Область техники, к которой относится изобретение
Настоящее изобретение предоставляет искусственную целлюлосому для ферментативного расщепления эластичных субстратов. В частности, настоящее изобретение предоставляет комплекс, содержащий основной каркас, имеющий по меньшей мере четыре участка связывания, способных связывать ферментные компоненты, вследствие чего по меньшей мере два участка связывания обладают по существу одинаковой специфичностью связывания; и по меньшей мере три разных ферментных компонента случайным образом связываются с по меньшей мере четырьмя участками связывания. Кроме того, настоящее изобретение относится к способу получения этого комплекса. Помимо этого, настоящее изобретение относится к применению указанных комплексов, а также разных ферментных компонентов для ферментативного расщепления эластичных субстратов, таких как целлюлоза.
Уровень техники
Целлюлоза является широко распространенным возобновляемым источником в области биотехнологии для изготовления различных видов биотоплива и структурных компонентов для химической промышленности. С наступлением приближающегося второго поколения Белой Биотехнологии (промышленной биотехнологии) целлюлоза из лигноцеллюлозной биомассы рассматривается в качестве самого крупного источника сахара, используемого для ферментации в промышленном масштабе. В настоящее время глюкоза для ферментации в промышленном масштабе производится главным образом из крахмала. Использование целлюлозы для производства сахара могло бы по меньшей мере удвоить урожай сельскохозяйственных продуктов в расчете на гектар. Кроме того, может быть увеличена площадь пахотных земель благодаря возможности выращивания большего разнообразия энергетических растений, в том числе таких, которые растут на обедненной почве, при неблагоприятных климатических условиях, а также в засушливых, влажных, холодных или обогащенных солями условиях.
Однако целлюлоза представляет собой прочный материал, устойчивый как к ферментативной, так и химико-физической деградации. Целлюлоза состоит из длинных волокон, составленных из линейных молекул, не имеющих ответвлений. С химической точки зрения она представляет собой в высокой степени гомогенный β-1,4-глюкан, который образует регулярные кристаллы формы Iα и Iβ1. Однако эти кристаллы не являются идеально структурированными - через более или менее регулярные промежутки они прерываются аморфными областями. Следовательно, структурные особенности целлюлозы являются разнообразными, и необходимы ферменты, катализирующие расщепление различного типа.
Для деградации целлюлозных материалов в современном технологическом процессе ферментативного гидролиза целлюлозы необходимо использовать большое количество ферментов, как правило, грибкового происхождения. Как правило, эти ферменты нельзя использовать повторно. Гидролиз является довольно неэффективным в силу ряда причин: неоднородности материала, комплексообразования с гемицеллюлозой, пектином и лигнином, кристалличности целлюлозы, недоступности для ферментов из-за плотной упаковки в клеточных стенках и кристаллах и т.д. Это увеличивает количество различных видов ферментативной активности, необходимых для деградации, а также снижает скорость реакции и, следовательно, выход процесса. Скорость реакции может быть увеличена путем повышения температуры реакции, но платой за это является стабильность ферментов. Особенно сложным этот вопрос является для группы ферментов целлюлаз, которые по своей сути являются медленно реагирующими ферментами, которые работают с кристаллическим, т.е. нерастворимым субстратом. Разнообразие видов воздействия целлюлазы на целлюлозу объясняет многообразие различных ферментов, задействованных в комплексах целлюлаз: структурной гетерогенностью волокна целлюлозы (кристаллическое или аморфное, края или плоскости и т.д.), типом кристалла (Iα или Iβ), видом активности (процессивная экзоглюканаза или целлобиогидролаза, процессивная или непроцессивная эндоглюканаза (β-глюкозидаза). Для эффективной деградации кристаллического субстрата эти ферменты должны функционировать гармонично, несмотря на то, что все они расщепляют идентичные β-1,4-глюкозидные связи.
Коммерчески доступные целлюлазы, обычно используемые в Белой Биотехнологии, содержат смесь растворимых ферментов грибов. Наиболее успешными продуцентами наряду с другими являются Trichoderma longibrachiatum, Т. reesei (= Hypocrea jecorina), Т. viride (= Т. harzianum или Hypocrea atroviridis), Aspergillus niger, Phanerochaete chrysosporium, Chrysosporium lucknowense и Penicillium janthinellum. Некоторые смеси целлюлаз получают из двух микроорганизмов, например, из T. longibrachiatum и A. niger или из T. longibrachiatum и Т. reesei. Эти грибы продуцируют в культуральной жидкости большое количество экзопротеинов, в частности после интенсивного усовершенствования штаммов путем отбора и при помощи генной инженерии. Целлюлазы содержат эндоглюканазы, целлобиогидролазы (экзоглюканазы) и β-глюкозидазы, в некоторых штаммах - также несколько гемицеллюлаз.
Для этой новой области технологии прогнозируется резкое увеличение спроса на ферментативные составы для гидролиза устойчиво продуцируемых возобновляемых источников сахара, таких как лигноцеллюлозный материал. Другими областями использования целлюлаз являются в основном области, связанные с продуктами питания, текстилем, моющими средствами, бумажной промышленностью и добавками для кормов животным. Хотя целлюлазы уже широко представлены на рынке, в целом ожидается, что будущие области применения будут в основном относиться к производству целлюлаз для только что зарождающегося и потенциально гораздо более объемного рынка, связанного с расщеплением биомассы, в секторах химической биотехнологии, связанных и производством биотоплива и крупнотоннажным производством.
Однако различные структурные характеристики кристаллов целлюлозы и тот факт, что они по своей природе являются нерастворимыми, требуют наличия одновременно нескольких различных видов активности, таких как процессивные и непроцессивные целлюлазы. По отдельности эти ферменты обладают более низкой активностью; и только в сочетании они демонстрируют высокую активность. Эта разница между одним и множеством ферментов в смеси называется синергизмом, при котором активность смеси превышает сумму активностей ферментов, используемых по отдельности. Однако такую синергию можно наблюдать у ферментов грибов в смеси, в которой растворены единичные ферменты, не состоящие в “комплексе”, т.е. упакованные вместе в результате адсорбции, или в полипептиде. Синергия между растворимыми единичными ферментами исключает наличие по меньшей мере двух совместно работающих видов активности на одном участке субстрата. В растворимых системах это возможно только в случае высокой концентрации ферментов в смеси. Примерами ограничений, присущих целлюлазам грибов, являются относительно высокие концентрации, необходимые для ферментативного гидролиза, ограниченная термостабильность и высокий уровень неудачного связывания. Для получения более высокой производительности эти ограничения необходимо преодолеть.
Коммерчески доступные целлюлазы в основном относятся к ферментам грибов. Однако бактериальные ферментативные системы также интенсивно изучались. Растворимые ферменты Thermomonospora bispora или других термофильных аэробных бактерий обсуждались в качестве добавок к смесям целлюлаз грибов. Были описаны некоторые анаэробные бактерии, чьи внеклеточные ферментативные системы обладают более высокой удельной активностью и процессивностью по отношению к целлюлозе. Большинство из последних продуцируют большой внеклеточный ферментативный комплекс, в котором единичные ферменты связаны на основном каркасе, так называемом скаффолдине или Cip (интегрирующем целлюлосому белке). Эти комплексы удерживаются вместе благодаря сильным белок-белковым взаимодействиям, которые являются видоспецифичными. Эти комплексы называются целлюлосомами. Известно, что целлюлосомы продуцирует относительно небольшое количество бактерий. В настоящее время их список включает:
в типе Firmicutes:
Lachnospiraceae - Butyrivibrio fibrisolvens, Ruminococcus flavefaciens, R. albus;
Clostridiaceae - Clostridium cellulovorans, С cellobioparum, С papyrosolvens, С josui, C. cellulolyticum, С thermocellum, С sp. C7, Bacteroides sp. P-1, B. cellulosovens, Acetivibrio cellulolyticus;
в типе Actinobacteria:
Nocardiopsaceae - Thermobifida (Thermonospora) fusca;
в типе Fibrobacteres:
Fibrobacteriaceae - Fibrobacter succinogenes.
Некоторого улучшения в анализе эффективности целлюлосомы можно достичь, используя строго анаэробные, термофильные бактерии вида Clostridium thermocellum, которые являются микроорганизмами, обладающими самой высокой скоростью роста на трудно разлагаемом субстрате, кристаллической целлюлозе, поскольку они имеют ферментные системы3, которые являются одними из самых эффективных в отношении деградации целлюлозы. Не вдаваясь в теорию, следует отметить, что накапливаются некоторые доказательства того, что такая более высокая эффективность по сравнению с другими целлюлолитическими системами связана с формированием огромного ферментного комплекса, который, однако, нельзя получать в промышленных масштабах. Этот комплекс имеет диаметр, равный примерно 18 нм, а его масса превышает 2×106 Да4. Для ферментативно активных клонов более или менее случайным образом путем скрининга геномных библиотек, полученных из С. Thermocellum, было клонировано около 30 генов, связанных с содержащими докерины целлюлосомами5,6. Кроме того, был идентифицирован белок скаффолдин CipA, содержащий 9 модулей когезина типа I, с которыми особым образом стыкуются ферменты и другие белковые компоненты посредством своих 7 модулей докерина типа I7. Посредством взаимодействия когезин типа II-докерин белок CipA прикрепляется к соединенным с клеточной стенкой белкам OlpB или SdbA и возможно другим8,9. Предполагается, что в формировании структуры этого огромного комплекса участвует неферментный компонент CspP. Тем не менее, не так много известно о структуре этого комплекса и о том, как он собран. Исследованные целлюлосомы, полученные из других бактерий, также содержат белок скаффолдин, которые чаще всего имеют другую архитектуру.
В геномной последовательности11 бактерии Clostridium thermocellum с депозитным номером АТСС 2740512 было идентифицировано 72 целлюлосомальных гена. Наиболее преобладающие целлюлосомальные компоненты были идентифицированы при помощи анализа протеом выделенных целлюлосом и при помощи мРНК анализа. Однако при этом не было ясно, какие компоненты являются необходимыми для расщепления целлюлозы, и какова их роль в формировании комплекса.
До сих пор была возможна только частичная реконструкция целлюлосомы путем создания небольших третичных комплексов мини-скаффолдина, объединенного с двумя ферментными компонентами, полученными рекомбинантным способом, и был продемонстрирован выраженный синергетический эффект14. Мутант, выделенный другой исследовательской группой, названный AD2, не абсорбировался целлюлозой, и не были охарактеризованы его молекулярный механизм действия, способность расщеплять целлюлозу и формировать целлюлосому15,16. Из мутировавших культур С. thermocellum были выделены мутанты, не образующие целлюлосому, которые не адсорбируются на кристаллической целлюлозе17.
Другие группы сконструировали основной каркас, например, из трех модулей дивергентного когезина для направленного эквимолярного связывания такого же количества разных ферментных компонентов (например, в WO 2010057064). Структура комплекса целлюлосомы собирается на поверхности дрожжевых клеток при помощи сконструированного “консорциума дрожжей”. В отличие от комплекса по изобретению, задействовано только нестатистическое (упорядоченное) связывание, при этом нельзя подобрать соотношение компонентов, как это возможно по настоящему изобретению.
В WO 2010012805 углеводсвязывающий модуль (СВМ) и X-модуль из целлюлосомального скаффолдина используют для обеспечения более эффективного продуцирования и секреции белков в рекомбинантном хозяине. В этом случае ген целлюлазы является генетически слитым с полипептидной цепью, в которой модули СВМ и X используются в качестве модулей “хелперов” для экспрессии, а не с целью получения оптимизированного расщепления целлюлозы.
В US20090035811 содержащие когезин белки и обладающие ферментативной активностью целлюлазы продуцируются in vivo дрожжевыми клетками и остаются прикрепленными к клеточной поверхности. Это приводит к повышенной плотности ферментных комплексов на относительно большой клеточной поверхности. Не показано, как можно манипулировать составом ферментных компонентов и их соотношением. Продуцирующий целлюлосому организм (дрожжи) нельзя с легкостью адаптировать к композиции, подходящей для другого субстрата.
В отличие от несвязанной с клеткой системы по настоящему изобретению, недостаток связанных с дрожжевыми клетками целлюлосом, описанных в области техники, заключается в том, что они связываются с одним специфичным продуктом в зависимости от организма, для которого разработаны. Их эффективность не может быть оптимизирована путем изменения соотношения компонентов. Кроме того, нативные целлюлосомы, такие как целлюлосомы, связанные с дрожжевыми клетками, нельзя получать в промышленном масштабе.
Все три способа не обеспечивают активность целлюлазы, превышающую активность нативной целлюлосомы, вероятно в связи с тем, что состав комплекса является не совсем оптимальным, и не может быть адаптирован к особенностям субстрата.
Тем не менее, необходимо изучить in vitro сборку целлюлосомы и ее единичных компонентов в отношении роли отдельных генов в процессе гидролиза целлюлозы и расщеплении волокон. Однако такие попытки до сих пор не увенчались успехом в связи с непреодолимыми трудностями, связанными с получением компонентов, не находящихся в своем нативном состоянии - прочная связь в комплексе препятствует легкому отделению при помощи мягких, неденатурирующих методов.
Ферментативное расщепление нерастворимого и кристаллического материала, такого как кристаллическая целлюлоза и гетерогенная гемицеллюлоза, по-прежнему является неэффективным, медленным и требует высокой концентрации ферментов, что делает применение в промышленном масштабе дорогостоящим и малоэффективным при использовании ферментных препаратов, существующих на сегодняшний день.
