Область техники, к которой относится изобретение
Настоящее изобретение относится к способу получения лилий, содержащих синий пигмент (делфинидин) в лепестках, путем введения в лилии инородного гена. Конкретнее, настоящее изобретение относится к способу получения лилий, содержащих синий пигмент в лепестках, путем введения гена флавоноид 3',5'-гидроксилазы (F3'5'H), полученного из колокольчика, который необходим для синтеза делфинидина, в лилии, и введения фрагмента гена флавоноид 3'-гидроксилазы (F3'H) лилии в целях подавления экспрессии эндогенного F3'H, который действует на синтез красного пигмента (цианидина), и индукции синтеза делфинидина в лепестках лилии; и к лилии, полученной данным способом.
Предшествующий уровень техники
Ранее сообщалось о способах индукции выработки делфинидина в растительных клетках путем введения инородного гена в гвоздики (см. Патентный Документ 1), хризантемы (см. Патентный Документ 1), розы (см. Патентный Документ 1 и Патентный Документ 2), фаленопсисы (см. Патентный Документ 3) и цимбидиумы (см. Патентный Документ 3).
Однако еще не сообщалось о случаях успешно полученных лилий с синими лепестками путем синтеза делфинидина в лепестках лилий под действием введенного инородного гена.
Примеры способов, которые используют для улучшения сортов растений, включают (1) перекрестное скрещивание, путем скрещивания пестика и тычинки, (2) спонтанную искусственную мутацию, и (3) генетическую рекомбинацию. Применение технологии генетической рекомбинации позволяет экспрессировать полезный ген в растении без стеснения генетическими ограничениями, имеющимися у видов-мишеней, что позволяет придать новый признак растению, которым растение исходно не обладало. Генетически модифицированные растения, полученные технологией генетической рекомбинации, в настоящее время широко культивируются по всему миру.
Примеры компонентов растительных пигментов включают антоцианин, каротеноид и беталаин.
Антоцианины являются представителями группы вторичных метаболитов, которые называются флавоноидами, и, как показано на фиг.1, синтез которых проходит начиная с фенилаланина до кумароил-КоА под действием множества ферментов. Цвет антоцианинов зависит от их структуры. То есть, при увеличении количества гидроксильных групп В кольца антоцианидинов, которое является хромофором антоцианинов, синий цвет становиться более интенсивным, и основные антоцианидины в форме делфинидинов, цианидинов, и пералгонидинов имеют увеличенное количество гидроксильных групп, в указанном порядке.
Антоцианины, полученные из делфинидина, накапливаются во многих цветах голубого цвета. Кроме того, известно, что при увеличении количества ароматических ацильных групп (таких как кумароильные группы или кафеольные группы), которые модифицируют антоцианины, цвет антоцианинов становится все более и более синим (то есть, максимальное поглощение сдвигается в более длинную длину волны), а стабильность антоцианов растет. В частности, антоцианины, в которых связаны множественные ароматические ацильные группы, называемые ацилированными антоцианинами, содержатся в синих лепестках горечавки и мотылькового горошка (см. непатентный документ 1).
Ферменты, вовлеченные в биосинтез антоцианина и гены, которые кодируют эти ферменты, ранее были выделены из множества растений (см. непатентный документ 1). Цвет антоцианинов зависит от таких факторов, как рН вакуолей, в которых они локализованы, наличия других флавоноидов и ионов металлов. То есть, антоцианины выглядят красными, если рН вакуолей низкий, и становятся синими, если рН вакуолей становится нейтральным. Кроме того, флавоны и флавонолы, известные как ко-пигменты для антоцианинов, оказывают воздействие, которое делает антоцианины синими. Также, ионы железа и алюминия, как известно, являются причиной того, что антоцианины становятся синими, так как координируют гидроксильные группы В-кольца антоцианинов.
Хотя растения образуют цветы различных расцветок, только несколько видов способны давать цветы всех расцветок. Это происходит потому, что пигменты, которые могут быть синтезированы видами, задаются генетически. Например, нет синих или фиолетовых сортов роз, гвоздик, хризантем, лилий и гербер. Основная причина этого заключается в том, что эти растения не имеют гена флавоноид 3',5'-гидроксилазы (F3'5'H) необходимого для синтеза делфинидина. По этой причине уже было несколько попыток получения голубых цветов путем индукции выработки делфинидина вследствие экспрессии гена F3'5'H.
При экспрессии гена F3'5'H в петуниях, утративших F3'5'H, выросло количество антоцианинов, полученных из делфинидина (см. непатентный документ 2 и непатентный документ 3). Кроме того, при экспрессии гена F3'5'H в табаке (Nicotiana tabacum), который накапливает цианидин, синтезируется делфинидин и цветы принимают голубоватый оттенок (см. непатентный документ 3). В случае экспрессии в табаке гена F3'5'H, выделенного из колокольчика, эустомы или петунии, ген F3'5'H, полученный из колокольчика, вырабатывает делфинидин с наилучшей эффективностью (см. непатентный документ 4). Кроме того, в случае экспрессии гена в F3'5'H мотылькового горошка или вербены в вербене, в случае экспрессии гена, выделенного из мотылькового горошка, получается больший объем выработки, и цвет цветка подвергается четко выраженному изменению (см. непатентный документ 5). При экспрессии в розах гена, выделенного из петунии, горечавки, мотылькового горошка, цинерарии, эустомы, анютиных глазок и лаванды, только в случае экспрессии гена F3'5'H, выделенного из анютиных глазок делфинидин вырабатывался на уровне, равном 10% общего количества антоцианидинов, что говорит о том, что выделенный из анютиных глазок ген F3'5'H подходящим образом функционирует в розах (см. Патентный Документ 4).
Генетически модифицированная хризантема, которая накапливает 50% и большее количество делфинидина, и в которой цвет цветка изменился на синий путем транскрипции выделенного из анютиных глазок гена F3'5'H с промотора хальконсинтазы розы, была получена для сортов хризантем «Improved Reagan» и «Dark Splendid Reagan» (см. Патентный Документ 5). При экспрессии гена F3'5'H, выделенного из таких растений как колокольчик, вербена, цинерария, анютины глазки, петуния или лобелия, с промотора, выделенного из гена флавонон 3-гидроксилазы хризантемы и 5'-нетранслируемой области гена алкогольдегидрогеназы табака, гены F3'5'H из колокольчика, вербены, цинерарии и анютиных глазок функционировали сравнительно хорошо, и делфинидин вырабатывался на уровне, равном 25% от общего количества антоцианидинов. В хризантемах выделенный из колокольчика ген F3'5'H функционирует наилучшим образом, и делфинидин вырабатывался на уровне 75% от общего количества антоцианидинов (см. непатентный документ 16).
Из этих описанных в литературе примеров очевидно, что несмотря на то, что уже были описаны примеры выработки делфинидина путем экспрессии гетерологичного (инородного) гена в растениях, для выработки делфинидина в лепестках потребуется изрядное количество экспериментов методом проб и ошибок и повторных экспериментов, поскольку трудно спрогнозировать какой из генов F3'5'H в действительности будет экспрессироваться в лепестках представляющего интерес растения, при введении гена F3'5'H, выделенного из определенного растения, в представляющее интерес растение под контролем определенного промотора.
Кроме того, результаты всех исследований, представленные в вышеупомянутых примерах, были получены для двудольных растений. Существуют немалые различия между двудольными и однодольными растений даже среди высших растений.
Во-первых, двудольные и однодольные растения имеют различные морфологические различия. Это говорит о том, что в ходе своего происхождения однодольные растения подвергались значительной морфологической эволюции. Так, однодольные растения характеризуются: наличием одной семядоли, при этом основа семядоли образует оболочку, которая покрывает другие части проростка, сосудисто-волокнистыми тяжами случайно присутствующими в поперечном разрезе стебля, после того как стебель завершает свой рост он не становится толще (камбий не развивается), сосуды имеют форму параллельных линий (параллельные сосуды), стержневой корень не развивается до больших размеров, но скорее имеет форму мочковатых корней, количество листочков околоцветника, тычинок и плодолистиков кратно 3, нет различия между лепестками и чашелистиками (см. http://www.fukuoka-edu.ac.jp/~fukuhara /keitai/9-1.html).
Более того, поскольку однодольные и двудольные растения значимо отличаются по контролю генной экспрессии (см. непатентный документ 6 и непатентный документ 7), возникают трудности при культивировании генетически модифицированных однодольных растений. Однако на однодольных растениях кукурузе и рисе было проведено изрядное количество научных исследований и разработок, а генетически модифицированная кукуруза и уже была коммерциализирована во многих странах.
Таким образом, хотя существует ряд сообщений, описывающих временную экспрессию гетерологичного гена (чужеродного) F3'5'H в лепестках однодольных цветущих растений (см. Патентный Документ 3), и существует несколько сообщений, описывающих разработку генетически модифицированных однодольных цветущих растений, но все еще отсутствуют сообщения, описывающие создание однодольных цветущих растений, в которые был бы введен ген F3'5'H, а именно создание однодольных цветущих растений, в которых цвет цветка был изменен посредством аккумуляции делфинидина в его лепестках в результате введения гетерологичного гена F3'5'H.
Помимо этого в непатентном документе 8 описано, что при выведении синих лилий путем введения гена, относящегося к синтезу синего пигмента, в красные лилии, были получены лилии, в которых содержание синего пигмента составляло половину или больше от всего пигмента. Однако, тканью или частью растения лилии, в которой в непатентном документе 8 был проанализирован синий пигмент, был лишь черешок. Существует множество растений, в которых антоцианины не синтезируются в цветах даже в том случае, когда они синтезируются в листьях (например, в арабидопсисе) и существуют известные случаи, при которых молекулярные виды антоцианинов в листьях и антоцианины в цветках отличаются. Например, хотя антоцианины, которые содержатся только в листьях Sarrecenia purpurea и Sarracenia psitticina, происходят из цианидина, антоцианины содержащиеся в их цветках, происходят из цианидина и делфинидина (см. непатентный документ 17). Кроме того, контроль синтеза антоцианина весьма вероятно отличается в зависимости от ткани. Например, несмотря на то, что ген с, придает белый цвет вьюнкам пурпурным, их стебли окрашиваются антоцианинами, а семена являются черными. С другой стороны, ген c, придает белый цвет цветам, хотя стебли не окрашиваются и семена являются белыми.
