Заявляемое изобретение относится к биотехнологии, в частности к созданию систем экспонирования для молочнокислых бактерий, и представляет собой способ применения белка S-слоя из штамма Lactobacillus brevis АТСС 367 для создания слитых белков, способных к специфической адсорбции на поверхности клеточных стенок.
В состав клеточной стенки прокариот, представителей доменов эубактерий и архебактерий, входит внешний белковый S-слой. S-слой формируется одним или несколькими белками с молекулярными массами в пределах 40-200 кДа, которые создают на поверхности клетки структурированную оболочку с различными типами симметрии (Дебабов, 2004). Белки S-слоя соединяются друг с другом и с клеточной стенкой через нековалентные взаимодействия и обладают свойством агрегироваться в регулярные структуры как в растворе, так и на твердой подложке in vitro. Для белков, формирующих S-слой, характерно разнообразие аминокислотных последовательностей, наличие значительного количества полярных кислых и неполярных аминокислот и низким расчетным значением изоэлектрической точки. В геноме конкретного штамма может присутствовать несколько генов белков S-слоя и характер их экспрессии может сильно различаться: активная экспрессия одного и молчание остальных, последовательная смена активности генов в процессе роста или формирования S-слоя несколькими белками (Hynonen and Palva, 2013). Благодаря способности к самопроизвольной сборке в упорядоченные, обладающие симметрией структуры, белки S-слоя интенсивно исследуют на предмет практического применения.
Белки поверхностного слоя S-слоя обнаружены и охарактеризованы у многих представителей рода Lactobacillus: L. acidophilus, L. amylolylicus, L. amylovorus, L. brevis, L. buchncri. L. crispalus, L. gallinarum, L. gigeriorum, L. helvelicus, L. hilgardii, L. kefircmofaciens, L. pastcurii, L. ultunensis (Hynonen and Palva, 2013). Аминокислотные последовательности белков S-слоя лактобацилл характеризуются выраженной гетерогенностью, характерной в том числе для штаммов одного вида ( and Palva, 2005; Hagen et al., 2005). Белки S-слоя лактобацилл отличаются малой массой (25-71 кДа), высокими значениями изоэлектрической точки (9,4-10,4) и состоят из двух домена - домена самосборки и домена присоединения к клеточной стенке. Также для белков S-слоя лактобацилл характерно отсутствие SLH-мотива, который отвечает за присоединение к клеточной стенке S-белков Bacillus anthracis, Geobacillus stearothermophilus, Lysinibacillus sphaericus и Clostridium thermocellum, что говорит о различии механизмов адсорбции на клеточной стенке (Lupas et al., 1994). Доменная структура охарактеризована для ряда белков S-слоя лактобацилл и условно делит их на две группы. Так, белки SA L. acidophilus АТСС 4356 (Smit et al., 2002) и CbsA L. crispatus JCM 5810 (Antikainen et al., 2002) характеризуются значительной идентичностью C-концевого домена, который отвечает за присоединение к клеточной стенке, и вариабельностью N-концевого домена, который вовлечен в процесс самосборки. Механизм адсорбции белков SA и CbsA описывается взаимодействием между полярными основными аминокислотными группами домена присоединения и негативно заряженными полимерами клеточной стенки, в качестве которых выступают тейхоевые кислоты (Antikainen et al., 2002; Sun et al., 2012). Доменная структура белка S-слоя SlpA L. brevis ATCC 8287 характеризуется зеркально обращенными регионами: N-концевой домен отвечает за присоединение к клеточной стенке, а С-концевой домен участвует в самосборке (
and Palva, 2005; Hagen et al., 2005). Белки S-слоя лактобацилл отличаются малой массой (25-71 кДа), высокими значениями изоэлектрической точки (9,4-10,4) и состоят из двух домена - домена самосборки и домена присоединения к клеточной стенке. Также для белков S-слоя лактобацилл характерно отсутствие SLH-мотива, который отвечает за присоединение к клеточной стенке S-белков Bacillus anthracis, Geobacillus stearothermophilus, Lysinibacillus sphaericus и Clostridium thermocellum, что говорит о различии механизмов адсорбции на клеточной стенке (Lupas et al., 1994). Доменная структура охарактеризована для ряда белков S-слоя лактобацилл и условно делит их на две группы. Так, белки SA L. acidophilus АТСС 4356 (Smit et al., 2002) и CbsA L. crispatus JCM 5810 (Antikainen et al., 2002) характеризуются значительной идентичностью C-концевого домена, который отвечает за присоединение к клеточной стенке, и вариабельностью N-концевого домена, который вовлечен в процесс самосборки. Механизм адсорбции белков SA и CbsA описывается взаимодействием между полярными основными аминокислотными группами домена присоединения и негативно заряженными полимерами клеточной стенки, в качестве которых выступают тейхоевые кислоты (Antikainen et al., 2002; Sun et al., 2012). Доменная структура белка S-слоя SlpA L. brevis ATCC 8287 характеризуется зеркально обращенными регионами: N-концевой домен отвечает за присоединение к клеточной стенке, а С-концевой домен участвует в самосборке ( et al., 2008). Рецепторы клеточной стенки, с которыми взаимодействует SlpA, также отличаются - селективное удаление тейхоевых кислот не сказывается на адсорбции SlpA на клеточной стенке; предполагается, что SlpA взаимодействует с нейтральными полисахаридами клеточной стенки посредством водородных связей ( et al., 2008; Dohm et al., 2011).
et al., 2008). Рецепторы клеточной стенки, с которыми взаимодействует SlpA, также отличаются - селективное удаление тейхоевых кислот не сказывается на адсорбции SlpA на клеточной стенке; предполагается, что SlpA взаимодействует с нейтральными полисахаридами клеточной стенки посредством водородных связей ( et al., 2008; Dohm et al., 2011).
Разнообразие белков S-слоя у штаммов Lactobacillus brevis исследовано в работе (Yasui et al., 1995). Показано, что у 36 из 41 исследованных штаммов обнаруживаются белки S-слоя с молекулярными массами от 38 до 55 кДа. Вестерн-блот анализ демонстрирует гетерогенность белков S-слоя у различных штаммов Lactobacillus brevis. Биохимически и генетически белки S-слоя охарактеризованы у штаммов Lactobacillus brevis АТСС 8287 (Vidgren et al., 1992) и ATCC 14869 (Jakava-Viljanen el al., 2002). Зрелый белок S-слоя штамма ATCC 8287 SlpA состоит из 435 аминокислотных остатков и имеет массу 46 кДа. Штамм АТСС 14869 экспрессирует два белка поверхностного слоя - SlpB и SlpD, хотя геном содержит три гена белков S-слоя slpB, slpC, and slpD.
Создание систем экспонирования белковых доменов на клеточной стенке лактобацилл является активно разрабатываемой прикладной областью с большим биотехнологическим потенциалом, который включает создание систем направленной доставки антигенов, систем диагностики, скрининга пептидных библиотек и созданием биокатализаторов (Leenhouts et al., 1999; Yu et al., 2013; Hynonen and Palva, 2013). Различные белки, ассоциированные с клеточной стенкой лактобацилл, исследованы на пригодность для создания систем экспонирования: LPXTG-белки (Marraffini et al., 2006), грансмембранный белок PgsA (Narita et al., 2006), белки, содержащие LysM мотив (Okano et al., 2008), белки S-слоя ( et al., 2002). Использование белков S-слоя основано как на создании слитых с целым белком чужеродных конструкций, так и на использовании функциональных частей белков S-слоя - доменов присоединения к клеточной стенке.
