ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет заявки США серийный номер 62/236,226, поданной 2 октября 2015 года, которая включена в данный документ посредством ссылки.
Область ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ изобретениЕ
Изобретение относится к получению сахара D-тагатозы. Более конкретно, изобретение относится к способам получения D-тагатозы посредством ферментативного преобразования сахаридов (напр., полисахаридов, олигосахаридов, дисахаридов, сахарозы, D-глюкозы и D-фруктозы) в D-тагатозу.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
D-тагатоза (далее тагатоза) представляет собой низкокалорийный, натуральный подсластитель, который имеет 92% сладости сахарозы, но только 38% калорий. Это природный моносахарид гексоза, которая присутствует только в небольших количествах во фруктах, какао и молочных продуктах. Тагатоза была одобрена в качестве пищевой добавки Управлением по контролю за продуктами и лекарствами (FDA) в 2003 году, которое определило ее как общепризнанно безопасную (GRAS). Однако вследствие высоких сбытовых цен на тагатозу ее использование в качестве подсластителя было ограничено. Тагатоза обладает множеством преимуществ для здоровья: она некариогенная; она низкокалорийная; она имеет очень низкий гликемический индекс, равный 3; она снижает гликемический индекс глюкозы на 20%; она может снижать уровни глюкозы в крови; она помогает предотвратить сердечно-сосудистые заболевания, инсульт и другие сосудистые заболевания путем повышения уровня холестерина липопротеинов высокой плотности (HDL); и это проверенный пребиотик и антиоксидант. Lu et al., Tagatose, a New Antidiabetic and Obesity Control Drug, Diabetes Obes. Metab. 10(2): 109-34 (2008). В связи с этим тагатоза явно имеет множество применений в фармацевтической, биотехнологической, научной, пищевой промышленности, производстве напитков, диетических добавок и бакалейной промышленности.
В настоящее время тагатозу получают преимущественно путем гидролиза лактозы лактазой или кислотным гидролизом с образованием D-глюкозы и D-галактозы (WO 2011150556, CN 103025894, US 5002612, US 6057135 и US 8802843). D-галактозу затем изомеризуют в D-тагатозу либо химически гидроксидом кальция в щелочных условиях, либо ферментативно с помощью L-арабинозоизомеразы в условиях нейтрального pH. Конечный продукт выделяют посредством комбинации фильтрации и ионообменной хроматографии. Этот способ выполняется в нескольких резервуарах или биореакторах. В целом, этот способ страдает из-за дорогостоящего отделения других сахаров (напр., D-глюкозы, D-галактозы и негидролизированной лактозы) и низкого выхода продукта. Разрабатывается несколько способов путем ферментации микробных клеток, но ни один из них не зарекомендовал себя в качестве практической альтернативы вследствие их зависимости от дорогостоящего сырья (напр., галактитола и D-псикозы), низких выходов продукта и дорогостоящего разделения.
Необходимо разработать экономически эффективный синтетический путь для высокоэффективного получения тагатозы, в котором по меньшей мере одна стадия способа включает энергетически выгодную химическую реакцию. Кроме того, существует потребность в производстве тагатозы, в котором этапы способа могут проводиться в одном резервуаре или биореакторе. Существует также необходимость в способе получения тагатозы, который может быть проведен при относительно низкой концентрации фосфата, при этом фосфат может повторно использоваться, и/или способ не требует использования аденозинтрифосфата (АТФ) в качестве источника фосфата. Существует также потребность в способе получения тагатозы, который не требует использования дорогостоящего кофермента никотинамида аденозиндинуклеотида (НАД (Н)) на любой из стадий реакции.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретения, описанные в данном документе, относятся к способам получения тагатозы. В различных аспектах способы включают преобразование фруктозо-6-фосфата (F6P) в тагатозо-6-фосфат (T6P), катализируемое эпимеразой; и преобразование Т6Р в тагатозу, катализируемое фосфатазой. Изобретения также относятся к тагатозе, полученной любым из способов, описанных в данном документе.
В некоторых аспектах изобретения способ получения тагатозы также включает стадию преобразования глюкозо-6-фосфата (G6P) в F6P, при этом стадию катализирует фосфоглюкозоизомераза (PGI). В других аспектах способ синтеза тагатозы также включает стадию преобразования глюкозо-1-фосфата (G1P) в G6P, и данную стадию преобразования катализирует глюкозофосфомутаза (PGM).
В различных аспектах способ получения тагатозы может включать преобразование сахарида в G1P, катализируемое по меньшей мере одним ферментом; преобразование G1P в G6P, катализируемое глюкозофосфомутазой (PGM); преобразование G6P в F6P, катализируемое фосфоглюкозоизомеразой (PGI); преобразование F6P в тагатозо-6-фосфат (T6P), катализируемое эпимеразой; и преобразование полученного T6P в тагатозу, катализируемое фосфатазой.
Сахариды, используемые в любом из способов, могут быть выбраны из группы, состоящей из крахмала или его производных, целлюлозы или ее производных и сахарозы. Крахмал или его производные могут представлять собой амилозу, амилопектин, растворимый крахмал, амилодекстрин, мальтодекстрин, мальтозу или глюкозу. В некоторых аспектах изобретения способ получения тагатозы включает преобразование крахмала в производное крахмала посредством ферментного гидролиза или посредством кислотного гидролиза крахмала. В других аспектах производное крахмала может быть получают посредством ферментного гидролиза крахмала, катализируемого изоамилазой, пуллуланазой, альфа-амилазой или комбинацией двух или более этих ферментов. Способ получения тагатозы, в определенных аспектах, также может включать добавление 4-глюканотрансферазы (4GT).
В различных аспектах, способ получения тагатозы может включать преобразование фруктозы в F6P, катализируемое по меньшей мере одним ферментом; преобразование F6P в тагатозо-6-фосфат (T6P), катализируемое эпимеразой; и преобразование полученного T6P в тагатозу, катализируемое фосфатазой. В других вариантах осуществления способ получения тагатозы включает преобразование сахарозы во фруктозу, катализируемое по меньшей мере одним ферментом; преобразование фруктозы в F6P, катализируемое по меньшей мере одним ферментом; преобразование F6P в тагатозо-6-фосфат (T6P), катализируемое эпимеразой; и преобразование полученного T6P в тагатозу, катализируемое фосфатазой.
В других аспектах изобретения G6P, подлежащий использованию в способе получения тагатозы, может быть получен посредством преобразования глюкозы в G6P, катализируемого по меньшей мере одним ферментом. В свою очередь, глюкоза может быть получена посредством преобразования сахарозы в глюкозу, катализируемого по меньшей мере одним ферментом.
В некоторых аспектах изобретения эпимераза, используемая для преобразования F6P в T6P, представляет собой фруктозо-6-фосфатэпимеразу. Фруктозо-6-фосфатэпимераза может кодироваться полинуклеотидом, содержащим нуклеотидную последовательность, имеющую по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% идентичность последовательности с SEQ ID NOS.: 1, 3, 5, 7, 9 или 10. В различных аспектах фруктозо-6-фосфатэпимераза содержит аминокислотную последовательность, имеющую по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% идентичность последовательности с SEQ ID NOS.: 2, 4, 6, 8 или 11.
В различных аспектах изобретения фосфатаза, используемая для преобразования T6P в тагатозу, представляет собой тагатозо-6-фосфатфосфатазу. Тагатозо-6-фосфатфосфатаза может кодироваться полинуклеотидом, содержащим нуклеотидную последовательность, имеющую по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% идентичность последовательности с SEQ ID NO.: 12, 14 или 16. В некоторых аспектах изобретения тагатозо-6-фосфатфосфатаза содержит аминокислотную последовательность, имеющую по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% идентичность последовательности с SEQ ID NO.: 13, 15 или 17.
В различных аспектах способ изобретения должен проводиться при температуре, варьирующей от приблизительно 40°C до приблизительно 70°C, при рН, варьирующем от приблизительно от приблизительно 5,0 до приблизительно 8,0, и/или в течение от приблизительно 8 часов до приблизительно 48 часов. В некоторых аспектах стадии способа получения тагатозы проводят в одном биореакторе. В других аспектах стадии проводят во множестве биореакторов, расположенных последовательно.
В других аспектах изобретения стадии способа получения тагатозы проводят без наличия ATP, без наличия NAD(H), при концентрации фосфата от приблизительно 0 мМ до приблизительно 150 мМ, фосфат используется повторно, и/или по меньшей мере одна стадия способа включает энергетически выгодную химическую реакцию.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Эти чертежи иллюстрируют некоторые аспекты некоторых из вариантов осуществления изобретения и не должны использоваться для ограничения или установки границ изобретения.
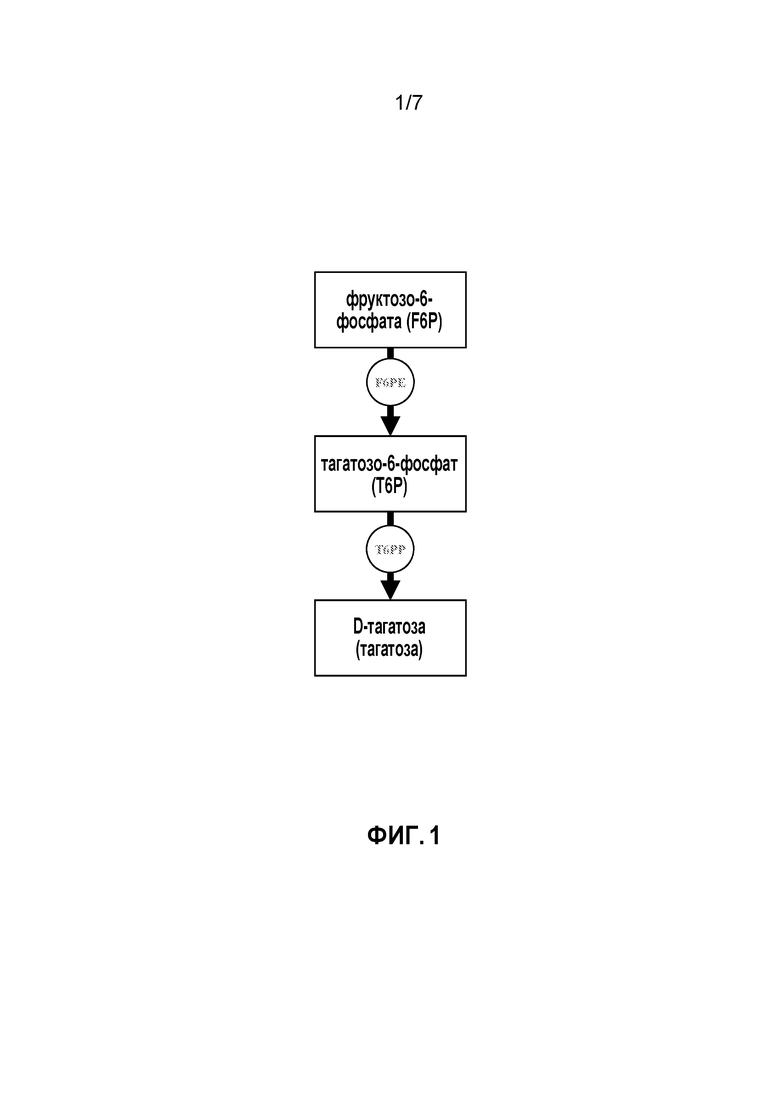
ФИГ. 1 представляет собой схематичное изображение, иллюстрирующее ферментативный путь преобразования фруктозо-6-фосфата в тагатозо-6-фосфат и затем в D-тагатозу (тагатозу).
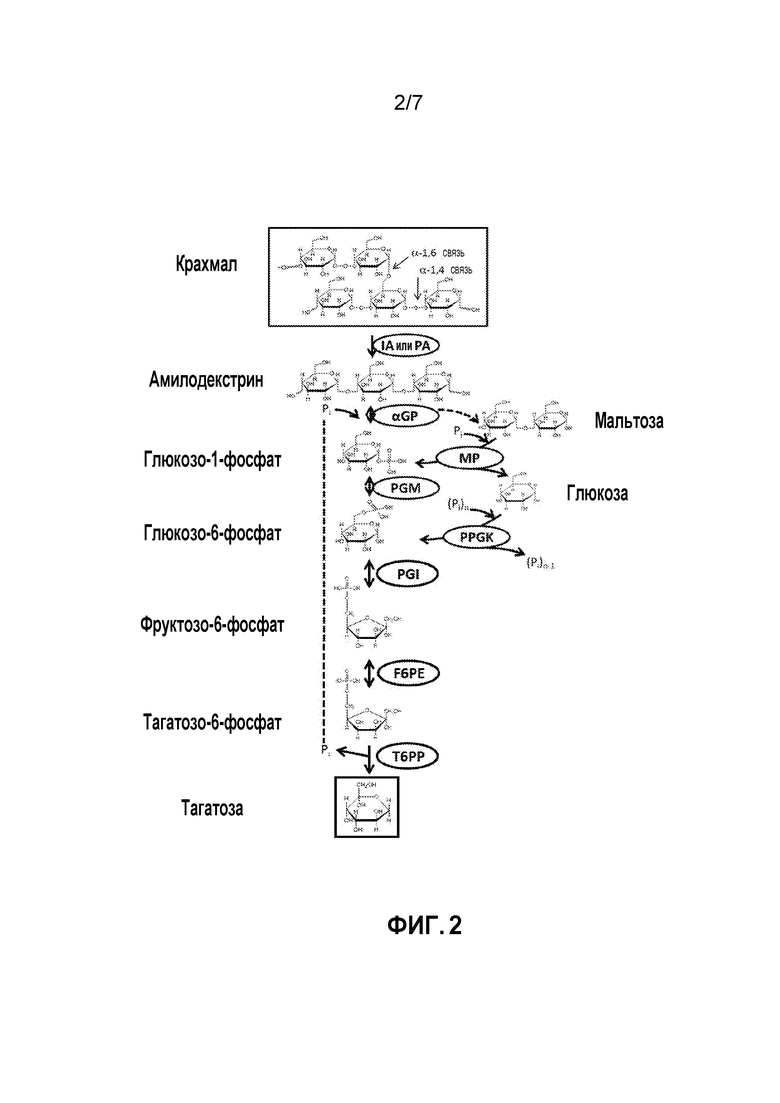
ФИГ. 2 представляет собой схематичное изображение, иллюстрирующее ферментативный путь преобразования крахмала или полученных из него продуктов в тагатозу. Используются следующие сокращения: αGP, альфа-глюканофосфорилаза или крахмальная фосфорилаза; PGM, глюкозофосфомутаза; PGI, фосфоглюкоизомераза; F6PE, фруктозо-6-фосфатэпимераза; T6PP, тагатозо-6-фосфатфосфатаза; IA, изоамилаза; PA, пуллуланаза; MP, мальтозофосфорилаза; PPGK, полифосфатглюкокиназа.
