверяют специфичность методом блот-гибридизации. Используют тотальную РНК ротавирусов различных субгрупп - РНК ротавируса человека длинного ЭФ-типа (II c/гр), РНК ротавируса человека короткого ЭФ-типа (I c/гр) и РНК ротавируса обезьян SAII, имеющую длинный ЭФ-тип, но относящуюся к 1 с/гр. РНК разгоняют в ПААГ при 10 мМ на гель в течение 18 ч в буферной системе Лэммли, переносят на нейлоновый фильтр Z-probe методом щелочного переноса и проводят гибридизацию с Р32-АТР меченой полноразмерной копией шестого гена - dVR6 фрагментом. Устанавливают, что dVR6-фрагмент гибридизуется с соответствующим сегментом РНК РВЧ длинного ЭФ-типа, не гибридизуется с РНК РВЧ короткого ЭФ-типа, но гибридизуется с РНК SAII (фиг.2). Результаты позволяют сделать вывод о недостаточной специфичности dVR6-фрагмента и невозможности использования полноразмерной копии шестого гена для субгрупповой дифференциации ротавирусов.
П р и м е р 3. Секвенирование dVR6-фрагмента
А. Получение производных фрагмента dVR6, укороченных с Hind III-конца
ДНК плазмиды pVR6 (2 мкг) гидролизуют эндонуклеазой рестрикции Hind III (5 ед. ) в буфере У2 в объеме реакционной смеси 40 мкл в течение 2 ч при 37оС. Препарат экстрагируют равным объемом фенола, осаждают 2,5 объемами этанола и растворяют полученный осадок в 10 мкл буфера ТЕ.
Полученный препарат обрабатывают нуклеазой Bal31 (2 ед.) в 20 мкл буфера, содержащего 20 мМ Трис-HCl pH 7,3, 0,6 M NaCl, 12,5 мМ MgCl2, 12,5 мМ CaCl2, при 30оС в течение 10 мин. Реакцию останавливают добавлением ЭДТА до 20 мМ, препарат экстрагируют фенолом, осаждают 2,5 объемами этанола, осадок промывают 70о этанолом и растворяют в 20 мкл ТЕ. Гидролизуют ДНК эндонуклеазой рестрикции EcoRI (5 ед.) в 30 мкл буфера У2 при 37оС 2 ч. Препарат подвергают электрофорезу в 0,9%-ном агарозном геле. Набор укороченных с Hind III конца фрагментов ДНК вырезают и элюируют из геля с помощью электроэлюции.
Полученные фрагменты ДНК клонируют в плазмиде рИС9 (предварительно гидролизованной эндонуклеазами EcoRI и Sma I) по известной методике. Клоны анализируют на наличие плазмидной ДНК с помощью электрофореза в агарозном геле и отбирают клоны, несущие плазмиды с делециями фрагмента dVR6 размером 200, 300, 400 и 500 н.п.
Б. Получение производных фрагмента dVR6, укороченных с EcoRI-конца.
Аналогично получают делеции фрагмента dVR6 c EcoRI-конца, для чего ДНК плазмиды pVR6 первоначально гидролизуют эндонуклеазой рестрикции EcoRI.