Таким образом, сообщение, в котором описывается, что содержание делфинидина составляет половину всех пигментов в черешке лилий в результате введения гетерологичного гена, не означает, что был разработан способ культивирования синих лилий, который позволяет выращивать лилии, в которых накопление делфинидина в лепестках является результатом введения гетерологичного гена.
Кроме того, в целях изменения цвета цветов на синий, требуется содержание делфинидина 50% или больше, предпочтительно 69% или больше, более предпочтительно 70% или больше, еще более предпочтительно 80% или больше, еще более предпочтительно 90% или больше, еще более предпочтительно до 95% или больше, еще более предпочтительно до 99% или больше, и наиболее предпочтительно 100% от общего количества антоцианидинов. Поскольку для достижения этого зачастую недостаточно просто сверхэкспрессировать ген F3'5'H, необходимо дополнительное изобретательство, в том числе дополнительные манипуляции с генами.
Например, в случае гвоздик, при экспрессии как гена F3'5'H, так и гена петунии дигидрофлованол 4-редуктазы (DFR) в белых гвоздиках, дефектных по гену DFR, содержание делфинидина становится почти 100% и получаются генетически модифицированные гвоздики, в которых окраска цветов изменена на синий (см. патентный документ 6 и фиг.1 настоящей заявки). Также при экспрессии в розах выделенного из анютиных глазок гена F3'5'H и выделенного из ириса гена DFR в дополнение к подавлению экспрессии гена DFR розы, содержание делфинидина становится почти 100% и получаются генетически модифицированные розы, в которых окраска цветов изменена на синий (см. Патентный Документ 2). Кроме того, в хризантемах содержание делфинидина повышается сверхэкспрессией выделенного из анютиных глазок гена F3'5'H и подавлением экспрессии эндогенного гена F3'5'H хризантемы (см. Патентный документ 5).
Документы предшествующего уровня техники
Патентные документы
Патентный Документ 1: Международная публикация No. WO 94/28140 (PCT/AU94/00265, Нерассмотренная патентная публикация Японии (Перевод РСТ Заявки) No. H8-511683)
Патентный Документ 2: Международная публикация No. WO 2005/017147 (PCT/JP2004/001958)
Патентный документ 3: Нерассмотренная патентная публикация Японии No. 2008-252250
Патентный Документ 4: Международная публикация No. WO 2004/020637 (PCT/AU03/01111)
Патентный Документ 5: Международная публикация No. WO 2009/062253 (PCT/AU2008/001694)
Патентный Документ 6: Международная публикация No. WO 96/36716 (PCT/AU96/00296)
Патентный Документ 7: Международная публикация No. WO 2007/094521 (PCT/JP2007/053342)
Патентный Документ 8: Международная публикация No. WO 2008/156214 (PCT/JP2008/061603)
Патентный документ 9: Патентная Заявка Японии No. 2008-276029, которая будет опубликована в ближайшее время.
Непатентные документы
Непатентный документ 1: Plant J. (2008) 54, 737-749
Непатентный документ 2: Nature (1993) 366, 276-279
Непатентный документ 3: FEBS Lett. (1999) 461, 241-245
Непатентный документ 4: Biosci. Biotechnol. Biochem. (2003) 67(1), 161-165
Непатентный документ 5: Plant Biotechnology (2006) 23, 5-11
Непатентный документ 6: Protein, Nucleic Acid and Enzyme (1989) 34, 1873-1878
Непатентный документ 7: Trends in Genetics (1988) 4, 13-18
Непатентный документ 8: 2006 Annual Report No. 1-26 of the Niigata Agricultural Research Institute, (32) Project for the Development of Top Brand Agricultural Products, Cultivation of Blue Lilies
Непатентный документ 9: Plant Cell Reports (2004) 22, 415-421
Непатентный документ 10: Plant Cell Reports (2004) 22, 359-364
Непатентный документ 11: Theor. App. Genet. (1999) 99, 383-390
Непатентный документ 12: Plant Cell Physiol. (1996) 37, 49-59
Непатентный документ 13: Clontech, Molecular Breeding (2003) 11, 287-293
Непатентный документ 14: J. Japan Soc. Hort. Sci. (2008) 77, 94-102
Непатентный документ 15: Plant J. (2008) 54, 949-962
Непатентный документ 16: Abstracts of the 2010 Annual Meeting of the Japanese Society of Plant Physiologists, Poster No. P1C012(600),
http://www.jspp.org/13member/2010abstract/pdf/07.pdf
Непатентный документ 17: Hort. Science (2001) 36, 384.
Сущность изобретения
Проблема, решаемая изобретением
Почти во всех растениях в образования синего цвета участвует делфинидин, пигмент на основе антоцианина. Хотя F3'5'H является существенным для синтеза делфинидина, этот фермент отсутствует в лилиях. Следовательно, синих лилий не существует. Для культивирования синих лилий путем введения гена, требуется получение в лилиях высокой экспрессии F3'5'H. С другой стороны, содержание делфинидина не увеличивается в лилиях, синтезирующих цианидин, до тех пор, пока не подавляется синтез цианидина.
Целью настоящего изобретения является предложение способа получения лилий, которые синтезируют в клетках лепестков синий пигмент делфинидин с более высоким относительным содержанием, путем обеспечения высокого уровня экспрессирующего F3'5'H, выделенного из определенных видов, вместе с подавлением эндогенного цианидина лилий.
Способы решения проблем
В результате проведения интенсивных исследований для решения вышеупомянутых задач, авторы настоящего изобретения обнаружили, что при введении выделенного из колокольчика гена F3'5'H в сорт восточной лилии «Acapulco», которая имеет розовый цвет и в которой цианидин отвечает почти за весь присутствующий пигмент на основе антоциана, и при подавлении синтеза цианидина с помощью RNAi путем введения фрагмента гена F3'H лилии, 80% из всего пигмента на основе антоциана в клетках лепестков становится делфинидином, что приводит, таким образом, к осуществлению настоящего изобретения.
Подробнее настоящее изобретение представлено ниже.
[1] Способ получения лилий, содержащих в лепестках делфинидин, который включает следующие стадии:
введение в лилии выделенного из колокольчиков гена флавоноид-3',5'-гидроксилазы (F3'5'H), включающего нуклеотидную последовательность, представленную в SEQ ID NO:1 или SEQ ID NO:11, или нуклеотидную последовательность, идентичную, по меньшей мере, на 90% последовательности с нуклеотидной последовательностью, представленной в SEQ ID NO:1 или SEQ ID NO:11, которая также кодирует полипептид, обладающий активностью F3'5'H,
введение фрагмента гена флавоноид-3'-гидроксилазы (F3'H), выделенного из лилии, включающего нуклеотидную последовательность, представленную в SEQ ID NO:3 или SEQ ID NO:16, или нуклеотидную последовательность, идентичную, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:3 или SEQ ID NO:16, которая также кодирует полипептид, обладающий активностью F3'H, и
синтез делфинидина в результате деятельности введенного гена F3'5'H, с одновременным подавлением экспрессии эндогенного гена F3'H, который участвует в синтезе цианидина в лепестках лилий.
[2] Способ, описанный в [1], в котором подавление экспрессии эндогенного F3'H проводится способом RNAL
[3] Способ, описанный в [1] или [2] выше, в котором выделенный из колокольчика ген F3'5'H состоит из нуклеотидной последовательности, идентичной, по меньшей мере, на 95% нуклеотидной последовательности, представленной в SEQ ID NO:1 или SEQ ID NO:11, которая также кодирует полипептид, имеющий активность F3'5'H.
[4] Способ, описанный в [1] или [2], в котором выделенный из колокольчика ген F3'5'H состоит из нуклеотидной последовательности, представленной в SEQ ID NO:1 или SEQ ID NO:11.
[5] Способ, описанный в любом из [1] - [4] выше, в котором фрагмент выделенного из лилии гена F3'H состоит из нуклеотидной последовательности, идентичной, по меньшей мере, на 95% нуклеотидной последовательности, представленной в SEQ ID NO:3 или SEQ ID NO:16, которая также кодирует полипептид, имеющий активность F3'H.
[6] Способ, описанный в любом из [1] - [4], в котором фрагмент выделенного из лилии гена F3'H состоит из нуклеотидной последовательности, представленной в SEQ ID NO:3 или SEQ ID NO:16.
[7] Способ, описанный в любом из [1] - [6] выше, дополнительно включающий: введение фрагмента выделенного из лилии гена дигидрофлавонол 4-редуктазы (DFR), содержащего нуклеотидную последовательность, представленную в SEQ ID NO:13 или нуклеотидную последовательность, идентичную, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:13, которая также кодирует полипептид, имеющий активность DFR, и подавление экспрессии эндогенного DFR, которая оказывает влияние на синтез цианидина в лепестках лилий, и одновременное введение выделенного из ириса инородного гена DFR.
[8] Способ, описанный в [7] выше, в котором подавление экспрессии эндогенного DFR проводится способом RNAi.
[9] Способ, описанный в любом из [1] - [8], дополнительно включающий: применение последовательности промотора DFR лилий, представленного в SEQ ID NO:24.
[10] Лилия, ее ткань, ее часть, полученное из нее вегетативным размножением растение или ее потомок, которые содержат делфинидин в лепестках, полученные согласно способу, описанному в любом из [1] или [9] выше.
[11] Лилия, ее ткань, ее часть, полученное из нее вегетативным размножением растение, или ее потомок, описанные в [10], где ткань или часть является срезанным цветком.