Показана возможность использовать SlpA в качестве системы экспонирования эпитопов на поверхности клетки (Avall-Jaaskelainen et al., 2002) и способность связываться с фибронектином эпителиальных клеток человека (Hynonen et al., 2002). Авторами статьи (Avall-Jaaskelainen et al., 2002) успешно опробована стратегия создания системы экспонирования эпитопов, основанная на встраивании аминокислотной последовательности чужеродного антигена в последовательность домена самосборки белка SlpA Lactobacillus brevis АТСС 8287. Химерная конструкция, состоящая из белка S-слоя и полиовирусного эпитопа VP1, экспрессируемая в генетически модифицированном Lactobacillus brevis, успешно формирует S-слой и экспонирует антиген на поверхности клетки. Данное направление получило дальнейшее развитие и были определены критичные регионы для встраивания чужеродных эпитопов в последовательность белка SlpA, в частности определены регионы внешней поверхности сетки S-слоя, внутренней поверхности сетки S-слоя, внутренний глобулярный регион и регион белок-белкового интерфейса (Vilen et al., 2009). Главное достоинство предложенного подхода состоит в том, что белок SlpA не теряет своего базового свойства по самосборке в составе S-слоя. Недостатками подхода является ограниченность сайтов для встраивания эпитопа и размер вставки (полиовирусный эпитоп VP1 представлен размером в 11 аминокислотных остатков), что требует индивидуального подхода к каждому случаю создания конструкций и не гарантирует успеха в случае крупных вставок.
Другой подход состоит в использовании домена присоединения к клеточной стенке белка S-слоя. Показано, что слитый белок, содержащий домен LcsB белка SipB из Lactobacillus crispalus и маркер GFP, способен иммобилизоваться на клеточных стенках штаммов различных представителей рода Lactobacillus (Hu et al., 2011). Данный подход, реализованный на основе высокоаффинного домена CBD из лизоцима LysA2 фага А2 Lactobacillus casei и антигена Е7, привел к пониманию принципиальной возможности создания вакцины против вируса папилломы человека, состоящего из слитого белка, иммобилизованного на поверхности клеток генетически не измененного штамма Lactobacillus casei (Ribelles et al., 2012; Ribelles et al., 2013).
Ближайшим аналогом заявляемого изобретения является система экспонирования слитых белков на клеточной стенке лактобацилл, основанная на использовании С-концевого домена присоединения к клеточной стенке белка SlpB Lactobacillus crispatus К2-4-3 (Hu et al., 2011). Наработанный с помощью системы гетерологичной экспрессии на основе E.coli слитый белок GFP-LcsB способен присоединяться к клеточной стенке различных видов лактобацилл in vitro. Заявляемое изобретение отличается от ближайшего аналога специфичной адсорбцией слитых белков на поверхности клеток Lactobacillus brevis или Lactobacillus acidophilis и незначительной адсорбцией на клеточные стенки других представителей рода Lactobacillus, что позволит создавать мультикомпонентные системы.
Задача заявляемого изобретения состоит в расширении арсенала белков, пригодных для применения в качестве компонента системы для экспонирования слитых белков на поверхности клеток молочнокислых бактерий.
Задача решена путем применения в качестве компонента системы экспонирования слитых белков на поверхности клеток молочнокислых бактерий полипептида, соответствующего SEQ ID NO 1.
Полипептид, соответствующий SEQ ID NO 1, представляет собой N-концевой домен присоединения к клеточной стенке белка S-слоя lvis_2083 штамма Lactobacillus brevis АТСС 367, и, будучи компонентом слитого белка, сообщает последнему свойство специфической адсорбции на клеточной стенке штаммов рода Lactobacillus.
Процесс получения слитых белков, способных к специфической адсорбции на клеточной стенке штаммов рода Lactobacillus, состоит из нескольких этапов:
- амплификации нуклеотидной последовательности, содержащей часть гена, кодирующую N-концевой домен присоединения к клеточной стенке лактобацилл белка lvis_2083;
- амплификации нуклеотидной последовательности, кодирующей белок антигенного компонента вакцины Е7-18 или флюоресцентного маркера eGFP,
- конструирования плазмидной ДНК (плазмиды), предназначенной для гетерологичной экспрессии слитого белка, состоящего из N-концевого домена присоединения к клеточной стенке белка lvis_2083, спейсерной последовательности и домена антигенного компонента вакцины или флюоресцентного маркера;
- конструирования рекомбинантного штамма-продуцента слитого белка на основе штамма Escherichia coli,
- экспрессии и очистки рекомбинантного слитого белка;
- адсорбции рекомбинантного слитого белка на культуре штаммов Lactobacillus brevis или Lactobacillus acidophilis.
Фигура 1. Рекомбинантная плазмида pET-lvis_2083_eGFP.
Фигура 2. Флюоресценция белка Lvis_2083_eGFP, адсорбированного на поверхности клеток штаммов: А - Lactobacillus brevis АТСС 367, Б - Lactobacillus brevis АТСС 8287.
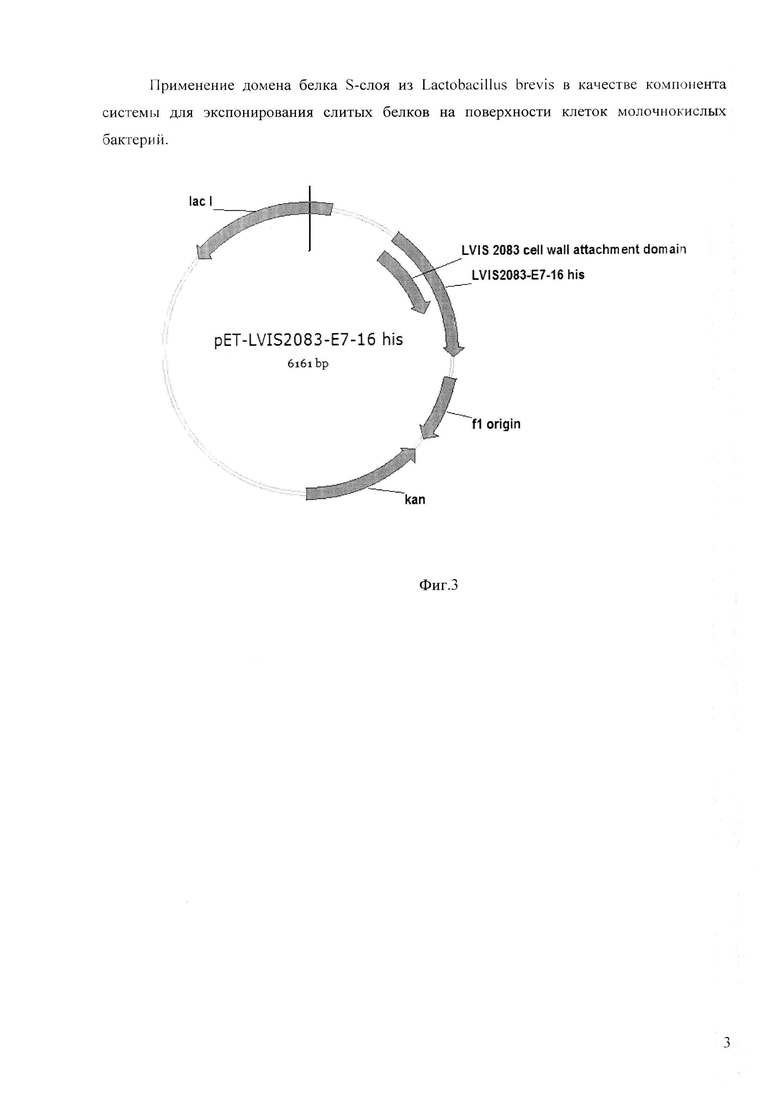
Фигура 3. Рекомбинантная плазмида pET-lvis_2083_E7-18.
