Изобретение относится к рекомбинантному плазмидному вектору pET32-WNV-DIII, обеспечивающему синтез и секрецию рекомбинантного домена III структурного гликопротеина Е вируса лихорадки Западного Нила в клетках E.coli, штамму клеточной линии E.coli BL21(DE3)-DIII-WNV и рекомбинантному белку DIII-WNV, предназначенному для получения иммунобиологических препаратов, и может быть использовано в генной инженерии, биотехнологии и медицине для создания профилактических вакцин против лихорадки Западного Нила, а также в клинической практике для серологического тестирования сывороток крови пациентов на содержание иммуноглобулинов классов IgM и IgG, специфичных к вирусу лихорадки Западного Нила.
Уровень техники
Вирус Западного Нила (WNV) - флавивирус, принадлежащий антигенному комплексу японского энцефалита, переносимый комарами. Вирус широко распространен в Африке, Европе, Австралии, Азии, США, Канаде, Мексике и странах Карибского бассейна, а также в некоторых районах Центральной и Южной Америки [1]. Маркеры WNV выявлены в 61 субъекте Российской Федерации за период с 1999 по 2020 годы [2]. Специфического лечения или вакцины против WNV не существует [3].
Основной мишенью протективных антител против флавивиурсов является поверхностный белок Е. Этот белок флавивирусов (м.м. 50-53 кДа, длина 500 а.о.) формирует димеры на поверхности зрелых вирионов при физиологическом рН. При кислом рН (<6,5) внутри эндосомы происходят необратимые конформационные изменения, включая перестройку белка Е в тримерную форму [4]. Предполагают, что необычно широкий тропизм флавивирусов, их способность реплицироваться в клетках и тканях различных видов комаров, клещей, птиц и млекопитающих связаны со структурными особенностями поверхностного белка Е [5], состоящего из трех доменов (domains - DI, DII и DIII, соответственно). DI расположен центрально и содержит 120 а.о. в трех сегментах (1-51, 137-189 и 285-302 а.о.). Два участка белка Е (52-136, 190-284 а.о.), образующие петли между сегментами DI формируют DII, который в пространстве имеет «пальцевидную структуру», на кончике которой находится последовательность 98-113 а.о. (пептид или петля слияния, fusion peptide) [6]. DIII, расположенный на противоположной стороне от DI, принимает семицепочечную иммуноглобулиноподобную складку и участвует в клеточном прикреплении. Короткие, гибкие линкерные области соединяют домены и допускают конформационные изменения, связанные с созреванием и слиянием вируса [7].
В ряде исследований показано, что основным регионом белка Е, на который вырабатываются нейтрализующие антитела является домен DIII [8, 3]. Поэтому в случае WNV было решено в качестве основного иммуногена использовать рекомбинантный белок, включающий область DIII этого вируса.
Ближайшие аналоги
Известные патенты РФ (RU 2787531 C1, RU 2687435 C1, RU 2413769 C1, RU 2716975 C1, RU 2729403 C1), в которых описан способ получения разных рекомбинантных белков, таких как противоопухолевый модифицированный белок DR5-B человека, гибридный белок альфа-фактор некроза опухолей-тимозин альфа 1 (TNF-Thy), химерный белок прохимозина Vicugna pacos с использованием рекомбинантного плазмидного вектора рЕТ32. В данных патентах используется схожий с нами плазмидный вектор рЕТ32 для создания стабильного продуцента на основе бактериальных клеток E.coli. Отличие состоит в том, что авторы получали другие рекомбинантные белки.
Также известна заявка на изобретение (RU 2018141183 А, МПК C12N 15/00, опубл. 22.05.2020 г.) [12], в которой описана рекомбинантная плазмидная ДНК-конструкция pET32a-NS1-ZIKV, обеспечивающая экспрессию гена NS1, кодирующего неструктурный белок NS1 вируса Зика, предназначенная для получения рекомбинантного белка NS1-ZIKV. Отличие состоит в том, что авторы получали неструктурный белок NS1 вируса Зика.
Известны композиции и способы, относящиеся к антигенам белкового домена iii оболочки флавируса (международная заявка WO 2004016586, МПК G01N 33/53, опубл. 26.02.2004 г.) [13]. Настоящее изобретение касается способов и композиций, включающих антигены домена III белка оболочки флавивируса, для обнаружения вируса и обнаружения антител против вируса. Такие способы и композиции можно использовать для обнаружения вирусов серокомплекса КЭ или инфекции вируса Западного Нила у субъекта, пациента, животного или биологической жидкости. Изобретение также касается наборов для реализации таких способов. В некоторых вариантах реализации наборы содержат рекомбинантный антиген домена III оболочечного белка вируса Западного Нила, полипептид домена III белка оболочки вируса Западного Нила получен из штамма Западного Нила 382-99, EthAn4766, 385-99, Kunjin MRM16, Goblum, TL443, DalAnMg798 или 804994.
Известным аналогом (прототипом) является флавивирусная мультивалентная вакцина домена iii (международная заявка WO 2012045063, МПК А61К 39/12, опубл. 05.04.2012 г. ) [14]. Настоящее изобретение относится к четырехвалентной вакцине против вируса Денге, способам индукции нейтрализующего иммунного ответа против флавивируса, такого как вирус Денге 1-4. Изобретение также относится к способам изготовления вакцины путем введения в клетку-хозяина трансгена, который кодирует белок, содержащий полипептиды домена III Денге, или с помощью очищенных полипептидов dIII, специфичных к серотипам денге 1-4, вирусу желтой лихорадки, вирусу Западного Нила, вирусу японского энцефалита.
Наиболее близким по технической сущности аналогом (прототипом) заявляемого изобретения является заявка на патент RU 2018141179, МПК С07К 14/00, опубл. 22.05.2020 г. [15]. Создана рекомбинантная плазмида pET32a-DomIII-E-ZIKV для получения фрагмента гена Е, кодирующего домен III структурного гликопротеина Е вируса Зика в бактериальной системе E.coli и рекомбинантный белок Dom III-E-ZIKV, предназначенный для индукции специфического иммунитета против вируса Зика. Однако полученный фрагмент гена Е, кодирующего домен III структурного гликопротеина Е вируса Зика обладает недостаточной вируснейтрализующей активностью против вируса Западного Нила (WNV).
