Изобретение относится к области биотехнологии, а именно к способам и технологии нейтрализации вируса гепатита С, а именно к антителам против вируса гепатита С, и может использоваться в медицине, фармацевтической промышленности и смежных отраслях науки и техники.
Гепатит С является широко распространенным в мире опасным вирусным заболеванием. Согласно проведенным статистическим исследованиям, в мире вирусом гепатита С (HCV) заражены более 170 миллионов человек, то есть почти в 5 раз больше, чем вирусом иммунодефицита человека (ВИЧ). HCV-инфекция вышла на первое место в ряду причин печеночно-клеточного рака; кроме того, она стала наиболее частой причиной трансплантации печени у взрослых в западных странах (Mohd Hanafiah K, Groeger J, Flaxman AD, Wiersma ST. Hepatology. 2013, 57(4):1333-1342). HCV-инфекция во многих случаях протекает без клинических проявлений, несмотря на воспаление и фиброз печени, которые носят стойкий и прогрессирующий характер и в конечном итоге приводят к циррозу печени, печеночной недостаточности или раку печени (Zoulim F, Chevallier М, Maynard M, Trepo С.Rev Med Virol. 2003, 13(1):57-68). Проблема усугубляется тем, что вирус HCV очень часто остается в организме инфицированных больных навсегда - доля инфицированных, у которых вирус вызывает хроническую инфекцию, составляет до 85%. Все эти причины делают исследования, направленные на диагностику и лечение гепатита С, весьма актуальными и практически важными.
Вирус гепатита С (Hepatitis С Virus, HCV) был впервые идентифицирован в 1989 г. (Chen SL, Morgan TR. Int J Med Sci. 2006, 3(2):47-52), однако масштаб проблемы, которую он представляет для здравоохранения, и нагрузки, создаваемой вызываемым им заболеванием, в полной мере осознан лишь в последние годы, когда появились достаточно надежные методы его диагностики. В настоящее время считается, что распространение данного вируса происходило в течение нескольких десятилетий или даже дольше, причем случаи заражения в недавнем прошлом были часто связаны с внутривенным введением лекарственных препаратов и другими неустановленными факторами/способами передачи инфекции (Aceijas С, Rhodes Т. Int J Drug Policy. 2007, (5):352-358).
HCV - это небольшой (40-60 нм в диаметре), оболочечный, одноцепочечный РНК-содержащий вирус рода Hepacivirus семейства Flaviviridae. Как геном вируса HCV, так и белки, кодируемые его генами, полностью охарактеризованы (Kato N. Microb Comp Genomics. 2000, 5(3): 129-151). Положительная цепь РНК вирусного генома состоит из 5′-нетранслируемой области, одной открытой рамки считывания длиной около 9 000 нуклеотидов и короткой 3′- нетранслируемой области (фиг.1). Открытая рамка считывания кодирует один полипептид длиной приблизительно 3 010 аминокислот, так называемый «полипротеин», который во время и после трансляции расщепляется в нескольких точках под действием сигнальных хозяйских пептидаз, а также протеаз, кодируемых HCV (Niepmann M. Curr Top Microbiol Immunol. 2013, 369:143-166). В результате этого расщепления образуются по крайней мере три структурных белка (Core, E1 и Е2) и шесть неструктурных (NS) белков (NS2, NS3, NS4A, NS4B, NS5A и NS5B).
Е1 и Е2 представляют собой два сильно гликозилированных белка вирусной оболочки, которые могут взаимодействовать с плазматическими мембранами гепатоцитов и других клеток, опосредуя прикрепление вируса и его проникновение в клетку. Белки Е1 и Е2 образуют друг с другом гете-ромерные комплексы, однако вопрос о том, должны ли эти белки ассоциировать друг с другом для связывания с клеточными мембранами, окончательно не решен. Гликопротеин Е1 (аминокислоты 192-383 полипротеина) служит шапероном для правильной укладки белка Е2 во время его синтеза, облегчает связывание Е2 с поверхностными рецепторами клеток и, как считается, участвует в слиянии вируса с клеточной мембраной (Poenisch M, Bartenschlager R. Semin Liver Dis. 2010, 30(4):333-347). Гликопротеин Е2 (аминокислоты 384-746 полипротеина) содержит гипервариабельный участок в N-концевой области (аминокислоты 384-411). NS-белки представляют собой ферменты или вспомогательные факторы, которые катализируют и регулируют репликацию РНК-генома HCV. Соге-белок взаимодействует с вирусной РНК и образует нуклеокапсид (Piceiro D, Martinez-Salas Е. Viruses. 2012, 4(10):2233-2250).
В настоящее время предполагается следующая модель жизненного цикла HCV (Kim CW, Chang KM. Clin Mol Hepatol. 2013, 19(1):17-25; Pawlotsky JM, Chevaliez S, McHutchison JG. Gastroenterology. 2007, 132(5); 1979-1998). Заражение вирусом начинается с его прикрепления к клетке за счет специфического взаимодействия между молекулами, расположенными на поверхности клетки-мишени, и белками оболочки вируса. Прикрепление HCV к поверхности клетки опосредуется связыванием гликопротеина Е2 с рецептором тетраспанинового семейства CD81, экспрессирующимся на гепатоцитах и В-лимфоцитах, после чего возникает более специфическое взаимодействие с одним или несколькими «входными» рецептороми, среди которых идентифицированы: рецептор липопротеинов низкой плотности, скавенджер-рецептор SR-BI, клаудин-1 и окклю-дин. На следующем этапе вирус входит в клетку, по-видимому, за счет рецептор-опосредованного эндоцитоза. Плюс-цепь РНК вирусного генома подвергается трансляции в цитоплазме, и образующийся полипептид процессируется на мембране эндоплазматического ретикулума. Там же происходит сборка репликазы и направляемый ею синтез минус-цепи РНК, которая затем используется в качестве матрицы для синтеза молекул плюс-цепи РНК. Плюс-цепь РНК инкапсулируется структурными белками и заключается в оболочку путем почкования в просвет эндоплазматического ретикулума. В конечном итоге, инфекционные вирионы высвобождаются за счет транспорта через комплекс Гольджи.
Вследствие высокой частоты репликации и отсутствия функции редактирования у репликазы вируса, геном HCV характеризуется высокой генетической изменчивостью. HCV подразделяется на шесть основных генотипов, нуклеотидные последовательности которых различаются примерно на 30%; при этом в мире насчитывается более 30 подтипов этого вируса. В США и Европе доминируют подтипы 1а и 1b, в то время как в большинстве азиатских стран часто встречается подтип lb (Doyle JS, Hellard ME, Thompson AJ. Best Pract Res Clin Gastroenterol. 2012, 26(4):413-427; Hara K, Rivera MM, Koh C, Sakiani S, Hoofhagle JH, Heller T. J Clin Microbiol. 2013, 51(5):1485-1489; Bukh J, Miller RH, Purcell RH. Semin Liver Dis. 1995, 15(1):41-63).
Вирус HCV заражает человека главным образом через гепатоциты, однако имеются данные, что HCV способен также инфицировать другие органы и клетки, в особенности, клетки лимфоидных органов. Подобная внепеченочная инфекция может вносить свой вклад в опосредованный иммунной системой патогенез хронического заболевания печени и/или в развитие аутоиммунных заболеваний, в том числе смешанной кри-оглобулинемии и гломерулонефрита (Inokuchi M, Ito Т, Uchikoshi M, Shimozuma Y, Morikawa K, Nozawa H, Shimazaki Т, Hiroishi К, Miyakawa Y, Imawari M. J Med Virol. 2009, 82(12):2064-2072; Morsica G, Tambussi G, Sitia G, Novati R, Lazzarin A, Lopaico L, Mukenge S. Blood. 1999, 94(3): 1138-1139).
Современные методы лечения HCV-инфекции далеко не оптимальны. Интерферон-альфа в комбинации с рибавирином, которые применяются для лечения гепатита С в настоящее время, позволяют получить желаемый ответ на лечение лишь у 50% от леченых больных; при этом значительная часть больных вообще не могут применять эти препараты из-за возникающих у них нежелательных явлений (Wong W, Terrault N. Clin. Gastroenterol. Hepatol. 2005, 3(6):507-520.). При этом ощущается острая потребность в более эффективных средствах для предотвращения рецидива гепатита С у пациентов, которым проведена трансплантация печени (повторная HCV-инфекция у пациентов, которым была проведена трансплантация печени в связи с терминальной печеночной недостаточностью, обусловленной HCV-инфекцией, встречается практически в 100% случаев). Отмечено, что пациенты, перенесшие трансплантацию, переносят лечение интерфероном-альфа в комбинации с рибавирином плохо. Более того, интерферон-альфа может в ряде случаев усугублять реакцию отторжения трансплантата (Escudero A, Rodraguez F, Serra MA, Del Oimo JA, Montes F, Rodrigo JM. J Gastroenterol Hepatol. 2008, 23(6):861-866; Abe H, Aida Y, Ishiguro H, Yoshizawa K, Seki N, Miyazaki T, Itagaki M, Sutoh S, Ika M, Kato K, Shimada N, Tsubota A, Aizawa Y. J Med Virol. 2013, 85(9):1523-1533).
Хотя в последние несколько лет осложнения и смертность, связанные с гепатитом С, удалось значительно снизить за счет применения направленных противовирусных препаратов, в частности ингибиторов протеазы и обратной транскриптазы, разработка новых лекарственных средств против HCV наталкивается на серьезные препятствия, к числу которых относятся способность вируса к хронической персистенции, генетическая диверсификация, возникающая при репликации в хозяйском организме, появление мутантных форм вируса, устойчивых к лекарственным препаратам. Кроме того, надо учитывать, что противовирусные лекарственные препараты могут вызывать негативные побочные эффекты.
В настоящее время многие биотехнологические и фармацевтические компании осуществляют поиск новых методов направленного воздействия на уже известные вирусные мишени, разрабатывают профилактические и/или терапевтические вакцины против HCV, а также усовершенствуют уже имеющиеся методы лечения на основе интерферона. В клинической разработке находятся средства лечения гепатита С, которые должны быть более безопасными и эффективными; некоторые из них представляют собой продукты модификации интерферона-альфа. К числу ингибиторов, находящихся во II фазе клинической разработки, относятся вещества, мишенями которых являются сайт внутреннего связывания рибосом (IRES) HCV, протеаза NS3 и полимераза NS5B. Клиническая разработка одного из возможных ингибиторов протеазы NS3 была остановлена, поскольку он оказывал токсическое действие на сердце млекопитающих, что говорит о возможных проблемах, которые могут встать при разработке лекарственных препаратов этого класса в будущем.
Недавно разработанные ингибиторы NS3-протеазы вируса HCV (те-лапревир и цепревир) позволяют увеличить частоту стойкого вирусологического ответа, однако их можно применять лишь у пациентов, инфицированных HCV генотипа 1; кроме того, у пациентов, получающих эти лекарственные препараты, быстро возникает устойчивость вируса к ним. (Kwo PY, Lawitz EJ, McCone J, SchiffER, Vierling JM, Pound D, Davis MN, Galati JS, Gordon SC, Ravendhran N, Rossaro L, Anderson FH, Jacobson IM, Rubin R, Koury K, Pedicone LD, Brass CA, Chaudhri E, Albrecht JK. Lancet. 2010, 376(9742):705-716; McHutchinson J.G., McHutchison JG, Manns MP, Muir AJ, Terrault NA, Jacobson IM, Afdhal NH, Heathcote EJ, Zeuzem S, Reesink HW, Garg J, Bsharat M, George S, Kauffman RS, Adda N, Di Bisceglie AM. N Engl J Med. 2010, 362 (14):1292-1303).
Для связывания и ингибирования микро РНК печени miR-122, которая необходима для репликации вируса, предлагается использовать фосфоротиоат-модифицированный олигонуклеотид (препарат миравирсен) (Janssen HL, Reesink HW, Lawitz EJ, Zeuzem S, Rodriguez-Torres M, Patel K, van der Meer AJ, Patick AK, Chen A, Zhou Y, Persson R, King BD, Kauppinen S, Levin AA, Hodges MR. N Engi J Med. 2013, 368(18):1685-1694). Недостатком подобных препаратов, направленных на снижение уровня miR-122, является возможность образования и ускоренное развитие злокачественных новообразований (Koberle V, Kronenberger В, Pleli Т, Trojan J, Imelmann E, Peveling-Oberhag J, Welker MW, Elhendawy M, Zeuzem S, Piiper A, Waidmann O. Eur J Cancer. 2013, dot: 10.10167j.ejca.2013.06.002).
Среди появившихся за последние годы новых и более эффективных методов лечения наиболее перспективным является иммунотерапия с использованием терапевтических моноклональных антител, специфичных к белкам HCV (Sautto GA, Diotti RA, Clementi M. New Microbiol. 2012, 35(4):387-397; Haberstroh A, Schnober EK, Zeisel MB, Carolla P, Barth H, Blum HE, Cosset FL, Koutsoudakis G, Bartenschlager R, Union A, Depla E, Owsianka A, Patel AH, Schuster C, Stoll-Keller F, Doffoel M, Dreux M, Bau-mert TF. Gastroenterology. 2008, 135(5):1719-1728). Данные разработки базируются на имеющихся сообщениях, что в организме больных, инфицированных HCV, образуются антитела, направленные против вируса. Эти антитела могут принадлежать к различным классам (подклассам) иммуноглобулинов и обладать различной антигенной специфичностью. Функциональные эффекты таких антител варьируют от простого связывания, не оказывающего влияния на вирус, до реальной нейтрализации и уничтожения вируса или предотвращения его размножения Piazza M, Sagliocca L, Tosone G, Guadagnino V, Stazi MA, Orlando R, Borgia G, Rosa D, Abrignani S, Palumbo F, Manzin A, Clementi M. Arch Intern Med. 1997, 157(14): 1537-1544). При этом элиминации вируса сопутствует быстрое (уже на ранней стадии инфекции) появление нейтрализующих антител широкого спектра действия. Напротив, антитела от пациентов, у которых наблюдается прогрессирование инфекции в хроническую фазу, нейтрализуют вирус лишь в небольшой степени, либо вообще не обладают нейтрализующей способностью (Pestka JM, Zeisel MB, Blaser E, Schurmann P, Bartosch B, Cosset FL, Patel AH, Meisel H, Baumert J, Viazov S, Rispeter K, Blum HE, RoggendorfM, Baumert TF. Proc Nati Acad Sci USA. 2007, 104(14):6025-6030). Можно предположить, что неэффективность антительного ответа против HCV обусловлена общим дисбалансом гуморального иммунного ответа (Di Lorenzo С., Angus A.G., Patel A.H. Vimses. 2011, 3(11):2280-2300).
Терапевтический потенциал антител в значительной степени зависит от их конкретных мишеней - эпитопов белков вируса, с которыми они способны реагировать. Хотя антитела против неструктурных белков HCV зарегистрированы у людей, больных гепатитом С, такого рода антитела обычно не оказывают существенного влияния на вирус. В то же время антитела, направленные против структурных белков, и прежде всего против белков оболочки Е1 и Е2, могут обладать нейтрализующей активностью, блокируя проникновение вируса в клетки или запуская механизмы опосредованной антителами клеточной цитотоксичности (ADCC) и комплемент-зависимого лизиса (CDL), приводящие к его уничтожению. Исходя из этого, можно ожидать, что одним из перспективных подходов к созданию новых препаратов для лечения гепатита С является получение и испытание человеческих моноклональных антител против консервативных участков структурных вирусных белков на основе природных антител, выработавшихся естественным путем у HCV-инфицированных больных (Clementi N, De Marco D, Mancini N, Solforosi L, Moreno GJ, Gubareva LV, Mishin V, Di Pietro A, Vicenzi E, Siccardi AG, Clementi M, Burioni R. PLoS One. 2011, 6(12):e28001. dot: 10.1371; Solforosi L, Mancini N, Canducci F, Clementi N, Sautto GA, Diotti RA, Clementi M, Burioni R. New Microbiol. 2012, 35(3):289-294). При этом можно ожидать, что моноклональные антитела, действуя непосредственно на вирус, будут снижать его уровень в крови и предотвращать повторную инфекцию новых клеток печени (Burioni R, Perotti M, Mancini N, Clementi M.J. Hepatol. 2008, 49(2):299-300). Кроме того, введение специфических моноклональных антител против HCV или смесей таких антител может восстановить баланс в пользу иммунной системы организма и воспрепятствовать распространению вируса.
