Изобретение относится к биотехнологии и касается способа выделения и очистки рекомбинантного белка GAGE1 человека (p4GAGEl), пригодного для научно-исследовательских работ.
Белки семейства GAGE относятся к раково-тестикулярным антигенам и насчитывают около 16 членов на данный момент. Белки семейства GAGE могут быть потенциальными антигенами для иммунотерапии широкого спектра онкологических заболеваний, так как экспрессируются в различных типах опухолей, но не в нормальных тканях, а также вызывают иммунный ответ [Van den Eynde В, Peeters О, De Backer О, Gaugler В, Lucas S, Boon T. A new family of genes coding for an antigen recognized by autologous cytolytic T-lymphocytes on a human melanoma. J Exp Med, 1995; 182; c. 689-698]. Для оценки прогностических и терапевтических свойств антигенов необходима проверка их экспрессии на уровне белка. Создание продуцентов рекомбинантных белков это быстрый способ получения чистого белка, пригодного для иммунизации с целью получения специфических антител. Чистый рекомбинантный белок также может быть использован для детекции гуморального иммунного ответа в сыворотке крови онкологических больных и для нагрузки дендритных клеток при создании дендритноклеточной вакцины. Кроме того, чистый рекомбинантный белок может быть основой противоопухолевой иммуногенной композиции.
Все белки семейства GAGE очень похожи между собой по аминокислотному составу. Филогенетически ген GAGE1 признан самым древним в кластере генов семейства GAGE, и его белок имеет самый большой размер в семействе [Gjerstorff MF, Ditzel HJ. An overview of the GAGE cancer/testis antigen family with the inclusion of newly identified members. Tissue Antigens. 2008; 71(3): c. 187-192]. Полноразмерный рекомбинантный белок GAGE1 человека содержит практически все эпитопы, которые можно выделить при процессинге других белков семейства GAGE. Использование GAGE1 в научно-прикладных исследованиях будет иметь ряд преимуществ по сравнению с другими белками. Благодаря большему количеству эпитопов можно стимулировать развитие B- и T-клеток, обладающих широким репертуаром рецепторов, позволяющих распознать как GAGE1, так и другие белки семейства GAGE.
Наиболее близким к заявляемому способу является способ получения рекомбинантного белка GAGE12 [Gjerstorff MF, Besir Н, Larsen MR, Ditzel HJ. Expression, purification and characterization of the cancer-germline antigen GAGE 121: a candidate for cancer immunotherapy. Protein Expr Purif 2010; 73: 217-222], принятый за прототип. Способ состоит в том, что рекомбинантный белок GAGE 12 человека экспрессируют в дрожжевом продуценте Pichia pastoris (GS115). Далее получают биомассу клеток, которую ресуспендируют в соответствующем буфере и выделяют целевой белок с помощью трех последовательных хроматографических очисток: аффинной, ионобменной и гель-фильтрации. После первого этапа аффинной хроматографии получают белок с 60% электрофоретической чистотой. Общий выход чистого рекомбинантного белка составляет 10,5 мг/л исходной культуры продуцента после первого этапа хроматографической очистки и 6 мг/л культуры после дополнительных двух этапов хроматографической очистки.
Недостатками способа являются невысокий общий выход белка, а также присутствие олигомерных форм белка при его экспрессии, что требует многоэтапной хроматографической очистки.
Задача заявляемого изобретения состоит в создании способа получения рекомбинантного белка GAGE1 человека более высокой степени очистки и более высокого общего выхода белка с использованием одного этапа хроматографической очистки.
Технический результат заявляемого способа состоит в получении рекомбинантного белка GAGE1 человека с чистотой более 95% и общим выходом 24,5 мг/л культуры продуцента с использованием одного этапа хроматографической очистки.
Поставленная задача решается использованием бактериального продуцента рекомбинантного белка и введением предварительного этапа отмывки клеток продуцента от части неспецифических белков.
Сущность заявляемого способа получения рекомбинантного белка GAGE1 человека состоит в использовании рекомбинантной плазмиды pGAGE1, размером 5763 пар нуклеотидов, кодирующей рекомбинантный белок p4GAGE1. Эта рекомбинантная плазмида представляет собой плазмидный вектор, состоящий из следующих элементов: последовательности кодирующей рекомбинантный белок p4GAGE1 с сайтами узнавания для рестриктаз EcoRI и HindIII, промотора Т7, гена устойчивости к канамицину, последовательности инициации репликации pBR322(ori), гена, кодирующего lac-индуктор, и последовательностей, необходимых для инициации трансляции белка в клетках Е. coli.