Техническим результатом является получение фрагмента DIII белка Е вируса лихорадки Западного Нила (DIII-WNV) со степенью чистоты 90% и конечным выходом 35 мг с 1 л культуры для получения диагностических средств и иммунобиологических препаратов, обладающих более высокой вируснейтрализующей активностью против WNV.
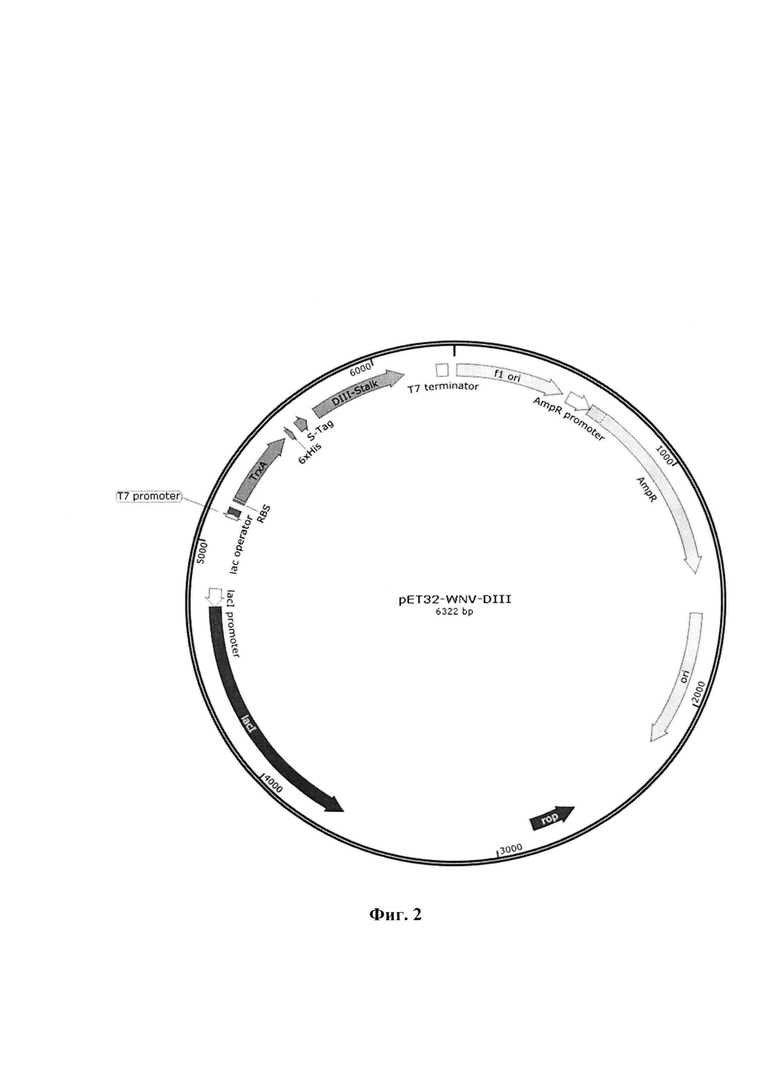
Указанный технический результат достигается созданием плазмидного вектора pET32-WNV-DIII, обеспечивающего экспрессию и секрецию фрагмента DIII белка Е вируса лихорадки Западного Нила (DIII-WNV) в бактериальных клетках, имеющего размер 6322 п.н., нуклеотидную последовательность SEQ ID No. 1 (фиг. 1 и приложение) и содержащего в соответствии с физической и генетической картой, представленной на Фиг. 2, следующие элементы:
- участок начала репликации ori (координаты с 1630 по 2218 п.н);
- lacI (координаты с 3648 по 4730 п.н.), последовательность, кодирующая репрессор лактозного оперона;
- последовательность, кодирующая promoter lacI (координаты с 4731 по 4808 п.н.);
- Т7 promoter (координаты с 5121 по 5139 п.н.), бактериальный промотор Т7 позволяют экспрессировать рекомбинантный белок в Е. coli;
- lac operator (координаты с 5140 по 5164 п.н.), последовательность, кодирующая лактозный оперон, контролирует экспрессию генов;
- последовательность, кодирующая RBS, (координаты с 5195 по 5200 п.н.) эффективный сайт связывания рибосомы из гена бактериофага Т7;
- TrxA (координаты с 5209 по 5535 п.н.) ген кодирующий белок тиоредоксин;
- последовательность, кодирующая 6xHis (координаты с 5557 по 5574 п.н. и с 6166 по 6183 п.н.) полигистидиновый тэг для очистки рекомбинантного белка с помощью металл-хеллатной хроматографии;
- DIII-Stalk (координаты с 5704 по 6113 п.н.), фрагмент генома вируса лихорадки Западного Нила кодирующий DIII поверхностного белка Е и имеющий нуклеотидную последовательность SEQ ID NO: 2 (фиг. 1 и приложение);
- Т7 terminator (координаты с 5704 по 6113 п.н.) последовательность из бактериофага Т7, которая обеспечивает эффективную терминацию транскрипции;
- f1 ori (координаты с 12 по 467 п.н.), точка начала репликации одноцепочечной ДНК;
- AmpR promoter (координаты с 494 по 598 п.н.), бактериальный промотор гена устойчивости к ампициллину;
- AmpR (координаты с 599 по 1459 п.н.), ген устойчивости к антибиотику ампициллину, позволяющий проводить амплификацию плазмиды в E.coli.
Указанный технический результат достигается также созданием рекомбинантного штамма клеточной линии E.coli BL21(DE3)-DIII-WNV, содержащего плазмидный вектор рЕТЗ2-WNV-DIII по п. 1, включающий нуклеотидную последовательность SEQ ID NO: 2 (см. фиг. 1 и приложение) гена, кодирующую фрагмент DIII гликопротеина Е вируса лихорадки Западного Нила и продуцирующего рекомбинантный белок DIII-WNV.
Указанный технический результат достигается также получением рекомбинантного белка DIII-WNV гликопротеина Е WNV, продуцируемого рекомбинантной клеточной линией E.coli BL21(DE3)- DIII-WNV по п. 2, имеющего аминокислотную последовательность SEQ ID NO: 3 (см. фиг. 1 и приложение) и предназначенного для получения иммунобиологических препаратов.
Изобретение поясняется графическими материалами, представленными на фиг. 1-4, и таблицами 1, 2.
