Настоящая заявка является родственной и испрашивает приоритет предварительной заявки США с серийным номером 61/597486, поданной 10 февраля 2012, описание которой включено в настоящий документ в качестве ссылки в полном объеме.
Список последовательностей
Настоящая заявка содержит список последовательностей, предлагаемый в формате ASCII при посредстве EFS-Web и включенный в настоящий документ в качестве ссылки в полном объеме. Указанная копия ASCII, созданная 28 января 2013, озаглавлена P4733R1WO_PCTSequenceListing.txt и имеет размер 7501 байт.
Область изобретения
Настоящее изобретение относится к новым рекомбинантным белкам и белковым комплексам, включающим гетеромультимеры (например, одноцепочечные антитела, многоцепочечные антитела и комплексы иммуноадгезин-антитело), которые обладают моно- или мультиспецифичностью, а также к способам их конструирования и получения. Настоящее изобретение также относится к новому применению технологий, используемых для получения моно- или мультиспецифичных гетеромультимеров. Гетеромультимеры, полученные с помощью описанных в настоящем описании способов, можно использовать в качестве терапевтического средства для лечения любого заболевания или патологического состояния, а также для любого другого применения, в котором желательно использовать антитело.
Уровень техники изобретения
Разработка технологий получения антител или других гетеромультимеров с разными характеристиками связывания (например, моноспецифичных или мультиспецифичных), которые можно использовать и масштабировать в коммерческих и терапевтических целях, является труднодостижимой. Многие способы проходили испытания, однако почти все они имеют существенные недостатки, включающие, в числе прочих проблем, плохую растворимость или невозможность экспрессии в клетках млекопитающих, а также низкий выход образующегося гетеродимера при техническом переходе к промышленному получению, или иммуногенность, короткий период полужизни in vivo или нестабильность гетеродимера (например, Hollinger et al., (1993) PNAS 90:6444-6448; US 5,932,448; US 6,833,441; US 5,591,828; US7, 129,330; US 7,507,796; Fischer et al., (2007) Pathobiology 74:3-14; Booy (2006) Arch. Immunol. Ther. Exp. 54:85-101; Cao et al., (2003) 55: 171-197; and Marvin et al., (2006) Current Opinion in Drug Discovery & Development 9(2): 184-193). Таким образом, существует потребность в усовершенствованных технологиях и процессах для получения антител или других гетеромультимеров с различными характеристиками связывания.
Сущность изобретения
Настоящее изобретение предлагает гетеромультимеры (например, новые одноцепочечные антитела (scAb), многоцепочечные антитела (mcAb) и комплексы иммуноадгезин-антитело), а также способы конструирования, промышленного получения и применения гетеромультимеров. В одном из аспектов изобретение описывает гетеромультимерное одноцепочечное антитело, содержащее линкер (tether) гетеродимеризации (HD), который соединяет первый вариабельный домен тяжелой цепи (VH) со вторым доменом VH, где гетеромультимер содержит один или несколько константных доменов тяжелой цепи (CH), выбранных из первого домена CH2, первого домена CH3, второго домена CH2 и второго домена CH3. В одном из вариантов осуществления гетеромультимер содержит по меньшей мере одну пару константных доменов тяжелой цепи. В другом варианте осуществления гетеромультимер может содержать шарнирный домен, расположенный между доменами VH и CH2 на одной или обоих тяжелых цепях. В другом варианте осуществления гетеромультимер содержит первый и/или второй домен CH1. Один или два домена CH1 связаны по C-концу с одним или обоими доменами VH и по N-концу с одним или обоими шарнирными доменами, или с одним или обоими доменами CH2 в отсутствии шарнирных доменов. В конкретных вариантах осуществления гетеромультимер также может содержать один или два вариабельных домена легкой цепи (VL), которые связаны по N-концу с первым и/или вторым доменом VH посредством одного или двух линкеров CLH (линкер, соединяющий родственные цепи LC и HC). В некоторых вариантах осуществления гетеромультимер также содержит один или два константных домена легкой цепи (CL), каждый из которых связан по C-концу с одним или обоими доменами VL и непосредственно по N-концу с одним или обоими линкерами CLH.
В другом аспекте изобретение описывает гетеромультимерное одноцепочечное антитело, содержащее один полипептид, в состав которого входят следующие домены, располагающиеся относительно друг друга в направлении от N-конца к С-концу в следующем порядке: VL1-CL1-CLH линкер1-VH1-CH11-шарнир1-CH21-CH31-HD линкер-VL2-CL2-CLH линкер2-VH2-CH12-шарнир2-CH22-CH32.
В другом аспекте изобретение описывает гетеромультимер, представляющий собой многоцепочечное антитело, которое содержит три полипептидные цепи, где первая и вторая полипептидные цепи являются идентичными и каждая образует легкую цепь (LC), а третья полипептидная цепь образует первую тяжелую цепь (HC) и вторую HC. Каждая из первой и второй полипептидных цепей содержит домены VL и CL. Третья полипептидная цепь содержит два домена VH, линкер HD, один или два шарнирных домена и один или несколько константных доменов тяжелой цепи, выбранных из группы, включающей первый домен CH1, первый домен CH2, первый домен CH3, второй домен CH1, второй домен CH2 и, второй домен CH3, где компоненты второй полипептидной цепи располагаются относительно друг друга в направлении от N-конца к С-концу в следующем порядке: VH1-необязательный CH11-необязательный шарнир1-необязательный CH21-необязательный CH31-HD линкер-VH2-необязательный CH12-необязательный шарнир2-необязательный CH22-необязательный CH32.
В другом аспекте изобретение описывает гетеромультимер, представляющий собой многоцепочечное антитело, которое содержит две полипептидные цепи, где первая полипептидная цепь образует первую легкую цепь (LC), а вторая полипептидная цепь образует первую тяжелую цепь (HC), вторую LC и вторую HC. Первая полипептидная цепь содержит первый домен VL и CL. Вторая полипептидная цепь содержит два домена VH, линкер HD, второй домен VL, второй домен CL, линкер CLH, один или два шарнирных домена, а также один или несколько константных доменов тяжелой цепи, выбранных из группы, включающей первый домен CH1, первый домен CH2, первый домен CH3, второй домен CH1, второй домен CH2 и второй домен CH3, где компоненты второй полипептидной цепи располагаются относительно друг друга в направлении от N-конца к С-концу в следующем порядке: VH1-необязательный CH11-необязательный шарнир1-необязательный CH21-необязательный CH31-HD линкер-VL2-CL2-CLH линкер-VH2-необязательный CH12-необязательный шарнир2-необязательный CH22-необязательный CH32.
В другом аспекте изобретение относится к гетеромультимеру, представляющему собой многоцепочечное антитело, которое содержит две полипептидные цепи, где первая полипептидная цепь образует первую LC, первую HC и вторую HC, а вторая полипептидная цепь образует вторую LC. Первая полипептидная цепь содержит два домена VH, HD-линкер, первый домен VL, первый домен CL, CLH линкер, один или два шарнирных домена, и один или несколько константных доменов тяжелой цепи, выбранных из группы, включающей первый домен CH1, первый домена CH2, первый домен СН3, второй домен CH1, второй домен СН2 и второй домен СН3, где компоненты второй полипептидной цепи расположены относительно друг друга в направлении от N-конца к С-концу следующим образом: VL1-CL1-CLH линкер-VH1-необязательно CH11-необязательно шарнир1-необязательно CH21-необязательно CH31-HD линкер-VH2-необязательно CH12-необязательно шарнир2-необязательно CH22- необязательно CH32. Вторая полипептидная цепь содержит вторую VL и домен CL.
В другом аспекте настоящее изобретение относится к гетеромультимеру, содержащему две полипептидные цепи, где первый полипептид содержит иммуноадгезин, содержащий адгезин и один или несколько константных доменов тяжелой цепи (например, CH21 и/или CH31), второй полипептид образует половину антитела, которая включает домен VH и один или несколько константных доменов тяжелой цепи (например, CH1, CH22 и/или CH32), причем первая и вторая полипептидные цепи связаны друг с другом посредством HD линкера, образуя единую полипептидную цепь. Компоненты гетеромультимера расположены относительно друг друга в направлении от N-конца к С-концу следующим образом: адгезин-необязательно CH21-необязательно CH31-HD линкер-VH-необязательно CH1-необязательно CH22-необязательно CH32. HD линкер облегчает взаимодействие между одним или несколькими константными доменами иммуноадгезина и половиной антитела. В одном из вариантов осуществления CLH линкер облегчает взаимодействие между компонентами легкой цепи и тяжелой цепи половины антитела с получением гетеромультимера, содержащего компоненты, расположенные друг относительно друга в направлении от N-конца к С-концу следующим образом: адгезин-необязательно CH21-необязательно СН31-HD линкер-VL-CL-CLH линкер-VH-необязательно CH1-необязательно CH22-необязательно CH32. В другом варианте осуществления домены VL и CL легкой цепи половины антитела предоставляются вторым полипептидом, который связан с тяжелой цепью половины антитела первой полипептидной цепи, с образованием родственной пары легкая цепь-тяжелая цепь. В другом варианте осуществления иммуноадгезиновый фрагмент гетеромультимера может содержать аминокислотный спейсер, находящийся между компонентами адгезина и константным доменом тяжелой цепи. В одном из вариантов осуществления спейсер содержит остатки глицина (G) и серина (S), например, повторы GGS. В другом варианте осуществления длина спейсера находится в диапазоне 10-80 аминокислот, например, в диапазоне 20-40 остатков.
Гетеромультимер по настоящему изобретению может содержать HD линкер размером 15-100 аминокислот в длину. В конкретном варианте осуществления длина HD линкера составляет 30-39 аминокислот, например, 30, 31, 32, 33, 34, 35, 36, 37, 38 или 39 аминокислот. В одном из вариантов осуществления линкер содержит остатки глицина (G) и серина (S). В другом варианте осуществления линкер содержит повторы GGS. В предпочтительном варианте осуществления линкер содержит 8-9 повторов GGS (SEQ ID NO:19).
Гетеромультимер по настоящему изобретению также может содержать один или несколько линкеров CLH. В одном из вариантов осуществления длина одного или каждого из нескольких линкеров CLH находится в диапазоне 10-80 аминокислот. В конкретном варианте осуществления длина одного или каждого из нескольких линкеров CLH находится в диапазоне 20-40 аминокислот. В одном из вариантов осуществления линкер содержит остатки глицина (G) и серина (S). В другом варианте осуществления линкер содержит повторы GGS.
В другом варианте осуществления один или несколько из линкеров HD и CLH по настоящему изобретению могут отщепляться под действием одной или нескольких из следующих эндопептидаз: фурин, урокиназа, тромбин, тканевый активатор плазминогена (tРА), гененаза, Lys-C, Arg-С, Asp-N, Glu-C, фактор Ха, протеаза вируса травления табака (ТРВ), энтерокиназа, протеаза риновируса человека С3 (HRV С3) или кининогеназа. В предпочтительном варианте осуществления по меньшей мере один из линкеров отщепляется под действием фурина. В другом варианте осуществления по меньшей мере один из линкеров по настоящему изобретению расщепляется по двум участкам на N- и С-концах линкера или вблизи N- и С-концов линкера. В случае HD линкера, предпочтительно, один из двух участков расщепления представляет собой участок расщепления фурином, а другой участок расщепления представляет собой участок расщепления Lys-C. В случае CLH линкера, предпочтительно, и N- и С-концевые участки расщепления одного или нескольких линкеров CLH расщепляются под действием фурина. В некоторых вариантах осуществления участок расщепления фурином содержит аминокислотную последовательность RKRKRR (SEQ ID NO:9). В некоторых других вариантах осуществления участок расщепления фурином содержит аминокислотную последовательность RHRQPR (SEQ ID NO:10). В одном из вариантов осуществления расщепление под действием эндопептидазы происходит in situ. В конкретном варианте осуществления используют эндопептидазу, полученную путем рекомбинантной экспрессии в клетке-хозяине. В другом варианте осуществления расщепление под действием эндопептидазы проводят путем добавления эндопептидазы после очистки.
Гетеромультимер по настоящему изобретению может содержать один или несколько (например, два) линкеров CLH, каждый из которых содержит один или несколько участков расщепления под действием одной или нескольких из следующих специфических экзопептидаз: карбоксипептидаза А, карбоксипептидаза В, карбоксипептидаза В плазмы (также известная как карбоксипептидаза U или активируемый тромбином ингибитор фибринолиза (TAFI)), карбоксипептидаза D, карбоксипептидаза E (также известная как конвертаза энкефалина или карбоксипептидаза Н), карбоксипептидаза М, карбоксипептидаза N или карбоксипептидаза Z. В предпочтительном варианте осуществления гетеромультимер по настоящему изобретению расщепляется под действием экзопептидазы карбоксипептидазы В. В одном из вариантов осуществления расщепление под действием экзопептидазы происходит in situ. В конкретном варианте осуществления используют экзопептидазу, полученную путем рекомбинантной экспрессии в клетке-хозяине. В другом варианте осуществления расщепление под действием экзопептидазы происходит при добавлении экзопептидазы после очистки. В настоящем описании карбоксипептидаза B может относиться к классу карбоксипептидаз или она может представлять собой конкретную карбоксипептидазу. Как класс, карбоксипептидаза B включает все специфические карбоксипептидазы, кроме карбоксипептидазы А. В качестве специфической карбоксипептидазы карбоксипептидаза B также известна как карбоксипептидаза U или TAFI. Специалист в данной области может легко различить карбоксипептидазу В как класс и карбоксипептидазу В как специфическую экзопептидазу, в зависимости от контекста, в котором используется термин.
В следующем варианте осуществления гетеромультимер по настоящему изобретению может содержать один или несколько шарнирных доменов, включающих, без ограничения, шарнирные домены, которые содержат остатки от Glu216 до Pro230 IgG1 человека. В некоторых вариантах осуществления один или оба шарнирных доменов содержат мутацию, удаляющую участок расщепления эндопептидазой Lys-C. В одном из примеров мутация, удаляющая участок расщепления эндопептидазой Lys-C, представляет собой замену K222A (система нумерации EU).
Гетеромультимер по настоящему изобретению может быть моноспецифическим. В одном из вариантов осуществления моноспецифический гетеромультимер по настоящему изобретению содержит две половины антител, которые связываются с одним и тем же эпитопом-мишенью, но с разной аффинностью. В другом варианте осуществления моноспецифичный гетеромультимер по настоящему изобретению содержит половину антитела, связанную с иммуноадгезином, где все компоненты специфичны к одному и тому же партнеру по связыванию или эпитопу.
Гетеромультимер по настоящему изобретению может быть биспецифическим или мультиспецифическим. В одном из вариантов осуществления гетеромультимер способен связывать по меньшей мере два антигена. В другом варианте осуществления гетеромультимер способен связывать по меньшей мере два эпитопа на одном и том же антигене. В следующем варианте осуществления биспецифический или мультиспецифический гетеромультимер по настоящему изобретению содержит половину антитела, связанную с иммуноадгезином, каждый из которых является специфичным к разным партнерам по связыванию или эпитопам.
В другом варианте осуществления гетеромультимер по настоящему изобретению содержит константный участок, конъюгированный с цитотоксическим средством.
В другом варианте осуществления гетеромультимер может содержать два константных домена тяжелой цепи (например, два домена СН3), содержащих выступ или полость, где выступ или полость одного константного домена тяжелой цепи (например, домена CH31) могут быть расположены в полости или выступе, соответственно, второго константного домена тяжелой цепи (например, домена CH32). Предпочтительно, два константных домена совмещаются по граничной поверхности, содержащей выступ и полость. В следующем варианте осуществления гетеромультимер может содержать граничную поверхность по меньшей мере одного константного домена легкой цепи и одного константного домена тяжелой цепи (например, граничная поверхность CL/CH1), где константный домен легкой цепи (например, домен CL) и константный домен тяжелой цепи (например, домен CH1) взаимодействуют между собой по меньшей мере отчасти, посредством взаимодействия выступ-полость.
В другом варианте осуществления гетеромультимер по настоящему изобретению содержит мутацию домена CH2, либо во фрагменте CH21, либо во фрагменте CH22, которая приводит к изменению эффекторных функций антитела. В предпочтительном варианте осуществления мутация домена CH2 представляет собой мутацию N297. В некоторых вариантах осуществления мутация N297 представляет собой мутацию N297A. В некоторых других вариантах осуществления домен СН2 необязательно содержит мутацию D256A.
В другом аспекте настоящее изобретение предлагает способы получения гетеромультимера. В другом аспекте настоящее изобретение относится к полинуклеотидам, кодирующим гетеромультимеры по настоящему изобретению. В дополнительных аспектах изобретение предлагает векторы, содержащие полинуклеотиды по настоящему изобретению, а также клетки-хозяева, содержащие указанные векторы. В одном из вариантов осуществления клетка-хозяин представляет собой клетку млекопитающего. В предпочтительном варианте осуществления клетка млекопитающего представляет собой клетку CHO. В другом варианте осуществления клетка-хозяин является прокариотической клеткой. В следующем варианте осуществления прокариотическая клетка представляет собой клетку E. coli. В другом аспекте настоящее изобретение предлагает способ получения гетеромультимера, который включает культивирование клетки-хозяина, которая содержит вектор, содержащий полинуклеотиды, кодирующие гетеромультимер, в культуральной среде. Предпочтительно, гетеромультимер извлекают из клетки-хозяина или из среды, в которой культивируют клетку-хозяина.
В другом аспекте настоящее изобретение относится к одноцепочечным антителам, содержащим один полипептид, содержащий нижеследующие домены, расположенные относительно друг друга в направлении от N-конца к C-концу в следующем порядке: VL1-CL1-CLH линкер1-VH1-CH11-шарнир1-CH21-CH31-HD линкер-VL2-CL2-CLH линкер2-VH2-CH12-шарнир2-CH22-CH32, где каждый из CLH линкера1, CLH линкера2 и HD линкера содержит аминокислотную последовательность, расщепляемую фуриновой эндопептидазой. В некоторых вариантах осуществления данного аспекта расщепляемая фурином последовательность содержит аминокислотную последовательность RKRKRR (SEQ ID NO:9), тогда как в других вариантах осуществления расщепляемая фурином последовательность содержит аминокислотную последовательность RHRQPR (SEQ ID NO:10). В родственных аспектах изобретение предлагает полинуклеотидные молекулы, кодирующие одноцепочечное антитело по настоящему изобретению, векторы, содержащие полинуклеотиды, и клетки-хозяева, содержащие векторы. В некоторых вариантах осуществления клетка-хозяин представляет собой клетку млекопитающего, включающую, без ограничения, клетку СНО. В некоторых других вариантах осуществления клетка-хозяин представляет собой прокариотическую клетку, включающую, без ограничения, клетку E.coli. В другом родственном аспекте настоящее изобретение предлагает способы получения одноцепочечного антитела, включающие культивирование клетки-хозяина, содержащей вектор, в культуральной среде. В некоторых вариантах осуществления способ дополнительно включает стадию извлечения указанного одноцепочечного антитела из указанной клетки-хозяина или указанной культуральной среды.
Другие признаки и преимущества настоящего изобретения будут очевидны из нижеследующего подробного описания, чертежей и формулы изобретения.
Краткое описание чертежей
На фиг. 1 представлена схематическая диаграмма, демонстрирующая структуру типичного гетеромультимерного одноцепочечного антитела, содержащего три расщепляемых линкера. Участки расщепления эндопептидазой обозначены треугольниками. Также изображена необязательная мутация K222A.
На фиг. 2 представлена схематическая диаграмма, демонстрирующая расположение в типичном гетеромультимерном одноцепочечном антителе участков LC, HC, линкеров и участков расщепления. Примерами участков расщепления являются RKRKRRG(GGS)6GRSRKRR (SEQ ID NO:14) и (GGS)(8-10)RSRKRR(SEQ ID NO:15-17). Примером участка расщепления фурином является RXRXRR (SEQ ID NO:8).
На фиг. 3А приведен пример гетеромультимерного одноцепочечного антитела после расщепления фурином. Остатки в скобках (RKRKRR (SEQ ID NO:9) и RKRKR (SEQ ID NO:18)) обозначают остатки, которые могут быть удалены под действием эндогенных экзопептидаз перед очисткой на колонке с белком А с получением неровных C-концов. Также изображена необязательная мутация K222A.
На фиг. 3В приведен пример гетеромультимерного одноцепочечного антитела после обработки фурином, Lys-C и экзопептидазой (например, карбоксипептидазой В). Также изображена необязательная мутация K222A.
На фиг. 4 приведен пример конъюгированного гетеромультимерного одноцепочечного антитела до расщепления с целью удаления линкеров. Также изображен необязательно конъюгированный фрагмент, например, такой как токсин, антибиотик и т.д., и необязательная мутация K222A.
На фиг. 5А представлена схематическая диаграмма, демонстрирующая структуру типичного гетеромультимерного мультицепочечного антитела, содержащего один расщепляемый линкер. Два несвязанных линкером LC могут экспрессироваться независимо от полипептида, содержащего линкер. Несвязанные линкером LC могут экспрессироваться в одной клетке, или в разных клетках, как и связанные тяжелые цепи. Несвязанные линкером LC могут экспрессироваться из одной плазмиды, или из разных плазмид. Также изображен необязательно конъюгированный фрагмент, такой как токсин, антибиотик и т.д., и необязательная мутация K222A.
На фиг. 5В представлена схематическая диаграмма, демонстрирующая структуру типичного гетеромультимерного мультицепочечного антитела, содержащего два расщепляемых линкера. Линкер HD связывает первый HC и второй HC косвенно через связанный линкером LC. Несвязанный линкером LC может экспрессироваться независимо от полипептида, содержащего линкер. Несвязанные линкером LC могут экспрессироваться в одной клетке, или в разных клетках, как и связанные тяжелые цепи. Несвязанные линкером LC могут экспрессироваться из одной плазмиды, или из разных плазмид. Также изображена необязательная мутация K222A.
На фиг. 5C представлена схематическая диаграмма, демонстрирующая структуру типичного гетеромультимерного мультицепочечного антитела, содержащего два расщепляемых линкера. Линкер HD непосредственно связывает первый HC и второй HC. Несвязанный линкером LC может экспрессироваться независимо от полипептида, содержащего линкер. Несвязанные линкером LC могут экспрессироваться в одной клетке, или в разных клетках, как и связанные тяжелые цепи. Несвязанные линкером LC могут экспрессироваться из одной плазмиды, или из разных плазмид. Также изображена необязательная мутация K222A.
На фиг. 6А-6D приведены графики, демонстрирующие результаты анализа октета. (A) Показаны все графики, изображенные на чертежах B-D. (B) Типичное гетеромультимерное одноцепочечное антитело связывается с антигеном 1 и с антигеном 2 одновременно. (C и D) Антитело 1 и антитело 2 не вступают в перекрестное взаимодействие с антигенами друг друга, но связываются со своим соответствующим антигеном. На оси Х откладывают время в секундах. На оси Y откладывают относительное поглощение. Подробное описание можно найти в примере 3.
Подробное описание изобретения
Нежелательная гомодимеризация тяжелых цепей обычно происходит при образовании моноспецифических или мультиспецифических (например, биспецифических) антител или других гетеромультимеров, обладающих разными связующими свойствами и содержащих несколько полипептидных цепей. Авторы настоящего изобретения обнаружили, что данную общую проблему можно преодолеть путем образования одноцепочечных моноспецифических или мультиспецифических гетеромультимеров, сборка которых управляется одним линкером или несколькими линкерами. Без связи с какой-либо теорией, авторы полагают, что HD линкер обеспечивает связывание разных компонентов Fc тяжелой цепи с высокой степенью точности и эффективности, что позволяет получить функциональный гетеромультимер, содержащий две половины молекулы (например, две половины антитела), которые связываются с одной и той же мишенью, или с разными мишенями с одинаковой или разной аффинностью. Гетеромультимер, содержащий связанные компоненты тяжелой цепи, может дополнительно содержать разные компоненты легкой цепи с образованием функционального одноцепочечного моноспецифического или мультиспецифического гетеромультимера (например, антитела) с полным комплектом тяжелых и легких цепей. Дополнительные линкеры по настоящему изобретению можно использовать для связи легких и тяжелых цепей гетеромультимера, обеспечивая правильное соединение отдельной легкой цепи с родственной тяжелой цепью.
Применение описанных в настоящем описании способов получения гетеромультимеров позволяет получать практически гомогенные популяции моноспецифических или мультиспецифических гетеромультимеров, полученных с использованием одной или нескольких полипептидных последовательностей. Гетеромультимеры, полученные с помощью описанных в настоящем описании способов, можно использовать для распознавания нескольких мишеней в патогенном пути или для совместной локализации конкретной мишени (например, опухолевой клетки) и средства, направленного против мишени (например, Т-клетки). Кроме того, описанные в настоящем описании гетеромультимеры имеют преимущество, заключающееся в том, что они позволяют избежать необходимости применения комбинированной терапии, направленной на два антигена, и риска, связанного с введением индивиду двух или более лекарственных средств.
I. Определения
Термин "антитело" в настоящем документе используется в самом широком смысле и относится к любой молекуле иммуноглобулина (Ig), содержащей две тяжелые цепи и две легкие цепи, а также к любым фрагментам, мутантам или вариантам такой молекулы, при условии, что они обладают желательной биологической активностью (например, эпитопсвязывающей активностью). Примеры антител включают моноклональные антитела, поликлональные антитела, биспецифические антитела, мультиспецифические антитела и фрагменты антител.
Систему нумерации Kabat, как правило, используют в применении к остаткам вариабельного домена (примерно остатки 1-107 легкой цепи и остатки 1-113 тяжелой цепи) (например, Kabat et al., Sequences of Immunological Interest. 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)). "Систему нумерации EU" или "индекс EU" обычно используют в применении к остаткам константного домена тяжелой цепи иммуноглобулина (например, индекс EU описан в Kabat et al., выше). "Индекс EU по Kabat" относится к EU нумерации остатков антитела человека IgG1. Если не указано иначе, номера остатков вариабельных доменов антител приводятся в соответствии с системой нумерации Kabat. Если не указано иначе, номера остатков константных доменов тяжелых цепей антител приводятся в соответствии с системой нумерации EU.
Основное встречающееся в природе 4-цепочечное антитело представляет собой гетеротетрамерный гликопротеин, состоящий из двух идентичных легких цепей (LC) и двух идентичных тяжелых цепей (HC) (антитело IgM состоит из 5 основных гетеротетрамерных элементов и дополнительного полипептида, называемого J-цепь, и, следовательно, содержит 10 антигенсвязывающих участков, тогда как секретируемые антитела IgA могут полимеризоваться с образованием поливалентных комплексов, содержащих 2-5 основных 4-цепочечных элементов наряду с J цепью). В случае IgG размер 4-цепочечного элемента составляет, как правило, примерно 150000 Дальтон. Каждая LC связана с HC одной ковалентной дисульфидной связью, а две HC связаны друг с другом посредством одной или нескольких дисульфидных связей в зависимости от изотипа HC. Все НС и LC также содержат расположенные через одинаковые промежутки внутрицепочечные дисульфидные мостики. Каждая НС содержит на N-конце вариабельный домен (VH), за которым следуют три константных домена (CH1, CH2, CH3) в случае α и γ цепей и четыре Cj домена в случае изотипов μ и ε. Каждая LC содержит на N-конце вариабельный домен (VL), за которым следует константный домен (CL), расположенный на другом конце. VL можно совместить с VH, а CL можно совместить с первым константным доменом тяжелой цепи (CH1). CH1 может быть соединен со вторым константным доменом тяжелой цепи (CH2) посредством шарнирного участка. Считается, что определенные аминокислотные остатки образуют граничную поверхность между вариабельными доменами легкой цепи и тяжелой цепи. В результате конъюгации VH и VL образуется единый антигенсвязывающий участок. Описание структуры и свойств разных классов антител можно найти, например, в Basic and Clinical Immunology, 8 th edition, Daniel P. Stites, Abba I. Terr and Tristram G. Parslow (eds.), Appleton & Lange, Norwalk, CT, 1994, page 71 and Chapter 6.
"Шарнирным участком" обычно называют участок от Glu216 до Pro230 IgG1 человека (Burton, Molec. Immunol. 22: 161-206 (1985)). Шарнирные участки других изотипов IgG можно выравнивать с последовательностью IgG1, помещая первый и последний остатки цистеина, образующие S-S связи между тяжелыми цепями, в одинаковых положениях.
"Нижний шарнирный участок" Fc-участка обычно определяют как последовательность остатков, расположенную непосредственно на C-конце шарнирного участка, т.е. остатки с 233 по 239 участка Fc. До настоящего изобретения считали, что связывание FcgammaR, как правило, обуславливается аминокислотными остатками в нижнем шарнирном участке Fc-участка IgG.
"Домен CH2" Fc-участка IgG человека, как правило, простирается примерно от остатка 231 до остатка 340 IgG. Домен CH2 уникален тем, что он не способен точно спариваться с другим доменом. Более того, между двумя доменами CH2 интактной нативной молекулы IgG расположены две N-связанные разветвленные углеводные цепи. Существует предположение, что углевод служит заменой спаривания доменов и помогает стабилизировать домен CH2. Burton, Molec. Immunol. 22: 161-206 (1985).
"Домен СН3" включает последовательность остатков, расположенных на С-конце домена CH2 Fc-участка (т.е. он простирается примерно от аминокислотного остатка 341 до аминокислотного остатка 447 IgG).
Легкую цепь (LC), полученную из позвоночного любого вида, можно отнести к одному из двух четко различающихся типов, называемых каппа и лямбда, в зависимости от аминокислотных последовательностей их константных доменов. На основании аминокислотных последовательностей константных доменов тяжелых цепей (СН) иммуноглобулины можно отнести к разным классам или изотипам. Существует пять классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, тяжелые цепи которых обозначают α, δ, γ, ε и μ, соответственно. Классы γ и α дополнительно подразделяют на подклассы на основе сравнительно незначительных различий в последовательности CH и функции, например, выделяют следующие подклассы: IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2.
Термин "вариабельный" относится к тому факту, что последовательности некоторых сегментов вариабельных доменов разных антител значительно различаются. Домен V опосредует связывание антигена и определяет специфичность конкретного антитела к конкретному антигену. Однако вариабельность неравномерно распределена на протяжении участков вариабельных доменов длиной 110 аминокислот. Наоборот, V-домены содержат относительно постоянные сегменты размером 15-30 аминокислот, называемые каркасные участки (FR), которые разделены короткими предельно изменчивыми участками, называемыми "гипервариабельные участки", каждый из которых содержит 9-12 аминокислот в длину. Каждый из вариабельных доменов нативных тяжелых и легких цепей содержит четыре FR, как правило, имеющих бета-складчатую конфигурацию, соединенных тремя гипервариабельными участками, которые образуют петли, соединяющие бета-складчатые структуры, а в некоторых случаях являются частью данной бета-складчатой структуры. Гипервариабельные области каждой цепи удерживаются вместе в непосредственной близости посредством FR, и, наряду с гипервариабельными участками другой цепи, вносят вклад в образование антигенсвязывающего центра антител (см. Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991). Константные домены непосредственно не участвуют в связывании антитела с антигеном, но выполняют разные эффекторные функции, например, отвечают за участие антитела в антителозависимой клеточной цитотоксичности (ADCC).
Термин "гипервариабельный участок" или "HVR" в настоящем описании относится ко всем участкам вариабельного домена антитела, которые характеризуются гипервариабельностью последовательностей и/или образуют структурно определенные петли ("гипервариабельные петли"). Как правило, нативные четырехцепочечные антитела содержат шесть HVR; три в VH (H1, Н2, Н3), и три в VL (L1, L2, L3). HVR, как правило, содержат аминокислотные остатки гипервариабельных петель и/или "участков, определяющих комплементарность" (CDR), причем последние обладают наивысшей вариабельностью последовательностей и/или участвуют в распознавании антигена. HVR в соответствии с настоящим описанием содержит любое число остатков, расположенных в положениях 24-36 (L1), 46-56 (L2), 89-97 (L3), 26-35B (H1), 47-65 (H2) и 93-102 (H3). Таким образом, HVR содержит остатки описанных ранее в положениях в (А), (В) и (С): (А) 24-34 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987)); (В) 24-34 из L1, 50-56 из L2, 89-97 из L3, 31-35B из H1, 50-65 из Н2, и 95-102 из Н3 (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)); (С) 30-36 (L1), 46-55 (L2), 89-96 (L3), 30-35 (H1), 47-58 (H2), 93-100a-j (H3) (MacCallum et al. J. Mol. Biol. 262:732-745 (1996)). Если не указано иначе, остатки HVR и другие остатки вариабельного домена (например, остатки FR) в настоящем описании нумеруют в соответствии с Kabat et al., выше.
Используется ряд контуров HVR, которые входят в объем настоящего описания. Чаще всего используют участки, определяющие комплементарность (CDR), в соответствии с Kabat, установленные по вариабельности последовательности (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Servce, National Institutes of Health, Bethesda, MD, 1991). В соответствии с Chothia данные участки определяют по местоположению структурных петель (Chothia и Lesk J. Mol. Biol. 196: 901-917 (1987)). HVR AbM представляют собой компромисс между HVR Kabat и структурными петлями Chothia, и используются программным обеспечением для моделирования антител Oxford Molecularʹs AbM. "Контактные" HVR определяют на основе анализа доступных сложных кристаллических структур. Остатки каждого из указанных HVR приведены ниже.