В настоящее время известно, что несколько препаратов на основе моноклональных антител против белка Е2 проходят фазы I или II клинических испытаний. В частности, в клинических исследованиях фазы I человеческого моноклонального антитела XTL-002 было показано, что более чем у половины из 15 пациентов с гепатитом С, которым была проведена однократная внутривенная инфузия XTL-002, отмечалось значительное снижение вирусной нагрузки, величина которого составляла от 2 до 100 раз; при этом серьезных побочных эффектов у них отмечено не было (http://www.xtlbio.com/news/news_item.asp-id=26.htm). Два клинических исследования по предотвращению повторной HCV-инфекции у пациентов с хронической инфекцией во время и после трансплантации печени показали умеренное и кратковременное снижение виремии после инфузии человеческого антитела XTL68 против белка Е2 вируса HCV (Galun E, Terrault NA, Eren R, Zauberman A, Nussbaum O, Terkieltaub D, Zohar M, Buchnik R, Ackerman Z, Safadi R, Ashur Y, Misrachi S, Liberman Y, Rivkin L, Dagan S. J.Hepatol. 2007, 46(1):37-44; Schiano TD, Charlton M, Younossi Z, Galun E, Pruett T, Tur-Kaspa R, Eren R, Dagan S, Graham N, Williams PV, Andrews J. Liver Transpl. 2006, 12(9): 1381-1389). Еще одно клиническое исследование по предотвращению повторной инфекции трансплантата печени при помощи моноклональных антител начато в Страсбурге, Франция (Fafi-Kremer S, Fofana I, Soulier E, Carolla P, Meuleman P, Leroux-Roels G, Patel AH, Cosset FL, Pessaux P, Doffoel M, Wolf P, Stoll-Keller F, Baumert TF. J Exp Med. 2010 207(9):2019-2031).
С использованием трансгенных мышей было разработано гуманизированное моноклональное антитело MBL-HCV1, направленное против высококонсервативного линейного эпитопа оболочечного гликопротеина Е2 вируса HCV (аминокислоты 412-423) и способное in vitro нейтрализовать HCV различных генотипов (Broering TJ, Garrity KA, Boatright NK, Sloan SE, Sandor F, Thomas WD Jr, Szabo G, Finberg RW, Ambrosino DM, Babcock GJ. J. Virol. 2009, 83(23): 12473-12482). В исследовании по выбору диапазона доз, которое проводили на шимпанзе, однократная инъекция этого антитела в дозе 250 мг/кг предотвращала острую HCV-инфекцию (Morin TJ, Broering TJ, Leav BA, Blair BM, Rowley KJ, Boucher EN, Wang Y, Cheslock PS, Knauber M, Olsen DB, Ludmerer SW, Szabo G, Finberg RW, Purcell RH, Lan-ford RE, Ambrosino DM, Molrine DC, Babcock GJ. PLoS Pathog. 2012; 8(8):e1002895. doi: 10.1371).
Было предпринято рандомизированное исследование с целью изучения эффекта многократных инъекций MBL-HCV1 на элиминацию HCV у пациентов, перенесших трансплантацию печени. Завершенное в 2009 г. исследование I фазы, в котором участвовал 31 здоровый доброволец, показало, что антитело хорошо переносится и не вызывает серьезных побочных эффектов. Испытуемые, инфицированные вирусом HCV генотипа 1а, в исследовании I фазы получали 11 инфузий антитела в дозе 50 мг/кг/сут (до 14 дней после трансплантации печени). У испытуемых, получавших антитело, снижение вирусной нагрузки относительно исходного значения было заметно сильнее, чем у испытуемых, получавших плацебо. Кроме того, в группе с антителом срок восстановления вирусной нагрузки был значительно больше, чем в группе с плацебо (18,7 дней против 2,4 дней). Вместе с тем, как и в случае монотерапии другими препаратами для лечения HCV-инфекции, у пациентов, получавших антитело, возникали устойчивые варианты вируса. Это может быть обусловлено тем, что эпитоп для данного антитела расположен очень близко к гипервариабельному участку белка Е2 HCV (Chung RT, Gordon FD, Curry MP, Schiano TD, Emre S, Corey K, Markmann JF, Herd M, Pomposelli JJ, Pomfret EA, Florman S, Schilsky M, Broering TJ, Finberg RW, Szabo G, Zamore PD, Khettry U, Babcock GJ, Ambrosino DM, Leav B, Leney M, Smith HL, Molrine DC. American J. of Transpl. 2013, 13(4): 1047-1054).
Компания Crucell, Ltd. разработала программу комбинирования разных антител против вируса гепатита С. В настоящее время ею проводится оценка большой панели полностью человеческих моноклональных антител против вируса гепатита С, при этом особое внимание уделяется определенным участкам белка Е2 HCV (http://www.crucell.com/R_and_D-Clinical_Development-HepatitisC).
Человеческие моноклональные антитела против HCV получают, главным образом, на основе технологии рекомбинантных ДНК с использованием библиотек вариабельных доменов легких и тяжелых цепей человеческих иммуноглобулинов или при помощи гибридных клеточных культур, являющихся результатом слияния лимфоцитов крови инфицированного человека и подходящей «партнерской» линии клеток (Karpas A, Dremucheva A, Czepulkowski BH. Proc Nati Acad Sci USA. 2001, 98(4): 1799-1804). Альтернативным подходом может быть «очеловечивание» мышиных моноклональных антител против HCV.
Вместе с тем, проведенный анализ просмотренных литературных источников показал, что имеющиеся разработки не носят универсального характера и в настоящее время не позволяют получать достаточно надежные результаты по диагностике HCV и лечению вызываемого им заболевания.
Наиболее близкой по технической сущности и достигаемому эффекту является группа изобретений (US 20120039846, 2012), включающая в себя композиции для профилактики и лечения HCV-инфекции. В состав композиции входит одно или несколько человеческих моноклональных антител, направленных против конформационных эпитопов белка Е2 оболочки вируса гепатита С, а именно моноклональное антитело НС-11, секретируемое линией гибридомных клеток, хранящейся в коллекции АТСС под инвентарным номером РТА-9418, или его фрагмент, включающие CDR1-участок тяжелой цепи, содержащий последовательность GATFSSFI, CDR2-участок тяжелой цепи, содержащий последовательность IIPMFGTA, CDR3-участок тяжелой цепи, содержащий последовательность AMEVPGFCRGGSCSGYMDV, CDR1-участок легкой цепи, содержащий последовательность HSVSSSN, CDR2-участок легкой цепи, содержащий последовательность GAS и CDR3-участок легкой цепи, содержащий последовательность QQYGSSPIT; моноклональное антитело НС-1, секретируемое линией гибридомных клеток, хранящейся в коллекции АТСС под инвентарным номером РТА-9416, или его фрагмент, включающие CDR1-участок тяжелой цепи, содержащий последовательность GGTYNSEV, CDR2-участок тяжелой цепи, содержащий последовательность FIPMFGTA, CDR3-участок тяжелой цепи, содержащий последовательность AKVLQVGGNLVVRPL, CDR1-участок легкой цепи, содержащий последовательность QTISSTH, CDR2-участок легкой цепи, содержащий последовательность GVS и CDR3-участок легкой цепи, содержащий последовательность HQYGNSPQT; моноклональное антитело НС-3, секретируемое линией гибридомных клеток, хранящейся в коллекции АТСС под инвентарным номером РТА-9417, или его фрагмент, включающие CDR1-участок тяжелой цепи, содержащий последовательность GFSLSTTGVG, CDR2-участок тяжелой цепи, содержащий последовательность IYWDDDK, CDR3-участок тяжелой цепи, содержащий последовательность ALNSYRSGTILYRELELRGLFYI, CDR1-участок легкой цепи, содержащий последовательность QSISSW, CDR2-участок легкой цепи, содержащий последовательность ESS и CDR3-участок легкой цепи, содержащий последовательность QQYESSSWT; моноклональное антитело СВН-23, секретируемое линией гибридомных клеток, хранящейся в коллекции АТСС под инвентарным номером РТА-9419, или его фрагмент, включающие CDR1-участок тяжелой цепи, содержащий последовательность GGTFSSYA, CDR2-участок тяжелой цепи, содержащий последовательность IVPMFGTE, CDR3-участок тяжелой цепи, содержащий последовательность ARHENIYGTPFDY, CDR1-участок легкой цепи, содержащий последовательность HSITRY, CDR2-участок легкой цепи, содержащий последовательность AAS, и CDR3-участок легкой цепи, содержащий последовательность QQSYSTLLT.
Наряду с антителами в данную группу изобретений входят клеточные линии, их продуцирующие, а также фармацевтические композиции на их основе. Эпитопы перечисленных антител имеют конформационный характер. Авторами показано, что эпитоп для антитела НС-11 включает аминокислоты Gly530 и Asp535 белка Е2, контактирующие с антителом; эпитоп для антитела НС-1 включает контактирующие аминокислоты Trp529, Gly530 и Asp535; эпитоп для антитела НС-3 включает контактирующие аминокислоты Arg657, Asp658, Phe679, Leu692, Ile696, Asp698. (Здесь и далее нумерация аминокислот соответствует последовательности полипротеина вируса гепатита С изолята Н77 геротипа 1a - референсный номер NP_671491.1 в международной базе данных NCBI Protein).
Недостатком данной группы изобретений является то, что предлагаемые композиции не покрывают все эпитопы белка Е2, к которым возможно образование антител, нейтрализующих вирус, в связи с чем можно ожидать их недостаточную эффективность с точки зрения блокирования размножения вируса.
Задачей, решаемой в рамках заявляемой группы изобретений, являлось повышение эффективности нейтрализации вируса гепатита С. При решении данной задачи исходили из того, что к настоящему времени охарактеризован ряд эпитопов белка Е2 для человеческих моноклональных антител, обладающих нейтрализующей HCV активностью, а именно: линейные эпитопы с координатами 412-423 (W02006100449, 2006); 480-494, 613-621 (W02004087760, 2004); конформационные непрерывные эпитопы с координатами 396-424, 412-424, 436-447, 523-540 (US 20110311550, 2011); конформационные прерывистые эпитопы, содержащие аминокислоты Leu641, Thr648, Pro512, Leu580, Pro591, Arg588 (WO 2010035292, 2010), а также указанные выше эпитопы, представленные в изобретении (US 20120039846, 2012). Проведенный анализ показал, что некоторые участки аминокислотной последовательности белка Е2 не содержат описанных эпитопов. В частности, неизвестны антитела с эпитопами на участках 448-479, 494-511, 541-579, 592-612, 622-640. Можно было предполагать, что антитела к таким эпитопам будут обладать новыми свойствами и потенциально могут оказаться более эффективными для профилактики и лечения гепатита С.
Технической задачей являлось расширение круга антител, пригодных для воздействия на вирус гепатита С, путем создания человеческих моно-клональных антител к белку Е2 оболочки вируса, обладающих нейтрализующей активностью и направленных к новым, ранее не описанным эпитопам, и повышение надежности связывания вирусов гепатита С. При этом под термином «эпитоп» в тексте настоящего изобретения понимается та часть полипептидной молекулы белка Е2, которая распознается каким-либо из описываемых моноклональных антител и вступает с ним в непосредственный контакт. Эпитоп может представлять собой непрерывную аминокислотную последовательность, а может быть прерывистым, то есть составленным из ряда аминокислот, далеких друг от друга по порядковому номеру, но близкорасположенных в пространственной структуре белка.
Также различают линейные и конформационные эпитопы. В первом случае распознавание эпитопа антителом определяется только аминокислотной последовательностью эпитопа, во втором случае оно также зависит от конформационной структуры эпитопа в составе белковой молекулы.
Технический результат достигался созданием антител, способных связываться с эпитопами, далее обозначенными как Э1 и/или Э2 и/или Э3, которые включают в себя аминокислотные последовательности вариабельных участков их тяжелых (VH) и легких (VL) цепей:
последовательность участка VH антитела RYB1, SEQ ID NO:5;
последовательность участка VL антитела RYB1, SEQ ID NO:9;
последовательность участка VH антитела RYB2, SEQ ID NO:13;
последовательность участка VL антитела RYB2, SEQ ID NO:17;
последовательность участка VH антитела RYB3, SEQ ID NO:21;
последовательность участка VL антитела RYB3, SEQ ID NO:25.
При этом для ингибирования вируса используется, как правило, композиция, состоящая из трех антител, получивших наименования RYB 1, RYB2 и RYB3, в соотношении (% масс.) RYB1:RYB2:RYB3=20-40:20-40:20-40. Лучшие результаты достигались при соотношении ингредиентов композиции в соотношении 1:1:1. Снижение концентрации одного из ингредиентов до содержания в смеси менее 20% нежелательно в связи с возрастающей вероятностью понижения эффективности воздействия на вирус из-за вариабельности его генотипа, хотя определенный, но менее выраженный эффект достигается при использовании для блокирования размножения вируса в качестве одного из вариантов воздействия вышеупомянутых антител в меньшей концентрации или вообще индивидуально.
Установлено, что эпитопом антитела RYB1, в дальнейшем обозначенным как Э1, является определенная конформация последовательности аминокислот, заключенной в пределах участка HPEATYSRCG (589-598, SEQ ID NO:30) и содержащей аминокислоты Ser595 и Arg596. При этом наиболее значимым является центральный фрагмент этого участка из 6 аминокислот: EATYSR (591-596, SEQ ID NO:31). Данный эпитоп в литературе не описан.
Эпитопом антитела RYB2, в дальнейшем обозначенным как Э2, является определенная конформация последовательности аминокислот, заключенной в пределах участка VCGPVYCF (502-509, SEQ ID NO:32) и содержащей аминокислоту Tyr507. При этом наиболее значимым является центральный фрагмент этого участка из 6 аминокислот: CGPVYC (503-508, SEQ ID NO:33). Данный эпитоп в литературе не описан.
Эпитопом антитела RYB3, в дальнейшем обозначенным как Э3, является определенная конформация последовательности аминокислот, заключенной в пределах участка HPEATYSRCGSGPWITP (589-605, SEQ ID NO:34) и содержащей аминокислоты Ser595, Arg596 и Ser599. При этом наиболее значимым является один из внутренних фрагментов этого участка YSRCGS (594-599, SEQ ID NO:35) или SRCGSG (595-600, SEQ ID NO:36). Данный эпитоп в литературе не описан.
Указанные антитела были получены в результате создания гибридных клеточных линий (гибридом) BIONA-RYB1, BIONA-RYB2 и BIONA-RYB3, продуцирующих вышеуказанные антитела. Заявляемые гибридомы депонированы 17.07.2013 во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) под номерами, соответственно, Н-142, Н-143 и Н-144 и обладают следующими характеристиками.
Штамм BIONA-RYB1 (регистрационный номер Н-142)
Название антитела: RYB1.
Класс/суб. класс антитела: human IgG1 (kappa).
Иммуноген, используемый для получения антитела: вирус гепатита С (естественное инфицирование).
Специфичность антитела: белок Е2 оболочки вируса гепатита С.
Известная перекрестная реакция: все генотипы вируса гепатита С.
Возраст штамма: 12 мес.
Место происхождения: г.Москва, ООО «БионА Фарма», штамм родословной не имеет и ранее не депонировался
Партнеры для гибридизации клеток: мононуклеарные клетки, выделенные из селезенки пациента, умершего от гепатита С, и принадлежащая ООО «БионА Фарма» гибридная (мышь/человек) миеломная клеточная линия BIONA-X.
Культуральные свойства штамма, маркерные признаки: суспензионная культура лимфоцитоподобных клеток, маркеры штамма не определяли.
Рекомендуемые условия для замораживания. Клетки штамма осаждают центрифугированием 15 мин при 200 g, ресуспендируют в сыворотке плода коровы, содержащей 10% диметилсульфоксида, до концентрации 3×106 клеток/мл, разливают в пластиковые ампулы для криоконсервирования (COSTAR-CORNING) объемом 2 мл и помещают в контейнер STRATAGENE для медленного замораживания, предварительно охлажденный до 4°С. Контейнер выдерживают в низкотемпературном холодильнике при -70°С 24 часа, после чего ампулы с клетками переносят в жидкий азот. Возможно программное замораживание со снижением температуры на 1°С в минуту до -4°С с переносом в жидкий азот. Размораживание быстрое, при 37°С. Содержимое ампулы после размораживания переносят в 10 мл бессывороточной среды DMEM, осаждают центрифугированием, ресуспендируют в 5 мл среды для культивирования и переносят в культуральный флакон. Выживаемость клеток, определяемая по включению трипанового синего, составляет более 80%.
