Область техники, к которой относится изобретение
Настоящее изобретение относится к способам разработки сконструированных Т-клеток для иммунотерапии, точнее к способам модификации Т-клеток путем инактивации генов иммунных контрольных точек, предпочтительно по меньшей мере двух, выбранных из разных метаболических путей, чтобы обойти механизмы ингибирования и повысить иммунную активность Т-клеток. Такой способ включает применение специфических редкощепящих эндонуклеаз, в частности TALE-нуклеаз (TAL-эффекторных нуклеаз), и полинуклеотидов, кодирующих такие полипептиды, для точного нацеливания и отбора ключевых генов в Т-клетках, которые могут быть получены от доноров или из культуры первичных клеток. Настоящее изобретение открывает путь к высокоэффективным адаптивным стратегиям иммунотерапии для лечения рака и вирусных инфекций.
Уровень техники
Адаптивная иммунотерапия, которая включает перенос аутологических антиген-специфичных Т-клеток, полученных ex vivo, представляет многообещающую стратегию в лечении вирусных инфекций и рака. Т-клетки, используемые для адаптивной иммунотерапии, могут быть получены или размножением антиген-специфичных Т-клеток, или перенаправлением Т-клеток за счет генетической инженерии (Park и др., 2011). Перенос специфичных к вирусному антигену Т-клеток представляет хорошо разработанную процедуру, используемую для лечения трансплантат-ассоциированных вирусных инфекций и редких связанных с вирусом злокачественных заболеваний. Показано, что аналогичным образом выделение и перенос специфичных к опухолям Т-клеток, успешно в лечении меланомы.
Новые специфические свойства Т-клеток были успешно созданы путем генетического переноса трансгенных рецепторов Т-клеток или химерных антигенных рецепторов (CAR) (Jena и др., 2010). CAR являются синтетическими рецепторами, включающими часть молекулы нацеливания, которая ассоциирована с одним или несколькими сигнальными доменами в одной гибридной молекуле. В общем, связывающая часть молекулы CAR состоит из антигенсвязывающего домена одноцепочечного антитела (single-chain antibody - scFv), включающего легкие и вариабельные фрагменты моноклонального антитела, соединенного гибким линкером. Связывающие части молекулы, основанные на доменах рецептора или лиганда, также успешно применяют. Сигнальные домены для первой генерации CAR производны от цепей цитоплазматической области CD3zeta или Fc рецептора gamma. CAR позволяют успешно перенаправить Т-клетки против антигенов, экспрессируемых на поверхности раковых клеток разных злокачественных образований, включая лимфомы и плотные опухоли (Jena и др., 2010).
Опосредованный Т-клетками иммунитет включает множество последовательных стадий, регулируемых балансом между ко-стимулирующими и ингибирующими сигналами, которые точно регулируют иммунный ответ. Ингибирующие сигналы, называемые иммунными контрольными точками, являются критическими для поддержания аутотолерантности, а также для ограничения иммуноопосредованного повреждения коллатеральных тканей. Способность опухолей кооптировать такие ингибирующие метаболические пути представляет важный механизм иммунной устойчивости и ограничивает успешность иммунотерапии. Одним из обнадеживающих подходов к активированию Т-клеточного иммунного ответа является блокада таких иммунных контрольных точек (Pardoll, 2012). Иммунные контрольные точки представляют существенные преграды для активирования функционального клеточного иммунитета при заболевании раком, и антагонистические антитела, специфичные в отношении ингибирующих лигандов на Т-клетках, включая CTLA4 и белок запрограммированной смерти 1 (PD-1), являются примерами направленных агентов, проходящих оценку в клиниках.
Цитотоксический-Т-лимфоцит-ассоциированный антиген 4 (Cytotoxic-T-lymphocyte-associated antigen 4 - CTLA-4; также обозначаемый CD 152) снижает регуляцию амплитуды активации Т-клеток, и лечение антителами-антагонистами CTLA4 (ипилимумаб) показало преимущественное выживание пациентов с меланомой (Robert и Mateus, 2011). Белок запрограммированной смерти клеток (programmed cell death protein 1 - PD1 или PDCD1, также обозначаемый CD279) представляет другую весьма многообещающую мишень для иммунотерапии (Pardoll и Drake 2012; Pardoll 2012). В отличие от CTLA-4, PD1 ограничивает эффекторные функции Т-клеток в периферической ткани в момент воспалительного ответа на инфекцию и ограничивает аутоиммунность. Первое клиническое испытание PD1 антитела показывает некоторые случаи регрессии опухолей (Brahmer и др., 2010). Множественные дополнительные иммунные белки контрольной точки представляют перспективные мишени для терапевтической блокады, что следует из предшествующих исследований.
Хотя недостаточно ясны точные метаболические пути, по которым передается сигнал белков иммунных контрольных точек, предполагают, что белки иммунных контрольных точек используют другие механизмы для ингибирования активирования Т-клеток и потенциально не являются излишними. Установлено, что применение по меньшей двух разных антител для блокирования множественных лигандов или рецепторов иммунных контрольных точек производит синергические противоопухолевые активности. Тем не менее, степень побочных эффектов повышается среди пациентов, которых лечат комбинированной терапией антител CTLA-4/PD-1 (Wolchok, Kluger и др.).
Чтобы избежать побочных эффектов при лечении опухолей антителами, в настоящем изобретении конструируют высоко активные Т-клетки для иммунотерапии путем инактивации по меньшей мере двух генов, кодирующих белки иммунных контрольных точек. В отличие от блокады иммунных контрольных точек, опосредованной обработкой антителами, контрольные точки сигнальных метаболических путей блокируют непосредственно в сконструированных Т-клетках, используемых для иммунотерапии, при этом побочных эффектов из-за неспецифических эффектов антител на иммунную систему пациента удается избежать.
В частности это стало возможным за счет инактивации генов с помощью специфических TALE-нуклеаз, направленных против белков по меньшей мере двух иммунных контрольных точек, например, PD1 и CTLA-4. Инактивация генов иммунных контрольных точек в Т-лимфоцитах допускает существенную пролиферацию и активность интродуцированных лимфоцитов. Таким образом, ожидают, что такие модифицированные Т-клетки будут высоко активны в крови пациентов, где они смогут нацеливаться на опухолевые клетки или инфицированные клетки.
В дополнение к указанной выше концепции генетически модифицированных Т-клеток, которые могут быть высокоактивными, с помощью конструкции специфических TALE-нуклеаз одновременно инактивируют разные гены в Т-клетках, тем самым, получая двойные мутанты. Собственно говоря, двойное генное нацеливание с помощью двухцепочечных разрывов в Т-клетках было очень далеко от успешного решения из-за трудностей с получением и поддержанием Т-клеток в культуре в течение времени из-за свойственной им низкой степени трансформации и утраты при процедуре селекции. Эти осложнения делают маловероятным успешное получение таких клеток.
Таким образом, одной из важных задач, решаемых в настоящем изобретении, является конструирование специфических TALE-нуклеаз, допускающих повышенные степени двухцепочечных разрывов (DSB) в Т-клетках, которые хорошо переносятся клетками (особенно при ко-трансфекции) и способны направленно отбирать гены по настоящему изобретению. С помощью применения редкощепящих эндонуклеаз, например, описанных в настоящем изобретении TALE-нуклеаз, возможность получения дважды инактивированных генов в трансфецированных Т-клетках существенно повышается, и соответственно в настоящее время появляется возможность получать сконструированные Т-клетки, доступные от донора на регулярной основе по стандартным процедурам.
Краткое описание изобретения
Одна из задач, решаемых в настоящем изобретении, относится к разработке способов конструирования Т-клеток, пригодных для целей иммунотерапии. Способы по настоящему изобретению допускают более точную модификацию генома клеток, релевантных для иммунотерапии, путем инактивирования или замены генов, кодирующих белки иммунных контрольных точек. В некоторых вариантах осуществления настоящего изобретения модифицированные клетки, релевантные для иммунотерапии, дополнительно включают экзогенные рекомбинантные полинуклеотиды, кодирующие химерные антигенные рецепторы (Chimeric Antigen Receptors - CAR) для специфического распознавания клеток. Другая задача, решаемая в настоящем изобретении, относится к полипептидам и полинуклеотидам, которые кодируют редкощепящие эндонуклеазы для точного нацеливания на указанные выше гены, кодирующие белки иммунных контрольных точек, тем самым допуская генетическую модификацию Т-клеток для иммунотерапии. Настоящее изобретение предусматривает более точные специфические целевые последовательности в этих генах и TALE-нуклеазы, сконструированные для соответствующего нацеливания на эти гены.
Настоящее изобретение также относится к выделенным клеткам или линиям клеток, включающим какие-либо из белков, полипептидов или векторов, описанных в настоящем изобретении. В некоторых вариантах осуществления настоящего изобретения Т-клетки по настоящему изобретению включают инактивированные гены иммунных контрольных точек для их применения в иммунотерапии.
В предпочтительном варианте осуществления настоящего изобретения применяют модифицированные Т-клетки в качестве терапевтического продукта, в идеале «имеющегося в продаже». В этом отношении Т-клетки по настоящему изобретению могут быть дополнительно генетически модифицированы, например, путем инактивации по меньшей мере одного гена, кодирующего компонент TCR (Т cell receptor - Т-клеточный рецептор), для получения аллогенных клеток и тем самым пригодных для лечения различных пациентов из образца одного донора.
В другом варианте осуществления настоящее изобретение относится к способу лечения или предупреждения рака или инфекций у пациента путем введения сконструированных Т-клеток, получаемых описанными выше способами.
Краткое описание фигур и таблиц
Помимо перечисленных признаков объектов, настоящее изобретение дополнительно предусматривает другие признаки, которые могут проявляться исходя из последующего описания и прилагаемых иллюстраций. Более полная оценка настоящего изобретения и его преимуществ поясняется приводимым ниже фигурами и подробным описанием.
Фиг. 1. Схематическое описание нормального взаимодействия между Т-клетками и антиген-презентирующими клетками.
Фиг. 2. Схематическое описание генетически модифицированных терапевтических Т-клеток по настоящему изобретению и опухолевых клеток пациента.
Фиг. 3. Схематическое описание многоцепочечного CAR.
Фиг. 4. Схематическое описание разных версий многоцепочечных CAR. А. Схематическое описание рецептора FcεRI. Б, В. Разные версии многоцепочечных CAR (от csm1 до csm10), включающих scFv и стволовую область CD8, гибридизированных с трансмембранным доменом FcεRI alpha цепи. По меньшей мере один из доменов, 41ВВ, CD28 и/или CD3 zeta, может быть гибридизирован с FcεRI alpha, beta и/или gamma цепью.
Фиг. 5. Схематическое описание одного из примеров способа конструирования аллогенных клеток человека для иммунотерапии.
Фиг. 6. Концентрация в клетках/мл живых CD52-положительных или CD52-отрицательных клеток после лечения анти-CD52 антителом (САМРАТН1-Н) с комплементом или контролями.
Фиг. 7. Сравнение прямого светорассеяния (forward side scatter - FSC) - индикатора размера клеток, между TCR-положительными и TCR-отрицательными клетками, или между CD52- положительными и CD52-отрицательными клетками, и неактивированными клетками в качестве контроля.
Фиг. 8. Анализ методом жидкостной цитометрии экспрессии CD107a (маркера дегрануляции) на целевых CD52 и TCRalpha инактивированных Т-клетках. Экспрессию CD107 анализируют на CD52+TCRαβ+ клетках (первая колонка), CD52-TCRαβ- клетках (вторая колонка), CD52-TCRαβ+ клетках (третья колонка) и CD52+TCRαβ- клетках (четвертая колонка) до (А) и после инкубации с клетками Дауди (Б); (В) представляет анализ методом жидкостной цитометрии Т-клеток, дополнительно трансфецированных CAR и инкубированных с клетками Дауди; (Г) представляет анализ методом жидкостной цитометрии Т-клеток, трансфецированных CAR, но не инкубировавшихся с клетками Дауди, и (Д) представляет анализ методом жидкостной цитометрии Т-клеток, трансфецированных CAR и обработанных РМА/иономицином (положительный контроль).
Фиг. 9. Глубокий сиквенс-анализ потенциала CD52 и TRAC TALE-нуклеаз в отношении мишеней вне сайта.
Фиг. 10. Анализ PDCD1 и CTLA-4 геномного локуса путем исследования с применением Т7-эндонуклеазы. Стрелки указывают на расщепленные ПЦР продукты.
Фиг. 11. Схематическое описание некоторых примеров preTalpha конструкций.
Фиг. 12. Анализ методом жидкостной цитометрии эффективности трансдукции (% BFP + клеток) и активности конструкций FL, Δ18, Δ48 pTalpha (% CD3 экспрессии на поверхности) на TCR alpha инактивированных клетках Jurkat.
Фиг. 13. Схематическое описание лентивирусной конструкции, кодирующей pTalpha белок (preTCRα).
Фиг. 14. А. Протокол проведения эксперимента. Б. Анализ методом жидкостной цитометрии TCR alpha/beta, CD3 экспрессии и BFP экспрессии на TCRalpha инактивированных Т-клетках (KО), трансдуцированных или BFP-2A-pTalphaΔ48 (KО/Δ48), или контрольным BFP лентивирусным вектором (KO/BFP), до и после очистки. В. Анализ методом жидкостной цитометрии TCR alpha/beta и CD3 экспрессии на очищенных TCR alpha инактивированных клетках, трансдуцированных (BFP положительных) или не трансдуцированных (BFP отрицательных) BFP-2A-pTalphaΔ48 лентивирусным вектором. NEP (non electroporated) обозначает не подвергнутые электропортации клетки TRAC ТALE-нуклеазами.
Фиг. 15А, Б. Анализ методом жидкостной цитометрии экспрессии раннего маркера активации CD69 (А), позднего маркера активации CD25 (Б) через 24 и 48 ч после реактивации гранулами анти-CD3/CD28, соответственно, на неэлектропорированных клетках (NEP) и TCRalpha инактивированных клетках (KО), трансдуцированных BFP-2A-pTα-Δ48 лентивирусным вектором (рТα-Δ48), BFP-2A-pTα-Δ48.41BB лентивирусным вектором (pTα-Δ48.ВВ) или контрольным BFP вектором (BFP). pTα-Δ48 гистограммы соответствуют сигналу, выявляемому в TCR-инактивированных клетках, экспрессирующих pTα-Δ48 (BFP + клетки), в то время как KО гистограммы соответствуют TCRalpha инактивированным клеткам, которые не экспрессируют pTα-Δ48 (BFP- клетки). pTα-Δ48.ВВ гистограммы соответствуют сигналу, выявляемому в TCR-инактивированных клетках, экспрессирующих pTα-Δ48.41 ВВ (BFP + клетки), в то время как KО гистограммы соответствуют TCRalpha инактивированным клеткам, которые не экспрессируют pTα-Δ48.41 ВВ (BFP- клетки). NEP (не электропорированные) гистограммы соответствуют сигналу, выявляемому в неконструированных клетках. В. Анализ методом жидкостной цитометрии размера клеток через 72 ч после реактивации анти-CD3/CD28 гранулами на не электропорированных клетках (NEP) и TCRalpha инактивированных клетках (KО), трансдуцированных BFP-2A-pTα-Δ48 лентивирусным вектором (pTα-Δ48), BFP-2A-pTα-Δ48.41BB лентивирусным вектором (pTα-Δ48.ВВ) или контрольным BFP вектором (BFP). Величины, указанные в верхней части каждой графы, соответствуют среднему геометрическому значению флуоресценции каждой популяции.
Фиг. 16. Анализ роста TCR alpha инактивированных клеток (KО), трансдуцированных pTalpha-Δ48 (pTaΔ48) или контрольным вектором BFP (BFP) при поддержке IL2 или IL2 с анти-CD3/CD28 гранулами в разное время (х-ось). Число BFP + клеток оценивают в разное время для каждого состояния и кратность индукции этих клеток (у-ось) оценивают относительно величины, полученной на вторые сутки после реактивации. Результаты получают от двух независимых доноров. Для второго донора также определяют рост клеток, трансдуцированных pTalpha-Δ48.41BB (pTa-Δ48.ВВ) и pTalpha- (pTa-FL) полной длины.
Фиг. 16. Анализ роста TCR alpha инактивированных клеток (KО), трансдуцированных pTalpha-Δ48 (pTaΔ48) или контрольным вектором BFP (BFP) при поддержке IL2 или IL2 с анти-CD3/CD28 гранулами в разное время (х-ось). Число BFP + клеток оценивают в разное время для каждого состояния и кратность индукции этих клеток (у-ось) оценивают относительно величины, полученной на вторые сутки после реактивации. Результаты получают от двух независимых доноров. Для второго донора также определяют рост клеток, трансдуцированных pTalpha-Δ48.41BB (pTa-Δ48.ВВ) и pTalpha- (pTa-FL) полной длины.
Фиг. 17. Анализ методом жидкостной цитометрии GFP-положительных клеток среди МКПК, подвергнутых электропорации по пяти разным программам Cytopulse. Верхняя линия соответствует трансфекции 6×106 клеток на кювету, а нижняя линия соответствует трансфекции 3×106 клеток на кювету.
Фиг. 18. Анализ методом жидкостной цитометрии гибели очищенных Т-клеток, используя прижизненный краситель (eFluor-450) и GFP-положительные клетки среди жизнеспособной популяции после электропорации GFP мРНК, GFP ДНК и контрольной pUC ДНК. NEP соответствует клеткам, которые поддерживают в буфере для электропорации, но они не подвергались электропорации, и NT соответствует клеткам, не подвергавшимся электропорации, поддерживаемым в культуральной среде.
Фиг. 19. Анализ методом жидкостной цитометрии TCR alpha/beta и CD3 экспрессии на первичных Т-клетках человека после электропорации мРНК TRAC TALE-нуклеазы (вверху). Глубокий сиквенс-анализ геномной ДНК, экстрагированной из первичных Т-клеток человека после электропорации мРНК TRAC TALE-нуклеазы (внизу).
Фиг. 20. А. Анализ методом жидкостной цитометрии CAR экспрессии (анти-F(ab')2) после электропорации Т-клеток мРНК, или без мРНК, кодирующей одноцепочечный CAR. Б. Анализ методом жидкостной цитометрии CD107а экспрессии (маркера дегрануляции) на электропортированных Т-клетках, совместно культивируемых с клетками Дауди.
Фиг. 21. А. Представление мРНК, кодирующей многоцепочечный CAR. Б. Анализ методом жидкостной цитометрии CAR экспрессии (анти-F(ab')2) на жизнеспособных Т-клетках, подвергнутых электропорации полицистронной мРНК, или без нее, кодирующей многоцепочечный CAR. В. Анализ методом жидкостной цитометрии CD107a экспрессии (маркера дегрануляции) на электропорированных Т-клетках, совместно культивируемых с клетками Дауди.
Фиг. 22. Экспрессия многоцепочечных CAR в Т-клетках человека после электропорации полицистронных мРНК.
Фиг. 23. Экспрессия мульти-субъединичных CAR обусловлена экспрессией трех цепей: α, β и γ.
Фиг. 24. Т-клетки человека, временно экспрессирующие многоцепочечные CAR, дегранулируют после совместного культивирования с целевыми клетками. А: от csm1 до csm5 CAR конструкции. Б: от csm6 до csm10 CAR конструкции.
Фиг. 25. Т-клетки человека, временно экспрессирующие многоцепочечные CAR, секретируют цитокины после совместного культивирования с целевыми клетками (Т-клетки против клеток Дауди или K562). А: высвобождение IL8. Б: высвобождение IFNγ. В: высвобождение IL5.
Фиг. 26. Т-клетки человека, временно экспрессирующие многоцепочечные CAR (конструкции от csm1 по csm10), лизируют целевые клетки.
Фиг. 27. Инактивирование CTLA4 в первичных Т-клетках, измеренное путем внутриклеточного окрашивания с применением флуоресцентного антитела и жидкостного цитометрического анализа.
Фиг. 28. Распределение флуоресцентных Т-клеток, экспрессирующих CTLA4, при трансфекции TALEN T1, Т2 и Т3. Пропорция клеток, экспрессирующих CTLA4, резко снижается относительно контрольных клеток.
Фиг. 29. Инактивация PD1 в первичных Т-клетках, измеренная путем внутриклеточного окрашивания флуоресцентным антителом с применением метода жидкостной цитометрии. Пропорция клеток, экспрессирующих PD1, резко снижается относительно контрольных клеток.
Фиг. 30. Диаграмма, показывающая частоту делеций, наблюдаемых в Т-клетках при трансфекции TALEN Т01 и Т03, нацеленных на ген PD1.
Фиг. 31. Диаграмма, показывающая, что цитотоксическая активность возрастает в Т-клетках с разрушенным PD1, как в эксперименте, описанном в примере 3.
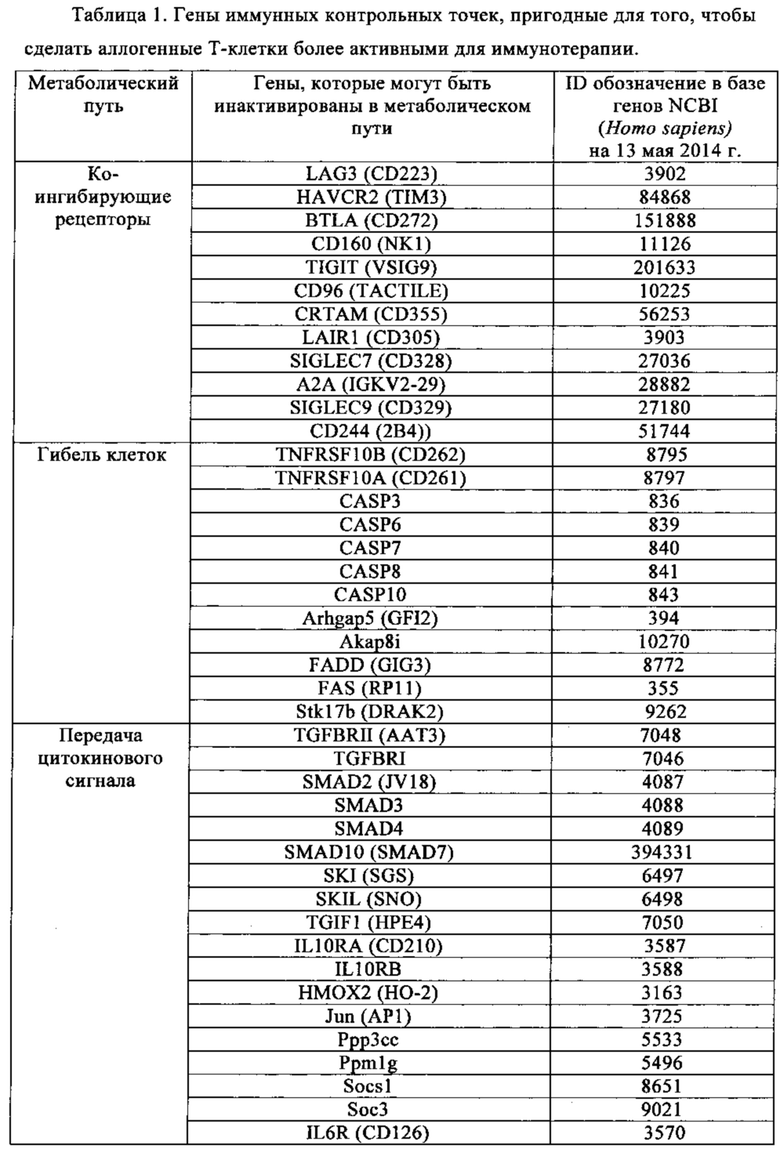
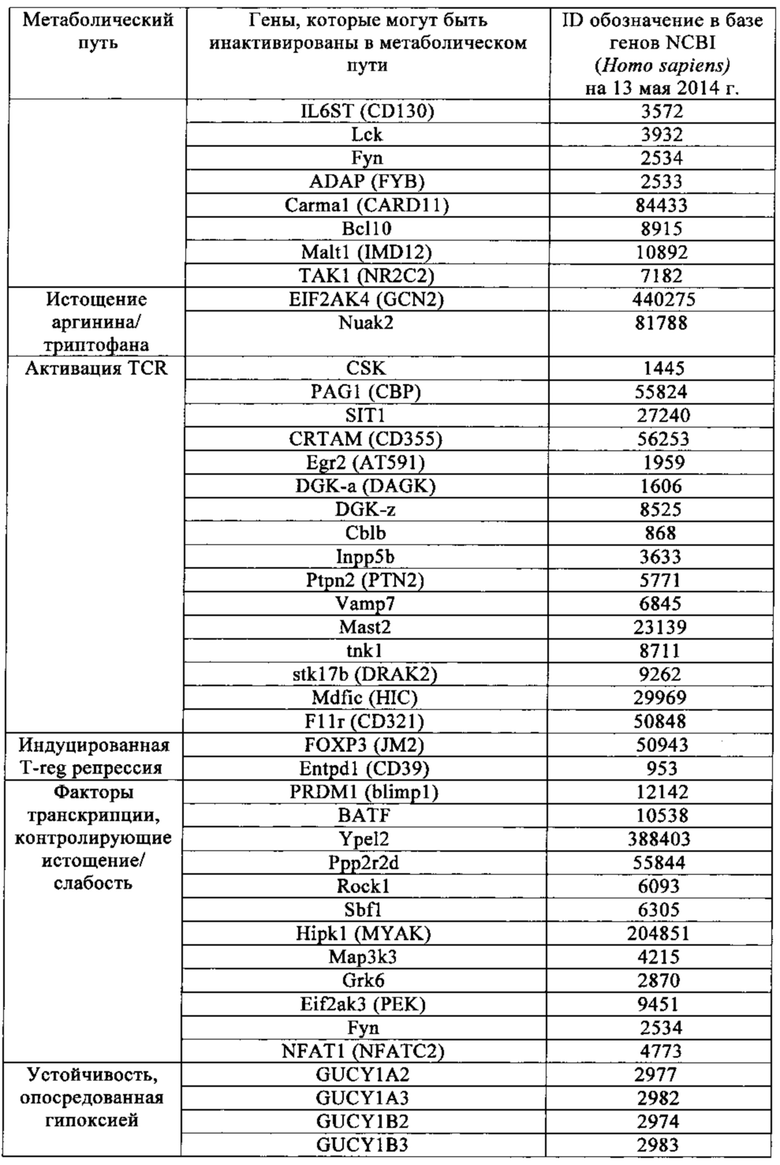
Таблица 1. Перечень генов иммунных контрольных точек, идентифицированных в настоящем изобретении в качестве пригодных для получения аллогенных Т-клеток более активных для иммунотерапии.
Таблица 2. Описание GR TALE-нуклеаз и последовательностей целевых сайтов TALE-нуклеаз в гене человека GR.
Таблица 3. Расщепляющая активность GR TALE-нуклеаз в дрожжах. Величины находятся в диапазоне от 0 до 1. Максимальная величина равна 1.
Таблица 4. Процент направленного мутагенеза в сайтах-мишенях эндогенной TALE-нуклеазы в клетках 293.
Таблица 5. Процент направленного мутагенеза в эндогенных сайтах-мишенях TALE-нуклеазы в первичных Т-лимфоцитах.
Таблица 6. Описание CD52, TRAC и TRBC TALE-нуклеаз и последовательностей сайтов-мишеней TALE-нуклеаз в соответствующих генах человека.
Таблица 7. Дополнительные целевые последовательности TRAC и CD52 TALE-нуклеаз.
Таблица 8. Процент инсерций/делеций для нацеливания TALE-нуклеаз на мишени CD52_T02, TRAC_T01, TRBC_T01 и TRBC_T02.
Таблица 9. Процент СD52-отрицательных, TCR-отрицательных и CD52/TCR-дважды отрицательных Т-лимфоцитов после трансфекции соответствующих TALE-нуклеаза-экспрессирующих полинуклеотидов.
Таблица 10. Процент TCR-отрицательных Т-лимфоцитов после трансфекции TRBC TALE-нуклеаза-экспрессирующих полинуклеотидов.
Таблица 11. Описание CTLA4 и PDCD1 TALE-нуклеаз и последовательностей сайтов-мишеней TALE-нуклеаз в соответствующих генах человека.
Таблица 12. Описание подгруппы конструкций pTalpha.
Таблица 13. Активность разных pTalpha конструкций в TCR alpha инактивированных клетках Jurkat. Активность измеряют методом жидкостной цитометрии CD3 экспрессии на TCR alpha инактивированных клетках Jurkat, трансфецированных разными конструкциями preTalpha.
Таблица 14. Разные программы Cytopulse, применяемые для определения минимального напряжения, требуемого для электропорации в Т-клетки, производные от мононуклеарных клеток периферической крови (МКПК).
Таблица 15. Программа Cytopulse, используемая для электропорации очищенных Т-клеток.
Таблица 16. Перечень генов, кодирующих белки иммунных контрольных точек.
Подробное описание изобретения
Если специально не указано иначе, все технические и научные термины имеют то же значение, что и обычно употребляемое специалистами в области генной терапии, биохимии, генетики и молекулярной биологии.
Все методы и материалы, близкие или равные используемым в настоящем изобретении, могут применяться на практике или при тестировании настоящего изобретения, наряду с методами и материалами, описанными для применения в настоящем изобретении. Все публикации, патентные заявки, патенты и другие источники, упоминаемые в настоящем изобретении, включены в него в виде ссылок на их целостную сущность. Особенно это касается объекта изобретения, раскрываемого в каждом из источников, который относится к объекту изобретения в примыкающих предложениях, параграфах или разделах, в которых источник цитируется. В случае конфликта настоящее описание, включая определения, превалирует. Кроме того, все материалы, методы и примеры являются только иллюстрацией и не ограничивают настоящее изобретение, если не указано иначе.
В практике настоящего изобретения применяют, если не указано иначе, традиционные методы клеточной биологии, культур клеток, молекулярной биологии, трансгенной биологии, микробиологии, рекомбинантной ДНК и иммунологии, которые известны специалистам в данной области. Эти методы подробно описаны в литературе. См., например, Current Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA); Molecular Cloning: A Laboratory Manual, 3-е изд., (Sambrook и др., 2001, Cold Spring Harbor, Нью-Йорк: Cold Spring Harbor Laboratory Press); Oligonucleotide Synthesis (под ред. M.J. Gait, 1984); Mullis и др., US4683195; Nucleic Acid Hybridization (под ред. В. D. Harries и S.J. Higgins, 1984); Transcription And Translation (под ред. В.D. Hames и S.J. Higgins, 1984); Culture Of Animal Cells (R. I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); серии, Methods In ENZYMOLOGY (под ред. J. Abelson и M. Simon, Academic Press, Inc., Нью-Йорк), в особенности Т. 154, 155 (под ред. Wu и др.) и Т. 185 "Gene Expression Technology" (под ред. D. Goeddel); Gene Transfer Vectors For Mammalian Cells (под ред. J.H. Miller и M.P. Calos, 1987, Cold Spring Harbor Laboratory); Immunochemical Methods In Cell And Molecular Biology (под ред. Mayer и Walker, Academic Press, Лондон, 1987); Handbook Of Experimental Immunology, T. I-IV (под ред. D.M. Weir и С.С. Blackwell, 1986); Manipulating the Mouse Embryo (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Нью-Йорк, 1986).
Главная задача настоящего изобретения заключается в разработке способов для новых адаптивных стратегий лечения рака и инфекций.
Высоко активные Т-клетки для иммунотерапии
Для решения первой задачи, выдвинутой для решения в настоящем изобретении, были выявлены новые гены, кодирующие так называемые «иммунные контрольные точки», функция которых может быть отдельно нарушена для повышения эффективности Т-клеток для адаптивной иммунотерапии. Ниже в табл. 1 перечислены упомянутые новые целевые гены, которые более конкретно вовлечены в функции ко-ингибирующего рецептора, гибели клеток, передачи сигнала через цитокин, истощения аргинина/триптофана, передачи сигнала через TCR, индуцированной T-reg репрессии, факторов транскрипции, контролирующие истощение или анемию, и опосредованной гипоксией устойчивости. Ранее упоминалось, что нацеливание на одну из этих более поздних функций путем нарушения или контроля экспрессии одного из этих генов может быть осуществлено как часть аутологичной или аллогенной схемы лечения.
Точнее настоящее изобретение относится к сконструированным Т-клеткам, в которых по меньшей мере один из генов, идентичный с одним из генов в табл. 1, нарушен или репрессирован с целью применения в иммунотерапии.
Опосредованный Т-клетками иммунитет содержит множество последовательных стадий, включая клональную селекцию антигенспецифичных клеток, их активацию и пролиферацию во вторичной лимфоидной ткани, их движение к сайтам антигена и воспалению, достижение прямой эффекторной функции и предоставление помощи (через цитокины и мембранные лиганды) множеству эффекторных иммунных клеток. Каждая и этих стадий регулируется противовесом стимулирующего и ингибирующего сигнала, который тонким образом настраивает ответ. Хотя механизм ингибирования сигналов в большинстве случаев не обнаружен, установлено, что компоненты отрицательного регулирования могут ослаблять ответ Т-клеток более чем по одному пути. Например, CTLA-4 может конкурировать с CD28 за костимулирующие лиганды. Напротив, молекулы, содержащие ингибирующий мотив, например PD1, могут рекрутировать фосфатазы (например, такие как SHP-1, SHP-2 или SHIP) к TCR-проксимальным сигнальным комплексам и ослаблять передачу сигнала. И PD-1, и CTLA-4, могут ингибировать передачу сигнала через серин-треониновую киназу Akt, но предположительно делают это по разным молекулярным механизмам. В итоге инактивация одного из генов может быть компенсирована другим отрицательным регуляторным метаболическим путем. Чтобы обойти это, в настоящем изобретении наряду с решением другой задачи стремились инактивировать по меньшей мере, два гена, кодирующие белки иммунных контрольных точек, чтобы в достаточной степени увеличить активность Т-клеток для иммунотерапии. На основе классификации, показанной в табл. 1, были выбраны различные гены, кодирующие белки, которые предпочтительно участвуют в различных ингибирующих метаболических путях и отрицательно регулируют иммунный ответ.
Настоящее изобретение относится к способу получения Т-клеток для иммунотерапии путем инактивирования в Т-клетках по меньшей мере двух генов, кодирующих белки иммунных контрольных точек для существенного повышения активности Т-клеток для иммунотерапии. В частности, настоящее изобретение относится к способу получения Т-клеток для иммунотерапии, включающему:
(а) модифицирование Т-клеток путем инактивации по меньшей мере двух генов, кодирующих белки иммунных контрольных точек,
(б) размножение указанных клеток.
Специалистам в данной области известно, что термин «иммунные контрольные точки» означает группу молекул, экспрессируемых Т-клетками. Эти молекулы эффективно служат в качестве «тормозов» для снижения модуляции или ингибирования иммунного ответа. К молекулам иммунных контрольных точек относятся, но ими перечень не ограничивается, белок запрограммированной смерти 1 (Programmed Death 1 - PD-1, также обозначаемый PDCD1 или CD279, номер доступа: NM_005018), антиген цитотоксического Т-лимфоцита 4 (Cytotoxic T-Lymphocyte Antigen 4 - CTLA-4, также обозначаемый CD152, GenBank номер доступа: AF414120.1), белок LAG3 (также обозначаемый CD223, номер доступа: NM_002286.5), Tim3 (также обозначаемый HAVCR2, GenBank номер доступа: JX049979.1), BTLA (также обозначаемый CD272, номер доступа: NM_181780.3), BY55 (также обозначаемый CD160, GenBank номер доступа: CR541888.1), TIGIT (также обозначаемые VSTM3, номер доступа: NM_173799), LAIR1 (также обозначаемый CD305, GenBank номер доступа: CR542051.1), (Meyaard, Adema и др. 1997)), SIGLEC10 (GeneBank номер доступа: AY358337.1), 2В4 (также обозначаемый CD244, номер доступа: NM_001166664.1), РРР2СА, РРР2СВ, PTPN6, PTPN22, CD96, CRTAM, SIGLEC7 (Nicoll, Ni et al. 1999), SIGLEC9 (Zhang, Nicoll и др., 2000; Ikehara, Ikehara и др., 2004), TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT1, FOXP3, PRDM1, BATF (Quigley, Pereyra и др., 2010), GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3, которые непосредственно ингибируют иммунные клетки. Например, CTLA-4 является белком на поверхности клетки, экспрессируемым на определенных CD4 и CD8 Т-клетках; при связывании собственными лигандами (В7-1 и В7-2) на антиген-презентирующих клетках, подавляются активация и эффекторная функция Т-клеток. Таким образом, настоящее изобретение относится к способу конструирования Т-клеток специально для иммунотерапии, включая генетически модифицированные Т-клетки, путем инактивации по меньшей мере одного белка, участвующего в иммунной контрольной точке, в частности PD1 и/или CTLA-4.
Приведенная ниже табл. 1, которая не является исчерпывающей, показывает гены иммунных контрольных точек, которые могут быть инактивированы согласно изложенному в настоящем изобретении для того, чтобы улучшить эффективность и соответствие сконструированных Т-клеток. Ген иммунных контрольных точек предпочтительно выбран из генов, обладающих идентичностью по отношению к тем генам, которые перечислены в этой таблице и участвуют в функции коингибирующего рецептора, гибели клеток, передаче цитокиновых сигналов, истощении аргинина и триптофана, активации TCR, индуцированной T-reg репрессии, контроля факторами транскрипции истощения или слабости, и устойчивости, опосредованной гипоксией.
В предпочтительном варианте осуществления настоящего изобретения указанные два гена, кодирующие белки иммунных контрольных точек, выбраны из группы, включающей: CTLA4, РРР2СА, РРР2СВ, PTPN6, PTPN22, PDCD1, LAG3, HAVCR2, BTLA, CD160, TIGIT, CD96, CRTAM, LAIR1, SIGLEC7, SIGLEC9, CD244, TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT1, FOXP3, PRDM1, BATF, GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3.