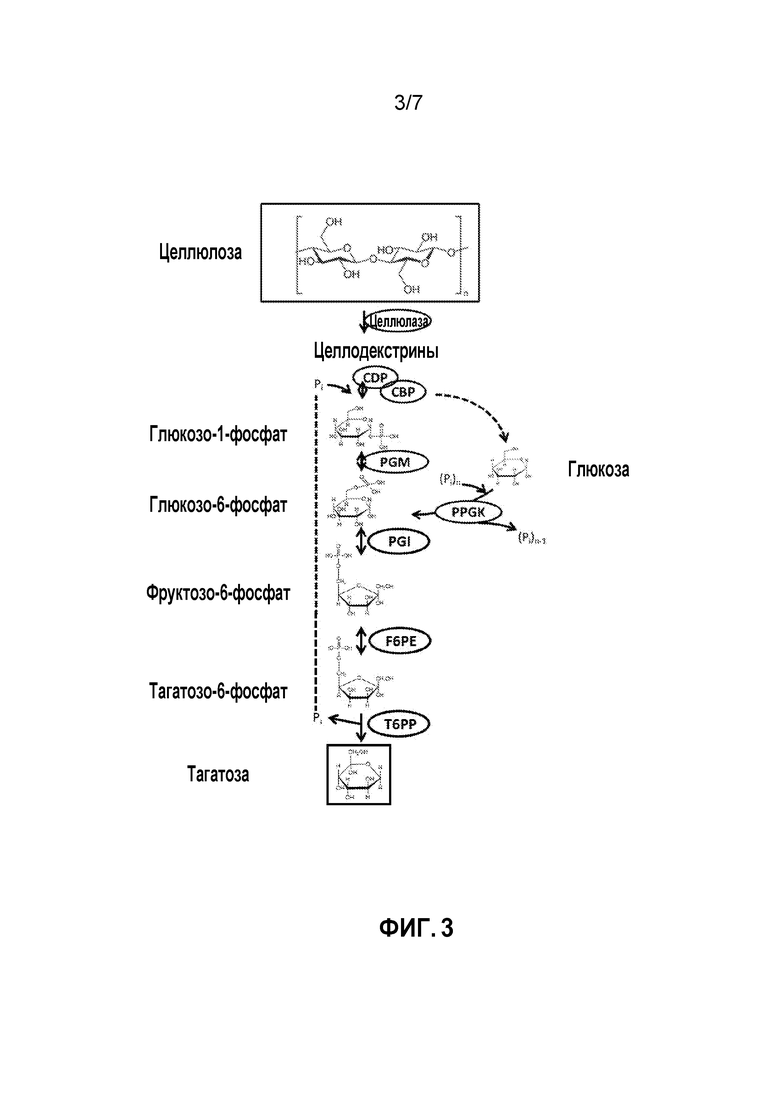
ФИГ. 3 демонстрирует ферментативный путь преобразования целлюлозы или полученных из нее продуктов в тагатозу. CDP, целлодекстринфосфорилаза; CBP, целлобиозофосфорилаза; PPGK, полифосфатглюкокиназа; PGM, глюкозофосфомутаза; PGI, фосфоглюкоизомераза; F6PE, фруктозо-6-фосфатэпимераза; T6PP, тагатозо-6-фосфатфосфатаза.
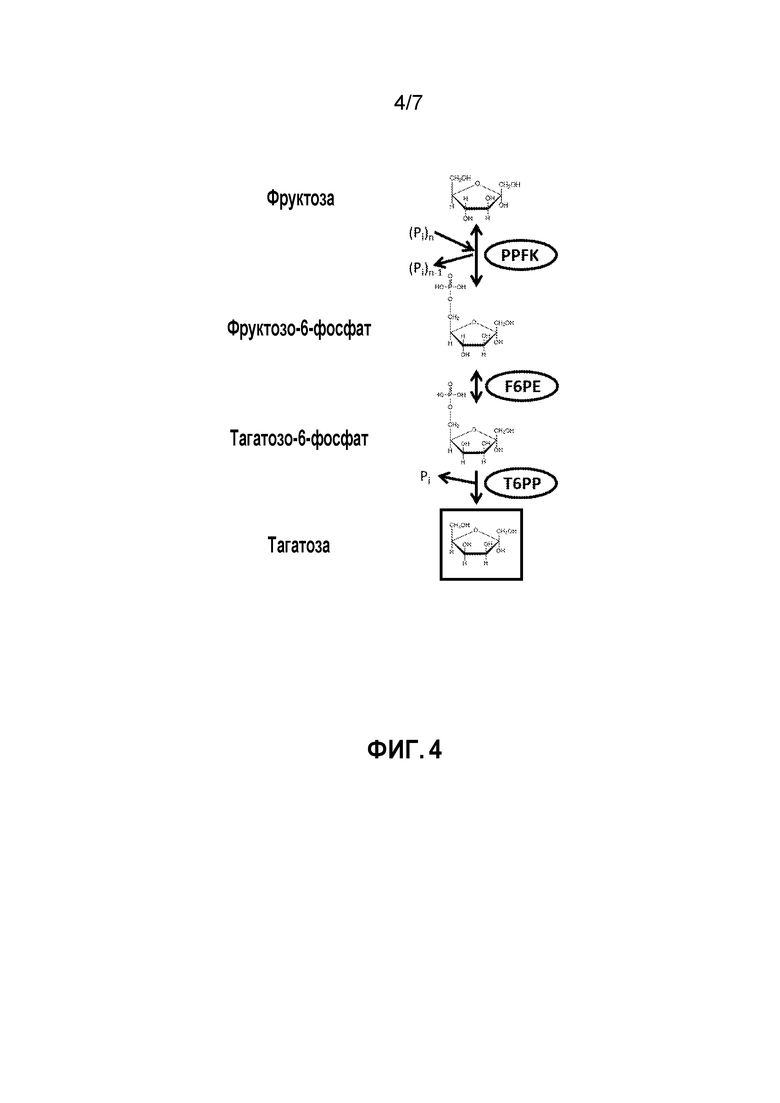
ФИГ. 4 представляет собой схематичное изображение, иллюстрирующее ферментативный путь преобразования фруктозы в тагатозу. PPFK, полифосфатфруктокиназа; F6PE, фруктозо-6-фосфатэпимераза; T6PP, тагатозо-6-фосфатфосфатаза.
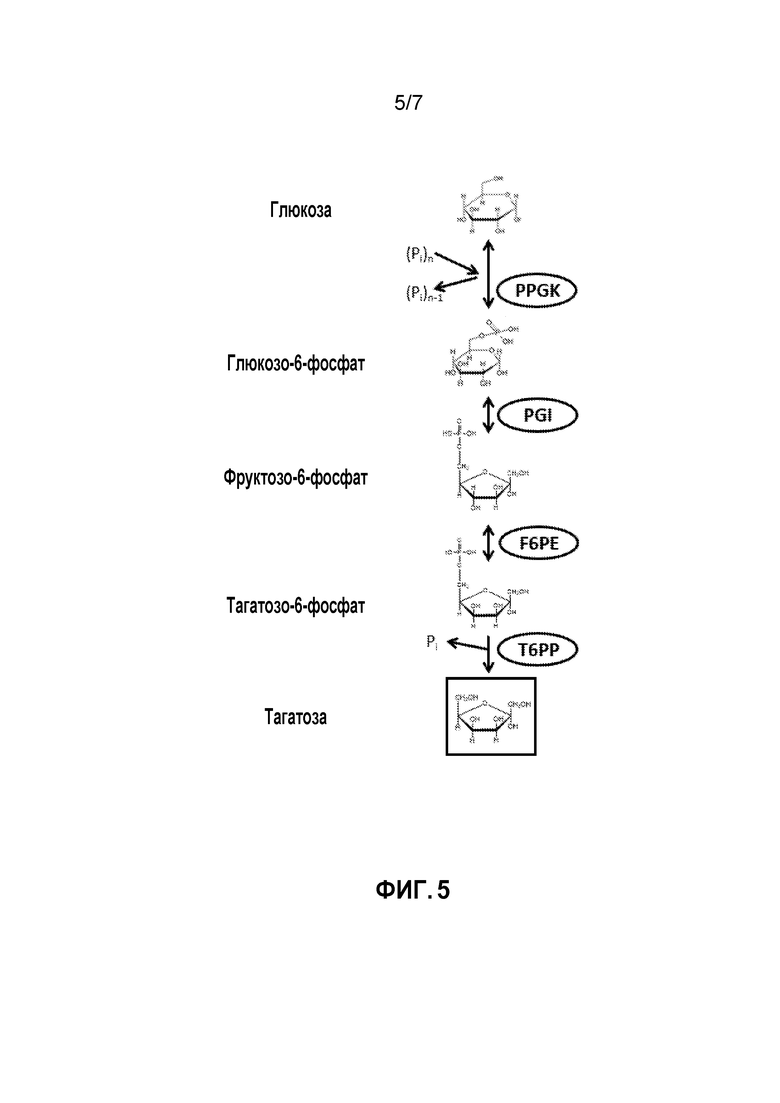
ФИГ. 5 представляет собой схематичное изображение, иллюстрирующее ферментативный путь преобразования глюкозы в тагатозу. PPGK, полифосфатглюкокиназа; PGI, фосфоглюкоизомераза; F6PE, фруктозо-6-фосфатэпимераза; T6PP, тагатозо-6-фосфатфосфатаза.
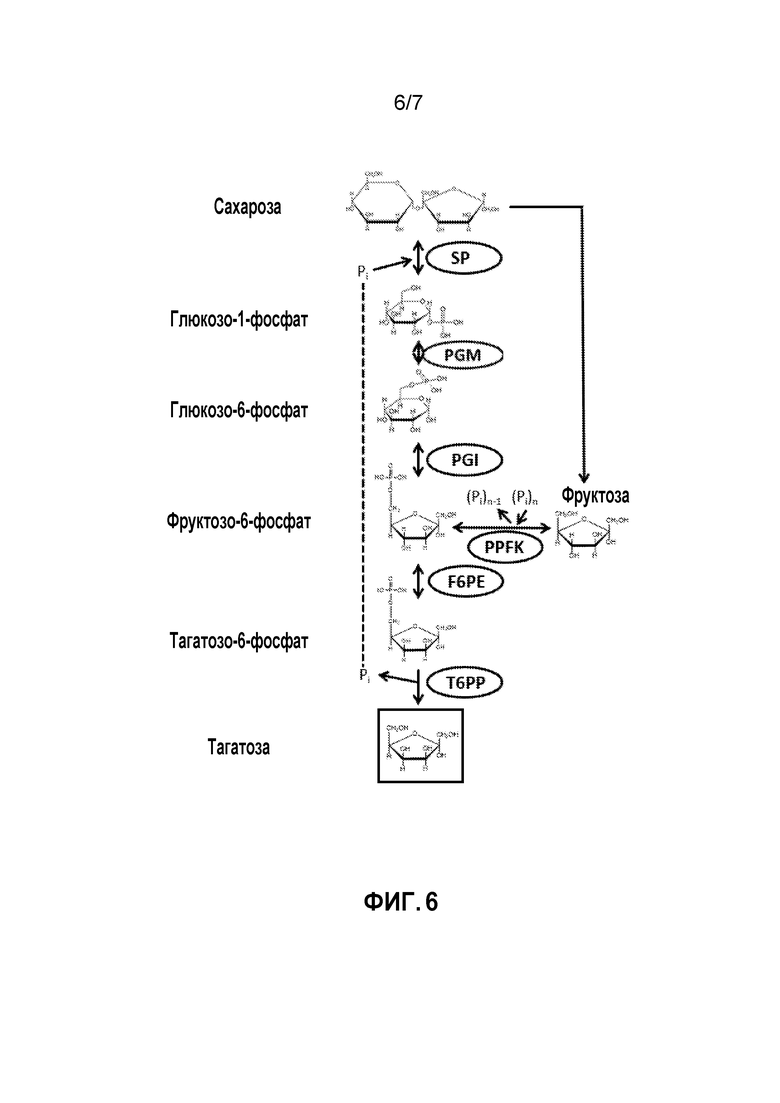
ФИГ. 6 демонстрирует ферментативный путь преобразования сахарозы или полученных из нее продуктов в тагатозу. SP, сахарозофосфорилаза; PPFK, полифосфатфруктокиназа; PGM, глюкозофосфомутаза; PGI, фосфоглюкоизомераза; F6PE, фруктозо-6-фосфатэпимераза; T6PP, тагатозо-6-фосфатфосфатаза.
