Область техники
Изобретение относится к области медицины и может быть использовано в фармацевтической промышленности для производства лекарственного средства для лечения заболевания, сопровождающегося отеком макулы вследствие повышенной экспрессии фактора роста эндотелия сосудов - VEGF-A.
Уровень техники
Потеря зрения, связанная с патогенетическим механизмом неоангиогенеза, в основе которого лежит повышенная экспрессия VEGF-A и ряда других факторов роста, становится все более актуальной проблемой во всем мире, к числу таких заболеваний относятся влажная форма возрастной макулярной дегенерации, диабетическая ретинопатия, миопическая ХНВ, отек макулы вследствие окклюзии вен сетчатки и др. По данным Всемирной Организации Здравоохранения, возрастная макулярная дегенерация сетчатки (ВМД) является одной из наиболее частых причин слепоты и ухудшения зрения у лиц старше 59 лет в экономически развитых странах. Так, примерно у 10% пациентов в возрасте от 66 до 74 лет выявляется ВМД, у пациентов в возрасте от 75 до 85 лет частота данного заболевания возрастает до 30%. Возрастная макулярная дегенерация сетчатки подразделяется на два типа - экссудативная (влажная форма) и неэкссудативная (сухая форма). Сухая форма составляет около 90% случаев ВМД и характеризуется медленным снижением остроты зрения, появлением коллоидных образований (друз) на сетчатке и деградацией пигментного эпителия, известны случаи перехода сухой формы ВМД во влажную [Solomon SD, Lindsley K, Vedula SS, Krzystolik MG, Hawkins BS. Anti-vascular endothelial growth factor for neovascular age-related macular degeneration. Cochrane Database Syst Rev. 2014 Aug 29; 8:CD005139]. Влажная форма составляет около 10% ВМД, характеризуется быстрым развитием болезни, неоваскуляризацией сетчатки, геморрагиями и чаще приводит к потере зрения [Comprehensive Ophthalmology. Ed. by A.K. Khurana. New Age International (P) Ltd., Publishers, New Delhi, 2007]. Заболеваемость ВМД в РФ составляет 15:1000 населения и в 54,4% случаев является причиной серьезных нарушений зрения, а в 22,9% случаев - причиной слепоты [Агаркова Д.И. 2013. Математическая алгоритмизация диагностики возрастной макулярной дегенерации. Фундаментальные исследования №5, с. 17-22.; Rein DB, Wittenborn JS, Zhang X, Honeycutt AA, Lesesne SB, Saaddine J; Vision Health Cost-Effectiveness Study Group. Forecasting age-related macular degeneration through the year 2050: the potential impact of new treatments. Arch Ophthalmol. 2009 Apr; 127(4):533-40].
Диабетическая ретинопатия (ДР) является одним из основных осложнений при сахарном диабете (СД) первого и второго типа, число больных которым неуклонно с каждым годом нарастает. Так, по оценкам экспертов ВОЗ, в 1995 г. больных СД было 135 млн, а уже в 2001 году их число достигло 175,4 млн, к 2025 году это число возрастет до 300 миллионов, а к 2030 году достигнет 366 млн человек. Показатель распространенности ДР среди больных СД составляет 45,8%. Самой тяжелой формой поражения глаз является пролиферативная диабетическая ретинопатия, характеризующаяся неудержимым ростом патологических тканевых структур в сетчатке, которые появляются примерно у 10-40% всех больных СД, что, несмотря на лечение, быстро и неуклонно приводит к потере зрения в 2% случаев и в 10% случаев - к тяжелым нарушениям зрения. За последнее десятилетие отмечается рост частоты ДР, которая в настоящее время стала основной причиной необратимой слепоты, особенно у лиц трудоспособного возраста, что создает серьезные медико-социальные проблемы во многих странах мира.
Важнейшим патогенетическим фактором влажной формы ВМД, ДР и отека макулы вследствие окклюзии вен сетчатки является фактор роста эндотелия сосудов - VEGF-A (vascular endothelial growth factor). К семейству VEGF относятся несколько лигандов, отличающихся структурно и функционально: VEGF-A, VEGF-B, VEGF-C, VEGF-D и P1GF. VEGF-A является основным фактором, стимулирующим рост и новообразование сосудов, он обладает митогенным действием и способствует миграции эндотелиоцитов. VEGF-A, в свою очередь, объединяет несколько структурно родственных белков, являющихся продуктами альтернативно сплайсированной мРНК - VEGF165, VEGF121 и VEGF110. Основными изоформами VEGF-A, определяющими его активность, являются изоформы VEGF165 и VEGF121. VEGF-B также способствует ангиогенезу, VEGF-C участвует в лимфоангиогенезе во время эмбрионального развития, а также в функционировании дифференцированных лимфатических эндотелиоцитов взрослых. VEGF-D стимулирует рост лимфатического и сосудистого эндотелия. P1GF участвует в ангиогенезе, заживлении ран и воспалительном ответе, [Ohr М. and Kaiser P. Aflibercept in wet age-related macular degeneration: a perspective review. Therapeutic Advances in Chronic Disease. 2012. 3(4), 153-161].
Лекарственные средства, ингибирующие связывание VEGF-A с его рецепторами, такие как ранибизумаб (Lucentis®) и Eyelea® являются основными и наиболее эффективными лекарственными препаратами, применяемыми для терапии влажной формы ВМД, миопической ХНВ, диабетической ретинопатии и отека макулы вследствие окклюзии вен сетчатки. Интравитреальное (ИВТ) применение таких препаратов не только замедляет развитие этих заболеваний, но может и несколько улучшить зрение [BLA 125156; Summary Review 125387 Orig1s000.]. Первым препаратом для анти-VEGF терапии в виде интравитреальных инъекций, сертифицированным в России для применения в офтальмологии, был ранибизумаб, совершивший настоящую революцию в лечении ВМД и ставший «золотым стандартом». В июне 2006 года он был утвержден американским агентством по контролю за лекарственными средствами (FDA) как уникальное средство для лечения возрастной макулярной дегенерации, а в 2008 году был зарегистрирован и в России.
Хотя существующие в данное время препараты для ИВТ инъекций обладают высокой эффективностью, но сама техника выполнения данной манипуляции зачастую сопровождается значительными осложнениями со стороны органа зрения для пациентов, приводим выдержку из инструкции по применению луцентиса: «очень часто - интраокулярное воспаление, воспаление стекловидного тела, отслойка стекловидного тела, кровоизлияние в сетчатку глаза, нарушения зрения, боль в глазу, деструкция стекловидного тела, кровоизлияние в конъюнктиву, раздражение глаза, ощущение постороннего тела в глазу, повышенное слезоотделение, блефарит, сухость глаз, гиперемия глаза, ощущения зуда в глазу, повышение внутриглазного давления; часто - дистрофия сетчатки глаза, нарушения функций сетчатки глаза, отслаивание сетчатки глаза, разрыв сетчатки, отслаивание пигментного эпителия сетчатки, отрыв пигментного эпителия сетчатки, снижения остроты зрения, кровоизлияние в стекловидное тело, нарушение функции стекловидного тела, увеит, ирит, иридоциклит, катаракта, субкапсулярная катаракта, помутнение задней капсулы, точечный кератит, повреждение роговицы, воспаления передней камеры глаза, затуманенное зрение, геморрагии в месте инъекции, кровоизлияние в глаз, конъюнктивит, аллергический конъюнктивит, выделения из глаза, фотопсия, фотофобия, ощущение дискомфорта в глазу, отек век, боль в веке, гиперемия конъюнктивы, слепота, эндофтальмит, ползучая язва роговицы, кровотечение в переднюю камеру глаза, кератопатия, спайки радужки, отложения на роговице, отек роговицы, образование линий растяжения (стрии) на роговице, боль в участке инъекции, покраснение в участке инъекции, повышенная чувствительность глаза, раздражение века». Вследствие этого современная тенденция компаний-разработчиков новых антиангиогенных лекарственных средств для ИВТ инъекций - создание препаратов с увеличенным временем полувыведения из стекловидного тела, что позволило бы снизить частоту ИВТ инъекций, тем самым снизив вероятность возникновения нежелательных побочных явлений и повысив качество жизни пациентов, принимающих данные препараты [Wells J. et. Al. 2016. Two-Year Results from a Comparative Effectiveness Randomized Clinical Trial. Ophthalmology, 1-9.].
Луцентис® (ранибизумаб) является Fab-фрагментом гуманизированного моноклонального антитела против VEGF-A человека (IgG1(κ)) и эффективно блокирует взаимодействие основных изоформ VEGF-A - VEGF165, VEGF121 и VEGF110 с рецепторами VEGF первого и второго типов, [Hutton-Smith L.A. et. al. 2016. A Mechanistic Model of the Intravitreal Pharmacokinetics of Large Molecules and the Pharmacodynamic Suppression of Ocular Vascular Endothelial Growth Factor Levels by Ranibizumab in Patients with Neovascular Age-Related Macular Degeneration. Molecular Pharmaceutics. DOI:10.1021/acs.molpharmaceut.Sb00849]. Eyelea® является химерным белком на основе доменов рецепторов VEGF (3-го домена VEGFR1 и 2-го домена VEGFR2) и Fc-фрагмента антитела класса IgG1, [Wells J. 2016. Aflibercept, Bevacizumab, or Ranibizumab for Diabetic Macular Edema. Two-Year Results from a Comparative Effectiveness Randomized Clinical Trial. American Academy of Ophthalmology]. Молекулярная масса Луцентис® - 50 кДа, молекулярная масса Eyelea® - 115 кДа, включая олигосахаридные цепи. Луцентис® и Eyelea® назначаются в следующем режиме - вначале три инъекции подряд (1 раз в месяц), далее - по показаниям. По данным, опубликованным в журнале американского общества офтальмологов, по результатам 2-х летних сравнительных рандомизированных исследований эффективности препаратов Eyelea®, Авастин® и Луцентис®, медиана количества инъекций на 2-й год терапии (по показаниям) составила 5, 6 и 6, соответственно, а в течение 2-х лет - 15, 16 и 15, соответственно, при сопоставимой эффективности препаратов, [Wells J. et. Al. 2016. Two-Year Results from a Comparative Effectiveness Randomized Clinical Trial. Ophthalmology, 1-9].