HVR могут содержать следующие "расширенные HVR": 24-36 или 24-34 (L1), 46-56 или 50-56 (L2) и 89-97 или 89-96 (L3) в VL и 26-35 (H1), 50-65 или 49-65 (Н2) и 93-102, 94-102 или 95-102 (Н3) в VH. Для каждого из указанных определений остатки вариабельного домена пронумерованы в соответствии с Kabat et al., см. выше.
"Каркасные участки" (FR) содержат остатки вариабельного домена, отличающиеся от остатков CDR. Каждый вариабельный домен обычно содержит четыре FR, обозначаемых FR1, FR2, FR3 и FR4. Если CDR определяют в соответствии с Kabat, остатки FR легкой цепи располагаются в положениях 1-23 (LCFR1), 35-49 (LCFR2), 57-88 (LCFR3) и 98-107 (LCFR4), а остатки FR тяжелой цепи располагаются в положениях 1-30 (HCFR1), 36-49 (HCFR2), 66-94 (HCFR3) и 103-113 (HCFR4). Если CDR содержат аминокислотные остатки гипервариабельной петли, то остатки FR легкой цепи располагаются в положениях 1-25 (LCFR1), 33-49 (LCFR2), 53-90 (LCFR3) и 97-107 (LCFR4), а остатки FR тяжелой цепи располагаются в положениях 1-25 (HCFR1), 33-52 (HCFR2), 56-95 (HCFR3) и 102-113 (HCFR4). В некоторых случаях, если CDR содержит аминокислоты, соответствующие определению Kabat, и аминокислоты гипервариабельной петли, остатки FR корректируют соответствующим образом. Например, если CDRH1 содержит аминокислоты H26-H35, остатки FR1 тяжелой цепи находятся в положениях 1-25, а остатки FR2 находятся в положениях 36-49.
"Консенсусная каркасная последовательность человека" представляет собой каркасный участок, который содержит аминокислотные остатки, наиболее часто встречающиеся в выборке каркасных участков VL или VH иммуноглобулинов человека. Как правило, выборку последовательностей VL или VH иммуноглобулинов человека получают из подгруппы последовательностей вариабельного домена. Как правило, подгруппа последовательностей представляет собой подгруппу, определенную по системе Kabat. В одном из вариантов осуществления в случае VL подгруппа представляет собой каппа I в соответствии с определением Kabat. В одном из вариантов осуществления в случае VH подгруппа представляет собой подгруппу III в соответствии с определением Kabat.
Одним из примеров "интактного" антитела является антитело, которое содержит антигенсвязывающий участок, а также CL и константные домены тяжелой цепи, CH1, CH2 и CH3. Константные домены могут содержать нативные последовательности (например, они могут представлять собой константные домены с нативными последовательностями человека), или варианты указанных аминокислотных последовательностей.
"Фрагмент антитела" представляет собой часть интактного антитела, предпочтительно, антигенсвязывающий или вариабельный участок интактного антитела. Примеры фрагментов антител включают фрагменты Fab, Fabʹ, F(аbʹ)2 и Fv; диатела (Db); тандемные диатела (taDb), линейные антитела (см., например, патент США № 5641870, Пример 2; Zapata et al., Protein Eng. 8 (10): 1057-1062 (1995)); одноплечие антитела, антитела с одним вариабельным доменом, минитела, молекулы одноцепочечных антител и мультиспецифические антитела, полученные из фрагментов антител (например, включающие, без ограничения, Db-Fc, taDb-Fc, taDb-СН3 и (ScFv)4-Fc).
Фрагмент "Fab" представляет собой антигенсвязывающий фрагмент, полученный в результате расщепления антитела папаином, который содержит целую L-цепь вместе с вариабельным доменом Н-цепи (VH) и первый константный домен тяжелой цепи (CH1). Расщепление антител папаином дает два идентичных фрагмента Fab. Обработка антитела пепсином дает один большой фрагмент F(аbʹ)2, который примерно соответствует двум фрагментам Fab, связанным дисульфидными связями, характеризуется двухвалентной антигенсвязывающей активностью и сохранением способности к перекрестному связыванию антигена. Фрагменты Fabʹ отличаются от фрагментов Fab несколькими дополнительными остатками на карбоксильном конце домена CH1, включающими один или несколько остатков цистеина из шарнирного участка антитела. Fabʹ-SH в настоящем описании используют как обозначение Fabʹ, в котором цистеиновый остаток (цистеиновые остатки) константного домена несет свободную тиольную группу. Фрагменты антитела F(аbʹ)2 исходно были получены как пары фрагментов Fabʹ, соединенные шарнирными остатками цистеина. Также известны другие способы химического соединения фрагментов антител.
Фрагмент "Fc" представляет собой остаточный фрагмент антитела, полученный в результате расщепления папаином, обозначение которого отражает способность фрагмента легко кристаллизоваться. Фрагмент Fc содержит карбокси-концевые участки обоих Н-цепей, удерживаемые вместе дисульфидными связями. Эффекторные функции антител определяются последовательностями фрагмента Fc; данный фрагмент также представляет собой участок, распознаваемый рецепторами Fc (FcR), присутствующими на определенных типах клеток.
В настоящем описании термин "Fc-участок" используется для определения С-концевого участка тяжелой цепи иммуноглобулина и включает нативные последовательности Fc-участка и вариантные последовательности Fc-участка. Хотя границы Fc-участка тяжелой цепи иммуноглобулина могут варьировать, Fc-участок тяжелой цепи IgG человека обычно определяют как простирающийся от аминокислотного остатка Cys226, или от Pro230, до карбокси-конца. С-концевой лизин (остаток 447 в соответствии с системой нумерации EU) Fc-участка можно удалить, например, в процессе получения или очистки антитела, или путем применения рекомбинантной нуклеиновой кислоты, кодирующей тяжелую цепь антитела. Соответственно, композиция интактных антител может содержать популяции антител, в которых удалены все остатки K447, популяции антител, в которых не удален ни один остаток K447, а также популяции, включающие смесь антител, содержащих и не содержащих остаток K447.
"Функциональный Fc-участок" способен выполнять "эффекторную функцию" нативной последовательности Fc-участка. Примеры "эффекторных функций" включают связывание C1q; CDC; cвязывание рецептора Fc; ADCC; фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора; BCR) и т.д. Такие эффекторные функции, для выполнения которых обычно требуется объединение Fc-участка со связывающим доменом (например, вариабельным доменом антитела), можно тестировать с помощью разных анализов, как описано, например, в приведенных в настоящем описании определениях.
"Нативная последовательность Fc-участка" включает аминокислотную последовательность, идентичную аминокислотной последовательности Fc-участка, встречающегося в природе. Нативная последовательность Fc-участка человека включает нативную последовательность Fc-участка IgG1 человека (аллотипы не А и А); нативную последовательность Fc-участка IgG2 человека; нативную последовательность Fc-участка IgG3 человека; и нативную последовательность Fc-участка IgG4 человека, а также их варианты, встречающиеся в природе.
"Вариант Fc-участка" включает аминокислотную последовательность, которая отличается от нативной последовательности Fc-участка в результате модификации по меньшей мере одной аминокислоты, предпочтительно, в результате одной или нескольких аминокислотных замен. Предпочтительно, вариант Fc-участка содержит по меньшей мере одну аминокислотную замену по сравнению с нативной последовательностью Fc-участка или Fc-участка исходного полипептида, например, примерно от одной до десяти аминокислотных замен и, предпочтительно, приблизительно от одной до пяти аминокислотных замен в нативной последовательности Fc-участка или в Fc-участке исходного полипептида. В соответствии с настоящим описанием вариант Fc-участка, предпочтительно, по меньшей мере примерно на 80% гомологичен нативной последовательности Fc-участка и/или Fc-участку исходного полипептида, и, наиболее предпочтительно, по меньшей мере примерно на 90% гомологичен им, более предпочтительно, по меньшей мере примерно на 95% гомологичен им.
В настоящем описании термин "Fc-комплекс" относится к двум доменам CH2 Fc-участка, взаимодействующим вместе, и/или к двум доменам CH3 Fc-участка, взаимодействующим вместе, где домены СН2 и/или домены СН3 соединяются посредством связей и/или взаимодействий (например, вандерваальсовых, гидрофобных, гидрофильных взаимодействий), отличных от пептидных связей.
В настоящем описании термин "компонент Fc" относится к шарнирному участку, домену СН2 или домену СН3 Fc-участка.
В настоящем описании термин "CH-компонент Fc" или "FcCH" относится к полипептиду, содержащему домен СН2, домен СН3, или домены СН2 и СН3 Fc-участка.
"Fv" представляет собой димер, содержащий один вариабельный домен тяжелой цепи и один вариабельный домен легкой цепи, прочно связанные нековалентной связью. После укладки двух указанных доменов образуются шесть гипервариабельных петель (3 петли в каждой из Н- и L-цепи), которые предоставляют аминокислотные остатки, участвующие в связывании антигена, и отвечают за специфичность связывания антигена с антителом. Однако даже один вариабельный домен (или половина Fv, содержащая только три CDR, специфичные к антигену) обладает способностью распознавать и связывать антиген, хотя часто с более низкой аффинностью, чем целый участок связывания.
В настоящем описании термин "иммуноадгезин" обозначает молекулы, которые сочетают в себе специфичность связывания гетерологичного белка ("адгезин") с эффекторными функциями константных доменов иммуноглобулина. Структура иммуноадгезинов включает гибрид аминокислотной последовательности, обладающей желательной специфичностью связывания, причем указанная аминокислотная последовательность отличается от участка антитела, отвечающего за распознавание и связывание антигена (т.е. является "гетерологичный" по отношению к константному участку антитела), и последовательности константного домена иммуноглобулина (например, последовательности СН2 и/или СН3 IgG). Константные домены адгезина и иммуноглобулина могут необязательно разделяться аминокислотным спейсером. Примеры последовательностей адгезина включают непрерывные аминокислотные последовательности, которые содержат фрагмент рецептора или лиганда, способный связываться с представляющим интерес белком. Последовательности адгезина также могут представлять собой последовательности, способные связывать представляющий интерес белок, но отличающиеся от последовательностей рецептора или лиганда (например, последовательности адгезина, присутствующие в пептидоантителах). Такие полипептидные последовательности можно выбрать или идентифицировать с помощью разных методов, включающих методы фаговых дисплеев и методы сортинга с высокой пропускной способностью. Последовательность константного домена иммуноглобулина, входящую в состав иммуноадгезина, можно получить из любого иммуноглобулина, такого как IgG1, IgG2, IgG3, или подтипы IgG4, IgA (в том числе Iga1 и IgA2), IgE, IgD или IgM.
В настоящем описании термин аминокислотный "спейсер" обозначает аминокислотную последовательность из двух или более аминокислот в длину, которая не расщепляется, например, в результате ауторасщепления, ферментативного или химического расщепления. Спейсер может содержать нейтральные, полярные или неполярные аминокислоты. Аминокислотный спейсер может содержать, например, от 2 до 100 аминокислот в длину, например, 10-80 или 20-40 аминокислот в длину, например, 3, 5, 10, 15, 20, 25, 30, 35 или 40 аминокислот в длину. В некоторых вариантах осуществления аминокислотный спейсер может содержать остатки глицина (G) и серина (S), например, в составе повторов GGS. В некоторых вариантах осуществления аминокислотный спейсер может содержать остатки треонина (Т) и гистидина (H). Примерами спейсеров являются THT (SEQ ID NO:1), GGGSTHT (SEQ ID NO:2) и GGGSGGGSTHT (SEQ ID NO:3).
"Одноцепочечные Fv", также сокращенно называемые "sFV" или "scFv", представляют собой фрагменты антител, которые содержат домены антител VH и VL, соединенные в одной полипептидной цепи. Предпочтительно, полипептид sFV необязательно содержит полипептидный линкер между доменами VH и VL, который обеспечивает формирование структуры sFV, необходимой для связывания антигена. Обзор по sFV можно найти в Pluckthun, The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994); Malmborg et al., J. Immunol. Methods 183:7-13, 1995.
Термин "диатела" относится к небольшим фрагментам антител, полученным путем конструирования фрагментов sFV (см. предыдущий абзац), содержащих короткие линкеры (примерно 5-10 остатков) между доменами VH и VL, которые способствуют спариванию V-доменов между цепями, но не внутри одной цепи, с образованием двухвалентного фрагмента, то есть фрагмента, содержащего два антигенсвязывающих участка. Биспецифические диатела представляют собой гетеродимеры, содержащие два "пересекающихся" sFv фрагмента, в которых домены VH и VL двух антител, присутствующих на разных полипептидных цепях. Диатела более подробно описаны, например, в ЕР 404097; WO 93/11161; и Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993).
Выражение "однодоменные антитела" (sdAb) или "антитела, содержащие один вариабельный домен (SVD)," как правило, относится к антителам, в которых один вариабельный домен (VH или VL) отвечает за связывание с антигеном. Другими словами, одному вариабельному домену не нужно взаимодействовать с другим вариабельным доменом, чтобы распознать целевой антиген. Примеры однодоменных антител включают антитела, полученные из верблюдовых (лам и верблюдов) и хрящевых рыб (например, акул), а также антитела, полученные рекомбинантными методами из антител человека и мыши (Nature (1989) 341: 544-546; Dev Comp Immunol (2006) 30: 43-56; Trend Biochem Sci (2001) 26: 230-235; Trends Biotechnol (2003) 21:484-490; WO 2005/035572; WO 03/035694; Febs Lett (1994) 339: 285-290; WO 00/29004; WO 02/051870).
В настоящем описании термин "половина антитела" относится к одному плечу антитела и включает по меньшей мере домен VH и один домен CH. В некоторых вариантах осуществления половину антитела можно присоединить к иммуноадгезину с образованием гетеромультимера по настоящему изобретению. В других вариантах осуществления первую половину антитела можно присоединить ко второй половине антитела с идентичной или неидентичной аминокислотной последовательностью (например, отличающейся по меньшей мере на один аминокислотный остаток) с образованием симметричного или асимметричного гетеромультимера, соответственно.
Термин "одноцепочечное антитело", как используется в настоящем описании, в самом широком смысле и конкретно включает моноспецифическое или мультиспецифическое (например, биспецифическое) антитело, которое изначально получают в виде одной непрерывной полипептидной цепи. Такие одноцепочечные антитела включают, без ограничения, антитело, содержащее два связанных HC, которые могут быть разными или одинаковыми и могут содержать два разных или одинаковых домена VH, линкер HD, соединяющий два HC, и по меньшей мере один константный домен тяжелой цепи, выбранный из двух разных или идентичных доменов СН2 и двух разных или идентичных доменов СН3. Одноцепочечные антитела могут дополнительно содержать один или два домена СН1, которые могут быть разными или одинаковыми. В некоторых вариантах осуществления одноцепочечные антитела содержат один или два шарнирных участка, которые связывают один домен HC (например, VH или CH1) со вторым доменом, непосредственно примыкающим к первому (например, CH2). В других вариантах осуществления одноцепочечные антитела могут содержать одну или две связанных LC, которые могут быть разными или одинаковыми, причем каждая из них может содержать два разных или одинаковых домена VL и CL, каждый из которых связан с конкретным линкером HC или CLH. Как описано в настоящем документе, одноцепочечные антитела, для получения которых необязательно используют технологию совмещений выступов и впадин, обеспечивающую гетеродимеризацию HC/HC или HC/LC, могут содержать отщепляемые линкеры. Одноцепочечные антитела представляют собой гетеромультимеры по настоящему изобретению.
В настоящем описании термин "многоцепочечное антитело" относится к антителу, состоящему из двух LC и двух HC, где две HC экспрессируются в составе одного полипептида, а по меньшей мере одна LC экспрессируется в виде отдельного полипептида. Независимо экспрессированные LC связываются с родственными HC, образуя антитело, содержащее два функциональных плеча. Многоцепочечные антитела могут быть моноспецифическими или мультиспецифическими. Многоцепочечные антитела, для получения которых необязательно используют технологию совмещений выступов и впадин, обеспечивающую гетеродимеризацию HC/HC или HC/LC, могут содержать отщепляемые линкеры.
Термин "совмещение выступов и впадин" или методика "KnH" в настоящем описании относится к метододике проведения спаривания двух полипептидов in vitro или in vivo путем помещения выступа (выпуклости), присутствующего в одном полипептиде, и полости (впадины), присутствующей в другом полипептиде, на граничную поверхность, где происходит их взаимодействие. Например, KnH помещают на поверхности связывания Fc:Fc, граничной поверхности CL:CH1 или граничной поверхности VH/VL антитела (например, US2007/0178552, WO 96/027011, WO 98/050431 и Zhu et al. (1997) Protein Science 6: 781-788). Данный способ, предпочтительно, используют для спаривания двух разных тяжелых цепей при получении мультиспецифических антител. Например, мультиспецифичные антитела, содержащие KnH в Fc-участках, может дополнительно содержать отдельные вариабельные домены, связанные с каждым Fc-участком, или они могут необязательно содержать разные вариабельные домены тяжелой цепи, способные спариваться с подобными или разными вариабельными доменами легкой цепи. Метод KnH также можно использовать для спаривания двух разных внеклеточных доменов рецепторов, или любых других полипептидных последовательностей, которые содержат разные последовательности, способные распознавать мишени.
Термин "мультиспецифическое антитело" используется в самом широком смысле и конкретно охватывает антитело, которое обладает полиэпитопной специфичностью. Такие мультиспецифичные антитела включают, без ограничения, антитела, содержащие вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), где блок VH/VL обладает полиэпитопной специфичностью, антитела, содержащие два или более доменов VL и VH, где разные блоки VH/VL связываются с разными эпитопами, антитела, содержащие два или более отдельных вариабельных доменов, где разные вариабельные домены связываются с разными эпитопами, полноразмерные антитела, фрагменты антител, такие как Fab, Fv, dsFv, ScFv, диатела, биспецифические диатела и триатела, фрагменты антител, связанные ковалентно или нековалентно. Термин "полиэпитопная специфичность" относится к способности специфически связывать два или более разных эпитопов на одной и той же мишени, или на разных мишенях. Термин "моноспецифический" относится к способности связывать только один антиген. В одном из вариантов осуществления моноспецифический гетеромультимер связывает два разных эпитопа на одной мишени/одном антигене. Согласно одному варианту осуществления мультиспецифическое антитело представляет собой антитело IgG, которое связывается с каждым эпитопом с аффинностью, находящейся в диапазоне от 5 мкМ до 0,001 пМ, от 3 мкМ до 0,001 пМ, от 1 мкМ до 0,001 пМ, от 0,5 мкМ до 0,001 пМ, или от 0,1 мкМ до 0,001 пМ.
Антитела по настоящему изобретению могут включать "химерные" антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям антител, полученных из конкретных видов, или принадлежащих к конкретному классу или подклассу антител, тогда как остальная часть цепи (цепей) идентична или гомологична соответствующим последовательностям антител, полученных из другого вида, или принадлежащих к другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют желательную биологическую активность (патент США № 4816567 и Morrison et al., Proc. Natl. Acad. Sci. USA 81: 6851-6855 (1984)). Химерные антитела, представляющие интерес с точки зрения настоящего описания, включают приматизированные антитела, содержащие антигенсвязывающие последовательности вариабельного домена, полученного от отличного от человека примата (такого как старосветская мартышка, человекообразная обезьяна и т.д.), и последовательности человеческого константного участка.
"Гуманизированные" формы нечеловеческих антител (например, антител грызунов) представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из нечеловеческого антитела. По большей части гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки гипервариабельного участка реципиента заменены остатками гипервариабельного участка антитела (донорное антитело) отличного от человека вида, такого как мышь, крыса, кролик или отличный от человека примат, обладающего желательной специфичностью, аффинностью и емкостью. В некоторых случаях остатки каркасного участка (FR) иммуноглобулина человека заменяют соответствующими нечеловеческими остатками. Кроме того, гуманизированные антитела могут содержать остатки, которые не встречаются в реципиентном антителе или в донорном антителе. Такие модификации осуществляют с целью дополнительного улучшения характеристик антитела. Как правило, гуманизированное антитело содержит по существу все, или по меньшей мере один, чаще всего два вариабельных домена, в которых все или почти все гипервариабельные петли соответствуют гипервариабельным петлям иммуноглобулина, не принадлежащего человеку, а все или почти все FR получены из последовательности иммуноглобулина человека. Гуманизированное антитело необязательно содержит по меньшей мере часть константного участка иммуноглобулина (Fc), как правило иммуноглобулина человека. Дополнительную информацию можно найти в Jones et al., Nature 321: 522-525 (1986); Riechmann et al., Nature 332: 323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2: 593-596 (1992).
Термин "комплекс" или "комплексный" в настоящем описании относится к ассоциации двух или более молекул, которые соединяются друг с другом посредством связей и/или взаимодействий (таких как вандерваальсовы, гидрофобные, гидрофильные взаимодействия), отличных от пептидных связей. В одном из вариантов осуществления комплекс представляет собой гетеромультимерную молекулу. Следует понимать, что термин "белковый комплекс" или "полипептидный комплекс" в соответствии с настоящим изобретением включает комплексы, которые содержат небелковый компонент, конъюгированный с белком, входящим в состав белкового комплекса (например, небелковый компонент может включать, без ограничения, химические молекулы, такие как токсин или средство, облегчающее детекцию).
Используемый в настоящем описании термин "гетеромультимер" или "гетеромультимерный" описывает молекулу, содержащую два или более полипептидов с разными последовательностями, которые соединяются друг с другом посредством непептидной ковалентной связи (такой как дисульфидная связь), и/или посредством нековалентных взаимодействий (таких как водородные связи, ионные связи, вандерваальсовы или гидрофобные взаимодействия). Кроме того, данное определение охватывает изначально связанные мультимерные полипептиды (например, полученные в виде одной непрерывной полипептидной цепи). В соответствии с настоящим описанием гетеромультимеры включают, например, одноцепочечные антитела и многоцепочечные антитела, а также мультимеры, содержащие одну или несколько половин антител, связанных с одним или несколькими иммуноадгезинами. Гетеромультимеры включают полипептиды и/или полипептидные комплексы, в которых линкер HD может присутствовать или отсутствовать.
Антитело по настоящему изобретению, "которое связывает" представляющий интерес антиген, представляет собой антитело, которое связывает антиген с аффинностью, достаточной для того, чтобы антитело можно было использовать в качестве диагностического и/или терапевтического средства, направленного на белок, или клетку, или ткань, экспрессирующую антиген, и которое практически не вступает в перекрестные реакции с другими белками. В таких вариантах осуществления степень связывания антитела с белком "не мишенью" составляет примерно менее 10% от связывания антитела с конкретным белком-мишенью, в соответствии с результатами анализа методом сортинга клеток с возбуждением флуоресценции (FACS), или радиоиммунопреципитации (РИА), или ELISA. Что касается связывания антитела с молекулой-мишенью, термин "специфически связывающий" или "специфически связывающийся с" или "специфичный к" в применении к конкретному полипептиду или эпитопу на конкретной полипептидной мишени, относится к связыванию, которое измеримо отличается от неспецифического взаимодействия (например, неспецифическое взаимодействие может представлять собой связывание с бычьим сывороточным альбумином или казеином). Специфическое связывание можно измерить, например, путем определения связывания молекулы по сравнению со связыванием контрольной молекулы. Например, специфическое связывание можно определить путем конкурентного анализа в присутствии контрольной молекулы, подобной мишени, например, в избытке немеченой мишени. В данном случае специфическое связывание имеет место, если связывание меченой мишени с зондом конкурентно ингибируется избытком немеченой мишени. Термин "специфически связывающий" или "специфически связывающийся с" или "специфичный к" в применении к конкретному полипептиду или эпитопу на конкретной полипептидной мишени, в соответствии с настоящим описанием может использоваться в применении, например, к молекуле, характеризующейся Kd комплекса с мишенью, составляющей примерно от 1 мкМ до 1 фМ, альтернативно примерно от 200 нМ до 1 фМ, альтернативно примерно от 200 нМ до 1 пМ, альтернативно примерно от 150 нМ до 1 фМ, альтернативно примерно от 150 нМ до 1 пМ, альтернативно примерно от 100 нМ до 1 фМ, альтернативно примерно от 100 нМ до 1 пМ, альтернативно примерно от 60 нМ до 1 фМ, альтернативно примерно от 60 нМ до 1 пМ, альтернативно примерно от 50 нМ до 1 фМ, альтернативно примерно от 50 нМ до 1 пМ, альтернативно примерно от 30 нМ до 1 фМ, альтернативно примерно от 30 нМ до 1 пМ, альтернативно примерно от 20 нМ до 1 фМ, альтернативно примерно от 20 нМ до 1 пМ, альтернативно примерно от 10 нМ до 1 фМ, альтернативно примерно от 10 нМ до 1 пМ, альтернативно примерно от 8 нМ до 1 фМ, альтернативно примерно от 8 нМ до 1 пМ, альтернативно примерно от 6 нМ до 1 фМ, альтернативно примерно от 6 нМ до 1 пМ, альтернативно примерно от 4 нМ до 1 фМ, альтернативно примерно от 4 нМ до 1 пМ, альтернативно примерно от 2 нМ до 1 фМ, альтернативно примерно от 2 нМ до 1 пМ, альтернативно примерно от 1 нМ до 1 фМ, альтернативно примерно от 1 нМ до 1 пМ. В одном из вариантов осуществления термин "специфическое связывание" относится к взаимодействию молекулы, которая связывается с конкретным полипептидом, или эпитопом на конкретном полипептиде, и практически не связывается с любым другим полипептидом или полипептидным эпитопом.
Термин "аффиность связывания", как правило, относится к силе общей суммы нековалентных взаимодействий между одним участком связывания молекулы (например, антитела) и его партнером по связыванию (например, антигеном). Если не указано иначе, используемый в настоящем описании термин "аффинность связывания" относится к характеристической аффинности связывания, которая отражает в соотношении 1:1 взаимодействие между членами пары по связыванию (например, антителом и антигеном). Аффинность молекулы X в отношении ее партнера Y, как правило, можно выразить в виде константы диссоциации (Kd). Например, Kd может составлять примерно 200 нМ, 150 нМ, 100 нМ, 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 8 нМ, 6 нМ, 4 нМ, 2 нМ, 1 нМ или менее. Аффинность можно измерить с помощью обычных методов, известных в данной области, включающих описанные в настоящем документе. Антитела с низкой аффинностью обычно связывают антиген медленно и, как правило, образовавшиеся комплексы легко распадаются, тогда как антитела с высокой аффинностью обычно связывают антиген быстрее и, как правило, образовавшиеся комплексы дольше сохраняются. Разные методы измерения аффинности связывания хорошо известны в данной области и любой из них можно использовать в целях настоящего изобретения.
В одном из вариантов осуществления "Kd" или "значение Kd" согласно данному изобретению измеряют методом поверхностного плазмонного резонанса с помощью BIAcoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc, Piscataway, NJ) при 25°C, используя чипы с иммобилизованным антигеном CM5 при ~10 единицах ответа (RU). Антитела по настоящему изобретению могут иметь аффинность к мишеням, соответствующую значениям Kd примерно от 1 мкМ до 1 фМ, альтернативно примерно от 200 нМ до 1 фМ, альтернативно примерно от 200 нМ до 1 пМ, альтернативно примерно от 150 нМ до 1 фМ, альтернативно примерно от 150 нМ до 1 пМ, альтернативно примерно от 100 нМ до 1 фМ, альтернативно примерно от 100 нМ до 1 пМ, альтернативно примерно от 60 нМ до 1 фМ, альтернативно примерно от 60 нМ до 1 пМ, альтернативно примерно от 50 нМ до 1 фМ, альтернативно примерно от 50 нМ до 1 пМ, альтернативно примерно от 30 нМ до 1 фМ, альтернативно примерно от 30 нМ до 1 пМ, альтернативно примерно от 20 нМ до 1 фМ, альтернативно примерно от 20 нМ до 1 пМ, альтернативно примерно от 10 нМ до 1 фМ, альтернативно примерно от 10 нМ до 1 пМ, альтернативно примерно от 8 нМ до 1 фМ, альтернативно примерно от 8 нМ до 1 пМ, альтернативно примерно от 6 нМ до 1 фМ, альтернативно примерно от 6 нМ до 1 пМ, альтернативно примерно от 4 нМ до 1 фМ, альтернативно примерно от 4 нМ до 1 пМ, альтернативно примерно от 2 нМ до 1 фМ, альтернативно примерно от 2 нМ до 1 пМ, альтернативно примерно от 1 нМ до 1 фМ, альтернативно примерно от 1 нМ до 1 пМ. Чтобы измерить значения Kd методом поверхностного плазмонного резонанса, биосенсорные чипы из карбоксиметилированного декстрана (CMS, BIAcore Inc.) активируют гидрохлоридом N-этил-Nʹ-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разбавляют 10 мМ раствором ацетата натрия, рН 4,8, до концентрации 5 мкг/мл (~0,2 мкМ) и затем вводят со скоростью потока 5 мкл/мин до достижения примерно 10 единиц ответа (RU) присоединенного белка. После введения антигена добавляют 1М этаноламин, чтобы блокировать непрореагировавшие группы. Чтобы измерить кинетические параметры, двукратные серийные разведения Fab (например, от 0,78 нМ до 500 нМ) вводят в PBS, содержащем 0,05% твин 20 (PBST) при 25°C со скоростью потока примерно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) рассчитывают с использованием простой модели связывания Ленгмюра "один к одному" (программное обеспечение для обработки результатов BIAcore, версия 3.2) путем одновременной аппроксимации сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kd) рассчитывают как отношение koff/kon. См., например, Chen et al., J. Mol. Biol. 293:865-881 (1999). Если скорость ассоциации превышает 106 М-1с-1 по данным поверхностного плазмонного резонанса, указанного выше, то ее можно определить методом гашения флуоресценции, в котором измеряют увеличение или уменьшение интенсивности излучения флуоресценции (возбуждение = 295 нм; излучение = 340 нм, полоса пропускания 16 нм) антителом против антигена (в виде Fab) в концентрации 20 нм в PBS, рН 7,2, при 25°C в присутствии возрастающих концентраций антигена, с помощью спектрометра, такого как спектрофотометр с остановленным потоком (Aviv Instruments) или спектрофотометр 8000 серии SLM-Aminco (ThermoSpectronic) с перемешиваемой красной кюветой.
"On-скорость", или "скорость ассоциации", или "скорость образования комплекса", или "kon", в соответствии с настоящим изобретением также можно определить с помощью описанного выше метода поверхностного плазмонного резонанса, с помощью BIAcoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc, Piscataway, NJ) при 25°C, используя чипы с иммобилизованным антигеном CM5 при ~10 единицах ответа (RU). Коротко говоря, биосенсорные чипы из карбоксиметилированного декстрана (СМ5, BIAcore Inc) активируют гидрохлоридом N-этил-Nʹ-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разбавляют 10 мМ раствором ацетата натрия, рН 4,8, до концентрации 5 мкг/мл (~0,2 мкМ) и затем вводят со скоростью потока 5 мкл/мин до достижения примерно 10 единиц ответа (RU) присоединенного белка. После введения антигена добавляют 1М этаноламин, чтобы блокировать непрореагировавшие группы. Чтобы измерить кинетические параметры, двукратные серийные разведения Fab (например, от 0,78 нМ до 500 нМ) вводят в PBS, содержащем 0,05% твин 20 (PBST) при 25°C со скоростью потока примерно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) рассчитывают с использованием простой модели связывания Ленгмюра "один к одному" (программное обеспечение для обработки результатов BIAcore, версия 3.2) путем одновременной аппроксимации сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kd) рассчитывают как отношение koff/kon. См., например, Chen et al., J. Mol. Biol. 293:865-881 (1999). Однако, если скорость ассоциации превышает 106 М-1с-1 по данным поверхностного плазмонного резонанса, указанного выше, то ее можно определить методом гашения флуоресценции, в котором измеряют увеличение или уменьшение интенсивности излучения флуоресценции (возбуждение = 295 нм; излучение = 340 нм, полоса пропускания 16 нм) антителом против антигена (в виде Fab) в концентрации 20 нМ в PBS, рН 7,2, при 25°C в присутствии возрастающих концентраций антигена, с помощью спектрометра, такого как спектрофотометр с остановленным потоком (Aviv Instruments) или спектрофотометр 8000 серии SLM-Aminco (ThermoSpectronic) с перемешиваемой кюветой.
Если не указано иначе, термины "биологически активный", "биологическая активность" и "биологические характеристики", используемые в применении к полипептиду по настоящему изобретению, такому как антитело, его фрагмент или производное, обозначают способность связываться с биологической молекулой.
Гетеромультимеры по настоящему изобретению, как правило, очищают практически до гомогенности. Фразы "практически гомогенный", "практически в гомогенном виде" и "практически гомогенность" используют для того, чтобы указать, что продукт практически не содержит побочных веществ, образующихся в результате нежелательных сочетаний полипептидов.