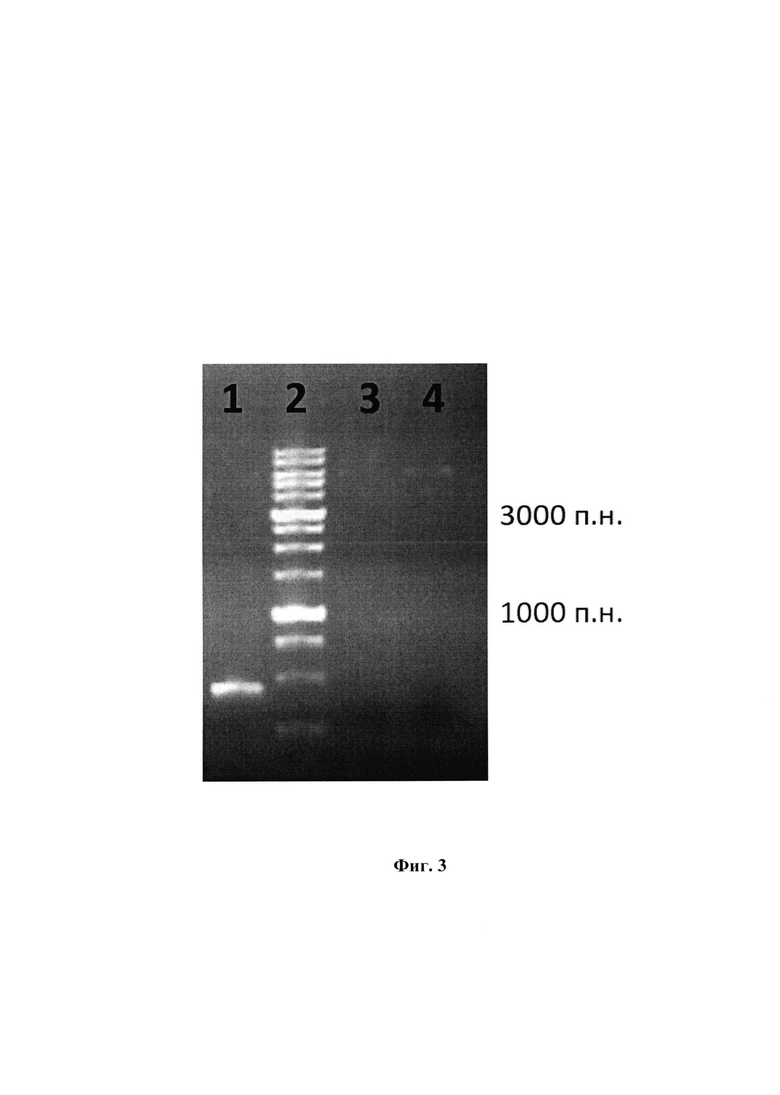
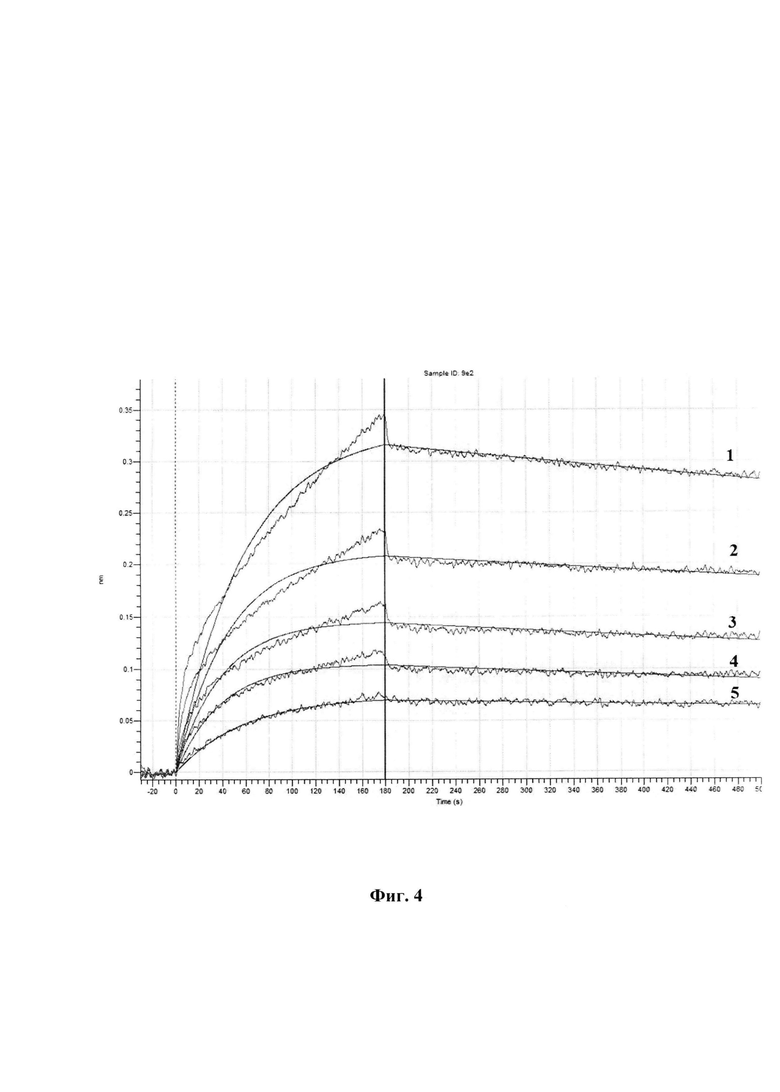
На фиг. 1 представлены последовательности: SEQ ID NO: 1 - нуклеотидная последовательность плазмидного вектора pET32-WNV-DIII; SEQ ID NO: 2 - нуклеотидная последовательность гена, кодирующая фрагмент DIII полипротеина Е вируса лихорадки Западного Нила; SEQ ID NO: 3 - аминокислотная последовательность рекомбинантного фрагмента DIII полипротеина Е вируса лихорадки Западного Нила. На фиг. 2 изображена физическая и генетическая карта плазмидного вектора pET32-WNV-DIII. На фиг. 3 представлена электрофереграмма продуктов ферментативного гидролиза ПЦР продукта WNV-DIII и плазмидного вектора рЕТ32а, где: 1 - ПЦР продукт WNV-DIII, 2 - маркер молекулярных весов (M12, Сибэнзим), 3, 4 - плазмидный вектор рЕТ32а. На фиг. 4 изображены кривые биослойной интерферометрии связывания возрастающих концентраций антитела 9Е2 с иммобилизованными WNV-DIII, где: 1 - концентрация антитела 166,7 нМ; 2 - концентрация антитела 83,35 нМ; 3 - концентрация антитела 41,8 нМ; 4 - концентрация антитела 20,7 нМ; 5 - концентрация антитела 10,4 нМ.