Рекомендуемые условия для культивирования. Среда DMEM с добавлением 10% сыворотки плода коровы, 4 мМ L-глутамина, 1 мМ пирувата-Na, 100 IU/мл пенициллина, 100 мкг/мл стрептомицина и концентрата аминокислот и витаминов для базальной среды Игла. Для выращивания штамма используют стандартные культуральные флаконы. Во флакон площадью 25 см2 в 5 мл среды засевают 1×106 клеток. Культивирование проводят при 37°С в атмосфере 5,6% CO2. Пассаж 1:5 производят при достижении плотности культуры 1×106 клеток/мл среды путем замены 4 мл клеточной суспензии на 4 мл свежей культуральной среды.
Контаминация: бактерии, грибы и микоплазма в культуре при длительном наблюдении визуально не обнаружены.
Производимый продукт: моноклональное полностью
человеческое антитело RYB1 класса IgG1 (kappa), специфичное к белку Е2 оболочки вируса гепатита С.
Область применения штамма: создание терапевтического средства для лечения гепатита С.
Активность (продуктивность) штамма (с указанием условий культивирования), а также другие производственные показатели: при культивировании в стандартных условиях в течение 3 сут концентрация IgG1 человека в культуральной среде достигает значения 20 мкг/мл.
Способ определения активности штамма с указанием метода: измерение IgG1 человека в культуральной кондиционированной среде методом количественного иммуноферментного анализа (ELISA) при помощи соответствующего набора реагентов (например, ELH-IGG 1-001 Human IgG1 ELISA kit, Raybiotech, США).
Штамм BIONA-RYB2 (регистрационный номер Н-143)
Название антитела: RYB2.
Класс/суб. класс антитела: human IgG1 (kappa).
Иммуноген, используемый для получения антитела: вирус гепатита С (естественное инфицирование).
Специфичность антитела: белок Е2 оболочки вируса гепатита С.
Известная перекрестная реакция: все генотипы вируса гепатита С.
Возраст штамма: 12 мес.
Место происхождения: г. Москва, ООО «БионА Фарма», штамм родословной не имеет и ранее не депонировался.
Партнеры для гибридизации клеток: мононуклеарные клетки, выделенные из селезенки пациента, умершего от гепатита С, и принадлежащая ООО «БионА Фарма» гибридная (мышь/человек) миеломная клеточная линия BIONA-X.
Культуральные свойства штамма, маркерные признаки: суспензионная культура лимфоцитоподобных клеток, маркеры штамма не определяли.
Рекомендуемые условия для замораживания. Клетки штамма осаждают центрифугированием 15 мин при 200 g, ресуспендируют в сыворотке плода коровы, содержащей 10% диметилсульфоксида, до концентрации 3×106 клеток/мл, разливают в пластиковые ампулы для криоконсервирования (COSTAR-CORNING) объемом 2 мл и помещают в контейнер STRATAGENE для медленного замораживания, предварительно охлажденный до 4°С. Контейнер выдерживают в низкотемпературном холодильнике при -70°С 24 часа, после чего ампулы с клетками переносят в жидкий азот. Возможно программное замораживание со снижением температуры на 1°С в минуту до -4°С с переносом в жидкий азот.Размораживание быстрое, при 37°С. Содержимое ампулы после размораживания переносят в 10 мл бессывороточной среды DMEM, осаждают центрифугированием, ресуспендируют в 5 мл среды для культивирования и переносят в культуральный флакон. Выживаемость клеток, определяемая по включению трипанового синего, составляет более 85%.
Рекомендуемые условия для культивирования. Среда DMEM с добавлением 10% сыворотки плода коровы, 4 мМ L-глутамина, 1 мМ пирувата-Na, 100 IU/мл пенициллина, 100 мкг/мл стрептомицина и концентрата аминокислот и витаминов для базальной среды Игла. Для выращивания штамма используют стандартные культуральные флаконы. Во флакон площадью 25 см2 в 5 мл среды засевают 1×106 клеток. Культивирование проводят при 37°С в атмосфере 5,6% СО2. Пассаж 1:5 производят при достижении плотности культуры 1×106 клеток/мл среды путем замены 4 мл клеточной суспензии на 4 мл свежей культуральной среды.
Контаминация: бактерии, грибы и микоплазма в культуре при длительном наблюдении визуально не обнаружены.
Производимый продукт: моноклональное полностью человеческое антитело RYB2 класса IgG1 (kappa), специфичное к белку Е2 оболочки вируса гепатита С.
Область применения штамма: создание терапевтического средства для лечения гепатита С.
Активность (продуктивность) штамма (с указанием условий культивирования), а также другие производственные показатели: при культивировании в стандартных условиях в течение 3 сут. концентрация IgG1 человека в культуральной среде достигает значения 25 мкг/мл.
Способ определения активности штамма с указанием метода: измерение IgG1 человека в культуральной кондиционированной среде методом количественного иммуноферментного анализа (ELISA) при помощи соответствующего набора реагентов (например, ELH-IGG1-001 Human IgG1 ELISA kit, Raybiotech, США).
Штамм BIONA-RYB3 (регистрационный номер Н-144)
Название антитела: RYB3.
Класс/суб. класс антитела: human IgG1 (kappa).
Иммуноген, используемый для получения антитела: вирус гепатита С (естественное инфицирование).
Специфичность антитела: белок Е2 оболочки вируса гепатита С.
Известная перекрестная реакция: все генотипы вируса гепатита С.
Возраст штамма: 12 мес.
Место происхождения: г. Москва, ООО «БионА Фарма», штамм родословной не имеет и ранее не депонировался.
Партнеры для гибридизации клеток: мононуклеарные клетки, выделенные из селезенки пациента, умершего от гепатита С, и принадлежащая ООО «БионА Фарма» гибридная (мышь/человек) миеломная клеточная линия BIONA-X.
Культуральные свойства штамма, маркерные признаки: полусуспензионная культура лимфоцитоподобных клеток, маркеры штамма не определяли. Рекомендуемые условия для замораживания. Клетки штамма осаждают центрифугированием 15 мин при 200 g, ресуспендируют в сыворотке плода коровы, содержащей 10% диметилсульфоксида, до концентрации 3×106 клеток/мл, разливают в пластиковые ампулы для криоконсервирования (COSTAR-CORNING) объемом 2 мл и помещают в контейнер STRATAGENE для медленного замораживания, предварительно охлажденный до 4°С. Контейнер выдерживают в низкотемпературном холодильнике при -70°С 24 часа, после чего ампулы с клетками переносят в жидкий азот. Возможно программное замораживание со снижением температуры на 1°С в минуту до -4°С с переносом в жидкий азот. Размораживание быстрое, при 37°С. Содержимое ампулы после размораживания переносят в 10 мл бессывороточной среды DMEM, осаждают центрифугированием, ресуспендируют в 5 мл среды для культивирования и переносят в культуральный флакон. Выживаемость клеток, определяемая по включению трипанового синего, составляет более 70%.
Рекомендуемые условия для культивирования. Среда DMEM с добавлением 10% сыворотки плода коровы, 4 мМ L-глутамина, 1 мМ пирувата-Na, 100 IU/мл пенициллина, 100 мкг/мл стрептомицина и концентрата аминокислот и витаминов для базальной среды Игла. Для выращивания штамма используют стандартные культуральные флаконы. Во флакон площадью 25 см2 в 5 мл среды засевают 1×106 клеток. Культивирование проводят при 37°С в атмосфере 5,6% СО2. Пассаж 1:5 производят при достижении плотности культуры 1×106 клеток/мл среды путем переноса среды с плавающими клетками в стерильную пробирку, снятия прикрепленных клеток трипсином, объединения с плавающими клетками и отбора пятой части объединенной клеточной суспензии для нового посева.
Контаминация: бактерии, грибы и микоплазма в культуре при длительном наблюдении визуально не обнаружены.
Производимый продукт: моноклональное полностью человеческое антитело RYB3 класса IgG1 (kappa), специфичное к белку Е2 оболочки вируса гепатита С.
Область применения штамма: создание терапевтического средства для лечения гепатита С.
Активность (продуктивность) штамма (с указанием условий культивирования), а также другие производственные показатели: при культивировании в стандартных условиях в течение 3 сут концентрация IgG1 человека в культуральной среде достигает значения 30 мкг/мл.
Способ определения активности штамма с указанием метода: измерение IgG1 человека в культуральной кондиционированной среде методом количественного иммуноферментного анализа (ELISA) при помощи соответствующего набора реагентов (например, ELH-IGG1-001 Human IgG1 ELISA kit, Raybiotech, США).
Существо заявляемых изобретений иллюстрируется следующими графическими материалами.
На фиг.1 показана структура генома вируса гепатита С. В прямоугольную рамку заключена область, кодирующая полипротеин. Шпилечные участки соответствуют 5'NTR- и 3'NTR-нетранслируемым областям, включающим сайт внутреннего связывания рибосом (IRES). Обозначены участки генома, кодирующие белковые продукты, образующиеся при расщеплении полипротеина клеточными и вирусными пептидазами. Указаны известные функции каждого белкового продукта. Координаты кодирующих участков вирусного генома для вируса изолята Н77 (генотип 1а): нуклеотиды 342-914 - структурный белок Core, нуклеотиды 342-828 - белок gp2 (образуется в результате сдвига рамки трансляции), нуклеотиды 915-1490 - структурный белок Е1, нуклеотиды 1491-2579 - структурный белок Е2, нуклеотиды 2580-2768 - белок р7 (локализуется на мембране), нуклеотиды 2769-3419 -белок NS2 (мембранный белок, ингибитор CIDE-B индуцированного апоптоза), нуклеотиды 3420-5312 - белок NS3 (протеаза/хеликаза), нуклеотиды 5313-5474 - белок NS4A (кофактор для протеазы NS3), нуклеотиды 5475-6257 - белок NS4B (мембранный белок-регулятор репликационного комплекса NS3-NS5B), нуклеотиды 6258-7601 - белок NS5A (способен к фосфорилированию, вероятно, опосредует чувствительность к интерферону), нуклеотиды 7602-93 74-белок NS5B (РНК-зависимая РНК-полимераза). Участки генома 1-341 и 9378-9646 являются, соответственно, 5'NTR - и 3'NTR-некодирующими.
На фиг.2 показаны результаты цитофлуориметрического анализа клеток HEK-Е1Е2 (темная гистограмма) после инкубации с кондиционированной средой одного из гибридомных клонов. Серая кривая - гистограмма иммунофлуоресценции контрольных нетрансфицированных клеток НЕК293 после инкубации с той же кондиционированной средой.
На фиг.3 приведена электрофореграмма образцов очищенных антител RYB1, RYB2 и RYB3 (по 7,5 мкг на дорожку), разделенных в 7,5%-ном полиакриламидном геле без меркаптоэтанола (3А) или в 10%-ном полиакриламидном геле с меркаптоэтанолом (3В) в присутствии додецилсульфата натрия и окрашенных Coomassie Brilliant Blue R-250.
На фиг.4 показана взаимная конкуренция антител RYB1, RYB2 и RYB3 за связывание с клетками HEK-Е1Е2, экспрессирующими белковый комплекс Е1-Е2 HCV. На графиках представлены средние значения флуоресценции клеток после проведения реакции связывания со смесями FITC-меченых и немеченых антител в указанных соотношениях. Неспецифическое связывание измерено на клетках HEK293.
На фиг.5 приведены масс-спектры комплексов рекомбинантного белка Е2 с антителами RYB1, RYB2 и RYB3, где А: чистый белок Е2; B-D: препараты антител, соответственно, RYB1, RYB2 и RYB3; E-G: комплексы антител с белком Е2.
На фиг.6 продемонстрировано расположение молекул сшивающего реагента на пептидных продуктах протеолиза белка Е2 в окрестности эпитопа для антитела RYB1 до и после образования комплексов с антителом. Темные прямоугольники соответствуют пептидам, присутствующим среди продуктов протеолиза как чистого белка Е2, так и его комплексов с антителом RYB1. Темной звездочкой отмечены аминокислоты, связанные со сшивающим реагентом. Светлые прямоугольники соответствуют пептидам, присутствующим среди продуктов протеолиза чистого белка Е2, но отсутствующим после образования комплекса с антителом. Светлой звездочкой отмечены аминокислоты, связанные со сшивающим реагентом в случае протеолиза чистого белка Е2. Вдоль горизонтальной оси приведена последовательность аминокислот белка Е2 на рассматриваемом участке.
На фиг.7 показано расположение молекул сшивающего реагента на пептидных продуктах протеолиза белка Е2 в окрестности эпитопа для антитела RYB2 до и после образования комплексов с антителом. Темные прямоугольники соответствуют пептидам, присутствующим среди продуктов протеолиза как чистого белка Е2, так и его комплексов с антителом RYB2. Темной звездочкой отмечены аминокислоты, связанные со сшивающим реагентом. Светлые прямоугольники соответствуют пептидам, присутствующим среди продуктов протеолиза чистого белка Е2, но отсутствующим после образования комплекса с антителом. Светлой звездочкой отмечены аминокислоты, связанные со сшивающим реагентом в случае протеолиза чистого белка Е2. Вдоль горизонтальной оси приведена последовательность аминокислот белка Е2 на рассматриваемом участке.
На фиг.8 показано расположение молекул сшивающего реагента на пептидных продуктах протеолиза белка Е2 в окрестности эпитопа для антитела RYB3 до и после образования комплексов с антителом. Темные прямоугольники соответствуют пептидам, присутствующим среди продуктов протеолиза как чистого белка Е2, так и его комплексов с антителом RYB3. Темной звездочкой отмечены аминокислоты, связанные со сшивающим реагентом. Светлые прямоугольники соответствуют пептидам, присутствующим среди продуктов протеолиза чистого белка Е2, но отсутствующим после образования комплекса с антителом. Светлой звездочкой отмечены аминокислоты, связанные со сшивающим реагентом в случае протеолиза чистого белка Е2. Вдоль горизонтальной оси приведена последовательность аминокислот белка Е2 на рассматриваемом участке.
На фиг.9 приведены масс-спектры комплексов рекомбинантного белка Е2 с антителами RYB1, RYB2 и RYB3 в присутствии избытка пептидных продуктов протеолиза белка Е2.
Сущность и промышленная применимость заявляемых изобретений иллюстрируются следующими примерами.
Пример 1. Получение клонов гибридных клеток (гибридом).
Для получения гибридомных клеток-продуцентов природных полностью человеческих моноклональных антител против белков оболочки вируса гепатита С использовали клинические материалы от семи пациентов, умерших от гепатита С. У всех этих пациентов первичный диагноз был поставлен на основании присутствия у них антител против антигенов HCV. Антитела определяли коммерчески доступным методом твердофазного иммуноферментного анализа (ELISA). Никаких критериев исключения, связанных с возрастом, расой или стадией заболевания на момент смерти, не применяли. Сопутствующие инфекции, такие как ВИЧ, простой герпес, вирус папилломы человека и венерические заболевания, допускались. Наличие в анамнезе наркомании, злокачественного новообразования печени и других заболеваний также допускалось. Единственными критериями исключения были септический шок и любое аутоиммунное заболевание, в том числе волчанка, сахарный диабет и др. Все данные, по которым можно идентифицировать умершего пациента, были удалены; клиническая информация включала только сведения о возрасте и поле и историю болезни.
Селезенку иссекали и помещали в чашку Петри диаметром 100 мм со стерильной средой RPMI 1640, дополнительно содержащей 4 мМ L-глутамин, заменимые аминокислоты (из 100-кратного концентрата), витамины (из 100-кратного концентрата), 1 мМ пируват натрия и 50 мкг/мл гентамицин. Кусочки селезенки разделяли при помощи хирургических щипцов и ножниц. Разделенную ткань пропускали через металлическое сито (ячейки 50 меш) с использованием стеклянного пестика. По 10 мл полученной суспензии переносили в стерильные конические пробирки объемом 15 мл, содержащие 5 мл среды Histopaque 1.077 для разделения лимфоцитов (Sigma-Aldrich, США) и центрифугировали 20 мин при 400 g. Образовавшееся на границе между слоями мутное кольцо мононуклеарных клеток отбирали при помощи пастеровской пипетки и разводили в 10 раз стандартной бессывороточной средой RPMI 1640. Клетки центрифугировали 10 мин при 300g и дважды промывали средой.