Поэтому целью настоящего изобретения является предоставление новых ферментных составов, способных к ферментативному расщеплению эластичных субстратов, таких как кристаллическая целлюлоза и гетерогенная гемицеллюлоза с более высокой эффективностью.
Другой целью настоящего изобретения является предоставление новых ферментных составов, а также эффективных и экономичных in vitro способов и применений этих ферментных составов, которые преодолевают недостатки ферментных смесей предшествующего уровня техники, в частности, необходимости для ферментативного расщепления более низких концентраций ферментов, увеличенной термостабильности ферментных составов, и увеличенной скорости связывания фермента.
Краткое описание изобретения
Согласно настоящему изобретению, сильное увеличение активности может быть достигнуто благодаря комплексу по изобретению, как описано более подробно ниже. Продемонстрировано повышение активности такой ферментной системы, ответственной либо за непрерывную цепь каталитических событий, либо за синергическое воздействие на эластичные субстраты, в том числе, без ограничения, кристаллическую целлюлозу и гетерогенную гемицеллюлозу. Авторы изобретения смогли показать, что реконструированные in vitro комплексы по изобретению проявляют более высокую активность в отношении кристаллической целлюлозы, чем нативные целлюлосомы, выделенные из бактерий.
Авторы настоящего изобретения выделили мутантов С. thermocellum, которые не формируют комплексы, и вместо этого секретируют нативные целлюлосомальные компоненты, не находящиеся в комплексе. Эти белки изначально использовали для реконструкции искусственной целлюлосомы, обладающей повышенной активностью. Используя этот мутант, может быть впервые исследована роль синергизма при помощи in vitro реконструкции комплексов.
Таким образом, изобретение предоставляет связанный с частицами или не содержащий частиц комплекс, включающий: (а) основной каркас, содержащий по меньшей мере четыре участка связывания, причем по меньшей мере два участка связывания обладают по существу одинаковой специфичностью связывания; и (b) ферментный компонент, связанный in vitro с каждым из указанных четырех участков связывания, где по меньшей мере три из указанных ферментных компонента являются разными ферментными компонентами. Молярное соотношение в указанном комплексе модулей когезина в основном каркасе (а) к сумме содержащих докерин ферментных компонентов (b) составляет 1:1. По меньшей мере, три ферментных компонента (b) в основном каркасе предпочтительно присутствуют в комплексе в молярном отношении друг к другу от 1:1 до 1:50, 1:1,5 до 1:30, предпочтительно от 1:1,8 до 1:15.
В предпочтительном варианте комплекс представляет собой комплекс без частиц или изолированный комплекс, который не связан с живой клеткой, особенно предпочтительно не связан с дрожжевой клеткой.
Предпочтительно, указанные ферментные компоненты случайным образом связываются in vitro с по меньшей мере четырьмя участками связывания.
Используемый здесь термин “in vitro” означает отделенный от живой клетки или организма.
Термин “комплекс” или “ферментный комплекс”, используемый здесь, означает координирование и объединение компонентов, связанных химическим или биологическим взаимодействием. Указанные комплексы могут быть связаны вместе, формируя комплекс более высокого порядка (также синонимично используемый здесь как “искусственная целлюлосома” или “целлюлазный комплекс”), состоящий из одного или нескольких содержащих когезин основных каркасов, предпочтительно содержащих когезин каркасных белков (обозначенных также здесь “мини-скаффолдин”), и одного или более содержащих докерин ферментных или неферментных компонентов, о чем более подробно описано ниже. Кроме того, искусственная целлюлосома состоит из одного или нескольких содержащих докерин основных каркасов, предпочтительно содержащих докерин каркасных белков (также обозначенных здесь “мини-скаффолдин”), и одного или более содержащих когезин ферментных или неферментных компонентов. Используемый здесь термин “смесь ферментов”, напротив, относится к растворимым ферментам, производимым в промышленном масштабе.
Используемый здесь термин “связанный с частицами” комплекс означает, что комплекс по изобретению связан с частицами, которые служат в качестве материала-носителя. Подходящими частицами являются, например, наночастицы. Наночастицы, используемые в этом методе, не превышают 2000 нм, предпочтительно, их средний диаметр составляет менее 100 нм. Они могут состоять из органического или неорганического вещества, такого как кремний, оксид металла, золото, полистирол и другие органические полимеры, и других веществ неживой природы или гибридов из разных веществ (такие, как наночастицы ядро-оболочка). Предпочтительно используются ферромагнитные наночастицы, которые обладают суперпарамагнитным поведением. Более предпочтительно их ядро покрыто полимерной оболочкой, такой как оболочка из полистирола, а поверхность частиц химически модифицирована для обеспечения возможности образования химических связей с биомолекулами. Предпочтительно модификации содержат свободные карбоксильные группы (СООН) или свободные аминогруппы (NH2) для реакций присоединения посредством сшивающих агентов для присоединения белков или химических молекул остова.
В качестве альтернативы, поверхности наночастиц, предпочтительно модифицированные аминными или карбоксильными функциональными группами, могут быть ковалентно сшиты предпочтительно с гетеробифункциональной молекулой, такой как полиэтиленгликолевый линкер, и, наконец, с нитрилотриуксусной кислотой (NTA) посредством глутаральдегида (аминная модификация) или EDC/сульфо-NHS (карбоксильная модификация), соответственно, согласно уровню техники. Молекулы мини-скаффолдина остова прикрепляются к остаткам NTA путем их слияния с полигистидином на концах белков (предпочтительно маркированных 6xHis) методом аффинного связывания с никелем, известным в уровне техники.
Наночастицы, покрытые основными каркасами, смешивают с ферментами, как описано при реконструкции комплексов.
Используемый здесь термин “без частиц” означает комплекс по изобретению, который не связан с клеткой или изолирован, соответственно.
Используемый здесь термин “основной каркас” относится к субстрату, используемому в качестве остова для комплекса, который предоставляет участки, подходящие для связывания ферментативных или неферментативных белковых компонентов. Остов (основание) может быть белковым остовом, каркасным остовом (скаффолдином) или полимерной органической молекулой с множеством участков связывания. Каркасный остов может состоять из одного или нескольких мини-скаффолдинов.
Термин “имеющий по существу такую же специфичность связывания”, используемый в отношении участков, связывающих ферментные компоненты, относится к специфичности связывания между модулями когезина и докерина такой, что друг с другом связываются только пары когезин-докерин, обладающие идентичной специфичностью связывания. Это можно проверить, например, путем смешивания пары белков и оценки показателя пробега в гель-электрофорезе в нативном состоянии.
В одном из вариантов изобретение относится к комплексу, согласно данному здесь определению, в котором основной каркас является линейным. Линейный основной каркас может быть синтетическим или биологическим. Синтетический основной каркас может быть, например, синтетическим полимерным носителем или линейным органическим полимером с функциональными группами, способными связывать белки. Белки могут быть ферментами, которые должны быть включены в комплекс, или белками, содержащими один или несколько модулей, участвующих во взаимодействии когезин-докерин (которые связывают ферментные компоненты). Биологический основной каркас может быть белком, содержащим встречающиеся в природе участки связывания (когезины) для докеринов, слитых с ферментными компонентами или связывающими модулями естественным образом или методами генной инженерии.
В другом варианте осуществления изобретения ферментные компоненты связаны с линейным основным каркасом посредством взаимодействия когезин-докерин. В предпочтительном варианте осуществления изобретения основной каркас комплекса по изобретению имеет по меньшей мере четыре участка связывания когезина для докеринов.
Предпочтительно остов комплекса по изобретению состоит из одного или нескольких белков, где один или несколько белков представляют собой белки остова, которые связаны между собой посредством химического взаимодействия или взаимодействия когезин-докерин, специфичность связывания которого отличается от таковой для взаимодействия ферментов с остовом, причем специфичность связывания для взаимодействия с образованием перекрестных связей отличается от специфичности связывания ферментов. Более предпочтительно, один или несколько белков сшиты посредством взаимодействия когезин-докерин, специфичность связывания которого отличается от специфичности связывания взаимодействия когезин-докерин, связывающего ферментные компоненты.
Используемый здесь термин “взаимодействие когезин-докерин” относится к взаимодействию между когезином и докерином. Докерин представляет собой белковый модуль, обнаруженный в компонентах целлюлосомы, предпочтительно, один модуль в каждом ферментном компоненте целлюлосомы. Партнером по связыванию докерина является модуль когезина, который, как правило, представляет собой повторяющуюся модульную часть белка основного каркаса в целлюлосоме. Это взаимодействие имеет важное значение для конструирования комплекса целлюлосомы. Такая же система когезин-докерин в комплексе по изобретению используется для связывания различных ферментных компонентов в комплекс. Один или несколько белков основного каркаса по изобретению могут быть сшиты посредством взаимодействия когезин-докерин, при котором пара когезин-докерин имеет специфичность связывания, отличающуюся от таковой для пары когезин-докерин, используемой для связывания ферментных компонентов. Связывание компонентов в полисахарид, наряду с другими методами, может быть определено по замедлению миграции полос белка в гель-электрофорезе в нативном состоянии или путем измерения количества белков после разделения жидкой фракции и фракции твердых частиц, содержащей частицы целлюлозы, стандартным методом для определения концентрации белка, известным специалисту в данной области.
В другом варианте основной каркас комплекса по изобретению получают, используя некаталитический каркасный белок, выделенный из разлагающих клетчатку микроорганизмов, формирующих целлюлосому, или их генетически модифицированных производных.
Используемый здесь “целлюлолитический микроорганизм, формирующий целлюлосому” относится к таким бактериям и грибам, которые формируют внеклеточные комплексы (называемые целлюлосомами), в которых ферменты связаны с основным каркасом посредством взаимодействия когезин-докерин. Далее, целлюлолитические микроорганизмы, формирующие целлюлосому, которые могут быть использованы в настоящем изобретении, представляют собой: бактерии, такие как Acetivibrio cellulolyticus, Bacterioides cellulosolvens, Butyrivibrio fibrisolvens, Clostridium acetobutylicum, С. cellulolyticum, С. cellulovorans, С. cellobioparum, С. josui, С. papyrosolvens, С. thermocellum, С. sp. C7, С. sp. P-1, Fibrobacter succinogenes, Ruminococcus albus, Р. flavefaciens, и грибковых микроорганизмов, таких как Piromyces sp. E2.
В другом варианте осуществления основной каркас получают из некаталитического каркасного белка CipA из Clostridium thermocellum или его генетически модифицированных производных.
Используемый здесь термин “генетически модифицированные производные” означает, что белок основного каркаса комплекса по изобретению является генетически модифицированным, например, основной каркас является генетически модифицированным производным, полученным из CipA-белка С. thermocellum или основной каркас является генетически слитым с модулем докерина или His-маркированной последовательностью, или изменено количество или порядок встречающихся в природе модулей (в CipA), или изменена нуклеотидная последовательность путем введения или удаления сайтов рестрикции, адаптации использования кодонов или изменения аминокислотных остатков в определенных позициях.
В предпочтительном варианте основной каркас комплекса по изобретению содержит УСМ-c1-c1-d3, представленный в SEQ ID NO:24, c3-с1-c1-d2, представленный в SEQ ID NO:22, c2-c1-с1, представленный в SEQ ID NO:26, или их производные, содержащие в своих модулях когезина аминокислотную последовательность с более 60% идентичностью.
Термин “идентичность последовательности”, известный специалистам в данной области определяет степень родства между двумя или более нуклеотидными или полипептидными молекулами, для которых определяют сходство их последовательностей. Процент “идентичности” определяется процентом идентичных областей двух или более последовательностей с учетом зазоров или других особенностей этих последовательностей.
Идентичность между родственными полипептидами может быть определена известными методами. Как правило, используются специальные компьютерные программы с алгоритмами, в которых учитываются специальные требования. Согласно предпочтительным процедурам определения идентичности сначала генерируют наибольшее согласие между изучаемыми последовательностями. Компьютерные программы для определения гомологии между двумя последовательностями, включают, без ограничений, программный пакет GCG, в том числе GAP (Devereux J et al., (1984); Genetics Computer Group University of Wisconsin, Madison (WI); BLASTP, BLASTN и FASTA (Altschul S et al., (1990)). Программу BLAST X можно приобрести в Национальном центре биотехнологической информации (NCBI) и через другие источники (BLAST Handbook, Altschul S et al., NCB NLM NIH Bethesda MD 20894; Altschul S et al.,1990). Для определения идентичности последовательностей также может быть использован хорошо известный алгоритм Смита Ватермана (Smith Waterman).
Предпочтительные параметры для сравнения последовательностей включают следующие:
Алгоритм: Needleman S.B. and Wunsch, C.D. (1970)
Матрица сравнения: BLOSUM62 от Henikoff S. и Henikoff J.G. (1992).
Штраф за пропуск в последовательностях: штраф при длине пропуска 12: 2
Программа GAP также подходит для использования с указанными выше параметрами. Приведенные выше параметры являются стандартными параметрами (параметрами по умолчанию) для сравнения аминокислотных последовательностей, в которых пропуски на концах не уменьшают значение идентичности. При сравнении очень коротких последовательностей с эталонной последовательностью, может возникнуть необходимость увеличить ожидаемое значение до 100000, а в некоторых случаях уменьшить длину слова (размера слова) до 2.
Также можно использовать модельные алгоритмы, штрафы за открытие пропусков, штрафы за удлинение пропусков и сравнение матриц, в том числе указанных в Program Handbook, Wisconsin Package, версия 9, сентябрь 1997. Выбор будет зависеть от проводимого сравнения, а также от того, будет ли это сравнение проводиться между парами последовательностей, для которого предпочтительными являются программы GAP или Best Fit, или между одной последовательностью и большой базой данных последовательностей, для которого предпочтительными являются программы FASTA или BLAST. Согласие 60%, определенное при помощи указанных алгоритмов, описывается как 60% идентичность. То же относится к более высокой степени идентичности.