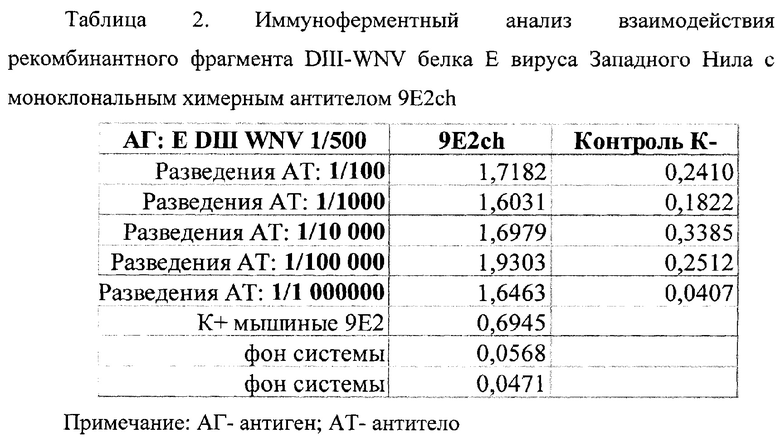
В таблице 1 представлены олигонуклеотидные праймеры использованные для амплификации фрагментов ДНК кодирующих домен III белка Е WNV. В таблице 2. представлен иммуноферментный анализ взаимодействия рекомбинантного фрагмента DIII-WNV белка Е вируса Западного Нила с моноклональным химерным антителом 9E2ch.
Для лучшего понимания сущности предлагаемого изобретения ниже приведены примеры его осуществления. Все стандартные генно-инженерные и микробиологические манипуляции, а также амплификацию и секвенирование ДНК проводили по известным методикам [9].
Пример 1. Конструирование плазмидного pET32-WNV-DIII для синтеза и секреции рекомбинантного белка DIII-WNV в бактериальных клетках
Рекомбинантную плазмиду (фиг. 2), рЕТ32-WNV-DIII содержащую последовательность, кодирующую фрагмент DIII белка Е вируса лихорадки Западного Нила, получали путем встройки продукта ПЦР в акцепторную плазмиду рЕТ32а. В качестве матрицы использовали плазмиду pGH-WNV-DIII, содержащую синтезированную нуклеотидную последовательность, кодирующую фрагмент DIII белка Е вируса лихорадки Западного Нила изолят VLG-4 [GenBank: AF317203] SEQ ID No. 2. При расчете в последовательности олигонуклеотидных праймеров были включены сайты рестрикций BamHI и HinIIIl (таблица 1).

Для амплификации последовательности использовали Pfu-полимеразу, фактический размер ампликона составил 465 п.н. Полученный ампликон и акцепторную плазмиду обрабатывали эндонуклеазами рестрикции в буферах и условиях, рекомендованных производителем (SibEnzyme, Россия). Продукты гидролиза подвергали электрофоретическому разделению (Фиг. 3). ДНК-фрагменты, соответствующие вставкам и вектору, очищали от агарозного геля при помощи набора QIAquick Gel Extraction Kit (QIAGEN Sciences, США).
Выделенные продукты гидролиза лигировали в соответствии с общепринятыми рекомендациями. Лигазной смесью трансформировали компетентные клетки Е. coli штамм BL21. Целостность рамки считывания подтверждалась при помощи ПЦР и секвенирования. В результате была получена плазмида рЕТ32-WNV-DIII (Фиг. 2).
Пример 2. Получение штамма-продуцента рекомбинантной клеточной линии E.coli BL21(DE3)-DIII-WNV, продуцирующий рекомбинантный белок DIII-WNV
Штамм рекомбинантной клеточной линии E.coli BL21(DE3)-DIII-WNV получен на основе клеточной линии E.coli штамма BL21(DE3) с использованием разработанной конструкции рЕТ32-WNV-DIII. К «компетентным» клеткам BL21(DE3) добавляли 10 мкл лигазной смеси, инкубировали на льду в течение 30 минут. После этого клетки подвергали «температурному шоку» при 42°С в течение 45 сек. Охлаждали клетки на льду в течение 2 минут, затем добавляли 200 мкл среды "SOC" и инкубировали при 37°С в течение 60 минут. По окончании инкубации трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (Lysogeny broth) (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл).
Клетки E.coli штамма BL21, трансформированные рекомбинантной плазмидой pET32-WNV-DIII, селективно культивировали в 100 мл жидкой питательной среды LB с добавлением индуктора. В качестве индуктора для синтеза целевого рекомбинантного белка использовали изопропил-β-D-1-тиогалактопиранозид (ИПТГ) в конечной концентрации 1 мМ, варьируя условия культивирования (18°С - 18 часов, 24°С - 16 часов, 37°С - 2 и 4 часа) были подобраны оптимальные условия для наработки рекомбинантного белка DIII-WNV.
Пример 3. Очистка рекомбинантного белка DIII-WNV с помощью металл-хелатной аффинной хроматографии
Бактериальная масса после индукции осаждалась центрифугированием, после чего осадки ресуспензировали в лизирующем буфере с последующей дезинтеграции ультразвуком.
Очистку белка проводили с использованием металл-хелатной аффинной хроматографии с использованием сорбента Ni-IMAC сефарозы (GE Helthcare, США). Клеточный лизат наносился на уравновешенную колонку базовым буфером содержащий 30 мМ NaH2PO4, 500 мМ NaCl, 20 мМ имидазола, и 8 М мочевины с рН 7,4, с последующей промывкой колонки от несвязавшихся белков при помощи базового буфера. Элюция проводилась посредством увеличения концентрации имидазола до 500 мМ.
Полученные фракции анализировались в белковом SDS PAGE электрофорезе (фиг. 3), где: 1 - ПЦР продукт WNV-DIII, 2 - маркер молекулярных весов (M12, Сибэнзим), 3,4-плазмидный вектор рЕТ32а.
Степень чистоты препарата составила не менее 90%. Конечный выход продукта в среднем составил 35 мг с 1 л культуры.