Клетки BIONA-X, представляющие собой специализированную гибридную (мышь/человек) миеломную клеточную линию, созданную ООО «БионА Фарма» для получения человеческих гибридом, выращивали в среде RPMI 1640 без антибиотиков, содержащей 10%-ную фетальную телячью сыворотку (Hyclone), 4 мМ L-глутамин, 1 мМ пируват натрия, заменимые аминокислоты и витамины (полная среда). Перед слиянием клетки культивировали в присутствии 20 мкг/мл 8-азагуанина (Sigma-Aldrich, США) во избежание реверсии НАТ-чувствительных клеток к дикому типу. Клетки в экспоненциальной фазе роста доводили до плотности 10% от монослоя.
Клетки линии BIONA-X и мононуклеарные клетки селезенки промывали 3 раза в бессывороточной среде RPMI 1640 при помощи центрифугирования 5 мин при 300 g, смешивали друг с другом в отношении 1:5 (BIONA-X: клетки селезенки) и центрифугировали 10 минут при 300 g. Супернатант удаляли, клеточный осадок ресуспендировали в 100-300 мкл (в зависимости от объема клеток) среды DMEM, к смеси клеток добавляли 100-300 мкл раствора полиэтиленгликоль-1500/диметилсульфоксид (1:1), нагретого до комнатной температуры, а затем в течение 3 минут встряхивали пробирку, слегка постукивая по ней. После этого в пробирку добавляли 15 мл смеси (1:1) сбалансированного солевого раствора Хэнкса и фосфатно-солевого буфера (PBS). Добавление осуществляли следующим образом: 10 мл медленно в течение 10 минут, затем 5 мл в течение 5 минут. Затем добавляли 10 мл полной среды в течение 5 минут и, наконец, еще 5 мл полной среды в течение 1 минуты. Общий объем составлял 30 мл. Затем в пробирку добавляли 600 мкл 50-кратного концентрата раствора НТ (гипоксантин-тимидин) (Cellgro, США) и 20-30 мкл диметилсульфоксида. Суспензию клеток перемешивали в пробирке, переносили в чашку Петри (100×15 мм) и инкубировали при 37°С в CO2-инкубаторе в течение ночи. После этого клетки собирали, центрифугировали при 300 g в течение 10 минут и ресуспендировали в полной среде, к которой был добавлен раствор HAT (гипоксантин-аминоптерин-тимидин) из 50-кратного концентрата (Cellgro, США).
Клетки после слияния высевали в 96-луночные планшеты в объеме 200 мкл (примерно по 250000 клеток на лунку) в ростовой среде RPMI 1640 без антибиотиков, содержащей 10%-ную фетальную телячью сыворотку (Hyclone), 4 мМ L-глутамин, 1 мМ пируват натрия, заменимые аминокислоты и витамины. Суммарно было засеяно 65 планшетов. Два раза в неделю 50% среды отбирали и заменяли ее на свежую. Первую неделю клоны клеток культивировали с добавлением к ростовой среде раствора HAT, следующие две недели - с добавлением раствора НТ, после чего проводили тестирование на секрецию иммуноглобулинов человека.
Тестирование гибридомных клонов на секрецию человеческих иммуноглобулинов класса IgG проводили следующим образом. Планшеты для твердофазного иммуноферментного анализа покрывали поликлональными козьими антителами против человеческих иммуноглобулинов класса IgG, специфичными к Fc-фрагментам (Sigma-Aldrich, США). Для этого антитела вносили в лунки планшетов в 100 мкл карбонатного буфера (0,1 М карбонат натрия, рН 9.0) в количестве 100 нг на лунку. Планшеты закрывали герметичными крышками и инкубировали в течение ночи при 4°С. Затем планшеты дважды промывали раствором PBS, остающиеся капли удаляли, после чего в лунки добавляли по 200 мкл блокирующего раствора (0,4% сухого обезжиренного молока в PBS) и инкубировали 2 ч. По окончании инкубации лунки 3 раза промывали PBS, а затем в каждую лунку вносили по 50 мкл гибридомного супернатанта, разведенного блокирующим раствором в отношении 1:1. Отрицательным контролем служила полная среда для культивирования клеток. Положительным контролем служила человеческая сыворотка в разведении 1:2000.
Планшеты инкубировали в течение 2 часов при комнатной температуре, промывали 4 раза PBS и в лунки вносили меченые пероксидазой хрена поликлональные козьи антитела против человеческих иммуноглобулинов класса IgG (Sigma-Aldrich, США), разведенные в блокирующем растворе в отношении 1:2000. После 1 ч инкубации при комнатной температуре планшеты 4 раза промывали PBS и в каждую лунку добавляли субстрат пероксидазы (ортофенилендиамин в фосфат-цитратном буфере с перекисью водорода). Цветную реакцию останавливали добавлением 20 мкл 10%-ной соляной кислоты. Колориметрические измерения проводили в планшет-ридере Infinite F50 (Тесаn, Австрия) при 492 нм.
Положительным результатом тестирования считалось значение оптической плотности, по крайней мере в 3 раза превышающее уровень оптической плотности в лунках с отрицательным контролем. Из 6240 первичных клонов гибридом 100% показали положительный результат, и, следовательно, секретировали в культуральную среду человеческие иммуноглобулины класса IgG.
Пример 2. Получение клеток НЕК293, стабильно трансфицированных генами Е1-Е2 HCV.
Нуклеотидную последовательность, кодирующую полипротеин вируса гепатита С изолята Н77 (генотип 1а), извлекали из международной базы данных NCBI Nucleotide (http://www.ncbi.nlm.nih.gov/nuccore). Последовательность имеет референсный номер NC_004102.1. Для получения генетической конструкции, кодирующей гены Е1-Е2 HCV, использовали фрагмент этой последовательности (нуклеотиды 735-2579, SEQ ID NO:1), кодирующий аминокислоты 132-746 полипротеина. К 5 В-концу фрагмента присоединяли последовательность GCAGGTACCGCCGCCGCCATGAATTCC (SEQ ID NO:2), содержащую сайт узнавания рестриктазы Acc65I, участок связывания рибосом и кодирующую аминокислоты Met, Asn и Ser. К 3 В-концу фрагмента присоединяли последовательность TAATCTAGAGCG (SEQ ID NO:3), кодирующую сигнал терминации трансляции и содержащую сайт узнавания рестриктазы XbaI. В результате получали нуклеотидную последовательность, содержащую участок связывания рибосом и кодирующую аминокислоты Met, Asn, Ser, фрагмент из 60 С-концевых аминокислот белка Core (аминокислоты 132-191 полипротеина HCV), белок Е1 (аминокислоты 192-383 полипротеина) и белок Е2 (аминокислоты 384-746 полипротеина). Фрагмент двухцепочечной ДНК с указанной нуклеотидной последовательностью (SEQ ID No:4) синтезировали химическим путем, расщепляли рестриктазами Асс651 и XbaI и встраивали в эукариотический экспрессионный вектор pcDNA3.1-neo (Invitrogen, США) по сайтам этих же рестриктаз с образованием плазмиды pcDNA3.1-E1E2.
В зараженных вирусом гепатита С клетках белки Core, E1 и Е2 образуются из полипротеина HCV путем его расщепления клеточными сигнальными пептидазами и транслоцируются в эндоплазматический ретикулум. Правильность фолдинга белков Е1 и Е2 и образования комплекса между ними в большой степени зависит от их совместной транслокации и процессинга (Cocquerel L, Meunier JC, Op de Beeck A, Bonte D, Wychowski C, Dubuisson J. J. en. Virology. 2001, 82(7):1629-1635). Поэтому можно было предположить, что кодируемый плазмидой pcDNA3.1-E1E2 фрагмент полипротеина, содержащий С-концевую часть Core, E1 и Е2, будет расщепляться в трансфицированных плазмидой клетках подобным же образом. При этом белки Е1 и Е2 могут сформировать друг с другом гетеромерные комплексы, сходные по структуре с белковыми комплексами оболочки HCV. Трансмембранные домены белков Е1 и Е2 удерживают комплексы в эндоплазматическом ретикулуме (Flint, M., and J. A. McKeating J. Gen. Virol. 1999, 80 (8): 1943-1947), что приводит к необходимости пермеабилизации клеток перед проведением флуориметрического иммуноанализа.
Клетки HEK293 почки эмбриона человека культивировали в среде DMEM, дополнительно содержащей 10%-ную сыворотку плода коровы (Invitrogen, США), 2,2 мМ L-глутамин, 100 ед./мл пенициллин и 100 мг/мл стрептомицин, при 37°С в CO2-инкубаторе в присутствии 5% CO2. Для проведения трансфекции клетки сеяли в лунки 24-луночной плашки (50000 клеток на лунку). На следующий день клетки трансфицировали плазмидой pcDNA3.1-E1E2 с использованием реагента липофектамин 2000 (Invitrogen, США) в соотношении 1 мкг плазмидной ДНК и 1 мкл реагента на лунку в соответствии с протоколом производителя. Через 24 часа культуральную среду меняли на свежую и инкубировали клетки еще сутки. После этого клетки снимали с подложки трипсином, готовили серийные разведения клеточной суспензии и высевали в лунки 96-луночных плашек. Для проведения селекции трансфицированных клонов к культуральной среде добавляли антибиотик G418 (Invitrogen, США) до конечной концентрации 1 мг/мл. Через 2-3 недели клетки из лунок, содержащих не более одного клеточного клона, переносили в лунки 24-луночной плашки, подращивали, переносили в культуральные флаконы и далее замораживали.
Клетки трансфицированных клонов подвергали тестированию на экспрессию белковых комплексов Е1-Е2. Для этого клетки клонов выращивали во флаконах Т75 (Coming-Costar, США) до состояния свежего монослоя (5-6 млн клеток). Флаконы с клетками ополаскивали 5 мл холодного PBS, добавляли 5 мл холодного раствора 20 мМ EDTA, приготовленного на PBS, и инкубировали на льду 20 минут. Затем флаконы встряхивали, убеждались, что все клетки открепились, добавляли 5 мл PBS и собирали клеточный осадок центрифугированием 15 мин при 200 g.
Для пермеабилизации клеток осадок промывали два раза в 10 мл PBS и ресуспендировали в 1 мл метанола, предварительно охлажденного до -20°С. К суспензии добавляли еще 9 мл охлажденного метанола и инкубировали при -20°С 20 мин, периодически взбалтывая. По окончании инкубации клетки осаждали центрифугированием при 400 g, промывали PBS, ресуспендировали в 10 мл PBS и разносили в эппендофовские пробирки для тестирования кондиционированных сред гибридомных клонов, по 105 клеток на пробирку.
К суспензии пермеабилизованных клеток добавляли 1,5 мл раствора 0,3%-ного бычьего сывороточного альбумина (BSA) в PBS, инкубировали 5 мин, клетки осаждали центрифугированием и ресуспендировали в 20 мкл раствора 20 мкг/мл мышиных моноклональных антител BDI167 против антигена Е2 HCV (Abeam, США), приготовленного на PBS с 0,3% BSA. Клетки инкубировали 30 мин при комнатной температуре, затем отмывали в 1,5 мл раствора 0,3% BSA в PBS, инкубировали 30 мин при комнатной температуре с 20 мкл ПТС-меченых аффинно-очищенных антител кролика к иммуноглобулинам мыши (Becton Dickinson, США) в концентрации 2 мкг/мл, отмывали в 1,5 мл раствора 0,3% BSA в PBS и суспендировали в 400 мкл 1%-ного формальдегида в PBS. Флуоресценцию клеток анализировали на проточном цитофлуориметре FACS-Calibur (BD Biosciences, США). Клон клеток, показавший наиболее высокий уровень экспрессии комплекса Е1-Е2, был отобран для дальнейшего использования и назван HEK-Е1Е2.
Пример 3. Отбор гибридомных клонов, продуцирующих человеческие моноклональные антитела к белкам оболочки HCV.
Для отбора гибридом, производящих человеческие антитела, способные распознавать белковый комплекс Е1-Е2, использовали клетки HEK-Е1Е2, получение которых описано в примере 2. Контрольными клетками служили нетрансфицированные клетки родительской культуры HEK293. Клетки выращивали и пермеабилизовывали, как описано в примере 2.
Тестирование гибридомных кондиционированных сред проводили следующим образом. К суспензии пермеабилизованных клеток HEK-Е1Е2 или HEK293 добавляли 1,5 мл 0,3% BSA в PBS, инкубировали 5 мин, клетки осаждали центрифугированием и ресуспендировали в 20 мкл образца кондиционированной среды. Клетки инкубировали 30 мин при комнатной температуре, затем отмывали в 1,5 мл раствора 0,3% BSA в PBS, инкубировали 30 мин при комнатной температуре с 20 мкл FITC-меченых аффинно-очищенных антител кролика к иммуноглобулинам человека (OOO фирмы «ИМТЕК», Москва) в концентрации 3,3 мкг/мл, отмывали в 1,5 мл раствора 0,3% BSA в PBS и суспендировали в 400 мкл 1%-ного формальдегида в PBS. Флуоресценцию клеток анализировали на проточном цитофлуориметре FACS-Calibur (BD Biosciences, США). Положительным результатом анализа считалось среднее значение флуоресценции клеток HEK-Е1Е2, по крайней мере в 3 раза превышающее среднее значение флуоресценции для контрольных клеток НЕК293, инкубированных с тем же образцом кондиционированной среды.
На фиг.2 приведен типичный результат цитофлуориметрического анализа, в котором выявлено присутствие в кондиционированной среде человеческих антител, распознающих белковый комплекс Е1-Е2 в трансфицированных клетках. Из 6240 проанализированных гибридомных клонов 56 показали положительный результат в тесте на связывание с клетками HEK-Е1Е2 и были отобраны для повторного клонирования. После нескольких раундов клонирования большинство клеточных линий потеряло способность секретировать антитела, распознающие комплекс Е1-Е2. В конечном счете были получены три линии клеток гибридом, стабильно продуцирующих человеческие антитела против белкового комплекса Е1-Е2. Этим линиям гибридом были присвоены названия BIONA-RYB1, BIONA-RYB2 и BIONA-RYB3, а производимым ими антителам, соответственно, RYB1, RYB2HRYB3.
Пример 4. Получение очищенного препарата человеческих моноклональных антител RYB1.
Клетки линии гибридомы BIONA-RYB1 культивировали в 8 картриджах биореакторов холлоуфайберного типа «FiberCell Duet Pump» (FiberCell Systems Inc., США) в бессывороточной среде «HyClone SFM4MAb» (HyClone, США). Для первоначальной загрузки использовали 5·108 клеток на картридж. Через 4 недели после начала культивирования производили первый сбор кондиционированной среды, содержащей моноклональные антитела. Объем сбора составлял 50 мл с одного картрижда, при этом концентрация антител в среде варьировала от 0.01 мг/мл до 0.05 мг/мл. Кондиционированную среду собирали в течение 3 месяцев с интервалом в 6 суток, так что полный объем собранной среды составил 6,0 л. Партии среды замораживали и хранили при -20°С.
Для проведения хроматографической очистки антител кондиционированную среду размораживали и концентрировали на установке Amicon с использованием MWCO 100 kDa мембраны (Millipore, США). Перед кон-ентрированием установку и мембрану промывали стерильной апирогенной водой для устранения эндотоксинов. Из 6 литров среды получали 800 мл концентрата, содержащего 90 мг человеческих антител. Концентрат разбавляли равным объемом буфера для связывания и наносили на колонку с 2 мл сорбента «protein A/G plus agarose» (Pierce, США) в течение 3 ч. Колонку промывали 20 мл буфера для связывания, антитела элюировали 10 мл буфера для элюции и собирали фракции по 0.5 мл в пробирки с нейтрализующим буфером (100 мкл 1 М Tris-HCl, pH 8.0). Фракции, содержащие белок, объединяли и диализовали против PBS.
Препарат очищенных антител RYB1 (76 мг) анализировали при помощи электрофореза в полиакриламидном геле в присутствии додецил-сульфата натрия. Анализ показал (фиг.3.), что посторонние белковые примеси в полученном препарате практически отсутствуют.
Пример 5. Получение очищенного препарата человеческих моноклональных антител RYB2.
