Включение ссылкой
Данная заявка притязает на приоритет по предварительной заявке США с порядковым номером No.61/180755 от 22 мая 2009 года.
Область техники, к которой относится изобретение
Настоящее изобретение относится к способу поддержания и продуцирования плазмид в грамотрицательной бактерии без использования селекционного давления антибиотиком. Кроме того, изобретение относится к продуцируемым безлекарственным плазмидам, а также к составам и/или композициям, содержащим безлекарственные плазмиды, и к составам и/или композициям, содержащим белок или иммуноген, полученный с помощью безлекарственной плазмиды, а также к способам введения таких составов и/или композиций организму-хозяину. Изобретение относится к грамотрицательным бактериям, содержащим безлекарственные плазмиды.
Предшествующий уровень техники
В настоящее время нет плазмидных ДНК-векторов, которые бы считались безопасными, мощными и эффективными при применении в медицине. Присутствие генов устойчивости к антибиотикам в доставляемых плазмидах является одним из препятствий для применения современной генной терапии и ДНК-вакцин.
Плазмиды представляют собой внехромосомные молекулы ДНК, которые отделены от хромосомной ДНК и способны к репликации независимо от хромосомной ДНК (Lipps G. (editor). (2008). Plasmids: Current Research and Future Trends. Caister Academic Press. ISBN 978-1-904455-35-6). В естественной среде плазмиды обычно встречаются в бактериях, но иногда их можно обнаружить и в эукариотических организмах. Предполагается, что плазмиды представляют собой переносимые генетические элементы или репликоны, способные к автономной репликации внутри соответствующего организма-хозяина. Плазмиды являются депротеинизированной ДНК и не кодируют генов, необходимых для упаковки генетического материала для переноса к новому организму-хозяину. Таким образом, перенос плазмид от хозяина к хозяину требует прямого, механического переноса путем конъюгации или изменений в экспрессии гена в организме хозяине, что дает возможность намеренного поглощения генетического элемента путем трансформации (Lipps G. 2008).
Использование плазмидной ДНК (пДНК) для генной терапии и вакцинации представляет собой новую технологию со значительным потенциалом в здравоохранении людей и животных (Mairhofer, J. et al., Biotechnol. J., 3, 83-89, 2008). Кроме того, плазмиды служат в качестве важных инструментов в генетических и биотехнологических лабораториях, где они широко используются для умножения количества или экспрессии конкретных генов (Russell, David W.; Sambrook, Joseph (2001), Molecular cloning: a laboratory manual. Cold Spring Harbor, N.Y; Cold Spring Harbor Laboratory).
Плазмиды обеспечивают механизм горизонтального переноса гена внутри популяции микробов и, как правило, обеспечивают селективное преимущество в определенном состоянии окружающей среды. Например, плазмиды могут нести гены, которые обеспечивают устойчивость к природным антибиотикам. Также кроме антибиотиков присутствуют альтернативные маркеры. Например, белки, продуцируемые плазмидой, могут функционировать в качестве токсинов, которые также обеспечивают преимущество в определенном состоянии окружающей среды. Плазмиды также могут обеспечивать бактерии способностью фиксировать элементарный азот или разрушать трудно деградируемые органические соединения, что дает преимущество в условиях дефицита питательных веществ (Lipps G., 2008).
Для отбора с помощью генов устойчивости к антибиотикам, получают плазмиды, в которые вставлен ген, образующий белок, который делает клетки устойчивыми к конкретному антибиотику. Затем с помощью процесса, называемого трансформацией, плазмиды вставляют в бактерии. После этого бактерии подвергают воздействию определенного антибиотика. В присутствии антибиотика выживают только бактерии, которые поглощают копии плазмид, так как плазмида делает их устойчивыми к антибиотику.
Представляющие интерес гены, могут быть доставлены с помощью плазмиды, содержащей маркер устойчивости к антибиотику. Такой ген, как правило, вставляют в сайт множественного клонирования (MCS, или полилинкер). Гены устойчивости к антибиотикам экспрессируются и экспрессируемые белки выводят антибиотик из строя. Таким образом, антибиотики действуют в качестве фильтра для селекции только модифицированных бактерий. Поэтому для выделения представляющей интерес плазмиды эти бактерии могут быть выращены в больших количествах, собраны и лизированы (часто с помощью метода щелочного лизиса).
В настоящее время большой успех антибиотиков в медицине становится проблемой. Многие бактерии, включая патогены инфекционных заболеваний, уже устойчивы к антибиотикам и не могут более контролироваться конкретными антибиотиками. В области медицины людей и животных антибиотики применяли слишком часто. Кроме того, еще более значимым является тот факт, что в течение продолжительного времени антибиотики добавляли в корм животных в качестве вещества, повышающего продуктивность. Эта практика сейчас признана в основном противозаконной, но широко распространенные антибиотики придали преимущество выживания тем бактериям, которые обладали соответствующим геном устойчивости. Кроме того, гены устойчивости в бактериях часто локализованы на мобильных элементах ДНК, которые могут быть передаваться между различными видами.
В отличие от этих данных, существуют данные о том, что бактерии могут поглощать маркерные гены, что в результате приводит к появлению патогенов, против которых прописываемые в настоящее время антибиотики не эффективны. Существует перенос генов в микроорганизмы окружающей среды, например, патогенов (Murphy, D.В., Epstein, S.L., Guidance for Industry: Guidance for human somatic cell therapy and gene therapy, Food and Drug Administration, Rockville 1998). Другим вопросом безопасности является возможная интеграция гена устойчивости к антибиотику в человеческую хромосому (Smith, H.A., Klinman, D.M., Curr. Opin. Biotechnol. 12, 299-203, 2001).
Кроме того, такие гены оказывают значительное влияние на процесс продуцирования плазмиды, поскольку конститутивная экспрессия этих генов накладывает необязательную метаболическую нагрузку на бактериальные клетки-хозяева (Cranenburgh, R.M., et al., Nucleic Acids Res. 2001, 29, e26; Rozkov, A., et al., Enzyme Microb. Technol. 2006, 39, 47-50). Уменьшение размера плазмид путем исключения этих генов приведет к повышению стабильности и увеличению выхода пДНК, получаемой в процессе ферментации (Smith, M.A., et al., Can. J. Microbiol. 1998, 44, 351-355).
Таким образом, исходя из существующего уровня техники, существует безусловная потребность избегать применения генов устойчивости к антибиотикам в конечном (коммерческом) продукте (вакцине в виде депротеинизированной ДНК), поскольку согласно текущим рекомендациям надзорных органов существует потенциальный риск их получения населением/потребителями. Управление по контролю за продуктами и лекарствами (FDA) и Всемирная Организация Здравоохранения (ВОЗ) регулируют применение маркеров устойчивости к антибиотикам для гарантии качества ДНК-вакцин и для профилактики инфекционных заболеваний. Аналогично, директива Европейского союза об осмотрительном введении в окружающую среду генетически измененных организмов, которая действует с 2002 года, требует «постепенной отмены использования маркеров устойчивости к антибиотикам в генетически модифицированных организмах, которые могут нанести вред здоровью человека или окружающей среде». Недостатки традиционных маркеров стали очевидны даже в исследовательской практике. Например, существует потребность в получении систем доставки без антибиотиков для коммерческих применений технологии бактофекции. Технология бактофекции представляет собой доставку плазмидной ДНК в эукариотические клетки с помощью инвазивных бактерий. Кроме того, существует техническая потребность в уменьшении необязательных метаболических затрат во время выращивания, что позволит достигать более высоких значений OD и более высокого выхода плазмидной ДНК.
Для того чтобы решить проблемы, касающиеся распространения генов устойчивости к антибиотикам в кишечных бактериях человека, были разработаны альтернативные стратегии селекции, включая ауксотрофную комплементацию, титрацию репрессора, схемы селекции противоядия/яда на белковой основе и использование маркеров селекции на основе РНК (см. Williams J.A. et al., Plasmid DNA vaccine vector design: Impact on efficacy, safety and upstream production, Biotechnol Adv (2009), doi:10.1016/j.biotechadv.l009.02.003).
Cranenburgh, R.M. et al. сообщили о создании двух новых штаммов Escherichia coli (Dt1lacdapD и DH1lacP2dapD), которые облегчают селекцию без антибиотика и стабильное поддержание рекомбинантных плазмид в комплексной среде. Они содержат жизненно-важный хромосомный ген, dapD, под контролем оператора/промотора lac (Cranenburgh, R. M., et al., 2001). Если не добавлять IPTG (который индуцирует экспрессию dapD) или DAP, эти клетки лизируются, однако при трансформации штаммов мультикопийной плазмидой, содержащей оператор lac, последний конкурентно титрует репрессор LacI и делает возможной экспрессию dapD с промотора lac. Таким образом, с помощью селекции титрацией репрессора трансформанты могут быть просто выделены и размножены на любой среде по их способности к росту. Для плазмиды не требуются гены устойчивости к антибиотикам или другие последовательности, экспрессирующие белки, и антибиотики не являются обязательными для селекции плазмид.
Mairhofer et al. недавно занимались разработкой штаммов бактериальных клеток-хозяев, которые служат для селекции и поддержания плазмид без применения в плазмиде каких-либо маркеров селекции или любых других дополнительных последовательностей. Несколько бактериальных штаммов модифицировали так, чтобы ингибитор репликации РНК I плазмиды мог подавлять трансляцию значимых для роста генов посредством реакции РНК-антисмысловая РНК (Mairhofer, J. et al, Biotechnol. J., 3, 83-89, 2008). Жизненно-важный ген (murA) модифицировали так, чтобы белок-репрессор (tetR) мог препятствовать его экспрессии (Mairhofer, J. et al., 2008). Только в присутствии плазмиды и, следовательно, РНК I, tetR выключался, а murA экспрессировался. Авторы настоящего изобретения сообщали о том, что различные коммерчески доступные плазмиды можно селектировать с помощью различных модифицированных штаммов Escherichia coli. Кроме того, авторы разработали минимальную плазмиду, лишенную какого-либо маркера селекции.
Цитата или указание любого документа в данной заявке не является признанием того, что такой документ доступен в качестве предшествующего уровня техники по отношению к настоящему изобретению.
Сущность изобретения
Предлагаются плазмиды без устойчивости к антибиотикам, а также грамотрицательные бактерии, содержащие плазмиды без устойчивости к антибиотикам. Также предлагаются композиции, содержащие плазмиды без устойчивости к антибиотикам, содержащие гетерологичный ген, кодирующий иммуноген или белок, или предлагаются композиции, содержащие иммуноген или экспрессирующие белок с помощью плазмид без устойчивости к антибиотикам. Грамотрицательные бактерии конструировали так, чтобы они содержали один или несколько гетерологичных полину клеотидов в несущественном участке бактериальной хромосомы. Плазмиды без устойчивости к антибиотикам содержат полинуклеотид, кодирующий репрессор, который регулирует экспрессию гетерологичного полинуклеотида на бактериальной хромосоме. Плазмиды без устойчивости к антибиотикам могут дополнительно содержать один или несколько полинуклеотидов, кодирующих иммуноген или белок.
Способы по изобретению включают способы получения плазмид без устойчивости к антибиотикам, а также способы переноса чужеродных генов в клетки млекопитающих с помощью плазмид без устойчивости к антибиотикам. Эти и другие воплощения описаны в следующем подробном описании или являются очевидными из него и охвачены им.
Краткое описание чертежей
Следующее подробное описание, данное в качестве примера, но не предназначенное для ограничения изобретения конкретными описанными воплощениями, может быть истолковано в сочетании с следующими чертежами, включенными в настоящий документ ссылкой, в которых:
Фигура 1 описывает первый компонент безлекарственной модели, который заключается в клонировании гена sacB, кодирующего левансахаразу в грамотрицательных бактериях, который приводит к быстрой смерти трансформированных бактерий при посеве на среду, содержащую сахарозу.
Фигура 2 описывает второй и третий компоненты безлекарственной модели или модели без устойчивости к антибиотику, которые представляют собой область иммунитета фага λ, где cI кодирует репрессор cI; его кодирует белок Cro; N кодирует транскрипционный анти-терминатор; cII кодирует активатор cI; cIII кодирует протеазный ингибитор cIII; OL 1, 2 и 3 и OR 1, 2 и 3 являются операторами; PL и PR являются правосторонними и левосторонними промоторами; PRE является промотором для первоначального синтеза репрессора; PRM является промотором для поддержания репрессора.
Фигура 3 описывает теоретический обзор «безлекарственной» модели, где активность репрессора cI, экспрессируемого с плазмиды pDL1 (англ. DrugLess Plasmid), ингибирует транскрипцию токсичного продукта гена sacB, управляемую промотором Pr, которые находятся на хромосоме клетки-хозяина, и где выживаемость клеток-хозяев Е. coli в присутствии сахарозы гарантирована достаточным уровнем экспрессии плазмидой белка-репрессора cI из фага λ.
Фигура 4 описывает «чувствительность к сахарозе» инактивированной системы, где температурный переход на 42°С ингибирует активность связывания репрессора cI с промотором Pr из фага λ и делает клетки-хозяева чувствительными к сахарозе.
Фигура 5 демонстрирует комбинацию штамма клеток-хозяев с плазмидой pDL1 без устойчивости к антибиотику, полученную трансформацией pDL1, и роста в среде, содержащей сахарозу, которая приводит к получению безлекарственной плазмиды.
Фигуры 6А-6С иллюстрируют план эксперимента. Фиг.6А дает общий план; ФИГ 6В демонстрирует родительский штамм ΔpurN (или ΔedA), который устойчив к канамицину и конститутивно экспрессирует SacB+, что придает ему чувствительность к сахарозе, при инкубации клеток при 30°С-37°С. Когда родительский штамм ΔpurN (или ΔedA) трансформируют плазмидой рРВ829 или рРВ838, то эти клетки приобретают способность выживать и расти в присутствии сахарозы, при их инкубации при 30-37°С; ФИГ 6С демонстрирует родительский штамм ΔpurN (или ΔedA), трансформированный либо плазмидой рРВ829, либо рРВ838 (плазмиды с cI), который способен расти в среде LB, содержащей либо канамицин, либо сахарозу, либо хлорамфеникол. Переход температуры с 30-37°С на 42°С приводит к смерти клеток при посеве в присутствии сахарозы.
Фигура 7 описывает две плазмиды (рРВ829 и рРВ838), маркированные генами, кодирующими хлорамфениколацетилтрансферазу (CatR и ген λcI. рВ838 представляет собой производное pMCS5, в которой ген хлорамфениколацетилтрансферазы (cat) заменяет ген устойчивости к ампициллину. Эта плазмида содержит ген cI, расположенный под контролем слабого промотора гена устойчивости к канамицину (Р1). рРВ829 представляет собой производное плазмиды pVR1012, содержащей ген cat. Плазмиды содержат ген cI, который расположен под контролем слабого промотора (Р1). При введении в родительский штамм Е. coli ΔpurN (или ΔedA), данные плазмиды сделают возможным рост клеток в присутствии сахарозы. Хлорамфеникол позволяет продемонстрировать достоверность модели.
Фигуры 8А-Е иллюстрируют создание штаммов клеток-хозяев. Фиг.8А и 8В иллюстрируют получение кассеты λPr::sacB Ωkan, вставляемой с помощью аллельной замены гена edA или purN, соответственно, в хромосоме Е. coli. Фигура 8С иллюстрирует создание полной кассеты sacB (ΔpurNΩλPr::sacB cat) с помощью ПЦР и объединяющего ПЦР, используемой для аллельной замены гена edA в клетках-хозяевах ΔpurNΩλPr::sacB Km штамма Е. coli для получения штамма Е. coli с удвоенной кассетой sacB.
На Фигурах 9А, 9В и 9С показано применение ПЦР для демонстрации создания штамма клеток-хозяев, а именно для демонстрации делеции edA и purN в Е. coli. Фиг.9А частично иллюстрирует два продукта ПЦР (соответственно 151 и 729 п.о.), которые очищали и использовали в качестве матрицы во второй стадии ПЦР с праймерами РВ 1186 и РВИ 89 и с ДНК-полимеразой «Phusion». Фиг.9 В иллюстрирует контрольную ПЦР, демонстрирующую введение кассеты λPr::sacB+ΩKan в хромосому Е. coli. Фиг.9С иллюстрирует корректность вставки кассеты λPr::sacB+ΩKan в локус с помощью аллельной замены генов edA и/или purN. Фигура 9D иллюстрирует использование ПЦР для демонстрации создания штамма клеток-хозяев, несущих кассету с дублированным sacB, а именно путем делеции обоих генов, edA и purN в Е. coli с помощью аллельной замены. Контрольный ПЦР демонстрирует, что обе кассеты, λPr::sacB+ΩKan и λPr::sacB+ΩCat, корректно введены в определенный локус на хромосоме Е. coli, что подтверждено секвенированием. На Фигуре 9D показаны: Дорожки 1: BJ5183 дт; 2: BJ5138 ΔpurN; 3: BJ5138 ΔedA; 4: BJ5183 ΔpurN ΔedA, не разведено, 1 мкл; 5: BJ5183 ΔpurN ΔedA, разведено 1/10; 6: Вода, 0 п.о. (контроль); А: BJ5183 дт; В: BJ5138 ΔpurN; С: BJ5138 ΔedA; D: BJ5183 ΔpurN ΔedA, не разведено, 3 мкл; Е: BJ5183 ΔpurN ΔedA, не разведено, 1 мкл; F: BJ5183 ΔpurN ΔedA разведено 1/10; G: BJ5183 ΔpurN ΔedA, разведено, 1/50; Н: Вода, 0 п.о.(контроль).
Фигура 10А демонстрирует, что штамм клеток-хозяев (ΔpurN λPr::sacB+Ωkan) является высокочувствительным (отсутствие роста) к сахарозе, что клетки, трансформированные плазмидой рРВ829 или рРВ838 хорошо растут в присутствии сахарозы при 30°С, что экспрессия гена sacB полностью подавлялась продуктом гена cI, синтезированным с каждой из плазмид с cI, и что поддержание плазмиды было на 100% эффективным, что было ясно продемонстрировано с помощью контрольного эксперимента, осуществленного параллельно с помощью аналогичных клеток, которые выращивали в присутствии хлорамфеникола. Повышение температуры до 42°С вызывало инактивацию продукта гена cI и смерть клеток, посеянных на LB-агар, содержащий либо сахарозу, либо хлорамфеникол. Эксперименты по части надежности поддержания данной плазмиды осуществляли после двух последовательных пассажей клеток, растущих в присутствии сахарозы или хлорамфеникола. Фигуры 10 В-С демонстрирует, что штамм Е. coli, несущий кассеты с дублированным sacB (ΔpurN λPr::sacB+Ωkan ΔedA λPr::sacB+ΩCat), является высокочувствительным (отсутствие роста) в присутствии сахарозы при инкубации при 30°С и 42°С, хотя при этом встречаются спонтанные мутанты посеянных на среде с сахарозой штаммов Е. coli, содержащих одну кассету sacB. Фиг.10С также демонстрирует самую низкую концентрацию сахарозы, необходимую для штамма Е. coli, содержащего две кассеты sacB, по сравнению со штаммом Е. coli, содержащим одну кассету sacB (от 10% до 2% конечной концентрации).
Фигура 11 иллюстрирует стабильность плазмид рРВ829 и рРВ838 в растущих клетках.
Фигура 12А демонстрирует отсутствие спонтанной мутации в кассете λPr::sacB+, содержащей один sacB, при 30°С. Фигуры 12В-С демонстрируют, что присутствие двух кассет sacB (а не одной) придает большую надежность в отношении чувствительности к сахарозе (оставаясь оптимальной при 2% сахарозы) при температурах в интервале от 30°С до 37°С. Наличие одной кассеты sacB на хромосоме Е. coli было менее стабильным при концентрации сахарозы в интервале от 2% до 4% при аналогичных температурах. В данном анализе с посевом на чашку подтвердили, что частота мутаций под влиянием сахарозы штамма Е. coli с двумя кассетами sacB была не детектируемой при самой низкой концентрации сахарозы (2%) при инкубации при 37°С, в то время как частота мутаций штамма Е. coli с одной кассетой sacB колебалась в интервале от 3,8×10-6 до 5×10-5. Ряд независимых экспериментов показал, что частота мутаций штамма Е. coli с двумя кассетами sacB составила около 5×10-10 при инкубации при 37°С в присутствии 2% сахарозы.
Фигуры 13А-В демонстрирует экспериментальные процедуры, дающие возможность скрининга трансформированных клеток с помощью плазмид cI, маркированных хлорамфениколом (Cat) без использования антибиотика в качестве фактора селекционного давления на чашках с агаром.
Фигуры 14А-С иллюстрируют эффективность экспериментальной процедуры для селекции трансформированных клеток только с плазмидой с cI (рРВ829 и рРВ838) без применения антибиотика в качестве фактора селекционного давления.
Фигуры 15А-В иллюстрируют стабильность и выход плазмид рРВ829 и рРВ838 в растущих клетках E.coli, которые содержат одну копию кассеты sacB. Фигуры 15С-Е иллюстрируют стабильность и выход плазмиды рРВ885 без cat в обоих штаммах Е. coli с одной и с двумя кассетами sacB (ΔpurN λPr::sacB+Ωkan и ΔpurN λPr::sacB+Ωkan ΔedA λPr::sacB+Ωkancat, соответственно). Фигура 15F иллюстрирует эффективность трансфекции плазмид (экспрессия GFP) в клетках СНО, плазмиды без устойчивости к антибиотику (рРВ896/сахароза в качестве фактора селекционного давления) и плазмиды с устойчивостью к антибиотику (pCG105/Cat в качестве фактора селекционного давления). Не наблюдали никакого очевидного различия на примере экспрессии белка GFP между этими двумя селективными давлениями (Сахароза против Cat в качестве антибиотика).
Фигура 16 демонстрирует карту плазмиды pCRblunt + РВ1184-1185.
Фигура 17 демонстрирует рестрикционную карту и характеристики рРВ828.
Фигура 18 демонстрирует рестрикционную карту и характеристики рРВ829.
Фигура 19 демонстрирует рестрикционную карту и характеристики рРВ838.
Фигура 20 демонстрирует плазмидную карту и характеристики рРВ844.
Фигура 21 демонстрирует плазмидную карту и характеристики рРВ845.
Фигура 22 демонстрирует плазмидную карту и характеристики рРВ846.
Фигура 23 демонстрирует плазмидную карту и характеристики рРВ847.
Фигура 24 демонстрирует плазмидную карту и характеристики рРВ885. На карте элементов рРВ885 показаны: две CDS: репрессор cI: 324-1037; AP(R): 2321-2990 (комплементарная цепь), исходное положение (комплементарная цепь 1626..2483); две последовательности с точкой начала репликации: последовательность точки начала репликации из pUC: 1500-2173 (комплементарная цепь), исходное положение (комплементарная цепь 805..1478); последовательность точки начала репликации из f1: 2991-3301 (исходное положение 2615-3053).
Фигура 25 демонстрирует плазмидную карту рРВ896. На карте элементов рРВ896 показаны: три CDS: GFP: 1675-2397 (исходное положение 1676…2398); репресор cI: 4089-4802; Cm(R): 4828-5484 (комплементарная цепь), комплементарная цепь (950-1606); один интрон: интрон А: 805-1625 (исходное положение 805-1625); один эукариотический промотор: промотор CMV: 1-683; одна последовательность точки начала репликации: 3133-3640 (исходное положение 2425..2932); один терминатор: терминатор транскрипции BGH/PolyA: 2405-2951 (исходное положение 1697..2243); один 5' UTR: CMV IE 5' UTR: 684-804 (исходное положение 684..804).
Фигура 26 демонстрирует выравнивание последовательностей белков репрессора cI и процент идентичности последовательности.
Фигура 27 демонстрирует выравнивание последовательностей белков sacB и процент идентичности последовательности.
Фигура 28 представляет собой таблицу, демонстрирующую SEQ ID NO, соответствующие полинуклеотидам и белкам.
Фигура 29 демонстрирует выравнивание последовательностей полинуклеотидов, кодирующих белки репрессора cI, и процент идентичности на уровне нуклеотидов.
Фигура 30 демонстрирует выравнивание последовательностей полинуклеотидов, кодирующих белки sacB, и процент идентичности на уровне нуклеотидов.
Фигура 31 демонстрирует последовательности промотора Р1, нативного промотора cI, промотора λPr, промотора λPr+5' UTR+ген sacB, промотора P1+ген cI, и нативного промотора cI+ген cI.
Подробное описание
Следует отметить, что в данном описании и, особенно, в формуле изобретения термины, такие как «содержит», «содержащий» и т.п. могут иметь значение, которое придается термину в патентном законе США; например, они могут означать «включает», «включенный», «включающий» и т.п.; и что термины, такие как «состоящий в основном» и «состоит в основном» имеют значение приписанное им в патентном законе США, например, они позволяют элементам не явным образом упоминаться, но исключать элементы, которые обнаружены в предшествующем уровне техники или которые влияют на основную или новую характеристику данного изобретения.
Если иное не определено, все технические и научные термины, использованные здесь, имеют тот же смысл, который вкладывает в них обычный специалист в области, к которой принадлежит данное изобретение. Термины в единственном числе включают их значения во множественном числе, если контекстом ясно не указано иное. Аналогичным образом, слово «или» предназначено для включения «и» если контекст с очевидностью не указывает на иное.
Термин «животное» предназначен для обозначения млекопитающих, птиц и тому подобных. Животное или организм-хозяин включает млекопитающих и человека. Животное может быть выбрано из группы, состоящей из лошадиных (пример, лошадь), псовых (пример, собака, волк, лиса, койот, шакал), кошачьих (пример, лев, тигр, домашняя кошка, дикая кошка, другие большие кошки, и другие животные семейства кошачьих, включая гепарда и рысь), овец (пример, овца), крупного рогатого скота (пример, корова), свиней (пример, свинья), птиц (пример, курица, утка, гусь, индейка, перепел, фазан, попугай, зяблик, ястреб, ворона, страус, эму и казуар), приматов (пример, древесный примат, долгопят, обезьяна, гиббон, человекообразные обезьяны), и рыб. Термин «животное» также включает индивидуальное животное во всех стадиях развития, включая стадии эмбриона и плода.
Термины «белок», «пептид», «полипептид» и «полипептидный фрагмент» используются взаимозаменяемо для обозначения полимеров любой длины из аминокислотных остатков. Полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты или аналоги аминокислот, и он может прерываться химическими компонентами, отличными от аминокислот. Термины также охватывают аминокислотный полимер, который был модифицирован в естественной среде или в результате экспериментального вмешательства; например, путем образования дисульфидной связи, гликозилирования, липидирования, ацетилирования, фосфорилирования или с помощью любой другой манипуляции или модификации, такой как конъюгация с помощью мечения или с помощью биоактивного компонента.
Термин «нуклеиновая кислота» или «полинуклеотид» используются взаимозаменяемо и обозначают РНК или ДНК, линейные или разветвленные, одноцепочечные или двухцепочечные или их гибриды. Термин также охватывает РНК/ДНК гибриды. Далее представлены частные примеры полинуклеотидов: ген или фрагмент гена, экзоны, интроны, мРНК, тРНК, рРНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенная ДНК любой последовательности, выделенная РНК любой последовательности, нуклеотидные зонды и праймеры. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и аналоги нуклеотидов, урацил, другие сахара и группы присоединения, такие как фторрибоза и тиолат, а также ответвления нуклеотидов. Последовательность нуклеотидов может быть дополнительно модифицирована после полимеризации, как, например, путем конъюгации с компонентом мечения. Другие типы модификаций, включенные в данное определение, являются кэпированием, заменой одного или нескольких природных нуклеотидов на аналог и введением посредством прикрепления полинуклеотида к белкам, ионам металлов, компонентам мечения, другим полинуклеотидам и к твердой подложке. Полинуклеотиды могут быть получены путем химического синтеза или могут быть выделены из микроорганизма.
Термины «безлекарственная плазмида» или «плазмида без устойчивости к антибиотику» используются взаимозаменяемо и обозначают ДНК-плазмиду, которая не содержит гена для селекции с помощью антибиотика.
Термин «ген» используется широко для обозначения какого-либо сегмента полинуклеотида, ассоциированного с биологической функцией. Таким образом, гены включают интроны и экзоны, как в геномной последовательности, или только кодирующие последовательности, как в кДНК, и/или регуляторные последовательности, необходимые для экспрессии. Например, ген также относится к фрагменту нуклеиновой кислоты, которая экспрессирует мРНК или функциональную РНК, или кодирует конкретный белок, и которая включает регуляторные последовательности.
Объект изобретения, раскрытый в настоящем документе, относится к новому способу для получения ДНК-векторов на основе бактериальных плазмид путем преодоления применения генов устойчивости к антибиотикам для получения безопасных вакцин и иммуногенных композиций.
Объект изобретения, раскрытый в настоящем документе, демонстрирует новую модель для поддержания высокого количества копий плазмид в грамотрицательной бактерии-хозяине, причем новая модель основана на трех компонентах. Первый компонент представляет собой грамотрицательную бактерию, которая экспрессирует продукт, токсичный для бактерии в определенных условиях культивации, где токсичный ген вставлен в несущественный участок бактериальной хромосомы. Вторым компонентом является ген, присутствующий на хромосоме грамотрицательной бактерии, где ген кодирует токсичный продукт под контролем конститутивного промотора, который может подвергаться сильной регулировке. Третьим компонентом является экспрессия на плазмиде конкретного репрессора, которую регулирует промотор, функционально связанный с токсичным геном на хромосоме.
Грамотрицательные бактерии, рассматриваемые в настоящем изобретении, включают в частности Avibacterium, Brucella, Escherichia coli, Haemophilus (например, Haemophilus suis), Salmonella (например. Salmonella enteridis, Salmonella typhimurium, Salmonella infantis), Shigella, Pasteurella, и Rimeirella.
В одном воплощении, токсичный ген представляет собой структурный ген sacB, который кодирует левансахаразу. В другом воплощении, токсичный ген представляет собой структурный ген sacB из Bacillus subtilis, который кодирует левансахаразу Bacillus subtilis. Экспрессия sacB в его естественном окружении является безопасной для грамположительной бактерии, но при экспрессии в грамотрицательной бактерии приводит к быстрой смерти трансформированной бактерии при посеве на среду с сахарозой.
В одном аспекте в настоящем изобретении предлагается белок sacB (левансахараза). В другом аспекте в настоящем изобретении предлагается белок sacB, содержащий последовательность, представленную в SEQ ID NO:4, 60, 62, 64, 66, 68, 70, 72 или 74, и ее вариант или фрагмент. В другом аспекте в настоящем изобретении предлагается белок sacB, имеющий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, содержащим последовательность, представленную в SEQ ID NO:4, 60, 62, 64, 66, 68, 70, 72 или 74. Еще в одном аспекте, в настоящем изобретении предлагаются фрагменты и варианты белка sacB, идентифицированные выше (SEQ ID NO:4, 60, 62, 64, 66, 68, 70, 72 или 74), которые легко могут быть получены специалистом в данной области с помощью хорошо известных методов молекулярной биологии. Варианты являются гомологичными полипептидами, содержащими аминокислотную последовательность с идентичностью, составляющей, по меньшей мере, около 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99% по отношению к полипептидам по изобретению, конкретно, к аминокислотной последовательности, представленной в SEQ ID NO:4, 60, 62, 64, 66, 68, 70, 72 или 74.
В другом аспекте в настоящем изобретении предлагается полинуклеотид, такой как ген sacB, кодирующий белок sacB (левансахаразу), например, полинуклеотид, кодирующий белок sacB, содержащий последовательность, представленную в SEQ ID NO:4, 60, 62, 64, 66, 68, 70, 72 или 74. Еще в одном аспекте настоящего изобретения предлагается полинуклеотид, кодирующий белок sacB, имеющий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% or 99% идентичности последовательности с полипептидом, содержащим последовательность, представленную в SEQ ID NO:4, 60, 62, 64, 66, 68, 70, 72 или 74, или консервативный вариант, аллельный вариант, гомолог или фрагмент, содержащий, по меньшей мере, восемь или, по меньшей мере, десять последовательно расположенных аминокислот одного из этих полипептидов, или комбинация этих полипептидов. В другом аспекте, в настоящем изобретении предлагается полинуклеотид, содержащий нуклеотидную последовательность, представленную в SEQ ID NO:3, 61, 63, 65, 67, 69, 71, или 73, или ее вариант. В еще одном аспекте, в настоящем изобретении предлагается полинуклеотид, обладающий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичностью последовательности по отношению к полинуклеотиду, содержащему последовательность, изложенную в SEQ ID NO:3, 61, 63, 65, 67, 69, 71, или 73, или к его варианту.
Другие токсичные гены, которые можно применить в настоящем изобретении, включают в частности гены Listeria или Staphylococcus. Токсичным геном может быть ДНК, которая экспрессирует токсичный продукт гена (токсичный белок или РНК), или может быть токсичной сама по себе. Примеры таких токсичных продуктов генов хорошо известны в данной области и включают, в частности, эндонуклеазы рестрикции (например, Dpnl), CcdA/CcdB (Maki S. et al. J. Mol. Biol. 256: 473-482, 1996) и гены, которые убивают организм-хозяин в отсутствии функции подавления, например, kicB. В ином случае, токсичный ген можно селектировать in vitro, например, с помощью сайта рестрикции.
При использовании в данном документе термин «гомолог» включает ортологи, аналоги и паралоги. Аналоги, ортологи и паралоги полипептида дикого типа могут отличаться от полипептида дикого типа посттрансляционными модификациями, различиями в аминокислотной последовательности, или и тем и другим. В частности, гомологи изобретения будут, как правило, демонстрировать, по меньшей мере, 80-85%, 85-90%, 90-95%, или 95%, 96%, 97%, 98%, 99% идентичности последовательности, со всем или с частью полипептида дикого типа или с полинуклеотидной последовательностью, и будут демонстрировать похожие функции.
«Варианты» предназначены для обозначения по существу похожих последовательностей. Для полинуклеотидов, вариант содержит делецию и/или вставку одного или нескольких нуклеотидов в одном или нескольких участках внутри нативного полинуклеотида и/или замену одного или нескольких нуклеотидов в одном или нескольких участках в нативном полинуклеотиде. При использовании в настоящем документе, «нативный» полинуклеотид или полипептид содержит, соответственно, природную нуклеотидную или аминокислотную последовательность. Варианты конкретного полинуклеотида по изобретению (т.е. эталонного полинуклеотида) также могут оцениваться путем сравнения процента идентичности последовательности между полипептидом, кодируемым вариантным полинуклеотидом, и полипептидом, кодируемым эталонным полинуклеотидом. «Вариантный» белок предназначен для обозначения белка, выделенного из нативного белка путем делеции или вставки одной или нескольких аминокислот по одному или нескольким участкам в нативном белке, и/или путем замены одной или нескольких аминокислот по одному или нескольким участкам в нативном белке. Вариантные белки, охваченные настоящим изобретением, являются биологически активными, то есть они обладают способностью вызывать иммунный ответ.
Варианты включают аллельные варианты. Термин «аллельный вариант» относится к полинуклеотиду или полипептиду, содержащему полиморфизмы, которые приводят к изменениям в аминокислотных последовательностях белка, и которые существуют в естественной популяции (например, вирусные виды или разновидности). Такие естественные аллельные вариации, могут, как правило, приводить к 1-5% дисперсии в полинуклеотиде или полипептиде. Аллельные варианты могут быть идентифицированы секвенированием интересующей последовательности нуклеиновой кислоты во множестве различных видов, что может быть легко осуществлено при использовании гибридизационных зондов для идентификации одинаковых генных локусов в этих видах. Любая и все такие нуклеотидные вариации и обусловленные ими аминокислотные полифорфизмы или вариации, которые являются результатом природных аллельных вариаций, и которые не изменяют функциональную активность интересующего гена, попадают в рамки данного изобретения.
Термин «консервативная вариация» обозначает замену аминокислотного остатка другим похожим остатком, или замену нуклеотида в последовательности нуклеиновой кислоты, так чтобы кодируемый аминокислотный остаток не изменялся или был бы другим биологически похожим остатком. В этом отношении, в особенности предпочтительными заменами будут, как правило, консервативные по своей природе замены, описанные выше.
Полинуклеотиды изобретения включают последовательности, которые являются вырожденными из-за вырожденности генетического кода, например, из-за оптимизации частоты использования кодонов для конкретного хозяина. При использовании в данном документе «оптимизированный» относится к полинуклеотиду, который изменен с помощью генной нженерии для повышения его экспрессии в данных видах. Для обеспечения оптимизированных полинуклеотидов, кодирующих полипептиды по настоящему изобретению, последовательности ДНК полипептидов могут быть модифицированы для 1) включения кодонов, которые предпочтительно используются в высоко экспрессирующихся генах в конкретных видах; 2) изменения содержания А+Т или G+C в составе нуклеотидных оснований, чтобы оно совпадало с по существу обнаруженным в указанных видах; 3) для образования инициирующей последовательности указанных видов; или 4) для исключения последовательностей, которые вызывают дестабилизацию, некорректное полиаденилирование, деградацию и терминацию РНК, или таких, которые образуют шпилечную вторичную структуру или сайты сплайсинга РНК. Повышенная экспрессия полипептида по настоящему изобретению, такого как белок sacB или белок-репрессор cI в указанных видах может достигаться путем применения распределения частоты используемых кодонов у эукариотов и прокариотов или в конкретных видах. Термин «частота предпочтительного использования кодонов» относится к предпочтению, демонстрируемому конкретной клеткой-хозяином в применении нуклеотидных кодонов для определения данной аминокислоты. Существуют 20 аминокислот, большая часть которых определена более чем одним кодоном. Следовательно, все вырожденные нуклеотидные последовательности включены в описание при условии, что аминокислотная последовательность полипептида, кодируемая нуклеотидной последовательностью, является функционально неизмененной.
«Идентичность» в отношении последовательностей может относится к числу позиций с идентичными нуклеотидами или аминокислотами разделенное на число нуклеотидов или аминокислот в более короткой из двух последовательностей где выравнивание двух последовательностей может быть определено в соответствии с алгоритмом Вилбура и Липмана (Wilbur and Lipman). Когда последовательности РНК называются похожими, или обладающими степенью идентичности последовательности или гомологией с последовательностью ДНК, тимидин (Т) в последовательности ДНК рассматривается как равный урацилу (U) в последовательности РНК. Таким образом, последовательности РНК находятся в пределах объема притязаний изобретения и могут быть получены из последовательностей ДНК, если тимидин (Т) в последовательностях ДНК рассматривается как равные урацилу (U) в последовательностях РНК. Идентичность последовательности или сходство последовательности двух аминокислотных последовательностей, или идентичность последовательности двух нуклеотидных последовательностей может быть определена с помощью пакета программного обеспечения «Vector NTI» (Invitrogen, 1600 Faraday Ave., Карлсбад, Калифорния).
В другом воплощении, промотор, который функционально связан с токсичным геном по настоящему изобретению, представляет собой промотор из фага λ. Фаг λ является умеренным фагом, который обитает в Е. coli. После того как фаг попадает внутрь своего хозяина, он может интегрироваться в ДНК хозяина. В этом состоянии фаг λ называют профагом и он становится резидентом внутри генома хозяина, не вызывая большого вреда хозяину. Таким образом, профаг дублируется при каждом клеточном делении организма-хозяина. ДНК профага, которая экспрессируется в данном состоянии, кодирует белки, которые следят за признаками стресса в клетке-хозяине. Стресс может быть результатом истощения, ядов (типа антибиотиков), и других факторов, которые могут повредить или разрушить организм-хозяин. При обнаружении стрессовых условий, профаг снова становится активным, вырезает сам себя из ДНК клетки-хозяина и входит в свой литический цикл. Реактивированный фаг разбирает ДНК хозяина и перепрограммирует его белковый синтез для продуцирования множества копий новых фагов. Когда все ресурсы организма-хозяина истощены из-за сборки новых фагов, клетка лизируется (клеточная мембрана разрушается), и высвобождаются новые фаги.
Генная система репрессора фага лямбда состоит из (слева направо на хромосоме): гена cI, OR3, OR2, OR1, гена его, ген cI кодирует репрессор λ («белок-репрессор cI»). Участок генома, который кодирует белок-репрессор cI, известен как участок, связанный с иммунитетом. Белок-репрессор cI одновременно является позитивным и негативным регулятором транскрипции генов. Он позволяет фагу λ оставаться «латентным» на хромосоме его бактерии-хозяина. Лизогенное состояние бактериофага поддерживается путем связывания регуляторного белка cI с операторами OR (правый оператор) и OL (левый оператор) из промоторов λ, Pl и Pr, соответственно, предотвращая транскрипцию белков, необходимых для литической фазы. Бактериальные клетки, несущие лизогенный фаг λ, обладают иммунитетом по отношению к следующей инфекции фага λ. Белок-репрессор cI ингибирует литическое развитие любых дополнительных инфицирующих фаговых частиц.
В одном аспекте воплощения, промотор содержит полинуклеотид, имеющий последовательность, представленную в SEQ ID NO:5. В другом аспекте, промотор содержит полинуклеотид, имеющий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полинуклеотидом, имеющим последовательность, представленную в SEQ ID NO:5.
В одном воплощении, белок-репрессор по настоящему изобретению представляет собой белок-репрессор cI, такой как репрессор с1857, который является термочувствительным. Фаг λ, несущий с1857 в качестве лизогена, будет расти при температуре ниже 39°С, но при повышении температуры будет индуцировать литический рост. При 30°С, белок-репрессор cI активен и связывается с правым и левым операторами инфицирующего фага. Это предотвращает транскрипцию любых фаговых белков и таким образом предотвращается лизис. Однако при 42°С, репрессор cI инактивируется и не может связываться с операторами промоторов.
В одном аспекте в настоящем изобретении предлагается белок-репрессор cI. В другом аспекте в настоящем изобретении предлагается белок-репрессор cI, содержащий последовательность, представленную в SEQ ID NO:2, 44, 46, 48, 50, 52, 54, 56, или 58, и ее вариант или фрагмент. В другом аспекте в настоящем изобретении предлагается белок-репрессор cI, имеющий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, содержащим последовательность, представленную в SEQ ID NO:2, 44, 46, 48, 50, 52, 54, 56, или 58. Еще в одном аспекте, в настоящем изобретении предлагаются фрагменты и варианты белка-репрессора cI, идентифицированные выше (SEQ ID NO:2, 44, 46, 48, 50, 52, 54, 56, или 58), которые легко могут быть получены специалистом в данной области с помощью хорошо известных методов молекулярной биологии. Варианты являются гомологичными полипептидами, содержащими аминокислотную последовательность с идентичностью, составляющей, по меньшей мере, около 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99% по отношению к полипептидам по изобретению, конкретно, к аминокислотной последовательности, представленной в SEQ ID NO:2, 44, 46, 48, 50, 52, 54, 56, или 58.
В другом аспекте, в настоящем изобретении предлагается полинуклеотид, такой как ген cI, кодирующий белок-репрессор cI, такой как полинуклеотид, кодирующий белок-репрессор cI, содержащий последовательность, представленную в SEQ ID NO:2, 44, 46, 48, 50, 52, 54, 56, или 58. В еще одном аспекте настоящего изобретения предлагается полинуклеотид, кодирующий белок-репрессор cI, имеющий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% or 99% идентичности последовательности с полипептидом, содержащим последовательность, представленную в SEQ ID NO:2, 44, 46, 48, 50, 52, 54, 56, или 58, или консервативный вариант, аллельный вариант, гомолог или фрагмент, содержащий, по меньшей мере, восемь или, по меньшей мере, десять последовательно расположенных аминокислот одного из этих полипептидов, или комбинация этих полипептидов. В другом аспекте настоящего изобретения предлагается полинуклеотид, содержащий нуклеотидную последовательность, представленную в SEQ ID NO:1, 45, 47, 49, 51, 53, 55, 57, или 59. В еще одном аспекте настоящего изобретения предлагается полинуклеотид, обладающий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичностью последовательности по отношению к полинуклеотиду, содержащему последовательность, изложенную в SEQ ID NO:1, 45, 47, 49, 51, 53, 55, 57, или 59, или его варианту.
В одном воплощении промотор, управляющий транскрипцией гена cI, представляет себя нативный промотор гена cI. В другом воплощении, промотор, управляющий транскрипцией гена cI, является слабым промотором гена устойчивости к канамицину (Р1). В другом воплощении промотор содержит полинуклеотид, имеющий последовательность, представленную в SEQ ID NO:29, 83, или 30. В другом воплощении, промотор содержит полинуклеотид, имеющий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полинуклеотидом, имеющим последовательность, представленную в SEQ ID NO:29, 83, или 30.
Другие пары промотор/репрессор, которые могут применяться в способах по настоящему изобретению, включают в частности промотор carB, контролируемый парой репрессор-антирепрессор CarA-CarS, промоторы и репрессоры гена гормона роста, Lac-репрессор (lacl) и промотор Ptrc-2.
Мутантная пара промотор/репрессор в качестве субстрата для мутагенеза может быть отобрана среди мутантов, доступных в настоящее время и описанных в данной области. Исследования связывания мутантных репрессоров с их операторами [См., например. Nelson and Sauer, Cell, 42:549 (1985); Nelson and Sauer, J. Mol. Biol. 192:27 (1986); and Gussin, et al, Lambda II, (Hendrix, Roberts, Stahl, and Weisberg, eds) Cold Spring Harbor Press, Cold Spring Harbor, N.Y., p.93-123 (1983)] делают возможным отбор известных последовательностей мутантного промотора и/или мутантного репрессора, которые могут обеспечить снижение, но не прекращение транскрипции. Отобранный промоторный участок включают в гетерологичную экспрессирующую плазмиду и затем изменяют путем соответствующего сайт-направленного мутагенеза (см., например, Morinaga, et al., Biotechnology, 2:636, 1984). В ином случае последовательность, кодирующую репрессор, изменяют аналогичным способом. Полученную в результате мутантную пару репрессор/промотор анализируют на предмет возможности стимуляции секреции гетерологичного белка путем сравнения с экспрессией плазмиды дикого типа, не подвергнутой сайт-направленному мутагенезу.
Объект изобретения, раскрытый в настоящем документе, обеспечивает безлекарственную концепцию или концепцию без устойчивости к антибиотику, где активность репрессора (такого как репрессор cI), экспрессируемого плазмидой, ингибирует транскрипцию токсичного продукта гена (такого как продукт гена sacB), помещенного под контроль промотора фага λ, локализованного на хромосоме клетки-хозяина, и где выживаемость клеток-хозяев в присутствии субстрата, такого как сахароза, гарантируется достаточным уровнем экспрессии плазмидой белка-репрессора плазмиды (такого как белок-репрессор cI фага λ). Рост штамма клеток-хозяев, содержащих плазмиду без устойчивости к антибиотику, в присутствии сахарозы гарантирует эффективную систему для поддержания и продуцирования ДНК-плазмид.
Термин «плазмида» охватывает любую единицу транскрипции ДНК, содержащую полинуклеотид в соответствии с изобретением, и элементы, необходимые для ее in vivo экспрессии в клетке или клетках требуемого хозяина или мишени; и, в связи с этим отмечается, что суперскрученная или не суперскрученная, кольцевая плазмида, а также линейная форма, предназначены для включения в объем притязаний изобретения.
Одно воплощение настоящего изобретения относится к безлекарственной плазмиде, содержащей один или несколько генов репрессоров, функционально связанных с одним или несколькими промоторами. В одном аспекте безлекарственная плазмида содержит полинуклеотид, кодирующий белок-репрессор cI. В другом аспекте безлекарственная плазмида содержит промотор, функционально связанный с полинуклеотидом, кодирующим белок-репрессор cI. Промотор может представлять собой нативный промотор гена cI или слабый промотор гена устойчивости к канамицину. В другом аспекте безлекарственная плазмида дополнительно содержит гетерологичный полинуклеотид, кодирующий иммуноген или белок. В другом аспекте безлекарственная плазмида содержит промотор, функционально связанный с гетерологичным полинуклеотидом, кодирующим иммуноген или белок. Любой подходящий промотор, известный в данной области, можно применить в безлекарственных плазмидах согласно настоящему изобретению, включая бактериальные, дрожжевые, грибные промоторы, промоторы насекомых, млекопитающих и растительные промоторы. Промотор может быть, в частности, немедленно-ранним промотором цитомегаловируса (CMV-IE) происходящим из клеток человека или мыши, или необязательно с другим происхождением, например, может быть суперпромотором из клеток крысы или морской свинки (Ni, M. et al., Plant J.7, 661-676, 1995). Промотор CMV-IE может содержать действительную промоторную часть, которая может связана или не связана с энхансерной частью. Можно сослаться на ЕР-А-260148, ЕР-А-323597, Пат. США No.5168062, 5385839 и 4968615, а также на заявку РСТ No WO 87/03905. Другие промоторы из грамотрицательных бактерий представляют собой также подходящие промоторы, выделенные из Avibacterium, Brucella, Escherichia coli, Haemophilus (например, Haemophilus suis), Salmonella (e.g., Salmonella enteridis, Salmonella typhimurium, Salmonella infantis), Shigella, Pasteurella и Rimeirella.
Плазмиды могут содержать другие элементы контроля экспрессии. Особенно предпочтительно включить стабилизирующую последовательность, например, интронную последовательность, например, первый интрон hCMV-IE (заявка РСТ No.WO 89/01036), II интрон кроличьего β-глобинового гена (van Ooyen et al., 1979). В другом воплощении, плазмида может содержать 3' UTR. 3' UTR может быть в частности 3' UTR нопалин-синтазы агробактерии.
Что касается сигнала полиаденилирования (полиА) для плазмид и вирусных векторов отличных от поксвирусных, то возможно применение сигнала поли(А) гена бычьего гормона роста (bGH) (см. Пат. США No.5122458), или сигнал поли(А) кроличьего гена β-глобина или сигнала поли(А) вируса SV40.
«Клетка-хозяин» означает прокариотическую или эукариотическую клетку, которая была генетически изменена, или является способной к генетическому изменению путем введения экзогенного полинуклеотида, такого как рекомбинантная плазмида или вектор. Когда речь идет о генетически измененных клетках, термин относится к первоначально измененной клетке и к ее потомкам.
Другое воплощение настоящего изобретения относится к штамму грамотрицательных бактерий-хозяев, содержащих безлекарственную плазмиду. Один аспект воплощения относится к штамму грамотрицательных бактерий-хозяев, содержащих безлекарственную плазмиду и гетерологичный полинуклеотид, вставленный в один или несколько несущественных участков хромосомы организма-хозяина, и где гетерологичный полинуклеотид функционально связан с промотором, который может сильно регулироваться репрессором. В другом аспекте воплощения гетерологичный полинуклеотид, вставленный в хромосому организма-хозяина, представляет собой ген sacB. В другом воплощении ген sacB функционально связан с промотором, таким как правый промотор фага λ. Признается, что ген sacB и промотор могут быть вставлены во множестве копий, например, в двух или трех копиях, в хромосому организма-хозяина. Несущественный участок штамма клеток-хозяев может представлять собой ген deA или purN на хромосоме E. coli. Гены deA и purN не являются жизненно необходимыми, и удаление этих генов не оказывает влияния на скорость роста бактерий (Kim, J. et al., Biochemistry. 46, 44:12501-12511). В одном аспекте одну копию кассеты с геном sacB и промотора вставляют в локус deA или в локус purN путем аллельной замены. В другом аспекте, две копии кассеты гена sacB и промотора вставляют в локус deA или в локус purN. В еще одном аспекте две копии кассеты гена sacB и промотора вставляют в хромосому организма-хозяина, где одну копию кассеты гена sacB и промотора вставляют в локус deA, а другую копию вставляют в локус purN. Кассета гена sacB и промотора относится к полинуклеотиду, содержащему ген sacB, функционально связанному с промотором. В одном аспекте кассета гена sacB и промотора включает ген sacB и правый промотор фага λ. В другом аспекте, кассета гена sacB и промотора содержит полинуклеотид, содержащий последовательность, представленную в SEQ ID NO:75.
Объект изобретения, раскрытый в настоящем документе, дополнительно относится к вакцине или к композиции, содержащей безлекарственные плазмиды, содержащие гетерологичный ген, кодирующий иммуноген или белок, или к вакцине или композиции, содержащей иммуноген или белок, экспрессируемый с помощью безлекарственных плазмид. Вакцина или композиция дополнительно может содержать фармацевтически приемлемый носитель.
В одном воплощении, иммуноген выбирают из кошачьего патогена, такого как в частности кошачий вирус герпеса (FHV), кошачий калицивирус (FCV), кошачий вирус лейкоза (FeLV), кошачий вирус иммунодефицита (FIV), вирус бешенства и так далее, и из их комбинации.
В другом воплощении, иммуноген выбирают из патогена собак, в частности вирус бешенства, вирус герпеса собак (CHV), парвовируса собак (CPV), коронавируса собак, Leptospira canicola, Leptospira icterohaemorragiae, Leptospira grippotyphosa, Borrelia burgdorferi, Bordetella bronchiseptica и так далее, и из их комбинаций.
Еще в одном воплощении, иммуноген выбирают из лошадиного патогена, такого как вирус герпеса лошадей (типа 1 или типа 4), вирус гриппа лошадей, столбняка, вирус западного нила, Streptococcus equi, Rhodococcus equi и так далее или из их комбинаций.
Еще в одном воплощении, иммуноген выбирают из коровьих патогенов, таких как вирус бешенства, коровий ротавирус, коровий вирус парагриппа типа 3 (bPIV-3), коровий коронавирус, коровий вирус вирусной диарреи (BVDV), вирус ящура (FMDV), коровий респираторно-синцитиальный вирус (BRSV), Коровий вирус инфекционного ринотрахеита (IBR), Escherichia coli, Pasteurella multocida, Pasteurella haemolytica, сальмонелла, Cryptosporidium и так далее, и из их комбинаций.