Выражаемая как чистота, практическая гомогенность означает, что количество побочных продуктов не превышает 10%, 9%, 8%, 7%, 6%, 4%, 3%, 2% или 1% по массе, или оно составляет менее 1% по массе. В одном из вариантов осуществления количество побочного продукта ниже 5%.
Термин "биологическая молекула" относится к молекулам нуклеиновых кислот, белков, углеводов, липидов и их сочетаний. В одном из вариантов осуществления биологическая молекула существует в природе.
Термин "выделенный", используемый в применении к разным гетеромультимерам, описанным в настоящем документе, обозначает гетеромультимер, идентифицированный и отделенный от и/или выделенный из клетки или клеточной культуры, из которой он экспрессируется. Загрязняющие компоненты его природной среды представляют собой вещества, которые обычно препятствуют диагностическим или терапевтическим применениям полипептида, и включают ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления гетеромультимер очищают (1) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с помощью секвенатора с вращающейся чашкой, или (2) до гомогенности методом SDS-PAGE в невосстанавливающих или восстанавливающих условиях с окрашиванием Кумасси синим или, предпочтительно, серебром. Выделенные гетеромультимеры включают антитела in situ внутри рекомбинантных клеток, поскольку по меньшей мере один компонент полипептидной природной среды отсутствует. Однако чаще всего выделенные полипептиды получают, используя по меньшей мере одну стадию очистки.
Термин "связанный" или "связывает" в соответствии с настоящим описанием относится либо к простой пептидной связи, соединяющей первую и вторую аминокислотные последовательности, либо к мостику, который включает третью аминокислотную последовательность, представляющую собой пептид, связанный с первой и второй аминокислотными последовательностями и находящийся между ними. Например, мостик может представлять собой аминокислотный линкер, связанный с С-концом одной аминокислотной последовательности и N-концом другой аминокислотной последовательности.
Используемый в настоящем описании термин "линкер" обозначает аминокислотную последовательность, содержащую две или более аминокислоты в длину. Линкер может состоять из нейтральных, полярных или неполярных аминокислот. Длина линкера может составлять, например, от 2 до 100 аминокислот, например, от 2 до 50 аминокислот, например, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 аминокислот. Линкер может быть "отщепляемым", например, он может отщепляться в результате ауторасщепления, либо ферментативного или химического расщепления. Участки расщепления аминокислотных последовательностей, а также ферменты и химические вещества, способные осуществлять расщепление по таким участкам, хорошо известны в данной области и, кроме того, описаны в настоящем документе.
Используемый в настоящем описании термин "HD линкер" или "линкер гетеродимеризации" относится к аминокислотному линкеру, который соединяет два полипептида, содержащих разные константные домены (CH) тяжелой цепи. Как правило, два полипептида, содержащих домены CH, соединяют путем связывания домена CH2 или СН3 первого полипептида с доменом VL, который сам является составной частью второго CH-содержащего полипептида. В некоторых вариантах осуществления HD линкер соединяет домен СН3 первого полипептида непосредственно с доменом VH второго CH-содержащего полипептида. Как правило, эффективными являются HD линкеры, содержащие 15-100 аминокислот, такие как HD линкеры, содержащие 20-40 аминокислот, 25-40 аминокислот и 30-40 аминокислот. В конкретных вариантах осуществления HD линкер содержит от 30 до 39 аминокислот в длину (например, 30, 31, 32, 33, 34, 35, 36, 37, 38 или 39 аминокислот в длину). HD линкер может быть "отщепляемым", например, он может отщепляться в результате ауторасщепления, либо ферментативного или химического расщепления, которое проводят с использованием методов и реагентов, стандартно используемых в данной области.
Используемый в настоящем описании термин "CLH линкер" или "линкер, соединяющий родственные цепи LC и HC" обозначает аминокислотный линкер, который соединяет легкую цепь с родственной тяжелой цепью. CLH линкер, как правило, включает аминокислоты, которые соединяют домен CL легкой цепи с доменом VH тяжелой цепи. В некоторых вариантах осуществления CLH линкер соединяет домен VL легкой цепи непосредственно с доменом VH тяжелой цепи. Как правило, эффективными являются CLH линкеры, содержащие 10-80 аминокислот, такие как CLH линкеры, содержащие 20-40 аминокислот, 25-40 аминокислот, 30-40 аминокислот и 30-35 аминокислот (например, 30, 31, 32, 33, 34 или 35 аминокислот). Одноцепочечные антитела по настоящему изобретению могут содержать несколько линкеров CLH, которые могут иметь одинаковые или разные последовательности и/или длины. В предпочтительных вариантах осуществления одноцепочечное антитело содержит два линкера (CLH линкер1 и CLH линкер2), каждый из которых соединяет легкую цепь с родственной тяжелой цепью. CLH линкер может быть "отщепляемым", например, он может отщепляться в результате ауторасщепления, либо ферментативного или химического расщепления, которое проводят с использованием методов и реагентов, стандартно используемых в данной области.
Ферментативное расщепление "линкера" или "соединяющей последовательности" может включать использование эндопептидазы, такой как, например, урокиназа, Lys-C, Asp-N, Arg-C, V8, Glu-C, химотрипсин, трипсин, пепсин, папаин, тромбин, тканевый активатор плазминогена (ТРА), гененаза, фактор Ха, TEV (цистеин-протеаза вируса травления табака), энтерокиназа, HRV C3 (протеаза C3 риновируса человека), кининогеназа, а также субтилизиноподобные конвертазы пробелков (например, фурин (PC1), PC2 или PC3), или двухосновная N-аргинин-конвертаза. В желательном варианте осуществления ферментативное расщепление протекает под действием эндопептидазы фурин. Химическое расщепление можно проводить с использованием, например, гидроксиламина, N-хлорсукцинимида, N-бромсукцинимида или цианбромида.
Используемый в настоящем описании термин "участок расщепления фуриновой эндопептидазой" относится к аминокислотной последовательности X1-X2-X3-аргинин (SEQ ID NO:6), где X1 обозначает остаток основной аминокислоты (природной или неприродной, модифицированной или немодифицированной), а каждый из Х2 и Х3 может обозначать остаток любой аминокислоты (природной или неприродной, модифицированной или немодифицированной), который может отщепляться по С-концу под действием фуриновой эндопептидазы. Фуриновая эндопептидаза отщепляет остаток аргинина по С-концу. В некоторых вариантах осуществления участок расщепления фурином содержит аминокислотную последовательность RXRXYR, где Y обозначает K или R, а Х обозначает любой аминокислотный остаток (SEQ ID NO:7), более конкретно указанная аминокислотная последовательность представляет собой RXRXRR (SEQ ID NO:8). В некоторых вариантах осуществления участок расщепления фурином содержит аминокислотную последовательность RKRKRR (SEQ ID NO:9). В некоторых других вариантах осуществления участок расщепления фурином содержит аминокислотную последовательность RHRQPR (SEQ ID NO:10). В следующих вариантах осуществления участок расщепления фурином содержит аминокислотную последовательность RSRKRR (SEQ ID NO:11).
"Участок расщепления эндопептидазой Lys-C" в соответствии с данным описанием представляет собой остаток лизина в аминокислотной последовательности, который может отщепляться по С-концу под действием эндопептидазы Lys-C. Эндопептидаза Lys-C отщепляет остаток лизина по С-концу.
Ферментативное расщепление "линкера" или "соединяющей последовательности" также можно проводить с использованием экзопептидазы, такой как, например, карбоксипептидаза А, карбоксипептидаза B, карбоксипептидаза D, карбоксипептидаза E (также называемая карбоксипептидаза Н), карбоксипептидаза М, карбоксипептидаза N или карбоксипептидаза Z, которая позволяет удалить остаточные последовательности распознавания эндопептидазой после расщепления эндопептидазой.
Термин "эффекторные функции" антитела относится к биологической активности, присущей Fc-участку (содержащему нативную или модифицированную аминокислотную последовательность) антитела, и изменяются в зависимости от изотипа антитела. Примеры эффекторных функций антител включают: связывание C1q и комплемент-зависимую цитотоксичность; связывание рецептора Fc; антителозависимую клеточно-опосредованную цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (таких как B-клеточный рецептор); и активацию В-клеток.
Термин "антителозависимая клеточно-опосредованная цитотоксичность" или "ADCC" относится к разновидности цитотоксичности, при которой секретируемый Ig связывается с рецепторами Fc (FcR), присутствующими на некоторых цитотоксических клетках (таких как природные клетки-киллеры (NK), нейтрофилы, и макрофаги), делая эти цитотоксические эффекторные клетки способными специфически связываться с антиген-несущей клеткой-мишенью с последующим ее уничтожением цитотоксическими средствами. Антитела являются "оружием" цитотоксических клеток и абсолютно необходимы для такого уничтожения. Основные клетки, опосредующие ADCC, NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гемопоэтических клетках показана в таблице 3 на странице 464 в Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Для определения ADCC-активности представляющей интерес молекулы in vitro можно использовать анализ ADCC, например, описанный в патенте США № 5500362 или 5821337. Эффекторные клетки, подходящие для применения в таких анализах, включают мононуклеарные клетки периферической крови (PBMC) и натуральные киллерные (NK) клетки. Альтернативно, или дополнительно, ADCC-активность представляющей интерес молекулы можно определить in vivo, например, у животной модели, по способу, описанному в Clynes et al., Proc. Natl. Acad. Sci. USA 95:652-656 (1998).
Термин "рецептор Fc", или "FcR", относится к рецептору, способному связываться с Fc-участком антитела. Предпочтительный FcR представляет собой нативную последовательность FcR человека. Кроме того, предпочтительный FcR обладает способностью связываться с антителом IgG (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII, и FcγRIII, в том числе аллельные варианты и альтернативно сплайсированные формы таких рецепторов. Рецепторы FcγRII включают FcγRIIA ("активирующий рецептор") и FcyRIIB ("ингибирующий рецептор"), которые имеют сходные аминокислотные последовательности, различающиеся, прежде всего, по цитоплазматическим доменам. Активирующий рецептор FcyRIIA содержит иммунорецепторный тирозинсодержащий мотив активации (ITАМ) в цитоплазматическом домене. Ингибирующий рецептор FcyRIIB содержит иммунорецепторный тирозинсодержащий мотив ингибирования (ITIM) в цитоплазматическом домене (см. обзор M. in Daeron, Annu. Rev. Immunol. 15: 203-234 (1997)). FcR описаны в Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-492 (1991); Capel et al., Immunomethods 4: 25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126: 330-41 (1995). Описанный в настоящем описании термин "FcR" также охватывает другие FcR, в том числе те, которые могут быть идентифицированы в будущем. Данный термин также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG к плоду (Guyer et al., J. Immunol. 117:587 (1976) and Kim et al., J. Immunol. 24:249 (1994)).
"Эффекторные клетки человека" представляют собой лейкоциты, которые экспрессируют один или несколько FcR и выполняют эффекторные функции. Предпочтительно, клетки экспрессируют по меньшей мере FcγRIII и выполняют ADCC-эффекторную функцию. Примеры лейкоцитов человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (PBMC), природные киллеры (NK), моноциты, цитотоксические Т-клетки и нейтрофилы; причем предпочтительными являются PBMC и NK-клетки. Эффекторные клетки можно выделить из природного источника, например, из крови.
Термин "комплемент-зависимая цитотоксичность" или "CDC" относится к лизису клетки-мишени в присутствии комплемента. Активация классического пути комплемента инициируется связыванием первого компонента системы комплемента (C1q) с антителами (соответствующего подкласса), связанными со своим специфическим антигеном. Активацию комплемента можно определить с помощью анализа CDC, например, описанного в Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996).
Термин "терапевтически эффективное количество" относится к количеству гетеромультимера, антитела, фрагмента антитела или производного, эффективного для лечения заболевания или расстройства у индивида. В случае опухоли (например, раковой опухоли) терапевтически эффективное количество гетеромультимера, антитела или фрагмента антитела (например, мультиспецифического антитела или фрагмента антитела) может уменьшать число раковых клеток; уменьшать размер первичных опухолей; ингибировать (т.е. замедлять до некоторой степени и, предпочтительно, останавливать) инфильтрацию раковых клеток в периферические органы; ингибировать (т.е. замедлять до некоторой степени и, предпочтительно, останавливать) метастазирование опухоли; ингибировать в некоторой степени рост опухоли; и/или облегчать в некоторой степени один или несколько симптомов, связанных с расстройством. В зависимости от степени, в которой гетеромультимер, антитело или фрагмент антитела может предотвращать рост существующих раковых клеток и/или уничтожать существующие раковые клетки, эффект может быть цитостатическим и/или цитотоксическим. В случае противораковой терапии эффективность in vivo можно измерить, например, путем определения продолжительности жизни, времени до прогрессирования заболевания (ТТП), скорости ответа (RR), продолжительности ответа и/или качества жизни.
Под "уменьшением или подавлением" подразумевают способность вызывать общее снижение, составляющее, предпочтительно, 20% или более, более предпочтительно, 50% или более, и наиболее предпочтительно, 75%, 85%, 90%, 95% или более. Уменьшение или подавление может относиться к симптомам расстройства, подлежащего лечению, наличию или размеру метастазов, размеру первичной опухоли или размеру или количеству кровеносных сосудов при ангиогенных нарушениях.
Используемый в настоящем описании термин "цитотоксическое средство" относится к веществу, которое ингибирует или предотвращает функционирование клетки и/или вызывает разрушение клетки. Данный термин охватывает радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, Ra223, Р32, и радиоактивные изотопы Lu), химиотерапевтические средства, например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие средства, ферменты и их фрагменты, такие как нуклеолитические ферменты, антибиотики и токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, в том числе их фрагменты и/или варианты, а также разные описанные в настоящем описании противоопухолевые, противораковые и химиотерапевтические средства. В данном документе также описаны другие цитотоксические средства. Противоопухолевое средство вызывает разрушение опухолевых клеток.
"Химиотерапевтическое средство" представляет собой химическое соединение, используемое для лечения злокачественного новообразования. Примеры химиотерапевтических средств включают алкилирующие средства, такие как тиотепа и циклофосфамид CYTOXAN®; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, в том числе алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогенины (в особенности, буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, MARTNOL®); бета-лапахон; лапахол; колхицины; бетулиновая кислота; камптотецин (в том числе синтетический аналог топотекан (HYCAMTIN®), СРТ-11 (иринотекан, CAMPTOSAR®), ацетилкамптотецин, скополектин и 9-аминокамптотецин); бриостатин; каллистатин; СС-1065 (в том числе его синтетические аналоги адозелесин, карзелесин и бизелесин); подофиллотоксин; подофиллиновая кислота; тенипозид; криптофицины (в том числе криптофицин 1 и криптофицин 8); доластатин; дуокармицин (в том числе его синтетические аналоги KW-2189 и CB1-TM1); элеутеробин; панкратистатин; саркодиктиина; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид оксида мехлоретамина, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урацилиприт; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, в частности калихеамицин гамма 1 (см., например, Agnew, Chem. Intl. Ed. Engl. 33:183-186 (1994)); динемицин, в том числе динемицин A; эсперамицин; а также неокарциностатиновый хромофор и антибиотические хромофоры, родственные хромопротеину энедиину), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин Адриамицин® (в том числе морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидина, убенимекс, зиностатин, зорубицина; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; пуриновые аналоги, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолона пропионат, эпитиостанол, мепитиостан, тестолактон; антиметаболиты гормонов надпочечников, такие как аминоглютетимид, митотан, трилостан; компенсатор фолиевой кислоты, такой как фролиновая кислота; ацеглатон; альдофосфамида гликозид; аминолевулиновая кислота; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазиквон; элфорнитин; эллиптиния ацетат; эпотилон; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраерин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновая кислота; триазикон; 2,2ʹ,2"-трихлортриэтиламин; трихотецены (в частности, токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндезин (ELDISINE®, FILDESIN®); дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ара-С"); тиотепа; таксоиды, например, паклитаксел TAXOL® (Bristol-Myers Squibb Oncology, Princeton, NJ), не содержащий кремофор ABRAXANE®, композиция паклитаксела, содержащая наночастицы, полученные из альбумина (American Pharmaceutical Partners, Schaumberg, EL) и доксетаксел TAXOTERE® (Rhone-Poulenc Rorer, Antony, France); хлорамбуцил; гемцитабин (GEMZAR®), 6-тиогуанин; меркаптопурин, метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин (Velban®); платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин (Oncovin®); оксалиплатин; лейкововин; винорелбин (Navelbine®); новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметиломитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин (XELODA®); фармацевтически приемлемые соли, кислоты или производные любого из указанных выше средств; а также сочетания двух или более из указанных выше средств, такие как CHOP, сокращенное название комплексной терапии, включающей циклофосфан, доксорубицин, винкристин и преднизолон, и FOLFOX, сокращенное название схемы лечения, включающей применение оксалиплатина (ELOXATINTM) в сочетании с 5-FU и лейкововином.
Данное определение также охватывает анти-гормональные средства, которые регулируют, уменьшают, блокируют или ингибируют эффекты гормонов, способных стимулировать рост опухоли, и часто применяются для системного лечения, или лечения всего тела. Они сами могут являться гормонами. Примеры таких средств включают антиэстрогены и селективные модуляторы рецептора эстрогена (SERM), такие как, например, тамоксифен (в том числе тамоксифен NOLVADEX®), EVISTA® ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен Fareston®; антипрогестероны; понижающие регуляторы рецептора эстрогена (ERD); средства, которые действуют, подавляя или останавливая функционирование яичников, например, агонисты рилизинг-фактора лютеинизирующего гормона (LHRH), такие как лейпролида ацетат Lupron® и Eligard®, ацетат гозерелина, бузерелина ацетат и триптерелин; другие антиандрогены, такие как флутамид, нилутамид и бикалютамид; ингибиторы ароматазы, которые ингибируют фермент ароматазу, регулирующий продукцию эстрогена в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглютетимид, мегестролацетат MEGASE®, экземестан AROMASIN®, форместан, фадрозол, ворозол RIVISOR®, летрозол FEMARA® и анастрозол ARIMIDEX®. Кроме того, такое определение химиотерапевтических средств включает бисфосфонаты, такие как клодронат (например, BONEFOS® или OSTAC®), этидронат DIDROCAL®, NE-58095, золедроновая кислота/золедронат Zometa®, алендронат FOSAMAX®, памидронат Aredia®, тилудронат SKELID® или ризедронат Actonel®; а также троксацитабин (1,3-диоксоланнуклеозидный аналог цитозина); антисмысловые олигонуклеотиды, особенно те, которые ингибируют экспрессию генов, связанных с сигнальными путями, участвующими в аберрантной пролиферации клеток, такие как, например, PKC-альфа, Raf, H-Ras, и рецептор эпидермального фактора роста (EGF-R); вакцины, такие как вакцина THERATOPE® и вакцины для генной терапии, например, вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; ингибитор топоизомеразы 1 LURTOTECAN®; антагонист GnRH ABARELIX®; лапатиниба дитозилат (двойной низкомолекулярный ингибитор тирозинкиназ ErbB-2 и EGFR, также известный как GW572016); а также фармацевтически приемлемые соли, кислоты или производные любого из перечисленных выше средств.
Термин "средство, ингибирующее рост" в настоящем описании относится к соединениям или композициям, которые ингибируют рост клетки in vitro или in vivo. Так, средство, ингибирующее рост, может представлять собой средство, которое значительно снижает процент клеток в S-фазе. Примеры средств, ингибирующих рост, включают средства, блокирующие развитие клеточного цикла (в момент, отличный от S-фазы), такие как средства, которые индуцируют остановку G1 и М-фазы. Классические блокаторы M-фазы включают алкалоиды барвинка (например, винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Средства, которые останавливают G1, также могут вызвать остановку S-фазы, их примеры включают средства, алкилирующие ДНК, такие как тамоксифен, преднизон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил и ara-C. Дополнительную информацию можно найти в The Molecular Basis of Cancer, под редакцией Mendelsohn and Israel, глава 1, озаглавленная "Cell cycle regulation, oncogenes, and antineoplastic drugs", Murakami et al. (WB Saunders: Philadelphia, 1995), особенно, на стр. 13. Таксаны (паклитаксел и доцетаксел) представляют собой противоопухолевые препараты, полученные из тиса. Доцетаксел (TAXOTERE®, Rhone-Poulenc Rorer), полученный из европейского тиса, представляет собой полусинтетический аналог паклитаксела (TAXOL®, Bristol-Myers Squibb). Паклитаксел и доцетаксел стимулируют сборку микротрубочек из димеров тубулина и стабилизируют микротрубочки путем предотвращения деполимеризации, приводя к ингибированию митоза в клетках.
Используемый в настоящем описании термин "противораковая терапия" относится к обработке, которая уменьшает или ингибирует развитие злокачественного новообразования у индивида. Примеры противораковой терапии включают цитотоксическую лучевую терапию, а также введение индивиду терапевтически эффективного количества цитотоксического средства, химиотерапевтического средства, средства, ингибирующего рост, вакцины против рака, ингибитора ангиогенеза, пролекарства, цитокина, антагониста цитокина, кортикостероида, иммунодепрессанта, противорвотного средства, антитела или фрагмента антитела или анальгетика.
Термин "молекула-мишень" относится к молекуле, которая может связываться с белковым комплексом по настоящему изобретению (предпочтительно, с аффинностью, превышающей аффинность, соответствующую Kd 1 мкМ согласно анализу Скэтчарда). Примеры молекул-мишеней включают, без ограничения, растворимые белки сыворотки и их рецепторы, такие как цитокины и рецепторы цитокинов, адгезины, факторы роста и их рецепторы, гормоны, вирусные частицы (например, белок RSVF, CMV, StaphA, вирус гриппа, вирус гепатита С), микроорганизмы (например, белки бактериальных клеток, грибковые клетки), адгезины, белки CD и их рецепторы.
Термин "индивид" относится к позвоночным, таким как млекопитающие, например, к человеку. Млекопитающие включают, без ограничения, сельскохозяйственных животных (таких как коровы), спортивных животных, домашних животных (таких как кошки, собаки и лошади), приматов, мышей и крыс.
Если не указано иначе, коммерчески доступные реагенты, указанные в примерах, используют в соответствии с инструкциями производителя. Источником клеток, идентифицированных в нижеследующих примерах и на протяжении всего описания с помощью номеров доступа ATCC, является Американская коллекция типовых культур, Manassas, VA. Если не указано иначе, в настоящем изобретении используют стандартные технологии рекомбинантных ДНК, например, описанные в данном документе выше, а также в следующих руководствах: Sambrook et al., supra; Ausubel et al., Current Protocols in Molecular Biology (Green Publishing Associates and Wiley Interscience, NY, 1989); Innis et al., PCR Protocols: A Guide to Methods and Applications (Academic Press, Inc., NY, 1990); Harlow et al., Antibodies: A Laboratory Manual (Cold Spring Harbor Press, Cold Spring Harbor, 1988); Gait, Oligonucleotide Synthesis (IRL Press, Oxford, 1984); Freshney, Animal Cell Culture, 1987; Coligan et al., Current Protocols in Immunology, 1991.
На протяжении данного описания и формулы изобретения термин "содержать", или его вариации, такие как "содержит" или "содержащий", следует понимать как включение указанного объекта или группы объектов, но не исключение любого другого объекта или другой группы объектов.
Термин "пролекарство", используемый в данной заявке, относится к предшественнику или производному фармацевтически активного вещества, которое необязательно может быть менее цитотоксичным по отношению к опухолевым клеткам, чем исходное лекарственное средство, и может активироваться или превращаться в более активную исходную форму под действием ферментов. См., например, Wilman, “Prodrugs in Cancer Chemotherapy” Biochemical Society Transactions, 14, pp. 375-382, 615th Meeting Belfast (1986) and Stella et al., “Prodrugs: A Chemical Approach to Targeted Drug Delivery,” Directed Drug Delivery, Borchardt et al., (ed.), pp. 247-267, Humana Press (1985). Пролекарства включают, без ограничения, фосфатсодержащие пролекарства, тиофосфатсодержащие пролекарства, сульфатсодержащие пролекарства, пептидсодержащие пролекарства, пролекарства, модифицированные D-аминокислотами, гликозилированные пролекарства, бета-лактамсодержащие пролекарства, пролекарства, содержащие необязательно замещенный феноксиацетамид, или пролекарства, содержащие необязательно замещенный фенилацетамид, 5-фторцитозиновые и другие 5-фторуридиновые пролекарства, которые могут превращаться в более активное цитотоксическое свободное лекарственное средство. Примеры цитотоксических лекарственных средств, которые можно дериватизировать с получением пролекарственной формы для применения в настоящем изобретении, включают, без ограничения, описанные выше химиотерапевтические средства.
Термин "цитокин" является общим термином, включающим белки, которые высвобождаются одной популяцией клеток и действуют на другие клетки как межклеточные медиаторы. Примерами таких цитокинов являются лимфокины, монокины и традиционные полипептидные гормоны. К числу цитокинов относятся гормоны роста, такие как гормон роста человека (HGH), N-метионил-гормон роста человека и бычий гормон роста; паратгормон; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (ФСГ), тиреотропный гормон (ТТГ) и лютеинизирующий гормон (LH); эпидермальный фактор роста (EGF); печеночный фактор роста; фактор роста фибробластов (FGF); пролактин; плацентарный лактоген; факторы некроза опухоли альфа и бета; муллериан-ингибирующее вещество; гонадотропин-ассоциированный пептид мыши; ингибин; активин; фактор роста эндотелия сосудов; интегрин; тромбопоэтин (ТРО); факторы роста нервов, такие как NGF-α; фактор роста тромбоцитов; трансформирующие факторы роста (TGF), такие как TGF-альфа и TGF-β; инсулиноподобные факторы роста I и II; эритропоэтин (ЕРО); остеоиндуктивные факторы; интерфероны, такие как интерфероны альфа, бета и гамма, колониестимулирующие факторы (CSF), такие как макрофагальный CSF (M-CSF); гранулоцитарно-макрофагальный CSF (GM-CSF); и гранулоцитарный CSF (G-CSF); интерлейкины (ILS), такие как IL-1, IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12; IL-18, фактор некроза опухолей, такой как TNF-α или TNF-β; и другие полипептидные факторы, включающие LIF и лиганд kit (KL). В данном описании термин цитокин включает белки из природных источников или из культуры рекомбинантных клеток, а также биологически активные эквиваленты цитокинов с природной последовательностью.
Под "антагонистом цитокина" подразумевают молекулу, которая частично или полностью блокирует, подавляет или нейтрализует биологическую активность по меньшей мере одного цитокина. Например, антагонисты цитокинов могут ингибировать активность цитокинов путем ингибирования экспрессии и/или секреции цитокинов, или путем связывания цитокина или рецептора цитокина. Антагонисты цитокинов включают антитела, гетеромультимеры, пептиды с синтетическими или нативными последовательностями, иммуноадгезины и низкомолекулярные антагонисты, способные связываться с цитокинами или рецепторами цитокинов. Антагонист цитокина необязательно конъюгируют или гибридизуют с цитотоксическим средством.
Термин "иммуносупрессивное средство" в данном описании относится к веществам, которые действуют, подавляя или экранируя иммунную систему индивида, подлежащего лечению. Данный термин охватывает вещества, которые подавляют продуцирование цитокинов, осуществляют понижающую регуляцию экспрессии аутоантигена или подавляют ее, или маскируют антигены МНС. Примеры иммуносупрессивных средств включают такие средства как 2-амино-6-арил-5-замещенные пиримидины (см. патент США № 4665077); микофенолятмофетил, такой как CELLCEPT®; азатиоприн (IMURAN®, Azasan®/6-меркаптопурин; бромокриптин, даназол; дапсон; глутаральдегид (который маскирует антигены МНС, как описано в патенте США № 4120649); антиидиотипические антитела против антигенов МНС и фрагментов МНС; циклоспорин А; стероиды, такие как кортикостероиды и глюкокортикостероиды, например, преднизон, преднизолон, такой как Pediapred® (преднизолонфосфат натрий) или ORAPRED® (пероральный раствор преднизолонфосфата натрия), метилпреднизолон и дексаметазон; метотрексат (для перорального или подкожного введения) (Rheumatrex®, TrexallTM); гидроксихлорохин/хлорохин; сульфасалазин; лефлуномид; антагонисты цитокинов или рецепторов цитокинов, включающие антитела против интерферонов γ, β, или α, антитела против фактора некроза опухоли α (инфликсимаб или адалимумаб), антитела против TNFα-иммуноадгезина (ENBREL®, этанерцепт), антитела против фактора некроза опухоли β, антитела против интерлейкина 2 и антитела против рецептора IL-2; антитела против LFA-1, включающие антитела против CD11a и антитела против CD18; антитела против L3T4; гетерологичный антилимфоцитарный глобулин; поликлональные или pan-Т антитела, или моноклональные антитела против CD3 или антитела против CD4/CD4a; растворимый пептид, содержащий домен связывания LFA-3 (WO 90/08187); стрептокиназа; TGF-β; стрептодорназа; РНК или ДНК хозяина; FK506; RS-61443; дезоксиспергуалин; рапамицин; Т-клеточный рецептор (Cohen et al., патент США № 5114721); фрагменты Т-клеточного рецептора (Offner et al. Science 251: 430-432 (1991), WO 90/11294; Ianeway, Nature 341: 482 (1989); и WO 91/01133); антитела против Т-клеточного рецептора (ЕР 340109), такие как Т10В9; циклофосфамид (Cytoxan®); дапсон; пеницилламин (CUPRIMINE®); плазмаферез; или иммуноглобулин для внутривенного введения (IVIG). Их можно использовать по отдельности или в сочетании друг с другом, в частности, можно использовать сочетание стероида с другим иммунодепрессантом, или такие сочетания используют с последующим применением поддерживающей дозы нестероидного средства, чтобы уменьшить потребность в стероидах.
Термин "анальгетик" относится к лекарственному средству, которое действует, ингибируя или подавляя боль у индивида. Примеры анальгетиков включают нестероидные противовоспалительные препараты (НПВП), такие как ибупрофен (Motrin®), напроксен (Naprosyn®), ацетилсалициловая кислота, индометацин, сулиндак и толметин, в том числе их соли и производные, а также разные другие препараты, используемые для уменьшения возникающих колющих болей, такие как противосудорожные средства (габапентин, фенитоин, карбамазепин) или трициклические антидепрессанты. Конкретные примеры включают ацетаминофен, аспирин, амитриптилин (ELAVIL®), карбамазепин (TEGRETOL®), фенилтоин (DILANTIN®), габапентин (NEURONTIN®), (E)-N-ванилил-8-метил-6-нонеамид (капсаицин®), или блокатор нервов.
Термин "кортикостероид" относится к любому из нескольких синтетических или природных веществ, имеющих общую химическую структуру стероидов, которые имитируют или усиливают эффекты природных кортикостероидов. Примеры синтетических кортикостероидов включают преднизон, преднизолон (в том числе метилпреднизолон), дексаметазона триамцинолон и бетаметазон.
Термин "противораковая вакцина" в настоящем описании относится к составу, который стимулирует у индивида противораковый иммунный ответ. Противораковые вакцины обычно содержат источник веществ или клеток, связанных с раком (антиген), который может быть аутологичным (полученным из того же организма) или аллогенным (полученным из другого организма) по отношению к индивиду, наряду с другими компонентами (такими как вспомогательные вещества), обеспечивающими дополнительное стимулирование и усиление иммунного ответа на антиген. Противораковые вакцины могут стимулировать иммунную систему индивида, вызывая образование антител против одного или нескольких специфических антигенов, и/или Т-клеточных киллеров, которые атакуют раковые клетки, содержащие эти антигены.
Термин "противорвотное средство" представляет собой соединение, которое уменьшает или предотвращает тошноту у индивида. Противорвотные соединения включают, например, антагонисты рецепторов нейрокинина-1, антагонисты рецепторов 5НТ3 (такие как ондансетрон, гранисетрон, трописетрон и затисетрон), агонисты рецепторов GABAB, такие как баклофен, кортикостероиды, такие как дексаметазон, Kenalog®, Aristocort® или NASALIDE®, антидопаминергические средства, фенотиазины (например, прохлорперазин, флуфеназин, тиоридазин и мезоридазин), дронабинол, метроклопрамид, домперидон, галоперидол, циклизин, лоразепам, прохлорперазин и левомепромазин.
II. Конструирование одноцепочечных и многоцепочечных, содержащих линкеры антител и других гетеромультимеров
Описанные в настоящем описании гетеромультимеры, в том числе моноспецифические и мультиспецифические (например, биспецифические антитела), можно сконструировать с использованием одного или нескольких линкеров.
