Группа изобретений относится к биотехнологии, генной и белковой инженерии и предназначена для получения генетически слитых гибридных белков, состоящих из одноцепочечного антитела мыши против белка Е вируса клещевого энцефалита и фрагментов люциферазы NanoLuc, при связывании которых с белками Е вируса клещевого энцефалита происходит их комплементация и возникновение биолюминесценции в присутствии субстрата - целентеразина.
Вирус клещевого энцефалита (ВКЭ), представитель семейства Flaviviridae - это высокопатогенный инфекционный агент, вызывающий серьезные поражения нервной системы. Современная эпидемическая ситуация в отношении КЭ характеризуется значительным ростом заболеваемости как Российской Федерации, так и в мире [1, 2]. Экстренная профилактика клещевого энцефалита у невакцинированных укушенных пациентов включает введение препарата «Иммуноглобулин человека против клещевого энцефалита», полученного из сыворотки донорской крови. Применение продуктов донорской крови всегда сопровождается биологическим риском. Кроме того, не исключены случаи дефицита препарата «Иммуноглобулин человека против клещевого энцефалита» в период эпидемиологического сезона. Показано, что в среднем, лишь около 5% клещей являются носителями ВКЭ. Поэтому быстрое, высокочувствительное выявление этого вируса необходимо для экспресс-диагностики для того, чтобы избежать необоснованного применения препарата «Иммуноглобулин человека против клещевого энцефалита» для экстренной профилактики и, как следствие, снижения количества случаев возможных осложнений.
Искусственная люцифераза NanoLuc представляет собой небольшой (19 кДа), одноцепочечный полипептид. Фермент катализирует окисление субстрата - целентеразина (или его синтетических аналогов), молекулярным кислородом с образованием СО2 и молекулы целентерамида в возбужденном состоянии. Переход этого соединения в основное состояние сопровождается выделением света в видимой области спектра (λmax=455 нм). Ген, кодирующий этот белок, был получен в результате множественного мутагенеза гена, кодирующего малую субъединицу люциферазы глубоководной креветки Oplophurus gracilirostris, и получен рекомбинантный белок, обладающий высокой биолюминесцентной активностью [3], а также фрагменты этого белка, обладающие способностью комплементации с восстановлением биолюминесцентной активности [4]. При пассивном смешивании растворов этих фрагментов восстановление биолюмиесцентной активности не происходит.Эффект восстановления биолюминесцентной активности возникает в результате сближения и комплементации этих фрагментов при связывании добавленных полипептидных доменов с молекулой-мишенью.
В литературе описаны гибридные белки на основе этих фрагментов NanoLuc люциферазы, слитых с другими полипептидами, например, с антителами или их антигенсвязывающими фрагментами [5, 6], антигенами [7], взаимодействующими полипептидами [8, 9] и пр. [см., напр., обзор 10]
Наиболее близким аналогом - прототипом к заявляемой группе изобретений - рекомбинантных плазмидных ДНК pFLAG-sc14D5a-NLuc-r1 и pFLAG-NLuc-r2-sc14D5a, обеспечивающие синтез гибридных белков sc14D5a-NLuc-r1 и NLuc-r2-sc 14D5a, является рекомбинантная плазмидная ДНК pFLAG-sc14D5a-Rm7, обеспечивающая синтез гибридного белка sc14d5a- Rm7, и гибридный белок sc14D5a-Rm7, связывающий белок Е вируса клещевого энцефалита и обладающий биолюминесцентной активностью при добавлении субстрата - целентеразина [11, 12]. Недостатком прототипа является то, что гибридный белок sc14D5a-Rm7 пригоден для выявления вируса клещевого энцефалита твердофазным биолюминесцентным иммуноанализом сэндвич типа, который включает ряд стадий промывки и инкубирования образца и может проводиться только в лабораторных условиях. Предлагаемые в заявке белки sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a излучают свет только при наличии в образце нескольких близко расположенных ВКЭ-ассоциированных антигенов (фрагментов белка Е), с которыми они связываются за счет аффинности домена sc14D5a, вызывая комплементацию фрагментов люциферазы и возникновение целентеразин-зависимой биолюминесценции. Такой анализ проводится однофазно, не требует ряда стадий промывки и инкубирования образца и может быть проведен во внелабораторных условиях.
Техническим результатом группы изобретений является получение двух гибридных белков, обладающих способностью связывать белки Е вируса клещевого энцефалита с комплементацией фрагментов люциферазы NanoLuc и возникновением биолюминесцентной активности в присутствии субстрата - целентеразина, как потенциально пригодной высокочувствительной системы для выявления вируса клещевого энцефалита в биологических образцах однофазным биолюминесцентным анализом.
Указанный технический результат достигается путем конструирования рекомбинантных плазмидных ДНК pFLAG-sc14D5a-NLuc-r1 и pFLAG-NLuc-r2-sc14D5a, содержащих уникальный ген одноцепочечного антитела мыши, направленного против белка Е вируса клещевого энцефалита, и гены фрагментов люциферазы NLuc-r1 и NLuc-r2; экспрессией целевых гибридных белков в трансформированных упомянутыми плазмидными ДНК клетках Escherichia coli Rosetta-gami2 с последующими выделением из цитоплазматической фракции и очисткой металл-хелатной хроматографией гибридных белков sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a, которые обладают способностью связывать белок Е вируса клещевого энцефалита, сопровождающейся комплементацией фрагментов люциферазы с возникновением целентеразин-зависимой биолюминесцентной активности.
Сущность группы изобретений заключается в следующем:
Генно-инженерными методами получают плазмидные ДНК pFLAG-sc14D5a-NLuc-r1 и pFLAG-NLuc-r2-sc14D5a, несущих фрагмент ДНК, кодирующий одноцепочечное антитело мыши, направленное против белка Е вируса клещевого энцефалита, коннекторный пептид (Gly2Ser)5 либо (Gly2Ser)4, в одной рамке считывания; гибридные белки sc14D5a-NLuc-r1 либо NLuc-r2-sc14D5a, состоящие из одноцепочечного антитела мыши, направленного против белка Е вируса клещевого энцефалита, коннекторного пептида (Gly2Ser)5, либо (Gly2Ser)4 и фрагменты люциферазы NLuc-r1 либо NLuc-r2, не обладающие целентеразин-зависимой биолюминесцентной активностью, полученные в результате бактериального синтеза в Escherichia coli Rosetta-gami2, трансформированных сконструированными плазмидами; указанные белки способны восстанавливать целентеразин-зависимую биолюминесцентную активность в результате комплементации при сближении, которое обеспечивает миниантитело sc14D5a при связывании с близко расположенными белками Е.