Осуществления изобретения
Согласно предложенному в настоящем изобретении способу получения лилий, содержащих в лепестках делфинидин, синий пигмент, присутствующий в клетках лепестков лилий в форме делфинидина, может быть аккумулирован с более высоким относительным содержанием, что, таким образом, позволяет получить ранее не существовавшие лилии, которые имеют синие цветы.
Краткое описание чертежей
На фиг.1 изображен путь биосинтеза антоцианинов.
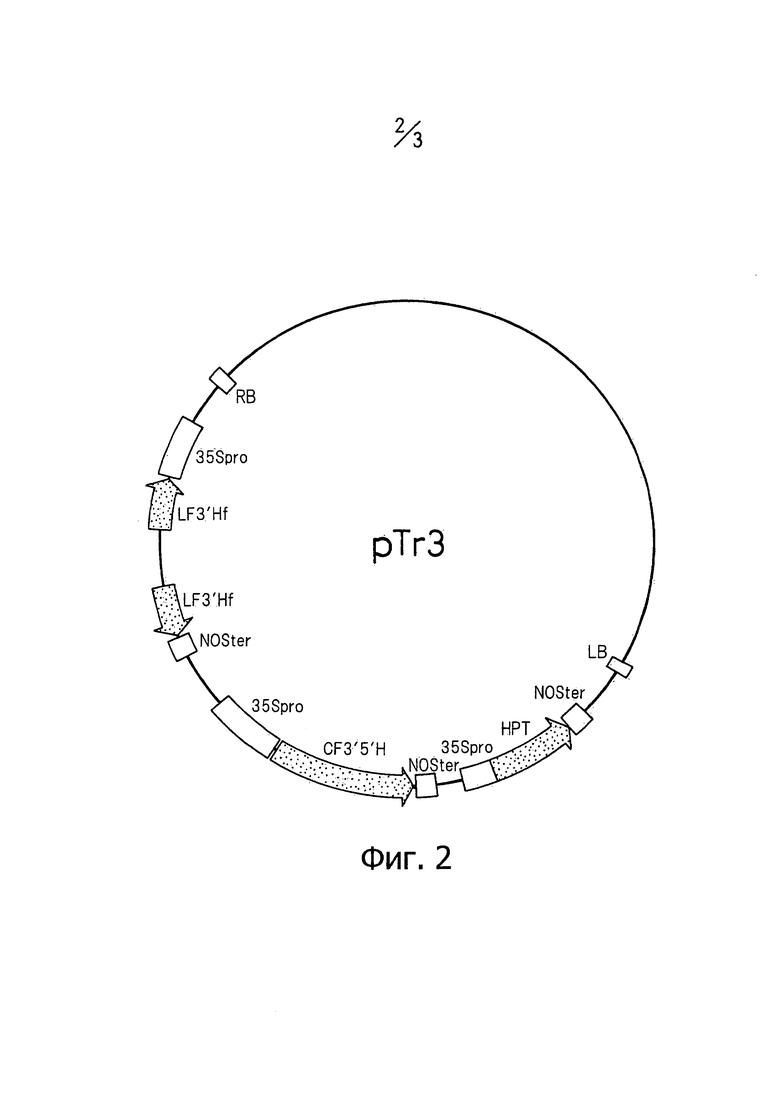
На фиг.2 изображена карта бинарного вектора Tr3.
На фиг.3 изображены карты бинарных векторов Tr7 - Tr11.
Воплощения изобретения
Ниже представлено подробное описание настоящего изобретения.
Настоящее изобретение относится к способу получения лилий, содержащих в лепестках делфинидин, включающему следующие стадии:
введение в лилии выделенного из колокольчиков гена флавоноид-3',5'-гидроксилазы (F3'5'H), включающего нуклеотидную последовательность, представленную в SEQ ID NO:1 или SEQ ID NO:11, или нуклеотидную последовательность, идентичную, по меньшей мере, на 90% последовательности с нуклеотидной последовательностью, представленной в SEQ ID NO:1 или SEQ ID NO:11, которая также кодирует полипептид, обладающий активностью F3'5'H,
введение фрагмента выделенного из лилии гена флавоноид-3'-гидроксилазы (F3'H), включающего нуклеотидную последовательность, представленную в SEQ ID NO:3 или SEQ ID NO:16, или нуклеотидную последовательность, идентичную, по меньшей мере, на 90% последовательности с нуклеотидной последовательностью, представленной в SEQ ID NO:3 или SEQ ID NO:16, которая также кодирует полипептид, обладающий активностью F3'H, и
синтез делфинидина в результате действия введенного гена F3'5'H, с одновременным подавлением экспрессии эндогенного гена F3'H, который участвует в синтезе цианидина в лепестках лилий.
Примеры вышеупомянутой нуклеотидной последовательности, идентичной, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:1 или SEQ ID NO:11, которая также кодирует полипептид, обладающий активностью флавоноид-3'5'-гидроксилазы (F3'5'H), включают ДНК, идентичную, по меньшей мере, на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% и 99% или более ДНК, состоящей из нуклеотидной последовательности, представленной в SEQ ID NO:1, при расчете с помощью пакета аналитических программ, таких как «BLAST» или «FASTA».
Аналогично, примеры вышеупомянутой нуклеотидной последовательности, идентичной, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:3 или SEQ ID NO:16, которая также кодирует полипептид, обладающий активностью флавоноид-3'-гидроксилазы (F3'H), включают ДНК, идентичную, по меньшей мере, на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% и 99% или более ДНК, состоящей из нуклеотидной последовательности, представленной в SEQ ID NO:3, при расчете с помощью пакета аналитических программ, таких как «BLAST» или «FASTA».
Нуклеотидная последовательность, идентичная, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:1 или SEQ ID NO:11, которая обладает активностью флавоноид-3',5'-гидроксилазы (F3'5'H), также может представлять собой нуклеотидную последовательность, в которой один или несколько нуклеотидов удалены, заменены или вставлены в нуклеотидную последовательность, представленную в SEQ ID NO:1 или SEQ ID NO:11, которая также кодирует полипептид, обладающий активностью флавоноид-3',5'-гидроксилазы (F3'5'H). В данном документе, «один или несколько» в ДНК или в нуклеотидной последовательности означает, что 1-20, предпочтительно, 1-10, более предпочтительно, 1-5, еще более предпочтительно, 1-4, и еще более предпочтительно, 1-3 оснований в нуклеотидной последовательности, представленной в SEQ ID NO:1, удалены, заменены или вставлены, при условии наличия вышеупомянутой ферментной активности. Кроме того, термины «делеция», «замена» и «вставка» обозначают образование нуклеотидной последовательности, которая кодирует белок, обладающий свойствами, аналогичными свойствам белка (или полипептида) (SEQ ID NO:2), кодируемого нуклеотидной последовательностью, представленной в SEQ ID NO:1.
Аналогично, нуклеотидная последовательность, идентичная, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:3 или SEQ ID NO:16, которая кодирует пептид обладающий активностью флавоноид-3'-гидроксилазы (F3'H), также может представлять собой нуклеотидную последовательность, в которой один или несколько нуклеотидов удалены, заменены или вставлены в нуклеотидную последовательность, представленную в SEQ ID NO:3 или SEQ ID NO:16, которая также кодирует полипептид, обладающий активностью флавоноид-3'-гидроксилазы (F3'H). В данном документе, «один или несколько» в ДНК или в нуклеотидной последовательности означает, что 1-20, предпочтительно, 1-10, более предпочтительно, 1-5, еще более предпочтительно, 1-4, и еще более предпочтительно, 1-3 нуклеотидов в нуклеотидной последовательности, представленной в SEQ ID NO:3 или в SEQ ID NO:16, удалены, заменены или вставлены, при условии сохранения в результате вышеупомянутой ферментной активности. Кроме того, термины «делеция», «замена» и «вставка» обозначают образование нуклеотидной последовательности, которая кодирует белок, обладающий свойствами, аналогичными свойствам белка (или полипептида) (SEQ ID NO:4 или SEQ ID NO:17), кодируемого нуклеотидной последовательностью, представленной в SEQ ID NO:1 или в SEQ ID NO:16.
Нуклеотидная последовательность, идентичная, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:1 или SEQ ID NO:11, которая также кодирует полипептид, обладающий активностью флавоноид-3',5'-гидроксилазы (F3'5'H), может представлять собой нуклеотидную последовательность, способную гибридизоваться в жестких условиях с комплементарной цепью нуклеотидной последовательности, представленной в SEQ ID NO:1 или SEQ ID NO:11, которая также кодирует полипептид, обладающий активностью флавоноид-3'5'-гидроксилазы (F3'5'H).
Аналогично, нуклеотидная последовательность, идентичная, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:3 или SEQ ID NO:16, которая также кодирует полипептид, обладающий активность флавоноид-3'-гидроксилазы (F3'H), может представлять собой нуклеотидную последовательность, способную гибридизоваться в жестких условиях с комплементарной цепью нуклеотидной последовательности, представленной в SEQ ID NO:3, которая также кодирует полипептид, обладающий активностью флавоноид-3'-гидроксилазы (F3'H).