Применение линкеров позволяет получить относительно чистую популяцию гетеромультимеров, содержащих разные тяжелые и/или легкие цепи в одном гетеромультимере. В частности, как описано выше, антитела обычно содержат две идентичные тяжелые цепи, каждая из которых спаривается с одной из двух идентичных легких цепей. Применение технологии линкеров по настоящему изобретению позволяет осуществлять димеризацию двух разных тяжелых цепей антитела при получении одного гетеромультимерного антитела, содержащего одну полипептидную цепь. Линкеры могут соединять первую тяжелую цепь со второй тяжелой цепью, как правило, путем соединения домена СН3 первой тяжелой цепи с доменом VL легкой цепи, которая сама связана со второй тяжелой цепью посредством второго линкера. В гетеромультимерном одноцепочечном антителе третий линкер может соединять вторую легкую цепь непосредственно с первой тяжелой цепью, что приводит к образованию гетеромультимера, который содержит две разные тяжелые цепи, каждая из которых может спариваться с родственной ей легкой цепью. В других вариантах осуществления линкеры могут соединять первую тяжелую цепь со второй тяжелой цепью непосредственно путем связывания С-конца константного домена первой тяжелой цепи с N-концом вариабельного домена второй тяжелой цепи. Разные пары тяжелых и легких цепей в таком гетеромультимере могут иметь разную специфичность связывания в связи с наличием разных пар родственных тяжелых и легких цепей и, следовательно, гетеромультимер можно рассматривать как мультиспецифическое антитело. С помощью технологии линкеров по настоящему изобретению также можно получить гетеромультимер (например, многоцепочечное антитело), который может содержать три связанные полипептидные цепи, где одна полипептидная цепь содержит два HC, соединенных непосредственно друг с другом с помощью линкера HD, как описано выше, а две другие полипептидные цепи идентичны и образуют LC, которые соединяются с двумя HC, образуя две функциональные пары родственных HC/LC. В других вариантах осуществления применение линкеров HD и CLH позволяет получить мультиспецифическое антитело, содержащее две полипептидные цепи, где первая полипептидная цепь содержит первый LC, а другая полипептидная цепь содержит два HC и второй LC, который связан с первым HC через HD линкер и со вторым HC через CLH линкер. В других вариантах осуществления применение линкеров HD и CLH позволяет получить мультиспецифическое антитело, содержащее две полипептидные цепи, где первая полипептидная цепь содержит первый LC и два HC, где один HC соединяется с первым LC непосредственно через CLH линкер и со вторым HC непосредственно через HD линкер, а второй полипептид образует второй LC, который соединен с HC, не связанным с первым LC. В других вариантах осуществления применение технологии линкеров по настоящему изобретению позволяет получить гетеромультимер, в котором иммуноадгезин связан с половиной антитела посредством HD линкера. Для конструирования гетеромультимеров по настоящему изобретению технологию линкеров можно использовать отдельно или в сочетании с методикой выступ-впадина ("KnH"). Гетеромультимеры, содержащие линкеры, полученные с использованием, или без использования метода KnH, а также рекомбинантные способы получения гетеромультимеров подробно описаны ниже.
А. Линкеры для гетеромультимеров
Настоящее изобретение относится к гетеромультимерам, полученным с использованием линкеров. Например, гетеромультимер может содержать линкер, который соединяет С-конец константного домена тяжелой цепи иммуноглобулина с N-концом вариабельного домена тяжелой цепи иммуноглобулина. В других вариантах осуществления гетеромультимер дополнительно содержит один или несколько (например, два) дополнительных линкера, обеспечивающих правильное соединение тяжелой цепи с родственной легкой цепью (т.е. соединение тяжелой цепи с легкой цепью, с которой она связана посредством линкера). Такой гетеромультимер можно сконструировать с использованием, или без использования дополнительного домена гетеродимеризации, например, с применением методики KnH.
Как показано на схеме, приведенной на фиг. 1, типичный гетеромультимер представляет собой мультиспецифическое одноцепочечное антитело, которое содержит две разные тяжелые цепи (HC1 и HC2) и две разные легкие цепи (LC1 и LC2), где HC1 и LC1 образуют первую родственную пару, а HC2 и LC2 образуют вторую родственную пару. Типичный гетеромультимер содержит три линкера. Первый линкер (HD линкер) соединяет HC1 с LC2, а второй линкер (CLH линкер1) и третий линкер (CLH линкер2) соединяют LC1 с HC1 и LC2 с HC2, соответственно. Таким образом, линкеры, обеспечивающие соединение двух разных тяжелых цепей с родственными легкими цепями, позволяют получить гетеромультимерное одноцепочечное антител, обладающее мультиспецифичностью.
В конкретном варианте осуществления HD линкер является достаточно длинным для того, чтобы охватить расстояние между С-концом НС1 и N-концом LC2, (или, в случае отсутствия LC2, HC2) в собранном гетеромультимере (фиг. 1), обеспечивая надлежащее соединение первой и второй тяжелых цепей, но достаточно коротким, чтобы предотвратить межмолекулярную гомодимеризацию НС1 и/или НС2. Расстояния между С-концом НС1 и N-концом LC2 (или, в отсутствие LC2, НС2) могут различаться среди собранных гетеромультимеров, и, следовательно, длина HD линкеров также может варьировать в разных гетеромультимерах. Как правило, эффективными являются HD линкеры длиной 15-100 аминокислот, например, HD линкеры длиной 20-40 аминокислот, 25-40 аминокислот и 30-40 аминокислот. В конкретных вариантах осуществления HD линкер содержит от 36 до 39 аминокислот в длину (например, 36, 37, 38 или 39 аминокислот в длину).
В конкретном варианте осуществления каждый из линкеров CLH является достаточно длинным для того, чтобы охватить расстояние между N-концом тяжелой цепи и С-концом родственной легкой цепи в собранном гетеромультимере (фиг. 1), обеспечивая правильное соединение легкой цепи и тяжелой цепи, но достаточно коротким, чтобы предотвратить нежелательную межцепочечную связь (т.е., соединение легкой цепи с неродственной тяжелой цепью, с которой она не соединена посредством линкера). Расстояния между N-концом тяжелой цепи и С-концом родственной легкой цепи могут различаться между антителами, или даже внутри самого антитела (т.е. расстояние между N-концом тяжелой цепи и С-концом родственной легкой цепи первой родственной пары отличается от соответсвующего расстояния у второй родственной пары). Соответственно, длины HD линкеров могут варьировать среди гетеромультимеров по настоящему изобретению, а длина CLH линкера1 может отличаться от длины CLH линкера2 в одном гетеромультимере. Как правило, эффективными являются CLH линкеры длиной 10-80 аминокислот, например, CLH линкеры длиной 20-40 аминокислот, 25-40 аминокислот, 30-40 аминокислот и 30-35 аминокислот (например, 30, 31, 32, 33, 34 или 35 аминокислот). Гетеромультимеры по настоящему изобретению могут содержать два линкера (CLH линкер1 и CLH линкер2), соединеняющие первую легкую цепь с первой тяжелой цепью и вторую легкую цепь со второй тяжелой цепью.
Чтобы линкеры HD или CLH оставались гибкими и не образовывали вторичные структуры, используют линкеры, содержащие остатки глицина (G) и серина (S). Линкеры могут состоять только из остатков G и S, но могут также содержать и другие остатки, при условии, что линкеры остаются гибкими и обеспечивают внутрицепочечные соединения, описанные выше. В конкретных вариантах осуществления линкеры HD или CLH содержат повторы GGS (фиг. 2). В некоторых вариантах осуществления линкер HD содержит по меньшей мере 1 повтор GGS. Типичный HD линкер, описанный в настоящем документе, содержит 8-9 повторов GGS (SEQ ID NO:19) и участки расщепления эндопептидазой (например, участки расщепления фурином и Lys-C) на N- и С-концах (фиг. 2). В другом варианте осуществления линкеры CLH содержат не менее 1 повтора GGS. Типичный CLH линкер, описанный в настоящем документе, содержит 6 повторов GGS (SEQ ID NO:20) и участки расщепления эндопептидазой (например, участки расщепления фурином) на N- и С-концах (фиг. 2).
В. Отщепление линкеров гетеромультимера
После сборки гетеромультимеров по настоящему изобретению линкеры могут более не требоваться, и в некоторых вариантах осуществления могут отщепляться от гетеромультимеров. Для удаления линкеров можно использовать участки расщепления, которые расположены в линкерах, или в непосредственной близости от линкеров, но не присутствуют в компонентах последовательностей гетеромультимера, отличающихся от линкера, или не доступны для расщепления в используемых условиях.
На фиг. 2 показано расположение типичных участков расщепления в трех линкерах, которые могут присутствовать в типичном гетеромультимерном одноцепочечном антителе. Как правило, участки расщепления в линкерах расположены на С- и N-концах последовательностей линкера, или близко к С- и N-концам последовательностей линкера, или внутри последовательности антитела, или близко к месту, где соединяются антитело и линкер. Если один или несколько линкеров расщепляются с помощью эндопептидазы Lys-C (например, по остатку лизина на С-конце константного домена тяжелой цепи), может потребоваться модификация последовательности гетеромультимера с целью удаления участков расщепления эндопептидазой Lys-C. Примером такой модификации является замена лизина в шарнирном участке на аланин (например, K222A, система нумерации Kabat; K222A, система нумерации EU в типичных описанных в настоящем описании гетеромультимерах). Могут потребоваться модификации других участков расщепления, которые проводят подобным образом, если для использования в настоящем изобретении выбирают разные расщепляющие средства.
Расщепление аминокислотных последовательностей по конкретным участкам является стандартной процедурой в данной области и может включать ферментативное расщепление, химическое расщепление или ауто-процессинг. Например, линкер можно отщепить от белка с помощью эндопептидазы. Типичные эндопептидазы включают, без ограничения, фурин, урокиназу, Lys-C, Asp-N, Arg-C, V8, Glu-C, тромбин, тканевый активатор плазминогена (tРА), гененазу (вариант протеазы субтилизина BPNʹ), фактор Ха, TEV (цистеин-протеаза вируса травления табака), энтерокиназу, C3 HRV (протеаза C3 риновируса человека), кининогеназу, химотрипсин, трипсин, пепсин и папаин, которые являются коммерчески доступными (например, от Boehringer Mannheim, Thermo Scientific или New England Biolabs). Субтилизиноподобные пропротеин-конвертазы, такие как фурин (PC1), PC2 и PC3, (Steiner (1991) in Peptide Biosynthesis and Processing (Fricker ed.) pp. 1-16, CRC Press, Boca Raton, FL; Muller et al., JBC 275:39213-39222, (2000)) и двухосновные N-аргинин-конвертазы (Chow et al., JBC 275: 19545-19551 (2000)), осуществляют расщепление по двухосновным участкам. Lys-C отщепляет на карбоксильном конце остатки лизина, V8 и Glu-C отщепляют на карбоксильном конце остатки глутаминовой кислоты, Arg-C отщепляет на карбоксильном конце остатки аргинина, Asp-N отщепляет на амино-конце остатки аспарагиновой кислоты, химотрипсин отщепляет на карбоксильном конце остатки тирозина, фенилаланина, триптофана и лейцина, а трипсин отщепляет на карбоксильном конце остатки аргинина и лизина. TEV расщепляет аминокислотную последовательность GluAsnLeuTyrPheGlnGly (SEQ ID NO:4) между остатками "Gln" и "Gly". Такие ферменты широко используются в данной области и методы их применения предоставляются производителями. На фиг. 3А показано типичное гетеромультимерное одноцепочечное антитело после расщепления фурином.
Альтернативно линкер можно отщепить от белка с помощью химического средства, такого как гидроксиламин. Гидроксиламин расщепляет пептидные связи аспарагин-глицин. Если гидроксиламин используют для отщепления линкера от белка, может потребоваться изменение нескольких остатков глицина или аспарагина, присутствующих в белке, чтобы избежать фрагментации белка.
В данной области известно много других химических веществ, способных расщеплять пептидные связи. Например, N-хлорсукцинимид отщепляет на С-конце остатки триптофана (Shechter et al., Biochemistry 15: 5071-5075 (1976)). N-бромсукцинимид и бромциан также расщепляют на С-конце остатки триптофана. Кроме того, 2-нитротиоцианобензойную кислоту или органофосфины можно использовать для расщепления белка по остаткам цистеина на N-конце (см., например, EP 0339217).
Также известно, что белки могут подвергаться аутопроцессингу. Например, белок Hedgehog подвергается процессингу по участку расщепления GlyAspTrpAsnAlaArgTrpCysPhe (SEQ ID NO:5) под действием протеолитической активности, присущей самому белку. Участок аутопротеолитического расщепления также можно ввести в состав линкерной или связующей последовательности.
После эндопептидазного расщепления гетеромультимеры по настоящему изобретению можно необязательно обработать одной или несколькими экзопептидазами до или после очистки гетеромультимера. На фиг. 3В показано гетеромультимерное одноцепочечное антитело после очистки и обработки фурином, Lys-C и экзопептидазой (например, карбоксипептидазой B). После обработки фурином линкер HD, еще присоединенный к домену СН31 гетеромультимера, удаляют путем обработки Lys-C. Оставшиеся последовательности, распознаваемые фурином, присоединенные к доменам CL1 и CL2, удаляют путем обработки экзопептидазой {например, карбоксипептидазой В).
С. Методика выступ-впадина
В состав гетеромультимеров по настоящему изобретению можно необязательно ввести домен гетеродимеризации с помощью технологии выступ-впадина (KnH) (см., например, патент США № 5731168, включенный в настоящее описание в качестве ссылки в полном объеме), которая обеспечивает создание граничной поверхности между двумя разными тяжелыми цепями антитела, инициируя их соединение. Как правило, способ включает введение "выступа" на граничной поверхности первой тяжелой цепи и соответствующей "полости" на граничной поверхности второй тяжелой цепи, так, чтобы выступ мог располагаться в полости, дополнительно стимулируя гетеродимеризацию тяжелых цепей.
Первая предпочтительная граничная поверхность включает по меньшей мере часть фрагмента СН3 первого константного домена тяжелой цепи (CH31) и по меньшей мере часть фрагмента СН3 второго константного домена тяжелой цепи (CH32). Выступы можно сконструировать путем замены аминокислот с маленькими боковыми цепями, присутствующими на граничной поверхности первого домена (например, CH31), на аминокислоты с крупными боковыми цепями (такими как тирозин или триптофан). Компенсаторные полости, размер которых идентичен или подобен размеру выступов, необязательно создают на граничной поверхности второго домена (например, CH32) путем замены аминокислот с крупными боковыми цепями на аминокислоты с меньшими боковыми цепями (такими как аланин или треонин). Вторая предпочтительная граничная поверхность включает по меньшей мере часть домена CL легкой цепи и домен CH1 тяжелой цепи, на которой взаимодействие выступ-впадина можно сконструировать по описанному выше способу. Если на граничной поверхности первого или второго домена присутствуют выступы или впадины, расположенные соответствующим образом и имеющие подходящие размеры, нужно только сконструировать соответствующие впадины или выступы на соседней граничной поверхности.
D. Моноспецифические или мультиспецифические гетеромультимеры, имеющие разные связывающие свойства
Следует понимать, что вариабельные домены таких гетеромультимеров можно получить с помощью нескольких способов. Например, вариабельные домены гетеромультимеров по настоящему изобретению могут быть такими же, как в существующих антителах, известных в данной области.
Как описано выше, HD линкер по настоящему изобретению можно использовать для получения гетеромультимеров с варьирующими связывающими свойствами. В некоторых вариантах осуществления настоящее изобретение можно использовать для получения одноцепочечного моноспецифического антитела, которое содержит две половины антител, обладающие разной аффинностью связывания к одному целевому эпитопу. В другом варианте осуществления настоящее изобретение можно использовать для получения одноцепочечного мультиспецифического антитела (антитела, которое связывается по меньшей мере с двумя антигенами или по меньшей мере с двумя эпитопами на одном антигене). Как правило, в природных антителах IgG вариабельные участки каждой пары тяжелых и легких цепей антитела являются идентичными. Применение линкеров по настоящему изобретению позволяет ввести в состав антитела две разные тяжелые цепи с получением антител, содержащих антигенсвязывающие домены с разной специфичностью связывания (например, к эпитопам-мишеням), или антигенсвязывающие домены с одинаковой специфичностью связывания, но с разной аффинностью связывания. В некоторых вариантах осуществления HD линкер инициирует соединение двух разных тяжелых цепей, кодируемых в одной полипептидной цепи, с получением одноцепочечных антител, содержащих только тяжелые цепи. В других вариантах осуществления CLH линкеры инициируют соединение тяжелой цепи и родственной ей легкой цепи. В некоторых вариантах осуществления соединение одной из тяжелых цепей с родственной ей легкой цепью происходит в отсутствии CLH линкера с образованием гетеромультимера по настоящему изобретению. Один или несколько линкеров необязательно содержат участки расщепления эндопептидазой (например, участки расщепления фурином и Lys-C), которые обеспечивают удаление одного или нескольких линкеров из гетеромультимера после сборки. Гетеромультимер по настоящему изобретению необязательно включает одну или несколько граничных поверхностей, содержащих выступы и полости, сконструированные с использованием методики KnH.
Как показано на фиг. 1, описанный выше типичный биспецифический гетеромультимер (в данном случае биспецифическое антитело) содержит HD линкер и два линкера CLH, каждый из которых может содержать два участка расщепления фурином на N- и С-конце. При получении типичного гетеромультимера можно использовать методику KnH, чтобы дополнительно содействовать соединению HC1 и HC2.
Последовательность гетеромультимеров с отщепляемыми линкерами не должна содержать участки расщепления эндопептидазой (эндопептидазами), используемой для отщепления линкеров, или, если участки расщепления присутствуют в последовательности гетеромультимера, отличной от последовательности линкера, участки расщепления не должны расщепляться в используемых условиях, если также не предполагается расщепление гетеромультимера. Последовательность гетеромультимера можно сканировать с целью определения присутствия каких-либо участков расщепления (например, участка расщепления фурином или эндопептидазой Lys-C) в последовательностях тяжелых или легких цепей гетеромультимера, которые необходимо удалить, чтобы избежать расщепления самого гетеромультимера при удалении линкеров.
Одноцепочечные моноспецифические или мультиспецифические гетеромультимеры (например, антитела) можно сконструировать с помощью способов, описанных в настоящем документе, в которых две разные тяжелые цепи гетеромультимера соединяют непосредственно друг с другом с помощью HD линкера. В одном из вариантов осуществления в одноцепочечном, содержащем только тяжелую цепь моноспецифическом или мультиспецифическом гетеромультимере может отсутствовать домен CH1 (домены VH непосредственно соединены по меньшей мере с одним доменом CH, необязательно посредством шарнирных участков). В другом варианте осуществления тяжелые цепи одноцепочечного моноспецифического или мультиспецифического гетеромультимера не содержат домены CH1, но связаны с соответствующими им легкими цепями, в которых отсутствуют домены CL, посредством линкеров CLH. Такие гетеромультимеры можно использовать для соединения двух разных антигенов или для связывания B- и Т-клеток. В следующем варианте осуществления линкеры можно использовать для связывания половины антитела с иммуноадгезином.
Кроме того, изобретение охватывает гетеромультимеры, обладающие моноспецифичностью или мультиспецифичностью. В одном из вариантов осуществления первая тяжелая цепь, связанная с родственной легкой цепью посредством CLH линкера, соединена со второй тяжелой цепью, родственная легкая цепь которой соединяется с ней независимо от линкера.
E. Гетеромультимеры, имеющие разные эффекторные функции
В некоторых вариантах осуществления гетеромультимеры (например, одноцепочечные или многоцепочечные антитела или комплексы антитело-иммуноадгезин) по настоящему изобретению, сконструированные с помощью описанных в настоящем описании способов, могут содержать мутации в домене СН2, приводящие к изменению эффекторной функции. Как правило, мутация домена CH2 представляет собой мутацию N297, приводящую к образованию гетеромультимера, обладающего измененным статусом гликозилирования. В некоторых вариантах осуществления мутация N297 представляет собой замену N297A. Изменяемые эффекторные функции включают, без ограничения, связывание C1q и комплемент-зависимую цитотоксичность, связывание Fc-рецептора, антителозависимую клеточную цитотоксичность, фагоцитоз, понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора) и активацию В-клеток.
F. Конъюгированные белковые комплексы
Как подробно описано ниже, HD линкер по настоящему изобретению также можно использовать для получения белковых комплексов таких гетеромультимеров (например, моноспецифических, биспецифических, мультиспецифических, одно- или многоцепочечных антител), в которых константный домен модифицирован путем конъюгации с цитотоксическим средством. Например, с помощью HD линкера можно получить гетеромультимеры, в которых один из константных участков тяжелой цепи (НС1 или НС2) содержит модификацию, обеспечивающую конъюгацию с цитотоксическим средством, а другой константный участок не содержит такую модификацию. В одном примере HC1 конъюгируют с цитотоксическим средством, а НС2 не конъюгируют. Схематическая диаграмма, на которой изображен пример конъюгированного гетеромультимера по настоящему изобретению, приведена на фиг. 4. Типичное гетеромультимерное одноцепочечное антитело включает две полноразмерные тяжелые цепи и родственные им легкие цепи, а также описанные выше линкеры HD и CLH. Как показано на рисунке 4, одну из тяжелых цепей конъюгируют с цитотоксическим средством (например, токсином или антибиотиком). Подобным образом, в альтернативной гетеромультимерной конструкции один из константных участков легкой цепи может быть конъюгирован с цитотоксическим средством, а другой константный участок легкой цепи - нет (например, LC1 конъюгируют с цитотоксическим средством, а LC2 - нет). Как показано на фиг. 5А, конъюгированный белковый комплекс может представлять собой многоцепочечное антитело по настоящему изобретению. Конъюгация с цитотоксическим средством не ограничивается конкретным многоцепочечным антителом, изображенным на фиг. 5А. Для такой конъюгации по константному участку также можно использовать такие многоцепочечные антитела, как антитела, изображенные на фиг. 5В и 5С, а также гетеромультимеры иммуноадгезин-антитело.
В одном из конкретных примеров константный участок гетеромультимера можно модифицировать путем введения электрофильных фрагментов, которые могут взаимодействовать с нуклеофильными заместителями, присутствующими на связующем реагенте, используемом для присоединения цитотоксического средства к гетеромультимеру, или на самом цитотоксическом средстве. Сахара гликозилированных антител можно окислить, например, периодатсодержащими окисляющими реагентами, с образованием альдегидных или кетоновых групп, которые могут взаимодействовать с аминогруппой связующего реагента или цитотоксического средства. Образующиеся в результате иминогруппы, представляющие собой основания Шиффа, могут образовывать стабильную связь, или их можно восстановить, например, боргидридсодержащими реагентами, с образованием стабильных связей, в которых участвует аминогруппа. Нуклеофильные группы на цитотоксическом средстве включают, без ограничения, такие группы, как амин, тиол, гидроксил, гидразид, оксим, гидразин, тиосемикарбазон, гидразинкарбоксилат, и арилгидразид, способные взаимодействовать с образованием ковалентных связей с электрофильными группами, присутствующими на антителах и связующих реагентах, такими как: (i) активированные сложные эфиры, такие как сложные эфиры NHS, эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; и (iii) альдегиды, кетоны, карбоксильные и малеимидные группы.
G. Другие свойства белков
Гетеромультимеры по настоящему изобретению могут содержать последовательности из любого источника, включающие последовательности человека и мыши, или их сочетания. Последовательности некоторых фрагментов белков (например, гипервариабельных участков) также могут представлять собой искусственные последовательности, такие как последовательности, идентифицированные путем скрининга библиотеки (например, библиотеки фагового дисплея), включающие случайные последовательности.
Если гетеромультимеры содержат последовательности, полученные из разных источников, они могут представлять собой "химерные" гетеромультимеры, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующей последовательности антитела, полученного от конкретного вида, или принадлежащего к конкретному классу или подклассу антител, тогда как остальная часть цепи (цепей) идентична или гомологична соответствующей последовательности антитела, полученного из другого вида, или принадлежащего к другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют желательную биологическую активность (патент США № 4816567 и Morrison et al., Proc. Natl. Acad. Sci. USA 81: 6851-6855 (1984)). Такие химерные гетеромультимеры могут содержать, например, мышиные вариабельные участки (или их фрагменты) и человеческие константные участки.
Химерные одноцепочечные гетеромультимеры необязательно могут представлять собой "гуманизированные" одноцепочечные гетеромультимеры (например, одноцепочечные антитела), которые содержат минимальную последовательность, полученную из нечеловеческого антитела. Гуманизированные гетеромультимеры обычно получают из антител человека (реципиентное антитело), в которых остатки гипервариабельного участка реципиента заменены остатками гипервариабельного участка антитела отличного от человека вида (донорное антитело), такого как мышь, крыса, кролик или отличный от человека примат, которое обладает желательными специфичностью, аффинностью и емкостью. В некоторых случаях остатки каркасного участка (FR) иммуноглобулина человека заменяют соответствующими нечеловеческими остатками. Кроме того, гуманизированные гетеромультимеры могут содержать остатки, которые не встречаются в реципиентном антителе или в донорном антителе. Такие модификации осуществляют для дополнительного улучшения характеристик гетеромультимера. Обычно гуманизированный гетеромультимер содержит по меньшей мере один, как правило, оба вариабельных домена, в которых все или практически все гипервариабельные петли соответствуют гипервариабельным петлям нечеловеческого иммуноглобулина, а все или практически все FR представляют собой последовательности иммуноглобулина человека. Гуманизированный гетеромультимер также может содержать по меньшей мере часть константного участка иммуноглобулина (Fc), как правило, иммуноглобулина человека. Дополнительную информацию можно найти в Jones et al., Nature 321: 522-525 (1986); Riechmann et al., Nature 332: 323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992).
Более подробно, гуманизированный гетеромультимер может содержать один или несколько аминокислотных остатков, полученный из источника, отличного от человека. Указанные нечеловеческие аминокислотные остатки часто называют "импортными" остатками, которые обычно получают из "импортного" вариабельного домена. Гуманизацию можно проводить по способу Winter с соавторами (Jones et al., Nature 321: 522-525 (1986); Riechmann et al., Nature 332:323-327 (1988); Verhoeyen et al., Science 239:1534-1536 (1988)), путем замены CDR грызунов или последовательностей CDR соответствующими последовательностями антитела человека. Соответственно, такие "гуманизированные" гетеромультимеры представляют собой химерные антитела (патент США № 4816567), в которых фрагмент, существенно меньший интактного вариабельного домена человека, заменяют на соответствующую последовательность отличного от человека вида. На практике гуманизированные гетеромультимеры, как правило, представляют собой антитела человека, в которых некоторые остатки CDR и, возможно, некоторые остатки FR заменены остатками аналогичных участков антител грызунов.
Выбор вариабельных доменов человека как легкой, так и тяжелой цепи, подходящих для получения гуманизированных гетеромультимеров, очень важен для снижения антигенности. В соответствии с так называемым методом "наилучшего приближения" последовательность вариабельного домена антитела грызуна скринируют против целой библиотеки известных последовательностей вариабельных доменов человека. Затем последовательность человека, наиболее приближающуюся к последовательности грызуна, используют в качестве каркасного участка человека (FR) для получения гуманизированного гетеромультимера (Sims et al., J. Immunol. 151:2296 (1993); Chothia et al., J. Mol. Biol. 196:901 (1987)). В другом способе используют конкретный каркасный участок, полученный из консенсусной последовательности всех антител человека из конкретной подгруппы легких или тяжелых цепей. Один и тот же каркасный участок можно использовать для получения нескольких разных гуманизированных гетеромультимеров (Carter et al., Proc. Natl. Acad. Sci. USA 89: 4285 (1992); Presta et al., J. Immnol. 151: 2623 (1993)).
Кроме того, важно, чтобы при гуманизации гетеромультимеров сохранялись высокая аффинность в отношении одного или нескольких антигенов-мишеней и другие желательные биологические свойства. Для достижения этой цели, в соответствии с иллюстративным способом гуманизированные гетеромультимеры получают путем анализа исходных последовательностей и разных концептуальных гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются доступными и известны специалистам в данной области. Существуют компьютерные программы, которые иллюстрируют и отображают возможные трехмерные конформационные структуры выбранных кандидатных последовательностей иммуноглобулина. Исследование этих изображений позволяет анализировать вероятную роль остатков в функционировании последовательностей иммуноглобулина-кандидата, т.е. определить остатки, которые влияют на способность иммуноглобулина-кандидата связываться с антигеном. Таким образом, можно выбрать остатки FR из реципиентной и импортной последовательностей и объединить их, чтобы получить гетеромультимер с желательными характеристиками, такими как повышенная аффинность по отношению к антигену-мишени (антигенам-мишеням). Как правило, на связывание антигена непосредственно и наиболее сильно влияют остатки CDR.
III. Векторы, клетки-хозяева, и рекомбинантные методы
В рекомбинантном способе получения гетеромультимера по настоящему изобретению выделяют кодирующую его нуклеиновую кислоту и встраивают ее в реплицируемый вектор с целью последующего клонирования (амплификации ДНК) или экспрессии. ДНК, кодирующую гетеромультимер, можно легко выделить и секвенировать с помощью традиционных способов (например, с использованием олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи гетеромультимера). Существуют разные векторы. Выбор вектора отчасти зависит от используемой клетки-хозяина. Как правило, предпочтительные клетки-хозяева имеют либо прокариотическое или эукариотическое (чаще всего используют клетки млекопитающих, однако можно использовать и клетки грибов (например, дрожжей), насекомых, растений, а также содержащие ядра клетки других многоклеточных организмов) происхождение. Следует иметь в виду, что для этой цели можно использовать константные участки любого изотипа, в том числе константные участки IgG, IgM, IgA, IgD и IgE, и что такие константные участки можно получить из организма человека или животного любого вида.
А. Получение гетеромультимеров с использованием прокариотических клеток-хозяев.
i. Конструирование вектора
Полинуклеотидные последовательности, кодирующие полипептидные компоненты гетеромультимера по настоящему изобретению, можно получить с помощью стандартных рекомбинантных методов. Желательные полинуклеотидные последовательности можно выделить из антитело-продуцирующих клеток, таких как гибридомные клетки, и секвенировать. Альтернативно полинуклеотиды можно синтезировать с использованием нуклеотидного синтезатора или методов ПЦР. После получения последовательности, кодирующие полипептиды, вставляют в рекомбинантный вектор, способный реплицироваться и экспрессировать гетерологичные полинуклеотиды в прокариотических хозяевах. В целях настоящего изобретения можно использовать многочисленные доступные векторы, известные в данной области. Выбор соответствующего вектора зависит, в основном, от размера нуклеиновых кислот, которые нужно вставить в вектор, и конкретной клетки-хозяина, подлежащей трансформации вектором. Каждый вектор содержит разные компоненты, в зависимости от своей функции (амплификация или экспрессия гетерологичных полинуклеотидов, или то и другое) и его совместимости с конкретной клеткой-хозяином, в которой он находится. Компоненты вектора обычно включают, без ограничения: точку начала репликации, ген селективного маркера, промотор, участок связывания рибосомы (RBS), сигнальную последовательность, вставку гетерологичной нуклеиновой кислоты и последовательность терминации транскрипции.
Как правило, в сочетании с конкретной клеткой-хозяином используют плазмидные векторы, содержащие репликон и контрольные последовательности, полученные из видов, совместимых с указанной клеткой-хозяином. Вектор обычно несет участок репликации, а также маркирующие последовательности, облегчающие фенотипический отбор в трансформированных клетках. Например, E.coli обычно трансформируют плазмидой pBR322, полученной из разновидностей E.coli. pBR322 содержит гены, которые кодируют устойчивость к ампициллину (Amp) и тетрациклину (Tet), и, следовательно, облегчают идентификацию трансформированных клеток. pBR322, ее производные, или другие микробные плазмиды или бактериофаги также могут содержать, изначально или после модификации, промоторы, которые можно использовать в микроорганизмах, выбранных для экспрессии эндогенных белков. Примеры производных pBR322, используемых для экспрессии конкретных гетеромультимерных антител подробно описаны в Carter et al., патент США N 5648237.
Кроме того, в сочетании с конкретными организмами-хозяевами в качестве трансформирующих векторов можно использовать фаговые векторы, содержащие репликон и контрольные последовательности, совместимые с указанными микроорганизмами-хозяевами. Например, бактериофаг, такой как λGEM-11TM, можно использовать для получения рекомбинантного вектора, предназначенного для трансформации восприимчивых клеток-хозяев, таких как E.coli LE392.
Вектор экспрессии по настоящему изобретению может содержать две или более пары промотор-цистрон, кодирующие все полипептидные компоненты. Промотор представляет собой нетранслируемую регуляторную последовательность, расположенную выше (5ʹ) цистрона и способную модулировать его экспрессию. Прокариотические промоторы обычно делят на два класса, индуцируемые и конститутивные. Индуцируемый промотор представляет собой промотор, который инициирует повышенные уровни транскрипции цистрона под его контролем в ответ на изменение условий культивирования, например, наличие или отсутствие питательного вещества или изменение температуры.
Хорошо известно большое число промоторов, распознаваемых разными потенциальными клетками-хозяевами. Выбранный промотор можно функционально связать с цистроном ДНК, кодирующим легкую или тяжелую цепь, путем выделения промотора из источника ДНК посредством рестрикционного ферментативного расщепления и вставки выделенной последовательности промотора в вектор по настоящему изобретению. Для непосредственной амплификации и/или экспрессии целевых генов можно использовать как промотор с нативной последовательностью, так и разные гетерологичные промоторы. В некоторых вариантах осуществления используют гетерологичные промоторы, так как обычно они обеспечивают более высокий уровень транскрипции и более высокие выходы экспрессированного целевого гена по сравнению с нативным промотором полипептида-мишени.