Пример 4. Иммуноферментный анализ взаимодействия рекомбинантного фрагмента DIII-WNV белка Е вируса Западного Нила с моноклональным химерным антителом 9E2ch
Антиген DIII гликопротеина Е WNV в разведение 1/500 сорбировали в лунках 96-луночного планшета в фосфатно-солевом буфере (PBS) (Greiner bio one, Германия) инкубировали в шейкер-термостате на 37±2°С 1 час при 600 об/мин. Затем отмывали буфером PBST (0.1% р-р Tween-20 в PBS) и блокировали 1% раствором казеина в буфере PBST в течение 1 часа в шейкер-термостате на 37±2°С. После этого вносили моноклональное химерное одноцепочечное антитело 9E2ch (патент на изобретение 2801532 С1, 10.08.2023.) в десятикратном последовательном разведении, начиная с 1:10, и инкубировали в течение 1 часа в шейкер-термостате на 37±2°С. Затем добавляли Goat Anti-Human IgG-Fc Secondary Antibody (HRP) (разбавленный в 1% блокирующем растворе казеина 1:1000), конъюгированный с пероксидазой хрена. В качестве отрицательного контроля использовали блокирующий буфер. После каждого этапа проводили отмывку несвязавшихся белков буфером PBST. Затем вносили раствор субстрата ТМВ (Amresco, США) и инкубировали в течение 10 минут, реакцию терминировали раствором 1Н HCl и сразу измеряли оптическую плотность при длине волны 450 нм на ИФА-ридере (ChroMate Awareness technology inc., США). Результаты ИФА представлены в таблице 2.
Пример 5. Анализ кинетики межмолекулярных взаимодействий мышиного антитела 9Е2 с доменом III белка Е WNV
Аналит (доменом III белка Е WNV) разводили до концентрации 20 мкг/мл, антитело (мышиное 9Е2 [10, 11]) разводили до концентрации 166,7 нМ; 83,35 нМ; 41,8 нМ; 20,7 нМ; 10,4 нМ добавляли в лунку микропланшета по 200 мкл. Далее планшет с образцами и сенсорами устанавливали в Octet K2. Сенсоры промывались в промывочном буфере (PBS) в течение 10 мин при перемешивании 1000 об/мин. Затем поверхность сенсоров обрабатывали 0.01М раствором NiCl2 в течение 300 сек при постоянном перемешивании 1000 об/мин. Затем 2 сенсора параллельно погружался в лунку с промывочным буфером и инкубировался в течение 60 сек при постоянном перемешивании 1000 об/мин. Затем первый сенсор погружается в лунку с аналитом, и инкубировался течение 300 сек при постоянном перемешивании 1000 об/мин. Отмывка от не связавшихся аналитов проводилась в промывочном буфере в течение 30 сек при постоянном перемешивании 1000 об/мин. Для измерения ассоциации сенсор погружался в лунку с антителом в концентрации 166,7 нМ, и инкубировался течение 180 сек при постоянном перемешивании 1000 об/мин. Для измерения диссоциации сенсор погружался в промывочный буфер и инкубировались течение 320 сек при постоянном перемешивании 1000 об/мин. Последующая цикличная регенерация проводилась при помощи регенерирующего 50 мМ раствора глицин-HCl рН 1,5 и промывочного буфера на протяжении 3 циклов и на каждом этапе сенсор инкубировался течение 10 сек при постоянном перемешивании 1000 об/мин. Затем процедура повторяется но с уменьшением концентрации антитела. Было проведено по три измерения при пяти разных концентрациях антитела с двухкратным разведением от 166,7 до 10,4 нМ. На Фиг 4 изображены кривые биослойной интерферометрии связывания возрастающих концентраций антитела 9Е2 с иммобилизованными WNV-DIII, где: 1 - концентрация антитела 166,7 нМ; 2 - концентрация антитела 83,35 нМ; 3 - концентрация антитела 41,8 нМ; 4 - концентрация антитела 20,7 нМ; 5 - концентрация антитела 10,4 нМ. В результате были получены значения KD равное 1,46±0,27×10-9.
ИФА (пример 4) и наличие межмолекулярных взаимодействий мышиного антитела 9Е2 с доменом III белка Е WNV (пример 5) доказывают его вируснейтрализующую активность.
Источники информации
1. Батурин А.А. и др. Роль птиц как потенциальных резервуаров вируса Западного Нила на территории Российской Федерации // Проблемы особо опасных инфекций. - 2012. - №. 4. - С. 18-21.
2. Монастырский М.В., Демина Ю.В. Влияние изменения климата на заболеваемость лихорадкой Западного Нила и расширение ареала вируса Западного Нила на территории Российской Федерации // Медицинский алфавит.- 2022. - №. 14. - С. 45-49.
3. Несмеянова В.С., Щербаков Д.Н., Казачинская Е.И. Использование моноклональных антител, специфичных к флавивирусам, для исследования их антигенной структуры и разработки терапевтических препаратов // Молекулярная генетика, микробиология и вирусология. - 2021. - Т. 39. - №. 2. - С. 3-13.
4. Heinz F.X. et al. Field effectiveness of vaccination against tick-borne encephalitis // Vaccine. - 2007. - T. 25. - №. 43. - C. 7559-7567.
5. Campbell G.L. et al. West nile virus // The Lancet infectious diseases. - 2002. - Т. 2. - №. 9. - C. 519-529.
6. Chen W.H. et al. Characterization and epitope mapping of Dengue virus type 1 specific monoclonal antibodies // Virology journal. - 2017. - T. 14. - №. 1. - C. 1-11.
7. Oliphant T. et al. Induction of epitope-specific neutralizing antibodies against West Nile virus // Journal of virology. - 2007. - T. 81. - №. 21. - С.11828-11839.
8. Shrestha B. et al. The development of therapeutic antibodies that neutralize homologous and heterologous genotypes of dengue virus type 1 // PLoS pathogens. - 2010. - T. 6. - №. 4. - C. e1000823, Chen W. H. et al. Characterization and epitope mapping of Dengue virus type 1 specific monoclonal antibodies // Virology journal. - 2017. - T. 14. - №. 1. - C. 1-11.