Моноклональные антитела RYB2 получали по технологии примера 4 с использованием гибридомы BIONA-RYB2. Полный объем собранной среды для гибридомы BIONA-RYB2 составил 5,8 л, выход очищенных антител RYB2 - 81 мг. Проведенный анализ показал (фиг.3), что посторонние белковые примеси в полученном препарате практически отсутствуют.
Пример 6. Получение очищенного препарата человеческих моноклональных антител RYB3.
Моноклональные антитела RYB3 получали по технологии примера 4 с использованием гибридомы BIONA-RYB3. Полный объем собранной среды для гибридомы BIONA-RYB3 составил 6,1 л, выход очищенных антител RYB3 - 85 мг. Проведенный анализ показал (фиг.3.), что посторонние белковые примеси в полученном препарате практически отсутствуют
Пример 7. Определение подклассов тяжелых и легких цепей антител.
Подклассы тяжелых цепей антител RYB1, RYB2 и RYB3 определяли с использованием набора «Human IgG Subclass Profile Kit» (Invitrogen, США). Мышиные Моноклональные антитела против человеческих иммуноглобулинов подклассов IgG1, IgG2, IgG3, IgG4 вносили в лунки иммунологического планшета согласно инструкции фирмы-производителя, затем вносили разбавленные препараты очищенных антител RYB1, RYB2 и RYB3, и инкубировали 30 мин. Затем лунки планшетов промывали, наносили меченые пероксидазой антитела против человеческих иммуноглобулинов класса IgG, инкубировали 30 мин, планшеты снова промывали и проявляли при помощи ТМВ-субстрата. После остановки реакции значения оптической плотности при 450 нм считывали при помощи планшет-ридера Infinite F50 (Tecan, Австрия). В результате такого анализа было установлено, что тяжелые цепи всех трех антител принадлежат подклассу IgG1.
Типы легких цепей определяли при помощи того же набора «Human IgG Subclass Profile Kit», однако в качестве меченых пероксидазой антител использовали козьи поликлональные антитела против легких цепей типа kappa или против легких цепей типа Lambda человеческих иммуноглобулинов (Bethyl Laboratories, США). Легкие цепи антител RYB1, RYB2 и RYB3 оказались принадлежащими типу kappa.
Пример 8. Определение аминокислотных последовательностей вариабельных участков тяжелых и легких цепей антител RYB1, RYB2 и RYB3.
Для определения аминокислотных последовательностей вариабельных участков антител из клеток гибридом BIONA-RYB1, BIONA-RYB2 и BIONA-RYB3 выделяли суммарную клеточную РНК, при помощи обратной транскриптазы получали кДНК и проводили амплификацию фрагментов кДНК, кодирующих вариабельные домены иммуноглобулинов человека. Амплифицированные фрагменты клонировали, подвергали секвенированию и по полученным последовательностям нуклеотидов методом трансляции in silico определяли последовательности аминокислот соответствующих участков антител RYB1, RYB2 и RYB3. Ниже приведено подробное описание процедуры и представлены результаты.
По 5·107 клеток каждой гибридомы собирали в фазе экспоненциального роста. Клетки промывали PBS и лизировали в 10 мл RLT-буфера из набора для выделения РНК «RNeasy Midi Kit» (Qiagen, США). Для уменьшения вязкости раствора лизаты продавливали по 1 разу через иглы калибра 29G, добавляли равный объем (10 мл) 70%-ного этанола и наносили на хроматографические колонки из набора для выделения РНК. Колонки промывали растворами RW1 и RPE из набора, после чего элюировали РНК в 4 мл Н2О. К элюатам добавляли по 100 мкл 5 М NaCl и 10,5 мл этанола и инкубировали в течение ночи при - 20°С для осаждения РНК. Осадки РНК собирали центрифугированием 30 мин при 4000 об/мин, растворяли в 900 мкл Н2О, добавляли по 100 мкл 10-кратного буфера для ДНК-азы (400 мМ Tris-HCl, рН 7,0, 100 мМ Nad, 100 мМ MgCl2, no 5 мкл рекомбинантной ДНК-азы I, 10 ед./мкл, свободной от примесей рибонуклеаз (Roche Applied Science, США) и инкубировали 2 ч при комнатной температуре для расщепления остаточной клеточной ДНК. Реакцию останавливали добавлением 40 мкл 0,5 М EDTA, добавляли 4 мл буфера RLT и 2,8 мл этанола и наносили на колонки из набора для выделения РНК для проведения второго раунда очистки, как описано выше. Элюированную РНК, свободную от примесей геномной ДНК, осаждали этанолом, собирали центрифугированием, растворяли в H2O и доводили концентрацию до 1 мг/мл. Концентрацию РНК измеряли спектрофотометрическим методом при длине волны 260 нм. Отсутствие деградации РНК контролировали при помощи электрофореза в агарозно-формальдегидном геле в присутствии бромистого этидия. Соотношение интенсивностей полос 28S и 18S рибосомной РНК составляло приблизительно 2:1.
Реакцию обратной транскрипции для РНК из клеток BIONA-RYB1, BIONA-RYB2 и BIONA-RYB3 проводили в объеме 10 мкл с использованием 2 мкг суммарной клеточной РНК, oligo(dT)-затравки и обратной транскриптазы «PowerScript» (Clontech, США) в течение 2 ч при 42°С согласно протоколу фирмы-производителя. Аликвоты полученных кДНК амплифицировали в присутствии наборов праймеров, специфичных к 5′- и 3′-концам нуклеотидных последовательностей, кодирующих вариабельные участки тяжелых (VH) и легких (VL) цепей иммуноглобулинов человека класса IgG1 kappa (Antibody Engineering: Methods and Protocols, edited by Benny K. C. Lo 2004, Humana Press Inc., p.282). Продукты амплификации клонировали в вектор pCR2.1 (Invitrogen, США) и полученные клоны (не менее 4 клонов для каждого вариабельного участка иммуноглобулина) подвергали секвенированию. По результатам секвенирования составляли нуклеотидные и аминокислотные последовательности вариабельных участков VH и VL антител RYB1, RYB2 и RYB3. Области, определяющие комплементарность (Complementarity Determining Region, CDR) в последовательностях VH и VL идентифицировали в соответствии с системой нумерации IMGT (Lefranc MP, Giudicelli V, Ginestoux С, Bodmer J, Muller W, Bontrop R, Lemaitre M, Malik A, Barbie V, Chaume D. Nucleic Acids Res. 1999, 27(1):209-212).
Таким образом были определены следующие аминокислотные последовательности, представленные в перечне последовательностей:
последовательность участка VH антитела RYB1, SEQ ID NO:5;
последовательность CDR1 участка VH антитела RYB1, SEQ ID NO:6;
последовательность CDR2 участка VH антитела RYB1, SEQ ID NO:7;
последовательность CDR3 участка VH антитела RYB1, SEQ ID NO:8;
последовательность участка VL антитела RYB1, SEQ ID NO:9;
последовательность CDR1 участка VL антитела RYB1, SEQ ID NO:10;
последовательность CDR2 участка VL антитела RYB 1, SEQ ID NO:11;
последовательность CDR3 участка VL антитела RYB1, SEQ ID NO:12;
последовательность участка VH антитела RYB2, SEQ ID NO:13;
последовательность CDR1 участка VH антитела RYB2, SEQ ID NO:14;
последовательность CDR2 участка VH антитела RYB2, SEQ ID NO:15;
последовательность CDR3 участка VH антитела RYB2, SEQ ID NO:16;
последовательность участка VL антитела RYB2, SEQ ID NO:17;
последовательность CDR1 участка VL антитела RYB2, SEQ ID NO:18;
последовательность CDR2 участка VL антитела RYB2, SEQ ID NO:19;
последовательность CDR3 участка VL антитела RYB2, SEQ ID NO:20;
последовательность участка VH антитела RYB3, SEQ ID NO:21;
последовательность CDR1 участка VH антитела RYB3, SEQ ID NO:22;
последовательность CDR2 участка VH антитела RYB3, SEQ ID NO:23;
последовательность CDR3 участка VH антитела RYB3, SEQ ID NO:24;
последовательность участка VL антитела RYB3, SEQ ID NO:25;
последовательность CDR1 участка VL антитела RYB3, SEQ ID NO:26;
последовательность CDR2 участка VL антитела RYB3, SEQ ID NO:27;
последовательность CDR3 участка VL антитела RYB3, SEQ ID NO:28.
Пример 9. Изучение взаимной конкуренции антител.
Для исследования взаимной конкуренции антитела RYB1, RYB2 и RYB3 метили флуоресцеин изотиоцианатом (FITC) с использованием набора «FluoroTag FITC Conjugation Kit» (Sigma-Aldrich, CIIIA) согласно инструкции компании-производителя. В предварительных экспериментах по цитофлуориметрическому анализу связывания FITC-меченых антител в различных концентрациях с пермеабилизованными клетками HEK-Е1Е2 определяли константы ассоциации Ka и концентрации антител, при которых реакция связывания выходит на насыщение. Для этого снимали зависимость средних значений флуоресценции клеток, инкубированных с мечеными антителами, от концентрации антител. Константы ассоциации и концентрации антител, достаточные для насыщения центров специфического связывания, которыми являются белковые комплексы Е1-Е2 HCV, за 2 часа инкубации, приведены в табл.1.
В эксперименте по изучению взаимной конкуренции составляли смеси FITC-меченых и немеченых антител в отношениях 1:0 (только меченое антитело), 1:5 и 1:20 во всех возможных комбинациях. В соответствии с данными предварительных экспериментов концентрацию меченых антител в смесях выбирали равной 5 мкг/мл для RYB1 и RYB2 и 2 мкг/мл для RYB3. В качестве отрицательного контроля использовали полностью человеческое моноклональное антитело Адалимумаб (торговое название «Ху-мира»), специфичное к фактору некроза опухолей альфа и принадлежащее подклассу IgG1. К суспендированным образцам пермеабилизованных клеток HEK-Е1Е2 добавляли 1,5 мл раствора 0,3% BSA в PBS, инкубировали 5 мин, клетки осаждали центрифугированием и ресуспендировали в 20 мкл смеси меченых и немеченых антител. Клетки инкубировали 2 часа при комнатной температуре, затем отмывали в 1,5 мл раствора 0,3% BSA в PBS и суспендировали в 400 мкл 1%-ного формальдегида в PBS. Флуоресценцию клеток анализировали на проточном цитофлуориметре FACS-Calibur (BD Biosciences, США) и определяли средние значения флуоресценции для каждого образца. Уровень флуоресценции, обусловленный неспецифическим связыванием меченых антител с клетками, определяли аналогичным образом с использованием нетрансфицированных клеток НЕК293 вместо клеток HEK-Е1Е2.
Результаты эксперимента представлены на фиг.4. Они показывают, что антитела RYB1, RYB2 и RYB3 эффективно конкурируют друг с другом за связывание с белковым комплексом Е1-Е2. Степени конкуренции антител в гетерологичных парах практически неотличимы от конкуренции в гомологичных парах. При этом человеческое антитело Адалимумаб не конкурирует ни с одним из исследуемых антител.
По-видимому, эпитопы данных антител взаимно перекрываются или расположены близко друг от друга в пространственной структуре белка Е2, так что связывание антитела с одним из эпитопов мешает взаимодействию другого эпитопа со своим антителом, что свидетельствует о правильности использования предлагаемого набора эпитопов для воздействия на вирус гепатита С полученными антителами.
Пример 10. Образование комплексов антител с рекомбинантным белком Е2.
Аминокислотную последовательность (SEQ ID NO:29), соответствующую последовательности белка Е2 вируса гепатита С изолята Н77 (генотип 1а), извлекали из международной базы данных NCBI Protein (http://www.ncbi.nlm.nih.gov/protein). Последовательность имеет референсный номер NP_751921.1 и содержит 363 аминокислотных остатка с координатами 384-746 по последовательности полипротеина HCV того же изолята (референсный номер NP_671491.1). Аминокислотную последовательность белка Е2 подвергали обратной трансляции in silico для получения нуклеотидной последовательности, кодирующей белок Е2 и оптимизированной по кодонному составу для экспресии в бактериях E.coli. На основании этой последовательности методом химического синтеза конструировали ген белка Е2 и встраивали в прокариотический экспрессионный вектор pET100/D-TOPO (Invitrogen, США) с образованием плазмиды рЕТ100-Е2. В процессе встраивания участки, кодирующие Xpress-эпитоп и 6xHis-tag, были из вектора удалены.
Плазмидой рЕТ100-Е2 трансформировали клетки E.coli штамма BL21 (ВЕ3), клоны бактерий-трансформантов выращивали при 37°С в шейкер-инкубаторе Innova 43R (New Brunswick Scientific, США) в стеклянных колбах объемом 2 л до оптической плотности OD600=0,5, экспрессию рекомбинантного продукта индуцировали добавлением IPTG до 1 мМ, после чего бактерии растили еще 150 мин. Рекомбинантный белок Е2 очищали из лизата бактерий при помощи стандартных хроматографических процедур.
Попытки продемонстрировать связывание антител RYB1, RYB2, RYB3 с рекомбинантным белком Е2 традиционным методом Western-анализа с проведением электрофореза в полиакриламидном геле в присутствии додецил-сульфата натрия (SDS) не дали положительных результатов. По-видимому, изменение конформации белка Е2 под действием SDS нарушает стуктуру эпитопов, распознаваемых антителами. Поэтому был применен альтернативный подход к наблюдению комплексов, основанный на масс-спектрометрии.
Для образования комплексов с антителами 5 мкл раствора очищеного белка Е2 с концентрацией 4 мкМ в PBS смешивали с 5 мкл раствора одного из антител RYB1, RYB2 или RYB3 с концентрацией 2 мкМ. К 9 мкл полученной смеси добавляли 1 мкл стабилизирующего реагента K200 из набора «К200 MALDI MS analysis kit» (CovalX, Швейцария) и инкубировали 3 ч при комнатной температуре. Обработка стабилизирующим реагентом приводит к ковалентной сшивке компонентов комплекса друг с другом (Bich С, Maedler S, Chiesa K, DeGiacomo F, Bogliotti N, Zenobi. Anal Chem. 2010 82(1): 172-179). При этом молекулярная масса комплекса увеличивается приблизительно на 4-5% за счет добавления молекул стабилизирующего реагента.
Образцы для проведения масс-спектрометрического анализа по технологии матрично-активированной лазерной десорбции/ионизации (MALDI) готовили следующим образом. По окончании инкубации к 1 мкл смеси, содержащей сшитые комплексы, добавляли 1 мкл матрицы, состоящей из перекристаллизованной синапиновой кислоты (10 мг/мл) в растворе ацетонитрил-вода (1:1, v/v), дополнительно содержащем 0,1%-ную триф-торуксусную кислоту. 1 мкл полученной смеси наносили на MALDI-планшет «МТР AnchorChip 384 TF» (Bmker Daltonik, Германия), где образец кристаллизовался при комнатной температуре.
Сразу после кристаллизации планшет с образцами помещали в приемное устройство время-пролетного масс-анализатора «Ultraflex III MALDI TOF TOF» (Bmker Daltonik, Германия), оснащенного детектором «НМ3 High-Mass system» (CovalX, Швейцария), позволяющим регистрировать микроагрегаты с молекулярной массой вплоть до 2 МДа с чувствительностью в наномолярном диапазоне. Масс-спектрометрический анализ проводили с использованием стандартного азотного лазера в линейном положительном диапазоне при следующих настраиваемых параметрах: Ion Source 1: 20 kV, Ion Source 2: 17 kV, Lens: 12 kV, Pulse Ion Extraction: 400 ns, HM3 Gain Voltage: 3.14 kV, HM3 Acceleration Voltage: 20 kV. Спектры усредняли по 300 событиям лазерной детекции. Для калибровки инструмента использовали кластеры инсулина, бычьего сывороточного альбумина и иммуноглобулина G. Обработку данных проводили при помощи программы «Complex Tracker analysis», версия 2.0 (CovalX, Швейцария). Все измерения повторяли трижды.