B. Определение нуклеотидной последовательности фрагмента dVR6.
Плазмиду pVR6, а также плазмиды с делеционными производными фрагмента dVR6 гидролизуют соответствующей эндонуклеазой рестрикции (Hind III или EcoRI). Концевое мечение фосфором 32 полученных препаратов ДНК осуществляют с помощью ДНК-полимеразы (фрагмент Кленова) по известной методике. Выделение и очистку меченого фрагмента ДНК проводят известным способом. Нуклеотидную последовательность меченых фрагментов определяют по методу Максама-Гилтерта. Полученная в результате полная нуклеотидная последовательность фрагмента dVR6 имеет следующий вид;
GGCTTTAAAACGAAGTCTTCGACATGGAGGTTCTGTACTCACTATCAAAAACTCTTAAAGATGCTA GGGACAAAATTGTTGAAGGTACATTATATTCTAATGTAAGCGATCTTATTCAGCAATTCAATCAAA TGATAGTAACTATGAATGGAAATGACTTTCAGACTGGAGGAATTGGTAATTTACCTGTTAGAAATT GGACTTTCGATTTTGGTCTATTAGGTACAACACTTTTGAACTTGGATGCTAATTATGTTGAGAATG CAAGAACTACAATTGAATATTTTATTGACTTTATTGATAATGTATGTATGGATGAAATGGCAAGAG AATCTCAAAGAAATGGAGTAGCGCCACAATCTGAAGCATTGAGAAAGTTAGCGGGAATAAAATTTA AGAGAATAAATTTCGATAATTCATCAGAATACATAGAAAATTGGAACTTACAAAATAGAAGACAGC GCACCGGATTTGTTTTCCATAAACCTAACATATTTCCATACTCAGCTTCATTTACTCTAATAAGAT CTCAACCAATGCATGATAATTTAATGGGAACCATGTGGCTTAATGCTGGATCAGAAATTCAAGTGG CTGGATTTGATTACTCATGCGCCATAAATGTACCAGCGAACATACAGCAATTTGAACATATCGTCC AGCTTAGGCGCGCACTGACTACAGCTACTATAACTTTATTACCTGATGCAGAGAGATTTAGTTTTC CAAGAGTAATTAATTCAGCTGATGGCGCGACTACTTGGTTCTTTAATCCAGTTATTCTAAGACCAA ATAATGTAGAGGTAGAATTTTTGTTGAATGGACAAATTATTAATACATATCAGGCTAGATTTGGTA CTATAATCGCAAGAAATTTTGATACAATTCGGTTTATATTTCAGTTGATGCGTCCACCTAATATGA CACCAGCTGTTAATGCACTGTTGCCACAAGCACAACCTTTTCAGCACCATGCAACAGTTGGACTTA CATTACGTATTGAATCTGCGGTTTGTGAATCAGTGCTTGCGGACGCGAATGAAACTCTGTTAGCAA ATGTGACCGCGGTGCGTCAAGAATATGCCATACCAGTTGGACCGATATTTCCACCAGACATGAATT GGACTGAATTGATTACTAACTATTCGCCATCTAGAGAAGATAACTTGCAACGCGTTTTCACGGTAG CTTCCATTAGAAGCATGTTGATTAAGTGAGGACCAGACTAAACATCTGGTATCCAATCTTAGTTAG CATGTAGCTACATCAAGTCATTCAGACTCTTCAAGTAAGGACATGATTTCATGTTCGCTACGTAGA GTAACTGTCTGAATGATGTAGTGAGAGGATGTGACC
П р и м е р 4. Анализ последовательности dVR6 c использованием программы "MicroGenic".
A. Оформление субгрупповой специфичности dVR6-фрагмента.
Проводят сравнительный анализ определенной последовательности dVR6-фрагмента с первичной структурой гена 6 ротавирусов.
Рассчитывают, что полноразмерная кДНК dVR6-фрагмента имеет 98,8% гомологии с шестым геном ротавируса человека шт. Wa, относящегося ко II-й серологической подгруппе, и 77% гомологии с кДНК шестого гена ротавируса обезьян шт. SAII, относящегося к I-й cубгруппе, 83,3% с кДНК шестого гена ротавируса субгрупп I и II (шт. F 1-14) и 88,1% с кДНК шестого гена ротавируса субгруппы ни I, ни II (шт. Н-2). Делают вывод, что клонированный dVR6-фрагмент определяет специфичность II-й субгруппы.
Б. Выбор вариабельного участка.
Проводят поиск вариабельных и консервативных участков шестого гена ротавирусов различных серологических подгрупп. Находят, что 5'-конец (1-750 п. н. ) наиболее консервативен у ротавирусов II-й и I-й субгрупп (гомология 82,4% ). Устанавливают, что участок 750-1000 п.н. наиболее вариабелен у ротавирусов I-й и II-й субгрупп (гомология 73%).