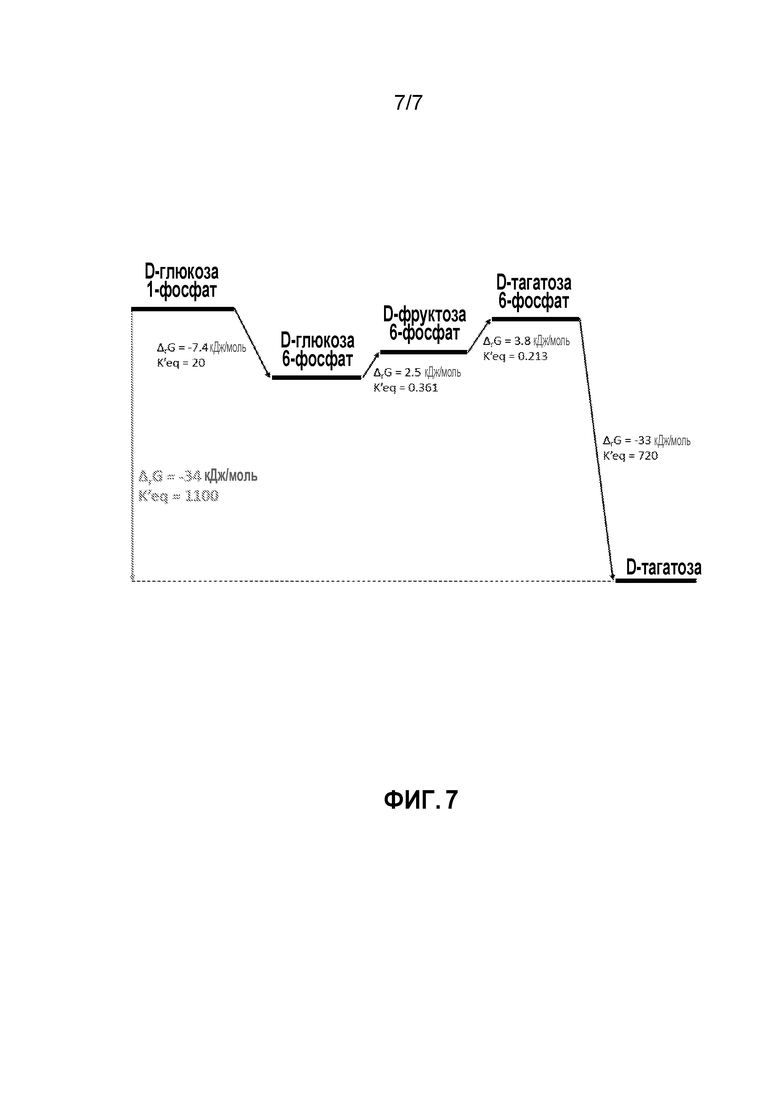
ФИГ. 7 демонстрирует энергию Гиббса реакции между промежуточными продуктами, исходя на образовании энергии Гиббса для преобразования глюкозо-1-фосфата в тагатозу.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретение предоставляет ферментативные пути или способы синтеза тагатозы с высоким выходом продукта, при этом значительно снижая затраты на разделение продукта и затраты на производство тагатозы.
Изобретение относится к способу получения тагатозы, при этом способ включает преобразование фруктозо-6-фосфата (F6P) в тагатозо-6-фосфат (T6P), катализируемое эпимеразой, и преобразование полученного T6P в тагатозу, катализируемое фосфатазой (напр., тагатозо-6-фосфатфосфатазой, T6PP). Этот способ в целом показан на ФИГ. 1. В некоторых вариантах осуществления, эпимераза, которая катализирует преобразование F6P в T6P, представляет собой фруктозо-6-фосфатэпимеразу (F6PE).
В способе изобретения могут использоваться эпимеразы, которые преобразуют F6P в T6P. В некоторых аспектах изобретения эпимеразы, подходящие для использования в способах преобразования F6P в T6P, содержат аминокислотную последовательность, которая имеет степень идентичности с аминокислотной последовательностью SEQ ID NOS.: 2, 4, 6, 8 или 11 (показано ниже), составляющую по меньшей мере 60%, предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и даже наиболее предпочтительно по меньшей мере 96%, 97%, 98%, 99% или 100%. Подходящие эпимеразы кодируются полинуклеотидом, содержащим нуклеотидную последовательность, которая имеет степень идентичности с нуклеотидной последовательностью SEQ ID NOS.: 1, 3, 5, 7, 9 или 10 (показано ниже), составляющую по меньшей мере 60%, предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и даже наиболее предпочтительно по меньшей мере 96%, 97%, 98%, 99% или 100%.
Изобретение также относится к эпимеразам, которые содержат аминокислотную последовательность, которая имеет степень идентичности с аминокислотной последовательностью SEQ ID NOS.: 2, 4, 6, 8 или 11, составляющую по меньшей мере 60%, предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и даже наиболее предпочтительно по меньшей мере 96%, 97%, 98%, 99% или 100%. В других аспектах изобретение относится к эпимеразам, которые кодируются полинуклеотидом, содержащим нуклеотидную последовательность, которая имеет степень идентичности с нуклеотидной последовательностью SEQ ID NOS.: 1, 3, 5, 7, 9 или 10, составляющую по меньшей мере 60%, предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и даже наиболее предпочтительно по меньшей мере 96%, 97%, 98%, 99% или 100%.
В способе изобретения могут использоваться фосфатазы, которые преобразуют T6P в тагатозу (D-тагатозу). В некоторых аспектах изобретения фосфатазы, которые могут использоваться для преобразования T6P в тагатозу (D-тагатозу), содержат аминокислотную последовательность, которая имеет степень идентичности с аминокислотной последовательностью SEQ ID NOS.: 12, 14 или 16 (показано ниже), составляющую по меньшей мере 60%, предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и даже наиболее предпочтительно по меньшей мере 96%, 97%, 98%, 99% или 100%. Тагатозофосфатазы кодируются полинуклеотидом, содержащим нуклеотидную последовательность, которая имеет степень идентичности с нуклеотидной последовательностью SEQ ID NOS.: 13, 15 или 17 (показано ниже), составляющую по меньшей мере 60%, предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и даже наиболее предпочтительно по меньшей мере 96%, 97%, 98%, 99% или 100%.
Изобретение также относится к фосфатазам, которые преобразуют T6P в тагатозу (D-тагатозу) и содержат аминокислотную последовательность, которая имеет степень идентичности с аминокислотной последовательностью SEQ ID NOS.: 12, 14 или 16, составляющую по меньшей мере 60%, предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и даже наиболее предпочтительно по меньшей мере 96%, 97%, 98%, 99% или 100%. В различных аспектах изобретение относится к фосфатазам, которые преобразуют T6P в тагатозу (D-тагатозу) и кодируются полинуклеотидом, содержащим нуклеотидную последовательность, которая имеет степень идентичности с нуклеотидной последовательностью SEQ ID NOS.: 13, 15 или 17, составляющую по меньшей мере 60%, предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и даже наиболее предпочтительно по меньшей мере 96%, 97%, 98%, 99% или 100%.
В некоторых вариантах осуществления способ получения тагатозы согласно изобретению также включает стадию ферментативного преобразования глюкозо-6-фосфата (G6P) в F6P, и эта стадия катализируется фосфоглюкозоизомеразой (PGI). В других вариантах осуществления способ получения тагатозы дополнительно включает стадию преобразования глюкозо-1-фосфата (G1P) в G6P, при этом стадия катализируется глюкозофосфомутазой (PGM). В дополнительных вариантах осуществления способ получения тагатозы также включает стадию преобразования сахарида в G1P, которая катализируется по меньшей мере одним ферментом.
Вследствие этого, способ получения тагатозы согласно изобретению может, например, включать следующие стадии: (i) преобразования сахарида в глюкозо-1-фосфат (G1P) с использованием одного или более ферментов; (ii) преобразования G1P в G6P с использованием глюкозофосфомутазы (PGM, EC 5.4.2.2); (iii) преобразования G6P в F6P с использованием фосфоглюкоизомеразы (PGI, EC 5.3.1.9); (iv) преобразования F6P в T6P посредством фруктозо-6-фосфатэпимеразы (F6PE) и (v) преобразования T6P в тагатозу посредством тагатозо-6-фосфатфосфатазы (T6PP). Пример способа, в котором сахарид представляет собой крахмал, показан на ФИГ. 2.
Как правило, соотношения ферментных единиц, используемых в описанном способе, составляют 1:1:1:1:1 (αGP:PGM:PGI:F6PE:T6PP). Чтобы оптимизировать выходы продукта, эти соотношения могут быть скорректированы в любом количестве комбинаций. Например, соотношение 3:1:1:1:1 можно использовать для максимизации концентрации фосфорилированных промежуточных продуктов, что приведет к увеличению активности последующих реакций. И наоборот, соотношение 1:1:1:1:3 может быть использовано для поддержания надежного снабжения фосфатом для αGP, что приведет к более эффективному фосфоролитическому расщеплению альфа-1,4-гликозидных связей. Соотношение ферментов, например, 3:1:1:1:3 может быть использовано для дополнительного увеличения скорости реакции. Вследствие этого соотношения ферментов, включая другие необязательные ферменты, обсуждаемые ниже, могут быть изменены для повышения эффективности получения тагатозы. Например, конкретный фермент может присутствовать в количестве приблизительно 2 х, 3 х, 4 х, 5 х и т. д. относительно количества других ферментов.
Одним из важных преимуществ способов является то, что стадии способа можно проводить в одном биореакторе или реакционном сосуде. В качестве альтернативы, стадии также можно проводить во множестве биореакторов или реакционных сосудов, которые расположены последовательно.
Фосфат-ионы, полученные посредством дефосфорилирования T6P T6PP, могут затем использоваться повторно на стадии способа преобразования сахарида в G1P, особенно когда всю стадию способа проводят в одном биореакторе или реакционном сосуде. Способность повторно использовать фосфат в раскрытых способах обеспечивает использование нестехиометрических количеств фосфата, которые сохраняют низкой концентрацию фосфатов реакции. Это влияет на общий путь и общую скорость способов, но не ограничивает активность отдельных ферментов и обеспечивает общую эффективность способов получения тагатозы.
Например, концентрация фосфатов реакции может варьировать от приблизительно 0 мМ до приблизительно 300 мМ, от приблизительно 0 мМ до приблизительно 150 мМ, от приблизительно 1 мМ до приблизительно 50 мМ, предпочтительно от приблизительно 5 мМ до приблизительно 50 мМ, или более предпочтительно от приблизительно 10 мМ до приблизительно 50 мМ. Например, концентрация фосфатов реакции может составлять приблизительно 0,1 мМ, приблизительно 0,5 мМ, приблизительно 1 мМ, приблизительно 1,5 мМ, приблизительно 2 мМ, приблизительно 2.5 мМ, приблизительно 5 мМ, приблизительно 6 мМ, приблизительно 7 мМ, приблизительно 8 мМ, приблизительно 9 мМ, приблизительно 10 мМ, приблизительно 15 мМ, приблизительно 20 мМ, приблизительно 25 мМ, приблизительно 30 мМ, приблизительно 35 мМ, приблизительно 40 мМ, приблизительно 45 мМ, приблизительно 50 мМ или приблизительно 55 мМ.
Вследствие этого, низкая концентрация фосфата приводит к понижению затрат на производство вследствие низкого общего фосфата и, таким образом, к понижению стоимости удаления фосфата. Это также предотвращает ингибирование T6PP высокими концентрациями свободного фосфата и уменьшает возможность загрязнения фосфатами.
Кроме того, способы, раскрытые в данном документе, могут проводиться без добавления ATP в качестве источника фосфата, т.е. без наличия ATP. Способы также могут проводиться без необходимости добавления NAD(H), т.е. без наличия NAD(H). Другие преимущества также включают то, что по меньшей мере одна стадия раскрытых способов получения тагатозы включает энергетически выгодную химическую реакцию (ФИГ. 7).
Примеры ферментов, используемых для преобразования сахарида в G1P, включают альфа-глюканофосфорилазу (αGP, EC 2.4.1.1), мальтозофосфорилазу (MP, EC 2.4.1.8), целлодекстринфосфорилазу (CDP, EC 2.4.1.49), целлобиозофосфорилазу (CBP, EC 2.4.1.20), целлюлозофосфорилазу, сахарозофосфорилазу (SP, EC 2.4.1.7) и их комбинацию. Выбор фермента или комбинации ферментов зависит от используемого в способе сахарида.
Сахариды, используемые для получения G1P, могут представлять собой полисахариды, олигосахариды и/или дисахариды. Например, сахарид может представлять собой крахмал, одно или более производных крахмала, целлюлозу, одно или более производных целлюлозы, сахарозу, одно или более производных сахарозы или их комбинацию.
Крахмал является наиболее широко используемым веществом сохранения энергии в природе и в основном хранится в семенах растений. Природный крахмал содержит линейную амилозу и разветвленный амилопектин. Примеры производных крахмала включают амилозу, амилопектин, растворимый крахмал, амилодекстрин, мальтодекстрин, мальтозу, фруктозу и глюкозу. Примеры производных целлюлозы включают предварительно обработанную биомассу, регенерированную аморфную целлюлозу, целлодекстрин, целлобиозу, фруктозу и глюкозу. Производные сахарозы включают фруктозу и глюкозу.
Производные крахмала могут быть получены посредством ферментного гидролиза крахмала или посредством кислотного гидролиза крахмала. Конкретно, ферментативный гидролиз крахмала может катализироваться или усиливаться изоамилазой (IA, EC. 3.2.1.68), которая гидролизирует α-1,6-глюкозидные связи; пуллуланазой (PA, EC. 3.2.1.41), которая гидролизирует α-1,6-глюкозидные связи; 4-α-глюканотрансферазой (4GT, EC. 2.4.1.25), которая катализирует трансгликозилирование коротких мальтоолигосахаридов, с получением более длинных мальтоолигосахаридов; или альфа-амилазой (EC 3.2.1.1), которая расщепляет α-1,4-глюкозидные связи.
Кроме того, производные целлюлозы могут быть получены посредством ферментного гидролиза целлюлозы, катализируемого смесями целлюлозы, кислотами, или предварительно обработанной биомассой.
В некоторых вариантах осуществления ферменты, используемые для преобразования сахарида в G1P, содержат αGP. На этой стадии, когда сахариды включают крахмал, G1P получают из крахмала с помощью αGP; когда сахариды включают растворимый крахмал, амилодекстрин или мальтодекстрин, G1P получают из растворимого крахмала, амилодекстрина или мальтодекстрина с помощью αGP.
Когда сахариды включают мальтозу, а ферменты включают мальтозофосфорилазу, G1P получают из мальтозы с помощью мальтозофосфорилазы. Если сахариды включают сахарозу, а ферменты включают сахарозофосфорилазу, G1P получают из сахарозы с помощью сахарозофосфорилазы.
В другом варианте осуществления, когда сахариды включают целлобиозу, а ферменты включают целлобиозофосфорилазу, G1P получают из целлобиозы с помощью целлобиозофосфорилазы.
В дополнительном варианте осуществления, когда сахариды содержат целлодекстрины, а ферменты включают целлодекстринфосфорилазу, G1P получают из целлодекстринов с помощью целлодекстринфосфорилазы.