Исходным генетическим материалом для конструирования рекомбинантной плазмиды pFLAG-sc14D5a-NLuc-r1, являются:
а) плазмидный вектор pFLAG-CTS (Sigma-Aldrich, США), обеспечивающий встраивание фрагментов ДНК, из модифицированного N-концевым пептидом НННННН одноцепочечного антитела мыши sc14D5a, направленного против белка Ε вируса клещевого энцефалита, коннекторного пептида (Gly2Ser)5 и большого фрагмента люциферазы NanoLuc (NLuc-r1), и его экспрессию под контролем tac-промотора;
б) ДНК плазмиды pFLAG- sc14D5a-NLuc кодирующей одноцепочечное антитело мыши и последовательность люциферазы NanoLuc.
Полученная в результате плазмида pFLAG-sc14D5a-NLuc-r1, характеризуется следующими признаками:
- имеет молекулярную массу 4.4 МДа и размер 6609 п.о.;
- кодирует гибридный белок sc14D5a-NLuc-r1, состоящий из модифицированного N-концевым пептидом НННННН одноцепочечного антитела мыши sc14D5a, направленного против белка Ε вируса клещевого энцефалита, коннекторного пептида (Gly2Ser)5 и большого фрагмента люциферазы NanoLuc (NLuc-r1);
- состоит из следующих элементов:
а) фрагмент ДНК NdeI-NotI размером 730 п.о., представляющий собой последовательность, кодирующую тяжелую и легкую цепи вариабельного домена моноклонального антитела sc14D5a, объединенные линкером (Gly4Ser)3 содержащий N-концевой пептид НННННН;
б) фрагмент ДНК размером 524 п.о., представляющий собой последовательность, кодирующую коннекторный пептид (Gly2Ser)5 и NLuc-r1 (большой фрагмент люциферазы NanoLuc);
- содержит генетические маркеры: amp - ген ампициллин-резистентности (ген β-лактамазы), определяющий устойчивость к ампициллину при трансформации Escherichia coli.
Исходным генетическим материалом для конструирования рекомбинантной плазмиды pFLAG-NLuc-r2-sc14D5a, является:
а) плазмидный вектор pFLAG-CTS (Sigma-Aldrich, США), обеспечивающий встраивание фрагментов ДНК, из малого фрагмента (NLuc-r2) люциферазы NanoLuc, соединенной коннекторным пептидом (Gly2Ser)4 с одноцепочечным антителом мыши, направленным против белка Ε вируса клещевого энцефалита содержащим С-концевой пептид НННННН, и его экспрессию под контролем tac-промотора;
б) ДНК плазмиды pFLAG-NLuc-r1-sc14D5a кодирующая последовательность гибридного белка, состоящего из большого фрагмента люциферазы NanoLuc (NLuc-r1) и одноцепочечного антитела мыши, направленного против белка Ε вируса клещевого энцефалита, содержащего С-концевой пептид НННННН.
Полученная в результате плазмида pFLAG-NLuc-r2-sc14D5a характеризуется следующими признаками:
- имеет молекулярную массу 4 МДа и размер 6159 п.о.;
- кодирует гибридный белок NLuc-r2-sc14D5a состоящий из малого фрагмента (NLuc-r2) люциферазы NanoLuc, коннекторного пептида (Gly2Ser)4 и одноцепочечного антитела мыши sc14D5a, направленного против белка Ε вируса клещевого энцефалита, содержащего С-концевой пептид НННННН
состоит из фрагмента ДНК NdeI-EcoRI размером 822 п.о., кодирующего малый фрагмент (NLuc-r2) люциферазы NanoLuc, коннекторный пептид (Gly2Ser)4 и одноцепочечное антитело мыши sc14D5a, направленного против белка Ε вируса клещевого энцефалита, содержащего С-концевой пептид НННННН;
- содержит генетические маркеры: amp - ген ампициллин-резистентности (ген β-лактамазы), определяющий устойчивость к ампициллину при трансформации Escherichia coli.
Для получения штаммов-продуцентов гибридных белков sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a компетентные клетки бактерий Escherichia coli Rosetta-gami2 (Δ(ara-leu)7697 ΔlacX74 ΔphoA PvuII phoR araD139 ahpC galE galK rpsL F'[lac+] lacIq pro] gor522:Tn10 trxB pRARE23, (CamR, StrR, TetR) (Novagen, США), трансформируют сконструированными плазмидами pFLAG-sc14D5a-NLuc-r1 и pFLAG-NLuc-r2-sc14D5a.
Рекомбинантные клетки Escherichia coli Rosetta gami2/pFLAG-sc14D5a-NLuc-r1 и Rosetta gami2/pFLAG-NLuc-r2-sc14D5a обеспечивают индуцируемый изопропилтиогалактозидом (ИПТГ) синтез гибридных белков sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a. Клетки данных штаммов содержат мутированные гены тиоредоксин редуктазы и глутатион редуктазы в результате чего существенно увеличивается образование дисульфидных связей в белках при экспрессии в цитоплазме. Индикацию экспрессии осуществляют с помощью гель-электрофореза в денатурирующих условиях (SDS-PAGE). Уровень экспрессии определяют с помощью денситометрии полиакриламидного геля, окрашенного Кумасси-R250 с использованием программного обеспечения AlfaImage, поставляемого с прибором AlphaImager (AlfaInnotech, США). Уровень экспрессии составляет около 0.5% суммарного клеточного белка.
Оба целевых белка локализованы в цитоплазматической фракции соответствующих клеток-продуцентов. Их выделение и очистку проводят с помощью металл-хелатной хроматографии и гель-фильтрации. Степень очистки определяют сканированием геля на AlphaImager (AlfaInnotech, США). Концентрацию очищенных гибридных белков определяют спектрофотометрически с помощью набора DC Protein Assay (Bio-Rad, США) по протоколу производителя.
Полученные гибридные белки обладают способностью связывать белок Е вируса клещевого энцефалита, которая была показана с помощью конкурентного биолюминесцентного твердофазного иммуноанализа.
Полученные гибридные белки порознь не обладают целентеразин-зависимой биолюминесцентной активностью.
Полученные гибридные белки обладают целентеразин-зависимой биолюминесцентной активностью в результате комплементации при связывании с модельной молекулой-мишенью, несущей несколько копий фрагмента белка Е вируса клещевого энцефалита.
При проведении биолюминесцентного микроанализа измерения проводят с помощью планшетного люминометра Mithras LB 940 (Berthold, Германия). Биолюминесценцию инициируют внесением свежеприготовленного раствора целентеразина в растворе 50 мМ Трис-HCl рН 7.0, 25 мМ NaCl, 1 мМ ЭДТА.
Заявляемая группа изобретений соответствует требованию единства изобретения, поскольку образует единый изобретательский замысел с получением единого технического результата, т.к. впервые получены плазмидные ДНК обеспечивающие продукцию в бактериальных клетках Escherichia coli гибридных белков sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a, и гибридные белки sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a, состоящие из одноцепочечного антитела мыши против белка Е вируса клещевого энцефалита и способного связывать белок Е, коннекторных пептидов (Gly2Ser)5 либо (Gly2Ser)4 и большого либо малого фрагментов люциферазы NanoLuc, содержащие N либо С- концевые пептиды НННННН, обладающих способностью связываться с белком Е и комплементацией с возникновением целентеразин-зависимой биолюминесцентной активности при соединении с модельными молекулами мишенями, несущими несколько фрагментов белка Е, что делает их перспективными для выявления вируса клещевого энцефалита однофазным биолюминесцентным анализом.