Проводят поиск сайтов рестрикции на кДНК dVR6. Устанавливают, что искомый участок последовательности может быть вырезан эндонуклеазой рестрикции FNU2 (Bsp50 1). Рестриктаза FNU2 вырезает фрагмент ДНК размером 281 п.н. , расположенный между 753-й и 1034-й парами оснований. Выбранный участок имеет 99% гомологии с соответствующей нуклеотидной последовательностью участка шестого гена ротавируса человека шт. Wa(II c/гр) и 74% гомологии с кДНК РНК ротавируса обезьян шт. SAII (1 c/гр). Выбранный участок обозначают dVR6-II3.
П р и м е р 5. Выделение фрагмента dVR6-II3 и проверка его специфичности.
Фрагмент dVR6 нарабатывают в препаративном количестве как описано выше. Проводят рестрикцию ДНК dVR6 c помощью эндонуклеазы Bsp50I в буфере У2 в течение 18 ч при 37оС. Фрагмент кДНК размером 281 пары нуклеотидов (dVR6-II3) выделяют методом электрофореза в легкоплавкой агарозе. Фрагмент dVR6-II3 метят Р32 в реакции ник-трансляции и проводят гибридизационный анализ по стандартной методике. Для этого на нейлоновый фильтр производства р/к Хийу Калур наносят в точку очищенную РНК ротавирусов человека длинного ЭФ-типа (II c/гр), РНК ротавирусов человека короткого типа (I c/гр) и РНК ротавируса обезьян шт. SAII, имеющую длинный ЭФ-тип, но относящуюся к I субгруппе, и используют в гибридизационном анализе. Устанавливают, что фрагмент d VR6-II3гибридизуется с РНК II-й субгруппы и не гибридизуется с РНК различных ЭФ-типов ротавирусов I-й субгруппы (фиг.3, где 1 - кДНК dVR6-II3; 2 - кДНК dVR6; 3 - РНК РВ4 (II c/гр, длинный ЭФ-тип; 4 - РНК РВЧ (I c/гр, короткий ЭФ-тип; 5 - РНК AII (I c/гр, длинный ЭФ-тип). Результаты гибридизационного анализа позволяют заключить, что фрагмент dVR6-II3специфически выявляет ротавирусы II-й субгруппы.
П р и м е р 6. Конструирование плазмиды pVR6-II3.
5 мкг векторной плазмиды pGEM4Z гидролизуют эндонуклеазой рестрикции Sma I в 50 мкл буфера У2 в течение ночи при 30оС, затем депротеинизируют 2-кратной экстракцией фенолом (рН 8,0). Фенол удаляют экстракцией равным объемом хлороформа и плазмидную ДНК осаждают спиртом.
Проводят восстановление тупых концов у фрагмента dVR6-II3. С этой целью ДНК dVR6-II3 обрабатывают фрагментом Кленова в буфере, содержащем 10 мМ Трис-HCl, pH 7,5, 50 мМ NaCl, 10 мМ MgCl2, 2 мМ спермидина и по 100 мкМ дЦТФ и дГТФ в течение 15 мин при комнатной температуре. Препарат депротеинизируют с использованием фенола.
Плазмидную ДНК и ДНК dVR6-II3 смешивают в эквивалентных количествах и осаждают 2,5 объемами этанола. Осадок ДНК растворяют в 30 мкл буфера, содержащего 10 мМ Трис-HCl (pH 7,5), 10 мМ MgCl2, 10 мМ ДТТ, 0,0025 мМ АТФ и 10 ед. акт. ДНК-лигазы Т4. Реакцию проводят в течение ночи при 20оС.
Результативность лигазной реакции проверяют методом электрофореза в 1% -ном агарозном геле.
П р и м е р 7. Создание штамма E. coli - продуцента фрагмента dVR6-II3.
В качестве резистентных клеток используют штамм E. coli TG1. Компетентные клетки готовят как описано выше для E. coli Z 85.