В альтернативном варианте осуществления преобразования сахарида в G1P, когда сахариды включают целлюлозу, а ферменты включают целлюлозофосфорилазу, G1P получают из целлюлозы с помощью целлюлозофосфорилазы.
Согласно изобретению, тагатоза может также быть получена из фруктозы. Пример способа показан на ФИГ. 4. Например, способ включает получение F6P из фруктозы и полифосфата, катализируемое полифосфатфруктокиназой (PPFK); преобразование F6P в T6P, катализируемое F6PE; и преобразование T6P в тагатозу, катализируемое T6PP. Фруктоза может быть получена, например, посредством ферментативного преобразования сахарозы.
В других вариантах осуществления тагатоза может быть получена из сахарозы. Пример такого способа показан на ФИГ.6. Способ предоставляет синтетический путь in vitro, который включает следующие ферментативные стадии: получение G1P из сахарозы и свободного фосфата, катализируемое сахарозофосфорилазой (SP); преобразование G1P в G6P, катализируемое PGM; преобразование G6P в F6P, катализируемое PGI; преобразование F6P в T6P, катализируемое F6PE; и преобразование T6P в тагатозу, катализируемое T6PP.
Фосфат-ионы, получаемые, когда T6P преобразуют в тагатозу, могут потом использоваться повторно на стадии преобразования сахарозы в G1P. Кроме того, как показано на ФИГ. 6, PPFK и полифосфат могут использоваться для увеличения выхода тагатозы за счет получения F6P из фруктозы, полученной посредством фосфоролитического расщепления сахарозы с помощью SP.
В некоторых вариантах осуществления способ получения тагатозы включает следующие стадии: получение глюкозы из полисахаридов и олигосахаридов посредством ферментного гидролиза или кислотного гидролиза, преобразование глюкозы в G6P, катализируемое по меньшей мере одним ферментом, получение фруктозы из полисахаридов и олигосахаридов посредством ферментного гидролиза или кислотного гидролиза и преобразование фруктозы в G6P, катализируемое по меньшей мере одним ферментом. Примеры полисахаридов и олигосахаридов перечислены выше.
В других вариантах осуществления, G6P получают из глюкозы и полифосфата натрия с помощью полифосфатглюкокиназы.
Настоящее раскрытие предоставляет способы преобразования сахаридов, таких как полисахариды и олигосахариды, в виде крахмала, целлюлозы, сахарозы и их производных продуктов, в тагатозу. В некоторых вариантах осуществления предоставлены искусственные (ненатуральные) не имеющие ATP ферментативные способы преобразования крахмала, целлюлозы, сахарозы и их производных продуктов в тагатозу с использованием не содержащих клеток ферментных коктейлей.
Как показано выше, для увеличения выхода G1P могут использоваться некоторые ферменты для гидролиза крахмала. Такие ферменты включают изоамилазу, пуллуланазу и альфа-амилазу. Кукурузный крахмал содержит множество ветвей, которые препятствуют действию αGP. Для деветвления крахмала можно использовать изоамилазу с получением линейного амилодекстрина. Предварительно обработанный изоамилазой крахмал может приводить к более высокой концентрации F6P в конечном продукте. Изоамилаза и пуллуланаза расщепляют альфа-1,6-гликозидные связи, что обеспечивает более полное разложение крахмала альфа-глюканофосфорилазой. Альфа-амилаза расщепляет альфа-1,4-гликозидные связи, вследствие этого альфа-амилазу используют для разложения крахмала на фрагменты для более быстрого преобразования в тагатозу.
Как показано на ФИГ. 2, для повышения выхода тагатозы посредством фосфоролитического расщепления продукта разложения мальтозы на G1P и глюкозу можно использовать мальтозофосфорилазу (MP). В качестве альтернативы, для повышения выхода тагатозы можно использовать 4-глюканотрансферазу (4GT) за счет повторного использования продуктов разложения глюкозы, мальтозы и мальтотриозы на более длинные мальтоолигосахариды; которые могут фосфоролитически расщепляться с помощью αGP для выхода G1P.
Кроме того, целлюлоза является самым богатым биоресурсом и является основным компонентом клеточных стенок растений. Непищевая лигноцеллюлозная биомасса содержит целлюлозу, гемицеллюлозу и лигнин, а также другие второстепенные компоненты. С помощью серии обработок можно получить чистую целлюлозу, включая Avicel (микрокристаллическую целлюлозу), регенерированную аморфную целлюлозу, бактериальную целлюлозу, фильтровальную бумагу и так далее. Частично гидролизованные целлюлозные субстраты включают водонерастворимые целлодекстрины, степень полимеризации которых превышает 7, водорастворимые целлодекстрины со степенью полимеризации 3-6, целлобиозу, глюкозу и фруктозу.
В некоторых вариантах осуществления целлюлоза и полученные из нее продукты могут быть преобразованы в тагатозу посредством ряда стадий. Пример такого способа показан на ФИГ. 3. Способ предоставляет способ синтеза in vitro, который включает следующие стадии: получение G1P из целлодекстрина и целлобиозы и свободного фосфата, катализируемое целлодекстринфосфорилазой (CDP) и целлобиозофосфорилазой (CBP), соответственно; преобразование G1P в G6P, катализируемое PGM; преобразование G6P в F6P, катализируемое PGI; преобразование F6P в T6P, катализируемое F6PE; и преобразование T6P в тагатозу, катализируемое T6PP. В данном способе фосфат-ионы могут использоваться повторно посредством стадии преобразования целлодекстрина и целлобиозы в G1P.
Для гидролиза твердой целлюлозы в водорастворимые целлодекстрины и целлобиозу могут использоваться некоторые ферменты. Такие ферменты включают эндоглюканазу и целлобиогидролазу, но не включают бета-глюкозидазу (целлобиазу).
Перед гидролизом целлюлозы и получением G1P целлюлоза и биомасса могут быть предварительно обработаны для повышения их химической активности и уменьшения степени полимеризации цепей целлюлозы. Способы предварительной обработки целлюлозы и биомассы включают предварительную обработку разбавленной кислотой, фракционирование лигноцеллюлозы на основе растворителя целлюлозы, разрушение целлюлозы аммиаком, промывку водой аммиака, обработку ионной жидкостью и частичный гидролиз с использованием концентрированных кислот, включая хлористоводородную кислоту, серную кислоту, фосфорную кислоту и их комбинации.
В некоторых вариантах осуществления в способ могут быть добавлены полифосфат и полифосфатглюкокиназа (PPGK), повышая таким образом выход тагатозы посредством фосфорилирования продукта разложения глюкозы в G6P, как показано на ФИГ. 3.
В других вариантах осуществления тагатоза может быть получена из глюкозы. Пример такого способа показан на ФИГ.5. Способ включает стадии получения G6P из глюкозы и полифосфата, катализируемого полифосфатглюкокиназой (PPGK); преобразования G6P в F6P, катализируемого PGI; преобразования F6P в T6P, катализируемого F6PE; и преобразования T6P в тагатозу, катализируемого T6PP.
В способе изобретения может использоваться любой подходящий биологический буфер, известный в данной области, такой как HEPES, PBS, BIS-TRIS, MOPS, DIPSO, Trizma и т.д. Реакционный буфер для всех вариантов осуществления может иметь pH, варьирующий от 5,0 до 8,0. Более предпочтительно, pH реакционного буфера может варьировать от приблизительно 6,0 до приблизительно 7,3. Например, pH реакционного буфера может составлять 6,0, 6,2, 6,4, 6,6, 6,8, 7,0, 7,2 или 7,3.
Реакционный буфер также может содержать катионы важнейших металлов. Примеры ионов металлов включают Mg2+ и Zn2+.
Температура реакции, при которой проводят стадии способа, может варьировать от 37 до 85°C. Более предпочтительно, стадии можно проводить при температуре, варьирующей от приблизительно 40°C до приблизительно 70°C. Температура может составлять, например, приблизительно 40°C, приблизительно 45°C, приблизительно 50°C, приблизительно 55°C или приблизительно 60°C. Предпочтительно, температура реакции составляет приблизительно 50°C.
Время реакции раскрытых способов при необходимости может регулироваться и может варьировать от приблизительно 8 часов до приблизительно 48 часов. Например, время реакции может составлять приблизительно 16 часов, приблизительно 18 часов, приблизительно 20 часов, приблизительно 22 часа, приблизительно 24 часа, приблизительно 26 часов, приблизительно 28 часов, приблизительно 30 часов, приблизительно 32 часа, приблизительно 34 часа, приблизительно 36 часов, приблизительно 38 часов, приблизительно 40 часов, приблизительно 42 часа, приблизительно 44 часа, приблизительно 46 часов или приблизительно 48 часов. Более предпочтительно, время реакции составляет приблизительно 24 часов.
В способах согласно изобретению можно достигать высоких выходов вследствие весьма благоприятной константы равновесия для всей реакции. Например, ФИГ. 7 демонстрирует энергию Гиббса реакции между промежуточными продуктами, исходя из образования энергии Гиббса для преобразования глюкозо-1-фосфата в тагатозу. Энергию Гиббса реакций получали, используя http://equilibrator.weizmann.ac.il/. Теоретически, можно достичь выхода до 99%, если исходный материал полностью превратится в промежуточный продукт.
В способах согласно изобретению используют недорогие исходные материалы, и затраты на производство снижают за счет снижения затрат, связанных с разделением сырья и продукта. Крахмал, целлюлоза, сахароза и их производные являются менее дорогостоящими исходными материалами, чем, например, лактоза. Когда тагатозу получают из лактозы, глюкозы и галактозы, а тагатозу отделяют посредством хроматографии, это приводит к более высоким затратам на производство.
Также, стадия преобразования T6P в тагатозу согласно изобретению представляет собой необратимую фосфатазную реакцию, независимо от исходного сырья. Вследствие этого, тагатоза производится с очень высоким выходом, и при этом эффективно минимизируются последующие затраты на разделение продукта.
В противоположность клеточным способам производства, изобретение включает в себя бесклеточное получение тагатозы, имеет относительно высокие скорости реакции вследствие элиминации клеточной мембраны, которая часто замедляет перенос субстрата/продукта в клетку и из нее. Оно также имеет конечный продукт, свободный от богатых питательными веществами ферментационных сред/клеточных метаболитов.
ПРИМЕРЫ
Материалы и Способы
Химические вещества
Все химические вещества, включая кукурузный крахмал, растворимый крахмал, мальтодекстрины, мальтозу, глюкозу, фильтровальную бумагу, имели марку технически чистые или выше и закуплены у Sigma-Aldrich (St. Louis, MO, USA) или Fisher Scientific (Pittsburgh, PA, USA), если не указано иное. Рестрикционные ферменты, T4 лигаза, и ДНК-полимераза Phusion, закупали у New England Biolabs (Ipswich, MA, USA). Олигонуклеотиды были синтезированы либо Integrated DNA Technologies (Coralville, IA, USA), либо Eurofins MWG Operon (Huntsville, AL, USA). Регенерированную аморфную целлюлозу, используемую при ферментативной очистке, получили у Avicel PH105 (FMC BioPolymer, Philadelphia, PA, USA) посредством ее растворения и восстановления, как описано в: Ye et al., Fusion of a family 9 cellulose-binding module improves catalytic potential of Clostridium thermocellum cellodextrin phosphorylase on insoluble cellulose. Appl. Microbiol. Biotechnol. 2011; 92:551-560. Для манипуляций с ДНК в качестве клетки-хозяина использовали Escherichia coli Sig10 (Sigma-Aldrich, St. Louis, MO, USA), а для экспрессии рекомбинантного белка в качестве клетки-хозяина применяли E. coli BL21 (DE3) (Sigma-Aldrich, St. Louis, MO, USA). Для роста клеток E. coli и экспрессии рекомбинантного белка использовали среды ZYM-5052, содержащие либо 100 мг л-1 ампициллина, либо 50 мг л-1 канамицина. Целлюлазу из Trichoderma reesei (Номер по каталогу:C2730) и пуллуланазу (Номер по каталогу:P1067) закупали у Sigma-Aldrich (St. Louis, MO, USA), а производила их Novozymes (Franklinton, NC, USA). Мальтозофосфорилаза (Номер по каталогу:M8284) была закуплена у Sigma-Aldrich.
Производство и очистка рекомбинантных ферментов
Штамм E. coli BL21 (DE3), содержащий плазмиду экспрессии белка, инкубировали в 1 л колбе Эрленмейера со 100 мл среды ZYM-5052, содержащей либо 100 мг л-1 ампициллина, либо 50 мг л-1 канамицина. Клетки выращивали при 37°C с качанием и вращением при 220 об/мин в течение 16-24 часов. Клетки собирали посредством центрифугирования при 12°C и однократно промывали либо 20 мМ HEPES (pH 7,5), содержащего 50 мМ NaCl и 5 мМ MgCl2 (термическое осаждение и целлюлозосвязывающий модуль), либо 20 мМ HEPES (pH 7,5), содержащего 300 мМ NaCl и 5 мМ имидазола (Ni очистка). Клеточные пеллеты повторно суспендировали в том же буфере и лизировали ультразвуком (научно-исследовательский звуковой клеточный дезинтегратор Фишера модель 500; 5 c импульс и 10 с отсутствие импульса, всего 21 мин с 50% амплитудой). После центрифугирования целевые белки в супернатантах очищали.
Для очистки различных рекомбинантных белков использовали три подхода. His-меченые белки очищали с помощью смолы Profinity IMAC Ni-Charged (Bio-Rad, Hercules, CA, USA). Белки слияния, содержащие целлюлозосвязывающий модуль (CBM) и саморасщепляющийся интеин, очищали посредством высокоаффинной адсорбции на регенерированной аморфной целлюлозе с большой площадью поверхности. Для очистки гипертермостабильных ферментов применяли термическое осаждение при 70-95°C в течение 5-30 мин. Чистоту рекомбинантных белков проверяли посредством электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE).