В предпочтительных вариантах осуществления изобретения производные имеют более 60% идентичность аминокислотной последовательности, предпочтительно более 70%, более предпочтительно более 80% или 90% идентичность аминокислотной последовательности в их модулях когезина.
В другом варианте осуществления изобретения основной каркас комплекса по изобретению содержит углеводсвязывающий модуль (CBM). Предпочтительно, углеводсвязывающий модуль представляет собой углеводсвязывающий модуль семейства CBM3 согласно классификации, приведенной в базе данных CAZy (http://www.cazv.orq/Carbohvdrate-Bindinq-Modules.html) для гена CipA Clostridium thermocellum, который интегрирован или прикреплен к линейному основному каркасу.
Используемый здесь термин “углеводсвязывающий модуль (СВМ)” относится к непрерывной аминокислотной последовательности, обладающей углеводсвязывающей активностью. CBM могут либо вводиться в комплекс (“мини-скаффолдины”) по изобретению, либо CBM присутствует в ферментном компоненте, либо, в качестве альтернативы, он может быть генетически связан с мини-скаффолдинами посредством слияния с белковым компонентом. Разные CBM могут распознавать разные полисахариды или полисахаридные структуры. CBM также может быть связан с мини-скаффолдинами в результате генетической модификации или химической реакции с функциональной группой основного каркаса. CBM вызывает “эффект ориентации”, т.е. усиливает связывание между комплексом и субстратом, что является особенно выгодным для нерастворимых субстратов. CBM определены как дискретно сложенный некаталитический полипептидный модуль, связывающийся с полисахаридом или комплексным углеводом. Они могут быть обнаружены или созданы методами генной инженерии с целью слияния по модульному принципу с ферментами или каркасными белками, или в виде генетического слияния, связанного с основным каркасом посредством взаимодействия когезин-докерин или другими средствами. В предпочтительном варианте они связаны с кристаллической целлюлозой и принадлежат к семейству 3 СВМ (CBM3). (См.: http://www.cazv.org/Carbohvdrate-Bindinq-Modules.html). Связывание с полисахаридом может быть определено, наряду с другими методами, по замедлению миграции белка в гель-электрофорезе в нативном состоянии, в котором полисахарид равномерно распределен в геле.
В другом варианте осуществления изобретение относится к комплексу, согласно данному здесь определению, в котором ферментный компонент содержит по меньшей мере модуль докерина и каталитический модуль фермента.
Термин “модуль” описывает отдельно складывающийся фрагмент в полипептиде, который, как в “Лего”, может быть использован для сборки белков с новыми характеристиками методами генной инженерии или в ходе естественной рекомбинации. Используемый здесь “каталитический модуль фермента” относится к белковому модулю, который обеспечивает каталитическую активность полипептида. Все ферменты целлюлосомы являются мультимодульными ферментами и состоят из каталитических и некаталитических модулей, по меньшей мере каталитического модуля и модуля докерина. Некаталитический модуль может быть докерином, когезином, CBM, гомологичным S-слою модулем или модулем с еще неизвестной функцией (часто называемым X-модулем).
В еще одном варианте осуществления изобретение относится к комплексу, согласно данному здесь определению, в котором ферментные компоненты выбирают из группы, состоящей из процессивных или непроцессивных эндо-β-1,4-глюканаз, процессивных экзо-β-1,4-глюканаз и гликозидаз из полисахаролитических или сахаролитических микроорганизмов или их генетически модифицированных производных.
Ферментные компоненты, объединенные в комплекс по изобретению, включают помимо прочего целлюлазы из целлюлосомы Clostridium thermocellum, например, компоненты CbhA, CelA, CelE, CelJ, CelK, CeIR, CelS или CelT термофильных бактерий Clostridium thermocellum, или термостабильных β-гликозидаз, например, β-глюкозидазы BgIB из термофильных бактерий Thermotoga neapolitana.
Заменяя на другие виды активности, удаляя или добавляя ферментные компоненты или изменяя их молярное соотношение можно расширять спектр и усиливать активность комплексов по отношению к другим субстратам. Новые компоненты могут включать β-глюкозидазы, гемицеллюлазы (ксиланазы, маннаназы, арабинофуранозидазы, глюкуронидазы, ксилан-эстеразы и др.), пектиназы, пектин-лиазы, амилазы и другие ферменты для гидролиза лигноцеллюлозной биомассы, другие полисахариды или комбинацию ферментов для биохимического пути синтеза.
Используемые здесь “полисахаролитические микроорганизмы” относятся к гидролитическим микроорганизмам, способным разлагать полисахариды, такие как амилолитические, пектинолитические, целлюлолитические или гемицеллюлолитические микроорганизмы. Используемые здесь “сахаролитические микроорганизмы” относятся к микроорганизмам, использующим углеводы в качестве основного источника углерода и энергии.
В еще одном варианте осуществления изобретение относится к комплексу, согласно данному здесь определению, в котором по меньшей мере три ферментных компонента выбирают из группы, состоящей из целлюлолитических и гемицеллюлолитических ферментов, полученных из других микроорганизмов.
Примерами полисахаридов являются ацетан, агар-агар, альгинат, амилопектин, арабинан, арабиногалактан, арабиноксилан, карбоксиметилцеллюлоза, целлюлоза, хитин, хитозан, хризоламинарин, курдлан, циклозофоран, декстран, декстрин, эмульсан, фруктан, галактан, галактоманнан, геллан, α-глюкан, β-глюкан, глюкуронан, глюкуроноксилан, гликоген, N-ацетил-гепарозан, гидроксиэтилцеллюлоза, индикан, инулин, кефиран, ламинарин, лентинан, леван, лихенин, лихенан, люпин, маннан, пахиман, пектиновый галактан, пектин, пентозан, плеуран, полигалактуроновая кислота, пуллулан, рамногалактуронан, шизофиллан, склероглюкан, крахмал, сукциногликан, велан, ксантан, ксилоглюкан, зимозан.
Примерами целлюлолитических микроорганизмов являются бактерии, такие как Acetivibrio cellulolyticus, A. cellulosolvens, Anaerocellum thermophilum, Bacteroides cellulosolvens, Butyrivibrio fibrisolvens, Caldicellulosiruptor saccharolyticus, Cs. lactoaceticus, Cs. kristjansonii, Clostridium acetobutylicum, С. aldrichii, С. celerescens, С. cellobioparum, С. cellulofermentas, С. cellulolyticum, С. cellulosi, С. cellulovorans, С. chartatabidum, С. herbivorans, С. hungatei, С. josui, С. papyrosolvens, С. sp. C7, С. sp. P-1, С. stercorarium, С. thermocellum, С. thermocopriae, С. thermopapyrolyticum, Fibrobacter succinogenes, Eubacterium cellulolyticum, Ruminococcus albus, R. flavefaciens, R. succinogenes, Achromobacter piechaudii, Actinoplanes aurantiaca, Bacillus circulans, Bacillus megaterium, Bacillus pumilus, Caldibacillus cellulovorans, Cellulomonas biazotea, Cm. cartae, Cm. cellasea, Cm. cellulans, Cm. fimi, Cm. flavigena, Cm. gelida, Cm. iranensis, Cm. persica, Cm. uda, Cellvibrio fulvus, Cv. Gilvus, Cv. Mixtus, Cv. vulgaris, Curtobacterium falcumfaciens, Cytophaga sp., Flavobacterium johnsoniae, Microbispora bispora, Micromonospora melonosporea, Myxobacter sp. AL-1, Pseudomonas fluorescens, Ps. mendocina, Streptomyces alboguseolus, Sm. antibioticus, Sm. aureofaciens, Sm. cellulolyticus, Sm. flavogriseus, Sm. lividans, Sm. nitrosporeus, Sm. olivochromogenes, Sm. reticuli, Sm. rochei, Sm. thermovulgaris, Sm. viridosporus, Sporocytophaga myxcoccoides, Thermoactinomyces sp. XY, Thermobifida alba, Tb. cellulolytica, Tb. fusca, Thermonospora curvata, Xanthomonas sp.; грибов, таких как Anaeromyces mucronatus, Aspergillus niger, Caesomyces comunis, Cyllamyces aberensis, Hypocrea sp., Neocallimastix frontalis, Orpinomyces sp., Phanerochaete chrysosporium, Piptoporus cinnabarinus, Piromyces sp., Piromyces equi, Piromyces sp. E2, Rhizopus stolonifer, Serpula lacrymans, Sporotrichum pulverulentum, Trichoderma (=Hypocrea) harzianum, T. koningii, T. longibrachiatum, T. pseudokoninii, T. reesei, T. viride.
В еще одном варианте комплекс по изобретению содержит по меньшей мере три ферментных компонента, полученных из содержащих докерин компонентов Clostridium thermocellum или компонентов Thermotoga mahtima, имеющих слитый с ними докерин. Комплекс по изобретению также может включать содержащие докерин ферментные компоненты или ферментные компоненты, имеющие слитый с ними докерин, полученные из других бактерий.
В предпочтительном варианте, ферментные компоненты содержат CelK-d1, представленный в SEQ ID NO:8, CelR-d1, представленный в SEQ ID NO:10, CelT-d1, представленный в SEQ ID NO:14, CelE-d1, представленный в SEQ ID NO:16, CelS-d1, представленный в SEQ ID NO:6 и BglB-d1, представленный в SEQ ID NO:4, или их производные или родственные гены, полученные из других бактерий, имеющие более чем 50%, предпочтительно более 60%, более чем 70%, более предпочтительно более чем 80%, более предпочтительно более чем 90%, и наиболее предпочтительно более чем 97% идентичность аминокислотной последовательности.
В другом варианте комплекс по изобретению включает основной каркас, содержащий белки CBM-c1-c1-d3, представленный в SEQ ID NO:24, c3-c1-c1-d2, представленный в SEQ ID NO:22, c2-c1-c1, представленный в SEQ ID NO:26, и ферментные компоненты, содержащие CelK-d1, представленный в SEQ ID NO:8, CelR-d1, представленный в SEQ ID NO:10, CelT-d1, представленный в SEQ ID NO:14, CelE-d1, представленный в SEQ ID NO:16, CelS-d1, представленный в SEQ ID NO:6, и BglB-d1, представленный в SEQ ID NO:4.
Кроме того, изобретение предоставляет способ получения комплекса, согласно приведенному здесь определению, содержащий стадии:
а) получения рекомбинантным способом по меньшей мере трех ферментных компонентов, согласно приведенному здесь определению;
b) получения рекомбинантным способом основного каркаса по любому из пп.1-8;
с) смешивания очищенных, частично очищенных или неочищенных компонентов а) и b) in vitro; и
d) связывания случайным образом ферментных компонентов с основным каркасом.
В другом варианте осуществления способ получения комплекса, согласно приведенному здесь определению, содержит стадию связывания полученного рекомбинантным способом основного каркаса или ферментных частиц с частицей-носителем. Подходящие частицы включают наночастицы, согласно приведенному здесь определению, предпочтительно наночастицы, покрытые полистиролом, такие как покрытые полистиролом ферромагнитные наночастицы или химически функционализированные суперпарамагнитные наночастицы и другие мелкие наночастицы, предпочтительно имеющие диаметр от 10 до 2000 нм, более предпочтительно от 30 до 250 нм, с карбоксильными или аминогруппами, прикрепленными к поверхности. Для связывания основных каркасов или ферментов к поверхности частиц используются химические реакции присоединения, известные в данной области техники.
Ферментные компоненты могут быть получены рекомбинантным способом путем клонирования генов и использования методов модификации, хорошо известных в данной области, например, ферментные компоненты, полученные методом генной инженерии, могут быть слиты с модулями докерина, когезина и/или другими некаталитическими модулями, необязательно, с последующей белковой инженерией этих компонентов для увеличения продуцирования рекомбинанта, например, путем оптимизации сигналов секреции, изменения белковых сегментов, уменьшающих успешную экспрессию или секрецию, или использования кодонов. На следующей стадии рекомбинантным способом были получены основные каркасы (“мини-скаффолдины”) по изобретению, необязательно слитые с модулями когезина и объединенные случайным образом для формирования комплекса путем смешивания ферментных компонентов с основными каркасами. Компоненты могут быть очищенными, частично очищенными или неочищенными, предпочтительно частично очищенными или неочищенными. Предпочтительно молярное отношение модулей когезина и докерина в смеси фермент-основной каркас составляет 1:1. По меньшей мере три ферментных компонента смешивают вместе in vitro в молярном отношении от 1:1 до 1:50, от 1:1,5 до 1:30, предпочтительно от 1:1,8 до 1:15.
Таким образом, ферментные компоненты случайным образом связываются с основным каркасом. Модули докерина и когезина взаимодействия когезин-докерин, согласно приведенному здесь определению, являются взаимозаменяемыми, что означает, что модули когезина могут присутствовать либо в основном каркасе, а модули докерина в ферментном компоненте, либо наоборот, модули когезина могут присутствовать в ферментном компоненте, а модули докерина - в основном каркасе.