Анализ чистого белка Е2, а также препаратов антител, взятых по-отдельности, показал (фиг.5, A-D), что ни белок Е2, ни антитела не образуют комплексов или агрегатов сами с собой. Следует отметить, что из-за обработки стабилизирующим реагентом наблюдаемая молекулярная масса рекомбинантного белка Е2, согласно расчету составляющая 40048 Да, вырастает в среднем до 44215 Да. Присутствующий в спектрах образцов антител дополнительный пик с молекулярной массой около 60 кДа соответствует, вероятнее всего, какому-то примесному белку. После смешивания белка Е2 с препаратами антител в спектре образцов появляются пики (фиг.5 E-G), соответствующие по массе молекулярным комплексам, содержащим одну молекулу антитела и одну или две молекулы белка Е2 (табл.2). Стехиометрическая структура комплексов находится в полном соответствии с двухвалентной природой моноклональных антител класса IgG. Таким образом, подтверждена способность антител RYB1, RYB2 и RYB3 образовывать комплексы с рекомбинантным белком Е2 в растворе.
Пример 11. Определение эпитопов антител.
Для установления эпитопной специфичности антител RYB1, RYB2 и
RYB3 с высоким разрешением комплексы антиген-антитело обрабатывали сшивающим реагентом, меченым дейтерием, после чего подвергали мультиферментному протеолизу. Анализ и идентификацию полученных пептидов проводили при помощи нано-жидкостной хроматографии с последующей масс-спектрометрией. Эпитопы определяли по исчезновению ряда меченых пептидов из спектра продуктов протеолиза антигена в случае образования комплекса с антителом. Ниже приведено подробное описание каждой из стадий процедуры.
На первой стадии проводили образование комплексов антиген-антитело в стехиометрическом соотношении 2:1 в следующих условиях: 5 мкл раствора рекомбинантного белка Е2 (см. пример 10) с концентрацией 4 мкМ смешивали с 5 мкл раствора одного из антител RYB1, RYB2 или RYB3 с концентрацией 2 мкМ и инкубировали 180 мин при 37°С. В качестве образца чистого антигена использовали 10 мкл раствора белка Е2 с концентрацией 2 мкМ. Образец чистого антигена также инкубировали 180 мин при 37°С.
На второй стадии к образцам комплексов антиген-антитело и образцу чистого антигена добавляли по 1 мкл заранее приготовленной смеси (1:1) сшивающих реагентов дисукцинилимидил суберата DSS-dO и меченного 12 атомами дейтерия дисукцинилимидил суберата DSS-dl2 (CovalX, Швейцария) с суммарной концентрацией 2 мг/мл в диметилформамиде. Образцы инкубировали 180 мин при комнатной температуре для образования сшивок.
На третьей стадии проводили восстановление дисульфидных связей и алкилирование образцов для облегчения последующего протеолиза. К образцам после реакции сшивки добавляли по 20 мкл 25 мМ бикарбоната аммония, рН 8,3, и 2,5 мкл 500 мМ дитиотреитола, инкубировали 60 мин при 55°С, затем добавляли 2,5 мкл 1 М иодоацетамида и икубировали еще 60 мин при комнатной температуре без доступа света.
На четвертой стадии образцы подвергали протеолизу при помощи трипсина или α-химотрипсина. Для обработки трипсином к образцам добавляли по 120 мкл буфера для протеолиза (100 мМ Трис-HCl, рН 7,8, 10 мМ CaCl2), 2 мкл раствора трипсина (1 мг/мл) и инкубировали ночь при 37°С. Для обработки α-химотрипсином к образцам добавляли по 120 мкл того же буфера для протеолиза, 2 мкл раствора α-химотрипсина (200 мкМ) и инкубировали ночь при 30°С.
На пятой стадии продукты протеолиза разделяли на нано-жидкостном хроматографе «Ultimate 3000» (Dionex, Германия) с использованием предварительной колонки «300-µm ID × 5-mm C4 РерМар» и основной колонки «75-µm ID x 5-cm C4 РерМар» в градиенте (95% Н2О, 5% ацетонитрил, 0,1% НСООН) - (20% Н2О, 80% ацетонитрил, 0,1% НСООН).
На шестой стадии материал хроматографических пиков подвергали масс-спектрометрическому анализу на приборе «LTQ Orbitrap XL» (Thermo Scientific, США), аппаратно соединенном с хроматографической системой. Поскольку сшивающий реагент, использованный для обработки комплексов, является смесью меченных дейтерием и немеченых молекул в соотношении 1:1, то каждый пептид, несущий по одной молекуле сшивающего реагента, представлен на масс-спектре двумя сигналами одинаковой интенсивности, отличающимися друг от друга ровно на 12 атомных единиц - по числу атомов дейтерия в молекуле DSS-dl2 (в случае однократно ионизированных пептидов). Этот факт позволяет произвести первичный отбор сигналов, соответствующих пептидам с одиночной молекулой сшивающего реагента. Дальнейшую идентификацию таких пептидов с определением их аминокислотной последовательности осуществляли на основании данных масс-спектрометрии с помощью программного комплекса «XQuest», версия 2.0, и базы данных «CovalX_110823_01.fasta» компании CovalX (Швейцария), используя последовательность аминокислот рекомбинантного белка Е2 (SEQ ID NO:29). При этом для каждого пептида устанавливали также положение аминокислоты, ковалентно связанной с молекулой сшивающего реагента.
На седьмой стадии проводили сравнение спектров продуктов протеолиза чистого белка Е2 и его стехиометрических комплексов с антителами. Отсутствие определенного пептида среди продуктов протеолиза комплекса указывает на тесный контакт некоторых аминокислот пептида с антителом, то есть на перекрывание последовательности пептида с искомым эпитопом для данного антитела. Следует отметить, что взаимодействие антигена с антителом само по себе не меняет спектр продуктов протеолиза, поскольку протеолитической реакции предшествует восстановление дисульфидных связей и алкилирование образцов, приводящее к разрушению комплексов антиген-антитело. Вместе с тем, образование комплеса антиген-антитело может воспрепятствовать химической реакции связывания молекул сшивающего реагента с теми аминокислотами антигена, которые находятся в непосредственной близости с антителом. В результате пептиды, содержащие такие аминокислоты, оказываются свободными от сшивающего реагента, не проходят первичный отбор и выпадают из наблюдения.
В табл.3 приведены последовательности продуктов протеолиза чистого белка Е2, несущих по одной молекуле сшивающего реагента. В табл.4-6 представлены пептиды из табл.3, отсутствующие среди продуктов протеолиза комплексов белка Е2 с антителами RYB1, RYB2 и RYB3.
Анализ расположения молекул сшивающего реагента на пептидных продуктах протеолиза белка Е2 до и после образования комплексов с антителом позволяет с высокой точностью локализовать участки Е2, вступившие в контакт с антителом. Так, в случае антитела RYB1 аминокислоты белка Е2 на участках от N-конца до лизина Lys588 включительно и, начиная с серина Ser599 до С-конца, сохраняют способность связываться со сшивающим реагентом независимо от образования комплекса с антителом (фиг.6). В то же время аминокислоты серин Ser595 и аргинин Arg596 теряют способность связываться со сшивающим реагентом после образования комплекса. На основании этих данных можно сделать вывод, что эпитоп белка Е2 для антитела RYB1 (эпитоп Э1) представляет собой непрерывную последовательность аминокислот, содержащую аминокислоты Ser595 и Arg596 и заключенную в пределах участка HPEATYSRCG (589-598) (SEQ ID NO:30). Наиболее вероятно, что эпитопом является центральный фрагмент этого участка из 6 аминокислот: EATYSR (591-596) (SEQ ID NO:31). Данный эпитоп в литературе не описан.
В случае антитела RYB2 аминокислоты белка Е2 на участках от N-конца до серина Ser501 включительно и, начиная с треонина Thr510 до С-конца, сохраняют способность связываться со сшивающим реагентом независимо от образования комплекса с антителом (фиг.7). В то же время аминокислота тирозин Tyr507 теряет способность связываться со сшивающим реагентом после образования комплекса. На основании этих данных можно сделать вывод, что эпитоп белка Е2 для антитела RYB2 (эпитоп Э2) представляет собой непрерывную последовательность аминокислот, содержащую аминокислоту Tyr507 и заключенную в пределах участка VCGPVYCF (502-509) (SEQ ID NO:32). Наиболее вероятно, что эпитопом является центральный фрагмент этого участка из 6 аминокислот: CGPVYC (503-508) (SEQ ID NO:33). Данный эпитоп в литературе не описан.
В случае антитела RYB3 аминокислоты белка Е2 на участках от N-конца до лизина Lys588 включительно и, начиная с аргинина Arg606 до С-конца, сохраняют способность связываться со сшивающим реагентом независимо от образования комплекса с антителом (фиг.8). В то же время аминокислоты серии Ser595, аргинин Arg596 и серии Ser599 теряют способность связываться со сшивающим реагентом после образования комплекса. На основании этих данных можно сделать вывод, что эпитоп белка Е2 для антитела RYB3 (эпитоп Э3) представляет собой непрерывную последовательность аминокислот, содержащую аминокислоты Ser595, Arg596 и Ser599 и заключенную в пределах участка HPEATYSRCGSGPWITP (589-605) (SEQ ID NO:34). Скорее всего, эпитопом является внутренний фрагмент этого участка из 6 аминокислот: YSRCGS (594-599) (SEQ ID NO:35) или SRCGSG (595-600) (SEQ ID NO:36). Данный эпитоп в литературе не описан.
Необходимо отметить, что эпитопы Э1 и Э3 содержат общие аминокислоты серии Ser595 и аргинин Arg596 и, следовательно, перекрываются, что согласуется с данными экспериментов по взаимной конкуренции антител (пример 9). Анализ полученных последовательностей эпитопов показывает (табл.7), что эпитоп Э2 абсолютно консервативен для всех генотипов вируса. Эпитопы Э1 и Э3 содержат как консервативные, так и варьирующие аминокислоты.
Пример 12. Определение конформационного характера эпитопов.
Невозможность наблюдать реакцию антител RYB1, RYB2 и RYB3 с белком Е2 методом Western-анализа наводит на мысль о том, что эпитопы для этих антител, установленные в примере 11, имеют конформационный характер. Для проверки этого предположения был поставлен ряд экспериментов по определению способности коротких пептидов, содержащих эпитопы, конкурировать с полноразмерным рекомбинантным белком Е2 за связывание с антителами. Поскольку короткие пептиды не сохраняют конформацию исходного белка, положительный результат экспериментов по конкуренции говорил бы о том, что распознавание эпитопов антителами не зависит от конформационной структуры эпитопов и определяется лишь их аминокислотными последовательностями (линейный характер эпитопов). Наоборот, отрицательный результат свидетельствовал бы в пользу значимости конформационной структуры для взаимодействия с данными антителами (конформационный характер эпитопов).
Набор пептидов получали протеолизом рекомбинантного белка Е2 при помощи иммобилизованного пепсина в следующих условиях. К 25 мкл раствора белка Е2 (см. пример 10) в PBS с концентрацией 13 мкМ добавляли 5 мкл суспензии агарозного носителя с иммобилизованным пепсином (Thermo Scientific, США) и инкубировали 30 мин при комнатной температуре. По окончании инкубации образцы центрифугировали и супернатант, содержащий продукты протеолиза, переносили в другие пробирки. Условия реакции были оптимизированы для получения основной массы пептидных продуктов в диапазоне 1000-3500 Да (8-30 аминокислотных остатков). Пепсин почти не обладает специфичностью в отношении гидролизуемых пептидный связей, поэтому следует ожидать, что представленность любых коротких последовательностей исходного белка, в частности эпитопов для антител RYB1, RYB2 и RYB3, среди продуктов протеолиза примерно одинакова и соответствует молярной концентрации исходного белка.
Эксперименты по конкурентному связыванию выполняли в следующих условиях. К 5 мкл образца пептидной смеси с концентрацией 13 мкМ (по исходному белку Е2) добавляли 5 мкл препарата одного из антител RYB1, RYB2 или RYB3 с концентрацией 4 мкМ и инкубировали 6 ч при 37°С. Концентрация пептидов в образце обеспечивала заведомый избыток последовательностей эпитопов по отношению к концентрации центров связывания антител (1,625:1). Затем к реакционной смеси добавляли 10 мкл раствора рекомбинантного белка Е2 с концентрацией 4 мкМ и проводили обработку стабилизирующим реагентом К200, как описано в примере 10. Далее образцы подвергали масс-спектрометрическому анализу по технологии MALDI при помощи время-пролетного масс-анализатора «Ultraflex III MALDI TOF TOF» как описано в том же примере.
Результаты экспериментов показали (фиг.9), что короткие пептиды, содержащие эпитопы, не ингибируют связывания полноразмерного белка Е2 с антителами. В присутствии избыточного количества пептидов образуются белковые комплексы, идентичные по молекулярной структуре комплексам белка Е2 с антителами, охарактеризованным в примере 6. На основании этих данных можно заключить, что связывание антител RYB1, RYB2 и RYB3 со своими эпитопами возможно только при определенных конформациях эпитопов в пространственной структуре молекулы белка Е2. Таким образом, эпитопы указанных антител имеют конформационный характер.
Пример 13. Исследование нейтрализующей активности антител.
Нейтрализующую активность антител, то есть способность антител RYB1, RYB2 и RYB3 препятствовать заражению клеток вирусом, изучали на модели клеточной культуры гепатоцеллюлярной карциномы человека Huh-7 с использованием псевдовирусных частиц HCVpp.Частицы HCVpp являются псевдолентивирусными частицами, несущими в качестве белков оболочки комплексы Е1-Е2 вируса гепатита С (Bartosch В, Dubuisson J, Cosset FL. J Exp Med. 2003, 197(5):633-642).
Псевдовирусные частицы HCVpp получали как описано ниже. Клетки HEK293-TN (System Biosciences, США), экспрессирующие Т-большой антиген вируса SV40, выращивали во флаконах Т225 (Coming, США) в среде DMEM с высоким содержанием глюкозы, дополнительно содержащей 10%-ную инактивированную фетальную бычью сыворотку (Invitrogen, США) и 2 мМ глутамин, до 70%-ной плотности монослоя. Непосредственно перед трансфекцией среду во флаконах меняли на свежую среду DMEM, содержащую 2%-ную фетальную бычью сыворотку. Трансфекцию клеток проводили смесью плазмид pCMVdeltaR8.2, pSIH-CMV-CopGFP-H1 (000 «МонА», Москва) и pcDNA3.1-E1E2 (см. пример 3) при помощи реагента Lipofectamine 2000 (Invitrogen, США). К 1,5 мл среды DMEM без сыворотки добавляли 225 мкл реагента Lipofectamine 2000, перемешивали пипетированием и по каплям добавляли к заранее приготовленному раствору смеси плазмид (54 мкг pCMVdeltaR8.2, 18 мкг pSIH-CMV-CopGFP-H1 и 18 мкг pcDNA3.1-E1E2 в 1.5 мл среды DMEM без сыворотки). Полученный раствор перемешивали, инкубировали 15 мин и вносили во флакон с клетками HEK293-TN. Клетки инкубировали 24 часа при 37°С в СO2-инкубаторе в атмосфере 5% CO2, после чего культуральную среду заменяли на свежую и инкубировали клетки еще 48 часов. По окончании инкубации культуральную среду, содержащую частицы HCVpp, собирали и фильтровали через ацетат-целлюлозные мембранные фильтры с размером пор 0,22 мкм (Corning, США). Порции среды замораживали и хранили при -80°С.
Плазмида pCMVdeltaR8.2 содержит гены gag-pol и rev вируса иммунодефицита человека (HIV), продукты экспрессии которых необходимы для упаковки псевдолентивирусных частиц и встраивания вирусного генома в геномную ДНК зараженной клетки-хозяина. Плазмида pSIH-CMV-CopGFP-H1 кодирует геномную РНК лентивирусного вектора на основе HIV и несет в своем составе ген зеленого флуоресцентного белка CopGFP из копеподы Pontellina plumata под контролем высокоэффективного промотора ранних генов цитомегаловируса. Плазмида pcDNA3.1-E1E2 кодирует белки Е1 и Е2 оболочки вируса гепатита С.При одновременной экспрессии в трансфицированных клетках генов трех вышеуказанных плазмид клетки продуцируют в культуральную среду псевдовирусные частицы HCVpp, внутренней частью которых является геномная РНК ленти-вектора в комплексе с продуктами генов gag-pol и rev, а оболочкой служит белковый комплекс Е1-Е2 HCV. Такие частицы могут заражать клетки Huh-7 по механизму, сходному с механизмом инфицирования клеток печени вирусом HCV. При этом ген CopGFP в составе лентивекторного генома встраивается в ДНК зараженной клетки и экспрессируется с образованием флуоресцентного продукта, что позволяет детектировать заражение флуориметрическим методом.