Элиминация препаратов из стекловидного тела при интравитреальном введении происходит через переднюю камеру глаза или через сосудистую оболочку задней камеры глаза, при этом выведение высокомолекулярных соединений из стекловидного тела происходит, в основном, через переднюю камеру глаза по механизму диффузии с потоком водянистой влаги и, в значительно меньшей степени, через сосудистую оболочку глаза вследствие существования гематоофтальмического барьера. Скорость диффузии зависит от молекулярной массы молекулы. Hutton-Smith L. и соавторы разработали математическую механистическую модель фармакокинетики макромолекул при интравитреальном введении, основные выкладки которой хорошо согласуются с экспериментальными данными доклинических и клинических исследований ранибизумаба и бевацизумаба, в которой показали, что увеличение времени экспозиции макромолекул в стекловидном теле можно достичь увеличением массы молекулы [Hutton-Smith L.A. et.al. 2016. A Mechanistic Model of the Intravitreal Pharmacokinetics of Large Molecules and the Pharmacodynamic Suppression of Ocular Vascular Endothelial Growth Factor Levels by Ranibizumab in Patients with Neovascular Age-Related Macular Degeneration. Molecular Pharmaceutics. DOI: 10.1021/acs.molpharmaceut.Sb00849].
Препараты для интравитреального введения должны обладать низким системным влиянием. Системная доступность макромолекул при интравитреальном введении вследствие наличия гематоофтальмического барьера снижена, но тем не менее небольшая доля препарата (около 1%) поступает в кровь [BLA 125156, Summary Review 125387Orig1s000], для уменьшения системного влияния разработчики ранибизумаба использовали Fab-формат молекулы вследствие его быстрой элиминации из кровотока, поскольку Fab не взаимодействует с FcRn. Fc-фрагмент антител стабилизирует молекулы, увеличивает период их полувыведения из кровотока, поэтому данный подход широко используется при создании химерных белков [Czajkowsky, D., Shao, Z., Pleass, R. 2012. Fc-fusion proteins: new developments and future perspectives. EMBO Mol. Med., 4, 1015-1028.], в том числе и для препарата Eyelea®, [Wells J. 2016. Aflibercept, Bevacizumab, or Ranibizumab for Diabetic Macular Edema. Two-Year Results from a Comparative Effectiveness Randomized Clinical Trial. American Academy of Ophthalmology]. Известны и другие способы модификации белков, увеличивающие как молекулярную массу молекул и физические размеры (гидродинамический объем), так и время их жизни в плазме крови. [Therapeutic Proteins: Strategies to Modulate Their Plasma Half-Lives, First Edition. Edited by Roland Kontermann. © 2012 Wiley-VCH Verlag GmbH & Co. KGaA.]. Одной из таких модификаций является химическая модификация белков с помощью полиэтиленгликолей (ПЭГ), [Veronese, F., Pasut, G. 2005. PEGylation, successful approach to drug delivery. Drag Discovery Today, v/10, №21, 1451-58]. Однако неблагоприятными свойствами пэгилированных белков являются химическая неоднородность модифицированных молекул вследствие неоднородности структуры модифицирующего агента и не избирательном (случайном) механизме этого процесса, а также накопление в организме производных ПЭГ, не подверженных биодеградации.
Существуют и другие способы модификации белков, которые основываются не на химической модификации, а на создании генетических конструкций (ГК), в результате трансляции которых экспрессируются целевые белки, слитые с модифицирующими полипептидами, придавая исходным белкам желаемые свойства. К числу таких полипептидов относятся:
- искусственные полипептиды, в которых содержатся сайты N-гликозилирования и преобладают повторы из остатков глицина, аспарагиновой кислоты и глутаминовой кислоты [US 20090298762];
- полианионные полипептиды из остатков глутаминовой кислоты (84 и 173 остатка) [US 20020169125, AU 2002252429, WO 02077036];
- С-концевой пептид (СТР) β-субъединицы хорионического гонадотропина человека, содержащего 28 аминокислотных остатков, формирующих четыре сайта О-гликозилирования [US 20120015437]; Одна или несколько копий СТР, присоединяемых к С-концу молекул, существенно удлиняют время жизни терапевтических белков в плазме крови [US 2012035101, US 2012015437, US 2012004286, US 2011286967, US 2010317585, US 2010081614, US 5759818]. Использование СТР-технологии ограничено необходимостью получать модифицированные белки исключительно в клетках животных, обеспечивающих корректное О-гликозилирование;
- неструктурированные полипептиды.
Способ пролонгирования терапевтических белковых молекул с использованием неструктурированных полипептидов предполагает генетическую гибридизацию белков с высоко растворимыми, химически и иммунологически инертными неструктурированными полипептидами (НП). Такие НП в водных растворах при физиологических солевых, температурных и буферных условиях устойчивы к фолдингу (формированию третичной структуры) и поддерживают стабильное неупорядоченное состояние типа "клубок", гидродинамический объем которого зависит только от числа аминокислотных остатков в составе НП, собственного размера (диаметра) модифицируемого белка и конформационной гибкости НП. В этой связи НП способны опосредовать значительное увеличение гидродинамического объема коньюгированных белков, вызывающее ослабление почечной фильтрации и удлиняющее время жизни белков в плазме крови, [Binder U., Skerra А. 2012. Half - Life Extension of Therapeutic Proteins via Genetic Fusion to Recombinant PEG Mimetics. Therapeutic Proteins: Strategies to Modulate Their Plasma Half-Lives, First Edition. Edited by Roland Kontermann. © 2012 Wiley-VCH Verlag GmbH & Co. KGaA].
Использование НП для пролонгирования белков имеет преимущества перед химической модификацией с помощью ПЭГ, в частности:
- белки, модифицированные по этой технологии, могут быть экспрессированы и внутриклеточно, и в секретированном виде в различных типах клеток, включая бактерии и дрожжи, что позволяет значительно удешевить и ускорить их получение и очистку;
- модифицирующие НП обладают крайне низкой иммуногенностью и подобно ПЭГ способны экранировать чужеродные белки от распознавания иммунной системой;
- модифицирующие НП являются биоразлагаемыми, что значительно снижает риск появления токсичности или накопления в каких-либо органах (например, в почках или печени) или клетках (например, в макрофагах);
- генетически кодируемая последовательность НП гарантирует неизменность и точность модификации белка.
В основных чертах эти характерные особенности свойственны трем известным типам НП, способным пролонгировать действие белков до 100 раз:
1) XTEN-полипептидам, содержащим случайное чередование 6 аминокислотных остатков аланина, глутаминовой кислоты, глицина, пролина, серина и треонина [ЕР 2402754, US 20110312881, WO 2011123830, WO 2011123813, US 20110151433, US 20100323956, WO 2010144502, WO 2010091122, CA 2748314, WO 2007103515, US 7846445, US 7855279];
2) PAS-полипептидам, содержащим остатки пролина, аланина и серина [WO 2011144756, NZ 580670, US 20100292130, ЕР 2173890, WO 2008155134;
3) НАР-полипептидам, являющимися глицин-богатыми последовательностями [WO 2007103515], содержащими остатки глицина (G) и серина (S).
Показано пролонгирование действия ИФН за счет использования НП PAS, наименьший размер которых составлял около 200 аминокислотных остатков [WO 2008155134 A1]. Показано пролонгирование действия ИФН альфа-2 человека за счет использования НАР-полипептидов S(G4S)16 и S(G4S)20 [RU 2515913 С1].
Однако неструктурированные полипептиды не только увеличивают продолжительность жизни в крови терапевтического белка, но могут привести к экранированию активных сайтов и снижению их целевой биологической активности. Кроме того, в уровне техники показано увеличение продолжительности жизни терапевтического белка, модифицированного с помощью неструктурированного полипептида только в крови.
Таким образом, до сих пор сохраняется насущная потребность в решении задачи создания препарата фрагмента антитела с увеличенным временем экспозиции в стекловидном теле и высокой аффинностью связывания с мишенью.
Раскрытие сущности изобретения
Настоящее изобретение решает задачу создания препарата фрагмента антитела с увеличенным временем экспозиции в стекловидном теле и высокой аффинностью связывания с мишенью.
Для создания препарата с увеличенным временем экспозиции в стекловидном теле была взята за основу концепция математической механистической модели фармакокинетики препаратов при интравитреальном введении - увеличение массы терапевтического белка. Однако было решено отойти от традиционного подхода увеличения массы терапевтического белка путем модификации его с помощью НП, ПЭГ. На примере ранибизумаба авторами настоящего изобретения была исследована возможность увеличения массы терапевтического фрагмента антитела за счет добавления F(ab’)-фрагмента. В результате был получен генно-инженерный конструкт F(ab’)2-ранибизумаба для эукариотической экспрессии, содержащий на тяжелой цепи hinje-участок с цистеиновыми остатками для образования дисульфидных связей. Молекулу F(ab’)2-ранибизумаб мы использовали как базисную структуру для разработки других вариантов молекул, модифицированных различными полипептидами с целью увеличения их массы и гидродинамического объема. В качестве полипептидов, модифицирующих F(ab’)2-ранибизумаб мы использовали либо Fc-фрагмент иммуноглобулина IgG1, либо СН3-домен Fc-фрагмента иммуноглобулина IgG1, либо неструктурированные гидрофильные полипептиды (НГП), слитые либо с hinje-участком, либо с СН3-доменом Fc-фрагмента иммуноглобулина IgG1, либо с легкой цепью молекулы ранибизумаба.
Многочисленные исследования полученного продукта показали, что F(ab')2-ранибизумаб имеет увеличенное время экспозиции в стекловидном теле и при этом характеризуется неожиданно высокой аффинностью к VEGF-A по сравнению с ранизумабом.
Таким образом, объектом настоящего изобретения является полипептид, содержащий F(ab')2-фрагмент терапевтического антитела, для введения в стекловидное тело для целей лечения и/или профилактики заболевания, в отношении которого указанное антитело имеет терапевтическую активность.
Указанным белком предпочтительно является ранибизумаб, а указанным заболеванием - любое заболевание, сопровождающееся отеком макулы вследствие повышенной экспрессии VEGF-A.
При этом специалисту в уровне техники очевидно, что для последующего увеличения массы и времени действия F(ab’)2-фрагмента терапевтического антитела по изобретению может быть дополнен другими полипептидными последовательностями. Например, такими полипептидными последовательностями может быть домен СН3 антитела и/или НП, включая НАР-полипептидные последовательности S(G4S)16 и S(G4S)20 и другие по НП, упомянутые в разделе «Уровень техники». Поэтому F(ab’)2-фрагмент терапевтического антитела по изобретению может дополнительно включать полипептидные последовательности, выбираемые, в том числе, из S(G4S)16, S(G4S)20, домена СН3 или их комбинации.