В некоторых вариантах осуществления настоящего изобретения указанные два гена, кодирующие белки иммунных контрольных точек, выбраны из группы, включающей: BTLA и BATF, CASP10 и BATF, CASP10 и BTLA, CASP3 и BATF, CASP3 и BTLA, CASP3 и СASP10, CASP6 и BATF, CASP6 и BTLA, CASP6 и CASP10, CASP6 и CASP3, CASP7 и BATF, CASP7 и BTLA, CASP7 и СASP10, CASP7 и CASP3, CASP7 и CASP6, CASP8 и BATF, CASP8 и BTLA, CASP8 и CASP10, CASP8 и CASP3, CASP8 и CASP6, CASP8 и CASP7, CD160 и BATF, CD160 и BTLA, CD160 и CASP10, CD160 и CASP3, CD160 и CASP6, CD160 и CASP7, CD160 и CASP8, CD244 и BATF, CD244 и BTLA, CD244 и CASP10, CD244 и CASP3, CD244 и CASP6, CD244 и CASP7, CD244 и CASP8, CD244 и CD160, CD96 и BATF, CD96 и BTLA, CD96 и CASP10, CD96 и CASP3, CD96 и CASP6, CD96 и CASP7, CD96 и CASP8, CD96 и CD160, CD96 и CD244, CRTAM и BATF, CRTAM и BTLA, CRTAM и СASP10, CRTAM и CASP3, CRTAM и CASP6, CRTAM и CASP7, CRTAM и CASP8, CRTAM и CD160, CRTAM и CD244, CRTAM и CD96, CSK и BATF, CSK и BTLA, CSK и СASP10, CSK и CASP3, CSK и CASP6, CSK и CASP7, CSK и CASP8, CSK и CD160, CSK и CD244, CSK и CD96, CSK и CRTAM, CTLA4 и BATF, CTLA4 и BTLA, CTLA4 и CASP10, CTLA4 и CASP3, CTLA4 и CASP6, CTLA4 и CASP7, CTLA4 и CASP8, CTLA4 и CD160, CTLA4 и CD244, CTLA4 и CD96, CTLA4 и CRTAM, CTLA4 и CSK, EIF2AK4 и BATF, EIF2AK4 и BTLA, EIF2AK4 и С ASP10, EIF2AK4 и CASP3, EIF2AK4 и CASP6, EIF2AK4 и CASP7, EIF2AK4 и CASP8, EIF2AK4 и CD160, EIF2AK4 и CD244, EIF2AK4 и CD96, EIF2AK4 и CRTAM, EIF2AK4 и CSK, EIF2AK4 и CTLA4, FADD и BATF, FADD и BTLA, FADD и СASP10, FADD и CASP3, FADD и CASP6, FADD и CASP7, FADD и CASP8, FADD и CD160, FADD и CD244, FADD и CD96, FADD и CRTAM, FADD и CSK, FADD и CTLA4, FADD и EIF2AK4, FAS и BATF, FAS и BTLA, FAS и CASP10, FAS и CASP3, FAS и CASP6, FAS и CASP7, FAS и CASP8, FAS и CD160, FAS и CD244, FAS и CD96, FAS и CRTAM, FAS и CSK, FAS и CTLA4, FAS и EIF2AK4, FAS и FADD, FOXP3 и BATF, FOXP3 и BTLA, FOXP3 и СASP10, FOXP3 и CASP3, FOXP3 и CASP6, FOXP3 и CASP7, FOXP3 и CASP8, FOXP3 и CD160, FOXP3 и CD244, FOXP3 и CD96, FOXP3 и CRTAM, FOXP3 и CSK, FOXP3 и CTLA4, FOXP3 и EIF2AK4, FOXP3 и FADD, FOXP3 и FAS, GUCY1A2 и BATF, GUCY1A2 и BTLA, GUCY1A2 и CASP10, GUCY1A2 и CASP3, GUCY1A2 и CASP6, GUCY1A2 и CASP7, GUCY1A2 и CASP8, GUCY1A2 и CD160, GUCY1A2 и CD244, GUCY1A2 и CD96, GUCY1A2 и CRTAM, GUCY1A2 и CSK, GUCY1A2 и CTLA4, GUCY1A2 и EIF2AK4, GUCY1A2 и FADD, GUCY1A2 и FAS, GUCY1A2 и FOXP3, GUCY1A3 и BATF, GUCY1A3 и BTLA, GUCY1A3 и СASP10, GUCY1A3 и CASP3, GUCY1A3 и CASP6, GUCY1A3 и CASP7, GUCY1A3 и CASP8, GUCY1A3 и CD160, GUCY1A3 и CD244, GUCY1A3 и CD96, GUCY1A3 и CRTAM, GUCY1A3 и CSK, GUCY1A3 и CTLA4, GUCY1A3 и EIF2AK4, GUCY1A3 и FADD, GUCY1A3 и FAS, GUCY1A3 и FOXP3, GUCY1A3 и GUCY1A2, GUCY1B2 и BATF, GUCY1B2 и BTLA, GUCY1B2 и CASP10, GUCY1B2 и CASP3, GUCY1B2 и CASP6, GUCY1B2 и CASP7, GUCY1B2 и CASP8, GUCY1B2 и CD160, GUCY1B2 и CD244, GUCY1B2 и CD96, GUCY1B2 и CRTAM, GUCY1B2 и CSK, GUCY1B2 и CTLA4, GUCY1B2 и EIF2AK4, GUCY1B2 и FADD, GUCY1B2 и FAS, GUCY1B2 и FOXP3, GUCY1B2 и GUCY1A2, GUCY1B2 и GUCY1A3, GUCY1B3 и BATF, GUCY1B3 и BTLA, GUCY1B3 и CASP10, GUCY1B3 и CASP3, GUCY1B3 и CASP6, GUCY1B3 и CASP7, GUCY1B3 и CASP8, GUCY1B3 и CD160, GUCY1B3 и CD244, GUCY1B3 и CD96, GUCY1B3 и CRTAM, GUCY1B3 и CSK, GUCY1B3 и CTLA4, GUCY1B3 и EIF2AK4, GUCY1B3 и FADD, GUCY1B3 и FAS, GUCY1B3 и FOXP3, GUCY1B3 и GUCY1A2, GUCY1B3 и GUCY1A3, GUCY1B3 и GUCY1B2, HAVCR2 и BATF, HAVCR2 и BTLA, HAVCR2 и СASP10, HAVCR2 и CASP3, HAVCR2 и CASP6, HAVCR2 и CASP7, HAVCR2 и CASP8, HAVCR2 и CD160, HAVCR2 и CD244, HAVCR2 и CD96, HAVCR2 и CRTAM, HAVCR2 и CSK, HAVCR2 и CTLA4, HAVCR2 и EIF2AK4, HAVCR2 и FADD, HAVCR2 и FAS, HAVCR2 и FOXP3, HAVCR2 и GUCY1A2, HAVCR2 и GUCY1A3, HAVCR2 и GUCY1B2, HAVCR2 и GUCY1B3, НМОХ2 и BATF, НМОХ2 и BTLA, НМОХ2 и CASP10, HMOX2 и CASP3, HMOX2 и CASP6, НМОХ2 и CASP7, НМОХ2 и CASP8, HMOX2 и CD160, НМОХ2 и CD244, НМОХ2 и CD96, НМОХ2 и CRTAM, НМОХ2 и CSK, НМОХ2 и CTLA4, НМОХ2 и EIF2AK4, НМОХ2 и FADD, НМОХ2 и FAS, НМОХ2 и FOXP3, НМОХ2 и GUCY1A2, НМОХ2 и GUCY1A3, НМОХ2 и GUCY1B2, НМОХ2 и GUCY1B3, НМОХ2 и HAVCR2, IL10RA и BATF, IL10RA и BTLA, IL10RA и CASP10, IL10RA и CASP3, IL10RA и CASP6, IL10RA и CASP7, IL10RA и CASP8, IL10RA и CD160, IL10RA и CD244, IL10RA и CD96, IL10RA и CRTAM, IL10RA и CSK, IL10RA и CTLA4, IL10RA и EIF2AK4, IL10RA и FADD, IL10RA и FAS, IL10RA и FOXP3, IL10RA и GUCY1A2, IL10RA и GUCY1A3, IL10RA и GUCY1B2, IL10RA и GUCY1B3, IL10RA и HAVCR2, IL10RA и НМОХ2, IL10RB и BATF, IL10RB и BTLA, IL10RB и CASP10, IL10RB и CASP3, IL10RB и CASP6, IL10RB и CASP7, IL10RB и CASP8, IL10RB и CD160, IL10RB и CD244, IL10RB и CD96, IL10RB и CRTAM, IL10RB и CSK, IL10RB и CTLA4, IL10RB и EIF2AK4, IL10RB и FADD, IL10RB и FAS, IL10RB и FOXP3, IL10RB и GUCY1A2, IL10RB и GUCY1A3, IL10RB и GUCY1B2, IL10RB и GUCY1B3, IL10RB и HAVCR2, IL10RB и НМОХ2, IL10RB и IL10RA, IL6R и BATF, IL6R и BTLA, IL6R и CASP10, IL6R и CASP3, IL6R и CASP6, IL6R и CASP7, IL6R и CASP8, IL6R и CD160, IL6R и CD244, IL6R и CD96, IL6R и CRTAM, IL6R и CSK, IL6R и CTLA4, IL6R и EIF2AK4, IL6R и FADD, IL6R и FAS, IL6R и FOXP3, IL6R и GUCY1A2, IL6R и GUCY1A3, IL6R и GUCY1B2, IL6R и GUCY1B3, IL6R и HAVCR2, IL6R и НМОХ2, IL6R и IL10RA, IL6R и IL10RB, IL6ST и BATF, IL6ST и BTLA, IL6ST и CASP10, IL6ST и CASP3, IL6ST и CASP6, IL6ST и CASP7, IL6ST и CASP8, IL6ST и CD160, IL6ST и CD244, IL6ST и CD96, IL6ST и CRTAM, IL6ST и CSK, IL6ST и CTLA4, IL6ST и EIF2AK4, IL6ST и FADD, IL6ST и FAS, IL6ST и FOXP3, IL6ST и GUCY1A2, IL6ST и GUCY1A3, IL6ST и GUCY1B2, IL6ST и GUCY1B3, IL6ST и HAVCR2, IL6ST и НМОХ2, IL6ST и IL10RA, IL6ST и IL10RB, IL6ST и IL6R, LAG3 и BATF, LAG3 и BTLA, LAG3 и CASP10, LAG3 и CASP3, LAG3 и CASP6, LAG3 и CASP7, LAG3 и CASP8, LAG3 и CD160, LAG3 и CD244, LAG3 и CD96, LAG3 и CRTAM, LAG3 и CSK, LAG3 и CTLA4, LAG3 и EIF2AK4, LAG3 и FADD, LAG3 и FAS, LAG3 и FOXP3, LAG3 и GUCY1A2, LAG3 и GUCY1A3, LAG3 и GUCY1B2, LAG3 и GUCY1B3, LAG3 и HAVCR2, LAG3 и НМОХ2, LAG3 и IL10RA, LAG3 и IL10RB, LAG3 и IL6R, LAG3 и IL6ST, LAIR1 и BATF, LAIR1 и BTLA, LAIR1 и CASP10, LAIR1 и CASP3, LAIR1 и CASP6, LAIR1 и CASP7, LAIR1 и CASP8, LAIR1 и CD160, LAIR1 и CD244, LAIR1 и CD96, LAIR1 и CRTAM, LAIR1 и CSK, LAIR1 и CTLA4, LAIR1 и EIF2AK4, LAIR1 и FADD, LAIR1 и FAS, LAIR1 и FOXP3, LAIR1 и GUCY1A2, LAIR1 и GUCY1A3, LAIR1 и GUCY1B2, LAIR1 и GUCY1B3, LAIR1 и HAVCR2, LAIR1 и HMOX2, LAIR1 и IL10RA, LAIR1 и IL10RB, LAIR1 и IL6R, LAIR1 и IL6ST, LAIR1 и LAG3, PAG1 и BATF, PAG1 и BTLA, PAG1 и CASP10, PAG1 и CASP3, PAG1 и CASP6, PAG1 и CASP7, PAG1 и CASP8, PAG1 и CD160, PAG1 и CD244, PAG1 и CD96, PAG1 и CRTAM, PAG1 и CSK, PAG1 и CTLA4, PAG1 и EIF2AK4, PAG1 и FADD, PAG1 и FAS, PAG1 и FOXP3, PAG1 и GUCY1A2, PAG1 и GUCY1A3, PAG1 и GUCY1B2, PAG1 и GUCY1B3, PAG1 и HAVCR2, PAG1 и HMOX2, PAG1 и IL10RA, PAG1 и IL10RB, PAG1 и IL6R, PAG1 и IL6ST, PAG1 и LAG3, PAG1 и LAIR1, PDCD1 и BATF, PDCD1 и BTLA, PDCD1 и CASP10, PDCD1 и CASP3, PDCD1 и CASP6, PDCD1 и CASP7, PDCD1 и CASP8, PDCD1 и CD160, PDCD1 и CD244, PDCD1 и CD96, PDCD1 и CRTAM, PDCD1 и CSK, PDCD1 и CTLA4, PDCD1 и EIF2AK4, PDCD1 и FADD, PDCD1 и FAS, PDCD1 и FOXP3, PDCD1 и GUCY1A2, PDCD1 и GUCY1A3, PDCD1 и GUCY1B2, PDCD1 и GUCY1B3, PDCD1 и HAVCR2, PDCD1 и HMOX2, PDCD1 и IL10RA, PDCD1 и IL10RB, PDCD1 и IL6R, PDCD1 и IL6ST, PDCD1 и LAG3, PDCD1 и LAIR1, PDCD1 и PAG1, PPP2CA и BATF, PPP2CA и BTLA, PPP2CA и CASP10, PPP2CA и CASP3, PPP2CA и CASP6, PPP2CA и CASP7, PPP2CA и CASP8, РРР2СА и CD160, РРР2СА и CD244, РРР2СА и CD96, РРР2СА и CRTAM, РРР2СА и CSK, РРР2СА и CTLA4, РРР2СА и EIF2AK4, РРР2СА и FADD, РРР2СА и FAS, РРР2СА и FOXP3, РРР2СА и GUCY1A2, РРР2СА и GUCY1A3, РРР2СА и GUCY1B2, РРР2СА и GUCY1B3, РРР2СА и HAVCR2, РРР2СА и НМОХ2, РРР2СА и IL10RA, РРР2СА и IL10RB, РРР2СА и IL6R, РРР2СА и IL6ST, РРР2СА и LAG3, РРР2СА и LAIR1, РРР2СА и PAG1, РРР2СА и PDCD1, РРР2СВ и BATF, РРР2СВ и BTLA, РРР2СВ и CASP10, РРР2СВ и CASP3, РРР2СВ и CASP6, РРР2СВ и CASP7, РРР2СВ и CASP8, РРР2СВ и CD160, РРР2СВ и CD244, РРР2СВ и CD96, РРР2СВ и CRTAM, РРР2СВ и CSK, РРР2СВ и CTLA4, РРР2СВ и EIF2AK4, РРР2СВ и FADD, РРР2СВ и FAS, РРР2СВ и FOXP3, РРР2СВ и GUCY1A2, РРР2СВ и GUCY1A3, РРР2СВ и GUCY1B2, РРР2СВ и GUCY1B3, РРР2СВ и HAVCR2, РРР2СВ и НМОХ2, РРР2СВ и IL10RA, РРР2СВ и IL10RB, РРР2СВ и IL6R, РРР2СВ и IL6ST, РРР2СВ и LAG3, РРР2СВ и LAIR1, РРР2СВ и PAG1, РРР2СВ и PDCD1, РРР2СВ и РРР2СА, PRDM1 и BATF, PRDM1 и BTLA, PRDM1 и CASP10, PRDM1 и CASP3, PRDM1 и CASP6, PRDM1 и CASP7, PRDM1 и CASP8, PRDM1 и CD160, PRDM1 и CD244, PRDM1 и CD96, PRDM1 и CRTAM, PRDM1 и CSK, PRDM1 и CTLA4, PRDM1 и EIF2AK4, PRDM1 и FADD, PRDM1 и FAS, PRDM1 и FOXP3, PRDM1 и GUCY1A2, PRDM1 и GUCY1A3, PRDM1 и GUCY1B2, PRDM1 и GUCY1B3, PRDM1 и HAVCR2, PRDM1 и HMOX2, PRDM1 и IL10RA, PRDM1 и IL10RB, PRDM1 и IL6R, PRDM1 и IL6ST, PRDM1 и LAG3, PRDM1 и LAIR1, PRDM1 и PAG1, PRDM1 и PDCD1, PRDM1 и PPP2CA, PRDM1 и PPP2CB, PTPN22 и BATF, PTPN22 и BTLA, PTPN22 и CASP10, PTPN22 и CASP3, PTPN22 и CASP6, PTPN22 и CASP7, PTPN22 и CASP8, PTPN22 и CD160, PTPN22 и CD244, PTPN22 и CD96, PTPN22 и CRTAM, PTPN22 и CSK, PTPN22 и CTLA4, PTPN22 и EIF2AK4, PTPN22 и FADD, PTPN22 и FAS, PTPN22 и FOXP3, PTPN22 и GUCY1A2, PTPN22 и GUCY1A3, PTPN22 и GUCY1B2, PTPN22 и GUCY1B3, PTPN22 и HAVCR2, PTPN22 и НМОХ2, PTPN22 и IL10RA, PTPN22 и IL10RB, PTPN22 и IL6R, PTPN22 и IL6ST, PTPN22 и LAG3, PTPN22 и LAIR1, PTPN22 и PAG1, PTPN22 и PDCD1, PTPN22 и РРР2СА, PTPN22 и РРР2СВ, PTPN22 и PRDM1, PTPN6 и BATF, PTPN6 и BTLA, PTPN6 и CASP10, PTPN6 и CASP3, PTPN6 и CASP6, PTPN6 и CASP7, PTPN6 и CASP8, PTPN6 и CD160, PTPN6 и CD244, PTPN6 и CD96, PTPN6 и CRTAM, PTPN6 и CSK, PTPN6 и CTLA4, PTPN6 и EIF2AK4, PTPN6 и FADD, PTPN6 и FAS, PTPN6 и FOXP3, PTPN6 и GUCY1A2, PTPN6 и GUCY1A3, PTPN6 и GUCY1B2, PTPN6 и GUCY1B3, PTPN6 и HAVCR2, PTPN6 и НМОХ2, PTPN6 и IL10RA, PTPN6 и IL10RB, PTPN6 и IL6R, PTPN6 и IL6ST, PTPN6 и LAG3, PTPN6 и LAIR1, PTPN6 и PAG1, PTPN6 и PDCD1, PTPN6 и РРР2СА, PTPN6 и РРР2СВ, PTPN6 и PRDM1, PTPN6 и PTPN22, SIGLEC7 и BATF, SIGLEC7 и BTLA, SIGLEC7 и CASP10, SIGLEC7 и CASP3, SIGLEC7 и CASP6, SIGLEC7 и CASP7, SIGLEC7 и CASP8, SIGLEC7 и CD160, SIGLEC7 и CD244, SIGLEC7 и CD96, SIGLEC7 и CRTAM, SIGLEC7 и CSK, SIGLEC7 и CTLA4, SIGLEC7 и EIF2AK4, SIGLEC7 и FADD, SIGLEC7 и FAS, SIGLEC7 и FOXP3, SIGLEC7 и GUCY1A2, SIGLEC7 и GUCY1A3, SIGLEC7 и GUCY1B2, SIGLEC7 и GUCY1B3, SIGLEC7 и HAVCR2, SIGLEC7 и НМОХ2, SIGLEC7 и IL10RA, SIGLEC7 и IL10RB, SIGLEC7 и IL6R, SIGLEC7 и IL6ST, SIGLEC7 и LAG3, SIGLEC7 и LAIR1, SIGLEC7 и PAG1, SIGLEC7 и PDCD1, SIGLEC7 и РРР2СА, SIGLEC7 и РРР2СВ, SIGLEC7 и PRDM1, SIGLEC7 и PTPN22, SIGLEC7 и PTPN6, SIGLEC9 и BATF, SIGLEC9 и BTLA, SIGLEC9 и CASP10, SIGLEC9 и CASP3, SIGLEC9 и CASP6, SIGLEC9 и CASP7, SIGLEC9 и CASP8, SIGLEC9 и CD160, SIGLEC9 и CD244, SIGLEC9 и CD96, SIGLEC9 и CRTAM, SIGLEC9 и CSK, SIGLEC9 и CTLA4, SIGLEC9 и EIF2AK4, SIGLEC9 и FADD, SIGLEC9 и FAS, SIGLEC9 и FOXP3, SIGLEC9 и GUCY1A2, SIGLEC9 и GUCY1A3, SIGLEC9 и GUCY1B2, SIGLEC9 и GUCY1B3, SIGLEC9 и HAVCR2, SIGLEC9 и НМОХ2, SIGLEC9 и IL10RA, SIGLEC9 и IL10RB, SIGLEC9 и IL6R, SIGLEC9 и IL6ST, SIGLEC9 и LAG3, SIGLEC9 и LAIR1, SIGLEC9 и PAG1, SIGLEC9 и PDCD1, SIGLEC9 и РРР2СА, SIGLEC9 и РРР2СВ, SIGLEC9 и PRDM1, SIGLEC9 и PTPN22, SIGLEC9 и PTPN6, SIGLEC9 и SIGLEC7, SIT1 и BATF, SIT1 и BTLA, SIT1 и CASP10, SIT1 и CASP3, SIT1 и CASP6, SIT1 и CASP7, SIT1 и CASP8, SIT1 и CD160, SIT1 и CD244, SIT1 и CD96, SIT1 и CRTAM, SIT1 и CSK, SIT1 и CTLA4, SIT1 и EIF2AK4, SIT1 и FADD, SIT1 и FAS, SIT1 и FOXP3, SIT1 и GUCY1A2, SIT1 и GUCY1A3, SIT1 и GUCY1B2, SIT1 и GUCY1B3, SIT1 и HAVCR2, SIT1 и НМОХ2, SIT1 и IL10RA, SIT1 и IL10RB, SIT1 и IL6R, SIT1 и IL6ST, SIT1 и LAG3, SIT1 и LAIR1, SIT1 и PAG1, SIT1 и PDCD1, SIT1 и РРР2СА, SIT1 и РРР2СВ, SIT1 и PRDM1, SIT1 и PTPN22, SIT1 и PTPN6, SIT1 и SIGLEC7, SIT1 и SIGLEC9, SKI и BATF, SKI и BTLA, SKI и CASP10, SKI и CASP3, SKI и CASP6, SKI и CASP7, SKI и CASP8, SKI и CD160, SKI и CD244, SKI и CD96, SKI и CRTAM, SKI и CSK, SKI и CTLA4, SKI и EIF2AK4, SKI и FADD, SKI и FAS, SKI и FOXP3, SKI и GUCY1A2, SKI и GUCY1A3, SKI и GUCY1B2, SKI и GUCY1B3, SKI и HAVCR2, SKI и HMOX2, SKI и IL10RA, SKI и IL10RB, SKI и IL6R, SKI и IL6ST, SKI и LAG3, SKI и LAIR1, SKI и PAG1, SKI и PDCD1, SKI и PPP2CA, SKI и PPP2CB, SKI и PRDM1, SKI и PTPN22, SKI и PTPN6, SKI и SIGLEC7, SKI и SIGLEC9, SKI и SIT1, SKIL и BATF, SKIL и BTLA, SKIL и CASP10, SKIL и CASP3, SKIL и CASP6, SKIL и CASP7, SKIL и CASP8, SKIL и CD160, SKIL и CD244, SKIL и CD96, SKIL и CRTAM, SKIL и CSK, SKIL и CTLA4, SKIL и EIF2AK4, SKIL и FADD, SKIL и FAS, SKIL и FOXP3, SKIL и GUCY1A2, SKIL и GUCY1A3, SKIL и GUCY1B2, SKIL и GUCY1B3, SKIL и HAVCR2, SKIL и HMOX2, SKIL и IL10RA, SKIL и IL10RB, SKIL и IL6R, SKIL и IL6ST, SKIL и LAG3, SKIL и LAIR1, SKIL и PAG1, SKIL и PDCD1, SKIL и PPP2CA, SKIL и PPP2CB, SKIL и PRDM1, SKIL и PTPN22, SKIL и PTPN6, SKIL и SIGLEC7, SKIL и SIGLEC9, SKIL и SIT1, SKIL и SKI, SMAD10 и BATF, SMAD10 и BTLA, SMAD10 и CASP10, SMAD10 и CASP3, SMAD10 и CASP6, SMAD10 и CASP7, SMAD10 и CASP8, SMAD10 и CD160, SMAD10 и CD244, SMAD10 и CD96, SMAD10 и CRTAM, SMAD10 и CSK, SMAD10 и CTLA4, SMAD10 и EIF2AK4, SMAD10 и FADD, SMAD10 и FAS, SMAD10 и FOXP3, SMAD10 и GUCY1A2, SMAD10 и GUCY1A3, SMAD10 и GUCY1B2, SMAD10 и GUCY1B3, SMAD10 и HAVCR2, SMAD10 и HMOX2, SMAD10 и IL10RA, SMAD10 и IL10RB, SMAD10 и IL6R, SMAD10 и IL6ST, SMAD10 и LAG3, SMAD10 и LAIR1, SMAD10 и PAG1, SMAD10 и PDCD1, SMAD10 и PPP2CA, SMAD10 и PPP2CB, SMAD10 и PRDM1, SMAD10 и PTPN22, SMAD10 и PTPN6, SMAD10 и SIGLEC7, SMAD10 и SIGLEC9, SMAD10 и SIT1, SMAD10 и SKI, SMAD10 и SKIL, SMAD2 и BATF, SMAD2 и BTLA, SMAD2 и CASP10, SMAD2 и CASP3, SMAD2 и CASP6, SMAD2 и CASP7, SMAD2 и CASP8, SMAD2 и CD160, SMAD2 и CD244, SMAD2 и CD96, SMAD2 и CRTAM, SMAD2 и CSK, SMAD2 и CTLA4, SMAD2 и EIF2AK4, SMAD2 и FADD, SMAD2 и FAS, SMAD2 и FOXP3, SMAD2 и GUCY1A2, SMAD2 и GUCY1A3, SMAD2 и GUCY1B2, SMAD2 и GUCY1B3, SMAD2 и HAVCR2, SMAD2 и HMOX2, SMAD2 и IL10RA, SMAD2 и IL10RB, SMAD2 и IL6R, SMAD2 и IL6ST, SMAD2 и LAG3, SMAD2 и LAIR1, SMAD2 и PAG1, SMAD2 и PDCD1, SMAD2 и РРР2СА, SMAD2 и РРР2СВ, SMAD2 и PRDM1, SMAD2 и PTPN22, SMAD2 и PTPN6, SMAD2 и SIGLEC7, SMAD2 и SIGLEC9, SMAD2 и SIT1, SMAD2 и SKI, SMAD2 и SKIL, SMAD2 и SMAD10, SMAD3 и BATF, SMAD3 и BTLA, SMAD3 и CASP10, SMAD3 и CASP3, SMAD3 и CASP6, SMAD3 и CASP7, SMAD3 и CASP8, SMAD3 и CD160, SMAD3 и CD244, SMAD3 и CD96, SMAD3 и CRTAM, SMAD3 и CSK, SMAD3 и CTLA4, SMAD3 и EIF2AK4, SMAD3 и FADD, SMAD3 и FAS, SMAD3 и FOXP3, SMAD3 и GUCY1A2, SMAD3 и GUCY1A3, SMAD3 и GUCY1B2, SMAD3 и GUCY1B3, SMAD3 и HAVCR2, SMAD3 и HMOX2, SMAD3 и IL10RA, SMAD3 и IL10RB, SMAD3 и IL6R, SMAD3 и IL6ST, SMAD3 и LAG3, SMAD3 и LAIR1, SMAD3 и PAG1, SMAD3 и PDCD1, SMAD3 и РРР2СА, SMAD3 и РРР2СВ, SMAD3 и PRDM1, SMAD3 и PTPN22, SMAD3 и PTPN6, SMAD3 и SIGLEC7, SMAD3 и SIGLEC9, SMAD3 и SIT1, SMAD3 и SKI, SMAD3 и SKIL, SMAD3 и SMAD10, SMAD3 и SMAD2, SMAD4 и BATF, SMAD4 и BTLA, SMAD4 и CASP10, SMAD4 и CASP3, SMAD4 и CASP6, SMAD4 и CASP7, SMAD4 и CASP8, SMAD4 и CD160, SMAD4 и CD244, SMAD4 и CD96, SMAD4 и CRTAM, SMAD4 и CSK, SMAD4 и CTLA4, SMAD4 и EIF2AK4, SMAD4 и FADD, SMAD4 и FAS, SMAD4 и FOXP3, SMAD4 и GUCY1A2, SMAD4 и GUCY1A3, SMAD4 и GUCY1B2, SMAD4 и GUCY1B3, SMAD4 и HAVCR2, SMAD4 и НМОХ2, SMAD4 и IL10RA, SMAD4 и IL10RB, SMAD4 и IL6R, SMAD4 и IL6ST, SMAD4 и LAG3, SMAD4 и LAIR1, SMAD4 и PAG1, SMAD4 и PDCD1, SMAD4 и РРР2СА, SMAD4 и РРР2СВ, SMAD4 и PRDM1, SMAD4 и PTPN22, SMAD4 и PTPN6, SMAD4 и SIGLEC7, SMAD4 и SIGLEC9, SMAD4 и SIT1, SMAD4 и SKI, SMAD4 и SKIL, SMAD4 и SMAD10, SMAD4 и SMAD2, SMAD4 и SMAD3, TGFBR1 и BATF, TGFBR1 и BTLA, TGFBR1 и CASP10, TGFBR1 и CASP3, TGFBR1 и CASP6, TGFBR1 и CASP7, TGFBR1 и CASP8, TGFBR1 и CD160, TGFBR1 и CD244, TGFBR1 и CD96, TGFBR1 и CRTAM, TGFBR1 и CSK, TGFBR1 и CTLA4, TGFBR1 и EIF2AK4, TGFBR1 и FADD, TGFBR1 и FAS, TGFBR1 и FOXP3, TGFBR1 и GUCY1A2, TGFBR1 и GUCY1A3, TGFBR1 и GUCY1B2, TGFBR1 и GUCY1B3, TGFBR1 и HAVCR2, TGFBR1 и HMOX2, TGFBR1 и IL10RA, TGFBR1 и IL10RB, TGFBR1 и IL6R, TGFBR1 и IL6ST, TGFBR1 и LAG3, TGFBR1 и LAIR1, TGFBR1 и PAG1, TGFBR1 и PDCD1, TGFBR1 и РРР2СА, TGFBR1 и РРР2СВ, TGFBR1 и PRDM1, TGFBR1 и PTPN22, TGFBR1 и PTPN6, TGFBR1 и SIGLEC7, TGFBR1 и SIGLEC9, TGFBR1 и SIT1, TGFBR1 и SKI, TGFBR1 и SKIL, TGFBR1 и SMAD10, TGFBR1 и SMAD2, TGFBR1 и SMAD3, TGFBR1 и SMAD4, TGFBR2 и BATF, TGFBR2 и BTLA, TGFBR2 и CASP10, TGFBR2 и CASP3, TGFBR2 и CASP6, TGFBR2 и CASP7, TGFBR2 и CASP8, TGFBR2 и CD160, TGFBR2 и CD244, TGFBR2 и CD96, TGFBR2 и CRTAM, TGFBR2 и CSK, TGFBR2 и CTLA4, TGFBR2 и EIF2AK4, TGFBR2 и FADD, TGFBR2 и FAS, TGFBR2 и FOXP3, TGFBR2 и GUCY1A2, TGFBR2 и GUCY1A3, TGFBR2 и GUCY1B2, TGFBR2 и GUCY1B3, TGFBR2 и HAVCR2, TGFBR2 и HMOX2, TGFBR2 и IL10RA, TGFBR2 и IL10RB, TGFBR2 и IL6R, TGFBR2 и IL6ST, TGFBR2 и LAG3, TGFBR2 и LAIR1, TGFBR2 и PAG1, TGFBR2 и PDCD1, TGFBR2 и РРР2СА, TGFBR2 и РРР2СВ, TGFBR2 и PRDM1, TGFBR2 и PTPN22, TGFBR2 и PTPN6, TGFBR2 и SIGLEC7, TGFBR2 и SIGLEC9, TGFBR2 и SIT1, TGFBR2 и SKI, TGFBR2 и SKIL, TGFBR2 и SMAD10, TGFBR2 и SMAD2, TGFBR2 и SMAD3, TGFBR2 и SMAD4, TGFBR2 и TGFBR1, TGIF1 и BATF, TGIF1 и BTLA, TGIF1 и CASP10, TGIF1 и CASP3, TGIF1 и CASP6, TGIF1 и CASP7, TGIF1 и CASP8, TGIF1 и CD160, TGIF1 и CD244, TGIF1 и CD96, TGIF1 и CRTAM, TGIF1 и CSK, TGIF1 и CTLA4, TGIF1 и EIF2AK4, TGIF1 и FADD, TGIF1 и FAS, TGIF1 и FOXP3, TGIF1 и GUCY1A2, TGIF1 и GUCY1A3, TGIF1 и GUCY1B2, TGIF1 и GUCY1B3, TGIF1 и HAVCR2, TGIF1 и HMOX2, TGIF1 и IL10RA, TGIF1 и IL10RB, TGIF1 и IL6R, TGIF1 и IL6ST, TGIF1 и LAG3, TGIF1 и LAIR1, TGIF1 и PAG1, TGIF1 и PDCD1, TGIF1 и РРР2СА, TGIF1 и PPP2CB, TGIF1 и PRDM1, TGIF1 и PTPN22, TGIF1 и PTPN6, TGIF1 и SIGLEC7, TGIF1 и SIGLEC9, TGIF1 и SIT1, TGIF1 и SKI, TGIF1 и SKIL, TGIF1 и SMAD10, TGIF1 и SMAD2, TGIF1 и SMAD3, TGIF1 и SMAD4, TGIF1 и TGFBR1, TGIF1 и TGFBR2, TIGIT и BATF, TIGIT и BTLA, TIGIT и CASP10, TIGIT и CASP3, TIGIT и CASP6, TIGIT и CASP7, TIGIT и CASP8, TIGIT и CD160, TIGIT и CD244, TIGIT и CD96, TIGIT и CRTAM, TIGIT и CSK, TIGIT и CTLA4, TIGIT и EIF2AK4, TIGIT и FADD, TIGIT и FAS, TIGIT и FOXP3, TIGIT и GUCY1A2, TIGIT и GUCY1A3, TIGIT и GUCY1B2, TIGIT и GUCY1B3, TIGIT и HAVCR2, TIGIT и HMOX2, TIGIT и IL10RA, TIGIT и IL10RB, TIGIT и IL6R, TIGIT и IL6ST, TIGIT и LAG3, TIGIT и LAIR1, TIGIT и PAG1, TIGIT и PDCD1, TIGIT и PPP2CA, TIGIT и PPP2CB, TIGIT и PRDM1, TIGIT и PTPN22, TIGIT и PTPN6, TIGIT и SIGLEC7, TIGIT и SIGLEC9, TIGIT и SIT1, TIGIT и SKI, TIGIT и SKIL, TIGIT и SMAD10, TIGIT и SMAD2, TIGIT и SMAD3, TIGIT и SMAD4, TIGIT и TGFBR1, TIGIT и TGFBR2, TIGIT и TGIF1, TNFRSF10A и BATF, TNFRSF10A и BTLA, TNFRSF10A и CASP10, TNFRSF10A и CASP3, TNFRSF10A и CASP6, TNFRSF10A и CASP7, TNFRSF10A и CASP8, TNFRSF10A и CD160, TNFRSF10A и CD244, TNFRSF10A и CD96, TNFRSF10A и CRTAM, TNFRSF10A и CSK, TNFRSF10A и CTLA4, TNFRSF10A и EIF2AK4, TNFRSF10A и FADD, TNFRSF10A и FAS, TNFRSF10A и FOXP3, TNFRSF10A и GUCY1A2, TNFRSF10A и GUCY1A3, TNFRSF10A и GUCY1B2, TNFRSF10A и GUCY1B3, TNFRSF10A и HAVCR2, TNFRSF10A и HMOX2, TNFRSF10A и IL10RA, TNFRSF10A и IL10RB, TNFRSF10A и IL6R, TNFRSF10A и IL6ST, TNFRSF10A и LAG3, TNFRSF10A и LAIR1, TNFRSF10A и PAG1, TNFRSF10A и PDCD1, TNFRSF10A и PPP2CA, TNFRSF10A и PPP2CB, TNFRSF10A и PRDM1, TNFRSF10A и PTPN22, TNFRSF10A и PTPN6, TNFRSF10A и SIGLEC7, TNFRSF10A и SIGLEC9, TNFRSF10A и SIT1, TNFRSF10A и SKI, TNFRSF10A и SKIL, TNFRSF10A и SMAD10, TNFRSF10A и SMAD2, TNFRSF10A и SMAD3, TNFRSF10A и SMAD4, TNFRSF10A и TGFBR1, TNFRSF10A и TGFBR2, TNFRSF10A и TGIF1, TNFRSF10A и TIGIT, TNFRSF10B и BATF, TNFRSF10B и BTLA, TNFRSF10B и CASP10, TNFRSF10B и CASP3, TNFRSF10B и CASP6, TNFRSF10B и CASP7, TNFRSF10B и CASP8, TNFRSF10B и CD160, TNFRSF10B и CD244, TNFRSF10B и CD96, TNFRSF10B и CRTAM, TNFRSF10B и CSK, TNFRSF10B и CTLA4, TNFRSF10B и EIF2AK4, TNFRSF10B и FADD, TNFRSF10B и FAS, TNFRSF10B и FOXP3, TNFRSF10B и GUCY1A2, TNFRSF10B и GUCY1A3, TNFRSF10B и GUCY1B2, TNFRSF10B и GUCY1B3, TNFRSF10B и HAVCR2, TNFRSF10B и HMOX2, TNFRSF10B и IL10RA, TNFRSF10B и IL10RB, TNFRSF10B и IL6R, TNFRSF10B и IL6ST, TNFRSF10B и LAG3, TNFRSF10B и LAIR1, TNFRSF10B и PAG1, TNFRSF10B и PDCD1, TNFRSF10B и PPP2CA, TNFRSF10B и PPP2CB, TNFRSF10B и PRDM1, TNFRSF10B и PTPN22, TNFRSF10B и PTPN6, TNFRSF10B и SIGLEC7, TNFRSF10B и SIGLEC9, TNFRSF10B и SIT1, TNFRSF10B и SKI, TNFRSF10B и SKIL, TNFRSF10B и SMAD10, TNFRSF10B и SMAD2, TNFRSF10B и SMAD3, TNFRSF10B и SMAD4, TNFRSF10B и TGFBR1, TNFRSF10B и TGFBR2, TNFRSF10B и TGIF1, TNFRSF10B и TIGIT, TNFRSF10B и TNFRSF10A.
При инактивировании гена подразумевают, что целевой ген не экспрессируется в форму функционального белка. Ген может быть изменен или делегирован. В одном из вариантов осуществления настоящего изобретения указанный белок может быть мутированным и сохраняющим активность как доминантная отрицательная форма. В другом предпочтительном варианте осуществления настоящего изобретения генетическая модификация способа зависит от экспрессии в клетках, предназначенных для конструирования, одной редкощепящей эндонуклеазы таким образом, что указанная редкощепящая эндонуклеазы специфически катализирует расщепление в одном целевом гене, тем самым инактивируя указанный целевой ген. Разрывы в цепи нуклеиновой кислоты, вызванные редкощепящей эндонуклеазой, обычно подвергаются репарации через различные механизмы гомологичной рекомбинации или негомологичного соединения концов (non-homologous end joining - NHEJ). Однако NHEJ не является совершенным процессом репарации, что часто приводит к изменениям в последовательности ДНК по сайту расщепления. Механизмы включают воссоединение остатков двух концов ДНК путем прямого повторного лигирования (Critchlow и Jackson, 1998) или за счет так называемого основанного на микрогомологии соединения концов (Ма и др., 2003). Репарация путем соединения негомологичных концов (NHEJ) часто приводит к малым инсерциям или делециям и может применяться для создания специфических генных нокаутов. Указанная модификация может представлять замещение, делецию или вставку по меньшей мере одного нуклеотида. Клетки, в которых происходит мутагенез путем расщепления, т.е. мутагенез - следствие NHEJ, могут быть идентифицированы и/или отобраны методом, хорошо известным специалистам в этой области.
В одном из вариантов осуществления настоящего изобретения указанный способ конструирования клеток включает по меньшей мере одну из следующих стадий:
(а) получения Т-клеток, предпочтительно из культуры клеток или из образца крови;
(б) интродукции в указанные Т-клетки редкощепящей эндонуклеазы, способной селективно инактивировать путем расщепления ДНК, предпочтительно путем двухцепочечного разрыва, по меньшей мере двух генов, кодирующих белки иммунных контрольных точек, и
(в) размножения указанных клеток.
Указанные гены белков иммунных контрольных точек предпочтительно выбраны из перечня, согласно описанному выше.
В другом варианте осуществления настоящего изобретения указанная редкощепящая эндонуклеаза может быть мегануклеазой, цинкпальцевой нуклеазой, CRISPR/Cas9 или TALE-нуклеазой. В предпочтительном варианте осуществления настоящего изобретения указанная редкощепящая эндонуклеаза является TALE-нуклеазой. Под TALE-нуклеазой подразумевается гибридный белок, состоящий из ДНК-связывающего домена, происходящего от эффектора, подобного активатору транскрипции (Transcription Activator Like Effector - TALE), и одного каталитического домена нуклеазы для расщепления целевой последовательности нуклеиновой кислоты (Boch и др., 2009; Moscou и Bogdanove, 2009; Christian и др., 2010; Cermak и др., 2011; Geissler и др., 2011; Huang и др., 2011; Li и др., 2011; Mahfouz и др., 2011; Miller и др., 2011; Morbitzer и др., 2011; Mussolino и др., 2011; Sander и др., 2011; Tesson и др., 2011; Weber и др., 2011; Zhang и др., 2011; Deng и др., 2012; Li и др., 2012; Mahfouz и др., 2012; Маk и др., 2012).
В настоящем изобретении сконструированы новые TALE-нуклеазы для точного нацеливания на релевантные гены для адаптивных стратегий иммунотерапии. Предпочтительными TALE-нуклеазами по настоящему изобретению являются те, которые распознают и расщепляют целевую последовательность, выбранную из группы, включающей: SEQ ID NO: 77 и SEQ ID NO: 78 (PD1), с SEQ ID NO: 74 по SEQ ID NO: 76 (CTLA-4). Настоящее изобретение также относится к полипептидам TALE-нуклеаз, которые включают аминокислотную последовательность, выбранную из группы, включающей последовательности с SEQ ID NO: 79 по SEQ ID NO: 88.
В другом варианте осуществления настоящего изобретения добавочный каталитический домен может быть дополнительно интродуцирован в клетку с указанной редкощепящей эндонуклеазой для повышения мутагенеза с целью усиления их способности инактивировать целевые гены. В частности указанный дополнительный каталитический домен является ферментом, процессирующим конец ДНК. Примерами, которые не ограничивают настоящее изобретение, являются ферменты, процессирующие конец ДНК: 5-3' экзонуклеазы, 3-5' экзонуклеазы, 5-3' щелочные экзонуклеазы, 5' флэп-эндонуклеазы, геликазы, фосфатазы, гидролазы и независящие от матрицы ДНК-полимеразы. Примерами таких каталитических доменов, которые не ограничивают настоящее изобретение, являются белковый домен или каталитически активное производное белкового домена, выбранные из группы, включающей: hExoI (EXO1_HUMAN), Yeast ExoI (EXO1_YEAST), E.coli ExoI, Human TREX2, Mouse TREX1, Human TREX1, Bovine TREX1, Rat TREX1, TdT (terminal deoxynucleotidyl transferase - терминальная дезоксинуклеотидилтрансфераза), Human DNA2, Yeast DNA2 (DNA2_YEAST). В предпочтительном варианте осуществления настоящего изобретения указанный дополнительный каталитический домен обладает 3'-5'-экзонуклеазной активностью, и в более предпочтительном варианте осуществления настоящего изобретения указанным дополнительным каталитическим доменом является TREX, более предпочтительно каталитический домен TREX2 (WO 2012/058458). В другом предпочтительном варианте осуществления настоящего изобретения указанный каталитический домен кодируется одноцепочечным полипептидом TREX (WO 2013/009525). Указанный дополнительный каталитический домен может быть гибридизирован с нуклеазным гибридным белком или химерным белком по настоящему изобретению необязательно пептидным линкером.
Известно, что эндонуклеотические разрывы стимулируют степень гомологической рекомбинации. Например, в другом варианте осуществления настоящего изобретения стадия генетической модификации способа дополнительно включает стадию интродукции в клетки экзогенной нуклеиновой кислоты, включающей по меньшей мере последовательность, гомологичную части целевой последовательности нуклеиновой кислоты, таким образом, что гомологичная рекомбинация происходит между целевой последовательностью нуклеиновой кислоты и экзогенной нуклеиновой кислотой. В некоторых вариантах осуществления настоящего изобретения указанная экзогенная нуклеиновая кислота включает первую и вторую части, гомологичные области 5' и 3' целевой последовательности нуклеиновой кислоты, соответственно. В вариантах осуществления настоящего изобретения указанная экзогенная нуклеиновая кислота также включает третью часть, расположенную между первой и второй частями, которая не имеет гомологии с областями 5' и 3' целевой последовательности нуклеиновой кислоты. После расщепления целевой последовательности нуклеиновой кислоты гомологическая рекомбинация стимулируется между целевой последовательностью нуклеиновой кислоты и экзогенной нуклеиновой кислотой. Предпочтительно гомологичные последовательности, состоящие по меньшей мере из 50 п.о., предпочтительно более чем из 100 п.о. и более предпочтительно более чем из 200 п.о., применяют в указанном матриксе донора. Следовательно, экзогенная нуклеиновая кислота содержит предпочтительно от 200 п.о. до 6000 п.о., более предпочтительно от 1000 п.о. до 2000 п.о. В самом деле, общие гомологи нуклеиновой кислоты локализованы в областях, фланкирующих вниз по цепи и вверх по цепи сайт разрыва, и интродуцируемая последовательность нуклеиновой кислоты должна быть локализована между этими двумя плечами.
В частности, указанная экзогенная нуклеиновая кислота последовательно включает первую область, гомологичную последовательностям, расположенным выше по цепи от указанного расщепления, последовательность для инактивации одного целевого гена, выбранного из группы, состоящей из генов иммунных контрольных точек, и вторую область, гомологичную последовательностям, расположенным ниже по цепи от указанного расщепления. Стадия указанной интродукции полинуклеотида может быть одновременно, до или после интродукции или экспрессии указанной редкощепящей эндонуклеазы. В зависимости от расположения целевой последовательности нуклеиновой кислоты, в которой происходит расщепление, такая экзогенная нуклеиновая кислота может применяться для нокаута гена, например, если экзогенная нуклеиновая кислота расположена в открытой рамке считывания указанного гена, или для интродукции новых последовательностей или целевых генов. Инсерции последовательностей с помощью такой экзогенной нуклеиновой кислоты могут применяться для модификации имеющегося целевого гена путем коррекции или замещения указанного гена (примером, который не ограничивает настоящее изобретение, является замещение аллеля), или путем повышения или понижения регуляции экспрессии целевого гена (примером, который не ограничивает настоящее изобретение, является замещение промотора). Выше отмечалось, что стадия дополнительной модификации генома может быть стадией инактивации, включающей:
(а) интродукцию в указанные клетки по меньшей мере одной редкощепящей эндонуклеазы таким образом, что указанная редкощепящая эндонуклеаза специфически катализирует расщепление в одной целевой последовательности генома указанной клетки,
(б) необязательно интродукцию в указанные клетки экзогенной нуклеиновой кислоты, последовательно включающей первую область, гомологичную последовательностям выше по цепи от указанного расщепления, последовательность, которую встраивают в геном указанной клетки, и вторую область, гомологичную последовательностям, расположенным ниже по цепи от указанного сайта расщепления,
причем указанная интродуцированная экзогенная нуклеиновая кислота инактивирует ген и интегрирует по меньшей мере одну экзогенную полинуклеотидную последовательность, кодирующую по меньшей мере один целевой рекомбинантный белок. В другом варианте осуществления настоящего изобретения указанную экзогенную полинуклеотидную последовательность интегрируют в ген, выбранный из группы, состоящей из генов иммунных контрольных точек.
В одном из вариантов осуществления настоящего изобретения указанный способ конструирования клеток дополнительно включает стадию модификации генома. С помощью дополнительной стадии модификации генома может быть осуществлена интродукция в клетки для конструирования одного целевого белка. Указанным целевым белком могут быть, например, химерный антигенный рецептор (Chimeric Antigen Receptor - CAR), многоцепочечный CAR, биспецифическое антитело, согласно описанию настоящего изобретения, но этими примерами перечень не ограничивается. Указанный способ конструирования клеток также может дополнительно включать интродукцию редкощепящей эндонуклеазы, способной селективно инактивировать путем расщепления ДНК ген, кодирующий мишень для указанного иммуносупрессирующего агента, гены TCR, согласно описанию настоящего изобретения.
Настоящее изобретение также относится к TALE-нуклеазам. В основном настоящее изобретение относится к TALE-нуклеазе, включающей:
(а) Эффектор, подобный активатору транскрипции (Transcription Activator Like Effector - TALE) ДНК-связывающий домен, который был сконструирован для связывания целевой последовательность в генах, выбранных из группы, состоящей из генов иммунных контрольных точек;
(б) Домен или полудомен расщепления.
Предпочтительными TALE-нуклеазами по настоящему изобретению являются нуклеазы, распознающие и расщепляющие целевую последовательность, выбранную из группы, включающей: SEQ ID NO: 77 и SEQ ID NO: 78 (PD1), а также с SEQ ID NO: 74 по SEQ ID NO: 76 (CTLA-4).
Указанные TALE-нуклеазы предпочтительно включают полипептидную последовательность, выбранную из группы, включающей последовательности с SEQ ID NO: 79 по SEQ ID NO: 88 для расщепления соответствующей целевой последовательности с SEQ ID NO: 74 по SEQ ID NO: 78.
Поскольку некоторая вариабельность может возникнуть от генома, от которого происходят эти полипептиды, а также учитывая возможность замещения некоторых из аминокислот в составе этих полипептидов без значительной потери активности (функциональные варианты), настоящее изобретение включает варианты указанных выше полипептидов, которые по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% и еще более предпочтительно по меньшей мере на 95% идентичны последовательностям, предусмотренным в настоящей патентной заявке.
Таким образом, настоящее изобретение представляет полипептиды, включающие полипептидную последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90%, 95% 97% или 99% идентична аминокислотной последовательности, выбранной из группы, включающей последовательности с SEQ ID NO: 79 по SEQ ID NO: 88.
К области охвата настоящего изобретения также относятся полинуклеотиды, векторы, кодирующие описанные выше редкощепящие эндонуклеазы по настоящему изобретению.
К области охвата настоящего изобретения также относятся выделенные клетки или линии клеток, которые применимы для получения указанным способом конструированных клеток, в частности Т-клеток, в которых был инактивирован по меньшей мере один ген, выбранный из группы, состоящей из генов иммунных контрольных точек, предпочтительно генов, выбранных из группы: CTLA4, РРР2СА, РРР2СВ, PTPN6, PTPN22, PDCD1, LAG3, HAVCR2, BTLA, CD160, TIGIT, CD96, CRTAM, LAIR1, SIGLEC7, SIGLEC9, CD244, TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT1, FOXP3, PRDM1, BATF, GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3, более предпочтительно инактивированы два из указанных выше генов. Согласно настоящему изобретению гены предпочтительно инактивированы по меньшей мере одной редкощепящей эндонуклеазой. В настоящем изобретении установлено, что применение TALE-нуклеаз особенно полезно для достижения двойной инактивации в Т-клетках. Настоящее изобретение охватывает выделенные Т-клетки, включающие по меньшей мере два полинуклеотида, кодирующие по меньшей мере первую и вторую TALE-нуклеазы, предпочтительно TALE-нуклеазы нацеливают против по меньшей мере двух генов, кодирующих белки иммунных контрольных точек, например, CTLA4, РРР2СА, РРР2СВ, PTPN6, PTPN22, PDCD1, LAG3, HAVCR2, BTLA, CD160, TIGIT, CD96, CRTAM, LAIR1, SIGLEC7, SIGLEC9, CD244, TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT1, FOXP3, PRDM1, BATF, GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3. В другом варианте осуществления настоящего изобретения указанные выделенные клетки также включают одну дополнительную геномную модификацию. В другом варианте осуществления настоящего изобретения указанная дополнительная геномная модификация является интеграцией по меньшей мере одной экзогенной полинуклеотидной последовательности.
Не аллореактивные и высоко активные Т-клетки для иммунотерапии
Действующий протокол лечения пациентов, в котором применяют адаптивную иммунотерапию, основан на переносе аутологичных клеток. Аутологическая терапия сталкивается со значительными техническими и логистическими препятствиями для практического применения, ее разработка нуждается в дорогостоящих специализированных средствах, требуется участие высококвалифицированных специалистов, терапия должна быть разработана в короткие сроки после постановки пациенту диагноза, и во многих случаях предварительное лечение пациента приводит к ухудшению иммунной функции, при этом лимфоциты пациента могут плохо функционировать и присутствовать в малых количествах. В идеале хотелось бы использовать стандартизированную терапию, в которой аллогенные терапевтические клетки могут быть предварительно изготовлены, подробно охарактеризованы, и доступны для немедленного введения пациентам. Однако специфичность эндогенных TCR аллогенных клеток, распознающих ткань хозяина как чужеродную, приводит к состоянию «трансплантат против хозяина» (graft versus host - GvHD), которое может привести к тяжелому повреждению тканей и смерти. Таким образом, для эффективного применения аллогенных клеток инактивирование TCRalpha или TCRbeta может привести к элиминации TCR с поверхности Т-клеток, предупреждая распознавание аллоантигена и соответственно GvHD.
Таким образом, конкретная задача, решаемая в настоящем изобретении, может относиться к способу конструирования аллогенных Т-клеток, особенно для иммунотерапии.
В одном из вариантов осуществления настоящего изобретения способ включает:
(а) получение Т-клетки,
(б) модифицирование Т-клеток путем инактивирования по меньшей мере:
- первого гена, кодирующего белок иммунной контрольной точки, и
- второго гена, кодирующего компонент рецептора Т-клеток (T-cell receptor - TCR)
(в) размножение указанных клеток.
В предпочтительном варианте осуществления настоящего изобретения стадия генетической модификации настоящего способа основывается на инактивации одного гена, предпочтительно двух генов, выбранных из группы, включающей CTLA4, РРР2СА, РРР2СВ, PTPN6, PTPN22, PDCD1, LAG3, HAVCR2, BTLA, CD160, TIGIT, CD96, CRTAM, LAIR1, SIGLEC7, SIGLEC9, CD244, TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT, FOXP3, PRDM1, BATF, GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3, TCR alpha и TCR beta. В другом варианте осуществления настоящего изобретения стадия генетической модификации основывается на инактивации двух генов, выбранных из группы, включающей CTLA4 и TCR alpha, CTLA4 и TCR beta, РРР2СА и TCR alpha, РРР2СВ и TCR beta, PTPN6 и TCR alpha, PTPN6 и TCR beta, PTPN22 и TCR alpha, PTPN22 и TCR beta, PDCD1 и TCR alpha, PDCD1 и TCR beta, LAG3 и TCR alpha, LAG3 и TCR beta, HAVCR2 и TCR alpha, HAVCR2 и TCR beta, BTLA и TCR alpha, BTLA и TCR beta, CD160 и TCR alpha, CD160 и TCR beta, TIGIT и TCR alpha, TIGIT и TCR beta, CD96 и TCR alpha, CD96 и TCR beta, CRTAM и TCR alpha, CRTAM и TCR beta, LAIR1 и TCR alpha, LAIR1 и TCR beta, SIGLEC7 и TCR alpha, SIGLEC7 и TCR beta, SIGLEC9 и TCR alpha, SIGLEC9 и TCR beta, CD244 и TCR alpha, CD244 и TCR beta, TNFRSF10B и TCR alpha, TNFRSF10B и TCR beta, TNFRSF10A и TCR alpha, TNFRSF10A и TCR beta, CASP8 и TCR alpha, CASP8 и TCR beta, CASP10 и TCR alpha, CASP10 и TCR beta, CASP3 и TCR alpha, CASP3 и TCR beta, CASP6 и TCR alpha, CASP6 и TCR beta, CASP7 и TCR alpha, CASP7 и TCR beta, FADD и TCR alpha, FADD и TCR beta, FAS и TCR alpha, FAS и TCR beta, TGFBRII и TCR alpha, TGFBRII и TCR beta, TGFRBRI и TCR alpha, TGFRBRI и TCR beta, SMAD2 и TCR alpha, SMAD2 и TCR beta, SMAD3 и TCR alpha, SMAD3 и TCR beta, SMAD4 и TCR alpha, SMAD4 и TCR beta, SMAD10 и TCR alpha, SMAD10 и TCR beta, SKI и TCR alpha, SKIL и TCR beta, TGIF1 и TCR alpha, TGIF1 и TCR beta, IL10RA и TCR alpha, IL10RA и TCR beta, IL10RB и TCR alpha, IL10RB и TCR beta, HMOX2 и TCR alpha, HMOX2 и TCR beta, IL6R и TCR alpha, IL6R и TCR beta, IL6ST и TCR alpha, IL6ST и TCR beta, EIF2AK4 и TCR alpha, EIF2AK4 и TCR beta, CSK и TCR alpha, CSK и TCR beta, PAG1 и TCR alpha, PAG1 и TCR beta, SIT1 и TCR alpha, SIT1 и TCR beta, FOXP3 и TCR alpha, FOXP3 и TCR beta, PRDM1 и TCR alpha, PRDM1 и TCR beta, BATF и TCR alpha, BATF и TCR beta, GUCY1A2 и TCR alpha, GUCY1A2 и TCR beta, GUCY1A3 и TCR alpha, GUCY1A3 и TCR beta, GUCY1B2 и TCR alpha, GUCY1B2 и TCR beta, GUCY1B3 и TCR alpha, GUCY1B3 и TCR beta. В другом варианте осуществления настоящего изобретения стадия генетической модификации настоящего способа основывается на инактивации более двух генов. Генетическая модификация предпочтительно производится ex-vivo.
В одном из вариантов осуществления настоящего изобретения указанный способ конструирования клеток включает по меньшей мере одну из следующих стадий:
(а) получения Т-клетки, предпочтительно из культуры клеток или из образца крови;
(б) интродукции в указанную Т-клетку редкощепящей эндонуклеазы, способной селективно инактивировать за счет расщепления ДНК, предпочтительно за счет двухцепочечного расщепления соответствующим образом:
- указанного гена, кодирующего белок иммунной контрольной точки, и
- по меньшей мере одного гена, кодирующего компонент рецептора Т-клеток (TCR);
(в) размножения указанных клеток.
В более предпочтительном варианте осуществления настоящего изобретения указанный способ включает:
(а) получение Т-клеток, предпочтительно из культуры клеток или из образца крови;
(б) трансформацию указанных Т-клеток нуклеиновой кислотой, кодирующей редкощепящую эндонуклеазу, способную селективно инактивировать путем расщепления ДНК, предпочтительно за счет следующего двухцепочечного расщепления:
- указанного гена, кодирующего белок иммунной контрольной точки, и
- по меньшей мере одного гена, кодирующего компонент рецептора Т-клеток (TCR);
(в) экспрессию указанных редкощепящих эндонуклеаз в указанных Т-клетках;
(г) сортировку трансформированных Т-клеток, которые не экспрессируют TCR на своей поверхности;
(д) размножение указанных клеток.
В другом предпочтительном варианте осуществления настоящего указанная редкощепящая эндонуклеаза специфически нацеливается по меньшей мере на один ген, предпочтительно на два гена, выбранных из группы, включающей: CTLA4, РРР2СА, РРР2СВ, PTPN6, PTPN22, PDCD1, LAG3, HAVCR2, BTLA, CD160, TIGIT, CD96, CRTAM, LAIR1, SIGLEC7, SIGLEC9, CD244, TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT, FOXP3, PRDM1, BATF, GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3, TCR alpha и TCR beta. В другом варианте осуществления настоящего изобретения более двух редкощепящих эндонуклеаз может экспрессироваться в клетках для конструирования с целью нацеливания и/или инактивирования более двух генов.
В другом варианте осуществления настоящего изобретения указанная редкощепящая эндонуклеаза может быть мегануклеазой, цинкпальцевой нуклеазой или TALE-нуклеазой. В предпочтительном варианте осуществления настоящего изобретения указанной редкощепящей эндонуклеазой является TALE-нуклеаза.
В настоящем изобретении новые TALE-нуклеазы конструируют для точного нацеливания на релевантные для адаптивных стратегий иммунотерапии гены. Предпочтительными TALE-нуклеазами по настоящему изобретению являются те, которые распознают и расщепляют целевую последовательность, выбранную из группы, включающей: SEQ ID NO: 77 и SEQ ID NO: 78 (PD1), с SEQ ID NO: 74 по SEQ ID NO: 76 (CTLA-4), SEQ ID NO: 37, с SEQ ID NO: 57 по SEQ ID NO: 60 (TCRalpha), SEQ ID NO: 38 или SEQ ID NO: 39 (TCRbeta). Настоящее изобретение также относится к полипептидам TALE-нуклеаз, которые включают аминокислотную последовательность, выбранную из группы, включающей последовательности с SEQ ID NO: 79 по SEQ ID NO: 88 и с SEQ ID NO: 41 по SEQ ID NO: 46.
Настоящее изобретение также относится к полипептидам, содержащим аминокислотную последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90%, 95% 97% или 99% идентична аминокислотной последовательности из группы, включающей последовательности с SEQ ID NO: 79 по SEQ ID NO: 88. Настоящее изобретение также относится к полинуклеотидам, векторам, кодирующим описанные выше редкощепящие эндонуклеазы по настоящему изобретению. Указанный способ может быть объединен с каким-либо из различных способов, изложенных в настоящем описании.
В предпочтительном варианте осуществления настоящего изобретения инактивация генов из группы, включающей гены иммунных контрольных точек, TCR alpha и TCR beta, может быть выполнена с точной геномной локализацией путем нацеливания специфической TALE-нуклеазы, при котором указанная TALE-нуклеаза катализирует расщепление, причем экзогенная нуклеиновая кислота последовательно включает по меньшей мере область гомологии и последовательность для инактивации одного целевого гена, выбранного из группы, включающей гены иммунных контрольных точек, TCR alpha and TCR, которые интегрированы за счет гомологической рекомбинации. В другом варианте осуществления настоящего изобретения несколько генов могут быть, последовательно или одновременно, инактивированы с помощью нескольких TALE-нуклеаз, которые соответствующим образом специфически нацеливаются на один определенный ген и несколько специфических полинуклеотидов для специфической генной инактивации.
Стадия дополнительной модификации генома также может заключаться в инактивации другого гена, выбранного из группы, включающей гены иммунных контрольных точек, TCR alpha и TCR beta. Выше отмечалось, что стадия дополнительной модификации генома может быть стадией инактивирования, включающей:
(а) интродукцию в указанные клетки по меньшей мере одной редкощепящей эндонуклеазы таким образом, что указанная редкощепящая эндонуклеаза специфически катализирует расщепление в одной целевой последовательности генома указанной клетки.
(б) необязательно интродукцию в указанные клетки экзогенной нуклеиновой кислоты, последовательно включающей первую область, гомологичную последовательностям выше по цепи от указанного расщепления, последовательность, которую встраивают в геном указанной клетки, и вторую область, гомологичную последовательностям, расположенным ниже по цепи от указанного сайта расщепления,
причем указанная интродуцированная экзогенная нуклеиновая кислота инактивирует ген и интегрирует по меньшей мере одну экзогенную полинуклеотидную последовательность, кодирующую по меньшей мере один целевой рекомбинантный белок. В другом варианте осуществления настоящего изобретения указанная экзогенная полинуклеотидная последовательность объединяется с геном, выбранным из группы, состоящей из генов иммунных контрольных точек, TCR alpha и TCR beta.
В одном из вариантов осуществления настоящего изобретения указанный способ конструирования клеток дополнительно включает стадию модификации генома. С помощью дополнительной стадии модификации генома может быть осуществлена интродукция в клетки для конструирования одного целевого белка. Указанным целевым белком могут быть, например, pTalpha или его функциональный вариант, химерный антигенный рецептор (Chimeric Antigen Receptor - CAR), многоцепочечный CAR, биспецифическое антитело, согласно описанию настоящего изобретения, но этими примерами перечень не ограничивается. Указанный способ конструирования клеток также может дополнительно включать интродукцию редкощепящей эндонуклеазы, способной селективно инактивировать путем расщепления ДНК ген, кодирующий мишень для указанного иммуносупрессирующего агента согласно описанию настоящего изобретения.
К области охвата настоящего изобретения также относятся полинуклеотиды, векторы, кодирующие описанные выше редкощепящие эндонуклеазы по настоящему изобретению.
К области охвата настоящего изобретения также относятся выделенные клетки или линии клеток, которые применимы для получения указанным способом сконструированных клеток, в частности Т-клеток, в которых были инактивированы по меньшей мере два гена, выбранные из группы, включающей: CTLA4 и TCR alpha, CTLA4 и TCR beta, РРР2СА и TCR alpha, РРР2СВ и TCR beta, PTPN6 и TCR alpha, PTPN6 и TCR beta, PTPN22 и TCR alpha, PTPN22 и TCR beta, PDCD1 и TCR alpha, PDCD1 и TCR beta, LAG3 и TCR alpha, LAG3 и TCR beta, HAVCR2 и TCR alpha, HAVCR2 и TCR beta, BTLA и TCR alpha, BTLA и TCR beta, CD160 и TCR alpha, CD160 и TCR beta, TIGIT и TCR alpha, TIGIT и TCR beta, CD96 и TCR alpha, CD96 и TCR beta, CRTAM и TCR alpha, CRTAM и TCR beta, LAIR1 и TCR alpha, LAIR1 и TCR beta, SIGLEC7 и TCR alpha, SIGLEC7 и TCR beta, SIGLEC9 и TCR alpha, SIGLEC9 и TCR beta, CD244 и TCR alpha, CD244 и TCR beta,TNFRSF10B и TCR alpha, TNFRSF10B и TCR beta, TNFRSF10A и TCR alpha, TNFRSF10A и TCR beta, CASP8 и TCR alpha, CASP8 и TCR beta, CASP10 и TCR alpha, CASP10 и TCR beta, CASP3 и TCR alpha, CASP3 и TCR beta, CASP6 и TCR alpha, CASP6 и TCR beta, CASP7 и TCR alpha, CASP7 и TCR beta, FADD и TCR alpha, FADD и TCR beta, FAS и TCR alpha, FAS и TCR beta, TGFBRII и TCR alpha, TGFBRII и TCR beta, TGFRBRI и TCR alpha, TGFRBRI и TCR beta, SMAD2 и TCR alpha, SMAD2 и TCR beta, SMAD3 и TCR alpha, SMAD3 и TCR beta, SMAD4 и TCR alpha, SMAD4 и TCR beta, SMAD10 и TCR alpha, SMAD10 и TCR beta, SKI и TCR alpha, SKIL и TCR beta, TGIF1 и TCR alpha, TGIF1 и TCR beta, IL10RA и TCR alpha, IL10RA и TCR beta, IL10RB и TCR alpha, IL10RB и TCR beta, HMOX2 и TCR alpha, HMOX2 и TCR beta, IL6R и TCR alpha, IL6R и TCR beta, IL6ST и TCR alpha, IL6ST и TCR beta, EIF2AK4 и TCR alpha, EIF2AK4 и TCR beta, CSK и TCR alpha, CSK и TCR beta, PAG1 и TCR alpha, PAG1 и TCR beta, SIT1 и TCR alpha, SIT1 и TCR beta, FOXP3 и TCR alpha, FOXP3 и TCR beta, PRDM1 и TCR alpha, PRDM1 и TCR beta, BATF и TCR alpha, BATF и TCR beta, GUCY1A2 и TCR alpha, GUCY1A2 и TCR beta, GUCY1A3 и TCR alpha, GUCY1A3 и TCR beta, GUCY1B2 и TCR alpha, GUCY1B2 и TCR beta, GUCY1B3 и TCR alpha, GUCY1B3 и TCR beta.