Эксперимент по измерению нейтрализующей активности антител выполняли следующим образом. Готовили растворы антител RYB1, RYB2 и RYB3 с концентрацией 1 мг/мл. Композицию антител RYB1:RYB2:RYB3=1:1:1 получали путем смешивания 200 мкл раствора RYB1, 200 мкл раствора RYB2 и 200 мкл раствора RYB3. Композицию антител RYB1:RYB2:RYB3=20:40:40 получали путем смешивания 100 мкл раствора RYB1, 200 мкл раствора RYB2 и 200 мкл раствора RYB3. Композицию антител RYB1:RYB2:RYB3=40:20:40 получали путем смешивания 200 мкл раствора RYB1, 100 мкл раствора RYB2 и 200 мкл раствора RYB3. Наконец, композицию антител RYB1:RYB2:RYB3=40:40:20 получали путем смешивания 200 мкл раствора RYB1, 200 мкл раствора RYB2 и 100 мкл раствора RYB3.
К 1,9 мл культуральной среды, содержащей псевдовирусные частицы HCVpp, добавляли 100 мкл раствора антитела RYB1, RYB2 или RYB3 с концентрацией 1 мг/мл или 100 мкл одной из приготовленных композиций и инкубировали 1 час при 37°С. Суммарная концентрация антител во всех образцах инкубационной смеси составляла 50 мкг/мл. В качестве отрицательного контроля использовали человеческое моноклональное антитело Адалимумаб, которое добавляли также до конечной концентрации 50 мкг/мл. Клетки Huh-7 выращивали в лунках 6-луночных культуральных планшетов (Coming, США) в среде DMEM, дополнительно содержащей 10%-ную инактивированную фетальную бычью сыворотку (Invitrogen, США) и 2 мМ глутамин, до состояния 30%-ной плотности монослоя. Для заражения клеток культуральную среду в лунках планшетов заменяли на проинкубированные смеси частиц HCVpp с антителами или их композициями и инкубировали 3 часа при 37°С в СО2-инкубаторе. По окончании инкубации меняли среду в планшетах на свежую, культивировали клетки еще 4 дня и подвергали флуориметрическому анализу на проточном цитофлуориметре FACS-Calibur (BD Biosciences, США). Эффективность заражения определяли как процент флуоресцирующих клеток в образце. Результаты анализа представлены в табл.16. Как можно видеть, антитела RYB1, RYB2 и RYB3 проявляют значительную нейтрализующую активность (ингибирование заражения в 22-36 раз), что открывает возможности их широкого применения в клинической практике. При этом наиболее высокий результат по нейтрализации демонстрирует композиция трех антител в пропорции 1:1:1 (ингибирование заражения в 53,6 раза), а уменьшение содержания какого-либо антитела в композиции до 20% приводит к некоторому снижению нейтрализующей активности.
Таким образом, в экспериментах по изучению нейтрализующей активности антител на модельной системе клеток гепатоцеллюлярной карциномы человека Huh-7 продемонстрирована способность антител и их композиций эффективно ингибировать заражение клеток псевдовирусными частицами HCVpp. Степень ингибирования при оптимальном выборе композиции достигает примерно 53 раз, что свидетельствует о потенциальной пригодности данных антител и композиций на их основе для клинического применения при профилактике и лечении гепатита С.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ ДЛЯ ДИАГНОСТИКИ ГЕПАТИТА C ЧЕЛОВЕКА (ВАРИАНТЫ), СПОСОБ И НАБОР ДЛЯ ОБНАРУЖЕНИЯ АНТИТЕЛ К ВИРУСУ ГЕПАТИТА C ЧЕЛОВЕКА | 1991 |
|
RU2130969C1 |
| ОЛИГОМЕРНАЯ ЧАСТИЦА, ИНДУЦИРУЮЩАЯ ИММУНИТЕТ ПРОТИВ ВИРУСА ГЕПАТИТА С, СПОСОБ ПОЛУЧЕНИЯ ОЛИГОМЕРНОЙ ЧАСТИЦЫ, КОМПОЗИЦИЯ, СПЕЦИФИЧЕСКОЕ АНТИТЕЛО, НАБОР (ВАРИАНТЫ), ИММУНОЛОГИЧЕСКИЙ АНАЛИЗ И ВАКЦИНА ПРОТИВ ВИРУСА ГЕПАТИТА С | 1999 |
|
RU2247729C2 |
| НОВЫЕ АНТИТЕЛА ПРОТИВ ВИРУСА ГЕПАТИТА B И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2814471C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ ВИРУСА БЕШЕНСТВА И ИХ СМЕСЬ | 2019 |
|
RU2822457C2 |
| АНТИТЕЛА ПРОТИВ ВИРУСА ГЕПАТИТА В И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2803082C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К BTN2, И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2821703C2 |
| ХИМЕРНЫЕ АНТИГЕНЫ ДЛЯ ВАКЦИНЫ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2013 |
|
RU2639504C2 |
| ГУМАНИЗИРОВАННОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ Aβ и ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2783528C1 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА К ВИРУСУ ГРИППА А И ПУТИ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2803850C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |

















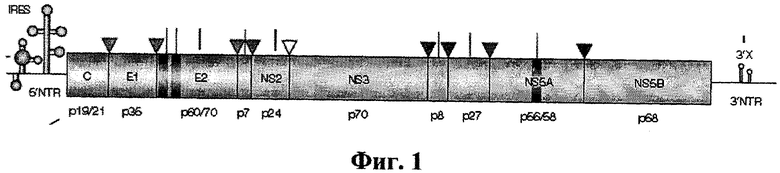
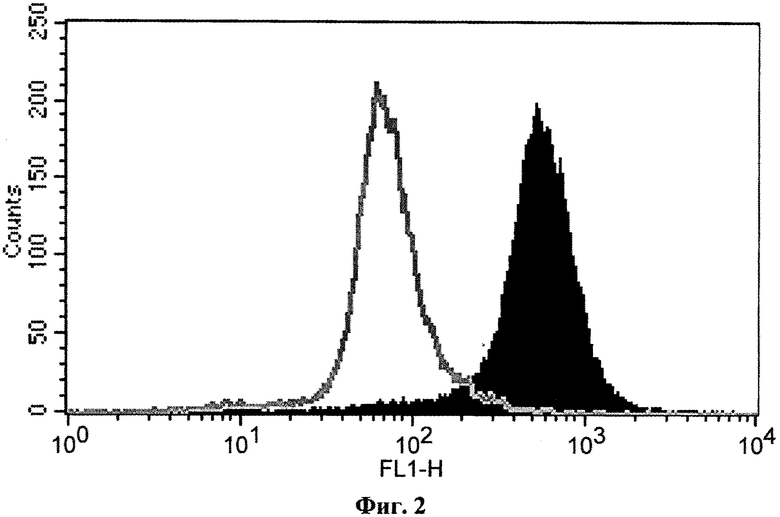
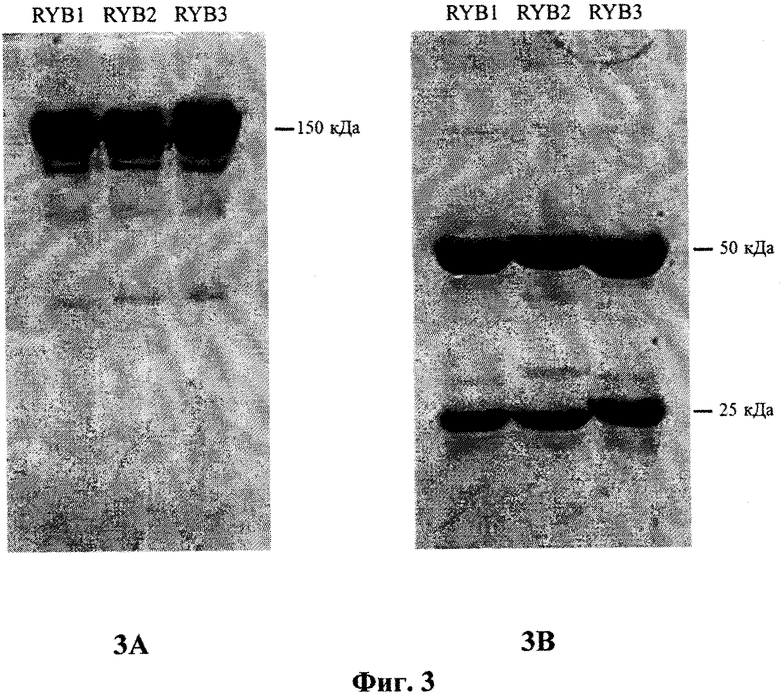
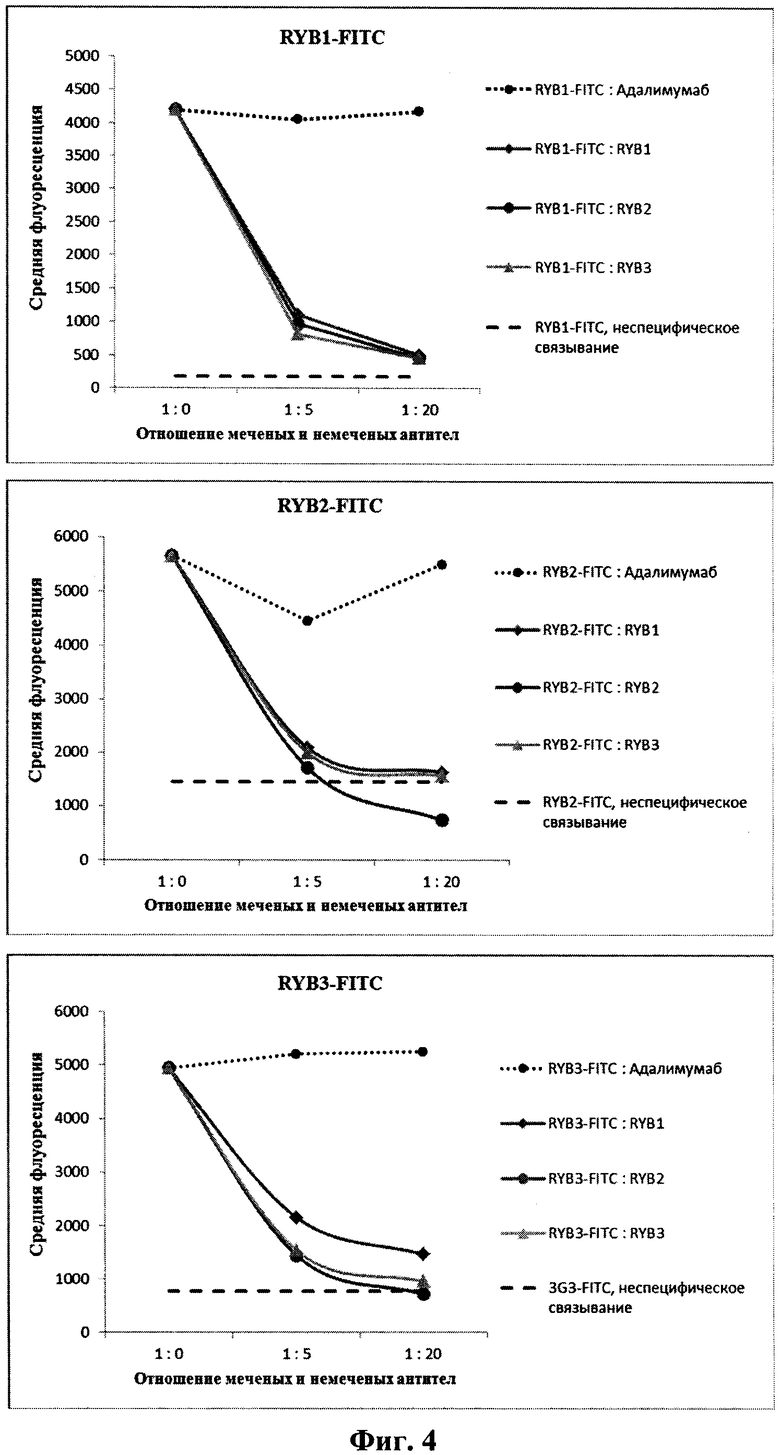
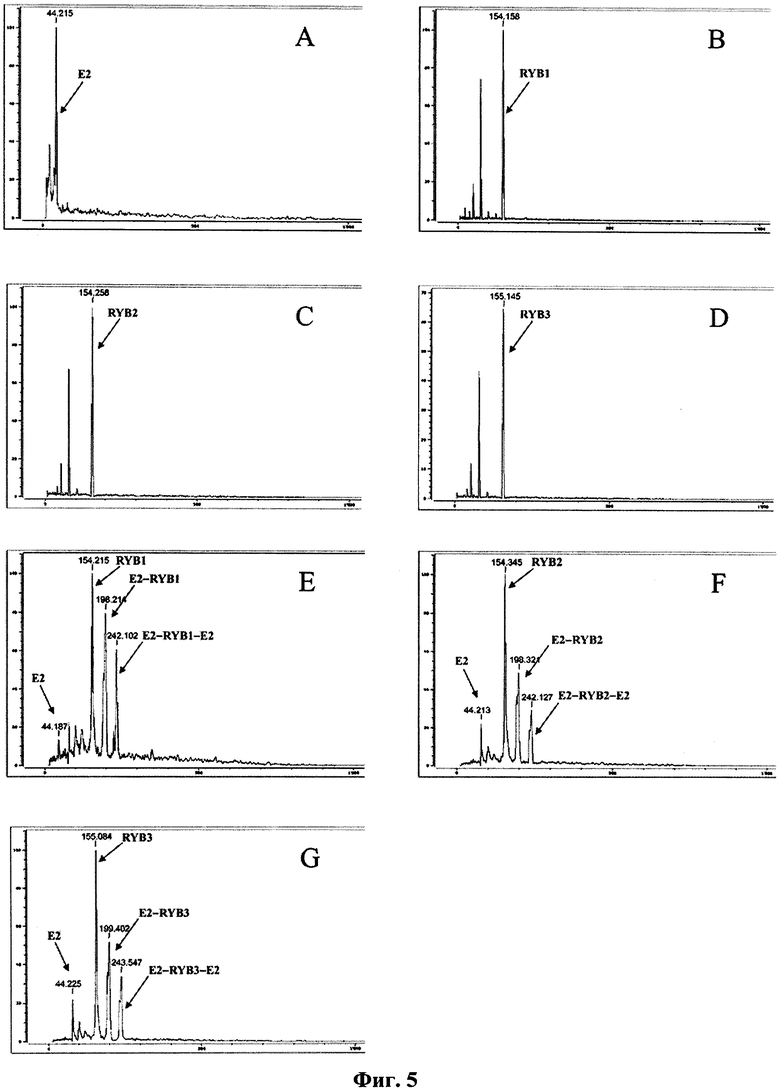
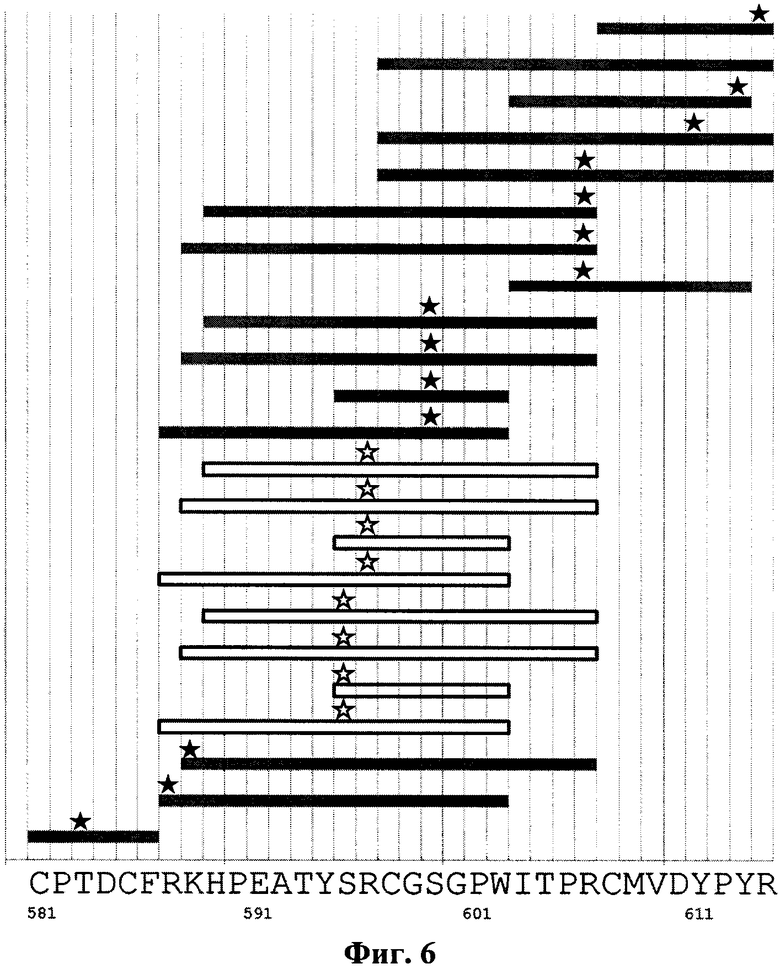
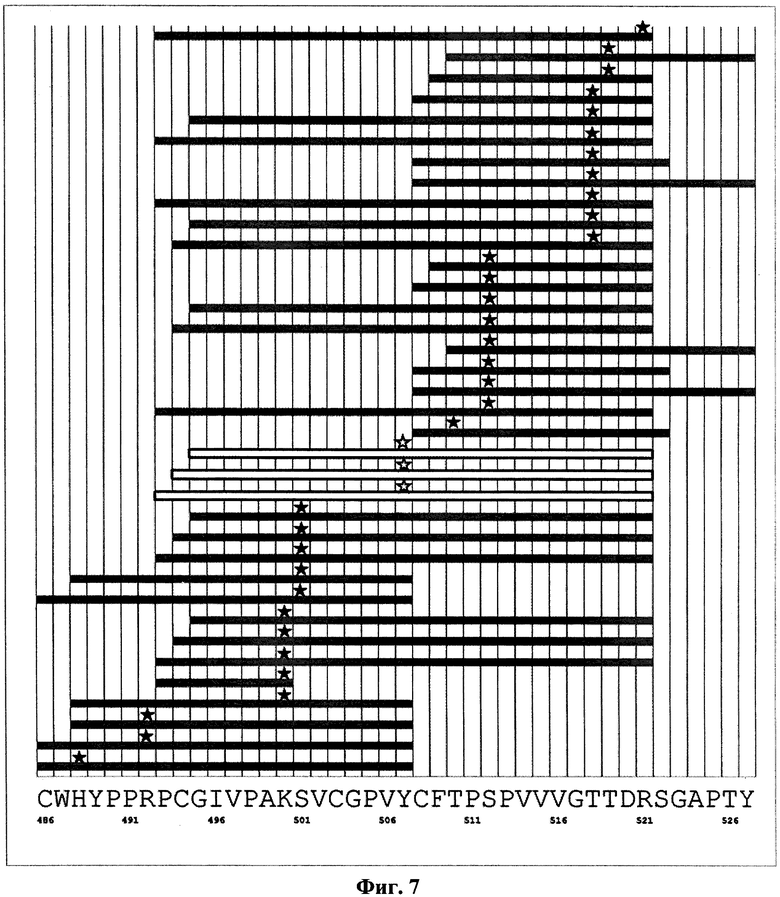
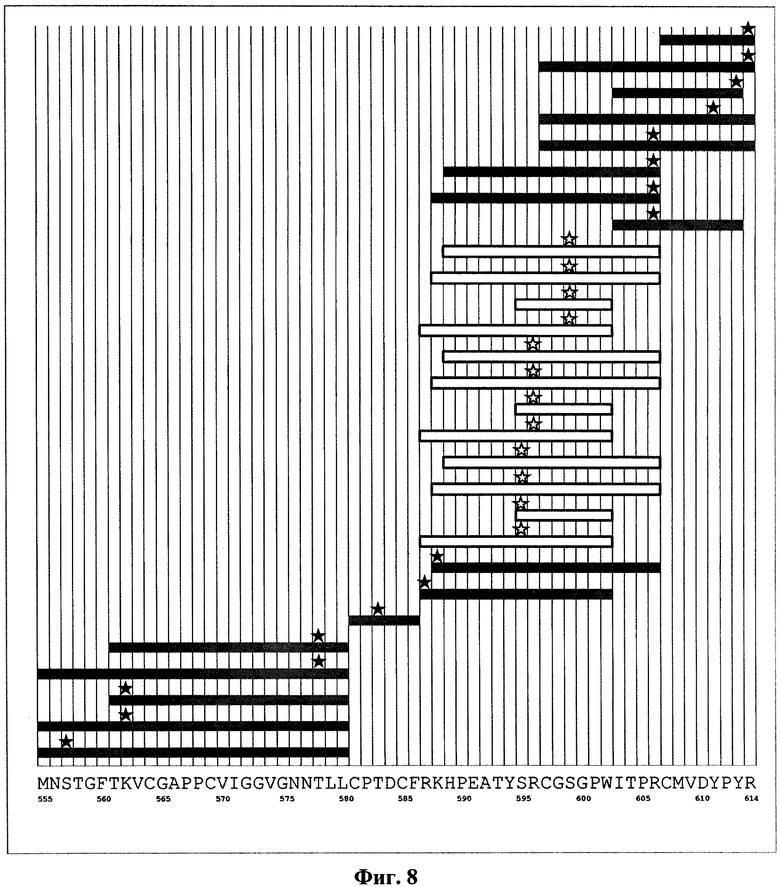
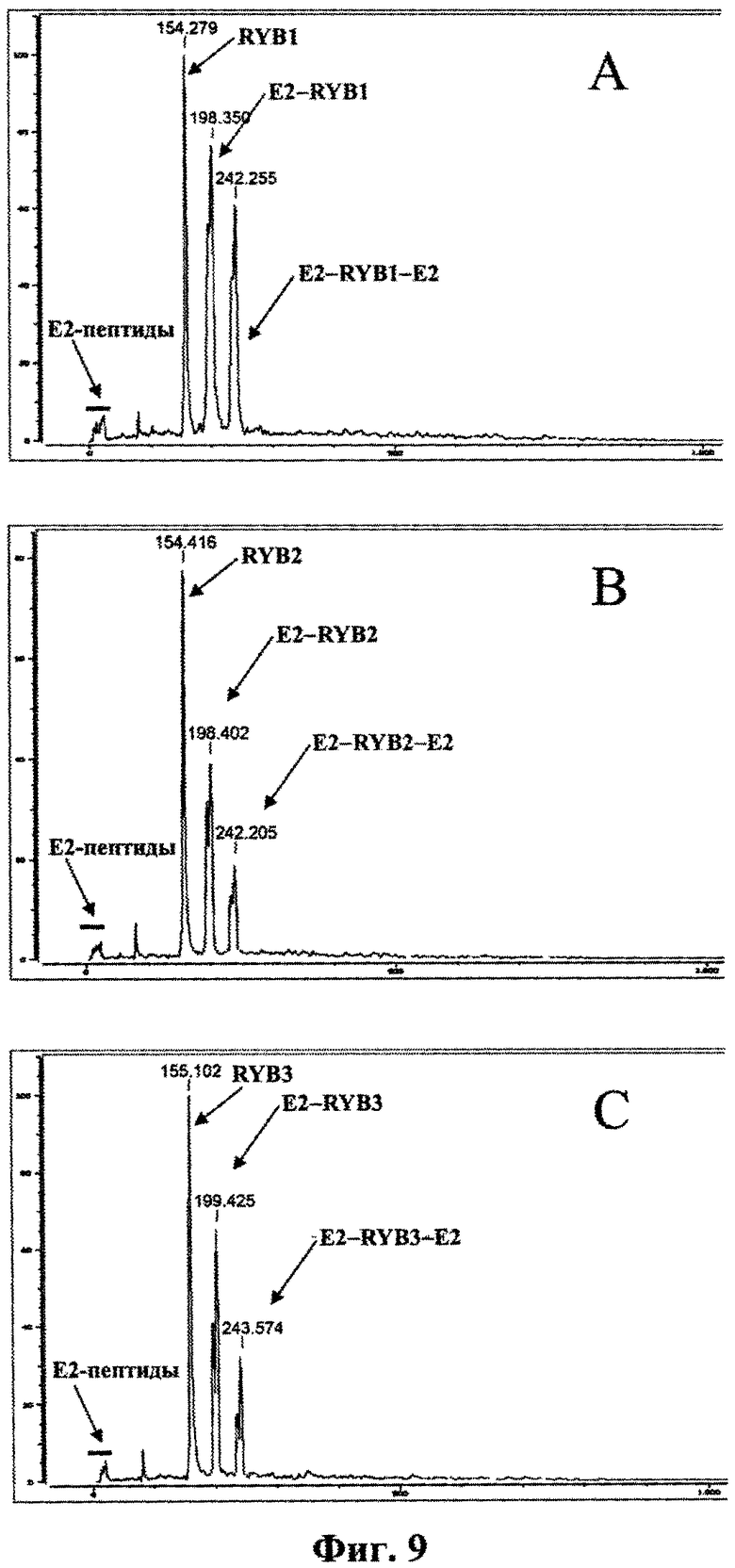
Изобретение относится к области биотехнологии. Раскрыты способы и технологии нейтрализации вируса гепатита С, а также антитела против вируса гепатита С. Предлагается использовать для профилактики и лечения гепатита С полностью человеческие моноклональные антитела RYB1, RYB2 и RYB3, а также композиции на их основе. Указанные антитела получают путем культивирования гибридом BIONA-RYB1, BIONA-RYB2 и BIONA-RYB3. Эффективность антител обусловлена тем, что они связывают эпитопы, соответственно, Э1, Э2, Э3 белка Е2 оболочки вируса гепатита С. Продемонстрирована нейтрализующая активность антител на модельной системе заражения клеток человека в культуре. Показано, что использование заявляемой группы изобретений позволяет повысить надежность связывания антителами вируса гепатита С. Предложенная группа изобретений может использоваться в медицине, фармацевтической промышленности и смежных отраслях науки и техники. 8 н. и 1 з.п. ф-лы, 9 ил., 8 табл., 13 пр.