Еще в одном воплощении, иммуноген выбирают из свиных патогенов, таких как, в частности, вирус свиного гриппа (SIV), свиной цирковирус типа 2 (PCV-2), свиной вирус репродуктивно-респираторного синдрома (PRRS), вирус псевдобешенства (PRV), свиной парвовирус (PPV), FMDV, Mycoplasma hyopneumoniae, Erysipelothrix rhusiopathiae, Pasteurella multocida, Bordetella bronchiseptica, Escherichia coli, вирус катаральной лихорадки, вирус африканской болезни лошадей, лихорадки долины Рифт, вируса Нипах и так далее, и из их комбинаций.
Термины «антиген» или «иммуноген» используются взаимозаменяемо и обозначают вещество, которое индуцирует специфичный иммунный ответ у животного-хозяина. Антиген может содержать рекомбинантный вектор, содержащий вставку с иммуногенными свойствами; часть или фрагмент ДНК, способной индуцировать иммунный ответ при презентировании животному-хозяину; белок, полипептид, пептид, эпитоп, гаптен или любую их комбинацию. В ином случае, иммуноген или антиген могут содержать токсин или антитоксин.
При использовании в настоящем документе, термины «фармацевтически приемлемый носитель» и «фармацевтически приемлемое средство переноса» взаимозаменяемы и обозначают жидкое средство переноса для содержания вакцинных антигенов, которые можно инъецировать организму-хозяину без побочных эффектов. Подходящие фармацевтически приемлемые носители, известные в данной области, включают в частности стерильную воду, солевой раствор, глюкозу, декстрозу или буферные растворы. Носители могут включать вспомогательные агенты, включающие в частности разбавители, стабилизаторы (т.е., сахара и аминокислоты), консерванты, увлажнители, эмульгаторы, рН-буферные агенты, добавки усиления вязкости, красители и так далее.
Катионные липиды, содержащие четвертичную аммонийную соль, которые предпочтительно, но не исключительно, подходят для плазмид, предпочтительно представляют собой те, которые имеют следующую формулу:
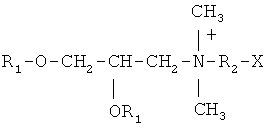
в которой R1 является насыщенным или ненасыщенным неразветвленным алифатическим радикалом, содержащим 12-18 атомов углерода, R2 является другим алифатическим радикалом, содержащий 2 или 3 атома углерода, а Х является аминогруппой или гидроксильной группой, например, DMRIE. В другом воплощении катионный липид может быть ассоциирован с нейтральным липидом, например, DOPE.
Среди этих катионных липидов предпочтение отдается DMRIE (N-(2-гидроксиэтил)-N,N-диметил-2,3-бис(тетрадедецилокси)-1-пропан аммоний; WO 96/34109), предпочтительно ассоциированный с нейтральным липидом, предпочтительно DOPE (диолеоил-фосфатидил-этанол амин; Behr, 1994), с образованием DMRIE-DOPE.
Предпочтительно, если смесь плазмиды с адъювантом готовится для немедленного приема и предпочтительно, если смесь готовится одновременно с введением препарата или сразу перед введением, препарата; например, смесь плазмида-адъювант готовится сразу перед или до введения, предпочтительно так, чтобы было достаточно времени перед введением для того, чтобы в смеси образовался комплекс, например, от около 10 до около 60 минут до введения, как например, приблизительно 30 минут до введения.
Если присутствует DOPE, то молярное соотношение DMRIE:DOPE составляет предпочтительно около 95:около 5 - около 5:около 95, более предпочтительно, около 1:около 1, например, 1:1.
С DMRIE или с DMRIE-DOPE массовое соотношение адъювант:плазмида может составлять от около 50:около 1 до около 1:около 10, как например, около 10:около 1 и около 1:около 5, и предпочтительно, около 1:около 1, и около 1:около 2, например, 1:1 и 1:2.
В другом воплощении, фармацевтически или ветеринарно приемлемый носитель, наполнитель или основа может представлять собой эмульсию вода-в-масле. Примеры подходящих эмульсий вода-в-масле включают вакцинные эмульсии вода-в-масле на основе масла, которые стабильны и являются жидкими при 4°С содержат: от 6 до 50 об./об.% антиген-содержащей водной фазы, предпочтительно, от 12 до 25 об./об.%, от 50 до 94 об./об.% масляной фазы, содержащей в целом или частично неметаболизируемое масло (например, минеральное масло, такое как парафиновое масло) и/или метаболизируемое масло (например, растительное масло, или жирную кислоту, полиол или эфиры спирта), от 0,2 до 20 p/v % поверхностно-активных веществ, предпочтительно, от 3 до 8 p/v %, последние в целом или частично или в смеси с эфирами полиглицерина, причем указанные эфиры полиглицерина предпочтительно представляют собой (поли)рицинолеаты глицерина, или полиоксиэтилен рициновые масла или еще гидрогенизированные полиоксиэтилен рициновые масла. Примеры поверхностно-активных веществ, которые могут использоваться в эмульсии вода-в-масле, включают эфиры этоксилированного сорбита (например, эфир полиоксиэтилен (20) сорбита и моноолеиновой кислоты (Tween 80®), доступен у «AppliChem, Inc.», Чешир, Коннектикут) и эфиры сорбита (например, эфир сорбита и моноолеиновой кислоты (Span 80®), доступен от Sigma Aldrich, Сент-Луис, Миссури). Кроме того, относительно эмульсии вода-в-масле см. также Пат. США No.6919084, например. Пример 8 оттуда, включенный в настоящий документ ссылкой. В некоторых воплощениях, антиген-содержащая водная фаза содержит солевой раствор, содержащий один или несколько буферных агентов. Примером подходящего буферного раствора является фосфатно-солевой буферный раствор. В предпочтительном воплощении, эмульсия вода-в-масле может представлять собой тройную эмульсию вода/масло/вода (В/М/В) (см., например, Пат. США No.6358500, включенный в настоящий документ ссылкой). Примеры других подходящих эмульсий описаны, например, в Пат. США No.7371395, включенном в настоящий документ ссылкой).
Вакцины или композиции могут быть введены в дозах и методами, хорошо известными специалистам в области медицины или ветеринарии, принимая во внимание такие факторы как возраст, пол, масса, вид и состояние животного-реципиента и путь введения. Путь введения может быть чрескожным введением через слизистую оболочку (например, пероральное, назальное, ректальное, вагинальное), или может быть введением через парентеральный путь (внутрикожное, внутримышечное, подкожное, внутривенное или внутрибрюшинное введение). Вакцина или композиция может вводиться индивидуально или может вводиться совместно или последовательно с другими типами лечения или терапиями. Формы введения могут включать суспензии, сиропы или эликсиры, и препараты для парентерального, подкожного, внутрикожного, внутримышечного или внутривенного введения (например, введения с помощью инъекций), такие как стерильные суспензии или эмульсии. Вакцина или композиция может вводиться в виде спрея или может смешиваться с пищей и/или водой, или может доставляться в виде предварительной смеси с использованием подходящего носителя, разбавителя или наполнителя, такого как стерильная вода, физиологический солевой раствор, глюкоза и так далее. Композиции могут содержать вспомогательные вещества, такие как увлажнители или эмульгаторы, буферные агенты для регулирования рН, адъюванты, добавки, усиливающие гелеобразование или вязкость, консерванты, ароматизаторы, красители и т.п., в зависимости от пути введения и требуемого препарата. Стандартные фармацевтические тексты, такие как «Remington's Pharmaceutical Sciences», 1990, могут быть использованы в качестве справки для получения подходящих препаратов, без проведения излишних экспериментов.
Объект изобретения, раскрытый в настоящем документе, дополнительно относится к способу получения безлекарственной плазмиды, включающему стадии 1) создания клеток-хозяев в виде штамма грамотрицательных бактерий, содержащих гетерологичный полинуклеотид, вставленный путем аллельной замены в один или несколько несущественных участков хромосомы организма-хозяина; 2) конструирования ДНК-плазмиды, содержащей полинуклеотид, кодирующей белок-репрессор cI, 3) трансформации штамма бактериальных клеток-хозяев ДНК-плазмидой, содержащей ген, кодирующий белок-репрессор cI, 4) роста трансформированного штамма бактериальных клеток-хозяев в присутствии сахарозы при температуре в интервале от 30°С до 42°С; и 5) извлечения ДНК-плазмиды.
В одном аспекте воплощения, ДНК-плазмида дополнительно содержит гетерологичный полинуклеотид, кодирующий иммуноген или белок, где гетерологичный полинуклеотид функционально связан с промотором. Промотор может быть промотором, функциональным в прокариотических или эукариотических клетках, такой как промотор CMV.
Объект изобретения, раскрытый в настоящем документе, дополнительно относится к способу получения белка или иммуногена с использованием безлекарственной плазмиды, включающему стадии 1) создания штамма грамотрицательных бактерий-хозяев, содержащих гетерологичный полинуклеотид, вставленный путем аллельной замены в один или несколько несущественных участков хромосомы организма-хозяина; 2) конструирования ДНК-плазмиды, содержащей полинуклеотид, кодирующей белок-репрессор cI, и ген, кодирующий иммуноген или белок; 3) трансформации штамма бактерий-хозяев ДНК-плазмидой, содержащей полинуклеотид, кодирующий белок-репрессор cI, и ген, кодирующий иммуноген или белок; 4) культивирование (выращивание) трансформированного штамма бактерий-хозяев в присутствии сахарозы при температуре в интервале от 30°С до 42°С; и 5) извлечения иммуногена или белка.
В одном аспекте воплощения, ген, кодирующий иммуноген или белок, функционально связан с промотором, функционирующим в прокариотических клетках. Промотор может представлять собой промотор из грамотрицательных бактерий, включающий в частности промоторы, выделенные из Avibacterium, Brucella, Escherichia coli, ш (например, Haemophilus suis), Salmonella (e.g., Salmonella enteridis, Salmonella typhimurium, Salmonella infantis), Shigella, Pasteurella, и Rimeirella.
В другом аспекте воплощения, гетерологичный полинуклеотид, вставленный в хромосому организма-хозяина, кодирует белок sacB. В другом аспекте, концентрация сахарозы может варьировать в интервале от 0% до около 20%, от около 1% до около 20%, от около 1% до около 15%, от около 1% до около 10%. Еще в одном аспекте, концентрация сахарозы может варьироваться в интервале от около 1% до около 10%, от около 1% до около 2%, от около 2% до около 3%, от около 3% до около 4%, от около 4% до около 5%, от около 5% до около 6%, от около 6% до около 7%, от около 7% до около 8%, от около 8% до около 9%, или от около 9% до около 10%. Еще в одном аспекте, концентрация сахарозы может составлять около 1%, около 2%, около 3%, около 4%, около 5%, около 6%, около 7%, около 8%, около 9%, или около 10%.
В одном аспекте воплощения, температура может варьировать в интервале от около 30°С до около 42°С. В другом аспекте, температура может варьировать в интервале от около 30°С до около 41°С, от около 30°С до около 39°С, от около 30°С до около 38°С, от около 30°С до около 37°С.Еще в одном аспекте, температура может составлять около 30°С, около 31°С, около 32°С, около 33°С, около 34°С, около 35°С, около 36°С, около 37°С, около 38°С, около 39°С, около 40°С, или около 41°С.
Термин извлечение включает в частности накопление, экстракцию, сбор или очистку из культуральной среды.
«Выделенный» биологический компонент (такой как полинуклеотид, ДНК-плазмида, белок или органелла) обозначает компонент, который был по существу отделен или очищен от других биологических компонентов в клетке организма, в которой эти компоненты существуют в естественном состоянии, например, от другой хромосомной и внехромосомной ДНК и РНК, белков и органелл. Полинуклеотиды (включая ДНК-плазмиды) и белки, которые были «выделены», включают полинуклеотиды и белки, очищенные с помощью стандартных методов очистки, например, см. Russell, David W.; Sambrook, Joseph (2001), Molecular cloning: a laboratory manual. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory. Термин также охватывает полинуклеотиды и белки, полученные с помощью рекомбинантной технологии, а также с помощью химического синтеза.
При использовании в данном документе термин «очищенный» не требует абсолютной чистоты; скорее он подразумевается как относительный термин. Таким образом, например, препарат очищенного полипептида или полинуклеотида является препаратом, в котором полипептид или полинуклеотид находится в более обогащенном состоянии, чем полипептид или полинуклеотид в его естественной среде. То есть полипептид или полинуклеотид отделяется от клеточных компонентов. Термин «по существу очищенный» предназначен для такого состояния, когда удалено, по меньшей мере, 60%, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, или, по меньшей мере, 98%, или более, клеточных компонентов или материалов. Аналогично этому, полипептид или полинуклеотид могут быть частично очищенными. Термин «частично очищенный» предназначен для такого состояния, когда удалено менее чем 60% клеточных компонентов или материалов.
В настоящем изобретении дополнительно предлагаются способы индукции иммунного или защитного ответа у животного, включающие введение животному иммунологической композиции, вакцины или композиции согласно изобретению. Вызванные иммунные ответы в частности представляют собой антительный и/или клеточный иммунный ответ и конкретно ответ гамма-интерферона.
Конкретно, в настоящем изобретении предлагаются способы иммунизации против симптомов или профилактики симптомов или уменьшения симптомов, вызванных инфекцией животного патогенным организмом (например, инфекцией с помощью вируса, бактерий, грибов или простейших паразитов). Способ по настоящему изобретению применяется для позвоночных животных, включающих в частности людей, псовых (пример, собака), кошачьих (пример, кошка); лошадиных (пример, лошадь), крупный рогатый скот (пример, корова) и свиней (пример, домашняя свинья), а также для птиц, включающих в частности кур, индеек, гусей, перепелов, фазанов, попугаев, зябликов, ястребов, ворон и бескилевых (страусов, эму и казуаров и так далее). Способ по настоящему изобретению также применяется для обеспечения ДНК-вакцин для рыб.
В конкретном аспекте изобретения, эти способы состоят из вакцинации беременных самок перед родами путем введения вакцинной композиции, полученной согласно изобретению. Эти способы дополнительно включают индукцию защитных антител, вызванную протоколом вакцинации, и перенос этих защитных антител от вакцинированных беременных самок к их потомству. Перенос таких материнских антител впоследствии защищает потомство от заболевания.
Дозировка вакцинной композиции, полученной согласно настоящему изобретению, будет зависеть от вида, породы, возраста, размера, истории вакцинации и состояния здоровья животного, подвергаемого вакцинации. Другие факторы типа концентрации антигена, дополнительных компонентов вакцин, и пути введения (т.е., подкожное, внутрикожное, пероральное, внутримышечное или внутривенное введение) также будут способствовать эффективному дозированию. Дозировка вакцины для введения легко определяется на основе концентрации антигена вакцины, пути введения и возраста и состояния животного, подвергаемого вакцинации. Каждая партия антигенов может быть индивидуально откалибрована. В ином случае, систематические испытания иммуногенности различных дозировок, а также исследования Минимальной Защитной Дозы (англ. Minimum Protective Dose, MPD) и другие процедуры скрининга могут использоваться для определения эффективной дозировки вакцинной композиции согласно настоящему изобретению без проведения излишних экспериментов. Из примеров, представленных ниже, легко понять, какая должна быть приблизительная дозировка и какой приблизительно объем будет соответствовать для вакцинной композиции, описанной в настоящем документе. Критический фактор заключается в том, что дозировка обеспечивает, по меньшей мере, частичный защитный эффект против природной инфекции, что становится очевидным в результате снижения смертности и заболеваемости из-за этой природной инфекции. Подходящий объем аналогично легко оценивается специалистом в данной области. Например, у птиц объем дозы может составлять от около 0,1 мл до около 0,5 мл и предпочтительно от около 0,3 мл до около 0,5 мл. Для животных, относящихся к различным видам кошачьих, псовых и лошадиных, объем дозы может составлять от около 0,2 мл до около 3 мл, предпочтительно, от около 0,3 мл до около 2 мл и более, предпочтительно, от около 0,5 мл до около 1 мл. Для крупного рогатого скота и свиней объем дозы может составлять от около 0,2 мл до около 5 мл, предпочтительно, от около 0,3 мл до около 3 мл и более, предпочтительно, от около 0,5 мл до около 2 мл.
Повторные вакцинации могут осуществляться предпочтительно с интервалом периодичности для усиления исходного иммунного ответа, или когда истек продолжительный период времени после последней дозы. В одном воплощении настоящего изобретения, вакцинную композицию вводят в виде парентеральной инъекции (т.е. подкожно, внутрикожно или внутримышечно). Композиция может вводиться в виде однократной дозы или, в альтернативных воплощениях, может вводиться в виде повторяющихся доз от около двух до около пяти, которые вводят с интервалом от около двух до около шести недель, предпочтительно, от около двух до около пяти недель. Однако специалисту в данной области понятно, что количество доз и временные интервалы между вакцинациями зависят от ряда факторов, включающих в частности возраст вакцинируемого животного; состояние животного; путь иммунизации; количество антигена, доступного на одну дозу; и т.п. Для исходной вакцинации, период, как правило, будет продолжительнее, чем неделя и предпочтительно составит от около двух до около пяти недель. Для ранее вакцинированных животных, вторичная вакцинация перед беременности или во время нее может осуществляться с интервалом около года.
Настоящее изобретение также предполагает введение вакцинной композиции с использованием безигольного инъектора, такого как «Pigjet®», «Avijet®», «Dermojet®» или «Biojector®» (Bioject, Орегон, США). Специалист в данной области способен скорректировать технические параметры инъектора с учетом ряда факторов, таких как вид животного, подвергаемого вакцинации; возраст и масса животного и т.п. без проведения излишних экспериментов.
Настоящее изобретение дополнительно относится к набору реактивов, содержащему первый флакон, содержащий активный ингредиент, такой как иммуноген или фармацевтическую композицию, и второй флакон, содержащий разбавитель, полученный согласно настоящему изобретению. Иммуноген может быть представлен в лиофилизированной форме, в сухой форме или в виде водного раствора, описанного в настоящем документе.
Далее изобретение будет описано с помощью представленных ниже неограничивающих примеров.
ПРИМЕРЫ
Пример 1: Концепция поддержания плазмиды без устойчивости к антибиотику
Следующий пример демонстрирует новую концепцию для поддержания высокого количества копий плазмид в клетке-хозяине, относящейся к грамотрицательным бактериям, которая-основана на трех компонентах:
1. Грамотрицательная бактерия-хозяин, которая в определенных условиях культивации экспрессирует продукт, токсичный для грамотрицательной бактерии, где токсичный ген вставлен в один или несколько несущественных участков хромосомы бактериальной клетки-хозяина.
2. Наличие на бактериальной хромосоме гена, кодирующего токсичный продукт под контролем конститутивного промотора, который можно строго регулировать; и
3. Экспрессия конкретного репрессора с плазмиды, который регулирует промотор, функционально связанный с токсичным геном на хромосоме клетки-хозяина.
В этой конструкции, присутствие плазмиды это то, что контролирует размножение клеток-хозяев, относящихся к грамотрицательным бактериям, когда они находятся в определенных условиях культивирования (например, в присутствии сахарозы).
В первом компоненте, грамотрицательную бактерию трансформируют для экспрессии гена sacB, кодирующего левансахаразу (см. Фиг.1). Второй и третий компоненты относятся к продукту гена репрессора cI и к правому промотору фага λ. Ген cI бактериофага Х кодирует репрессор λ.
Участок генома, который кодирует белок-репрессор cI, известен как участок, связанный с иммунитетом. Участок, связанный с иммунитетом, показан на Фиг.2. Плазмидная система, экспрессирующая продукт гена cI, должна ингибировать транскрипцию гена sacB, помещенного под контроль промотора λ. Такая репрессия должна придавать устойчивость к сахарозе и давать возможность как для поддержания плазмиды, так и для ее размножения в клетках-хозяевах (см. Фиг.1).
На Фиг.3 дан теоретический обзор «безлекарственной» концепции, где активность репрессора cI, экспрессируемого плазмидой, ингибирует транскрипцию токсичного продукта гена sacB, помещенного под контроль промотора Pr фага λ, локализованного на хромосоме клетки-хозяина, и где выживаемость клеток-хозяев Е. coli в присутствии сахарозы гарантирована с помощью достаточного уровня экспрессии белка-репрессора cI с плазмиды. Репрессор cI, связывающийся с его специфическим промотором, оптимален при температурах от 30°С до 37°С (см. Фиг.3).
В конкретном случае (таком как при использовании термочувствительного репрессора cI857), репрессор cI не будет связываться с промотором при температурах выше, чем 40°С. Таким образом, переключение на 42°С ингибирует активность связывания репрессора cI с промотором Pr фага λ, и, как следствие, должно придавать клеткам-хозяевам чувствительность к сахарозе (более конкретно, к субпродуктам левана). Инкубация при температуре 42°С в присутствии сахарозы является специальным условием для подтверждения функциональности цельной системы и является составной частью «доказательства от противного» в поддержку полной функциональности системы (см. ФИГ 4 и ФИГ 6С). Действительно, родительские клетки λPr::sacBΩkan, трансформированные либо плазмидой рРВ829, либо рРВ838, растущие в присутствии либо Cat, либо Сахарозы, были не жизнеспособны при их посеве на LB-агар с сахарозой при инкубации при 42°С. Это говорит о том, что жизнеспособность родительских клеток зависит от поддержания плазмиды, содержащей ген cI.
Пример 2: Получение Штамма Клеток-Хозяев Е. coli. Несущих λ Промотор/Ген Левансахаразы
Получали два сконструированных штамма клеток-хозяев, содержащих кассету λPr::sacBΩkan, где кассету, содержащую ген sacB под контролем промотора лямбда (λPr), метили геном устойчивости к канамицину и затем вводили в хромосому Е. coli с помощью аллельной замены или гена edA или purN. Эти гены не являются жизненно важными, и их делеция не влияет на скорость роста. Трансформированные клетки становятся высоко чувствительными к сахарозе.
На Фиг 8А-В показано, что кассета λPr::sacBΩkan, которую вставляют в хромосому Е. coli аллельной заменой гена edA или purN, соответственно, состоит из шести независимых компонентов, которые были амплифицированы с помощью ПЦР перед их сшивкой посредством объединяющей ПЦР.
На Фиг 8D показано, что кассету λPr::sacBΩkan вставляли в хромосому Е. coli с помощью аллельной замены гена edA или purN. Вставку кассет осуществляли путем трансформации Е. coli с использованием линейной ДНК-матрицы, содержащей делегированный ген, фланкированный в 5'- и 3'-области гомологичным участком хромосомного локуса, и путем переноса в специальный рекомбинационный штамм Е. coli, такой как мутанты recD, recB, recC, sbcA. Штаммом Е. coli для создания родительского штамма являлся штамм BJ5183 от Stratagene (ref# 200154), с генотипом endA1 sbcBC recBC galK met thi-1 bioT hsdR (Strr). Показано, что исходный фенотип Е. coli дикого типа это KanS, CatS и СахарозаR. После трансформации штамма, фенотип изменился на KanR, CatS и СахарозаS. Последний фенотип соответствовал одному из родительских штаммов, использованных для дальнейших экспериментов, связанных с демонстрацией функциональности плазмиды без устойчивости к антибиотику.
На Фиг 8С показано, что кассета λPr::sacBΩkan, которую вставляют аллельной заменой гена edA в хромосому Е. coli Prλ::sacBΩkan, состоит из шести независимых компонентов, которые были амплифицированы с помощью ПЦР перед их сшивкой посредством объединяющей ПЦР. Целью внедрения новой кассеты λPr::sacBΩcat, затрагивающей делецию гена edA путем аллельной замены, было создание двойного делеционного мутанта ΔedA ΔpurN, экспрессирующего две кассеты sacB.
На Фиг 8Е показано, что когда родительский штамм трансформировали плазмидой cI (рРВ829 или рРВ838), то клетки (родительский штамм+плазмида с cI [рРВ829 или рРВ838]) становились устойчивыми к канамицину (KanR), хлорамфениколу (CatR) и сахарозе (СахарозаR). Фигура иллюстрирует сосуществование созданного штамма клеток-хозяев (ΔpurN λPr::sacB+Ωkan) или (ΔedA λPr::sacB+Ωkan) и плазмиды с устойчивостью к медикаменту для доказательства концепции, когда ген sacB помещен под контроль промотора лямбда и конструкт введен в хромосому Е. coli путем аллельной замены гена edA или purN. Трансформированные клетки становятся высоко чувствительными к сахарозе. Когда содержащие репрессор cI плазмиды, меченные геном устойчивости к хлорамфениколу (рРВ838 или рРВ829), вводили в штаммы ΔpurN λPr::sacB+Ωkan или ΔedA λPr::sacB+Ωkan, то трансформированные клетки становились устойчивыми к канамицину, хлорамфениколу и сахарозе.
Промотор λ (λPr) амплифицировали с помощью праймеров РВ1232 и РВ1233 для делеции purN.
Праймер РВ1232 (SEQ ID NO:6):
(CCGAACAACGCGTGGTTATCGACACCGCAAGGGATAAATATCTAACACCG) и
РВ1233 праймер (SEQ ID NO:7):
(CAAACTTTTTGATGTTCATATCCATCTGATCCTCTTCAAAAGGCCACCTG)
Промотор λ (λPr) амплифицировали с использованием праймеров РВ1234 и РВ1233 для делеции edA.
РВ1234 (SEQ ID NO:8):
(GACGACAAATTTGTAATCAGGCGAGAGCACCGCAAGGGATAAATATCTAACACCG) Амплификацию промотора λ (λPr) (SEQ ID NO:5) осуществляли с использованием плазмиды pLDRS (ATCC# 77357) в качестве ДНК матрицы. Ген sacB (SEQ ID NO:3) амплифицировали с помощью праймеров
РВ1192 (SEQ ID NO:9)
(ATGGATATGAACATCAAAAAGTTTGC) и
РВ1193 (SEQ ID NO:10)
(AAACAAATAGGGGTTCCGCGCACATTTATTTGTTAACTGTTAATTGTCCTTG)
с использованием pNB350 (от «Merial») в качестве ДНК матрицы, когда применяли объединяющую ПЦР для создания кассеты λPr::sacBΩkan (см. Фиг 8А-В). Для создания кассеты λPr::sacBΩcat использовали обратный праймер РВ1320 (SEQ ID NO:80) (GCCGATCAACGTCTCATTTTCGCCGTTAACAGATCTTTATTTGTTAACTGTTAATTGT CCTTG) вместо РВ1193. Промотор bla амплифицировали с помощью праймеров РВ1194 (SEQ ID NO:11) (ATGTGCGCGGAACCCCTATTTG) и РВ1195 (SEQ ID NO:12) (GACGTTTCCCGTTGAATATGGCTCATACTCTTCCTTTTTCAATATTATTGAAGC) с использованием pCMVβ в качестве ДНК матрицы. Ген устойчивости к канамицину амплифицировали с помощью праймеров РВ1196 (SEQ ID NO:13) (ATGAGCCATATTCAACGGGAAACG) и РВ1197 (SEQ ID NO:14) (GAAAAACGCCAGCGGCAGAGCTGGCGCTTAGAAAAACTCATCGAGCATCAAATG) с использованием плазмиды pLL14 (содержащей слабый Промотор гена устойчивости к Канамицину, от «Merial») в качестве ДНК матрицы. Ген устойчивости к хлорамфениколу (cat), помещенный под контроль нативного промотора cat, амплифицировали с помощью праймероы РВ1321 (SEQ ID NO:81) (AGATCTGTTAACGGCGAAAATGAG) и РВ1322 (SEQ ID NO:82) (AAAACGCTACAAAAATGCCCGATCCTTTACGCCCCGCCCTGCCACTCATCGC) с использованием рРВ829 в качестве ДНК матрицы. 5'-фланкирующий участок гена purN амплифицировали с использованием праймеров РВ1237 (SEQ ID NO:15) (TTTGCGGCCGCTGGTGGTGGTCGCCATGTGCGTTAATGACC) и РВ1199 (SEQ ID NO:16) (TATTCGATAACCACGCGTTGTTCGG) с использованием геномной ДНК штамма Е. coli SCSI в качестве ДНК матрицы. 3'-фланкирующий участок гена purN амплифицировали с помощью праймеров РВ1200 (SEQ ID NO:17) (GCGCCAGCTCTGCCGCTGGCGTTTTTC) и РВ1238 (SEQ ID NO:18) (TTTGGATCCGCTGGTGGATATCATCAAGGCAGTAACGCAGAATG) с использованием геномной ДНК штамма Е. coli SCSI в качестве ДНК матрицы. 5'-фланкирующий участок гена edA амплифицировали с помощью праймеров РВ1235 (SEQ ID NO:19) (TTTGCGGCCGCTGGTGGTTGAGAACCAGGTGATTGAAGCGCC) и РВ1209 (SEQ ID NO:20) (CTCTCGCCTGATTACAAATTTGTCGTC) с использованием геномной ДНК штамма Е. coli SCSI в качестве ДНК матрицы. 3'-фланкирующий участок гена edA амплифицировали с помощью праймеров РВ1210 (SEQ ID NO:21) (AGGATCGGGCATTTTTGTAGCGT) и РВ1236 (SEQ ID NO:22) (TTTCTAGAGCTGGTGGCGACTACCGTGAATCCTGGCAACC) с использованием плазмиды pLDR8 в качестве ДНК матрицы.
Данные шесть индивидуальных ПЦР-продуктов кассеты λPr::sacBΩkan или λPr::sacBΩcat, которые следовало перенести в хромосому Е. coli, сшивались вместе после объединяющей ПЦР (Фиг 8А-В-С). Данные шесть индивидуальных амплифицированных фрагментов очищали с помощью набора реагентов для очистки ПЦР-продуктов «PCR clean-up kit» (Geneclean turbo kit; MP Biomedicals, Калифорния, США) и проводили второй раунд ПЦР со всеми 6 фрагментами и без праймеров. Условия ПЦР были следующими: смесь ПЦР (конечный объем 25 мкл), 1 мкл каждого ПЦР-фрагмента, 2 мкл dNTP (2,5 мМ каждого, 5 мкл 5×ПЦР-буфера, 11,5 мкл стерильной дистиллированной воды, 0,5 мкл ДНК-полимеразы «Phusion» (Finenzymes, Финляндия); Циклы ПЦР: 98°С 30 с, (98°С 10 с, 60°С 30 с, 72°С 5 мин)*15 циклов и 72°С 10 мин. Третий раунд ПЦР-амплификации осуществляли с использованием 1 мкл очищенного продукта второго раунда с использованием праймеров РВ1235 и РВ1236 для purN (аллельная замена) и праймеров РВ1237 и РВ1238 для edA (аллельная замена). Условия ПЦР были следующими: ПЦР-смесь (конечный объем 50 мкл), 1 мкл второго ПЦР-фрагмента, 1 мкл dNTP (2,5 мМ каждого, 10 мкл 5×ПЦР-буфера, 37 мкл стерильной дистиллированной воды, 0,5 мкл прямого праймера, 0,5 мкл обратного праймера, 0,5 мкл ДНК-полимеразы «Phusion» (Finenzymes, Финляндия); Циклы ПЦР: 98°С 30 с, (98°С 15 с, 60°С 30 с, 72°С 4 мин)*35 циклов и 72°С 10 мин. Конечные соединенные ПЦР ампликоны (4007 п.о. для аллельной замена purN, и 3407 п.о. для аллельной замены edA) проверяли на агарозном геле и очищали перед трансформацией в E.coli. Каждый ПЦР-соединенный ампликон контролировали рестрикционным анализом.
Линейный ДНК-фрагмент (λPr::sacBΩkan), кодирующий ген sacB, помещенный под контроль промотора λPr (SEQ ID NO:75) и ген устойчивости к канимицину Kan, помещенный под контроль промотора bla, фланкированный двумя длинными участками, гомологичными ДНК-последовательностям, ограничивающим локус-мишень (edA или purN), интегрировали в хромосому Е. coli с помощью электропорации. Получали приблизительно 300 трансформантов-кандидатов (ΔpurN λPr::sacBΩkan или одиночный мутант ΔedA λPr::sacBΩkan) на чашке с LB-агаром, содержащей канамицин в качестве фактора селекционного давления. 20 колоний случайным образом отбирали и очищали с помощью посева штрихом на чашки, содержащие канамицин, а с помощью ПЦР подтверждали, вставку кассеты λPr::sacB+Ωkan в локус edA или purN (см. Фиг 9В). ПЦР с помощью праймеров РВ1196 и РВ1197 осуществляли для контроля вставки Канамицина в хромосому. ПЦР с использованием праймеров РВ1192 и РВ1193 осуществляли для подтверждения присутствия гена sacB в хромосоме. ПЦР с использованием праймеров РВ1204 (SEQ ID NO:23) (GTGGTGCTTATTTCCGGCAACGG) и РВ1205 (SEQ ID NO:24) (CCAGCCACGCGGCGTTTTCGTGC) осуществляли для подтверждения отсутствия гена purN в хромосоме. ПЦР с помощью праймеров РВ1214 (SEQ ID NO:25) (GACCACCGGCCCGGTTGTACCGG) и РВ1215 (SEQ ID NO:26) (CGGACCCGCGATCGCCTGCAGG) осуществляли для подтверждения отсутствия гена edA) (см. Фиг 9В). ПЦР с использованием праймеров РВ1213 (SEQ ID NO:27) (GGTGGATGGCGTCCATTTCTGTGC) и РВ 1196 осуществляли для подтверждения правой и корректной вставки кассеты в локус edA. ПЦР с использованием праймеров РВ1212 (SEQ ID NO:28) (CAAAAGTGTTAAGCGGTAACCTG) и РВ1233 осуществляли для подтверждения корректной вставки левой кассеты в локус edA. ПЦР с использованием праймеров РВ1203 и РВ1196 осуществляли для подтверждения корректной вставки правой кассеты в локус purN. ПЦР с использованием праймеров РВ1202 и РВ1233 осуществляли для подтверждения левой и корректной вставки кассеты в локус purN (см. Фиг 9С). Все контроли ПЦР подтверждали интеграцию в корректный локус. Кроме того, функциональность гена sacB также подтверждали, как представлено на Фиг 10. Созданный родительский штамм ΔpurN λPr::sacBΩkan был не способен расти на чашке с LB-агаром, содержащим сахарозу (ФИГ 10).
Линейный фрагмент ДНК (λPr::sacBΩcat), кодирующий ген sacB (помещенный под контроль промотора λPr), и ген устойчивости к CAT (помещенный под контроль своего собственного промотора cat), фланкированный двумя длинными участками, гомологичными ДНК-последовательностям, ограничивающим локус-мишень (edA), интегрировали в хромосому штамма ΔpurN λPr::sacBΩkan E.coli с помощью электропорации. Получали приблизительно 200 трансформантов-кандидатов (двойной мутант ΔpurN λPr::sacBΩkan ΔedA λPr::sacBΩcat) на чашке с LB-агаром, содержащей хлорамфеникол в качестве фактора селекционного давления. 20 колоний случайным образом отбирали и очищали с помощью посева штрихом на чашки, содержащие хлораменикол, и с помощью ПЦР подтверждали вставку кассеты λPr::sacBΩcat в локус edA (см. Фиг 9С) с помощью праймеров РВ1212 и РВ1213 (см. Фиг 9С). Продукт ПЦР контролировали с помощью секвенирования для подтверждения идентичности λPr::sacBΩcat.
Созданный штамм клеток-хозяев представлен на Фиг.8D. На Фиг.8Е показано сосуществование штамма клеток-хозяев (одна кассета sacB [λPr::sacBΩkan]) и безлекарственной плазмиды. Исходным генотипом E. coli является KanS, CatS, СахарозаR. Интеграцией кассеты в хромосому E. coli ΔpurN λPr::sacBΩkan получали новый генотип, KanR, CatS, СахарозаS. Введение плазмиды, несущей репрессор cI, приводило к получению конечного генотипа KanR, CatR, СахарозаR.
Пример 3: Получение Плазмид, Несущих Кассету, Экспрессирующую Репрессор cI (плазмиды рРВ838-рРВ844-рРВ847-рРВ885-рРВ896)
Краткое изложение Конструирования Плазмид
Получали плазмиды на основе рРВ828, такие как рРВ829 и рРВ844-рРВ847, содержащие открытую рамку считывания cI фага λ под контролем или его собственного промотора или слабого промотора гена устойчивости к канамицину (Р1).
Плазмиду рРВ838, содержащую ген cI, помещенный под контролем слабого промотора гена устойчивости к Канамицину (Р1), получали с помощью промежуточного плазмидного вектора pMCS5, в котором геном хлорамфениколацетилтрансферазы (cat) заменяли ген устойчивости к ампициллину (Фиг.19).
Плазмиды рРВ844 и рРВ845 содержат ген cI (помещенный под контролем промотора Р1), клонированный в вектор рРВ828, который представляет собой производное pVR1012 (pVR1020 или плазмида 1012, VICAL Inc.; Luke et al., 1997; Hartikka et al., 1996, см., например, Пат. США No.5846946 и 6451769), содержащее ген cat. Плазмиды отличаются по ориентации гена cI в плазмиде (ФИГ 20-21).
Плазмиды рРВ846 и рРВ847 содержат ген cI (помещенный под контроль своего собственного (нативного) промотора SEQ ID NO:30), клонированный в вектор рРВ828, который представляет собой производное pVR1012, содержащее ген cat. Плазмиды отличаются по ориентации гена cI в плазмиде (ФИГ 22-23).
рРВ885 получали из плазмиды рРВ838 [ген cI, помещенный под контроль слабого промотора гена устойчивости к Канамицину (Р1)], в которой ген хлорамфениколацетилтрансферазы (cat) удаляют с помощью рестрикционного расщепления с помощью AvrII (ФИГ 24).
Плазмиды на основе рРВ829, такие как рРВ896, содержат ген cI, помещенный под слабый промотор гена устойчивости к Канамицину (Р1). Плазмида рРВ896 содержит маркерный ген GFP, помещенный под контроль эукариотического промотора CMV (ФИГ 25).
Эти плазмиды использовали в качестве векторов для клонирования гетерологичного полинуклеотида, представляющего интерес гена (трансгена) в безлекарственной системе. Каждая плазмида несет ген устойчивости к антибиотику (cat), который может быть вырезан и удален с помощью уникального фермента рестрикции, чтобы сделать возможным размножение плазмиды в подходящем штамме Е. coli sacB+ в присутствии сахарозы без фактора селекционного давления (такого как хлорамфеникол).
Ген устойчивости к антибиотику (cat) в этих плазмидах использовали только для доказательства правильности концепции. Ген cat в конечном счете удаляют в конечных безлекарственных плазмидах (см. ФИГ 13-15 и Пример 4).
Последовательность гена cI из плазмиды pLDR8 (ATCC номер # 77357) продуцирует мутантный белок-репрессор с1857 (NCBI: AB248924, вектор для клонирования pND707, Love, CA. et al., Gene. 17; 176 (1-2): 49-53; 1996).
А. Конструирование плазмиды рРВ829, содержащей открытую рамку считывания репрессора cI, помещенного под контроль слабого промотора Р1
Плазмида рРВ828: (плазмида на основе pVR1012+ген Cat) (ФИГ 17) Получали плазмиды на основе рРВ828, такие как рРВ829, содержащие открытую рамку считывания репрессора cI фага λ под контролем слабого промотора гена устойчивости к канамицину (Р1) и гена Cat, который можно было вырезать по уникальному сайту рестрикции EcoRI. Получали экспрессирующий вектор на основе pVR1012, содержащий ген хлорамфениколацетилтрансферазы (cat), с образованием плазмиды рРВ828. Ген cat из рРВ627 (пропиетарный материал «Merial») представлял собой производное из pNB335 (пропиетарный материал «Merial»), которая сама является производным исходной плазмиды pSW23T (плазмида для клонирования с учетным номером в GenBank No.AY733066). Фрагмент ДНК, соответствующий гену cat (798 п.о.) (SEQ ID NO:31 для ДНК, SEQ ID NO:32 для белка), получали ПЦР с помощью праймеров РВ1184 (SEQ ID NO:33) (GAATTCCGGTCCGGGCGAAAATGAGACGTTGATCGGCS) [Содержащего сайт EcoRI (подчеркнут)] и РВ 1185 (SEQ ID NO:34)
(CCTAGGCTGTGTTAATTAAGGCGCGCCGAATTCCGGTCCGTTACGCCCCGCCCTGCCACTCATCGC) [содержащего сайт EcoRI (подчеркнут)], с использованием pNB350 в качестве матрицы и ДНК-полимеразы «Phusion™ High-Fidelity» (Finnzymes, 02150 Эспо, Финляндия). Полученный а результате продукт ПЦР (832 п.о.) дотировали в вектор pCR blunt Topo (Invitrogen, CA, США) с получением плазмиды pCRblunt/PB1184-1185 (4351 п.о.) (ФИГ 16). Идентичность pCRblunt/PB1184-1185 подтверждали анализом рестрикцией (расщепление PvuII). ДНК-фрагмент, соответствующий гену Cat, получали путем ферментативного расщепления плазмиды pCRblunt/PB1184-1185 с помощью Ecl136II и EcoRV. Фрагмент длиной 897 п.о. лигировали в плазмиду на основе pVR1012, ранее расщепленную с помощью MscI и StuI (3303 п.о.) с получением плазмиды рРВ828 (ФИГ 17). Идентичность рРВ828 подтверждали анализом рестрикцией (расщепление с помощью PvuII и NcoI для определения размера вставки и с помощью EcoRI для подтверждения того, что ген cat правильно удален).
Плазмида рРВ829: (плазмида рРВ828+ген Р1::cI) (ФИГ 18) Конструкт, содержащий ген cI под контролем промотора Р1 (слабый промотор гена устойчивости к Канамицину), получали сшивающей ПЦР. Фрагмент ДНК, соответствующий слабому промотору Р1, получали ПЦР с помощью праймеров РВ1186 (SEQ ID NO:35) (TCATACCAGGCCTAGGTGATACGCCTATTTTTATAGGTTAATG) и РВ1187 (SEQ ID NO:36) (AACACCCCTTGTATTACTGTTTATG), с использованием в качестве матрицы pLL14 (см патентную заявку «Merial» US 2005/0164946) и ДНК-полимеразы «Phusion». Фрагмент ДНК, соответствующий гену cI (SEQ ID NO:1), получали ПЦР с помощью праймеров РВ1188 (SEQ ID NO:37)
(AATACAAGGGGTGTTATGAGCACAAAAAAGAAACCATTAACA) [содержащего комплементарный участок последовательности Р1 в 5'-области (подчеркнут)] и РВ1189 (SEQ ID NO:38)
(CCGGAATTCGGCGCGTCAGCCAAACGTCTCTTCAGGCCACTG) с использованием в качестве матрицы pLDR8 (АТСС #77357) и ДНК-полимеразы «Phusion». Два продукта ПЦР (соответственно, 151 и 729 п.о.) очищали и использовали в качестве матрицы во второй стадии ПЦР с праймерами РВ1186 и РВ1189 и с применением ДНК-полимеразы «Phusion» (Finnzymes, Финляндия) (см. Фиг.24). Два Продукта ПЦР (соответственно, 151 и 729 п.о.) очищали и лигировали с плазмидой рРВ828, расщепленной AscI и AvrII с получением плазмиды рРВ829 (4970 п.о.) (ФИГ 9А). Лигирование проводили с помощью набора «In-Fusion PCR Cloning Kit» (кат. No.740590.250) от «Clontech» (Takara Bio Europe, Сен-Жермен-эн-Лайе, Франция). Клоны рРВ829 отбирали после анализа расщеплением с помощью HindII. Целостность последовательности гена cI и промотора Р1 подтверждали секвенированием плазмиды рРВ829.
Плазмида рРВ896: (рРВ829 плазмида+ген Р1::cI+GFP) (ФИГ 25) Маркерный ген GFP помещали под контроль эукариотического промотора CMV (SEQ ID NO:43). Ген GFP получали путем рестрикционного расщепления плазмиды pCG105 (плазмида, полученная на основе рРВ828, содержащая ген GFP, помещенный под контроль эукариотического промотора CMV, пропиетарный материал «Merial»), с помощью Notl/BglII. Этот вырезанный ген GFP клонировали в плазмиду рРВ829, предварительно разрезанную с помощью Notl/BglII с получением плазмиды рРВ896.
В. Конструирование промежуточной плазмиды рРВ838, и плазмиды рРВ844-рРВ845, содержащих ген cI под контролем промотора Р1 (слабый промотор гена устойчивости к канамицину)
Плазмида рРВ837.1: (плазмида pMCS5+ген Cat)
Фрагмент ДНК, соответствующий гену Cat, получали ПЦР с помощью праймеров РВ1182 (SEQ ID NO:39)
(AGATCTGTTAACGGCGAAAATGAGACGTTGATCGGC) [содержит сайт BglII (подчеркнут) в 5'-области] и РВ1183 (SEQ ID NO:40) (GTCGACGTTAACTTACGCCCCGCCCTGCCACTCATCGC) [содержит сайт Sail (подчеркнут) в 5'-области], с использованием в качестве матрицы рРВ791 [производное плазмиды pNB350] и ДНК-полимеразы «Phusion».
Праймеры ПЦР разрабатывали на основании последовательности cat из pNB350. Три независимых продукта ПЦР (779 п.о.) собирали и лигировали в вектор pCRII с получением плазмиды pCRII+РВ1182-1183 (4734 п.о.). Клоны pCRII+РВ1182-1183 отбирали после расщепления SalI/BglII. Плазмиду pMCS5 (MoBiTec, Германия) расщепляли с помощью SalI/BglII с целью получения фрагмента ДНК А (2950 п.о.). pCRII+РВ1182-1183 расщепляли с помощью SalI/BglII, и фрагмент SalI-BglII выделяли из агарозного геля (797 п.о.: фрагмент В). Фрагменты А и В лигировали для получения плазмид рРВ837.1 (3747 п.о.).
рРВ837.2: (pMCS5+Cat) без промотора bla
Получали плазмиду рРВ837.2, которая не содержит участок промотора bla.
Плазмиду рРВ837.1 расщепляли NaeI и XmnI, фрагмент NaeI-XmnI 3300 п.о. выделяли из агарозного геля (фрагмент А). Фрагмент А лигировали с получением плазмиды рРВ837.2 (3300 п.о.). Клон рРВ837.2 отбирали после расщепления с помощью BglI.
Плазмида рРВ838 (pMCS5+Cat без промотора bla)+Р1::cI (ФИГ 22)
Конструкт, содержащий ген cI под управлением промотора Р1 (слабый промотор гена устойчивости к канамицину), получали сшивающей ПЦР. Фрагмент ДНК, соответствующий слабому промотору Р1, получали ПЦР с помощью праймеров РВ1186 и РВ1187, с использованием pLL14 в качестве матрицы и ДНК-полимеразы «Phusion». Фрагмент ДНК, соответствующий гену cI, получали ПЦР с помощью праймеров РВ1188 и РВ1189, с использованием pLDR8 (ATCC #77357) в качестве матрицы и ДНК-полимеразы «Phusion» (Finnzymes, Финляндия). Два продукта ПЦР (соответственно, 151 и 729 п.о.) очищали и использовали в качестве матрицы во второй стадии ПЦР с праймерами РВ1186 и РВ1189 с использованием ДНК-полимеразы «Phusion». Два независимых продукта ПЦР (880 п.о.) собирали и лигировали в вектор pCRII blunt с получением плазмиды pCRblunt+РВ1186-1189 (4400 п.о.). Клоны pCRII blunt+РВ1186-1189 отбирали после расщепления EcoRV/SpeI. pCRII blunt+РВ1186-1189 расщепляли EcoRV и SpeI с целью получения EcoRV-SpeI фрагмента А (936 п.о.). рРВ837.2 расщепляли EcoRV и SpeI, и выделяли фрагмент EcoRV-SpeI из агарозного геля (3464 п.о.: фрагмент В). Фрагменты А и В лигировали с получением плазмид рРВ838 (4216 п.о.). Клон рРВ838 отбирали после расщепления HindIII. Целостность последовательности гена cI и промотора Р1 подтверждали с помощью секвенирования.
рРВ885 плазмида (pMCSS+Cat без промотора bla)+Р1::cI (ФИГ 24)
Эту плазмиду получали на основе плазмиды рРВ838, где ген хлорамфениколацетилтрансферазы (cat) удаляли с помощью рестрикционного расщепления с помощью AvrII (ФИГ 24). Плазмида рРВ885 содержит ген cI под контролем слабого промотора гена устойчивости к канамицину (Р1). Плазмиду рРВ885 выделяли как описано в примере 4 (см. ниже), после трансформации штамма клеток-хозяев Е. coli ΔpurN λPr::sacBΩkan путем прямой селекции на чашке с LB-агаром, содержащим 10% сахарозу.
Плазмиды рРВ844-рРВ845 (ФИГ 23-24)
Фрагмент ДНК, соответствующий гену cI под контролем промотора Р1, получали ПЦР с помощью праймеров РВ1186 и РВ1263 (SEQ ID NO:41)
(TCAGCCAAACGTCTCTTCAGGCCAC), с использованием плазмиды рРВ838 в качестве матрицы и ДНК-полимеразы «Phusion». Четыре независимых ПЦР-продукта РВ1186-РВ1263 (864 п.о.) собирали и лигировали в вектор pCRblunt с получением плазмиды pCRblunt+РВ1186-1263 (4383 п.о.). Клоны pCRblunt+РВ1186-1263 селектировали после расщепления HindIII.
Плазмиду pCRblunt+РВ1186-1263 расщепляли EcoRI, и выступающие концы достраивали с помощью полимеразы фрагмент Кленова для получения фрагмента с тупыми концами (929 п.о.: фрагмент А). Плазмиду рРВ828 расщепляли с помощью XmnI, и полученный в результате фрагмент XmnI-XmnI очищали (фрагмент В: 4135 п.о.). Фрагменты А и В лидировали для получения плазмид рРВ844 или рРВ845. Кандидаты скринировали рестрикцией BamHI/HmdIII. В плазмиде рРВ844 (см. Фиг.20), ген cI находится в той же ориентации, что и представляющий интерес трансген, будучи клонированным в MCS (сайт множественного клонирования) вектора под контролем промотора CMV (SEQ ID NO:43). В плазмиде рРВ845 (см. Фиг.21) cI находится в противоположной ориентации.
С. Получение плазмид РВ846-рРВ847 с геном cI и его нативными промоторами
Фрагмент ДНК, соответствующий гену cI, под его собственным промотором (нативный промотор cI: SEQ ID NO:30) получали ПЦР с помощью праймеров РВ1266 (SEQ ID NO:42) (GCTGACTCATACCAGGCACGCACGGTGTTAGATATTTATCCC) и РВ1263, с использованием плазмиды pLDR8 в качестве матрицы и ДНК-полимеразы «Phusion».
Плазмиды рРВ846-рРВ847
Четыре независимых продукта ПЦР РВ1266-РВ1263 (777 п.о.) собирали и лигировали в вектор pCRblunt для получения плазмиды pCRblunt+РВ1266-1263 (4296 п.о.). Клоны pCRblunt+РВ 1266-1263 отбирали после рестрикции HindIII.
Плазмиду pCRblunt+РВ1266-1263 расщепляли с помощью EcoRI, и выступающие концы достраивали с помощью полимеразы фрагмент Кленова для получения фрагмента с тупыми концами (842 п.о.: фрагмент А). Плазмиду рРВ828 расщепляли с помощью XmnI, и очищали фрагмент XmnI-XmnI (фрагмент В: 4135 п.о.). Фрагменты А и В лигировали для получения плазмид рРВ846 (4999 п.о.) или рРВ847. Кандидаты скринировали с помощью рестрикции BamHI/HindIII. В плазмиде рРВ846 (см. Фиг.22), ген cI находится в той же ориентации, что и ген, представляющий интерес, будучи клонированным в сайт MCS вектора под контролем промотора CMV. В плазмиде рРВ847 (см. Фиг.23) ген cI находится в противоположной ориентации.
D. РЕЗУЛЬТАТЫ КОНСТРУИРОВАНИЯ
На Фиг.7 представлены конструкты рРВ838 и рРВ829, несущие репрессор cI, которые использовали для получения безлекарственных плазмид. Обе плазмиды маркировали с помощью гена устойчивости к Хлорамфениколу (CatR). Ген Cat в этих плазмидах требовался только для определения доказательство правильности концепции, и в конечном счете его можно полностью удалить (см. Фиг.13-15 и Пример 4).
Пример 4: Трансформация штамма клеток-хозяев Е. coli плазмидами с cI и прямая селекция на чашках с агаром и сахарозой
Была эффективно продемонстрирована экспериментальная процедура, которая позволяет осуществить на чашках с агаром скрининг клеток трансформированных маркированными хлорамфениколом (Cat) плазмидами с cI без применения антибиотика в качестве фактора селекционного давления (см. ФИГ 13-15). Компетентные клетки Е. coli λPr::sacB+ ΔpurNΩkan трансформировали 1 мкг либо плазмиды рРВ838, либо плазмиды рРВ829 и инкубировали в течение 3 часов при 30°С в жидкой среде LB. Аликвоту 100 мкл разведенных (10-3 и 10-4) трансформированных клеток (с плазмидами рРВ838 и рРВ829) высевали на чашку с LB-агаром, содержащим 10% сахарозу. Данные разведения в интервале от 10-3 до 10-4 оказались подходящими для отбора истинных трансформированных клеток (например, содержащих плазмиду с cI), так как 98-100% клеток, устойчивых к сахарозе, растущих на чашке с LB-агаром с сахарозой, были CatR. Предполагается, что данные трансформированные клетки содержат плазмиду с cI (рРВ829 или рРВ838).
На ФИГ 13А показано, что компетентные клетки Е. coli λPr::sacB+ ΔpurNΩkan трансформировали с помощью 1 мкг либо плазмиды рРВ838, либо плазмиды рРВ829 и инкубировали в течение 3 часов при 30°С в жидкой среде LB перед дот-спот анализом (dot-spotting) (4 мкл для каждого дот-спот анализа). Фигура демонстрирует, Log-разведения (в интервале от неразведенных до разведения 6Log) на чашках с LB-агаром, содержащих или хлорамфеникол (15 мкг/мл) или сахарозу (10%). Не трансформированные компетентные клетки штамма Е. coli λPr::sacB+ применяли в качестве стандартных клеток, растущих в аналогичных условиях. Как ожидалось, стандартные клетки не росли на селективных чашках с cat и сахарозой. Тем не менее, благодаря продолжительному периоду инкубации (3 дня), в этих условиях появлялись некоторые спонтанные мутанты, устойчивые к сахарозе. Уровень спонтанных мутаций определяет исходную точку, рассматриваемую для скрининга истинных трансформированных клеток. Наблюдали различия в два Log для трансформированных клеток, растущих на cat или сахарозе в условиях данного эксперимента. Компетентная клетка по определению является клеткой с временно ослабленной клеточной стенкой. Инкубация трансформированных клеток в жидкой среде LB после электропорации позволяет осуществить регенерацию как вышеупомянутой ослабленной клеточной стенки, так и регенерацию пор, индуцированных в клеточной стенке и клеточной мембране после электропорации. Так как компетентные клетки генетически чувствительны к сахарозе благодаря вставке кассеты λPr::sacB+ ΔpurNΩkan, то выход трансформированных клеток в присутствии сахарозы был уменьшен в два Log по сравнению с выходом клеток на Cat. Эти наблюдения подтверждались при рассеве 100 мкл трансформированных клеток на селективных чашках (см. ФИГ 13В).
На ФИГ 13В показано, что 100 мкл трансформированных (плазмидами рРВ838 и рРВ829) и не трансформированных стандартных клеток рассевали с использованием подходящего Log разведения на чашки с LB-агаром, содержащим либо cat, либо сахарозу для определения степени эффективности трансформации на cat и на сахарозе, и осуществляли стандартное рассевание на сахарозе для лучшего определения исходного уровня спонтанных мутаций, рассматриваемых для успешного скрининга истинных трансформированных клеток, способных расти на чашках с LB-агаром и сахарозой. Согласно предыдущему эксперименту дот-спот анализа клеток (см. ФИГ 13А), спонтанные мутанты, устойчивые к сахарозе, не детектировали при рассевании 10-3 трансформированных клеток на чашки с LB-агаром, содержащим сахарозу. Как показано ранее (см. ФИГ 13А), между селекцией на cat и на сахарозе в данных условиях существует различие в 2Log в эффективности трансформации. Разведения в интервале от 10-3 до 10-4 при рассеве на LB-агар, содержащий сахарозу, оказались подходящими для селекции истинных трансформированных клеток (например, содержащих плазмиду с cI).
На ФИГ 14А-С продемонстрирована эффективность экспериментальной процедуры селекции трансформированных клеток только с плазмидой с cI (рРВ829 и рРВ838) без применения антибиотика в качестве фактора селекционного давления.
ФИГ 14А демонстрирует экспериментальный анализ реплик чашек для подтверждения скрининга истинных трансформированных клеток, содержащих плазмиду с cI. Подобно тому, что показано на ФИГ 13В, родительские компетентные клетки λPr::sacB+ ΔpurNΩkan трансформировали плазмидой с cI (рРВ829 и рРВ838), и подходящие Log-разведения клеток рассевали на селективные чашки (Cat или Сахароза) с LB-агаром перед инкубацией в течение 3 дней при 30°С. После этого с чашек, демонстрирующих титр КОЕ (колониеобразующие единицы) в интервале от 20 до 150, делали реплики на новых чашках со свежим LB-агаром, содержащим либо Cat, либо сахарозу или их комбинацию перед инкубацией в течение 3 дней при 30°С. Теоретически, устойчивые к Cat колонии должны соответствовать клеткам, несущим плазмиды с cI.
На Фиг 14В представлены экспериментальные результаты, подтверждающие, что после анализа реплик с чашек значения КОЕ CatR были идентичны на чашках с LB-агаром и сахарозой, и на чашках с LB-агаром и Cat/сахарозой. Как показано на ФИГ 14В (панель В), КОЕ CatR i) все были устойчивы к сахарозе и ii) все устойчивы к комбинации Cat и сахарозы, что означает, что все родительские клетки после трансформации несут плазмиды с cI. На панели С показано, что разведения 10-3 и 10-4 трансформированных клеток, отобраных непосредственно на сахарозе, также были устойчивы к сахарозе и к комбинации сахарозы и Cat, что также означает, что эти устойчивые к сахарозе КОЕ соответствуют трансформированным клеткам, содержащим плазмиду с cI. Действительно, когда нетрансформированные родительские клетки высевали на чашку с LB-агаром, содержащим сахарозу, то устойчивые к сахарозе КОЕ при репликах на среду с комбинацией сахарозы и Cat не росли, как и ожидалось, так как устойчивость к хлорамфениколу представляет собой фенотипическую особенность, привнесенную плазмидой рРВ829 или рРВ838.
На Фиг 14С представлены статистические данные, полученные при анализе чешек с репликами (см. ФИГ 14В), а более конкретно, процент колоний, содержащих плазмиду с cI, которые отобрались без применения антибиотика. В условиях эксперимента, процент колоний, содержащих плазмиду, находится в интервале от 98% до 100%.
Пример 5: Различные условия культивирования в присутствии или в отсутствии сахарозы, демонстрирующие, что плазмиду с cI можно поддерживать
Клетки Е. coli, экспрессирующие sacB, ΔpurN λPr::sacB+ΩKan, выращивали в присутствии или в отсутствии сахарозы с целью контроля функциональности кассеты λPr::sacB+, вставленной в хромосому Е. coli.
На Фигуре 4 дано описание «чувствительности к сахарозе» инактивированной системы, где температурный переход на 42°С ингибирует активность связывания репрессора cI с промотором Pr фага λ и придает клеткам-хозяевам чувствительность к сахарозе. Белок репрессора cI, экспрессируемый плазмидой, ингибирует транскрипцию токсичного продукта гена sacB, помещенного под контроль промотора Pr фага λ, который локализован на хромосоме клеток-хозяев. Выживаемость клеток-хозяев Е. coli в присутствии сахарозы гарантировалась путем достаточного уровня экспрессии плазмиды белка-репрессора cI фага λ. На Фиг.5 дано описание объединения штамма клеток-хозяев с плазмидой pDL1 без устойчивости к антибиотику в присутствии сахарозы для наращивания безлекарственной плазмиды. Экспериментальный план для доказательства правильности выбранной концепции дополнительно иллюстрируется на ФИГ 6А, 6В, и 6С.
Физиологические характеристики клеток-хозяев, экспрессирующих sacB, ΔpurN Ω λPr::sacB+ΩKan при росте в присутствии или в отсутствии сахарозы, описаны в Таблице 1.
Физиологические характеристики клеток-хозяев, экспрессирующих sacB (λPr::sacB+), вместе или без плазмиды, при росте в присутствии или в отсутствии сахарозы, описаны в Таблице 2.