Сопоставительный анализ с прототипом позволил выявить совокупность существенных по отношению к техническому результату отличительных признаков для каждого из заявляемых объектов группы, изложенных в формулах. Следовательно, каждый из объектов группы изобретений соответствует критерию «новизна».
Аналоги - прототипные белки, способные связывать белок Е вируса клещевого энцефалита и обладающие способностью к самосборке фрагментов люциферазы с возникновением биолюминесцентной активности, отсутствуют.
Конструирование, получение и использование гибридных белков для иммуноанализа вируса клещевого энцефалита с применением фрагментов искусственной люциферазы NanoLuc или их аналогов в качестве репортеров в литературе не описано, что позволяет сделать вывод о соответствии заявляемого решения критерию «изобретательский уровень».
Изобретение иллюстрируется следующими фигурами:
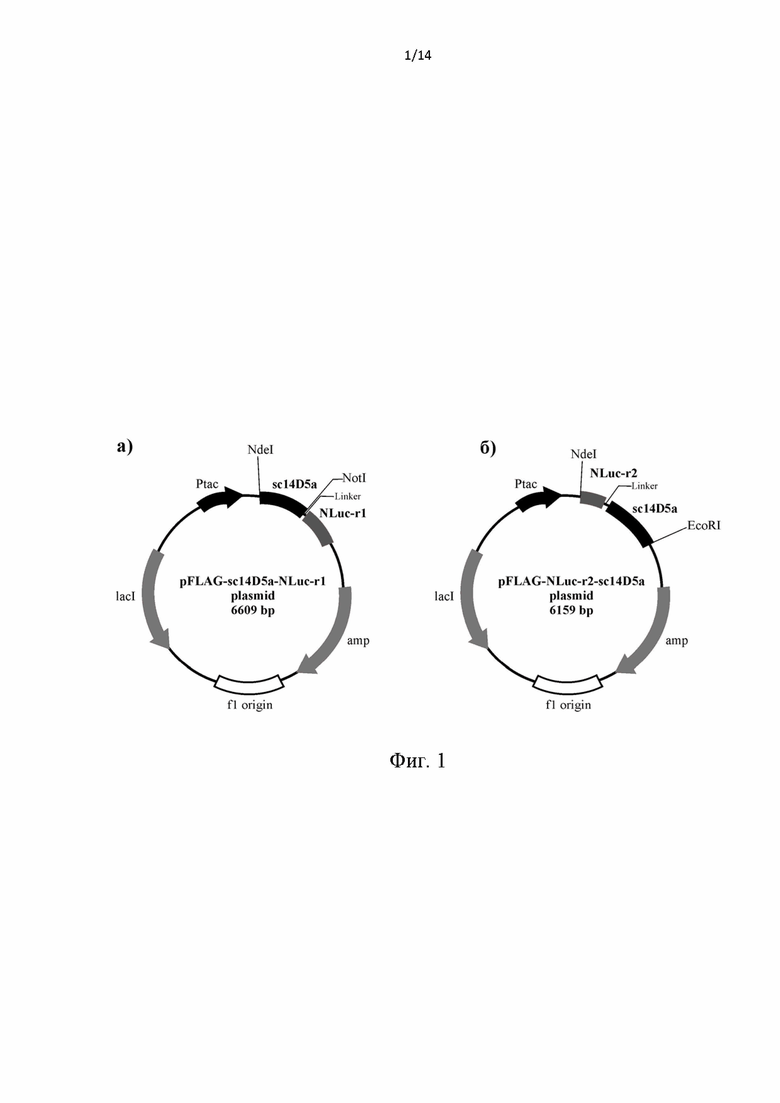
Фиг. 1. Общая схема структурной организации плазмиды pFLAG-sc14D5a-NLuc-r1 (а) и плазмиды pFLAG-NLuc-r2-sc14D5a (б). Обозначения: 14D5a- scNLuc-r1 - ген, кодирующий одноцепочечное антитело мыши, способное связывать белок Е вируса клещевого энцефалита, содержащее N концевой пептид НННННН и большой фрагмент люциферазы NanoLuc; NLuc-r2- sc14D5 - ген, кодирующий малый фрагмент люциферазы NanoLuc и одноцепочечное антитело мыши, способное связывать белок Е вируса клещевого энцефалита, содержащее С концевой пептид НННННН.
Ptac - область промотора; amp - ген устойчивости к ампициллину; указаны некоторые сайты рестрикции.
Фиг. 2. Нуклеотидная последовательность кодирующая гибридный белок sc14D5a-NLuc-r1 SEQ ID No.1.
Фиг. 3. Аминокислотная последовательность гибридного белка sc14D5a-NLuc-r1 SEQ ID No.2.
Фиг. 4. Нуклеотидная последовательность кодирующая гибридный белок NLuc-r2-sc14D5a SEQ ID No.3.
Фиг. 5. Аминокислотная последовательность гибридного белка NLuc-r2-sc14D5a SEQ ID No.4.