Промоторы, подходящие для использования в прокариотических хозяевах, включают промотор PhoA, промоторные системы β-галактамазы и лактозы, промоторную систему триптофана (trp) и гибридные промоторы, такие как промоторы tac или trc. Однако можно использовать и другие промоторы, способные функционировать в бактериях (такие как другие известные бактериальные или фаговые промоторы). Их нуклеотидные последовательности опубликованы, что позволяет квалифицированным специалистам лигировать их с цистронами, кодирующими целевые легкие и тяжелые цепи (Siebenlist et al., (1980) Cell 20:269), используя линкеры или адаптеры, обеспечивающие необходимые участки рестрикции.
В одном из аспектов настоящего изобретения каждый цистрон в рекомбинантном векторе содержит компонент секреторной сигнальной последовательности, который управляет перемещением экспрессированных полипептидов через мембрану. Как правило, сигнальная последовательность может представлять собой компонент вектора, или она может быть частью вставленной в вектор ДНК, кодирующей целевой полипептид. Сигнальная последовательность, подходящая для целей настоящего изобретения, должна представлять собой последовательность, которая может распознаваться и подвергаться процессингу (т.е. расщепляется сигнальной пептидазой) в клетке-хозяине. В случае прокариотических клеток-хозяев, которые не способны распознавать и процессировать сигнальную последовательность, нативную по отношению к гетерологичному полипептиду, сигнальную последовательность заменяют прокариотической сигнальной последовательностью, например, выбранной из группы, включающей щелочную фосфатазу, пенициллиназу, Ipp или лидерные последовательности термостабильного энтеротоксина II (STII), LamB, Phoe, PelB, OmpA и MBP. В одном из вариантов осуществления изобретения сигнальные последовательности, используемые в обоих цистронах системы экспрессии, представляют собой сигнальные последовательности STII, или их варианты.
В другом аспекте продукцию гетеромультимеров по настоящему изобретению осуществляют в цитоплазме клетки-хозяина, и при этом присутствие секреторных сигнальных последовательностей в пределах каждого цистрона не требуется. В данном случае экспрессия, укладка и сборка легких и тяжелых цепей гетеромультимера с образованием функционального гетеромультимера происходит в цитоплазме. В цитоплазме некоторых штаммов-хозяев (таких как штаммы E. coli trxB) присутствуют условия, благоприятные для образования дисульфидных связей, и, следовательно, обеспечивающие правильную укладку и сборку экспрессированных белковых субъединиц (Proba и Pluckthun, Gene, 159:203 (1995)).
Прокариотические клетки-хозяева, подходящие для экспрессии гетеромультимеров по настоящему изобретению, включают Archaebacteria и Eubacteria, такие как грамотрицательные или грамположительные организмы. Примеры полезных бактерий включают Escherichia (например, E. coli), палочковидные бактерии (например, B. subtilis), энтеробактерии, виды Pseudomonas (например, P. aeruginosa), Salmonella Typhimurium, Serratia marcescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla или Paracoccus. В одном из вариантов осуществления используют грамотрицательные клетки. В одном из вариантов осуществления настоящего изобретения в качестве хозяев используют клетки E.coli. Примеры штаммов E.coli включают штамм W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp 1190-1219; номер депозита в АТСС 27325) и его производные, в том числе штамм 33D3, имеющий генотип W3110 ΔfhuA (∆tonA) ptr3 lac Iq lacL8 ∆ompT∆(nmpc-fepE) degP41 kanR (патент США № 5639635). Также можно использовать другие штаммы и их производные, такие как E. coli 294 (АТСС 31446), E.coli B, E.coli λ1776 (АТСС 31537) и E. coli RV308 (АТСС 31608). Эти примеры являются иллюстративными, но не ограничивающими. Способы конструирования производных вышеуказанных бактерий, имеющих определенные генотипы, известны в данной области и описаны, например, в Bass et al., Proteins 8:309-314 (1990). Как правило, при выборе подходящих бактерий учитывают способность репликона реплицироваться в клетках бактерии. Например, разновидности E. coli, Serratia или Salmonella можно использовать в качестве хозяев, если для доставки репликона используют хорошо известные плазмиды, такие как pBR322, pBR325, pACYC177 или pKN410. Обычно клетка-хозяин секретирует минимальные количества протеолитических ферментов и может потребоваться добавление к клеточной культуре дополнительных ингибиторов протеазы.
ii. Получение гетеромультимера
Клетки-хозяева трансформируют вышеописанными векторами экспрессии и культивируют в традиционных питательных средах, модифицированных соответствующим образом для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих желательные последовательности.
Трансформация означает введение ДНК в прокариотического хозяина таким образом, чтобы ДНК воспроизводилась, либо как внехромосомный элемент, либо как элемент, интегрированный в хромосому. В зависимости от используемой клетки-хозяина трансформацию проводят с использованием стандартных методов, соответствующих таким клеткам. Обработку кальцием с использованием хлорида кальция, как правило, проводят в случае применения бактериальных клеток, клеточные стенки которых представляют собой усиленные барьеры. В другом способе трансформации используют полиэтиленгликоль/ДМСО. Еще одним способом трансформации является электропорация.
Прокариотические клетки, используемые для получения гетеромультимеров по настоящему изобретению, выращивают в среде, известной в данной области и подходящей для культивирования выбранных клеток-хозяев. Примеры подходящих сред включают бульон Luria (LB), а также необходимые питательные добавки. В некоторых вариантах осуществления среда также содержит средство селекции, выбранное в зависимости от конструкции вектора экспрессии, и позволяющее селективно выращивать прокариотические клетки, содержащие вектор экспрессии. Например, в среду, используемую для выращивания клеток, экспрессирующих ген устойчивости к ампициллину, добавляют ампициллин.
Любые необходимые добавки, помимо источников углерода, азота и неорганического фосфата, также могут быть включены в состав среды в соответствующих концентрациях, по отдельности или в виде смесей с другими добавками или средами, такими как комплексный источник азота. Культуральная среда может необязательно содержать одно или несколько восстанавливающих средств, выбранных из группы, включающей глутатион, цистеин, цистамин, тиогликолят, дитиоэритритол и дитиотреитол.
Прокариотические клетки-хозяева культивируют при подходящих температурах. Например, температура, подходящая для выращивания E. Coli, предпочтительно, находится в диапазоне примерно от 20°C до 39°C, более предпочтительно, примерно от 25°C до 37°C, еще более предпочтительно, она составляет примерно 30°C. РН среды может иметь любое значение в диапазоне примерно от 5 до 9, и зависит, в основном, от организма-хозяина. E.coli выращивают при рН в диапазоне примерно от 6,8 до 7,4, более предпочтительно, при рН, составляющем примерно 7,0.
Если в векторе экспрессии по настоящему изобретению используют индуцируемый промотор, экспрессию белка индуцируют в условиях, подходящих для активации промотора. В одном из аспектов для контролирования транскрипции полипептидов по настоящему изобретению используют промоторы PhoA. Соответственно, чтобы обеспечить индукцию, трансформированные клетки-хозяева культивируют в ограниченной по фосфату среде. Предпочтительно, ограниченная по фосфату среда представляет собой среду C.R.A.P (см., например, Simmons et al., J. Immunol. Methods (2002), 263: 133-147). Как известно в данной области, в зависимости от конструкции используемого вектора можно использовать множество других индукторов.
В одном из вариантов осуществления экспрессированные гетеромультимеры по настоящему изобретению секретируются в периплазму клеток-хозяев, из которой их извлекают. Извлечение белка обычно включает разрушение микроорганизма, как правило, посредством осмотического шока, ультразвука или лизиса. После разрушения клеток клеточный дебрис или целые клетки можно удалить путем центрифугирования или фильтрации. Гетеромультимеры можно дополнительно очистить, например, путем аффинной хроматографии на смоле. Кроме того, белки могут транспортироваться в культуральную среду, из которой их можно выделить. Из культуры можно удалить клетки с получением культурального супернатанта, который фильтруют и концентрируют с целью дальнейшей очистки белков. Затем экспрессированные полипептиды можно выделить и идентифицировать с помощью хорошо известных методов, таких как электрофорез в полиакриламидном геле (PAGE) и вестерн-блоттинг.
В одном из аспектов настоящего изобретения получение гетеромультимера проводят в крупном масштабе с использованием процесса ферментации. Для получения рекомбинантных белков можно использовать разные крупномасштабные способы ферментации с применением подпитываемой культуры. Емкость крупномасштабных способов ферментации составляет по меньшей мере 1000 литров, предпочтительно, примерно от 1000 до 100000 литров. Такие ферментеры оборудуют турбинными мешалками, чтобы распределять кислород и питательные вещества, в особенности глюкозу (предпочтительный источник углерода/энергии). Термин мелкомасштабная ферментация, как правило, относится к ферментации в ферментере, объемная емкость которого составляет не более чем примерно 100 литров, и может варьировать примерно от 1 литра до 100 литров.
В процессе ферментации индукцию экспрессии белка обычно проводят после выращивания клеток в подходящих условиях с достижением желаемой плотности, например, соответствующей OD550 примерно 180-220, когда клетки находятся в ранней стационарной фазе. В зависимости от конструкции вектора можно использовать ряд индукторов, как известно в данной области техники и описано выше. Период выращивания клеток до индукции может быть более коротким. Как правило, клетки индуцируют в течение примерно 12-50 часов, хотя можно использовать более длинные или более короткие периоды индукции.
Чтобы улучшить выход и качество гетеромультимеров по настоящему изобретению, некоторые условия ферментации можно изменить. Например, чтобы улучшить надлежащие сборку и сворачивание секретируемых гетеромультимеров осуществляют совместную трансформацию прокариотических клеток-хозяев дополнительными векторами, обеспечивающими сверхэкспрессию белков-шаперонов, таких как белки Dsb (DsbA, DsbB, DsbC, DsbD и/или DsbG) или FkpA (пептидилпролил цис,транс-изомераза с шаперонной активностью). Показано, что белки-шапероны облегчают правильную укладку и растворимость гетерологичных белков, продуцируемых в бактериальных клетках-хозяевах (Chen et al., (1999) J. Biol. Chem. 274: 19601-19605; Georgiou et al., патент США No. 6083715; Georgiou et al., патент США No. 6027888; Bothmann and Pluckthun (2000) J. Biol. Chem. 275: 17100-17105; Ramm and Pluckthun, (2000) J. Biol. Chem. 275: 17106-17113; Arie et al., (2001) Mol. Microbiol. 39:199-210).
Чтобы минимизировать протеолиз экспрессированных гетерологичных белков (особенно чувствительных к протеолизу), для осуществления настоящего изобретения можно использовать некоторые штаммы-хозяева с дефицитом протеолитических ферментов. Например, штаммы клеток-хозяев можно модифицировать путем введения мутации (мутаций) в гены, кодирующие известные бактериальные протеазы, такие как протеаза III, OmpT, DegP, Tsp, протеаза I, протеаза Mi, протеаза V, протеаза VI, и их сочетания. Существует несколько штаммов E. coli с дефицитом протеазы, которые описаны, например, в Joly et al., (1998), Proc. Natl. Acad. Sci. USA 95:2773-2777; Georgiou et al., патенте США № 5264365; Georgiou et al., патенте США № 5508192; Hara et al., Microbial Drug Resistance, 2:63-72 (1996).
В одном из вариантов осуществления штаммы E.coli, дефицитные по протеолитическим ферментам и трансформированные плазмидами, обеспечивающими сверхэкспрессию одного или нескольких белков-шаперонов, используют в качестве клеток-хозяев в системе экспрессии по настоящему изобретению.
В другом варианте осуществления в клетке E.coli дополнительно экспрессируют эндопептидазу (например, фурин), чтобы отщепить один или несколько линкеров, присутствующих на представляющем интерес гетеромультимере, до очистки. В следующем варианте осуществления эукариотическая клетка-хозяин может экспрессировать эндопептидазу (например, фурин) и экзопептидазу (например, карбоксипептидазу B), где эндопептидаза отщепляет один или несколько линкеров от гетеромультимера, а экзопептидаза разрушает оставшиеся после действия эндопептидазы участки расщепления до очистки.
iii. Очистка гетеромультимера
Можно использовать стандартные методы очистки белков, известные в данной области. Следующие процедуры являются примерами подходящих способов очистки: фракционирование на иммуноаффинной или ионообменных колонках, осаждение этанолом, ВЭЖХ на обращенной фазе, хроматография на диоксиде кремния или на катионообменной смоле, такой как DEAE, хроматофокусирование, SDS-PAGE, использование колонок с гидрофобным взаимодействием (HIC), осаждение сульфатом аммония и гель-фильтрация с использованием, например, Sephadex G-75.
В одном из аспектов белок A, иммобилизованный на твердой фазе, используют для иммуноаффинной очистки полноразмерных гетеромультимерных продуктов по настоящему изобретению. Белок A представляет собой белок клеточной стенки Staphylococcus aureus размером 41 кДа, который связывается с высокой аффиностью с Fc-участком антитела. Lindmark et al., (1983) J. Immunol. Meth. 62: 1-13. Твердая фаза, на которой иммобилизуют белок A, предпочтительно, представляет собой колонку, содержащую поверхность из стекла или оксида кремния, более предпочтительно, колонку, содержащую стекло с контролируемым размером пор, или колонку с кремневой кислотой. В некоторых случаях колонку покрывают реагентом, таким как глицерин, чтобы предотвратить неспецифическую адгезию примесей.
На первой стадии очистки препарат, полученный из описанной выше культуры клеток, наносят на твердую фазу с иммобилизованным белком A, чтобы обеспечить специфическое связывание представляющего интерес гетеромультимера с белком А. Затем твердую фазу промывают, чтобы удалить неспецифически связанные с ней примеси. Представляющий интерес гетеромультимер можно извлечь из твердой фазы путем элюирования раствором, содержащим хаотропное средство или слабый детергент. Примеры хаотропных средств включают, без ограничения, мочевину, гуанидин-HCl, перхлорат лития, гистидин и аргинин. Примеры слабых детергентов включают, без ограничения, твин (например, твин-20), тритон (например, тритон X-100), NP-40 (нонилфеноксилполиэтоксилэтанол), Nonidet P-40 (октилфеноксилполиэтоксилэтанол) и додецилсульфат натрия (SDS). Разбавление гетеромультимера в растворе, содержащем хаотропное средство или слабый детергент, после элюирования с колонки (такой как колонка mAbSure), позволяет сохранить стабильность гетеромультимера после элюирования. В некоторых вариантах осуществления один или несколько участков расщепления, оставшихся после обработки гетеромультимеров по настоящему изобретению эндопептидазой, можно удалить после очистки путем добавления экзопептидаз (например, карбоксипептидазы В). В других вариантах осуществления один или несколько линкеров, присутствующих в гетеромультимерах по настоящему изобретению, можно расщепить и обработать эндопептидазами (например, фурином) и экзопептидазами (например, карбоксипептидазой В) после очистки вместо предварительной очистки.
В. Получение гетеромультимеров с использованием эукариотических клеток-хозяев
Как правило, векторы содержат, без ограничения, один или несколько из следующих компонентов: сигнальная последовательность, точка начала репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции.
i. Сигнальная последовательность
Вектор, предназначенный для применения в эукариотической клетке-хозяине, может содержать сигнальную последовательность или другой полипептид, содержащий специфический участок расщепления на N-конце зрелого белка или представляющего интерес полипептида. Выбранная гетерологичная сигнальная последовательность может представлять собой последовательность, способную распознаваться и процессироваться (например, расщепляться сигнальной пептидазой) в клетке-хозяине. При экспрессии в клетках млекопитающих можно использовать доступные сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например, сигнал gD простого герпеса. ДНК, содержащую такой участок-предшественник, лигируют в рамке считывания с ДНК, кодирующей антитело.
ii. Компонент, представляющий собой точку начала репликации
Как правило, наличие компонента, представляющего собой точку начала репликации, не является необходимым для векторов экспрессии, используемым в клетках млекопитающих. Например, обычно используют точку начала репликации SV40, но только потому, что оно содержит ранний промотор.
iii. Компонент, представляющий собой ген, облегчающий селекцию
Векторы экспрессии и клонирования могут содержать ген селекции, также называемый селектируемый маркер. Типичные гены селекции кодируют белки, которые (а) придают устойчивость к антибиотикам или другим токсинам, таким как ампициллин, неомицин, метотрексат или тетрациклин, (b) при необходимости пополняют ауксотрофные дефициты, или (c) обеспечивают необходимые питательные вещества, недоступные из сложных сред.
В одном из примеров схемы селекции используют средство, останавливающее рост клетки-хозяина. Клетки, успешно трансформированные гетерологичным геном, продуцируют белок, придающий устойчивость к лекарственному средству и, следовательно, выживают в условиях селекции. В примерах такой доминантной селекции используют лекарственные средства неомицин, микофеноловую кислоту и гигромицин.
Другие примеры селективных маркеров, подходящих для клеток млекопитающих, включают маркеры, которые позволяют идентифицировать клетки, содержащие нуклеиновую кислоту, кодирующую гетеромультимер, такие как гены DHFR, тимидинкиназы, металлотионеина-I и -II, предпочтительно, металлотионеина приматов, аденозиндезаминазы, орнитиндекарбоксилазы и т.д.
Например, вначале идентифицируют клетки, трансформированные геном селекции, таким как ген DHFR, путем культивирования всех трансформантов в культуральной среде, которая содержит метотрексат (Мtx), конкурентный антагонист DHFR. В случае применения DHFR дикого типа подходящей клеткой-хозяином является клетка яичника китайского хомячка (СНО), дефицитная по активности DHFR (например, АТСС CRL-9096).
Альтернативно селекцию клеток-хозяев (в частности, хозяев дикого типа, которые содержат эндогенный DHFR), трансформированных или со-трансформированных последовательностями ДНК, кодирующими гетеромультимер дикого типа, белок DHFR и другой селектируемый маркер, такой как аминогликозид-3ʹ-фосфотрансфераза (АРН), можно проводить путем выращивания клеток в среде, содержащей средство селекции, соответствующее селектируемому маркеру, такое как аминогликозидный антибиотик, например, канамицин, неомицин или G418. См., например, патент США № 4965199.
iv. Промоторный компонент
Векторы экспрессии и клонирования обычно содержат промотор, который распознается организмом-хозяином и функционально связан с последовательностью (последовательностями) нуклеиновой кислоты, кодирующей гетеромультимер. Известны промоторные последовательности эукариот. Практически все эукариотические гены имеют участок, обогащенный AT, расположенный на расстоянии примерно 25-30 оснований выше участка инициации транскрипции. Другая последовательность, расположенная на 70-80 оснований выше начала транскрипции многих генов, представляет собой участок CNCAAT (SEQ ID NO:12), где N может быть любым нуклеотидом. На 3ʹ-конце большинства эукариотических генов находится последовательность ААТААА (SEQ ID NO:13), которая может представлять собой сигнал добавления последовательности поли A к 3ʹ-концу кодирующей последовательности. Все указанные последовательности соответствующим образом вставляют в эукариотические векторы экспрессии.
Транскрипция векторов, кодирующих гетеромультимеры, в клетках-хозяевах млекопитающих может контролироваться, например, промоторами, полученными из геномов вирусов, таких как, например, вирус полиомы, вирус оспы кур, аденовирус (такой как аденовирус 2), вирус бычьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и обезьяний вирус 40 (SV40), гетерологичными промоторами млекопитающих, такими как промотор актина или промотор иммуноглобулина, или промоторами теплового шока, при условии, что такие промоторы совместимы с системами клеток-хозяев.
Ранние и поздние промоторы вируса SV40 можно легко получить в виде фрагмента рестрикции SV40, который также содержит начало репликации вируса SV40. Немедленный ранний промотор цитомегаловируса человека можно получить в виде фрагмента рестрикции HindIII E. Система экспрессии ДНК в хозяевах-млекопитающих с использованием бычьего папилломавируса в качестве вектора описана в патенте США № 4419446. Модификация данной системы описана в патенте США № 4601978. Альтернативно длинный концевой повтор вируса саркомы Рауса можно использовать в качестве промотора.
v. Компонент, представляющий собой энхансерный элемент
Транскрипцию ДНК, кодирующей гетеромультимерный полипептид (полипептиды), в высших эукариотах можно увеличить путем вставки в вектор энхансерной последовательности. В настоящее время известно много энхансерных последовательностей генов млекопитающих (таких как гены глобина, эластазы, альбумина, α-фетопротеина и инсулина). Кроме того, можно использовать энхансер из вируса эукариотических клеток. Примеры энхансеров включают энхансер SV40 на поздней стороне точки начала репликации (100-270 п.о.), энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней стороне точки начала репликации и энхансеры аденовируса. Описание элементов, увеличивающих активацию эукариотических промоторов, также приведено в Yaniv, Nature 297: 17-18 (1982). Энхансер можно вставить в вектор, присоединив его по 5ʹ- или 3ʹ-положению к последовательности, кодирующей гетеромультимерный полипептид, при условии, что достигается усиление, однако обычно его располагают на 5ʹ-участке промотора.
vi. Компонент, отвечающий за терминацию транскрипции
Кроме того, векторы экспрессии, используемые в эукариотических клетках-хозяевах, зачастую содержат последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Как правило, такие последовательности можно получить из 5ʹ- и иногда из 3ʹ-нетранслируемых участков эукариотических или вирусных ДНК или кДНК. Указанные участки содержат нуклеотидные сегменты, транскрибируемые в виде полиаденилированных фрагментов в нетранслируемой части мРНК, кодирующей гетеромультимер. Один из подходящих компонентов терминации транскрипции представляет собой участок полиаденилирования бычьего гормона роста. См. WO 94/11026 и раскрытый в нем вектор экспрессии.
vii. Выбор и трансформация клеток-хозяев
Клетки-хозяева, подходящие для клонирования или экспрессии ДНК в векторах по настоящему изобретению, включают клетки высших эукариот, описанные в настоящем документе, в том числе клетки-хозяева позвоночных. Размножение клеток позвоночных в культуре (культуре ткани) стало рутинной процедурой. Примеры подходящих линий клеток-хозяев млекопитающих включают линию клеток почки обезьяны CV1, трансформированных SV40 (COS-7, ATCC CRL 1651); линию клеток почки эмбриона человека (клетки 293 или 293, субклонированные для выращивания в суспензионной культуре, Graham et al., J. Gen. Virol. 36:59 (1977)); клетки почки детеныша хомячка (ВНК, АТСС CCL 10); клетки яичника китайского хомячка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки Сертоли мыши (TM4, Mather, Biol. Reprod. 23: 243-251 (1980)); клетки почки обезьяны (CV1 АТСС CCL 70); клетки почки зеленой африканской мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HeLa, АТСС CCL 2); клетки почки собаки (MDCK, АТСС CCL 34); клетки печени крысы buffalo (BRL 3A, АТСС CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; клетки FS-4; и линия клеток гепатомы человека (Нер G2).
Клетки-хозяева трансформируют описанными выше векторами экспрессии или клонирования с целью получения гетеромультимера и культивируют в обычных питательных средах, модифицированных соответствующим образом для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих целевые последовательности.
viii. Культивирование клеток-хозяев
Клетки-хозяева, используемые для получения гетеромультимера по настоящему изобретению, можно культивировать в разных средах. Для культивирования клеток-хозяев можно использовать коммерчески доступные среды, такие как среда Hamʹs F10 (Sigma), минимальная эссенциальная среда ((MEM), (Sigma), RPMI-1640 (Sigma) и модифицированная по Дульбекко среда Игла ((DMEM), Sigma). Кроме того, в качестве среды для культивирования клеток-хозяев можно использовать любую из сред, описанных в Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal. Biochem. 102: 255 (1980), патентах США №№ 4767704; 4657866; 4927762; 4560655; или 5122469; WO 90/03430; WO 87/00195; или в патенте США Re. 30985. Любую из указанных сред при необходимости можно дополнить гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, соли кальция, магния и фосфаты), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство гентамицинTM), микроэлементами (определяемыми как неорганические соединения, конечные концентрации которых обычно находятся в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. В состав сред также можно включить в соответствующих концентрациях любые другие необходимые добавки, известные специалистам в данной области. Для культивирования используют условия, такие как температура, рН и т.п., ранее используемые в применении к клетке-хозяину, выбранной для экспрессии, и хорошо известные специалистам в данной области.
ix. Очистка гетеромультимера
При использовании рекомбинантных способов гетеромультимер может продуцироваться внутри клетки, либо он может секретироваться непосредственно в среду. Если гетеромультимер продуцируется внутри клетки, на первой стадии дебрис, содержащий частицы, такие как клетки-хозяева или фрагменты, полученные в результате лизиса, удаляют, например, методом центрифугирования или ультрафильтрации. Если гетеромультимер секретируется в среду, супернатанты таких систем экспрессии, как правило, сначала концентрируют с использованием коммерчески доступного фильтра для концентрирования белков, например, с помощью устройства для ультрафильтрации Amicon или Millipore Pellicon. На любой из предыдущих стадий можно добавить ингибитор протеазы, такой как PMSF, чтобы подавить протеолиз, и антибиотики для предотвращения роста случайных загрязняющих организмов.
Полученную из клеток гетеромультимерсодержащую композицию можно очистить, например, методом хроматографии на гидроксилапатите, гель-электрофореза, диализа и аффинной хроматографии, причем аффинная хроматография является предпочтительным методом очистки. Возможность применения белка A в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, присутствующего в гетеромультимере. Белок A можно использовать для очистки гетеромультимеров, содержащих тяжелые цепи человека γ1, γ2 или γ4 (Lindmark et al., J. Immunol. Meth. 62: 1-13 (1983)). Белок G рекомендуется использовать для очистки всех изотипов мыши и γ3 человека (Guss et al., EMBO J. 5: 15671575 (1986)). В качестве матрицы, к которой присоединяют аффинный лиганд, наиболее часто используют агарозу, однако можно использовать и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, обеспечивают более высокую скорость потока и более короткое время обработки, чем агароза. Если гетеромультимер содержит домен СН3, для очистки используют смолу Bakerbond ABXTM (J.T. Baker, Phillipsburg, NJ). В зависимости от антитела, подлежащего извлечению, можно использовать и другие методы очистки белков, такие как фракционирование на ионообменной колонке, осаждение этанолом, ВЭЖХ на обращенной фазе, хроматография на диоксиде кремния, хроматография на гепаринсодержащей SEPHAROSETM, хроматография на анион- или катион-обменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония.
В одном из вариантов осуществления эукариотическая клетка-хозяин дополнительно экспрессирует эндопептидазу (например, фурин), которая может отщеплять один или несколько линкеров от представляющего интерес гетеромультимера перед очисткой (например, во время транспорта через аппарат Гольджи). В другом варианте осуществления эукариотическая клетка-хозяин может экспрессировать эндопептидазу (например, фурин или Lys-C) и экзопептидазу (например, карбоксипептидазу В), где эндопептидаза отщепляет один или несколько линкеров от гетеромультимера, а экзопептидаза разрушает участки, оставшиеся после расщепления эндопептидазой, перед очисткой.
Представляющий интерес гетеромультимер извлекают из твердой фазы колонки путем элюирования в раствор, содержащий хаотропное средство или мягкий детергент. Под "хаотропным средством" подразумевают водорастворимое вещество, способное разрушать трехмерную структуру белка (например, антитела), препятствуя стабилизации внутримолекулярных взаимодействий (таких как водородные связи, Ван-дер-Ваальсовы или гидрофобные взаимодействия). Примеры хаотропных средств включают, без ограничения, мочевину, гуанидин-HCl, перхлорат лития, гистидин и аргинин. Термин "мягкий детергент" обозначает водорастворимое вещество, которое разрушает трехмерную структуру белка или белкового комплекса (такого как гетеромультимер), препятствуя стабилизации внутримолекулярных взаимодействий (таких как водородные связи, Ван-дер-Ваальсовы или гидрофобные взаимодействия), но которое не вызывает необратимого разрушения белковой структуры, приводящего к потере биологической активности (то есть, которое не вызывает денатурации белка). Примеры мягких детергентов включают, без ограничения, твин (например, твин-20), тритон (например, тритон X-100), NP-40 (нонилфеноксилполиэтоксилэтанол), Nonidet P-40 (октилфеноксилполиэтоксилэтанол) и додецилсульфат натрия (SDS).
После любой предварительной стадии (стадий) очистки смесь, содержащую представляющий интерес гетеромультимер и загрязняющие вещества, можно подвергнуть гидрофобной хроматографии при низких значениях рН, используя для элюирования буфер, имеющий рН в диапазоне примерно 2,5-4,5, которую, предпочтительно, проводят при низкой концентрации соли (такой как примерно 0-0,25 М).
В некоторых вариантах осуществления один или несколько остаточных участков расщепления эндопептидазой в обработанных эндопептидазой гетеромультимерах по настоящему изобретению после очистки можно обработать и разрушить экзопептидазами (например, карбоксипептидазой В). В других вариантах осуществления один или несколько линкеров, входящих в состав гетеромультимеров по настоящему изобретению, можно расщепить и обработать эндопептидазой (такой как фурин) и экзопептидазой (такой как карбоксипептидаза В) после очистки, а не до очистки.
х. Получение гетеромультимера с использованием бакуловирусов
Рекомбинантный бакуловирус можно получить путем совместной трансфекции плазмиды, кодирующей гетеромультимер или фрагмент гетеромультимера, и ДНК вируса BaculoGoldTM (Pharmingen) в клетку насекомого, такую как клетка Spodoptera frugiperda (например, клетка Sf9, ATCC CRL 1711), или клетка Drosophila melanogaster S2 с использованием, например, липофектина (поставляется GIBCO-BRL). В конкретном примере последовательность гетеромультимера вставляют в бакуловирусный вектор экспрессии перед эпитопным маркером. Такие эпитопные маркеры включают маркеры поли-His. Можно использовать разные плазмиды, в том числе плазмиды, полученные из коммерчески доступных плазмид, таких как pVL1393 (Novagen) или pAcGP67B (Pharmingen). Коротко говоря, последовательность, кодирующую гетеромультимер-антитело, или ее фрагмент, можно амплифицировать методом ПЦР с использованием праймеров, комплементарных 5ʹ- и 3ʹ-участкам. 5ʹ-праймер может содержать фланкирующие (выбранные) участки рестрикции. Затем продукт можно расщепить выбранными ферментами рестрикции и субклонировать в векторе экспрессии.
После трансфекции вектором экспрессии клетки-хозяева (например, клетки Sf9) инкубируют в течение 4-5 дней при 28°С, высвобожденный вирус собирают и используют для последующих амплификаций. Инфицирование вирусом и экспрессию белка можно проводить по способу, описанному, например, в OʹReilley et al. (Baculovirus expression vectors: A Laboratory Manual. Oxford: Oxford University Press (1994)).
Затем экспрессированный поли-His-меченный гетеромультимер можно очистить, например, методом Ni2+-хелатирующей аффинной хроматографии следующим образом. Экстракты можно получить из клеток Sf9, инфицированных рекомбинантным вирусом, по способу, описанному Rupert et al. (Nature 362: 175-179 (1993)). Коротко говоря, клетки Sf9 промывают, ресуспендируют в буфере для обработки ультразвуком (25 мл HEPES рН 7,9; 12,5 мМ MgCl2, 0,1 мМ ЭДТА, 10% глицерина, 0,1% NP-40; 0,4 М КСl) и обрабатывают ультразвуком дважды в течение 20 секунд на льду. Обработанные ультразвуком образцы очищают центрифугированием, супернатант разбавляют в 50 раз буфером для загрузки (50 мМ фосфат; 300 мМ NaCl, 10% глицерина, рН 7,8) и фильтруют через фильтр с размером пор 0,45 мкм. Готовят колонку с Ni2+-NTA-агарозой (поставляемой Qiagen) с объемом слоя 5 мл, затем промывают ее 25 мл воды и уравновешивают 25 мл буфера для загрузки. Отфильтрованный экстракт клеток наносят на колонку со скоростью 0,5 мл в минуту. Колонку промывают до фонового уровня A280 буфером для загрузки, после чего начинают сбор фракций. Затем колонку промывают вторым буфером для промывания (50 мМ фосфат; 300 мМ NaCl, 10% глицерин, рН 6,0), который элюирует неспецифически связанный белок. После повторного достижения фонового уровня A280 колонку элюируют в градиенте имидазола от 0 до 500 мМ, растворенного во втором буфере для промывания. Собирают аликвоты фракций объемом один мл и анализируют их методом SDS-PAGE с окрашиванием серебром или Вестерн-блоттинга с использованием Ni2+-NTA, конъюгированного со щелочной фосфатазой (Qiagen). Фракции, содержащие элюированный His10-меченный (SEQ ID NO:21) гетеромультимер, объединяют и диализуют против буфера для загрузки.