Пример 1. Получение слитого белка lvis_2083_eGFP
Для получения слитого белка lvis_2083_eGFP создают генетическую конструкцию, состоящую из (5'->3') последовательностей, кодирующих структурную часть гена lvis_2083, глицин-сериновый спейсер, зеленый флуоресцентный белок и полигистидиновую метку. В качестве флуоресцентного белка используют последовательность улучшенного зеленого флуоресцентного белка EGFP (Enhanced Green Fluorescent Protein), созданного на основе GFP, выделенного из медузы Aequorea victoria. Домены слитого белка соединяют с помощью глицин-серинового спейсера следующей последовательности: Gly-Gly-Ser-Gly-Gly-Ser-Gly-Gly-Ser-Gly, который удобен тем, что позволяет вписать сайт узнавания рестриктазы BamHI без изменения последовательности аминокислот. Для последующей очистки слитого белка используют метод металл-аффинной хроматографии на Ni-NTA агарозе, для чего к С-конецу слитого белка присоединяют последовательность из 6 аминокислотных остатков гистидина (6*His tag).
Фрагмент гена Lvis_2083, кодирующий полипептид с последовательностью SEQ ID NO 1, получают методом полимеразной цепной реакции (ПЦР), для чего выбраны праймеры, соответствующие началу и концу области ДНК, кодирующую N-концевой домен присоединения к клеточной стенке, следующего состава (5'->3'; подчеркнуты сайты эндонуклеаз рестрикции NdeI и BamHI): NdeI-LVlS_2083 - 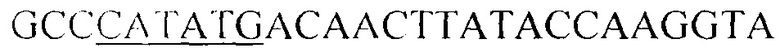 и R-BamHI-LVlS_2083 -
и R-BamHI-LVlS_2083 - 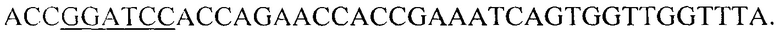 В обратный праймер R-BamHI-LVIS_2083 вписана часть последовательности глицин-серинового спейсера. Матрицей для ПЦР служит геномная ДНК штамма Lactobacillus brevis АТСС 367, выделенная по стандартной методике. В результате получают продукт длиной 630 по.
В обратный праймер R-BamHI-LVIS_2083 вписана часть последовательности глицин-серинового спейсера. Матрицей для ПЦР служит геномная ДНК штамма Lactobacillus brevis АТСС 367, выделенная по стандартной методике. В результате получают продукт длиной 630 по.
Структурную часть гена eGFP получают методом полимеразной цепной реакции, для чего выбраны праймеры следующего состава (5'->3'; подчеркнуты сайты эндонуклеаз рестрикции BamHI и XhoI): F-BamHI-eGFP - 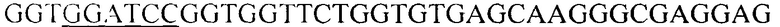 и R-XhoI-eGFP -
и R-XhoI-eGFP -  В прямой праймер F-BamHI-eGFP вписана часть последовательности глицин-серинового спейсера. Матрицой для ПЦР служит плазмидчая ДНК, содержащая синтезированную последовательность белка EGFP. В результате получают продукт длиной 744 по.
В прямой праймер F-BamHI-eGFP вписана часть последовательности глицин-серинового спейсера. Матрицой для ПЦР служит плазмидчая ДНК, содержащая синтезированную последовательность белка EGFP. В результате получают продукт длиной 744 по.
ПЦР проводили в 50 мкл реакционной смеси, содержащей 0,05-0,5 мкг ДНК, 10 пкМ каждого праймера, 25 мМ TAPS-HCl, рН 9.3, 50 mM KCl, 2 mM MgCl2, 1 mM β-меркаптоэтанола, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0,5 ед. высокоточной Phusion-ДНК-полимеразы. Реакцию амплификации проводят по следующей программе: первичная денатурация 96°С 2 мин; 30 циклов: 96°С - 10 сек, 59°С - 15 сек, 72°С - 30 с; финальная элонгация 72°С - 8 минут. Очистку полученных амплификацией фрагментов ДНК проводят с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия).
Плазмиду pET-2083_Gl/Ser_eGFP конструируют путем клонирования полученных фрагментов в вектор рЕТ-24а.
Гидролиз плазмиды рЕТ-24а и полученных фрагментов ДНК проводят рестриктазами NdeI, XhoI и BamHI (Thermo scientific, EU) при 37°C в буфере производителя в течение 1 часа. Очистку полученных фрагментов ДНК проводят с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия). Цитирование проводят в реакционной смеси объемом 20 мкл, содержащей 20 нг ДНК вектора, 20 нг ДНК фрагмента, 1 ед. Т4 ДНК лигазы (СибЭнзим, Россия), 50 mM Tris-HCl (рН 7.8 при 25°С), 10 mM MgCl2, 10 mM DTT, 1 mM АТР. Реакцию лигирования проводят при температуре 12-20°С в течение 3 часов. Полученной лигазной смесью в количестве 5 мкл трансформируют компетентные клетки штамма Escherichia coli XL1 blue MRF' (Stratagene, США) (генотип - Δ(mcrA)183 Δ(mcrCB-hsdSMR-mrr) 173 end Al supE44 thi-1 recAl gyrA96 relAl lac [F' proAB lacIqZΔM15 Tn10 (Tetr)]) по стандартной методике (Sambrook et al., 2001). После трансформации клоны отбирают на агаризованной среде LB (вес. %: триптон - 1, дрожжевой экстракт - 0,5, хлорид натрия - 1, агар - 1,5, остальное - вода), содержащей антибиотик канамицин в концентрации 25 мкг/мл. Плазмидную ДНК полученных трансформантов анализируют путем гидролиза рестриктазами NdeI и XhoI. В результате отбирают клоны, содержащие NdeI/XhoI фрагменты размером, равным исходному фрагменту вставки. Правильность клонирования подтверждают секвенированием вставки методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology. США).
В результате получают рекомбинантную плазмиду pET-2083_Gl/Ser_eGFP размером 6581 пар оснований, содержащую наряду с генами вектора рЕТ-24а, также ген слитого белка lvis_2083_eGFP (Фиг. 1).
С целью получения штамма-продуцента слитого белка lvis_2083_eGFP клетки штамма Escherichia coli BL21-CodonPlus (DE3)-RIPL трансформируют плазмидой рЕТ-2083_GI/Ser_eGFP. Трансформацию проводят 10 нг плазмидной ДНК методом химической трансформации по стандартной методике (Sambrook et al., 2001). Селекцию грансформантов проводят на LB-aгape, содержащем канамицин (25 мкг/мл), хлорамфеникол (34 мкг/мл) и стрептомицин (25 мкг/мл). В результате получают штамм Escherichia coli/pET-2083_Gl/Ser_eGFP, синтезирующий слитой белок lvis_2083_eGFP.