Используемые ферменты и анализ их активности
Использовали альфа-глюканофосфорилазу (αGP) из Thermotoga maritima (Uniprot ID G4FEH8). Активность оценивали в 50 мМ натрий-фосфатного буфера (pH 7,2) содержащего 1 мМ MgCl2, 5 мМ DTT и 30 мМ мальтодекстрина при 50°C. Реакцию останавливали посредством фильтрации фермента концентратором Vivaspin 2 (10000 MWCO) (Vivaproducts, Inc., Littleton, MA, USA). Глюкозо-1-фосфат (G1P) измеряли с использованием набора для анализа глюкозогексокиназа/G6PDH (Sigma Aldrich, Номер по каталогу GAHK20-1KT), обогащенного 25 Ед/мл глюкозофосфомутазы. Единица (U) описана как мкмоль/мин.
Использовали глюкозофосфомутазу (PGM) из Thermococcus kodakaraensis (Uniprot ID Q68BJ6). Активность измеряли в 50 мМ буфера HEPES (pH 7,2), содержащего 5 мМ MgCl2 и 5 мМ G1P при 50°C. Реакцию останавливали посредством фильтрации фермента концентратором Vivaspin 2 (10000 MWCO). Продукт глюкозо-6-фосфат (G6P) определяли, используя набор для анализа гексокиназа/G6PDH (Sigma Aldrich, номер по каталогу GAHK20-1KT).
Использовали два различных источника фосфоглюкоизомеразы (PGI) из Clostridium thermocellum (Uniprot ID A3DBX9) и Thermus thermophilus (Uniprot ID Q5SLL6). Активность измеряли в 50 мМ буфера HEPES (pH 7,2), содержащего 5 мМ MgCl2 и 10 мМ G6P, при 50°C. Реакцию останавливали посредством фильтрации фермента концентратором Vivaspin 2 (10000 MWCO). Продукт, фруктозо-6-фосфат (F6P), определяли, используя сопряженный ферментный анализ фруктозо-6-фосфаткиназа (F6PK)/пируватдегидрогеназа (PK)/лактатдегидрогеназа (LD), в котором уменьшение поглощения при 340 нм указывает на получение F6P. Эти 200 мкл реакционной смеси содержали 50 мМ HEPES (pH 7,2), 5 мМ MgCl2, 10 мМ G6P, 1,5 мМ ATP, 1,5 мМ фосфоенолпирувата, 200 мкМ NADH, 0,1 Ед PGI, 5 Ед PK и 5 Ед LD.
Использовали фруктозо-6-фосфатэпимеразу (F6PE) из Dictyoglomus thermophilum (Uniprot ID B5YBD7). Активность измеряли в 50 мМ буфера HEPES (pH 7,2), содержащего 5 мМ MgCl2 и 10 мМ F6P при 50°C. Реакцию останавливали посредством фильтрации фермента концентратором Vivaspin 2 (10000 MWCO). Продукт, тагатозо-6-фосфат (T6P), определяли, используя тагатозо-6-фосфатфосфатазу и определяя высвобождение свободного фосфата. Для определения высвобождения свободного фосфата 500 мкл раствора, содержащего 0,1 M цинка ацетата и 2 мМ аммония молибдата (pH 5), добавляли к 50 мкл реакционной смеси. Их перемешивали и затем добавляли 125 мкл 5% аскорбиновой кислоты (pH 5). Этот раствор смешивали, затем инкубировали при 30°С в течение 20 мин. Для определения высвобождения свободного фосфата считывали поглощение при 850 нм.
Термофильный F6PE из Anaerolinea thermophila UNI-1 (Uniprot ID: E8N0N6)
Нуклеотидная последовательность (SEQ ID NO.: 1)
ATGTTCGGCTCGCCTGCTCCCCTGCTGGATATGGTCACCGCGCAGAAACAGGGCATGGCGCGGGGTATCCCATCCATTTGTTCGGCACATCCGGTGGTGCTGAGTGCCGCCTGCCATCTTGCCCGCCGGAGCGGCGCGCCCCTGCTCATCGAAACCACCTGCAATCAGGTCAACCACCAAGGTGGGTACAGCGGCATGACCCCCGCCGATTTTGTCCGCTTTCTGCGCGAAATTCTGGAACGGGAAGGTATTCCCCCGCAACAGGTCATCCTGGGCGGGGATCACCTGGGTCCTTACCCCTGGCGGAAAGAGCCTGCCGAAACCGCCATAGCACAAGCGCTGGAAATGGTGCGGGCATACGTGCAGGCAGGCTACACCAAAATTCATCTGGACGCTTCCATGCCCTGCGCCGATGACGACCCCGAGCGTCCCCTGCCGCTGGAGCGCATAGCCCGACGGGCGGCGCAGTTGTGCGCCGCCGCCGAAGCCGCCGCGGGAGCGGTTCAGCCGGTGTACGTAATTGGCAGTGAGGTGCCCCCGCCCGGCGGCGCGCAGGGTCAGGAGGCAAGACTTCACGTCACCACTCCGCAGGAAGCCCAAGCCGCGCTGGATGCCTTTCGGGAAGCCTTTCTGCAGGCAGGCTTGACTCCCGTTTGGGAGCGGGTCATTGCGCTGGTAGTCCAGCCGGGGGTGGAGTTTGGCGTGGACAGCATTCACGCCTATCAGCGCGAAGCCGCCCGCCCGCTGAAGACCTTCATCGAGGGCGTGCCCGGCATGGTGTATGAAGCCCACTCGACCGATTACCAGACCCGTGCCTCCCTGCGTGCGCTGGTGGAAGACCACTTTTCCATTCTCAAGGTTGGTCCGGCACTAACCTTTGCCTACCGCGAAGCCGTGTTCGCCCTGGAACACATCGAACGGGAAATATTGGGCAGGCAGGATATGCCTCTCTCCCGCCTGAGTGAAGTCCTCGACGAGGTGATGCTGAACGATCCACGCCACTGGCAGGGATACTTTGCCGGCGCTCCCGCCGAACAGGCGCTGGCGCGCCGCTACAGTTTCAGCGACCGCATTCGCTATTACTGGCACCATCCCGCCGCGCAGGAAGCCGTGCGGAGACTGCTCGCCAACCTGATCGAAACCCCGCCGCCGCTGAGTTTGCTCAGCCAGTACCTGCCGCGCGAGTATGAGATGGTGCGCGCGGGGGAAATCTCCAGCCACCCGCAGGACCTGATTCGGGCACATATCCAGCACACGCTGGAAGATTACGCTGCGGCGTGCGGGTAA
Аминокислотная последовательность (SEQ ID NO.: 2)
MFGSPAPLLDMVTAQKQGMARGIPSICSAHPVVLSAACHLARRSGAPLLIETTCNQVNHQGGYSGMTPADFVRFLREILEREGIPPQQVILGGDHLGPYPWRKEPAETAIAQALEMVRAYVQAGYTKIHLDASMPCADDDPERPLPLERIARRAAQLCAAAEAAAGAVQPVYVIGSEVPPPGGAQGQEARLHVTTPQEAQAALDAFREAFLQAGLTPVWERVIALVVQPGVEFGVDSIHAYQREAARPLKTFIEGVPGMVYEAHSTDYQTRASLRALVEDHFSILKVGPALTFAYREAVFALEHIEREILGRQDMPLSRLSEVLDEVMLNDPRHWQGYFAGAPAEQALARRYSFSDRIRYYWHHPAAQEAVRRLLANLIETPPPLSLLSQYLPREYEMVRAGEISSHPQDLIRAHIQHTLEDYAAACG
Термофильный F6PE из Caldicellulosiruptor kronotskyensis (Uniprot ID: E4SEH3)
Нуклеотидная последовательность (SEQ ID NO.: 3)
ATGAGTCCTCAAAATCCATTGATTGGTTTATTTAAGAATAGAGAAAAAGAGTTTAAGGGTATTATTTCAGTTTGTTCTTCAAATGAAATAGTCTTAGAAGCAGTTTTAAAAAGAATGAAAGATACAAACCTACCAATTATTATTGAAGCCACAGCGAACCAGGTAAATCAATTTGGCGGGTATTCTGGGTTGACACCGTCTCAGTTCAAAGAACGAGTTATAAAAATTGCTCAAAAAGTTGATTTTCCACTTGAGAGAATAATTCTTGGTGGGGACCATCTTGGACCATTTGTGTGGCGTGACCAGGAACCAGAAATTGCTATGGAGTATGCTAAGCAAATGATAAAAGAATACATAAAAGCAGGTTTTACCAAAATTCACATCGACACGAGTATGCCTTTAAAAGGGGAGAACAGCATAGATGATGAAATAATTGCTAAAAGAACTGCTGTGCTCTGCAGGATTGCGGAGGAGTGTTTTGAGAAGATTTCTATAAACAATCCCTATATTACAAGGCCAGTTTATGTGATAGGAGCTGATGTGCCACCTCCCGGCGGAGAGTCTTCTATTTGTCAAACAATTACTACTAAAGATGAATTAGAAAGAAGTTTAGAATATTTCAAAGAAGCATTTAAAAAGGAAGGAATTGAGCATGTATTCGATTATGTAGTTGCTGTTGTTGCAAATTTTGGAGTTGAATTTGGGAGCGATGAAATTGTTGATTTTGATATGGAAAAAGTAAAGCCGCTAAAAGAACTTTTGGCAAAGTACAATATAGTATTTGAAGGCCATTCTACAGATTATCAAACAAAAGAAAACTTAAAAAGAATGGTCGAATGTGGTATTGCAATTTTAAAGGTTGGTCCTGCTCTAACATTTACATTGCGCGAAGCGTTAGTAGCACTTAGTCATATTGAAGAAGAAATTTATAGCAATGAAAAGGAGAAACTGTCAAGATTTAGAGAAGTTTTATTGAATACTATGCTAACATGCAAAGATCACTGGAGTAAATATTTTGATGAGAATGATAAGTTAATTAAGTCAAAGCTCCTATATAGCTATCTTGACAGATGGAGATACTATTTTGAAAACGAGAGTGTGAAAAGTGCTGTTTATTCTCTTATTGGAAATTTAGAGAATGTTAAAATTCCACCTTGGCTTGTAAGTCAGTATTTTCCTTCTCAGTACCAAAAGATGAGAAAAAAAGATTTAAAAAACGGTGCTGCCGACCTAATATTGGATAAAATAGGGGAAGTCATTGACCATTATGTTTATGCGGTAAAAGAATAA
Аминокислотная последовательность (SEQ ID NO.: 4)
MSPQNPLIGLFKNREKEFKGIISVCSSNEIVLEAVLKRMKDTNLPIIIEATANQVNQFGGYSGLTPSQFKERVIKIAQKVDFPLERIILGGDHLGPFVWRDQEPEIAMEYAKQMIKEYIKAGFTKIHIDTSMPLKGENSIDDEIIAKRTAVLCRIAEECFEKISINNPYITRPVYVIGADVPPPGGESSICQTITTKDELERSLEYFKEAFKKEGIEHVFDYVVAVVANFGVEFGSDEIVDFDMEKVKPLKELLAKYNIVFEGHSTDYQTKENLKRMVECGIAILKVGPALTFTLREALVALSHIEEEIYSNEKEKLSRFREVLLNTMLTCKDHWSKYFDENDKLIKSKLLYSYLDRWRYYFENESVKSAVYSLIGNLENVKIPPWLVSQYFPSQYQKMRKKDLKNGAADLILDKIGEVIDHYVYAVKE
Термофильный F6PE из Caldilinea aerophila (Uniprot ID: I0I507)
Нуклеотидная последовательность (SEQ ID NO.: 5)