На ФИГ 10-11 представлены результаты, подтверждающие, что генетически созданные штаммы клеток-хозяев были высоко устойчивы к сахарозе при 30-37°С, в то время как рост не наблюдался при 42°С. На Фиг.10 показано, что второй пассаж штамма клеток-хозяев Е. coli ΔpurN λPr::sacB+Ωkan, несущего либо плазмиду рРВ829, либо плазмиду рРВ838, хорош рос в присутствии сахарозы при 32°С. Экспрессия гена sacB полностью подавлялась продуктом гена cI, синтезируемого обеими плазмидами с cI. Поддержание плазмиды было на 100% эффективным (в сравнении с контролем на CatR) и клетки оказались способны выжить в присутствии сахарозы. При 42°С продукт гена cI инактивировался. Плазмиды не могли поддерживаться и в присутствии сахарозы клетки погибали. Это говорит о том, что выживаемость клеток в присутствии сахарозы имела место благодаря присутствию функциональной плазмиды с cI.
Пример 6: Условия культивирования, демонстрирующие, что плазмиды с cI стабильны в течение около 5 пассажей без селекционного давления антибиотиком и демонстрирующие отсутствие спонтанных мутаций в отношении устойчивости к сахарозе.
После получения безлекарственных плазмид, их тестировали на стабильность и устойчивость на протяжении нескольких пассажей в свежей жидкой культуральной среде, содержащей сахарозу или Cat.
На Фиг.11 показана стабильность плазмид рРВ829 и рРВ838 в растущих клетках, включая отсутствие обычных генетических перестроек во время селекции на хлорамфениколе или на сахарозе. Сравнимые выходы плазмид и их поддержание в культурах, растущих в присутствии сахарозы и хлорамфеникола, наблюдали даже после пяти последовательных пассажей. Системы с устойчивостью к антибиотику и без устойчивости к антибиотику были, по меньшей мере, эквивалентны в отношении поддержания плазмид и селекции растущих клеток.
На Фиг.12 показано отсутствие спонтанных мутаций в кассете λPr..sacB, вставленной в родительский штамм Е. coli, при росте клеток при 30°С. Это наблюдение очевидно демонстрирует стабильность плазмиды с cI в таком генетическом окружении как в Е. coli, при росте клеток в присутствии сахарозы (см. Фиг 10). Данный эксперимент для определения частоты мутаций осуществляли с помощью родительского штамма (с делецией purN и с экспрессией гена sacB). Было продемонстрировано, что мутации не появлялись при 30°С через 18 ч инкубации. Клональные культуры родительских клеток, несущих кассету sacB+ в своей хромосоме, росли в среде LB, содержащей сахарозу. Клетки собирали при OD 1. и аликвоты по 100 мкл высевали на чашки с LB-агаром, содержащим сахарозу. Так как присутствие гена sacB вызывает клеточную смерть, то частоту спонтанных мутаций оценивали путем подсчета устойчивых к сахарозе колоний после инкубации в течение ночи при 30°С. Стандартные клетки, несущие кассету sacB+, но трансформированные с помощью плазмиды cI (рРВ829 или рРВ838) стали устойчивыми к сахарозе, и для подсчета культуру с DO 1 их следует развести в 5 Log перед высеванием (100 мкл) на чашки с LB-агаром, содержащим сахарозу. После культивирования в течение ночи (18-часовая инкубация), в родительских клеточных культурах (ΔpurN λPr::sacBΩkan) не детектировали колоний, устойчивых к сахарозе, в то время как культуры ΔpurN λPr::sacBΩkan с разведением 5Log, трансформированные плазмидами рРВ829 или рРВ838, росли в присутствии сахарозы. Потребовалось тридцать шесть часов для детектирования некоторых устойчивых к сахарозе колоний (ΔpurN λPr::sacBΩkan) при инкубации клеток при 37°С.
Спонтанные мутации происходили с частотой 1 мутация/1,3×105 клеток через 36 часов при инкубации при 37°С. Поддержание плазмиды с cI в клетках, растущих в присутствии сахарозы, было эффективно после ночной инкубации при 30°С, в то время как рост материнского штамма не детектировался.
Пример 7: Улучшение Родительского штамма клеток-хозяев Е. coli для уменьшения частоты спонтанных мутаций по сахарозе, путем добавления второй кассеты sacB (ΔedA λPr::sacBΩcat) в вышеупомянутый штамм Е. coli с одной кассетой sacB (ΔpurN λPr::sacBΩkan).
Обе фигуры, ФИГ 8С и ФИГ 9D, иллюстрируют создание нового родительского штамма клеток-хозяев Е. coli, несущего две кассеты sacB (ΔpurN λPr::sacBΩkan ΔedA λPr::sacBΩcat).
На Фигурах 10В-С показано, что штамм Е. coli, несущий продублированные кассеты с sacB (ΔpurN λPr::sacB+Ωkan ΔedA λPr::sacB+Ωcat), является высокочувствительным (отсутствие роста) в присутствии сахарозы при инкубации при 30°С и 42°С, хотя при посеве на среду, содержащую 10% сахарозы, случаются спонтанные мутации штаммов Е. coli, с одной кассетой sacB (ΔpurN λPr::sacBΩkan).
Фиг.10С также демонстрирует самую низкую концентрацию сахарозы (конечная концентрация 2%), требуемую для штамма Е. coli, содержащего две кассеты sacB, по сравнению с штаммом Е. coli, содержащим одну кассету sacB (конечная концентрация 10%). Кроме того, очевидно, что штамм Е. coli, содержащий две кассеты sacB, гораздо более чувствителен к сахарозе, что продемонстрировано при низкой % сахарозы и при самой высокой температуре, такой как 42°С. Это предполагает, что температура около 37°С и концентрация сахарозы около 2% (или чуть ниже) может соответствовать оптимальным условиям для поддержания плазмид, несущих ген репрессора cI в растущих клетках.
Фигуры 12В-С демонстрируют, что присутствие двух кассет sacB (а не одной) придает большую устойчивость по отношению к чувствительности к сахарозе (оставаясь оптимальной при 2% сахарозы) при температурах в интервале от 30°С до 37°С. Клетки E. coli, трансформированные плазмидами, содержащими две кассеты sacB, BJ5183 ΔpurNΩ λPr::sacBKm ΔedAΩ λPr::sacB Cat клон #1 и BJ5183 ΔpurNΩ λPr::sacBKm ΔedAΩ λPr::sacB Cat клон #2, и клетки E. coli, трансформированные плазмидами, содержащими одну кассету sacB, BJ5183 ΔpurNΩ λPr::sacBKm клон #16 и BJ5183 ΔedAΩ λPr::sacBKm клон #11, растили до OD600 около 1. Около 100 мкл каждой культуры высевали на чашки с LB-агаром, содержащим 0%, 2% и 4% сахарозы (разведение 10-5) для подсчета КОЕ. Чашки инкубировали при 30°С-37°С. Инкубация при 37°С использовали для оценки устойчивости родительского штамма, экспрессирующего две кассеты sacB, в присутствии 2% и 4% сахарозы. ФИГ 12В-С и Таблицы 3-7 ниже демонстрируют, что Е. coli с одной кассетой sacB в хромосоме, были менее устойчивы при концентрации сахарозы в интервале от 2 до 4% при одинаковых температурах, таких как 30°С и 37°С. В данном анализе с рассеванием подтвердили, что частота мутаций по сахарозе штамма Е. coli с двумя кассетами sacB была не детектируемой при самой низкой концентрации сахарозы (2%) при инкубации при 37°С, в то время как частота мутаций штамма Е. coli с одной кассетой sacB колебалась в интервале от 3,8×10-6 до 5×10-5. Ряд независимых экспериментов продемонстрировал, что частота мутаций штамма Е. coli с двумя кассетами sacB составила около 5×10-10 при инкубации при 37°С в присутствии 2% сахарозы.