Использование изобретения позволяет повысить удобство анти- VEGF-A терапии при отеке макулы за счет увеличенного времени экспозиции в стекловидном теле и высокой аффинности связывания анти-VEGF-A полипептида с мишенью, что позволит значительно сократить количество введений терапевтического средства.
Краткое описание чертежей
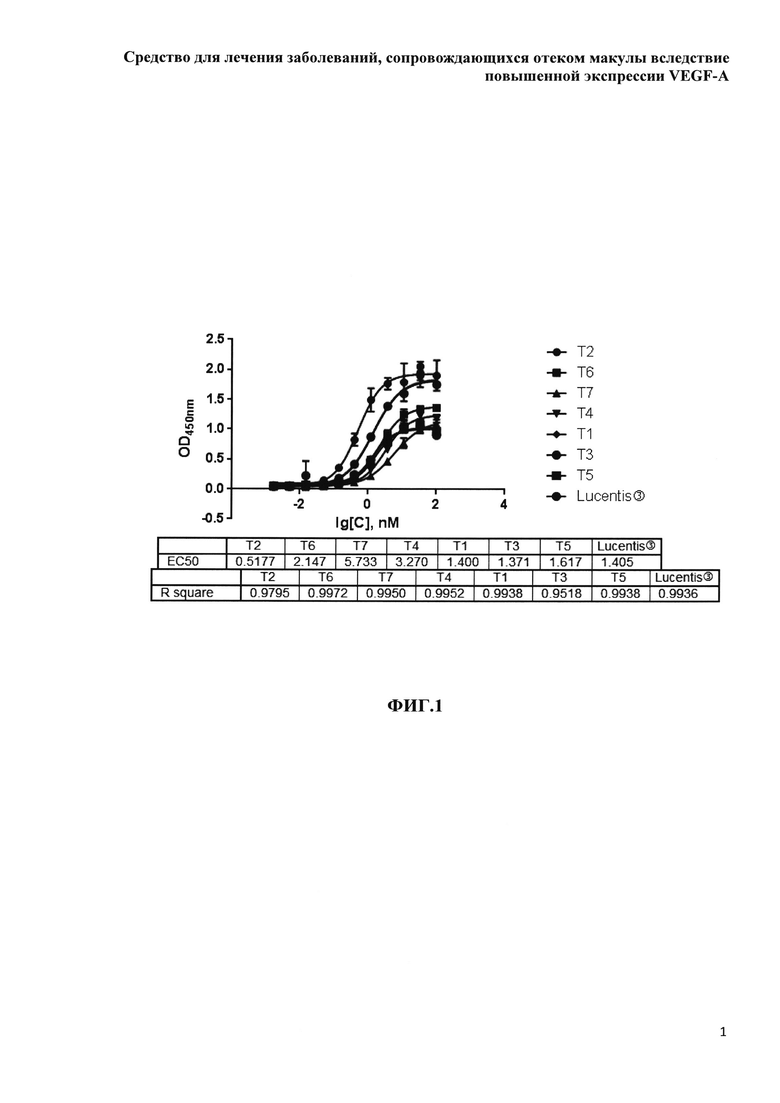
Фиг. 1. Связывание белков на основе F(ab’)2-фрагмента ранибизумаба с рекомбинантным человеческим эндотелиальным фактором роста rhVEGF121, определенное методом ИФА.
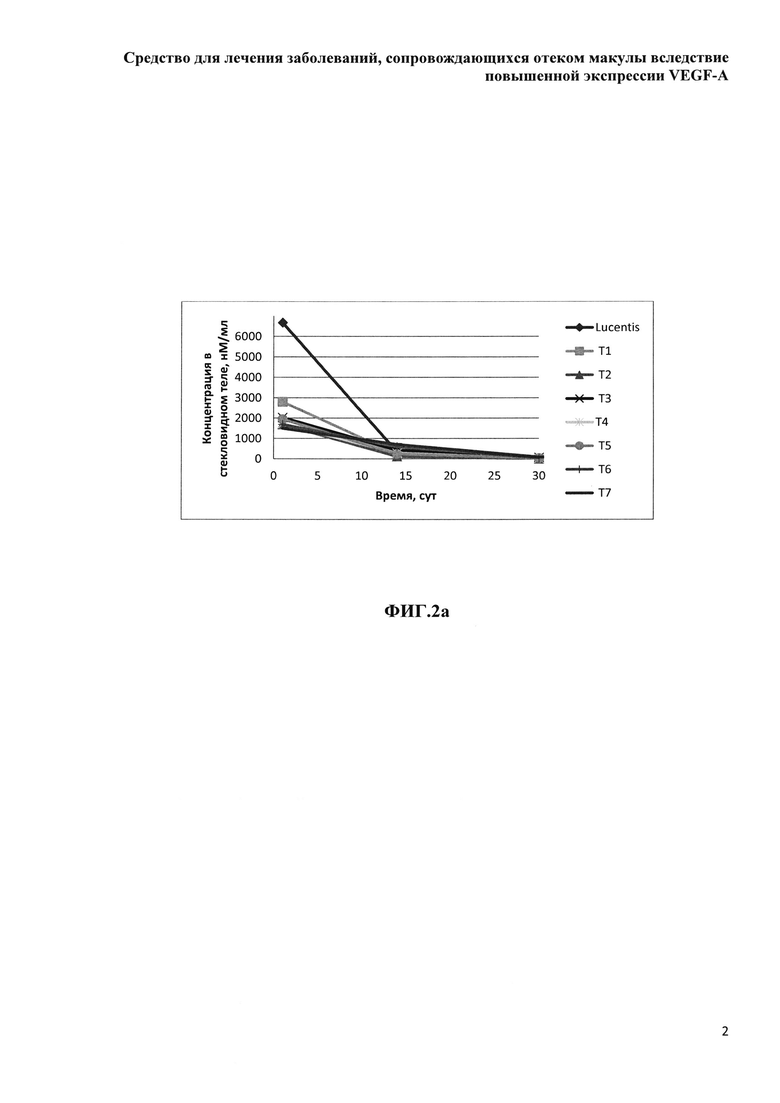
Фиг. 2а. Зависимость медианы концентрации определяемого белка от дня отбора образцов стекловидного тела.
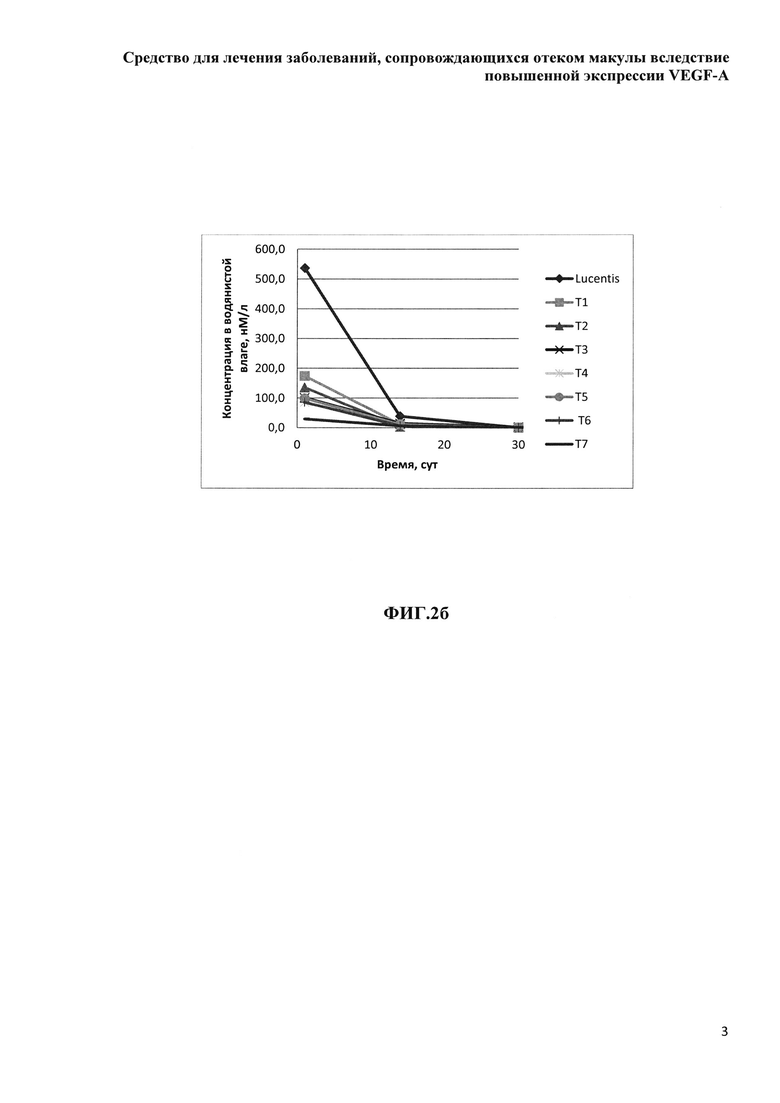
Фиг. 2б. Зависимость медианы концентрации определяемого белка от дня отбора образцов водянистой влаги.