В данном документе «жесткие условия» по отношению к нуклеотидной последовательности или ДИК (последовательности) обозначают условия, описанные, например, в «Molecular Cloning: A Laboratory Manual (2nd Edition)» edited by Sambrook et al. Cold Spring Harbor Laboratory Press), хотя температура и концентрация солей во время гибридизации, а также, предпочтительно, во время промывки могут соответствующим образом регулироваться в зависимости от ДНК, которая гибридизуется с комплементарной цепью ДНК, состоящей из SEQ ID NO:1 или 11 или SEQ ID NO:3 или 16. Более конкретно, эти условия относится к стадии, состоящей из (i) инкубации в течение ночи при 42°C с зондом в растворе, содержащем 6xSSC (состав 1xSSC: 0,15 М NaCl, 0,015 М цитрата натрия, рН 7), 0,5% SDS, 5х раствор Денхардта, 100 µг/мл денатурированной фрагментированной ДНК спермы лосося и 50% формальдегид, и (ii) удаления неспецифично загибридизовавшегося зонда путем отмывки, где с целью дальнейшего повышения точности проводят отмывку в условиях более низкой ионной силы, например, в 2xSSC и более жестко в 0,1xSSC, и/или отмывку проводят при высокой температуре, например, при 40°С ниже, чем значение Тпл используемой нуклеиновой кислоты, более жестко при температуре на 30°С ниже, еще более жестко при температуре на 25°С ниже, и еще более жестко при температуре на 10°С ниже. Более специфично, хотя и возможны вариации в зависимости от значения Тпл используемой нуклеиновой кислоты, отмывку проводят при 25°С или выше, более жестко при 37°С или выше, еще более жестко при 42°С или выше, еще более жестко при 50°С или выше, и еще более предпочтительно, при 60°С или выше. Например, Тпл определяется из формулы Тпл=81,5+16,6(log[Na+])+0,41(%G+C)-(600/N) (где N обозначает длину олигонуклеотида, и %G+C обозначает содержания остатков гуанина и цитозина в олигонуклеотиде). Условия гибридизации могут быть установлены, например, согласно вышеупомянутой публикации. Кроме того, процедура, приведенная в настоящем описании, может быть подходящим образом проведена с учетом сведений из вышеупомянутой публикации.
Подавление экспрессии эндогенной флавоноид-3'-гидролазы (F3'H), участвующей в синтезе цианидина в лепестках лилий, путем введения фрагмента, выделенного из лилий гена F3'H, состоящего из последовательности, представленной в SEQ ID NO:3 или SEQ ID NO:16, или последовательности, идентичной, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:3 или SEQ ID NO:16, которая также кодирует полипептид, обладающий активностью F3'H, может проводиться супрессией антисмысловой последовательностью, супрессией с использованием смысловой последовательности (ко-супрессии) или RNAi и так далее, причем способ RNAi является предпочтительным.
Настоящее изобретение также относится к вышеупомянутому способу, дополнительно включающему введение фрагмента выделенного из лилии гена дигидрофлавонол-4-редуктазы (DFR), содержащего нуклеотидную последовательность, представленную в SEQ ID NO:13 или нуклеотидную последовательность, идентичную, по меньшей мере, на 90% нуклеотидной последовательности, представленной в SEQ ID NO:13, которая также кодирует полипептид, имеющий активность DFR, подавление экспрессии эндогенного DFR, который участвует в синтезе цианидина в лепестках лилий, и одновременное введение выделенного из ириса чужеродного гена DFR. Подавление экспрессии эндогенного DFR предпочтительно проводят способом RNAi.
Настоящее изобретение также относится к вышеупомянутому способу, который дополнительно включает стадию применения промотора DFR из лилий, представленного в последовательности SEQ ID NO:24.
Кроме того, определения делеции, замены или вставки одного или нескольких нуклеотидов в нуклеотидной последовательности, идентичности последовательности и гибридизации, приведенные относительно SEQ ID NO:1, 11, 3 и 16, аналогично применяются к SEQ ID NO:13 и 24, и помимо нуклеотидных последовательностей, представленных в SEQ ID NO:13 и 24, например, нуклеотидная последовательность, идентичная, по меньшей мере, на 90%, 95% или 99%, также входит в объем настоящего изобретения.
Лилия, содержащая в лепестках делфинидин, полученная согласно способу получения лилий, содержащих в лепестках делфинидин по настоящему изобретению, ее ткань, ее часть, вегетативно полученное из нее растение или ее потомство, также входят в объем настоящего изобретения. Предпочтительно, если ее ткань или часть являются срезанным цветком.
Примеры
Хотя нижеследующее представляет более подробное пояснение настоящего изобретения с помощью примеров, подразумевается, что объем настоящего изобретения не ограничен этими примерами.
Справочный пример 1. Экспрессия в лилиях гена F3'5'H из барвинка
Плазмиду pNAVFH (см. Непатентный Документ 9, бинарный вектор для экспрессии гена F3'5'H барвинка в растениях) вводили в штамм агробактерий ЕНА101. Сорт лилий «Acapulco» (розовые цветы), трансформировали с помощью этих генетически модифицированных агробактерий. Хотя трансформацию лилий проводили способом, описанным, например, в Непатентном документе 10, используемый способ этим не ограничивается.
Требуется от двух до трех лет после получения растительной тканевой культуры трансформированных лилий для исследования того, функционирует ли введенный ген F3'5'H в лилиях и индуцирует ли аккумуляцию делфинидина. Анализ присутствия антоцианов в культивируемых черешках, был сфокусирован на наблюдении антоцианового окрашивания в черешках, что, по меньшей мере, позволяло быстро определить, что введенный ген не функционирует или функционирует лишь в небольшой степени. Из полученных растительных тканевых культур трансформированных лилий было получено 19 культиваров. Собирали приблизительно по 0,5 г черешков, в которых наблюдали антоциановое окрашивание, и подвергали анализу антоцианов с использованием способа, описанного в Патентном Документе 2. В результате не обнаружили культиваров, в которых бы детектировался делфинидин.
Справочный пример 2. Экспрессия в лилиях гена F3'5'H из цинерарии Фрагмент ДНК, длиной приблизительно 1,7 тыс.п.о., полученный гидролизом плазмиды pSPB2774, содержащей выделенную из цинерарии F3'5'H кДНК Ci5a18 (см. Патентный документ 9), по SmaI и XhoI, фрагмент ДНК, содержащий 35S-промотор вируса мозаики цветной капусты (также обозначенный как «355-промотор»), полученный гидролизом плазмиды pBI1221 no HindIII и BamHI, и фрагмент ДНК, содержащий репликон фрагмента ДНК, полученного гидролизом плазмиды pSPB176 (см. Патентный документ 7) по HindIII и SalI, лигировали с получением плазмиды pSPB3472. Фрагмент ДНК, полученный гидролизом данной плазмиды по AscI и PacI (содержащей последовательность, в которой лигировали 35S-промотор, Ci5a18 и терминатор нопалин-синтазы), лигировали с фрагментом ДНК, полученным гидролизом #493 по AscI и PacI с получением плазмиды pSPB3376. Затем с помощью агробактерий полученную плазмиду вводили в лилии, как описано в Справочном примере 1, с получением растительной тканевой культуры для 65 культиваров трансформированных лилий. Хотя в 23 из этих культиваров в черешках листьев аккумулировался антоциан, при этом не было получено ни одного культивара, в котором бы аккумулировался делфинидин.
Справочный пример 3. Экспрессия в лилиях гена F3'5'H из горечавки Фрагмент ДНК, длиной приблизительно 1,7 тыс.п.о., полученный гидролизом плазмиды pGHF48, содержащей кДНК F3'5'H из горечавки (см. Патентный Документ 4), лигировали с плазмидой pSPB176, гидролизованной по BamHI и SalI с получением плазмиды pSPB3329. Участок промотора удаляли гидролизом по HindIII и BamHI. Затем введением фрагмента ДНК, полученного гидролизом pBI1221 no HindIII и BamHI (содержащего 35S-промотор), получали плазмиду pSPB3473. Фрагмент ДНК, полученный гидролизом данной плазмиды по AscI и PacI (содержащий последовательность, в которой лигировали 35S-промотор, кДНК F3'5'H горечавки и терминатор нопалин-синтазы), лигировали с фрагментом ДНК, полученным гидролизом pSPB493 по AscI и PacI с получением плазмиды pSPB3378. Затем с помощью агробактерий полученный конструкт вводили в лилии, как описано в Справочном примере 1, с получением растительной тканевой культуры для 65 линий трансформированных лилий. Черешки листьев, в которых аккумулировался антоциан, получали из 26 из этих культиваров. Процент накопления делфинидина (соотношение количества аккумулированного делфинидина к общему количеству антоциана) достигал всего лишь максимум 1%.
Справочный пример 4. Экспрессия в лилиях гена F3'5'H из лаванды
Фрагмент ДНК, длиной приблизительно 1,7 тыс.п.о., полученный гидролизом плазмиды pGHF48, содержащей кДНК F3'5'H из лаванды (см. Патентный Документ 4), по BamHI и XhoI, лигировали с плазмидой pSPB176, гидролизованной по BamHI и SalI с получением плазмиды pSPB2772. Извлекали фрагмент ДНК, полученный гидролизом данной плазмиды по BamHI и PacI, содержащий последовательность, в которой были лигированы кДНК F3'5'H лаванды и терминатор нопалин-синтазы. Данный фрагмент ДНК лигировали с фрагментом, содержащим репликон фрагмента ДНК, полученного гидролизом плазмиды pSPB3472 (3377) по BamHI и PacI, с получением плазмиды pSPB3471. Фрагмент, ДНК полученный гидролизом данной плазмиды по AscI и PacI (содержащий последовательность, в которой были лигированы 35S-промотор, кДНК F3'5'H горечавки и терминатор нопалин-синтазы), лигировали с фрагментом ДНК, полученным гидролизом pSPB493 по AscI и PacI, с получением плазмиды pSPB3376. Затем с помощью агробактерий полученный конструкт вводили в лилии, как описано в Справочном примере 1, с получением растительной тканевой культуры для 23 культиваров трансформированных лилий. Черешки листьев, в которых аккумулировался антоциан, получали из 20 из этих культиваров. Процент накопления делфинидина (соотношение количества аккумулированного делфинидина к общему количеству антоциана) достигал всего лишь максимум 1%.