Исходным посевным материалом для экспрессии служит культура Escherichia coli/pET-2083_Gl/Ser_eGFP, выращенная при температуре 30°С в течение 18 часов на среде LB (вес. %: триптон - 1, дрожжевой экстракт - 0,5, хлорид натрия - 1, остальное - вода), содержащей 25 мг/мл канамицина, 34 мкг/мл хлорамфеникола и 25 мкг/мл стрептомицина. Процесс биосинтеза ведут к колбах Эрленмейера объемом 750 мл, содержащих 100 мл среды LB с канамицином (25 мг/л), хлорамфениколом (34 мкг/мл) и стрептомицином (25 мкг/мл). Посевной материал вносят в ферментационную среду в количестве, необходимом для создания его концентрации около 1 об. %. Процесс культивирования продуцента ведут при температуре 30°С на круговой качалке со скоростью вращения 130 об/мин до достижения оптической OD600=0,8-1,0 ОЕ (около 3-4 часов культивирования), после чего в культуральную жидкость добавляют IPTG до концентрации 0,5 мМ. Ферментацию продолжают при тех же условиях в течение еще 5 часов. Биомассу клеток отделяют центрифугированием при 3000 об/мин промывают 0,1М натрий-фосфатным буфером и вновь центрифугируют. Контроль экспрессии осуществляют с помощью денатурирующего электрофореза в полиакриламидном геле.
При сравнении спектра белков у штаммов Escherichia coli/pET-2083 Gl/Ser_eGFP и Escherichia coli BL21-CodonPlus (DE3)-R1PL обнаруживали в цитозольной растворимой фракции появление дополнительной белковой полосы с молекулярной массой 50,7 кДа, что соответствует расчетной молекулярной массе рекомбинантного слитого белка lvis_2083_eGFP.
Полученную биомассу ресуспендируют в 50 мМ фосфатном буфере, рН 8,0, содержащем 150 мМ хлорида натрия и 10 мМ имидазола. Клетки разрушают обработкой ультразвуком, проводя процедуру на льду. Целые клетки и нерастворимую фракцию отделяют центрифугированием (16000 об/мин, 25 мин). К супернатанту добавляют равный объем 50% суспензии Ni-NTA агарозы (Qiagen, EU), предварительно уравновешенной вышеуказанным буфером, и интенсивно перемешивают в течение 1 часа. Колонку заполняют полученной суспензией, промывают промывочным буфером (50 мМ фосфатный буфер, 150 мМ хлорида натрия, 20 мМ имидазола) и элюируют целевой белок с колонки буфером для смыва (50 мМ фосфатный буфер, 150 мМ хлорида натрия, 400 мМ имидазола). Полученные фракции, содержащие слитый белок lvis_2083_eGFP, объединяют и подвергают диализу против дистиллированной воды.
Пример 2. Адсорбция слитого белка Lvis_2083_eGFP на поверхности клеток Lactobacillus brevis
Адсорбцию слитого белка lvis_2083_eGFP проводят на живых подготовленных культурах штаммов Lactobacillus brevis АТСС 367 или Lactobacillus brevis АТСС 8287. 5 мл. свежей ночной культуры центрифугируют при 4000 об/мин 5 минут при комнатной температуре. Осадок трижды промывают буфером для связывания (25 mM TrisHCl рН=7.4, 150 mM NaCl). К осадку добавляют 0.7 мл 2М GuHCl, тщательно перемешивают и инкубируют при комнатной температуре 30 минут. Осаждают клетки центрифугированием при 4000 об./мин. 5 минут, супернатант удаляют и добавляют к осадку 0.8 мл. 5М LiCl, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут. Затем осаждают клетки центрифугированием при 4000 об/мин 5 минут, супернатант удаляют, а осадок промывают трижды от остатков реагентов буфером для связывания по 1.5 мл. Получившийся осадок ресуспензируют в 300 мкл буфера для связывания. Для адсорбции на живой культуре применяют очищенный слитый белок lvis_2083_eGFP или клеточный экстракт штамма продуцента с индуцированной экспрессией слитого белка lvis_2083_eGFP. К 100 мкл. подготовленных клеток добавляют либо 30 мкл. раствора очищенного белка (50-100 нг белка), либо 50-100 мкл. клеточного экстракта, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут с периодическим перемешиванием. Смесь центрифугируют при 4000 об./мин. 10 минут, супернатант используют как контроль. Осадок дважды промывается буфером для связывания. Осадок клеток анализируют методом флюоресцентной микроскопии и методом смыва поверхностного слоя детергентом. Микроскопию проводят на универсальном моторизированном исследовательском микроскопе OLYMPUS ВХ61, при увеличении 1000* с использованием иммерсионного объектива UPlanSApo 100*/1.40 oil и канала FITC для детекции флюоресценции eGFP. Смыв поверхностного слоя проводят 50-100 мкл 2% раствора SDS и инкубируют при 70°С в течение 10 минут. Супернатант анализируют методом SDS-PAGE.
В результате наблюдают специфическую сорбцию полученного слитого белка Lvis_20S3_eGFP на подготовленной клеточной стенке штаммов Lactobacillus brevis АТСС 8287 или Lactobacillus brevis АТСС 367. Контрольный целый белок eGFiJ не сорбируется клеточной культурой. По данным флюоресцентной микроскопии протестированные культуры штаммов АТСС 8287 и АТСС 367 представляют собой скопление характерных палочковидных бактерий с четкой зеленой флюоресценцией клеточной поверхности (Фиг. 2). Контрольная культура с добавлением только рекомбинантного eGFP не проявляла свойств флюоресценции клеточной поверхности. Для экспериментов применяют очищенный препарат рекомбинантного белка или суммарный клеточный лизат рекомбинантного штамма-продуцента, что подтверждает специфичность адсорбции и позволяет упростить процедуру очистки.
Пример 3. Адсорбция слитого белка Lvis_2083_eGFP на поверхности клеток штамма Lactobacillus acidophilis АТСС 4356
Адсорбцию слитого белка lvis_2083_eGFP проводят на живой подготовленной культуре штамма Lactobacillus acidophilis АТСС 4356. 5 мл свежей ночной культуры центрифугируют при 4000 об/мин 5 минут при комнатной температуре. Осадок трижды промывают буфером для связывания (25 mM TrisHCl рН=7.4, 150 mM NaCl). К осадку добавляют 0.7 мл. 2М GuHCl, тщательно перемешивают и инкубируют при комнатной температуре 30 минут. Осаждают клетки центрифугированием при 4000 об/мин 5 минут, супернатант удаляют и добавляют к осадку 0.8 мл 5М LiCl, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут. Затем осаждают клетки центрифугированием при 4000 об/мин 5 минут, супернатант удаляют, а осадок промывают трижды от остатков реагентов буфером для связывания по 1.5 мл. Получившийся осадок ресуспензируют в 300 мкл буфера для связывания. Для адсорбции на живой культуре применяют очищенный слитый белок lvis_2083_eGFP или клеточный экстракт штамма продуцента с индуцированной экспрессией слитого белка lvis_2083_eGFP. К 100 мкл подготовленных клеток добавляют либо 30 мкл раствора очищенного белка (50-100 нг белка), либо 50-100 мкл клеточного экстракта, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут с периодическим перемешиванием. Смесь центрифугируют при 4000 об/мин 10 минут, супернатант используют как контроль. Осадок дважды промывается буфером для связывания. Осадок клеток анализируют методом флюоресцентной микроскопии и методом смыва поверхностного слоя детергентом. Микроскопию проводят на универсальном моторизированном исследовательском микроскопе OLYMPUS ВХ61, при увеличении 1000* с использованием иммерсионного объектива UPlanSApo 100*/1.40 oil и канала FITC для детекции флюоресценции eGFP. Смыв поверхностного слоя проводят 50-100 мкл 2% раствора SDS и инкубируют при 70 °С в течение 10 минут. Супернатант анализируют методом SDS-PAGE.