Предпочтительно, по меньшей мере 3 из целлюлосомальных компонентов, например, компонентов CbhA, CelA, CelE, CelJ, CelK, CelR, CelS или CelT термофильных бактерий Clostridium thermocellum, или термостабильных β-гликозидаз, например, β-глюкозидазы BglB, полученной из термофильных бактерий Thermotoga neapolitana, объединят на молекуле остова, предпочтительно в молярном отношении от 0,05 до 1,5 частей каждого из BglB, CbhA, CelE, CelJ или CelT; в молярном отношении от 0,1 до 3,0 частей каждого из CelK и CelR, и в молярном отношении от 0,2 до 6,0 каждого из CelA и CelS. В другом варианте осуществления молярное отношение составляет от 0,1 до 1,0 частей для каждого из BglB, CbhA, CelE, CelJ или CelT, при молярном отношении от 0,2 до 1,0 частей каждого из CelK и CelR и молярном отношении от 0,5 до 1,0 для каждого из CelA и CelS. В наиболее предпочтительном варианте осуществления молярное отношение составляет от 0,06 до 0,6 частей для каждого из BglB, CbhA, CelE, CelJ или CelT, при молярном отношении от 0,1 до 1,8 частей каждого из CelK и CelR и молярном отношении от 0,3 до 2,0 каждого из CelA и CelS.
В другом варианте осуществления изобретение относится к способу получения комплекса по изобретению, согласно приведенному здесь определению, в котором общее количество основных каркасов на стадии с) и общее количество ферментных компонентов смешивают вместе в молярном отношении 1 модуль когезина на 1 ферментный компонент, и по меньшей мере три ферментных компонента смешивают вместе in vitro в молярном отношении от 1:1 до 1:50, от 1:1,5 до 1:30, предпочтительно от 1:1,8 до 1:15, предпочтительно от 1:1 до 1:15.
В другом варианте осуществления изобретение относится к комплексу, полученному описанным здесь способом.
“Молярное отношение” в той мере, в какой оно относится к отношению основных каркасов к ферментным компонентам, вычисляют в виде молярного отношения, при котором 1 участок связывания предпочтительно модуля когезина, который содержится в основном каркасе соответствует 1 участку связывания предпочтительно модуля докерина, который содержится в ферментном компоненте.
В другом варианте осуществления изобретение относится к способу ферментативного гидролиза полисахаридных субстратов, содержащему стадии:
а) смешивания комплекса по изобретению с нерастворимой целлюлозой; и необязательно
b) выделения продуктов распада.
Смешивание комплекса по изобретению с нерастворимой целлюлозой предпочтительно проводят в водной среде при оптимальных или почти оптимальных рН и температуре. Оптимальный или почти оптимальный рН равен 6,5±0,5. Оптимальная температура находится в диапазоне 25-65°С, предпочтительно 30-65°C, наиболее предпочтительно около 55°C.
В другом варианте осуществления изобретение относится к применению комплекса по изобретению для ферментативного гидролиза полисахаридных субстратов.
В предпочтительном варианте осуществления изобретения, полисахаридный субстрат, как описано здесь выше, представляет собой кристаллическую целлюлозу или субстрат, содержащий кристаллическую целлюлозу.
Краткое описание чертежей
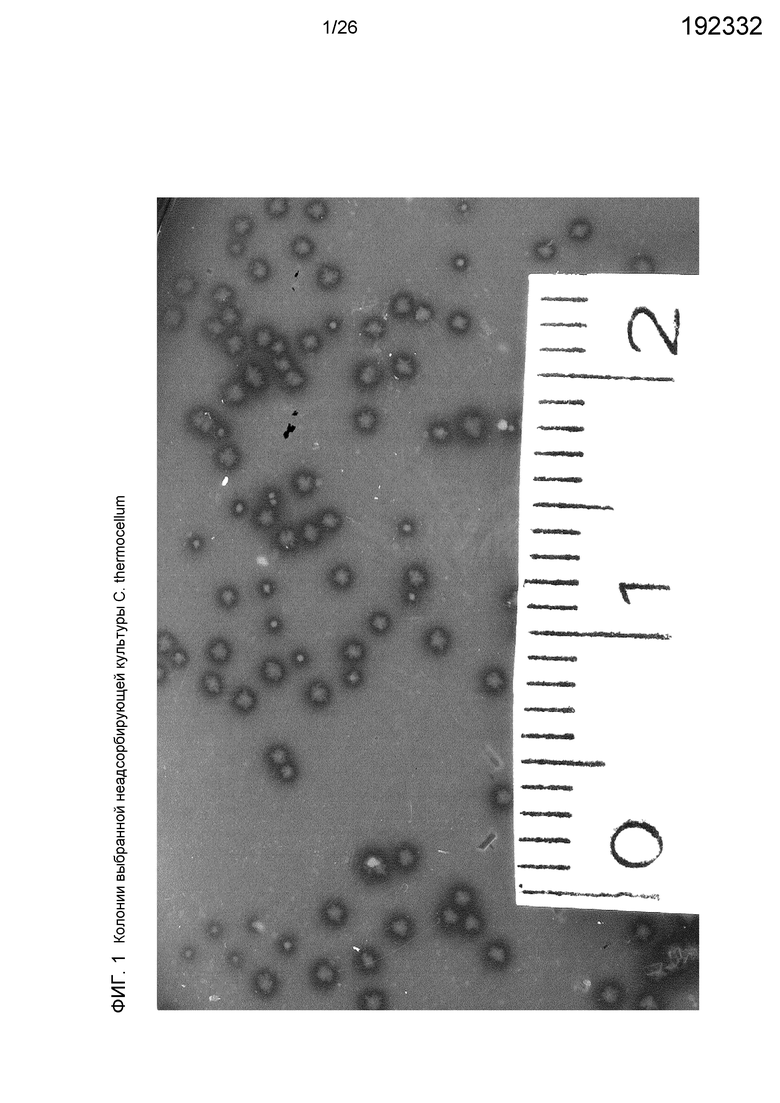
На фиг.1 показаны колонии выбранной неадсорбирующей культуры С. thermocellum. На мутной агаровой поверхности видны колонии, окруженные темным (=осветленным) гало гидролизованной целлюлозы. На масштабной линейке показаны отметки, равные 1 и 2 см.
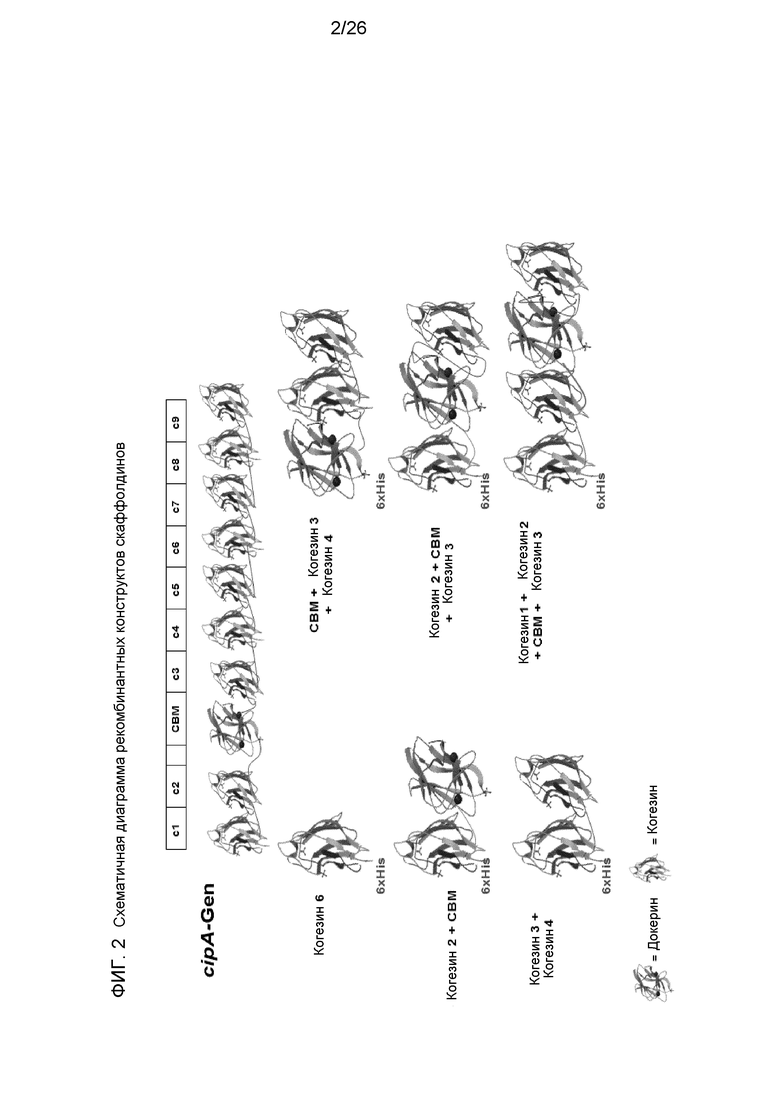
На фиг.2 показана аппроксимированная 3D-структура (с любезного разрешения H. Gilbert) рекомбинантных каркасных структур. Производные CipA-белка С. thermocellum (верхний ряд: с1 = когезин 1 и т.д.; CBM = углеводвязывающий модуль).
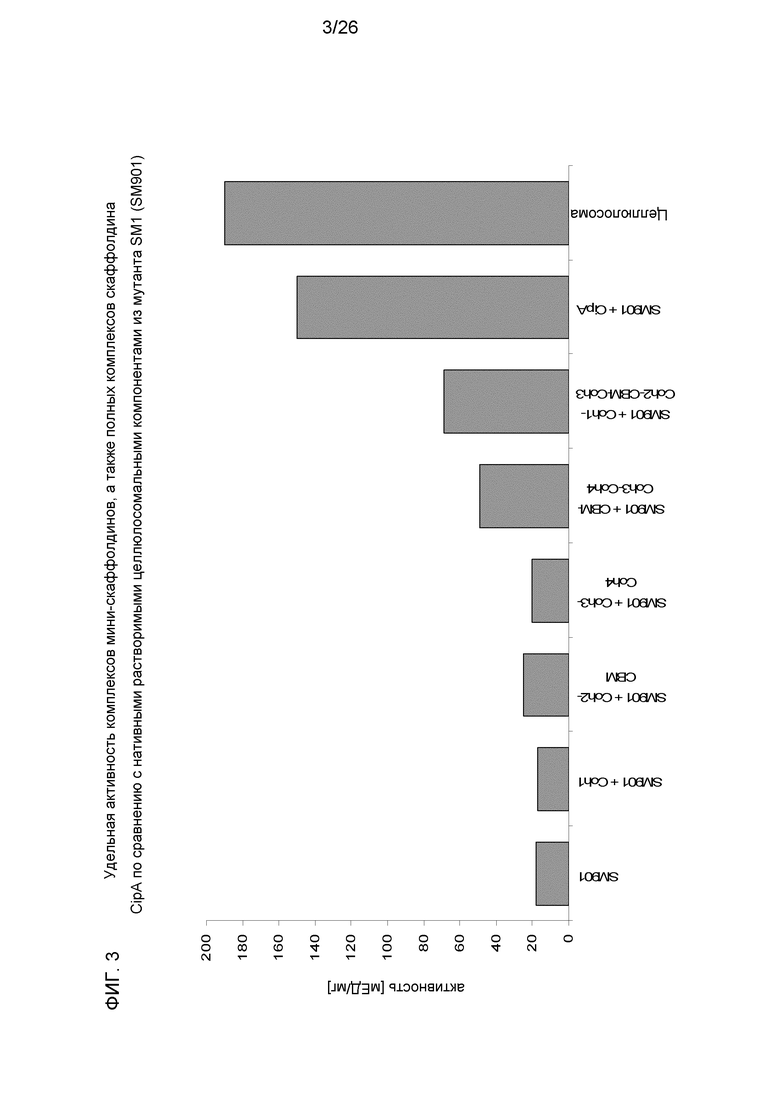
На фиг.3 показана удельная активность [мЕД/мг в эквивалентах глюкозы] комплекса с нативными растворимыми целлюлосомальными компонентами из мутантных SM1 (SM901) и мини-скаффолдинов, а также полного нативного скаффолдина CipA С. thermocellum. Контроль: нативная целлюлосома, полученная из С. thermocellum. Активность определяли на 0,5% Avicel (T=60°C, pH 6,0). (Coh: когезин, CBM: углеводсвязывающий модуль).
На фиг.4 показан комплекс наночастица-линкер-скаффолдин-целлюлаза (NLSC). Для простоты скаффолдин показан только с 2 когезинами (вместо 9). Для наночастиц и различных молекул масштаб не соблюден.