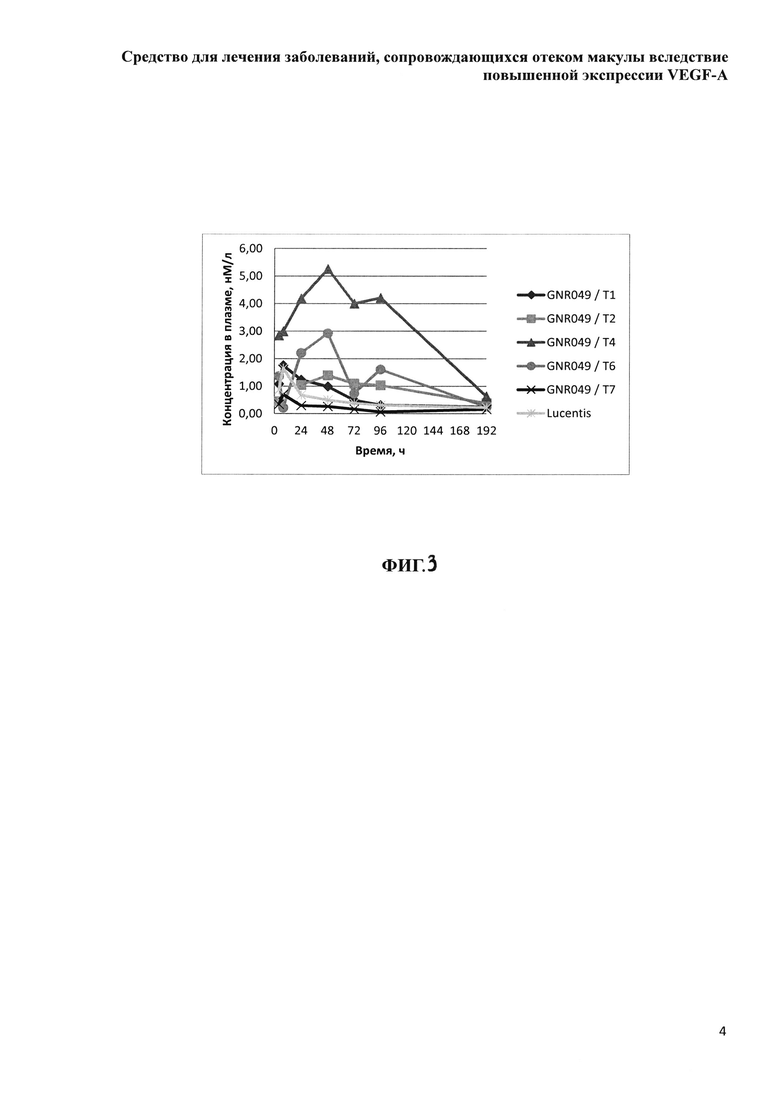
Фиг. 3. Зависимость медианы концентрации определяемого белка от времени отбора образцов крови.
Осуществление изобретения
Этапы получения и характеризации заявляемых гибридных белков на основе F(ab')2-ранибизумаба.
Этап 1. Получение генно-инженерных конструкций для эукариотической экспрессии. Используя рутинные методы генетической инженерии, осуществляли клонирование синтетических последовательностей ДНК, кодирующих тяжелую и легкую цепи F(ab')2-фрагмента ранибизумаба (Lucentis®) в плазмидные векторы, предназначенные для экспрессии белков в эукариотических клетках.
Используя рутинные методы генетической инженерии, осуществляли корректное слияние последовательностей ДНК, кодирующих тяжелую цепь F(ab’)2-фрагмента ранибизумаба (Lucentis®) и третий константный домен тяжелой цепи (СН3 домен) иммуноглобулина человека подкласса 1 класса IgG, полученный методом ПЦР с использованием соответствующих оригинальных праймеров.
Конструирование плазмидных ДНК, предназначенных для экспрессии в эукариотических клетках белков Lucentis-HC-НГП, Lucentis-LC-НГП и Lucentis-HC-СН3-НГП, слитых с неструктурированным гидрофильным пептидом (НГП), осуществляли с помощью рутинных методов генетической инженерии с использованием вспомогательных плазмид, содержащих последовательности, кодирующие соответствующие белки, и корректного слияния последовательностей данных белков с последовательностями, кодирующими НГП. Целевые экспрессионные плазмиды получали путем перемещения фрагментов вспомогательных плазмид, кодирующих белки Lucentis-HC-НГП, Lucentis-LC-НГП и Lucentis-HC-СН3-НГП, в плазмидные векторы, предназначенные для экспрессии белков в эукариотических клетках.
Этап 2. Получение клонов-продуцентов белков на основе эукариотических клеток. Приготовленные ГИК использовали для электропорации эукариотических клеток и создания клонов-продуцентов на их основе.
Этап 3. Культивирование клонов-продуцентов и выделение белков.
Культивирование клонов-продуцентов целевых белков на основе F(ab’)2-фрагмента ранибизумаба осуществляли в одноразовом волновом биореакторе в режиме фед-батч (периодическое культивирование с подпиткой).
Выделение белков осуществляли в нескольких стадий - стадия аффинной хроматографии с использованием сорбентов Capto L (GE Healthcare) или KappaSelect (GE Healthcare), стадии ионообменной хроматографии на гидроксиапатите (CHT-I, Bio-Rad) и SP-sepharose (GE Healthcare).
Этап 4. Изучение физико-химических и функциональных свойств in vitro и in vivo.
Характеризацию выделенных белков на основе F(ab’)2-фрагмента ранибизумаба осуществляли методами ПААГ-электрофореза с додецилсульфатом натрия, ВЭЖХ гель-фильтрации, ИФА, плазмонного поверхностного резонанса и в тесте ингибирования VEGF-A-индуцированной пролиферации клеток HUVEC (первичной культуры эндотелиоцитов из пупочной вены человека).
Этап 5. Оценка фармакокинетических параметров белков на основе F(ab’)2-фрагмента ранибизумаба при интравитреальном введении кроликам.
Целью исследования была сравнительная оценка параметров фармакокинетики и токсикокинетики белков на основе F(ab’)2-фрагмента ранибизумаба и препарата Lucentis® (Novartis) после однократного интравитреального введения кроликам породы Советская шиншилла.
Далее осуществление изобретения показано на конкретных примерах, что не должно восприниматься как ограничение в отношении созданного изобретения. Специалисту в уровне техники ясно, что без затрат изобретательского творчества могут быть разработаны дополнения и изменения относительно раскрытого в ниже представленных примерах в рамках настоящего изобретения.
Пример 1. Конструирование рекомбинантной плазмиды pRA1654, кодирующей тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®).
Синтезируют фрагмент ДНК, имеющий открытые «липкие» концы, идентичные «липким» концам, создаваемым эндонуклеазами рестрикции HindIII на 5'-конце и XbaI на 3'-конце, и кодирующий полипептид

соответствующий тяжелой цепи F(ab)2-фрагмента ранибизумаба (Lucentis®) с N-концевой лидерной последовательностью (курсив) фактора свертываемости крови FVII, удаляемой из состава целевого белка в процессе транспорта из цитоплазмы клеток клона-продуцента.
Рекомбинантную плазмиду pRA1654 конструируют на основе плазмидного вектора для экспрессии белков в эукариотических клетках, который имеет в своем составе следующие структурные элементы:
структурные элементы CMV промотора (CMV enhancer, EF-1α promoter, EF-1α intron A); сигнал полиаденилирования бычьего гормона роста (bGH poly(A) signal); ориджин репликации высококопийных плазмид (ori ColE1/pMB1/pBR322/pUC); ген, кодирующий β-лактамазу, обеспечивающую устойчивость к ампициллину, карбенициллину и подобным антибиотикам (AmpR, AmpR promoter); ориджин репликации бактериофага f1 (f1 ori); ранний промотор вируса SV40 (SV40 promoter); ориджин репликации вируса SV40 (SV40 ori); ген PuroR, кодирующий пуромицин N-ацетилтрансферазу, обеспечивающую устойчивость к пуромицину (альтернативно может содержат HygR, кодирующий гигромицин В фосфотрансферазу, обеспечивающую устойчивость к гигромицину В, или NeoR/KanR, кодирующую аминогликозид фосфотрансферазу из Tn5, обеспечивающую устойчивость к неомицину, канамицину и G418 (Geneticin®); синтетический сигнал полиаденилирования (poly(A) signal); а также уникальные сайты узнавания эндонуклеаз рестрикции HindIII и XbaI, расположенные последовательно.
С этой целью синтетический фрагмент ДНК клонируют в выбранном векторе, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную плазмиду pRA1654, содержащую клонированный синтетический фрагмент ДНК, кодирующий тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®)
Пример 2. Конструирование рекомбинантной плазмиды pRA1655, кодирующей легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®).
Синтезируют фрагмент ДНК, имеющий открытые «липкие» концы, идентичные «липким» концам, создаваемым эндонуклеазами рестрикции HindIII на 5'-конце и XbaI на 3'-конце, и кодирующий полипептид

соответствующий легкой цепи F(ab)2-фрагмента ранибизумаба (Lucentis®) с N-концевой лидерной последовательностью (курсив) фактора свертываемости крови FVII, удаляемой из состава целевого белка в процессе транспорта из цитоплазмы клеток клона-продуцента.
Рекомбинантную плазмиду pRA1655 конструируют на основе плазмидного вектора для экспрессии белков в эукариотических клетках, который имеет в своем составе структурные элементы, упомянутые в Примере 1.
С этой целью синтетический фрагмент ДНК клонируют в выбранном векторе, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную плазмиду pRA1655, содержащую клонированный синтетический фрагмент ДНК, кодирующий легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®).
Пример 3. Конструирование рекомбинантной плазмиды рМТ1605, кодирующей тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) с СН3 доменом IgG1 человека.
Для получения рекомбинантной плазмиды рМТ1605, кодирующей тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) с СН3 доменом IgG1 человека, проведены 2 полимеразных цепных реакции.
Реакция 1. В качестве матрицы использована рекомбинантная плазмида pRA1654 (получение описано в Примере 1), в качестве затравок синтеза использованы синтетические олигонуклеотиды:
St3_for
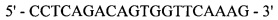
и
оМТ907
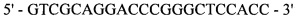 ;
;
полученный фрагмент с внесенным сайтом узнавания эндонуклеазой рестрикции SmaI на 3'-конце кодирует тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®).
Реакция 2. В качестве матрицы использована лабораторная рекомбинантная плазмида, кодирующая третий константный домен тяжелой цепи иммуноглобулина человека подкласса 1 класса IgG (СН3 домен IgG1 человека), в качестве затравок использованы синтетические олигонуклеотиды:
oMT908
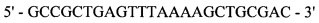
и
oRA405
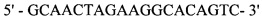 ;
;
полученный фрагмент с внесенным сайтом узнавания эндонуклеазой рестрикции DraI на 5'-конце кодирует СН3 домен IgG1 человека.
Рекомбинантную плазмиду рМТ1605 конструируют на основе плазмидного вектора для экспрессии белков в эукариотических клетках, который имеет в своем составе структурные элементы, упомянутые в Примере 1.
Фрагмент, полученный в ходе Реакции 1, расщепляют с использованием эндонуклеаз рестрикции HindIII и SmaI; а фрагмент, полученный в ходе Реакции 2, с использованием DraI и XbaI. Полученные фрагменты очищают с помощью спин-колонок и используют в трехкомпонентной (вектор + вставка 1 + вставка 2) реакции лигирования с выбранным вектором, расщепленным по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования HindIII и XbaI были восстановлены. В результате получают рекомбинантную плазмиду рМТ1605, содержащую клонированный фрагмент ДНК, кодирующий тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) слитую с СН3 доменом IgG1 человека, кодирующий полипептид

соответствующий тяжелой цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) слитой с СН3 доменом IgG1 человека с N-концевой лидерной последовательностью (курсив) фактора свертываемости крови FVII, удаляемой из состава целевого белка в процессе транспорта из цитоплазмы клеток клона-продуцента.