В результате наблюдают специфическую сорбцию полученного слитого белка Lvis_2083_eGFP на подготовленной клеточной стенке бактерий штамма Lactobacillus acidophilis АТСС 4356. Контрольный целый белок eGFP не сорбируется клеточной культурой. По данным флюоресцентной микроскопии протестированная культура штамма Lactobacillus acidophilis АТСС 4356 представляют собой скопление характерных палочковидных бактерий с четкой зеленой флюоресценцией клеточной поверхности. Контрольная культура с добавлением только рекомбинантного eGFP не проявляла свойств флюоресценции клеточной поверхности.
Пример 4. Адсорбция слитого белка Lvis_2083_eGFP на поверхности клеток штаммов рода Lactobacillus
Адсорбцию слитого белка lvis_2083_eGFP проводят на живых подготовленных культурах штаммов Lactobacillus brevis АТСС 367, Lactobacillus plantarum АТСС 8014, Lactobacillus delbrueckii ATCC 11842, Lactobacillus acidophilus ATCC 4356, Lactobacillus casci ATCC 393, Laclococcus lactis ATCC 19435. Штаммы Lactobacillus brevis ATCC 367 и Lactobacillus acidophilus ATCC 4356 выступают в качестве контроля. 5 мл свежей ночной культуры центрифугируют при 4000 об/мин 5 минут при комнатной температуре. Осадок трижды промывают буфером для связывания (25 mM TrisHCl рН=7.4, 150 mM NaCl). К осадку добавляют 0.7 мл 2М GuHCl, тщательно перемешивают и инкубируют при комнатной температуре 30 минут. Осаждают клетки центрифугированием при 4000 об/мин 5 минут, супернатант удаляют и добавляют к осадку 0.8 мл. 5М LiCl, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут. Затем осаждают клетки центрифугированием при 4000 об/мин 5 минут, супернатант удаляют, а осадок промывают трижды от остатков реагентов буфером для связывания по 1.5 мл. Получившийся осадок ресуспензируют в 300 мкл буфера для связывания. Для адсорбции на живой культуре применяют очищенный слитый белок lvis_2083_eGFP или клеточный экстракт штамма продуцента с индуцированной экспрессией слитого белка lvis_2083 eGFP. К 100 мкл подготовленных клеток добавляют либо 30 мкл. раствора очищенного белка (50-100 нг белка), либо 50-100 мкл клеточного экстракта, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут с периодическим перемешиванием. Смесь центрифугируют при 4000 об/мин 10 минут, супернатант используют как контроль. Осадок дважды промывается буфером для связывания. Осадок клеток анализируют методом флюоресцентной микроскопии и методом смыва поверхностного слоя детергентом. Микроскопию проводят на универсальном моторизированном исследовательском микроскопе OLYMPUS ВХ61, при увеличении 1000* с использованием иммерсионного объектива UPlanSApo 100*/1.40 oil и канала F1TC для детекции флюоресценции eGFP. Смыв поверхностного слоя проводят 50-100 мкл 2% раствора SDS и инкубируют при 70°С в течение 10 минут. Супернатант анализируют методом SDS-PAGE.
В результате наблюдают специфическую сорбцию полученного слитого белка Lvis_2083_eGFP на подготовленных культурах штаммов Lactobacillus brevis АТСС 367 и Lactobacillus acidophilis АТСС 4356. Слитый белок незначительно сорбируется на подготовленных культурах штаммов Lactobacillus plantarum АТСС 8014, Lactobacillus delbrueckii АТСС 11842, Lactobacillus casei ATCC 393 и Lactococcus lactis ATCC 19435. Метод флюоресцентной микроскопии подтверждает полученные данные адсорбционных экспериментов по штаммам Lactobacillus brevis АТСС 367 и Lactobacillus acidophilis АТСС 4356, и не выявляет значимой флюоресценции культур остальных штаммов.
Пример 5. Получение слитого белка lvis_2083_E7-18
Для получения слитого белка lvis_2083_E7-18 создают генетическую конструкцию, состоящую из (5'->3') последовательностей, кодирующих структурную часть гена lvis_2083, глицин-сериновый спейсер, антиген Е7 вируса папилломы человека (ВПЧ) тип-18 и полигистидиновую метку.
Фрагмент гена Lvis_2083, кодирующий полипептид с последовательностью SEQ ID NO 1, получают методом полимеразной цепной реакции (ПЦР), для чего выбраны праймеры, соответствующие началу и концу области ДНК, кодирующую N-концевой домен присоединения к клеточной стенке, следующего состава (5'->3': подчеркнуты сайты эндонуклеаз рестрикции NdeI и BamHI): NdeI-LVIS_2083 - 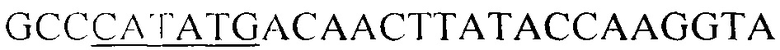 и R-BamHI-LVlS_2083 -
и R-BamHI-LVlS_2083 - 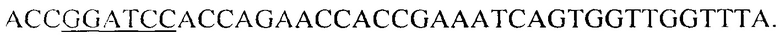 В обратный праймер R-BamHI-LVIS_2083 вписана часть последовательности глицин-серинового спейсера. Матрицей для ПЦР служит геномная ДНК штамма Lactobacillus brevis АТСС 367, выделенная по стандартной методике. В результате получают продукт длиной 630 по.
В обратный праймер R-BamHI-LVIS_2083 вписана часть последовательности глицин-серинового спейсера. Матрицей для ПЦР служит геномная ДНК штамма Lactobacillus brevis АТСС 367, выделенная по стандартной методике. В результате получают продукт длиной 630 по.
Структурную часть гена антигена Е7 ВПЧ тип-18 получают методом полимеразной цепной реакции, для чего были выбраны праймеры следующего состава (5'->3'; подчеркнуты сайты эндонуклеаз рестрикции BamHI и XhoI): SP_E7(_18_)_BamHI - 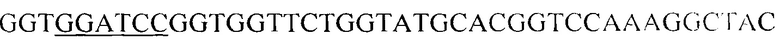 и ASP_E7(_18_)_in_frame_XhoI -
и ASP_E7(_18_)_in_frame_XhoI -  В прямой праймер SP_E7(_18_)_BamHI вписана часть последовательности глицин-серинового спейсера. Матрицой для ПЦР служит плазмидная ДНК, содержащая синтезированную последовательность гена антигена Е7 ВПЧ тип-18 (RU 2546917). В результате получают продукт длиной 345 по.
В прямой праймер SP_E7(_18_)_BamHI вписана часть последовательности глицин-серинового спейсера. Матрицой для ПЦР служит плазмидная ДНК, содержащая синтезированную последовательность гена антигена Е7 ВПЧ тип-18 (RU 2546917). В результате получают продукт длиной 345 по.
ПЦР проводят в 50 мкл реакционной смеси, содержащей 0,05-0,5 мкг ДНК, 10 пкМ каждого праймера, 25 мМ TAPS-HCl, рН 9.3, 50 mM KCl, 2 mM MgCl2, 1 mM β-меркаптоэтанола, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0,5 ед. высокоточной Phusion-ДНК-полимеразы. Реакцию амплификации проводят по следующей программе: первичная денатурация 96°С 2 мин; 30 циклов: 96°С - 10 сек, 59°С - 15 сек, 72°С - 30 с; финальная элонгация 72°С - 8 минут. Очистку полученных амплификацией фрагментов ДНК проводят с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия).