Справочный пример 5. Экспрессия в лилиях гена F3'5'H из мотылькового горошка
Сообщалось, что ген F3'5'H из мотылькового горошка (учетный номер базы данных ДНК АВ234897), введенный в вербену функционирует лучше и приводит в результате к более высокому содержанию делфинидина и к более существенному изменению цвета цветка, чем в случае введения гена F3'5'H самой вербены (см., например Непатентный Документ 5). Плазмиду pSPB748 (см., Непатентный Документ 5) гидролизовали по EcoRI, затупляли концы и гидролизовали по BamHI с последующим извлечением фрагмента ДНК, содержащего кДНК F3'5'H из мотылькового горошка. С другой стороны получали фрагмент ДНК гидролизом pSPB176 по SalI, затуплением концов и гидролизом по BamHI. Эти два фрагмента ДНК дотировали с получением плазмиды pSPB3166. Фрагмент ДНК, содержащий кДНК F3'5'H, полученный гидролизом pSPB3166 по AscI и PacI, вставляли в фрагмент ДНК, полученный гидролизом бинарного вектора pSBP3166, который содержит ген, обеспечивающий устойчивость к гигромицину (см. Непатентный Документ 11), по AscI и PacI. Полученный в результате бинарный вектор назвали pSBP3169. В растениях, трансформированных данным бинарным вектором, ген F3'5'H мотылькового горошка транскрибировался под контролем 35S-промотора вируса мозаики цветной капусты, к которому был добавлен энхансер (см., Непатентный Документ 12).
Затем с помощью агробактерий полученный конструкт вводили в лилии, как описано в Справочном примере 1, с получением растительной тканевой культуры для 23 культиваров трансформированных лилий. Ожидалось, что после введения гена F3'5'H из мотылькового горошка лилии начнут вырабатывать делфинидина. Однако, хотя антоциан аккумулировался в черешках листьев у 6 из этих культиваров, не было получено ни одного культивара, в котором бы аккумулировался делфинидин.
Справочный пример 6. Экспрессия в лилиях гена F3'5'H из фиалки
При введении в розы генов F3'5'H, выделенных из нескольких растений, только ген F3'5'H, выделенный из фиалки (учетный номер базы данных ДНК АВ332097), обеспечивал предпочтительную аккумуляцию делфинидина и вызывал существенное изменение цвета цветка (см., Патентный Документ 4). Кроме того, данный ген также вызывает аккумуляцию делфинидина в лепестках хризантем и приводит к изменению цвета цветка на синий (см., Патентный Документ 5). На основании данных фактов, предположили, что выделенный из фиалки ген F3'5'H подходит для выработки делфинидина в различных растениях.
Фрагмент ДНК, полученный гидролизом плазмиды pSPB575, содержащей кДНК F3'5'H фиалки (см., Патентный Документ 8), по AscI и PacI, и фрагмент ДНК, полученный гидролизом плазмиды pSPB3166 по AscI и PacI, дотировали с получением бинарного вектора pSBP3138. В растениях, трансформированных данным бинарным вектором, ген F3'5'H фиалки транскрибировался под контролем 35S-промотора вируса мозаики цветной капусты, к которому был добавлен энхансер.
Затем с помощью агробактерий полученный конструкт вводили в лилии, как описано в Справочном примере 1, с получением растительной тканевой культуры для 25 культиваров трансформированных лилий. Ожидалось, что после введения в лилии гена F3'5'H из фиалки, делфинидин будет вырабатываться. Однако, хотя из 12 из этих культиваров были получены черешки листьев, в которых аккумулировался антоциан, не было получено ни одного культивара, в котором бы аккумулировался делфинидин.
Пример 1
(1) Выделение Гена F3'5'H из геномной ДНК колокольчика
Хромосомную ДНК экстрагировали из листьев колокольчика (Campanula medium) с использованием «DNeasy» («Qiagen Corp.»). Нуклеотиды CFH1L: 5'-CATGTCTATAGACATATCCACCCTCTTCTATGAAC-3' (SEQ ID NO:5, подчеркиванием выделен старт-кодон) и cCFHIL: 5'-GCCTAGACAGTGTAAGAACTTGGAGGCAATCTTGG-3' (SEQ ID NO:6) синтезировали на основании базовой последовательности кДНК F3'5'H колокольчика (Campanula medium) (учетный номер базы данных ДНК D14590). ПЦР проводили используя в качестве матрицы 100 нг хромосомной ДНК и вышеупомянутые нуклеотиды в качестве праймеров. Для проведения ПЦР использовали «Takara LA PCR» («Takara Bio Inc.») в условиях, рекомендованных производителем. Полученная базовая последовательность представлена в SEQ ID NO:1. Аминокислотная последовательность SEQ ID NO:2, кодируемая данной базовой последовательностью, является той же, что и зарегистрированная в базе данных ДНК под номером No. D14590.
(2) Конструирование Бинарного Вектора, Содержащего Ген F3'5'H из Геномной ДНК Колокольчика
Плазмиду рВ1221 (см. Непатентный документ 13) гидролизовали по SmaI и SacI с последующим затуплением концов с помощью соответствующего набора реактивов (Takara Bio Inc.) и лигировали. Полученную в результате плазмиду гидролизовали по BamHI и затупляли. Затем проводили реакцию данного фрагмента ДНК с ДНК-полимеразой в присутствии дезокситмидинтрифосфатов для добавления одного основания дезокситимидина к 3'-концу данного фрагмента ДНК. Этот фрагмент ДНК лигировали с ПЦР-фрагментами, выделенными из вышеупомянутой хромосомной ДНК колокольчика. Среди полученных в результате плазмид, плазмиду, в которой старт-кодон гена F3'5'H колокольчика был вставлен в направлении, вблизи к последовательности 35S-промотора, обозначали как p35S-CFH-1. Данную плазмиду гидролизовали по PvuII и разделяли с помощью электрофореза на агарозном геле для извлечения фрагмента ДНК, содержащего ген F3'5'H колокольчика.
Плазмиду pNAVFH (см. Непатентный Документ 9, бинарный вектор для экспрессии гена F3'5'H барвинка в растениях) гидролизовали по HindIII и XbaI, затупляли и дефосфорилировали. Затем этот материал разделяли с помощью электрофореза на агарозном геле с последующим извлечением фрагмента ДНК, имеющего наибольшую молекулярную массу. Данный фрагмент ДНК лигировали с вышеупомянутым фрагментом ДНК, содержащим ген F3'5'H колокольчика, и полученный в результате бинарный вектор назвали pNARI-CFH.
(3) Выделение Фрагмента Гена F3'H Лилий (Конструирование pCR2.1-AP24-1)
Олигонуклеотиды F3H2: 5'-YTSGCYGGWGTWTTYAACRTHGG-3', эквивалентный LAGWNIG из последовательности Arabidopsis thaliana (SEQ ID NO:7), и cF3H4: 5'-GGRTCNCGRGCWATGGCCCA-3', эквивалентный NIWAIARDP последовательности Arabidopsis thaliana (SEQ ID NO:8), синтезировали на основании раннее раскрытых аминокислотных последовательностях F3'H Arabidopsis thaliana (регистрационный номер базы данных ДНК No. AF271651), астры (AF313488), сои (AF499731), периллы (АВ045593), риса (АС021892) и торении (АВ057672). В данном документе, Y представляет собой смесь С и Т, W представляет собой смесь А и Т, S представляет собой смесь С и G, R представляет собой смесь А и G, Н представляет собой смесь А, Т и С, и N представляет собой смесь G, А, Т и С. РНК экстрагировали из лепестков сорта лилии «Acapulco» (Yamaki Noen Co., Ltd.), с использованием набора реактивов «RNeasy» («Qiagen Corp.»). ОТ-ПЦР проводили с использованием РНК в качестве матрицы и с использованием F3H2 и олиго(dT) в качестве праймеров. ОТ-ПЦР проводили с использованием набора реактивов для ОТ-ПЦР («Invitrogen Corp.»). ПЦР проводили, как описано ранее, с использованием в качестве матрицы амплифицированного фрагмента ДНК и с использованием F3H2 и cF3H4 в качестве праймеров. Полученный в результате фрагмент ДНК обозначили как SEQ ID NO:3. Фрагмент ДНК с данной последовательностью, затем лигировали с плазмидой pCR2.1 («Invitrogen Corp.»). Полученную плазмиду назвали pCR2.1-AP24-l.
(4) Конструирование Плазмиды pNARI-Tr3
Плазмиду pCR2.1-AP24-l гидролизовали по SpeI и XbaI, затупляли и разделяли с помощью электрофореза на агарозном геле. Фрагмент ДНК длиной 665 п.о., извлекали из полученных фрагментов ДНК. Данный фрагмент ДНК лигировали с фрагментом ДНК, полученным гидролизом плазмиды pBI221 no ScaI и MluI, с последующим затуплением концов и дефосфорилированием. Полученную плазмиду назвали p35S-GUSL-AP24.
Плазмиду pCR2.1-AP24-1 гидролизовали по SpeI и NotI, затупляли и разделяли с помощью электрофореза на агарозном геле. Фрагмент ДНК длиной 647 п.о. извлекали из полученных в результате фрагментов ДНК. Данный фрагмент ДНК лигировали с фрагментом ДНК, полученным гидролизом плазмиды p35S-GUSL-AP24 no SmaI, с последующим дефосфорилированием, и полученную в результате плазмиду назвали p35S-F3'HTr2.
Плазмиду p35S-F3'HTr2 гидролизовали по PvuII и разделяли с помощью электрофореза на агарозном геле с последующим извлечением фрагмента ДНК размером приблизительно 3,3 тыс. п.о. С другой стороны, плазмиду pNARI-CFH гидролизовали по ClaI и NheI, затупляли, дефосфорилировали и разделяли с помощью электрофореза на агарозном геле с последующим извлечением фрагмента ДНК, содержащего репликон. Извлеченный фрагмент ДНК лигировали с вышеупомянутым фрагментом ДНК размером приблизительно 3,3 тыс. п.о. Полученную в результате плазмиду или бинарный вектор назвали pNARI-Tr3 (который также можно проще обозначить как «Tr3»). Как показано на фиг.2, данный бинарный вектор содержит в качестве маркера селекции ген фосфотрансферазы гигромицина на Т-ДНК, и он предназначен для подавления экспрессии гена F3'H лилий с помощью конститутивно экспрессирующегося гена F3'5'H колокольчика и транскрибирования двухцепочечной РНК гена F3'H лилий.
