ОБЛАСТЬ ТЕХНИКИ
Настоящая заявка относится к рекомбинантному микроорганизму, обладающему улучшенным уровнем внутриклеточной энергии, и к способу продуцирования L-аминокислот с помощью микроорганизма.
УРОВЕНЬ ТЕХНИКИ
Для продуцирования желаемого вещества с помощью микроорганизма применяли главным образом специфичные для желаемого вещества подходы, такие как усиление экспрессии генов, кодирующих ферменты, участвующие в продуцировании желаемого вещества, или удаление ненужных генов. Например, посредством повышения активности пути биосинтеза L-аминокислоты был разработан ряд применимых штаммов, включающих E. coli, способных к продуцированию желаемой L-аминокислоты с высоким выходом. Для продуцирования применимых желаемых веществ с высоким выходом с помощью микроорганизмов требуется образование и поддержание достаточного количества энергии.
Для биосинтеза веществ, таких как белки, нуклеиновые кислоты и т.п., in vivo используется энергия, запасаемая в форме NADH, NADPH и ATP (аденозин-5'-трифосфата). В частности, ATP представляет собой носитель энергии, который транспортирует химическую энергию, образующуюся в метаболических реакциях, для различных видов активности организмов.
ATP образуется главным образом в ходе метаболических процессов у микроорганизмов. Основными внутриклеточными путями образования ATP являются субстратное фосфорилирование, которое происходит путем гликолиза, или окислительное фосфорилирование, в ходе которого образуется ATP посредством электронтранспортной системы с использованием восстанавливающей способности, накапливаемой в NADH и т.п. путем гликолиза. Образованный ATP расходуется на такие виды активности in vivo, как биосинтез, движение, передача сигналов и деление клеток. Таким образом, промышленные микроорганизмы, применяемые для продуцирования применимых желаемых веществ, обычно характеризуются высоким потреблением ATP. Соответственно, проводили исследования для улучшения производительности путем повышения уровней внутриклеточной энергии при массовом продуцировании применимых желаемых веществ (Biotechnol Adv (2009) 27:94-101).
Железо является одним из элементов, существенных для поддержания гомеостаза микроорганизмов, и E. coli использует различные пути поглощения железа (Mol Microbiol (2006) 62:120-131). Одним из путей поглощения железа является поглощение железа посредством каналов комплекса FhuCDB, образованного белками FhuC, FhuD и FhuB. Недавно было выявлено, что в присутствии избытка L-триптофана в клетках белок TrpR, регулирующий экспрессию генов, участвующих в биосинтезе L-триптофана, образует комплекс с L-триптофаном, и этот комплекс, в свою очередь, связывается с регуляторной областью оперона fhuCDB, что позволяет предположить возможность существования корреляции между поглощением железа посредством белкового комплекса FhuCDB и биосинтезом L-триптофана. Однако, функция белкового комплекса FhuCDB в биосинтезе L-триптофана, а также его влияние на поглощение железа еще не были выяснены (Nat Chem Biol (2012) 8:65-71).
Авторы настоящего изобретения исследовали способы улучшения уровней ATP и повышения производительности в отношении применимых желаемых веществ, таких как L-аминокислоты, и они обнаружили, что уровни внутриклеточного ATP можно улучшить путем инактивации функции белкового комплекса FhuCDB посредством делеции гена fhuCDB, и в результате этого производительность в отношении желаемых веществ можно повысить, что тем самым довершает настоящую заявку.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ТЕХНИЧЕСКАЯ ЗАДАЧА
Целью настоящей заявки является обеспечение микроорганизма, обладающего улучшенным уровнем внутриклеточного ATP.
Другой целью настоящей заявки является обеспечение способа продуцирования желаемого вещества с помощью микроорганизма, обладающего улучшенным уровнем внутриклеточного ATP.
ТЕХНИЧЕСКОЕ РЕШЕНИЕ
В одном аспекте в настоящей заявке представлен микроорганизм, обладающий улучшенным уровнем внутриклеточного ATP.
В конкретном варианте осуществления настоящей заявки микроорганизм может представлять собой микроорганизм, в котором виды активности одного или нескольких из белка FhuC, белка FhuD и белка FhuB, образующих систему поглощения железа, были инактивированы, и, следовательно, который обладает повышенным уровнем внутриклеточного ATP по сравнению с немодифицированным штаммом.
Термин “FhuCDB”, используемый в данном документе, означает компонент системы поглощения железа (системы fhu), включающий в себя продукты экспрессии fhuA, fhuC, fhuD и fhuB, организованных в один оперон. fhuA кодирует многофункциональный OMP FhuA (79 кДа), действующий в качестве рецептора для комплекса феррихром-железо, фагов, бактериальных токсинов и антибиотиков. FhuA специфичен для комплекса Fe3+-феррихром и действует в качестве лиганд-специфичного управляемого канала (Protein Sci 7, 1636-1638). Другие белки системы fhu, а именно FhuD, FhuC и FhuB, также существенны для функций системы поглощения железа. Периплазматический белок FhuD и ассоциированные с цитоплазматической мембраной белки FhuC и FhuB образуют комплекс FhuCDB, функцией которого является транспорт феррихрома и других гидроксаматных соединений Fe3+ (комплексов Fe3+-аэробактин, Fe3+-копроген) через цитоплазматическую мембрану из периплазматического пространства в цитоплазму (J Bacteriol 169, 3844-3849). При поглощении железа посредством комплекса FhuCDB расходуется одна молекула ATP, и для этого процесса поглощения железа энергию предоставляет белковый комплекс TonB-ExbB-ExbD (FEBS Lett 274, 85-88).
FhuC кодирует ассоциированный с цитоплазматической мембраной белок размером 29 кДа и вместе с FhuD и FhuB образует канал для поглощения железа. FhuC может иметь аминокислотную последовательность SEQ ID NO: 5, и, в частности, FhuC может кодироваться нуклеотидной последовательностью SEQ ID NO: 1.
FhuD кодирует ассоциированный с цитоплазматической мембраной белок размером 31 кДа и вместе с FhuC и FhuB образует канал для поглощения железа. FhuD может иметь аминокислотную последовательность SEQ ID NO: 6, и, в частности, FhuD может кодироваться нуклеотидной последовательностью SEQ ID NO: 2.
FhuB кодирует ассоциированный с цитоплазматической мембраной белок размером 41 кДа и вместе с FhuC и FhuB образует канал для поглощения железа. FhuB может иметь аминокислотную последовательность SEQ ID NO: 7, и, в частности, FhuB может кодироваться нуклеотидной последовательностью SEQ ID NO: 3.
Более конкретно, хотя белки обладают идентичными видами активности, между объектами существуют небольшие различия в аминокислотных последовательностях. Таким образом, FhuC, FhuD и FhuB могут соответственно иметь SEQ ID NO: 5, 6 и 7, но не ограничиваться ими. Иными словами, FhuC, FhuD и FhuB в настоящей заявке могут представлять собой варианты, имеющие аминокислотные последовательности с заменой, делецией, вставкой, добавлением или инверсией одной или более аминокислот в одном или более положениях аминокислотных последовательностей, и они могут иметь последовательности, обладающие 70% или более высокой, 80% или более высокой, 90% или более высокой или 95% или более высокой гомологией с аминокислотными последовательностями SEQ ID NO: 5, 6 и 7 соответственно. Кроме того, в нуклеотидных последовательностях могут быть произведены различные модификации в кодирующей области при условии, что они не изменяют аминокислотные последовательности белков, экспрессируемых с кодирующей области, по причине вырожденности кодонов или с учетом предпочтительности кодонов для организма, в котором они подлежат экспрессии. Вышеописанная нуклеотидная последовательность приведена только в качестве примера различных нуклеотидных последовательностей, получаемых с помощью способа, хорошо известного специалистам в данной области, но не ограниченных ей.
Термин “гомология”, используемый в данном документе, относится к степени идентичности оснований или аминокислотных остатков после выравнивания обеих последовательностей для наилучшего совпадения определенных сравнимых областей в аминокислотной или нуклеотидной последовательности гена, кодирующего белок. Если гомология является достаточно высокой, то продукты экспрессии соответствующих генов могут обладать идентичной или сходной активностью. Процентное значение идентичности последовательностей можно определить с помощью известной программы для сравнения последовательностей, например, BLASTN (NCBI), CLC Main Workbench (CLC bio), MegAlign™ (DNASTAR Inc) и т.п.
Термин “микроорганизм”, используемый в данном документе, относится к прокариотическому микроорганизму или эукариотическому микроорганизму, обладающему способностью к продуцированию применимого желаемого вещества, такого как L-аминокислоты. Например, микроорганизм, обладающий улучшенным уровнем внутриклеточного ATP, может принадлежать к роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacteria, роду Pseudomonas, роду Leptospira, роду Salmonellar, роду Brevibacteria, роду Hypomononas, роду Chromobacterium или роду Norcardia микроорганизмов, или грибов, или дрожжей. В частности, микроорганизм может принадлежать к роду микроорганизмов Escherichia, и более конкретно, микроорганизм может представлять собой E. coli.
“Немодифицированный штамм”, как используется в данном документе, относится к микроорганизму, который не является модифицированным с помощью методики молекулярной биологии, такой как мутация или рекомбинация. В частности, немодифицированный штамм относится к микроорганизму до повышения уровня внутриклеточного ATP, в котором уровень внутриклеточного ATP повышают путем инактивации одного или нескольких из FhuC, FhuD и FhuB, образующих систему поглощения железа, комплекс FhuCDB, тем самым снижая расход внутриклеточного ATP. Иными словами, немодифицированный штамм относится к исходному микроорганизму, от которого происходит рекомбинантный микроорганизм.