Плазмиду pET-2083_Gl/Ser_lvis_2083_E7-18 конструируют путем клонирования полученных фрагментов в вектор рЕТ-24а.
Гидролиз плазмиды рЕТ-24а и полученных фрагментов ДНК проводят рестриктазами NdeI, XhoI и BamHI (Thermo scientific, EU) при 37°C в буфере производителя в течение 1 часа. Очистку полученных фрагментов ДНК проводят с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия). Лигирование проводят в реакционной смеси объемом 20 мкл, содержащей 20 нг ДНК вектора, 20 нг ДНК фрагмента, 1 ед. Т4 ДНК лигазы (СибЭнзим, Россия). 50 mM Tris-HCl (pH 7.8 при 25°С), 10 mM MgCl2, 10 mM DTT, 1 mM АТР. Реакцию лигирования проводят при температуре 12-20°С в течение 3 часов. Полученной лигазной смесью в количестве 5 мкл трансформируют компетентные клетки штамма Escherichia coli XL1 blue MRF' (Stratagene, США) (генотип - Δ(mcrA)183 Δ(mcrCB-hsdSMR-mrr)173 endAl supE44 thi-1 recAl gyrA96 relAl lac [F' proAB lacIqZΔM15 Tn10 (Tetr)]) по стандартной методике (Sambrook et al., 2001). После трансформации клоны отбирают на агаризованнной среде LB (вес. %: триптон - 1, дрожжевой экстракт - 0,5, хлорид натрия - 1, агар - 1,5, остальное - вода), содержащей антибиотик канамицин в концентрации 25 мкг/мл. Плазмидную ДНК полученных трансформантов анализируют путем гидролиза рестриктазами NdeI и XhoI. В результате отбирают клоны, содержащие NdeI/XhoI фрагменты размером, равным исходному фрагменту вставки. Правильность клонирования подтверждают секвенированием вставки методом Сэнгера на генетическом анализаторе АВ1 3500 (Life technology, США).
В результате получают рекомбинантную плазмиду pET-2083_Gl/Ser_ lvis_20S3_E7-18 размером 6182 пар оснований, содержащую наряду с генами вектора рЕТ-24а также ген слитого белка Ivis_2083_E7-18 (Фиг. 3).
С целью получения штамма-продуцента слитого белка lvis_2083_E7-18 клетки штамма Escherichia coli BL21-CodonPlus (DE3)-RIPL трансформируют плазмидой рЕТ-2083_Gl/Ser_lvis_2083_E7-18. Трансформацию проводят 10 нг плазмидной ДНК методом химической трансформации по стандартной методике (Sambrook et al., 2001). Селекцию трансформантов проводят на LB-aгape, содержащем канамицин (25 мкг/мл), хлорамфеникол (34 мкг/мл) и стрептомицин (25 мкг/мл). В результате получают штамм Escherichia coli/pET-2083_Gl/Ser_lvis_2083_E7-18, синтезирующий слитый белок lvis_2083_E7-18.
Исходным посевным материалом для экспрессии служит культура Escherichia coli/pET-2083_Gl/Ser_lvis_2083_E7-18, выращенная при температуре 30°С в течение 18 часов на среде LB, содержащей 25 мг/мл канамицина, 34 мкг/мл хлорамфеникола и 25 мкг/мл стрептомицина. Процесс биосинтеза ведут в колбах Эрленмейера объемом 750 мл, содержащих 100 мл среды LB с канамицином (25 мг/л), хлорамфениколом (34 мкг/мл) и стрептомицином (25 мкг/мл). Посевной материал вносят в ферментационную среду в количестве, необходимом для создания его концентрации около 1 об. %. Процесс культивирования продуцента ведут при температуре 30°С на круговой качалке со скоростью вращения 130 об/мин до достижения оптической OD600=0,8-1.0 ОЕ (около 3-4 часов культивирования), после чего в культуральную жидкость добавляют IPTG до концентрации 0,5 мМ. Ферментацию продолжают при тех же условиях в течение еще 5 часов. Биомассу клеток отделяют центрифугированием при 3000 об/мин промывают 0,1М натрий-фосфатным буфером и вновь центрифугируют. Контроль экспрессии осуществляют с помощью денатурирующего электрофореза в полиакриламидном геле.
При сравнении спектра белков у штаммов Escherichia coli/pET-2083_Gl/SerJvis_2083_E7-L8 и Escherichia coli BL21-CodonPlus (DE3)-RIPL обнаруживали в цитозольной растворимой фракции появление дополнительной белковой полосы с молекулярной массой 36,0 кДа, что соответствует расчетной молекулярной массе рекомбинантного слитого белка Ivis_2083_E7-18.
Полученную биомассу ресуспендируют в 50 мМ фосфатном буфере, рН 8,0, содержащем 150 мМ хлорида натрия и 10 мМ имидазола. Клетки разрушают обработкой ультразвуком, проводя процедуру на льду. Целые клетки и нерастворимую фракцию отделяют центрифугированием (16000 об/мин, 25 мин). К супернатанту добавляют равный объем 50% суспензии Ni-NTA агарозы (Qiagen, EU), предварительно уравновешенной вышеуказанным буфером, и интенсивно перемешивают в течение 1 часа. Колонку заполняют полученной суспензией, промывают промывочным буфером (50 мМ фосфатный буфер, 150 мМ хлорида натрия. 20 мМ имидазола) и элюируют целевой белок с колонки буфером для смыва (50 мМ фосфатный буфер. 150 мМ хлорида натрия, 400 мМ имидазола). Полученные фракции, содержащие слитый белок lvis_2083_E7-18, объединяют и подвергают диализу против дистиллированной воды.
Пример 6. Адсорбция слитого белка lvis_2083_E7-18 на поверхности клеток Lactobacillus brevis
Адсорбцию слитого белка lvis_2083_E7-18 проводят на живых подготовленных культурах штаммов Lactobacillus brevis АТСС 367 или Lactobacillus brevis АТСС 8287. 5 мл свежей ночной культуры центрифугируют при 4000 об/мин 5 минут при комнатной температуре. Осадок трижды промывают буфером для связывания (25 mM TrisHCl рН=7.4, 150 mM NaCl). К осадку добавляют 0.7 мл 2М GuHCl, тщательно перемешивают и инкубируют при комнатной температуре 30 минут. Осаждают клетки центрифугированием при 4000 об/мин 5 минут, отбрасывают супернатант и добавляют к осадку 0.8 мл 5М LiCl, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут. Затем осаждают клетки центрифугированием при 4000 об/мин 5 минут, отбрасывают супернатант и осадок промывают трижды от остатков реагентов буфером для связывания по 1.5 мл. Получившийся осадок ресуспензируют в 300 мкл буфера для связывания. Для адсорбции на живой культуре применяют очищенный слитый белок lvis_2083_E7-18 или клеточный экстракт штамма продуцента с индуцированной экспрессией слитого белка lvis_2083_E7-18. К 100 мкл подготовленных клеток добавляют либо 30 мкл раствора очищенного белка (50-100 нг белка), либо 50-100 мкл клеточного экстракта, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут с периодическим перемешиванием. Смесь центрифугируют при 4000 об/мин 10 минут, супернатант используют как контроль. Осадок дважды промывается буфером для связывания. Осадок клеток анализируют методом смыва поверхностного слоя детергентом. Смыв поверхностного слоя проводят 50-100 мкл 2% раствора SDS и инкубируют при 70 °С в течение 10 минут. Супернатант анализируют методом SDS-PAGE.