(5) Трансформация лилий
Каждый из бинарных векторов pNARI-CFH и pNARI-Tr3 вводили в штамм агробактерий ЕНА101. Сорт лилий «Acapulco» (розовые цветы) трансформировали с использованием данных генетически модифицированных агробактерий, как описано в Справочном примере 1. В случае использования агробактерий, содержащих pNARI-CFH, получали растительные тканевые культуры 31 линии лилий. Анализ антоцианидинов в черешках, в которых аккумулировался антоциан, выявил, что делфинидин аккумулировался на уровне 5% или более от суммарного количества антоцианинов в 4 культиварах. Максимальное содержание делфинидина составило 18%.
С другой стороны, в случае использования агробактерий, содержащих pNART-Tr3, получали растительные тканевые культуры 40 линий лилий. Анализ антоцианидинов в черешках, в которых аккумулировался антоциан, выявил, что делфинидин аккумулировался на уровне 5% или более от суммарного количества антоцианинов в 13 культиварах, как показано в таблице 1 ниже. Когда нескольким из этих генетически модифицированных лилий позволили зацвести в парнике для культивирования генетически модифицированных растений, содержание делфинидина в лепестках достигло максимум 80%, и в отличие от исходного цвета данного сорта, имеющей номер по цветовой шкале N74D, согласно классификации Британского Королевского общества садоводства, цвет генетически модифицированных лилий стал синим по сравнению с исходным цветом, что соответствует номеру 81C по цветовой шкале.
Кроме того, также в генетически модифицированных растениях детектировали пеларгонидин, который не наблюдался в растении-хозяине. Предполагается, что это эффект подавления гена F3'H.
Аналогичным образом трансформировали с использованием агробактерий, содержащих pNARI-Tr3, лилии «Tiara» (Yamaki Noen Co., Ltd.) (также обозначаемая как «TI»), сорт лилий R094-19 (Niigata Agricultural Research Institute) (также обозначаемая как «RO») и МР8 (предоставлена Mr. Miwa Fujii), и получили 38, 82 и 12 генетически модифицированных культиваров, соответственно. Анализ антоцианидинов в черешках листа, в которых аккумулировались антоцианины в данных культиварах, выявил, что среднее содержание делфинидина составило 24%, 8% и 11%, соответственно, а максимальное значение содержание делфинидина составило 68%, 53% и 35%, соответственно.
Анализ значений и цвета цветков для цветущих лилий TI («Tiara») и RO (R094-19), представлен в таблице 2 ниже. Максимальное содержание делфинидина в лепестках цветущей генетически модифицированного сорта «Tiara», составило 74% (индивидуальный no. TI Tr3 No. 42). Максимальное содержание делфинидина в лепестках цветущей генетически модифицированной R094-19, составило 15% (индивидуальный по. RO Tr3 No. 9).
Кроме того, антоцианины, присутствующие в основном сорте «Acapulco», и в генетически модифицированной лилии (также обозначаемой как «Acapulco Tr3-5»), анализировали с помощью LC-FTICR-MS (см. Непатентный Документ 14 и Непатентный Документ 15). Использование данного метода позволяет провести точное измерение масс-спектра антоцианинов и получить MS/MS-спектры с помощью тандемной масс-спектрометрии. В основном сорте «Acapulco» пики, соответствующие цианидину, детектировали при m/z 595,166612 (MS/MS m/z: 287,1 (10%), 432,8 (0.9%), 449,1 (14.7%) (данные в скобках обозначают относительные значения пиковой интенсивности), связанного с 1 эквивалентом рамнозы и глюкозы, и содержали основные антоцианины. С другой стороны, в случае «Acapulco Tr3-5», наблюдались два пика, соответствующие делфинидину, детектируемого при m/z 611,160529 (MS/MS mz: 303,1 (100%), 449.2 (1,1%), 465,1 (10,8%)) и 611,160902 (MS/MS m/z: 303,1 (100%), 449,1 (0.7%), 465,1 (12.3%)), связанного с 1 эквивалентом рамнозы и глюкозы, которые отсутствовали в основном сорте «Acapulco». Кроме того, наблюдали увеличение пика, соответствующего пеларгонидину, который основном сорте «Acapulco» находился на чрезвычайно низком уровне, связанному с одним эквивалентом рамнозы и глюкозы (детектировали m/z: 579,171847 (MS/MS m/z: 271,1 (100%), 417,1 (0.8%), 433,1 (18,4%)).
Пример 2. Экспрессия в лилиях кДНК F3'5'H из колокольчика
(1) Выделение кДНК F3'5'H колокольчика
Два типа праймеров, CamF1: 5'-GTGAAGCCACCATGTCTATAG-3' (SEQ ID NO:9) и CamR1: 5'-GCATTTGCCTAGACAGTGTAAG-3' (SEQ ID NO:10), синтезировали на основании транслируемой последовательности кДНК F3'5'H колокольчика (Campanula medium), зарегистрированной в базе данных ДНК GenBank (учетный номер в базе данных ДНК D14590). С помощью набора реактивов «RNeasy Mini Plant Kit» («Qiagen Corp.») экстрагировали РНК из лепестков бутонов коммерчески доступных колокольчиков, а с помощью набора реактивов для ОТ-ПЦР синтезировали 1-ую цепь ДНК. Затем проводили ПЦР с праймерами, при этом в качестве матрицы использовали 1-ую цепь ДНК. Полученные в результате фрагменты ДНК клонировали в pCR-TOPO II. Среди этих клонов определили, что основная последовательность кДНК клона #4 (обозначенного pSPB2561) соответствует SEQ ID NO:11. В отличие от F3'5'H, зарегистрированного в базе данных ДНК под учетным номером D14590, и состоящим из 523 остатков (SEQ ID NO:2), F3'5'H, кодируемая кДНК-клоном #4, состояла из 521 остатка (SEQ ID NO:12), и демонстрировала 95% идентичность к F3'5'H, зарегистрированной под номером D14590.
Ген F3'5'H генома колокольчика содержится в бинарном векторе, введенном в лилии в примере 1. Данная последовательность (SEQ ID NO:1) содержит интрон. Конструировали бинарные векторы, в которых данный геномный ген F3'5'H заменяли последовательностью, выделенной из вышеупомянутого клона #4 (не содержащего интрон). Бинарный вектор, не содержащий двухцепочечной ДНК F3'5'H, назвали pNARI-cCFH (также обозначенный как «сС»), тогда как бинарный вектор, содержащий двухцепочечную ДНК F3'H, назвали pNARI-cTr3 (также обозначенный как «cTr32»).
Как показано в таблице 3 ниже, с помощью агробактерий, содержащих pNARI-cCFH, получили растительные тканевые культуры для 42 культиваров генетически модифицированных лилий. Получали черешки, в которых накапливаются антоцианины. Анализ антоцианидинов в черешках, где накапливаются антоцианины, выявил, что делфинидин аккумулировался на уровне 5% или более от суммарного количества антоцианидинов в 16 культиварах. Максимальное содержание делфинидина составило 36%.
Как показано в следующих таблицах 4-6, сорта лилий «Acapulco», R094-19, «Tiara» и МР8 трансформировали агробактериями, содержащими pNARI-cTr3, и получали 123, 46, 33 и 2 культивара генетически модифицированных растений, соответственно. Среднее содержание делфинидина в черешках, в которых накапливаются антоцианины, составило 10%, 15%, 20% и 4%, соответственно, а максимальное содержание делфинидина составило 63% (индивидуальный по. АР cTr3 No. 80), 48%, 54% и 7%, соответственно. Результаты анализа суммированы в Таблицах 4-6 ниже. При цветении генетически модифицированных разновидностей «Acapulco» анализ антоцианидинов в лепестках показал, что содержание делфинидина в культиваре, обозначенном как No. 80-1, составило 73%. Цвет его цветка был аналогичен цвету генетически модифицированной разновидности «Acapulco» (описанной в примере 1), который демонстрировал содержание делфинидина 80%.
Пример 3. Конструирование Tr3M и Трансформация Лилий]
Терминатор гена F3'5'H колокольчика, содержащийся в Т-ДНК pNARI-Tr3, представляющий собой терминатор, выделенный из гена нопалин-синтазы, заменяли на терминатор, выделенный из гена манопин-синтазы (Патентный документ 1). Полученный в результате вектор назвали pTr3M. Лилии сорта «Acapulco», R094-19, «Tiara» и МР8 трансформировали агробактериями, содержащими данный вектор, и получали, 6, 19, 71 и 11 культиваров генетически модифицированных растений, соответственно. Среднее содержание делфинидина в черешках, в которых накапливаются антоцианины, составило 38%, 34%, 7% и 22%, соответственно, а максимальное содержание делфинидина составило, соответственно, 72%, 70%, 54% и 65%.
Пример 4. Конструирование Tr7 и Трансформация Лилий
(1) Получение гена Дигидрофлавонол-4-Редуктазы (DFR) из Лилий
кДНК-библиотеку, выделенную из лепестков лилий сорта «Aktiva» (Yamaki Noen Co., Ltd.), получали гидролизом вектора Lambda ZAP II no EcoRI и XhoI, и с помощью дефосфорилированного вектора Uni-ZAP XR для направленного клонирования («Agilent Technologies Inc.»). Способ, описанный в WO 01/092509, может использоваться, например, для получения кДНК-библиотеки. В результате скрининга полученной кДНК-библиотеки с использованием меченой кДНК DFR герани (учетный номер GenBank No. AB534774, как описано в Plant Biotechnology (2010) 27, 375-383) получили плазмиду обозначенную как pSPB3165, которая содержит кДНК DFR лилии (SEQ ID NO:13) между сайтами ферментов рестрикции EcoRI и XhoI вектора pBluescriptII. Скрининг кДНК-библиотеки можно проводить, например, согласно способу, описанному в WO 81/092509.