Пример 8: Поддержание плазмиды и выход плазмиды, полученные в отсутствии селекционного давления антибиотиком (плазмида с cI поддерживается с высоким количеством копий в штамме клеток-хозяев)
Клетки-хозяева Е. coli, экспрессирующие sacB и несущие плазмиду с cI (рРВ829 или рРВ838) выращивали в отсутствии селекционного давления антибиотиком (например, в присутствии сахарозы) или в присутствии Cat в качестве контроля.
На Фиг.11 выход плазмиды, определенный на 2 и 5 пассажах. Плазмиды экстрагировали из культур, поддерживаемых при 30°С, и подвергали гель-электрофорезу. Рестрикционное расщепление HindII обозначает, что отсутствуют обычные генетические перестройки, происходящие или в рРВ829 или рРВ838 во время размножения и селекции или с Cat или с Сахарозой. Кроме того, аналогичное поддержание плазмид с cI наблюдали во время селекции плазмид с cI под селекционным давлением либо Cat, либо сахарозы.
Фиг.15А-В иллюстрируют стабильность и эффективность поддержания плазмид с cI (рРВ829 и рРВ838, содержащих одну копию кассеты с sacB) в культурах, включая отсутствие генетической перестройки после селекции на хлорамфениколе или сахарозе. Наблюдали эффективное поддержание плазмид и стабильность плазмид с cI, выращенных в присутствии или сахарозы или хлорамфеникола. Кроме того, выход плазмид (количество копий) был гораздо выше (приблизительно 700 на клетку), если штамм клеток-хозяев, трансформированных либо рРВ838, либо рРВ829, выращивали при 30°С в присутствии сахарозы в качестве фактора селекционного давления, по сравнению с клетками, культивируемыми в присутствии Cat в качестве фактора селекционного давления. Поддержание плазмид и выход плазмид были высоко эффективны, а количество копий было в два раза выше по сравнению с клетками, культивируемыми в присутствии Cat. Данный хороший выход плазмид при культивировании клеток-хозяев в присутствии сахарозы был получен благодаря слабому промотору (Р1), который контролирует экспрессию cI. Действительно, экспрессия репрессора cI под контролем слабого промотора гена устойчивости к канамицину (Р1) оказывала положительный эффект на выход плазмид лучшим противодействием токсичности гена sacB, помещенного под контроль промотора Pr фага λ.
Ранее было показано (см. ФИГ 11А), что системы с устойчивостью к антибиотикам и без устойчивости к антибиотикам эквивалентны по части поддержания плазмид и селекции растущих клеток, несущих плазмиду с cI. На ФИГ 15В продемонстрировано, что выход плазмид (количество копий) был гораздо выше (приблизительно 700 на клетку), при росте родительского штамма клеток-хозяев, трансформированных либо рРВ838, либо рРВ829, в присутствии сахарозы, играющей роль фактора селекционного давления, по сравнению с клетками, культивируемыми в присутствии Cat, играющего роль фактора селекционного давления. Поддержание плазмид и выход плазмид были высоко эффективны, а количество копий было в два раза выше по сравнению с клетками, культивируемыми при селекции антибиотиком, таким как Cat.
Пример 9: Поддержание и выход плазмид при отсутствии гена устойчивости к антибиотику в плазмиде с cI: сравнительный анализ между штаммами Е. coli, содержащими одну или две кассеты с sacB
Фигуры 15С-Е иллюстрируют стабильность и выход плазмиды рРВ885 без устойчивости к cat в штаммах Е. coli, содержащих одну и две кассеты с sacB (ΔpurN λPr::sacB+Ωkan и ΔpurN λPr::sacB+Ωkan ΔedA λPr::sacB+ΩkanCat, соответственно). Оба родительских штамма ΔpurNΩ λPr::sacBkm (одна кассета с sacB) и ΔpurNΩ λPr::sacBKm ΔedAΩ λPr::sacBCat (две кассеты с sacB) трансформировали рРВ885, плазмидой с cI без устойчивости к cat, и инкубировали в течение 3 часов в среде LB перед посевом на чашку с сахарозой, а затем культивировали (3 пассажа) в жидкой ростовой среде LB, содержащей сахарозу. Выход плазмид определяли на 3 пассаже в среде LB, содержащей 5% и 10% сахарозы, для штаммов Е. coli с двумя и одной кассетой с sacB, соответственно. Плазмиды рРВ885 экстрагировали из культур, поддерживаемых при 37°С, и подвергали гель-электрофорезу. Рестрикционный анализ с PvuII показал отсутствие каких-либо явных генетических перестроек в рРВ885 во время размножения и селекции с помощью сахарозы (см. ФИГ 15D). На этом геле показано отсутствие каких-либо явных генетических перестроек после селекции с помощью 10% и 5% сахарозы в обоих штаммах клеток-хозяев Е. coli, содержащих одну и две кассеты с sacB, соответственно.
Фигура 15Е иллюстрирует выход плазмиды рРВ885 без cat в обоих штаммах Е. coli с одной и с двумя кассетами с sacB (ΔpurN λPr::sacB+Ωkan и ΔpurN λPr::sacB+Ωkan ΔedA λPr::sacB+ΩkanCat, соответственно). Интересно, что в клетках с двумя кассетами с sacB выход плазмид (количество копий) был гораздо выше (приблизительно 278 на клетку) по сравнению с выходом плазмид (приблизительно 142 на клетку), полученным с использованием штамма Е. coli с одной кассетой с sacB. Выход плазмид, полученный в данном эксперименте, оказался ниже, чем выход плазмид, полученный в предыдущем сравнительном исследовании (см. ФИГ 15В). Это произошло главным образом из-за того, что i) для идентификации лучшего варианта после трансформации с рРВ885 необходимо оценить больше клонов и ii) кроме того, условия роста штамма Е. coli с двумя кассетами с sacB, трансформированного рРВ885, плазмидой без cat, не были оптимальными для культивирования. Действительно, мы показали, что оптимальными условиями роста для данного штамма является температура около 37°С и концентрация сахарозы около 2% (или чуть ниже). Однако этот результат подтверждает, что в условиях оптимальной устойчивости выход плазмид был лучше в штамме Е. coli с двумя кассетами с sacB при сравнении со штаммом Е. coli с одной кассетой с sacB.
Пример 10: Трансфекция клеток СНО плазмидой с cI без устойчивости к антибиотику
Фигура 15F и Таблица 8 ниже иллюстрируют эффективность трансфекции (по экспрессии GFP) в клетках СНО плазмиды без устойчивости к антибиотику (рРВ896/сахароза в качестве фактора селекционного давления) и плазмиды с устойчивостью к антибиотику (pCG105/Cat в качестве фактора селекционного давления). По экспрессии белка GFP не наблюдали какого-либо очевидного различия между этими двумя факторами селекционного давления (Сахароза в сравнении с антибиотиком Cat). Плазмиду pCG105 (полученная из рРВ828 плазмида, содержащая ген GFP, помещенный под контроль промотора CMV [от «Merial»]) и плазмиду рРВ896 (рРВ829 плазмида+ген Р1::cI+GFP) использовали для трансфекции в клетки СНО для оценки функциональности концепции плазмид без устойчивости к антибиотику по сравнению с функциональностью существующих плазмид с устойчивостью к антибиотику. Этот эксперимент осуществляли с помощью плазмид, выделенных из культуры штамма клеток-хозяев Е. coli, содержащих одну кассету с sacB. pCG105 наращивали в Е. coli с использованием канамицина в качестве фактора селекционного давления, а рРВ896 наращивали в Е. coli с использованием 10% сахарозы в качестве фактора селекционного давления. ДНК-плазмиды экстрагировали и дозировали в одинаковой концентрации.
ФИГ 15F иллюстрирует in vitro экспрессию белка GFP, кодируемого pCG105 и рРВ896, после временной трансфекции клеток СНО-К1 с помощью «Lipofectamine 2000». Клетки СНО-К1 с конфлюентностью 90% в чашках диаметром 6 см трансфецировали 5 мкг плазмиды и 10 мкл «Lipofectamine» согласно инструкции производителя. После трансфекции клетки культивировали в среде MEM-glutamax, содержащей 1% SVF, в течение 24 часов. Культуру собирали и анализировали с помощью флуоресцентной микроскопии (ФИГ 15G) и проточной цитометрии (FACS Calibur [Becton Dickinson]) (Таблица 8).
На ФИГ 15F показано, что похожую эпифлуоресценцию GFP визуализировали во всех трансфецированных клетках СНО-К1. Плазмиды pCG105 и рРВ96 под селекционным давлением хлорамфениколом и сахарозой трансфецировали в клетки СНО-К1 со сравнимой эффективностью.
В Таблице 8 показана эффективность трансфекции СНО плазмидами pCG105 и рРВ896. Как указано в данной таблице, сравнительный % клеток СНО, экспрессирующих белок GFP, определяли, когда СНО трансфецировали плазмидой pCG105 (59% экспрессии GFP) и рРВ896 (48% экспрессии GFP). Эти результаты удостоверяют правильность применения системы плазмид без устойчивости к антибиотику с селекцией на сахарозе в приложениях, связанных с трансфекцией клеток СНО и с ДНК-вакцинами.