ATGTCAACACTTCGCCACATCATTTTGCGACTGATCGAGCTGCGTGAACGAGAACAGATCCATCTCACGCTGCTGGCCGTCTGTCCCAACTCGGCGGCGGTGCTGGAGGCAGCGGTGAAGGTCGCCGCGCGCTGCCACACGCCGATGCTCTTCGCTGCCACGCTCAATCAAGTCGATCGCGACGGCGGCTACACCGGTTGGACGCCTGCGCAATTCGTCGCCGAGATGCGTCGCTATGCCGTCCGCTATGGCTGCACCACCCCGCTCTATCCTTGCCTGGATCACGGCGGGCCGTGGCTCAAAGATCGCCATGCACAGGAAAAGCTACCGCTCGACCAGGCGATGCATGAGGTCAAGCTGAGCCTCACCGCCTGTCTGGAGGCCGGCTACGCGCTGCTGCACATCGACCCCACGGTCGATCGCACGCTCCCGCCCGGAGAAGCGCCGCTCGTGCCGATCGTCGTCGAGCGCACGGTCGAGCTGATCGAACATGCCGAACAGGAGCGACAGCGGCTGAACCTGCCGGCGGTCGCCTATGAAGTCGGCACCGAAGAAGTACATGGCGGGCTGGTGAATTTCGACAATTTTGTCGCCTTCTTGGATTTGCTCAAGGCAAGGCTTGAACAACGTGCCCTGATGCACGCCTGGCCCGCCTTCGTGGTGGCGCAGGTCGGCACTGACCTGCATACAACGTATTTTGACCCCAGTGCGGCGCAACGGCTGACTGAGATCGTGCGCCCTACCGGTGCACTGTTGAAGGGGCACTACACCGACTGGGTCGAAAATCCCGCCGACTATCCGAGGGTAGGCATGGGAGGCGCCAACGTTGGTCCAGAGTTTACGGCGGCCGAGTTCGAGGCGCTGGAAGCGCTGGAACGGCGGGAACAACGGCTGTGCGCCAACCGGAAATTGCAGCCCGCCTGTTTTTTGGCTGCACTGGAAGAGGCAGTAGTCGCTTCAGATCGTTGGCGGAAGTGGCTCCAGCCCGATGAGATCGGCAAGCCCTTTGCAGAATTAACGCCCGCACGCCGGCGCTGGCTCGTGCAGACCGGGGCACGCTACGTCTGGACTGCGCCGAAAGTTATCGCCGCACGCGAACAGCTCTATGCGCACCTCTCCCTTGTGCAGGCGGATCCACATGCCTACGTGGTAGAGTCAGTCGCCCGGTCAATCGAGCGCTATATCGATGCCTTCAACTTATACGACGCCGCTACATTGCTTGGATGA
Аминокислотная последовательность (SEQ ID NO.: 6)
MSTLRHIILRLIELREREQIHLTLLAVCPNSAAVLEAAVKVAARCHTPMLFAATLNQVDRDGGYTGWTPAQFVAEMRRYAVRYGCTTPLYPCLDHGGPWLKDRHAQEKLPLDQAMHEVKLSLTACLEAGYALLHIDPTVDRTLPPGEAPLVPIVVERTVELIEHAEQERQRLNLPAVAYEVGTEEVHGGLVNFDNFVAFLDLLKARLEQRALMHAWPAFVVAQVGTDLHTTYFDPSAAQRLTEIVRPTGALLKGHYTDWVENPADYPRVGMGGANVGPEFTAAEFEALEALERREQRLCANRKLQPACFLAALEEAVVASDRWRKWLQPDEIGKPFAELTPARRRWLVQTGARYVWTAPKVIAAREQLYAHLSLVQADPHAYVVESVARSIERYIDAFNLYDAATLLG
Термофильный F6PE из Caldithrix abyssi (Uniprot ID: H1XRG1)
Нуклеотидная последовательность (SEQ ID NO.: 7)
ATGAGTCTGCATCCTTTAAATAAATTAATCGAGCGACACAAAAAAGGAACGCCGGTCGGTATTTATTCCGTCTGTTCGGCCAATCCCTTTGTTTTGAAAGCGGCCATGCTACAGGCGCAAAAGGATCAGTCTTTGCTACTTATTGAGGCCACTTCCAACCAGGTAGATCAATTCGGCGGTTACACCGGCATGCGGCCCGAAGATTTTAAAACAATGACGCTTGAACTGGCAGCCGAAAACAATTACGATCCACAGGGATTAATCCTGGGCGGCGACCATCTGGGGCCCAACCGCTGGACAAAACTGAGCGCCTCCCGGGCCATGGACTACGCCAGAGAGCAGATTGCCGCTTATGTTAAAGCCGGCTTTTCCAAAATCCACTTAGACGCCACCATGCCCTTGCAAAACGATGCCACAGATTCCGCCGGCCGCCTTCCAGTCGAAACAATCGCTCAACGTACCGCAGAATTATGCGCCGTGGCCGAACAAACTTACCGGCAGAGCGACCAACTCTTTCCGCCGCCTGTTTACATTGTCGGCAGCGACGTGCCCATCCCGGGCGGCGCGCAAGAAGCGCTGAACCAGATCCATATTACGGAGGTAAAAGAGGTTCAACAGACCATTGATCACGTGCGGCGGGCCTTTGAAAAAAACGGCCTGGAAGCGGCTTACGAAAGAGTTTGCGCCGTTGTCGTGCAGCCAGGCGTTGAATTCGCCGATCAAATCGTTTTTGAATACGCTCCCGACAGAGCGGCGGCCTTAAAAGATTTTATTGAAAGCCATTCGCAGCTGGTTTATGAAGCGCACTCTACTGATTACCAGACCGCACCTCTTTTGCGCCAGATGGTAAAAGATCACTTTGCCATTTTAAAGGTCGGGCCTGCGCTCACCTTTGCCCTGCGCGAAGCCATTTTTGCTCTGGCCTTTATGGAAAAAGAGCTTTTGCCATTGCACAGAGCGCTCAAACCTTCTGCCATTCTGGAAACGCTGGACCAAACGATGGACAAAAACCCTGCTTACTGGCAAAAGCATTACGGCGGAACAAAGGAAGAAGTACGCTTTGCGCAGCGGTTTAGCCTGAGCGACCGCATTCGTTACTACTGGCCGTTTCCAAAGGTTCAAAAGGCCCTGCGCCAATTGCTAAAAAACTTGCAACAAATTTCCATTCCTCTAACTTTGGTAAGCCAGTTCATGCCAGAGGAATACCAACGTATTCGCCAAGGAACGTTAACCAACGATCCGCAGGCGCTGATTTTGAACAAAATTCAAAGCGTATTAAAGCAATACGCGGAGGCGACGCAAATTCAAAACTCTTTGACATTCACGCAAAATCAAAATTCATTAGCAATGGAGCGACTATGA
Аминокислотная последовательность (SEQ ID NO.: 8)
MSLHPLNKLIERHKKGTPVGIYSVCSANPFVLKAAMLQAQKDQSLLLIEATSNQVDQFGGYTGMRPEDFKTMTLELAAENNYDPQGLILGGDHLGPNRWTKLSASRAMDYAREQIAAYVKAGFSKIHLDATMPLQNDATDSAGRLPVETIAQRTAELCAVAEQTYRQSDQLFPPPVYIVGSDVPIPGGAQEALNQIHITEVKEVQQTIDHVRRAFEKNGLEAAYERVCAVVVQPGVEFADQIVFEYAPDRAAALKDFIESHSQLVYEAHSTDYQTAPLLRQMVKDHFAILKVGPALTFALREAIFALAFMEKELLPLHRALKPSAILETLDQTMDKNPAYWQKHYGGTKEEVRFAQRFSLSDRIRYYWPFPKVQKALRQLLKNLQQISIPLTLVSQFMPEEYQRIRQGTLTNDPQALILNKIQSVLKQYAEATQIQNSLTFTQNQNSLAMERL
Термофильный F6PE из Dictyoglomus thermophilum (Uniprot ID: B5YBD7)
Нуклеотидная последовательность (SEQ ID NO.: 9)
ATGTGGCTTAGTAAAGATTATTTGAGAAAAAAGGGAGTTTATTCTATATGTAGCTCTAATCCATATGTGATTGAGGCAAGTGTTGAATTTGCTAAGGAGAAGAATGATTATATTTTAATTGAGGCGACACCTCATCAGATAAACCAGTTTGGTGGATATTCAGGTATGACTCCCGAAGATTTTAAAAACTTTGTAATGGGAATAATAAAAGAAAAGGGAATAGAAGAGGATAGGGTGATTCTTGGAGGGGACCATTTAGGCCCTCTCCCTTGGCAAGATGAACCTTCTTCTTCTGCAATGAAAAAGGCAAAAGACCTTATAAGGGCCTTTGTGGAGAGTGGTTATAAGAAGATACACCTTGATTGTAGTATGTCTCTTTCTGATGATCCTGTAGTGCTCTCTCCCGAGAAGATAGCAGAAAGGGAGAGGGAACTTCTTGAGGTTGCAGAAGAGACTGCTAGAAAGTACAATTTTCAGCCTGTGTATGTGGTGGGAACTGATGTACCGGTAGCTGGAGGAGGCGAAGAGGAAGGTATTACCTCAGTGGAGGATTTTAGAGTAGCAATCTCCTCTTTAAAAAAATATTTTGAGGATGTTCCAAGGATATGGGATAGGATAATTGGTTTTGTAATAATGCTTGGTATAGGTTTTAATTATGAAAAAGTGTTTGAGTATGACAGGATTAAGGTGAGAAAAATTTTAGAGGAGGTAAAGAAAGAGAATCTTTTTGTTGAAGGTCACTCTACTGACTATCAGACAAAACGTGCATTGAGAGATATGGTAGAGGATGGAGTAAGAATTCTTAAGGTTGGTCCTGCTTTAACAGCAAGTTTTAGAAGGGGAGTATTTTTATTAAGTAGCATTGAGGATGAGCTTATATCGGAAGATAAAAGGTCTAATATTAAGAAAGTTGTGCTTGAGACTATGTTAAAAGATGATAAATATTGGAGAAAGTATTATAAGGATTCAGAAAGATTAGAATTAGATATTTGGTACAACTTACTTGATAGGATTAGATATTATTGGGAATATAAAGAGATAAAAATAGCTTTAAATAGGCTTTTTGAAAATTTTTCGGAAGGGGTTGATATTAGATACATCTATCAATATTTTTATGATTCGTATTTTAAAGTAAGAGAAGGAAAAATAAGAAATGATCCAAGGGAGCTAATAAAGAATGAAATAAAGAAGGTCTTGGAGGACTATCACTATGCTGTAAACTTATAA
Кодон-оптимизированная нуклеотидная последовательность (SEQ ID NO.: 10)
ATGTGGCTGAGCAAGGACTACCTGCGTAAGAAGGGCGTTTACAGCATTTGCAGCAGCAACCCGTATGTTATTGAAGCGAGCGTGGAGTTCGCGAAGGAGAAAAACGATTACATCCTGATTGAAGCGACCCCGCACCAGATCAACCAATTTGGTGGCTATAGCGGCATGACCCCGGAGGACTTCAAGAACTTTGTTATGGGCATCATTAAGGAAAAAGGTATCGAGGAAGACCGTGTGATTCTGGGTGGCGATCACCTGGGTCCGCTGCCGTGGCAGGATGAGCCGAGCAGCAGCGCGATGAAGAAAGCGAAAGACCTGATCCGTGCGTTCGTTGAAAGCGGTTACAAGAAAATTCACCTGGATTGCAGCATGAGCCTGAGCGACGATCCGGTGGTTCTGAGCCCGGAGAAGATCGCGGAACGTGAGCGTGAACTGCTGGAAGTTGCGGAGGAAACCGCGCGTAAATACAACTTTCAACCGGTGTATGTGGTGGGTACCGATGTTCCGGTTGCGGGTGGCGGTGAGGAAGAGGGTATCACCAGCGTGGAGGACTTCCGTGTTGCGATTAGCAGCCTGAAGAAATACTTTGAAGACGTTCCGCGTATTTGGGATCGTATCATTGGTTTCGTGATCATGCTGGGCATTGGTTTCAACTACGAGAAGGTGTTTGAATATGATCGTATCAAAGTGCGTAAAATTCTGGAAGAGGTTAAGAAAGAGAACCTGTTTGTGGAAGGCCACAGCACCGACTATCAGACCAAGCGTGCGCTGCGTGACATGGTGGAGGATGGCGTTCGTATCCTGAAAGTGGGTCCGGCGCTGACCGCGAGCTTCCGTCGTGGTGTGTTTCTGCTGAGCAGCATCGAGGACGAACTGATTAGCGAGGATAAACGTAGCAACATTAAGAAAGTGGTTCTGGAAACCATGCTGAAGGACGATAAATACTGGCGTAAGTACTATAAAGACAGCGAGCGTCTGGAACTGGATATCTGGTACAACCTGCTGGACCGTATTCGTTACTACTGGGAGTACAAGGAAATCAAGATTGCGCTGAACCGTCTGTTCGAGAACTTTAGCGAAGGCGTTGATATCCGTTACATCTACCAATACTTCTACGACAGCTACTTCAAAGTGCGTGAGGGTAAAATCCGTAACGACCCGCGTGAACTGATTAAGAACGAGATTAAGAAAGTGCTGGAAGACTACCATTATGCGGTGAACCTGTAA
Аминокислотная последовательность (SEQ ID NO.: 11)
MWLSKDYLRKKGVYSICSSNPYVIEASVEFAKEKNDYILIEATPHQINQFGGYSGMTPEDFKNFVMGIIKEKGIEEDRVILGGDHLGPLPWQDEPSSSAMKKAKDLIRAFVESGYKKIHLDCSMSLSDDPVVLSPEKIAERERELLEVAEETARKYNFQPVYVVGTDVPVAGGGEEEGITSVEDFRVAISSLKKYFEDVPRIWDRIIGFVIMLGIGFNYEKVFEYDRIKVRKILEEVKKENLFVEGHSTDYQTKRALRDMVEDGVRILKVGPALTASFRRGVFLLSSIEDELISEDKRSNIKKVVLETMLKDDKYWRKYYKDSERLELDIWYNLLDRIRYYWEYKEIKIALNRLFENFSEGVDIRYIYQYFYDSYFKVREGKIRNDPRELIKNEIKKVLEDYHYAVNL
Применяли тагатозо-6-фосфатфосфатазу (T6PP) из Archaeoglobus fugidis (Uniprot ID O29805). Активность измеряли в 50 мМ буфера HEPES (pH 7,2), содержащего 5 мМ MgCl2 и 10 мМ T6P при 50°C. Реакцию останавливали посредством фильтрации фермента концентратором Vivaspin 2 (10000 MWCO). Производство тагатозы определяли, определяя высвобождение свободного фосфата, как описано для F6PE.