Пример 4. Конструирование рекомбинантной плазмиды рМТ1616, кодирующей тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) с СН3 доменом IgG1 человека, слитую с НГП.
Для получения рекомбинантной плазмиды рМТ1616, кодирующей тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) с СН3 доменом IgG1 человека слитую с НГП, получена вспомогательная плазмида №1.
Рекомбинантную вспомогательную плазмиду №1 конструируют на основе любого лабораторного плазмидного вектора, содержащего уникальные сайты узнавания эндонуклеаз рестрикции HindIII и XbaI, а также такие структурные элементы как ориджин репликации высококопийных плазмид (ori ColE1/pMB1/pBR322/pUC); ген, кодирующий β-лактамазу, обеспечивающую устойчивость к ампициллину, карбенициллину и подобным антибиотикам (AmpR, AmpR promoter); ориджин репликации бактериофага f1 (f1 ori); и не содержащую сайта узнавания эндонуклеазы рестрикции SapI, например, модифицированную плазмиду pBK685.
С этой целью рекомбинантную плазмиду рМТ1605, кодирующей тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) слитую с СН3 доменом IgG1 человека, расщепляют с использованием эндонуклеаз рестрикции HindIII и XbaI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-1. Фрагмент-1 клонируют в выбранном векторе, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную вспомогательную плазмиду №1а или рМТ1608, содержащую клонированный фрагмент ДНК, кодирующий тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) с СН3 доменом IgG1 человека.
Реакция 3. Для внесения сайта узнавания эндонуклеазы рестрикции SapI в области 3'-конца за 4 п.н. до стоп-кодона проведена ПЦР. В качестве матрицы использована рекомбинантная плазмида рМТ1608; в качестве затравок использованы синтетические олигонуклеотиды:
oMT909
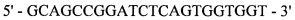
и
оМТ910
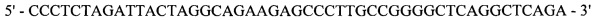 ;
;
полученный фрагмент с внесенным сайтом узнавания эндонуклеазы рестрикции SapI на 3'-конце кодирует модифицированную на С-конце тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) с СН3 доменом IgG1 человека.
Фрагмент ДНК, полученный в ходе Реакции 3, расщепляют с использованием эндонуклеаз рестрикции HindIII и XbaI; очищают с помощью спин-колонок и используют в реакции лигирования с выбранным вектором (pBK685), расщепленным по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды (вспомогательная плазмида №16) сайты клонирования HindIII и XbaI были восстановлены.
Для получения рекомбинантной вспомогательной плазмиды №1 вспомогательную плазмиду №16 расщепляют по сайту узнавания эндонуклеазы рестрикции SapI. Одновременно, рекомбинантную плазмиду, кодирующую НГП расщепляют по сайту узнавания эндонуклеазы рестрикции SapI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-2. Фрагмент-2 клонируют в вспомогательную плазмиду №16, расщепленную по сайту SapI. Полученная рекомбинантная вспомогательная плазмида №1 содержит клонированный фрагмент ДНК, кодирующий тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) с СН3 доменом IgG1 человека слитую с НГП.
Рекомбинантную плазмиду рМТ1616 конструируют на основе плазмидного вектора для экспрессии белков в эукариотических клетках, который имеет в своем составе структурные элементы, упомянутые в Примере 1.
С этой целью рекомбинантную вспомогательную плазмиду №1 расщепляют по сайтам узнавания эндонуклеазами рестрикции HindIII и XbaI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-3. Фрагмент-3 клонируют в выбранном векторе для эукариотической экспрессии, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную плазмиду рМТ1616, содержащую клонированный фрагмент ДНК, кодирующий тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) с СН3 доменом IgG1 человека, слитую с НГП.
Рекомбинантная плазмида рМТ1616 кодирует полипептид

соответствующий тяжелой цепь F(ab)2-фрагмента ранибизумаба (Lucentis®) слитой с СН3 доменом IgG1 человека, слитой с НГП (жирный), с N-концевой лидерной последовательностью (курсив) фактора свертываемости крови FVII, удаляемой из состава целевого белка в процессе транспорта из цитоплазмы клеток клона-продуцента.
Пример 5. Конструирование рекомбинантной плазмиды рМТ1612, кодирующей тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП.
Для получения рекомбинантной плазмиды рМТ1612, кодирующей тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП, получена вспомогательная плазмида №2.
Рекомбинантную вспомогательную плазмиду №2 получают также, как и рекомбинантную вспомогательную плазмиду №1, описанную в Примере 4.
С этой целью рекомбинантную плазмиду pRA1654 из Примера 1, кодирующей тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), расщепляют с использованием эндонуклеаз рестрикции HindIII и XbaI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-4. Фрагмент-4 клонируют в векторе pBK685, описанном в Примере 4, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную вспомогательную плазмиду №2а (рМТ1606), содержащую клонированный фрагмент ДНК, кодирующий тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®).
Реакция 4. Для внесения сайта узнавания эндонуклеазы рестрикции SapI в области 3'-конца за 4 п.н. до стоп-кодона проведена ПЦР. В качестве матрицы использована рекомбинантная плазмида рМТ1606; в качестве затравок использованы синтетические олигонуклеотиды:
оМТ909
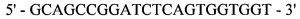
и
oMT911
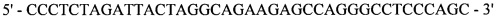 ,
,
полученный фрагмент с внесенным сайтом узнавания эндонуклеазы рестрикции SapI на 3'-конце кодирует модифицированную на С-конце тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®).
Фрагмент ДНК, полученный в ходе Реакции 4, расщепляют с использованием эндонуклеаз рестрикции HindIII и XbaI; очищают с помощью спин-колонок и используют в реакции лигирования с выбранным вектором рВК685, расщепленным по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды (вспомогательная плазмида №2б) сайты клонирования HindIII и XbaI были восстановлены.
Для получения рекомбинантной вспомогательной плазмиды №2 вспомогательную плазмиду №2б расщепляют по сайту узнавания эндонуклеазы рестрикции SapI. Одновременно, рекомбинантную плазмиду, кодирующую НГП расщепляют по сайту узнавания эндонуклеазы рестрикции SapI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-5. Фрагмент-5 клонируют во вспомогательную плазмиду №2б, расщепленную по сайту SapI. Полученная рекомбинантная вспомогательная плазмида №2 содержит клонированный фрагмент ДНК, кодирующий тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП.
Рекомбинантную плазмиду рМТ1612 конструируют на основе плазмидного вектора для экспрессии белков в эукариотических клетках, который имеет в своем составе структурные элементы, упомянутые в Примере 1.
С этой целью рекомбинантную вспомогательную плазмиду №2 расщепляют по сайтам узнавания эндонуклеазами рестрикции HindIII и XbaI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-6. Фрагмент-6 клонируют в выбранном векторе для эукариотической экспрессии, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную плазмиду рМТ1612, содержащую клонированный фрагмент ДНК, кодирующий тяжелую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП.
Рекомбинантная плазмида рМТ1612 кодирует полипептид

соответствующий тяжелой цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитой с НГП (жирный), с N-концевой лидерной последовательностью (курсив) фактора свертываемости крови FVII, удаляемой из состава целевого белка в процессе транспорта из цитоплазмы клеток клона-продуцента.
Пример 6. Конструирование рекомбинантных плазмид рМТ1613, рМТ1614 и рМТ1615, кодирующих легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП.
Для получения рекомбинантной плазмиды рМТ1613, рМТ1614 и рМТ1615, кодирующих легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП, получена вспомогательная плазмида №3.
Рекомбинантную вспомогательную плазмиду №3 получают также, как и рекомбинантные вспомогательные плазмиды №1 и №2, описанные в Примерах 4 и 5.
С этой целью рекомбинантную плазмиду pRA1655 из Примера 2, кодирующей легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), расщепляют с использованием эндонуклеаз рестрикции HindIII и XbaI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-7. Фрагмент-7 клонируют в векторе pBK685, описанном в Примере 4, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную вспомогательную плазмиду №3а (рМТ1607), содержащую клонированный фрагмент ДНК, кодирующий легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®).
Реакция 5. Для внесения сайта узнавания эндонуклеазы рестрикции SapI в области 3'-конца за 4 п.н. до стоп-кодона проведена ПЦР. В качестве матрицы использована рекомбинантная плазмида рМТ1607; в качестве затравок использованы синтетические олигонуклеотиды:
оМТ909
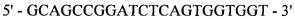
и
oMT912
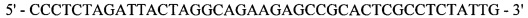 ,
,
полученный фрагмент с внесенным сайтом узнавания эндонуклеазы рестрикции SapI на 3'-конце кодирует модифицированную на С-конце легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®).
Фрагмент ДНК, полученный в ходе Реакции 5, расщепляют с использованием эндонуклеаз рестрикции HindIII и XbaI; очищают с помощью спин-колонок и используют в реакции лигирования с выбранным вектором pBK685, расщепленным по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды (вспомогательная плазмида №3б) сайты клонирования HindIII и XbaI были восстановлены.
Для получения рекомбинантной вспомогательной плазмиды №3 вспомогательную плазмиду №3б расщепляют по сайту узнавания эндонуклеазы рестрикции SapI. Одновременно, рекомбинантную плазмиду, кодирующую НГП расщепляют по сайту узнавания эндонуклеазы рестрикции SapI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-8. Фрагмент-8 клонируют во вспомогательную плазмиду №3б, расщепленную по сайту SapI. Полученная рекомбинантная вспомогательная плазмида №3 содержит клонированный фрагмент ДНК, кодирующий легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП.
Рекомбинантную плазмиду рМТ1613 конструируют на основе плазмидного вектора для экспрессии белков в эукариотических клетках, который имеет в своем составе структурные элементы, упомянутые в Примере 1.
С этой целью рекомбинантную вспомогательную плазмиду №3 расщепляют по сайтам узнавания эндонуклеазами рестрикции HindIII и XbaI и образовавшиеся фрагменты рестрикции разделяют в геле. Меньший по размеру фрагмент ДНК элюируют из геля и называют фрагмент-9. Фрагмент-9 клонируют в выбранном векторе для эукариотической экспрессии, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную плазмиду рМТ1613, содержащую клонированный фрагмент ДНК, кодирующий легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП.