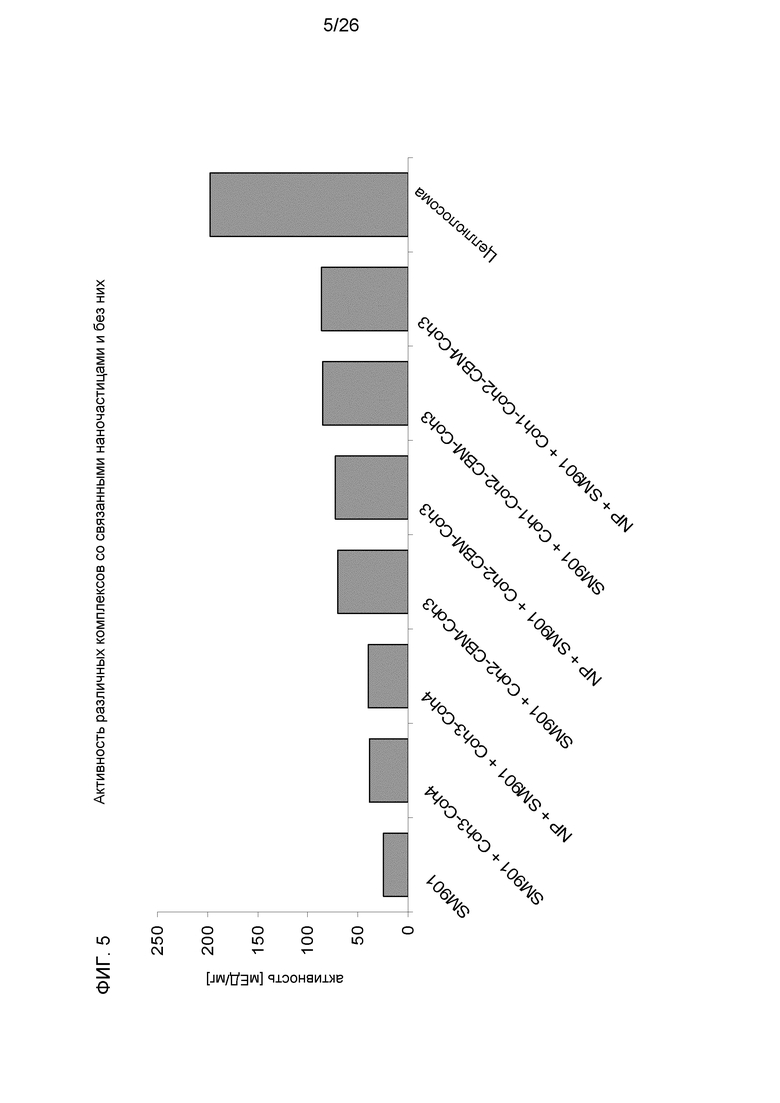
На фиг.5 показана активность различных комплексов со связанными наночастицами и без них. SM901: мутантные целлюлосомальные компоненты без скаффолдина.
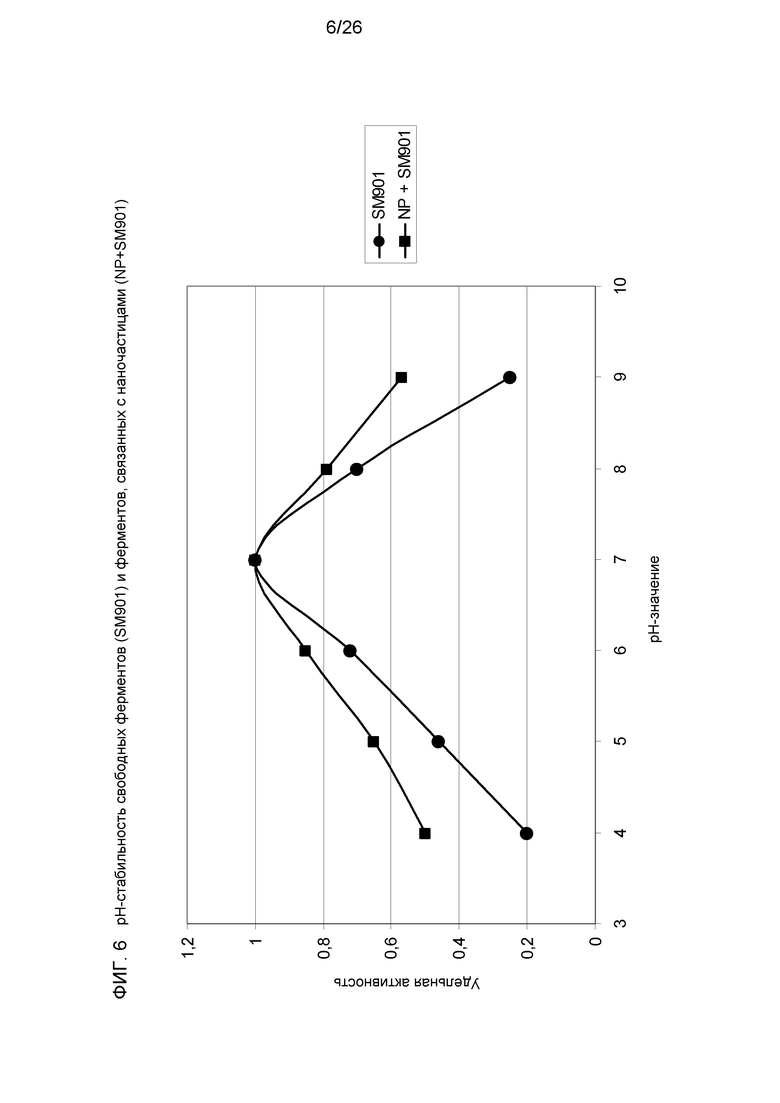
На фиг.6 приведено сравнение рН-стабильности свободных ферментов (SM901) и ферментов, связанных с наночастицами (NP+SM901).
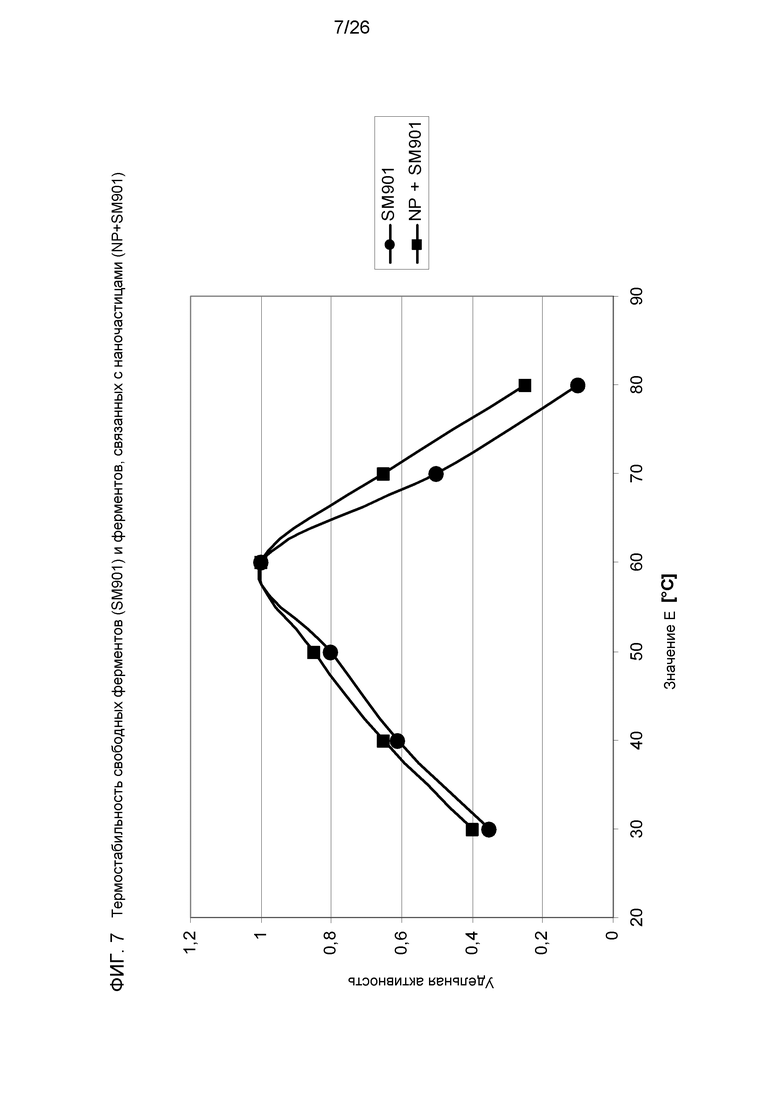
На фиг.7 приведено сравнение термостабильности свободных ферментов (SM901) и ферментов, связанных с наночастицами (NP+SM901).
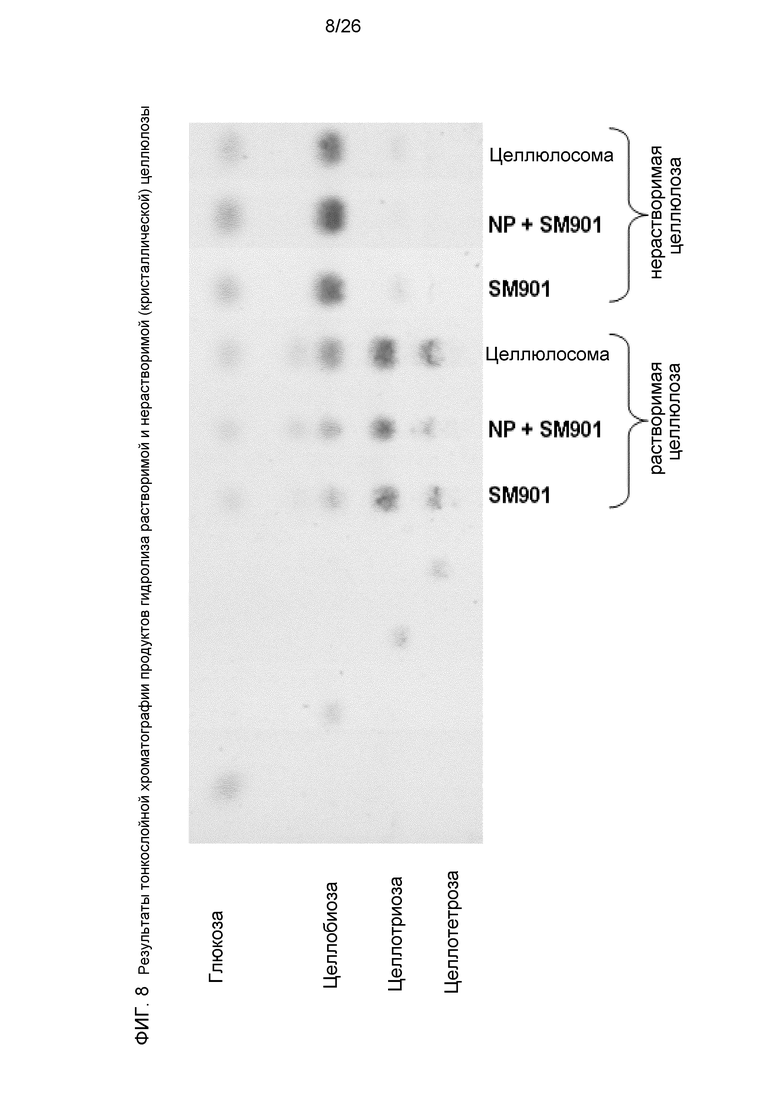
На Фиг.8 показаны результаты тонкослойной хроматографии продуктов гидролиза растворимой (СМС) и нерастворимой (кристаллической) целлюлозы (Avicel), использованной в качестве субстрата. Показано сравнение растворимых ферментов (SM901), искусственных комплексов и нативной целлюлосомы.
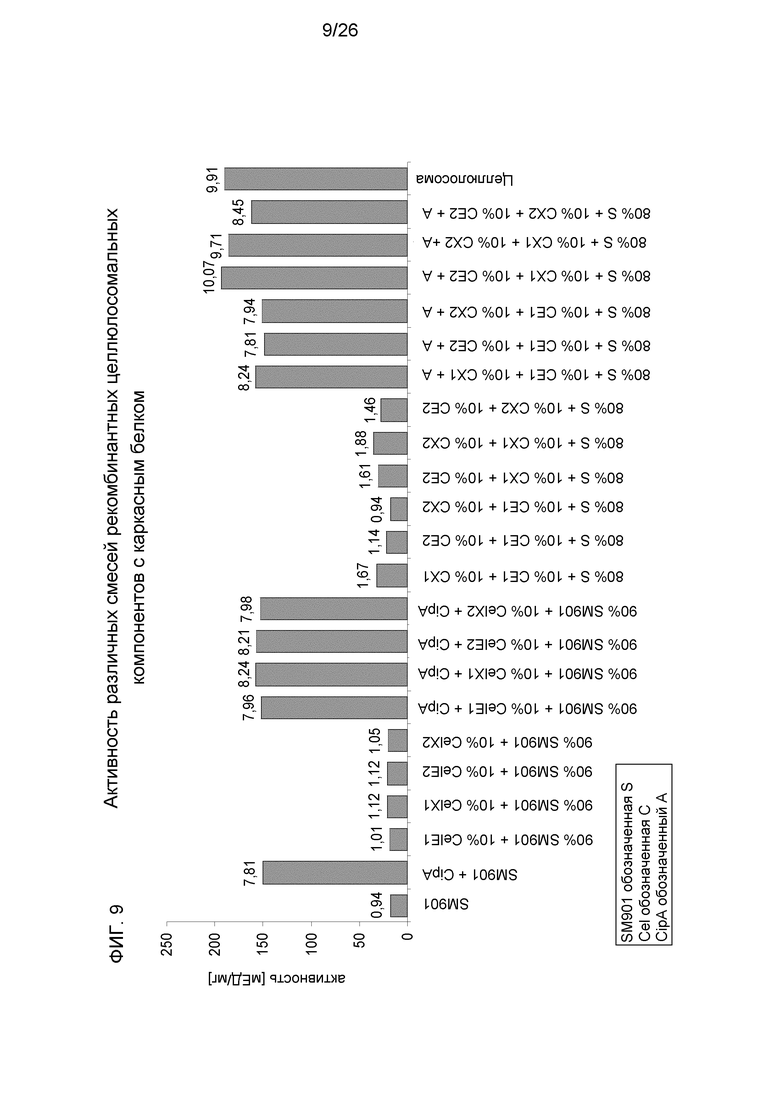
На фиг.9 показана активность различных смесей рекомбинантных целлюлосомальных компонентов с каркасным белком. В качестве субстрата используется кристаллическая целлюлоза.