В конкретном варианте осуществления настоящей заявки микроорганизм может содержать один или несколько из FhuC, FhuD и FhuB в инактивированном состоянии, и комбинацию FhuC, FhuD и FhuB в инактивированном состоянии, и, в частности, все из FhuC, FhuD и FhuB в инактивированном состоянии.
Термин “инактивация”, используемый в данном документе, означает, что активность соответствующего белка устранена или ослаблена посредством мутации в результате делеции, замены или вставки части или всего гена, кодирующего соответствующий белок, посредством модификации последовательности, регулирующей экспрессию, для снижения экспрессии гена, посредством модификации последовательности хромосомного гена для ослабления или устранения активности белка или посредством их комбинаций.
В частности, делецию части или всего гена, кодирующего белок, можно осуществлять путем замены полинуклеотида, который кодирует эндогенный целевой белок, в хромосоме полинуклеотидом, в котором имеет место делеция частичной последовательности, либо маркерным геном посредством инсерционного вектора на основе бактериальной хромосомы. Кроме того, можно индуцировать мутацию с помощью мутагена, такого как химические вещества или UV-свет, получая таким образом мутанта, имеющего делецию соответствующего гена, но не ограничиваясь этим.
Термин “последовательность, регулирующая экспрессию”, используемый в данном документе, т.е. нуклеотидная последовательность, регулирующая экспрессию гена, относится к сегменту, способному к повышению или снижению экспрессии конкретного гена у субъекта, и может включать в себя промотор, сайт связывания фактора транскрипции, сайт связывания рибосом, последовательность, регулирующую терминацию транскрипции и трансляции, но не ограничивается этим.
В частности, модификацию последовательности, регулирующей экспрессию, для того, чтобы вызвать снижение экспрессии гена, можно осуществлять путем индуцирования мутаций в последовательности, регулирующей экспрессию, посредством делеции, вставки, консервативной или неконсервативной замены в нуклеотидной последовательности или их комбинации для дополнительного ослабления активности последовательности, регулирующей экспрессию, или путем замены последовательности, регулирующей экспрессию, последовательностью, обладающей более слабой активностью, но не ограничиваясь этим.
В конкретном варианте осуществления настоящей заявки микроорганизм может представлять собой микроорганизм из рода Escherichia, обладающий улучшенной производительностью в отношении желаемого вещества по сравнению с немодифицированным штаммом. У микроорганизма из рода Escherichia согласно настоящей заявке один или несколько белков, образующих комплекс FhuCDB, инактивированы для инактивации пути поглощения железа, и, таким образом, при поглощении железа посредством данного пути снижается расход ATP. В результате этого микроорганизм из рода Escherichia имеет улучшенный уровень внутриклеточного ATP по сравнению с немодифицированным штаммом, и, следовательно, микроорганизм обладает улучшенной производительностью в отношении желаемого вещества.
Термин “микроорганизм, обладающий улучшенной производительностью” относится к микроорганизму, обладающему улучшенной производительностью в отношении желаемого вещества по сравнению с немодифицированным штаммом или исходной клеткой до модификации.
Термин “желаемое вещество”, используемый в данном документе, включает вещество, продуцируемое количество которого повышают путем повышения уровня внутриклеточного ATP у микроорганизма, без ограничения. Желаемое вещество может, в частности, представлять собой L-аминокислоту и, более конкретно, L-треонин или L-триптофан.
В конкретном варианте осуществления микроорганизм может представлять собой E. coli, обладающую улучшенной производительностью в отношении L-триптофана, где один или несколько из FhuC, FhuD и FhuB E. coli, обладающей производительностью в отношении L-триптофана, были инактивированы, и обладающую улучшенным уровнем внутриклеточного ATP по сравнению с немодифицированным штаммом. E. coli, обладающую производительностью в отношении L-триптофана, можно получить путем повышения экспрессии гена L-триптофанового оперона, устранения ингибирования по принципу обратной связи конечным продуктом L-триптофаном или устранения ингибирования и аттенюации гена L-триптофанового оперона на уровне транскрипции, но не ограничиваясь этим.
В конкретном варианте осуществления настоящей заявки микроорганизм может представлять собой E. coli, обладающую улучшенной производительностью в отношении L-треонина, где один или несколько из FhuC, FhuD и FhuB E. coli, обладающей производительностью в отношении L-треонина, были инактивированы, и обладающую улучшенным уровнем внутриклеточного ATP по сравнению с немодифицированным штаммом. E. coli, обладающую производительностью в отношении L-треонина, можно получить путем повышения экспрессии гена L-треонинового оперона, устранения ингибирования по принципу обратной связи конечным продуктом L-треонином или устранения ингибирования и аттенюации гена L-треонинового оперона на уровне транскрипции, но не ограничиваясь этим.
В другом аспекте в настоящей заявке обеспечивается способ продуцирования L-аминокислот, при этом способ включает культивирование микроорганизма из рода Escherichia, обладающего улучшенным уровнем внутриклеточного ATP, в среде и извлечение L-аминокислот из культуральной среды или микроорганизма.
Термин “микроорганизм из рода Escherichia, обладающий улучшенным уровнем внутриклеточного ATP”, используемый в данном документе, является таким же, как описанный выше.
В способе продуцирования L-аминокислот в соответствии с конкретным вариантом осуществления настоящей заявки культивирование микроорганизма, обладающего производительностью в отношении L-аминокислот, можно осуществлять в соответствии с надлежащими условиями среды и культивирования, известными из уровня техники. Процедуры культивирования могут без труда корректироваться специалистами в данной области в соответствии с выбранным микроорганизмом. Примеры процедур культивирования включают периодический тип, непрерывный тип и тип культивирования с подпиткой, но не ограничиваются этим.
Среда, применяемая для культивирования, должна соответствовать требованиям для культивирования конкретного микроорганизма. Культуральные среды для различных микроорганизмов описаны в литературе (“Manual of Methods for General Bacteriology”, American Society for Bacteriology, Washington D.C., USA, 1981). Эти среды содержат ряд источников углерода, источников азота и следовых элементов. Источники углерода включают углеводы, такие как глюкоза, лактоза, сахароза, фруктоза, мальтоза, крахмал и целлюлоза; липиды, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; а также органические кислоты, такие как уксусная кислота. Эти источники углерода можно использовать в отдельности или в комбинации, но не ограничиваясь этим. Источник азота включает органические источники азота, такие как пептон, дрожжевой экстракт, подлива, солодовый экстракт, жидкий кукурузный экстракт (CSL) и бобовая мука, а также неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота можно использовать в отдельности или в комбинации, но не ограничиваясь этим. В дополнение, среда может включать дигидрофосфат калия, гидрофосфат дикалия и их соответствующие натрийсодержащие соли в качестве источника фосфора, но не ограничиваться этим. Также среда может включать металл, такой как сульфат магния или сульфат железа. Кроме того, также можно добавлять аминокислоты, витамины и соответствующие предшественники.
Дополнительно, для поддержания культуры в аэробных условиях в культуру можно вводить кислород или кислородсодержащий газ (например, воздух). Температура культуры может обычно составлять 20°C-45°C и, в частности, 25°C-40°C. Культивирование можно продолжать, пока продуцирование L-аминокислот, таких как L-треонин или L-триптофан, не достигнет желаемого уровня, и, в частности, время культивирования может составлять от 10 часов до 100 часов.
Способ продуцирования L-аминокислот в соответствии с конкретным вариантом осуществления настоящей заявки может дополнительно включать извлечение L-аминокислот из полученных таким образом культуральной среды или микроорганизма. Извлечение L-аминокислот можно осуществлять с помощью соответствующего способа, известного из уровня техники, в зависимости от способа культивирования микроорганизма согласно настоящей заявке, например, периодического типа, непрерывного типа или типа культивирования с подпиткой, для очистки или извлечения желаемых L-аминокислот из культуры микроорганизма, но не ограничиваясь этим.