1. Способ нейтрализации вируса гепатита C путем контакта с антителами, связывающимися с эпитопами белка Е2 оболочки вируса, отличающийся тем, что контакт осуществляют, вводя в систему хотя бы одно антитело, способное взаимодействовать хотя бы с одним эпитопом, выбранным из группы Э1, Э2, Э3, в которой Э1 белка Е2 изолята Н77 вируса гепатита C представляет собой конформацию непрерывной последовательности аминокислот, заключенной в пределах участка HPEATYSRCG (SEQ ID NO: 30) и содержащей аминокислоты Ser и Arg этого участка, Э2 белка Е2 изолята Н77 вируса гепатита C представляет собой конформацию непрерывной последовательности аминокислот, заключенной в пределах участка VCGPVYCF (SEQ ID NO: 32) и содержащей аминокислоту Tyr этого участка, Э3 белка Е2 изолята Н77 вируса гепатита С представляет собой конформацию непрерывной последовательности аминокислот, заключенной в пределах участка HPEATYSRCGSGPWITP (SEQ ID NO: 34) и содержащей обе аминокислоты Ser и аминокислоту Arg этого участка и выбранное из группы RYB1, RYB2, RYB3, в которой антитело RYB1 имеет последовательность аминокислот вариабельного участка тяжелой цепи (VH) SEQ ID NO: 5 и последовательность аминокислот вариабельного участка легкой цепи (VL) SEQ ID NO: 9, антитело RYB2 имеет последовательность аминокислот вариабельного участка тяжелой цепи (VH) SEQ ID NO: 13 и последовательность аминокислот вариабельного участка легкой цепи (VL) SEQ ID NO: 17, антитело RYB3 имеет последовательность аминокислот вариабельного участка тяжелой цепи (VH) SEQ ID NO: 21 и последовательность аминокислот вариабельного участка легкой цепи (VL) SEQ ID NO: 25.
2. Полностью человеческое моноклональное антитело RYB1 с последовательностью аминокислот вариабельного участка тяжелой цепи (VH) SEQ ID NO: 5 и последовательностью аминокислот вариабельного участка легкой цепи (VL) SEQ ID NO: 9, обладающее иммунологической специфичностью к эпитопу Э1 белка Е2 изолята Н77 вируса гепатита C, представляющему собой конформацию непрерывной последовательности аминокислот, заключенной в пределах участка HPEATYSRCG (SEQ ID NO: 30) и содержащей аминокислоты Ser и Arg этого участка, и секретируемое клетками гибридной клеточной линии BIONA-RYB1, депонированной во Всероссийской Коллекции Промышленных Микроорганизмов под номером Н-142.
3. Полностью человеческое моноклональное антитело RYB2 с последовательностью аминокислот вариабельного участка тяжелой цепи (VH) SEQ ID NO: 13 и последовательностью аминокислот вариабельного участка легкой цепи (VL) SEQ ID NO: 17, обладающее иммунологической специфичностью к эпитопу Э2 белка Е2 изолята Н77 вируса гепатита C, представляющему собой конформацию непрерывной последовательности аминокислот, заключенной в пределах участка VCGPVYCF (SEQ ID NO: 32) и содержащей аминокислоту Tyr этого участка, и секретируемое клетками гибридной клеточной линии BIONA-RYB2, депонированной во Всероссийской Коллекции Промышленных Микроорганизмов под номером Н-143.
4. Полностью человеческое моноклональное антитело RYB3 с последовательностью аминокислот вариабельного участка тяжелой цепи (VH) SEQ ID NO: 21 и последовательностью аминокислот вариабельного участка легкой цепи (VL) SEQ ID NO: 25, обладающее иммунологической специфичностью к эпитопу Э3 белка Е2 изолята Н77 вируса гепатита C, представляющему собой конформацию непрерывной последовательности аминокислот, заключенной в пределах участка HPEATYSRCGSGPWITP (SEQ ID NO: 34) и содержащей обе аминокислоты Ser и аминокислоту Arg этого участка, и секретируемое клетками гибридной клеточной линии BIONA-RYB3, депонированной во Всероссийской Коллекции Промышленных Микроорганизмов под номером Н-144.
5. Композиция для нейтрализации вируса гепатита С на основе антител, обладающих иммунологической специфичностью к эпитопам Э1, Э2 и Э3 белка Е2, отличающаяся тем, что она содержит в качестве антител полностью человеческие моноклональные антитела RYB1, RYB2 и RYB3 в массовом соотношении 20-40:20-40:20-40.
6. Композиция по п.5, отличающаяся тем, что соотношение антител RYB1, RYB2 и RYB3 в композиции составляет 1:1:1.
7. Гибридная клеточная линия BIONA-RYB1, депонированная во Всероссийской Коллекции Промышленных Микроорганизмов под номером Н-142 - продуцент полностью человеческих моноклональных антител к эпитопу Э1 белка Е2 изолята Н77 вируса гепатита С, представляющему собой конформацию непрерывной последовательности аминокислот, заключенной в пределах участка HPEATYSRCG (SEQ ID NO: 30) и содержащей аминокислоты Ser и Arg этого участка.
8. Гибридная клеточная линия BIONA-RYB2, депонированная во Всероссийской Коллекции Промышленных Микроорганизмов под номером Н-143 - продуцент полностью человеческих моноклональных антител к эпитопу Э2 белка Е2 изолята Н77 вируса гепатита C, представляющему собой конформацию непрерывной последовательности аминокислот, заключенной в пределах участка VCGPVYCF (SEQ ID NO: 32) и содержащей аминокислоту Tyr этого участка.
9. Гибридная клеточная линия BIONA-RYB3, депонированная во Всероссийской Коллекции Промышленных Микроорганизмов под номером Н-144 - продуцент полностью человеческих моноклональных антител к эпитопу Э3 белка Е2 изолята Н77 вируса гепатита C, представляющему собой конформацию непрерывной последовательности аминокислот, заключенной в пределах участка HPEATYSRCGSGPWITP (SEQ ID NO: 34) и содержащей обе аминокислоты Ser и аминокислоту Arg этого участка.
| US 2012039846 A1 (UNIV LELAND STANFORD JUNIOR), 16.02.2012 | |||
| US 2012288510 A1 (UNIV MASSACHUSETTS), 15.11.2012 | |||
| АНТИТЕЛА ПРОТИВ ВИРУСА ГЕПАТИТА С | 2008 |
|
RU2462475C2 |