Рекомбинантную плазмиду рМТ1614 конструируют на основе плазмидного вектора для экспрессии белков в эукариотических клетках, который отличается от вектора, имеющего в своем составе структурные элементы, упомянутые в Примере 1, тем, что элемент PuroR, кодирующий пуромицин N-ацетилтрансферазу, обеспечивающую устойчивость к пуромицину, заменен на HygR, кодирующий гигромицин В фосфотрансферазу, обеспечивающую устойчивость к гигромицину В.
Для получения целевого экспрессионного вектора рМТ1614 фрагмент-9 клонируют в выбранном векторе для эукариотической экспрессии, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную плазмиду рМТ1614, содержащую клонированный фрагмент ДНК, кодирующий легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП.
Рекомбинантную плазмиду рМТ1615 конструируют на основе плазмидного вектора для экспрессии белков в эукариотических клетках, который отличается от вектора, имеющего в своем составе структурные элементы, упомянутые в Примере 1, тем, что элемент PuroR, кодирующий пуромицин N-ацетилтрансферазу, обеспечивающую устойчивость к пуромицину, заменен на NeoR/KanR, кодирующую аминогликозид фосфотрансферазу из Tn5, обеспечивающую устойчивость к неомицину, канамицину и G418 (Geneticin®).
Для получения целевого экспрессионного вектора рМТ1615 фрагмент-9 клонируют в выбранном векторе для эукариотической экспрессии, расщепленном по сайтам HindIII и XbaI таким образом, чтобы в составе результирующей плазмиды сайты клонирования были восстановлены. В результате получают рекомбинантную плазмиду рМТ1615, содержащую клонированный фрагмент ДНК, кодирующий легкую цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитую с НГП.
Рекомбинантные плазмиды рМТ1613, рМТ1614 и рМТ1615 кодируют полипептид

соответствующий легкой цепь F(ab)2-фрагмента ранибизумаба (Lucentis®), слитой с НГП (жирный), с N-концевой лидерной последовательностью (курсив) фактора свертываемости крови FVII, удаляемой из состава целевого белка в процессе транспорта из цитоплазмы клеток клона-продуцента.
Пример 7. Получение клонов-продуцентов белков на основе эукариотических клеток.
Приготовленные ГИК использовали для создания клонов-продуцентов на основе эукариотических клеток. Для этого смешивали препараты плазмид, описанных в примерах 1-6, кодирующих легкую и тяжелую цепи соответствующего белка, и трансфецировали эукариотические клетки методом электропорации. Проводили селекцию трансфецированных клеток методом культивирования на селективном антибиотике. После получение стабильных пулов клеток-продуцентов соответствующих белков производили их клонирование, культуральную жидкость одиночных клонов тестировали методом твердофазного ИФА на иммобилизованном рекомбинантном человеческом VEGF-A и выбирали клоны с максимальной продуктивностью целевых белков на основе F(ab’)2-фрагмента ранибизумаба.
Пример 8. Культивирование клонов-продуцентов и выделение белков.
Культивирование клонов-продуцентов целевых белков на основе F(ab’)2-фрагмента ранибизумаба осуществляли в одноразовом волновом биореакторе в режиме фед-батч (периодическое культивирование с подпиткой). В качестве базовой питательной среды использовали SFM4CHO (HyClone, США), в качестве подпитки - BalanCD СНО Feed 2 (Irvine Scientific, США). Продолжительность культивирования составляла 10 суток. По окончании культивирования суспензии клеток проводили через 3-х-ступенчатую осветляющую фильтрацию и проводили выделение целевых белков из культуральной жидкости.
На первой стадии очистки всех вариантов белков на основе F(ab’)2-фрагмента ранибизумаба применяли аффинную хроматографию с использованием сорбентов Capto L (GE Healthcare) или KappaSelect (GE Healthcare). Оба сорбента специфично связывают легкую цепь иммуноглобулинов типа IgG-каппа за ее вариабельную (Capto L) или константную (KappaSelect) часть. Сорбенты уравновешивали фосфатно-солевым буфером, наносили осветленную культуральную жидкость, промывали уравновешивающим буфером от несвязавшихся примесей, затем элюировали целевые белки буфером, содержащим 50 мМ гистидина, pH 2,5. Для всех белков показали схожую эффективность выделения при использовании обоих сорбентов.
На второй стадии очистки использовали мультимодальный сорбент гидроксиапатит (CHT-I, Bio-Rad), который часто применяется для очистки антител от родственных примесей и остаточных белков клеток-продуцентов. Сорбент предварительно уравновешивали раствором 50 мМ гистидина, pH 7,0, затем наносили элюаты белков после аффинной стадии, разведенные водой до проводимости 3-5 мСм/см, промывали сорбент уравновешивающим буфером, затем буфером, содержащим 50 мМ гистидин, 1 М хлорид натрия, pH 7,0. Целевые белки элюировали линейным градиентом фосфата натрия (pH 7,0) до концентрации 300 мМ. Все полученные на этой хроматографической стадии фракции анализировали с помощью ВЭЖХ гель-фильтрации и ПААГ-электрофореза с додецилсульфатом натрия для выбора фракций, содержащих целевые формы белков. Очистка на гидроксиапатите позволяла эффективно удалить низкомолекулярные родственные примеси, а также остаточные примесные белки.
При необходимости проводили доочистку целевого белка путем фильтрации через катионообменный сорбент SP-sepharose (GE Healthcare) при pH 5,5 и проводимости 4-6 мСм/см, а также концентрирование и диафильтрацию образцов в требуемый буфер на концентраторах Amicon Ultra (Millipore) с отсечкой по молекулярной массе 30-50 кДа. По окончании выделения чистота белка составляла не менее 90% целевой формы белка по данным электрофореза и ВЭЖХ гель-фильтрации.
Пример 9. Физико-химические свойства белков, определяемые методом ВЭЖХ гель-фильтрации.
Всего было получено 7 разных белков на основе F(ab’)2-фрагмента ранибизумаба, проводили характеризацию выделенных белков методом ВЭЖХ гель-фильтрации. Для этого использовали систему ВЭЖХ Alliance 2695 (Waters), с детектором UV/Visible Detector 2487 (Waters) и колонку Superdex200 (Increase 10/300 GL, Lot. 10226065, No. 0128; GE Healthcare). В качестве подвижной фазы использовали фосфатно-солевой изотонический буфер. В таблице 1 даны условное обозначение белков, их номинальная и кажущаяся молекулярная масса, полученная при ВЭЖХ гель-фильтрации.
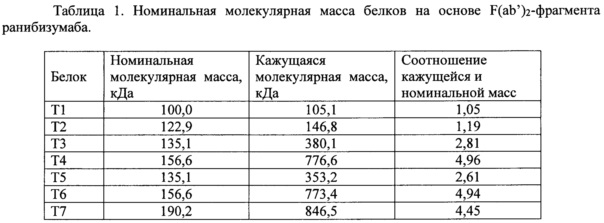
Варианты белков на основе F(ab')2-фрагмента ранибизумаба T3, T4, T5, T6 и T7 имеют по данным ВЭЖХ гель-фильтрации большую кажущуюся молекулярную массу, превышающую их номинальную молекулярную массу от 2,61 до 4,96 раз, что обусловлено наличием в их структуре НГП.
Пример 10. Взаимодействие белков с рекомбинантным VEGF-A (rhVEGF121) человека, определяемое методом ИФА.
Препараты белков на основе F(ab’)2-фрагмента ранибизумаба были охарактеризованы методом ИФА по связыванию с рекомбинантным человеческим фактором роста эндотелия сосудов - rhVEGF121 производства ООО «МБЦ «Генериум». Для этого rhVEGF121 производства ООО «МБЦ «Генериум» вносили в лунки ИФА планшетов по 100 μl в концентрации 1 μg/ml в карбонат-бикарбонатном буфере (pH 9,5) и сорбировали в течение ночи при +4С. Остаточные свободные центры связывания пластика блокировали раствором PBS-Tha («ПанЭко», Кат. № Э4100-110) в течение 1 часа при комнатной температуре, после чего в лунки планшетов вносили по 100 мкл растворов белков, разведенных PBS-0,05%Tw20 последовательными 3-кратными разведениями от 100 до 0,002 nM. Планшеты инкубировали в течение 1 часа, отмывали несвязавшиеся антитела, детекцию комплексов белков с rhVEGF121 проводили с помощью биотинилированных поликлональных крысиных антител к ранибизумабу производства ООО «МБЦ «Генериум» и конъюгированного с пероксидазой хрена стрептавидина (Abcam, ab7403). В качестве субстрата для пероксидазы хрена использовали ТМБ (США, Партия: TMBPD10222-2-10273-А), реакцию останавливали раствором 2М H2SO4. Развитие реакции оценивали по оптической плотности, измеренной при длине волны 450 нм. Результаты исследования представлены на рисунке 1.
В таблице 2 приведены усредненные данные по аффинности GNR049-кандидатов (ЕС50, nM), полученные с помощью ИФА.
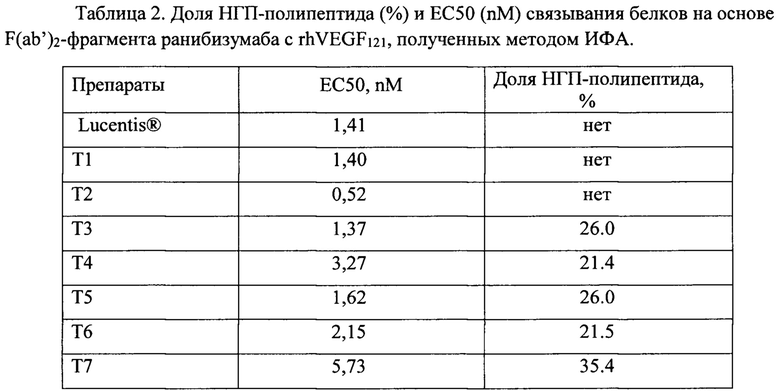
Все белки на основе F(ab’)2-фрагмента ранибизумаба взаимодействуют с рекомбинантным человеческим эндотелиальным фактором роста rhVEGF121, при этом варианты белков, несущие НГП, дают меньшую оптическую плотность при насыщающих концентрациях в сравнении с белками без НГП, а кажущийся аффинитет белков, несущих НГП, уменьшается с возрастанием доли НГП-полипептида в молекулярной массе.