Данные Примеры демонстрируют, что контроль токсичного гена, локализованного на хромосоме, может быть достигнут с помощью репрессора, локализованного на плазмиде. Система полностью функциональна в отсутствии селекционного давления антибиотиком. Система стабильна в течение около пяти (5) пассажей клеток-хозяев. Система дает возможность в клетках-хозяевах достигать высокого количества копий плазмид на клетку. Система полностью совместима с применением минимальной синтетической культуральной среды.
Обладая, таким образом, подробно описанными предпочтительными воплощениями настоящего изобретения, следует понимать, что изобретение, описанное в абзацах выше, не ограничено определенными деталями, вышеизложенными в описании, поскольку возможно множество очевидных вариаций без выхода за рамки смысла и сущности настоящего изобретения.
Все процитированные документы или ссылки настоящего документа («документы, процитированные в настоящем документе») и все процитированные документы или ссылки документов, процитированных в настоящем документе, вместе с какими-либо инструкциями производителя, описаниями, техническими описаниями изделия, технологическими картами для любых продуктов, упомянутых в настоящем документе, или в любом документе, включенном ссылкой в настоящий документ, настоящим включены в данный документ ссылкой, и могут быть использованы при практическом осуществлении изобретения.






















































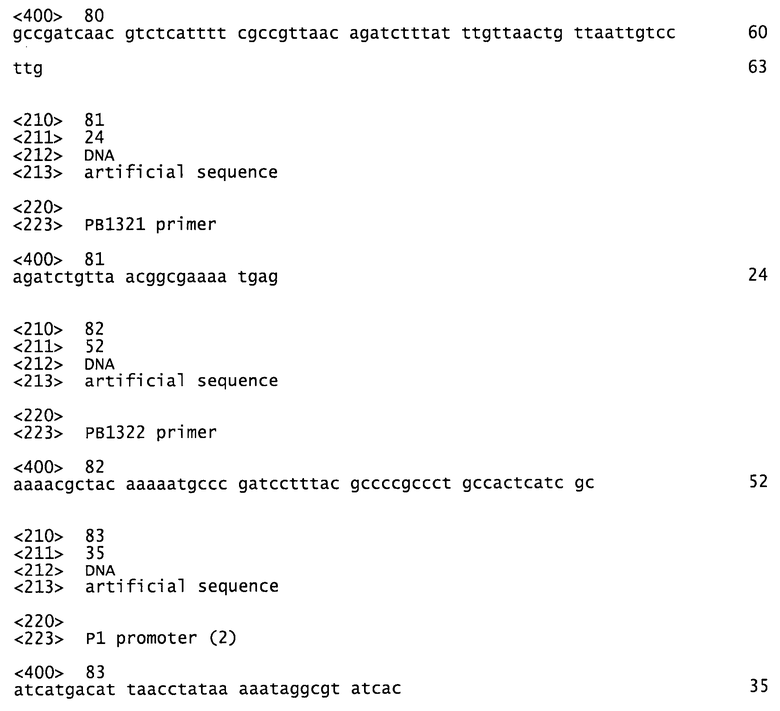
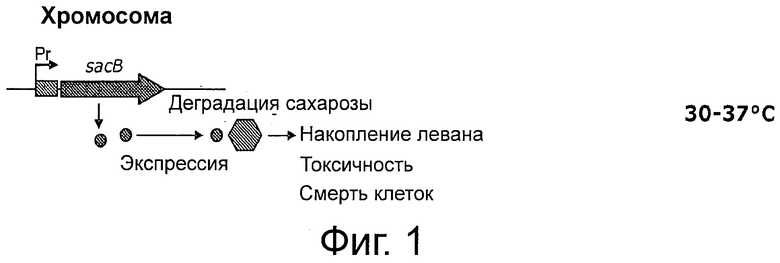
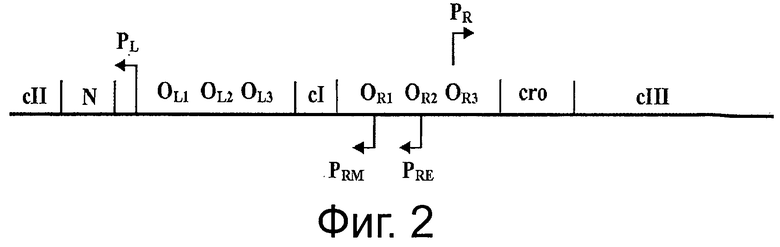