По настоящему изобретению эти гены предпочтительно инактивируют по меньшей мере одной редкощепящей эндонуклеазой. В настоящем изобретении установлено, что применение TALE-нуклеаз особенно предпочтительно для достижения двойного инактивирования в Т-клетках. Настоящее изобретение охватывает выделенную Т-клетку, включающую по меньшей мере два полинуклеотида, кодирующих по меньшей мере первую и вторую TALE-нуклеазу, предпочтительно первую TALE-нуклеазу, направленную против гена, кодирующего TCR, и вторую, направленную против гена, кодирующего белок иммунных контрольных точек, например, CTLA4, РРР2СА, РРР2СВ, PTPN6, PTPN22, PDCD1, LAG3, HAVCR2, BTLA, CD160, TIGIT, CD96, CRTAM, LAIR1, SIGLEC7, SIGLEC9, CD244, TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT1, FOXP3, PRDM1, BATF, GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3. В другом варианте осуществления настоящего изобретения указанная выделенная клетка также включают одну дополнительную геномную модификацию. В другом варианте осуществления настоящего изобретения указанная дополнительная геномная модификация представляет интеграцию по меньшей мере одной экзогенной полинуклеотидной последовательности.
Устойчивые к иммуносупрессии Т-клетки
Аллогенные клетки быстро отторгаются иммунной системой хозяина. Было показано, что аллогенные лейкоциты, имеющиеся в необлученных компонентах крови, могут сохраняться не более 5-6 суток (Boni и др., 2008). Таким образом, для предотвращения отторжения аллогенных клеток иммунную систему хозяина следует до некоторой степени эффективно подавить. Однако в случае применения адаптивной иммунотерапии применение иммуносупрессорных лекарственных средств также может оказывать вредное воздействие на интродуцированные терапевтические Т-клетки. Следовательно, для эффективного применения адаптивного иммунотерапевтического подхода в таких условиях интродуцированные клетки могут нуждаться в устойчивости к иммуносупрессивному лечению. Таким образом, в конкретном варианте осуществления настоящего изобретения способ в соответствии с настоящим изобретением дополнительно включает стадию модификации Т-клеток, чтобы сделать их устойчивыми к иммунодепрессивному агенту, предпочтительно путем инактивации по меньшей мере одного гена, кодирующего мишень для иммунодепрессивного агента. Иммунодепрессивный агент является агентом, который подавляет иммунную функцию одним из нескольких механизмов действия. Другими словами, иммунодепрессивный агент выступает в качестве соединения, которое демонстрирует способность уменьшать степень иммунного ответа. Примерами иммунодепрессивных агентов, не ограничивающими рамок охвата настоящего изобретения, являются ингибитор кальциневрина, мишень рапамицина, блокатор интерлейкин-2 α-цепи, ингибитор инозинмонофосфатдегидрогеназы, ингибитор редуктазы дигидрофолиевой кислоты, кортикостероиды или иммуносупрессивные антиметаболиты. Классические цитотоксические иммунодепрессанты действуют путем ингибирования синтеза ДНК. Другие могут действовать через активацию Т-клеток или путем ингибирования активации клеток-хелперов. Способ в соответствии с настоящим изобретением позволяет придать устойчивость к иммуносупрессии Т-клеткам для иммунотерапии за счет инактивации мишени иммунодепрессивного агента в Т-клетках. Примерами мишеней для иммуносупрессирующих агентов, не ограничивающими рамок охвата настоящего изобретения, могут быть рецепторы для иммуносупрессирующих агентов, например, CD52, глюкокортикоидные рецепторы (GR), представитель семейства генов FKBP и представитель семейства генов циклофилина.
В предпочтительном варианте осуществления настоящего изобретения генетическая модификация способа основана на экспрессии в предоставленных для конструирования клетках одной редкощепящей эндонуклеазы таким образом, что редкощепящая эндонуклеаза специфически катализирует расщепление в одном целевом гене, тем самым инактивируя указанный целевой ген. Указанная редкощепящая эндонуклеаза может быть мегануклеазой, цинковопальцевой нуклеазой или TALE-нуклеазой. Предпочтительными TALE-нуклеазами по настоящему изобретению являются те, которые распознают и расщепляют целевую последовательность, выбранную из группы, включающей последовательности: с SEQ ID NO: 1 по SEQ ID NO: 6 (GR), SEQ ID NO: 40, с SEQ ID NO: 61 по SEQ ID NO: 65 (CD52). Указанные TALE-нуклеазы предпочтительно включают полипептидную последовательность, выбранную из последовательностей с SEQ ID NO: 7 по SEQ ID NO: 18, SEQ ID NO: 47 и SEQ ID NO: 48.
Поскольку может возникнуть некоторая вариабельность, исходя из свойств конкретного генома, от которого эти полипептиды производны, а также принимая во внимание возможность замены некоторых из аминокислот в этих полипептидах без существенной потери активности (функциональные варианты), изобретение охватывает варианты указанных выше полипептидов, которые по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% и еще более предпочтительно по меньшей мере на 95% идентичны последовательностям, предусмотренным в настоящем изобретении.
Таким образом, настоящее изобретение относится к полипептидам, включающим полипептидную последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90%, 95% 97% или 99% идентична аминокислотной последовательности, выбранной из группы, включающей последовательности с SEQ ID NO: 7 по SEQ ID NO: 18, SEQ ID NO: 47 и SEQ ID NO: 48.
В рамки охвата настоящего изобретения также входят полинуклеотиды, векторы, кодирующие описанные выше редкощепящие эндонуклеазы по настоящему изобретению.
PreTalpha
Из-за природы отбора цепей TCRbeta через спаривание с preTalpha при развития тимуса, в Т-клетках, в которых инактивировали TCRalpha, гетерологичная интродукция трансгена pTalpha может привести к формированию preTCR. Такой pTCR может выступать в качестве средства активации или стимуляции Т-клеток без зависимости от МНС, таким образом, например, допуская непрерывное распространение альфа/бета Т-клеток с последующей инактивацией TCRalpha. Важно отметить, что комплекс pTCR демонстрирует сходную биохимическую композицию с TCR в отношении ассоциации субъединиц CD3 (Carrasco и др., 2001). Кроме того, в отличие от TCR, pre-TCR передача сигнала частично может быть независимой от лиганда. Кристаллическая структура внеклеточного домена pTCR обеспечивается структурной основой для возможной лиганд-независимой передачи сигнала pTCR. Показано, что pTCR формирует димер голова-хвост, в котором ассоциированы два гетеродимера pTalpha-TCRbeta (Pang и др., 2010).
Другой вариант осуществления настоящего изобретения дополнительно включает интродукцию в указанные Т-клетки pTalpha (также обозначаемого preTCRα) или его функционального варианта и размножение указанных клеток, необязательно через стимуляцию комплекса CD3. В предпочтительном варианте осуществления настоящего изобретения способ включает:
(а) Трансформацию указанных клеток нуклеиновой кислотой, кодирующей по меньшей мере фрагмент pTalpha для поддержки экспрессии CD3 на поверхности;
(б) Экспрессию указанного pTalpha в указанных клетках;
(в) Размножение указанных клеток, необязательно через стимуляцию комплекса CD3.
Настоящее изобретение также относится к способу получения Т-клеток для иммунотерапии, включающему стадии способа размножения Т-клеток.
В одном из вариантов осуществления настоящего изобретения полинуклеотидная последовательность pTalpha может быть интродуцирована случайным образом или через гомологичную рекомбинацию, в частности инсерция может быть ассоциирована с инактивацией гена TCR alpha.
Согласно настоящему изобретению применяют различные функциональные варианты pTalpha. Понятие «функциональный вариант» пептида в контексте настоящего изобретения означает молекулу, которая по существу схожа с целым пептидом или с его фрагментом. «Фрагментом» pTalpha или его функциональным вариантом в настоящем изобретении обозначают какое-либо подмножество молекулы, представленное более короткими пептидами. Предпочтительный pTalpha или его функциональный вариант может быть pTalpha полной длины или усеченной С-концевой версией pTalpha. С-концевой усеченный pTalpha утратил с С-конца один или более остатков. Примером усеченной С-концевой версии pTalpha, который не ограничивает рамок охвата настоящего изобретения, является белок, утративший остатки 18, 48, 62, 78, 92, 110 или 114 с С-конца (последовательности с SEQ ID NO: 107 по SEQ ID NO: 114). Кроме того, варианты аминокислотной последовательности пептида могут быть получены за счет мутаций в ДНК, кодирующей пептид. Такие функциональные варианты включают, например, делеции, или инсерции, или замещения остатков в аминокислотной последовательности. Какая-либо комбинация делеции, инсерции и замещения также может быть произведена для достижения итоговой конструкции, при условии, что итоговая конструкция обладает требуемой активностью, в частности восстанавливает функциональный комплекс CD3. В предпочтительном варианте осуществления настоящего изобретения по меньшей мере одну мутацию интродуцируют в различные версии pTalpha согласно описанному выше для воздействия на димеризацию. В качестве примера, который не ограничивает рамок охвата настоящего изобретения, мутантными остатками могут быть по меньшей мере W46R, D22A, K24А, R102A или R117A в белке человека pTalpha или выровненные положения, используя метод CLUSTALW применительно к семейству pTalpha или гомологичному представителю. Предпочтительно pTalpha или его вариант согласно описанному выше включает мутантный остаток W46R (SEQ ID NO:123) или мутантные остатки D22A, K24А, R102A и R117A (SEQ ID NO: 124). В одном из вариантов осуществления настоящего изобретения указанный pTalpha или его варианты также гибридизованы с сигнал-трансдуцирующим доменом, например, CD28, ОХ40, ICOS, CD27, CD137 (4-1ВВ) и CD8 (с SEQ ID NO: 115 по SEQ ID NO: 120), но эти примеры не ограничивают области охвата настоящего изобретения. Внеклеточный домен pTalpha или описанные выше варианты могут быть гибридизованы с фрагментом белка TCRalpha, особенно с трансмембранным и внутриклеточным доменом TCRalpha (SEQ ID NO: 122). Варианты pTalpha также могут быть гибридизованы с внутриклеточным доменом TCRalpha (SEQ ID NO: 121).
В другом варианте осуществления настоящего изобретения указанные версии pTalpha гибридизуют с внеклеточным лиганд-связывающим доменом, и более предпочтительно pTalpha или его функциональный вариант гибридизуют с одноцепочечным фрагментом антитела (single chain antibody fragment - scFV), включающим легкий (VL) и тяжелый (VH) вариабельный фрагмент целевого антигенспецифического моноклонального антитела, соединенного гибким линкером. В качестве примера, который не ограничивает рамок охвата настоящего изобретения, аминокислотную последовательность pTalpha или его функционального варианта выбирают из группы, включающей последовательности от SEQ ID NO: 107 по SEQ ID NO: 124.
Поскольку некоторая вариабельность может возникнуть от свойств генома, от которого происходят эти полипептиды, а также учитывая возможность замещения некоторых из аминокислот в составе этих полипептидов без значительной потери активности (функциональные варианты), настоящее изобретение включает варианты указанных выше полипептидов, которые по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90% и еще более предпочтительно по меньшей мере на 95% идентичны последовательностям, предусмотренным в настоящей патентной заявке.
Таким образом, настоящее изобретение представляет полипептиды, включающие полипептидную последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90%, 95% 97% или 99% идентична аминокислотной последовательности, выбранной из группы, включающей последовательности с SEQ ID NO: 107 по SEQ ID NO: 124.
TCR alpha-недостаточной Т-клеткой называют выделенную Т-клетку, которая утратила экспрессию функциональной TCR alpha цепи. Это может быть достигнуто разными средствами, например, путем конструирования Т-клетки таким образом, что она не экспрессирует функционального TCR alpha на своей поверхности, или путем конструирования Т-клетки таким образом, что она продуцирует очень слабо функциональную TCR alpha цепь на своей поверхности, или путем конструирования Т-клетки для экспрессии мутантной или усеченной формы TCR alpha цепи; эти примеры не ограничивают рамок охвата настоящего изобретения.
TCR alpha-недостаточные клетки уже не могут быть размножены из-за комплекса CD3. Поэтому для решения данной проблемы и для осуществления пролиферации TCR alpha-недостаточных клеток, pTalpha или его функциональный вариант интродуцируют в указанные клетки, тем самым восстанавливая функциональный CD3 комплекс. В предпочтительном варианте осуществления настоящего изобретения способ дополнительно включает интродукцию в указанные Т-клетки редкощепящих эндонуклеаз, способных селективно инактивировать путем расщепления ДНК один ген, кодирующий компонент рецептора Т-клеток (T-cell receptor - TCR). В одном из вариантов осуществления настоящего изобретении указанными редкощепящими эндонуклеазами являются TALE-нуклеазы. В качестве примеров, не ограничивающих рамок охвата настоящего изобретения, TALE-нуклеаза направлена против одной из целевых последовательностей TCRalpha, выбранных из группы, включающей последовательности SEQ ID NO: 37 и с SEQ ID NO: 57 по SEQ ID NO: 60. Предпочтительно TALE-нуклеазы выбирают из группы, включающей SEQ ID NO: 41 и SEQ ID NO: 42.
В одном из вариантов осуществления настоящего изобретения указанный способ размножения TCRalpha-недостаточных Т-клеток включает дополнительную стадию модификации генома. С помощью дополнительной стадии модификации генома может быть осуществлена интродукция в клетки для конструирования одного целевого белка. Примером, не ограничивающим рамок охвата настоящего изобретения, может быть указанный целевой белок химерный антигенный рецептор (Chimeric Antigen Receptor - CAR), в частности CAR, включающий аминокислотную последовательность SEQ ID NO: 73, многоцепочечный CAR, особенно многоцепочечный CAR, включающий аминокислотную последовательность SEQ ID NO: 125 биспецифического антитела, редкощепящие эндонуклеазы, нацеливающиеся на PDCD1 или CTLA-4, особенно нацеливающиеся на последовательности нуклеиновой кислоты с SEQ ID NO: 74 по SEQ ID NO: 78, или редкощепящая эндонуклеаза, нацеливающаяся на мишень для иммуносупрессирующего агента согласно описанию настоящего изобретения.
В рамках охвата настоящего изобретения также описаны полипептиды, кодирующие pTalpha, особенно описанные выше функциональные варианты. В предпочтительном варианте осуществления настоящее изобретение относится к pTalpha или его функциональным вариантам, гибридизованным с сигнал-трансдуцирующим доменом, например, CD28, ОХ40, ICOS, CD137 и CD8. Точнее, настоящее изобретение относится к функциональному варианту pTalpha, включающему аминокислотную последовательность, выбранную из группы, состоящей из последовательностей с SEQ ID NO: 107 по SEQ ID NO: 124. В рамках охвата настоящего изобретения также описаны полинуклеотиды, векторы, кодирующие pTalpha или его функциональные варианты, описанные выше.
К области охвата настоящего изобретения также относятся выделенные клетки или линии клеток, пригодные для осуществления указанного способа. В частности указанные выделенные клетки или линии клеток получают путем интродукции в указанные клетки pTalpha или его функционального варианта для поддержания экспрессии CD3 на поверхности. В предпочтительном варианте осуществления настоящего изобретения указанную выделенную клетку или линию клеток дополнительно модифицируют путем инактивирования гена TCRalpha. Этот ген предпочтительно инактивируют по меньшей мере одной редкощепящей эндонуклеазой. В предпочтительном варианте осуществления настоящего изобретения указанная редкощепящая эндонуклеаза является TALE-нуклеазой.
Многоцепочечный химерный антигенный рецептор (Chimeric Antigen Receptor - CAR)
В другом варианте осуществления настоящее изобретение относится к многоцепочечному химерному антигенному рецептору (CAR), в высокой степени адаптированному к получению и размножению сконструированных Т-клеток по настоящему изобретению. Многоцепочечный CAR включает по меньшей мере два из следующих компонентов:
(а) один полипептид, включающий трансмембранный домен FcεRI alpha цепи и внеклеточный лиганд-связывающий домен,
(б) один полипептид, включающий часть N- или С-концевого цитоплазматического хвоста и трансмембранный домен FcεRI beta цепи и/или
(в) два полипептида, каждый из которых включает часть внутрицитоплазматического хвоста и трансмембранный домен FcεRI gamma цепи, благодаря чему чего разные полипептиды спонтанно мультимеризуются вместе с формированием двумерного, трехмерного или тетрамерного CAR.
Пример тетрамерного CAR приведен на фиг. 3. Различные версии многоцепочечных CAR представлены на фиг. 4. Одним из примеров является многоцепочечный CAR, включающий аминокислотную последовательность SEQ ID NO: 125. Понятие «часть» относится к подмножеству молекул, являющихся более короткими пептидами. В другом варианте функциональные варианты аминокислотной последовательности полипептида могут быть получены путем мутаций ДНК, кодирующей полипептид. К таким функциональным вариантам относятся, например, формы с делециями, или инсерциями, или замещениями остатков в аминокислотной последовательности. Какая-либо комбинация делеции, инсерции и замещения также может быть сделана для получения итоговой конструкции, предусматривая, что итоговая конструкция проявляет требуемую активность, особенно специфическую клеточную иммунную активность в отношении мишени.
В предпочтительном варианте осуществления настоящего изобретения указанным внеклеточным лиганд-связывающим доменом является scFv. Помимо scFv также может применяться другой связывающий домен для предопределенного нацеливания лимфоцитов, например, фрагменты камелидизированных однодоменных антител или лиганды рецепторов, такие как полипептид - фактор роста эндотелия сосудов, интегрин-связывающий пептид, херегулин или IL-13 мутеин, антитело-связывающие домены, гипервариабельные петли антитела или CDR; указанные примеры не ограничивают рамок охвата настоящего изобретения.
В предпочтительном варианте осуществления настоящего изобретения указанный полипептид по п. (а) дополнительно включает стволовую область между указанным внеклеточным лиганд-связывающим доменом и указанным трансмембранным доменом. Понятие «стволовая область» в контексте настоящего изобретения обычно означает какой-либо олиго- или полипептид, функция которого заключается в связывании трансмембранного домена с внеклеточным лиганд-связывающим доменом. В частности стволовую область применяют для обеспечения большей гибкости и доступности по отношению к внеклеточному лиганд-связывающему домену. Стволовая область может включать до 300 аминокислот, предпочтительно от 10 до 100 аминокислот и наиболее предпочтительно от 25 до 50 аминокислот. Стволовая область может быть производной от всех или части природных молекул, например, от всех или части внеклеточной области CD8, CD4 или CD28, или от всех или части константной области антитела. В другом варианте осуществления настоящего изобретения стволовая область может быть синтетической последовательностью, которая соответствует природной стволовой последовательности, или может быть полностью синтетической стволовой последовательностью.
В предпочтительном варианте осуществления настоящего изобретения указанный полипептид по пунктам (а), (б) и/или (в) дополнительно включает по меньшей мере один сигнал-трансдуцирующий домен. В наиболее предпочтительном варианте осуществления настоящего изобретения указанный сигнал-трансдуцирующий домен выбран из группы, включающей CD28, ОХ40, ICOS, CD137 и CD8.
В предпочтительном варианте осуществления настоящего изобретения указанный С-концевой цитоплазматический хвост фрагмента цепи FcεRI alpha, beta и/или gamma дополнительно включает TNFR-ассоциированные фактор-2 (TRAF2) связывающие мотивы. В наиболее предпочтительном варианте осуществления настоящего изобретения указанный С-концевой цитоплазматический хвост FcεRI alpha, beta и/или gamma цепи замещен внутрицитоплазматическим хвостом ко-стимулирующего представителя семейства TNFR. Цитоплазматический хвост ко-стимулирующего представителя семейства TNFR содержит TRAF2-связывающие мотивы, состоящие из крупного консервативного мотива (P/S/A)X(Q/E)E) или минорного мотива (PXQXXD), где X означает какую-либо аминокислоту. Белки TRAF рекрутируются к внутриклеточным хвостам многих TNFR в ответ на тримеризацию рецептора.
В другом предпочтительном варианте осуществления настоящего изобретения указанный внутрицитоплазматический домен цепи FcεRI alpha, beta и/или gamma замещен внутрицитоплазматическим доменом цепи TCR zeta (также называемым CD3 zeta). В другом предпочтительном варианте осуществления настоящего изобретения указанный внутрицитоплазматический домен цепи FcεRI alpha, beta и/или gamma включает по меньшей мере один дополнительный активирующий тирозиновый мотив иммунных рецепторов (immunoreceptor tyrosine-based activation motif - ITAM). Мотивы ITAM являются хорошо охарактеризованными сигнальными мотивами, обнаруженными во внутрицитоплазматическом хвосте различных рецепторов, которые служат связующими сайтами для тирозиновых киназ класса syk/zap70. К примерам ITAM, применяемым в настоящем изобретении, относятся производные TCRzeta, FCRgamma, FCRbeta, CD3gamma, CD3delta, CD3epsilon, CD5, CD22, CD79a, CD79b и CD66d.
В качестве примера, который не ограничивает рамок охвата настоящего изобретения, на фиг. 4 приведены разные версии многоцепочечных CAR.
В предпочтительном варианте осуществления настоящего изобретения многоцепочечный CAR включает аминокислотную последовательность SEQ ID NO: 125. Настоящее изобретение относится к полипептидам, включающим полипептидную последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90%, 95% 97% или 99% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 125.
В рамки охвата настоящего изобретения также включены полинуклеотиды и векторы, кодирующие описанные выше многоцепочечные CAR по настоящему изобретению.
В рамках охвата настоящего изобретения предпочтительный вариант осуществления настоящего изобретения относится к способу получения Т-клеток для иммунотерапии, включающему интродукцию в указанные Т-клетки разных полипептидов, образующих указанный многоцепочечный CAR, и размножение указанных клеток.
В другом варианте осуществления настоящего изобретения указанный способ дополнительно включает стадию генетической модификации указанных клеток путем инактивации по меньшей мере одного гена, экспрессирующего один компонент TCR и/или мишень для иммуносупрессирующего агента. В предпочтительном варианте осуществления настоящего изобретения указанный ген выбран из группы, состоящей из TCR alpha, TCR beta, CD52 и GR. В предпочтительном варианте осуществления настоящего изобретения указанный способ дополнительно включает интродукцию в указанные Т-клетки редкощепящей эндонуклеазы, способной селективно инактивировать путем расщепления ДНК указанные гены. В более предпочтительном варианте осуществления настоящего изобретения указанной редкощепящей эндонуклеазой является TALE-нуклеаза. К предпочтительным TALE-нуклеазам по настоящему изобретению относятся те TALE-нуклеазы, которые распознают и расщепляют последовательность-мишень, выбранную из группы, состоящей из последовательностей: с SEQ ID NO: 1 по SEQ ID NO: 6 (GR), SEQ ID NO: 37, с SEQ ID NO: 57 по SEQ ID NO: 60 (TCR alpha), SEQ ID NO: 38 или SEQ ID NO: 39 (TCR beta), SEQ ID NO: 40, с SEQ ID NO: 61 по SEQ ID NO: 65 (CD52).
В одном из вариантов осуществления настоящего изобретения указанный способ дополнительно включает стадию модификации генома. С помощью дополнительной стадии модификации генома может быть произведена интродукция в клетки для конструирования одного целевого белка. Указанный целевой белок может быть биспецифическим антителом, редкощепящей эндонуклеазой, нацеливающейся на PDCD1 или CTLA-4, pTalpha или их функциональными вариантами, описанными в настоящем изобретении, однако эти примеры не ограничивают области охвата настоящего изобретения.
Настоящее изобретение также относится к выделенным клеткам или линиям клеток, пригодным для получения указанным способом сконструированных клеток. В частности указанные выделенные клетки включают экзогенные полинуклеотидные последовательности, кодирующие полипептиды, составляющие указанный многоцепочечный CAR.
Биспецифические антитела
В еще одном варианте осуществления настоящего изобретения сконструированные Т-клетки, полученные разными ранее описанными способами, могут быть дополнительно обработаны биспецифическими антителами. Указанные Т-клетки могут быть обработаны биспецифическими антителами ex vivo до введения пациенту или in vivo после введения пациенту. Указанные биспецифические антитела включают две вариабельные области с разными антигенными свойствами, в результате чего сконструированные клетки вступают в непосредственную близость с целевым антигеном. В качестве примера, который не ограничивает рамок охвата настоящего изобретения, указанное биспецифическое антитело направляется непосредственно против опухолевого маркера и антигена лимфоцита, например CD3, и имеет возможность перенаправить и активировать какие-либо Т-клетки в кровяном русле против опухолей.
Способы доставки
Разные описанные выше способы включают интродукцию целевого белка, например, редкощепящей эндонуклеазы, в клетку. Примером, который не ограничивает рамок охвата настоящего изобретения, является интродукция целевого белка в качестве трансгена, предпочтительно кодируемого по меньшей мере одним плазмидным вектором. Полипептиды могут быть синтезированы in situ в клетке в качестве результата интродукции в клетку полинуклеотидов, кодирующих указанные полипептиды. В другом варианте осуществления настоящего изобретения указанные полипептиды могут быть выработаны вне клетки и затем интродуцированы в нее. Методы интродукции полинуклеотидной конструкции в клетки животных известны в данной области, их примерами, которые не ограничивают рамок охвата настоящего изобретения, являются методы стабильной трансформации, в которых полинуклеотидную конструкцию интегрируют в геном клетки, методы кратковременной трансформации, в которых полинуклеотидную конструкцию не интегрируют в геном клетки, и методы, основанные на вирусах. Указанные полинуклеотиды могут быть интродуцированы в клетку, например, с помощью рекомбинантных вирусных векторов (например, ретровирусных, аденовирусных), липосом и других. Например, к методам кратковременной трансформации относятся микроинъекции, электропорация или бомбардировка частицами. Указанные полинуклеотиды могут быть включены в векторы, точнее в плазмиды или вирусы, имея в виду, что они будут экспрессироваться в клетках. Указанный плазмидный вектор может включать селективный маркер, который обеспечивает идентификацию и/или отбор клеток, получивших указанный вектор. Разные трансгены могут быть включены в один вектор. Указанный вектор может включать последовательность нуклеиновой кислоты, кодирующую последовательность рибосомального проскока, например, последовательность, кодирующую пептид 2А. Пептиды 2А, идентифицированные в подгруппе Aphthovirus пикорнавирусов, вызывают рибосомальный «проскок (скип)» от одного кодона к другому без формирования пептидной связи между двумя аминокислотами, кодируемыми кодонами (Donnelly и др., 2001; Donnelly и др., 1997; Doronina и др., 2008; Atkins и др., 2007). Понятие «кодон» означает три нуклеотида в мРНК (или в смысловой цепи молекулы ДНК), которые транслируются рибосомой в один аминокислотный остаток. Таким образом, полипептиды могут быть синтезированы из одной смежной открытой рамки считывания в мРНК, где полипептиды разделены олигопептидной последовательностью 2А, имеющейся в рамке. Такие механизмы рибосомальных проскоков известны в данной области, а также известно, что они используются в нескольких векторах для экспрессии нескольких белков, кодируемых одной мРНК. В качестве примера, который не ограничивает рамок охвата настоящего изобретения, пептиды 2А используют для экспрессии в клетке редкощепящей эндонуклеазы и фермента, процессирующего конец ДНК.
В более предпочтительном варианте осуществления настоящего изобретения полинуклеотиды, кодирующие полипептиды по настоящему изобретению, могут быть молекулами мРНК, которую интродуцируют непосредственно в клетки, например, с помощью электропорации. В настоящем изобретении определяют оптимальное условие электропорации мРНК в Т-клетки.
В настоящем изобретении применяют технологию cytoPulse, которая позволяет с помощью пульсирующих электрополей кратковременно делать живые клетки проницаемыми для поступления вещества в клетки. Эта технология, основанная на применении колебательных волн для электропорации от генератора PulseAgile (фирма ВТХ Havard Apparatus, 84 October Hill Road, Holliston, Массачусетс 01746, США), обеспечивает точный контроль длительность импульса, его интенсивности, а также интервалу между импульсами (US 6010613, WO 2004083379). Все эти параметры могут быть модифицированы для достижения наилучших условий для высоко эффективной трансфекции при минимальной гибели. По существу, первые высокие импульсы электрического поля формируют поры, последующие низкие импульсы электрического поля позволяют перемещать полинуклеотид в клетку.
Активация и размножение Т-клеток
Независимо от того, была уже проведена модификация Т-клеток или нет, Т-клетки могут быть активированы и размножены в основном методами, описанными, например, в патентах US 6352694, 6534055, 6905680, 6692964, 5858358, 6887466, 6905681, 7144575, 7067318, 7172869, 7232566, 7175843, 5883223, 6905874, 6797514, 6867041 и в патентной заявке US 20060121005. Т-клетки могут размножать in vitro или in vivo. Как правило, Т-клетки по настоящему изобретению размножают контактом с агентом, который стимулирует комплекс CD3 TCR и ко-стимулирующую молекулу на поверхности Т-клеток для создания сигнала активации Т-клетки. Например, такие химикаты, как ионофор кальция А23187, форбол 12-миристат 13-ацетат (РМА), или митогенные лектины типа фитогемагглютинина (РНА), могут применять для создания сигнала активации для Т-клетки. Примерами, не ограничивающими рамок охвата настоящего изобретения, может быть стимуляция Т-клеток in vitro, например, за счет контакта с анти-CD3 антителом, или его антигенсвязывающим фрагментом, или анти-CD2 антителом, иммобилизованным на поверхности, или путем контакта с активатором протеинкиназы С (например, с бриостатином) в сцеплении с ионофором кальция. Для ко-стимуляции дополнительной молекулы на поверхности Т-клеток применяют лиганд, который связывает дополнительную молекулу. Например, популяция Т-клеток может контактировать с анти-CD3 антителом и анти-CD28 антителом в условиях, пригодных для стимуляции пролиферации Т-клеток. Для стимуляции пролиферации либо CD4+ Т-клеток, либо CD8+ Т-клеток, применяют анти-CD3 антитело и анти-CD28 антитело. Например, агенты, обеспечивающие каждый сигнал, могут быть в растворе или соединены с поверхностью. Специалисты в данной области могут оценить, что соотношение частиц и клеток может зависеть от размера частиц относительно клеток-мишеней.
К условиям, пригодным для культуры Т-клеток, относится соответствующая среда (например, минимальная поддерживающая среда, или среда RPMI 1640, или X-vivo 5, (фирма Lonza)), которая может содержать факторы, требуемые для пролиферации и жизнеспособности. К таким факторам относятся сыворотка (например, фетальная сыворотка теленка или сыворотка человека), интерлейкин-2 (IL-2), инсулин, IFN-g, 1L-4, 1L-7, GM-CSF, -10,-2, 1L-15, TGFp и TNF- или какие-либо другие дополнительные факторы для роста клеток, известные специалистам в данной области. К другим дополнительным факторам для роста клеток относятся, но ими перечень не ограничивается, поверхностно-активное вещество, плазманат, и восстанавливающие агенты, например, N-ацетил-цистеин и 2-меркаптоэтанол. К средам относятся RPMI 1640, A1M-V, DMEM, MEM, a-MEM, F-12, X-Vivo 1 и X-Vivo 20, Optimizer, в которые добавляют аминокислоты, пируват натрия и витамины, не добавляют или добавляют требуемое количество сыворотки (или плазмы), или определенный набор гормонов, и/или количество цитокина (цитокинов), достаточное для роста и размножения Т-клеток. Антибиотики, например, пенициллин и стрептомицин, добавляют только в экспериментальные культуры, но не в те культуры клеток, которые вводят инфузией субъекту. Целевые клетки поддерживают в условиях, требуемых для сохранения роста, например, при соответствующей температуре (например, 37°С) и атмосфере (например, воздух плюс 5% СО2). Т-клетки, подвергнутые различной стимуляции, могут проявлять разные свойства.
Сконструированные Т-клетки
В рамки охвата настоящего изобретения также включена выделенная Т-клетка, полученная одним из ранее описанных способов. Указанная Т-клетка по настоящему изобретению может быть производной стволовой клетки. Стволовые клетки могут быть стволовыми клетками взрослых, эмбриональными стволовыми клетками, в частности стволовыми клетками, не являющимися стволовыми клетками человека, стволовыми клетками пуповинной крови, клетками-предшественницами, стволовыми клетками костного мозга, индуцированными мультипотентными стволовыми клетками, тотипотентными стволовыми клетками или кроветворными стволовыми клетками. Образцом клеток человека являются клетки человека CD34+. Указанные выделенные клетки также могут быть древовидными клетками, клетками - природными киллерами, В-клетками или Т-клетками, выбранными из группы, состоящей из воспалительных Т-лимфоцитов, цитотоксических Т-лимфоцитов, регуляторных Т-лимфоцитов или лимфоцитов Т-хелперов. В другом варианте осуществления настоящего изобретения указанная клетка может быть производной от группы клеток, включающей CD4+ Т-лимфоциты и CD8+ Т-лимфоциты. Перед размножением и генетической модификацией клеток по настоящему изобретению источник клеток может быть получен от субъекта одним из многочисленных методов, перечень которых не ограничен. Т-клетки могут быть получены от ряда источников, которыми перечень не ограничивается и к которым относятся мононуклеарные клетки периферической крови, костный мозг, ткань лимфоузла, ткань тимуса, ткань из места инъекции, асцит, плевральное истечение, ткань селезенки и опухоли. В некоторых вариантах осуществления настоящего изобретения может применяться какое-либо количество доступных линий Т-клеток, известных и доступных специалистам в данной области. В другом варианте осуществления настоящего изобретения указанные клетки могут быть получены от здорового донора, от пациента с диагнозом рак, или от пациента с диагнозом инфекция. В другом варианте осуществления настоящего изобретения указанная клетка является частью смешанной популяции клеток, обладающих разными фенотипическими характеристиками. В рамки охвата настоящего изобретения также включена линия клеток, полученная от трансформированных Т-клеток ранее описанным методом. В другом предпочтительном варианте осуществления настоящее изобретение относится к выделенной Т-клетке, включающей по меньшей мере два инактивированных гена, кодирующих белки иммунных контрольных точек, точнее по меньшей мере два гена, выбранных из группы, включающей: CTLA4, РРР2СА, РРР2СВ, PTPN6, PTPN22, PDCD1, LAG3, HAVCR2, BTLA, CD160, TIGIT, CD96, CRTAM, LAIR1, SIGLEC7, SIGLEC9, CD244, TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT1, FOXP3, PRDM1, BATF, GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3.
В другом варианте осуществления настоящего изобретения указанная выделенная клетка по настоящему изобретению может дополнительно включать один инактивированный ген, выбранный из группы, состоящей из CD52, GR, TCR alpha и TCR beta, и/или экспрессирует CAR, многоцепочечный CAR и/или pTalpha трансген.
Применение в терапии
В другом варианте осуществления настоящего изобретения выделенную клетку, полученную разными методами, или линию клеток, производную от указанной выделенной клетки согласно описанному ранее, можно использовать в качестве лекарственного средства. В другом варианте осуществления настоящего изобретения указанное лекарственное средство можно использовать для лечения рака или инфекций у пациента, нуждающегося в этом. В другом варианте осуществления настоящего изобретения указанная выделенная клетка по настоящему изобретению или линия клеток, производная от указанной выделенной клетки, может применяться для получения лекарственного средства для лечения рака или вирусной инфекции у пациента, нуждающегося в этом.
Другой объект настоящего изобретения относится к способам лечения пациента, нуждающегося в этом, причем указанный способ включает по меньшей мере одну из следующих стадий:
(а) получения Т-клетки каким-либо из ранее описанных способов;
(б) введения указанных трансформированных Т-клеток указанному пациенту.
В одном из вариантов осуществления настоящего изобретения указанные Т-клетки по настоящему изобретению могут претерпевать in vivo надежное размножение Т-клеток и могут сохраняться на протяжении длительного времени.
Указанное лечение может улучшать состояние больного, быть лечебным или профилактическим. Оно может быть либо частью аутологической иммунотерапии, либо частью аллогенной иммунотерапии. Понятие «аутологические» означает, что клетки, линии клеток или популяция клеток, используемые для лечения пациентов, происходят от указанного пациента или от совместимого донора человеческого лейкоцитарного антигена (Human Leucocyte Antigen - HLA). Понятие «аллогенные» означает, что клетки или популяция клеток, используемые для лечения пациентов, происходят не от указанного пациента, а от донора.
Клетки, которые могут применяться в раскрываемых способах, описаны в предыдущем разделе. Указанное лечение могут применять для лечения пациентов с диагнозами рак, вирусная инфекция, аутоиммунные расстройства или состояние «трансплантат против хозяина» (GvHD). К заболеваниям раком, которые можно вылечить, относятся неваскуляризированные опухоли или еще недостаточно васкуляризированные опухоли, а также васкуляризованные опухоли. К заболеваниям раком могут относиться несолидные опухоли (например, онкологические заболевания системы кроветворения, лейкозы и лимфомы) или солидные опухоли. К типам рака, поддающимся лечению CAR по настоящему изобретению, относятся, но ими не ограничиваются, карцинома, бластома и саркома, а также некоторые формы лейкоза или лимфоидных злокачественных заболеваний, доброкачественные и злокачественные опухоли, и такие злокачественные заболевания, как саркомы, карциномы и меланомы. К этому перечню также относятся опухоли/рак взрослых и опухоли/рак детей.
Лечение рака могут проводить в комбинации с одной или несколькими другими терапиями, выбранными из группы, включающей лечение антителами, химиотерапию, терапию цитокинами, терапию древовидными клетками, генную терапию, терапию гормонами, терапию светом лазера и терапию радиацией.
Введение клеток или популяции клеток по настоящему изобретению может быть выполнено каким-либо традиционным путем, включая аэрозольную ингаляцию, инъекцию, проглатывание, трансфузию, имплантацию или трансплантацию. Описанные в настоящем изобретении композиции могут вводить пациенту подкожно, внутрикожно, внутрь опухоли, внутрь узлов, интрамедуллярно, внутримышечно, внутривенной или внутрилимфатической инъекцией, или внутрибрюшинно. В одном из вариантов осуществления настоящего изобретения композиции клеток по настоящему изобретению предпочтительно вводят внутривенной инъекцией.
Введение клеток или популяции клеток по настоящему изобретению может заключаться во введении 104-109 клеток/кг массы тела, предпочтительно от 105 до 106 клеток/кг массы тела, включая все целые величины количества клеток в данных диапазонах. Клетки или популяции клеток могут вводить в виде одной или более доз. В другом варианте осуществления настоящего изобретения указанные эффективные количества клеток вводят в виде однократной дозы. В другом варианте осуществления настоящего изобретения указанное эффективное количество клеток вводят в виде более чем одной дозы на протяжении определенного периода времени. Сроки введения находятся в пределах, установленных врачом, и зависят от клинического состояния пациента. Клетки или популяции клеток могут получать от какого-либо источника, например, из банка крови или от донора. Хотя индивидуальные потребности варьируют, определение оптимальных диапазонов эффективных количеств полученных клеток для определенного заболевания или состояния находится в ведении специалиста в данной области. Эффективным является количество, обеспечивающее терапевтическую или профилактическую пользу. Вводимая доза зависит от возраста, здоровья и массы тела реципиента, типа сопутствующего лечения, если таковое имеется, частоты лечения и природы требуемого эффекта.
В другом варианте осуществления настоящего изобретения указанное эффективное количество клеток или композиции, включающей эти клетки, вводят парентерально. Указанное введение может быть внутривенным введением. Указанное введение может быть выполнено инъекцией непосредственно в опухоль.
В некоторых вариантах осуществления настоящего изобретения пациенту вводят клетки в сочетании (например, до, одновременно или после) с каким-либо числом соответствующих методов лечения, включая, но ими не ограничиваясь, лечение агентами, например, противовирусными средствами цидофовиром и интерлейкином-2, цитарабином (также называемым ARA-C) или натализумабом для пациентов с рассеянным склерозом, или лечение эфализумабом пациентов с псориазом, или другие формы лечения для пациентов с прогрессирующей мультифокальной лейкоэнцефалопатией (ПМЛ). В других вариантах осуществления настоящего изобретения Т-клетки по настоящему изобретению могут применять в комбинации с химиотерапией, радиацией, иммуносупрессивными агентами, например, с циклоспорином, азатиоприном, метотрексатом, микофенолятом и FK506, антителами или другими иммуноабляционными агентами, например, САМРАТН, анти-CD3 антителами или другими антителами, предназначенными для лечения, цитоксином, фладарабином, циклоспорином, FK506, рапамицином, микофеноловой кислотой, стероидами, FR901228, цитокинами и облучением. Эти лекарственные средства ингибируют или кальций-зависимую фосфатазу кальценеврин (циклоспорин и FK506), или ингибируют p70S6 киназу, которая важна для передачи сигнала, индуцированного фактором роста (рапамицин) (Liu и др., 1991; Henderson и др., 1991; Bierer и др., 1993). В другом варианте осуществления настоящего изобретения композиции клеток по настоящему изобретению вводят пациенту в сочетании (например, до, одновременно или после) с трансплантатом костного мозга, Т-клеточной абляционной терапией, используя химиотерапевтические агенты, например, флударабин, наружную дистанционную лучевую терапию (НДЛТ), циклофосфан или антитела, например, ОKТ3 или САМРАТН. В другом варианте осуществления настоящего изобретения композиции клеток по настоящему изобретению назначают после В-клеточной абляционной терапии, например, агентами, которые реагируют с CD20, например, ритуксаном. Например, в одном из вариантов осуществления настоящего изобретения субъекты могут пройти стандартное лечение высокими дозами химиотерапевтических средств с последующей трансплантацией стволовых клеток периферической крови. В определенных вариантах осуществления настоящего изобретения после трансплантации субъекты получают инфузию размноженных иммунных клеток по настоящему изобретению. В еще одном из вариантов осуществления настоящего изобретения размноженные клетки вводят до или после хирургического вмешательства.
Пример способа конструирования аллогенных клеток человека для иммунотерапии
Для лучшего понимания настоящего изобретения на фиг. 5 приводят пример способа конструирования аллогенных клеток человека для иммунотерапии. Способ включает комбинацию одной или нескольких из следующих стадий:
1. Получения Т-клеток из культуры клеток, или из образца крови от индивидуального пациента, или из банка крови, и активирование указанных Т-клеток, используя анти-CD3/С28 активаторные гранулы. Гранулы обеспечивают как первичные, так и ко-стимулирующие сигналы, необходимые для активирования и размножения Т-клеток.
2. (а) Трансдукции указанных клеток pTalpha или функциональным вариантом его трансгена для поддержки экспрессии CD3 на поверхности и осуществления размножения клеток через стимуляцию комплекса CD3. Предполагают, что разрушение комплекса TCR приводит к элиминации комплекса TCR и удалению аллореактивности (GvHD), но может разрушать размножение аллогенных клеток из-за утраты CD3 сигнального компонента. Ожидают, что трансдуцированные клетки экспрессируют цепь pTalpha или ее функциональный вариант. Такая цепь pTalpha спаривается с цепью TCRbeta и сигнальными компонентами CD3 для формирования комплекса preTCR и таким образом восстановления функционального комплекса CD3 и поддержки активации или стимуляции инактивированных клеток TCRalpha. Трансдукция Т-клеток pTalpha лентивирусным вектором может быть реализована до или после инактивации TCRalpha.
(б) Трансдукции указанных клеток многоцепочечными CAR позволяют перенаправить Т-клетки против антигенов, экспрессированных на поверхности целевых клеток разных злокачественных образований, включая лимфомы и солидные опухоли. Для улучшения функции ко-стимулирующего домена в настоящем изобретении сконструирован многоцепочечный CAR, производный от FcεRI, согласно ранее описанному. Трансдукция может быть осуществлена до или после инактивации TCRalpha и других генов, например, генов CD52.
3. Конструирования неаллореактивных и устойчивых к иммуносупрессии Т-клеток:
(а) Возможно инактивировать TCR alpha в указанных клетках для элиминации TCR с поверхности клетки и предупреждения распознавания ткани хозяина в качестве чужеродной аллогенным TCR и таким образом избежать GvHD.
(б) Также возможно инактивировать один ген, кодирующий мишень для иммуносупрессивного агента для компенсации устойчивости указанных клеток к иммуносупрессивной обработке для предупреждения отторжения трансплантата без воздействия на трансплантированные Т-клетки. В этом примере мишенью иммуносупрессивных агентов является CD52 и иммуносупрессивным агентом является гуманизированное моноклональное анти-CD52 антитело.
В настоящем изобретении установлено, что применение TALE-нуклеазы, обеспечивающее более высокую степень двунитевых разрывов (DSB) в Т-клетках, особенно предпочтительно для достижения указанной выше двойной инактивации в Т-клетках. Предпочтительно гены TCRalpha и CD52 инактивируют путем электропорации Т-клеток мРНК, кодирующей указанные гены, на которые нацеливается TALE-нуклеаза. В настоящем изобретении установлено, что применение мРНК, приводящее к высокой степени трансформации, менее губительно для Т-клеток и поэтому является критическим в процессе конструирования Т-клеток. Затем инактивированные Т-клетки сортируют, используя магнитные гранулы. Например, Т-клетки, экспрессирующие CD52, удаляют фиксацией на плотной поверхности, и инактивированные клетки не подвергаются стрессу при пропуске через колонку. Такой мягкий метод повышает концентрацию должным образом сконструированных Т-клеток.
4. Размножения in vitro сконструированных Т-клеток перед введением пациенту или in vivo после введения пациенту путем стимуляции комплекса CD3. Перед стадией введения пациенты могут подвергаться иммуносупрессивной обработке, например, САМРАТН1-Н, гуманизированным моноклональным антителом анти-CD52.
5. Необязательной обработки указанных клеток биспецифическими антителами ex vivo перед введением пациенту или in vivo после введения пациенту для доставки сконструированных клеток к целевому антигену.
Другие определения
- В контексте настоящего изобретения понятия «нуклеиновая кислота» или «полинуклеотиды» относятся к нуклеотидам и/или полинуклеотидам, например, к дезоксирибонуклеиновой кислоте (ДНК) или рибонуклеиновой кислоте (РНК), олигонуклеотидам, фрагментам, полученным полимеразной цепной реакцией (ПЦР), и фрагментам, полученным за счет какой-либо из следующих активностей: лигированием, расщеплением, действием эндонуклеазы и действием экзонуклеазы. Молекулы нуклеиновой кислоты могут состоять из мономеров, являющихся природными нуклеотидами (например, ДНК и РНК), или аналогов природных нуклеотидов (например, энантиомерных форм природных нуклеотидов), или из комбинации обоих. Модифицированные нуклеотиды могут иметь изменения в остатках сахаров и/или в частях пиримидиновых или пуриновых оснований. Модификации сахаров включают, например, замещение одной или более гидроксильных групп на галогены, алкильные группы, амины и азидо-группы, или сахара могут функционировать в качестве эфиров или сложных эфиров. Кроме того, весь остаток сахара может быть заменен на структуры, схожие стерически и электронно, например, на аза-сахара и карбоциклические аналоги сахаров. К примерам модификаций в основаниях относятся алкилированные пурины и пиримидины, ацилированные пурины или пиримидины или другие хорошо известные гетероциклические заместители. Мономеры нуклеиновых кислот могут быть связаны фосфодиэфирной связью или аналогами таких связей. Нуклеиновые кислоты могут быть одноцепочечными или двухцепочечными.
- Понятия «целевая ДНК», «последовательность целевой ДНК», «целевая последовательность ДНК», «целевая последовательность нуклеиновой кислоты», «целевая последовательность» или «процессируемый сайт» означают полинуклеотидную последовательность, на которую может нацеливаться и которую может процессировать редкощепящая эндонуклеаза по настоящему изобретению. Эти обозначения относятся к специфическому расположению ДНК, предпочтительно геномному расположению в клетке, но также к части генетического материала, который может существовать независимо относительно основного генетического материала, например, в составе плазмид, эписом, вирусов, транспозонов или органелл, например, митохондрий, однако приведенными примерами перечень не ограничивается. Примерами мишеней TALE-нуклеаз, которые не ограничивают рамок охвата настоящего изобретения, являются геномные последовательности нацеливания, обычно состоящие из двух последовательностей длиной 17 п.о. (называемых полумишенями), разделенных спейсером из 15 п.о. Каждую полумишень распознают повторы TALE-нуклеаз, которые кодируются плазмидами под контролем промотора EF1-alpha или промотора Т7, перечисленные в табл. 2, 6, 7 и 11 в качестве примеров, не ограничивающих настоящее изобретение. Целевая последовательность нуклеиновой кислоты выражена последовательностью 5' к 3' одной цепи указанной мишени, показанной в табл. 1, 5, 6 и 10.
- Понятие «трансген» означает последовательность нуклеиновой кислоты (кодирующей, например, один или несколько полипептидов), которая частично или полностью гетерологична, т.е. чужеродна, для клетки-хозяина, в которую она интродуцирована, но которая может быть сконструирована для изменения генома клетки, в которую она инсертирована (например, последовательность инсертирована по месту, которое отличается от места расположения природного гена, или ее инсерция приводит к нокауту). Трансген может включать одну или несколько последовательностей регуляции транскрипции и какую-либо другую нуклеиновую кислоту, например, интроны, которые могут быть необходимы для оптимальной экспрессии выбранной нуклеиновой кислоты, кодирующей полипептид. Полипептид, кодируемый трансгеном, может быть не экспрессирован, или экспрессирован, но биологически неактивен, в клетках, в которые трансген инсертирован.
- Понятие «геном» означает совокупный генетический материал, содержащийся в клетке, например, геном ядра, геном хлоропластов, геном митохондрий
- Понятие «химерный антигенный рецептор (CAR)» означает молекулы, которые объединяют связывающий домен против компонента, присутствующего на целевых клетках, например, специфичность на основе антитела в отношении желаемого антигена (например, опухолевого антигена) с Т-клеточный-рецептор-активирующим внутриклеточным доменом для получения химерного белка, который проявляет специфичную клеточную иммунную активность против мишени. Обычно CAR состоит из внеклеточного одноцепочечного антитела (scFvFc), гибридизированного с внутриклеточным сигнальным доменом Т-клеточного антигенного комплекса zeta цепи (scFvFc:ζ) и способен, при экспрессии в Т-клетках, переориентировать распознавание антигена, основанное на специфичности моноклональных антител. Одним из примеров CAR, используемым в настоящем изобретении, является CAR, направленный против антигена CD19 и способный включать в качестве примера, не ограничивающего настоящее изобретение, аминокислотную последовательность SEQ ID NO: 73.
- Понятия «вектор доставки» или «векторы доставки» означают какой-либо вектор доставки, который может применяться в настоящем изобретении для обеспечения контакта с клеткой («контактирование») или доставки внутрь клеток или субклеточных компартментов («интродукция») агентов/химических соединений и молекул (белков или нуклеиновых кислот), необходимых по настоящему изобретению. К ним относятся, но ими перечень не ограничивается, липосомальные векторы доставки, вирусные векторы доставки, лекарства - векторы доставки, химические носители, полимерные носители, липоплексы, полиплексы, дендримеры, микропузырьки (эхоконтрастные препараты), наночастицы, эмульсии или другие соответствующие векторы переноса. Такие векторы доставки позволяют доставлять молекулы, химические соединения, макромолекулы (гены, белки) или другие векторы, например, плазмиды, пептиды, разработанные фирмой Diatos. В этих случаях векторы доставки являются молекулярными носителями. Понятия «вектор доставки» или «векторы доставки» также означают методы доставки для осуществления трансфекции.
- Понятия «вектор» или «векторы» означают молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой связан вектор. В контексте настоящего изобретения к векторам относятся, но ими перечень не ограничивается, вирусный вектор, плазмида, вектор РНК, или линейная или кольцевая молекула ДНК или РНК, которая может состоять из хромосомных, не хромосомных, полусинтетических или синтетических нуклеиновых кислот. Предпочтительными являются те векторы, которые способны к автономной репликации (эписомальный вектор) и/или экспрессии нуклеиновых кислот, с которыми они связаны (векторы экспрессии). Специалистам в данной области известно большое количество применимых и коммерчески доступных векторов.