ПРЕИМУЩЕСТВЕННЫЕ ЭФФЕКТЫ ИЗОБРЕТЕНИЯ
В соответствии с настоящей заявкой, в случае, когда применяют микроорганизм из рода Escherichia, обладающий улучшенным уровнем внутриклеточного ATP по сравнению с немодифицированным штаммом, и способ продуцирования желаемого вещества с его помощью, высокий уровень внутриклеточного ATP усиливает экспрессию генов, биосинтез, транспорт веществ и т.п., тем самым обеспечивая эффективное продуцирование применимых желаемых веществ, включающих белки, L-аминокислоты и т.п.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
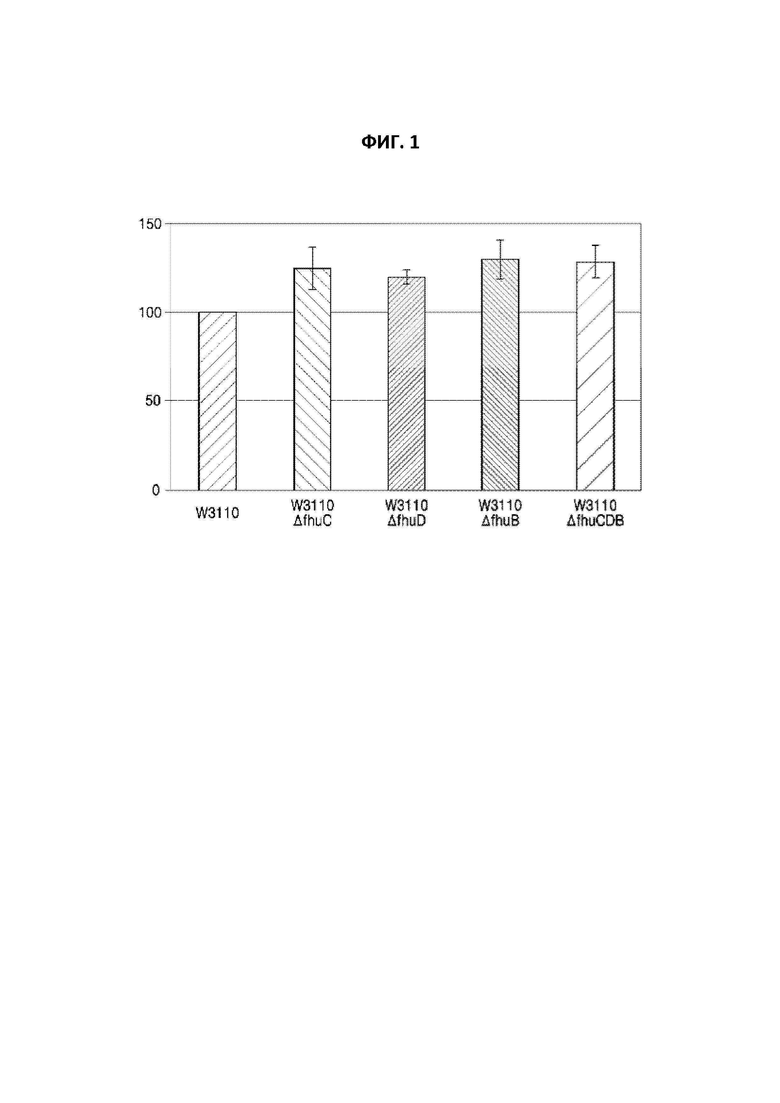
На фиг. 1 показаны уровни внутриклеточного ATP у E. coli в соответствии с конкретным вариантом осуществления настоящей заявки по сравнению с немодифицированным штаммом;
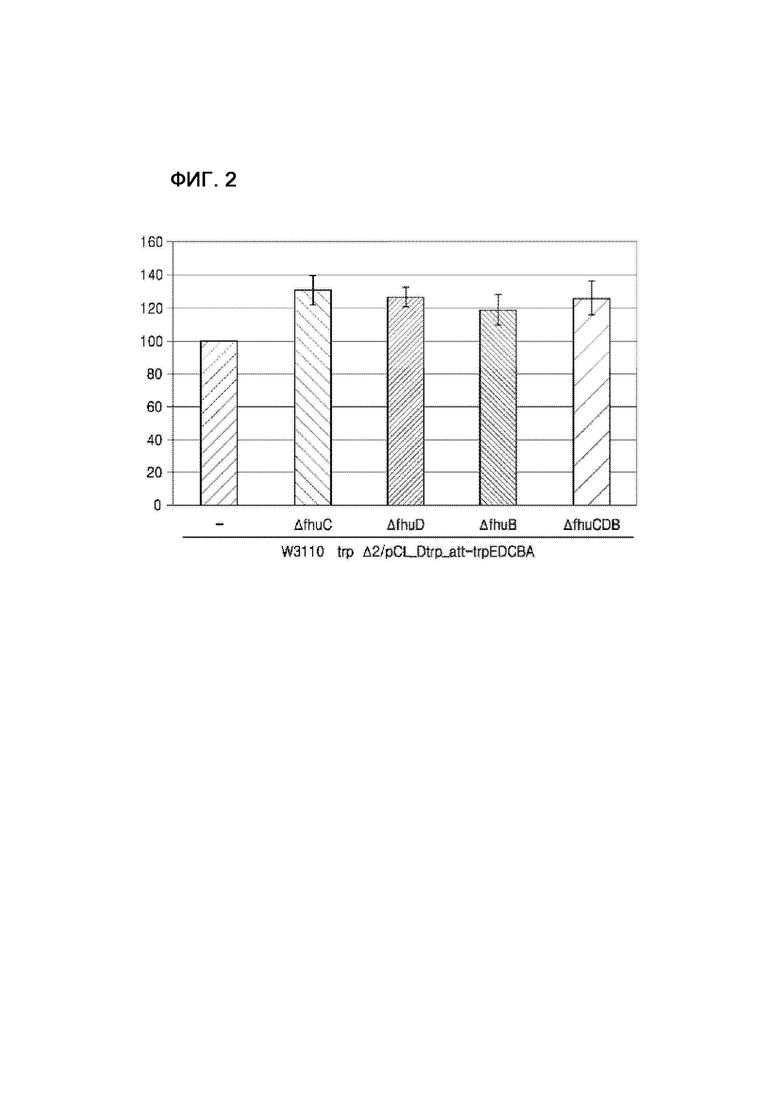
на фиг. 2 показаны уровни внутриклеточного ATP у происходящей от дикого типа E. coli, обладающей производительностью в отношении L-триптофана, в соответствии с конкретным вариантом осуществления настоящей заявки по сравнению с немодифицированным штаммом;
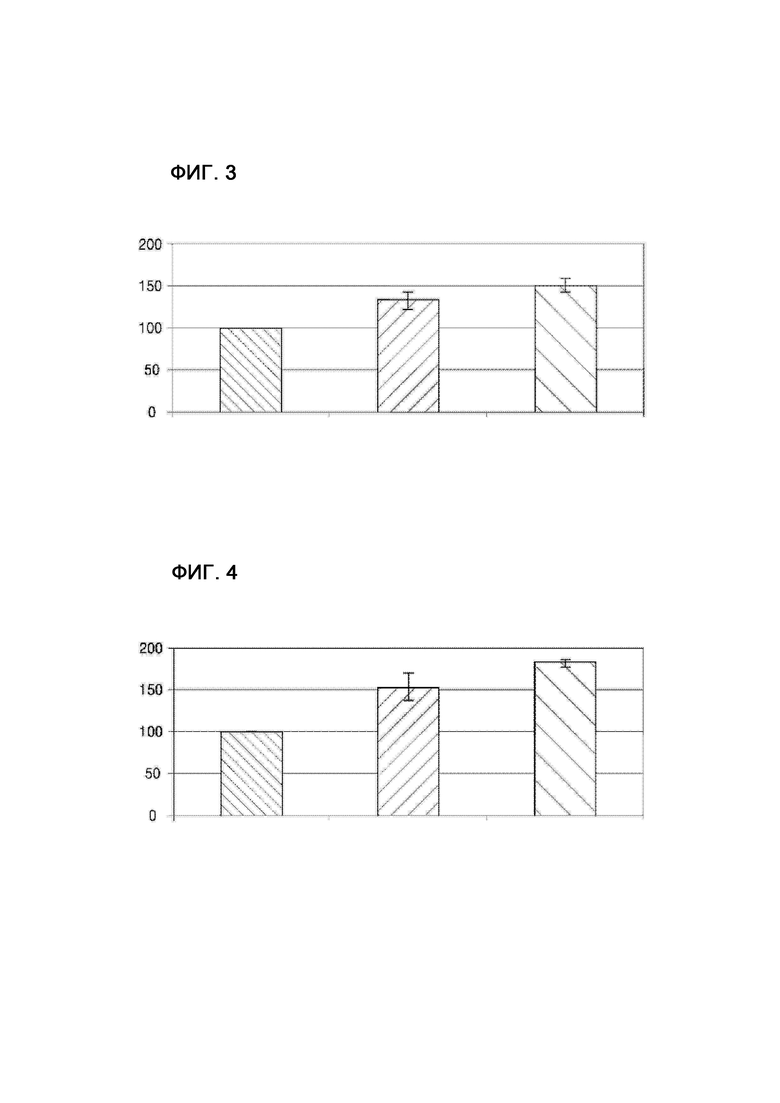
на фиг. 3 показаны уровни внутриклеточного ATP у E. coli, обладающей производительностью в отношении L-треонина, в соответствии с конкретным вариантом осуществления настоящей заявки по сравнению с немодифицированным штаммом;
на фиг. 4 показаны уровни внутриклеточного ATP у E. coli, обладающей производительностью в отношении L-триптофана, в соответствии с конкретным вариантом осуществления настоящей заявки по сравнению с немодифицированным штаммом;
на фиг. 5 показана производительность в отношении L-треонина у E. coli, обладающей производительностью в отношении L-треонина, в соответствии с конкретным вариантом осуществления настоящей заявки по сравнению с немодифицированным штаммом; и
на фиг. 6 показана производительность в отношении L-триптофана у E. coli, обладающей производительностью в отношении L-триптофана, в соответствии с конкретным вариантом осуществления настоящей заявки по сравнению с немодифицированным штаммом.
ПРИНЦИП ИЗОБРЕТЕНИЯ
Далее настоящая заявка будет описана более подробно со ссылками на примеры. Однако данные примеры приведены только в целях иллюстрации, и ограничение объема настоящей заявки данными примерами не подразумевается.
Пример 1. Получение E. coli W3110 дикого типа с инактивированными белками, кодируемыми генами fhuC , fhuD и fhuB
В этом примере гены fhuC, fhuD и fhuB E. coli W3110 дикого типа (ATCC® 39936™) соответственно подвергали делеции путем гомологичной рекомбинации.