(2) Получение кДНК F3'H лилии
Вышеупомянутую кДНК-библиотеку лепестков лилий подвергали скринингу с использованием фрагмента ДНК F3'H, содержащегося в pCR2.1-AP24-1, описанном в примере 1. Плазмиду, содержащую кДНК F3'H, назвали pSBP3175. Так как данная кДНК не является полноразмерной, то отсутствующую последовательность на 5'-конце амплифицировали с использованием набора реактивов «Gene Racer Kit» («Invitrogen Corp.») согласно способу, рекомендованному производителем. Синтетический олигонуклеотид LiRG-R3: 5'-TGCTCCGCTATCAACTTATC-3' (SEQ ID NO:15) использовали для реакции ПЦР. Полученный в результате фрагмент ДНК клонировали в pCR-TOPOII с получением плазмиды pSPB3194. Фрагмент ДНК размером приблизительно 700 п.о., полученный из данной плазмиды гидролизом по PstI и SspI, вставляли между сайтами PstI и SspI плазмиды pSPB3175 с получением плазмиды pSPB3198. Синтезированная кДНК F3'H лилии (SEQ ID NO:16) содержится в данной плазмиде.
(3) Конструирование вектора
Конструировали Бинарный вектор Tr7 для подавления экспрессии генов F3'H и DFR лилий и для экспрессии гена F3'5'H колокольчика и гена DFR ириса.
Кассету для подавления экспрессии гена DFR лилии конструировали способом, описанным ниже. Фрагмент ДНК размером приблизительно 0,5 тыс. п.о., полученный гидролизом плазмиды pSPB3165 no EcoRI и PstI (содержащей последовательность, кодирующую фрагмент, расположенный поблизости от N-конца кДНК DFR лилии), клонировали между сайтами EcoRI и PstI плазмиды pBluescriptSK- («Agilent Technologies Inc.»). Плазмиду pSPB3168 получали лигированием фрагмента ДНК размером приблизительно 0,5 тыс. п.о., полученного гидролизом данной плазмиды по BamHI и SalI (содержащего последовательность, кодирующую фрагмент, расположенный поблизости от N-конца кДНК DFR лилии), фрагмента ДНК размером приблизительно 3,5 тыс. п.о., полученного гидролизом плазмиды pSPB906 (описанной в WO 2005/017147) по BamHI и EcoRI, и фрагмента ДНК размером приблизительно 300 п.о., полученного гидролизом плазмиды pSPB176 по SalI и EcoRI (содержащего терминатор нопалин-синтазы). Плазмиду pSPB3178 получали лигированием фрагмента ДНК размером приблизительно 0,8 тыс. п.о., полученного гидролизом вышеупомянутой плазмиды pSPB3165 по BamHI и EcoRV (содержащего последовательность, кодирующую часть N-концевого участка кДНК DFR), с плазмидой pSPB3168, гидролизованной по BamHI и SmaI. Плазмида содержит кассету с 35S-промотором, к которому добавили энхансер, для обеспечения конститутивной транскрипции двухцепочечной РНК, соответствующей кДНК DFR лилии. Плазмиду, в которой 35S-промотор, к которому был добавлен энхансер плазмиды pSPB3178, заменяли на последовательность 35S-промотора, обозначали как pSPB3456.
Плазмиду, в которой последовательность 35S-промотора, к которому был добавлен энхансер плазмиды pSPB912 (описанной в WO 2005/017147), заменяли на последовательность 35S-промотора, назвали pSPB3419. Данную плазмиду использовали для конститутивной экспрессии кДНК DFR ириса. Кассету для подавления экспрессии гена F3'H лилии конструировали способом, описанным ниже. После гидролиза pSPB3198 по HaeII, ее дополнительно гидролизовали по BamHI с последующим извлечением фрагмента ДНК, содержащего 5'-область кДНК F3'H. Затем, pSPB3194 гидролизовали по HaeII с последующим затуплением, затем гидролизовали по XhoI и извлекали фрагмент ДНК, содержащий 5'-область кДНК F3'H. Эти фрагменты ДНК лигировали с pSPB912, гидролизованной по BamHI и SalI с получением плазмиды pSPB3200. Плазмиду, в которой последовательность 35S-промотора, к которому был добавлен энхансер плазмиды pSPB3200, заменяли на последовательность 355-промотора, назвали pSPB3455. Транскрипционные кассеты, соответственно, извлекали из pSPB3456, pSPB3419 и pSPB3455 и использовали для конструирования Tr7.
Tr7 содержит пять транскрипционных кассет на Т-ДНК, которые, если двигаться от правой границы последовательности, состоят из (i) последовательности 35S-промотора, последовательности для транскрибирования двухцепочечной РНК кДНК DFR лилии и терминатора нопалин-синтазы, (ii) последовательности 35S-промотора, последовательности гена F3'5'H колокольчика, содержащего интрон, и терминатора нопалин-синтазы (таких же как и в Tr3), (iii) последовательности 35S-промотора, последовательности гена DFR ириса и терминатора нопалин-синтазы, (iv) последовательности 35S-промотора, последовательности для транскрибирования двухцепочечной РНК с кДНК F3'H лилии и терминатора нопалин-синтазы (таких же как и в Tr3), и (v) последовательности 35S-промотора, последовательности гена фосфотрансферазы гигромицина, и терминатора нопалин-синтазы (таких же как и в Tr3). Карта Tr7 показана на фиг.3.
(4) Получение Генетически Модифицированных Лилий
Лилии сорта «Acapulco», R094-19, «Tiara» и МР8 трансформировали агробактериями, содержащими Tr7, и получали, соответственно, 22, 80, 32 и 11 культиваров генетически модифицированных растений. Среднее содержание делфинидина в черешках, в которых накапливаются антоцианины, составило 15%, 13%, 6% и 28%, соответственно, а максимальное содержание делфинидина составило 78%, 88%, 56% и 87%, соответственно.
Генетически модифицированный культивар R094, R094-19 Tr7 No. 52-3, имел содержание делфинидина 99%.
Пример 5. Конструирование Tr3 8 и Трансформация Лилий
Получали бинарный вектор Tr8, в котором порядок экспрессирующей кассеты на Т-ДНК отличается от порядка бинарного вектора Tr7. Tr8 содержит пять транскрипционных кассет на Т-ДНК, которые, если двигаться от правой границы последовательности, включают (i) последовательности 35S-промотора, последовательности гена F3'5'H колокольчика, содержащего интрон, и терминатора нопалин-синтазы (таких же как и в Tr3), (ii) последовательности 35S-промотора, последовательности гена DFR ириса и терминатора нопалин-синтазы, (iii) последовательности 35S-промотора, последовательности для транскрибирования двухцепочечной РНК кДНК DFR лилии и терминатора нопалин-синтазы (таких же как и в Tr7), (iv) последовательности 35S-промотора, последовательности для транскрибирования двухцепочечной РНК с кДНК F3'H лилии и терминатора нопалин-синтазы (таких же как и в Tr7), и (v) последовательности 35S-промотора, последовательности гена фосфотрансферазы гигромицина, и терминатора нопалин-синтазы (такой же как в Tr3). Карта Tr8 показана на фиг.3. Лилии сорта «Acapulco», R094-19 и «Tiara» трансформировали агробактериями, содержащими Tr7, и получали, соответственно, 66, 6 и 87 культиваров генетически модифицированных растений. Среднее содержание делфинидина в черешках, в которых накапливаются антоцианины, составило 15%, 6% и 30%, соответственно, и максимальное содержание делфинидина составило, 51%, 6% и 70%, соответственно.
Пример 6. Конструирование Tr39 и Трансформация Лилий
Конструировали бинарный вектор Tr9, который экспрессирует кДНК F3'5'H колокольчика. Карта Tr9 показана на фиг.3. Tr9 содержит пять транскрипционных кассет на Т-ДНК, которые, если двигаться от правой границы последовательности, состоят из (i) терминатора нопалин-синтазы (такого же как и в Tr3), последовательности гена F3'5'H колокольчика и последовательности 35S-промотора, (ii) последовательности 35S-промотора, последовательности для транскрибирования двухцепочечной РНК кДНК DFR лилии и терминатора нопалин-синтазы (таких же как и в Tr7, но в другой ориентации), (iii) терминатора нопалин-синтазы, последовательности для транскрибирования двухцепочечной РНК с кДНК F3'H лилии и последовательности 35S-промотора (таких же как и в Tr7, но в другой ориентации), (iv) терминатора нопалин-синтазы, последовательности для транскрибирования двухцепочечной РНК с кДНК F3'H лилии (такой же как в Tr7, но в другой ориентации) и (v) последовательности 35S-промотора, последовательности гена фосфотрансферазы гигромицина, и терминатора нопалин-синтазы (такого же как и в Tr3). Разновидности лилий «Acapulco», R094-19 и «Tiara» трансформировали агробактериями, содержащими Tr7, и получали, соответственно, 8, 51 и 33 культиваров генетически модифицированных растений. Среднее содержание делфинидина в черешках, в которых накапливаются антоцианины, составило 2%, 12% (анализировали 44 черешка из 51 культивара) и 13%, соответственно, а максимальное содержание делфинидина составило 9%, 68% и 38%, соответственно.