К вирусным векторам относятся ретровирусы, аденовирусы, парвовирусы (например, аденоассоциированные вирусы), коронавирусы, вирусы, содержащие минус-цепь РНК, например, ортомиксовирус (например, вирус гриппа), рабдовирус (например, вирус бешенства и вирус везикулярного стоматита), парамиксовирус (например, вирус кори и вирус Сендай), вирусы, содержащие плюс-цепь РНК, например, пикорнавирус и альфавирус, вирусы двухцепочечной ДНК, включая аденовирус, герпесвирус (например, Herpes Simplex вирус, тип 1 и 2, вирус Эпштейна-Барра, цитомегаловирус) и поксвирус (например, вирус осповакцины, вирус оспы кур, канарипокс). К другим вирусам относятся, например, вирус Норволк, тогавирус, флавивирус, реовирусы, паповирус, гепаднавирусы и вирусы гепатита. К примерам ретровирусов относятся: вирус лейкоза-саркомы птиц, вирусы млекопитающих типа С, вирусы типа В, вирусы типа D, вирусы группы HTLV-BLV, лентивирус, спумавирус (Coffin в кн.: «Retroviridae: The viruses and their replication, In Fundamental Virology», под ред. Fields и др., 1996, 3-е изд., изд-во Lippincott-Raven Publishers, Филадельфия).
- Понятия «лентивирусный вектор» означает лентивирусные векторы на основе ВИЧ, которые весьма перспективны для доставки генов благодаря их относительно большой вместимости, пониженной иммуногенности и способности стабилизировать трансдукцию с высокой эффективностью в большом диапазоне клеток разных типов. Лентивирусные векторы обычно получают в результате кратковременной трансфекции трех (упаковка, оболочка и перенос) или более плазмид в клетки-продуценты. Подобно ВИЧ, лентивирусные векторы входят в целевые клетки через взаимодействие гликопротеинов на поверхности вируса с рецепторами на поверхности клетки. После вхождения в клетку РНК вируса подвергается обратной транскрипции, которая опосредуется комплексом вирусной обратной транскриптазы. Продуктом обратной транскрипции является двухцепочечная линейная вирусная ДНК, которая является объектом для интеграции вируса в ДНК инфицированных клеток. Понятие «интегративные лентивирусные векторы (LV)» относится к вирусам, которые являются примерами, но не ограничивают рамок охвата настоящего изобретения; они могут интегрировать в геном целевых клеток. Противоположное понятие «неинтегративные лентивирусные векторы (NILV)» относится к эффективным векторам генной доставки, которые не интегрируют в геном целевых клеток через действие вирусной интегразы.
- Векторы доставки и векторы могут быть ассоциированы или скомбинированы с какими-либо методами клеточной проницаемости, например, с сонопорацией, или электропорацией, или производными этих методов.
- Понятие «клетка» или «клетки» означает какие-либо живые клетки, первичные клетки и линии клеток, производные от этих организмов для культур in vitro.
- Понятие «первичная клетка» или «первичные клетки» означает клетки, взятые непосредственно от живой ткани (т.е. материал биопсии) для роста in vitro; эти клетки претерпели очень мало удвоений клеточной популяции, и, следовательно, они более пригодны по многим функциональным компонентам и свойствам тканей, от которых они производны, по сравнению с непрерывными опухолеродными или искусственно иммортализованными линиями клеток.
Примеры клеточных линий, которые не ограничивают рамок охвата настоящего изобретения, могут быть выбраны из группы, включающей клетки СНО-K1; клетки HEK293; клетки Сасо2; клетки U2-OS; клетки NIH 3Т3; клетки NSO; клетки SP2; клетки CHO-S; клетки DG44; клетки K-562, клетки U-937; клетки MRC5; клетки IMR90; клетки Jurkat; клетки HepG2; клетки HeLa; клетки НТ-1080; клетки НСТ-116; клетки Hu-h7; клетки Huvec; клетки Molt 4.
Все эти клеточные линии могут быть модифицированы способом, представленным в настоящем изобретении, для получения моделей клеточных линий для выработки, экспрессии, измерения, обнаружения, изучения целевого гена или белка; эти модели также можно использовать для скрининга представляющих интерес биологически активных молекул в исследовании и выработке и в различных областях, например, химической, в биотопливе, терапевтической и агрономической в качестве примеров, не ограничивающих рамок охвата настоящего изобретения.
- Понятие «мутация» означает замещение, делецию, инсерцию, которые затрагивают до одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати, двенадцати, тринадцати, четырнадцати, пятнадцати, двадцати, двадцати пяти, тридцати, сорока, пятидесяти или более нуклеотидов/аминокислот в последовательности полинуклеотида (кДНК, ген) или полипептида. Мутация может влиять на кодирующую последовательность гена или его регуляторную последовательность. Она также может воздействовать на структуру геномной последовательности или на структуру/стабильность кодированной мРНК.
- Понятие «вариант (варианты)» означает повторяющийся вариант, вариант, ДНК-связывающий вариант, вариант TALE-нуклеазы, полипептидный вариант, полученный мутацией или замещением по меньшей мере одного остатка в аминокислотной последовательности исходной молекулы.
- Понятие «функциональный вариант» означает каталитически активный мутант белка или белкового домена; такой мутант может иметь ту же активность, что и его исходный белок или белковый домен, или может обладать дополнительными свойствами, или повышенной или пониженной активностью.
- Понятие «ген» означает основную единицу наследственности, состоящую из сегмента ДНК, собранного линейно вдоль хромосомы, которая кодирует специфический белок или сегмент белка. Ген обычно включает промотор, 5' нетранслируемую область, одну или более кодирующих последовательностей (экзонов), необязательно интронов, 3' нетранслируемую область. Ген может дополнительно включать терминатор, энхансеры и/или сайленсеры.
- В контексте настоящего изобретения понятие «локус» означает специфическую физическую локализацию последовательности ДНК (например, гена) на хромосоме. Понятие «локус» может означать специфическую физическую локализацию целевой последовательности редкощепящей эндонуклеазы на хромосоме. Такой локус может включать целевую последовательность, которая распознается и/или расщепляется редкощепящей эндонуклеазой по настоящему изобретению. Очевидно, что целевой локус по настоящему изобретению может не только уточнить последовательность нуклеиновой кислоты, которая существует в основном массиве генетического материала (т.е. в хромосоме) клетки, но также часть генетического материала, которая может существовать независимо от указанного основного массива генетического материала, например, плазмиды, эписомы, вирусы, транспозоны или в органеллах, например, в митохондриях, однако приведенные примеры не ограничивают рамок охвата настоящего изобретения.
- Понятие «редкощепящая эндонуклеаза» означает какой-либо фермент дикого типа или его вариант, способный катализировать гидролиз (расщепление) связей между нуклеиновыми кислотами в молекуле ДНК или РНК, предпочтительно в молекуле ДНК. В частности, указанная эндонуклеаза, более предпочтительно редкощепящая эндонуклеаза, которая высоко специфична, распознающая сайты мишени нуклеиновой кислоты с 10 до 45 п.о. в длину, обычно варьируя от 10 до 35 п.о. в длину. Эндонуклеаза по настоящему изобретению распознает и расщепляет нуклеиновую кислоту в специфических полинуклеотидных последовательностях, также называемых «целевым последовательностями». Редкощепящая эндонуклеаза может распознать и произвести одно- или двухцепочечные разрывы в определенных полинуклеотидных последовательностях.
В одном из вариантов осуществления настоящего изобретения указанная редкощепящая эндонуклеаза по настоящему изобретению может быть эндонуклеазой Cas9. В самом деле, ранее новое средство конструирования генома было разработано, основываясь на РНК-направленной нуклеазе Cas9 (Gasiunas и др., 2012; Jinek и др., 2012; Corn и др., 2013; Mali и др., 2013) от типа II CRISPR (коротких палиндромных повторов, регулярно расположенных группами - Clustered Regularly Interspaced Short palindromic Repeats) адаптивной иммунной системы (см. обзор Sorek и др., 2013). Система Cas (CRISPR Associated) впервые была открыта у бактерий, она функционирует как защита от чужеродной ДНК, и вирусной, и плазмидной. CRISPR-опосредованное конструирование генома сначала развивалось путем отбора целевой последовательности, зачастую фланкированной мотивом короткой последовательности, называемой прото-спейсерным сопредельным мотивом (proto-spacer adjacent motif - РАМ). После отбора целевой последовательности специфическую crRNA конструируют комплементарной к этой последовательности. Транс-активирование crRNA (tracrRNA) необходимо для систем II типа CRISPR, спаренных с crRNA и связанных с поступившим белком Cas9. Cas9 действует в качестве молекулярного якоря, способствующего спариванию оснований tracRNA и cRNA (Deltcheva и др., 20111). В таком тройном комплексе двойная tracrRNA:crRNA структура действует в качестве направляющей РНК, которая направляет эндонуклеазу Cas9 к родственной целевой последовательности. Распознавание целевой последовательности с помощью комплекса Cas9-tracrRNA:crRNA инициируется сканированием целевой последовательности для выявления гомологии между целевой последовательностью и crRNA. Помимо комплементарности целевой последовательности-crRNA, нацеливание на ДНК требует наличия короткого мотива, смежного с протоспейсером (protospacer adjacent motif - РАМ). После спаривания двойной РНК и целевой последовательности Cas9 интродуцирует тупой двойной разрыв цепи на три основания выше по цепи от мотива РАМ (Garneau и др., 2010). В настоящем изобретении направляющая РНК может быть сконструирована, например, для специфического нацеливания гена, кодирующего компонент TCR. После спаривания направляющей РНК и целевой последовательности Cas9 индуцирует расщепление в гене TCR.
Редкощепящие эндонуклеазы могут быть хоминг-эндонуклеазой, также называемой мегануклеазой. Такие хоминг-нуклеазы известны в данной области (Stoddard, 2005). Хоминг-нуклеазы высокоспецифичны, они распознают целевые сайты ДНК размером от 12 до 45 п.о. в длину, обычно от 14 до 40 п.о. в длину. Хоминг-нуклеазы по настоящему изобретению могут, например, соответствовать LAGLIDADG эндонуклеазе, HNH эндонуклеазе, или GIY-YIG эндонуклеазе. Предпочтительной хоминг-эндонуклеазой по настоящему изобретению может быть вариант I-CreI. «Вариантом» эндонуклеазы, т.е. эндонуклеазы, которая не существует в природе, а получена методом генной инженерии или методом случайного мутагенеза, может связывать последовательности ДНК, отличающиеся от тех, которые распознают эндонуклеазы дикого типа (см. WO 2006/097854).
Указанная редкощепящая эндонуклеаза может быть модульной ДНК-связывающей нуклеазой. Понятие «модульная ДНК-связывающая нуклеаза» означает какой-либо гибридный белок, включающий по меньшей мере один каталитический домен эндонуклеазы и по меньшей мере один ДНК-связывающий домен или белок, точно определяющий целевую последовательность нуклеиновой кислоты. ДНК-связывающий домен, обычно РНК- или ДНК-связывающий домен, сформированный независимо складывающимся доменом полипептида, который содержит по меньшей мере один мотив, который распознает двойные или однонитевые разрывы последовательностей нуклеиновой кислоты. Такие связывающие домены часто включают домены спираль-петля-спираль, домены лейциновой молнии, домены «крылатая спираль», домены HMG-бокс, домены иммуноглобулина, домен В3 или сконструированный цинковопальцевый домен; данные примеры не ограничивают рамок охвата настоящего изобретения.
В предпочтительном варианте осуществления настоящего изобретения ДНК-связывающий домен является производным эффектора, подобного активатору транскрипции (Transcription Activator Like Effector - TALE), причем специфичность последовательности направляется сериями из 33-35 аминокислотных повторов, происходящих от бактериальных белков Xanthomonas или Ralstonia. Эти повторы по существу отличаются двумя положениями аминокислот, которые определяют взаимодействие с парой оснований (Boch, Scholze и др., 2009; Moscou и Bogdanove, 2009).
Каждая пара оснований в ДНК мишени контактирует с одним повтором, со специфичностью являясь результатом двух вариантов аминокислот в повторе (так называемый «повторяемый вариабельный дипептид»; repeat variable dipeptide - RVD). TALE-связывающие домены могут дополнительно включать N-концевой домен транслокации, ответственный за потребность в первом основании тимине (Т0) целевой последовательности, и С-концевой домен, который содержит сигналы ядерной локализации (nuclear localization signals - NLS). Домен связывания нуклеиновой кислоты TALE обычно соответствует сконструированному коровому TALE стволу, включающему множество TALE-повторяющихся последовательностей, каждый повтор, включающий RVD, специфичен каждому нуклеотидному основанию сайта распознавания TALE. В настоящем изобретении каждая TALE-повторяющаяся последовательность указанной срединной части ствола состоит из 30-42 аминокислот, более предпочтительно 33 или 34, в которых две критические аминокислоты (RVD), локализованные в положениях 12 и 13, определяют распознавание одного нуклеотида в указанной последовательности сайта связывания TALE; эквивалент двух критических аминокислот может быть локализован по положениям 12 и 13 специально в TALE-повторяющейся последовательности, длина которой превышает 33 или 34 аминокислоты. Предпочтительно дипептидами RVD, ассоциированными с распознаванием разных нуклеотидов, являются HD для распознавания С, NG для распознавания Т, NI для распознавания А, NN для распознавания G или А. В другом варианте осуществления настоящего изобретения критические аминокислоты 12 и 13 могут быть мутантными и замененными другими аминокислотными остатками для модулирования их специфичности по отношению к нуклеотидам А, Т, С и G и в частности для повышения их специфичности. Домен, связывающий TALE нуклеиновую кислоту, обычно включает от 8 до 30 TALE повторяющихся последовательностей. Более предпочтительно указанный коровый ствол по настоящему изобретению включает от 8 до 20 последовательностей повтора TALE; еще более предпочтительно 15 последовательностей повтора TALE. Он также может включать дополнительную единичную усеченную последовательность повтора TALE, составленную из 20 аминокислот, расположенных с С-конца указанной серии последовательностей повтора TALE, т.е. дополнительная С-концевая полу-TALE повторяющаяся последовательность.
Другие сконструированные ДНК-связывающие домены являются модульными специфическими доменами специфического основание-на-основание связывания нуклеиновой кислоты (modular base-per-base specific nucleic acid binding domains - MBBBD) (PCT/US2013/051783). Указанные белки MBBBD могут быть сконструированы, например, из вновь идентифицированных белков, а именно белков EAV36_BURRH, E5AW43_BURRH, E5AW45_BURRH и E5AW46_BURRH из ранее секвенированного генома эндосимбионтного гриба Burkholderia Rhizoxinica (Lackner и др., 2011). Белки MBBBD включают модули примерно из 31-33 аминокислот, которые специфичны к основаниям. Последовательности этих модулей менее чем на 40% идентичны с общими повторами TALE у Xanthomonas, несмотря на то, что они презентуют большую вариабельность последовательностей полипептидов. Если они составлены вместе, такие модульные полипептиды могут нацеливать специфические последовательности нуклеиновых кислот чрезвычайно схожим образом с TALE-нуклеазами Xanthomonas. В предпочтительном варианте осуществления настоящего изобретения указанный ДНК-связывающий домен является сконструированным BBBD-связывающим доменом, включающим от 10 до 30 модулей, предпочтительно от 16 до 20 модулей. Разные домены от указанных выше белков (модули, N- и С-концы) из Burkholderia и Xanthomonas применимы для конструирования новых белков или стволов, обладающих связывающими свойствами в отношении специфических последовательностей нуклеиновых кислот. В частности примерами, не ограничивающими рамок охвата настоящего изобретения, являются дополнительные N-концевые и С-концевые домены сконструированных MBBBD, которые могут быть производными от природных TALE-подобных AvrBs3, PthXol, AvrHahl, PthA, Tal1c.
Понятие «TALE-нуклеаза» или «MBBBD-нуклеаза» относятся к сконструированным белкам, образуемым в результате слияния ДНК-связывающего домена, обычно производного от эффекторных белков, подобных активатору транскрипции (Transcription Activator Like Effector - TALE), или MBBBD-связывающего домена, с эндонуклеазным каталитическим доменом. Такой каталитический домен предпочтительно является нуклеазным доменом и более предпочтительно доменом, имеющим эндонуклеазную активность, например как у I-TevI, ColE7, NucA и Fok-I. В предпочтительном варианте осуществления настоящего изобретения указанная нуклеаза является мономерной TALE-нуклеазой или MBBBD-нуклеазой. Мономерная нуклеаза отличается тем, что не требует димеризации для специфического распознавания и расщепления, например гибриды сконструированного ДНК-связывающего домена с каталитическим доменом I-TevI, описанным в WO 2012138927. В другом предпочтительном варианте осуществления настоящего изобретения указанная редкощепящая эндонуклеаза является димерной TALE-нуклеазой или MBBBD-нуклеазой, предпочтительно включающая ДНК-связывающий домен, гибридизированный с FokI. TALE-нуклеаза уже была описана и применялась для стимуляции нацеливания гена и генных модификаций. Такие сконструированные TALE-нуклеазы коммерчески доступны под торговой маркой TALEN™ (Cellectis, 8 rue de la Croix Jarry, 75013 Paris, France).
- Понятие «расщепление» относится к разрыву ковалентного каркаса полинуклеотида. Расщепление могут инициировать разными методами, включая, но ими не ограничиваясь, ферментативный или химический гидролиз фосфодиэфирной связи. Возможно как одноцепочечное, так и двухцепочечное расщепление, причем двухцепочечное расщепление может происходить в результате двух отдельных однонитевых расщеплений. Двухцепочечное расщепление ДНК, РНК или гибрида ДНК/РНК может привести к формированию тупых или смещенных концов.
- Понятие «гибридный белок» относится к продукту известного в данной области процесса, состоящего из соединения двух или более генов, которые исходно кодируют отдельные белки или их часть. Трансляция указанного «гибридного белка» приводит к образованию единого полипептида с функциональными свойствами, производными от каждого из исходных белков.
- Понятие «идентичность» относится к идентичности последовательностей двух молекул нуклеиновой кислоты или полипептидов. Идентичность может быть определена сравнением положения в каждой последовательности, которая может быть выровнена для сравнения. Если положение в сравниваемой последовательности занято тем же самым основанием, тогда молекулы идентичны по этому положению. Степень схожести или идентичности последовательностей нуклеиновых кислот или аминокислот является функцией ряда идентичных или сопряженных нуклеотидов в положениях, общих для последовательностей нуклеиновых кислот. Различные алгоритмы и/или программы выравнивания могут применяться для подсчета идентичности между двумя последовательностями, включая FASTA или BLAST, которые доступны в качестве части пакета анализа последовательности GCG (Висконсинский университет, Мэдисон, США) и могут применяться, например, при несоответствующем окружении. В настоящем изобретении рассматривают, например, полипептиды, обладающие, по меньшей мере 70%, 85%, 90%, 95%, 98% или 99% идентичностью по отношению к специфическим полипептидам, описанным в настоящем изобретении, и предпочтительно проявляющие в существенной степени те же функции, а также полинуклеотид, кодирующий такие полипептиды.
- Понятие «сигнал-трансдуцирующий домен» или «ко-стимулирующий лиганд» означает молекулу на антиген-презентирующей клетке, которая специфически связывает родственную ко-стимулирующую молекулу на Т-клетке, тем самым обеспечивая сигнал, который дополнительно к первичному сигналу обеспечивается, например, связыванием комплекса TCR/CD3 с молекулой МНС, нагруженной пептидом, опосредует ответ Т-клетки, включая, но ими не ограничиваясь, активирование пролиферации, дифференциации и др. К ко-стимулирующим лигандам относятся, но ими рамки охвата настоящего изобретения не ограничиваются, CD7, В7-1 (CD80), В7-2 (CD86), PD-L1, PD-L2, 4-1BBL, OX40L, индуцибельный костимулирующий лиганд (inducible costimulatory ligand - ICOS-L), межклеточную адгезивную молекулу ICAM, CD30L, CD40, CD70, CD83, HLA-G, MICA, M1CB, HVEM, рецептор лимфотоксина бета, 3/TR6, ILT3, ILT4, агонист или антитело, которое связывает лиганд Toll-рецептора и лиганд, который специфически связывает В7-Н3. К понятию «ко-стимулирующий лиганд» также относят наряду с другими антитело, которое специфически связывает ко-стимулирующую молекулу, присутствующую на Т-клетках, например, из числа молекул, которыми однако рамки охвата настоящего изобретения не ограничиваются, CD27, CD28, 4-IBB, ОХ40, CD30, CD40, PD-1, ICOS, функционально связанного антигена лимфоцитов 1 (lymphocyte function-associated antigen-1 - LFA-1), CD2, CD7, LTGHT, NKG2C, B7-H3, лиганда, который специфически связывает CD83.
Понятие «ко-стимулирующая молекула» относится к родственному связывающему партнеру на Т-клетке, который специфически связывает ко-стимулирующий лиганд, тем самым опосредуя ко-стимулирующий ответ клетки, например, пролиферацию, но не только ее. К ко-стимулирующим молекулам относятся, но ими перечень не ограничивается, молекула МНС класса I, BTLA и Toll-лигандный рецептор.
Понятие «ко-стимулирующий сигнал» в контексте настоящего изобретения относится к сигналу, который в комбинации с первичным сигналом, например, TCR/CD3 лигированием, приводит к Т-клеточной пролиферации и/или повышенной или пониженной регуляции ключевых молекул.
- Понятие «биспецифическое антитело» относится к антителу, которое имеет сайты связывания для двух разных антигенов в одной молекуле антитела. Специалистам в данной области следует учитывать, что другие молекулы дополнительно к канонической структуре антитела могут быть сконструированы с двойной связывающей специфичностью. Также следует учитывать, что связывание антигена биспецифическими антителами может быть одновременным или последовательным. Биспецифические антитела могут быть получены химическими методами (см., например, Kranz и др., 1981), методами «polydoma» (US 4474893) или методами рекомбинации ДНК, суть которых известна. Каждый связывающий домен включает по меньшей мере одну вариабельную область из тяжелой цепи антитела («VH или Н область»), причем VH область первого связывающего домена специфически связывается с лимфоцитарным маркером, например, CD3, и VH область второго связывающего домена специфически связывается с опухолевым антигеном; данный пример не ограничивает области охвата настоящего изобретения.
- Понятие «внеклеточный лиганд-связывающий домен» в контексте настоящего изобретения относится к олиго- или полипептиду, который способен связывать лиганд. Предпочтительно домен может взаимодействовать с молекулой на поверхности клетки. Например, внеклеточный лиганд-связывающий домен может быть выбран для распознавания лиганда, который действует как маркер на поверхности клеток, ассоциированный с определенной стадией заболевания. Таким образом, к маркерам на поверхности клетки, которые могут действовать в качестве лигандов, относятся те, которые ассоциированы с вирусным, бактериальным или паразитарным заражением, аутоиммунным заболеванием и раковыми клетками.
Понятие «субъект» или «пациент» в контексте настоящего изобретения относится ко всем представителям царства животных, включая обезьян и людей.
Изложенное выше описание настоящего изобретения представляет способ и процесс получения и применения, которые изложены таким образом, что любой специалист в данной области сможет осуществить и применить настоящее изобретение, подробно изложенное в настоящем описании, особенно в формуле изобретения, которая представляет часть настоящего описания.
Если в настоящем изобретении указывают численный предел или диапазон, то конечные или ограничительные точки включены и указаны. Кроме того, все значения и поддиапазоны в пределах количественных ограничений или диапазона специально включены, как если бы они были выписаны.
Приведенное выше описание представлено для того, чтобы специалист в данной области мог осуществить и использовать настоящее изобретение, поясненное конкретными примерами. Различные модификации вариантов осуществления настоящего изобретения будут очевидны специалистам в данной области техники, и общие принципы, изложенные в настоящем изобретении, могут быть применены к другим вариантам осуществления настоящего изобретения, не выходящим за пределы сущности и объема настоящего изобретения. Таким образом, настоящее изобретение не ограничивается приведенными примерами, и может применяться широко в соответствии с описанными в нем нормами и свойствами.
Приведенные ниже конкретные примеры поясняют изложенное описание, служат его иллюстрацией и никоим образом не ограничивают рамок охвата настоящего изобретения.
Примеры
Пример 1. TALE-нуклеазы. расщепляющие ген GR человека
Шесть гетеродимерных TALE-нуклеаз, которые нацеливаются на экзоны гена GR человека, были разработаны и получены. Приведенная ниже табл. 2 представляет последовательности-мишени, расщепленные каждой из TALE-нуклеаз. GR TALE-нуклеаза составлена из двух независимых элементов (называемых полу-ТАLЕ-нуклеазами), каждый из которых содержит повторяющуюся последовательность, сконструированную для связи и расщепления последовательностей-мишеней GR, состоящих из двух последовательностей длиной 17 п.о. (называемых полумишенями), разделенных спейсером из 15 п.о.
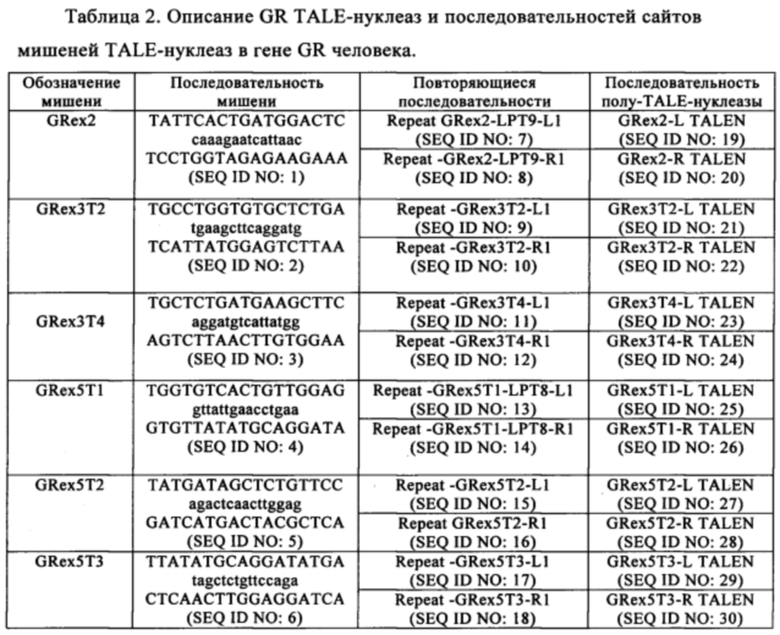
Аминокислотные последовательности N-концевого и С-концевого доменов и повтор основаны на AvrBs3 TALE (номер в GenBank: Х16130.1). N-концевой и С-концевой домены разделены двумя сайтами рестрикции BsmBI. Повторные матрицы (с SEQ ID NO: 7 по SEQ ID NO: 18), нацеливаемые на требуемые последовательности (с SEQ ID NO: 1 по SEQ ID NO: 6), были синтезированы, используя метод жесткой основы, состоящий из последовательных стадий рестрикции/лигирования/промывания (WO 2013/017950). Вкратце, первый блок (кодирование di-повтора) иммобилизуют на жесткой основе путем взаимодействия биотина/стрептавидина, второй блок (tri-повтор) затем лигируют с первым и после расщепления SfaNI присоединяют третий блок (tri-повтор). Процесс повторяют, используя блоки tri- или di-повторов для получения требуемой матрицы повторов. Затем продукт клонируют в классической плазмиде pAPG10 для амплификации в Е. coli и секвенируют. Последовательности полученных таким образом матриц повторов субклонируют в дрожжевом векторе экспрессии TALE, используя ферменты рестрикции BsmBI типа IIS для получения плазмиды и BbvI и SfaNI для инсерции последовательностей повторов. ДНК, кодирующую полу-ТАLЕ-нуклеазу, которая содержит TALE-производный ДНК-связывающий домен, гибридизированный с каталитическим доменом фермента рестрикции FokI, амплифицируют в Е. coli, восстанавливают стандартными микрометодами и секвенируют для оценки целостности вставки.
Активность GR TALE-нуклеаз в дрожжах
Нуклеазную активность шести GR-TALE-нуклеаз тестируют при 37°С и 30°С методом yeast SSA, ранее описанным авторами настоящего изобретения (WO 2004/067736; Epinat и др., 2003; Chames и др., 2005; Arnould и др., 2006; Smith и др., 2006), на мишенях, содержащих две TALE целевые последовательности, расположенные навстречу друг к другу на цепи ДНК и разделенные спейсером из 15 п.о., в результате чего образуются последовательности с SEQ ID NO: 1 по SEQ ID NO: 6. Все дрожжевые целевые репортерные плазмиды, содержащие целевые последовательности ДНК TALE-нуклеаз, конструируют согласно ранее описанному (WO 2004/067736; Epinat и др., 2003; Chames и др., 2005; Arnould и др., 2006; Smith и др., 2006). В дрожжах уровни расщепляющей активности TALE-нуклеаз отдельных клонов на мишенях представлены в табл. 3.
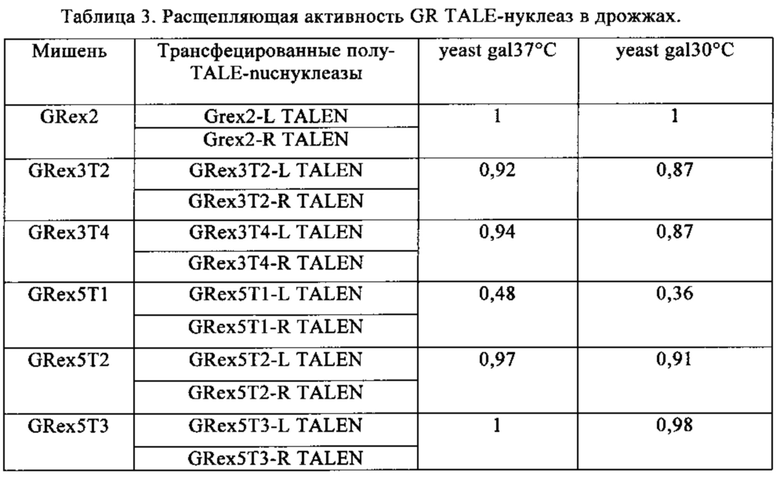
Величины находятся в диапазоне от 0 до 1. Максимальная величина равна 1.
Активность GR TALE-нуклеаз в клетках НЕK293
Каждую конструкцию TALE-нуклеазы субклонируют, используя фермент рестрикции в составе экспрессирующего вектора млекопитающих под контролем длинного промотора pEF1 alpha.
Один миллион клеток НЕK293 высевают за день до трансфекции. Клетки подвергают совместной трансфекции 2,5 мкг каждой из двух плазмид, кодирующих левую и правую половину GRex2, GRex3T2, GRex3T4, GRex5Tl, GRex5T2 или GRex5T3 TALE-нуклеазы, распознающей две полумишени целевых геномных последовательностей в гене GR под контролем промотора EF1alpha, используя 25 мкл липофектамина (фирма Invitrogen) по инструкциям производителя. В качестве контроля клетки подвергают совместной трансфекции 2,5 мкг каждой из двух плазмид, кодирующих левую и правую половину TALE-нуклеаз, нацеливающихся на сайт-мишень области константной цепи Т-клеточного рецептора альфа (TRAC_T01) ((TRAC_T01-L и -R TALE-нуклеаза (SEQ ID NO: 41 и SEQ ID NO: 42, TRAC_T01 сайт-мишень (SEQ ID NO: 37)) под контролем промотора EF1alpha. Двухцепочечный разрыв, произведенный TALE-нуклеазами в кодирующей последовательности GR, индуцирует негомологичное соединение концов (non homologous end joining - NHEJ), которое является механизмом, допускающим ошибки. Активность TALE-нуклеаз измеряют по частоте инсерций или делеций в геномном локусе нацеливания.
На 2 или 7 сутки после трансфекции клетки собирают и осуществляют локус-специфичную ПЦР по отношению к экстрагированной геномной ДНК, используя следующие праймеры: 5'-CCATCTCATCCCTGCGTGTCTCCGACTCAG-3' (форвардная адаптерная последовательность)-10N (TAG)- локус-специфичная форвардная последовательность для GR экзона 2: 5'-GGTTCATTTAACAAGCTGCC-3' (SEQ ID NO: 31), для GR экзона 3: 5'-GCATTCTGACTATGAAGTGA-3' (SEQ ID NO: 32) и для GR экзона 5: 5'-TCAGCAGGCCACTACAGGAGTCTCACAAG-3' (SEQ ID NO: 33) и реверсный праймер 5'-CCTATCCCCTGTGTGCCTTGGCAGTCTCAG-3' (реверсная адаптерная последовательность)-локус-специфичная реверсная последовательность для GR экзона 2: 5'-AGCCAGTGAGGGTGAAGACG-3' (SEQ ID NO: 34), для GR экзона 3: 5'-GGGCTTTGCATATAATGGAA-3' (SEQ ID NO: 35) и для GR экзона 5: 5'-CTGACTCTCCCCTTCATAGTCCCCAGAAC-3' (SEQ ID NO: 36).
Продукты ПЦР секвенируют с помощью системы секвенирования 454 (454 Life Sciences). Примерно 10000 последовательностей получают в качестве продукта ПЦР и затем анализируют на наличие сайт-специфической инсерции или делеции. Табл. 4 показывает процент последовательностей с инсерциями или делециями в целевом сайте TALE-нуклеазы от общего числа последовательностей в образце. Результаты репрезентативного эксперимента перечислены в табл. 4 для GRex2, GRex3T2 и GRex3T4.
Во всех исследованных случаях процент мутагенеза близок на 7 сутки к такому же показателю в образце на 2 сутки после трансфекции. Природу мутаций также анализируют, выявляя большинство делеций во всех случаях сравнения с инсерциями.
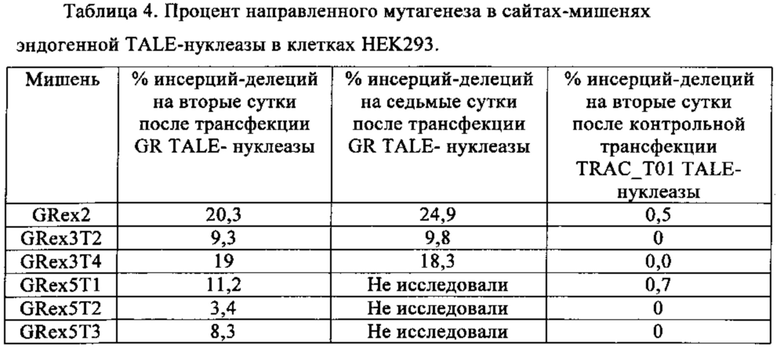
Активность GR TALE-нуклеаз в первичных Т-лимфоцитах
Каждую конструкцию TALE-нуклеазы субклонируют, используя фермент рестрикции в векторе экспрессии под контролем промотора Т7.
Молекулу мРНК, кодирующую TALE-нуклеазы, которые расщепляют GR геномные последовательности, синтезируют с каждой плазмиды, несущей кодирующие последовательности вниз по цепи от промотора Т7. Т-лимфоциты, выделенные из периферической крови, активируют в течение 5 суток, используя анти-CD3/CD28 активаторные гранулы (фирма Life technologies), и 5 миллионов клеток трансфецируют путем электропорации 10 мкг каждой из двух мРНК, кодирующих обе полу-TALE-нуклеазы, используя прибор CytoLVT-P (фирма BTX-Harvard apparatus). Т-клетки, трансфецированные 10 мкг каждой из двух мРНК, кодирующих обе полу-TALE-нуклеазы, нацеливающиеся на ген CD52 (CD52_T02-L и -R TALEN (SEQ ID NO: 55 и SEQ ID NO: 56), целевая последовательность CD52_T02 SEQ ID NO: 40) применяют в качестве контроля.
Через 3 и 7 суток после трансфекции геномную ДНК выделяют из трансфецированных клеток и проводят локус-специфическую ПЦР, используя описанные выше праймеры. Продукты ПЦР секвенируют с помощью системы секвенирования 454 (454 Life Sciences). Примерно 10000 последовательностей получают в расчете на ПЦР продукт и затем анализируют на наличие сайт-специфической инсерции или делеции; результаты показаны в табл. 5.
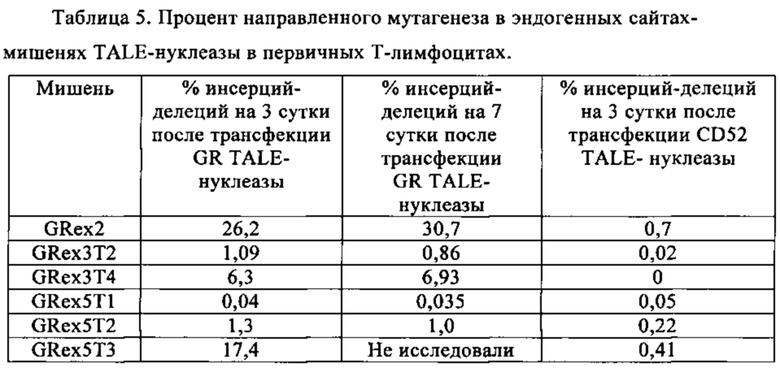
Пример 2. Расщепление TALE-нуклеазами гена человека CD52. константной цепи рецептора альфа Т-клеток человека (T-cell receptor alpha constant chain - TRAC) и константных цепей 1 и 2 рецептора бета Т-клеток (Т-cell receptor beta constant chains - TRBC)
Согласно описанию в примере 1, конструируют и получают гетеродимерные TALE-нуклеазы, нацеливающиеся соответствующим образом на гены CD52, TRAC и TRBC. Целевые геномные последовательности состоят из двух последовательностей длиной 17 п.о. (называемых полумишенями), разделенных спейсером из 11 или 15 п.о. Каждую полумишень распознают по повторам полу-TALE-нуклеаз, перечисленных в табл. 6. Геном человека содержит две функциональные бета цепи рецептора Т-клеток (TRBC1 и TRBC2). На протяжении развития альфа/бета Т-лимфоцитов одну из этих двух константных цепей выбирают в каждой клетке для сплайсирования с вариабельной областью TCR-beta и формируют функциональную полной длины цепь бета. Две TRBC мишени выбирают из последовательностей, сохраненных между TRBC1 и TRBC2 таким образом, что соответствующая TALE-нуклеаза может расщеплять обе цепи, TRBC1 и TRBC2, одновременно.
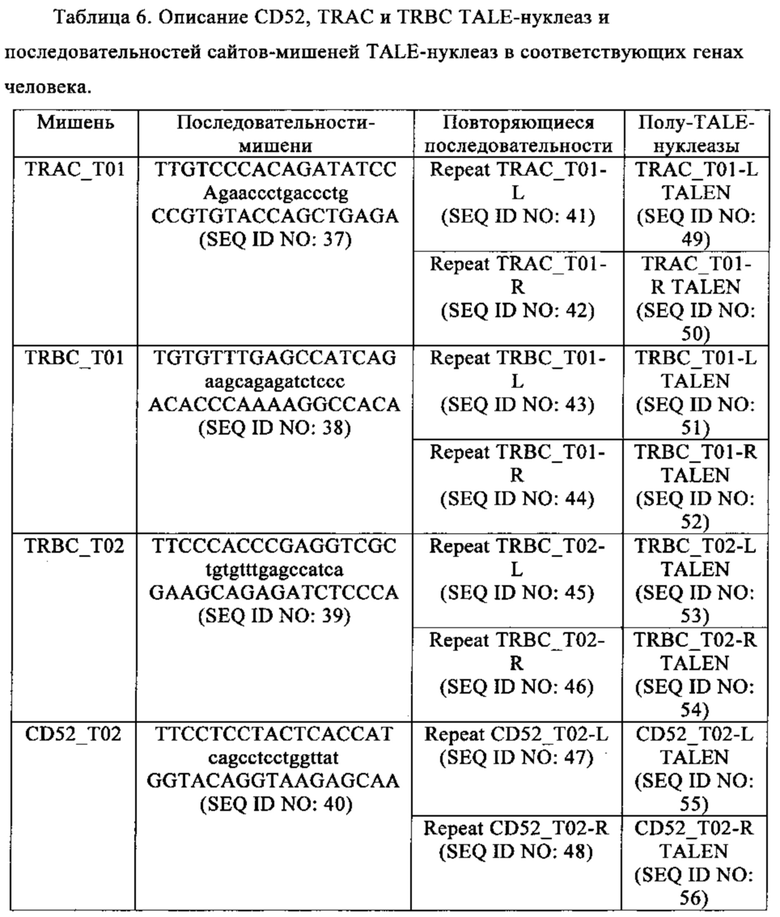
Сконструированы другие последовательности мишеней в генах TRAC и CD52, представленные в табл. 7.
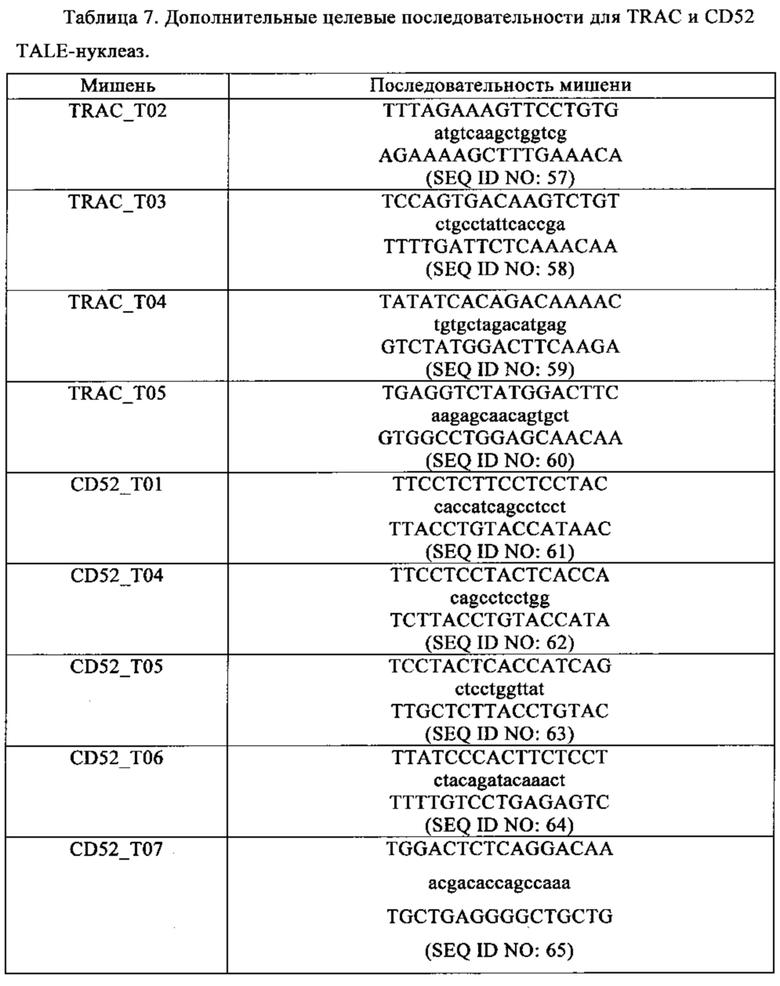
Активность CD52-TALE-нуклеазы, TRAC-TALE- нуклеазы и TRBC-TALE-нуклеазы в клетках НЕK293
Каждую конструкцию TALE-нуклеазы субклонируют, используя фермент рестрикции в составе экспрессирующего вектора млекопитающих под контролем pEF1alpha длинного промотора. Один миллион клеток НЕK293 высевают за день до трансфекции. Клетки подвергают совместной трансфекции 2,5 мкг каждой из двух плазмид, кодирующих TALE-нуклеазы, распознающие две полумишени в целевой геномной последовательности в гене CD52, область константной цепи рецептора альфа Т-клеток (TRAC) или область константной цепи рецептора бета Т-клеток (TRBC) под контролем промотора EF1-alpha, или 5 мкг контрольного вектора pUC (pCLS0003), используя 25 мкл липофектамина (фирма Invitrogen) по инструкциям производителя. Двухцепочечное расщепление, полученной под действием TALE-нуклеаз в CD52 или TRAC кодирующих последовательностях, репарируют в живых клетках путем негомологичного соединения концов (non homologous end joining - NHEJ), которое является механизмом, допускающим ошибки. Активность TALE-нуклеаз в живых клетках измеряют по частоте инсерций или делеций в геномном локусе нацеливания. Через 48 ч после трансфекции геномную ДНК выделяют из трансфецированных клеток и локус-специфическую ПЦР осуществляют, используя следующие праймеры: 5'-CCATCTCATCCCTGCGTGTCTCCGACTCAG (форвардная адаптерная последовательность)-10N (TAG)-локус-специфичная форвардная последовательность для CD52: 5'-CAGATCTGCAGAAAGGAAGC-3' (SEQ ID NO: 66), для TRAC: 5'-ATCACTGGCATCTGGACTCCA-3' (SEQ ID NO: 67), для TRBC1: 5'-AGAGCCCCTACCAGAACCAGAC-3' (SEQ ID NO: 68) или для TRBC2: 5'- GGACCTAGTAACATAATTGTGC-3' (SEQ ID NO: 69), и реверсный праймер 5'-CCTATCCCCTGTGTGCCTTGGCAGTCTCAG (реверсная адаптерная последовательность) - эндогенная локус-специфичная реверсная последовательность для CD52: 5'-CCTGTTGGAGTCCATCTGCTG-3' (SEQ ID NO: 70), для TRAC: 5'-CCTCATGTCTAGCACAGTTT-3' (SEQ ID NO: 71), для TRBC1 и TRBC2: 5'-ACCAGCTCAGCTCCACGTGGT-3' (SEQ ID NO: 72). Продукты ПЦР секвенируют с помощью системы секвенирования 454 (454 Life Sciences). Примерно 10000 последовательностей получают в расчете на ПЦР продукт и затем анализируют на наличие сайт-специфической инсерции или делеции; результаты показаны в табл. 8.
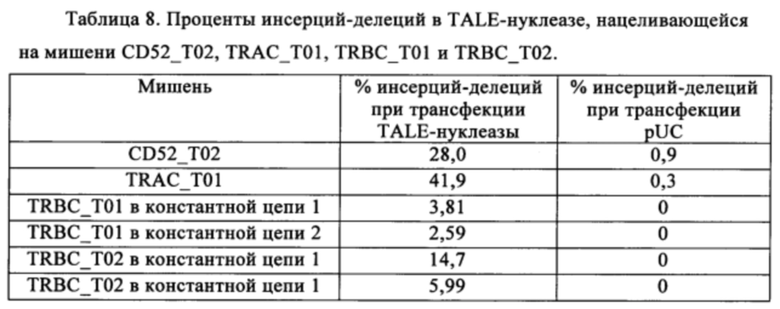
Активность CD52-TALE-нуклеазы. TRBC-TALE-нуклеазы и TRAC-TALE-нуклеазы в первичных Т-лимфоцитах
Каждую конструкцию TALE-нуклеазы субклонируют, используя фермент рестрикции в векторе экспрессии млекопитающих под контролем промотора Т7.
Молекулу мРНК, кодирующую TALE-нуклеазу, которая расщепляет CD52 TRAC и TRBC геномную последовательность, синтезируют с плазмиды, несущей кодирующие последовательности вниз по цепи от промотора Т7. Т-лимфоциты, выделенные из периферической крови, активируют в течение 5 суток, используя анти-CD3/CD28 активаторные гранулы (фирма Life technologies), и затем 5 миллионов клеток трансфецируют путем электропорации с 10 мкг каждой из 2 мРНК, кодирующих обе полу-ТАLЕ-нуклеазы (или некодирующую РНК в качестве контроля), используя прибор CytoLVT-P. В результате инсерций и делеций, индуцированных NHEJ, кодирующая последовательность CD52 и/или TRAC может быть вне рамки во фракции клеток, в результате чего получают нефункционирующие гены. Через 5 суток после электропорации клетки метят флуорохром-конъюгированным анти-CD52 или анти-TCR антителом жидкостной цитометрией на наличие CD52 или TCR на поверхности клеток. Поскольку все Т-лимфоциты, размноженные из периферической крови, в норме экспрессируют CD52 и TCR, пропорция CD52-отрицательных или TCR-отрицательных клеток является прямым показателем активности TALE-нуклеазы. В табл. 9 перечислены результаты репрезентативного эксперимента. В табл. 10 представлены результаты репрезентативного эксперимента по тестированию эффективности TRBC TALE-нуклеаз.
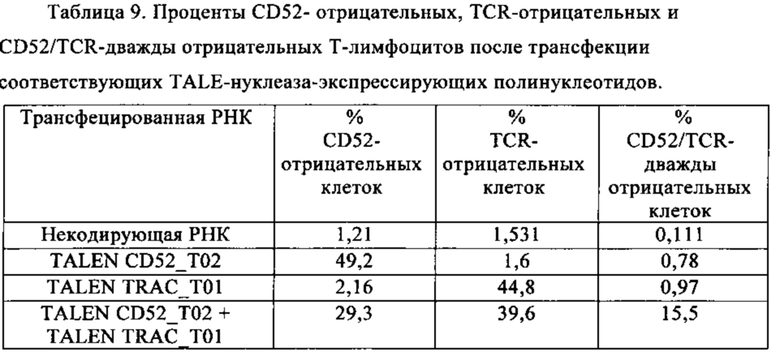

Функциональный анализ Т-клеток с целевым геном CD52
Целью инактивации гена CD52 является компенсация устойчивости Т-лимфоцитов к иммуносупрессии, опосредованной анти-CD52 антителом. Согласно описанному в предыдущем абзаце, в Т-лимфоциты трансфецируют мРНК, кодирующую TALE-нуклеазу, которая расщепляет CD52. Через 7 суток после трансфекции клетки обрабатывают 50 мкг/мл анти-CD52 моноклональным антителом (или IgG крысы в качестве контроля) с добавлением или без добавления 30% комплемента кролика (фирма Cedarlane). Через 2 ч инкубирования при 37°С клетки метят флуорохром-конъюгированным анти-CD52 антителом вместе с флуоресцентным прижизненным красителем (фирма eBioscience) и анализируют с помощью жидкостной цитометрии для измерения частоты CD52-положительных и CD52-отрицательных клеток среди живых клеток. Фиг. 6 показывает результат репрезентативного эксперимента, демонстрирующего, что CD52-отрицательные клетки полностью устойчивы к токсичности комплемент-опосредованного анти-CD52 антитела.
Функциональный анализ Т-клеток с целевым геном TRAC
Целью инактивации гена TRAC является компенсация неспособности Т-лимфоцитов стимулировать рецептор Т-клеток. Согласно описанному в предыдущем разделе, Т-лимфоциты трансфецируют мРНК, кодирующей TALE-нуклеазу, которая расщепляет TRAC или CD52. Через 16 суток после трансфекции клетки обрабатывают фитогемагглютинином (phytohemagglutinin - РНА; фирма Sigma-Aldrich) в концентрации до 5 мкг/мл, митогеном Т-клеток, действующим через рецептор Т-клеток. Клетки с функциональным Т-клеточным рецептором могут увеличиться в размере после обработки РНА. Через трое суток инкубирования клетки метят флуорохром-конъюгированным анти-CD52 или анти-TCR антителом и анализируют жидкостной цитометрией для сравнения распределения размера клеток между TCR-положительными и TCR-отрицательными клетками, или между CD52-положительными и CD52-отрицательными клетками. Фиг. 7 показывает, что TCR-положительные клетки существенно больше в размере после обработки РНА, тогда как TCR-отрицательные клетки имеют тот же размер, что и необработанные клетки, что свидетельствует о том, что инактивирование TRAC компенсирует неспособность реагировать на TCR-передачу сигнала. Напротив, CD52-положительные и CD52-отрицательные клетки увеличиваются в размере в той же степени.
Функциональный анализ Т-клеток с целевыми CD52 и TRAC генами
Для того, чтобы убедиться, что инженерия генома не влияет на способность Т-клеток проявлять противоопухолевую активность при обеспечении химерным антигенным рецептором (CAR), трансфецируют Т-клетки, на которые была нацелена CD52-TALE-нуклеаза и TRAC-TALE-нуклеаза с 10 мкг РНК, кодирующей анти-CD19 CAR (SEQ ID NO: 73). Через 24 ч Т-клетки инкубируют в течение 4 ч с CD19-экспрессирующими клетками Дауди. Повышение регуляции на поверхности клеток CD107a - маркера высвобождения цитотоксических гранул Т-лимфоцитами (процесс называют дегрануляцией), измеряют методом жидкостной цитометрии (Betts, Brenchley и др., 2003). Результаты, представленные на фиг. 8, показывают, что CD52-отрицательные/TCRαβ-отрицательные клетки и CD52-положительные/TCRαβ-положительные клетки имеют ту же способность к дегрануляции в ответ на РМА /иономицин (положительный контроль) или CD19+ клетки Дауди. Повышенная регуляция CD107 зависит от наличия CD19+. Эти данные показывают, что конструирование генома не оказывает негативного воздействия на способность Т-клеток повышать контролируемый противоопухолевый ответ.