Очистку гетеромультимера также можно проводить с использованием известных хроматографических методов, включающих, например, хроматографию на колонке с белком A или белком G. Представляющий интерес гетеромультимер можно извлечь из твердой фазы колонки путем элюирования в раствор, содержащий хаотропное средство или мягкий детергент. Примеры хаотропных средств и мягких детергентов включают, без ограничения, гуанидин-HCl, мочевину, перхлорат лития, аргинин, гистидин, SDS (додецилсульфат натрия), твин, тритон и NP-40, которые являются коммерчески доступными. Как описано выше, один или несколько линкеров, входящих в состав гетеромультимера, можно отщепить и обработать после очистки путем добавления эндопептидазы (такой как фурин или Lys-C) и экзопептидазы (такой как карбоксипептидаза В).
С. Метод очистки
Конкретный способ очистки, который можно использовать в применении к гетеромультимерам, содержащим HD линкер, описан ниже.
Содержащий линкеры гетеромультимер наносят на колонку с белком А (например, mAbSure) при 4°C
↓
Колонку промывают KPO4, затем PBS + 0,1% тритон X114
↓
Образец элюируют в Трис рН 8,0 (200 мМ)
↓
рН образца доводят до 8,0 и затем образец расщепляют Lys-C в течение 1,5 часов при 37°С
↓
Образец повторно очищают на колонке MabSURE
↓
рН образца доводят до 8,0 и затем образец расщепляют карбоксипептидазой В в течение 4 часов при 37°С
↓
Корректируют проводимость образца и наносят его на ионообменную колонку
↓
Фракции собирают, объединяют и диализуют против PBS
В описанном выше способе после начальной стадии очистки на колонке с белком A помимо аргинина можно использовать другие хаотропные средства или мягкие детергенты, которые включают, без ограничения, гуанидин-HCl, мочевину, перхлорат лития, гистидин, SDS (додецилсульфат натрия), твин, тритон и NP-40, которые являются коммерчески доступными. В некоторых вариантах осуществления один или несколько участков расщепления эндопептидазой, оставшихся в гетеромультимерах по настоящему изобретению после обработки эндопептидазой, можно обработать и разрушить после очистки с помощью экзопептидаз (например, карбоксипептидазы В). В других вариантах осуществления один или несколько линкеров, входящих в состав гетеромультимеров по настоящему изобретению, можно расщепить и обработать эндопептидазами (такими как фурин или Lys-C) и экзопептидазами (такими как карбоксипептидаза В) после очистки, а не до очистки.
IV. Конъюгированные белки
Настоящее изобретение также предлагает конъюгированные белки, такие как конъюгированные гетеромультимеры или иммуноконъюгаты (например, "конъюгаты антитело-лекарственное средство" или "ADC"), содержащие любой из описанных в настоящем описании гетеромультимеров (например, одноцепочечное моноспецифическое или мультиспецифическое антитело, содержащее HD линкер, многоцепочечный моноспецифический или мультиспецифический гетеромультимер, содержащий HD линкер), где один из константных участков легкой цепи или тяжелой цепи конъюгирован с химической молекулой, включающей краситель или цитотоксическое средство, такое как химиотерапевтическое средство, лекарственное средство, средство, ингибирующее рост, токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения, или его фрагменты) или радиоактивный изотоп (т.е. радиоконъюгат). В частности, как описано в настоящем документе, применение HD линкера позволяет получать гетеромультимеры, содержащие две разные тяжелые цепи (НС1 и НС2), а также две разные легкие цепи (LC1 и LC2). Иммуноконъюгат, сконструированный с использованием описанных в настоящем описании способов, может содержать цитотоксическое средство, конъюгированное с константным участком только одной из тяжелых цепей (НС1 или НС2) или только одной из легких цепей (LC1 или LC2). Кроме того, поскольку иммуноконъюгат может содержать цитотоксическое средство, присоединенное только к одной тяжелой или легкой цепи, количество цитотоксического средства, которое нужно вводить индивиду, уменьшается по сравнению с введением гетеромультимера, содержащих цитотоксическое средство на обеих тяжелых или легких цепях. Уменьшение количества цитотоксического средства, вводимого индивиду, снижает вредные побочные эффекты, связанные с применением цитотоксического средства.
Применение конъюгатов антитело-лекарственное средство для локальной доставки цитотоксических или цитостатических средств, то есть препаратов, уничтожающих или подавляющих опухолевые клетки, для лечения злокачественного новообразования (Syrigos and Epenetos, Anticancer Research 19:605-614 (1999); Niculescu-Duvaz and Springer, Adv. Drg. Del. Rev. 26: 151-172 (1997); патент США № 4975278) позволяет осуществлять направленную доставку фрагмента лекарственного средства к опухоли и внутриклеточное накопление в ней, где системное введение этих неконъюгированных лекарственных средств может привести к неприемлемым уровням токсичности в отношении нормальных клеток, а также опухолевых клеток, подлежащих устранению (Baldwin et al., Lancet (Mar. 15, 1986):603-605 (1986); Thorpe, (1985) "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review," in Monoclonal Antibodies ʹ84: Biological And Clinical Applications, A. Pinchera et al. (ed.s), pp. 475-506). Таким образом достигается максимальная эффективность при минимальной токсичности. Как описано, в указанных стартегиях можно использовать как поликлональные антитела, так и моноклональные антитела (Rowland et al., Cancer Immunol. Immunother. 21:183-187 (1986)). Лекарственные средства, используемые в описанных способах, включают дауномицин, доксорубицин, метотрексат и виндезин (Rowland et al., (1986), см. выше). Токсины, используемые для получения конъюгатов гетеромультимер-токсин, включают бактериальные токсины, такие как дифтерийный токсин, растительные токсины, такие как рицин, низкомолекулярные токсины, такие как гелданамицин (Mandler et al., Jour, of the Nat. Cancer Inst. 92(19): 1573-1581 (2000); Mandler et al., Bioorganic & Med. Chem. Letters 10:1025-1028 (2000); Mandler et al., Bioconjugate Chem. 13:786-791 (2002)), майтанзиноиды (EP 1391213; Liu et al., Proc. Natl. Acad. Sci. USA 93:8618-8623 (1996)) и калихеамицин (Lode et al., Cancer Res. 58:2928 (1998); Hinman et al., Cancer Res. 53:3336-3342 (1993)). Токсины могут оказывать цитотоксические и цитостатические эффекты посредством механизмов, включающих связывание тубулина, связывание ДНК или ингибирование топоизомеразы. Некоторые цитотоксические лекарственные средства, как правило, становятся неактивными или менее активными после конъюгирования с большими гетеромультимерными антителами или лигандами рецепторов белков.
Химиотерапевтические средства, которые можно использовать для получения иммуноконъюгатов, описаны в настоящем документе (например, выше). Подходящие для применения ферментативно активные токсины и их фрагменты включают А-цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, А-цепь экзотоксина (из Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки dianthin, белки Phytolaca Americana (PAP I, PAP II и PAP-S), ингибитор Momordica Charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. См., например, WO 93/21232, опубликованный 28 октября 1993 года. Ряд радионуклидов можно использовать для получения радиоконъюгированных антител. Их примеры включают 212Bi, 131I, 131In, 90Y и 186Re. Конъюгаты антитела и цитотоксического средства получают с использованием ряда бифункциональных белок-связывающих средств, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (например, диметиладипимидат НСl), активированные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получить по способу, описанному в Vitetta et al., Science 238: 1098 (1987). Углерод-14-меченая 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является примером хелатирующего средства, используемого для конъюгации радионуклида с антителом. См., например, WO94/11026.
Конъюгаты гетеромультимера с одним или несколькими низкомолекулярными токсинами, такими как калихеамицин, майтанзиноиды, доластатины, ауростатины, трихотецен и СС1065, а также производные указанных токсинов, которые обладают активностью токсина, также рассматриваются в настоящем описании.
А. Майтанзин и майтанзиноиды
В некоторых вариантах осуществления иммуноконъюгат содержит гетеромультимер по настоящему изобретению, конъюгированный с одной или несколькими молекулами майтанзиноидов.
Майтанзиноиды представляют собой ингибиторы митоза, которые действуют путем ингибирования полимеризации тубулина. Майтанзин был впервые выделен из восточно-африканского кустарника Maytenus serrata (патент США № 3896111). Впоследствии было обнаружено, что некоторые микроорганизмы также продуцируют майтанзиноиды, такие как майтанзинол и C-3 сложные эфиры майтанзинола (патент США № 4151042). Синтетический майтанзинол, его производные и аналоги описаны, например, в патентах США № 4137230; 4248870; 4256746; 4260608; 4265814; 4294757; 4307016; 4308268; 4,308,269; 4309428; 4313946; 4315929; 4317821; 4322348; 4331,598; 4361650; 4364866; 4424219; 4450254; 4362663 и 4371533.
Фрагменты майтанзиноидных лекарственных средств являются привлекательными с точки зрения получения конъюгатов гетеромультимерное антитело-лекарственное средство, поскольку они: (i) относительно доступны для получения путем ферментационной или химической модификации и дериватизации продуктов ферментации, (ii) поддаются дериватизации функциональными группами, подходящими для конъюгации с антителами посредством линкеров, отличных от дисульфидных, (iii) стабильны в плазме, и (iv) эффективны против разных опухолевых клеточных линий.
Иммуноконъюгаты, содержащие майтанзиноиды, способы их получения и их терапевтическое применение описаны, например, в патентах США № 5208020, 5416064 и в Европейском патенте ЕР 0425235 B1, описания которых специально включены в настоящий документ в качестве ссылки. Liu et al., Proc. Natl. Acad. Sci. USA 93: 8618-8623 (1996) описывают иммуноконъюгаты, содержащие мейтансиноид, обозначенный DM1, связанный с моноклональным антителом C242, направленным против колоректального рака человека. Обнаружено, что конъюгат является высоко токсичным по отношению к культуре клеток рака толстой кишки, и проявляет противоопухолевую активность в анализе роста опухоли in vivo. Chari et al., Cancer Research 52: 127-131 (1992) описывают иммуноконъюгаты, в которых майтанзиноид конъюгируют посредством дисульфидного линкера с мышиным антителом A7, способным связываться с антигеном на линии раковых клеток толстой кишки человека, или с другим мышиным моноклональным антителом TA.1, которое связывает онкоген HER-2/neu. Цитотоксичность конъюгата TA.1-майтанзиноид тестируют in vitro на линии клеток рака молочной железы человека SK-BR-3, экспрессирующих 3×105 поверхностных антигенов HER-2 на клетку. Конъюгат лекарственного средства достигает степени цитотоксичности, подобной цитотоксичности препарата свободного майтанзиноида, которую можно повысить путем увеличения количества молекул майтанзиноида на молекулу антитела. Конъюгат A7-майтанзиноид проявляет низкую системную цитотоксичность у мышей.
Конъюгаты майтанзиноида получают путем химического присоединения гетеромультимера к молекуле майтанзиноида без значительного снижения биологической активности гетеромультимера или молекулы майтанзиноида. См., например, патент США № 5208020 (описание которого специально включено в настоящий документ в качестве ссылки). Показано, что присоединение в среднем 3-4 молекул майтанзиноида к молекуле гетеромультимерного антитела приводит к повышению цитотоксичности в отношении клеток-мишеней, не оказывая негативного влияния на функцию или растворимость антитела, хотя можно ожидать, что присоединение даже одной молекулы токсина на молекулу антитела увеличивает цитотоксичность по сравнению с использованием “голого” антитела. Майтанзиноиды хорошо известны в данной области, их можно синтезировать с помощью известных способов или выделить из природных источников. Подходящие майтанзиноиды описаны, например, в патенте США № 5208020, а также в других патентах и непатентных публикациях, указанных выше. Предпочтительными майтанзиноидами являются майтанзинол и аналоги майтанзинола, модифицированные по ароматическому кольцу или по другим положениям молекулы майтанзинола, такие как разные сложные эфиры майтанзинола.
Существует много известных в данной области связывающих групп, которые можно использовать для получения конъюгатов майтанзиноидов, указанные группы включают, например, группы, описанные в патенте США № 5208020 или в патенте EP 0425235 B1, Chari et al., Cancer Research 52: 127-131 (1992), а также в заявке на патент США № 2005/0169933, описания которых специально включены в настоящий документ в качестве ссылки. Конъюгаты майтанзиноидов, содержащие линкерный компонент SMCC, можно получить по способу, описанному в публикации патентной заявки США № 2005/0169933. Связывающие группы включают дисульфидные группы, тиоэфирные группы, кислоточувствительные группы, фоточувствительные группы, пептидаза-чувствительные группы, или эстераза-чувствительные группы, описанные в вышеуказанных патентах, причем дисульфидные и тиоэфирные группы являются предпочтительными. Примеры других связывающих групп описаны в настоящем документе.
Конъюгаты гетеромультимера с майтанзиноидом можно получить с помощью ряда бифункциональных конденсирующих реагентов, используемых для синтеза белков, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил) циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат НСl), активированные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Особенно предпочтительные конденсирующие реагенты включают N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) (Carlsson et al., Biochem. J. 173: 723-737 (1978)) и N-сукцинимидил-4-(2-пиридилтио)пентановую кислоту (SPP), обеспечивающие образование дисульфидной связи.
Линкер можно присоединить к молекуле майтанзиноида в разных положениях, в зависимости от типа связи. Например, сложноэфирная связь может быть образована в результате взаимодействия с гидроксильной группой с использованием традиционных способов конденсации. Реакция может протекать по положению С-3, содержащему гидроксильную группу, положению C-14, модифицированному гидроксиметилом, положению С-15, модифицированному гидроксильной группой, и положению C-20, содержащему гидроксильную группу. В предпочтительном варианте осуществления связь образуется в положении С-3 майтанзинола или аналога майтанзинола.
B. Ауристатины и доластатины
В некоторых вариантах осуществления иммуноконъюгат содержит гетеромультимер по настоящему изобретению, конъюгированный с доластатинами или пептидными аналогами и производными доластатинов, ауристатинами (патенты США № 5635483 и 5780588). Показано, что доластатины и ауристатины препятствуют динамике микротрубочек, гидролизу ГТФ, а также делению ядер и клеток (Woyke et al., Antimicrob. Agents and Chemother. 45(12):3580-3584 (2001)), и обладают противоопухолевой (патент США № 5663149) и противогрибковой активностью (Pettit et al., Antimicrob. Agents Chemother. 42:2961-2965 (1998)). Лекарственный фрагмент, содержащий доластатин или ауристатин, можно присоединить к гетеромультимеру по N(амино)-концу или C(карбокси)-концу фрагмента пептидного лекарственного средства (WO 02/088172).
Примеры вариантов ауристатина включают присоединенные по N-концу монометилауристатиновые лекарственные фрагменты DE и DF, описанные в "Monomethylvaline Compounds Capable of Conjugation to Ligands," публикация заявки США № 2005/0238649, описание которой включено в настоящий документ в качестве ссылки в полном объеме.
Как правило, пептидные лекарственные фрагменты можно получить путем образования пептидной связи между двумя или более аминокислотами и/или пептидными фрагментами. Такие пептидные связи можно получить, например, методом жидкофазного синтеза (см. E. Schroder and K. Lubke, "The Peptides," volume 1, pp. 76-136, 1965, Academic Press), хорошо известного в области химии пептидов. Ауристатиновые/доластатиновые лекарственные фрагменты можно получить с помощью способов, описанных в: патентах США № 5635483 и 5780588; Pettit et al., J. Nat. Prod. 44:482-485 (1981); Pettit et al., Anti-Cancer Drug Design 13:47-66 (1998); Poncet, Curr. Pharm. Des. 5:139-162 (1999); and Pettit, Fortschr. Chem. Org. Naturst. 70:1-79 (1997). См. также Doronina, Nat. Biotechnol. 21(7):778-784 (2003); и "Monomethylvaline Compounds Capable of Conjugation to Ligands," публикация заявки США № 2005/0238649, включенная в настоящий документ в качестве ссылки в полном объеме (описывающая, например, линкеры и способы получения монометилвалиновых соединений, таких как MMAE и MMAF, конъюгированных с линкерами).
С. Калихеамицин
В других вариантах осуществления иммуноконъюгат содержит гетеромультимер по настоящему изобретению, конъюгированный с одной или несколькими молекулами калихеамицина. Антибиотики семейства калихеамицинов способны осуществлять двунитевые разрывы ДНК в субпикомолярных концентрациях. Способы получения конъюгатов, содержащих члены семейства калихеамицина описаны в патентах США № 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296 (все они принадлежат American Cyanamid Company). Структурные аналоги калихеамицина, подходящие для применения, включают, без ограничения, γ1I, α2I, α3I, N-ацетил-γ1I, PSAG и θI1 (Hinman et al., Cancer Research 53:3336-3342 (1993), Lode et al., Cancer Research 58:2925-2928 (1998), а также указанные выше патенты США, принадлежащие American Cyanamid). Другим противоопухолевым средством, с которым можно конъюгировать антитело, является QFA, который представляет собой антифолат. И калихеамицин, и QFA действуют внутри клетки и с трудом проходят через плазматическую мембрану. Следовательно, поглощение данных средств клетками путем антитело-опосредованной интернализации значительно усиливает их цитотоксическое действие.
D. Другие цитотоксические средства
Другие противоопухолевые средства, которые можно конъюгировать с гетеромультимерами по настоящему изобретению, или получать с помощью описанных в настоящем описании способов, включают BCNU, стрептозоцин, винкристин и 5-фторурацил, семейство средств, известных под общим названием комплекс LL-E33288, описанное в патентах США № 5053394 и 5770710, а также эсперамицины (патент США № 5877296).
Подходящие для применения ферментативно активные токсины и их фрагменты включают А-цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, А-цепь экзотоксина (из Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolaca americana (PAPI, PapII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены (см., например, WO 93/21232, опубликованной 28 октября 1993).
Настоящее изобретение также предлагает иммуноконъюгат, образованный гетеромультимером и соединением с нуклеолитической активностью (таким как рибонуклеаза или ДНК-эндонуклеаза, например, дезоксирибонуклеаза; ДНКаза).
С целью селективного разрушения опухоли в гетеромультимер можно ввести высокорадиоактивный атом. Для получения радиоконъюгированных гетеромультимеров можно использовать ряд радиоактивных изотопов. Их примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Если конъюгат используют для детекции, он может содержать радиоактивный атом для сцинтиграфических исследований, например Tc99m или I123, или спиновую метку для визуализации результатов ядерного магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), такую как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радиоактивные или другие метки можно ввести в состав конъюгата известными способами. Например, биосинтез пептида или химический синтез пептида из аминокислот проводят с использованием предшественников аминокислот, содержащих, например, фтор-19 вместо водорода. Такие метки, как Tc99m или I123, Re186, Re188 и In111, можно ввести в состав пептида через остаток цистеина. Иттрий-90 можно ввести через остаток лизина. Для включения йода-123 можно использовать способ IODOGEN (Fraker et al., Biochem. Biophys. Res. Commun. 80:49-57 (1978)). Другие способы подробно описаны в "Monoclonal Antibodies in Immunoscintigraphy" (Chatal, CRC Press 1989).
Конъюгаты гетеромультимера и цитотоксического средства можно получить с использованием ряда бифункциональных средств, связывающих белки, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат НСl), активированные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получить по способу, описанному Vitetta et al., Science 238: 1098 (1987). Углерод-14-меченная 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является примером хелатирующего средства, используемого для конъюгации радионуклида с гетеромультимером. См., например, WО94/11026. Линкер может представлять собой "расщепляемый линкер", облегчающий высвобождение цитотоксического лекарственного средства в клетке. Например, можно использовать кислоточувствительный линкер, пептидаза-чувствительный линкер, фоточувствительный линкер, диметиловый линкер или дисульфидсодержащий линкер (Chari et al., Cancer Research 52: 127-131 (1992); патент США № 5208020).
Соединения по настоящему изобретению специально включают, без ограничения, ADC, полученные с использованием поперечно-сшивающих реагентов: BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC, и сульфо-SMPB, а также SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Rockford, IL., США). См. стр. 467-498, 2003-2004, руководство по применению и каталог.
E. Получение конъюгированных гетеромультимеров
Конъюгированные гетеромультимеры по настоящему изобретению содержат гетеромультимер, конъюгированный с одним или несколькими фрагментами (например, фрагментами молекул лекарственного средства), например, с 1-20 фрагментами на антитело, необязательно присоединенными через линкер. Конъюгированные гетеромультимеры можно получить с помощью нескольких способов, с использованием реакций органической химии, условий и реагентов, известных специалистам в данной области, которые включают: (1) взаимодействие нуклеофильной группы гетеромультимерного антитела с двухвалентным связующим реагентом с образованием ковалентной связи, с последующей реакцией с представляющим интерес фрагментом; и (2) взаимодействие нуклеофильной группы фрагмента с двухвалентным связующим реагентом с образованием ковалентной связи, с последующей реакцией с нуклеофильной группой гетеромультимерного антитела. В настоящем документе описаны и другие способы получения конъюгированных гетеромультимеров.
Связующий реагент может состоять из одного или нескольких линкерных компонентов. Типичные линкерные компоненты включают 6-малеимидокапроил ("МС"), малеимидопропаноил ("MP"), валин-цитруллин ("val-cit"), аланин-фенилаланин ("аla-phe"), п-аминобензилоксикарбонил ("PAB"), N-сукцинимидил 4-(2-пиридилтио)пентаноат ("SPP"), N-сукцинимидил 4-(N-малеимидометил)циклогексан-1-карбоксилат ("SMCCʹ) и N-сукцинимидил (4-иодацетил)аминобензоат ("SIAB"). Другие линкерные компоненты известны в данной области и некоторые из них описаны в настоящем документе. См. также "Monomethylvaline Compounds Capable of Conjugation to Ligands," публикация заявки США № 2005/0238649, содержание которой включено в настоящий документ в качестве ссылки во всей полноте.
В некоторых вариантах осуществления линкер может содержать аминокислотные остатки. Типичные аминокислотные линкерные компоненты включают дипептид, трипептид, тетрапептид или пентапептид. Примеры дипептидов включают: валин-цитруллин (vc или val-cit), аланин-фенилаланин (af или ala-phe). Примеры трипептидов включают: глицин-валин-цитруллин (gly-val-cit) и глицин-глицин-глицин (gly-gly-gly). Аминокислотные остатки, которые содержат аминокислотный линкерный компонент, включают остатки, встречающиеся в природе, а также минорные аминокислоты и неприродные аналоги аминокислот, такие как цитруллин. Аминокислотные линкерные компоненты можно сконструировать и оптимизировать по селективности в отношении ферментативного расщепления под действием конкретных ферментов, таких как опухоль-связанная протеаза, катепсин B, C и D, или плазминовая протеазы.
Нуклеофильные группы, присутствующие на гетеромультимерах по настоящему изобретению, включают, без ограничения: (i) N-концевые аминогруппы, (ii) аминогруппы боковых цепей, например, боковой цепи лизина, (iii) тиольные группы боковых цепей, например, боковой цепи цистеина и (iv) гидроксильные группы или аминогруппы сахаров, если гетеромультимер является гликозилированным. Аминогруппы, тиольные группы и гидроксильные группы являются нуклеофильными и могут вступать в реакцию с электрофильными группами, присутствующими на линкерных фрагментах и линкерных реагентах, с образованием ковалентных связей, где электрофильные группы включают: (i) активированные сложные эфиры, такие как сложные эфиры NHS, эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидные группы. Некоторые гетеромультимеры содержат восстанавливаемые межцепочечные дисульфидные связи, т.е. цистеиновые мостики. Гетеромультимеры по настоящему изобретению можно активировать перед присоединением линкерных реагентов путем обработки восстановителем, таким как DTT (дитиотреитол). Следовательно, каждый цистеиновый мостик теоретически может образовать две реакционноспособные тиольные нуклеофильные группы. Другие нуклеофильные группы можно ввести в состав антител путем взаимодействия остатков лизина с 2-иминотиоланом (реагент Траута) с превращением амина в тиол. Реакционноспособные тиольные группы можно ввести в состав гетеромультимера (или его фрагмента) путем введения одного, двух, трех, четырех или более остатков цистеина (например, путем получения мутантных гетеромультимеров, содержащих один или несколько неприродных остатков цистеина).
Конъюгированные гетеромультимеры по настоящему изобретению также можно получить путем модификации гетеромультимера путем введения электрофильных групп, которые могут реагировать с нуклеофильными заместителями, присутствующими в линкерном реагенте или лекарственном или другом фрагменте. Сахара гликозилированных гетеромультимеров по настоящему изобретению можно окислить, например, периодатными окисляющими реагентами с получением альдегидных или кетоновых групп, которые могут взаимодействовать с аминогруппой линкерных реагентов или лекарственных или других фрагментов. Полученные в результате иминогруппы, представляющие собой основания Шиффа, могут образовывать стабильную связь, или их можно восстановить, например, боргидридсодержащими реагентами с образованием стабильных аминовых связей. В одном из вариантов осуществления взаимодействие углеводного фрагмента гликозилированного антитела с галактозооксидазой или метапериодатом натрия может приводить к образованию карбонильных (альдегидных и кетоновых) групп в белке, которые могут взаимодействовать с соответствующими группами лекарственного или другого фрагмента (Hermanson, Bioconjugate Techniques). В другом варианте осуществления белки, содержащие N-концевые остатки серина или треонина, могут взаимодействовать с метапериодатом натрия с образованием альдегида вместо первой аминокислоты (Geoghegan and Stroh, Bioconjugate Chem. 3: 138-146 (1992); патент США № 5362852). Такой альдегид может взаимодействовать с нуклеофилом лекарственного средства или линкера.
Подобным образом, нуклеофильные группы, присутствующие на фрагменте (например, на фрагменте лекарственного средства) включают, без ограничения, такие группы, как: амин, тиол, гидроксил, гидразид, оксим, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид, способные вступать в реакцию с образованием ковалентных связей с электрофильными группами на линкерных фрагментах и линкерных реагентах, включающими: (i) активированные сложные эфиры, такие как эфиры NHS, эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; и (iii) альдегидные, кетоновые, карбоксильные и малеимидные группы.
Альтернативно, слитый белок, содержащий гетеромультимер и цитотоксическое средство, можно получить, например, с помощью рекомбинантных способов или пептидного синтеза. Последовательность ДНК может содержать соответствующие участки, кодирующие две части конъюгата, либо примыкающие друг к другу, либо разделенные участком, кодирующим линкерный пептид, который не оказывает вредного воздействия на желательные свойства конъюгата. В следующем варианте осуществления гетеромультимер можно конъюгировать с "рецептором" (таким как стрептавидин) с целью предварительной направленности на опухоль, где конъюгат гетеромультимер-рецептор вводят индивиду с последующим удалением несвязанного конъюгата из кровотока с помощью очищающего реагента, и затем вводят "лиганд" (например, авидин), конъюгированный с цитотоксическим средством (таким как радионуклид).
V. Терапевтическое применение
Описанные в настоящем описании гетеромультимеры (например, HD линкерсодержащее одноцепочечное моноспецифическое или мультиспецифическое антитело, HD линкерсодержащее моноспецифическое или мультиспецифическое антитело, комплекс иммуноадгезин-антитело) можно использовать для терапевтического применения. Например, такие гетеромультимеры можно использовать для лечения любого заболевания, характеризующегося наличием подходящей мишени-кандидата для получения гетеромультимера, включающего пролиферативные заболевания, злокачественные новообразования, ангиогенные нарушения, воспалительные заболевания, аутоиммунные заболевания и расстройства иммунной системы.
Помимо терапевтического применения, гетеромультимеры по настоящему изобретению можно использовать для других целей, включающих диагностические методы, такие как методы диагностики любого заболевания или состояния, которое характеризуется наличием подходящей мишени-кандидата для генерирования антител, включающего указанные выше заболевания или состояния.
VI. Дозы, композиции и длительность лечения
Гетеромультимеры по настоящему изобретению можно использовать для получения композиций, дозировать и вводить с помощью способов, согласующихся с успешной медицинской практикой. Факторы, учитывающиеся в данном контексте, включают конкретное расстройства, подлежащее лечению, вид млекопитающего, подлежащего лечению, клиническое состояние конкретного индивида, причину расстройства, место доставки средства, способ введения, схему введения и другие факторы, известные практикующим врачам. "Терапевтически эффективное количество" вводимого белка определяется указанными факторами, и представляет собой минимальное количество, необходимое для предотвращения, облегчения или лечения конкретного расстройства (такого как пролиферативное заболевание, рак, ангиогенное расстройство, воспалительное расстройство, аутоиммунное заболевание или расстройство иммунной системы). Композиции на основе белков необязательно содержат одно или несколько средств, используемых в настоящее время для профилактики или лечения расстройства. Эффективное количество указанных других средств зависит от количества белков, присутствующих в композиции, типа заболевания или способа лечения и других факторов, описанных выше. Как правило, их вводят в дозах, описанных в настоящем документе, или составляющих примерно от 1 до 99% от ранее используемых доз.
В одном из вариантов осуществления настоящее изобретение можно использовать для увеличения продолжительности жизни человека, предрасположенного к конкретному расстройству, или у которого диагностировано конкретное расстройство, характеризующееся наличием подходящей мишени-кандидата для генерирования гетеромультимера. Продолжительность жизни определяется как время от первого введения препарата до смерти. Продолжительность жизни также можно измерить путем расслаивания отношения рисков (HR) в группе, получающей лечение, по сравнению с контрольной группой, в которой существует риск смерти индивида во время лечения.
В следующем варианте осуществления способы по настоящему изобретению значительно увеличивают процент ответивших в группе людей, предрасположенных к расстройству, или у которых диагностировано расстройство, и которых лечат с помощью одного или нескольких способов лечения данного расстройства. Процент ответивших определяют как процент ответивших на лечение среди индивидов, получающих лечение. В одном из аспектов комбинированное лечение по настоящему изобретению с использованием белков по настоящему изобретению и хирургии или другого способа лечения (такого как лучевая терапия или химиотерапия) значительно повышает процент ответивших в группе индивидов, получающих лечение, по сравнению с группой, получающей хирургическое лечение или другую форму монотерапии, причем увеличение соответствует величине р менее 0,005 по статистике хи-квадрат. Дополнительные способы измерения терапевтической эффективности при лечении раковых заболеваний описаны в публикации патентной заявки США № 20050186208.
Терапевтические композиции получают с помощью стандартных методов, известных в данной области, путем смешивания активного ингредиента, имеющего желательную степень чистоты, с необязательными физиологически приемлемыми носителями, наполнителями или стабилизаторами (Remingtonʹs Pharmaceutical Sciences (20th edition), ed. A. Gennaro, 2000, Lippincott, Williams & Wilkins, Philadelphia, PA). Приемлемые носители включают физиологический раствор или буферы, например, содержащие фосфат, цитрат и соли других органических кислот; антиоксиданты, в том числе аскорбиновая кислота; низкомолекулярные (содержащие приблизительно менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон, аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстрины; хелатирующие средства, такие как ЭДТА; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как TWEENTM, PLURONICSTM или ПЭГ.
Необязательно, но предпочтительно, композиция содержит фармацевтически приемлемую соль, предпочтительно, хлорид натрия, и, предпочтительно, приблизительно в физиологической концентрации. Композиции по настоящему изобретению необязательно содержат фармацевтически приемлемый консервант. В некоторых вариантах осуществления концентрации консерванта составляет от 0,1 до 2,0%, как правило, объемных. Подходящие консерванты включают консерванты, известные в области фармацевтики. Предпочтительными консервантами являются бензиловый спирт, фенол, м-крезол, метилпарабен и пропилпарабен. Композиции по настоящему изобретению дополнительно содержат фармацевтически приемлемое поверхностно-активное вещество в концентрации от 0,005 до 0,02%.
Описанная в настоящем описании композиция также может содержать несколько активных соединений, если это необходимо для лечения конкретного состояния, предпочтительно, соединения, активности которых дополняют друг друга и не оказывают друг на друга неблагоприятного влияния. Такие молекулы могут присутствовать в сочетании в количествах, которые являются эффективными для предполагаемой цели.
Активные ингредиенты также могут быть заключены в микрокапсулы, полученные, например, методом коацервации или путем межфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли(метилметакрилатные) микрокапсулы, соответственно, в коллоидные системы доставки лекарственного средства (такие как липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы описаны в Remingtonʹs Pharmaceutical Sciences, выше.
Можно получить препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие гетеромультимер, причем матрицы могут находиться в виде формованных изделий, например, пленок или микрокапсул. Примеры матриц, обеспечивающих замедленное высвобождение, включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США 3773919), сополимеры L-глутаминовой кислоты и этил-γ-L-глутамата, неразлагаемый этиленвинилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и лейпролида ацетата) и поли-D-(-)-3-гидроксимасляную кислоту. Тогда как такие полимеры, как этиленвинилацетат и молочная кислота-гликолевая кислота, способны высвобождать молекулы в течение более 100 дней, некоторые гидрогели высвобождают белки в течение более коротких периодов времени. Когда инкапсулированные гетеромультимеры изобретения остаются в организме в течение длительного времени, они могут денатурировать или агрегировать в результате воздействия влаги при 37°С, что приводит к потере биологической активности и возможным изменениям иммуногенности. Можно разработать рациональные стратегии стабилизации в зависимости от механизма действия. Например, если механизм агрегации включает образование межмолекулярной S-S связи посредством тиодисульфидного обмена, то стабилизацию можно осуществлять путем модификации сульфгидрильных остатков, лиофилизации из кислых растворов, контролирования содержания влаги, использования соответствующих добавок и разработки конкретных композиций полимерных матриц.