Гены fhuC, fhuD и fhuB, подлежащие делеции, соответственно имеют нуклеотидные последовательности SEQ ID NO: 1, 2 и 3, и эти гены существуют в форме оперона SEQ ID NO: 4.
Для делеции fhuC, fhuD и fhuB осуществляли одностадийную инактивацию с помощью рекомбиназы Red фага лямбда, разработанную Datsenko KA et al. (Proc Natl Acad Sci USA., (2000) 97:6640-6645). В качестве маркера для подтверждения вставки в ген использовали ген устойчивости к хлорамфениколу из pUCprmfmloxC, который получали путем лигирования промотора rmf с pUC19 (New England Biolabs (USA)) и лигирования с ним мутантной кассеты loxP-CmR-loxP, полученной из pACYC184 (New England Biolab) (заявка на корейский патент № 2009-0075549).
Вначале осуществляли первичную полимеразную цепную реакцию (далее в данном документе называемую ‘ПЦР’) с применением pUCprmfmloxC в качестве матрицы и комбинаций праймеров с SEQ ID NO: 8 и 9, 10 и 11, 12 и 13, и 8 и 13 с частью генов fhuC и fhuB и частичной последовательностью гена устойчивости к хлорамфениколу в гене pUCprmfmloxC в условиях 30 циклов денатурации при 94°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и элонгации при 72°C в течение 1 минуты с получением таким образом продуктов ПЦР ΔfhuC1st, ΔfhuD1st, ΔfhuB1st и ΔfhuCDB1st размером приблизительно 1,2 т.п.о.
После этого продукты ПЦР ΔfhuC1st, ΔfhuD1st, ΔfhuB1st и ΔfhuCDB1st размером 1,2 т.п.о., полученные посредством ПЦР, подвергали электрофорезу в 0,8% агарозном геле, а затем элюировали и применяли в качестве матрицы для вторичной ПЦР. Вторичную ПЦР осуществляли с применением элюированных продуктов первичной ПЦР в качестве матриц и комбинаций праймеров с SEQ ID NO: 14 и 15, 16 и 17, 18 и 19, 14 и 19, содержащих нуклеотидные последовательности размером 20 п.о. в 5'- и 3'-концевых областях продуктов ПЦР, полученных в первичной ПЦР, в условиях 30 циклов денатурации при 94°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и элонгации при 72°C в течение 1 минуты с получением таким образом продуктов ПЦР ΔfhuC, ΔfhuD, ΔfhuB и ΔfhuCDB размером приблизительно 1,3 т.п.о. Полученные таким образом продукты ПЦР подвергали электрофорезу в 0,8% агарозном геле, а затем элюировали и применяли в рекомбинации.
E. coli W3110, который трансформировали вектором pKD46 в соответствии со способом одностадийной инактивации, разработанным Datsenko KA et al. (Proc Natl Acad Sci USA., (2000) 97:6640-6645), получали в виде компетентного штамма, и трансформацию осуществляли путем введения фрагмента гена размером 1,3 т.п.о., полученного посредством первичной и вторичной ПЦР. Штаммы культивировали в среде LB, дополненной хлорамфениколом, и отбирали трансформантов, обладающих устойчивостью к хлорамфениколу. Делецию любого или всех из fhuC, fhuD и fhuB подтверждали по продуктам ПЦР размером приблизительно 4,4 т.п.о., приблизительно 4,3 т.п.о., приблизительно 3,3 т.п.о. и приблизительно 1,6 т.п.о., которые амплифицировали посредством ПЦР с применением геномов, полученных из отобранных штаммов, в качестве матриц и праймеров с SEQ ID NO: 20 и 21.
После удаления pKD46 из полученных таким образом первичных рекомбинантных штаммов, обладающих устойчивостью к хлорамфениколу, в первичные рекомбинантные штаммы, обладающие устойчивостью к хлорамфениколу, вводили вектор pJW168 (Gene, (2000) 247,255-264) для удаления маркерного гена устойчивости к хлорамфениколу из штаммов (Gene, (2000) 247, 255-264). Осуществляли ПЦР с применением праймеров с SEQ ID NO: 20 и 21 с получением продуктов ПЦР размером приблизительно 3,4 т.п.о., приблизительно 3,3 т.п.о., приблизительно 2,2 т.п.о. и приблизительно 0,6 т.п.о., что указывает на то, что полученные в конечном счете штаммы имели делецию любого или всех из генов fhuC, fhuD и fhuB. Штаммы соответственно обозначали как E. coli W3110_ΔfhuC, W3110_ΔfhuD, W3110_ΔfhuB и W3110_ΔfhuCDB.
Пример 2. Измерение уровней внутриклеточного ATP у E. coli с делециями генов fhuC , fhuD и fhuB , происходящих от E. coli дикого типа
В этом примере на практике измеряли уровни внутриклеточного ATP у штаммов, полученных в примере 1.
Для этой цели использовали “эффективный способ количественного определения синтетической активности клеточного ATP”, разработанный Kiyotaka Y. Hara et al., в котором использовали люциферазу (J Biom Scre, (2006) Vol. 11, No.3, pp310-17). Вкратце, E. coli W3110, представляющий собой немодифицированный штамм, применяемый в примере 1, и E. coli W3110_ΔfhuCDB, полученный посредством делеции генов, соответственно культивировали в течение ночи в жидкой среде LB, содержащей глюкозу. После культивирования образцы надосадочной жидкости удаляли путем центрифугирования, полученные таким образом клетки промывали с помощью 100 мМ Tris-Cl (pH 7,5) и затем обрабатывали PB-буфером (проникающий буфер: 40% [об./об.] глюкозы, 0,8% [об./об.] Triton X-100) в течение 30 минут для высвобождения внутриклеточного ATP из клеток. Затем образцы надосадочной жидкости удаляли путем центрифугирования, и к клеткам добавляли люциферин в качестве субстрата для люциферазы. Клеткам позволяли прореагировать в течение 10 минут. Формирование цвета под действием люциферазы измеряли с помощью люминометра для количественного определения уровней ATP. Результаты приведены на фиг. 1. Результаты на фиг. 1 регистрировали как среднее значение для трех повторяемых экспериментов.
Как показано на фиг. 1, уровни внутриклеточного ATP у E. coli W3110_ΔfhuC, W3110_ΔfhuD, W3110_ΔfhuB и W3110_ΔfhuCDB, полученных в примере 1, у которых любой или все из fhuC, fhuD и fhuB, происходящих от E. coli дикого типа, были подвергнуты делеции, были повышены по сравнению с немодифицированным штаммом E. coli W3110.
Пример 3. Получение происходящего от дикого типа штамма, продуцирующего L-триптофан, с инактивированными белками, кодируемыми генами fhuC , fhuD и fhuB , и измерение уровней внутриклеточного ATP
В этом примере любой или все из генов fhuC, fhuD и fhuB происходящего от дикого типа штамма E. coli W3110 trpΔ2/pCL-Dtrp_att-trpEDCBA, продуцирующего L-триптофан (публикация корейского патента № 10-2013-0082121), подвергали делеции путем гомологичной рекомбинации, как в примере 1, с получением штаммов W3110 trpΔ2_ΔfhuC/pCL-Dtrp_att-trpEDCBA, W3110 trpΔ2_ΔfhuD/pCL-Dtrp_att-trpEDCBA, W3110 trpΔ2_ΔfhuB/pCL-Dtrp_att-trpEDCBA и W3110 trpΔ2_ΔfhuCDB/pCL-Dtrp_att-trpEDCBA. У этих штаммов, полученных таким образом, уровни внутриклеточного ATP измеряли таким же образом, как в примере 2, и результаты приведены на фиг. 2.
Как показано на фиг. 2, уровни внутриклеточного ATP у штаммов, которые получали посредством делеции любого или всех из генов fhuC, fhuD и fhuB штамма дикого типа, продуцирующего L-триптофан, были повышены по сравнению с немодифицированным штаммом и контрольным штаммом.
Пример 4. Изучение титра происходящего от дикого типа штамма, продуцирующего L-триптофан, с инактивированными белками, кодируемыми генами fhuC , fhuD и fhuB
Как описано в примере 3, происходящий от дикого типа штамм W3110 trpΔ2/pCL-Dtrp_att-trpEDCBA, продуцирующий L-триптофан, и штаммы с улучшенными уровнями внутриклеточного ATP, полученные посредством делеции любого или всех из генов fhuC, fhuD и fhuB, подвергали титрованию с применением глюкозы в качестве источника углерода.
Каждый из штаммов инокулировали платиновой петлей в твердую среду LB и культивировали в инкубаторе при 37°C в течение ночи, а затем инокулировали платиновой петлей в 25 мл глюкозосодержащей титрационной среды, имеющей состав из таблицы 1. Затем штаммы культивировали в инкубаторе при 37°C и при 200 об./мин. в течение 48 часов. Результаты приведены в таблице 2. Все результаты регистрировали как среднее значение для трех повторяемых экспериментов.
[Таблица 1]
[Таблица 2]
* измерено через 48 часов
Как показано в таблице 2, было продемонстрировано, что у штаммов с улучшенными уровнями внутриклеточного ATP, полученных в примере 3 посредством делеции любого или всех из генов fhuC, fhuD и fhuB штамма дикого типа W3110 trpΔ2/pCL-Dtrp_att-trpEDCBA, продуцирующего L-триптофан, продуцирование L-триптофана повысилось на величину до приблизительно 63% по сравнению с немодифицированным штаммом trpΔ2/pCL-Dtrp_att-trpEDCBA. С учетом уровней внутриклеточного ATP, подтвержденных на фиг. 2, эти результаты по повышенным уровням внутриклеточного ATP указывают на то, что значения производительности штаммов в отношении L-триптофана были повышены.