9. Маниатис Т., Фрич Э, Сэмбрук Дж. Молекулярное клонирование, М.: Мир, 1984; Клонирование ДНК. Методы. Под ред. Д. Гловера, Пер. с англ., Москва, Мир, 1988; Saiki R.K. et al. Science. 1988, 239(4839):487-491; Sanger F. et al. Proc. Nat. Acad. Sci. 1977, 74:5463-5467
10. Казачинская E.И. Алексеенко, Т.П., Разумов, И.А., Протопопова, Е.В., Локтев, В.Б. ШТАММ ГИБРИДНЫХ КЛЕТОК ЖИВОТНОГО MUS MUSCULUS L. 9Е2, ИСПОЛЬЗУЕМЫЙ ДЛЯ ПОЛУЧЕНИЯ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ К БЕЛКУ Е ВИРУСА ЗАПАДНОГО НИЛА ШТАММ WNV/LEIV-VIG99-27889. - 2005.
11. Razumov I.A. Kazachinskaia, Е.I., Ternovoi, V.A., Protopopova, Е. V., Galkina, I.V., Gromashevskii, V.L., Loktev, V.В. Neutralizing monoclonal antibodies against Russian strain of the West Nile virus // Viral Immunology. - 2005. - Т. 18. - №. 3. - C. 558-568.
12. RU 2018141183 А, МПК C12N 15/00, опубл. 22.05.2020 г.
13. Международная заявка WO 2004016586, МПК G01N 33/53, опубл. 26.02.2004 г.
14. Международная заявка WO 2012045063, МПК А61К 39/12, опубл. 05.04.2012 г.
15. RU 2018141179, МПК С07К 14/00, опубл. 22.05.2020 г. (прототип).
Перечень последовательностей
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing originalFreeTextLanguageCode="ru"
nonEnglishFreeTextLanguageCode="ru" dtdVersion="V1_3"
fileName="Приложение_Плазм вектор, штамм E.coli и рек. белок
DIII-WNV.xml" softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2023-12-04">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>1234567</ApplicationNumberText>
<FilingDate>2023-12-04</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>123456</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>12345678</ApplicationNumberText>
<FilingDate>2023-12-01</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное бюджетное учреждение
науки «Государственный научный центр вирусологии и биотехнологии
«Вектор» Федеральной службы по надзору в сфере защиты прав
потребителей и благополучия человека (ФБУН ГНЦ ВБ «Вектор»
Роспотребнадзора)</ApplicantName>
<ApplicantNameLatin>Federalnoe byudzhetnoe uchrezhdenie nauki
"Gosudarstvennyj nauchnyj tsentr virusologii i biotekhnologii
"Vektor" Federalnoj sluzhby po nadzoru v sfere zashchity
prav potrebitelej i blagopoluchiya cheloveka (FBUN GNTS VB
"Vektor" Rospotrebnadzora) (RU)</ApplicantNameLatin>
<InventionTitle languageCode="ru">Рекомбинантный плазмидный вектор
pET32-WNV-DIII, обеспечивающий синтез и секрецию рекомбинантного
домена III структурного гликопротеина Е вируса лихорадки Западного
Нила в клетках E.coli, штамм клеточной линии E.coli
BL21(DE3)-DIII-WNV и рекомбинантный белок DIII-WNV, предназначенный
для получения иммунобиологических препаратов</InventionTitle>
<SequenceTotalQuantity>3</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>6322</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..6322</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tggcgaatgggacgcgccctgtagcggcgcattaagcgcggcgggtgtg
gtggttacgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctcctttcgctttcttccctt
cctttctcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttccgatt
tagtgctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgtagtgggccatcgccc
tgatagacggtttttcgccctttgacgttggagtccacgttctttaatagtggactcttgttccaaactg
gaacaacactcaaccctatctcggtctattcttttgatttataagggattttgccgatttcggcctattg
gttaaaaaatgagctgatttaacaaaaatttaacgcgaattttaacaaaatattaacgtttacaatttca
ggtggcacttttcggggaaatgtgcgcggaacccctatttgtttatttttctaaatacattcaaatatgt
atccgctcatgagacaataaccctgataaatgcttcaataatattgaaaaaggaagagtatgagtattca
acatttccgtgtcgcccttattcccttttttgcggcattttgccttcctgtttttgctcacccagaaacg
ctggtgaaagtaaaagatgctgaagatcagttgggtgcacgagtgggttacatcgaactggatctcaaca
gcggtaagatccttgagagttttcgccccgaagaacgttttccaatgatgagcacttttaaagttctgct
atgtggcgcggtattatcccgtattgacgccgggcaagagcaactcggtcgccgcatacactattctcag
aatgacttggttgagtactcaccagtcacagaaaagcatcttacggatggcatgacagtaagagaattat
gcagtgctgccataaccatgagtgataacactgcggccaacttacttctgacaacgatcggaggaccgaa
ggagctaaccgcttttttgcacaacatgggggatcatgtaactcgccttgatcgttgggaaccggagctg
aatgaagccataccaaacgacgagcgtgacaccacgatgcctgcagcaatggcaacaacgttgcgcaaac
tattaactggcgaactacttactctagcttcccggcaacaattaatagactggatggaggcggataaagt
tgcaggaccacttctgcgctcggcccttccggctggctggtttattgctgataaatctggagccggtgag
cgtgggtctcgcggtatcattgcagcactggggccagatggtaagccctcccgtatcgtagttatctaca
cgacggggagtcaggcaactatggatgaacgaaatagacagatcgctgagataggtgcctcactgattaa
gcattggtaactgtcagaccaagtttactcatatatactttagattgatttaaaacttcatttttaattt
aaaaggatctaggtgaagatcctttttgataatctcatgaccaaaatcccttaacgtgagttttcgttcc
actgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcgtaatctg
ctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctaccaactctt
tttccgaaggtaactggcttcagcagagcgcagataccaaatactgtccttctagtgtagccgtagttag
gccaccacttcaagaactctgtagcaccgcctacatacctcgctctgctaatcctgttaccagtggctgc
tgccagtggcgataagtcgtgtcttaccgggttggactcaagacgatagttaccggataaggcgcagcgg