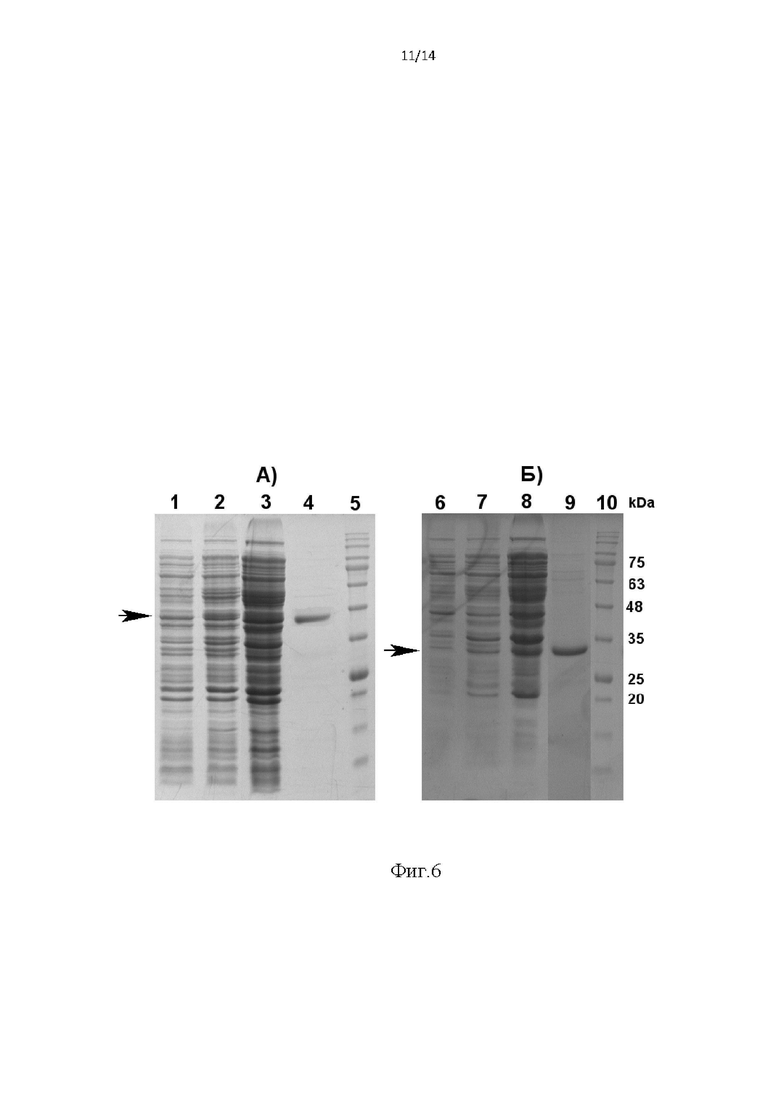
Фиг. 6. Электрофоретический анализ фракций при выделении целевых белков -sc14D5a-NLuc-r1 (А) и NLuc-r2-sc14D5a (Б) проводили в 12,5%-ном денатурирующем SDS-ПААГ. Дорожки:
1, 6 - лизат клеток Rosetta-gami/pFLAG- sc14D5a-NLuc-r1 и Rosetta-gami/pFLAG- NLuc-r2-sc14D5a до добавления ИПТГ;
2,7 - лизат клеток Rosetta-gami/pFLAG- sc14D5a-NLuc-r1 и Rosetta-gami/pFLAG- NLuc-r2-sc14D5a после добавления ИПТГ;
3, 8 - цитоплазматические фракции клеток Rosetta-gami/pFLAG- sc14D5a-NLuc-r1 и Rosetta-gami/pFLAG- NLuc-r2-sc14D5a;
4, 9 - препараты гибридных белков sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a после хроматографической очистки;
5, 10 - маркерные белки, мол. масса (kDa) показана цифрами.
Стрелками показаны полосы целевых белков.
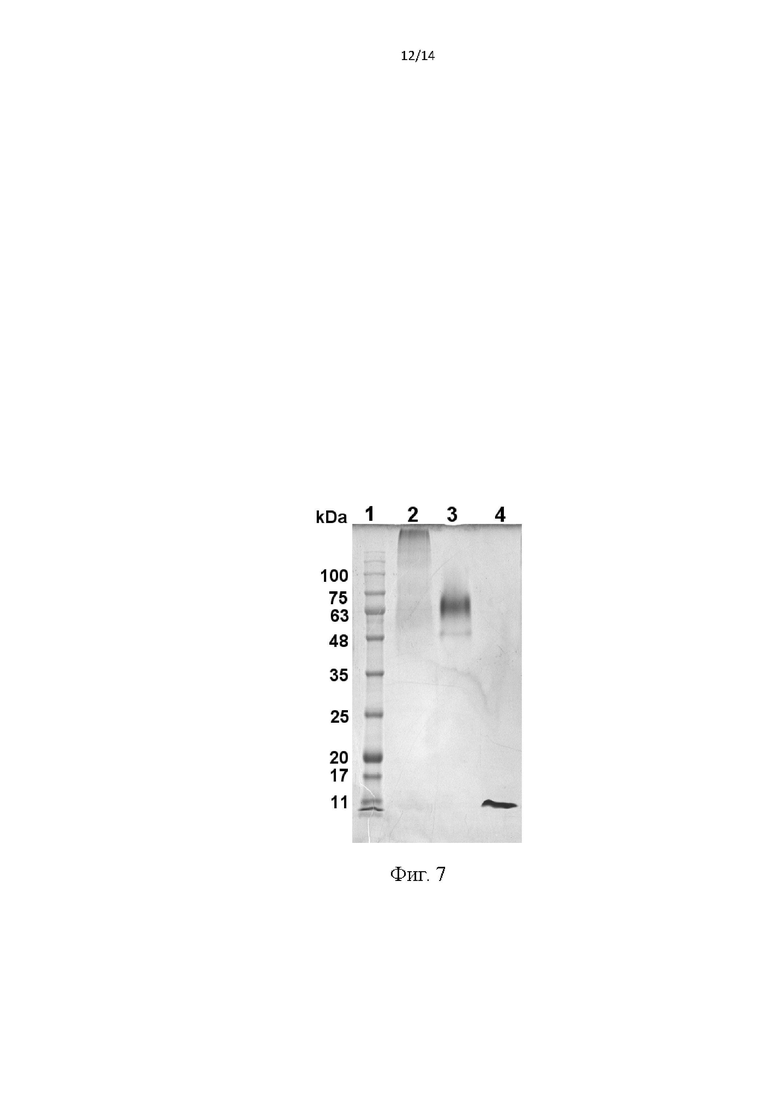
Фиг. 7. Электрофоретический анализ фракций при получении модельного белка-мишени проводили в 10%-ном SDS-ПААГ без денатурации белков. Дорожки: 1 - маркерные белки, мол масса (kDa) показана цифрами; 2 - образец молекулы-аддукта стрептавидина и биотинилированного производного фрагмента белка Е; 3 - исходный стрептавидин; 4 - исходный фрагмент белка Е.