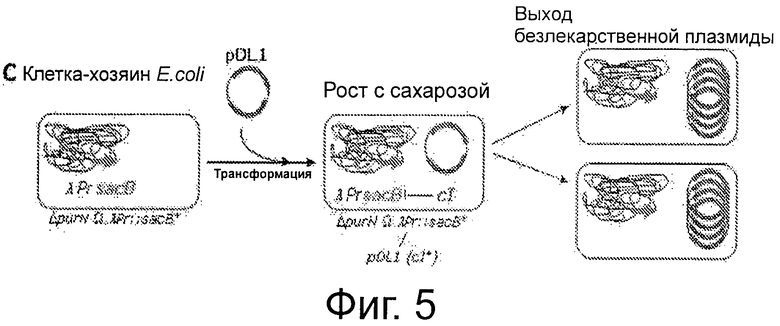
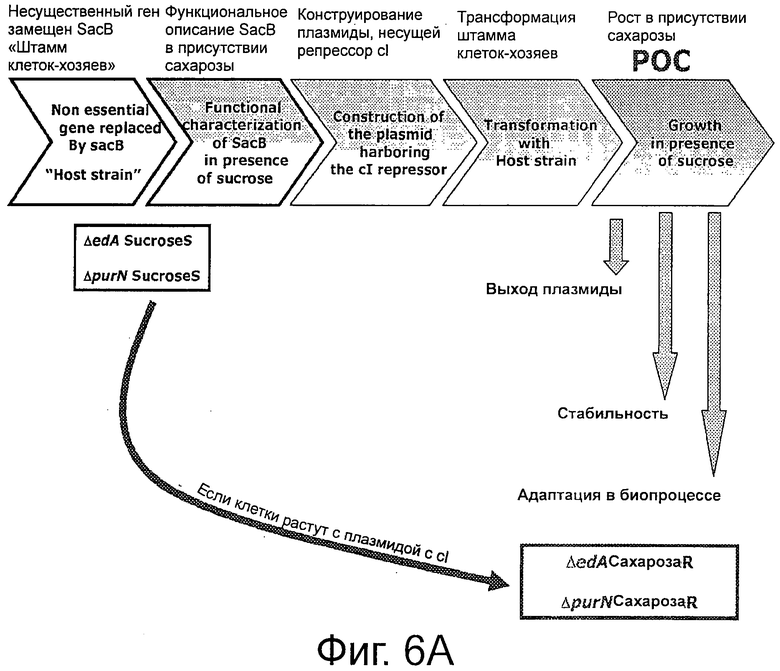
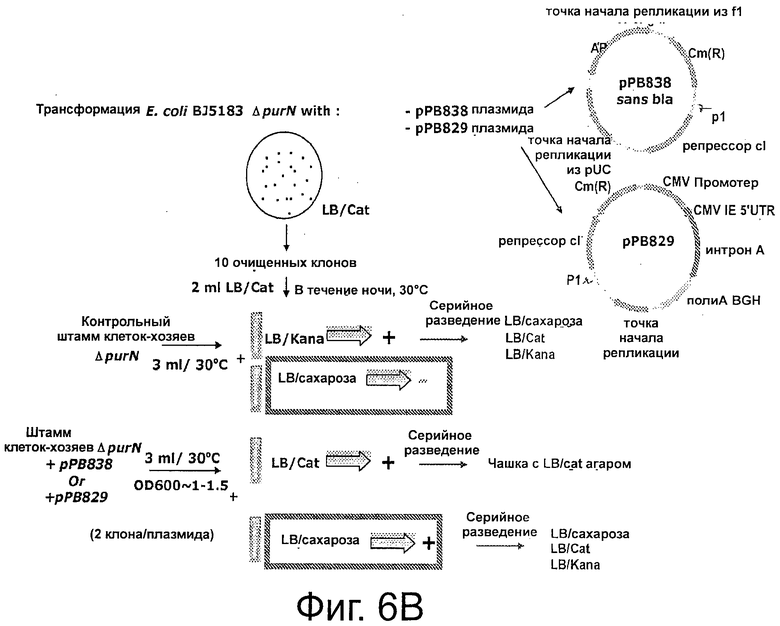
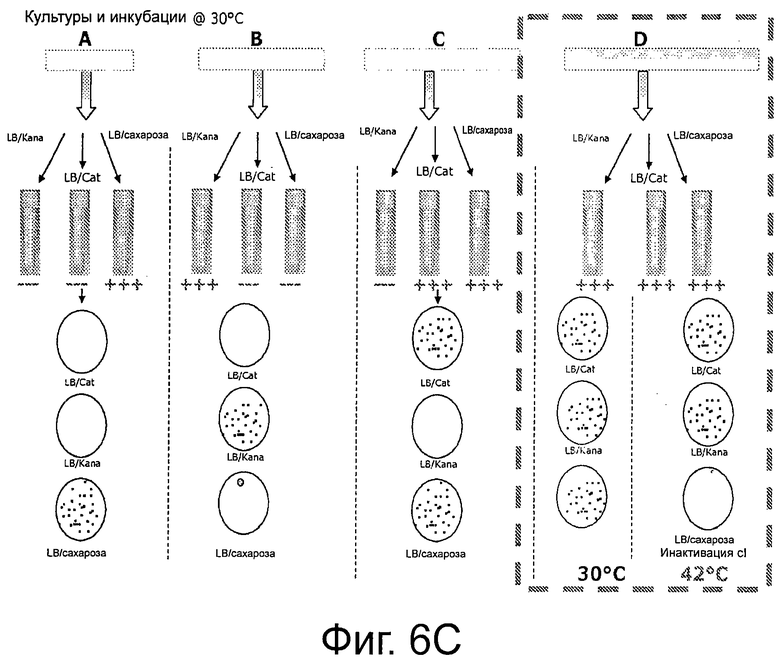
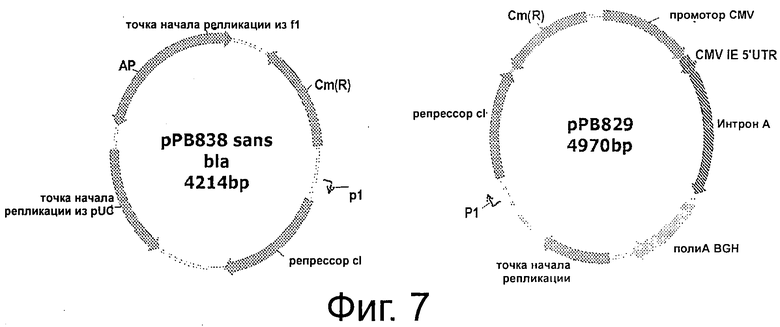

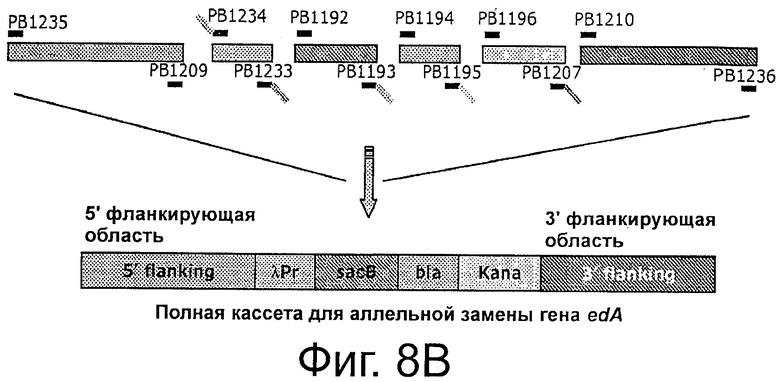
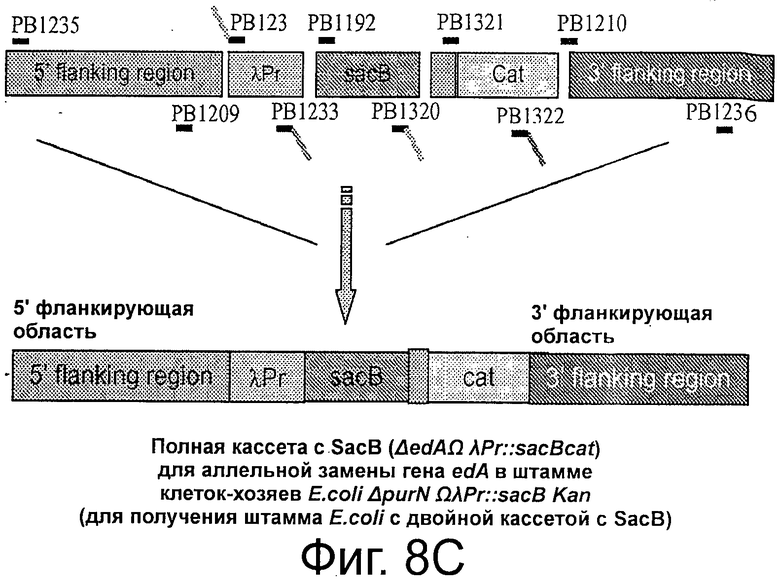
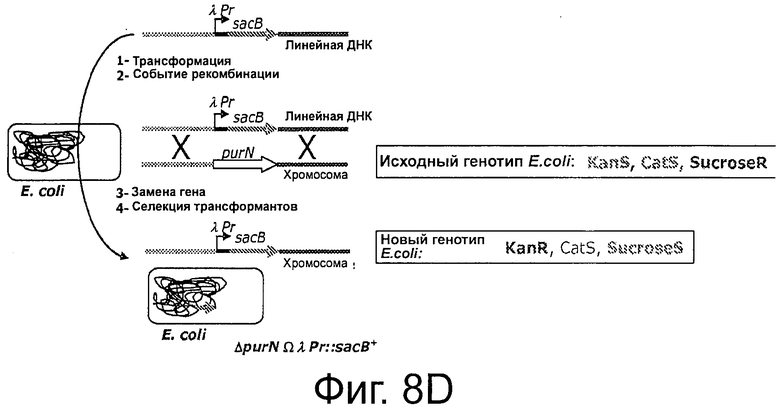
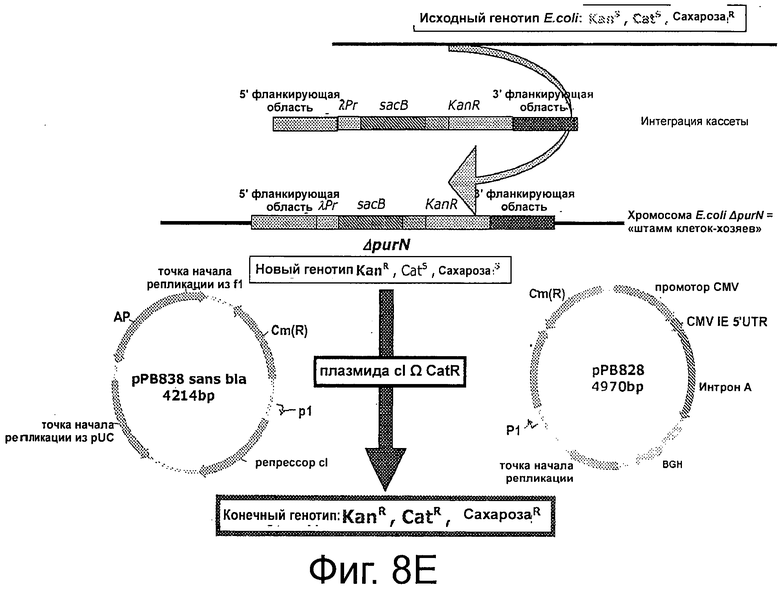
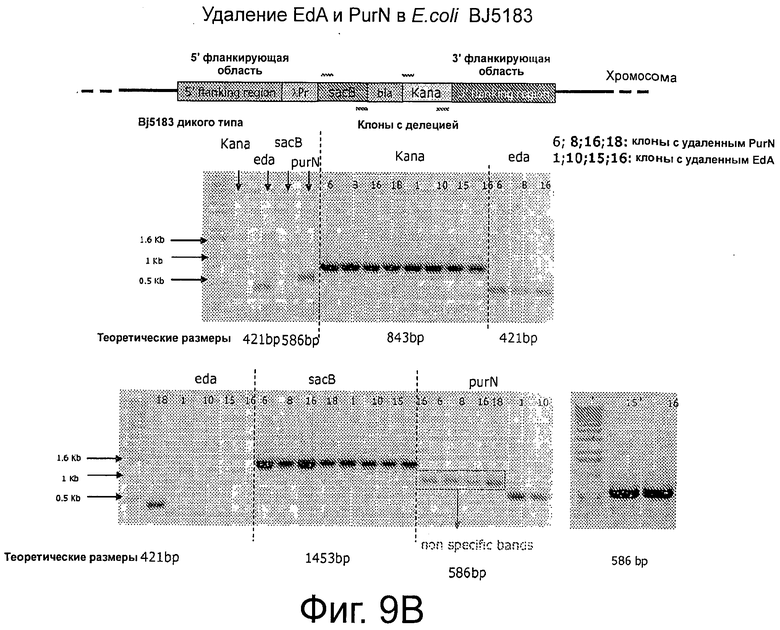

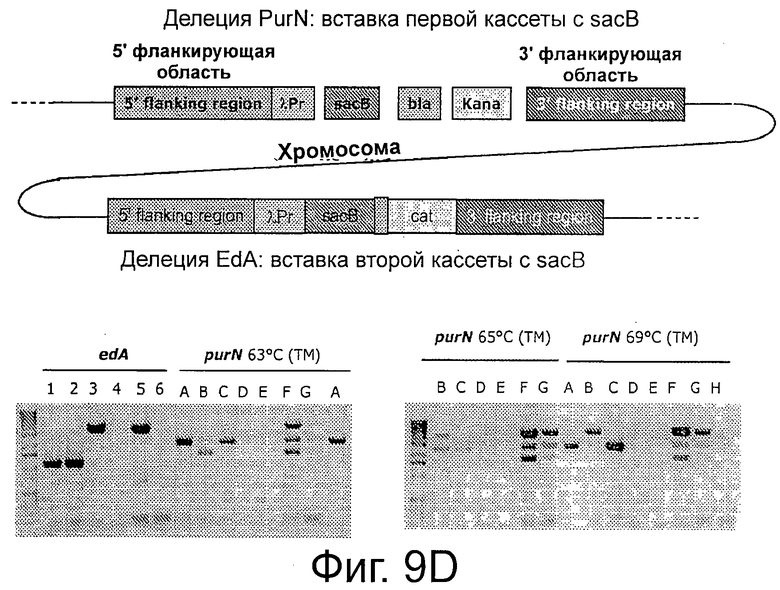
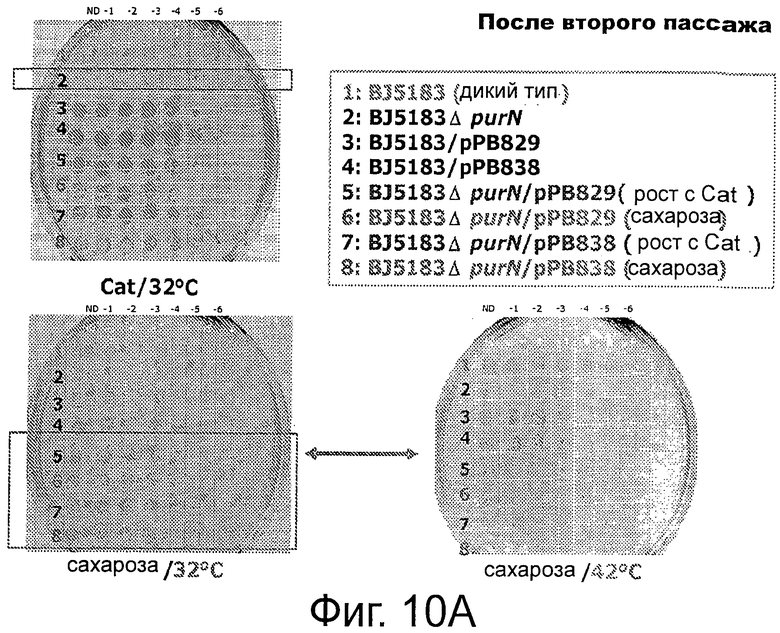
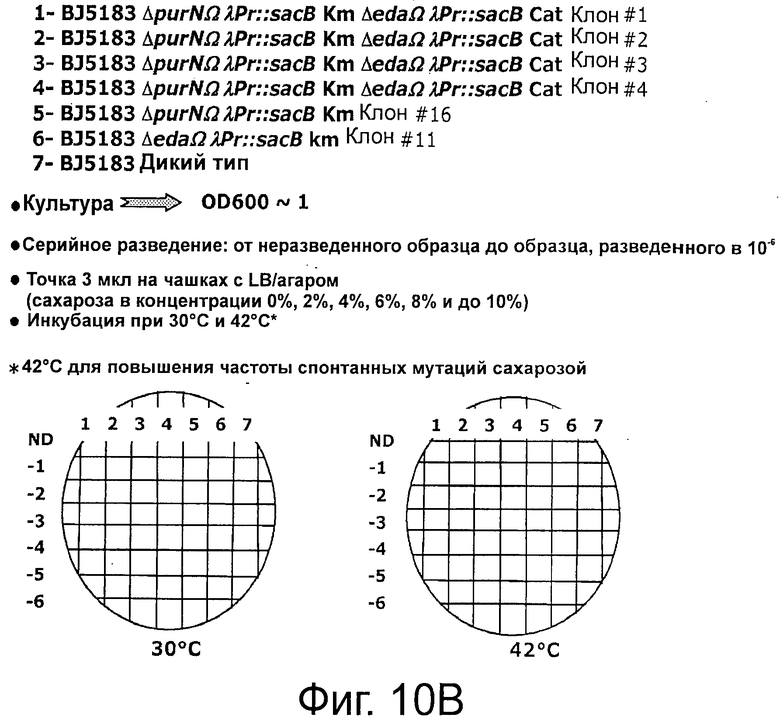
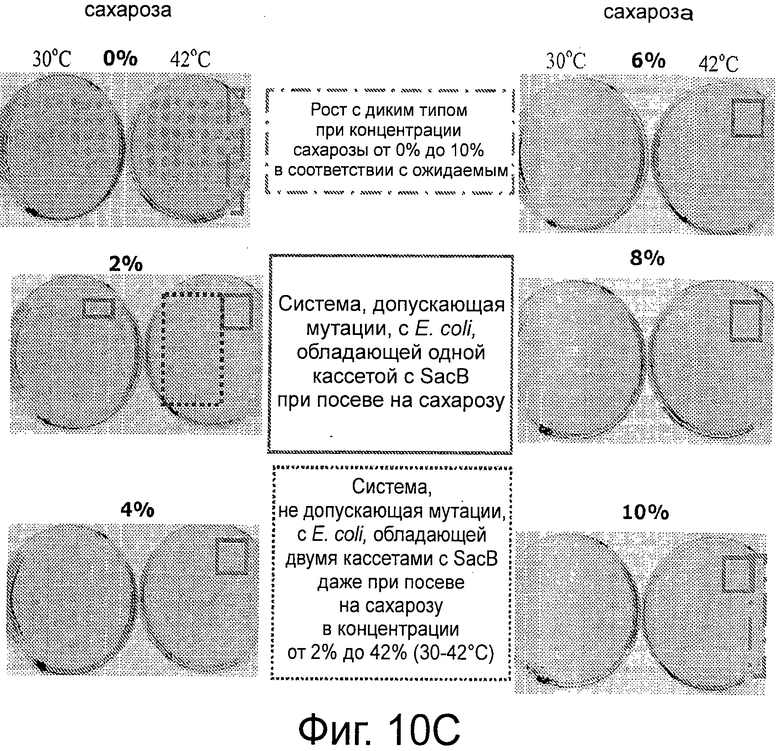
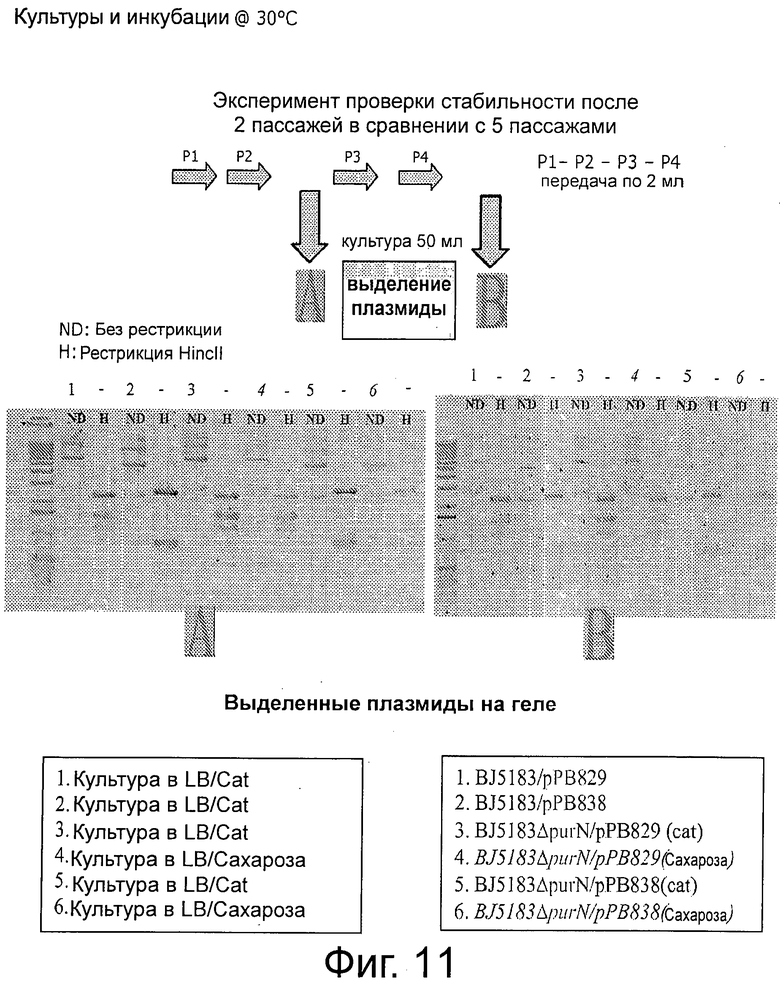
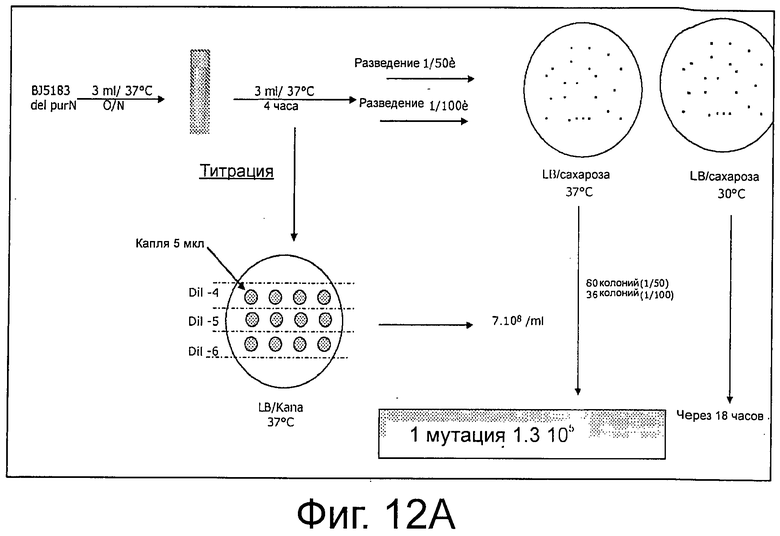
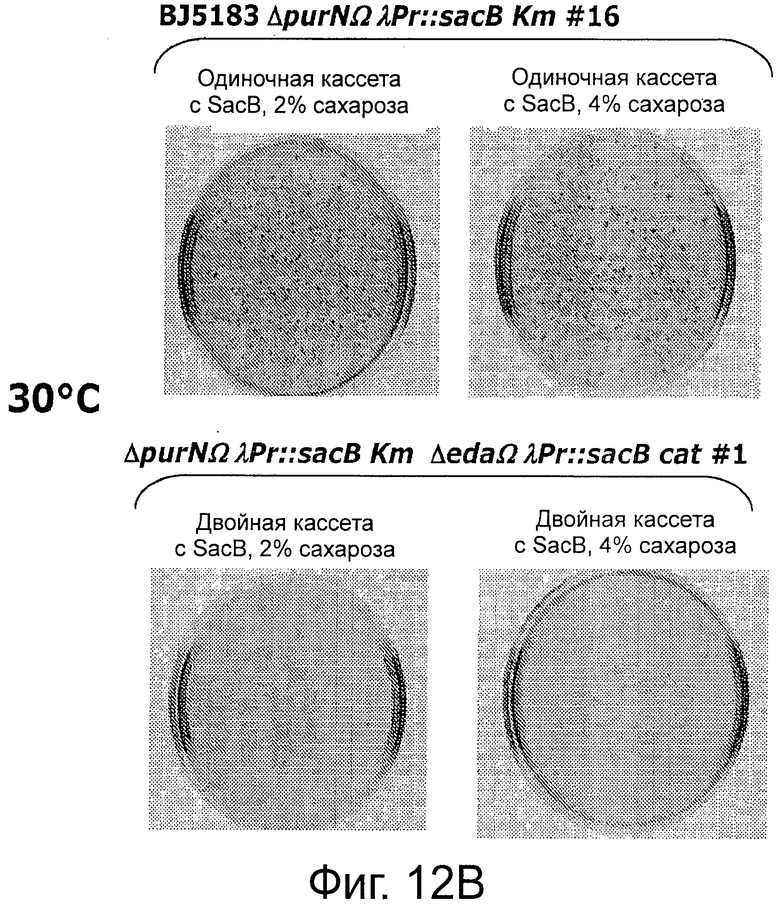
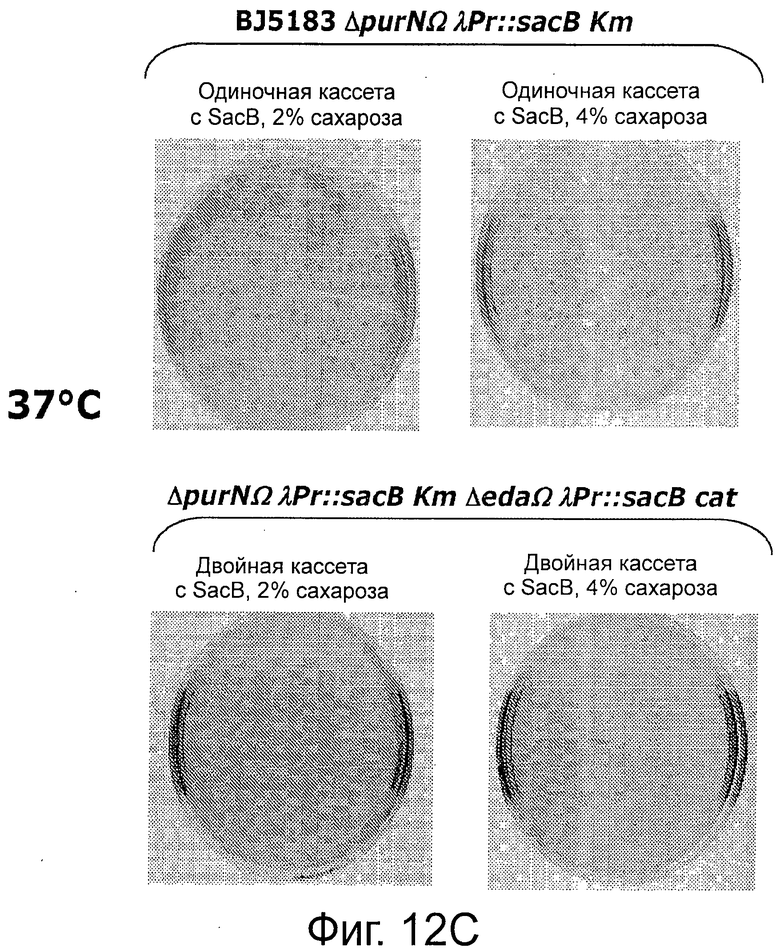
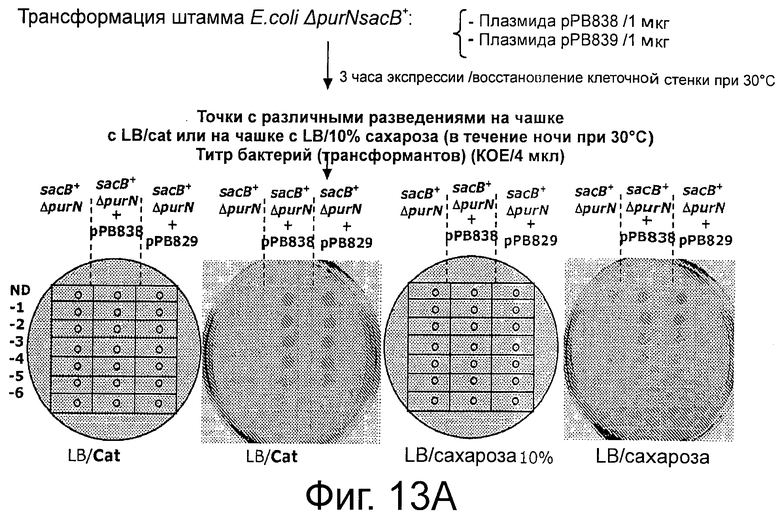