tcgggctgaacggggggttcgtgcacacagcccagcttggagcgaacgacctacaccgaactgagatacc
tacagcgtgagctatgagaaagcgccacgcttcccgaagggagaaaggcggacaggtatccggtaagcgg
cagggtcggaacaggagagcgcacgagggagcttccagggggaaacgcctggtatctttatagtcctgtc
gggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgtcaggggggcggagcctatggaaaa
acgccagcaacgcggcctttttacggttcctggccttttgctggccttttgctcacatgttctttcctgc
gttatcccctgattctgtggataaccgtattaccgcctttgagtgagctgataccgctcgccgcagccga
acgaccgagcgcagcgagtcagtgagcgaggaagcggaagagcgcctgatgcggtattttctccttacgc
atctgtgcggtatttcacaccgcatatatggtgcactctcagtacaatctgctctgatgccgcatagtta
agccagtatacactccgctatcgctacgtgactgggtcatggctgcgccccgacacccgccaacacccgc
tgacgcgccctgacgggcttgtctgctcccggcatccgcttacagacaagctgtgaccgtctccgggagc
tgcatgtgtcagaggttttcaccgtcatcaccgaaacgcgcgaggcagctgcggtaaagctcatcagcgt
ggtcgtgaagcgattcacagatgtctgcctgttcatccgcgtccagctcgttgagtttctccagaagcgt
taatgtctggcttctgataaagcgggccatgttaagggcggttttttcctgtttggtcactgatgcctcc
gtgtaagggggatttctgttcatgggggtaatgataccgatgaaacgagagaggatgctcacgatacggg
ttactgatgatgaacatgcccggttactggaacgttgtgagggtaaacaactggcggtatggatgcggcg
ggaccagagaaaaatcactcagggtcaatgccagcgcttcgttaatacagatgtaggtgttccacagggt
agccagcagcatcctgcgatgcagatccggaacataatggtgcagggcgctgacttccgcgtttccagac
tttacgaaacacggaaaccgaagaccattcatgttgttgctcaggtcgcagacgttttgcagcagcagtc
gcttcacgttcgctcgcgtatcggtgattcattctgctaaccagtaaggcaaccccgccagcctagccgg
gtcctcaacgacaggagcacgatcatgcgcacccgtggggccgccatgccggcgataatggcctgcttct
cgccgaaacgtttggtggcgggaccagtgacgaaggcttgagcgagggcgtgcaagattccgaataccgc
aagcgacaggccgatcatcgtcgcgctccagcgaaagcggtcctcgccgaaaatgacccagagcgctgcc
ggcacctgtcctacgagttgcatgataaagaagacagtcataagtgcggcgacgatagtcatgccccgcg
cccaccggaaggagctgactgggttgaaggctctcaagggcatcggtcgagatcccggtgcctaatgagt
gagctaacttacattaattgcgttgcgctcactgcccgctttccagtcgggaaacctgtcgtgccagctg
cattaatgaatcggccaacgcgcggggagaggcggtttgcgtattgggcgccagggtggtttttcttttc
accagtgagacgggcaacagctgattgcccttcaccgcctggccctgagagagttgcagcaagcggtcca
cgctggtttgccccagcaggcgaaaatcctgtttgatggtggttaacggcgggatataacatgagctgtc
ttcggtatcgtcgtatcccactaccgagatgtccgcaccaacgcgcagcccggactcggtaatggcgcgc
attgcgcccagcgccatctgatcgttggcaaccagcatcgcagtgggaacgatgccctcattcagcattt
gcatggtttgttgaaaaccggacatggcactccagtcgccttcccgttccgctatcggctgaatttgatt
gcgagtgagatatttatgccagccagccagacgcagacgcgccgagacagaacttaatgggcccgctaac
agcgcgatttgctggtgacccaatgcgaccagatgctccacgcccagtcgcgtaccgtcttcatgggaga
aaataatactgttgatgggtgtctggtcagagacatcaagaaataacgccggaacattagtgcaggcagc
ttccacagcaatggcatcctggtcatccagcggatagttaatgatcagcccactgacgcgttgcgcgaga
agattgtgcaccgccgctttacaggcttcgacgccgcttcgttctaccatcgacaccaccacgctggcac
ccagttgatcggcgcgagatttaatcgccgcgacaatttgcgacggcgcgtgcagggccagactggaggt
ggcaacgccaatcagcaacgactgtttgcccgccagttgttgtgccacgcggttgggaatgtaattcagc
tccgccatcgccgcttccactttttcccgcgttttcgcagaaacgtggctggcctggttcaccacgcggg
aaacggtctgataagagacaccggcatactctgcgacatcgtataacgttactggtttcacattcaccac
cctgaattgactctcttccgggcgctatcatgccataccgcgaaaggttttgcgccattcgatggtgtcc
gggatctcgacgctctcccttatgcgactcctgcattaggaagcagcccagtagtaggttgaggccgttg
agcaccgccgccgcaaggaatggtgcatgcaaggagatggcgcccaacagtcccccggccacggggcctg
ccaccatacccacgccgaaacaagcgctcatgagcccgaagtggcgagcccgatcttccccatcggtgat
gtcggcgatataggcgccagcaaccgcacctgtggcgccggtgatgccggccacgatgcgtccggcgtag
aggatcgagatcgatctcgatcccgcgaaattaatacgactcactataggggaattgtgagcggataaca
attcccctctagaaataattttgtttaactttaagaaggagatatacatatgagcgataaaattattcac
ctgactgacgacagttttgacacggatgtactcaaagcggacggggcgatcctcgtcgatttctgggcag
agtggtgcggtccgtgcaaaatgatcgccccgattctggatgaaatcgctgacgaatatcagggcaaact
gaccgttgcaaaactgaacatcgatcaaaaccctggcactgcgccgaaatatggcatccgtggtatcccg
actctgctgctgttcaaaaacggtgaagtggcggcaaccaaagtgggtgcactgtctaaaggtcagttga
aagagttcctcgacgctaacctggccggttctggttctggccatatgcaccatcatcatcatcattcttc
tggtctggtgccacgcggttctggtatgaaagaaaccgctgctgctaaattcgaacgccagcacatggac
agcccagatctgggtaccgacgacgacgacaaggccatggctgatatcggatccacaacctatggcgtct
gttcaaaggctttcaagtttcttgggactcccgcagacacaggtcacggcactgtggtgttggaattgca
gtacactggcacggatggaccttgcaaagttcctatctcgtcagtggcttcattgaacgacctaacgcca
gtgggcagattggtcactgtcaacccttttgtttcagtggccacggccaacgctaaggtcctgattgaat
tggaaccaccctttggagactcatacatagtggtgggcagaggagaacaacagatcaatcaccattggca
caagtctggaagtagcattggcaaagcctttacaaccaccctcaaaggagcgcagagactagccgctcta
ggagacacagcttgggactttggatcagttggaggggtgttcacctcagttgggaaggctgtccatcaag
tgtaaaagcttgcggccgcactcgagcaccaccaccaccaccactgagatccggctgctaacaaagcccg
aaaggaagctgagttggctgctgccaccgctgagcaataactagcataaccccttggggcctctaaacgg
gtcttgaggggttttttgctgaaaggaggaactatatccggat</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>410</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..410</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acaacctatggcgtctgttcaaaggctttcaagtttcttgggactcccg
cagacacaggtcacggcactgtggtgttggaattgcagtacactggcacggatggaccttgcaaagttcc
tatctcgtcagtggcttcattgaacgacctaacgccagtgggcagattggtcactgtcaacccttttgtt
tcagtggccacggccaacgctaaggtcctgattgaattggaaccaccctttggagactcatacatagtgg