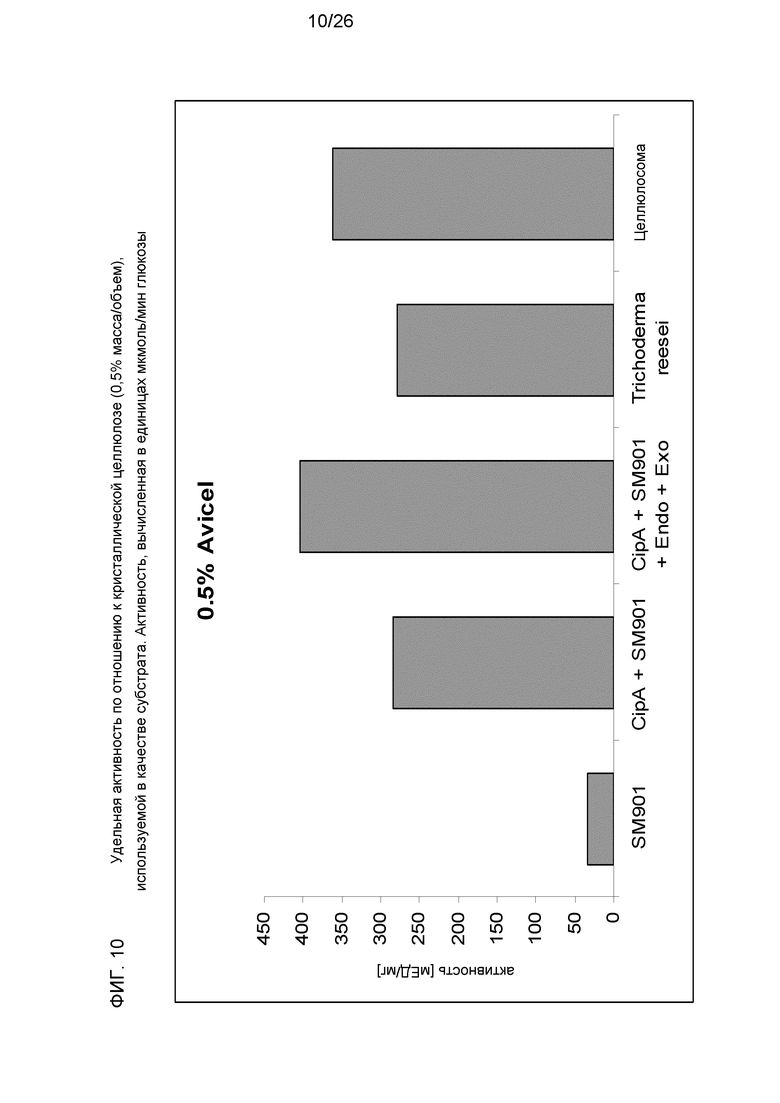
На фиг.10 показана удельная активность растворимых целлюлосомальных компонентов (SM901), тех же ферментов в виде комплекса (со скаффолдином), синтетической смеси с рекомбинантными компонентами (SM901 + CipA + Эндо + Экзо [NTC]), коммерчески доступного препарата на основе фермента Trichoderma reesei и нативной целлюлосомы из Clostridium thermocellum. Кристаллическая целлюлоза (0,5% масса/объем) использовалась в качестве субстрата. Активность вычисляли в единицах мкмоль/мин в эквивалентах глюкозы.
На фиг.11 показана нуклеотидная последовательность и аминокислотная последовательность основных каркасов CBM-c1-c1-d3, c3-c1-с1-d2, c2-c1-с1 и ферментных компонентов CelK-d1, CelR-d1, CelT-d1, CelE-d1, CelS-d1 и BglB-d1.
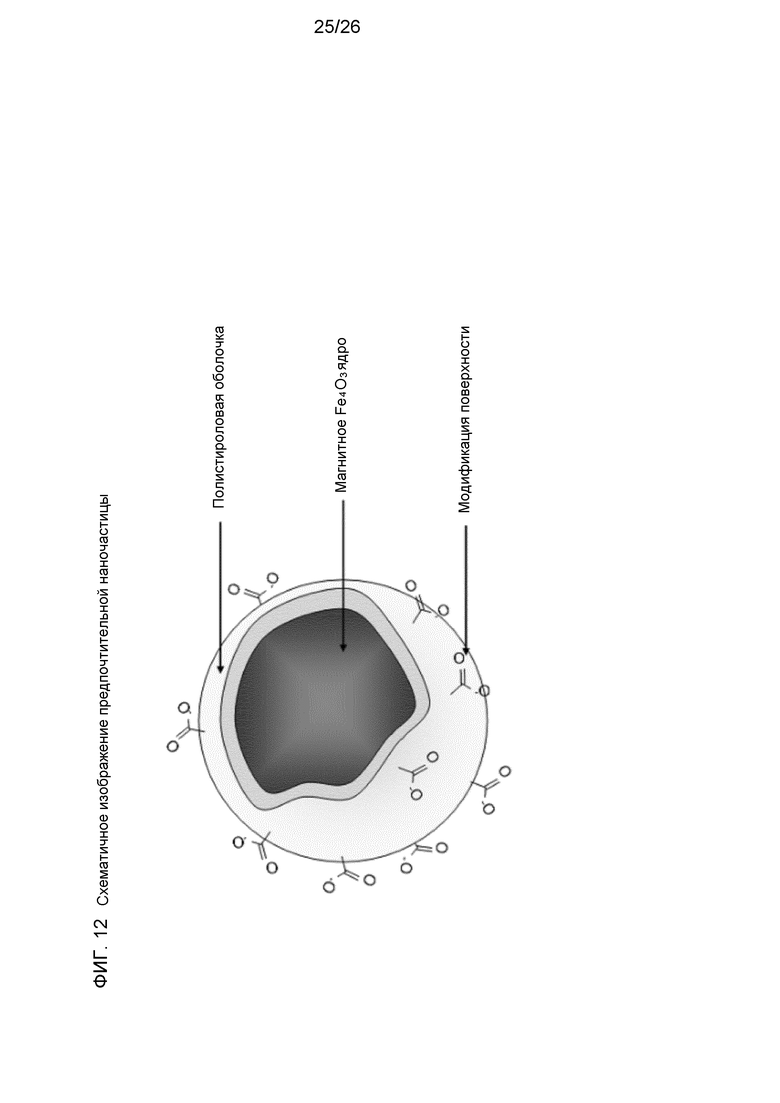
На фиг.12 схематично показана предпочтительная наночастица.
На фиг.13 показаны частицы, выделенные из раствора благодаря их суперпарамагнитным свойствам с использованием сильного дискового магнита. Реакционный раствор может быть легко удален со степенью извлечения из лунки преышающей 93%.
Примеры
Изобретение поясняется в приведенных ниже примерах, которые предназначены только для иллюстрации изобретения, и не должны рассматриваться как каким-либо образом ограничивающие изобретение.
В примерах показана гидролитическая деструкция целлюлозы с использованием системы, состоящей из 6 рекомбинантно экспрессированных целлюлаз, которые связаны с белковым носителем, основным каркасом, имитирующим скаффолдин CipA (интегрирующий целлюлосому белок). Основной каркас несет связывающие когезин модули, которые прочно и специфичным образом связаны с модулями докерина, формирующими С-конец ферментных компонентов. Это образование представляет собой собранный in vitro комплекс, сходный с целлюлосомой из термофильных анаэробных бактерий Clostridium thermocellum. Когезины и докерины связаны друг с другом случайным образом. Такие комплексы, при реконструкции in vitro при использовании правильных компонентов в правильном соотношении проявляют более высокую активность по отношению к кристаллической целлюлозе, чем нативные целлюлосомы, выделенные из этих бактерий.
Пример 1: Выделение мутанта, не формирующего целлюлосому
Из мутированных культур C. thermocellum были выделены мутанты (фиг.1). Шесть колоний с пониженной или отсутствующей способностью формировать прозрачное гало в целлюлозе вокруг колоний выбирали случайным образом. Один из мутантов C. thermocellum, SM1, полностью потерял способность продуцировать белок скаффолдин CipA или активный когезин. Ферменты от дикого типа (имеющего целлюлосомы) и мутант SM1 (без целлюлосом, без ферментов) тестировали по отношению к β-глюкану ячменя, CMC (оба в качестве контроля), а также микрокристаллической целлюлозе MN300. У обоих штаммов ферментативная активность по отношению к β-глюкану ячменя и CMC составила около 8,5 и 1,0 ЕД/мг белка, соответственно (табл. 1). Удельная активность по отношению к кристаллической целлюлозе, напротив, была существенно меньше у мутанта SM1, меньше в 15 раз, по сравнению с диким типом.
Мутант продуцировал целлюсомальные компоненты примерно в равных количествах по сравнению с диким типом, за исключением компонента CipA (скаффолдин CipA), который полностью отсутствовал. У мутанта полностью отсутствовали супрамолекулярные комплексы. Это указывает на неспособность мутанта правильно формировать целлюлосомы. В результате, около 50 целлюлосомальных белков оказались диспергируемыми, растворимыми, не находящимися в комплексе с белками, продуцированное количество и распределение которых было аналогично таковым, наблюдаемым для белков дикого типа.
Ферментативная активность концентрированных культуральных супернатантов мутанта SM1 и дикого типа (wt) по отношению к β-глюкану ячменя, CMC (оба растворимые) и целлюлозы MN300 (кристаллической)
Пример 2: Реконструкция целлюлосомы
А: Приготовление ферментных компонентов
Для реконструкции искусственной целлюлосомы был выбран мутант SM1 и мутантные супернатантные белки (SM901). Кроме того, были клонированы гены, кодирующие компоненты целлюлазы, и охарактеризованы их биохимические параметры, такие как рН и температурный оптимум, и активность по отношению к разным субстратам. Пять самых многообещающих ферментных компонентов с целлюлазной активностью отбирали согласно приведенным выше данным о составе целлюлосомы11. Кроме того, биохимически были охарактеризованы β-глюкозидазы, полученные из нескольких термофильных сахаролитических бактерий. β-глюкозидаза BglB из Thermotoga maritima была выбрана благодаря своей высокой термостойкости и высокой активности по отношению к целлодекстринам. Ген был слит с расположенным ниже модулем докерина, полученным из целлюлазы CelA С. thermocellum. Определяли оптимальные условия экспрессии. Ферменты, содержащие каталитические и некаталитические модули, включая модуль докерина, образованный таким образом, обозначены здесь CelK-d1, CelR-d1, CelT-dl, CelE-d1, CelS-d1 и BglB-d1. Аминокислотные последовательности CelK-d1, CelR-d1, CelT-d1, CelE-d1, CelS-d1 и BglB-d1 представлены в SEQ ID NO:4, 6, 8, 10, 14 и 16.
Для облегчения очистки они были клонированы с N-концевой His-меткой, с помощью которой возможна легкая очистка методом аффинной хроматографии. Для клонирования амплифицированных фрагментов ДНК в рамке считывания в пределах сайта рестрикции ниже промотор-операторной последовательности и 6xHis последовательности (например, pQE вектор из Quiagen) использовали методы уровня техники.
Баланс молярная стехиометрия компонентов поддерживался путем вычисления количества когезинов и докеринов.
B: Получение основного каркаса
Мини-скаффолдины, описанные ниже, были сконструированы путем объединения последовательностей когезина, докерина и CBM, полученных из гена CipA С. thermocellum, из целлюлосомальных компонентов C. thermocellum и С. josui. Последовательности оптимизировали для использования кодонов E.coli, т.е. самые редкие кодоны для почти неэкспрессируемых генов тРНК argU, ileY и leuW, которые узнают кодоны AGA/AGG, AUA и CUA, замещали синонимичными кодонами. Полученные таким образом последовательности были синтезированы и экспрессированы в плазмидных векторах E.coli, таких как pQE, способами, известными из уровня техники. В одном из вариантов осуществления использовали когезины 3-4 (тип I) и CBM3 из CipA, а также когезин c3 (тип II) и докерин d3 из olpB С. thermocellum, или c2 из cipA и d2 докерин из целлюлосомальных компонентов Clostridium josui. Использованные в примере основные каркасы обозначены здесь как CBM-c1-c1-d3, c3-c1-c1-d2, c2-c1-c1. Аминокислотные последовательности CBM-c1-c1-d3, c3-c1-c1-d2, c2-c1-c1 приведены в SEQ ID NO:22, 24 и 26.
C: Экспрессия, очистка и обогащение докерин-ферментных компонентов
Ферментные компоненты сначала были продуцированы мутантом SM1. Однако, С. thermocellum может продуцировать лишь ограниченное количество экзобелка из-за энергетических ограничений, связанных с анаэробным образом жизни. Даже улучшение штаммов не приводит к значительному увеличению количества экзобелков. Для замены смеси нативных ферментов из более, чем 30-ти компонентов на искусственную смесь целлюлаз, основные целлюлосомальные компоненты получали из рекомбинантного хозяина. Для простоты эксперимента в качестве хозяина использовали E.coli. Другие бактерии могут быть более подходящими благодаря меньшей себестоимости и высокой эффективности выхода рекомбинантных белков. Подходящим является любой промышленный штамм-производитель с высоким выходом данного белка.
Ферментные компоненты выделяли из рекомбинантного хозяина, и очищали аффинной хроматографией на His-tag колонках, или обогащали тепловой преципитацией белки E.coli, таким образом, рекомбинантные белки оставались в растворимой фазе. Тепловую преципитацию выполняли нагреванием раствора белков до 65°С в течение 10 мин, и осажденные белки E.coli удаляли центрифугированием способами, известными из уровня техники. Обогащение также было успешным в случае использования ультрафильтрации (отсечка 10 000 Да).