Геномная безопасность CD52-TALE-нуклеазы и TRAC-TALE-нуклеазы в первичных Т-лимфоцитах
Поскольку конструкции в настоящем изобретении включают нуклеазные субъединицы, важно установить, приводит ли множественная TALE-нуклеазная трансфекция к генотоксичности и расщеплению вне мишени в «близко прилежащих» целевых последовательностях или около ошибочного спаривания полу-TALE-нуклеаз. Чтобы оценить воздействие TRAC-TALE-нуклеазы и CD52-TALE-нуклеазы на целостность клеточного генома, в настоящем изобретении перечислены последовательности в геноме человека, которые обладают потенциалом для расщепления вне сайта. Для создания такого перечня идентифицируют все последовательности в геноме с числом замещений, доходящих до 4, при сравнении с исходными полумишенями, и затем идентифицируют пары потенциальных полумишеней с ориентацией голова-к-голове со спейсером друг от друга от 9 до 30 п.о. Этот анализ включает сайты, на которые потенциально нацелены гомодимеры молекулы полу-TALE-нуклеазы или гетеродимеры, сформированные одной CD52 полу-TALE-нуклеазой и одной TRAC полу-TALE-нуклеазой. Проводят оценку потенциала мишеней вне сайта, основываясь на специфических данных, принимая во внимание цену отдельных замещений и положение замещений (где несоответствия более устойчивы для оснований с 3' конца полумишени). Получены 173 уникальные последовательности с оценкой, отражающей вероятность расщепления. Отобраны 15 последовательностей с наивысшими оценками и проанализирована методом глубокого секвенирования частота мутаций, обнаруженных по этому локусу в Т-клетках, одновременно трансфецированных CD52 и TRAC TALE-нуклеазой, очищенных магнитным сепарированием и разделенных на CD52-отрицательные и TCRαβ-отрицательные. Результаты представлены на фиг. 9. Наибольшая частота инсерций/делеций составляет 7×10-4. Эти результаты делают мнимую мишень вне сайта по меньшей мере в 600 раз менее вероятной для формирования мутаций чем намеченные мишени. Таким образом, TALE-нуклеазные реагенты, используемые в этом исследовании, проявляют чрезвычайно высокую специфичность.
Пример 3. TALE-нуклеазы, расщепляющие ген человека CTLA4 и ген человека PDCD1.
В примере 1 показано, что гетеродимерные TALE-нуклеазы, нацеливающиеся соответствующим образом на гены PDCD1 и CTLA4, конструируют и нарабатывают. Целевые геномные последовательности, состоящие из двух последовательностей длиной 17 п.о. (называемые полумишенями), разделены спейсерами длиной 11 или 15 п.о. Каждую полумишень распознают с помощью повторов полу-ТАLЕ-нуклеаз, перечисленных в табл. 11.
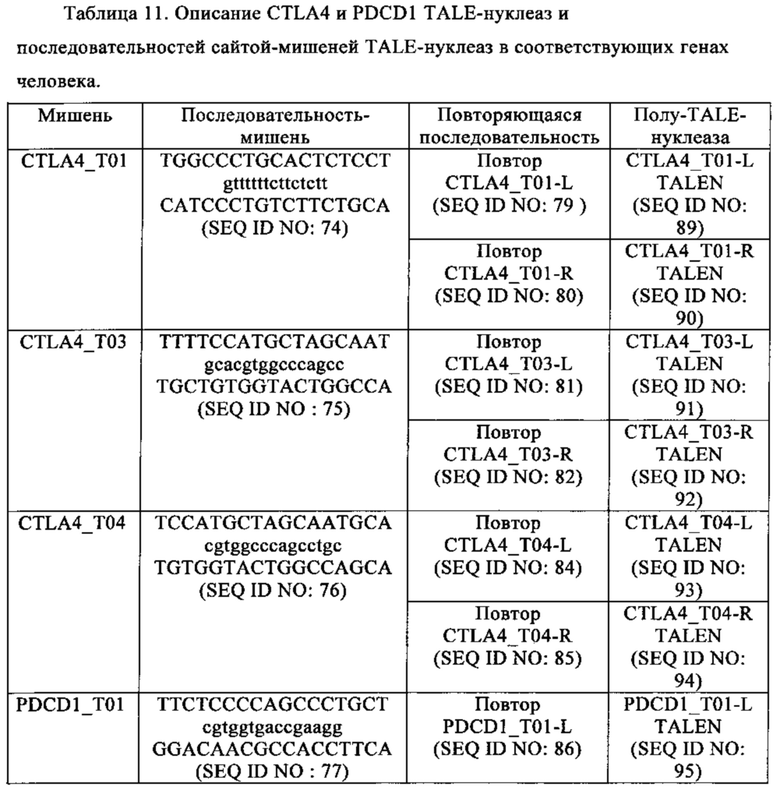

Активность CTLA4-TALE-нуклеазы и PDCD1-TALE-нуклеазы в клетках НЕK293
Каждую TALE-нуклеазную конструкцию субклонируют, используя фермент рестрикции в составе экспрессирующего вектора млекопитающих под контролем длинного промотора pEF1alpha. Миллион НЕK293 высевают за сутки до трансфекции. Клетки трансфецируют совместно 2,5 мкг каждой из двух плазмид, кодирующих TALE-нуклеазы, распознающие две полумишени в целевой геномной последовательности в генах PDCD1 и CTLA-4 под контролем промотора EF1-alpha, и 5 мкг контрольного вектора pUC (pCLS0003), используя 25 мкл липофектамина (фирма Invitrogen) по инструкциям производителя. Двухцепочечное расщепление, которое производят TALE-нуклеазы в кодирующих последовательностях PDCD1 или CTLA-4, репарируется в живых клетках путем негомологичного соединения концов (non homologous end joining - NHEJ), которое является механизмом, допускающим ошибки. Активность TALE-нуклеаз в живых клетках измеряют по частоте инсерций или делеций в целевом геномном локусе. Через 48 ч после трансфекции ДНК выделяют из трансфецированных клеток и осуществляют локус-специфичные ПЦР, используя следующие праймеры: 5'-CCATCTCATCCCTGCGTGTCTCCGACTCAG (форвардная адаптерная последовательность)- 10N (TAG)- локус-специфичная форвардная последовательность для CTLA4_T01: 5'-CTCTACTTCCTGAAGACCTG-3' (SEQ ID NO: 99), для CTLA4_T03/T04: 5'-ACAGTTGAGAGATGGAGGGG-3' (SEQ ID NO: 100), для PDCD1_T01: 5'-CCACAGAGGTAGGTGCCGC-3' (SEQ ID NO: 101) или для PDCD1_T03: 5'-GACAGAGATGCCGGTCACCA-3' (SEQ ID NO: 102) и реверсный праймер 5'-CCTATCCCCTGTGTGCCTTGGCAGTCTCAG (реверсная адаптерная последовательность)- - эндогенная локус-специфичная реверсная последовательность для CTLA4_T01: 5'-TGGAATACAGAGCCAGCCAA-3' (SEQ ID NO: 103), для CTLA4_T03/T04: 5'-GGTGCCCGTGCAGATGGAAT-3' (SEQ ID NO: 104), для PDCD1_T01: 5'-GGCTCTGCAGTGGAGGCCAG-3' (SEQ ID NO: 105) или для PDCD1_T03: 5'-GGACAACGCCACCTTCACCT-3' (SEQ ID NO: 106).
ПЦР продукты анализируют с помощью Т7-эндонуклеазы: вкратце, после денатурации и повторного отжига ПЦР продукта Т7-эндонуклеаза может специфически расщеплять несоответствующую ДНК, состоящую из цепей дикого типа и мутантных. Продукт расщепления затем разделяют электрофорезом в полиакриламидном геле. Наличие расщепляющегося продукта является показателем мутантных последовательностей, индуцированных действием TALE-нуклеазы. Результаты показаны на фиг. 10, где стрелки указывают на продукты расщепления ПЦР. Из результатов следует, что PDCD1_T1, PDCD1_T3, CTLA4_T1, CTLA4_T3 и CTLA4_T4 TALE-нуклеазы все проявляют мутагенную нуклеазную активность в соответствующих сайтах-мишенях.
CTLA4 инактивация в первичных Т-клетках
Первичные Т-клетки человека инактивируют гранулами CD3/28. Через 5 суток проводят электропорацию 5×106 клеток 20 мкг РНК, кодирующей одну из трех TALEN™ (T1, Т2 и Т3), сконструированных для гена CTLA4, или без РНК в качестве контроля. Через 3 суток после электропорации CTLA4 экспрессию измеряют путем внутриклеточного окрашивания, используя флуоресцентное антитело и метод жидкостной цитометрии (фиг. 27 и 28).
Все три TALEN™ индуцируют снижение регуляции экспрессии CTLA4, что коррелирует с их эффективностью в линиях клеток НЕK293 (Т1 более эффективен, чем Т3 и Т4).
Секвенирование геномной ДНК, выделенной из трансфецированных клеток с помощью технологии 454 (фирма Roche), выявил, что 96% аллелей CTLA4 мутированы в TALEN T1-обработанных клетках по сравнению с 0,1% в контрольном образце без TALEN.
Инактивация PD1 в первичных Т-клетках
Первичные Т-клетки человека активируют гранулами CD3/28. Через 5 суток проводят электропорацию 5×106 клеток 20 мкг РНК, кодирующей одну из двух последовательностей TALEN, специфичных в отношении гена человека PD1, или без РНК в качестве контроля. Через 10 суток клетки реактивируют и 3 суток после реактивации PD1 экспрессию измеряют окрашиванием поверхности, используя флуоресцентное антитело и метод жидкостной цитометрии (фиг. 29).
Обе TALEN индуцируют снижение регуляции экспрессии PD1.
Секвенирование геномной ДНК, выделенной из клеток, трансфецированных TALEN T1 и TALEN Т03 соответствующим образом, используя технологию 454 (фирма Roche), выявил, что 34% и 39% аллелей PD1 были соответствующим образом мутированы (результаты показаны на фиг. 30).
Повышенная противоопухолевая активность PD1-TALEN обработанных клеток:
Первичные Т-клетки человека активируют гранулами CD3/28. Через пять суток 5×106 клеток электропорируют 20 мкг РНК, кодирующей TALEN, специфичную в отношении гена человека PD1, или без РНК в качестве контроля. Через неделю клетки электропорируют мРНК, кодирующей химерный антигенный рецептор, специфичный в отношении CD19 человека, или без РНК в качестве отрицательного контроля. Через сутки их противоопухолевую активность измеряют в анализе клеточной цитотоксичности, используя CD19+ клетки Дауди (против K562 в качестве контроля) или клетки НСТ116, которые экспрессируют PD1 лиганд 1 (PDL1), трансдуцированный с вектором экспрессии CD19 (против исходных НСТ116 в качестве контроля). Цитотоксическую активность определяют путем сравнения жизнеспособности целевых клеток и контрольных клеток. Результаты показаны в диаграммах на фиг. 31. PD1 TALEN трансфекция восстанавливает цитотоксическую активность против PDL1-экспрессирующих клеток НСТ116 и улучшает цитотоксическую активность против клеток Дауди.
Пример 4. pTalpha допускает экспрессию CD3 на поверхности инактивированных TCR alpha Т-лимфоцитах:
Описание разных версий preTalpha:
Ген pTalpha человека кодирует трансмембранный гликопротеин, включающий внеклеточный Ig-подобный домен, гидрофобный трансмембранный домен и крупный С-концевой внутрицитоплазматический хвост. Разные версии, производные от pTalpha гликопротеина человека, были сконструированы, описаны в табл. 2 и представлены на фиг. 11.
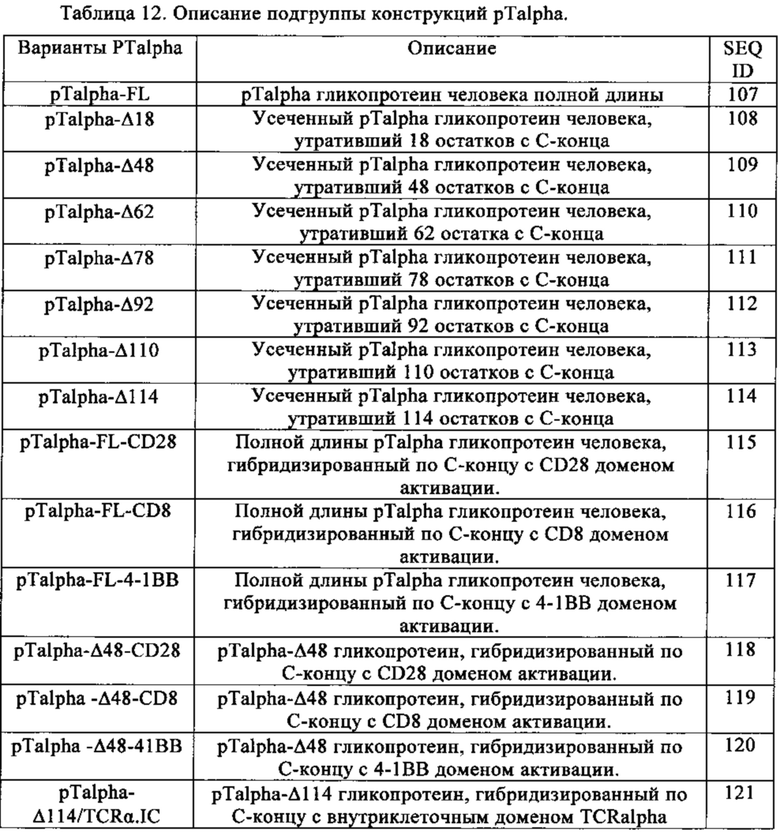

Разные исследованные preTalpha конструкции включают:
1. Мутанты pTalpha с делециями. Получают разные делеции во внутриклеточном цитоплазматическом хвосте белка человека pTalpha (который включает 114 аминокислот) (SEQ ID NO: 107). Исследуемые конструкции включали версию белка полной длины (FL) и мутанты, в которых аминокислоты 18, 48, 62, 78, 92, 110 и 114 были делетированы с С-конца белка (с SEQ ID NO: 108 по SEQ ID NO: 114).
2. Мутанты pTalpha. содержащие внутриклеточные домены активации.
Варианты FL и Δ48 гибридизируют с CD8, CD28 или 41ВВ внутриклеточными доменами активации по С-концам (с SEQ ID NO: 115 по SEQ ID NO: 120).
3. pTalpha/TCRα химерные мутанты: В одной из конструкций TCRα внутриклеточный домен (intracellular domain - IС) гибридизируют с бесхвостой версией (Δ114) pTalpha (SEQ ID NO: 121). Также получают вторую конструкцию, в которой внеклеточный домен pTalpha гибридизируют с трансмембранным (ТМ) и IС доменами от TCRα (SEQ ID NO: 122).
4. pTalpha мутанты димеризации: В литературе описаны мутации, способные изменять способность к олигомеризации/димеризации комплекса preTCR. Предполагают, что эти мутанты допускают экспрессию preTCR на поверхности Т-клеток без индукции конститутивной передачи сигнала (предположительно индукция происходит при олигомеризации preTCR). Мутации, интродуцированные в вариант pTalphaΔ48:
- 1× MUT: W46R (SEQ ID NO: 123)
- 4× MUT: D22A, K24А, R102A, R117A (SEQ ID NO: 124)
Активность разных preTalpha конструкций в TRAC-инактивированных клетках Jurkat.
Для скрининга различных вариантов pTalpha по способности восстанавливать экспрессию CD3 на поверхности TCRalpha-инактивированных клеток, получают линию клеток, в которой ген TCRalpha разорван с помощью нацеливания TALEN на TRAC. Клетки Jurkat (линия лейкозных Т-клеток) методом CytoPulse электропорации трансфецируют плазмидами, кодирующими TALEN, которая расщепляет TRAC, и клетки KО (TCRα/βNEG; CD3NEG) затем очищают методом отрицательной селекции, используя магнитные гранулы CD3. Популяцию клеток KО (JKT_KO×3 клетки) амплифицируют и используют для скрининга разных вариантов pTalpha. Скрининг проводят путем трансфекции одного миллиона JKT_KO×3 клеток 15 мкг плазмиды, кодирующей разные варианты pTalpha под контролем промотора EF1α, с последующим анализом жидкостной цитометрией экспрессии CD3 на поверхности клеток через 48 ч после трансфекции. На фиг. 12 показывают типичный пример эффективности трансфекции (% BFP + клеток) и активности FL, Δ18 и Δ48 pTalpha конструкций в JKT_KO×3 клетках на основании процента клеток CD3+, определенных жидкостной цитометрией. Результаты по разным конструкциям представлены по группам в табл. 13.
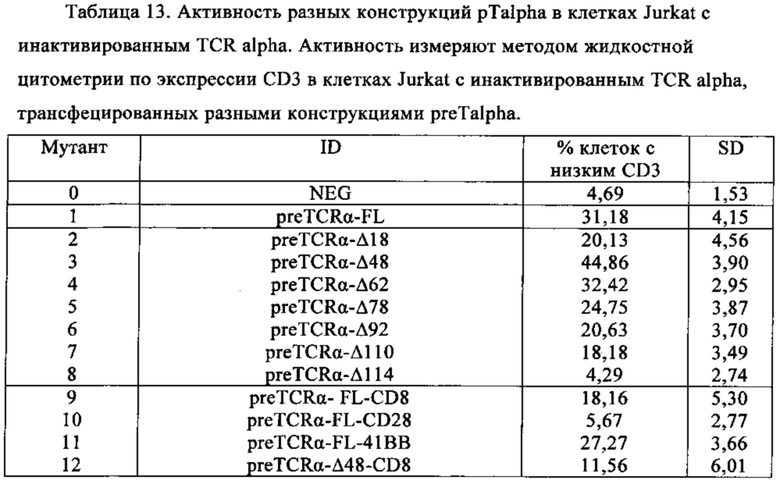
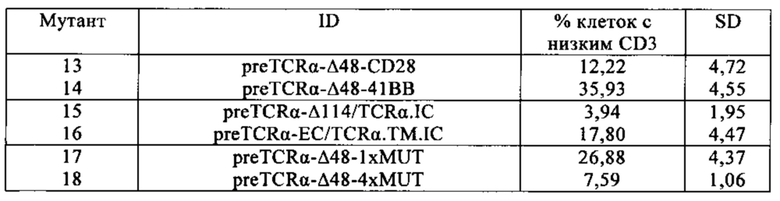
Активность pTalpha-FL и pTalpha-Δ48 в первичных Т-лимфоцитах с инактивированным TCR alpha
Для тестирования способности версий pTalpha-FL и pTalpha-Δ48 индуцировать экспрессию CD3 на поверхности Т-лимфоцитов с инактивированным TCR alpha, pTalpha-FL и pTalpha-Δ48 кодирующие последовательности клонируют в самоинактивирующем лентивирусном векторе pLV-SFFV-BFP-2A-PCTRA, который кодирует синий флуоресцентный белок (Blue Fluorescent protein - BFP) под контролем промотора SFFV и следующий за ним саморасщепляющий пептид Т2А (фиг. 13).
Т-лимфоциты, выделенные из периферической крови, активируют в течение 72 ч, используя анти-CD3/CD28 активирующие гранулы (фирма Life technologies), и 4,5 миллиона клеток подвергают трансфекции путем электропорации с помощью прибора CytoLVT-S (фирма BTX-Harvard Harbour) 10 мкг мРНК, кодирующей TALE-нуклеазу, которая нацеливается на область константной цепи TCR alpha (TRAC). Через 2 суток после электропорации Т-клетки трансдуцируют или LV-SFFV-BFP-2A- pTalpha-Δ48, или LV-SFFV-BFP-2А-контрольными лентивирусными векторами. CD3 отрицательные Т-клетки и Т-клетки с низким содержанием CD3 затем очищают, используя анти-CD3 магнитные гранулы (фирма Miltenyi Biotech). Протокол этого эксперимента представлен на фиг. 14А.
На фиг. 14Б представлен анализ методом жидкостной цитометрии TCRalpha/beta, экспрессию на поверхности клеток CD3 и экспрессию BFP на TCRalpha-инактивированных Т-клетках (KО), трансдуцированных BFP-2A-pTalphaΔ48 (KО/Δ48) или контрольным BFP лентивирусным вектором (KO/BFP) до и после очистки гранулами CD3. TCRalpha-инактивированные клетки, трансдуцированные BFP-T2A-pTalpha-Δ48 вектором (BFP+клетки), показывают повышенные уровни CD3 по сравнению с уровнями нетрансдуцированных клеток (BFP- клетки). Нет отличий среди клеток, трансдуцированных контрольным вектором BFP. Эти результаты показывают, что pTalpha опосредует восстановление экспрессии CD3 на поверхности TCRalpha инактивированных клеток. Напротив, TCRalpha/beta окрашивание сохраняет, как и ожидалось, неизменными клетки, трансдуцированные или не трансдуцированные вектором экспрессии pTalpha-Δ48.
pTalpha-опосредованная CD3 экспрессия поддерживает активацию TCR-недостаточных Т-клеток
Для определения способности pTalpha трансдуцировать сигналы активации клеток, исследуют сигналы маркеров ранней и поздней активации на TCR alpha - инактивированных Т-клетках, трансдуцированных pTalpha-Δ48 и pTalpha-Δ48.41ВВ. Т-клетки с инактивированным TCR alpha, трансдуцированные pTalpha-Δ48 и pTalpha-Δ48.41BB, получают из первичных Т-клеток человека согласно описанному в предыдущем разделе и на фиг. 14А.
Для обнаружения передачи сигнала через CD3 клетки реактивируют, используя гранулы с анти-CD3/CD28 покрытием через 3 суток после очистки TCR alpha инактивированных Т-клеток гранулами CD3 (фиг. 14А). Клетки окрашивают конъюгированными с флюорохромом анти-CD69 (маркер ранней активации) и анти-CD25 (маркер поздней активации) через 24 и 48 ч после реактивации, соответственно, и анализируют методом жидкостной цитометрии (фиг. 15А, Б). На фиг. 15А и Б показано, что TCR alpha-инактивированные клетки, экспрессирующие pTalpha-Δ48 (KО/pTα-Δ48) или pTalpha-Δ48.41BB (KО/pTα-Δ48.ВВ), проявляют повышенную регуляцию маркеров активации до уровней, схожих с уровнями, наблюдаемыми в TCRalpha/beta экспрессирующих клетках (клетки, не подвергшиеся электропорации; NEP - non electroporated).
Другим индикатором активации Т-клеток является увеличение размера клеток, которое иногда называют «губительным». Способность комплексов preTCR индуцировать «губительный» размер измеряют методом жидкостной цитометрии, определяя размер клеток через 72 ч после реактивации, используя анти-CD3/CD28- гранулы (фиг. 15В). Стимуляция анти-CD3/CD28 гранулами индуцировала сопоставимые повышения размера тех клеток, которые экспрессируют комплексы TCRalpha/beta, при сравнении с клетками, экспрессирующими pTalpha-Δ48 или pTalpha-Δ48.41BB. Анализируемые вместе, эти результаты означают, что комплексы preTCR компетентны для трансдукции сигналов, которые эффективно сочетаются с механизмами, опосредующими повышение регуляции маркеров активации.
pTalpha опосредованная CD3 экспрессия поддерживает размножение TCR-недостаточных первичных Т-клеток, используя стимулирующие анти-CD3/CD28 антитела
Для оценки способности комплексов preTCR поддерживать долгосрочную пролиферацию клеток, измеряют пролиферацию клеток, выработанных согласно описанному ранее. Через 10 суток после первоначальной активации клетки поддерживают в IL2 (нет реактивации) или в IL2 с анти-CD3/CD28 гранулами (реактивация). Для каждого состояния клетки подсчитывают и анализируют методом жидкостной цитометрии в разные моменты времени для оценки числа клеток BFP+. Рост TCRalpha инактивированных клеток (KО), трансдуцированных BFP или BFP-T2A-preTCRα-Δ48 векторами, сравнивают и оценивают кратную индукцию этих клеток относительно величины, полученной на 2 сутки после реактивации. На фиг. 16 показывают результаты, полученные от двух независимых доноров. В обоих случаях TCRalpha инактивированные клетки, экспрессирующие pTalpha-Δ48, обладают повышенным размножением по сравнению с TCRalpha инактивированными клетками, экспрессирующими только BFP контрольный вектор. Для второго донора TCRalpha инактивированные клетки, экспрессирующие pTalpha-Δ48.41BB или pTalpha полной длины, также включены и также проявляют повышенное размножение по сравнению с TCRalpha инактивированными клетками, экспрессирующими только BFP контрольный вектор.
Пример 5. Оптимизация трансфекции мРНК в Т-клетки. используя метод Cytopulse
Определение оптимизированной программы Cytopulse
Первую серию экспериментов проводят на неактивированных мононуклеарных клетках периферической крови (МКПК) для того, чтобы определить диапазон напряжения, при котором клетки могут быть трансфецированы. Пять протестированных разных программ представлены в табл. 14.
Электропорируют 3 или 6 миллионов клеток в кювете для электропорации с просветом 0,4 см (30 или 15×106 клеток/мл) с 20 мкг плазмид, кодирующих GFP, и контрольные плазмид pUC, используя разные программы Cytopulse. Через 24 ч после электропорации исследуют экспрессию GFP в электропорированных клетках методом жидкостной цитометрии для определения эффективности трансфекции. Полученные данные на фиг. 17 показывают минимальное напряжение, требуемое для электропорации плазмиды в МКПК производные Т-клетки. Эти результаты свидетельствуют, что программы cytopulse 3 и 4 допускают эффективную трансформацию Т-клеток (ЕР#3 и #4).
Электропорации мРНК в очищенные активированные Т-клетки
После определения наилучшей программы Cytopulse, которая позволяет осуществить эффективную электропорацию ДНК в Т-клетки, устанавливают применимость настоящего метода для электропорации мРНК.
5×106 очищенных Т-клеток, предварительно активированных 6 суток с PHA/IL2, ресуспендируют в буфере Т для цитопорации (фирма BTX-Harvard apparatus) и электропорируют в 0,4 см кюветах с 10 мкг мРНК, кодирующей GFP или 20 мкг плазмид, кодирующих GFP или pUC, используя предпочтительную программу цитопульса, определенную в предыдущем разделе (табл. 14).

Через 48 ч после трансфекции клетки окрашивают прижизненным красителем (eFluor-450) и определяют выживаемость клеток и процент жизнеспособных GFP+ клеток методом жидкостной цитометрии (фиг. 18).
Представленные на фиг. 18 результаты показывают, что электропорация РНК в оптимальных условиях, определенных в настоящем изобретении, не является токсичной и позволяет осуществить трансфекцию более чем 95% жизнеспособных клеток.
Полученные в совокупности результаты показывают, что Т-клетки могут быть эффективно трансфецированы как ДНК, так и РНК. В частности трансфекция РНК не влияет на выживаемость клеток и позволяет единообразить уровни экспрессии трансфецированных целевых генов в популяции клеток.
Эффективная трансфекция может быть достигнута раньше после клеточной активации независимо от применяемого метода активирования (PHA/IL-2 или CD3/CD28-покрытые-гранулы). В настоящем изобретении достигли эффективности в трансфекции клеток >95% через 72 ч после активирования. Кроме того, также может быть эффективной трансфекция Т-клеток после оттаивания и активирования по тому же протоколу электропорации.
Электропорация мРНК в первичные Т-клетки человека для функциональной экспрессии TALE-нуклеазы
После того, как было установлено, что электропорация мРНК приводит к эффективной экспрессии GFP в первичных Т-клетках человека, исследовали применимость настоящего способа для экспрессии других целевых белков. Эффекторные нуклеазы, подобные активаторам транскрипции (TALE-нуклеазы), являются сайт-специфичными нуклеазами, образованными в результате гибридизации TAL ДНК-связывающего домена с ДНК-расщепляющим доменом. Они представляют мощные инструменты редактирования генома, поскольку индуцируют двухцепочечные разрывы практически в любой последовательности ДНК, в которой это требуется. Такие двухцепочечные разрывы активируют соединение негомологичных концов (Non-homologous end-joining - NHEJ), подверженный ошибкам механизм репарации ДНК, потенциально приводящий к инактивации какого-либо требуемого целевого гена. В другом варианте, если адекватная репарирующая матрица одновременно интродуцирована в клетки, индуцированные TALE-нуклеазой разрывы ДНК могут быть репарированы путем гомологичной рекомбинации, вследствие этого открывая возможность модификации требуемой последовательности.
В настоящем описании применяют электропорацию мРНК для экспрессии TALE-нуклеазы, сконструированной для специфического расщепления последовательности в гене человека, кодирующем цепь альфа антигенного рецептора Т-клеток (T-cell antigen receptor - TRAC). Предполагают, что мутации, индуцированные в этой последовательности, приводят к инактивации гена и утрате комплекса TCRαβ с поверхности клетки. РНК TRAC TALE-нуклеазы или некодирующую РНК в качестве контроля трансфецируют в активированные первичные Т-лимфоциты человека, используя метод Cytopulse. Последовательность электропорации описана в табл. 15: она включает 2 импульса 1200В и затем 4 импульса 130В.
С помощью анализа методом жидкостной цитометрии экспрессии TRC на поверхности клеток через 7 суток после электропорации (фиг. 19, верхняя панель) установлено, что 44% Т-клеток утратили экспрессию TCRαβ. Исследование геномной ДНК трансфецированных клеток методом ПЦР амплификации локуса TRAC с помощью системы высокопродуктивного секвенирования 454. 33% секвенированных аллелей (727 из 2153) содержат инсерции или делеции в сайте расщепления TALE-нуклеазы. Фиг. 19 (нижняя панель) показывает примеры мутантных аллелей.
Полученные данные показывают, что электропорация мРНК методом Cytopulse приводит к функциональной экспрессии TRAC TALE-нуклеазы.
Электропорация Т-клеток моноцистронной мРНК, кодирующей анти-СР19 одноцепочечный химерный антигенный рецептор (CAR)
5×106 Т-клетки, предварительно активированные за несколько суток (за 3-5 суток) гранулами с анти-CD3/CD28 покрытием и IL2, ресуспендируют в буфере Т для цитопорации и проводят электропорацию в 0,4 см кюветах без мРНК или с 10 мкг мРНК, кодирующей одноцепочечный (SEQ ID NO: 73), используя программу, представленную в табл. 15.
Через 24 ч после электропорации клетки окрашивают фиксирующим прижизненным красителем eFluor-780 и РЕ-конъюгированным козьим антимышиным IgG F(ab')2 фрагментом, специфичным для оценки экспрессии CAR на поверхности живых клеток. Полученные данные на фиг. 20. А показывают, что огромное большинство живых Т-клеток, электропорированных описанной ранее моноцистронной мРНК, экспрессируют на своей поверхности CAR. Через 24 ч после электропорации Т-клетки культивируют совместно с клетками Дауди (CD19+) в течение 6 ч и анализируют методом жидкостной цитометрии для обнаружения экспрессии маркера дегранулирования CD107a на своей поверхности (Betts, Brenchley и др., 2003).
Представленные на фиг. 20 данные показывают, что большинство клеток, электропорированных описанной ранее моноцистронной мРНК, дегранулирует в присутствии целевых клеток, экспрессирующих CD19. Эти результаты четко показывают, что экспрессируемый на поверхности электропорированных Т-клеток CAR активен.
Электропорация Т-клеток полицистронной мРНК. кодирующей анти-CD 19 мультисубъединичный химерный антигенный рецептор (CAR)
5×106 Т-клеток, заранее активированных (за 3-5 суток) гранулами с покрытием анти-CD3/CD28 и IL2, подвергают электропорации в буфере Т для цитопорации и осуществляют электропорацию в 0,4 см кюветах без мРНК или с 45 мкг мРНК, кодирующей многоцепочечный CAR (SEQ ID NO: 125, кодируемая SEQ ID NO: 126, фиг. 21А и фиг. 4Б (csm4)), используя программу, представленную в табл. 15.
Через 24 ч после электропорации клетки окрашивают фиксирующим прижизненным красителем eFluor-780 и РЕ-конъюгированным козьим антимышиным IgG F(ab')2 фрагментом, специфичным для оценки экспрессии CAR на поверхности живых клетках. Представленные на фиг. 21 данные показывают, что огромное большинство живых Т-клеток, электропорированных ранее описанной полицистронной мРНК, экспрессируют CAR на поверхности.
Через 24 ч после электропорации Т-клетки культивируют совместно с клетками Даути (CD19+) в течение 6 ч и анализируют методом жидкостной цитометрии для обнаружения экспрессии на их поверхности маркера дегрануляции CD107. Представленные на фиг. 21 данные показывают, что большинство клеток, подверженных электропорации ранее описанной полицистронной мРНК, дегранулирует в присутствии целевых клеток, экспрессирующих CD19. Эти результаты четко показывают, что экспрессированный на поверхности подвергшихся электропорации Т-клеток CAR активен.
Литература
Arnould S., P. Chames, и др. «Engineering of large numbers of highly specific homing endonucleases that induce recombination on novel DNA targets.» J Mol Biol 355(3), 2006, cc. 443-458.
Betts M.R., J.M. Brenchley и др. «Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation.» J Immunol Methods 281(1-2), 2003, cc. 65-78.
Boch J., H. Scholze «Breaking the code of DNA binding specificity of TAL-type III effectors.» Science 326(5959), 2009, cc. 1509-1512.
Boni A., P. Muranski и др. «Adoptive transfer of allogeneic tumor-specific T cells mediates effective regression of large tumors across major histocompatibility barriers.» Blood 112(12), 2008, cc. 4746-4754.
Brahmer J.R., C.G. Drake и др. «Phase I study of single-agent anti-programmed death-1 (MDX-1106) in refractory solid tumors: safety, clinical activity, pharmacodynamics, and immunologic correlates.» J Clin Oncol 28(19), 2010, cc. 3167-3175.
Bucks С.М., J.A. Norton и др. «Chronic antigen stimulation alone is sufficient to drive CD8+ T cell exhaustion.» J Immunol 182(11), 2009, cc. 6697-6708.
Carrasco Y.R., A.R. Ramiro и др. «An endoplasmic reticulum retention function for the cytoplasmic tail of the human pre-T cell receptor (TCR) alpha chain: potential role in the regulation of cell surface pre-TCR expression levels.» J Exp Med 193(9), 2001, cc. 1045-1058.
Cermak Т., E.L. Doyle и др. «Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting.» Nucleic Acids Res 39(12), 2011, c. e82.
Chames P., J. C. Epinat и др. «In vivo selection of engineered homing endonucleases using double-strand break induced homologous recombination.» Nucleic Acids Res 33(20), 2005, c. e178.
Christian M., T. Cermak и др. «Targeting DNA double-strand breaks with TAL effector nucleases.» Genetics 186(2), 2010, cc. 757-761.
Cong L., F.A. Ran и др. «Multiplex genome engineering using CRISPR/Cas systems.» Science 339(6121), cc. 819-823.
Critchlow, S.E., S.P. Jackson и др. «DNA end-joining: from yeast to man.» Trends Biochem Sci 23(10), 1998, cc. 394-398.
Deltcheva E., K. Chylinski и др. «CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III.» Nature 471(7340), cc. 602-606.
Deng, D., C. Yan и др., (2012). "Structural basis for sequence-specific recognition of DNA by TAL effectors." Science 335(6069), 2012, cc. 720-723.
Dessarthe, В., A. Thedrez и др. «CRTAM receptor engagement by Necl-2 on tumor cells triggers cell death of activated Vgamma9Vdelta2 T cells.» J Immunol 190(9), 2013, cc. 4868-4876.
Epinat J. C., S. Arnould и др. «A novel engineered meganuclease induces homologous recombination in yeast and mammalian cells.» Nucleic Acids Res 31(11), 2003, cc. 2952-2962.
Garneau J.E., M.E. Dupuis и др. «The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA.» Nature 468(7320), 2010, cc. 67-71.
Gasiunas G., R. Barrangou и др. «Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria." Proc Natl Acad Sci USA 109(39), 2012, cc. E2579-2586.
Geissler R., H. Scholze и др. «Transcriptional activators of human genes with programmable DNA-specificity.» PLoS One 6(5), 2011, cc. e19509.
Grange M., G. Verdeil и др. «Active STAT5 regulates T-bet and eomesodermin expression in CD8 T cells and imprints a T-bet-dependent Tc1 program with repressed IL-6/TGF-beta1 signaling.» J Immunol 191(7), 2013, cc. 3712-3724.
Heinrich P.С., I. Behrmann и др. «Principles of interleukin (IL)-6-type cytokine signalling and its regulation.» Biochem J 374(часть 1), 2003, cc. 1-20.
Huang P., A. Xiao и др. «Heritable gene targeting in zebrafish using customized TALENs.» Nat Biotechnol 29(8), 2011, cc. 699-700.
Ikehara Y., S.K. Ikehara и др. «Negative regulation of T cell receptor signaling by Siglec-7 (p70/AIRM) and Siglec-9.» J Biol Chem 279(41), 2004, cc. 43117-43125.
Jena В., G. Dotti и др. «Redirecting T-cell specificity by introducing a tumor-specific chimeric antigen receptor.» Blood 116(7), 2010, cc. 1035-1044.
Jinek M., K. Chylinski и др. «A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity.» Science 337(6096), 2012, cc. 816-821.
Lackner G., N. Moebius и др. «Complete genome sequence of Burkholderia rhizoxinica, an Endosymbiont of Rhizopus microsporus.» J Bacteriol 193(3), 2011, cc. 783-784.
Li L., M. J. Piatek и др. «Rapid and highly efficient construction of TALE-based transcriptional regulators and nucleases for genome modification.» Plant Mol Biol 78(4-5), 2012, cc. 407-416.
Li Т., S. Huang и др. «Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes.» Nucleic Acids Res 39(14), 2011, cc. 6315-6325.
Lozano E., M. Dominguez-Villar и др. «The TIGIT/CD226 axis regulates human T cell function.» J Immunol 188(8), 2011, cc. 3869-3875.
Ma J.L., E.M. Kim и др. «Yeast Mre11 and Rad1 proteins define a Ku-independent mechanism to repair double-strand breaks lacking overlapping end sequences.» Mol Cell Biol 23(23), 2003, cc. 8820-8828.
Mahfouz M.M., L. Li и др. «Targeted transcriptional repression using a chimeric TALE-SRDX repressor protein.» Plant Mol Biol 78(3), 2012, cc. 311-321.
Mahfouz M.M., L. Li и др. «De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-strand breaks.» Proc Natl Acad Sci USA 108(6), 2011, cc. 2623-2628.
Мак A.N., P. Bradley и др. «The crystal structure of TAL effector PthXol bound to its DNA target.» Science 335(6069), 2012, cc. 716-719.
Mali P., L. Yang и др. «RNA-guided human genome engineering via Cas9." Science 339(6121), 2013, cc. 823-826.
Meyaard, L., G. J. Adema и др. «LAIR-1, a novel inhibitory receptor expressed on human mononuclear leukocytes.» Immunity 7(2), 1997, cc. 283-290.
Miller J.C., S. Tan и др. «A TALE nuclease architecture for efficient genome editing.» Nat Biotechnol 29(2), 2011, cc. 143-148.
Morbitzer R., P. Romer и др. «Regulation of selected genome loci using de novo-engineered transcription activator-like effector (TALE)-type transcription factors.» Proc Natl Acad Sci USA 107(50), 2011, cc. 21617-21622.
Moscou M.J., A. J. Bogdanove «A simple cipher governs DNA recognition by TAL effectors.» Science 326(5959), 2009, c. 1501.
Mussolino C., R. Morbitzer и др. «A novel TALE nuclease scaffold enables high genome editing activity in combination with low toxicity.» Nucleic Acids Res 39(21), 2011, cc. 9283-9293.
Nicoll G., J. Ni и др. «Identification and characterization of a novel siglec, siglec-7, expressed by human natural killer cells and monocytes.» J Biol Chem 274(48), 1999, cc. 34089-34095.
Pang S.S., R. Berry и др. «The structural basis for autonomous dimerization of the pre-T-cell antigen receptor.» Nature 467(7317), 2010, cc. 844-848.
Pardoll D., C. Drake «Immunotherapy earns its spot in the ranks of cancer therapy.» J Exp Med 209(2), 2012, cc. 201-209.
Pardoll D. M. «The blockade of immune checkpoints in cancer immunotherapy.» Nat Rev Cancer 12(4), 2012, cc. 252-264.
Park T.S., S.A. Rosenberg и др. «Treating cancer with genetically engineered T cells.» Trends Biotechnol 29(11), 2011, cc. 550-557.
Quigley M., F. Pereyra и др. «Transcriptional analysis of HIV-specific CD8+ T cells shows that PD-1 inhibits T cell function by upregulating BATF.» Nat Med 16(10), 2010, cc. 1147-1151.
Robert С., C. Mateus «Anti-CTLA-4 monoclonal antibody: a major step in the treatment of metastatic melanoma.» Med Sci (Paris) 27(10), 2011, cc. 850-858.
Rodriguez P.C., D.G. Quiceno и др. «L-arginine availability regulates T-lymphocyte cell-cycle progression.» Blood 109(4), 2007, cc. 1568-1573.
Sander J.D., L. Cade и др. «Targeted gene disruption in somatic zebrafish cells using engineered TALENs.» Nat Biotechnol 29(8), 2011, cc. 697-698.
Shin H., S.D. Blackburn и др. «A role for the transcriptional repressor Blimp-1 in CD8(+) T cell exhaustion during chronic viral infection.» Immunity 31(2), 2009, cc. 309-320.
Smith J., S. Grizot и др. «A combinatorial approach to create artificial homing endonucleases cleaving chosen sequences.» Nucleic Acids Res 34(22), 2006, c. el49.
Sorek, R., С.M. Lawrence и др. «CRISPR-mediated Adaptive Immune Systems in Bacteria and Archaea.» Annu Rev Biochem. 2013.
Stoddard B.L. «Homing endonuclease structure and function.» Q Rev Biophys 38(1), 2005, cc. 49-95.
Tesson L., C. Usal и др. «Knockout rats generated by embryo microinjection of TALENs.» Nat Biotechnol 29(8), 2011, cc. 695-696.
Thaventhiran J.E., A. Hoffmann и др. «Activation of the Hippo pathway by CTLA-4 regulates the expression of Blimp-1 in the CD8+ T cell.» Proc Natl Acad Sci USA 109(33), 2012, с. E2223-9.
Weber E., R. Gruetzner и др. «Assembly of designer TAL effectors by Golden Gate cloning.» PLoS One 6(5), 2011, c. e19722.
Wolchok, J.D., H. Kluger и др. «Nivolumab plus ipilimumab in advanced melanoma.» N Engl J Med 369(2), cc. 122-133.
Zhang F., L. Cong и др. «Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription.» Nat Biotechnol 29(2), 2011, cc. 149-153.