Способ получения рекомбинантного белка p4GAGE1 осуществляется следующим образом.
Синтезируют последовательность ДНК, кодирующую человеческий ген GAGE1. В качестве источника гена GAGE1 используют кДНК клеточной линии эритроидного миелолейкоза К562. Ген GAGE1 человека выделяют методом амплификации с помощью олигонуклеотидных праймеров. Получают последовательность ДНК, содержащую кодирующую последовательность гена GAGE1. Нуклеотидные последовательности праймеров, используемых при синтезе кодирующей последовательности гена GAGE1 представлены в Перечне последовательностей - SEQ ID NO: 1, 2.
Далее конструируют плазмидный вектор для экспрессии рекомбинантного белка. Для этого фрагмент ДНК, кодирующий соответствующий белок GAGE1, обрабатывают ферментами рестрикции HindIII и EcoRI и вставляют по сайтам рестрикции HindIII и EcoRI в вектор рЕТ-30a (Novagen), получая при этом вектор pGAGE1. Соответствие последовательности ДНК плазмидного вектора pGAGEl схеме конструирования подтверждают методом прямого секвенирования по Сэнгеру. ДНК последовательности, кодирующей рекомбинантный белок GAGE1 человека, представлена в Перечне последовательностей - SEQ ID NO: 3 и аминокислотная последовательность рекомбинантного белка GAGE1 человека - SEQ ID NO: 4.
Затем синтезируют рекомбинантный белок в клетках Е. coli. Клетки Е. coli штамма BL21(DE3) pLysS трансформируют плазмидой pGAGE1 и выращивают в течение 18 ч при 37°C в агаризованной среде LB с добавлением канамицина (20 мкг/мл) и хлорамфеникола (34 мкг/мл). Затем несколько колоний вносят в свежую среду ТВ с канамицином и хлорамфениколом в тех же концентрациях и подращивают 3-4 ч до плотности OD600=0.6, затем вносят изопропил-β-D-1-тиогалактопиранозид до конечной концентрации 0,5 мМ для индукции синтеза целевого белка. Растят биомассу клеток продуцента 20 часов для синтеза p4GAGE1 при 25°C с принудительной аэрацией. Центрифугируют полученную суспензию клеток при 5 тыс. об/мин в течение 10 мин, супернатант удаляют.
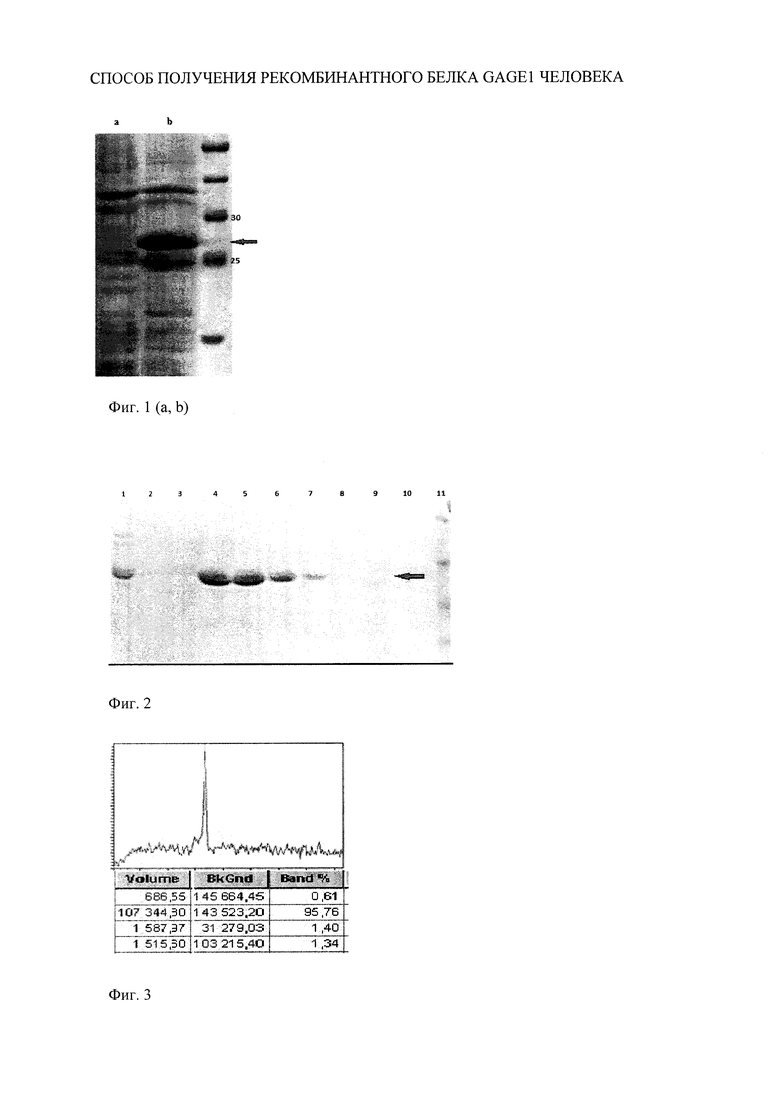
Способ иллюстрируется фиг. 1(а, b), 2 и 3.