Пример 5. Получение штамма, продуцирующего L-треонин, и штамма, продуцирующего L-триптофан, с инактивированными белками, кодируемыми генами fhuC , fhuD и fhuB
В этом примере гены fhuC, fhuD и fhuB штамма KCCM10812P, продуцирующего L-триптофан (корейский патент № 0792095), и штамма KCCM10541, продуцирующего L-треонин (корейский патент № 0576342), соответственно подвергали делеции путем гомологичной рекомбинации, как в примере 1.
Немодифицированный штамм E. coli KCCM10812P, обладающий производительностью в отношении L-триптофана, представляет собой штамм, происходящий от варианта E. coli (KFCC 10066), обладающего производительностью в отношении L-фенилаланина, и представляет собой рекомбинантный штамм E. coli, обладающий производительностью в отношении L-триптофана, характеризующийся тем, что хромосомная ауксотрофность по триптофану была уменьшена или устранена, гены pheA, trpR, mtr и tnaAB были подвергнуты аттенюации, а гены aroG и trpE были модифицированы.
Также, немодифицированный штамм E. coli KCCM10541P, обладающий производительностью в отношении L-треонина, представляет собой штамм, происходящий от E. coli KFCC10718 (публикация корейского патента № 1992-0008365), и представляет собой E. coli, обладающую устойчивостью к аналогу L-метионина, фенотипом ауксотрофности по метионину, устойчивостью к аналогу L-треонина, "растекающимся" фенотипом ауксотрофности по изолейцину, устойчивостью к аналогу L-лизина и устойчивостью к α-аминомасляной кислоте, а также производительностью в отношении L-треонина.
Гены fhuC, fhuD и fhuB, подлежащие делеции, подвергали делеции у E. coli KCCM10812P и E. coli KCCM10541P таким же образом, как в примере 1, соответственно. В результате этого получали штамм KCCM10541_ΔfhuCDB, продуцирующий L-треонин, и штамм KCCM10812P_ΔfhuCDB, продуцирующий L-триптофан.
Пример 6. Измерение уровней ATP у штамма, продуцирующего L-треонин, и штамма, продуцирующего L-триптофан, с инактивированными белками, кодируемыми генами fhuC , fhuD и fhuB
В этом примере на практике измеряли уровни внутриклеточного ATP у штаммов, полученных в примере 5.
Уровни внутриклеточного ATP измеряли таким же образом, как в примере 2. Результаты приведены на фиг. 3 и 4. Результаты из фиг. 3 и 4 регистрировали как среднее значение для трех повторяемых экспериментов. В качестве контрольных групп использовали штамм, продуцирующий L-треонин (E. coli KCCM10541P_ΔysaΔydaS), и штамм, продуцирующий L-триптофан (E. coli KCCM10812P_ΔysaΔydaS), с делециями ysa и ydaS, которые, как известно, обладают более высокими уровнями внутриклеточного ATP, чем немодифицированные штаммы E. coli KCCM10812P и E. coli KCCM10541P, используемые в примере 3 (корейский патент № 1327093).
Как показано на фиг. 3 и 4, штаммы с делециями fhuC, fhuD и fhuB, полученные из штамма, продуцирующего L-треонин, и штамма, продуцирующего L-триптофан, в примере 3, продемонстрировали повышенные уровни внутриклеточного ATP по сравнению с немодифицированными штаммами и контрольными штаммами.
Пример 7. Изучение титра штамма, продуцирующего L-треонин, с инактивированными белками, кодируемыми генами fhuC , fhuD и fhuB
Как описано в примере 5, штаммы с улучшенными уровнями внутриклеточного ATP, которые получали посредством делеции генов fhuC, fhuD и fhuB у микроорганизма, продуцирующего L-треонин, E. coli KCCM10541P (корейский патент № 0576342), подвергали титрованию с применением глюкозы в качестве источника углерода. Штамм, продуцирующий L-треонин, с делециями ysa и ydaS (E. coli KCCM10541P_ΔysaΔydaS) применяли в качестве контрольной группы для сравнения результатов титрования.
Каждый из штаммов культивировали в твердой среде LB в инкубаторе при 33°C в течение ночи, а затем инокулировали платиновой петлей в 25 мл глюкозосодержащей титрационной среды, имеющей состав из таблицы 3. Затем штаммы культивировали в инкубаторе при 33°C и при 200 об./мин. в течение 50 часов. Результаты приведены в таблице 4 и на фиг. 5. Все результаты регистрировали как среднее значение для трех повторяемых экспериментов.
[Таблица 3]
[Таблица 4]
* измерено через 30 часов
** измерено через 50 часов
Как показано в таблице 4, было продемонстрировано, что рекомбинантный штамм E. coli, продуцирующий L-треонин, полученный в соответствии с настоящей заявкой, демонстрировал физиологическую активность, сходную с таковой у немодифицированного штамма, и характеризовался продуцированием L-треонина, повышенным на величину до приблизительно 9% по сравнению с немодифицированным штаммом. С учетом уровней внутриклеточного ATP, подтвержденных на фиг. 3, эти результаты по повышенным уровням внутриклеточного ATP указывают на то, что значения производительности штаммов в отношении L-треонина были повышены.
Пример 8. Изучение титра штамма, продуцирующего L-триптофан, с инактивированными белками, кодируемыми генами fhuC , fhuD и fhuB
Как описано в примере 5, штаммы с улучшенными уровнями внутриклеточного ATP, которые получали посредством делеции генов fhuC, fhuD и fhuB у микроорганизма, продуцирующего L-триптофан, KCCM10812P (корейский патент № 0792095), подвергали титрованию с применением глюкозы в качестве источника углерода. Штамм, продуцирующий L-триптофан, с делециями ysa и ydaS (E. coli KCCM10812P_ΔysaΔydaS) применяли в качестве контрольной группы для оценивания титра таким же образом, как в примере 4.
Результаты титрования приведены в таблице 5 и на фиг. 6. Все результаты регистрировали как среднее значение для трех повторяемых экспериментов.
[Таблица 5]
* измерено через 33 часа
** измерено через 48 часов
Как показано в таблице 5, было продемонстрировано, что рекомбинантный штамм E. coli, продуцирующий L-триптофан, полученный в соответствии с настоящей заявкой, демонстрировал физиологическую активность, сходную с таковой у немодифицированного штамма, и характеризовался продуцированием L-триптофана, повышенным на величину до приблизительно 30% по сравнению с немодифицированным штаммом. С учетом уровней внутриклеточного ATP, подтвержденных на фиг. 4, эти результаты по повышенным уровням внутриклеточного ATP указывают на то, что значения производительности штаммов в отношении L-триптофана были повышены.
Рекомбинантный штамм CA04-2801 (KCCM10812P_ΔfhuCDB) согласно настоящей заявке был депонирован в Корейском центре культур микроорганизмов, международном органе по депонированию, 15 ноября 2013 г. под № доступа KCCM11474P.
На основании вышеприведенного описания специалистам в данной области будет понятно, что настоящую заявку можно реализовать в другой конкретной форме без изменения ее технической сущности или существенных характеристик. Таким образом, следует понимать, что вышеописанный вариант осуществления не является ограничивающим, но является иллюстративным во всех аспектах. Объем настоящей заявки определен прилагаемой формулой изобретения, а не предшествующим ей описанием, и, следовательно, предполагается, что все изменения и модификации, которые находятся в пределах формулы изобретения, или эквиваленты таких пределов охватываются, таким образом, формулой изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CHEILJEDANG CORP.