tgggcagaggagaacaacagatcaatcaccattggcacaagtctggaagtagcattggcaaagcctttac
aaccaccctcaaaggagcgcagagactagccgctctaggagacacagcttgggactttggatcagttgga
ggggtgttcac</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>136</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..136</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>TTYGVCSKAFKFLGTPADTGHGTVVLELQYTGTDGPCKVPISSVASLND
LTPVGRLVTVNPFVSVATANAKVLIELEPPFGDSYIVVGRGEQQINHHWHKSGSSIGKAFTTTLKGAQRL
AALGDTAWDFGSVGGVF</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---




Изобретение относится к области биотехнологии. Описана группа изобретений, включающая рекомбинантный плазмидный вектор pET32-WNV-DIII, обеспечивающий синтез и секрецию рекомбинантного домена III структурного гликопротеина Е вируса лихорадки Западного Нила в клетках бактерии E.coli, рекомбинантный штамм клеточной линии E.coli BL21(DE3)-DIII-WNV, продуцирующий рекомбинантный белок DIII-WNV и рекомбинантный белок DIII-WNV гликопротеина Е WNV, продуцируемый рекомбинантной клеточной линией E.coli BL21(DE3)-DIII-WNV, и предназначенный для получения иммунобиологических препаратов. В одном из вариантов реализации рекомбинантный плазмидный вектор содержит последовательность SEQ ID NO:1. Техническим результатом является получение фрагмента DIII белка Е вируса лихорадки Западного Нила (DIII-WNV) со степенью чистоты 90% и конечным выходом 35 мг с 1 л культуры для получения диагностических средств и иммунобиологических препаратов, обладающих более высокой вируснейтрализующей активностью против WNV. 3 н.п. ф-лы, 4 ил., 2 табл., 5 пр.
1. Рекомбинантный плазмидный вектор pET32-WNV-DIII, обеспечивающий синтез и секрецию рекомбинантного домена III структурного гликопротеина Е вируса лихорадки Западного Нила в клетках бактерии E.coli, имеющий размер 6322 п.н., нуклеотидную последовательность SEQ ID NO: 1 и содержащий в соответствии с физической и генетической картой следующие элементы:
- участок начала репликации ori, имеющий координаты с 1630 по 2218 п.н.;
- lacI - последовательность, кодирующая репрессор лактозного оперона, имеющая координаты с 3648 по 4730 п.н.;
- последовательность, кодирующая promoter lacI, имеющая координаты с 4731 по 4808 п.н.;
- Т7 promoter - бактериальный промотор Т7, позволяющий экспрессировать рекомбинантный белок в Е.coli и имеющий координаты с 5121 по 5139 п.н.;
- lac operator - последовательность, кодирующая лактозный оперон, контролирующая экспрессию генов и имеющая координаты с 5140 по 5164 п.н.;
- последовательность, кодирующая RBS - эффективный сайт связывания рибосомы из гена бактериофага Т7 и имеющая координаты с 5195 по 5200 п.н.;
- TrxA - ген, кодирующий белок тиоредоксин и имеющий координаты с 5209 по 5535 п.н.;
- последовательность, кодирующая 6xHis полигистидиновый тэг для очистки рекомбинантного белка с помощью металл-хеллатной хроматографии и имеющая координаты с 5557 по 5574 п.н. и с 6166 по 6183 п.н.;
- DIII-Stalk - фрагмент генома вируса лихорадки Западного Нила, кодирующий DIII поверхностный белок Е, имеющий координаты с 5704 по 6113 п.н. и нуклеотидную последовательность SEQ ID NO: 2;
- Т7 terminator - последовательность из бактериофага Т7, которая обеспечивает эффективную терминацию транскрипции и имеет координаты с 5704 по 6113 п.н.;
- f1 ori - точка начала репликации одноцепочечной ДНК, имеющая координаты с 12 по 467 п.н.;
- AmpR promoter - бактериальный промотор гена устойчивости к ампициллину, имеющий координаты с 494 по 598 п.н.;
- AmpR - ген устойчивости к антибиотику ампициллину, позволяющий проводить амплификацию плазмиды в E.coli и имеющий координаты с 599 по 1459 п.н.
2. Рекомбинантный штамм клеточной линии E.coli BL21(DE3)-DIII-WNV, содержащий плазмидный вектор pET32-WNV-DIII по п. 1, включающий нуклеотидную последовательность SEQ ID NO: 2 гена, кодирующего фрагмент DIII гликопротеина Е вируса лихорадки Западного Нила, и продуцирующий рекомбинантный белок DIII-WNV.
3. Рекомбинантный белок DIII-WNV гликопротеина Е WNV, продуцируемый рекомбинантной клеточной линией E.coli BL21(DE3)-DIII-WNV по п. 2, имеющий аминокислотную последовательность SEQ ID NO: 3 и предназначенный для получения иммунобиологических препаратов.
| Рекомбинантный плазмидный вектор pVEAL-M12B9ch, обеспечивающий стабильную экспрессию и секрецию химерного моноклонального антитела M12B9ch против ортопоксвирусов в клетках млекопитающих, и рекомбинантное химерное моноклональное scFv-Fc антитело M12B9ch, полученное с использованием указанного вектора pVEAL-M12B9ch | 2021 |
|
RU2790134C1 |
| RU 2018141179 A, 22.05.2020 | |||
| Chu J.H.J., Chiang C.C.S., Ng M.L | |||
| Immunization of flavivirus West Nile recombinant envelope domain III protein induced specific immune response and protection against West Nile virus infection //The Journal of Immunology, 2007, 178(5), p | |||
| Способ получения кодеина | 1923 |
|
SU2699A1 |
| Bahakoula S.P., Guo X., Dongxia Z | |||