На фиг. 1(a, b) представлена электрофореграмма белков бактериального продуцента P4GAGE1: а - культура бактерий без индукции синтеза белка, b - культура бактерий, продуцирующих рекомбинантный белок GAGE1 человека.
На фиг. 2 представлена электрофореграмма белков разных этапов очистки белка P4GAGE1 из бактериальной культуры: 1 - фракция белков, содержащих p4GAGE1, перед нанесением на колонку, 2, 3 - фракции промывки колонки, 4-8 - элюция очищенного P4GAGE1 с колонки.
На фигуре 3 представлена денситограмма очищенного рекомбинантного белка GAGE1 человека.
Клетки Е. coli, содержащие плазмиду pGAGE1, эффективно поддерживали экспрессию целевого рекомбинантного белка размером 27 кДа. Целевой белок экспрессировался в виде нерастворимых телец включения. Для очистки рекомбинантного белка p4GAGE1 из штамма-продуцента Е. coli, содержащего плазмиду pGAGE1 осадок клеток, полученный из культуры Е. coli на предыдущем этапе, ресуспендируют в буфере для обработки ультразвуком (50 мМ трис-HCl, рН=8,0, 20 мМ ЭДТА). Клетки подвергают двукратному замораживанию-оттаиванию. После этого клетки подвергают ультразвуковой дезинтеграции (22 кГц, 3×10 с), добавляют ингибиторы протеаз, рибонуклеазу до концентрации 2 ед./мл и дезоксирибонуклеазу до концентрации 300000 ед./мл и инкубируют в течение 10 мин при комнатной температуре. Фрагменты разрушенных клеток осаждают центрифугированием (100000 g, 4°C, 1 ч).
Далее проводят процедуру отмывки телец включения. Для этого осадок ресуспендируют в буфере, содержащем 50 мМ трис-HCl, рН=8,0, 0,2 мМ ЭДТА, 0,05М NaCl, и подвергают действию ультразвука, как описано выше, с последующим центрифугированием (10000 об/мин, 5 мин). Процедуру отмывки телец включения повторяют еще два раза. Такая процедура позволяет удалить часть белков продуцента и подготовить лизат смеси белков, обогащенный целевым рекомбинантным белком для последующей очистки методом металлохелатной аффинной хроматографии. Полученный препарат телец включения рекомбинантного белка растворяют в буфере, содержащем 50 мМ трис-HCl, pH=7,5, 6 М мочевину. Далее проводят металлохелатную аффинную хроматографию при умеренном давлении (BioLogic, "Bio-Rad") (скорость протока 2 мл/мин) на колонке с Ni+ агарозой (GE Healthcare), уравновешенной буфером, содержащим 50 мМ трис-HCl, рН=7,5, 6 М мочевину. После нанесения препарата колонку промывают буфером для уравновешивания, содержащим 50 мМ трис-HCl, рН=7,5, 6 М мочевину, 2% Твин-20, а затем тем же буфером, содержащим 50 мМ трис-HCl, рН=7,5, 6 М мочевину. Элюцию белка проводят буфером, содержащим 50 мМ трис-HCl, pH=7,5, 200 мМ имидазола, 6 М мочевину. Фракции анализируют с помощью электрофореза в полиакриламидном геле. На фиг.2 показано, что фракции элюции содержат очищенный белок. Содержащие целевой белок фракции объединяют и 2 раза проводят диализ против 100-кратного объема буфера, содержащего 5% глицерин, 10 мМ трис-HCl, рН 7,5, 0,1 мМ ЭДТА, 0,05 М NaCl. Затем 2 раза проводят диализ раствора, содержащего рекомбинантный белок, против 0,9% раствора NaCl и осуществляют стерилизующую фильтрацию через фильтр 0,22 мкм. Аликвоты хранят при -70°C либо подвергают лиофилизации.