Пример 11. Взаимодействие белков с рекомбинантным VEGF-A (rhVEGF121) человека, определяемое методом поверхностного плазмонного резонанса.
Кинетические характеристики взаимодействия белков на основе F(ab’)2-фрагмента ранибизумаба оценивали методом поверхностного плазмонного резонанса на приборе Biacore Т200. В первой серии экспериментов сравнивали кинетические характеристики взаимодействия коммерческого препарата Lucentis® (Novartis) и F(ab’)2-фрагмента ранибизумаба, полученного в ООО «МБЦ «Генериум». Для этого рекомбинантный человеческий белок rhVEGF121 производства ООО «МБЦ «Генериум» и рекомбинантный коммерческий VEGF-А кролика (Kingfisher Biotech, Inc., Cat. № RPO923U-100) ковалентно иммобилизовали на чип СМ-5, затем пропускали растворы Lucentis® (Novartis) и F(ab’)2-фрагмента ранибизумаба в буфере HBS-EP+. Рабочие концентрации 0.74, 2.22, 6.67, 20, 60 nM. Ассоциация 210 сек в буфере HBS-EP+. Диссоциация 14400 сек (4 часа) в буфере 20 mM Na-Acetate, 150 mM NaCl, 3 mM EDTA, 0.005% Tween20, pH 4.5. Финальная регенерация раствором 50 mM Gly-HCl, pH 2.5. В связи с использованием низкого pH при диссоциации, получены кажущиеся KD для вариантов Lucentis с human VEGF121. Кажущиеся KD рассчитывали из математической модели белок-белкового взаимодействия 1:1. Данные приведены в таблице 3.

Изменение формата молекулы с Fab-фрагмента на F(ab’)2-фрагмент при неизменной первичной структуре привело к повышению кажущейся аффинности примерно в 10 раз, столь значительное повышение аффинности F(ab’)2-фрагмента ранибизумаба явилось неожиданным.
Во второй серии экспериментов оценивали кинетические характеристики взаимодействия других вариантов белков на основе F(ab’)2-фрагмента ранибизумаба с рекомбинантным человеческим фактором роста эндотелия сосудов rhVEGF121 методом поверхностного плазмонного резонанса на приборе Biacore Т200 по приведенной выше методике. Данные приведены в таблице 4.
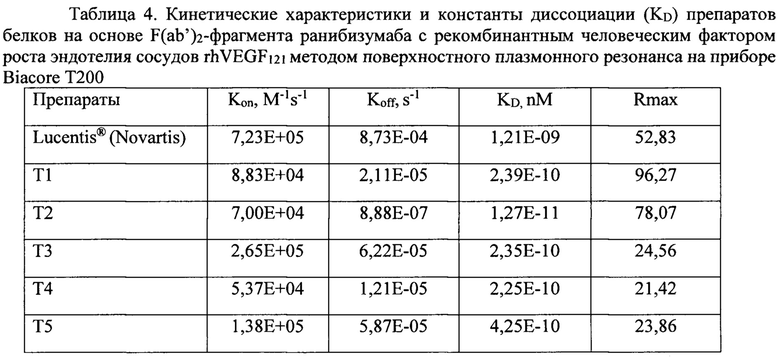
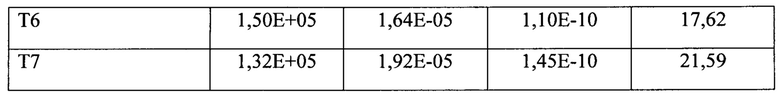
Как видно из данных, представленных в таблице 4, кинетические характеристики взаимодействия белков на основе F(ab')2-фрагмента ранибизумаба с рекомбинантным фактором роста сосудов человека rhVEGF121 - константы скорости ассоциации комплекса Ка (Kon) и скорости диссоциации комплекса Kd (Koff), также, как и равновесная константа диссоциации комплекса KD, у всех вариантов димерных молекул не уступают, и даже превосходят таковые характеристики для ранибизумаба. В то же время, RUmax для белков, несущих в своей структуре НГП, достоверно меньше, чем для белков без НГП, тогда как данный показатель должен расти с увеличением молекулярной массы, что мы и наблюдаем для мономера и димера ранибизумаба -для мономера RUmax составляет 52,83, а для димеров - 78,07 и 96,27. Достоверное снижение RUmax для белков с НГП может объясняться как малой долей активных молекул в белковом препарате, так и тем, что модифицированные НГП молекулы имеют больший гидродинамический объем, и при образовании комплекса с ковалентно иммобилизованным на подложке чипа rhVEGF121, НГП экранирует соседнюю иммобилизованную молекулу rhVEGF121, создавая эффект уменьшения плотности иммобилизованных молекул rhVEGF121, доступных для взаимодействия, что и приводит к снижению RUmax. Объяснение феномена снижения RUmax для препаратов белков с НГП при анализе на Biacore Т200 потребовало от нас постановки эксперимента с образованием комплекса каждого из белков с rhVEGF121 в растворе, поскольку в растворе отсутствуют стерические затруднения комплексообразования.
Пример 12. Образование комплекса белков на основе F(ab’)2-фрагмента ранибизумаба с рекомбинантным человеческим фактором роста эндотелия сосудов rhVEGF121 в растворе.
При анализе кинетических характеристик взаимодействия препаратов с rhVEGF121 на Biacore Т200 наблюдали значимое снижение RUmax для модифицированных НГП белков на основе F(ab’)2-фрагмента ранибизумаба, что могло свидетельствовать как о стерических затруднениях взаимодействия белков, несущих НГП, с иммобилизованным на подложке антигеном вследствие их большого гидродинамического радиуса, так и о том, что какая-то доля белка в препаратах не активна. Для этого проводили дополнительный анализ активности приготовленных препаратов, изучая их взаимодействие с rhVEGF121 в растворе при эквимолярном соотношении. Белки смешивали с rhVEGF121 в эквимолярном соотношении в растворе PBS, инкубировали в течение ночи при +4°С, затем проводили ВЭЖХ-гель-фильтрацию на колонке Superdex S200, эффективность комплексообразования оценивали по площади пика свободного rhVEGF121 в сравнении с соответствующим показателем для препарата Lucentis® (Novartis). Данные приведены в таблице 5.

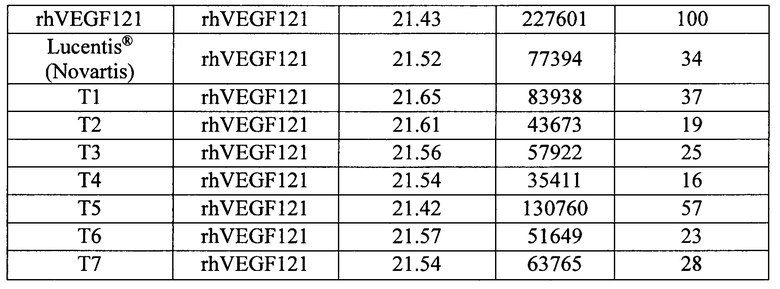
Как видно из данных, представленных в таблице 5, площадь пика свободного rhVEGF121 при эквимолярном соотношении Lucentis® (Novartis) и rhVEGF121 составляет 34% от площади пика rhVEGF121 без добавления какого-либо связывающего его агента. За исключением Т5, площадь пика свободного rhVEGF121 для которого составила 57%, соответствующий показатель для других белков на основе F(ab’)2-фрагмента ранибизумаба варьирует от 16% до 37%, что доказывает высокую антигенсвязывающую способность разработанных молекул, не уступающую таковой характеристике для луцентиса.
Пример 13. Ингибирование rhVEGF121-индуцированной пролиферации клеток HUVEC белками на основе Е(ab’)2-фрагмента ранибизумаба.
Было проведено сравнительное определение биологической активности препаратов белков на основе F(ab’)2-фрагмента ранибизумаба и препарата Lucentis® (Novartis) в тесте ингибирования rhVEGF121-индуцированной пролиферации первичной культуры человеческих эндотелиоцитов пупочной вены (HUVEC, Lonza, Cat. № СС2517). Для этого клетки HUVEC I пассажа (P1) (Lonza, Cat. № СС2517) размораживали и высевали в полную ростовую среду EGM-2 (Endothelial Growth Medium) во флакон Т25, предварительно покрытый раствором 0.1% желатина. После достижения клетками монослоя (Р2) их снимали с пластика с помощью обработки раствором аккутазы и высевали во флакон Т75, также предварительно покрытый раствором 0.1% желатина. Когда клетки достигали максимальной плотности (Р3) их высевали в покрытые желатином лунки 96-луночного плоскодонного планшета в количестве 10 тысяч клеток на лунку в полной ростовой среде EGM-2 (пассаж 3). На следующий день клетки HUVEC дважды аккуратно отмывали стерильным раствором PBS и инкубировали в среде ЕВМ с добавлением 0,2% FBS (среда для постановки теста) на протяжении 2.5-4 часов. Препараты белков на основе F(ab’)2-фрагмента ранибизумаба, Lucentis® и rhVEGF121 разводили в среде для постановки теста в соответствии со схемой эксперимента. Затем каждый из тестируемых препаратов в серии 8 последовательных 3-х-кратных разведений от 100 до 0.05 nM инкубировали с rhVEGF121 (125 ng/ml) в течение 2 часов, после чего вносили прединкубированные растворы в планшеты с клетками HUVEC согласно схеме эксперимента. На каждом планшете были дополнительно сделаны следующие контроли - лунки, содержащие только rhVEGF121 (125 ng/ml); лунки, содержащие полную ростовую среду, и лунки со средой для постановки теста. Все комбинации тестируемых веществ были добавлены к клеткам в трипликатах. Планшеты помещали в С02-инкубатор и инкубировали при 37С и 5% СО2 в течение 72 часов. После окончания инкубации для оценки жизнеспособности клеток в лунки планшетов добавляли реагент ХТТ (Roche, Cat. №11465015001) согласно инструкции производителя и дополнительно инкубировали планшеты при 37С и 5% СО2 в течение 4-6 часов. Оптическую плотность измеряли при длине волны 492 nM и референсной длине волны 690 nM. Процент ингибирования VEGF-A-индуцированной пролиферации клеток HUVEC вычисляли по формуле:
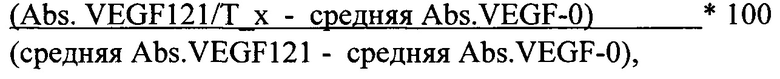
где "средняя Abs. VEGF121 - средняя Abs. VEGF-0" соответствует разности средней оптической плотности в лунках с добавлением rhVEGF121 (125 ng/ml) и средней оптической плотности в лунках со средой для постановки теста, т.е. максимальной пролиферации клеток HUVEC за время теста;
"Abs. VEGF121/Т-х" соответствует оптической плотности в лунке, содержащей исследуемый белок в определенной концентрации и rhVEGF121, а
"средняя Abs. VEGF-0" соответствует средней оптической плотности лунок, содержащих только тестовую среду без добавления ростовых добавок.