<120> МИКРООРГАНИЗМ, ОБЛАДАЮЩИЙ УЛУЧШЕННЫМ УРОВНЕМ ВНУТРИКЛЕТОЧНОЙ ЭНЕРГИИ, И
СПОСОБ ПРОДУЦИРОВАНИЯ L-АМИНОКИСЛОТЫ С ЕГО ПОМОЩЬЮ
<130> PX048855
<150> KR14/62593
<151> 2014-05-23
<160> 21
<170> PatentIn, версия 3.5
<210> 1
<211> 798
<212> ДНК
<213> Escherichia coli
<400> 1
atgcaggaat acacgaatca ttccgatacc acttttgcac tgcgtaatat ctcctttcgt 60
gtgcccgggc gcacgctttt gcatccgctg tcgttaacct ttcctgccgg gaaagtgacc 120
ggtctgattg gtcacaacgg ttctggtaaa tccactctgc tcaaaatgct tggccgtcat 180
cagccgccgt cggaagggga gattcttctt gatgcccaac cgctggaaag ctggagcagc 240
aaagcgtttg cccgcaaagt ggcttatttg ccgcagcagc ttcctccggc agaagggatg 300
accgtgcgtg aactggtggc gattggtcgt tacccgtggc atggcgcgct ggggcgcttt 360
ggggcggcag atcgcgaaaa agtcgaggaa gctatctcgc tggttggctt aaaaccgctg 420
gcgcatcggc tggtcgatag tctctctggc ggcgaacgtc agcgggcgtg gatcgccatg 480
ctggtggcgc aggatagccg ttgtctgttg ctcgacgaac cgacctcggc gctggatatc 540
gcccaccagg ttgatgtgct gtcgctggtg caccgtttaa gtcaggagcg tggcctgacg 600
gtcattgccg tgttgcacga tatcaatatg gcggcacgct actgtgatta tctggtcgcc 660
ctgcgcggcg gtgaaatgat tgctcaggga acgcctgcgg aaattatgcg cggcgaaacc 720
ctcgaaatga tttatggcat cccgatgggt attttgccgc atccggcggg tgctgcacct 780
gtgagttttg tttattga 798
<210> 2
<211> 891
<212> ДНК
<213> Escherichia coli
<400> 2
atgagcggct tacctcttat ttcgcgccgt cgactgttaa cggcgatggc gctttctccg 60
ttgttatggc agatgaatac cgcccacgcg gcggctattg atcccaatcg tattgtggcg 120
ctggagtggt tgccggtgga attactgctg gcgctcggca tcgtgcctta cggcgtggcg 180
gataccatca actatcgcct gtgggtcagc gaaccaccat tgccggactc agtgatcgac 240
gtcggtttgc gcacagaacc taaccttgaa ctgctgaccg aaatgaaacc atcgtttatg 300
gtctggtcgg caggatatgg cccttcacca gaaatgctgg ctcgtattgc gccgggtcgc 360
ggatttaact tcagtgacgg caaacagccg ttggcgatgg cgcgtaaatc gctgacggaa 420
atggcagatt tacttaacct gcaaagcgca gcggaaacgc atttagcgca atatgaagac 480
tttatccgca gcatgaaacc ccgctttgtg aagcgtggtg cgcgtccgtt attgctgacg 540
acgcttatcg atccgcgcca tatgctggtc ttcggtccaa acagcttgtt ccaggaaatt 600
cttgatgagt acggcatccc aaatgcctgg caaggggaaa ccaacttctg gggcagtacc 660
gccgtcagta tcgatcgtct ggcggcgtat aaagacgttg atgtgctctg ttttgatcac 720
gacaacagca aagacatgga tgcgctaatg gcaacgccgc tgtggcaggc catgccgttt 780
gtccgcgccg gacgctttca gcgcgtacct gcagtctggt tttatggtgc gacgctctcg 840
gcaatgcact ttgtgcgcgt tctggataac gccatcggag gtaaagcgtg a 891
<210> 3
<211> 1983
<212> ДНК
<213> Escherichia coli
<400> 3
gtgagtaaac gaattgcgct tttcccggcg ttattgctgg cgctgttagt gattgtcgct 60
acggcgctca cctggatgaa cttctcgcag gcgctgccgc gtagccagtg ggcgcaggct 120
gcctggtcgc cggatattga cgtcatcgag cagatgattt ttcactacag cttgttgccg 180
cgtctggcga tttcgctgct ggtgggcgcg ggtctggggc tggtgggcgt gctgtttcag 240
caagtgctgc gtaacccgct ggcggagccg acgacgcttg gcgttgctac aggcgcgcaa 300
ctggggatta ccgtcactac gctctgggcg atccctggtg cgatggcgag ccagtttgct 360
gcgcaggcag gggcttgtgt tgttggctta attgtctttg gcgtcgcgtg ggggaaacgg 420
ctgtcgccgg taacgctgat tctcgcgggg ttggtagtga gcctttattg cggcgcaatc 480
aatcagttac tggttatctt ccatcatgac caactgcaaa gcatgtttct gtggagcact 540
ggaacgctga cgcaaaccga ctggggcggc gttgagcgtt tatggccgca gctgctgggc 600
ggtgtgatgc tgacgttgct gctacttcgt ccgttaaccc tgatggggct tgatgatggc 660
gtggcgcgca atctcgggct ggccttgtcg cttgcgcgtc tggcagcgct gtcgctggcg 720
attgtcatca gtgcgctgct ggtgaacgct gtggggatta tcggctttat cgggttgttc 780
gcgccgctgc tggcaaaaat gctgggggcg cggcgtctgc tgccacgact gatgctggcg 840
tcgttgattg gtgcgctgat cctctggctt tccgatcaaa tcatcctctg gctgactcgc 900
gtgtggatgg aagtgtccac cggttcggtc actgcgttga tcggtgcgcc gctgctactg 960
tggctgttgc cgcgtttacg cagcattagc gcgccggata tgaaggtcaa cgatcgtgtc 1020
gcggctgaac gccaacatgt gctggcgttt gccctcgcgg gcggcgtgct gctgttgatg 1080
gctgtggtgg tggcgctgtc gtttggtcgt gatgcgcacg gctggacgtg ggcgagcggg 1140
gcgttgctcg aggatttaat gccctggcgc tggccgcgaa ttatggcggc gctgtttgcg 1200
ggcgtcatgc tggcggtggc gggctgtatt attcagcgac tgaccggaaa cccgatggca 1260
agcccggaag tgctggggat tagctccggc gcggcgtttg gcgtggtgtt gatgctgttt 1320
ctggtgccgg gtaatgcctt tggctggctg ttacctgcag gcagtctcgg cgcggcggtg 1380
acgctgttga tcattatgat cgccgccggc cgcggtggat tttccccaca ccgtatgtta 1440
ctggcgggga tggcgttaag caccgcgttc accatgcttt tgatgatgtt gcaggcaagt 1500
ggtgacccgc gaatggcgca agtgctgacc tggatttccg gttcgaccta caacgcgacc 1560
gatgcgcagg tctggcgcac cggaattgtg atggtgattt tgctggcgat taccccgctg 1620
tgccgccgct ggctgaccat tttaccgctg ggtggtgata ccgcccgagc cgtaggaatg 1680
gcgctgacgc cgacgcgaat tgcgctgctg ctgttagcgg cttgcctgac ggcgaccgcg 1740
acgatgacta ttggaccgtt gagttttgtt ggtttaatgg caccgcatat tgcgcggatg 1800
atgggctttc gacggacgat gccacacatc gtaatttcgg cgctggtggg tggtttactg 1860
ctggtgttcg ctgactggtg tgggcggatg gtgctgtttc cattccagat cccggcgggg 1920
ctgctgtcaa cctttatcgg cgcgccatat tttatctatt tgttgagaaa gcagagccgt 1980
taa 1983
<210> 4
<211> 3667
<212> ДНК
<213> Escherichia coli
<400> 4
atgcaggaat acacgaatca ttccgatacc acttttgcac tgcgtaatat ctcctttcgt 60
gtgcccgggc gcacgctttt gcatccgctg tcgttaacct ttcctgccgg gaaagtgacc 120
ggtctgattg gtcacaacgg ttctggtaaa tccactctgc tcaaaatgct tggccgtcat 180
cagccgccgt cggaagggga gattcttctt gatgcccaac cgctggaaag ctggagcagc 240
aaagcgtttg cccgcaaagt ggcttatttg ccgcagcagc ttcctccggc agaagggatg 300
accgtgcgtg aactggtggc gattggtcgt tacccgtggc atggcgcgct ggggcgcttt 360
ggggcggcag atcgcgaaaa agtcgaggaa gctatctcgc tggttggctt aaaaccgctg 420
gcgcatcggc tggtcgatag tctctctggc ggcgaacgtc agcgggcgtg gatcgccatg 480
ctggtggcgc aggatagccg ttgtctgttg ctcgacgaac cgacctcggc gctggatatc 540
gcccaccagg ttgatgtgct gtcgctggtg caccgtttaa gtcaggagcg tggcctgacg 600
gtcattgccg tgttgcacga tatcaatatg gcggcacgct actgtgatta tctggtcgcc 660
ctgcgcggcg gtgaaatgat tgctcaggga acgcctgcgg aaattatgcg cggcgaaacc 720
ctcgaaatga tttatggcat cccgatgggt attttgccgc atccggcggg tgctgcacct 780
gtgagttttg tttattgatg agcggcttac ctcttatttc gcgccgtcga ctgttaacgg 840
cgatggcgct ttctccgttg ttatggcaga tgaataccgc ccacgcggcg gctattgatc 900
ccaatcgtat tgtggcgctg gagtggttgc cggtggaatt actgctggcg ctcggcatcg 960
tgccttacgg cgtggcggat accatcaact atcgcctgtg ggtcagcgaa ccaccattgc 1020
cggactcagt gatcgacgtc ggtttgcgca cagaacctaa ccttgaactg ctgaccgaaa 1080
tgaaaccatc gtttatggtc tggtcggcag gatatggccc ttcaccagaa atgctggctc 1140
gtattgcgcc gggtcgcgga tttaacttca gtgacggcaa acagccgttg gcgatggcgc 1200
gtaaatcgct gacggaaatg gcagatttac ttaacctgca aagcgcagcg gaaacgcatt 1260
tagcgcaata tgaagacttt atccgcagca tgaaaccccg ctttgtgaag cgtggtgcgc 1320
gtccgttatt gctgacgacg cttatcgatc cgcgccatat gctggtcttc ggtccaaaca 1380
gcttgttcca ggaaattctt gatgagtacg gcatcccaaa tgcctggcaa ggggaaacca 1440
acttctgggg cagtaccgcc gtcagtatcg atcgtctggc ggcgtataaa gacgttgatg 1500
tgctctgttt tgatcacgac aacagcaaag acatggatgc gctaatggca acgccgctgt 1560