Zhang, J.Q., G. Nicoll и др. «Siglec-9, a novel sialic acid binding member of the immunoglobulin superfamily expressed broadly on human blood leukocytes.» J Biol Chem 275(29), 2000, cc. 22121-22126.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> СЕЛЛЕКТИС
<120> СПОСОБЫ КОНСТРУИРОВАНИЯ ВЫСОКО АКТИВНЫХ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ
<130> 435149WO (DI2013-31 PCT)
<160> 126
<170> PatentIn version 3.5
<210> 1
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> GRex2
<400> 1
tattcactga tggactccaa agaatcatta actcctggta gagaagaaa 49
<210> 2
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> GRex3T2
<400> 2
tgcctggtgt gctctgatga agcttcagga tgtcattatg gagtcttaa 49
<210> 3
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> GRex3T4
<400> 3
tgctctgatg aagcttcagg atgtcattat ggagtcttaa cttgtggaa 49
<210> 4
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> GRex5T1
<400> 4
tggtgtcact gttggaggtt attgaacctg aagtgttata tgcaggata 49
<210> 5
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> GRex5T2
<400> 5
tatgatagct ctgttccaga ctcaacttgg aggatcatga ctacgctca 49
<210> 6
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> GRex5T3
<400> 6
ttatatgcag gatatgatag ctctgttcca gactcaactt ggaggatca 49
<210> 7
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex2-LPT9-L1
<400> 7
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 8
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex2-LPT9-R1
<400> 8
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
340 345 350
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 9
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex3T2-L1
<400> 9
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
340 345 350
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 10
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex3T2-R1
<400> 10
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 11
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex3T4-L1
<400> 11
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 12
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex3T4-R1
<400> 12
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 13
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex5T1-LPT8-L1
<400> 13
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 14
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex5T1-LPT8-R1
<400> 14
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 15
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex5T2-L1
<400> 15
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 16
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex5T2-R1
<400> 16
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 17
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex5T3-L1
<400> 17
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 18
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор-GRex5T3-R1
<400> 18
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 19
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> GRex2-L TALEN
<400> 19
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 540
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 600
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 720
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 900
caggtggtgg ccatcgccag caatattggt ggcaagcagg cgctggagac ggtgcaggcg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1020
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaatgg cggtggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccccagcagg tggtggccat cgccagcaat aatggtggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1320
atcgccagca atattggtgg caagcaggcg ctggagacgg tgcaggcgct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caatggcggt 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 1740
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 1920
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 20
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> GRex2-R TALEN
<400> 20
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caatggcggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 720
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 840
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1020
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1140
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagccacga tggcggcaag 1260
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccccagcagg tggtggccat cgccagcaat ggcggtggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1440
atcgccagca atattggtgg caagcaggcg ctggagacgg tgcaggcgct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag ccacgatggc 1560
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1740
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1860
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 21
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> GRex3T2-L TALEN
<400> 21
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 600
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 720
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 900
caggtggtgg ccatcgccag caataatggt ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1020
agcaataatg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaatgg cggtggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccccagcagg tggtggccat cgccagcaat aatggtggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1320
atcgccagca atggcggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caataatggt 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 1740
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 1920
caggtggtgg ccatcgccag caataatggt ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 22
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> GRex3T2-R TALEN
<400> 22
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caatggcggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 660
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 720
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 840
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccggagca ggtggtggcc atcgccagca atattggtgg caagcaggcg 960
ctggagacgg tgcaggcgct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1020
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1140
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagccacga tggcggcaag 1260
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccggagcagg tggtggccat cgccagccac gatggcggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1440
atcgccagca atattggtgg caagcaggcg ctggagacgg tgcaggcgct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caatggcggt 1560
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 1680
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1740
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1860
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 23
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> GRex3T4-L TALEN
<400> 23
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 600
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 720
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 900
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1020
agcaataatg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagcaatat tggtggcaag 1140
caggcgctgg agacggtgca ggcgctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccccagcagg tggtggccat cgccagcaat ggcggtggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1320
atcgccagca ataatggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag caatattggt 1440
ggcaagcagg cgctggagac ggtgcaggcg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 1560
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 1740
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 1920
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 24
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> GRex3T4-R TALEN
<400> 24
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caatggcggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 720
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 840
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1020
caggtggtgg ccatcgccag caatattggt ggcaagcagg cgctggagac ggtgcaggcg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1140
agcaatattg gtggcaagca ggcgctggag acggtgcagg cgctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaataa tggtggcaag 1260
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccccagcagg tggtggccat cgccagcaat ggcggtggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1440
atcgccagca atggcggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag caatattggt 1560
ggcaagcagg cgctggagac ggtgcaggcg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 1680
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1740
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 1860
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 25
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> GRex5T1-L TALEN
<400> 25
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 600
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 720
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 900
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1020
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagcaatat tggtggcaag 1140
caggcgctgg agacggtgca ggcgctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccggagcagg tggtggccat cgccagccac gatggcggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1320
atcgccagca atggcggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caataatggt 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1740
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag caatattggt ggcaagcagg cgctggagac ggtgcaggcg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 26
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> GRex5T1-R TALEN
<400> 26
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccggag caggtggtgg ccatcgccag caatattggt 540
ggcaagcagg cgctggagac ggtgcaggcg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 720
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 840
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 1020
caggtggtgg ccatcgccag caataatggt ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1140
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagcaatat tggtggcaag 1260
caggcgctgg agacggtgca ggcgctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccccagcagg tggtggccat cgccagcaat ggcggtggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1440
atcgccagca atattggtgg caagcaggcg ctggagacgg tgcaggcgct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caatggcggt 1560
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 1680
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1740
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 1860
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccggagca ggtggtggcc atcgccagca atattggtgg caagcaggcg 1980
ctggagacgg tgcaggcgct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 27
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> GRex5T2-L TALEN
<400> 27
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 540
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 600
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 720
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccggagca ggtggtggcc atcgccagca atattggtgg caagcaggcg 840
ctggagacgg tgcaggcgct gttgccggtg ctgtgccagg cccacggctt gaccccccag 900
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1020
agcaatattg gtggcaagca ggcgctggag acggtgcagg cgctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaataa tggtggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccggagcagg tggtggccat cgccagccac gatggcggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1320
atcgccagca atggcggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag ccacgatggc 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1740
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 28
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> GRex5T2-R TALEN
<400> 28
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caataatggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 660
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 720
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 840
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 1020
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1140
agcaatattg gtggcaagca ggcgctggag acggtgcagg cgctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaataa tggtggcaag 1260
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccccagcagg tggtggccat cgccagcaat ggcggtggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1440
atcgccagcc acgatggcgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag caatattggt 1560
ggcaagcagg cgctggagac ggtgcaggcg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1740
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 1860
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 29
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> GRex5T3-L TALEN
<400> 29
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 600
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 720
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccggagca ggtggtggcc atcgccagca atattggtgg caagcaggcg 840
ctggagacgg tgcaggcgct gttgccggtg ctgtgccagg cccacggctt gaccccccag 900
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1020
agcaataatg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagccacga tggcggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccggagcagg tggtggccat cgccagcaat attggtggca agcaggcgct ggagacggtg 1260
caggcgctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1320
atcgccagca ataatggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caataatggt 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 1560
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 1740
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 1920
caggtggtgg ccatcgccag caataatggt ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 30
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> GRex5T3-R TALEN
<400> 30
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caataatggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 660
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 720
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 840
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 1020
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1140
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagccacga tggcggcaag 1260
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccggagcagg tggtggccat cgccagcaat attggtggca agcaggcgct ggagacggtg 1380
caggcgctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1440
atcgccagca atattggtgg caagcaggcg ctggagacgg tgcaggcgct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caataatggt 1560
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1740
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1860
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccggagca ggtggtggcc atcgccagca atattggtgg caagcaggcg 1980
ctggagacgg tgcaggcgct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 31
<211> 60
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер GR экзон 2
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 31
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn ggttcattta acaagctgcc 60
<210> 32
<211> 60
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер GR экзон 3
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 32
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn gcattctgac tatgaagtga 60
<210> 33
<211> 69
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер GR экзон 5
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 33
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn tcagcaggcc actacaggag 60
tctcacaag 69
<210> 34
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер GR экзон 2
<400> 34
cctatcccct gtgtgccttg gcagtctcag agccagtgag ggtgaagacg 50
<210> 35
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер GR экзон 3
<400> 35
cctatcccct gtgtgccttg gcagtctcag gggctttgca tataatggaa 50
<210> 36
<211> 59
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер GR экзон 5
<400> 36
cctatcccct gtgtgccttg gcagtctcag ctgactctcc ccttcatagt ccccagaac 59
<210> 37
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> TRAC_T01
<400> 37
ttgtcccaca gatatccaga accctgaccc tgccgtgtac cagctgaga 49
<210> 38
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> TRBC_T01
<400> 38
tgtgtttgag ccatcagaag cagagatctc ccacacccaa aaggccaca 49
<210> 39
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> TRBC_T02
<400> 39
ttcccacccg aggtcgctgt gtttgagcca tcagaagcag agatctccca 50
<210> 40
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> CD52_T02
<400> 40
ttcctcctac tcaccatcag cctcctggtt atggtacagg taagagcaa 49
<210> 41
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор TRAC_T01-L
<400> 41
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 42
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор TRAC_T01-R
<400> 42
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 43
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор TRBC_T01-L
<400> 43
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
340 345 350
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 44
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор TRBC_T01-R
<400> 44
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 45
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор TRBC_T02-L
<400> 45
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 46
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор TRBC_T02-R
<400> 46
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 47
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор CD52_T02-L
<400> 47
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
340 345 350
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 48
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор CD52_T02-R
<400> 48
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 49
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> TRAC_T01-L TALEN
<400> 49
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 600
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 720
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 900
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1020
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagcaatat tggtggcaag 1140
caggcgctgg agacggtgca ggcgctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccggagcagg tggtggccat cgccagccac gatggcggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1320
atcgccagca atattggtgg caagcaggcg ctggagacgg tgcaggcgct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caataatggt 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 1560
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 1740
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 50
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> TRAC_T01-R TALEN
<400> 50
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccggag caggtggtgg ccatcgccag ccacgatggc 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 720
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 840
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1020
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1140
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaataa tggtggcaag 1260
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccccagcagg tggtggccat cgccagcaat aatggtggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1440
atcgccagca atggcggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag caatattggt 1560
ggcaagcagg cgctggagac ggtgcaggcg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1740
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 1860
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 51
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> TRBC_T01-L TALEN
<400> 51
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 600
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 720
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 900
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1020
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaataa tggtggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccggagcagg tggtggccat cgccagcaat attggtggca agcaggcgct ggagacggtg 1260
caggcgctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1320
atcgccagca ataatggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag ccacgatggc 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1620
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1740
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag caatattggt ggcaagcagg cgctggagac ggtgcaggcg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 52
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> TRBC_T01-R TALEN
<400> 52
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caataatggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 720
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 840
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1020
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1140
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaatgg cggtggcaag 1260
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccccagcagg tggtggccat cgccagcaat ggcggtggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1440
atcgccagca atggcggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caataatggt 1560
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1740
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1860
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 53
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> TRBC_T02-L TALEN
<400> 53
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 600
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 720
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccggagca ggtggtggcc atcgccagca atattggtgg caagcaggcg 840
ctggagacgg tgcaggcgct gttgccggtg ctgtgccagg cccacggctt gaccccggag 900
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1020
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagccacga tggcggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccccagcagg tggtggccat cgccagcaat aatggtggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1320
atcgccagca atattggtgg caagcaggcg ctggagacgg tgcaggcgct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caataatggt 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 1740
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 54
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> TRBC_T02-R TALEN
<400> 54
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caataatggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 720
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 840
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1020
caggtggtgg ccatcgccag caatattggt ggcaagcagg cgctggagac ggtgcaggcg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1140
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagccacga tggcggcaag 1260
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccccagcagg tggtggccat cgccagcaat ggcggtggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1440
atcgccagcc acgatggcgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caatggcggt 1560
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1740
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1860
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 55
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> CD52_T02-L TALEN
<400> 55
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 600
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 720
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 900
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1020
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaatgg cggtggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccggagcagg tggtggccat cgccagcaat attggtggca agcaggcgct ggagacggtg 1260
caggcgctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1320
atcgccagcc acgatggcgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caatggcggt 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1620
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 1740
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag caatattggt ggcaagcagg cgctggagac ggtgcaggcg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 56
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> CD52_T02-R TALEN
<400> 56
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caatggcggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 720
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 840
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 1020
caggtggtgg ccatcgccag caatggcggt ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1140
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagcaatat tggtggcaag 1260
caggcgctgg agacggtgca ggcgctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccggagcagg tggtggccat cgccagccac gatggcggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1440
atcgccagcc acgatggcgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caatggcggt 1560
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1740
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 1860
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 57
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> TRAC_T02
<400> 57
tttagaaagt tcctgtgatg tcaagctggt cgagaaaagc tttgaaaca 49
<210> 58
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> TRAC_T03
<400> 58
tccagtgaca agtctgtctg cctattcacc gattttgatt ctcaaacaa 49
<210> 59
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> TRAC_T04
<400> 59
tatatcacag acaaaactgt gctagacatg aggtctatgg acttcaaga 49
<210> 60
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> TRAC_T05
<400> 60
tgaggtctat ggacttcaag agcaacagtg ctgtggcctg gagcaacaa 49
<210> 61
<211> 47
<212> ДНК
<213> Искусственная
<220>
<223> CD52_T01
<400> 61