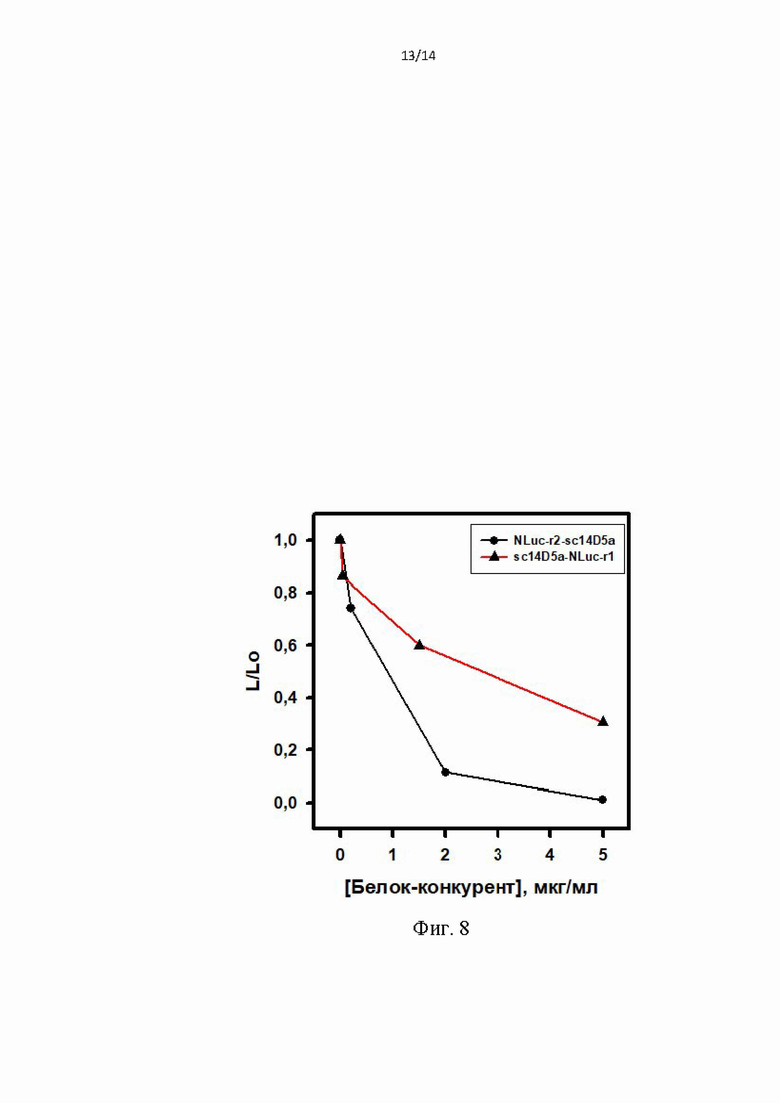
Фиг. 8. Зависимость биолюминесцентного сигнала гибридного белка sc14D5a-Rm7 от концентрации белков-конкурентов sc14D5a-NLuc-r1 (•) или NLuc-r2-sc14D5a (▲) при связывании с фрагментом белка Е, иммобилизованным на поверхности лунок.
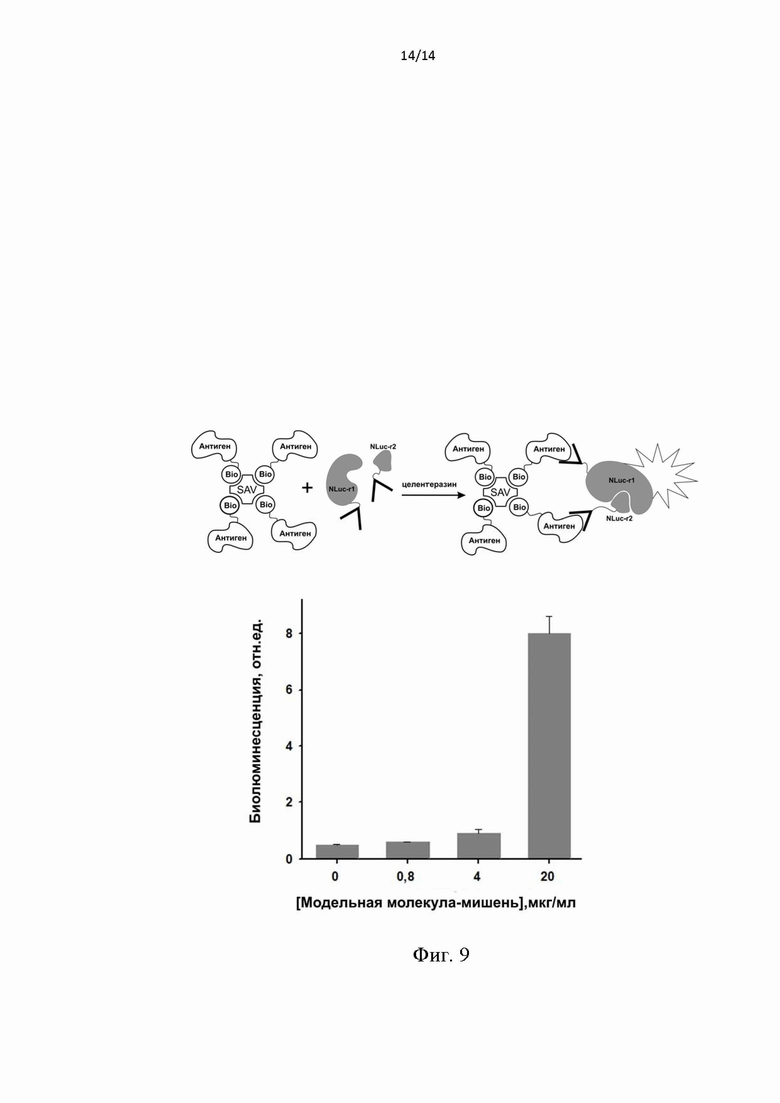
Фиг. 9. Зависимость биолюминесцентного сигнала в результате комплементации фрагментов люциферазы NLuc-r1 и NLuc-r2 от концентрации модельной мишени в образце при однофазном анализе. Обозначения: Bio-биотин, SAV - стрептавидин; Антиген - фрагмент белка Е. Образование комплекса с комплементацией фрагментов люциферазы представлена на схеме сверху.
Для лучшего понимания сущности предлагаемых изобретений, они иллюстрируются следующими примерами осуществления.
Пример 1. Конструирование плазмидной ДНК pFLAG- sc14D5a-NLuc-r1
Для получения конструкции была использована полученная нами ранее плазмида pFLAG-sc14D5a-NLuc кодирующая последовательность гибридного белка, состоящего из миниантитела к клещевому энцефалиту, соединенному с последовательностью полноразмерной люциферазы NanoLuc с гексагистидиновым фрагментом на С-конце.
Фрагмент ДНК кодирующий последовательность миниантитела к клещевому энцефалиту с гексагистидиновым фрагментом с N-конца (730 п.о.) с сайтами рестрикции 5'-(NdeI) и 3'- (NotI) концах получают ПЦР с использованием Tersus-ДНК-полимеразы (Евроген, Россия) и следующих праймеров: Up 5’-CTCCATATGCACCACCACCACCACCACGCCGAGGTGCAGCTGGTG-3’ Dn 5’-CCTGCGGCCGCACGTTTGATTTCCAGCTTGG-3’
Условия ПЦР: 95°С в течение 30 сек; 25 циклов (95°С - 30 с, 60°С - 30 с, 72°С - 1 мин); 72°С в течение 7 мин.
Продукт амплификации расщепляют ферментами рестрикции NdeI и NotI в течение 2 часов при 37°C, заключительная стадия - 20 минут при 65°C.
Обработку ДНК векторной плазмиды pFLAG- sc14D5a-NLuc проводят рестриктазами NdeI и NotI в аналогичных условиях, после чего дефосфорилируют щелочной фосфатазой CIP (NEB, США) в течение 1 часа при 37°C.
Далее ПЦР-фрагмент и линеаризованный вектор очищают электрофоретически в 1% агарозном геле с последующим выделением ДНК набором QIAquick™ Gel Extraction Kit (Quiagen, США) в соответствии с рекомендациями производителя.