ggcaggccat gccgtttgtc cgcgccggac gctttcagcg cgtacctgca gtctggtttt 1620
atggtgcgac gctctcggca atgcactttg tgcgcgttct ggataacgcc atcggaggta 1680
aagcgtgagt aaacgaattg cgcttttccc ggcgttattg ctggcgctgt tagtgattgt 1740
cgctacggcg ctcacctgga tgaacttctc gcaggcgctg ccgcgtagcc agtgggcgca 1800
ggctgcctgg tcgccggata ttgacgtcat cgagcagatg atttttcact acagcttgtt 1860
gccgcgtctg gcgatttcgc tgctggtggg cgcgggtctg gggctggtgg gcgtgctgtt 1920
tcagcaagtg ctgcgtaacc cgctggcgga gccgacgacg cttggcgttg ctacaggcgc 1980
gcaactgggg attaccgtca ctacgctctg ggcgatccct ggtgcgatgg cgagccagtt 2040
tgctgcgcag gcaggggctt gtgttgttgg cttaattgtc tttggcgtcg cgtgggggaa 2100
acggctgtcg ccggtaacgc tgattctcgc ggggttggta gtgagccttt attgcggcgc 2160
aatcaatcag ttactggtta tcttccatca tgaccaactg caaagcatgt ttctgtggag 2220
cactggaacg ctgacgcaaa ccgactgggg cggcgttgag cgtttatggc cgcagctgct 2280
gggcggtgtg atgctgacgt tgctgctact tcgtccgtta accctgatgg ggcttgatga 2340
tggcgtggcg cgcaatctcg ggctggcctt gtcgcttgcg cgtctggcag cgctgtcgct 2400
ggcgattgtc atcagtgcgc tgctggtgaa cgctgtgggg attatcggct ttatcgggtt 2460
gttcgcgccg ctgctggcaa aaatgctggg ggcgcggcgt ctgctgccac gactgatgct 2520
ggcgtcgttg attggtgcgc tgatcctctg gctttccgat caaatcatcc tctggctgac 2580
tcgcgtgtgg atggaagtgt ccaccggttc ggtcactgcg ttgatcggtg cgccgctgct 2640
actgtggctg ttgccgcgtt tacgcagcat tagcgcgccg gatatgaagg tcaacgatcg 2700
tgtcgcggct gaacgccaac atgtgctggc gtttgccctc gcgggcggcg tgctgctgtt 2760
gatggctgtg gtggtggcgc tgtcgtttgg tcgtgatgcg cacggctgga cgtgggcgag 2820
cggggcgttg ctcgaggatt taatgccctg gcgctggccg cgaattatgg cggcgctgtt 2880
tgcgggcgtc atgctggcgg tggcgggctg tattattcag cgactgaccg gaaacccgat 2940
ggcaagcccg gaagtgctgg ggattagctc cggcgcggcg tttggcgtgg tgttgatgct 3000
gtttctggtg ccgggtaatg cctttggctg gctgttacct gcaggcagtc tcggcgcggc 3060
ggtgacgctg ttgatcatta tgatcgccgc cggccgcggt ggattttccc cacaccgtat 3120
gttactggcg gggatggcgt taagcaccgc gttcaccatg cttttgatga tgttgcaggc 3180
aagtggtgac ccgcgaatgg cgcaagtgct gacctggatt tccggttcga cctacaacgc 3240
gaccgatgcg caggtctggc gcaccggaat tgtgatggtg attttgctgg cgattacccc 3300
gctgtgccgc cgctggctga ccattttacc gctgggtggt gataccgccc gagccgtagg 3360
aatggcgctg acgccgacgc gaattgcgct gctgctgtta gcggcttgcc tgacggcgac 3420
cgcgacgatg actattggac cgttgagttt tgttggttta atggcaccgc atattgcgcg 3480
gatgatgggc tttcgacgga cgatgccaca catcgtaatt tcggcgctgg tgggtggttt 3540
actgctggtg ttcgctgact ggtgtgggcg gatggtgctg tttccattcc agatcccggc 3600
ggggctgctg tcaaccttta tcggcgcgcc atattttatc tatttgttga gaaagcagag 3660
ccgttaa 3667
<210> 5
<211> 265
<212> БЕЛОК
<213> Escherichia coli
<400> 5
Met Gln Glu Tyr Thr Asn His Ser Asp Thr Thr Phe Ala Leu Arg Asn
1 5 10 15
Ile Ser Phe Arg Val Pro Gly Arg Thr Leu Leu His Pro Leu Ser Leu
20 25 30
Thr Phe Pro Ala Gly Lys Val Thr Gly Leu Ile Gly His Asn Gly Ser
35 40 45
Gly Lys Ser Thr Leu Leu Lys Met Leu Gly Arg His Gln Pro Pro Ser
50 55 60
Glu Gly Glu Ile Leu Leu Asp Ala Gln Pro Leu Glu Ser Trp Ser Ser
65 70 75 80
Lys Ala Phe Ala Arg Lys Val Ala Tyr Leu Pro Gln Gln Leu Pro Pro
85 90 95
Ala Glu Gly Met Thr Val Arg Glu Leu Val Ala Ile Gly Arg Tyr Pro
100 105 110
Trp His Gly Ala Leu Gly Arg Phe Gly Ala Ala Asp Arg Glu Lys Val
115 120 125
Glu Glu Ala Ile Ser Leu Val Gly Leu Lys Pro Leu Ala His Arg Leu
130 135 140
Val Asp Ser Leu Ser Gly Gly Glu Arg Gln Arg Ala Trp Ile Ala Met
145 150 155 160
Leu Val Ala Gln Asp Ser Arg Cys Leu Leu Leu Asp Glu Pro Thr Ser
165 170 175
Ala Leu Asp Ile Ala His Gln Val Asp Val Leu Ser Leu Val His Arg
180 185 190
Leu Ser Gln Glu Arg Gly Leu Thr Val Ile Ala Val Leu His Asp Ile
195 200 205
Asn Met Ala Ala Arg Tyr Cys Asp Tyr Leu Val Ala Leu Arg Gly Gly
210 215 220
Glu Met Ile Ala Gln Gly Thr Pro Ala Glu Ile Met Arg Gly Glu Thr
225 230 235 240
Leu Glu Met Ile Tyr Gly Ile Pro Met Gly Ile Leu Pro His Pro Ala
245 250 255
Gly Ala Ala Pro Val Ser Phe Val Tyr
260 265
<210> 6
<211> 296
<212> БЕЛОК
<213> Escherichia coli
<400> 6
Met Ser Gly Leu Pro Leu Ile Ser Arg Arg Arg Leu Leu Thr Ala Met
1 5 10 15
Ala Leu Ser Pro Leu Leu Trp Gln Met Asn Thr Ala His Ala Ala Ala
20 25 30
Ile Asp Pro Asn Arg Ile Val Ala Leu Glu Trp Leu Pro Val Glu Leu
35 40 45
Leu Leu Ala Leu Gly Ile Val Pro Tyr Gly Val Ala Asp Thr Ile Asn
50 55 60
Tyr Arg Leu Trp Val Ser Glu Pro Pro Leu Pro Asp Ser Val Ile Asp
65 70 75 80
Val Gly Leu Arg Thr Glu Pro Asn Leu Glu Leu Leu Thr Glu Met Lys
85 90 95
Pro Ser Phe Met Val Trp Ser Ala Gly Tyr Gly Pro Ser Pro Glu Met
100 105 110
Leu Ala Arg Ile Ala Pro Gly Arg Gly Phe Asn Phe Ser Asp Gly Lys
115 120 125
Gln Pro Leu Ala Met Ala Arg Lys Ser Leu Thr Glu Met Ala Asp Leu
130 135 140
Leu Asn Leu Gln Ser Ala Ala Glu Thr His Leu Ala Gln Tyr Glu Asp
145 150 155 160
Phe Ile Arg Ser Met Lys Pro Arg Phe Val Lys Arg Gly Ala Arg Pro
165 170 175
Leu Leu Leu Thr Thr Leu Ile Asp Pro Arg His Met Leu Val Phe Gly
180 185 190
Pro Asn Ser Leu Phe Gln Glu Ile Leu Asp Glu Tyr Gly Ile Pro Asn
195 200 205
Ala Trp Gln Gly Glu Thr Asn Phe Trp Gly Ser Thr Ala Val Ser Ile
210 215 220
Asp Arg Leu Ala Ala Tyr Lys Asp Val Asp Val Leu Cys Phe Asp His
225 230 235 240
Asp Asn Ser Lys Asp Met Asp Ala Leu Met Ala Thr Pro Leu Trp Gln
245 250 255
Ala Met Pro Phe Val Arg Ala Gly Arg Phe Gln Arg Val Pro Ala Val
260 265 270
Trp Phe Tyr Gly Ala Thr Leu Ser Ala Met His Phe Val Arg Val Leu
275 280 285
Asp Asn Ala Ile Gly Gly Lys Ala
290 295
<210> 7
<211> 660
<212> БЕЛОК
<213> Escherichia coli
<400> 7
Met Ser Lys Arg Ile Ala Leu Phe Pro Ala Leu Leu Leu Ala Leu Leu
1 5 10 15
Val Ile Val Ala Thr Ala Leu Thr Trp Met Asn Phe Ser Gln Ala Leu
20 25 30
Pro Arg Ser Gln Trp Ala Gln Ala Ala Trp Ser Pro Asp Ile Asp Val
35 40 45
Ile Glu Gln Met Ile Phe His Tyr Ser Leu Leu Pro Arg Leu Ala Ile
50 55 60
Ser Leu Leu Val Gly Ala Gly Leu Gly Leu Val Gly Val Leu Phe Gln
65 70 75 80
Gln Val Leu Arg Asn Pro Leu Ala Glu Pro Thr Thr Leu Gly Val Ala
85 90 95
Thr Gly Ala Gln Leu Gly Ile Thr Val Thr Thr Leu Trp Ala Ile Pro
100 105 110
Gly Ala Met Ala Ser Gln Phe Ala Ala Gln Ala Gly Ala Cys Val Val
115 120 125
Gly Leu Ile Val Phe Gly Val Ala Trp Gly Lys Arg Leu Ser Pro Val
130 135 140
Thr Leu Ile Leu Ala Gly Leu Val Val Ser Leu Tyr Cys Gly Ala Ile
145 150 155 160
Asn Gln Leu Leu Val Ile Phe His His Asp Gln Leu Gln Ser Met Phe
165 170 175
Leu Trp Ser Thr Gly Thr Leu Thr Gln Thr Asp Trp Gly Gly Val Glu
180 185 190
Arg Leu Trp Pro Gln Leu Leu Gly Gly Val Met Leu Thr Leu Leu Leu
195 200 205
Leu Arg Pro Leu Thr Leu Met Gly Leu Asp Asp Gly Val Ala Arg Asn
210 215 220
Leu Gly Leu Ala Leu Ser Leu Ala Arg Leu Ala Ala Leu Ser Leu Ala
225 230 235 240
Ile Val Ile Ser Ala Leu Leu Val Asn Ala Val Gly Ile Ile Gly Phe