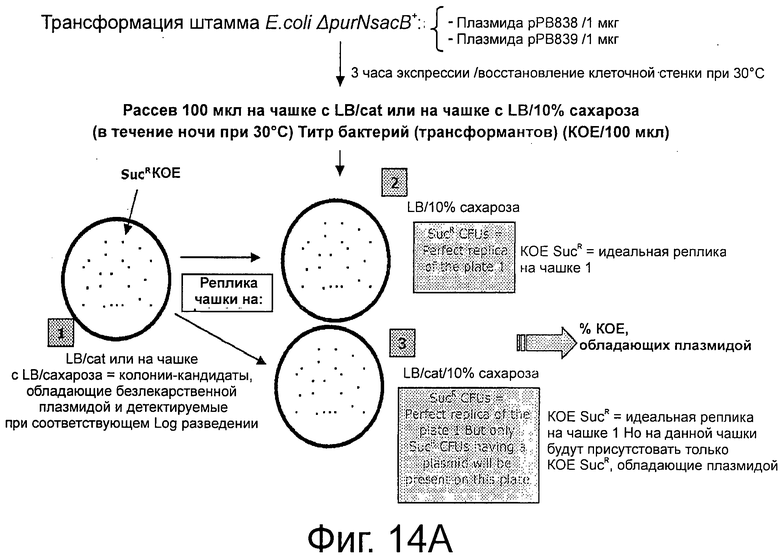
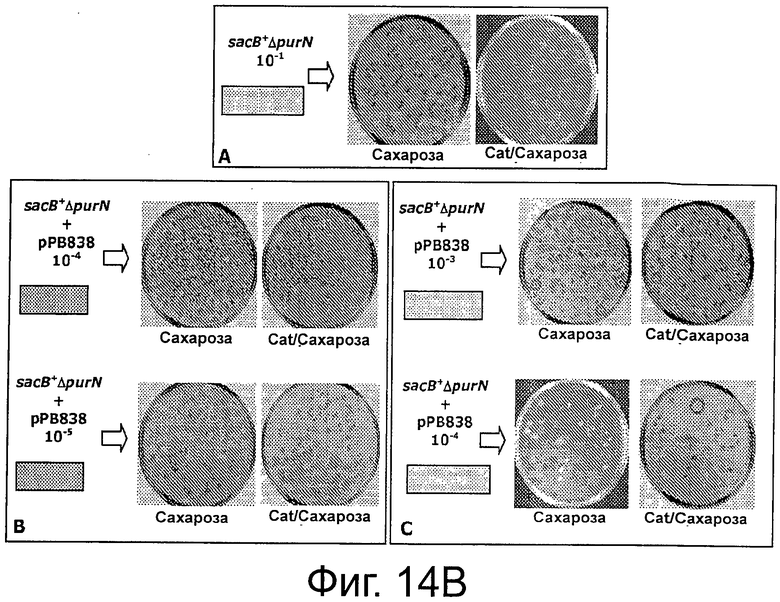
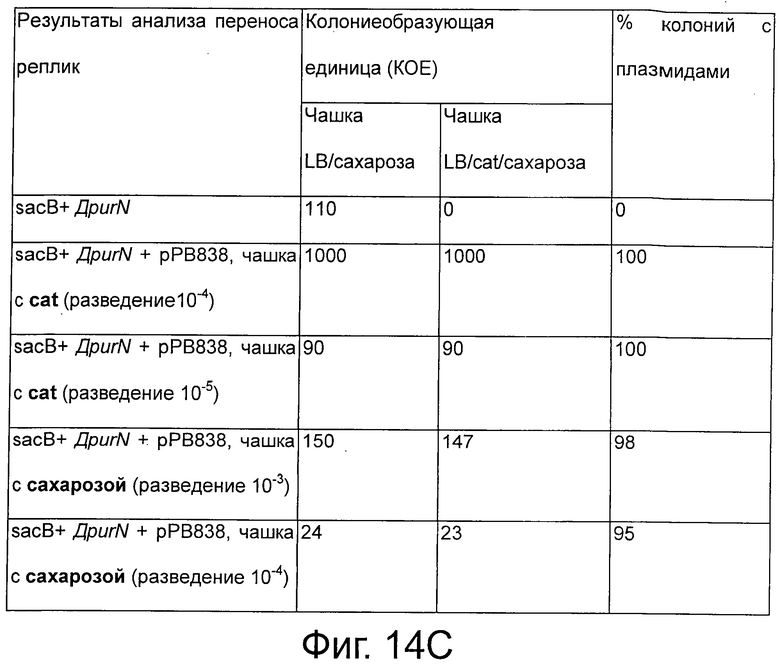
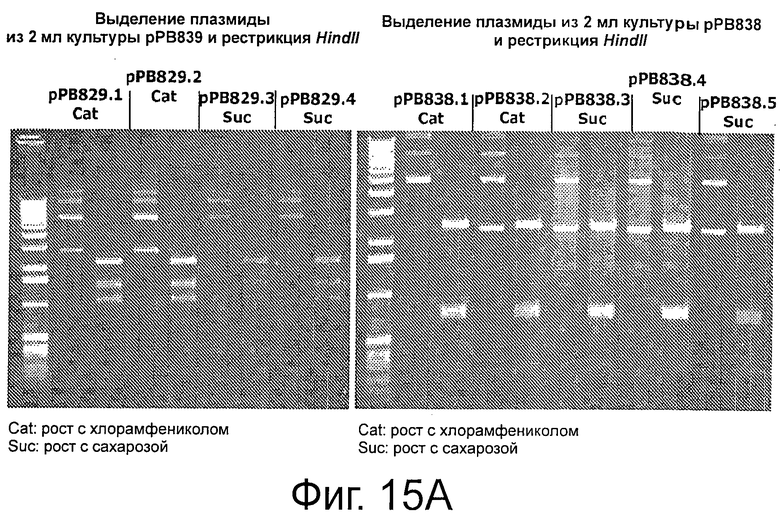
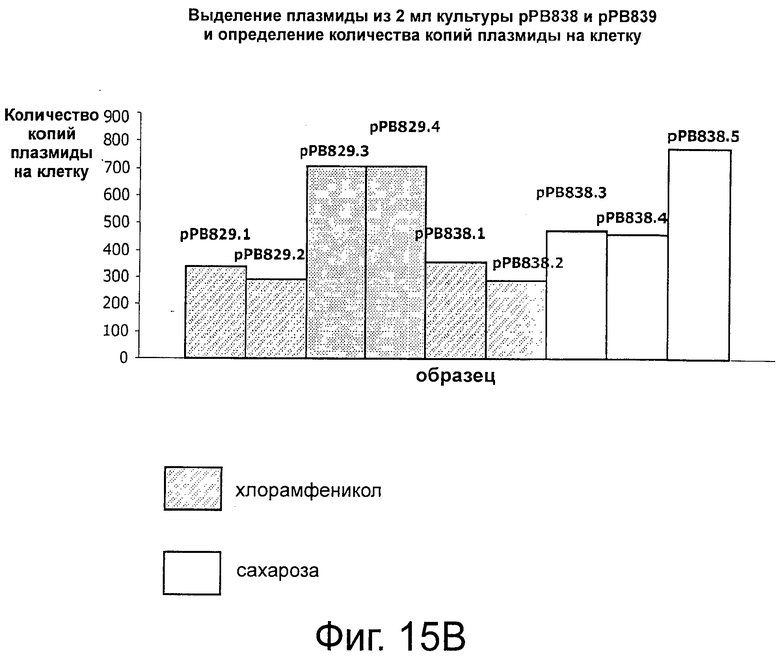
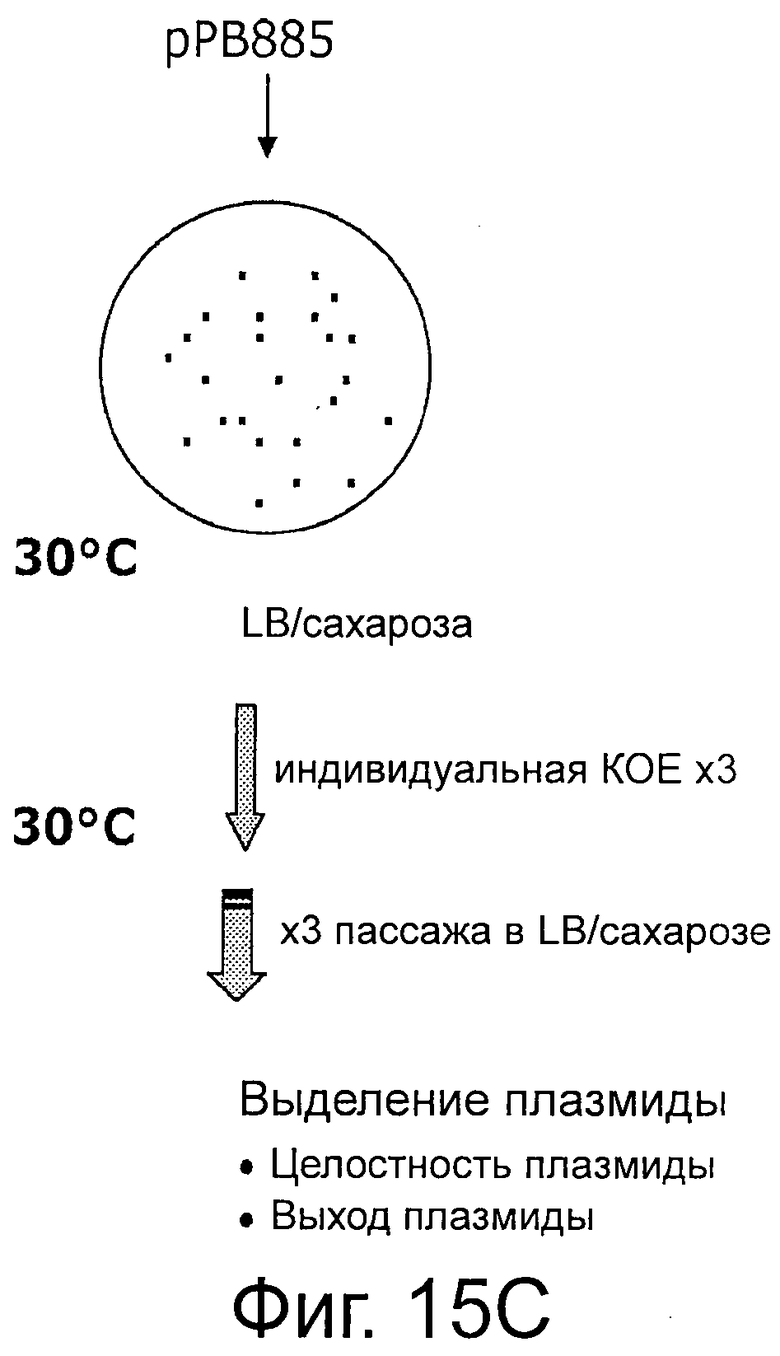
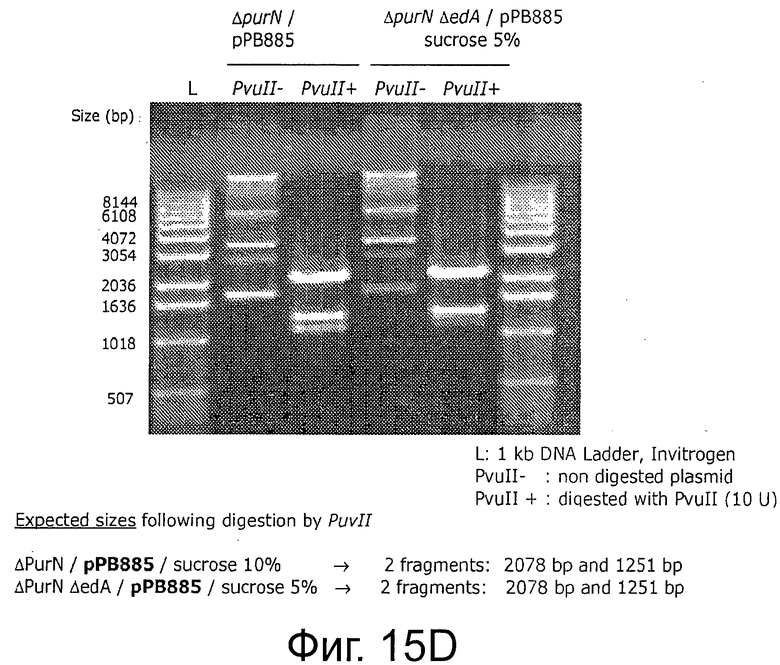
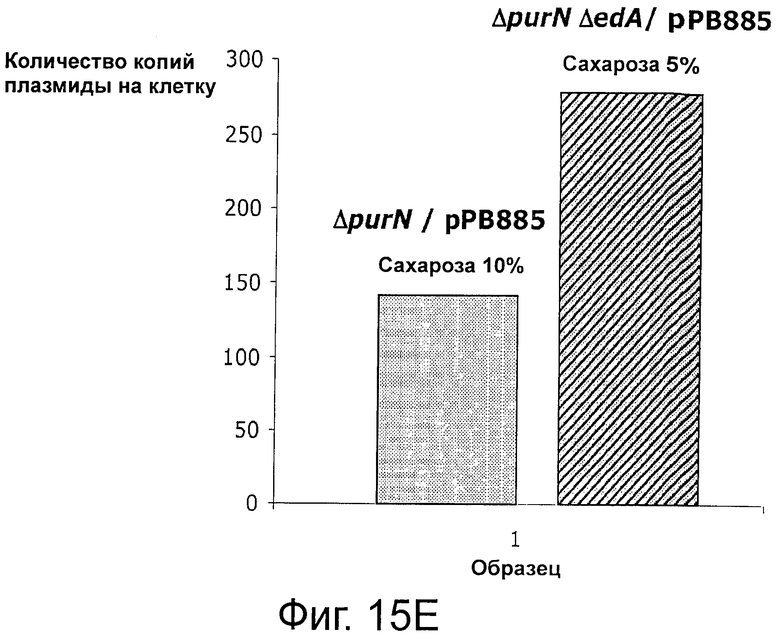

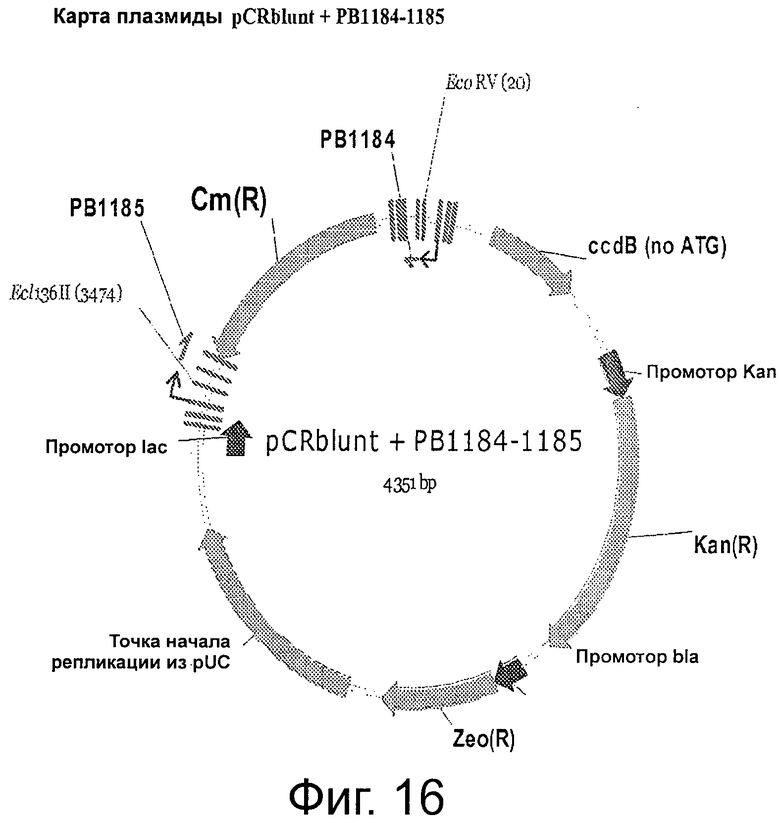

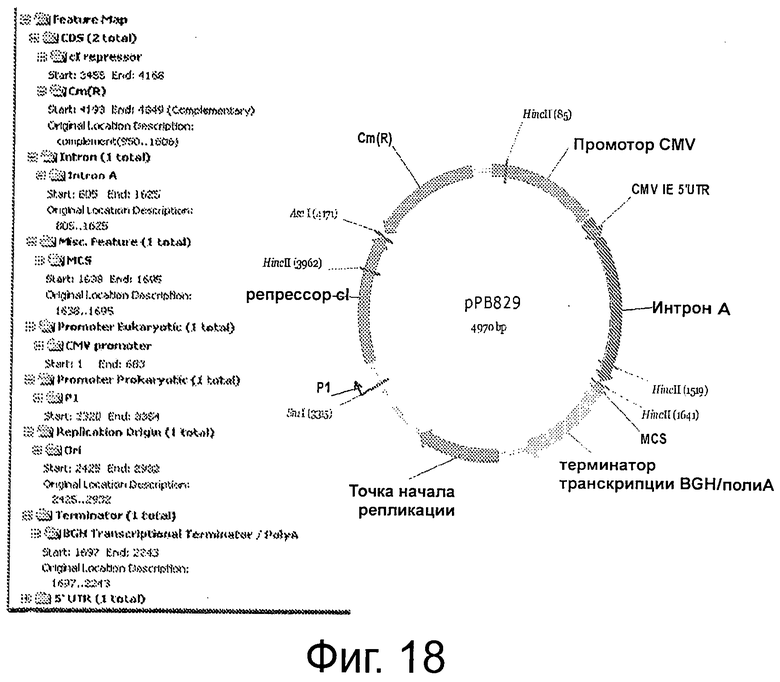
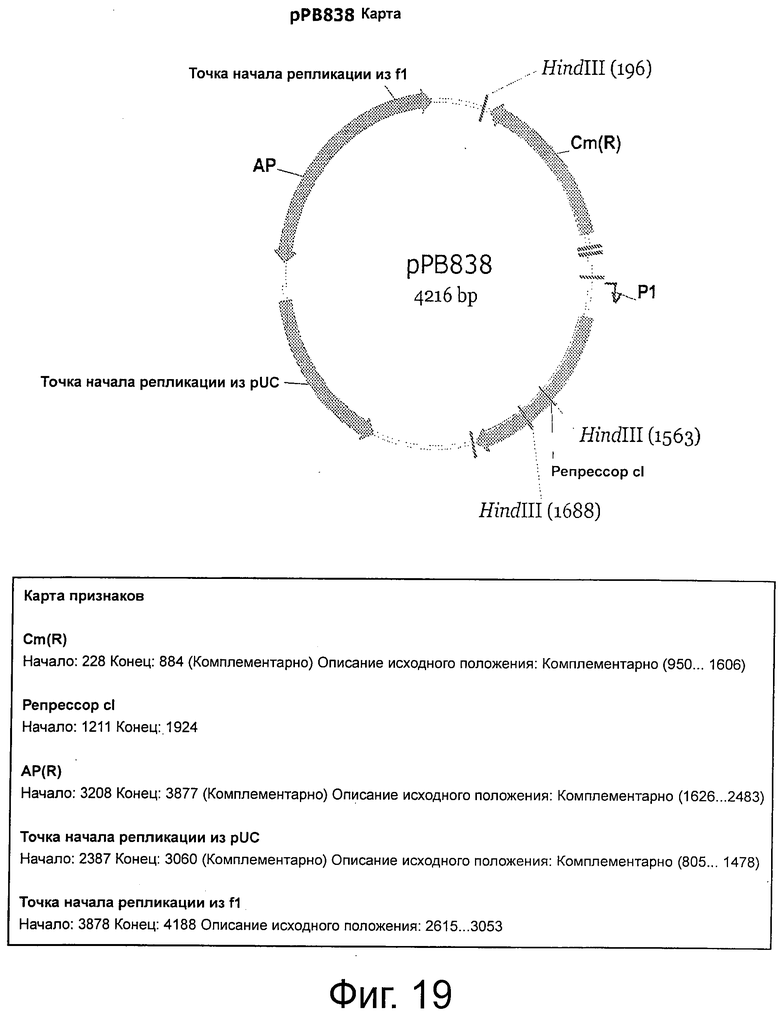
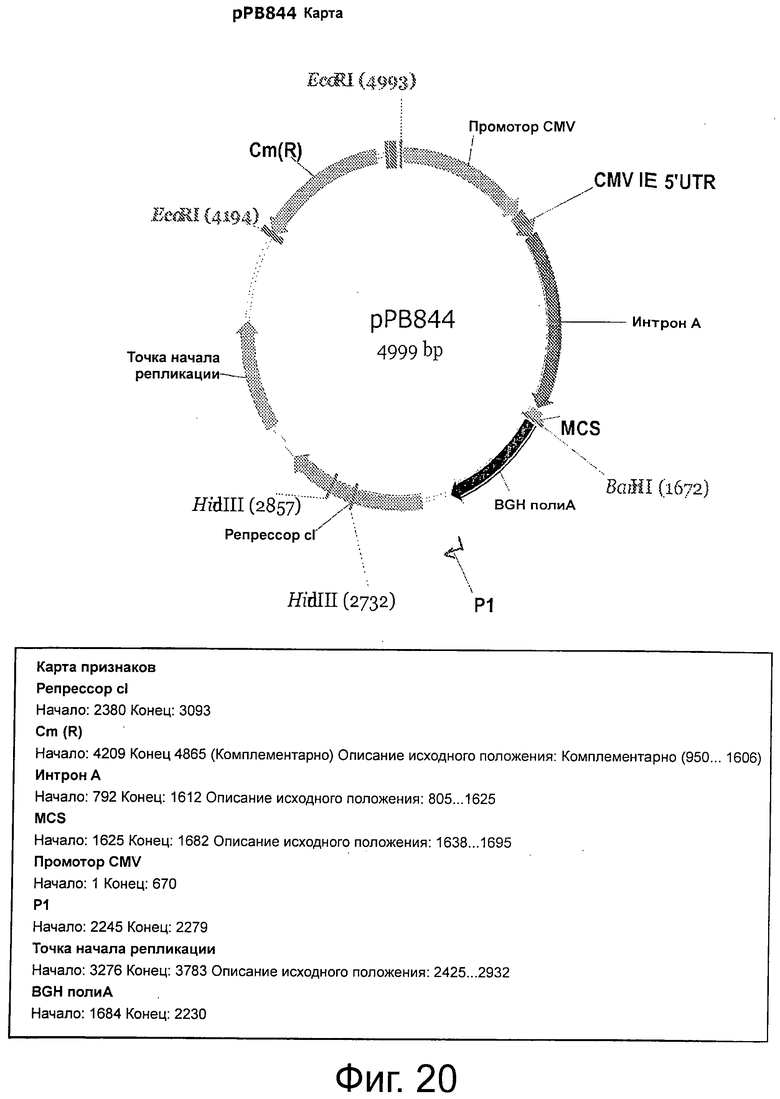
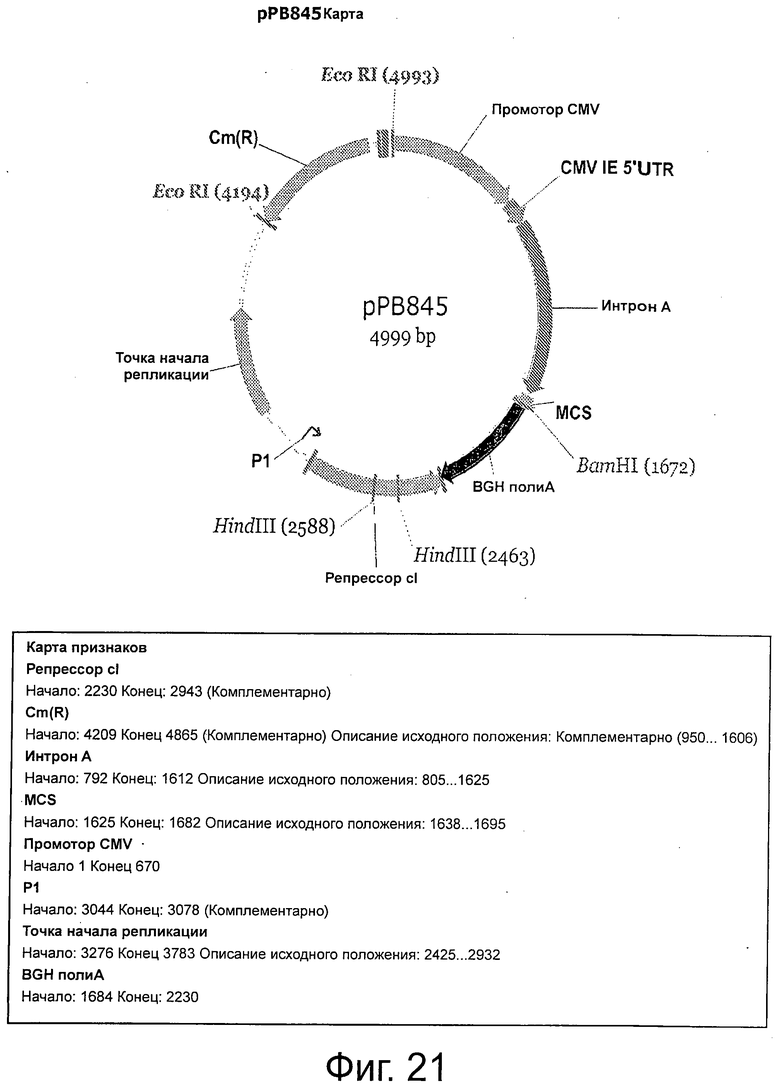