В результате наблюдают специфическую сорбцию слитого белка на основе домена присоединения к клеточной стенке lvis_2083_E7-18 на подготовленной клеточной стенке штаммов Lactobacillus brevis АТСС 8287 или Lactobacillus brevis АТСС 367. Контрольный целый белок Е7-18 не сорбируется клеточной культурой. Для экспериментов применяют очищенный препарат рекомбинантного белка или суммарный клеточный лизат рекомбинантного штамма-продуцента, что подтверждает специфичность адсорбции и позволяет упростить процедуру очистки.
Пример 7. Адсорбция слитого белка lvis_2083_E7-18 на поверхности клеток штамма Lactobacillus acidophilis АТСС 4356
Адсорбцию слитого белка lvis_2083_E7-18 проводят на живых подготовленных культурах штамма Lactobacillus acidophilis АТСС 43565 мл свежей ночной культуры центрифугируют при 4000 об/мин 5 минут при комнатной температуре. Осадок трижды промывают буфером для связывания (25 mM TrisHCl рН=7.4, 150 mM NaCl). К осадку добавляют 0.7 мл 2М GuHCl, тщательно перемешивают и инкубируют при комнатной температуре 30 минут. Осаждают клетки центрифугированием при 4000 об./мин. 5 минут, супернатант удаляют и добавляют к осадку 0.8 мл 5М LiCl, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут. Затем осаждают клетки центрифугированием при 4000 об/мин 5 минут, супернатант удаляют, а осадок промывают трижды от остатков реагентов буфером для связывания по 1.5 мл. Получившийся осадок ресуспензируют в 300 мкл буфера для связывания. Для адсорбции на живой культуре применяют очищенный слитый белок lvis_2083_E7-18 или клеточный экстракт штамма продуцента с индуцированной экспрессией слитого белка lvis 2083 Е7-18. К 100 мкл подготовленных клеток добавляют либо 30 мкл. раствора очищенного белка (50-100 нг белка), либо 50-100 мкл. клеточного экстракта, тщательно перемешивают и инкубируют при комнатной температуре в течение 30 минут с периодическим перемешиванием. Смесь центрифугируют при 4000 об/мин 10 минут, супернатант используют как контроль. Осадок дважды промывается буфером для связывания. Осадок клеток анализируют методом смыва поверхностного слоя детергентом. Смыв поверхностного слоя проводят 50-100 мкл. 2% раствора SDS и инкубируют при 70 °С в течение 10 минут. Супернатант анализируют методом SDS-PAGE.
В результате наблюдают специфическую сорбцию слитого белка lvis_2083_E7-18 на подготовленной клеточной стенке бактерий штамма Lactobacillus acidophilis АТСС 4356. Контрольный целый белок Е7-18 не сорбируется клеточной культурой.
Источники информации
1. Дебабов В.Г. // S-слои бактерий и архей как объект бионанотехнологий // Молекулярная биология, 2004.
2. Antikainen J., Anton L.,  J., Korhonen T.K // Domains in the S-layer protein CbsA of Lactobacillus crispatus involved in adherence to collagens, laminin and lipoteichoic acids and in self-assembly // Mol Microbiol., 2002.
J., Korhonen T.K // Domains in the S-layer protein CbsA of Lactobacillus crispatus involved in adherence to collagens, laminin and lipoteichoic acids and in self-assembly // Mol Microbiol., 2002.
3.  S., Palva A. // Lactobacillus surface layers and their applications // FEMS Microbiol Rev., 2005.
S., Palva A. // Lactobacillus surface layers and their applications // FEMS Microbiol Rev., 2005.
4. S.,  U., Ilk N., Pum D., Sleytr U.B., Palva A. // Identification and characterization of domains responsible for self-assembly and cell wall binding of the surface layer protein of Lactobacillus brevis ATCC 8287 // BMC Microbiol. 2008.
U., Ilk N., Pum D., Sleytr U.B., Palva A. // Identification and characterization of domains responsible for self-assembly and cell wall binding of the surface layer protein of Lactobacillus brevis ATCC 8287 // BMC Microbiol. 2008.
5. S.,  K., Kahala M., Miikkulainen-Lahti Т. Palva A. // Surface display of foreign epitopes on the Lactobacillus brevis S-layer // Appl. Environ. Microbiol., 2002.
K., Kahala M., Miikkulainen-Lahti Т. Palva A. // Surface display of foreign epitopes on the Lactobacillus brevis S-layer // Appl. Environ. Microbiol., 2002.
6. Dohm N., Petri A., Schlander M., Schlott В.,  H., Claus H. // Molecular and biochemical properties of the S-layer protein from the wine bacterium Lactobacillus hilgardii B706 //Arch Microbiol., 2011.
H., Claus H. // Molecular and biochemical properties of the S-layer protein from the wine bacterium Lactobacillus hilgardii B706 //Arch Microbiol., 2011.
7. Hagen K.E., Guan L.L., Tannock G.W., Korver D.R., Allison G.E. // Detection, characterization, and in vitro and in vivo expression of genes encoding S-protcins in Lactobacillus gallinarum strains isolated from chicken crops // Appl. Environ. Microbiol., 2005.
8. Hu S., Kong J., Sun Z., Han L., Kong W., Yang P. // Heterologous protein display on he cell surface of lactic acid bacteria mediated by the s-layer protein // Microb. Cell Fact., 2011.
9. U., Palva A. // Lactobacillus surface layer proteins: structure, function and applications // Appl. Microbiol. Biotechnol., 2013.
10. U.,  В., Palva A., Korhonen Т.К. // Identification by flagellum display of an epithelial cell- and fibronectin-binding function in the SlpA surface protean of Lactobacillus brevis //.1. Bacteriol., 2002.
В., Palva A., Korhonen Т.К. // Identification by flagellum display of an epithelial cell- and fibronectin-binding function in the SlpA surface protean of Lactobacillus brevis //.1. Bacteriol., 2002.
11. Jakava-Viljanen M.,  S., Messner P., Sleytr U.B., Palva A. // Isolation of three new surface layer protein genes (sip) from Lactobacillus brevis ATCC 14869 and characterization of the change in their expression under aerated and anaerobic conditions //.1 Bacteriol., 2002.
S., Messner P., Sleytr U.B., Palva A. // Isolation of three new surface layer protein genes (sip) from Lactobacillus brevis ATCC 14869 and characterization of the change in their expression under aerated and anaerobic conditions //.1 Bacteriol., 2002.
12. Leenhouts K., Buist G., Kok J: // Anchoring of proteins to lactic acid bacteria // Anionic Van Leeuwenhoek, 1999.
13. Lupas A, Engelhardt H., Peters J., Santarius U., Volker S., Baumeister W. // Domain struciure of the Acetogenium kivui surface layer revealed by electron crystallography and sequence analysis // J Bacteriol., 1994.
14. Marraffini L.A., Dedent A.C., Schneewind O. // Sortases and the art of anchoring proteins to the envelopes of gram-positive bacteria // Microbiol Mol Biol Rev., 2006.
15. Narita J., Okano K., Kitao Т., Ishida S., Sewaki Т., Sung M.H., Fukuda H., Kondo A:/ // Display of alpha-amylase on the surface of Lactobacillus casei cells by use of the PgsA anchor protein, and production of lactic acid from starch // Appl Environ Microbiol., 2006.