245 250 255
Ile Gly Leu Phe Ala Pro Leu Leu Ala Lys Met Leu Gly Ala Arg Arg
260 265 270
Leu Leu Pro Arg Leu Met Leu Ala Ser Leu Ile Gly Ala Leu Ile Leu
275 280 285
Trp Leu Ser Asp Gln Ile Ile Leu Trp Leu Thr Arg Val Trp Met Glu
290 295 300
Val Ser Thr Gly Ser Val Thr Ala Leu Ile Gly Ala Pro Leu Leu Leu
305 310 315 320
Trp Leu Leu Pro Arg Leu Arg Ser Ile Ser Ala Pro Asp Met Lys Val
325 330 335
Asn Asp Arg Val Ala Ala Glu Arg Gln His Val Leu Ala Phe Ala Leu
340 345 350
Ala Gly Gly Val Leu Leu Leu Met Ala Val Val Val Ala Leu Ser Phe
355 360 365
Gly Arg Asp Ala His Gly Trp Thr Trp Ala Ser Gly Ala Leu Leu Glu
370 375 380
Asp Leu Met Pro Trp Arg Trp Pro Arg Ile Met Ala Ala Leu Phe Ala
385 390 395 400
Gly Val Met Leu Ala Val Ala Gly Cys Ile Ile Gln Arg Leu Thr Gly
405 410 415
Asn Pro Met Ala Ser Pro Glu Val Leu Gly Ile Ser Ser Gly Ala Ala
420 425 430
Phe Gly Val Val Leu Met Leu Phe Leu Val Pro Gly Asn Ala Phe Gly
435 440 445
Trp Leu Leu Pro Ala Gly Ser Leu Gly Ala Ala Val Thr Leu Leu Ile
450 455 460
Ile Met Ile Ala Ala Gly Arg Gly Gly Phe Ser Pro His Arg Met Leu
465 470 475 480
Leu Ala Gly Met Ala Leu Ser Thr Ala Phe Thr Met Leu Leu Met Met
485 490 495
Leu Gln Ala Ser Gly Asp Pro Arg Met Ala Gln Val Leu Thr Trp Ile
500 505 510
Ser Gly Ser Thr Tyr Asn Ala Thr Asp Ala Gln Val Trp Arg Thr Gly
515 520 525
Ile Val Met Val Ile Leu Leu Ala Ile Thr Pro Leu Cys Arg Arg Trp
530 535 540
Leu Thr Ile Leu Pro Leu Gly Gly Asp Thr Ala Arg Ala Val Gly Met
545 550 555 560
Ala Leu Thr Pro Thr Arg Ile Ala Leu Leu Leu Leu Ala Ala Cys Leu
565 570 575
Thr Ala Thr Ala Thr Met Thr Ile Gly Pro Leu Ser Phe Val Gly Leu
580 585 590
Met Ala Pro His Ile Ala Arg Met Met Gly Phe Arg Arg Thr Met Pro
595 600 605
His Ile Val Ile Ser Ala Leu Val Gly Gly Leu Leu Leu Val Phe Ala
610 615 620
Asp Trp Cys Gly Arg Met Val Leu Phe Pro Phe Gln Ile Pro Ala Gly
625 630 635 640
Leu Leu Ser Thr Phe Ile Gly Ala Pro Tyr Phe Ile Tyr Leu Leu Arg
645 650 655
Lys Gln Ser Arg
660
<210> 8
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 1
<400> 8
ctcctttcgt gtgcccgggc gcacgctttt gcatccgctg tcgttaacct aggtgacact 60
atagaacgcg 70
<210> 9
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 2
<400> 9
caataaacaa aactcacagg tgcagcaccc gccggatgcg gcaaaatacc tagtggatct 60
gatgggtacc 70
<210> 10
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 3
<400> 10
cgactgttaa cggcgatggc gctttctccg ttgttatggc agatgaatac aggtgacact 60
atagaacgcg 70
<210> 11
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 4
<400> 11
tcacgcttta cctccgatgg cgttatccag aacgcgcaca aagtgcattg tagtggatct 60
gatgggtacc 70
<210> 12
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 5
<400> 12
gtgagtaaac gaattgcgct tttcccggcg ttattgctgg cgctgttagt aggtgacact 60
atagaacgcg 70
<210> 13
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 6
<400> 13
aggttgacag cagccccgcc gggatctgga atggaaacag caccatccgc tagtggatct 60
gatgggtacc 70
<210> 14
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 7
<400> 14
atgcaggaat acacgaatca ttccgatacc acttttgcac tgcgtaatat ctcctttcgt 60
gtgcccgggc 70
<210> 15
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 8
<400> 15
gccatcgccg ttaacagtcg acggcgcgaa ataagaggta agccgctcat caataaacaa 60
aactcacagg 70
<210> 16
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 9
<400> 16
cctgtgagtt ttgtttattg atgagcggct tacctcttat ttcgcgccgt cgactgttaa 60
cggcgatggc 70
<210> 17
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 10
<400> 17
aatcactaac agcgccagca ataacgccgg gaaaagcgca attcgtttac tcacgcttta 60
cctccgatgg 70
<210> 18
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 11
<400> 18
tcggcaatgc actttgtgcg cgttctggat aacgccatcg gaggtaaagc gtgagtaaac 60
gaattgcgct 70
<210> 19
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 12
<400> 19
ttaacggctc tgctttctca acaaatagat aaaatatggc gcgccgataa aggttgacag 60
cagccccgcc 70
<210> 20
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 13
<400> 20
gaatacgtcg ccagctgctt taacac 26
<210> 21
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер 14
<400> 21
tggtttgtcg gatgcggcgt gaac 24
<---

Изобретение относится к области биотехнологии. Изобретение относится к микроорганизму из рода Escherichia, обладающему способностью продуцировать L-аминокислоты и повышенным уровнем внутриклеточного ATP по сравнению с немодифицированным штаммом, где виды активности одного или нескольких белков, выбранных из аминокислотной последовательности SEQ ID NO: 5, аминокислотной последовательности SEQ ID NO: 6 и аминокислотной последовательности SEQ ID NO: 7, которые образуют систему поглощения железа, были инактивированы, и к способу продуцирования L-аминокислот, включающему культивирование микроорганизма из рода Escherichia по любому из пп. 1-5 в среде и извлечение L-аминокислот из культуральной среды или микроорганизма, где L-аминокислота представляет собой L-треонин или L-триптофан. Изобретение позволяет улучшить уровень внутриклеточного АТР. 2. н. и 5 з.п. ф-лы, 6 ил., 5 табл., 8 пр.
1. Микроорганизм из рода Escherichia, обладающий способностью продуцировать L-аминокислоты и повышенным уровнем внутриклеточного ATP по сравнению с немодифицированным штаммом, где виды активности одного или нескольких белков, выбранных из аминокислотной последовательности SEQ ID NO: 5, аминокислотной последовательности SEQ ID NO: 6 и аминокислотной последовательности SEQ ID NO: 7, которые образуют систему поглощения железа, были инактивированы.
2. Микроорганизм из рода Escherichia по п. 1, где виды активности всех белков, имеющих аминокислотные последовательности SEQ ID NO: 5, 6 и 7, были инактивированы.
3. Микроорганизм из рода Escherichia по п. 1, где микроорганизм представляет собой E. coli.
4. Микроорганизм из рода Escherichia по п. 1, где микроорганизм из рода Escherichia обладает улучшенной способностью к продуцированию L-аминокислоты по сравнению с немодифицированным штаммом.
5. Микроорганизм из рода Escherichia по п. 4, где L-аминокислота представляет собой L-треонин или L-триптофан.
6. Способ продуцирования L-аминокислот, при этом способ включает:
культивирование микроорганизма из рода Escherichia по любому из пп. 1-5 в среде и
извлечение L-аминокислот из культуральной среды или микроорганизма.
7. Способ по п. 6, где L-аминокислота представляет собой L-треонин или L-триптофан.
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ ТРЕОНИНА (ВАРИАНТЫ) | 2002 |
|
RU2244007C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2312893C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ltaE | 2005 |
|
RU2304166C2 |
| Привод кругового поступательного движения | 1990 |
|
SU1829965A3 |