ttcctcttcc tcctaccacc atcagcctcc tttacctgta ccataac 47
<210> 62
<211> 43
<212> ДНК
<213> Искусственная
<220>
<223> CD52_T04
<400> 62
ttcctcctac tcaccacagc ctcctggtct tacctgtacc ata 43
<210> 63
<211> 43
<212> ДНК
<213> Искусственная
<220>
<223> CD52_T05
<400> 63
tcctactcac catcagctcc tggttatttg ctcttacctg tac 43
<210> 64
<211> 47
<212> ДНК
<213> Искусственная
<220>
<223> CD52_T06
<400> 64
ttatcccact tctcctctac agatacaaac tttttgtcct gagagtc 47
<210> 65
<211> 47
<212> ДНК
<213> Искусственная
<220>
<223> CD52_T07
<400> 65
tggactctca ggacaaacga caccagccaa atgctgaggg gctgctg 47
<210> 66
<211> 60
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер CD52
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 66
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn cagatctgca gaaaggaagc 60
<210> 67
<211> 61
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер TRAC
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 67
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn atcactggca tctggactcc 60
a 61
<210> 68
<211> 62
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер TRBC1
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 68
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn agagccccta ccagaaccag 60
ac 62
<210> 69
<211> 62
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер TRBC2
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 69
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn ggacctagta acataattgt 60
gc 62
<210> 70
<211> 51
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер GR CD52
<400> 70
cctatcccct gtgtgccttg gcagtctcag cctgttggag tccatctgct g 51
<210> 71
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер GR TRAC
<400> 71
cctatcccct gtgtgccttg gcagtctcag cctcatgtct agcacagttt 50
<210> 72
<211> 51
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер GR TRBC1-2
<400> 72
cctatcccct gtgtgccttg gcagtctcag accagctcag ctccacgtgg t 51
<210> 73
<211> 495
<212> БЕЛОК
<213> Искусственная
<220>
<223> анти-CD19 CAR
<400> 73
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Ile
20 25 30
Lys Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr
35 40 45
Phe Thr Ser Tyr Val Met His Trp Val Lys Gln Lys Pro Gly Gln Gly
50 55 60
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Tyr Asn Asp Gly Thr Lys Tyr
65 70 75 80
Asn Glu Lys Phe Lys Gly Lys Ala Thr Leu Thr Ser Asp Lys Ser Ser
85 90 95
Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Gly Thr Tyr Tyr Tyr Gly Ser Arg Val Phe
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly
130 135 140
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Met
145 150 155 160
Thr Gln Ala Ala Pro Ser Ile Pro Val Thr Pro Gly Glu Ser Val Ser
165 170 175
Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu Asn Ser Asn Gly Asn Thr
180 185 190
Tyr Leu Tyr Trp Phe Leu Gln Arg Pro Gly Gln Ser Pro Gln Leu Leu
195 200 205
Ile Tyr Arg Met Ser Asn Leu Ala Ser Gly Val Pro Asp Arg Phe Ser
210 215 220
Gly Ser Gly Ser Gly Thr Ala Phe Thr Leu Arg Ile Ser Arg Val Glu
225 230 235 240
Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln His Leu Glu Tyr Pro
245 250 255
Phe Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Ser Asp Pro
260 265 270
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
275 280 285
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
290 295 300
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile
305 310 315 320
Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val
325 330 335
Ile Thr Leu Tyr Cys Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe
340 345 350
Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly
355 360 365
Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg
370 375 380
Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln
385 390 395 400
Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp
405 410 415
Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro
420 425 430
Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp
435 440 445
Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg
450 455 460
Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr
465 470 475 480
Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490 495
<210> 74
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> CTLA4_T01
<400> 74
tggccctgca ctctcctgtt ttttcttctc ttcatccctg tcttctgca 49
<210> 75
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> CTLA4_T03
<400> 75
ttttccatgc tagcaatgca cgtggcccag cctgctgtgg tactggcca 49
<210> 76
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> CTLA4_T04
<400> 76
tccatgctag caatgcacgt ggcccagcct gctgtggtac tggccagca 49
<210> 77
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> PDCD1_T01
<400> 77
ttctccccag ccctgctcgt ggtgaccgaa ggggacaacg ccaccttca 49
<210> 78
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> PDCD1_T03
<400> 78
tacctctgtg gggccatctc cctggccccc aaggcgcaga tcaaagaga 49
<210> 79
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор CTLA4_T01-L
<400> 79
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 80
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор CTLA4_T01-R
<400> 80
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 81
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор CTLA4_T03-L
<400> 81
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 82
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор CTLA4_T03-R
<400> 82
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 83
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор CTLA4_T04-L
<400> 83
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 84
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор CTLA4_T04-R
<400> 84
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 85
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор PDCD1_T01-L
<400> 85
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
100 105 110
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
165 170 175
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
195 200 205
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
340 345 350
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
370 375 380
Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
435 440 445
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 86
<211> 529
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор PDCD1_T01-R
<400> 86
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
65 70 75 80
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Lys Gln
275 280 285
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
290 295 300
Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly
305 310 315 320
Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln
325 330 335
Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly
340 345 350
Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu
355 360 365
Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser
370 375 380
Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro
385 390 395 400
Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile
405 410 415
Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu
420 425 430
Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val
435 440 445
Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
450 455 460
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
465 470 475 480
Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr
485 490 495
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
500 505 510
Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala Leu
515 520 525
Glu
<210> 87
<211> 530
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор PDCD1_T03-L
<400> 87
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
130 135 140
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Asn Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
420 425 430
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
435 440 445
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
450 455 460
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
465 470 475 480
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
485 490 495
Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
500 505 510
Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala
515 520 525
Leu Glu
530
<210> 88
<211> 529
<212> БЕЛОК
<213> Искусственная
<220>
<223> Повтор PDCD1_T03-R
<400> 88
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly
35 40 45
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
50 55 60
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His
65 70 75 80
Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
85 90 95
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
100 105 110
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
115 120 125
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala
130 135 140
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
145 150 155 160
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val
165 170 175
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
180 185 190
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln
195 200 205
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
210 215 220
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
225 230 235 240
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
245 250 255
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
260 265 270
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
275 280 285
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
290 295 300
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
305 310 315 320
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
325 330 335
Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Asn
340 345 350
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
355 360 365
Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val Ala Ile Ala
370 375 380
Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
385 390 395 400
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
405 410 415
Ile Ala Ser Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu
420 425 430
Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Gln Gln Val Val
435 440 445
Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
450 455 460
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
465 470 475 480
Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr
485 490 495
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
500 505 510
Gln Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala Leu
515 520 525
Glu
<210> 89
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> CTLA4_T01-L TALEN
<400> 89
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 600
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 720
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 900
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1020
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaataa tggtggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccggagcagg tggtggccat cgccagccac gatggcggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1320
atcgccagca atattggtgg caagcaggcg ctggagacgg tgcaggcgct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag ccacgatggc 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1620
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1740
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 90
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> CTLA4_T01-R TALEN
<400> 90
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caataatggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 720
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 840
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccggagca ggtggtggcc atcgccagca atattggtgg caagcaggcg 960
ctggagacgg tgcaggcgct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1020
caggtggtgg ccatcgccag caatattggt ggcaagcagg cgctggagac ggtgcaggcg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1140
agcaataatg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagcaatat tggtggcaag 1260
caggcgctgg agacggtgca ggcgctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccggagcagg tggtggccat cgccagccac gatggcggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1440
atcgccagca atattggtgg caagcaggcg ctggagacgg tgcaggcgct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caataatggt 1560
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1740
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 1860
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 91
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> CTLA4_T03-L TALEN
<400> 91
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 600
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 720
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 900
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1020
agcaatattg gtggcaagca ggcgctggag acggtgcagg cgctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaatgg cggtggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccccagcagg tggtggccat cgccagcaat aatggtggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1320
atcgccagcc acgatggcgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caatggcggt 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 1560
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 1740
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccggagca ggtggtggcc atcgccagca atattggtgg caagcaggcg 1860
ctggagacgg tgcaggcgct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag caatattggt ggcaagcagg cgctggagac ggtgcaggcg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 92
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> CTLA4_T03-RTALEN
<400> 92
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caataatggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 720
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 840
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccggagca ggtggtggcc atcgccagca atattggtgg caagcaggcg 960
ctggagacgg tgcaggcgct gttgccggtg ctgtgccagg cccacggctt gaccccccag 1020
caggtggtgg ccatcgccag caataatggt ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1140
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagcaatat tggtggcaag 1260
caggcgctgg agacggtgca ggcgctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccggagcagg tggtggccat cgccagccac gatggcggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccggagca ggtggtggcc 1440
atcgccagcc acgatggcgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag caatattggt 1560
ggcaagcagg cgctggagac ggtgcaggcg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1740
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1860
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 93
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> CTLA4_T04-L TALEN
<400> 93
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 600
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 720
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccccag 900
caggtggtgg ccatcgccag caataatggt ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1020
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaatgg cggtggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccggagcagg tggtggccat cgccagcaat attggtggca agcaggcgct ggagacggtg 1260
caggcgctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1320
atcgccagca ataatggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag ccacgatggc 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 1560
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1620
gtggccatcg ccagcaatat tggtggcaag caggcgctgg agacggtgca ggcgctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1740
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 94
<211> 2832
<212> ДНК
<213> Искусственная
<220>
<223> CTLA4_T04-R TALEN
<400> 94
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caggtggtgg ccatcgccag caataatggt 540
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 600
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 660
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 720
gtggccatcg ccagcaatgg cggtggcaag caggcgctgg agacggtcca gcggctgttg 780
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 840
aatggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 900
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 960
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1020
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1080
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1140
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1200
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagcaatat tggtggcaag 1260
caggcgctgg agacggtgca ggcgctgttg ccggtgctgt gccaggccca cggcttgacc 1320
ccccagcagg tggtggccat cgccagcaat aatggtggca agcaggcgct ggagacggtc 1380
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1440
atcgccagca atggcggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1500
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag caatattggt 1560
ggcaagcagg cgctggagac ggtgcaggcg ctgttgccgg tgctgtgcca ggcccacggc 1620
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 1680
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1740
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 1800
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagcaat 1860
attggtggca agcaggcgct ggagacggtg caggcgctgt tgccggtgct gtgccaggcc 1920
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 1980
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gacccctcag 2040
caggtggtgg ccatcgccag caatggcggc ggcaggccgg cgctggagag cattgttgcc 2100
cagttatctc gccctgatcc ggcgttggcc gcgttgacca acgaccacct cgtcgccttg 2160
gcctgcctcg gcgggcgtcc tgcgctggat gcagtgaaaa agggattggg ggatcctatc 2220
agccgttccc agctggtgaa gtccgagctg gaggagaaga aatccgagtt gaggcacaag 2280
ctgaagtacg tgccccacga gtacatcgag ctgatcgaga tcgcccggaa cagcacccag 2340
gaccgtatcc tggagatgaa ggtgatggag ttcttcatga aggtgtacgg ctacaggggc 2400
aagcacctgg gcggctccag gaagcccgac ggcgccatct acaccgtggg ctcccccatc 2460
gactacggcg tgatcgtgga caccaaggcc tactccggcg gctacaacct gcccatcggc 2520
caggccgacg aaatgcagag gtacgtggag gagaaccaga ccaggaacaa gcacatcaac 2580
cccaacgagt ggtggaaggt gtacccctcc agcgtgaccg agttcaagtt cctgttcgtg 2640
tccggccact tcaagggcaa ctacaaggcc cagctgacca ggctgaacca catcaccaac 2700
tgcaacggcg ccgtgctgtc cgtggaggag ctcctgatcg gcggcgagat gatcaaggcc 2760
ggcaccctga ccctggagga ggtgaggagg aagttcaaca acggcgagat caacttcgcg 2820
gccgactgat aa 2832
<210> 95
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> PDCD1_T01-L TALEN
<400> 95
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgacccccc agcaggtggt ggccatcgcc agcaatggcg gtggcaagca ggcgctggag 540
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 600
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 720
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 900
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgaccccgg agcaggtggt ggccatcgcc 1020
agccacgatg gcggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ggagcaggtg gtggccatcg ccagccacga tggcggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccggagcagg tggtggccat cgccagcaat attggtggca agcaggcgct ggagacggtg 1260
caggcgctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1320
atcgccagca ataatggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccggag caggtggtgg ccatcgccag ccacgatggc 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgaccccgg agcaggtggt ggccatcgcc agccacgatg gcggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 1620
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccccagcagg tggtggccat cgccagcaat 1740
ggcggtggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccccagca ggtggtggcc atcgccagca ataatggtgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 96
<211> 2829
<212> ДНК
<213> Искусственная
<220>
<223> PDCD1_T01-R TALEN
<400> 96
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gaccccccag caagtcgtcg caatcgccag caataacgga 540
gggaagcaag ccctcgaaac cgtgcagcgg ttgcttcctg tgctctgcca ggcccacggc 600
cttacccctg agcaggtggt ggccatcgca agtaacattg gaggaaagca agccttggag 660
acagtgcagg ccctgttgcc cgtgctgtgc caggcacacg gcctcacacc agagcaggtc 720
gtggccattg cctccaacat cggggggaaa caggctctgg agaccgtcca ggccctgctg 780
cccgtcctct gtcaagctca cggcctgact ccccaacaag tggtcgccat cgcctctaat 840
aacggcggga agcaggcact ggaaacagtg cagagactgc tccctgtgct ttgccaagct 900
catgggttga ccccccaaca ggtcgtcgct attgcctcaa acaacggggg caagcaggcc 960
cttgagactg tgcagaggct gttgccagtg ctgtgtcagg ctcacgggct cactccacaa 1020
caggtggtcg caattgccag caacggcggc ggaaagcaag ctcttgaaac cgtgcaacgc 1080
ctcctgcccg tgctctgtca ggctcatggc ctgacaccac aacaagtcgt ggccatcgcc 1140
agtaataatg gcgggaaaca ggctcttgag accgtccaga ggctgctccc agtgctctgc 1200
caggcacacg ggctgacccc ccagcaggtg gtggctatcg ccagcaataa tgggggcaag 1260
caggccctgg aaacagtcca gcgcctgctg ccagtgcttt gccaggctca cgggctcact 1320
cccgaacagg tcgtggcaat cgcctccaac ggagggaagc aggctctgga gaccgtgcag 1380
agactgctgc ccgtcttgtg ccaggcccac ggactcacac ctcagcaggt cgtcgccatt 1440
gcctctaaca acgggggcaa acaagccctg gagacagtgc agcggctgtt gcctgtgttg 1500
tgccaagccc acggcttgac tcctcaacaa gtggtcgcca tcgcctcaaa tggcggcgga 1560
aaacaagctc tggagacagt gcagaggttg ctgcccgtcc tctgccaagc ccacggcctg 1620
actccccaac aggtcgtcgc cattgccagc aacggcggag gaaagcaggc tctcgaaact 1680
gtgcagcggc tgcttcctgt gctgtgtcag gctcatgggc tgacccccca gcaagtggtg 1740
gctattgcct ctaacaatgg aggcaagcaa gcccttgaga cagtccagag gctgttgcca 1800
gtgctgtgcc aggcccacgg gctcacaccc cagcaggtgg tcgccatcgc cagtaacggc 1860
gggggcaaac aggcattgga aaccgtccag cgcctgcttc cagtgctctg ccaggcacac 1920
ggactgacac ccgaacaggt ggtggccatt gcatcccatg atgggggcaa gcaggccctg 1980
gagaccgtgc agagactcct gccagtgttg tgccaagctc acggcctcac ccctcagcaa 2040
gtcgtggcca tcgcctcaaa cggggggggc cggcctgcac tggagagcat tgttgcccag 2100
ttatctcgcc ctgatccggc gttggccgcg ttgaccaacg accacctcgt cgccttggcc 2160
tgcctcggcg ggcgtcctgc gctggatgca gtgaaaaagg gattggggga tcctatcagc 2220
cgttcccagc tggtgaagtc cgagctggag gagaagaaat ccgagttgag gcacaagctg 2280
aagtacgtgc cccacgagta catcgagctg atcgagatcg cccggaacag cacccaggac 2340
cgtatcctgg agatgaaggt gatggagttc ttcatgaagg tgtacggcta caggggcaag 2400
cacctgggcg gctccaggaa gcccgacggc gccatctaca ccgtgggctc ccccatcgac 2460
tacggcgtga tcgtggacac caaggcctac tccggcggct acaacctgcc catcggccag 2520
gccgacgaaa tgcagaggta cgtggaggag aaccagacca ggaacaagca catcaacccc 2580
aacgagtggt ggaaggtgta cccctccagc gtgaccgagt tcaagttcct gttcgtgtcc 2640
ggccacttca agggcaacta caaggcccag ctgaccaggc tgaaccacat caccaactgc 2700
aacggcgccg tgctgtccgt ggaggagctc ctgatcggcg gcgagatgat caaggccggc 2760
accctgaccc tggaggaggt gaggaggaag ttcaacaacg gcgagatcaa cttcgcggcc 2820
gactgataa 2829
<210> 97
<211> 2814
<212> ДНК
<213> Искусственная
<220>
<223> PDCD1_T03-L TALEN
<400> 97
atgggcgatc ctaaaaagaa acgtaaggtc atcgattacc catacgatgt tccagattac 60
gctatcgata tcgccgatct acgcacgctc ggctacagcc agcagcaaca ggagaagatc 120
aaaccgaagg ttcgttcgac agtggcgcag caccacgagg cactggtcgg ccacgggttt 180
acacacgcgc acatcgttgc gttaagccaa cacccggcag cgttagggac cgtcgctgtc 240
aagtatcagg acatgatcgc agcgttgcca gaggcgacac acgaagcgat cgttggcgtc 300
ggcaaacagt ggtccggcgc acgcgctctg gaggccttgc tcacggtggc gggagagttg 360
agaggtccac cgttacagtt ggacacaggc caacttctca agattgcaaa acgtggcggc 420
gtgaccgcag tggaggcagt gcatgcatgg cgcaatgcac tgacgggtgc cccgctcaac 480
ttgaccccgg agcaggtggt ggccatcgcc agcaatattg gtggcaagca ggcgctggag 540
acggtgcagg cgctgttgcc ggtgctgtgc caggcccacg gcttgacccc ggagcaggtg 600
gtggccatcg ccagccacga tggcggcaag caggcgctgg agacggtcca gcggctgttg 660
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 720
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 780
cacggcttga ccccccagca ggtggtggcc atcgccagca atggcggtgg caagcaggcg 840
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 900
caggtggtgg ccatcgccag ccacgatggc ggcaagcagg cgctggagac ggtccagcgg 960
ctgttgccgg tgctgtgcca ggcccacggc ttgacccccc agcaggtggt ggccatcgcc 1020
agcaatggcg gtggcaagca ggcgctggag acggtccagc ggctgttgcc ggtgctgtgc 1080
caggcccacg gcttgacccc ccagcaggtg gtggccatcg ccagcaataa tggtggcaag 1140
caggcgctgg agacggtcca gcggctgttg ccggtgctgt gccaggccca cggcttgacc 1200
ccccagcagg tggtggccat cgccagcaat ggcggtggca agcaggcgct ggagacggtc 1260
cagcggctgt tgccggtgct gtgccaggcc cacggcttga ccccccagca ggtggtggcc 1320
atcgccagca ataatggtgg caagcaggcg ctggagacgg tccagcggct gttgccggtg 1380
ctgtgccagg cccacggctt gaccccccag caggtggtgg ccatcgccag caataatggt 1440
ggcaagcagg cgctggagac ggtccagcgg ctgttgccgg tgctgtgcca ggcccacggc 1500
ttgacccccc agcaggtggt ggccatcgcc agcaataatg gtggcaagca ggcgctggag 1560
acggtccagc ggctgttgcc ggtgctgtgc caggcccacg gcttgacccc ccagcaggtg 1620
gtggccatcg ccagcaataa tggtggcaag caggcgctgg agacggtcca gcggctgttg 1680
ccggtgctgt gccaggccca cggcttgacc ccggagcagg tggtggccat cgccagccac 1740
gatggcggca agcaggcgct ggagacggtc cagcggctgt tgccggtgct gtgccaggcc 1800
cacggcttga ccccggagca ggtggtggcc atcgccagcc acgatggcgg caagcaggcg 1860
ctggagacgg tccagcggct gttgccggtg ctgtgccagg cccacggctt gaccccggag 1920
caggtggtgg ccatcgccag caatattggt ggcaagcagg cgctggagac ggtgcaggcg 1980
ctgttgccgg tgctgtgcca ggcccacggc ttgacccctc agcaggtggt ggccatcgcc 2040
agcaatggcg gcggcaggcc ggcgctggag agcattgttg cccagttatc tcgccctgat 2100
ccggcgttgg ccgcgttgac caacgaccac ctcgtcgcct tggcctgcct cggcgggcgt 2160
cctgcgctgg atgcagtgaa aaagggattg ggggatccta tcagccgttc ccagctggtg 2220
aagtccgagc tggaggagaa gaaatccgag ttgaggcaca agctgaagta cgtgccccac 2280
gagtacatcg agctgatcga gatcgcccgg aacagcaccc aggaccgtat cctggagatg 2340
aaggtgatgg agttcttcat gaaggtgtac ggctacaggg gcaagcacct gggcggctcc 2400
aggaagcccg acggcgccat ctacaccgtg ggctccccca tcgactacgg cgtgatcgtg 2460
gacaccaagg cctactccgg cggctacaac ctgcccatcg gccaggccga cgaaatgcag 2520
aggtacgtgg aggagaacca gaccaggaac aagcacatca accccaacga gtggtggaag 2580
gtgtacccct ccagcgtgac cgagttcaag ttcctgttcg tgtccggcca cttcaagggc 2640
aactacaagg cccagctgac caggctgaac cacatcacca actgcaacgg cgccgtgctg 2700
tccgtggagg agctcctgat cggcggcgag atgatcaagg ccggcaccct gaccctggag 2760
gaggtgagga ggaagttcaa caacggcgag atcaacttcg cggccgactg ataa 2814
<210> 98
<211> 2829
<212> ДНК
<213> Искусственная
<220>
<223> PDCD1_T03-R TALEN
<400> 98