Описанные в настоящем описании белки (например, HD линкерсодержащее одноцепочечное моноспецифическое или мультиспецифическое антитело, HD линкерсодержащий многоцепочечный моноспецифический или мультиспецифический гетеромультимер, комплекс иммуноадгезин-антитело), вводят человеку с помощью известных способов, таких как внутривенное введение в виде болюса или непрерывной инфузии в течение определенного периода времени, внутримышечное введение, внутрибрюшинное введение, интрацерброспинальное введение, подкожное введение, внутрисуставное введение, интрасиновиальное введение, интратекальное введение, пероральное введение, местное введение или введение ингаляцией. Местное введение может быть особенно желательным, если обширные побочные эффекты или токсичность связаны с антагонизмом к целевой молекуле, распознаваемой белком. Стратегию ex vivo также можно использовать для терапевтического применения. Стратегия ex vivo включает трансфекцию или трансдукцию клеток, полученных от индивида, полинуклеотидом, кодирующим гетеромультимер по настоящему изобретению. Затем трансфицированные или трансдуцированные клетки возвращают индивиду. Клетки могут относиться к любому из широкого диапазона видов, включающего, без ограничения, гемопоэтические клетки (например, клетки костного мозга, макрофаги, моноциты, дендритные клетки, Т-клетки или В-клетки), фибробласты, эпителиальные клетки, эндотелиальные клетки, кератиноциты или мышечные клетки.
В одном из примеров гетеромультимер по настоящему изобретению вводят местно, например, путем непосредственной инъекции, если расстройство или местонахождение опухоли позволяют это, причем инъекции можно периодически повторять. Белковый комплекс также можно доставлять индивиду системно или непосредственно вводить его в опухолевые клетки, например, его можно вводить в опухоль или ложе опухоли после хирургического удаления опухоли, чтобы предотвратить или уменьшить развитие местного рецидива или метастазирования.
VII. Промышленные изделия
Другой вариант осуществления изобретения включает промышленное изделие, содержащее один или несколько из описанных в настоящем описании гетеромультимеров, а также вещества, используемые для лечения или диагностики любого расстройства, характеризующегося наличием подходящей мишени для генерирования гетеромультимера. Изделие содержит контейнер и этикетку или листок-вкладыш, находящиеся на контейнере, или прикрепленные к контейнеру. Подходящие контейнеры включают, например, бутыли, флаконы, шприцы и т.д. Контейнеры могут быть изготовлены из разных материалов, таких как стекло или пластик. Контейнер содержит композицию, которая является эффективной для лечения состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенного раствора или флакон, имеющий пробку, прокалываемую иглой для подкожной инъекции). По меньшей мере одно активное средство в композиции представляет собой гетеромультимер по настоящему изобретению. На этикетке или вкладыше в упаковку указано, что композиция предназначена для лечения конкретного состояния. Этикетка или вкладыш в упаковку необязательно содержит инструкцию по введению композиции гетеромультимера индивиду. Изобретение также охватывает промышленные изделия и наборы для описанных в настоящем описании комбинированных методов лечения.
Вкладыш к упаковке относится к инструкциям, обычно включаемым в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, применению, дозировках, введении, противопоказаниях, и/или предупреждения, касающиеся использования таких терапевтических продуктов.
Кроме того, промышленное изделие может дополнительно содержать второй контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Он может дополнительно содержать другие вещества, необходимые с коммерческой и потребительской точки зрения, которые включают другие буферы, разбавители, фильтры, иглы и шприцы.
Изобретение также предлагает наборы, которые можно использовать для разных целей, например, для очистки антигена из клеток или иммунопреципитации антигена. Набор для выделения и очистки антигена может содержать гетеромультимер, соединенный с гранулами (например, сефарозы). Изобретение может предоставлять наборы, которые содержат гетеромультимеры для детекции и количественного определения антигена in vitro, например, методом ELISA или вестерн-блоттинга. Как и промышленное изделие, набор содержит контейнер и этикетку или листок-вкладыш, находящийся на контейнере, или прикрепленный к контейнеру. Контейнер содержит композицию, содержащую по меньшей мере один гетеромультимер по настоящему изобретению. В состав набора могут быть включены дополнительные контейнеры, которые содержат, например, разбавители и буферы, или контрольные гетеромультимеры. Этикетка или вкладыш в упаковку может содержать описание композиции, а также инструкции по предполагаемому применению in vitro, или по диагностическому применению.
VIII. Молекулы-мишени
Примеры молекул, на которые может быть направлен гетеромультимер (такой как одноцепочечное или многоцепочечное антитело, или комплекс иммуноадгезин-антитело) по настоящему изобретению, включают, без ограничения, растворимые сывороточные белки, рецепторные белки, связанные с мембранами белки (например, адгезины), цитокины, хемокины, факторы роста и гормоны.
Растворимые антигены или их фрагменты, необязательно конъюгированные с другими молекулами, можно использовать в качестве иммуногенов для получения гетеромультимеров. В случае трансмембранных молекул, таких как рецепторы, в качестве иммуногенов можно использовать их фрагменты (такие как внеклеточный домен рецептора). Альтернативно в качестве иммуногена можно использовать клетки, экспрессирующие трансмембранную молекулу. Такие клетки могут быть получены из природного источника (например, из линий раковых клеток) или могут представлять собой клетки, трансформированные рекомбинантными способами с целью экспрессии трансмембранной молекулы. Другие антигены и их формы, подходящие для получения гетеромультимеров, являются очевидными для специалистов в данной области.
Вышеприведенные письменное описание считается достаточным для того, чтобы специалист в данной области мог осуществить изобретение. Нижеследующие примеры предлагаются только с целью иллюстрации и не предназначены для ограничения каким-либо образом объема настоящего изобретения. Действительно, разные модификации изобретения, помимо показанных и описанных в настоящем документе, которые станут очевидными для специалистов в данной области из предшествующего описания, попадают в объем прилагаемой формулы изобретения. Описание, представленное в примерах, предлагается рассматривать в целом как часть изобретения.
Примеры
Пример 1. Конструирование векторов для экспрессии гетеромультимеров, содержащих HD линкер
Все HD линкерсодержащие гетеромультимерные конструкции одноцепочечных антител получают из существующих конструкций моноклональных антител IgG1 собственного производства. LC получают методом ПЦР с использованием 3ʹ-праймера, который простирается за пределы С-концевого участка LC, кодируя N-концевой участок CLH-линкера и заканчиваясь в участке BamHI. 5ʹ-праймер LC заканчивается в участке ClaI. Затем ПЦР-фрагмент расщепляют и клонируют в расщепленном аналогичным образом и дефосфорилированном векторе pRК (Genentech Inc.; Eaton et al., Biochemistry 25:8343-8347 (1986)). Затем методом ПЦР получают родственную HC с использованием 5ʹ-праймера, который простирается за пределы N-концевого участка HC (минус сигнальная последовательность), кодируя C-концевой участок CLH-линкера и заканчиваясь в участке BamHI. 3ʹ-праймер HC заканчивается в участке XbaI. Полученный методом ПЦР фрагмент HC расщепляют и клонируют в полученной аналогичным образом плазмиде, содержащей описанную выше родственную LC, с получением половины антитела, содержащей линкер. Участок BamHI располагают так, чтобы он кодировал аминокислотные остатки "GS", сохраняя целостность повторов GGS в линкере. Затем остаток лизина в положении K222 (система нумерации Kabat) заменяют на остаток аланина, используя набор для направленного мутагенеза QuikChange II XL, Stratagene.
Затем две вышеописанные половины антитела, содержащие CLH линкер, соединяют посредством следующего цикла ПЦР, где половину антитела, предназначенную для N-конца одноцепочечного антитела, амплифицируют с использованием такого же 5ʹ-праймера LC, как и в предыдущем цикле, и 3ʹ-праймер, который простирается за пределы С-концевого участка НС, кодируя N-концевой участок HD линкера и заканчиваясь в участке BspEI. Фрагмент ПЦР расщепляют и клонируют в расщепленном аналогичным образом и дефосфорилированном векторе pRK. Половину антитела, предназначенную для C-конца гетеромультимерного одноцепочечного антитела, амплифицируют с использованием такого же 3ʹ-праймера HC, как и при конструировании половин антитела, и 5ʹ-праймера, который простирается за пределы N-конца LC (минус сигнальная последовательность), кодируя С-концевой участок HD линкера и заканчиваясь в участке BspEI. Продукт ПЦР расщепляют и клонируют в расщепленной аналогичным образом и дефосфорилированной N-концевой конструкции вектора. Как и BamHI, BspEI можно расположить так, чтобы он поддерживал целостность повторов GGS в линкере.
Конструирование HD линкерсодержащих гетеромультимерных многоцепочечных антител проводят подобным образом, с единственным отличием, которое заключается в том, что N-конец "половины антитела" начинается с сигнальной последовательности HC, а родственную ей LC экспрессируют отдельно из второй плазмиды pRK.
Пример 2. Расщепление и очистка гетеромультимеров, содержащих HD линкер
Пример схемы, которую можно использовать для очистки гетеромультимеров по настоящему изобретению, описан ниже.
Гетеромультимеры загружают на колонку с белком А (например, mAbSure) при 4°C
↓
Колонку промывают KPO4, затем PBS + 0,1% Тритон X114
↓
Образец элюируют 100 мM раствором уксусной кислоты (pH 2,9)
↓
Значение рН в образце доводят до 8,0 с помощью 1 М Трис и проводят расщепление в течение 1,5 ч при 37°C, 1:500 (масс/масс) Lys-C
↓
Образец очищают на смоле mAbSure, чтобы удалить Lys-C
↓
Образец элюируют 100 мM раствором уксусной кислоты (рН 2,9) и нейтрализуют до рН 8 с помощью 1 М Трис
Образец расщепляют карбоксипептидазой В 1:5 (масс/масс) в течение 2,5 часов при 37°C
↓
После доведения рН образец наносят на катионообменную колонку в буфере, содержащем 20 мМ ацетат Na, рН 5,0, с градиентом до 20 мМ ацетат Na + 1М NaCl
↓
Фракции собирают, объединяют и диализуют против PBS
В частности, гетеромультимеры очищают от кондиционированной среды с использованием смолы mAbSure Select, GE Healthcare (Швеция), в течение ночи при 4°С. Колонку промывают двумя объемами колонки (CV) PBS (забуференный фосфатом физиологический раствор), затем 10 CV PBS + детергент 0,1% Тритон X114, а затем 10 CV калий-фосфатного буфера. Колонки элюируют 100 мМ уксусной кислотой (рН 2,9), после чего значение рН сразу доводят до 8,0 с помощью раствора трис (конечная концентрация 200 мМ), рН 8,0. HD линкеры удаляют из гетеромультимеров путем обработки эндопептидазой Lys-C (Wako Pure Chemical Laboratories) в соотношении 1:500 (масс/масс) при 37°С в течение 1-5 часов. Затем образец наносят на колонку, чтобы удалить фермент. Затем оставшиеся участки распознавания фурином удаляют с помощью карбоксипептидазы В свиньи (Roche Applied Sciences, Cat # 10103233001), 1:5 (масс/масс), в течение 1-5 часов. После расщепления образцы наносят на катионообменную колонку, чтобы удалить фермент. Пиковые фракции объединяют и диализуют против PBS в течение ночи, после чего проводят масс-спектрометрический анализ, чтобы подтвердить идентичность и чистоту.
Пример 3. Определение характеристик рекомбинантных гетеромультимеров
Чтобы определить, сохраняют ли типичные гетеромультимеры по настоящему изобретению, сконструированные методом гетеродимеризации с использованием HD линкера, связывающие свойства антител или адгезинов, из которых были получены их последовательности, проводят анализы связывания (фиг. 6А-6D). Данные анализы связывания проводят с использованием мастер-программы для определения кинетических параметров на системе ForteBio Octet. Все образцы тестируют в концентрации 25 мкг/мл, обеспечивающей насыщение зондов против IgG человека в повторных экспериментах и среди разных образцов. Зонды нагружают первым образцом в течение 10 минут и промывают PBS в течение 30 секунд. Все ассоциации второго и третьего образцов проводят в течение 15 минут с промыванием PBS в течение 30 секунд между проведением ассоциаций.
Другие варианты осуществления
Все патенты, патентные заявки, публикации патентных заявок, а также другие публикации, цитированные или упомянутые в настоящем описании, включены в качестве ссылки в той же степени, как если бы каждый независимый патент, каждая патентная заявка, каждая публикация патентной заявки или каждая другая публикация были конкретно и индивидуально указаны как включенные в качестве ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| СОДЕРЖАЩИЕ СУПЕРСПИРАЛЬ И/ИЛИ ПРИВЯЗКУ БЕЛКОВЫЕ КОМПЛЕКСЫ И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2573915C2 |
| УЛУЧШЕННАЯ СБОРКА БИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2012 |
|
RU2654567C2 |
| РЕКОМБИНАНТНЫЙ ИММУНОТОКСИН, НАЦЕЛЕННЫЙ НА МЕЗОТЕЛИН | 2012 |
|
RU2600067C2 |
| МОДИФИЦИРОВАННЫЙ ЭКЗОТОКСИН А ПСЕВДОМОНАД | 2014 |
|
RU2687143C2 |
| КОНСТРУКЦИИ МУЛЬТИСПЕЦИФИЧНЫХ АНТИТЕЛ | 2015 |
|
RU2710730C2 |
| УЛУЧШЕННЫЕ СПОСОБЫ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА | 2014 |
|
RU2711322C1 |
| БИСПЕЦИФИЧЕСКИЕ АСИММЕТРИЧНЫЕ ГЕТЕРОДИМЕРЫ, СОДЕРЖАЩИЕ АНТИ-CD3 КОНСТРУКЦИИ | 2013 |
|
RU2650868C2 |
| СПОСОБЫ КОЛИЧЕСТВЕННОГО ОПРЕДЕЛЕНИЯ ПОЛИПЕПТИДОВ | 2019 |
|
RU2782464C2 |
| ПОЛУЧЕНИЕ ГЕТЕРОМУЛЬТИМЕРНЫХ БЕЛКОВ С ИСПОЛЬЗОВАНИЕМ КЛЕТОК МЛЕКОПИТАЮЩИХ | 2015 |
|
RU2727012C2 |
| АНТИТЕЛА ПРОТИВ NOTCH2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2580029C2 |








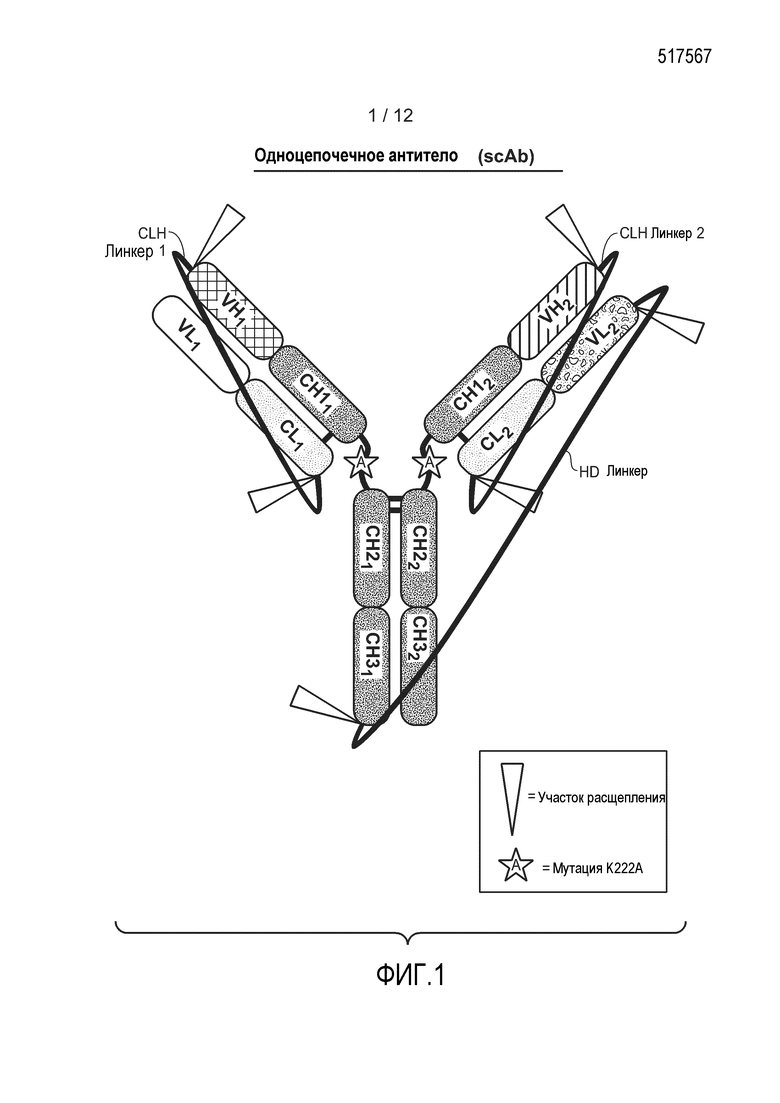
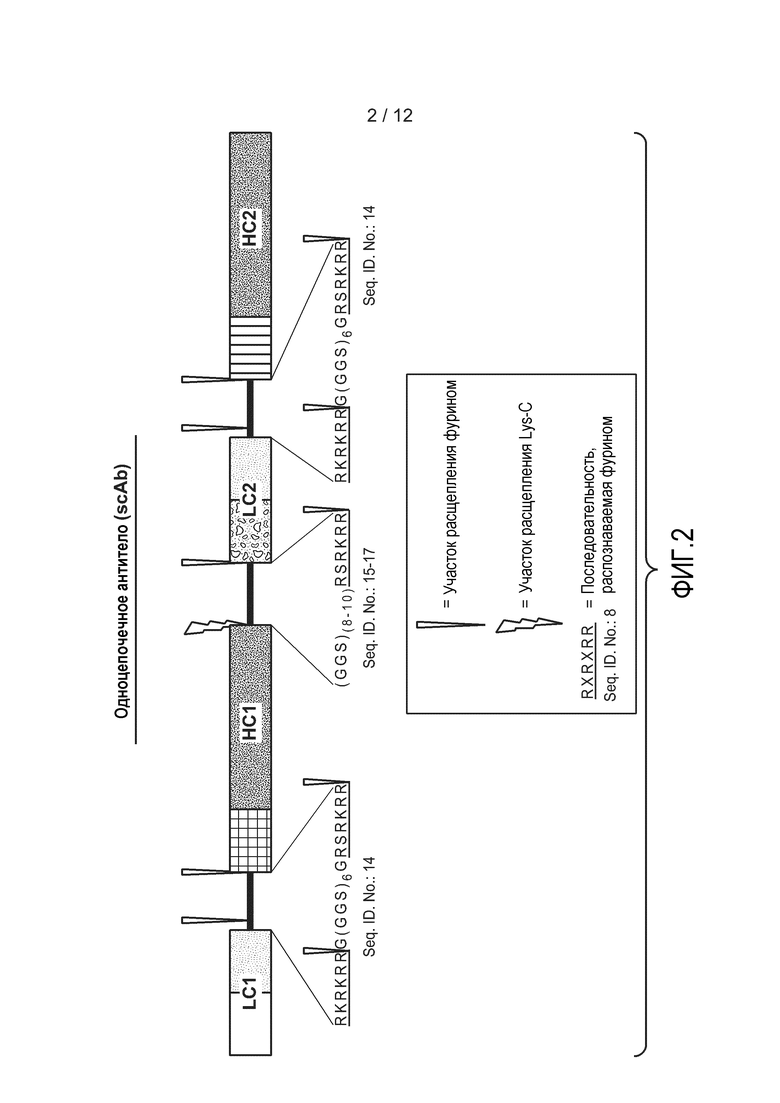
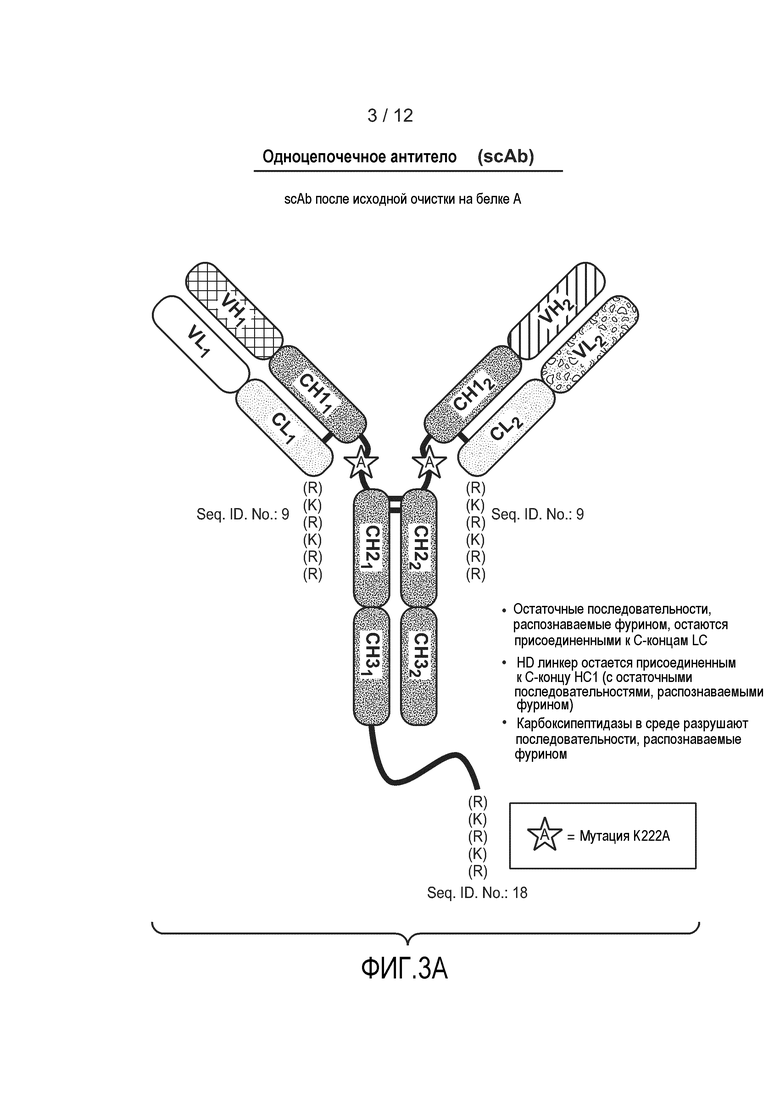
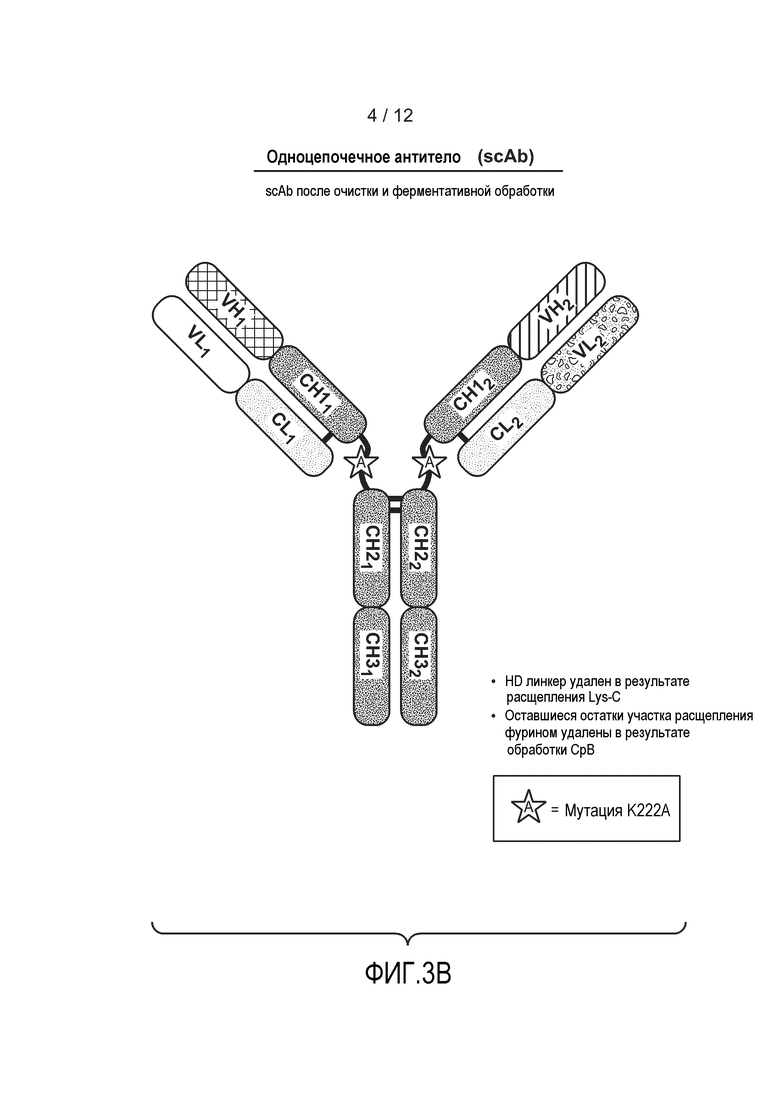

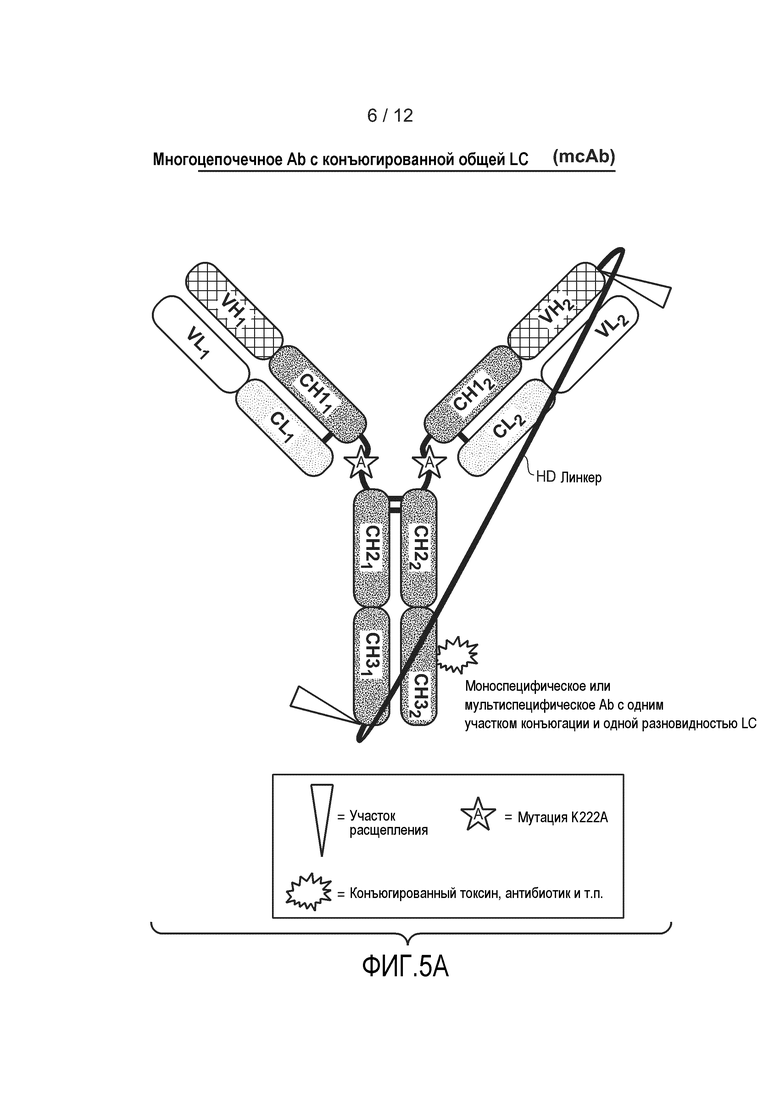
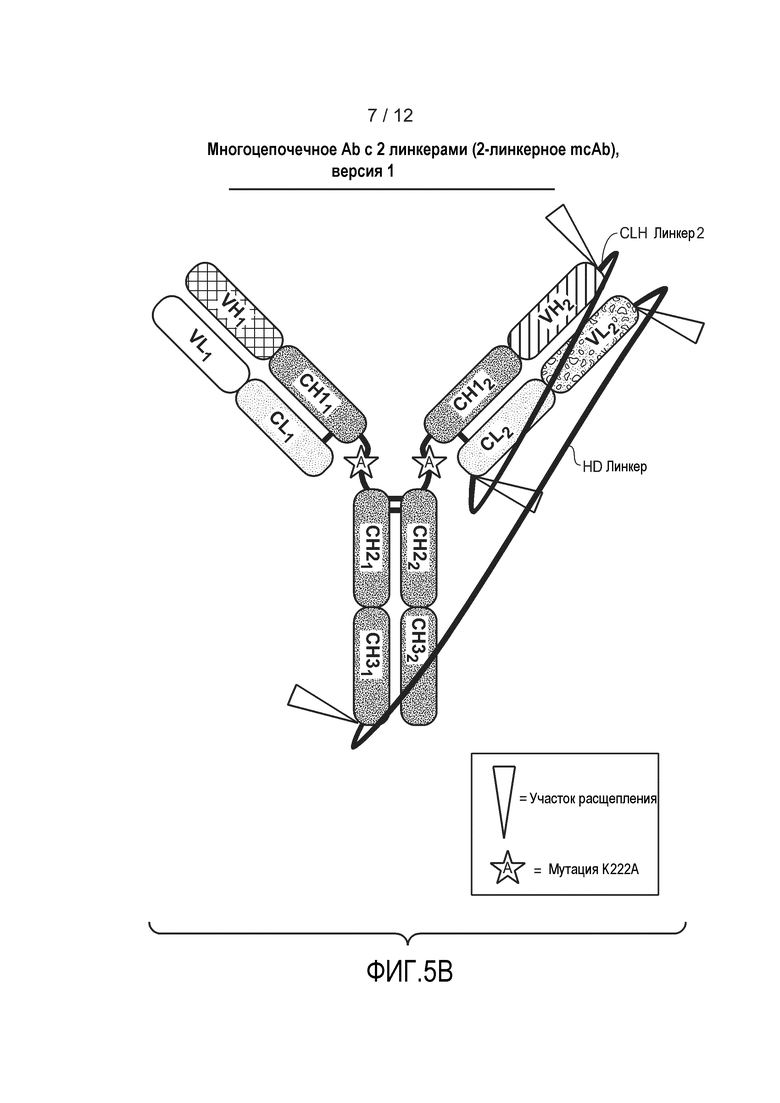
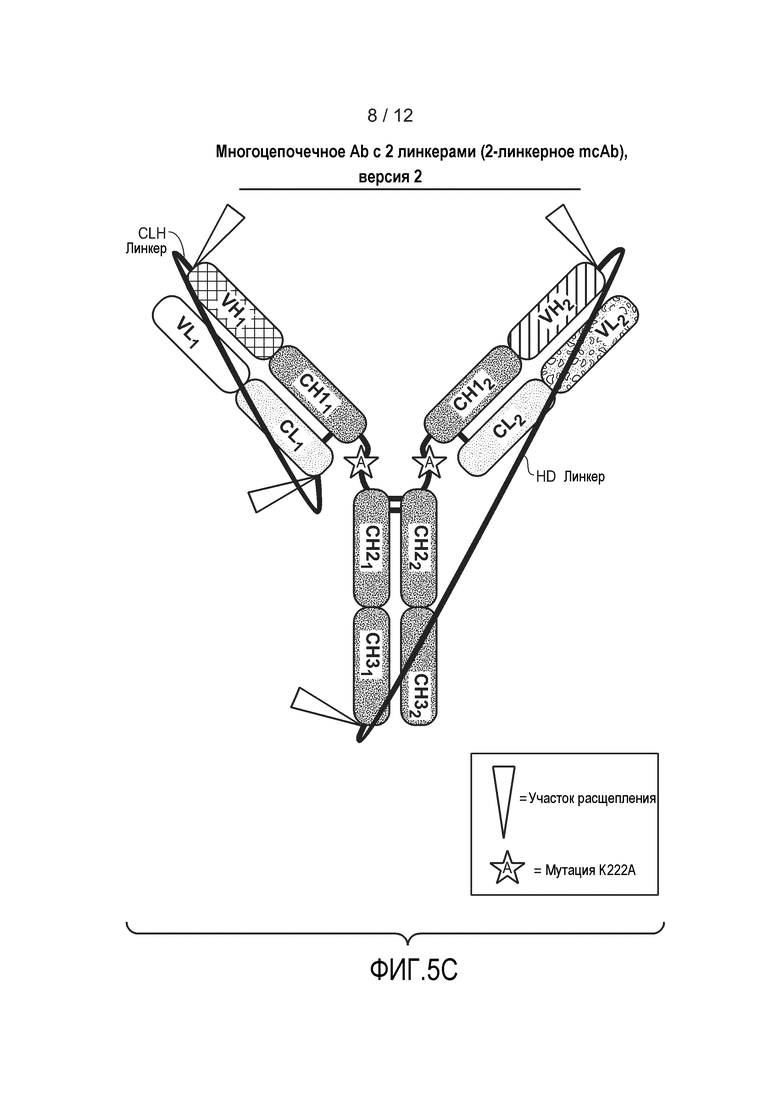
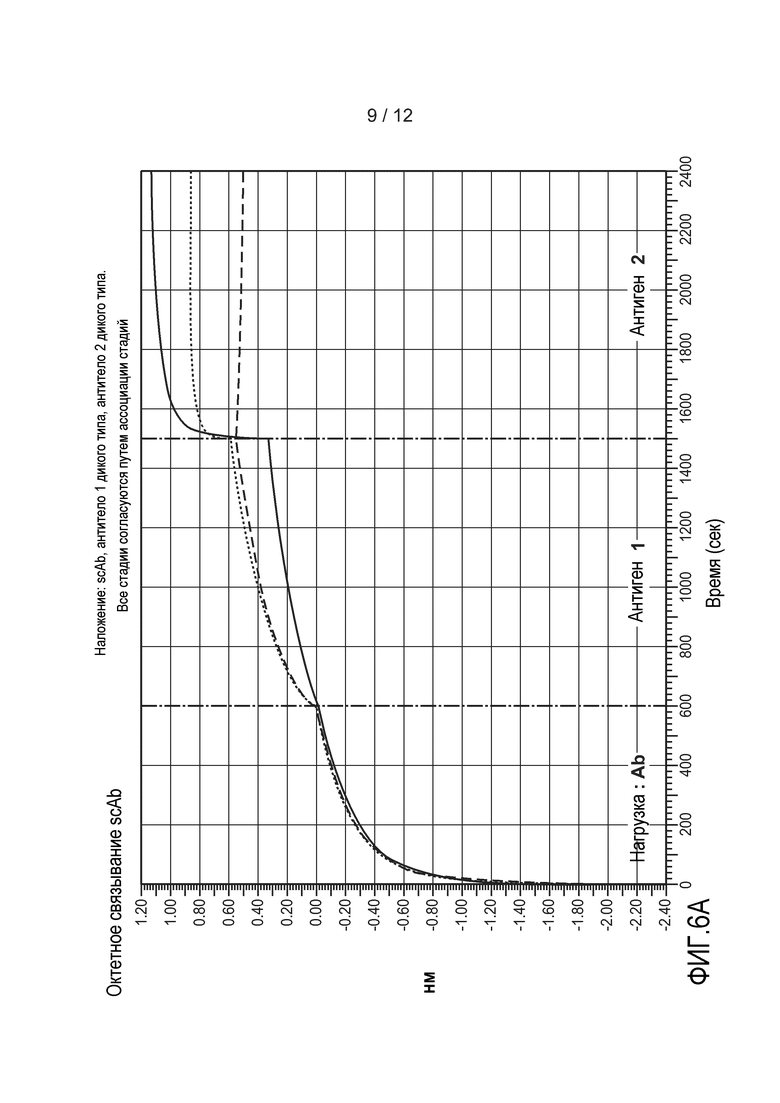
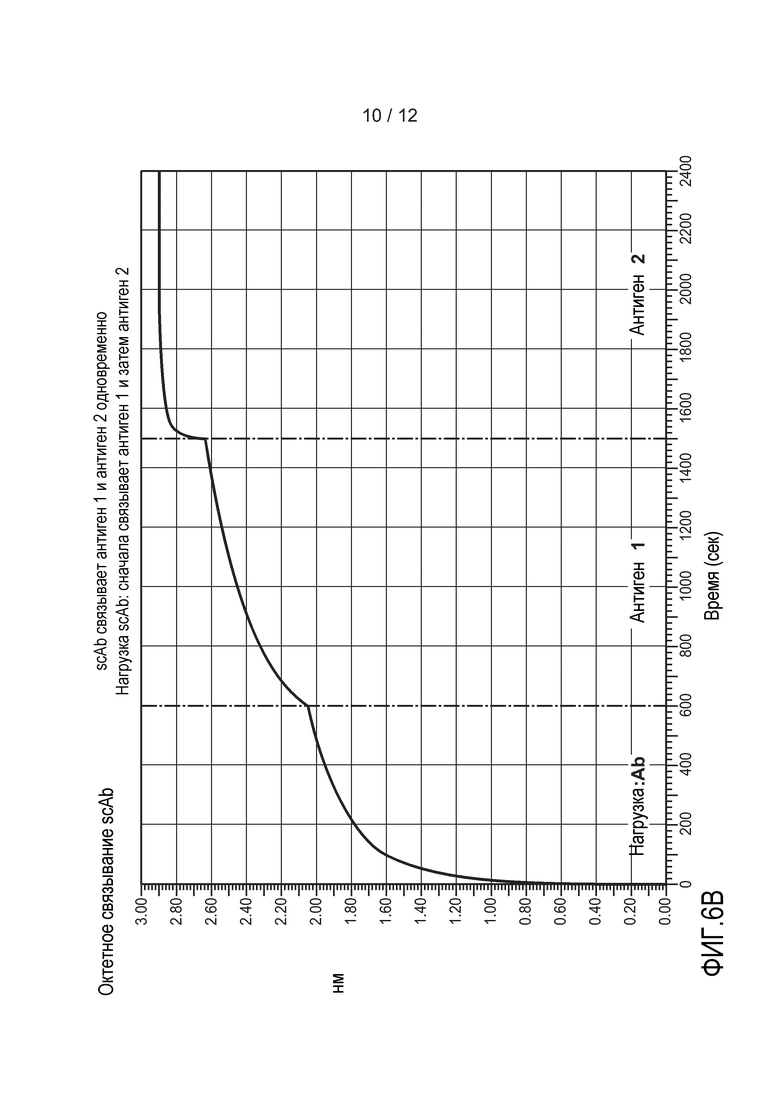
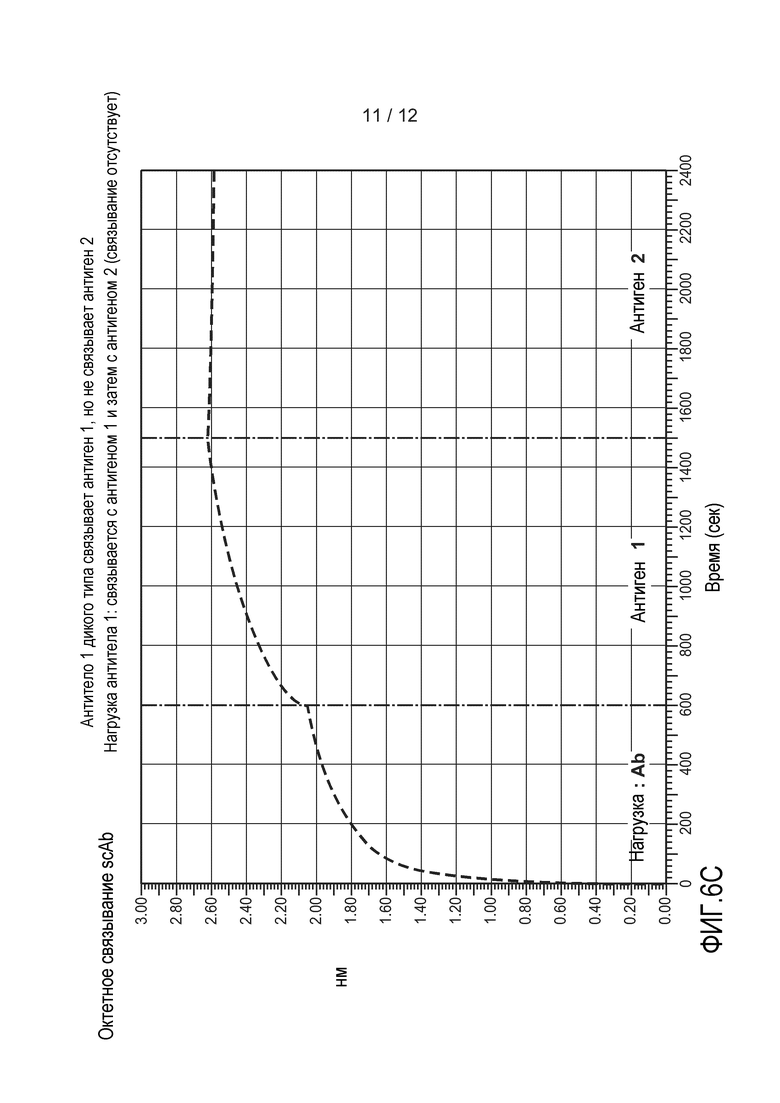
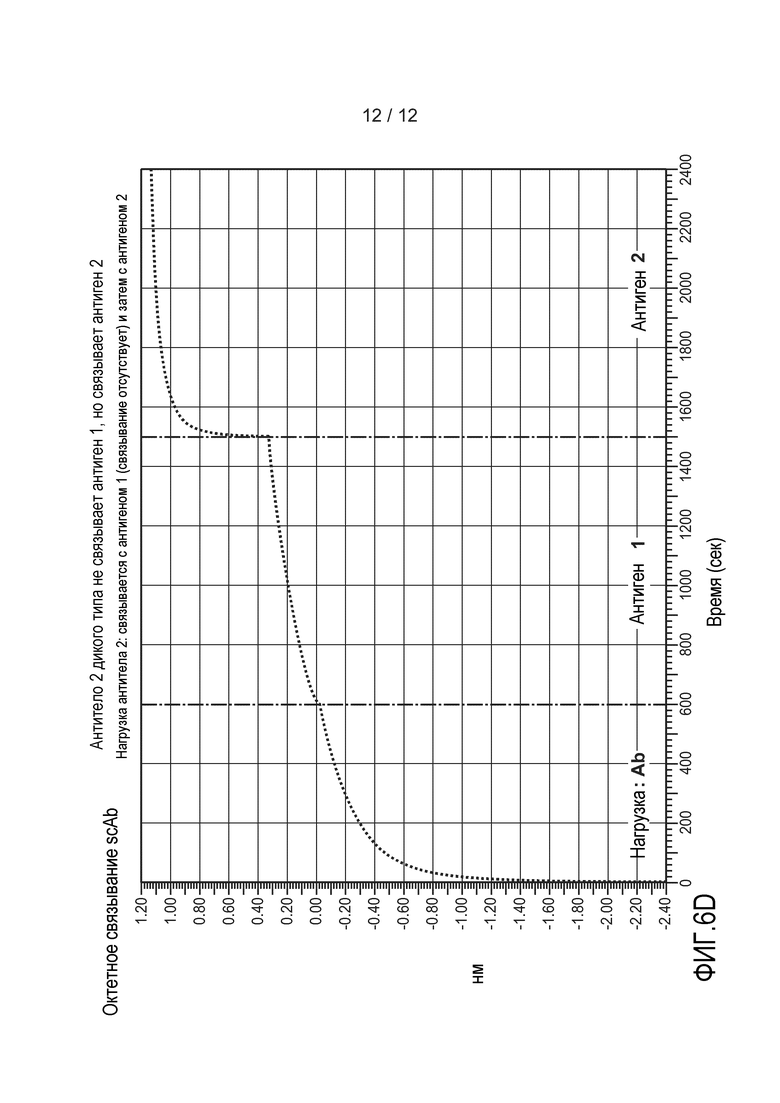
Изобретение относится к биотехнологии. Представлено одноцепочечное антитело для лечения или диагностики, содержащее один полипептид, содержащий следующие домены, расположенные относительно друг друга в направлении от N-конца к С-концу следующим образом: VL1-CL1-CLH линкер1-VH1-CH11-шарнир1-CH21-CH31-HD линкер-VL2-CL2-CLH линкер2-VH2-CH12-шарнир2-CH22-CH32, где каждый из CLH линкера1, CLH линкера2 и HD линкера содержит аминокислотную последовательность, расщепляемую эндопептидазой на N- и C-концах. Также представлен полинуклеотид, кодирующий одноцепочечное указанное антитело, вектор, содержащий такой полинуклеотид, клетка-хозяин, содержащая данный вектор, способ получения указанного одноцепочечного антитела с помощью указанной клетки-хозяина. Изобретение расширяет арсенал для получения антител. 10 н. и 102 з.п. ф-лы, 6 ил., 3 пр.
1. Одноцепочечное антитело для лечения, содержащее один полипептид, содержащий следующие домены, расположенные относительно друг друга в направлении от N-конца к С-концу следующим образом: VL1-CL1-CLH линкер1-VH1-CH11-шарнир1-CH21-CH31-HD линкер-VL2-CL2-CLH линкер2-VH2-CH12-шарнир2-CH22-CH32, где каждый из CLH линкера1, CLH линкера2 и HD линкера содержит аминокислотную последовательность, расщепляемую эндопептидазой на N- и C-концах.
2. Одноцепочечное антитело по п. 1, где указанный HD линкер имеет длину от 15 до 100 аминокислот.
3. Одноцепочечное антитело по п. 2, где указанный HD линкер имеет длину от 30 до 39 аминокислот.
4. Одноцепочечное антитело по п. 2, где указанный HD линкер содержит глициновые (G) и сериновые (S) остатки.
5. Одноцепочечное антитело по п. 4, где указанный HD линкер содержит GGS-повторы.
6. Одноцепочечное антитело по п. 5, где указанный HD линкер содержит 8-9 GGS-повторов (SEQ ID NO:19).
7. Одноцепочечное антитело по п. 1, где указанная аминокислотная последовательность HD линкера расщепляется указанной эндопептидазой in situ.
8. Одноцепочечное антитело по п. 7, где указанная аминокислотная последовательность в HD линкере расщепляется при добавлении указанной эндопептидазы после очистки.
9. Одноцепочечное антитело по п. 1, где указанный HD линкер содержит две аминокислотные последовательности, расщепляемые эндопептидазой на N- и С-концах HD линкера.
10. Одноцепочечное антитело по п. 7, где указанная эндопептидаза выбрана из группы, состоящей из фурина, урокиназы, тромбина, тканевого активатора плазминогена (tРА), гененазы, Lys-C, Arg-С, Asp-N, Glu-C, фактора Ха, протеазы вируса травления табака (ТРВ), энтерокиназы, протеазы риновируса человека С3 (HRV С3) и кининогеназы.
11. Одноцепочечное антитело по п. 10, где указанная эндопептидаза представляет собой фурин.
12. Одноцепочечное антитело по п. 11, где указанный HD линкер содержит два сайта, расщепляемые эндопептидазой на N- и С-концах указанного HD линкера, где один из указанных сайтов, расщепляемых эндопептидазой, представляет собой аминокислотную последовательность, расщепляемую фурином.
13. Одноцепочечное антитело по п. 12, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RKRKRR (SEQ ID NO:9).
14. Одноцепочечное антитело по п. 12, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RHRQPR (SEQ ID NO:10).
15. Одноцепочечное антитело по п. 10, где указанная эндопептидаза представляет собой Lys-C.