Пример 7. Конструирование Tr310 и Трансформация Лилий
Tr10 содержит пять транскрипционных кассет на Т-ДНК, которые, если двигаться от правой границы последовательности, состоят из (i) последовательности 35S-промотора, последовательности кДНК гена F3'5'H колокольчика, и терминатора нопалин-синтазы (таких же как и в Tr3), (ii) последовательности 35S-промотора, последовательности гена DFR ириса и терминатора нопалин-синтазы (таких же как и в Tr7), (iii) последовательности 35S-промотора, последовательности для транскрибирования двухцепочечной РНК с кДНК DFR лилии и терминатора нопалин-синтазы (таких же как и в Tr7), (iv) последовательности 35S-промотора, последовательности для транскрибирования двухцепочечной РНК с кДНК F3'H лилии и терминатора нопалин-синтазы (таких же как и в Tr7), и (v) последовательности 35S-промотора, последовательности гена фосфотрансферазы гигромицина, и терминатора нопалин-синтазы (таких же как и в Tr3). Карта Tr10 показана на фиг.3. Лилии сорта «Acapulco», R094-19, «Tiara» и МР8 трансформировали агробактериями, содержащими Tr10, и получали, соответственно, 41, 14, 78 и 10 культиваров генетически модифицированных растений. Среднее содержание делфинидина в черешках, в которых накапливаются антоцианины, составило 23%, 26%, 15% (анализировали черешки из 19 культиваров) и 36%, соответственно, а максимальное содержание делфинидина составило, 80%, 87%, 75% и 74%, соответственно.
Цвет цветка генетически модифицированного культивара «Acapulco», АР Tr10 No. 34-1, существенно изменился (номер по цветовой шкале N78A), и содержание делфинидина в лепестках составило 82%.
Пример 8. Конструирование Tr311 и Трансформация Лилий
(1) Выделение Промоторной Последовательности гена лилии DFR
Выделение промоторной последовательности гена лилии DFR было проведено с помощью обратной ПЦР. Четыре праймера, cLD2L: 5'-AGGGTCTCTCACAGTAGCCCGAACCGTGTAGCCA-3' (SEQ ID NO:18), LD1L: 5'-ACTAGATCTTCCTGGAGCTGATGAACGGCTAACC-3' (SEQ ID NO:19), cLDIL: 5'-AACATAGCCACTAGCTCCAGTAACCACCACGGGTC-3' (SEQ ID NO:20) и LD3L: 5'-AGCAAATCCAGCTTGTTCACTTGGATGACGTATGC-3' (SEQ ID NO:21) были синтезированы на основании кДНК последовательности DFR из сорта лилий «Montreux» (розовые цветы), зарегистрированной в базе данных ДНК GenBank (учетный номер базы данных ДНК АВ058641). ДНК выделяли из листьев лилий сорта «Montreux» (Yamaki Noen Co., Ltd.) с помощью набора реактивов «DNeasy Plant Mini Kit» («Qiagen Corp.»). После гидролиза полученной ДНК по EcoRI, получали циклическую ДНК с помощью реакции самолигирования фрагментов ДНК с использованием набора реактивов «DNA Ligation Kit» («Mighty Mix», «Takara Bio Inc.»). Первый раунд реакции ПЦР проводили с использованием «Takara LA Taq Hot Start Version» («Takara Bio Inc.») применяя в качестве матрицы циклическую ДНК и используя в качестве праймеров cLD2L и LD1L. При этом второй раунд ПНР-реакции проводили с использованием «Takara LA Taq Hot Start Version» с применением в качестве матрицы амплифицированной ДНК и cLD1L и LD3L в качестве праймеров с получением фрагмента ДНК, содержащего участок промотора DFR. Полученный фрагмент ДНК дотировали в плазмиду pCR2.1 («Invitrogen Corp.»). Полученную плазмиду назвали pCR-LDE. Кроме того, промоторную последовательность гена DFR лилии (SEQ ID NO:24) амплифицировали с использованием данной плазмиды в качестве матрицы и с использованием синтетических нуклеотидов AscI-HindIII-LDE6-F (5'-GGCGCGCCAAGCTTGAATTCTTAGCGGAAATTATCC-3' (SEQ ID NO:22) и XbaI-LDE6-R (5'-TCTAGAATCTCTCTCTCTCCCCCTC-3' (SEQ ID NO:23) в качестве праймеров. Фрагмент ДНК, содержащий эту последовательность, может быть извлечен обработкой по HindIII и XbaI.
(2) Конструирование вектора и Трансформация Лилии
Tr11 содержит пять транскрипционных кассет на Т-ДНК, которые, если двигаться от правой границы последовательности, состоят из (i) последовательности промотора гена DFR, последовательности кДНК гена F3'5'H колокольчика, и терминатора нопалин-синтазы, (ii) последовательности 35S-промотора, последовательности гена DFR ириса и терминатора нопалин-синтазы (таких же как и в Tr7), (iii) последовательности 35S-промотора, последовательности для транскрибирования двухцепочечной РНК с кДНК DFR лилии и терминатора нопалин-синтазы (таких же как и в Tr7), (iv) последовательности 35S-промотора, последовательности для транскрибирования двухцепочечной РНК с кДНК F3'H лилии и терминатора нопалин-синтазы (таких же как и в Tr7), и (v) последовательности 35S-промотора, последовательности гена фосфотрансферазы гигромицина, и терминатора нопалин-синтазы (таких же как и в Tr3). Карта Tr11 показана на фиг.3. Лилии сорта R094-19, «Tiara» и МР8 трансформировали агробактериями, содержащими Tr11, и получали, соответственно, 66, 120 и 18 культиваров генетически модифицированных растений. Среднее содержание делфинидина в черешках, в которых накапливаются антоцианины, составило 40%, 14% и 38%, соответственно, а максимальное содержание делфинидина составило 100% (анализировали 52 культивара), 54% (анализировали 14 культиваров) и 92% (анализировали 11 культиваров), соответственно.
Промышленная применимость
Согласно настоящему изобретению, делфинидин, не обнаруживаемый в природе в лилиях, может продуцироваться в них. Такие лилии, в которых окраска цветов изменяется на синий, могут быть получены подавлением экспрессии одного или несколько эндогенных генов флавоноидсинтазы в лилиях, и экспрессией гена F3'5'H или гена F3'5'H и гена DFR. Таким образом, настоящее изобретение предпочтительно может быть использовано в таких отраслях, как индустрия выращивания цветущих растений.
Краткое описание условных обозначений
RB Правая граница
LB Левая граница
35Spro 35S-промотор вируса мозаики цветной капусты
NOSter Терминатор нопалин-синтазы
LF3'hf фрагмент гена F3'H лилии
CF3'5'H, ген F3'5'H колокольчика
IrisDFR ген DFR ириса
cCF3'5'H, кДНК F3'5'H колокольчика
LDFR, DFR лилии
НРТ, ген устойчивости к гиграмицину.























Изобретение относится к области биохимии, в частности к способу получения лилий, содержащих в лепестках делфинидин. При этом способ включает введение в лилии гена флавоноид-3′,5′-гидроксилазы (F3′5′H) из колокольчиков, введение фрагмента гена флавоноид-3′-гидроксилазы (F3′H) из лилий, введение фрагмента гена дигидрофлавонол 4-редуктазы (DFR) из лилий, синтез делфинидина в результате деятельности введенного гена F3′5′H, с подавлением при этом экспрессии эндогенного гена F3′H, который участвует в синтезе цианидина в лепестках лилий, и получение лилий, которые содержат дельфинидин в лепестках. Изобретение позволяет эффективно получать лилии, содержащие в лепестках делфинидин. 4 з.п. ф-лы, 3 ил., 6 табл., 8 пр.
1. Способ получения лилий, содержащих в лепестках делфинидин, включающий следующие стадии:
введение в лилии гена флавоноид-3′,5′-гидроксилазы (F3′5′H) из колокольчиков, состоящего из нуклеотидной последовательности, представленной в SEQ ID NO: 1 или SEQ ID NO: 11, или нуклеотидной последовательности, идентичной, по меньшей мере, на 98% нуклеотидной последовательности, представленной в SEQ ID NO: 1 или SEQ ID NO: 11, которая также кодирует полипептид, обладающий активностью F3′5′H,
введение фрагмента гена флавоноид-3′-гидроксилазы (F3′H) из лилий, состоящего из нуклеотидной последовательности, представленной в SEQ ID NO: 3 или SEQ ID NO: 16, или нуклеотидной последовательности, идентичной, по меньшей мере, на 98% нуклеотидной последовательности, представленной в SEQ ID NO: 3 или SEQ ID NO: 16, которая также кодирует полипептид, обладающий активностью F3′H, и
введение фрагмента гена дигидрофлавонол 4-редуктазы (DFR) из лилий, состоящего из нуклеотидной последовательности, представленной в SEQ ID NO: 13, или нуклеотидной последовательности, идентичной, по меньшей мере, на 98% нуклеотидной последовательности, представленной SEQ ID NO: 13, которая также кодирует полипептид, имеющий активность DFR, подавление экспрессии эндогенного DFR, которая оказывает влияние на синтез цианидина в лепестках лилий, и одновременное введение выделенного инородного гена DFR из ириса,
синтез делфинидина в результате деятельности введенного гена F3′5′H, с подавлением при этом экспрессии эндогенного гена F3′H, который участвует в синтезе цианидина в лепестках лилий, и
получение лилий, которые содержат дельфинидин в лепестках.
2. Способ по п. 1, в котором подавление экспрессии эндогенного F3′H проводится способом RNAi.
3. Способ по п. 1, в котором полученный из колокольчика ген F3′5′H состоит из нуклеотидной последовательности, представленной в SEQ ID NO: 1 или SEQ ID NO: 11.
4. Способ по п. 1, в котором фрагмент полученного из лилии гена F3′H состоит из нуклеотидной последовательности, представленной в SEQ ID NO: 3 или SEQ ID NO: 16.
5. Способ по любому из пп. 1 или 2, дополнительно включающий: использование последовательности промотора DFR лилий, представленного в SEQ ID NO: 24.
| WO2005017147 A1, 24.02.2005 | |||
| TANAKA Y et al., Flower color modification by engineering of the flavonoid biosynthetic pathway: practical perspectives, Biosci Biotechnol Biochem | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| OKINAKA Y | |||
| et al., Selective Accumulation of Delphinidin Derivatives in Tobacco Using a Putative Flavonoid 3',5'-Hydroxylase cDNA from | |||