D: Образование комплекса ферментых компонентов с основным каркасом
Затем их связывали с рекомбинантными основными мини-каркасами, состоящими из различных когезинов с углеводсвязывающим модулем или без него, например, CBM-c1-c1. Комплекс может быть реконструирован из компонентов простым стехиометрическим добавлением к смеси ферментных компонентов (один несущий докерин компонент на один модуль когезина, присутствующий в смеси) в присутствии кальция, в одном из вариантов, 20 мМ CaCl2. Примерные структуры рекомбинантных конструкций мини-скаффолдина приведены на фиг.2. Образование комплекса происходит путем случайного объединения докерин-ферментных компонентов с мини-скаффолдинами, состоящими из различных когезинов, посредством взаимодействия когезин-докерин. Затем они были использованы для измерения эффекта комплексообразования либо с нативными ферментами, либо с ферментами, выделенными из рекомбинантных хозяев.
Пример 3: активность искусственного целлюлосомального комплекса
Смесь таких комплексов с СВМ и без него связывали через оптимизированную алифатическую молекулу-линкер с поверхностью полистирольных наночастиц. Такие структуры схематически показаны на фиг.3. Связывание различных комплексов мини-скаффолдина привело к увеличению активности гидролиза кристаллической целлюлозы, несмотря на стерические препятствия и некоторую потерю степей свободы ферментных компонентов в связи с плотным покрытием наночастиц (фиг.3 и 4). Кроме того, диапазон рН для ферментов увеличивается, если белки связаны с частицей (фиг.5). Это также верно и в отношении термостабильности целлюлаз (фиг.6). Оба эти результата являются важным преимуществом для технического применения.
Для проверки реализуемости такого подхода часть смеси компонентов SM901 заменяли одной или несколькими рекомбинантными целлюлазами. Несмотря на уменьшение в смеси компонентов SM901, в результате было показано, что при добавлении одного полученного рекомбинантным способом компонента активность комплекса по отношению к кристаллической целлюлозе слегка увеличивалась. При добавлении смеси рекомбинантных ферментов увеличение становилось существенным (фиг. 9). В некоторых смесях активность синтетических смесей была выше, чем активность нативной целлюлосомы. При полной замене и правильно сбалансированной стехиометрии синтетические комплексы демонстрировали еще более высокую активность.
Характер состава продуктов (глюкоза и целлодекстрины), полученных для растворимой, а также нерастворимой целлюлозы, был идентичным для свободных ферментов (SM901), искусственных комплексов и нативных целлюлосом (фиг.11). В случае использования в качестве субстрата нерастворимой целлюлозы, основным продуктом является целлобиоза с некоторым количеством глюкозы в качестве вторичного продукта. Целлобиоза должно быть дополнительно расщеплялась до глюкозы при добавлении к комплексу β-глюкозидазы. Ген β-глюкозидазы Thermotoga neapolitana, генетически слитый с модулем докерина, был успешно использован и увеличивал получение восстанавливающих сахаров примерно в 2 раза.
Таким образом, реконструкция целлюлосомы возможна. Следовательно, впервые была продемонстрирована возможность того, что, по-видимому, порядок расположения компонентов вдоль скаффолдина является случайным, и что in vitro активность реконструированной целлюлосомы по меньшей мере эквивалентна активности нативной целлюлосомы. Нативная целлюлосома не может производиться в промышленных масштабах.
Результаты
Результаты ясно показали, что разные когезины связывали компоненты одинаково и не различали целлюлосомальные компоненты, содержащие другие модули докерина. Связывание с когезином происходило случайным образом. Сборка между когезинами и докеринами была быстрой и спонтанной. После связывания компоненты прочно прикреплялись к скаффолдину. Увеличение числа когезинов в основном каркасе путем сшивки отдельных белков основного каркаса посредством взаимодействия когезин-докерин действительно приводило к увеличению активности по отношению к кристаллической целлюлозе (фиг.6). Далее, на фиг.6 показано, что добавление к комплексу СВМ определенного типа повышало активность (фиг.6). Кроме того, “полный” скаффолдин, образующий реконструированную целлюлосому, имел самую высокую активность, превышающую активность нативной целлюлосомы (еще до систематической оптимизации).
Пример 4: Необязательное связывание с наночастицами
Выбирали наночастицы со средним диаметром 0,110±0,007 мкм, ферромагнитным ядром с суперпарамагнитными свойствами и полистирольным покрытием, которые были химически функционализированы COOH-остатками на поверхности. Гетеробифункциональные молекулы-линкеры химически связывали с поверхностью, которая связывалась с выбранным основным каркасом; когезины каркаса можно было нагружать ферментами посредством белок-белковых взаимодействий с докеринами, прикрепленными к ферментным компонентам (нековалентное взаимодействие когезин-докерин). Схематический вид предпочтительной наночастицы показан на фиг.12.
Чтобы связать молекулы-линкеры с поверхностью наночастиц, активировали функциональные (свободные СООН-) группы. Растворимый в воде карбодиимид 1-этил-3-(3-диметиламинопропил)карбодиимида гидрохлорид (EDC) образует активные группы сложного эфира с карбоксильными группами при использовании водорастворимого соединения N-гидроксисульфосукцинимида (сульфо-NHS). EDC реагирует с карбоксилатной группой с образованием уходящей активной группы сложного эфира (O-ацилизомочевина). Сульфо-NHS сложные эфиры представляют собой активные гидрофильные группы, которые быстро реагируют с аминами на молекуле-мишени18. Однако в присутствии аминных нуклеофилов, которые могут атаковать карбонильную группу сложного эфира, сульфо-NHS группа быстро уходит, обеспечивая стабильную амидную связь с амином. Преимуществом добавления сульфо-NHS к EDC является повышение стабильности активного промежуточного соединения, которое в конечном итоге реагирует с атакующим амином. Продукт реакции EDC с карбоксилатными группами подвергается гидролизу в водном растворе в течение нескольких секунд. Образование промежуточного соединения сульфо-NHS сложного эфира в результате реакции гидроксильной группы на сульфо-NHS с комплексом активный EDC-сложный эфир увеличивает показатель полуактивности активированного карбоксилата19 до нескольких часов. Структура связанных молекул на поверхности наночастиц показана на фиг.4.
А. Активация
20 мг модифицированных карбоксильными группами наночастиц промывали три раза 2 мл буфера активации (50 мМ MES, 0,5 М NaCl, рН 6,0) в стеклянном флаконе путем отделения при помощи сильного NdFeB дискового магнита (1,41-1,45 Тл). Модифицированную поверхность частиц активировали добавлением свежеприготовленного раствора EDC и раствора сульфо-NHS до конечных концентраций 2 мМ и 5 мМ, соответственно. Реакцию смесей проводили в течение 15 минут при комнатной температуре.
B. Связывание с линкером
Частицы промывали два раза 2 мл реакционного буфера (0,1 M фосфат натрия, 0,5 М NaCl, рН 7,2) путем отделения от жидкости при помощи магнита. 5 мг О-(2-аминоэтил)-О-(2-карбоксиэтил)-полиэтиленгликоль 3000 гидрохлорида (NH2-PEG-COOH) растворяли в 100 мкл реакционного буфера в атмосфере азота и добавляли к активированным частицам. Ковалентная связь между активированными частицами и аминогруппами линкера на основе PEG образовывалась в пределах 3 часов при комнатной температуре. Буфер заменяли 2 мл буфера активации, и карбоксильную группу на конце ковалентно-связанного линкера активировали при помощи EDC и сульфо-NHS, как описано выше. После двух стадий промывки, используя 2 мл реакционного буфера, добавляли 10 мг Nα,Nα-бис(карбоксиметил)-L-лизин гидрата (NTA). Связывание NTA с активированными карбоксилатными группами линкера происходило в пределах 3 часов при комнатной температуре. Частицы промывали три раза, используя 2 мл дистиллированной воды. Добавляли 1 мл 1М NiSO4. Свободные ионы Ni2+ образовывали комплекс с карбоксилатными группами NTA, и формировали NTA-Ni. Через 5 минут частицы дважды промывали 2 мл дистиллированной воды, и дополнительно дважды промывали 2 мл 50 мМ MOPS, 0,1 М NaCl, 5 мМ CaCl2, рН 6,0.
С. Конъюгирование с His-мечеными белковыми носителями
Белки основного каркаса связывали с модифицированными наночастицами путем инкубирования описанных выше частиц с 1-1,5 мг белка в течение ночи в 2 мл 50 мМ MOPS, 0,1 М NaCl, 5 мМ CaCl2, рН 6,0.
Основные каркасы с разным количеством когезинов с углеводсвязывающим модулем (CBM) (CBM-c1-c1 и с1-с1) и без него иммобилизовали на наночастицах с модифицированной поверхностью.
Эффективность связывания белков с наночастицами определяли спектрофотометрическим измерением методом оптического поглощения (590 нм, анализ Брэдфорда (Bradford)) содержания белка до и после сшивки. Наночастицы, нагруженные белком, отделяли от реакционного раствора при помощи магнита. Эффективность связывания вычисляли путем вычитания количества оставшегося в реакционной смеси белка из первоначально использованного количества белка.
Количественное определение восстанавливающих сахаров проводили по меньшей мере три раза в линейной области реакции методом20, в котором используется 3,5-динитросалициловая кислота, из расчета, что 1 единица фермента освобождает 1 мкмоль эквивалента глюкозы в минуту.
Результаты
Было показано, что связывание на поверхности (иммобилизация) часто стабилизирует ферменты и их активность. Эксперименты с целлюлазами доказали, что непосредственная иммобилизация гидролитических ферментов (целлюлаз из С. thermocellum) на поверхности наночастиц уменьшает их удельную активность. Это означает, что непосредственно ковалентное связывание с поверхностью может оказывать влияние на активные или структурные домены ферментов. Поэтому для поддержания ферментативной активности было выбрано направленное специфичное связывание с целлюлосомальными основными каркасами.
Наилучшие результаты связывания всех протестированных реакций были получены в случае использования 2 ммоль EDC и 5 ммоль сульфо-NHS. Может быть достигнута эффективность связывания 80 мкг/мг наночастиц. Это соответствует вычисленному среднему количеству, равному примерно 1300 молекул основного каркаса на одну частицу. Между модифицированными карбоксильными группами частиц сшивка не наблюдалась. Кроме того, благодаря наличию свободных карбоксильных групп и образуемой в результате гидрофильной поверхности COOH-бусины ресуспендировали очень хорошо. Разделение показано на фиг.13.
Для восстановления магнитных наночастиц частицы промывали три раза ЭДТА (10 мМ) для удаления комплексных ионов Ni2+ и суспендировали в том же растворе в течение ночи при комнатной температуре. После трехразовой промывки деионизированной водой частицы снова нагружали Ni2+ и 6xHis-меченными белками основного каркаса (согласно рассчитанному эквимолярному отношению белков основного каркаса к ферментным компонентам). 1 мг свежемодифицированных COOH- группами наночастиц может быть нагружен примерно 80 мкг белка основного каркаса. В этом эксперименте такое же количество восстановленных наночастиц было способно связывать примерно 50 мкг (62,5% восстановления) белков скаффолдина, а дважды восстановленные наночастицы могли иммобилизовать примерно 25 мкг (31,2% восстановления) белков.
Целлюлазы были иммобилизованы на соединении наночастица-основной каркас путем когезин-докерин узнавания. Частицы нагружали мутантными ферментами SM901 и их удельную активность по отношению к растворимой, аморфной и нерастворимой целлюлозе определяли по получению восстанавливающих сахаров, используя в качестве реагента динитросалициловую кислоту, как описано в уровне техники. Эффективность деградации зависит от типа субстрата. Наиболее доступными для гидролиза являются β-глюкан ячменя, растворимый β-1,3-1,4-глюкан. Удельная активность β-глюканазы мутантных ферментов SM901 по отношению к β-глюкану ячменя составила около 8 ЕД/мг белка. Удельная активность для деградации карбоксиметилцеллюлозы составила примерно 1,1 ЕД/мг белка. Аморфная целлюлоза (набухшая в фосфорной кислоте целлюлоза) является более подверженной деградации, что дает удельную активность примерно 2,8 ЕД/мг белка. Сравнение с аналогичными комплексами, связанными и не связанными с наночастицами, показывает, что иммобилизация гидролитических ферментов посредством основных каркасов целлюлосомального типа не оказывает негативного влияния на скорость деградации для всех исследованных субстратов.
На кристаллической целлюлозе (MN300 или Avicel) свободные мутантные ферменты показали удельную активность, равную 30 и 12 мЕД/мг, соответственно. Очищенные нативные целлюлосомы с 9 когезинами и углеводсвязывающим модулем показали удельную активность, равную 423 мЕД/мг, по отношению к MN300, использованному в качестве субстрата и, равную 198 мЕД/мг, по отношению к Avicel. При образовании комплекса мутантных ферментов SM901 с основным каркасом, содержащим большее количество когезинов, гидролитическая активность усиливалась, достигая удельной активности 63 и 28 мЕД/мг, соответственно. По сравнению со свободными ферментами активность увеличилась в 2,1 и 2,3 раза. Если использовали комплекс с тремя когезинами и углеводсвязывающий модуль семейства 3 скорость деградации могла увеличиваться в 4,9 и 3,7 раз по сравнению с несвязанными гидролазами (фиг. 5).
Для сравнения, использовали белки основного каркаса без CBM или содержащие CBM (с1-с1 и CBM-c1-c1). Удельная активность возростала с 62 мЕД/мг до 102 мЕД/мг в случае использования в качестве субстрата MN300 и с 44 мЕД/мг до 108 мЕД/мг в случае использования в качестве субстрата Avicel, если в основном каркасе присутствовал CBM.