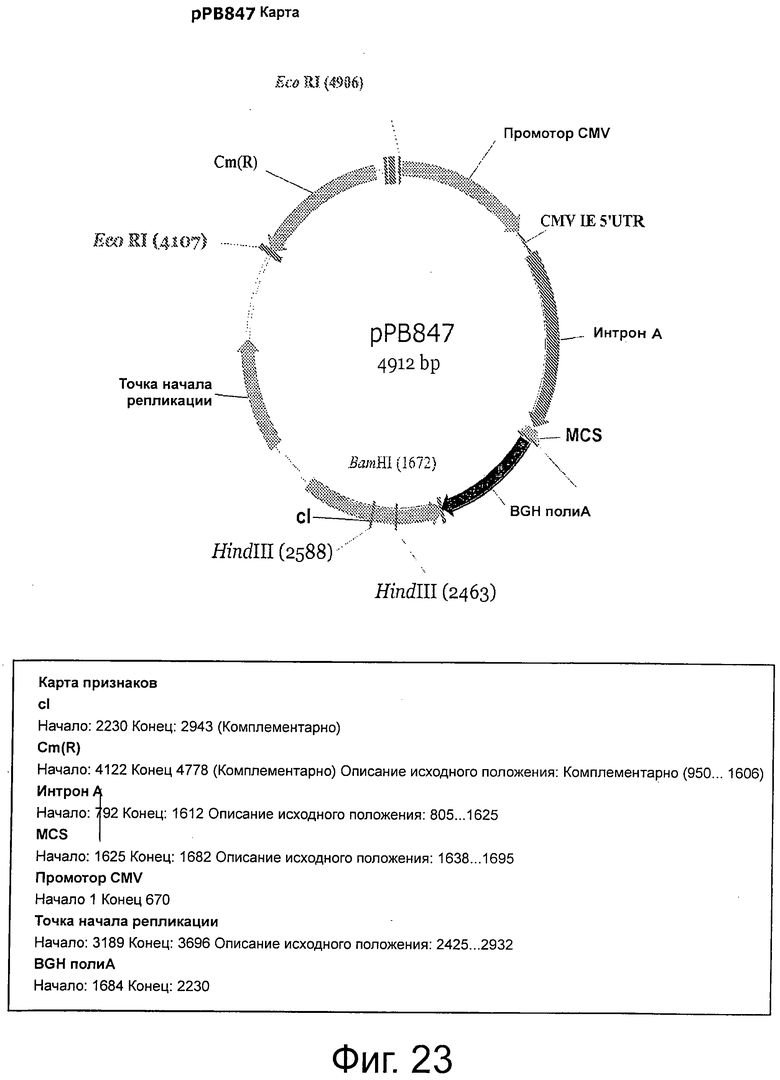
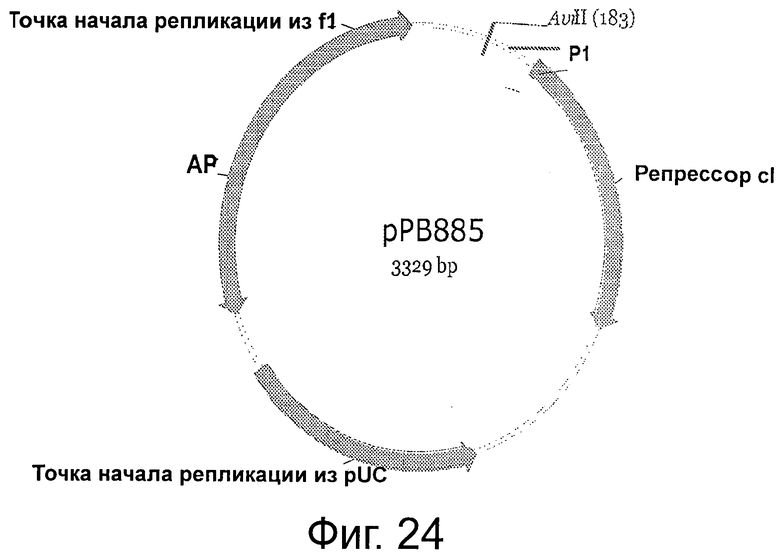
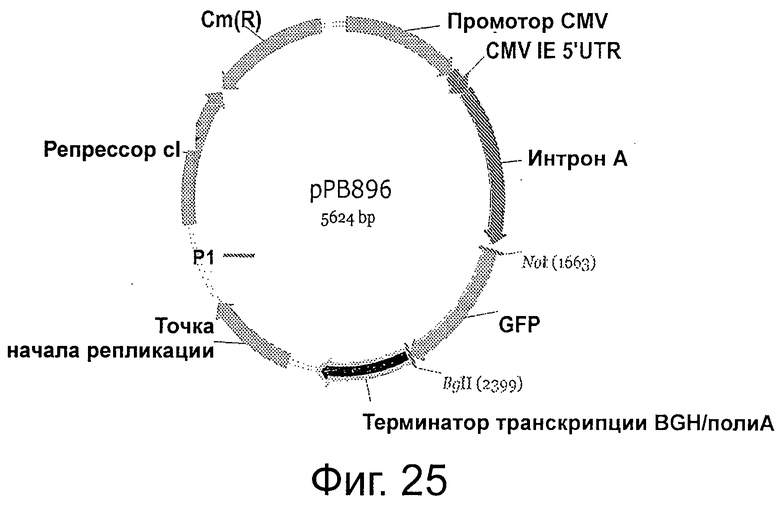

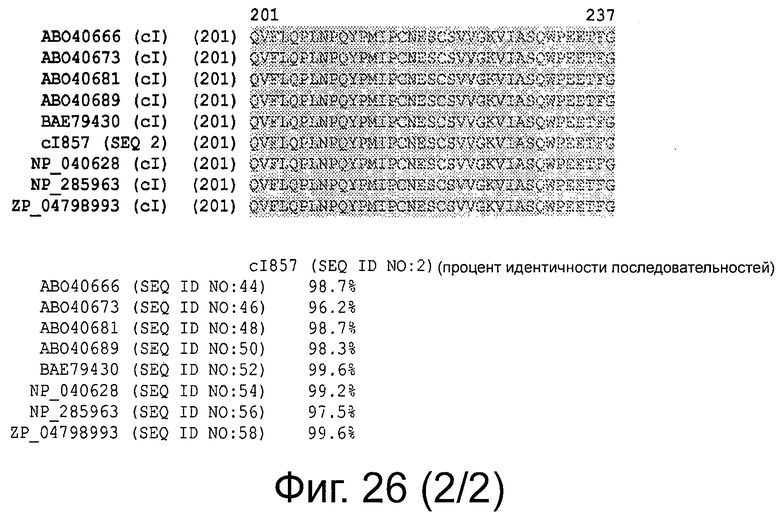
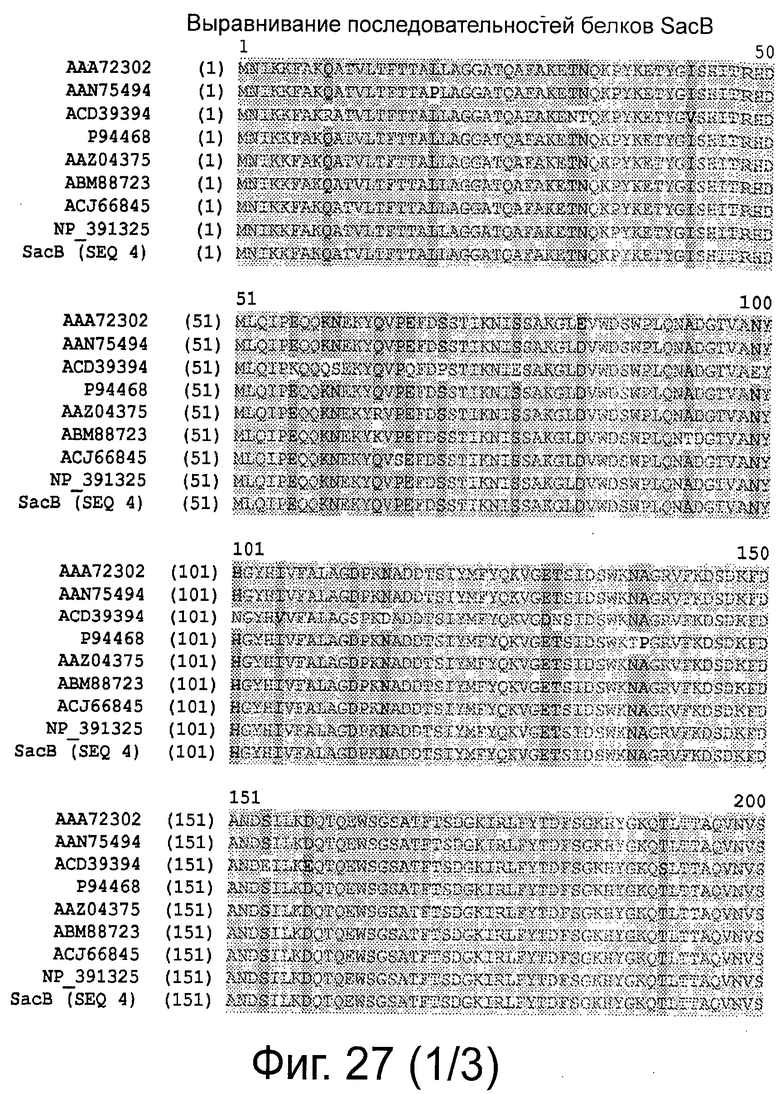



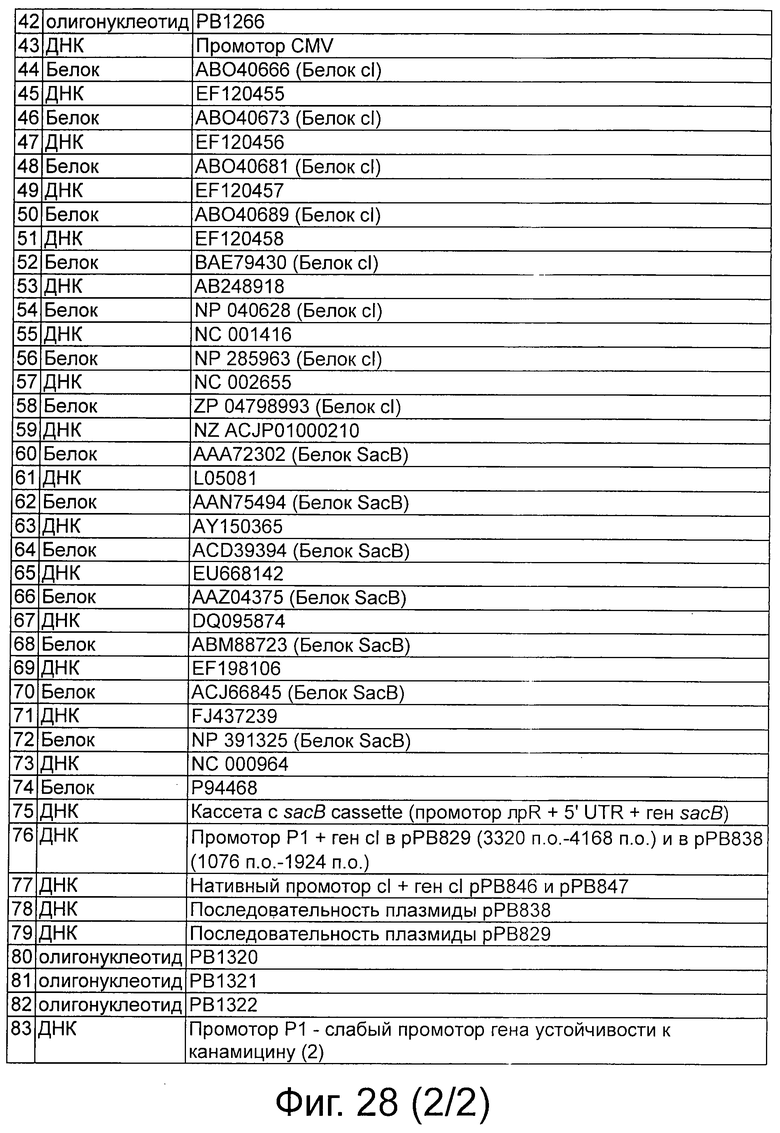





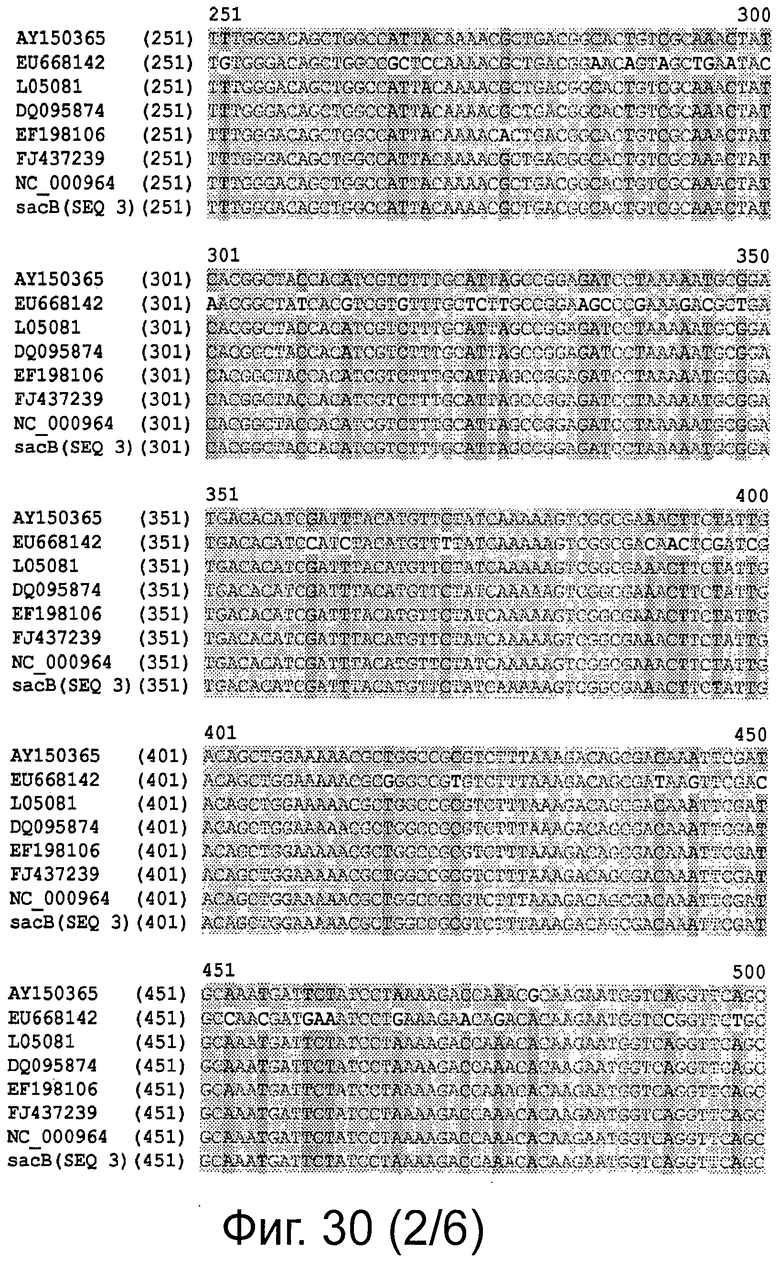



Настоящее изобретение относится к биотехнологии и представляет собой экспрессионную плазмиду без устойчивости к антибиотику, которая содержит полинуклеотид, кодирующий белок-репрессор cI. Экспрессия указанного белка-репрессора регулирует экспрессию токсичного генного продукта, встроенного в несущественный участок генома хозяина. Настоящее изобретение также раскрывает сконструированную клетку-хозяин, относящуюся к грамотрицательным бактериям, содержащую указанную плазмиду. Клетку-хозяин используют для получения плазмиды либо для получения некоего белка или иммуногена, если экспрессионная плазмида дополнительно содержит гены, кодирующие указанные белок или иммуноген. Способ получения экспрессионной плазмиды и способ получения белка или иммуногена проводят в несколько стадий. Сначала создают штамм клеток-хозяев, относящихся к грамотрицательным бактериям, встраиванием путем аллельной замены гена, кодирующего токсичный продукт, в несущественный участок хромосомы хозяина. Затем проводят конструирование ДНК-плазмиды, содержащей ген, кодирующий белок-репрессор cI, и, при необходимости, ген, кодирующий белок или иммуноген. После чего полученные клетки-хозяин трансформируют указанной плазмидой и выращивают в присутствии сахарозы при температуре 30-42 ºC. И, наконец, извлекают экспрессионную плазмиду, белок или иммуноген. Настоящее изобретение позволяет осуществлять контроль токсичного гена, локализованного на хромосоме, с помощью репрессора, локализованного на плазмиде при отсутствии селекционного давления антибиотиком. 4 н. и 29 з.п.ф-лы, 31 ил., 8 табл., 10 пр.
1. Экспрессионная плазмида без устойчивости к антибиотику, содержащая полинуклеотид, кодирующий белок-репрессор cI, который обладает, по меньшей мере, 96% идентичностью последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2, 44, 46, 48, 50, 52, 54, 56 или 58, где экспрессия белка-репрессора cI регулирует экспрессию токсичного генного продукта в хозяине.
2. Экспресионная плазмида без устойчивости к антибиотику по п. 1, где полинуклеотид функционально связан с промотором.
3. Экспресионная плазмида без устойчивости к антибиотику по п. 2, где промотор представляет собой промотор P1 или нативный промотор cI.
4. Экспресионная плазмида без устойчивости к антибиотику по п. 1, где полинуклеотид обладает, по меньшей мере, 90% идентичностью последовательности с полинуклеотидом, имеющим последовательность, представленную в SEQ ID NO: 1, 45, 47, 49, 51, 53, 55, 57 или 59.
5. Экспресионная плазмида без устойчивости к антибиотику по любому из пп. 1-4, где плазмида дополнительно содержит гетерологичный полинуклеотид, кодирующий иммуноген или белок.
6. Экспресионная плазмида без устойчивости к антибиотику по п. 5, где гетерологичный полинуклеотид функционально связан с промотором, функциональным в эукариотических или прокариотических клетках.
7. Экспресионная плазмида без устойчивости к антибиотику по п. 6, где промотор представляет собой промотор CMV или промотор, выделенный из грамотрицательной бактерии.
8. Экспресионная плазмида без устойчивости к антибиотику по п. 1, где полинуклеотид кодирует белок-репрессор cI, который обладает, по меньшей мере, 98% идентичностью последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2, 44, 46, 48, 50, 52, 54, 56 или 58.
9. Экспресионная плазмида без устойчивости к антибиотику по п. 1, где полинуклеотид обладает, по меньшей мере, 95% идентичностью последовательности с последовательностями, представленными в SEQ ID NO: 1, 45, 47, 49, 51, 53, 55, 57 или 59.
10. Экспресионная плазмида без устойчивости к антибиотику по п. 8 или 9, где полинуклеотид функционально связан с промотором.
11. Экспресионная плазмида без устойчивости к антибиотику по п. 10, где промотор представляет собой промотор P1 или нативный промотор cI.
12. Экспресионная плазмида без устойчивости к антибиотику по п. 1, где полинуклеотид кодирует белок-репрессор cI, имеющий последовательность, представленную в SEQ ID NO: 2, 44, 46, 48, 50, 52, 54, 56 или 58.
13. Экспресионная плазмида без устойчивости к антибиотику по п. 3, где промотор Р1 имеет последовательность, представленную в SEQ ID NO: 29 или SEQ ID NO: 83.
14. Экспресионная плазмида без устойчивости к антибиотику по п. 11, где промотор P1 имеет последовательность, представленную в SEQ ID NO: 29 или SEQ ID NO: 83.
15. Экспресионная плазмида без устойчивости к антибиотику по п. 3, где нативный промотор cI имеет последовательность, представленную в SEQ ID NO: 30.
16. Экспресионная плазмида без устойчивости к антибиотику по п. 11, где нативный промотор cI имеет последовательность, представленную в SEQ ID NO: 30.
17. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям, содержащая экспрессионную плазмиду без устойчивости к антибиотику, для поддержания и получения экспрессионной плазмиды без устойчивости к антибиотику и/или белка или антигена, где экспрессионная плазмида без устойчивости к антибиотику содержит полинуклеотид, кодирующий белок-репрессор cI, который обладает, по меньшей мере, 96% идентичностью последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2, 44, 46, 48, 50, 52, 54, 56 или 58, где экспрессия белка-репрессора cI регулирует экспрессию токсичного генного продукта, встроенного в несущественный участок генома хозяина.
18. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по п. 17, в которой полинуклеотид содержит ген sacB.
19. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по п. 18, в которой ген sacB кодирует белок SacB, обладающий по меньшей мере, 90% идентичностью последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 4, 60, 62, 64, 66, 68, 70, 72 или 74.
20. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по п. 17, в которой гетерологичный полинуклеотид функционально связан с промотором.
21. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по п. 20, в которой промотор является промотором из фага λ.
22. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по п. 17, в которой несущественный участок содержит ген deA или ген purN.
23. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по п. 18, в которой ген sacB кодирует белок SacB, обладающий, по меньшей мере, 90% идентичностью последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 4, и где ген sacB функционально связан с промотором, имеющим последовательность, представленную в SEQ ID NO: 5.
24. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по п. 18, где клетка содержит одну или несколько копий полинуклеотида, имеющего последовательность, представленную в SEQ ID NO: 75.
25. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям п. 24, где клетка содержит две копии полинуклеотида последовательности SEQ ID NO: 75, вставленного в ген deA и в ген purN хромосомы хозяина.
26. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по любому из пп.17-25, где клетка выбрана из группы, состоящей из Avibacterium, Brucella, Escherichia coli, Haemophilus (например, Haemophilus suis), Salmonella (например, Salmonella enteridis, Salmonella typhimurium, Salmonella infantis), Shigella, Pasteurella и Rimeirella.
27. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по п. 17, где экспрессионная плазмида без устойчивости к антибиотику содержит полинуклеотид, кодирующий белок-репрессор cI, который обладает, по меньшей мере, 98% идентичностью последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2, 44, 46, 48, 50, 52, 54, 56 или 58.
28. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по п. 17, где экспрессионная плазмида без устойчивости к антибиотику содержит полинуклеотид, кодирующий белок-репрессор cI, имеющий последовательность, представленную в SEQ ID NO:2, 44, 46, 48, 50, 52, 54, 56 или 58.
29. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по любому из пп. 17 или 27-28, в которой полинуклеотид функционально связан с промотором P1, имеющим последовательность, представленную в SEQ ID NO: 29 или SEQ ID NO: 83.
30. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по любому из пп. 17 или 27-28, в которой полинуклеотид функционально связан с нативным промотором cI, имеющим последовательность, представленную в SEQ ID NO: 30.
31. Сконструированная клетка-хозяин, относящаяся к грамотрицательным бактериям по любому из пп. 27 и 28, где клетка грамотрицательной бактерии выбрана из группы, состоящей из Avibacterium, Brucella, Escherichia coli, Haemophilus (например, Haemophilus suis), Salmonella (например, Salmonella enteridis, Salmonella typhimurium, Salmonella infantis), Shigella, Pasteurella и Rimeirella.
32. Способ получения экспресионной плазмиды без устойчивости к антибиотику по п.1, включающий стадии:
1) создания штамма клеток-хозяев, относящихся к грамотрицательным бактериям, содержащих гетерологичный полинуклеотид, содержащий ген, который кодирует токсичный продукт, под контролем промотора с сильной регуляцией, в частности промотора из фага λ, вставленный путем аллельной замены в один или несколько несущественных участков хромосомы хозяина;
2) конструирования ДНК-плазмиды, содержащей полинуклеотид, кодирующей белок-репрессор cI, который обладает, по меньшей мере, 96% идентичностью последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2, 44, 46, 48, 50, 52, 54, 56 или 58, где экспрессия белка-репрессора cI регулирует экспрессию токсичного генного продукта в хозяине;
3) трансформации штамма бактериальных клеток-хозяев ДНК-плазмидой, содержащей ген, кодирующей белок-репрессор cI;
4) роста трансформированного штамма бактериальных клеток-хозяев в присутствии сахарозы при температуре в интервале от 30°C до 42°C; и
5) извлечения ДНК-плазмиды.
33. Способ получения белка или иммуногена, включающий стадии:
1) создания штамма клеток-хозяев, относящихся к грамотрицательным бактериям, которые содержат гетерологичный полинуклеотид, содержащий ген, который кодирует токсичный продукт, под контролем промотора с сильной регуляцией, в частности промотора из фага λ, вставленный путем аллельной замены в один или несколько несущественных участков хромосомы организма-хозяина;
2) конструирования ДНК-плазмиды, содержащей полинуклеотид, кодирующей белок-репрессор cI, который обладает, по меньшей мере, 96% идентичностью последовательности с полипептидом, имеющим последовательность, представленную в SEQ ID NO: 2, 44, 46, 48, 50, 52, 54, 56 или 58, где экспрессия белка-репрессора cI регулирует экспрессию токсичного генного продукта в хозяине, и ген, кодирующий иммуноген или белок;
3) трансформации штамма бактериальных клеток-хозяев ДНК-плазмидой, содержащей полинуклеотид, кодирующий белок-репрессор cI, и ген, кодирующий иммуноген или белок;
4) роста трансформированного штамма бактериальных клеток-хозяев в присутствии сахарозы при температуре в интервале от 30°C до 42°C; и
5) извлечения иммуногена или белка.
| US 5147789 A, 15.09.1992 | |||
| GILADI H., et al., Selective stabilization by bacteriophage 434 repressor of the plasmid expressing bovine growth hormone in Escherichia coli, Applied and environmental microbiology, May 1988, pp.1297-1299 | |||
| WO 2006029985 A2, 23.03.2006 | |||
| Устройство для автоматического регулирования режима формовки полупроводниковых, в частности, селеновых элементов | 1957 |
|
SU109351A1 |
| VLADIMIR PELICIC, Expression of the Bacillus subtilis sacB | |||