16. Одноцепочечное антитело по п. 1, где указанный CLH линкер1 и/или CLH линкер2 содержит глициновые (G) и сериновые (S) остатки.
17. Одноцепочечное антитело по п. 16, где указанный CLH линкер1 и/или CLH линкер2 содержит GGS-повторы.
18. Одноцепочечное антитело по п. 1, где указанный CLH линкер1 и/или CLH линкер2 имеет длину от 10 до 80 аминокислот.
19. Одноцепочечное антитело по п. 18, где указанный CLH линкер1 и/или CLH линкер2 имеет длину от 20 до 40 аминокислот.
20. Одноцепочечное антитело по п. 1, где указанный CLH линкер1 и/или CLH линкер2 расщепляется указанной эндопептидазой in situ.
21. Одноцепочечное антитело по п. 20, где указанная аминокислотная последовательность в CLH линкере1 и/или CLH линкере2 расщепляется при добавлении указанной эндопептидазы после очистки.
22. Одноцепочечное антитело по п. 20, где указанная эндопептидаза выбрана из группы, состоящей из фурина, урокиназы, тромбина, тканевого активатора плазминогена (tРА), гененазы, Lys-C, Arg-С, Asp-N, Glu-C, фактора Ха, протеазы вируса травления табака (ТРВ), энтерокиназы, протеазы риновируса человека С3 (HRV С3) и кининогеназы.
23. Одноцепочечное антитело по п. 22, где указанная эндопептидаза представляет собой фурин.
24. Одноцепочечное антитело по п. 23, где указанный CLH линкер1 и/или CLH линкер2 содержит два сайта, расщепляемые эндопептидазой на N- и С-концах указанного CLH линкера1 или CLH линкера2, где оба сайта, расщепляемых эндопептидазой, расщепляются фурином.
25. Одноцепочечное антитело по п. 24, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RKRKRR (SEQ ID NO:9).
26. Одноцепочечное антитело по п. 24, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RHRQPR (SEQ ID NO:10).
27. Одноцепочечное антитело по п. 1, где шарнир1 и/или шарнир2 содержат от Glu216 до Pro230 IgG1 человека.
28. Одноцепочечное антитело по п. 27, где шарнир1 и/или шарнир2 содержат сайт, расщепляемый эндопептидазой Lys-C.
29. Одноцепочечное антитело по п. 28, где сайт, расщепляемый эндопептидазой Lys-C, содержит инактивирующую мутацию, удаляющую участок расщепления эндопептидазой Lys-C.
30. Одноцепочечное антитело по п. 29, где указанная инактивирующая мутация представляет собой замену K222A (система нумерации EU).
31. Одноцепочечное антитело по п. 1, где указанное одноцепочечное антитело представляет собой биспецифическое или мультиспецифическое одноцепочечное антитело.
32. Одноцепочечное антитело по п. 31, где указанное одноцепочечное антитело способно связываться по меньшей мере с двумя антигенами.
33. Одноцепочечное антитело по п. 31, где указанное одноцепочечное антитело способно связывать по меньшей мере два эпитопа на одном и том же антигене.
34. Одноцепочечное антитело по п. 1, где указанное одноцепочечное антитело содержит константные области, конъюгированные с функциональным фрагментом на по меньшей мере одном компоненте Fc.
35. Одноцепочечное антитело по п. 1, где указанное одноцепочечное антитело содержит один или более расщепляемых сайтов для дальнейшего расщепления экзопептидазой одного или более из указанных HD или CLH линкеров.
36. Одноцепочечное антитело по п. 35, где указанные один или более расщепляемых сайтов для экзопептидазы расщепляются экзопептидазой in situ.
37. Одноцепочечное антитело по п. 36, где указанные один или более расщепляемых сайтов для экзопептидазы расщепляются при добавлении экзопептидазы после очистки.
38. Одноцепочечное антитело по п. 35, где указанная экзопептидаза выбрана из группы, состоящей из карбоксипептидазы А, карбоксипептидазы B, карбоксипептидазы B плазмы, карбоксипептидазы D, карбоксипептидазы E, карбоксипептидазы М, карбоксипептидазы N и карбоксипептидазы Z.
39. Одноцепочечное антитело по п. 38, где указанная экзопептидаза расщепляет на остатке основной аминокислоты.
40. Одноцепочечное антитело по п. 38, где указанная экзопептидаза представляет собой карбоксипептидазу B.
41. Одноцепочечное антитело по п. 1, где каждый из указанных доменов CH31 и CH32 содержит выступ или впадину, где указанные выступ и полость указанного домена CH31 расположены в полости или выступе домена CH32 соответственно.
42. Одноцепочечное антитело по п. 41, где указанные домены CH31 и CH32 совмещаются по граничной поверхности между выступом и полостью.
43. Одноцепочечное антитело по п. 1, где указанный полипептид дополнительно содержит:
(a) выступ или полость в указанном домене CH11, или указанном домене CH12, или в обоих, и
(b) выступ или полость в указанном домене CL1, или указанном домене CL2, или в обоих, и где:
(c) указанный выступ или полость указанного домена CH11 располагается в указанной полости или выступе указанного домена CL1 соответственно,
(d) указанный выступ или полость указанного домена CH12 располагается в указанной полости или выступе указанного домена CL2 соответственно; или
(e) оба (c) и (d).
44. Одноцепочечное антитело по п. 43, где (a) указанные домены CH11 и CL1 домены совмещаются по граничной поверхности между выступом и полостью, (b) указанные домены CH12 и CL2 совмещаются по граничной поверхности между выступом и полостью или (a) и (b) вместе.
45. Одноцепочечное антитело по п. 1, где домен CH21 и/или CH22 содержат мутацию в домене CH2, которая приводит к изменению эффекторной функции антитела.
46. Одноцепочечное антитело по п. 45, где указанная мутация в домене CH2, которая приводит к изменению эффекторной функции антитела, представляет собой мутацию N297.
47. Одноцепочечное антитело по п. 1, где СLH линкер1 и/или CLH линкер2 содержит два сайта расщепления эндопептидазой, расположенные на N- и C-концах указанного СLH линкера1 или CLH линкера2, где один из указанных эндопептидазных сайтов представляет собой сайт расщепления фурином и один из указанных эндопептидазных сайтов представляет собой сайт расщепления Lys-C.
48. Одноцепочечное антитело по п. 12, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RXRXRR (SEQ ID NO:8), где X представляет собой любой аминокислотный остаток.
49. Одноцепочечное антитело по п. 24, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RXRXYR (SEQ ID NO:7), где Y представляет собой K или R и X представляет собой любой аминокислотный остаток.
50. Одноцепочечное антитело по п. 12, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RXRXYR (SEQ ID NO: 7), где Y представляет собой K или R и X представляет собой любой аминокислотный остаток.
51. Одноцепочечное антитело по п. 24, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RXRXYR (SEQ ID NO: 7), где Y представляет собой K или R и X представляет собой любой аминокислотный остаток.
52. Полинуклеотид, кодирующий одноцепочечное антитело по любому из пп. 1-51.
53. Вектор для получения одноцепочечного антитела по любому из пп. 1-51, содержащий полинуклеотид по п. 52.
54. Клетка-хозяин для получения одноцепочечного антитела по любому из пп. 1-51, содержащая вектор по п. 53.
55. Способ получения одноцепочечного антитела по любому из пп. 1-51, где указанный способ включает культивирование клетки-хозяина, которая содержит вектор, содержащий полинуклеотид, способный экспрессировать одноцепочечное антитело по любому из пп. 1-51, в культуральной среде в условиях, подходящих для экспрессии указанного одноцепочечного антитела.
56. Способ по п. 55, где указанный способ дополнительно включает выделение указанного одноцепочечного антитела из указанной клетки-хозяина или из указанной культуральной среды.
57. Одноцепочечное антитело для диагностики, содержащее один полипептид, содержащий следующие домены, расположенные относительно друг друга в направлении от N-конца к С-концу следующим образом: VL1-CL1-CLH линкер1-VH1-CH11-шарнир1-CH21-CH31-HD линкер-VL2-CL2-CLH линкер2-VH2-CH12-шарнир2-CH22-CH32, где каждый из CLH линкера1, CLH линкера2 и HD линкера содержит аминокислотную последовательность, расщепляемую эндопептидазой на N- и C-концах.
58. Одноцепочечное антитело по п. 57, где указанный HD линкер имеет длину от 15 до 100 аминокислот.
59. Одноцепочечное антитело по п. 58, где указанный HD линкер имеет длину от 30 до 39 аминокислот.
60. Одноцепочечное антитело по п. 58, где указанный HD линкер содержит глициновые (G) и сериновые (S) остатки.
61. Одноцепочечное антитело по п. 60, где указанный HD линкер содержит GGS-повторы.
62. Одноцепочечное антитело по п. 61, где указанный HD линкер содержит 8-9 GGS-повторов (SEQ ID NO:19).
63. Одноцепочечное антитело по п. 57, где указанная аминокислотная последовательность HD линкера расщепляется указанной эндопептидазой in situ.
64. Одноцепочечное антитело по п. 63, где указанная аминокислотная последовательность в HD линкере расщепляется при добавлении указанной эндопептидазы после очистки.
65. Одноцепочечное антитело по п. 57, где указанный HD линкер содержит две аминокислотные последовательности, расщепляемые эндопептидазой на N- и С-концах HD линкера.
66. Одноцепочечное антитело по п. 63, где указанная эндопептидаза выбрана из группы, состоящей из фурина, урокиназы, тромбина, тканевого активатора плазминогена (tРА), гененазы, Lys-C, Arg-С, Asp-N, Glu-C, фактора Ха, протеазы вируса травления табака (ТРВ), энтерокиназы, протеазы риновируса человека С3 (HRV С3) и кининогеназы.
67. Одноцепочечное антитело по п. 66, где указанная эндопептидаза представляет собой фурин.
68. Одноцепочечное антитело по п. 67, где указанный HD линкер содержит два сайта, расщепляемые эндопептидазой на N- и С-концах указанного HD линкера, где один из указанных сайтов, расщепляемых эндопептидазой, представляет собой аминокислотную последовательность, расщепляемую фурином.
69. Одноцепочечное антитело по п. 68, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RKRKRR (SEQ ID NO:9).
70. Одноцепочечное антитело по п. 68, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RHRQPR (SEQ ID NO:10).
71. Одноцепочечное антитело по п. 66, где указанная эндопептидаза представляет собой Lys-C.
72. Одноцепочечное антитело по п. 57, где указанный CLH линкер1 и/или CLH линкер2 содержит глициновые (G) и сериновые (S) остатки.
73. Одноцепочечное антитело по п. 71, где указанный CLH линкер1 и/или CLH линкер2 содержит GGS-повторы.
74. Одноцепочечное антитело по п. 57, где указанный CLH линкер1 и/или CLH линкер2 имеет длину от 10 до 80 аминокислот.
75. Одноцепочечное антитело по п. 74, где указанный CLH линкер1 и/или CLH линкер2 имеет длину от 20 до 40 аминокислот.
76. Одноцепочечное антитело по п. 57, где указанный CLH линкер1 и/или CLH линкер2 расщепляется указанной эндопептидазой in situ.
77. Одноцепочечное антитело по п. 76, где указанная аминокислотная последовательность в CLH линкере1 и/или CLH линкере2 расщепляется при добавлении указанной эндопептидазы после очистки.
78. Одноцепочечное антитело по п. 76, где указанная эндопептидаза выбрана из группы, состоящей из фурина, урокиназы, тромбина, тканевого активатора плазминогена (tРА), гененазы, Lys-C, Arg-С, Asp-N, Glu-C, фактора Ха, протеазы вируса травления табака (ТРВ), энтерокиназы, протеазы риновируса человека С3 (HRV С3) и кининогеназы.
79. Одноцепочечное антитело по п. 78, где указанная эндопептидаза представляет собой фурин.
80. Одноцепочечное антитело по п. 79, где указанный CLH линкер1 и/или CLH линкер2 содержит два сайта, расщепляемые эндопептидазой на N- и С-концах указанного CLH линкера1 или CLH линкера2, где оба сайта, расщепляемых эндопептидазой, расщепляются фурином.
81. Одноцепочечное антитело по п. 80, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RKRKRR (SEQ ID NO:9).
82. Одноцепочечное антитело по п. 80, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RHRQPR (SEQ ID NO:10).
83. Одноцепочечное антитело по п. 57, где шарнир1 и/или шарнир2 содержат от Glu216 до Pro230 IgG1 человека.
84. Одноцепочечное антитело по п. 83, где шарнир1 и/или шарнир2 содержат сайт, расщепляемый эндопептидазой Lys-C.
85. Одноцепочечное антитело по п. 84, где сайт, расщепляемый эндопептидазой Lys-C, содержит инактивирующую мутацию, удаляющую участок расщепления эндопептидазой Lys-C.
86. Одноцепочечное антитело по п. 85, где указанная инактивирующая мутация представляет собой замену K222A (система нумерации EU).
87. Одноцепочечное антитело по п. 57, где указанное одноцепочечное антитело представляет собой биспецифическое или мультиспецифическое одноцепочечное антитело.
88. Одноцепочечное антитело по п. 87, где указанное одноцепочечное антитело способно связываться по меньшей мере с двумя антигенами.
89. Одноцепочечное антитело по п. 88, где указанное одноцепочечное антитело способно связывать по меньшей мере два эпитопа на одном и том же антигене.
90. Одноцепочечное антитело по п. 57, где указанное одноцепочечное антитело содержит константные области, конъюгированные с функциональным фрагментом на по меньшей мере одном компоненте Fc.
91. Одноцепочечное антитело по п. 57, где указанное одноцепочечное антитело содержит один или более расщепляемых сайтов для дальнейшего расщепления экзопептидазой одного или более из указанных HD или CLH линкеров.
92. Одноцепочечное антитело по п. 91, где указанные один или более расщепляемых сайтов для экзопептидазы расщепляются экзопептидазой in situ.
93. Одноцепочечное антитело по п. 92, где указанные один или более расщепляемых сайтов для экзопептидазы расщепляются при добавлении экзопептидазы после очистки.
94. Одноцепочечное антитело по п. 91, где указанная экзопептидаза выбрана из группы, состоящей из карбоксипептидазы А, карбоксипептидазы B, карбоксипептидазы B плазмы, карбоксипептидазы D, карбоксипептидазы E, карбоксипептидазы М, карбоксипептидазы N и карбоксипептидазы Z.
95. Одноцепочечное антитело по п. 94, где указанная экзопептидаза расщепляет на остатке основной аминокислоты.
96. Одноцепочечное антитело по п. 94, где указанная экзопептидаза представляет собой карбоксипептидазу B.
97. Одноцепочечное антитело по п. 57, где каждый из указанных доменов CH31 и CH32 содержит выступ или впадину, где указанные выступ и полость указанного домена CH31 расположены в полости или выступе домена CH32 соответственно.
98. Одноцепочечное антитело по п. 97, где указанные домены CH31 и CH32 совмещаются по граничной поверхности между выступом и полостью.
99. Одноцепочечное антитело по п. 57, где указанный полипептид дополнительно содержит:
(a) выступ или полость в указанном домене CH11, или указанном домене CH12, или в обоих, и
(b) выступ или полость в указанном домене CL1, или указанном домене CL2, или в обоих, и где:
(c) указанный выступ или полость указанного домена CH11 располагается в указанной полости или выступе указанного домена CL1 соответственно,
(d) указанный выступ или полость указанного домена CH12 располагается в указанной полости или выступе указанного домена CL2 соответственно; или
(e) оба (c) и (d).
100. Одноцепочечное антитело по п. 99, где (a) указанные домены CH11 и CL1 домены совмещаются по граничной поверхности между выступом и полостью, (b) указанные домены CH12 и CL2, совмещаются по граничной поверхности между выступом и полостью, или (a) и (b) вместе.
101. Одноцепочечное антитело по п. 57, где домен CH21 и/или CH22 содержат мутацию в домене CH2, которая приводит к изменению эффекторной функции антитела.
102. Одноцепочечное антитело по п. 101, где указанная мутация в домене CH2, которая приводит к изменению эффекторной функции антитела, представляет собой мутацию N297.
103. Одноцепочечное антитело по п. 57, где СLH линкер1 и/или CLH линкер2 содержит два сайта расщепления эндопептидазой, расположенные на N- и C-концах указанного СLH линкера1 или CLH линкера2, где один из указанных эндопептидазных сайтов представляет собой сайт расщепления фурином и один из указанных эндопептидазных сайтов представляет собой сайт расщепления Lys-C.
104. Одноцепочечное антитело по п. 68, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RXRXRR (SEQ ID NO:8), где X представляет собой любой аминокислотный остаток.
105. Одноцепочечное антитело по п. 80, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RXRXYR (SEQ ID NO:7), где Y представляет собой K или R и X представляет собой любой аминокислотный остаток.
106. Одноцепочечное антитело по п. 68, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RXRXYR (SEQ ID NO: 7), где Y представляет собой K или R и X представляет собой любой аминокислотный остаток.
107. Одноцепочечное антитело по п. 80, где аминокислотная последовательность, расщепляемая фурином, содержит аминокислотную последовательность RXRXYR (SEQ ID NO: 7), где Y представляет собой K или R и X представляет собой любой аминокислотный остаток.
108. Полинуклеотид, кодирующий одноцепочечное антитело по любому из пп. 57-107.
109. Вектор для получения одноцепочечного антитела по любому из пп. 57-107, содержащий полинуклеотид по п. 108.
110. Клетка-хозяин для получения одноцепочечного антитела по любому из пп. 57-107, содержащая вектор по п. 109.
111. Способ получения одноцепочечного антитела по любому из пп. 57-107, где указанный способ включает культивирование клетки-хозяина, которая содержит вектор, содержащий полинуклеотид, способный экспрессировать одноцепочечное антитело по любому из пп. 57-107, в культуральной среде в условиях, подходящих для экспрессии указанного одноцепочечного антитела.
112. Способ по п.111, где указанный способ дополнительно включает выделение указанного одноцепочечного антитела из указанной клетки-хозяина или из указанной культуральной среды.
| ELS CONRATH K | |||
| et al | |||
| Camel single-domain antibodies as modular building units in bispecific and bivalent antibody constructs | |||
| JOURNAL OF BIOLOGICAL CHEMISTRY, 2001, vol.276, no.10, pp.7346-7350 | |||
| РЫБАЛЬСКИЙ Н.Г., СЕРОВА М | |||
| А., ИГНАТЬЕВА Г.А., СТАРЧЕУС А.П | |||
| Моноклональные антитела и гибридомы | |||
| М.: ВАСХНИЛ, 1989, стр.23-44. |