Наличие и чистоту целевого белка проверяют с помощью денатурирующего электрофореза в 10% полиакриламидном геле согласно стандартной методике. Денситометрию электрофореграммы очищенного белка проводят с помощью программы Phoretix 1D Advanced v. 5.10. На фиг. 3 показано, что получен белок с чистотой более 95%. Аминокислотная последовательность полученного чистого рекомбинантного белка подтверждается методом масс-спектрометрии и соответствует SEQ ID NO: 4.
Уровень продукции целевого белка (общий выход) в штамме Е. coli, трансформированном плазмидой pGAGE1, составляет 24,5 мг/л культуры продуцента, что в 6 раз выше, чем в приведенном способе прототипа.
Таким образом, предлагаемый способ позволяет получить рекомбинантный белок GAGE1 человека с более высокой степенью очистки, составляющей не менее 95%, и общим выходом белка 24,5 мг/л.




Изобретение относится к биотехнологии, конкретно к получению рекомбинантных белков человека, и может быть использовано для получения и очистки рекомбинантного белка GAGE1 человека. Способ получения рекомбинантного белка GAGE1 человека состоит из создания рекомбинантной плазмиды pGAGE1 на основе рЕТ-30а, экспрессии рекомбинатного белка и его хроматографической очистки и последующей ренатурации и стерилизующей фильтрации. Изобретение позволяет получить рекомбинантный белок человека GAGE1 с выходом очищенного белка 24,5 мг/л культуры E. coli штамма BL21(DE3)pLysS. 3 ил.
Способ получения рекомбинантного белка GAGE1 человека, состоящий из создания рекомбинантной плазмиды, экспрессии рекомбинатного белка и его хроматографической очистки, отличающийся тем, что полноразмерную кодирующую последовательность природного гена GAGE1 человека, полученную с помощью праймеров SEQ ID NO: 1 и 2, клонируют в плазмиду рЕТ-30а по сайтам рестрикции EcoRI и HindIII, получая рекомбинантную плазмиду pGAGE1 размером 5763 п. н., содержащую последовательность SEQ ID NO: 3, кодирующую рекомбинантный белок GAGE1, с сайтами узнавания для рестриктаз EcoRI и HindIII, промотора Т7, гена устойчивости к канамицину, последовательности инициации репликации pBR322(ori), и последовательностей, необходимых для инициации трансляции белка в клетках Е. coli., и получают рекомбинантный белок GAGE1 человека с аминокислотной последовательностью SEQ ID NO: 4, путем его экспрессии в бактериальных клетках E. coli штамма BL21(DE3)pLysS, содержащих плазмиду pGAGE1; при этом очистку рекомбинантного белка GAGE1 человека из штамма-продуцента осуществляют методом металлохелатной аффинной хроматографии с предварительной трехкратной отмывкой нерастворимых белков продуцента с помощью буфера 50 мМ трис-HCl, рН=8,0, 0,2 мМ ЭДТА, 0,05М NaCl и последующей ренатурацией и стерилизующей фильтрацией.
| ФИНАШУТИНА Ю | |||
| П | |||
| и др | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| GJERSTORFF M | |||
| F | |||
| et al | |||
| Expression, purification and characterization of the cancer-germline antigen GAGE12I: a candidate for cancer immunotherapy, Protein expression and purification, 2010 | |||
| V | |||
| Способ подготовки рафинадного сахара к высушиванию | 0 |
|
SU73A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Искусственный двухслойный мельничный жернов | 1921 |
|
SU217A1 |
| GJERSTORFF M | |||
| F | |||
| et al | |||
| An overview of the GAGE cancer/testis antigen family with the inclusion of newly identified members, Tissue Antigens, 2008, V | |||
| Контрольный стрелочный замок | 1920 |
|
SU71A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Индукционная катушка | 1920 |
|
SU187A1 |