Лигирование проводят в стандартном буфере. Полученной лигазной смесью трансформируют клетки Escherichia coli XL1Blue. С помощью полимеразной цепной реакции отбирают клоны, содержащие вставку нужного размера.
В полученной плазмиде вносили стоп кодон в последовательность люциферазы NanoLuc сайт-направленным мутагенезом, с использованием праймеров Up 5'-TTCGTGTCACCATCAATGGTTAAGTGACCGGCTGGCGTTTGTG-3', Dn 5'-CACAAACGCCAGCCGGTCACTTAACCATTGATGGTGACACGAA-3' и набор QuickChange site-directed mutagenesis (Stratagene, США) в соответствии с условиями производителя. Реакционную смесь после мутагенеза обрабатывали рестриктазой Dpn1 (NEB, США) для удаления матричной плазмиды и трансформировали ею клетки E. coli XL1Blue. Клоны, содержащие вставку нужного размера, отбирали с помощью ПЦР.
Полученную таким образом целевую плазмиду обозначают как pFLAG-sc14D5a-NLuc-r1. Схема плазмидной ДНК pFLAG-sc14D5a-NLuc-r1 представлена на Фиг. 1а. Последовательность клонированных фрагментов подтверждена секвенированием и представлена на Фиг. 2 и Фиг. 3.
Пример 2. Конструирование плазмидной ДНК pFLAG-NLuc-r2-sc14D5a
Для получения плазмидной ДНК была использована сконструированная нами ранее плазмида pFLAG- NLuc-r1-sc14D5a, кодирующая последовательность гибридного белка, состоящего из большого фрагмента люциферазы NanoLuc (NLuc-r1) и миниантитела к клещевому энцефалиту sc14D5a с гексагистидиновым фрагментом на С-конце.
Фрагмент ДНК кодирующий малый фрагмент люциферазы NanoLuc соединенный с миниантителом к клещевому энцефалиту с гексагистидиновым фрагментом на С-конце (822 п.о.) с сайтами рестрикции 5'-(NdeI) и 3'- (EcoRI) получают ПЦР с использованием Tersus-ДНК-полимеразы (Евроген, Россия) и следующих праймеров: Up 5'-GTTTGTGTGAACGCATTCTTGGGGGTTCAGGTGGTTCA-3’, Up2 5'-GTGCATATGGTGACCGGCTGGCGTTTGTGTGAACG-3’, Dn 5’-CTCGAATTCTCAGTGGTGGTGGTGGTGGTGCGCACGTTTGATTTCCAG-3’
Условия ПЦР: 95°С в течение 30 сек; 25 циклов (95°С - 30 с, 66°С - 30 с, 72°С - 1 мин); 72°С в течение 7 мин.
Продукт амплификации расщепляют ферментами рестрикции NdeI и EcoRI в течение 2 часов при 37°C, заключительная стадия - 20 минут при 65°C.
Обработку ДНК векторной плазмиды pFLAG-CTS (Sigma-Aldrich, США) проводят рестриктазами NdeI и EcoRI в аналогичных условиях, после чего дефосфорилируют щелочной фосфатазой CIP (NEB, США) в течение 1 часа при 37°C.
Далее ПЦР-фрагмент и линеаризованный вектор очищают электрофоретически в 1% агарозном геле с последующим выделением ДНК набором QIAquick™ Gel Extraction Kit (Quiagen, США) в соответствии с рекомендациями производителя.
Лигирование проводят в стандартном буфере. Полученной лигазной смесью трансформируют клетки Escherichia coli XL1Blue. С помощью полимеразной цепной реакции отбирают клоны, содержащие вставку нужного размера.
Полученную таким образом целевую плазмиду обозначают как pFLAG-NLuc-r2-sc14D5a. Схема плазмидной ДНК pFLAG-NLuc-r2-sc14D5a представлена на Фиг. 1б.
Последовательность клонированных фрагментов подтверждена секвенированием и представлена на Фиг. 4 и Фиг. 5.
Пример 3. Получение гибридных белков sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a
А) Клетки E.coli Rosetta-gami 2 (Novagen, США), трансформированные плазмидой pFLAG-sc14D5a-NLuc-r1, культивировали при активном перемешивании в LB среде, содержащей 200 мкг/мл ампициллина, при 37°C. При оптической плотности культуры ОД590 0,6-0,7 ее охлаждали до 23°C, вносили 0,5 мМ изопропил-β D-1-тиогалактопиранозида (ИПТГ; Диа-М, Россия) и продолжали культивирование при 23°C в течение 20 ч. Клетки осаждали центрифугированием, суспендировали в буфере A (5 мМ имидазол, 0,3 M NaCl, 50 мМ K-Na фосфатном буфере рН 7,0) и дезинтегрировали ультразвуком при охлаждении льдом. Смесь центрифугировали, супернатант наносили на колонку HisTrapTM HP (GE Healthcare), уравновешенную буфером А, и элюировали вещества с колонки линейным градиентом имидазола (50-250 мМ) в том же буфере. Фракции, содержащие целевой белок, объединяли, концентрировали и наносили на колонку HiTrap Desalting (GE Healthcare), уравновешенную 50 мМ K-Na фосфатном буфером рН 7,0, содержащим 150 мМ NaCl (PBS), элюировали этим же раствором. Фракции, содержащие целевой белок, объединяли.
Б) Клетки E.coli Rosetta-gami 2 (Novagen, США), трансформированные плазмидой pFLAG-NLuc-r2-sc14D5a, культивировали как описано выше. Выделение белка NLuc-r2-sc14D5a проводили как описано в А) для белка sc14D5a-NLuc-r1.
Результаты выделения и очистки представлены на Фиг. 6. На фигуре 6 видно (дорожки 2 и 7), что после индукции в клетках появляются новые белки, молекулярная масса которых около 45 кДа и 29 кДа, близка расчетной массе гибридных белков sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a, соответсвенно (показано стрелками). Белки находятся в цитоплазматической фракции (дорожка 3 и 8). После очистки цитоплазматической фракции металл-хелатной хроматографией и гель-фильтрации получены препараты, в которых целевые белки составляют 98% и 96% (дорожки 4 и 9 для sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a, соответственно). Дорожка 5 и 10 - маркеры молекулярных масс.
Пример 4. Получение модельной молекулы-мишени (аддукт стрептавидина с 4-я фрагментами белка Е)
Третий домен белка Е [13] инкубировали с 50-кратным молярным избытком N-гидроксисукцинимидного эфира биотинамидогексановой кислоты (Sigma- Aldrich), 2 часа, при комнатной температуре в 0,1 М NaHCO3. Избыток реагента отделяли гельфильтрацией на колонке HiTrap Desalting (GE Healthcare) уравновешенной PBS. Полученный белок инкубировали со стрептавидином [14] в мольном соотношении 1:4,5 в течение ночи при 5°C. Полученный аддукт использовали без дополинтельной очистки. Получение модельной молекулы контролировали с помощью гель-электрофореза без денатурации белков, как показано на Фиг. 7 Видно, что полученный аддукт (дорожка 2) представляет собой высокомолекулярный комплекс стрептавидина с фрагментами белка Е, отличающийся по мол. массе от исходных белков.