Иммобилизация гидролитических ферментов на связанных с наночастицами мини-скаффолдинах, не оказывала негативного влияния на скорость деградации растворимых и нерастворимых субстратов. Тем не менее, рН-стабильность и термостабильность увеличивалась значительно (фиг.6 и 7, соответственно).
Ссылки:
1 Schwarz, W. H., (2004). Cellulose „Struktur ohne Ende“, Naturwiss. Rundschau, 8:443-445.
2 Singhania, R. R., et al. (2010), “Advancement and comparative profiles in the production technologies using solid-state and submerged fermentation for microbial cellulases”; Enz. Microb. Technol. 46:541-549.
3 Lynd, L., Weimer, P. J., van Zyl, W. H., and Pretorius, I. S. (2002). Microbial cellulose utilization: fundamentals and biotechnology. Microbiol. Molec. Biol. Rev. 66, 506-577.
4 Shoham, Y., R. Lamed, and E. A. Bayer. 1999. The cellulosome concept as an efficient microbial strategy for the degradation of insoluble polysaccharides. Trends in Microbiol. 7:275-281.
5Bayer, E. A., Y. Shoham, and R. Lamed. 2000. Cellulose-decomposing bacteria and their enzyme systems. In M. Dworkin, S. Falkow, E. Rosenberg, K.-H. Schleifer, and E. Stackebrandt (eds.), The Prokaryotes: An Evolving Electronic Resource for the Microbiological Community, 3rd edition. Springer-Verlag, New York.
6 Schwarz, W. H. 2001. The cellulosome and cellulose degradation by anaerobic bacteria. Appl. Microbiol. Biotechnol. 56:634-649.
7 Mechaly, A., S. Yaton, R. Lamed, H.-P. Fierobe, A. Belaich, J.-P. Belaich, Y. Shoham, and E.A. Bayer. 2000. Cohesin-dockerin recognition in cellulosome assembly: Experiment versus hypothesis. Proteins 39:170-177.
8 Leibovitz, E., H. Ohayon, P. Gounon, and P. Béguin. 1997. Characterization and subcellular localization of the Clostridium thermocellum scaffoldin dockerin binding protein SdbA. J. Bacteriol. 179:2519-2523.
9 Bayer, E. A., L. J. W. Shimon, Y. Shoham, and R. Lamed. 1998. Cellulosomes - structure and ultrastructure. J. Struct. Biol. 124:221-234.
10 Zverlov, V. V., G. A. Velikodvorskaya, and W. H. Schwarz. 2003. Two new cellulosome components encoded downstream of celI in the genome of Clostridium thermocellum: the non-processive endoglucanase CelN and the possibly structural protein CseP. Microbiol. 149:515-524.
11 Zverlov, V. V., Kellermann, J., & Schwarz, W. H. (2005). Functional subgenomics of Clostridium thermocellum cellulosomal genes: Identification of the major catalytic components in the extracellular complex and detection of three new enzymes. Proteomics, 5. 3646-3653.
12 DOE Joint Genome Institute: http://genome.jgi-psf.org/microbial/
13 Gold N.D., & Martin V.J. (2007). Global view of the Clostridium thermocellum cellulosome revealed by quantitative proteomic analysis. J Bacteriol. 189:6787-95.
14 Fierobe, H.-P. et al. (2002). Degradation of cellulose substrates by cellulosome chimeras. J. Biol.Chem. 277: 49621-49630.
15 Bayer, E. A., R. Kenig, & R. Lamed (1983). Adherence of Clostridium thermocellum to cellulose. J. Bacteriol. 156: 818-827.
16 Bayer, E.A., Y. Shoham, J. Tormo, & R. Lamed (1996). The cellulosome: a cell surface organelle for the adhesion to and degradation of cellulose. In: Bacterial adhesion: molecular and ecological diversity, pp. 155-182. Wiley-Liss, Inc.
17 Zverlov, V.V., Klupp, M., Krauss, J., & Schwarz, W.H. (2008). Mutants in the scaffoldin gene cipA of Clostridium thermocellum with impaired cellulosome formation and cellulose hydrolysis: insertions of a new IS-element, IS1447, and implications for cellulase synergism on crystalline cellulose. J. Bacteriol. 190: 4321-4327.
18 Staros, J. V., Wright, R. W., Swingle, D. M., (1986). Enhancement by N-hydroxysulfosuccinimide of water-soluble carbodiimide-mediated coupling reactions. Anal. Biochem., 156:220-222.
19 Hermanson, G. T. (1995). Bioconjugate techniques. Academic Press.
20 Wood, T. M., and K. M. Bhat. 1988. Methods for measuring cellulase activities. Methods Enzymol. 160:87-112.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОНСТРУКЦИИ И СПОСОБЫ ДЛЯ ПРОДУЦИРОВАНИЯ И СЕКРЕЦИИ ФЕРМЕНТОВ, РАСЩЕПЛЯЮЩИХ КЛЕТОЧНУЮ СТЕНКУ | 2009 |
|
RU2525194C2 |
| ВИЧ ВАКЦИНА, ОСНОВАННАЯ НА НАПРАВЛЕННОСТИ МАКСИМИЗИРОВАННЫХ Gag И Nef НА ДЕНДРИТНЫЕ КЛЕТКИ | 2009 |
|
RU2539765C2 |
| СЛИТЫЕ БЕЛКИ, ВКЛЮЧАЮЩИЕ ФЕРМЕНТЫ, РАЗРУШАЮЩИЕ КЛЕТОЧНУЮ ОБОЛОЧКУ РАСТЕНИЙ, И ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2433140C2 |
| ЦЕЛЛЮЛОЗНЫЕ БЕЛКИ СЛИЯНИЯ И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2458127C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM CANESCENS CL14, ПРОДУЦИРУЮЩИЙ КОМПОНЕНТ ЦЕЛЛЛЮЛОСОМЫ CLOSTRIDIUM THERMOCELLUM, И СПОСОБ ЕГО КУЛЬТИВИРОВАНИЯ | 2016 |
|
RU2646132C1 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ D-ТАГАТОЗЫ | 2016 |
|
RU2749811C1 |
| ФЕРМЕНТНАЯ КОМПОЗИЦИЯ, СПОСОБНАЯ ЭФФЕКТИВНО РАЗЛАГАТЬ ЦЕЛЛЮЛОЗНЫЙ МАТЕРИАЛ | 2009 |
|
RU2529949C2 |
| НОВЫЙ РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM CANESCENS CS15, ПРОДУЦИРУЮЩИЙ ЦЕЛЛЮЛАЗУ CLOSTRIDIUM THERMOCELLUM, И СПОСОБ ЕГО КУЛЬТИВИРОВАНИЯ | 2015 |
|
RU2612158C1 |
| КОМПОЗИЦИЯ ФЕРМЕНТНОЙ СМЕСИ ДЛЯ ГИДРОЛИЗА СМЕСИ ЦЕЛЛЮЛОЗНЫХ И ГЕМИЦЕЛЛЮЛОЗНЫХ МАТЕРИАЛОВ (ВАРИАНТЫ) И СПОСОБЫ ЕЕ ИСПОЛЬЗОВАНИЯ (ВАРИАНТЫ) | 2009 |
|
RU2536256C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ D-АЛЛЮЛОЗЫ | 2017 |
|
RU2819709C2 |

















































































Предложены комплекс для ферментативного гидролиза полисахаридных субстратов, способ его получения и его использование. Комплекс содержит основной каркас и ферментные компоненты. Основной каркас содержит один или более белков, выбранных из группы, состоящей из CBM-c1-c1-d3, представленного в SEQ ID NO: 24, c3-c1-c1-d2, представленного в SEQ ID NO: 22, c2-c1-c1-c1, представленного в SEQ ID NO: 26, или их производных, имеющие более чем 60% идентичность аминокислотной последовательности в своих модулях когезина. Упомянутые белки могут быть сшиты вместе посредством взаимодействия когезин-докерин. Также основной каркас содержит по меньшей мере четыре участка связывания, представляющие собой когезинсвязывающие участки, по меньшей мере два из которых имеют по существу одинаковую специфичность связывания. Ферментные компоненты связаны с каждым из указанных выше участков связывания, причем по меньшей мере три из ферментных компонентов являются разными. Также предложены способ получения комплекса и применение этого комплекса для ферментативного гидролиза полисахаридных субстратов. 4 н. и 8 з.п. ф-лы, 13 ил., 1 табл., 4 пр.
1. Комплекс для ферментативного гидролиза полисахаридных субстратов, содержащий:
(a) основной каркас, содержащий один или более белков, выбранных из группы, состоящей из CBM-c1-c1-d3, представленного в SEQ ID NO: 24, c3-c1-c1-d2, представленного в SEQ ID NO: 22, c2-c1-c1-c1, представленного в SEQ ID NO: 26, или их производных, имеющие более чем 60% идентичность аминокислотной последовательности в своих модулях когезина, и содержащий по меньшей мере четыре участка связывания, причем упомянутые белки могут быть сшиты вместе посредством взаимодействия когезин-докерин, вследствие чего специфичность связывания для взаимодействия при сшивке отличается от специфичности связывания ферментов, причем упомянутые участки связывания представляют собой когезинсвязывающие участки, по меньшей мере два из которых имеют по существу одинаковую специфичность связывания; и
(b) ферментные компоненты, связанные с каждым из указанных по меньшей мере четырех участков связывания, где по меньшей мере три из указанных ферментных компонентов являются разными ферментными компонентами, причем ферментные компоненты выбирают из группы, состоящей из: процессивных или непроцессивных эндо-β-1,4-глюконаз, процессивных экзо-β-1,4-глюконаз и гликозидаз из полисахаролитических микроорганизмов.
2. Комплекс по п. 1, в котором основной каркас получают из некаталитического каркасного белка CipA из Clostridium thermocellum.
3. Комплекс по п. 1 или 2, в котором основной каркас содержит углеводсвязывающий модуль (СМВ).
4. Комплекс по п. 3, в котором углеводсвязывающий модуль является углеводсвязывающим модулем (СВМ3) из гена cipA Closthdium thermocellum, который интегрирован или прикреплен к линейному каркасу остова.
5. Комплекс по пп. 1, 2, или 4, в котором ферментный компонент содержит компонент, содержащий модуль докерина, Closthdium thermocellum.
6. Комплекс по пп. 1, 2, или 4, в котором ферментные компоненты содержат CelK-d1, представленный в SEQ ID NO: 8, CelR-d1, представленный в SEQ ID NO: 10, CelT-d1, представленный в SEQ ID NO: 14, CelE-d1, представленный в SEQ ID NO: 16, CelS-d1, представленный в SEQ ID NO: 6, и BglB-d1, представленный в SEQ ID NO: 4, или их производные, имеющие более чем 50% идентичность аминокислотной последовательности в своих модулях докерина.
7. Способ получения комплекса по любому из пп. 1-6, содержащий стадии:
a) получение рекомбинантным способом ферментных компонентов;
b) получение рекомбинантным способом основного каркаса;
c) смешивание очищенных, частично очищенных или неочищенных компонентов из стадий a) и b) in vitro при температуре от 25-65°C, предпочтительно 30-65°C, более предпочтительно примерно 55°C, таким образом, чтобы общее количество основных каркасов и ферментных компонентов составляло в молярном отношении 1 модуль когезина к 1 ферментному компоненту, и по меньшей мере три ферментных компонента составляли в молярном отношении друг к другу от 1:1 до 1:15, и
d) связывание случайным образом ферментных компонентов с основным каркасом при температуре от 25-65°C, предпочтительно 30-65°C, более предпочтительно примерно 55°C.
8. Способ ферментативного гидролиза полисахаридных субстратов, содержащий стадии:
a) смешивание комплекса по любому из пп. 1-6 с нерастворимой целлюлозой; и необязательно
b) выделение продуктов деградации.
9. Способ по п. 8, где смешивание комплекса по любому из пп. 1-6 с нерастворимой целлюлозой осуществляют в водной среде при pH равной 6,0-7,0 и температуре 25-65°C, предпочтительно 30-65°C, более предпочтительно примерно 55°C.
10. Способ по любому из пп. 8 или 9, где полисахаридным субстратом является кристаллическая целлюлоза или содержащий кристаллическую целлюлозу субстрат.
11. Применение комплекса по любому из пп. 1-6 для ферментативного гидролиза полисахаридных субстратов.
12. Применение по п. 11, где полисахаридным субстратом является кристаллическая целлюлоза или содержащий кристаллическую целлюлозу субстрат.
| FIEROBE H.-P | |||
| ET AL | |||
| Design and production of active cellulosome chimeras // The Journal of Biological Chemistry, 2001, vol | |||
| СПОСОБ СОСТАВЛЕНИЯ ЗВУКОВОЙ ЗАПИСИ | 1921 |
|
SU276A1 |
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |
| ТЕЛЕМЕХАНИЧЕСКОЕ УСТРОЙСТВО | 1930 |
|
SU21257A1 |
| FIEROBE H.-P | |||
| ET AL | |||
| Action of designer cellulosomes on homogeneous versus complex substrates // The Journal of Biological Chemistry, 2005, vol | |||
| СПОСОБ ПОЛУЧЕНИЯ ЧИСТОГО ГЛИНОЗЕМА И ЕГО СОЛЕЙ ИЗ СИЛИКАТОВ ГЛИНОЗЕМА, ПРОСТЫХ ГЛИН И. Т.П. | 1915 |
|
SU280A1 |
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Способ получения моноазокрасителей | 1927 |
|
SU16325A1 |
| US 20090035811 A1, | |||