К 50 мкл смеси гибридных молекул ДНК добавляют 100 мкл клеток, инкубируют 1 ч при 0оС, затем 2 мин при 42оС. После 10-кратного разведения бульоном 2хУТ клетки подращивают 2 ч с интенсивной аэрацией при 37оС и высевают на агаризованную среду 2хУТ, содержащую 0,004% бром-хлор-индолил-галактозидазы, 40 мМ изопропил-тиогалактозидазы и 100 мк/мл ампициллина.
Из выросших при 37оС клонов трансформантов белого цвета выделяют плазмидную ДНК щелочным способом.
Выделенные плазмидные ДНК расщепляют эндонуклеазой рестрикции Sma I, либо любой из пар эндонуклеаз, расположенных в полилинкере выше и ниже встроенного фрагмента кДНК (фиг.4) в буфере У2.
Полученные препараты анализируют в 1%-ном агарозном геле в трис-боратной буферной системе при 100v в течение 2 ч. Гель окрашивают бромистым этидием (0,5 мкг/мл) и анализируют в ультрафиолетовом свете.
По данным рестрикционного анализа отбирают штамм E. coli, продуцирующий рекомбинантную ДНК, содержащую фрагмент dVR6-II3. Физическая карта сконструированной плазмидной ДНК рVR6-II3 представлена на фиг.4.
Таким образом, в результате генноинженерных разработок создан штамм E. coli N ВКМ CR-354D, несущий рекомбинантную плазмиду - продуцент dVR6-II3 фрагмента ДНК-зонда для специфического выявления ротавирусов человека II-й субгруппы.
Фрагмент dVR6-II3 специфически выявляет РНК ротавирусов II-й субгруппы за счет того, что является наиболее вариабельным участком гена, кодирующего субгрупповой антиген у ротавирусов различных субгрупп.
Созданная плазмида pVR6-II3 позволяет нарабатывать фрагмент dVR6-II3. Уровень синтеза зонда в составе плазмиды в сконструированном штамме E. coli ВКМ CR-354D составляет при плотности культуры 5 х 108клеток/мл 1,5 мг/л.
Штамм Escherichia coli ВКМ CR-354D является продуцентом фрагмента dVR6-II3 ДНК-зонда для субгрупповой дифференциации ротавирусов.
dVR6-II3 фрагмент ДНК зонда для специфического выявления ротавирусов II-й субгруппы, рекомбинантная плазмида pVR6-II3, содержащая фрагмент dVR6-II3, штамм E. coli ВКМ CR-354D - продуцент зонда созданы и разработаны в Нижегородском НИИ эпидемиологии и микробиологии ГК РСФСР санитарно-эпидемиологического надзора и Институте биохимии и физиологии микроорганизмов АН СССР.


Изобретение относится к медицине, в частности к диагностике вирусных заболеваний человека, генетической инженерии и биотехнологии. Сущность изобретения состоит в том, что получена рекомбинантная плазмидная ДНК pVR6-II3 размером 3027 п. н. для специфического выявления ротавирусов II субгруппы, и содержит фрагмент ДНК плазмиды dVR6-II3 размером 281 п.н., гомологичный 753 - 1034 п.н. участку шестого гена ротавируса человека II субгруппы, фрагмент векторной плазмиды pGEM4z размером 2746 п.н. Предметом изобретения является также штамм бактерий - носитель фрагмента ДНК dVR6z для специфического выявления ротавирусов II субгруппы. 4 ил.
РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PVR6-IIз, ПРЕДНАЗНАЧЕННАЯ ДЛЯ СПЕЦИФИЧЕСКОГО ВЫЯВЛЕНИЯ РОТАВИРУСОВ II СУБГРУППЫ, ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - НОСИТЕЛЬ РЕКОМБИНАНТНОЙ ПЛАЗМИДНОЙ ДНК PVR6-β, ПРЕДНАЗНАЧЕННОЙ ДЛЯ ВЫЯВЛЕНИЯ РОТАВИРУСОВ II СУБГРУППЫ.
| ПОЛУАВТОМАТИЧЕСКАЯ ЛИНИЯ ДЛЯ ОТДЕЛКИ i" МЕБЕЛЬНЫХ ЩИТОВI | 0 |
|
SU331939A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