Пример 5. Конкурентный биолюминесцентный анализ связывания гибридных белков с фрагментом белка Е
В лунки планшета вносили по 100 мкл белка Е вируса клещевого энцефалита 1 мкг/мл в PBS и инкубировали 1 ч при 37°C. Промывали (PBS, содержащий 0,1% Tween 20, 5 мМ ЭДТА) и блокировали свободную поверхность лунки инкубированием со 150 мкл 1%-го раствора бычьего сывороточного альбумина в PBS в течение 1 ч при 37°C. После промывки, вносили по 100 мкл растворы PBS, содержащие 14D5-Rm7 (0,5 мкг/мл) и либо белок NLuc-r2-sc14D5a (5; 2; 0,2; 0 мкг/мл) либо белок sc14D5a-NLuc-r1 (5; 1; 0,1; 0 мкг/мл), инкубировали 1 ч при 23°C, затем промывали. Биолюминесценцию измеряли с помощью планшетного люминометра LB 940 Mithras (Berthold, Германия) сразу после впрыска 100 мкл раствора целентеразина (0,1 мкМ, в PBS), в течении 5 секунд. Белки NLuc-r2-sc14D5a либо sc14D5a-NLuc-r1 конкурируют с биолюминесцентным белком sc14D5-Rm7 и за связывание с белком Е: при повышении их концентрации в лунках, уменьшается количество связавшегося белка биолюминесцентного белка sc14D5-Rm7, что отражается в уменьшении биолюминесцентного сигнала от этих лунок. Зависимость относительного биолюминесцентного сигнала (отношение сигнала от рабочей лунки к сигналу от буферного раствора) от концентрации белков NLuc-r2-sc14D5a либо sc14D5a-NLuc-r1 показана на Фиг. 8. Этот пример демонстрирует наличие связывания целевых белков с антигеном за счет аффинности входящего в их состав домена sc14D5a.
Пример 6. Биолюминесцентный анализ модельной мишени однофазным методом.
В лунки планшета вносили раствор белков NLuc-r2-sc14D5a и sc14D5a-NLuc-r1 (2 мкг/мл каждый, 50 мкл в PBS) и модельной молекулы-мишени (20; 4; 0,8; 0 мкг/мл, 50 мкл в PBS), инкубировали 1 ч при 25°C, затем измеряли биолюминесцентную активность образовавшегося комплекса сразу после впрыска 50 мкл раствора целентеразина (0,3 мкМ, в PBS) с помощью планшетного люминометра LB 940 Mithras (Berthold, Германия), сигнал интегрировали в течении 7 секунд. Полученный результат приведен на Фиг. 9. Видно, увеличение биолюминесцентного сигнала от лунок в зависимости от количества модельной мишени. Это происходит благодаря образованию комплексов мишень-гибридные белки NLuc-r2-sc14D5a и sc14D5a-NLuc-r1 в результате чего происходит комплементация большого и малого фрагментов люциферазы, и полученный комплекс генерирует биолюминесцентный сигнал в присутсвии субстрата - целентеразина, аналогично полноразмерной люциферазе NanoLuc.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. T.G.T. Jaenson, M. Hjertqvist, T. Bergström, A. Lundkvist. Why is tick-borne encephalitis increasing? A review of the key factors causing the increasing incidence of human TBE in Sweden. Parasit Vectors. - 2012 - Vol.5. - Р.184-197.
2. Kunze U; ISW-TBE. Tick-borne encephalitis--a notifiable disease: report of the 15th Annual Meeting of the International Scientific Working Group on Tick-Borne Encephalitis (ISW-TBE) Ticks Tick Borne Dis. - 2013 - Vol.4(5). -P.363-365.
3. Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; Robers, M.B.; Benink, H.A.; Eggers, C.T.; Slater, M.R.; Meisenheimer, P.L.; Klaubert, D.H.; Fan, F.; Encell, L.P.; Wood, K.V. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem. Biol. 2012, 7, 1848-1857; DOI: 10.1021/cb3002478.
4. A.S. Dixon, M.K. Schwinn, M.P. Hall, K. Zimmerman, P. Otto, T.H. Lubben, B.L. Butler, B.F. Binkowski, T. Machleidt, T.A. Kirkland, M.G. Wood, C.T. Eggers, L.P. Encell, K.V. Wood. NanoLuc complementation reporter optimized for accurate measurement of protein interactions in cells. ACS Chem. Biol., 2016;11(2):400-408 doi: 10.1021/acschembio.5b00753.
5. Ohmuro-Matsuyama, Y.; Ueda, H. Homogeneous noncompetitive luminescent immunodetection of small molecules by ternary protein fragment complementation. Anal. Chem. 2018, 90, 3001-3004; DOI: 10.1021/acs.analchem.7b05140.
6. Soave, M.; Heukers, R.; Kellam, B.; Woolard, J.; Smith, M.J.; Briddon, S.J.; Hill, S.J. Monitoring allosteric interactions with CXCR4 using NanoBiT conjugated nanobodies. Cell Chem Biol. 2020;27(10):1250-1261.e5. doi: 10.1016/j.chembiol.2020.06.006.
7. Ranawakage, D.C.; Takada, T.; Kamachi, Y. HiBiT-qIP, HiBiT-based quantitative immunoprecipitation, facilitates the determination of antibody affinity under immunoprecipitation conditions. Sci Rep.2019, 9, 6895. doi: 10.1038/s41598-019-43319-y.
8. Cooley, R.; Kara, N.; Hui, N.S.; Tart, J.; Roustan, C.; George, R.; Hancock, D.C.; Binkowski, B.F.; Wood, K.V.; Ismail, M. Development of a cell-free split-luciferase biochemical assay as a tool for screening for inhibitors of challenging protein-protein interaction targets. Wellcome Open Res. 2020, 5: 20. doi: 10.12688/wellcomeopenres.15675.1.
9. LeBoulch,M.;Brossard,A.;LeDez,G.;Leon,S.;Rabut,G.Sensitive detection of protein ubiquitylation using a protein fragment complementation assay. J. Cell Sci. 2020, 133, jcs240093.
10. T. Azad, H.J. Janse van Rensburg, J. Morgan, R. Rezaei, M.J.F. Crupi, R. Chen, M. Ghahremani, M. Jamalkhah, N. Forbes, C. Ilkow, J.C. Bell. Luciferase-based biosensors in the era of the COVID-19 pandemic. ACS Nanosci. Au 2021, 1, 15-37 https://doi.org/10.1021/acsnanoscienceau.1c00009
11. Патент РФ №2565545.