Термофильный T6PP из Archaeoglobus fulgidus (Uniprot ID:O29805)
Нуклеотидная последовательность (SEQ ID NO.: 12)
ATGTTCAAACCAAAGGCCATCGCAGTTGACATAGATGGCACCCTCACCGACAGAAAGAGGGCTCTGAACTGCAGGGCTGTTGAAGCTCTCCGCAAGGTAAAAATTCCCGTGATTTTGGCCACTGGTAACATATCTTGTTTTGCGAGGGCTGCAGCAAAGCTGATTGGAGTCTCAGACGTGGTAATCTGCGAGAATGGGGGCGTGGTGAGGTTCGAGTACGATGGGGAGGATATTGTTTTAGGAGATAAAGAGAAATGCGTTGAGGCTGTGAGGGTGCTTGAGAAACACTATGAGGTTGAGCTGCTGGACTTCGAATACAGGAAGTCGGAAGTGTGCATGAGGAGGAGCTTTGACATCAACGAGGCGAGAAAGCTCATTGAGGGGATGGGGGTTAAGCTTGTGGATTCAGGCTTTGCCTACCACATTATGGATGCTGATGTTAGCAAGGGAAAAGCTTTGAAGTTCGTTGCCGAGAGGCTTGGTATCAGTTCAGCGGAGTTTGCAGTTATCGGCGACTCAGAGAACGACATAGACATGTTCAGAGTTGCTGGATTCGGAATTGCTGTTGCCAATGCCGATGAGAGGCTGAAGGAGTATGCTGATTTAGTTACGCCATCACCAGACGGCGAGGGGGTTGTTGAGGCTTTGCAGTTTCTGGGATTGTTGCGGTGA
Аминокислотная последовательность (SEQ ID NO.: 13)
MFKPKAIAVDIDGTLTDRKRALNCRAVEALRKVKIPVILATGNISCFARAAAKLIGVSDVVICENGGVVRFEYDGEDIVLGDKEKCVEAVRVLEKHYEVELLDFEYRKSEVCMRRSFDINEARKLIEGMGVKLVDSGFAYHIMDADVSKGKALKFVAERLGISSAEFAVIGDSENDIDMFRVAGFGIAVANADERLKEYADLVTPSPDGEGVVEALQFLGLLR
T6PP from Archaeoglobus profundus (Uniprot ID D2RHV2_ARCPA)
Нуклеотидную последовательность (SEQ ID NO.: 14)
GTGTTCAAGGCTTTGGTAGTTGATATAGACGGAACTTTGACGGATAAGAAGAGGGCAATAAACTGCAGAGCGGTCGAAGCACTTAGAAAACTAAAGATTCCTGTTGTCTTGGCAACCGGAAACATTTCATGCTTTGCAAGGGCTGTAGCTAAGATTATAGGTGTTTCCGATATTGTAATAGCTGAGAACGGAGGTGTTGTCAGATTCAGCTACGACGGAGAGGACATAGTTCTGGGGGATAGAAGTAAATGCTTAAGAGCTTTGGAGACACTTAGAAAACGCTTCAAAGTAGAGCTTCTCGACAACGAATATAGGAAGTCTGAGGTCTGCATGAGGAGGAACTTCCCTATAGAGGAAGCTAGAAAGATACTGCCAAAAGATGTTAGAATAGTCGATACAGGCTTCGCATACCACATAATCGATGCAAATGTCAGCAAGGGGAAGGCTTTGATGTTCATAGCCGATAAGCTTGGCTTGGACGTTAAGGATTTCATTGCGATAGGTGATTCCGAAAACGACATTGAAATGTTGGAAGTTGCAGGTTTTGGCGTTGCAGTTGCGAATGCGGATGAAAAGCTTAAGGAGGTAGCGGATTTGGTCACATCGAAGCCTAATGGAGACGGAGTTGTCGAAGCTCTTGAGTTCTTGGGACTCATTTAG
Аминокислотную последовательность (SEQ ID NO.: 15)
MFKALVVDIDGTLTDKKRAINCRAVEALRKLKIPVVLATGNISCFARAVAKIIGVSDIVIAENGGVVRFSYDGEDIVLGDRSKCLRALETLRKRFKVELLDNEYRKSEVCMRRNFPIEEARKILPKDVRIVDTGFAYHIIDANVSKGKALMFIADKLGLDVKDFIAIGDSENDIEMLEVAGFGVAVANADEKLKEVADLVTSKPNGDGVVEALEFLGLI
T6PP из Archaeoglobus veneficus (Uniprot ID F2KMK2_ARCVS)
Нуклеотидная последовательность (SEQ ID NO.: 16)
ATGCTCCGTCCAAAGGGTCTCGCCATTGACATCGACGGAACCATAACATACAGGAATCGAAGCCTGAACTGTAAGGCCGTTGAAGCTCTCAGGAAGGTAAAAATCCCTGTAGTTCTTGCAACTGGCAACATATCCTGTTTCGCAAGAACTGCTGCAAAGCTTATAGGCGTCTCAGACATTGTTATATGCGAAAATGGAGGTATTGTTCGATTCAGCTACGATGGCGACGACATAGTGCTTGGGGACATAAGCAAATGCCTTAAAGCGGCTGAAATTCTCAAAGAGTACTTTGAAATCGAATTCCTTGACGCTGAGTACAGGAAGTCGGAGGTCTGTCTTCGCAGAAACTTTCCTATTGAAGAGGCGAGGAAAATTCTTCACGATGCAAAGCTTGATGTTAAAATCGTCGATTCAGGTTTTGCGTACCACATAATGGATGCGAAGGTCAGCAAAGGAAGGGCTCTTGAGTACATAGCTGATGAACTTGGTATAAGTCCGAAGGAGTTCGCTGCAATTGGTGATTCTGAGAACGACATAGACCTGATTAAGGCTGCCGGCCTCGGTATTGCCGTTGGAGATGCTGACTTAAAGCTCAAAATGGAGGCCGACGTGGTAGTCTCGAAGAAGAATGGCGATGGAGTTGTTGAAGCACTTGAGCTTCTGGGCTTAATTTAA
Аминокислотная последовательность (SEQ ID NO.: 17)
MLRPKGLAIDIDGTITYRNRSLNCKAVEALRKVKIPVVLATGNISCFARTAAKLIGVSDIVICENGGIVRFSYDGDDIVLGDISKCLKAAEILKEYFEIEFLDAEYRKSEVCLRRNFPIEEARKILHDAKLDVKIVDSGFAYHIMDAKVSKGRALEYIADELGISPKEFAAIGDSENDIDLIKAAGLGIAVGDADLKLKMEADVVVSKKNGDGVVEALELLGLI
Рекомбинантная целлодекстринфосфорилаза и целлобиозофосфорилаза из C. thermocellum описаны у Ye et al. Спонтанное получение с высоким выходом водорода из целлюлозных материалов и воды катализировалось ферментными коктейлями. ChemSusChem 2009; 2:149-152. Их активность анализировали, как описано.
Рекомбинантная полифосфатглюкокиназа из Thermobifida fusca YX описана у Liao et al., One-step purification and immobilization of thermophilic polyphosphate glucokinase from Thermobifida fusca YX: glucose-6-phosphate выработке without ATP. Appl. Microbiol. Biotechnol. 2012; 93:1109-1117. Ее активность анализировали, как описано.
Рекомбинантная изоамилаза из Sulfolobus tokodaii описана у Cheng et al., Doubling power output of starch biobattery treated by the most thermostable isoamylase from an archaeon Sulfolobus tokodaii. Scientific Reports 2015; 5:13184. Ее активность анализировали, как описано.
Рекомбинантная 4-альфа-глюканолтрансфераза из Thermococcus litoralis описана у Jeon et al. 4-α-Glucanotransferase from the Hyperthermophilic Archaeon Thermococcus Litoralis. Eur. J. Biochem. 1997; 248:171-178. Ее активность измеряли, как описано.
Применяли сахарозофосфорилазу из Caldithrix abyssi (Uniprot H1XT50). Ее активность измеряли в 50 мМ буфера HEPES (pH 7,5) содержащем 10 мМ сахарозы и 12 мМ органического фосфата. Глюкозо-1-фосфат (G1P) измеряли с использованием набора для анализа глюкозогексокиназа/G6PDH, обогащенного 25 Ед/мл глюкозофосфомутазой, как и в случае альфа-глюканофосфорилазы.
Ферментные единицы, используемые в каждом примере ниже, могут быть увеличены или уменьшены, чтобы отрегулировать время реакции при необходимости. Например, если бы было нужно выполнить пример 9 за 8 ч вместо 24 ч, единицы ферментов нужно было бы увеличить примерно в 3 раза. И наоборот, если бы было нужно выполнить пример 9 за 48 часов вместо 24 часов, то ферментные единицы нужно было бы уменьшить примерно в 2 раза. Эти примеры иллюстрируют, как количество ферментных единиц можно использовать для увеличения или уменьшения времени реакции при сохранении постоянной производительности.
Пример 1
Для подтверждения технической возможности ферментативного биосинтеза фруктозо-6-фосфата из крахмала рекомбинантно экспрессировали три фермента: альфа-глюканофосфорилазу из T. maritima (Uniprot ID G4FEH8), глюкозофосфомутазу из Thermococcus kodakaraensis (Uniprot ID Q68BJ6) и фосфоизомеразу из Clostridium thermocellum (Uniprot ID A3DBX9). Рекомбинантные белки сверхэкспрессировали в E. coli BL21 (DE3) и очищали, как описано выше.
0,20 мл реакционной смеси, содержащей 10 г/л растворимого крахмала, 50 мМ забуференного фосфатом физиологического раствора с pH 7,2, 5 мМ MgCl2, 0,5 мМ ZnCl2, 0,01 ед αGP, 0,01 ед PGM и 0,01 ед PGI инкубировали при 50°C в течение 24 часов. Реакцию останавливали посредством фильтрации фермента концентратором Vivaspin 2 (10000 MWCO). Продукт, фруктозо-6-фосфат (F6P), определяли, используя сопряженный ферментный анализ фруктозо-6-фосфаткиназа (F6PK)/пируватдегидрогеназа (PK)/лактатдегидрогеназа (LD), в котором уменьшение поглощения при 340 нм указывает на производство F6P, как описано выше. Итоговая концентрация F6P после 24 часов составила 3,6 г/л.
Пример 2
Те же тесты, что и в примере 1 (с другими температурами реакции) проводили от 40 до 80°C. Было обнаружено, что 10 г/л растворимого крахмала производили 0,9 г/л F6P при 40°C и 3,6 г/л F6P при 80°C после 40 часовых реакций. Эти результаты свидетельствуют о том, что повышение температуры реакции для этого набора ферментов увеличивает выход F6P, но слишком высокая температура может немного ухудшить ферментную активность.
Пример 3
Было обнаружено, что при 80°C соотношение ферментов αGP: PGM: PGI, составляющее приблизительно 1:1:1, приводит к быстрой выработке F6P. Было отмечено, что соотношение ферментов не оказывало сильного влияния на итоговую концентрацию F6P, если время реакции было достаточно продолжительным. Однако, соотношение ферментов влияет на скорость реакции и общую стоимость ферментов, используемых в системе.
Пример 4
0,20 мл реакционной смеси, содержащей 10 г/л мальтодекстрина, 50 мМ забуференного фосфатом физиологического раствора с pH 7,2, 5 мМ MgCl2, 0,5 мМ ZnCl2, 0,01 ед αGP, 0,01 ед PGM и 0,01 ед PGI, инкубировали при 50°C в течение 24 часов. Реакцию останавливали посредством фильтрации фермента концентратором Vivaspin 2 (10000 MWCO). Продукт, фруктозо-6-фосфат (F6P), определяли, используя сопряженный ферментный анализ фруктозо-6-фосфаткиназа (F6PK)/пируватдегидрогеназа (PK)/лактатдегидрогеназа (LD), в котором уменьшение поглощения при 340 нм указывает на производство F6P, как описано выше. Итоговая концентрация F6P после 24 часов составила 3,6 г/л.
Пример 5
Для тестирования получения F6P от Avicel, Sigma, для гидролиза целлюлозы при 50°C применяли целлюлазу. Для удаления бета-глюкозидазы из коммерческой целлюлазы 10 единиц фильтровальной бумаги/мл целлюлазы смешивали с 10 г/л Avicel в ванне с ледяной водой в течение 10 мин. После центрифугирования при 4°C, супернатант, содержащий бета-глюкозидазу, декантировали. Avicel, который был связан с целлюлазой, содержащей эндоглюканазу и целлобиогидролазу, ресуспендировали в цитратном буфере (pH 4,8) для гидролиза при 50°C в течение трех дней. Гидролизат целлюлозы смешивали с 5 Ед/мл целлодекстринфосфорилазы, 5 Ед/л целлобиозофосфорилазы, 5 Ед/мл αGP, 5 Ед/мл PGM и 5 Ед/мл PGI в 100 мМ буфера HEPES (pH 7,2), содержащего 10 мМ фосфата, 5 мМ MgCl2 и 0,5 мМ ZnCl2. Реакцию проводили при 60°C в течение 72 часов и обнаруживали высокие концентрации F6P (небольшие количества глюкозы и отсутствие целлобиозы). F6P определяли с использованием сопряженного ферментного анализа, описанного выше. Глюкозу определяли с использованием набора для анализа гексокиназа/G6PDH, как описано выше.
Пример 6
Для увеличения выхода F6P из Avicel, Avicel предварительно обрабатывали концентрированной фосфорной кислотой для получения аморфной целлюлозы (RAC), как описано у Zhang et al. A transition from cellulose swelling to cellulose dissolution by o-phosphoric acid: evidence from enzymatic hydrolysis and supramolecular structure. Biomacromolecules 2006; 7:644-648. Для удаления бета-глюкозидазы из коммерческой целлюлазы 10 единиц фильтровальной бумаги/мл целлюлазы смешивали с 10 г/л RAC в ванне с ледяной водой в течение 5 мин. После центрифугирования при 4°C супернатант, содержащий бета-глюкозидазу, декантировали. RAC, который был связан с целлюлазой, содержащей эндоглюканазу и целлобиогидролазу, ресуспендировали в цитратном буфере (pH 4,8) для гидролиза при 50°C в течение 12 часов. Гидролизат RAC смешивали с 5 Ед/мл целлодекстринфосфорилазы, 5 ЕД/Л целлобиозофосфорилазы, 5 Ед/мл αGP, 5 Ед/мл PGM и 5 Ед/мл PGI в 100 мМ буфера HEPES (pH 7,2), содержащего 10 мМ фосфата, 5 мМ MgCl2 и 0,5 мМ ZnCl2. Реакцию проводили при 60°C в течение 72 часов. Получали высокие концентрации F6P и глюкозы, поскольку не добавляли ферменты для преобразования глюкозы в F6P. F6P определяли с использованием сопряженного ферментного анализа, описанного выше. Глюкозу определяли с использованием набора для анализа гексокиназа/G6PDH, как описано выше.
Пример 7
Для дополнительного увеличения выхода F6P из RAC добавляли полифосфатглюкокиназу и полифосфат. Для удаления бета-глюкозидазы из коммерческой целлюлазы 10 единиц фильтровальной бумаги/мл целлюлазы смешивали с 10 г/л RAC в ванне с ледяной водой в течение 5 мин. После центрифугирования при 4°C супернатант, содержащий бета-глюкозидазу, декантировали. RAC, который был связан с целлюлазой, содержащей эндоглюканазу и целлобиогидролазу, ресуспендировали в цитратном буфере (pH 4,8) для гидролиза при 50°C, инкубировали в цитратном буфере (pH 4,8) для гидролиза при 50°C в течение 12 часов. Гидролизат RAC смешивали с 5 Ед/мл полифосфатглюкокиназы, 5 Ед/мл целлодекстринфосфорилазы, 5 Ед/мл целлобиозофосфорилазы, 5 Ед/мл αGP, 5 Ед/мл PGM и 5 Ед/мл PGI в 100 мМ буфера HEPES (pH 7,2), содержащего 50 мМ полифосфата, 10 мМ фосфата, 5 мМ MgCl2 и 0,5 мМ ZnCl2. Реакцию проводили при 50°C в течение 72 часов. F6P обнаруживали в высоких концентрациях при наличии лишь небольших количеств глюкозы. F6P определяли с использованием сопряженного ферментного анализа, описанного выше. Глюкозу определяли с использованием набора для анализа гексокиназы/G6PDH, как описано выше.