atgggcgatc ctaaaaagaa acgtaaggtc atcgataagg agaccgccgc tgccaagttc 60
gagagacagc acatggacag catcgatatc gccgatctac gcacgctcgg ctacagccag 120
cagcaacagg agaagatcaa accgaaggtt cgttcgacag tggcgcagca ccacgaggca 180
ctggtcggcc acgggtttac acacgcgcac atcgttgcgt taagccaaca cccggcagcg 240
ttagggaccg tcgctgtcaa gtatcaggac atgatcgcag cgttgccaga ggcgacacac 300
gaagcgatcg ttggcgtcgg caaacagtgg tccggcgcac gcgctctgga ggccttgctc 360
acggtggcgg gagagttgag aggtccaccg ttacagttgg acacaggcca acttctcaag 420
attgcaaaac gtggcggcgt gaccgcagtg gaggcagtgc atgcatggcg caatgcactg 480
acgggtgccc cgctcaactt gacccccgag caagtcgtcg caatcgccag ccatgatgga 540
gggaagcaag ccctcgaaac cgtgcagcgg ttgcttcctg tgctctgcca ggcccacggc 600
cttacccctc agcaggtggt ggccatcgca agtaacggag gaggaaagca agccttggag 660
acagtgcagc gcctgttgcc cgtgctgtgc caggcacacg gcctcacacc agagcaggtc 720
gtggccattg cctcccatga cggggggaaa caggctctgg agaccgtcca gaggctgctg 780
cccgtcctct gtcaagctca cggcctgact ccccaacaag tggtcgccat cgcctctaat 840
ggcggcggga agcaggcact ggaaacagtg cagagactgc tccctgtgct ttgccaagct 900
catgggttga ccccccaaca ggtcgtcgct attgcctcaa acgggggggg caagcaggcc 960
cttgagactg tgcagaggct gttgccagtg ctgtgtcagg ctcacgggct cactccacaa 1020
caggtggtcg caattgccag caacggcggc ggaaagcaag ctcttgaaac cgtgcaacgc 1080
ctcctgcccg tgctctgtca ggctcatggc ctgacaccac aacaagtcgt ggccatcgcc 1140
agtaataatg gcgggaaaca ggctcttgag accgtccaga ggctgctccc agtgctctgc 1200
caggcacacg ggctgacccc cgagcaggtg gtggctatcg ccagcaatat tgggggcaag 1260
caggccctgg aaacagtcca ggccctgctg ccagtgcttt gccaggctca cgggctcact 1320
ccccagcagg tcgtggcaat cgcctccaac ggcggaggga agcaggctct ggagaccgtg 1380
cagagactgc tgcccgtctt gtgccaggcc cacggactca cacctgaaca ggtcgtcgcc 1440
attgcctctc acgatggggg caaacaagcc ctggagacag tgcagcggct gttgcctgtg 1500
ttgtgccaag cccacggctt gactcctcaa caagtggtcg ccatcgcctc aaatggcggc 1560
ggaaaacaag ctctggagac agtgcagagg ttgctgcccg tcctctgcca agcccacggc 1620
ctgactcccc aacaggtcgt cgccattgcc agcaacaacg gaggaaagca ggctctcgaa 1680
actgtgcagc ggctgcttcc tgtgctgtgt caggctcatg ggctgacccc cgagcaagtg 1740
gtggctattg cctctaatgg aggcaagcaa gcccttgaga cagtccagag gctgttgcca 1800
gtgctgtgcc aggcccacgg gctcacaccc cagcaggtgg tcgccatcgc cagtaacaac 1860
gggggcaaac aggcattgga aaccgtccag cgcctgcttc cagtgctctg ccaggcacac 1920
ggactgacac ccgaacaggt ggtggccatt gcatcccatg atgggggcaa gcaggccctg 1980
gagaccgtgc agagactcct gccagtgttg tgccaagctc acggcctcac ccctcagcaa 2040
gtcgtggcca tcgcctcaaa cggggggggc cggcctgcac tggagagcat tgttgcccag 2100
ttatctcgcc ctgatccggc gttggccgcg ttgaccaacg accacctcgt cgccttggcc 2160
tgcctcggcg ggcgtcctgc gctggatgca gtgaaaaagg gattggggga tcctatcagc 2220
cgttcccagc tggtgaagtc cgagctggag gagaagaaat ccgagttgag gcacaagctg 2280
aagtacgtgc cccacgagta catcgagctg atcgagatcg cccggaacag cacccaggac 2340
cgtatcctgg agatgaaggt gatggagttc ttcatgaagg tgtacggcta caggggcaag 2400
cacctgggcg gctccaggaa gcccgacggc gccatctaca ccgtgggctc ccccatcgac 2460
tacggcgtga tcgtggacac caaggcctac tccggcggct acaacctgcc catcggccag 2520
gccgacgaaa tgcagaggta cgtggaggag aaccagacca ggaacaagca catcaacccc 2580
aacgagtggt ggaaggtgta cccctccagc gtgaccgagt tcaagttcct gttcgtgtcc 2640
ggccacttca agggcaacta caaggcccag ctgaccaggc tgaaccacat caccaactgc 2700
aacggcgccg tgctgtccgt ggaggagctc ctgatcggcg gcgagatgat caaggccggc 2760
accctgaccc tggaggaggt gaggaggaag ttcaacaacg gcgagatcaa cttcgcggcc 2820
gactgataa 2829
<210> 99
<211> 60
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер CTLA4_T01
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 99
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn ctctacttcc tgaagacctg 60
<210> 100
<211> 60
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер CTLA4_T03/T04
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 100
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn acagttgaga gatggagggg 60
<210> 101
<211> 59
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер PDCD1_T01
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 101
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn ccacagaggt aggtgccgc 59
<210> 102
<211> 60
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер PDCD1_T03
<220>
<221> разные свойства
<222> (31)..(40)
<223> n is a or c or t or g
<400> 102
ccatctcatc cctgcgtgtc tccgactcag nnnnnnnnnn gacagagatg ccggtcacca 60
<210> 103
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер CTLA4_T01
<400> 103
cctatcccct gtgtgccttg gcagtctcag tggaatacag agccagccaa 50
<210> 104
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер CTLA4_T03/04
<400> 104
cctatcccct gtgtgccttg gcagtctcag ggtgcccgtg cagatggaat 50
<210> 105
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер PDCD1_T01
<400> 105
cctatcccct gtgtgccttg gcagtctcag ggctctgcag tggaggccag 50
<210> 106
<211> 50
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер PDCD1_T03
<400> 106
cctatcccct gtgtgccttg gcagtctcag ggacaacgcc accttcacct 50
<210> 107
<211> 281
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-FL
<400> 107
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val Trp Gly Glu Gly Ser Tyr Leu
225 230 235 240
Ser Ser Tyr Pro Thr Cys Pro Ala Gln Ala Trp Cys Ser Arg Ser Arg
245 250 255
Leu Arg Ala Pro Ser Ser Ser Leu Gly Ala Phe Phe Arg Gly Asp Leu
260 265 270
Pro Pro Pro Leu Gln Ala Gly Ala Ala
275 280
<210> 108
<211> 263
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta18
<400> 108
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val Trp Gly Glu Gly Ser Tyr Leu
225 230 235 240
Ser Ser Tyr Pro Thr Cys Pro Ala Gln Ala Trp Cys Ser Arg Ser Arg
245 250 255
Leu Arg Ala Pro Ser Ser Ser
260
<210> 109
<211> 233
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta48
<400> 109
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val
225 230
<210> 110
<211> 219
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta62
<400> 110
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg
210 215
<210> 111
<211> 203
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta78
<400> 111
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly
195 200
<210> 112
<211> 189
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta92
<400> 112
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg
180 185
<210> 113
<211> 171
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta110
<400> 113
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys
165 170
<210> 114
<211> 167
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta114
<400> 114
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu
165
<210> 115
<211> 344
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-FL-CD28
<400> 115
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val Trp Gly Glu Gly Ser Tyr Leu
225 230 235 240
Ser Ser Tyr Pro Thr Cys Pro Ala Gln Ala Trp Cys Ser Arg Ser Arg
245 250 255
Leu Arg Ala Pro Ser Ser Ser Leu Gly Ala Phe Phe Arg Gly Asp Leu
260 265 270
Pro Pro Pro Leu Gln Ala Gly Ala Ala Ala Ser Gly Gly Val Leu Ala
275 280 285
Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg
290 295 300
Ser Lys Arg Ser Arg Gly Gly His Ser Asp Tyr Met Asn Met Thr Pro
305 310 315 320
Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro
325 330 335
Arg Asp Phe Ala Ala Tyr Arg Ser
340
<210> 116
<211> 311
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-FL-CD8
<400> 116
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val Trp Gly Glu Gly Ser Tyr Leu
225 230 235 240
Ser Ser Tyr Pro Thr Cys Pro Ala Gln Ala Trp Cys Ser Arg Ser Arg
245 250 255
Leu Arg Ala Pro Ser Ser Ser Leu Gly Ala Phe Phe Arg Gly Asp Leu
260 265 270
Pro Pro Pro Leu Gln Ala Gly Ala Ala Ala Ser His Arg Asn Arg Arg
275 280 285
Arg Val Cys Lys Cys Pro Arg Pro Val Val Lys Ser Gly Asp Lys Pro
290 295 300
Ser Leu Ser Ala Arg Tyr Val
305 310
<210> 117
<211> 325
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-FL-41BB
<400> 117
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val Trp Gly Glu Gly Ser Tyr Leu
225 230 235 240
Ser Ser Tyr Pro Thr Cys Pro Ala Gln Ala Trp Cys Ser Arg Ser Arg
245 250 255
Leu Arg Ala Pro Ser Ser Ser Leu Gly Ala Phe Phe Arg Gly Asp Leu
260 265 270
Pro Pro Pro Leu Gln Ala Gly Ala Ala Gly Ser Lys Arg Gly Arg Lys
275 280 285
Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr
290 295 300
Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu
305 310 315 320
Gly Gly Cys Glu Leu
325
<210> 118
<211> 296
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta48-CD28
<400> 118
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val Ala Ser Gly Gly Val Leu Ala
225 230 235 240
Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg
245 250 255
Ser Lys Arg Ser Arg Gly Gly His Ser Asp Tyr Met Asn Met Thr Pro
260 265 270
Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro
275 280 285
Arg Asp Phe Ala Ala Tyr Arg Ser
290 295
<210> 119
<211> 263
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta48-CD8
<400> 119
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val Ala Ser His Arg Asn Arg Arg
225 230 235 240
Arg Val Cys Lys Cys Pro Arg Pro Val Val Lys Ser Gly Asp Lys Pro
245 250 255
Ser Leu Ser Ala Arg Tyr Val
260
<210> 120
<211> 277
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta48-41BB
<400> 120
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val Gly Ser Lys Arg Gly Arg Lys
225 230 235 240
Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr
245 250 255
Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu
260 265 270
Gly Gly Cys Glu Leu
275
<210> 121
<211> 172
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha-Delta114-TCRalpha.IC
<400> 121
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Arg Leu Trp Ser Ser
165 170
<210> 122
<211> 173
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha.EC-TCRalpha. TM. IC
<400> 122
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
145 150 155 160
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
165 170
<210> 123
<211> 233
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha.EC-Delta48-1xMUT
<400> 123
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Asp Gly Lys Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Arg Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Arg Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Arg Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val
225 230
<210> 124
<211> 233
<212> БЕЛОК
<213> Искусственная
<220>
<223> pTalpha.EC-Delta48-4xMUT
<400> 124
Met Ala Gly Thr Trp Leu Leu Leu Leu Leu Ala Leu Gly Cys Pro Ala
1 5 10 15
Leu Pro Thr Gly Val Gly Gly Thr Pro Phe Pro Ser Leu Ala Pro Pro
20 25 30
Ile Met Leu Leu Val Ala Gly Ala Gln Gln Met Val Val Val Cys Leu
35 40 45
Val Leu Asp Val Ala Pro Pro Gly Leu Asp Ser Pro Ile Trp Phe Ser
50 55 60
Ala Gly Asn Gly Ser Ala Leu Asp Ala Phe Thr Tyr Gly Pro Ser Pro
65 70 75 80
Ala Thr Asp Gly Thr Trp Thr Asn Leu Ala His Leu Ser Leu Pro Ser
85 90 95
Glu Glu Leu Ala Ser Trp Glu Pro Leu Val Cys His Thr Gly Pro Gly
100 105 110
Ala Glu Gly His Ser Ala Ser Thr Gln Pro Met His Leu Ser Gly Glu
115 120 125
Ala Ser Thr Ala Ala Thr Cys Pro Gln Glu Pro Leu Arg Gly Thr Pro
130 135 140
Gly Gly Ala Leu Trp Leu Gly Val Leu Arg Leu Leu Leu Phe Lys Leu
145 150 155 160
Leu Leu Phe Asp Leu Leu Leu Thr Cys Ser Cys Leu Cys Asp Pro Ala
165 170 175
Gly Pro Leu Pro Ser Pro Ala Thr Thr Thr Arg Leu Arg Ala Leu Gly
180 185 190
Ser His Arg Leu His Pro Ala Thr Glu Thr Gly Gly Arg Glu Ala Thr
195 200 205
Ser Ser Pro Arg Pro Gln Pro Arg Asp Arg Arg Trp Gly Asp Thr Pro
210 215 220
Pro Gly Arg Lys Pro Gly Ser Pro Val
225 230
<210> 125
<211> 848
<212> БЕЛОК
<213> Искусственная
<220>
<223> Многоцепочечный CAR
<400> 125
Met Ala Pro Ala Met Glu Ser Pro Thr Leu Leu Cys Val Ala Leu Leu
1 5 10 15
Phe Phe Ala Pro Asp Gly Val Leu Ala Glu Val Gln Leu Gln Gln Ser
20 25 30
Gly Pro Glu Leu Ile Lys Pro Gly Ala Ser Val Lys Met Ser Cys Lys
35 40 45
Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Val Met His Trp Val Lys Gln
50 55 60
Lys Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Tyr Asn
65 70 75 80
Asp Gly Thr Lys Tyr Asn Glu Lys Phe Lys Gly Lys Ala Thr Leu Thr
85 90 95
Ser Asp Lys Ser Ser Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Thr
100 105 110
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Gly Thr Tyr Tyr Tyr
115 120 125
Gly Ser Arg Val Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val
130 135 140
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
145 150 155 160
Ser Asp Ile Val Met Thr Gln Ala Ala Pro Ser Ile Pro Val Thr Pro
165 170 175
Gly Glu Ser Val Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu Asn
180 185 190
Ser Asn Gly Asn Thr Tyr Leu Tyr Trp Phe Leu Gln Arg Pro Gly Gln
195 200 205
Ser Pro Gln Leu Leu Ile Tyr Arg Met Ser Asn Leu Ala Ser Gly Val
210 215 220
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Ala Phe Thr Leu Arg
225 230 235 240
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln
245 250 255
His Leu Glu Tyr Pro Phe Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu
260 265 270
Lys Arg Ala Asp Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala
275 280 285
Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg
290 295 300
Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys
305 310 315 320
Asp Phe Phe Ile Pro Leu Leu Val Val Ile Leu Phe Ala Val Asp Thr
325 330 335
Gly Leu Phe Ile Ser Thr Gln Gln Gln Val Thr Phe Leu Leu Lys Ile
340 345 350
Lys Arg Thr Arg Lys Gly Phe Arg Leu Leu Asn Pro His Pro Lys Pro
355 360 365
Asn Pro Lys Asn Asn Arg Ala Glu Gly Arg Gly Ser Leu Leu Thr Cys
370 375 380
Gly Asp Val Glu Glu Asn Pro Gly Pro Met Asp Thr Glu Ser Asn Arg
385 390 395 400
Arg Ala Asn Leu Ala Leu Pro Gln Glu Pro Ser Ser Val Pro Ala Phe
405 410 415
Glu Val Leu Glu Ile Ser Pro Gln Glu Val Ser Ser Gly Arg Leu Leu
420 425 430
Lys Ser Ala Ser Ser Pro Pro Leu His Thr Trp Leu Thr Val Leu Lys
435 440 445
Lys Glu Gln Glu Phe Leu Gly Val Thr Gln Ile Leu Thr Ala Met Ile
450 455 460
Cys Leu Cys Phe Gly Thr Val Val Cys Ser Val Leu Asp Ile Ser His
465 470 475 480
Ile Glu Gly Asp Ile Phe Ser Ser Phe Lys Ala Gly Tyr Pro Phe Trp
485 490 495
Gly Ala Ile Phe Phe Ser Ile Ser Gly Met Leu Ser Ile Ile Ser Glu
500 505 510
Arg Arg Asn Ala Thr Tyr Leu Val Arg Gly Ser Leu Gly Ala Asn Thr
515 520 525
Ala Ser Ser Ile Ala Gly Gly Thr Gly Ile Thr Ile Leu Ile Ile Asn
530 535 540
Leu Lys Lys Ser Leu Ala Tyr Ile His Ile His Ser Cys Gln Lys Phe
545 550 555 560
Phe Glu Thr Lys Cys Phe Met Ala Ser Phe Ser Thr Glu Ile Val Val
565 570 575
Met Met Leu Phe Leu Thr Ile Leu Gly Leu Gly Ser Ala Val Ser Leu
580 585 590
Thr Ile Cys Gly Ala Gly Glu Glu Leu Lys Gly Asn Lys Val Pro Glu
595 600 605
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
610 615 620
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
625 630 635 640
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Gly Ser Gly Val Lys Gln
645 650 655
Thr Leu Asn Phe Asp Leu Leu Lys Leu Ala Gly Asp Val Glu Ser Asn
660 665 670
Pro Gly Pro Met Ile Pro Ala Val Val Leu Leu Leu Leu Leu Leu Val
675 680 685
Glu Gln Ala Ala Ala Leu Gly Glu Pro Gln Leu Cys Tyr Ile Leu Asp
690 695 700
Ala Ile Leu Phe Leu Tyr Gly Ile Val Leu Thr Leu Leu Tyr Cys Arg
705 710 715 720
Leu Lys Ile Gln Val Arg Lys Ala Ala Ile Thr Ser Tyr Glu Lys Ser
725 730 735
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
740 745 750
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
755 760 765
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
770 775 780
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
785 790 795 800
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
805 810 815
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
820 825 830
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
835 840 845
<210> 126
<211> 2547
<212> ДНК
<213> Искусственная
<220>
<223> Многоцепочечный CAR
<400> 126
atggctcctg ccatggaatc ccctactcta ctgtgtgtag ccttactgtt cttcgctcca 60
gatggcgtgt tagcagaggt gcagttgcag cagtcagggc cagagttgat taagcccgga 120
gcctccgtca agatgtcctg caaggccagc gggtacactt tcaccagcta cgtcatgcat 180
tgggtgaagc agaagccagg ccaggggctt gagtggattg ggtacatcaa cccctacaac 240
gacgggacca aatacaacga gaaattcaag ggcaaagcca cactcacctc cgataagtcc 300
tcctctaccg cctacatgga gctcagctcc ctgacctccg aggatagcgc tgtgtattac 360
tgcgcaaggg gcacatacta ctatggctct agggtgttcg actactgggg gcagggcact 420
actctcacag tgagctcagg cggaggaggc agtggcggag ggggaagtgg gggcggcggc 480
agcgatattg tcatgaccca ggcagcccct agtatccctg tgactccagg cgagagcgtg 540
agcatcagct gccggtccag caagagcctg ctgaacagta acggaaacac atacctctac 600
tggtttctgc agaggcccgg ccagagccct cagctgctga tttaccgcat gtcaaatctt 660
gcctctgggg tgcccgatag atttagtggg agcggatccg gcacagcttt tacattgcgg 720
atctccagag tcgaggccga agacgtgggg gtctattact gtatgcaaca cctggaatac 780
ccctttacct tcggagccgg cacaaagctg gagctgaagc gggctgacac cacaaccccc 840
gctccaaggc cccctacccc cgcaccaact attgcctccc agccactctc actgcggcct 900
gaggcctgtc ggcccgctgc tggaggcgca gtgcatacaa ggggcctcga tttcgcctgc 960
gattttttta tcccattgtt ggtggtgatt ctgtttgctg tggacacagg attatttatc 1020
tcaactcagc agcaggtcac atttctcttg aagattaaga gaaccaggaa aggcttcaga 1080
cttctgaacc cacatcctaa gccaaacccc aaaaacaaca gagccgaggg cagaggcagc 1140
ctgctgacct gcggcgacgt ggaggagaac ccaggcccca tggacacaga aagtaatagg 1200
agagcaaatc ttgctctccc acaggagcct tccagtgtgc ctgcatttga agtcttggaa 1260
atatctcccc aggaagtatc ttcaggcaga ctattgaagt cggcctcatc cccaccactg 1320
catacatggc tgacagtttt gaaaaaagag caggagttcc tgggggtaac acaaattctg 1380
actgctatga tatgcctttg ttttggaaca gttgtctgct ctgtacttga tatttcacac 1440
attgagggag acattttttc atcatttaaa gcaggttatc cattctgggg agccatattt 1500
ttttctattt ctggaatgtt gtcaattata tctgaaagga gaaatgcaac atatctggtg 1560
agaggaagcc tgggagcaaa cactgccagc agcatagctg ggggaacggg aattaccatc 1620
ctgatcatca acctgaagaa gagcttggcc tatatccaca tccacagttg ccagaaattt 1680
tttgagacca agtgctttat ggcttccttt tccactgaaa ttgtagtgat gatgctgttt 1740
ctcaccattc tgggacttgg tagtgctgtg tcactcacaa tctgtggagc tggggaagaa 1800
ctcaaaggaa acaaggttcc agagaaacgg ggccggaaga agctcctcta catttttaag 1860
cagcctttca tgcggccagt gcagacaacc caagaggagg atgggtgttc ctgcagattc 1920
cctgaggaag aggaaggcgg gtgcgagctg ggttctggcg tgaaacagac tttgaatttt 1980
gaccttctca agttggcggg agacgtggag tccaacccag ggcccatgat tccagcagtg 2040
gtcttgctct tactcctttt ggttgaacaa gcagcggccc tgggagagcc tcagctctgc 2100
tatatcctgg atgccatcct gtttctgtat ggaattgtcc tcaccctcct ctactgtcga 2160
ctgaagatcc aagtgcgaaa ggcagctata accagctatg agaaatcaag agtgaagttc 2220
tccaggagcg cagatgcccc cgcctatcaa cagggccaga accagctcta caacgagctt 2280
aacctcggga ggcgcgaaga atacgacgtg ttggataaga gaagggggcg ggaccccgag 2340
atgggaggaa agccccggag gaagaaccct caggagggcc tgtacaacga gctgcagaag 2400
gataagatgg ccgaggccta ctcagagatc gggatgaagg gggagcggcg ccgcgggaag 2460
gggcacgatg ggctctacca ggggctgagc acagccacaa aggacacata cgacgccttg 2520
cacatgcagg cccttccacc ccggtga 2547
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ КОНСТРУИРОВАНИЯ АЛЛОГЕННЫХ И ВЫСОКОАКТИВНЫХ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2736616C2 |
| CD19-СПЕЦИФИЧЕСКИЙ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР И ЕГО ПРИМЕНЕНИЯ | 2014 |
|
RU2727447C2 |
| ЦЕЛЕВАЯ ГЕННАЯ ИНТЕГРАЦИЯ ГЕНОВ-ИНГИБИТОРОВ NK ДЛЯ УЛУЧШЕННОЙ ИММУННОЙ КЛЕТОЧНОЙ ТЕРАПИИ | 2018 |
|
RU2797304C2 |
| КЛЕТКИ ДЛЯ ИММУНОТЕРАПИИ, СКОНСТРУИРОВАННЫЕ ДЛЯ НАЦЕЛИВАНИЯ НА АНТИГЕН, ПРИСУТСТВУЮЩИЙ ОДНОВРЕМЕННО НА ИММУННЫХ КЛЕТКАХ И НА ПАТОЛОГИЧЕСКИХ КЛЕТКАХ | 2015 |
|
RU2714258C2 |
| АНТИ-CLL1-СПЕЦИФИЧЕСКИЕ ОДНОЦЕПОЧЕЧНЫЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ (scCAR) ДЛЯ ИММУНОТЕРАПИИ РАКА | 2016 |
|
RU2731543C2 |
| CD123-СПЕЦИФИЧЕСКИЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ИММУНОТЕРАПИИ РАКА | 2015 |
|
RU2727290C2 |
| ВАРИАНТЫ, КОМПОЗИЦИИ И МЕТОДЫ ПРИМЕНЕНИЯ ХОМИНГ-ЭНДОНУКЛЕАЗЫ PD-1 | 2017 |
|
RU2781083C2 |
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| ВАРИАНТЫ, КОМПОЗИЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ ЭНДОНУКЛЕАЗЫ CBLB | 2018 |
|
RU2779097C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
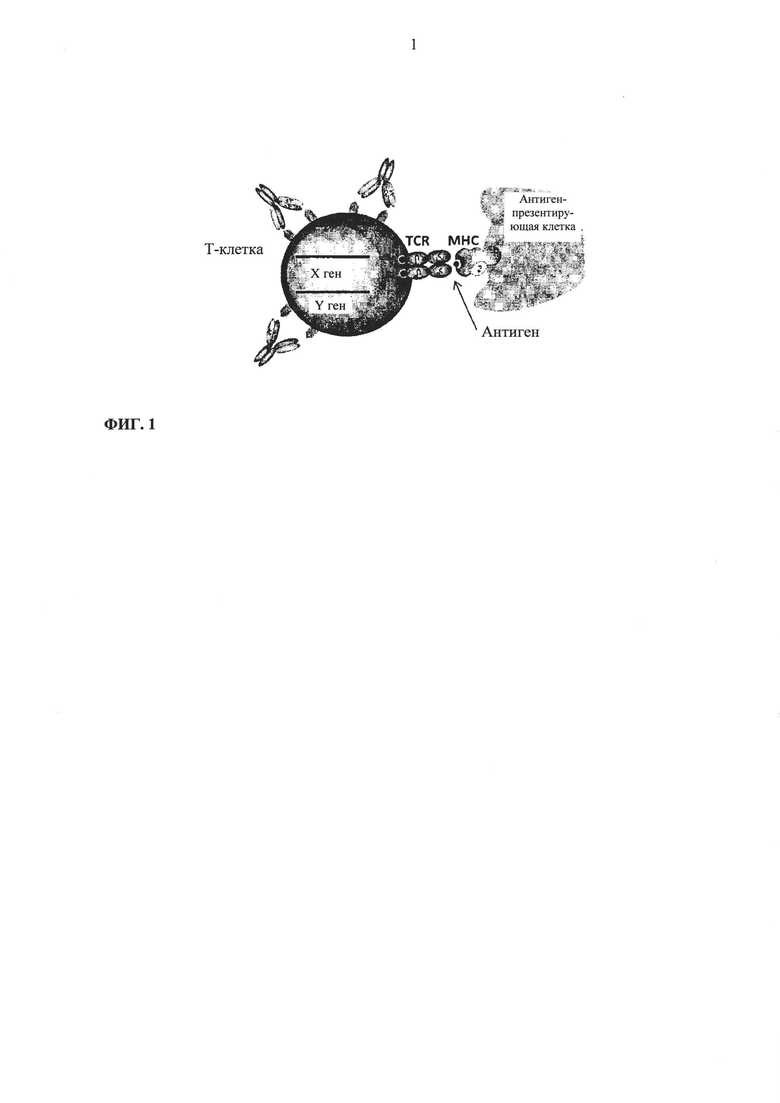
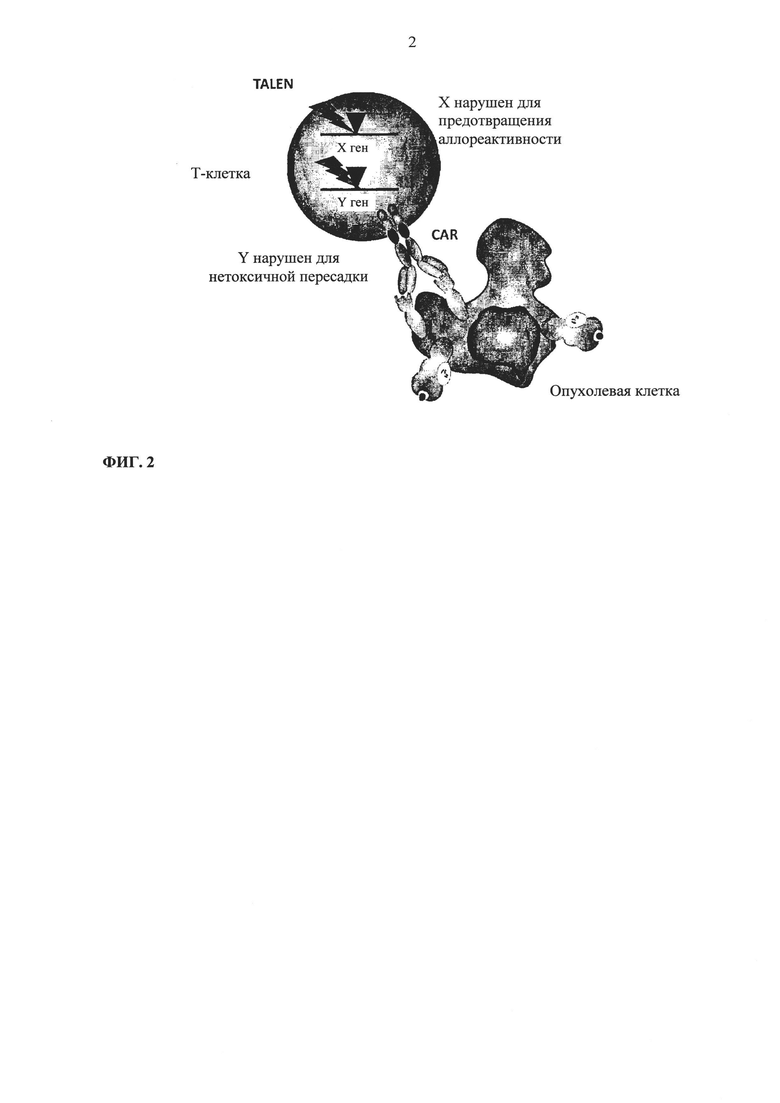

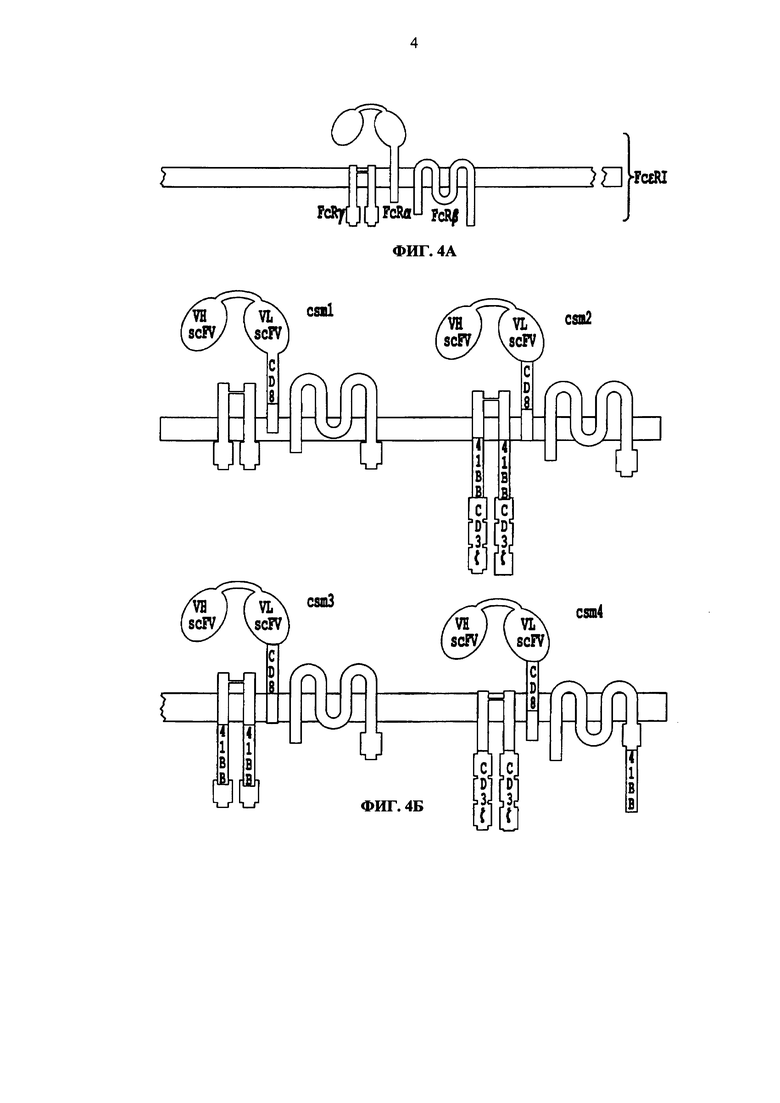
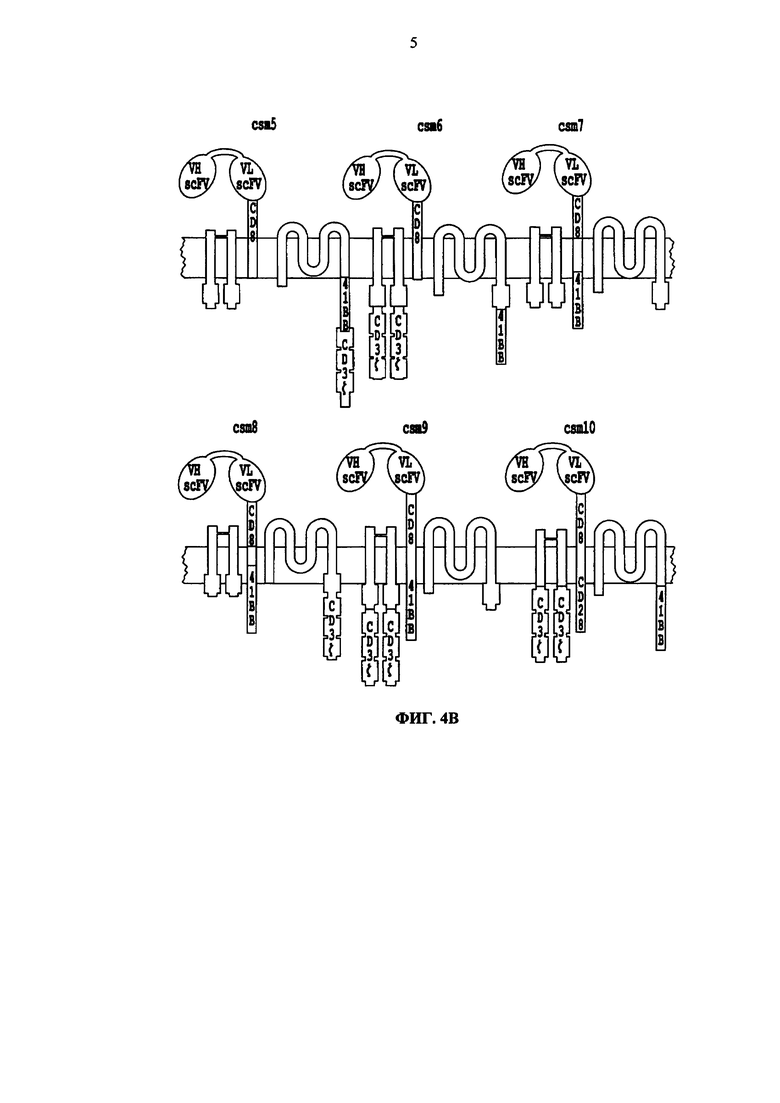
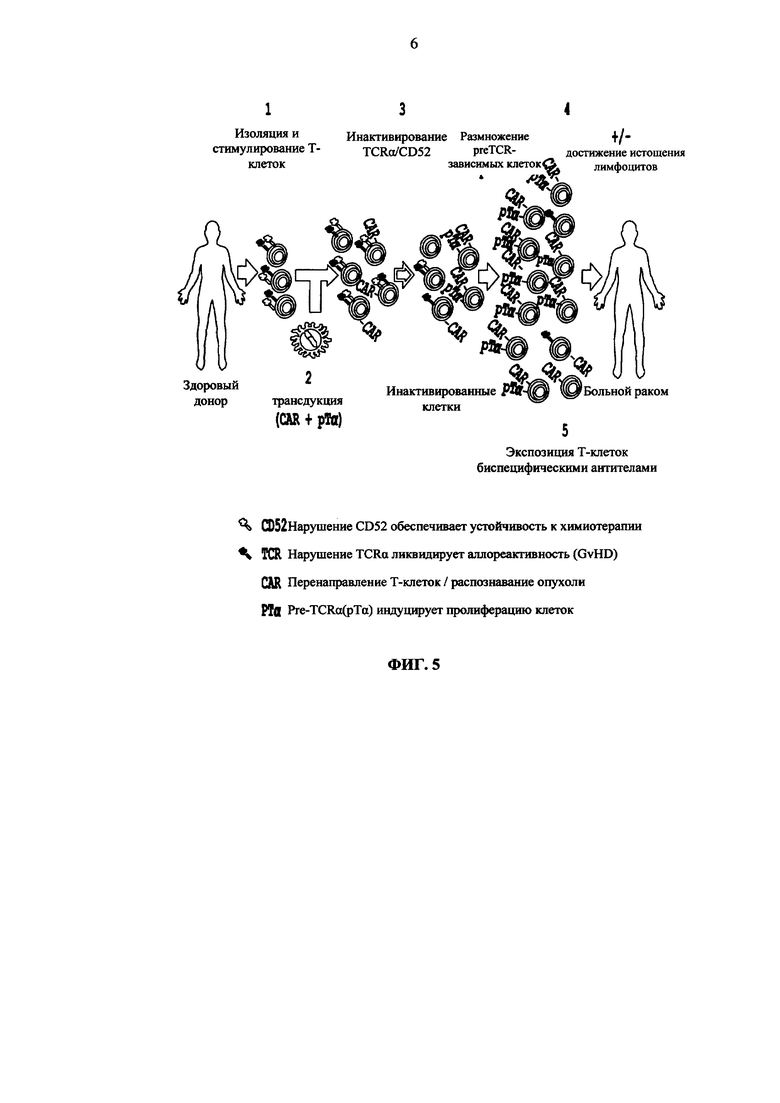
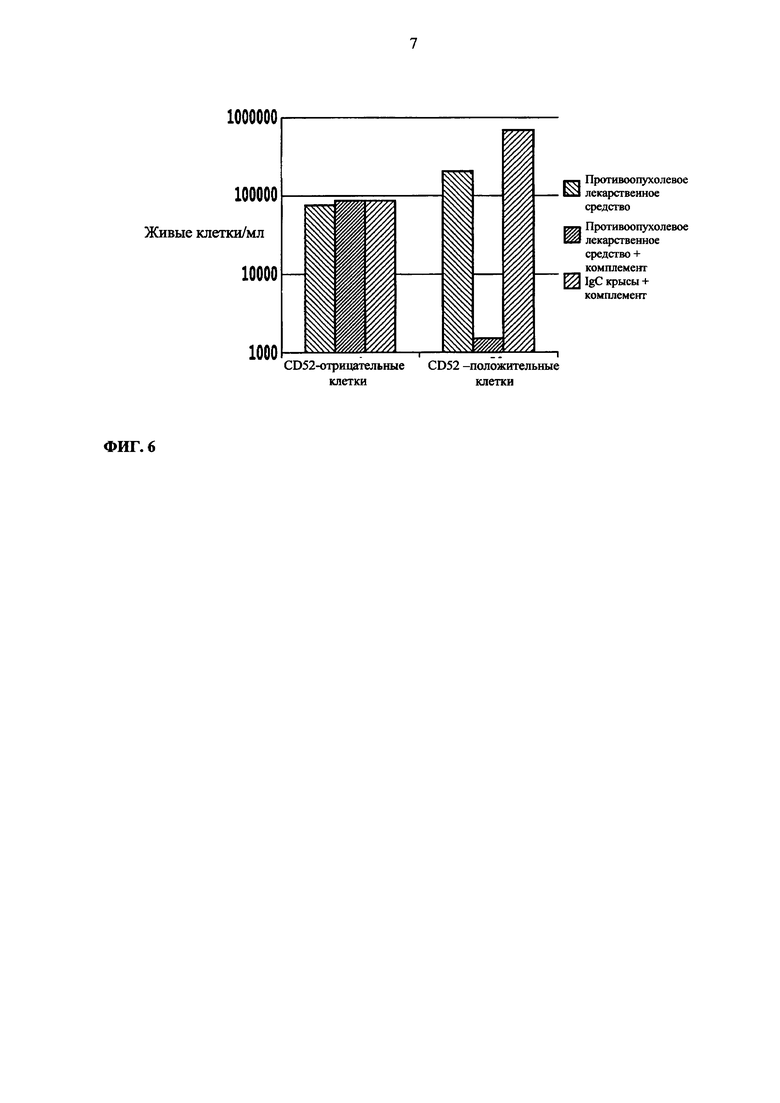
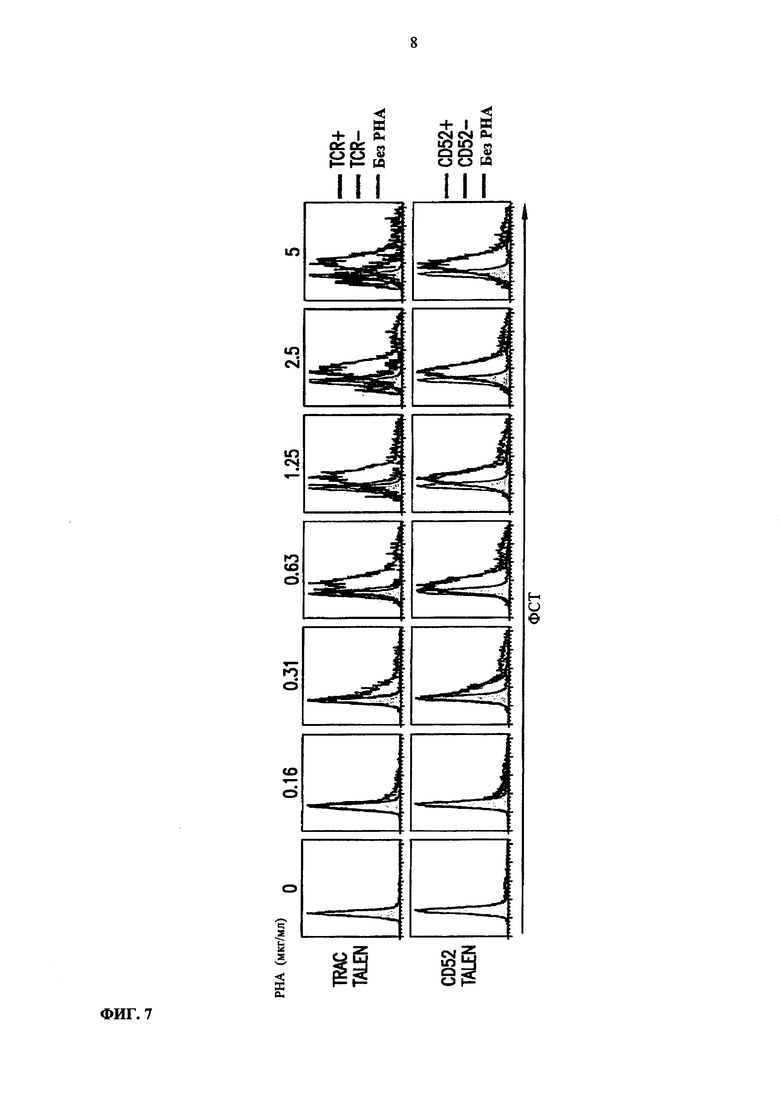
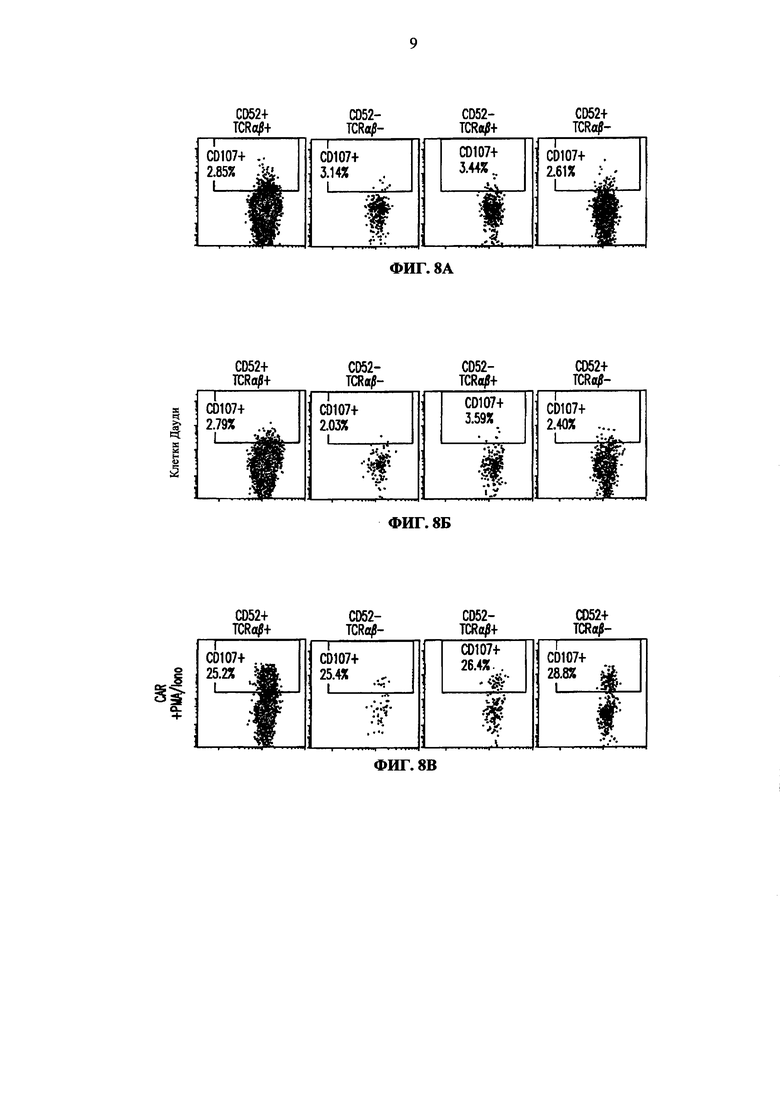
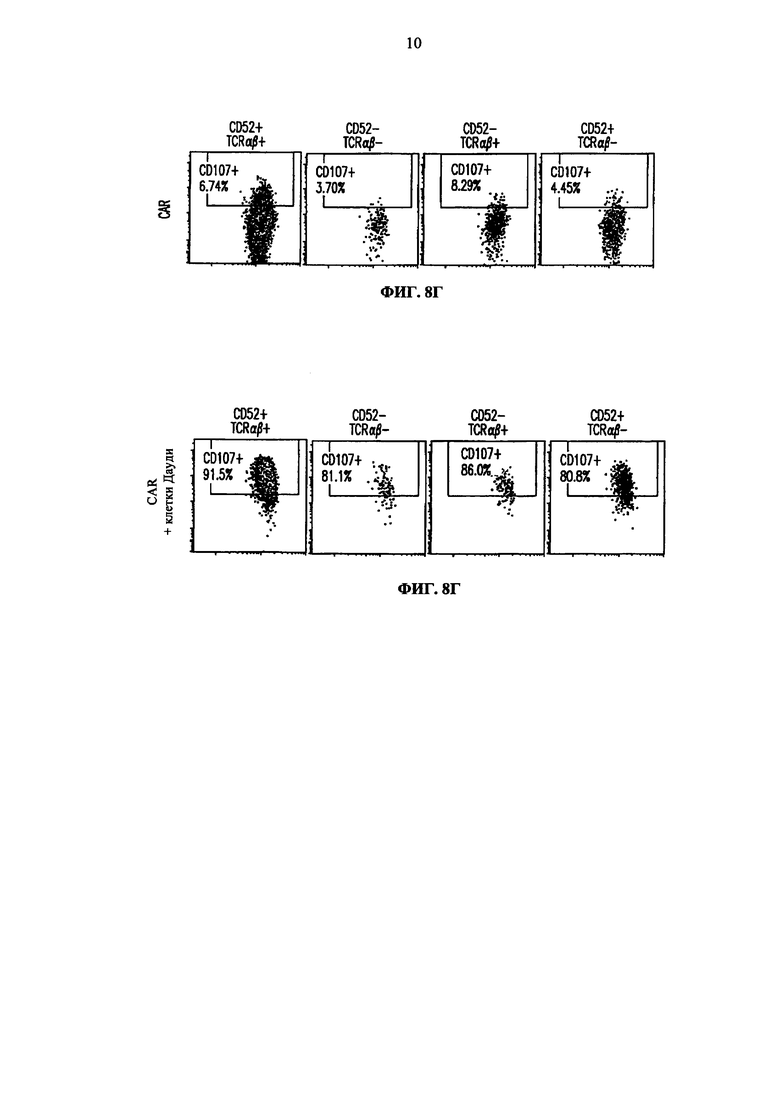

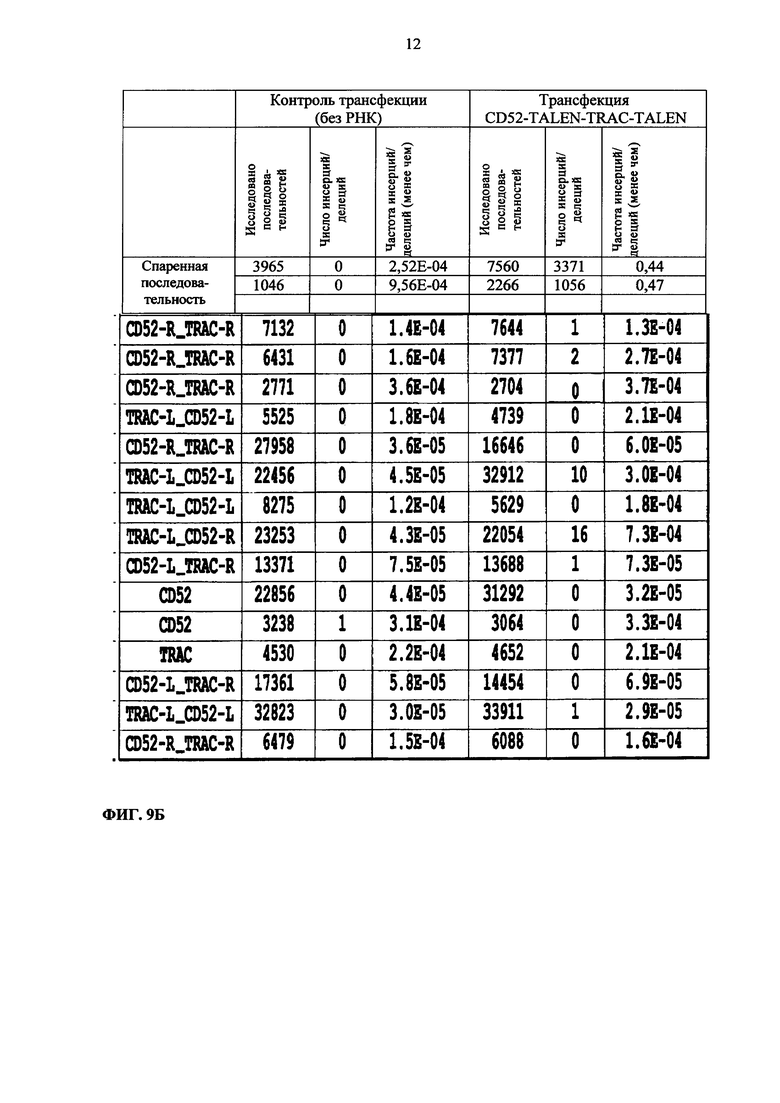
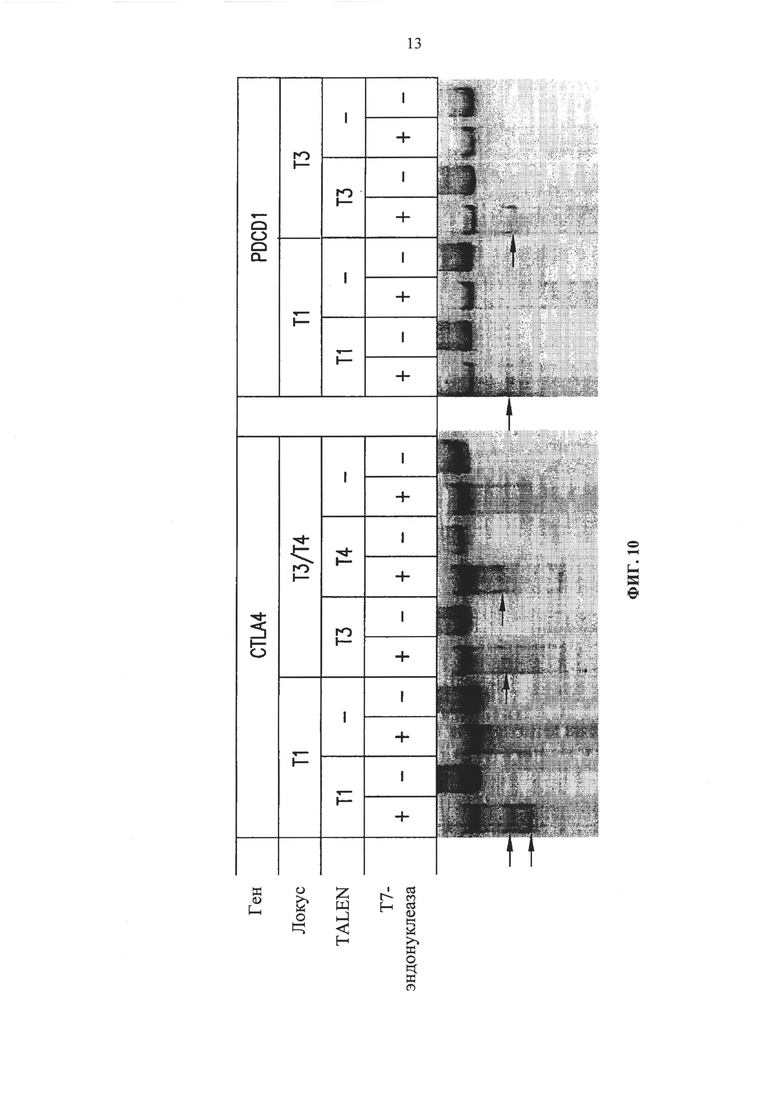
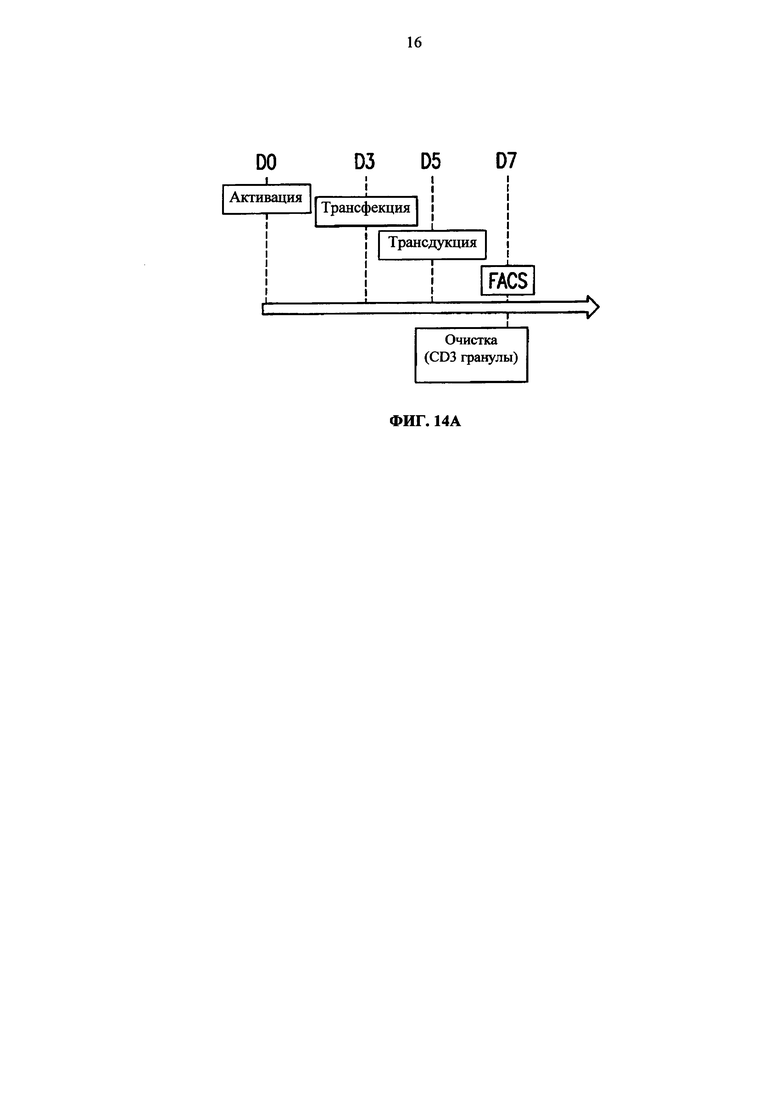
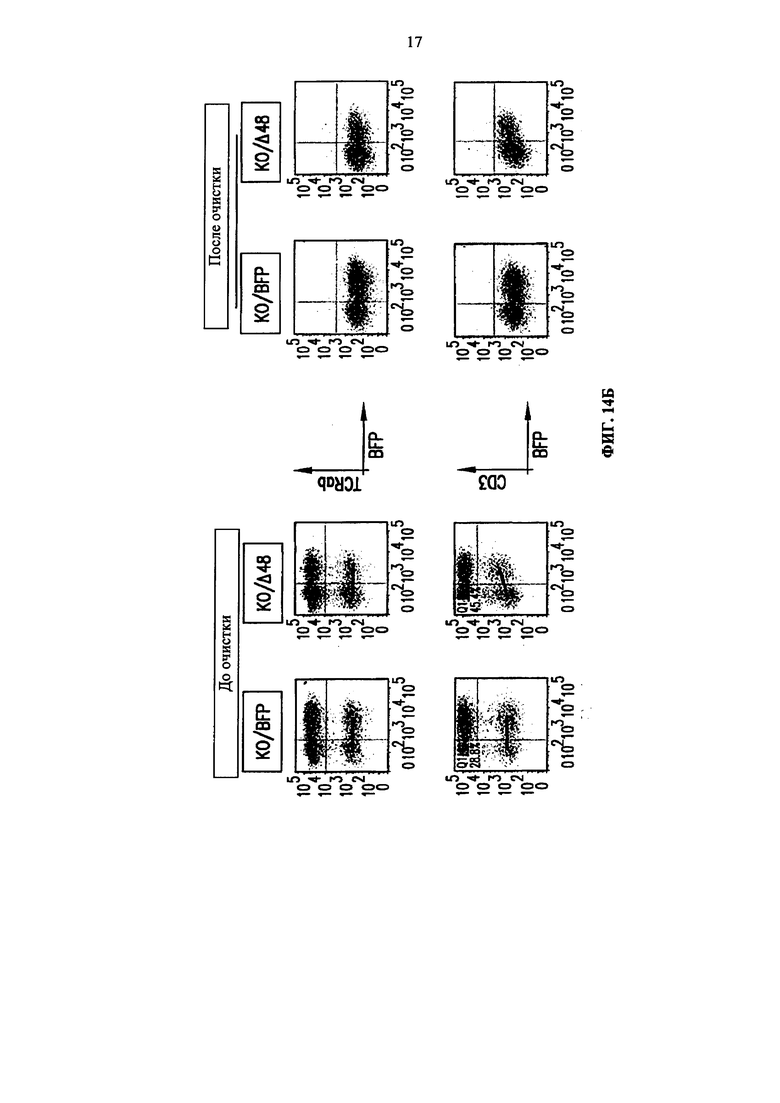
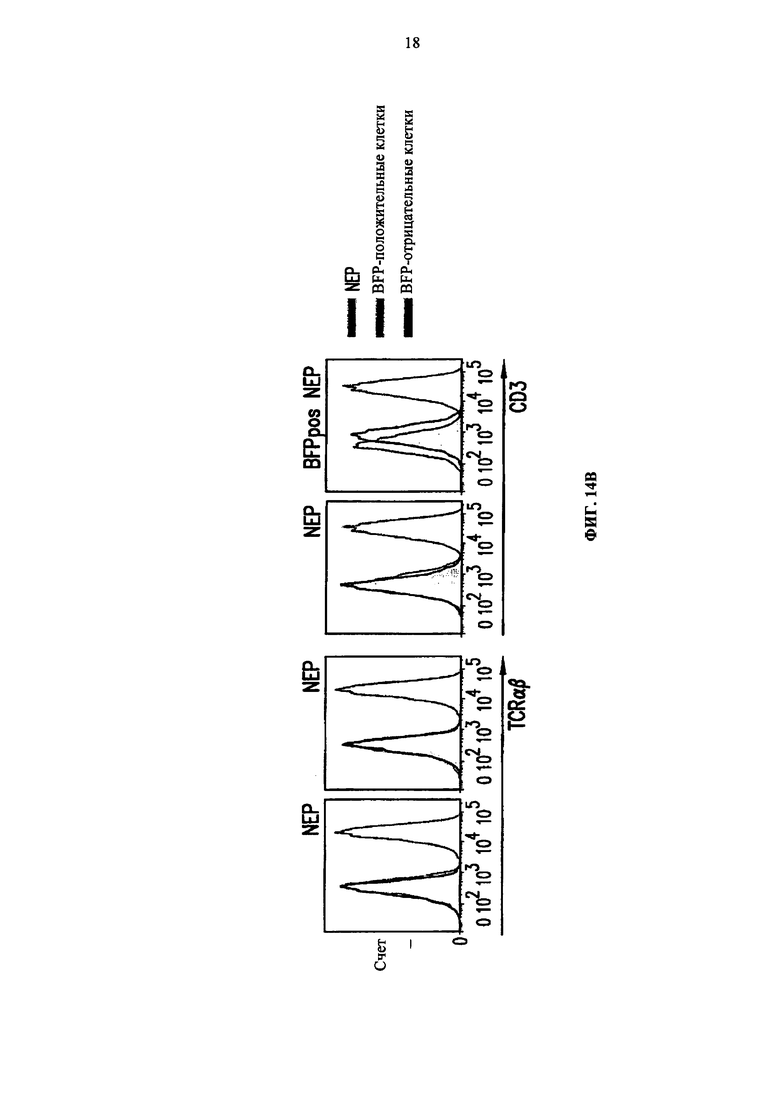
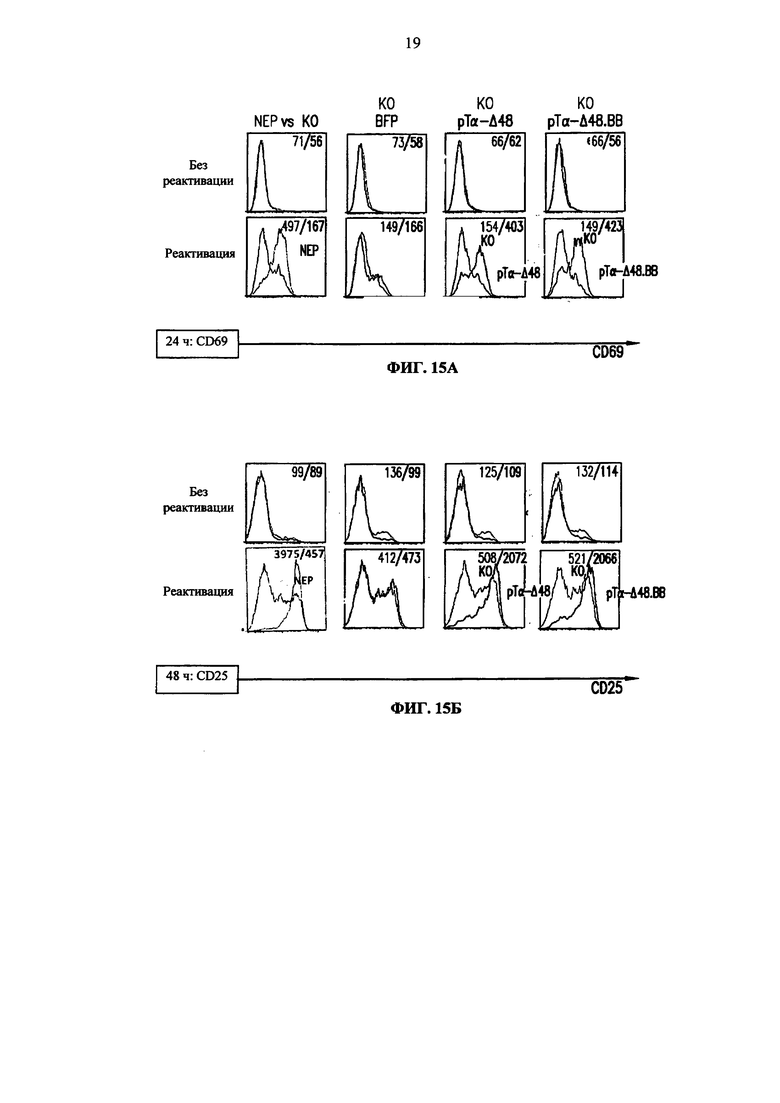
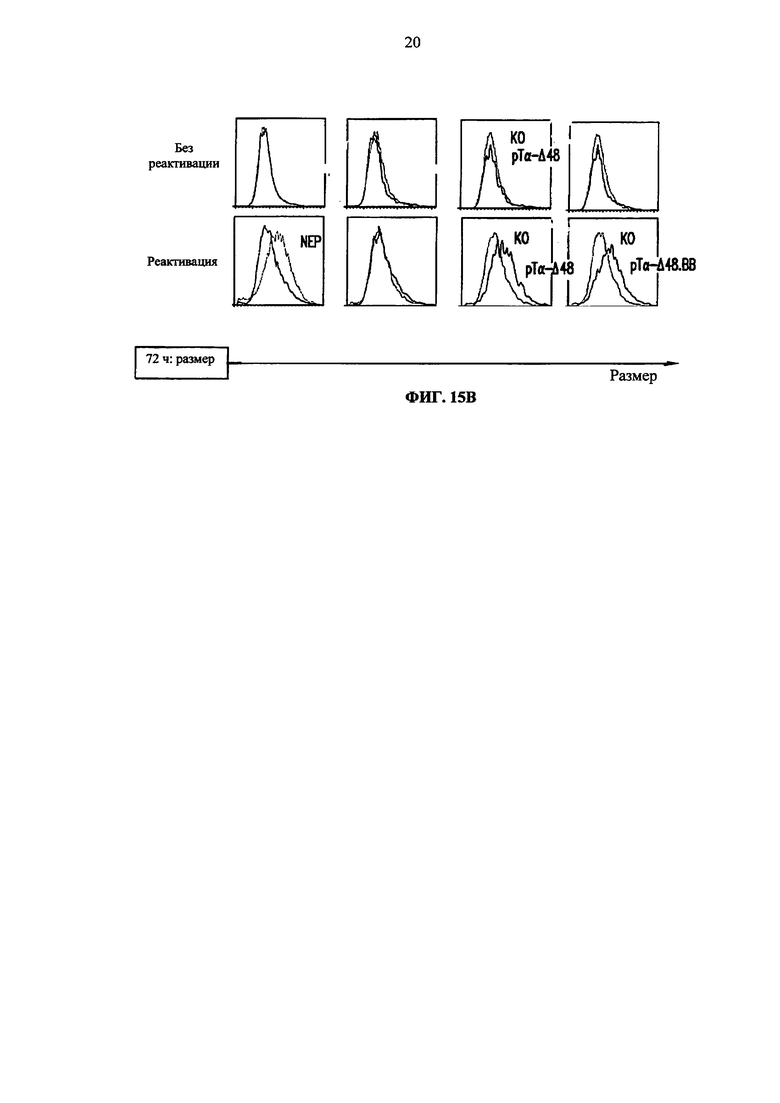
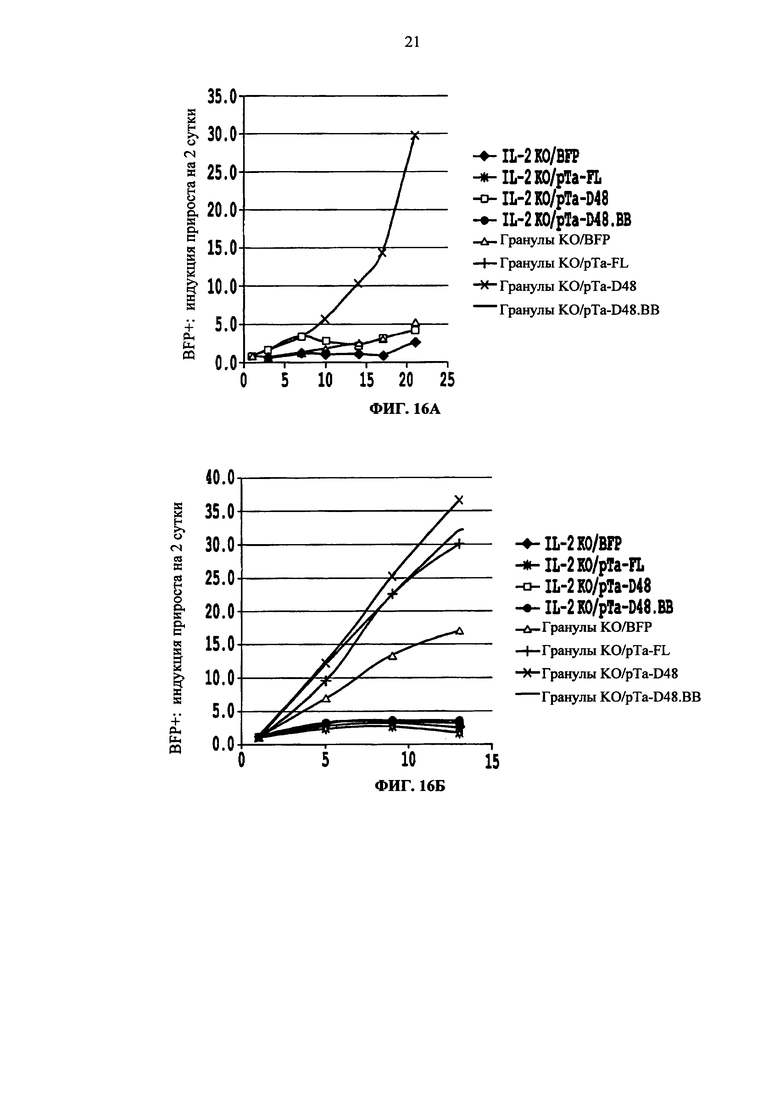
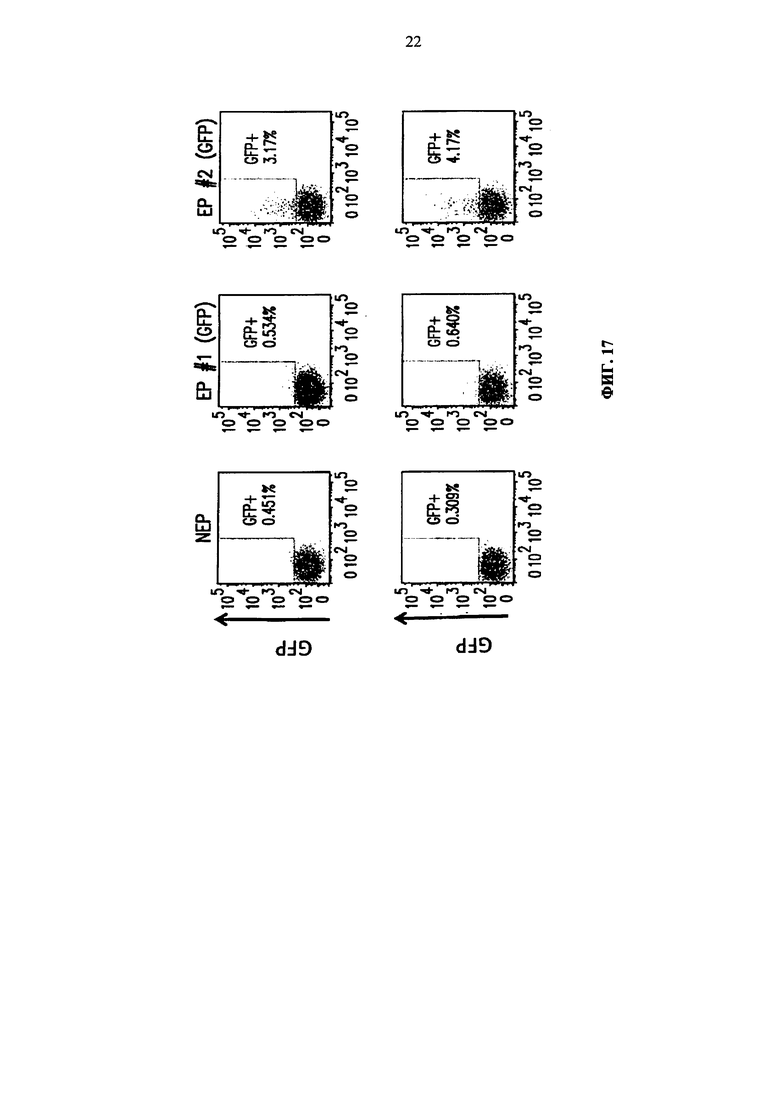
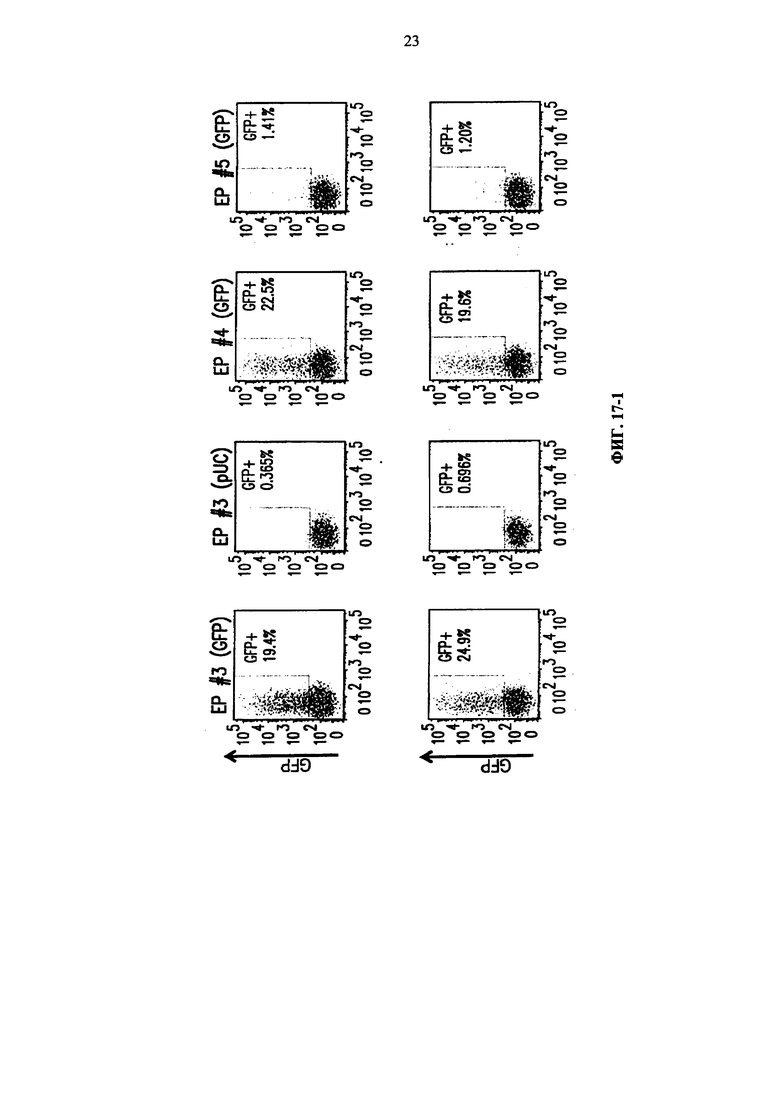
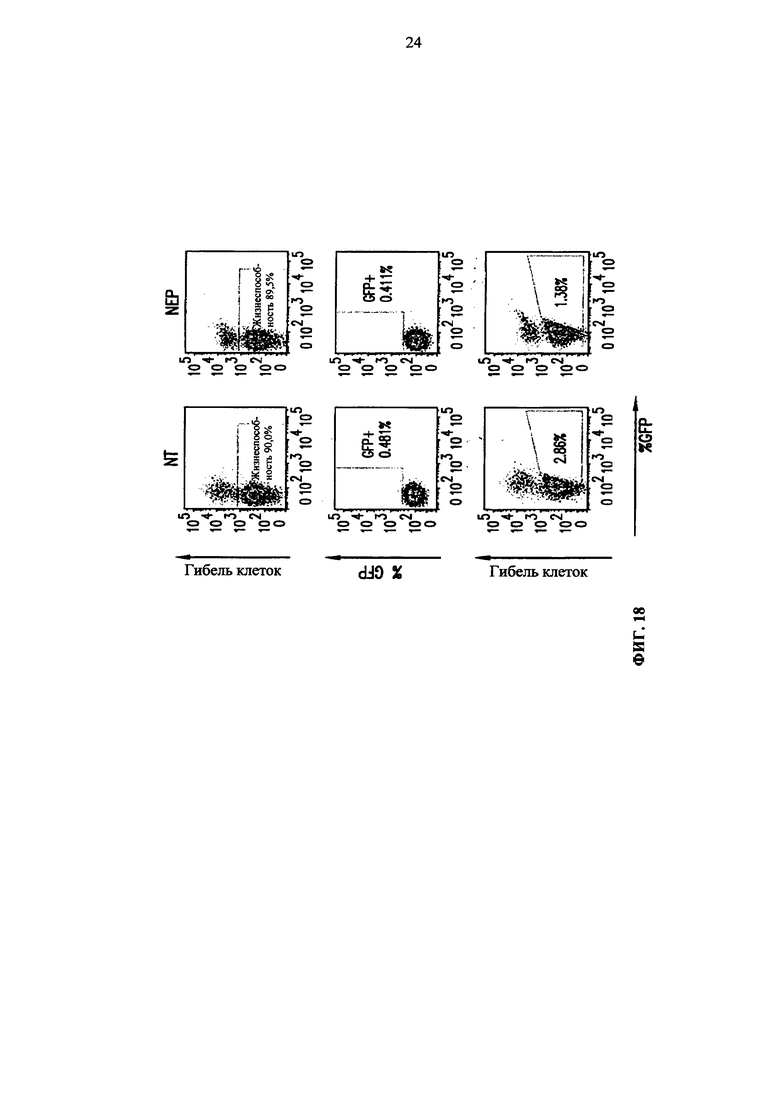
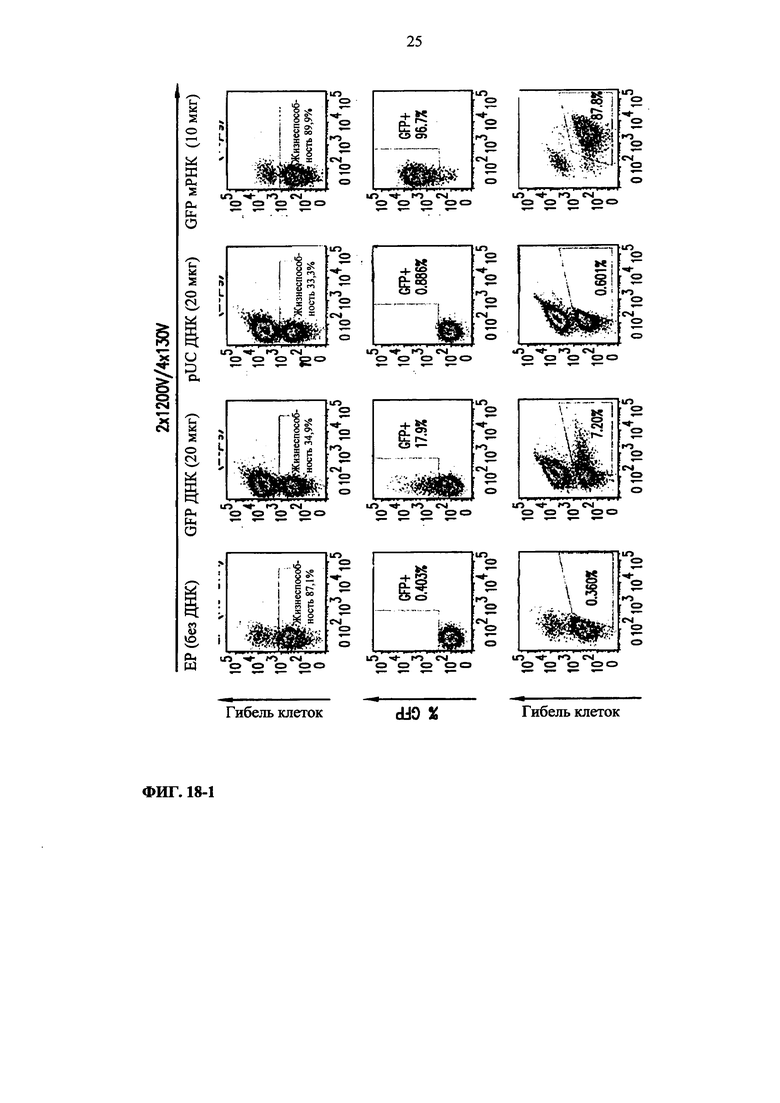
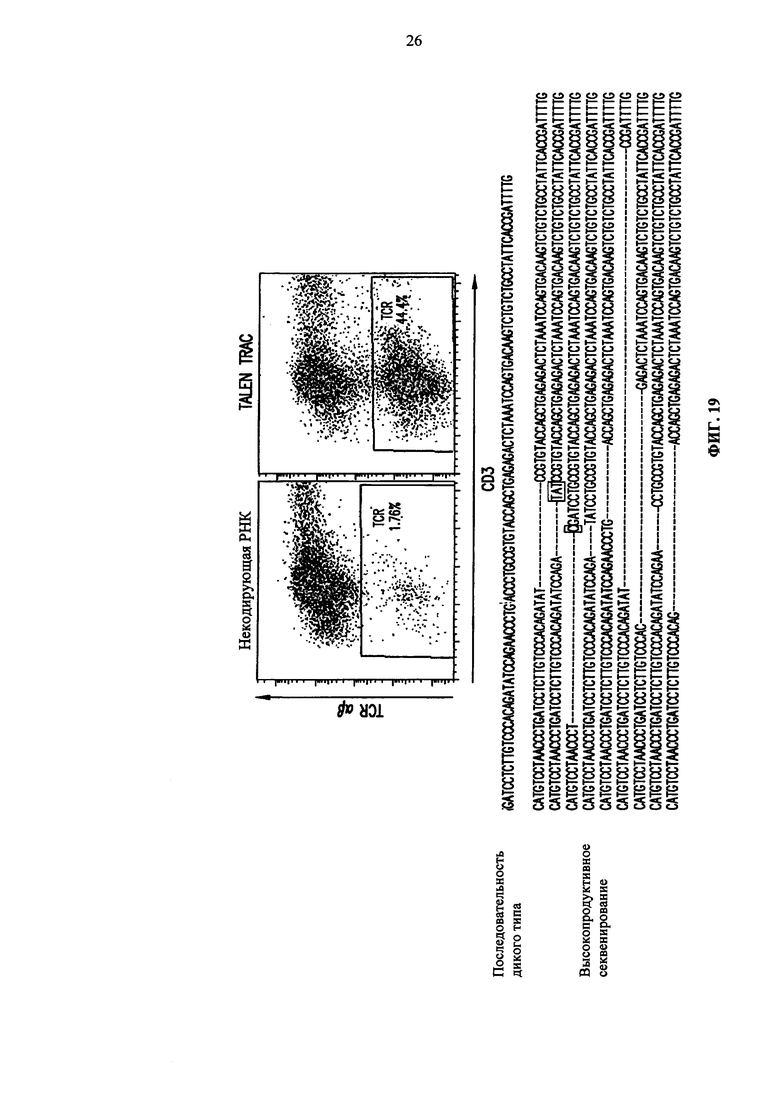
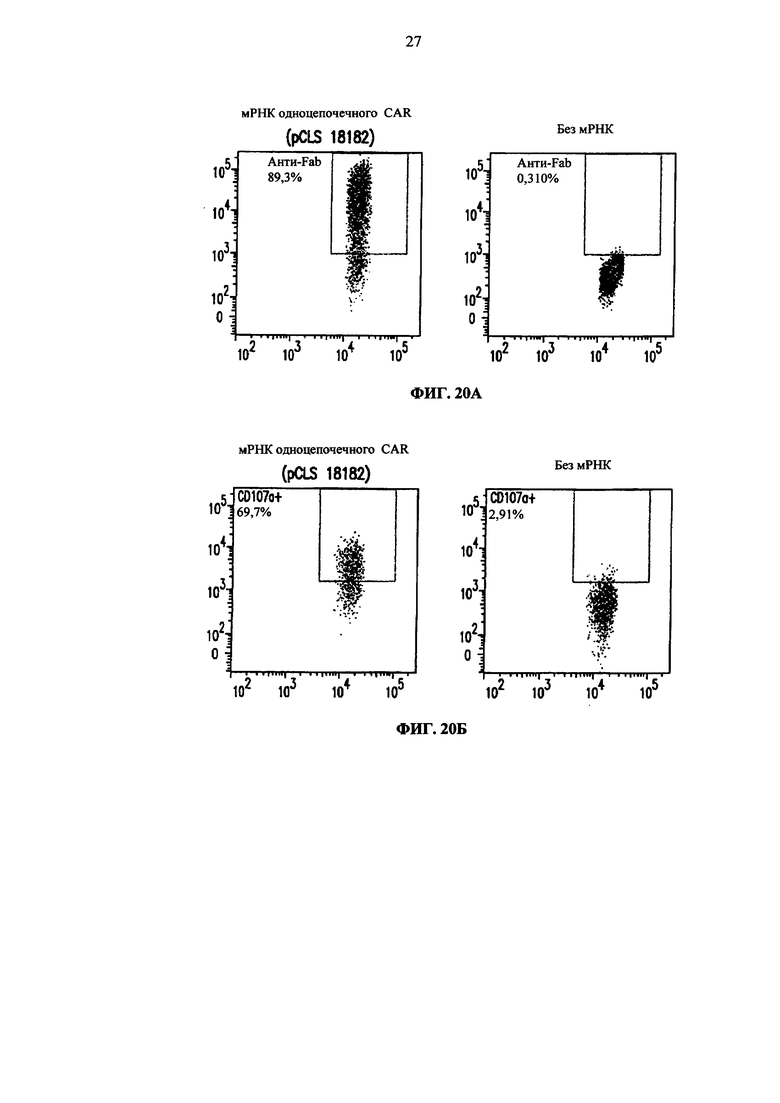
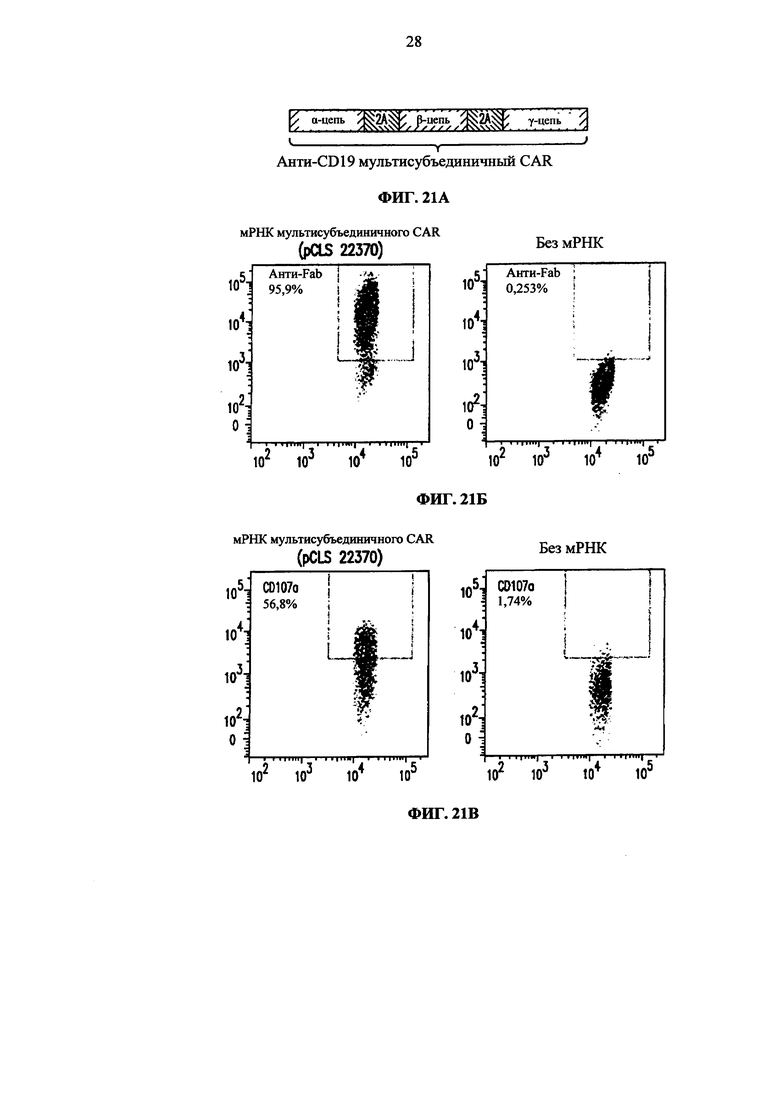
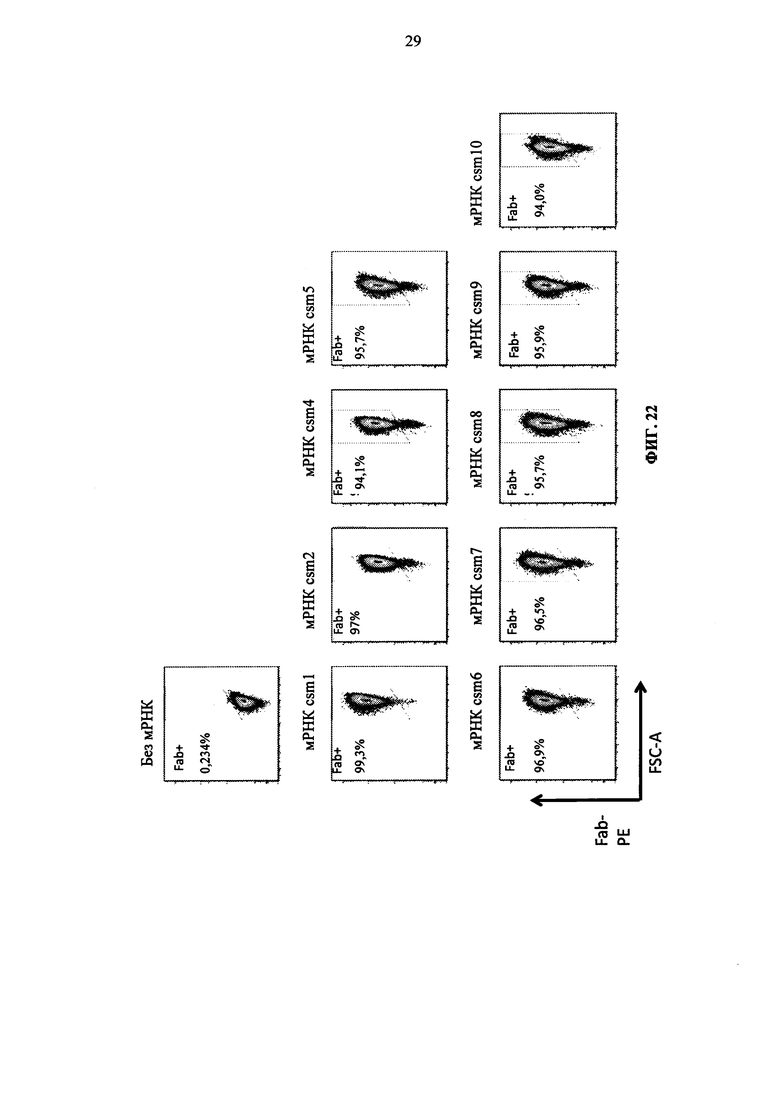
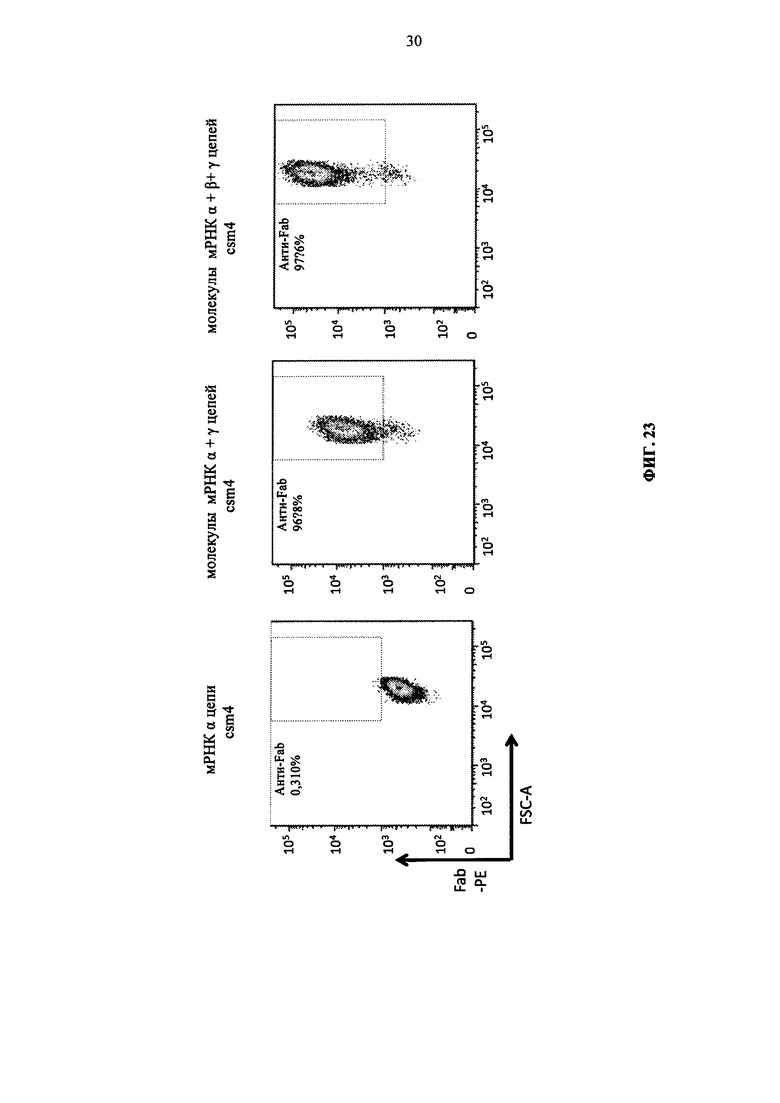
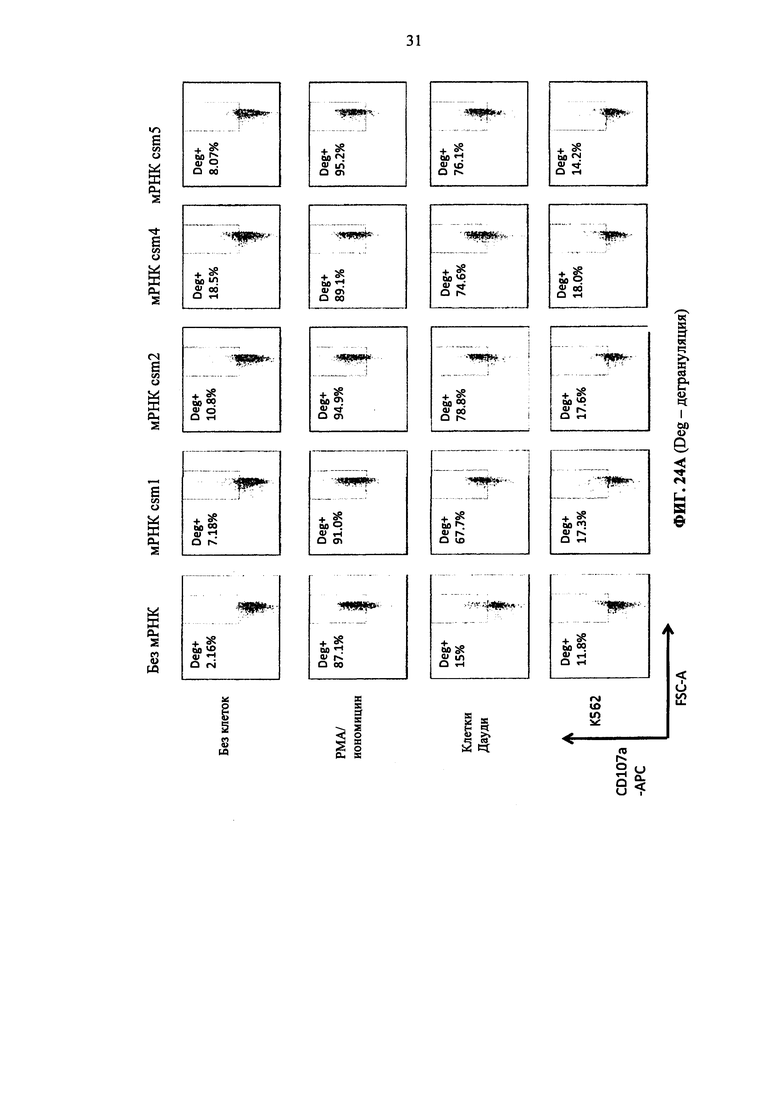
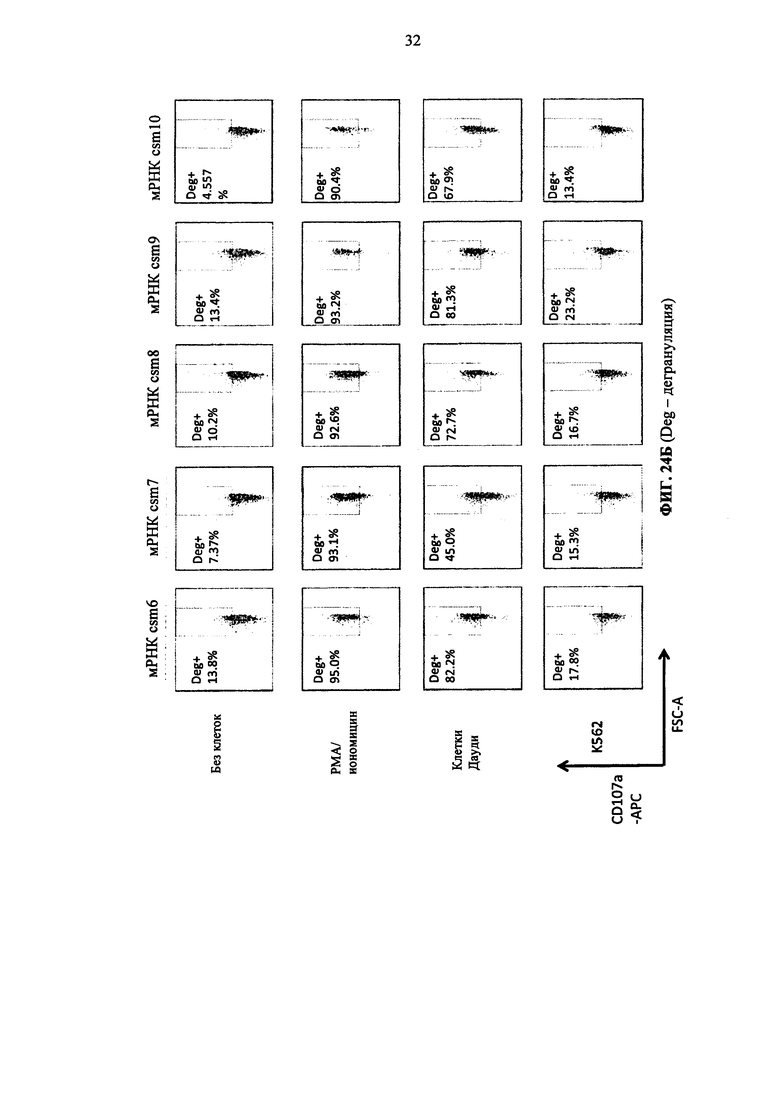
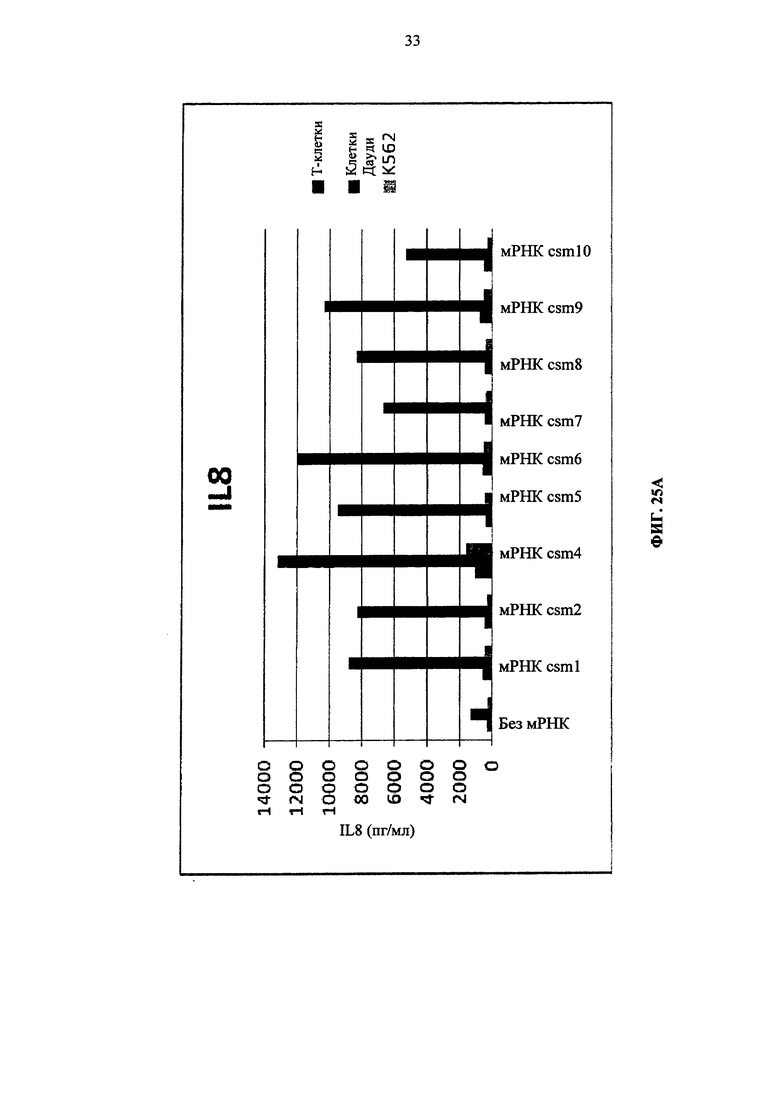
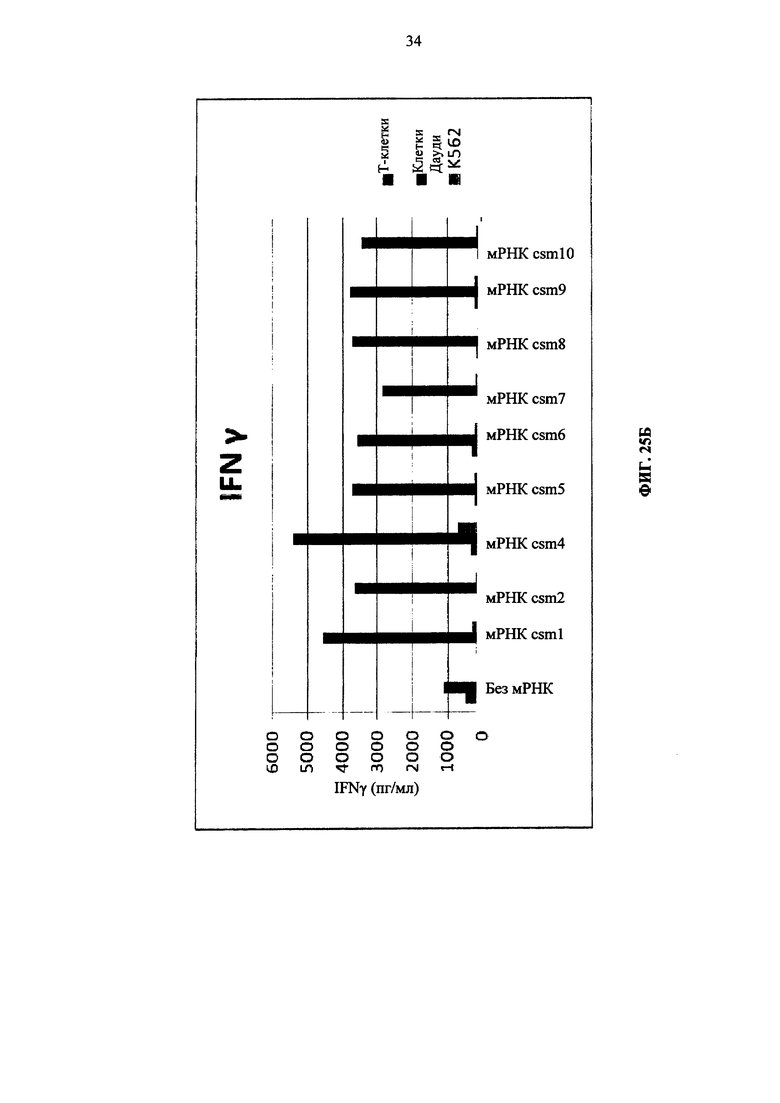
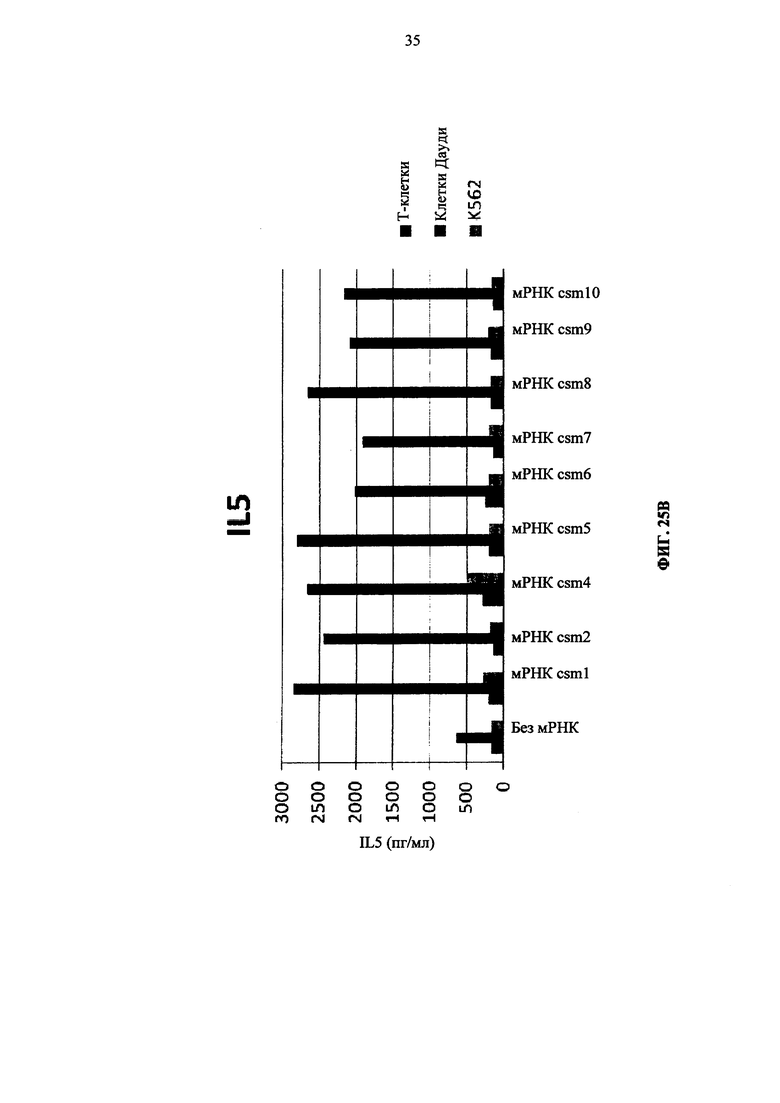
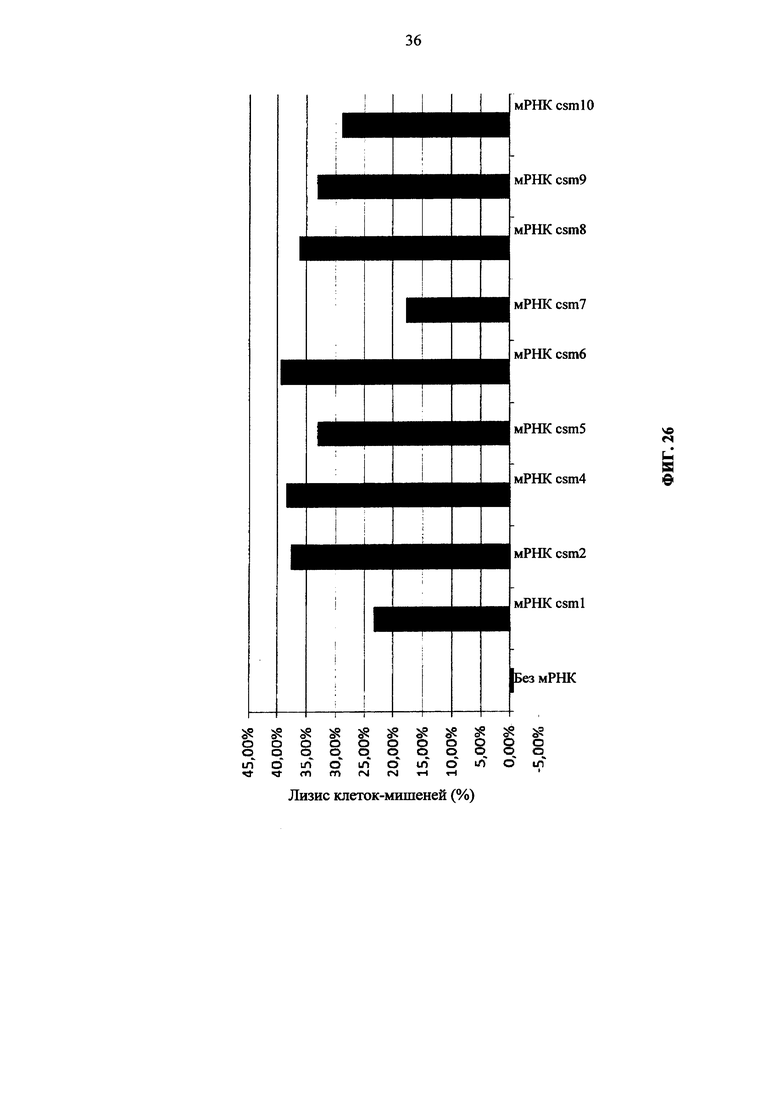
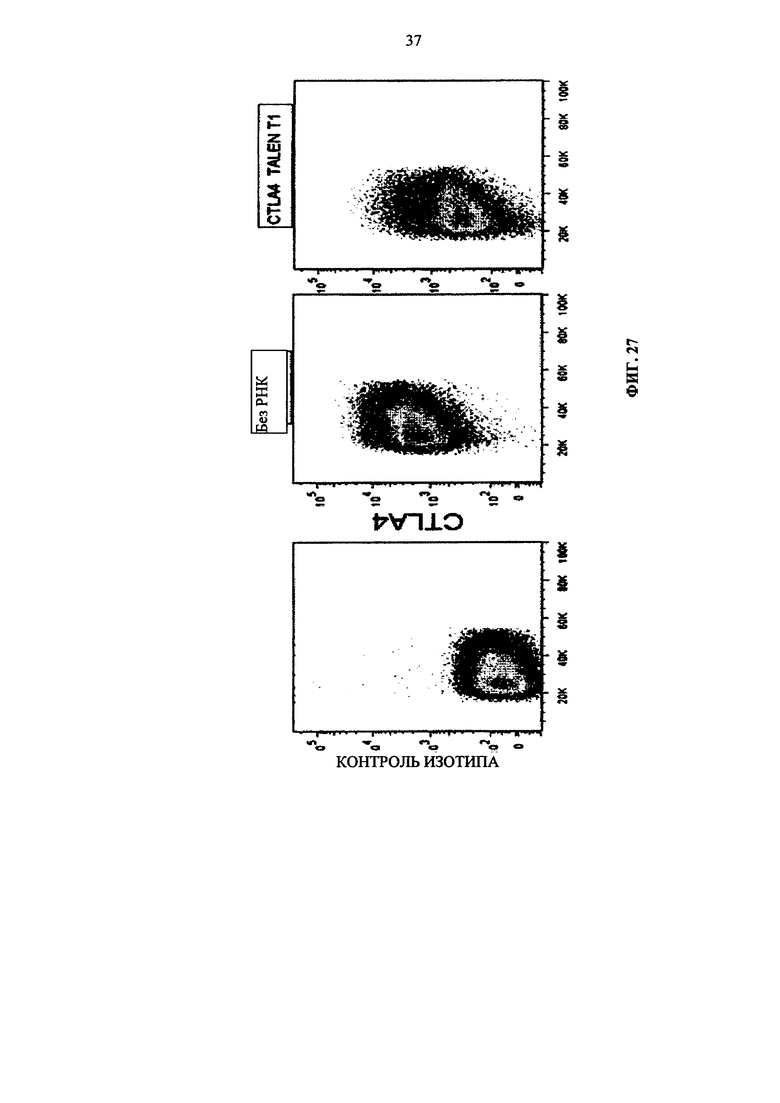
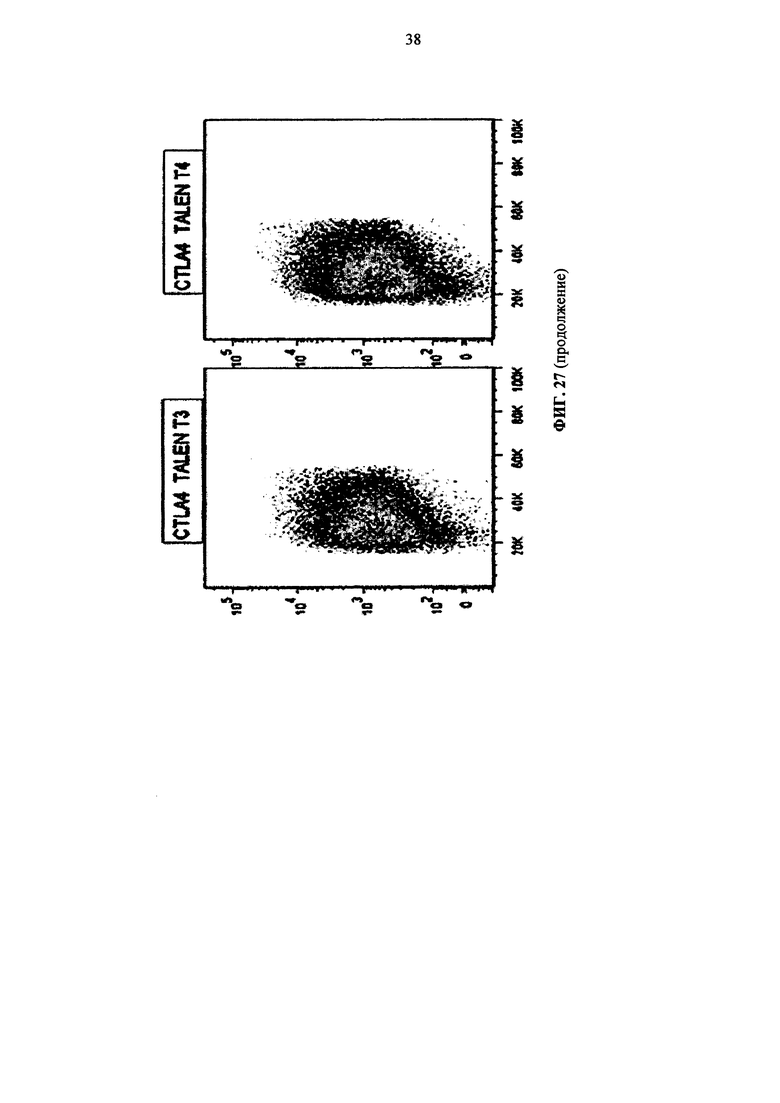
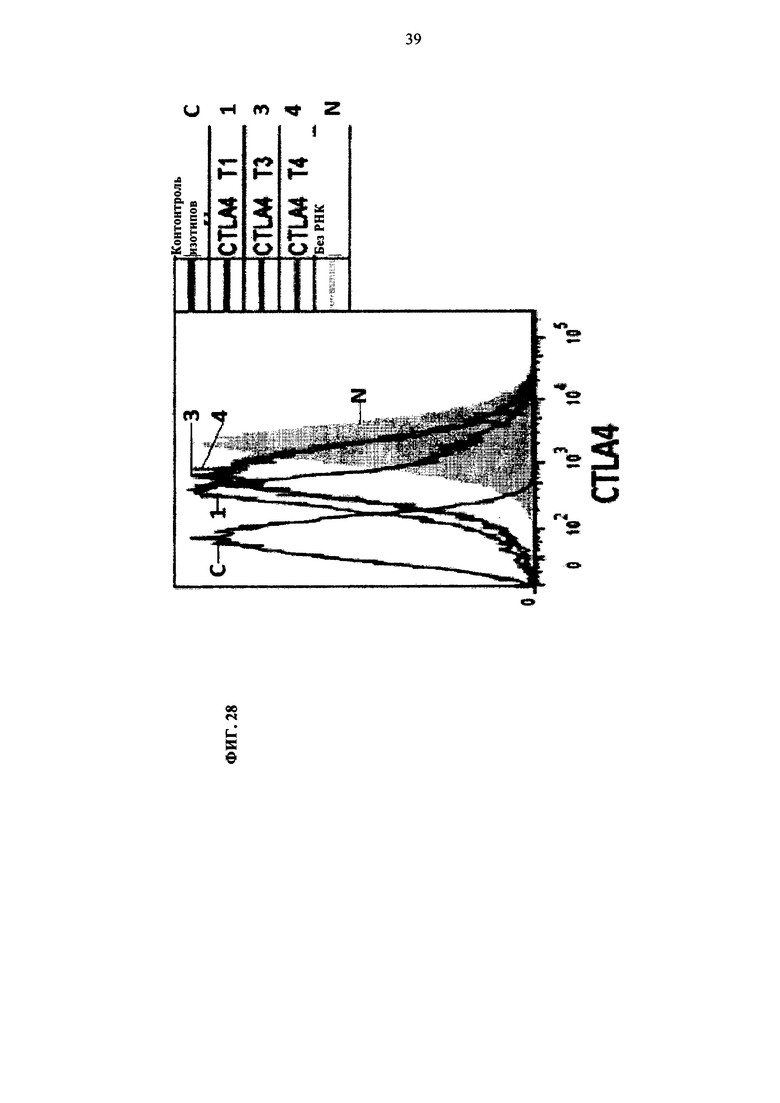
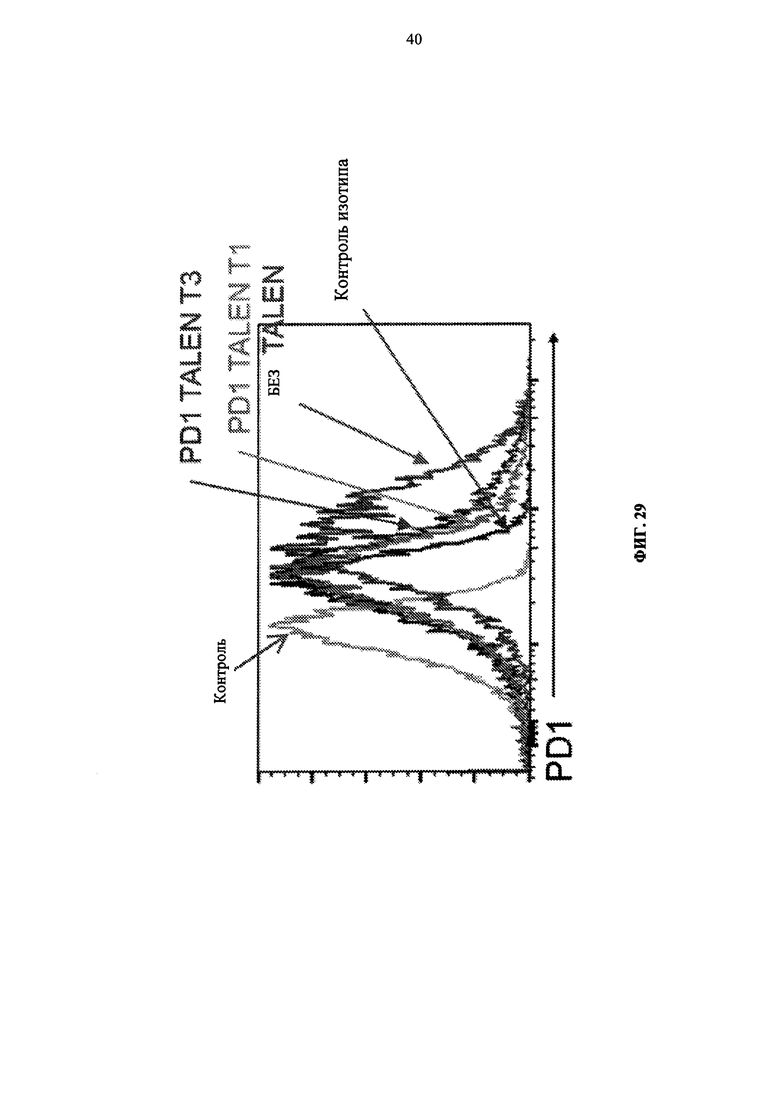
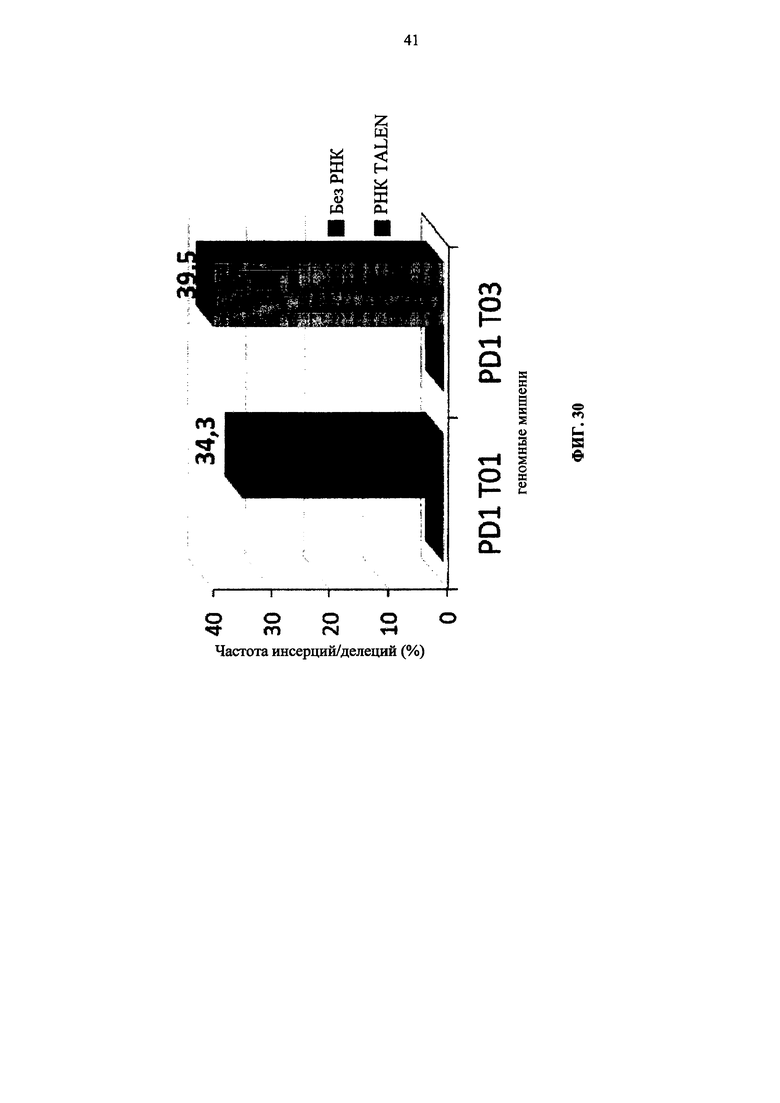
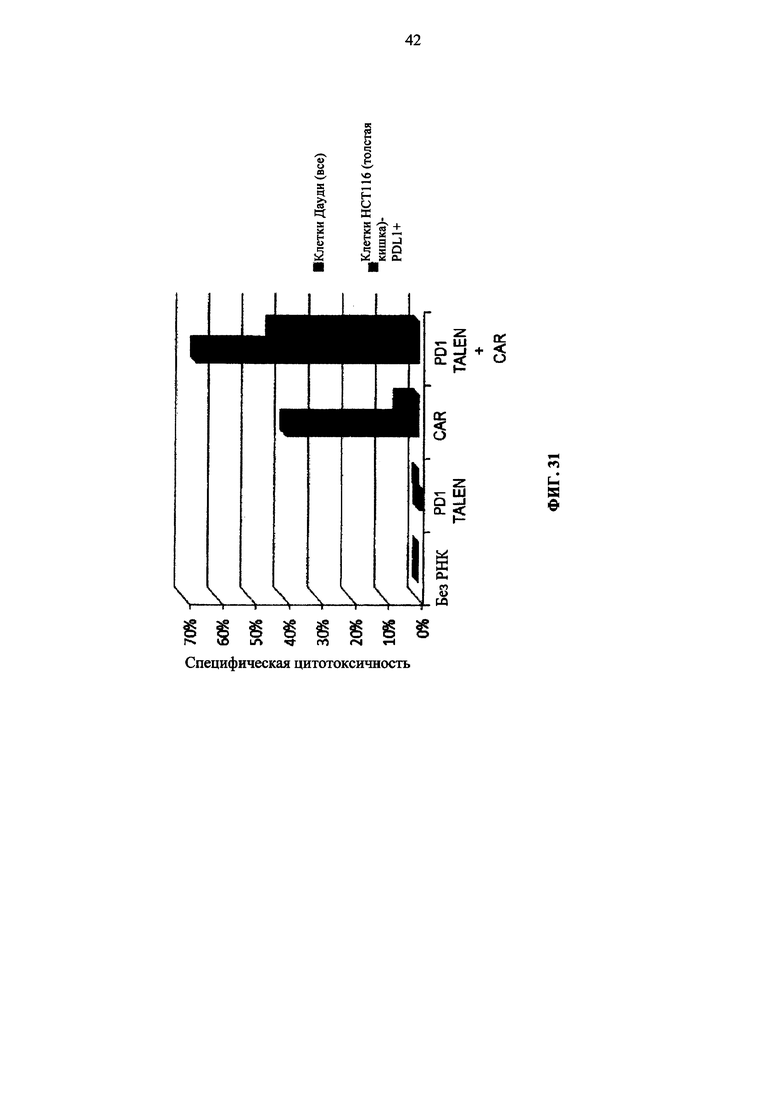
Данная группа изобретений относится к иммунологии. Предложен способ получения первичных Т-клеток, включающий генетическую модификацию Т-клеток с помощью редкощепящих эндонуклеаз, в частности TALE-нуклеаз, способных селективно инактивировать ген, кодирующий белок иммунной контрольной точки, и ген, кодирующий компонент Т-клеточного рецептора (TCR), и введение в Т-клетки нуклеиновой кислоты, кодирующей химерный антигенный рецептор против антигена к CD19. Описана получаемая таким способом Т-клетка, содержащая ее фармацевтическая композиция и способ лечения рака. Настоящее изобретение может найти применение в адоптивной иммунотерапии. 4 н. и 13 з.п. ф-лы, 31 ил., 15 табл., 5 пр.
1. Способ получения первичной Т-клетки/первичных Т-клеток для иммунотерапии при лечении рака, экспрессирующего CD19, включающий:
(а) получение и активацию Т-клеток из культур первичных клеток или образца крови;
(б) генетическую модификацию указанных Т-клеток с помощью введения в указанную Т-клетку редкощепящих эндонуклеаз, способных селективно инактивировать за счет расщепления ДНК по меньшей мере два гена:
- один ген, кодирующий белок иммунной контрольной точки, и
- один ген, кодирующий компонент Т-клеточного рецептора (TCR),
(в) введение в Т-клетки, полученные на стадии (б), нуклеиновой кислоты, кодирующей химерный антигенный рецептор против антигена к CD19 и экспрессирующей указанный химерный рецептор, и
(г) размножение указанных клеток, полученных на стадии (в).
2. Способ по п. 1, в которых указанный по меньшей мере один ген, кодирующий иммунную контрольную точку, выбран из группы, состоящей из CTLA4, РРР2СА, РРР2СВ, PTPN6, PTPN22, PD1, CD223, HAVCR2, BTLA, CD160, TIGIT, CD96, CRTAM, LAIR1, SIGLEC7, SIGLEC9, CD244, TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT, FOXP3, PRDM1, BATF, GUCY1A2, GUCY1A3, GUCY1B2 и GUCY1B3.
3. Способ по п. 1 или 2, в котором указанный ген, кодирующий иммунную контрольную точку, представляет собой PD1 и/или CTLA-4.
4. Способ по любому из пп. 1-3, где указанная по меньшей мере одна редкощепящая эндонуклеаза кодируется мРНК.
5. Способ по п. 4, который включает интродукцию одной редкощепящей эндонуклеазы или нескольких редкощепящих эндонуклеаз в указанную клетку на стадии (б) путем электропорации РНК.
6. Способ по любому из пп. 1-5, в котором по меньшей мере одной указанной редкощепящей эндонуклеазой является TALE-нуклеаза.
7. Способ по п. 6, в котором по меньшей мере одна указанная TALE-нуклеаза направлена против одной из генных целевых последовательностей PD1, выбранной из SEQ ID NO: 77 и SEQ ID NO: 78.
8. Способ по п. 6, в котором по меньшей мере одна TALE-нуклеаза направлена против одной из генных целевых последовательностей CTLA-4, выбранной из SEQ ID NO: 74 и SEQ ID NO: 76.
9. Способ по п. 1, где указанный химерный антигенный рецептор представляет собой мультицепочечный химерный антигенный рецептор.
10. Способ по п. 9, где указанный CAR содержит по меньшей мере один преобразующий сигнал домен, где указанный преобразующий сигнал домен представляет собой CD137 (4-1ВВ).
11. Способ по любому из пп. 1-10, где указанный CAR включает аминокислотную последовательность SEQ ID NO: 73.
12. Способ по п. 1, где указанная Т-клетка/указанные Т-клетки является/являются человеческой клеткой/человеческими клетками.
13. Выделенная Т-клетка, полученная способом по любому из пп. 1-12, для получения иммунотерапевтического агента для лечения рака, экспрессирующего CD19 антиген, у которой по меньшей мере два гена были инактивированы, при этом первый ген кодирует белок иммунной контрольной точки и второй ген кодирует компонент Т-клеточного рецептора (TCR) и дополнительно содержит экзогенную полинуклеотидную последовательность, кодирующую химерный антигенный рецептор, направленный против CD19 антигена.
14. Выделенная Т-клетка по п. 13 для применения в качестве лекарственного средства для лечения рака с помощью иммунотерапии.
15. Выделенная Т-клетка по п. 13 или 14 для лечения лимфомы или лейкемии.
16. Фармацевтическая композиция, включающая по меньшей мере одну выделенную Т-клетку по любому из пп. 13-15, для лечения рака, ассоциированного с CD19, с помощью иммунотерапии.
17. Способ лечения рака, ассоциированного с CD19, у пациента с помощью иммунотерапии, включающий:
(а) получение популяции модифицированных Т-клеток способом по любому из пп. 1-12 и
(б) введение указанных модифицированных Т-клеток указанному пациенту.
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| TORIKAI HIROKI et al | |||
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| WEBER J | |||
| "Immune checkpoint | |||