12. L.P. Burakova, A.N. Kudryavtsev, G.A. Stepanyuk, I.K. Baykov, V.V. Morozova, N.V. Tikunova, M.A. Dubova, V.N. Lyapustin, V.V. Yakimenko, L.A. Frank. Bioluminescent detection probe for tick-borne encephalitis virus immunoassay. Anal. Bioanal. Chem. 2015. V. 407(18). P. 5417-5423. doi: 10.1007/s00216-015-8710-6.
13. Байков И.К., Емельянова Л.А., Соколова Л.М., Карелина Е.М., Матвеев А.Л., Бабкин И.В., Хлусевич Я.А., Подгорный В.Ф., Тикунова Н.В. Анализ доменной специфичности протективного химерного антитела ch14D5a против гликопротеина Е вируса клещевого энцефалита. Вавиловский журнал генетики и селекции. 2018;22(4):459-467. DOI 10.18699/VJ18.383.
14. Башмакова, Е.Е. Разработка способа получения функционально активного рекомбинантного стрептавидина в клетках Escherichia coli / Е.Е. Башмакова, А.Н. Кудрявцев, Л.А. Франк. Журн. Сиб. федер. ун-та. Биология, 2020. 13(2). С. 218-229.









Изобретение относится к биотехнологии, генной и белковой инженерии и предназначено для получения генетически слитых гибридных белков. Предложены рекомбинантные плазмиды ДНК pFLAG-sc14D5a-NLuc-r1 и pFLAG-NLuc-r2-sc14D5a, содержащие уникальный ген одноцепочечного антитела мыши, направленного против белка Е вируса клещевого энцефалита, и гены фрагментов люциферазы NLuc-r1 и NLuc-r2. Предложен гибридный белок, экспрессирующийся в трансформированных упомянутыми плазмидными ДНК клетках Escherichia coli Rosetta-gami2 с последующими выделением из цитоплазматической фракции и очисткой металл-хелатной хроматографией гибридных белков sc14D5a-NLuc-r1 и NLuc-r2-sc14D5a. Изобретение позволяет получить два гибридных белка, обладающих способностью связывать белки Е вируса клещевого энцефалита с комплементацией фрагментов люциферазы NanoLuc и возникновением биолюминесцентной активности в присутствии субстрата - целентеразина, как потенциально пригодной высокочувствительной системы для выявления вируса клещевого энцефалита в биологических образцах однофазным биолюминесцентным анализом. 3 н.п. ф-лы, 9 ил., 6 пр.
1. Рекомбинантная плазмидная ДНК pFLAG-sc14D5a-NLuc-r1, обеспечивающая синтез гибридного белка sc14D5a-NLuc-r1 в клетках Escherichia coli, связывающих белок Е вируса клещевого энцефалита, и не обладающая биолюминесцентной активностью люциферазы NanoLuc;
имеющая в соответствии с физической и генетической картой плазмиды, приведенной на Фиг. 1а), размер 6609 п.о. и мол. м. 4.4 Md и содержащая:
- плазмидный вектор pFLAG-CTS, включающий сайт инициации репликации фага M13, промотор tac и уникальные сайты рестрикции NdeI (111), HindIII (171), XhoI (177), EcoRI (182), SmaI (187), XmaI (189), Asp718 (191), KpnI (195), BglII (197), SalI (203);
- NdeI-NotI - фрагмент, представляющий собой два участка размером 354 п.о. и 324 п.о. соответственно, кодирующие вариабельные домены тяжелой и легкой цепей моноклонального антитела sc14D5a против белка Е вируса клещевого энцефалита, между которыми находится коннектор размером 45 п.о., кодирующий линкерный пептид (Gly4Ser)3 и содержащий N-концевой пептид НННННН;
- фрагмент ДНК, представляющий собой последовательность, кодирующую коннекторный пептид (Gly2Ser)5 и большой фрагмент люциферазы NanoLuc - NLuc-r1.
2. Рекомбинантная плазмидная ДНК pFLAG-NLuc-r2-sc14D5a, обеспечивающая синтез гибридного белка NLuc-r2-sc14D5a в клетках Escherichia coli, связывающих белок Е вируса клещевого энцефалита, и не обладающая биолюминесцентной активностью люциферазы NanoLuc;
имеющая в соответствии с физической и генетической картой плазмиды, приведенной на Фиг. 1б), размер 6159 п.о. и мол.м. 4 Md и содержащая:
- плазмидный вектор pFLAG-CTS, включающий сайт инициации репликации фага M13, промотор tac и уникальные сайты рестрикции NdeI (111), HindIII (171), XhoI (177), EcoRI (182), SmaI (187), XmaI (189), Asp718 (191), KpnI (195), BglII (197), SalI (203);
- NdeI-EcoRI - фрагмент, кодирующий малый фрагмент люциферазы NanoLuc - NLuc-r2, коннекторный пептид (Gly2Ser)4, два участка размером 354 п.о. и 324 п.о. соответственно, кодирующие вариабельные домены тяжелой и легкой цепей моноклонального антитела sc14D5a против белка Е вируса клещевого энцефалита, между которыми находится коннектор размером 45 п.о., кодирующий линкерный пептид (Gly4Ser)3 и содержащий С-концевой пептид НННННН;
- генетические маркеры: amp’ - ген ампициллин резистентности (ген β-лактамазы), определяющий устойчивость к ампициллину при трансформации Escherichia coli.
3. Гибридный белок, пригодный для создания высокочувствительных иммунологических тест-систем для выявления вируса клещевого энцефалита однофазным анализом, связывающий белок Е вируса клещевого энцефалита и обладающий способностью комплементации с возникновением целентеразин-зависимой биолюминесцентной активности, при связывании с модельной молекулой-мишенью, несущей несколько копий фрагмента белка Е, имеющий аминокислотную последовательность, кодируемую нуклеотидной последовательностью SEQ ID NО: 1 или SEQ ID NО: 3.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pFLAG-sc14D5a-Rm7, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ГИБРИДНОГО БЕЛКА sc14D5a-Rm7, ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА sc14D5a-Rm7 И ГИБРИДНЫЙ БЕЛОК sc14D5a-Rm7, СВЯЗЫВАЮЩИЙ БЕЛОК Е ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА И ОБЛАДАЮЩИЙ БИОЛЮМИНЕСЦЕНТНОЙ АКТИВНОСТЬЮ | 2014 |
|
RU2565545C1 |
| ALEXANDER N | |||
| KUDRYAVTSEV, et al | |||
| A Test System for Tick-Borne Encephalitis Virus Detection Based on Bioluminescent Immunoassay, Journal of Siberian Federal University | |||
| Способ восстановления спиралей из вольфрамовой проволоки для электрических ламп накаливания, наполненных газом | 1924 |
|
SU2020A1 |
| МАТВЕЕВ А.Л., Проективное химерное антитело против вируса клещевого энцефалита: получение и | |||