Пример 8
Для подтверждения получения тагатозы из F6P, 2 г/л F6P смешивали с 1 Ед/мл фруктозо-6-фосфатэпимеразой (F6PE) и 1 Ед/мл тагатозо-6-фосфатфосфатазы (T6PP) в 50 мМ буфера HEPES (pH 7,2), содержащего 5 мМ MgCl2. Реакцию инкубировали в течение 16 часов при 50°C. 100% преобразование F6P в тагатозу наблюдали посредством HPLC (серия Agilent 1100) с использованием Agilent Hi-Plex H-column и рефрактометрического детектора. Образец обрабатывали в 5 мМ H2SO4 при 0,6 мл/мин.
Пример 9
Для подтверждения получения тагатозы из мальтодекстрина 0,20 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, 50 мМ забуференного фосфатом физиологического раствора с pH 7,2, 5 мМ MgCl2, 0,05 ед αGP, 0,05 ед PGM, 0,05 ед PGI, 0,05 ед F6PE и 0,05 ед T6PP инкубировали при 50°C в течение 24 часов. Реакцию останавливали посредством фильтрации фермента концентратором Vivaspin 2 (10000 MWCO). Тагатозу определяли и измеряли количественно с использованием Agilent 1100 серии ВЭЖХ с рефрактометрическим детектором и Agilent Hi-Plex H-column. Подвижная фаза составила 5 мМ H2SO4, которую обрабатывали при 0,6 мл/мин. Получали выход тагатозы, составивший 9,2 г/л. Это соответствует 92% теоретического выхода вследствие пределов разложения мальтодекстрина без ферментов, таких как изоамилаза или 4-глюкантрансфераза. Для количественной оценки выхода авторов изобретения использовались стандарты различных концентраций тагатозы.
Пример 10
Реакционную смесь, содержащую 200 г/л мальтодекстрина, 10 мМ ацетатного буфера (pH 5,5), 5 мМ MgCl2 и 0,1 г/л изоамилазы инкубировали при 80°C в течение 24 часов. Ее использовали для создания другой реакционной смеси, содержащей 20 г/л мальтодекстрина, обработанного изоамилазой, 50 мМ забуференного фосфатом физиологического раствора с pH 7,2, 5 мМ MgCl2, 0,05 ед αGP, 0,05 ед PGM, 0,05 ед PGI, 0,05 ед F6PE и 0,05 ед T6PP и инкубировали при 50°C в течение 24 часов. Производство тагатозы измеряли количественно, как в Примере 9. Выход тагатозы повысился до 16 г/л с предварительной обработкой мальтодекстрина изоамилазой. Это составляет 80% от теоретического выхода.
Пример 11
Для дополнительного увеличения выхода тагатозы из мальтодекстрина, в реакционную смесь, описанную в примере 9, добавили 0,05 ед 4-глюканотрансферазы (4GT).
0,2 мл реакционной смеси, содержащей 20 г/л мальтодекстрина, обработанного изоамилазой (см. пример 9), 50 мМ забуференного фосфатом физиологического раствора с pH 7,2, 5 мМ MgCl2, 0,05 ед αGP, 0,05 ед PGM, 0,05 ед PGI, 0,05 ед F6PE, 0,05 ед T6PP и 0,05 ед 4GT инкубировали при 50°C в течение 24 часов. Производство тагатозы измеряли количественно, как в примере 9. Выход тагатозы повысился до 17,7 г/л с добавлением 4GT к мальтодекстрину, обработанному IA. Это составляет 88,5% теоретического выхода.
Пример 12
Для определения диапазона концентраций забуференного фосфатом физиологического раствора (PBS) 0,20 мл реакционной смеси, содержащей 50 г/л мальтодекстрина; 6,25 мМ, 12,5 мМ, 25 мМ, 37,5 мМ или 50 мМ забуференного фосфатом физиологического раствора с pH 7,2; 5 мМ MgCl2; 0,1 ед αGP; 0,1 ед PGM; 0,1 ед PGI; 0,1 ед F6PE и 0,1 ед T6PP инкубировали при 50°C в течение 6 часов. Короткая продолжительность гарантирует, что завершение не было достигнуто, и вследствие этого можно было бы четко увидеть различия в эффективности. Производство тагатозы измеряли количественно, как в примере 9. Для реакционных смесей, содержащих либо 6,25 мМ, 12,5 мМ, 25 мМ, 37,5 мМ, либо 50 мМ забуференного фосфатом физиологического раствора с pH 7,2, получили выход, составивший 4,5 г/л, 5,1 г/л, 5,6 г/л, 4,8 г/л или 4,9 г/л тагатозы, соответственно (Таблица 1). Эти результаты показывают, что концентрация 25 мМ PBS с pH 7,2 идеально подходит для этих конкретных условий реакции. Важно отметить, что использование даже 6,25 мМ PBS при рН 7,2 приводит к значительному обороту вследствие повторного использования фосфата. Это демонстрирует, что раскрытые способы повторного использования фосфата способны сохранять уровни фосфата низкими даже при промышленных уровнях объемной производительности (напр., 200-300 г/л мальтодекстрина).
Таблица 1
Пример 13
Для определения диапазона рН каскадной реакции 0,20 мл реакционной смеси, содержащей 50 г/л мальтодекстрина; 50 мМ забуференного фосфатом физиологического раствора с pH 6,0, 6,2, 6,4, 6,6, 6,8, 7,0, 7,2, или 7,3; 5 мМ MgCl2; 0,02 ед αGP; 0,02 ед PGM; 0,02 ед PGI; 0,02 ед F6PE и 0,02 ед T6PP инкубировали при 50°C в течение 16 часов. Единицы уменьшали, обеспечивая, чтобы завершение не было достигнуто, и вследствие этого можно было бы четко увидеть различия в эффективности. Производство тагатозы измеряли количественно, как в примере 8. Для реакционных смесей, содержащих 50 мМ забуференного фосфатом физиологического раствора с pH 6,0, 6,2, 6,4, 6,6, 6,8, 7,0, 7,2 или 7,3, получили выход, составивший 4,0 г/л, 4,1 г/л 4,2 г/л, 4,1 г/л, 4,4 г/л, 4,1 г/л, 3,8 г/л или 4,0 г/л тагатозы (Таблица 2). Эти результаты показывают, что pH 6,8 является идеальной для этих конкретных условий реакции, хотя система работает при широком диапазоне рН.
Таблица 2
Пример 14
Чтобы исследовать увеличение масштаба 20 мл реакционной смеси, содержащей 50 г/л мальтодекстрина, обработанного изоамилазой (см. Пример 9), 50 мМ забуференного фосфатом физиологического раствора pH 7,2, 5 мМ MgCl2, 10 ед αGP, 10 ед PGM, 10 ед PGI, 10 ед F6PE и 10 ед T6PP инкубировали при 50°C в течение 24 часов. Производство тагатозы измеряли количественно, как в примере 8. Выход тагатозы составил 37,6 г/л в масштабе 20 мл и 50 г/л мальтодекстрина. Это составляет 75% теоретического выхода. Эти результаты показывают, что увеличение масштаба до больших объемов реакции не приведет к значительным потерям выхода.
Пример 15
Для дополнительного увеличения выхода тагатозы из мальтодекстрина 0,05 ед мальтозофосфорилазы добавили к реакционной смеси, описанной в примере 9.
Пример 16
Для дополнительного увеличения выхода тагатозы из мальтодекстрина к реакционной смеси, описанной в примере 9, добавили 0,05 ед полифосфатглюкокиназы и 75 мМ полифосфата.
Пример 17
Для получения тагатозы из фруктозы реакционную смесь, содержащую 10 г/л фруктозы, 50 мМ Трис-буфера с pH 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 ед фруктозополифосфаткиназы, 0,05 ед F6PE и 0,05 ед T6PP инкубировали при 50°C в течение 24 часов. Получение тагатозы количественно измеряли, как в примере 9.
Пример 18
Для получения тагатозы из глюкозы реакционную смесь, содержащую 10 г/л глюкозы, 50 мМ Трис-буфера с pH 7,0, 75 мМ полифосфата, 5 мМ MgCl2, 0,05 ед глюкозы полифосфаткиназа, 0,05 ед PGI, 0,05 ед F6PE и 0,05 ед T6PP инкубировали при 50°C в течение 24 часов. Получение тагатозы количественно измеряли, как в примере 9.
Пример 19
Для получения тагатозы из сахарозы реакционную смесь, содержащую 10 г/л сахарозы, 50 мМ забуференного фосфатом физиологического раствора с pH 7,0, 5 мМ MgCl2, 0,05 ед сахарозофосфорилаза, 0,05 PGM, 0,05 ед PGI, 0,05 ед F6PE и 0,05 ед T6PP инкубировали при 50°C в течение 24 часов. Получение тагатозы измеряли количественно, как в примере 9.
Пример 20
Для дополнительного увеличения выхода тагатозы из сахарозы к реакционной смеси в примере 15 добавили 75 мМ полифосфата и 0,05 полифосфатфруктокиназы. Получение тагатозы измеряли количественно, как в примере 9.
Изобретение относится к ферментативному способу получения тагатозы. Способ включает стадии (i) преобразование фруктозо-6-фосфата (F6P) в тагатозо-6-фосфат (T6P), катализируемое фруктозо-6-фосфатэпимеразой (F6PE); и (ii) преобразование полученного T6P в тагатозу, катализируемое тагатозо-6-фосфатфосфатазой (T6PP). При этом стадии (i) и (ii) способа проводят в одном реакционном сосуде. Изобретение обеспечивает синтез тагатозы с высоким выходом продукта. 19 з.п. ф-лы, 7 ил., 2 табл., 20 пр.
1. Ферментативный способ получения тагатозы, включающий стадии:
(i) преобразование фруктозо-6-фосфата (F6P) в тагатозо-6-фосфат (T6P), катализируемое фруктозо-6-фосфатэпимеразой (F6PE); и
(ii) преобразование полученного T6P в тагатозу, катализируемое тагатозо-6-фосфатфосфатазой (T6PP),
где стадии (i) и (ii) способа проводят в одном реакционном сосуде.
2. Способ по п.1, дополнительно включающий стадию преобразования глюкозо-6-фосфата (G6P) в F6P, при этом стадия катализируется фосфоглюкозоизомеразой (PGI).
3. Способ по п.2, дополнительно включающий стадию преобразования глюкозо-1-фосфата (G1P) в G6P, при этом стадия катализируется глюкозофосфомутазой (PGM).
4. Способ по п.3, дополнительно включающий стадию преобразования сахарида в G1P, при этом стадия катализируется по меньшей мере одним ферментом, при этом сахарид выбирают из группы, состоящей из крахмала или его производного, целлюлозы или ее производного и сахарозы.
5. Способ по п.4, в котором по меньшей мере один фермент выбирают из группы, состоящей из альфа-глюканофосфорилазы (αGP), мальтозофосфорилазы, сахарозофосфорилазы, целлодекстринфосфорилазы, целлобиозофосфорилазы и целлюлозофосфорилазы.
6. Способ по п.4, в котором сахарид представляет собой крахмал или его производное, выбранное из группы, состоящей из амилозы, амилопектина, растворимого крахмала, амилодекстрина, мальтодекстрина, мальтозы и глюкозы.
7. Способ по п.4 или 6, дополнительно включающий стадию преобразования крахмала в производное крахмала, при этом производное крахмала получают посредством ферментного гидролиза крахмала или посредством кислотного гидролиза крахмала.
8. Способ по п.6, в котором в способ включили 4-глюканотрансферазу (4GT).
9. Способ по п.7, в котором производное крахмала получают посредством ферментного гидролиза крахмала, катализируемого изоамилазой, пуллуланазой, альфа-амилазой или их комбинацией.
10. Способ по п.1, дополнительно включающий:
стадию преобразования фруктозы в F6P, при этом стадия катализируется по меньшей мере одним ферментом.
11. Способ по п.2, дополнительно включающий:
стадию преобразования глюкозы в G6P, при этом стадия катализируется по меньшей мере одним ферментом.
12. Способ по п.1, в котором фруктозо-6-фосфатэпимераза кодируется полинуклеотидом, содержащим нуклеотидную последовательность, имеющую по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% идентичность последовательности с SEQ ID NOS.: 1, 3, 5, 7, 9 или 10.
13. Способ по п.1, в котором фруктозо-6-фосфатэпимераза содержит аминокислотную последовательность, имеющую по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% идентичность последовательности с SEQ ID NOS.: 2, 4, 6, 8 или 11.
14. Способ по п.1, в котором тагатозо-6-фосфатфосфатаза кодируется полинуклеотидом, содержащим нуклеотидную последовательность, имеющую по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% идентичность последовательности с SEQ ID NOS.: 12, 14 или 16.
15. Способ по п.1, в котором тагатозо-6-фосфатфосфатаза содержит аминокислотную последовательность, имеющую по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 99% или 100% идентичность последовательности с SEQ ID NOS.: 13, 15 или 17.
16. Способ по любому из пп. 1-15, при этом стадии способа проводят при температуре, варьирующей от приблизительно 40°C до приблизительно 70°C, при рН, варьирующем от приблизительно от приблизительно 5,0 до приблизительно 8,0, и/или в течение от приблизительно 8 часов до приблизительно 48 часов.
17. Способ по любому одному из пп. 1-16, в котором стадии способа проводят в одном биореакторе или во множестве биореакторов, расположенных последовательно.
18. Способ по любому из пп. 1-17, в котором стадии способа проводят без содержания ATP, без содержания NAD(H), при концентрации фосфата от приблизительно 0 мМ до приблизительно 150 мМ, при этом фосфат используется повторно, и/или по меньшей мере одна стадия способа включает энергетически выгодную химическую реакцию.
19. Способ по п. 10, дополнительно включающий стадию превращения сахарозы во фруктозу, где стадия катализируется по меньшей мере одним ферментом.
20. Способ по п.11, дополнительно включающий стадию превращения сахарозы в глюкозу, где стадия катализируется по меньшей мере одним ферментом.
| KR 101480422 B1, 13.01.2015 | |||
| KR 1020060059622 А, 02.06.2006 | |||
| VAN DER MAAREL M.J.E.C | |||
| et al | |||
| Starch modification with microbial alpha-glucanotransferase enzymes | |||
| Carbohydr Polym | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| база | |||