16. Okano К, Zhang Q., Kimura S., Narita J., Tanaka Т., Fukuda H., Kondo A. // System using tandem repeats of the cA peptidoglycan-binding domain from Lactococcus lactis for display of both N- and C-terminal fusions on cell surfaces of lactic acid bacteria // Appl Environ Microbiol., 2008.
17. Ribelles P., Benbouziane В., Langella P.,  J.E.,
J.E.,  L.G. // Protection against human papillomavirus type 16-induced tumors in mice using non-genetically modi lied lactic acid bacteria displaying E7 antigen at its surface // Appl. Microbiol. Biotechnol., 2013.
L.G. // Protection against human papillomavirus type 16-induced tumors in mice using non-genetically modi lied lactic acid bacteria displaying E7 antigen at its surface // Appl. Microbiol. Biotechnol., 2013.
18. Ribelles P.,  I., J.E. // LysA2, the Lactobacillus casei bacteriophage A2 lysin is an endopeptidase active on a wide spectrum of lactic acid bacteria // Appl. Microbiol. Biotechnol., 2012.
I., J.E. // LysA2, the Lactobacillus casei bacteriophage A2 lysin is an endopeptidase active on a wide spectrum of lactic acid bacteria // Appl. Microbiol. Biotechnol., 2012.
19. Sambrook J. and Russell DW (2001) // Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press.
20. Smit E., Jager D., Martinez В., Tielen F.J., Pouwels P.H. // Structural and functional analysis of the S-layer protein crystallisation domain of Lactobacillus acidophilus ATCC 4356: evidence for protein-protein interaction of two subdomains // J Mol Biol., 2002
21. Sun Z., Kong Hu S., Kong W., Lu W., Liu W. // Characterization of a S-layer protein from Lactobacillus crispatus КЗ 13 and the domains responsible for binding to cell wall and adherence to collagen // Appl Microbiol Biotechnol., 2012.
22.  G., Palva 1., Pakkanen R., Lounatmaa K., Palva A. // S-layer protein gene of Lactobacillus brevis: cloning by polymerase chain reaction and determination of the nucleotide sequence // J Bacteriol., 1992.
G., Palva 1., Pakkanen R., Lounatmaa K., Palva A. // S-layer protein gene of Lactobacillus brevis: cloning by polymerase chain reaction and determination of the nucleotide sequence // J Bacteriol., 1992.
23. Vilen H.,  U., Badelt-Lichtblau H., Ilk N.,
U., Badelt-Lichtblau H., Ilk N., 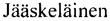 P., Torkkeli M.. Pah a A. // Surface location of individual residues of SlpA provides insight into the Lactobacillus brevis S-layer//J. Bacteriol., 2009.
P., Torkkeli M.. Pah a A. // Surface location of individual residues of SlpA provides insight into the Lactobacillus brevis S-layer//J. Bacteriol., 2009.
24. Yasir Т., Yoda K., Kamiya T. // Analysis of S-layer proteins of Lactobacillus brevis // FEMS Microbiol Lett., 1995.
25. Yu Q., Zhu L., Kang H., Yang Q. // Mucosal Lactobacillus vectored vaccines // Hum Vaccin Immunother., 2013.
26. RU 2546917 // Способ микробиологического синтеза гибридного белка e7-hsp70 (варианты) // Дата приоритета: 16.10.2013.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СЕКРЕТОРНОЙ ПРОДУКЦИИ БЕЛКА | 2002 |
|
RU2264463C2 |
| СПОСОБ ПОЛУЧЕНИЯ БИОЛОГИЧЕСКИ АКТИВНЫХ РЕКОМБИНАНТНЫХ ПРОТЕИНОВ | 2022 |
|
RU2807615C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА SAV-RGD | 2014 |
|
RU2577138C1 |
| БЕЛОК, ПОЛУЧЕННЫЙ ИЗ NEISSERIA MENINGITIDIS, ПРОЯВЛЯЮЩИЙ СВОЙСТВА АНТИГЕНА, АНТИТЕЛО, ФРАГМЕНТ НУКЛЕИНОВОЙ КИСЛОТЫ (ВАРИАНТЫ) И СОДЕРЖАЩАЯ ИХ КОМПОЗИЦИЯ | 1999 |
|
RU2233331C2 |
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКА E7-HSP70 И ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2012 |
|
RU2489481C1 |
| ГИБРИДНЫЙ БЕЛОК НА ОСНОВЕ L-АСПАРАГИНАЗЫ Wolinella succinogenes, ШТАММ Escherichia coli - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ГИБРИДНОГО БЕЛКА, ОБЛАДАЮЩЕГО ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТЬЮ | 2014 |
|
RU2562166C1 |
| ПРИМЕНЕНИЕ СЛИТОГО ПОЛИПЕПТИДА, СОДЕРЖАЩЕГО БЕЛОК MeCP2 И ДОМЕН ТРАНСДУКЦИИ БЕЛКА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ТАКОЙ БЕЛОК, И СПОСОБ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ СВЯЗАННОГО С РАЗВИТИЕМ НЕРВНОГО ЗАБОЛЕВАНИЯ | 2006 |
|
RU2432359C2 |
| ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE-BKM CR-349D-ПРОДУЦЕНТ ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА ЧЕЛОВЕКА | 1999 |
|
RU2150501C1 |
| БЕЛОК, СООТВЕТСТВУЮЩИЙ АНТИГЕННОМУ БЕЛКУ NEISSERIA MENINGITIDIS И ОБЛАДАЮЩИЙ ИММУНОГЕННЫМИ СВОЙСТВАМИ (ВАРИАНТЫ), СВЯЗЫВАЮЩЕЕСЯ С НИМ АНТИТЕЛО, НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ (ВАРИАНТЫ) И СОДЕРЖАЩАЯ ИХ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1998 |
|
RU2232191C2 |
| АНТИГЕНЫ NEISSERIA MENINGITIDIS | 1999 |
|
RU2343159C2 |



Изобретение относится к биотехнологии. Предложено применение полипептида, соответствующего SEQ ID NO 1, в качестве компонента системы для экспонирования слитых белков на поверхности клеток молочнокислых бактерий. Полипептид представляет собой N-концевой домен присоединения к клеточной стенке белка S-слоя lvis_2083 штамма бактерий Lactobacillus brevis ATCC 367. Изобретение обеспечивает расширить арсенала белков, пригодных для применения в качестве компонента системы для экспонирования слитых белков на поверхности клеток молочнокислых бактерий. 3 ил., 7 пр.
Применение полипептида, соответствующего SEQ ID NO 1, в качестве компонента системы для экспонирования слитых белков на поверхности клеток молочнокислых бактерий.
| HU S | |||
| et al | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| ВЕКТОР, ОБЕСПЕЧИВАЮЩИЙ ЭКСПРЕССИЮ АНТИГЕНА ВИРУСА SARS НА ПОВЕРХНОСТИ КЛЕТОК, И МИКРООРГАНИЗМЫ, ТРАНСФОРМИРОВАННЫЕ ЭТИМ ВЕКТОРОМ | 2004 |
|
RU2332457C2 |
| AVALL-JAASKELAINEN S | |||
| et al | |||
| Приспособление к электрическим измерительным приборам для получения некоторой части шкалы измененных отклонений подвижной системы | 1925 |
|
SU8287A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| HYNONEN U | |||
| et al | |||
| Lactobacillus surface layer proteins: structure, function and applications // Appl Microbiol Biotechnol, 2013, 97:5225-5243. | |||