Полученные данные обсчитывали в программе GraphPad Prism версии 6.0 с использованием 4-х параметрической логистической функции «доза-ответ - ингибирование», данные представлены в таблице 6.
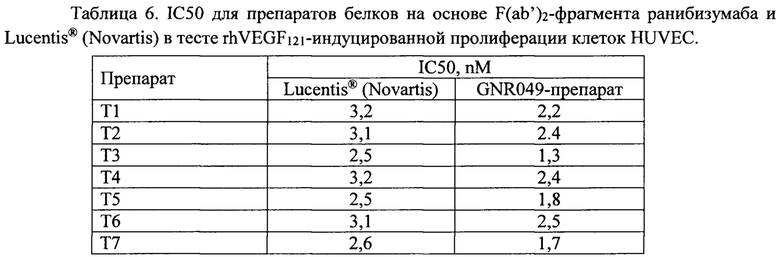
Как видно из данных, представленных в таблице 6, препараты белков на основе F(ab')2-фрагмента ранибизумаба не уступают по биологической активности препарату Lucentis® (Novartis) в тесте ингибирования rhVEGF121-индуцированной пролиферации клеток HUVEC in vitro. Данный тест обладает высокой предсказательной способностью антиангиогенной эффективности препаратов in vivo, в том числе, при введении человеку. Полумаксимальная концентрация ингибирования (IC50) rhVEGF121-индуцированной пролиферации препарата Lucentis® (Novartis) варьировала от 2,5 до 3,2 nM, среднее значение - 2,9 nM, стандартное отклонение - 0,3 nM. IC50 препаратов белков на основе F(ab’)2-фрагмента ранибизумаба изменялось от 1,3 nM до 2,5 nM, что говорит о том, что все белки обладают высокой антиангиогенной активностью, сопоставимой с таковым показателем для «золотого стандарта» в лечении офтальмологических заболеваний, патогенез которых обусловлен высокой активностью фактора роста эндотелия сосудов - ранибизумаба.
Пример 14. Доклинические исследования in vivo - исследование фармакокинетики и токсикокинетики белков на основе F(ab’)2-фрагмента ранибизумаба при однократном интравитреальном введении кроликам породы Советская шиншилла.
Целью исследования было сравнительное определение основных параметров фармакокинетики и токсикокинетики с оценкой офтальмотоксичности и возможного общего токсического действия белков на основе F(ab’)2-фрагмента ранибизумаба и препарата Lucentis® (Novartis) при однократном интравитреальном введении кроликам породы Советская шиншилла. Исследование было выполнено в ЗАО «НПО «Дом фармации» (Россия, Ленинградская область, Всеволожский муниципальный район, Кузьмоловское городское поселение) в соответствии с основными принципами надлежащей лабораторной практики и этическими нормами по охране животных, используемых в научных целях.
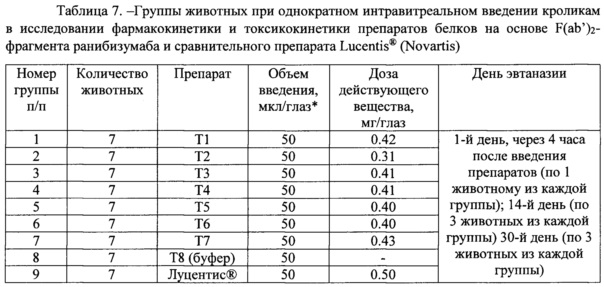
Задачи исследования включали изучение влияния тестируемых объектов на биометрические параметры исследуемых животных - смертность и динамику массы тела; изучение влияния тестируемых объектов на клиническое состояние структур глаза исследуемых животных; офтальмоскопическую оценку влияния тестируемых объектов на состояние глазного дна экспериментальных животных. В таблицах 7 и 8 представлены основные группы животных, доза препаратов для интравитреального введения и основные процедуры, выполненные в ходе исследования.
Материалы и методы. Исследование проводили на половозрелых самцах кроликов породы Советская шиншилла. Тестируемые объекты вводили животным интравитреально в дозе 50 мкл в каждый глаз животного. Объем введения тестируемых объектов для исследования фармакокинетики с оценкой офтальмотоксичности был выбран, опираясь на биоэтические принципы, учитывая травматичность и болезненность процедуры интравитреального введения, а также на основании данных о максимальных рекомендуемых объемах интравитреального введения кроликам. Был использован интравитреальный путь введения, так как этот способ введения является планируемым способом применения в клинической практике.


Интравитреальные инъекции осуществляли после наркотизации животных путем внутривенного введения им смеси препаратов Золетил®100 (Virbac, Франция) в дозе 1,5 мг/кг и Рометар® (Bioveta, Чехия) в дозе 1,5 мг/кг (СОП ЭЖ-20). Смесь анестетиков в данной дозе позволяла достичь оптимальной длительности наркоза для проведения инъекций в оба глаза животного и обеспечивала полное отсутствие роговичного рефлекса на всей продолжительности проведения манипуляций. Для местного обезболивания в конъюнктивальный мешок вносили раствор местного анестетика (Инокаин®, глазные капли, Промед Экспортс ПВТ. ЛТД). Очередность манипуляций при выполнении интравитреальной инъекции была следующей: 1) внесение раствора мидриатика (Ирифрин®, глазные капли, Промед Экспортс ПВТ. ЛТД) в конъюнктивальный мешок, ожидание 5 минут; 2) промывание поверхности роговицы стерильным физиологическим раствором; 3) внесение раствора местного анестетика (Инокаин®) в конъюнктивальный мешок, ожидание 3 минуты; 4) промывание поверхности роговицы стерильным физиологическим раствором; 5) внесение препарата Бетадин, 0,5% раствор (Pharmaceuticals PLC (Венгрия)); 6) проведение иглы одноразового инсулинового шприца через склеру на расстоянии 2-3 мм от лимба в направлении к экватору в верхнее-наружном квадранте; проведение иглы вглубь стекловидного тела на 2-3 мм параллельно хрусталику.
Осмотр животных в клетках содержания, с целью выявления смертности или признаков отклонения в состоянии здоровья, проводили ежедневно.
Лишение корма - животные были лишены утреннего кормления до процедуры введения и взвешивания животных. Доступ к воде не был ограничен.
Массу тела регистрировали в утренние часы непосредственно перед введением, далее еженедельно. Процедуру проводили на весах настольных медицинских МТ "Карапуз" 20 ВЖА-(5г,Р)бА, зав. №488688. Наименьший предел взвешивания - 100 г. Наибольший предел взвешивания - 20 кг. Цена поверочного деления 5 г. Класс точности 3.
Клинический осмотр видимых структур глаза проводили непосредственно перед началом исследования, далее - ежедневно. Осуществляли полуколичественную оценку параметров офтальмотоксичности в баллах. При наличии патологического признака присваивали 1 балл, при отсутствии признака - балл был равен 0. Оценивали следующие показатели офтальмотоксичности:
- Ксерофтальмия (сухость роговицы и конъюнктивы глаза, возникающая из-за нарушения слезоотделения);
- Кератит (воспаление роговицы глаза, сопровождающееся ее помутнением);
- Слезотечение;
- Нагноение;
- Отечность;
- Покраснение;
- Экзофтальм (смещение кпереди глазных яблок);
- Мидриаз (расширение зрачка);
- Миоз (значительное сужение зрачка, физиологическая рефлекторная реакция, которая возникает при освещении глаз светом большой яркости).
Офтальмоскопическую оценку состояния глазного дна осуществляли до начала исследования, на 2-й, 13-й и 29-й дни после инъекций препаратов путем прямой офтальмоскопии и осмотра переднего отдела глаза с помощью лупы (40 дптр) после введения в каждый глаз препарата Атропин (капли глазные 1% (Московский эндокринный завод (Россия)). Оценивали патологические изменения в области желтого пятна, центральной области сетчатки, диска зрительного нерва и сосудов сетчатки.
Отбор крови (по 1 мл) осуществляли в стерильные пробирки с ЭДТА через 4, 8, 24, 48, 72, 96, 192 и 384 часов после введения препаратов. Образцы крови центрифугировали для получения плазмы. Каждый образец плазмы делили на две части, помещали в промаркированные криоустойчивые пробирки, маркировка каждой пробирки включала номер экспериментальной группы, номер животного, вид биообразца, дату и время забора материала. Все полученные образцы замораживали, хранили при температуре -70° и транспортировали Спонсору исследования для проведения дальнейших анализов в сухом льду.
Эвтаназию животных в соответствии с Директивой 2010/63/EU Европейского Парламента и Совета Европейского Союза по охране животных, используемых в научных целях от 22 сентября 2010 г., проводили путем передозировки средствами для наркоза. Эвтаназию осуществляли с помощью препаратов Золетил® и Рометар®, после введения животных в глубокий наркоз осуществляли внутривенное введение препарата Лидокаин (Дальхимфарм (Россия)). Данный вид эвтаназии является гуманным и не противоречит нормативным документам, сопровождается минимумом боли, страдания и дистресса, и был проведен компетентными сотрудниками.
После эвтаназии животных производили забор полного объема стекловидного тела и водянистой влаги. Каждый биологический образец делили на две части, помещали в маркированные криоустойчивые пробирки, маркировка каждой пробирки включала в себя номер экспериментальной группы, номер животного, вид биообразца, дату и время забора материала. Все полученные образцы замораживали, хранили при температуре -70° и транспортировали Спонсору исследования для проведения дальнейших анализов в сухом льду.
Статистический анализ данных массы тела животных - данные проверяли на соответствие закону нормального распределения с помощью критерия Шапиро-Уилка. В случае нормального распределения считали среднее значение и стандартную ошибку среднего, которые вместе со значением n (количество наблюдений) представлены в итоговых таблицах. Для оценки данных с признаками нормального распределения использовали однофакторный дисперсионный анализ (ANOVA), предназначенный для сравнения нескольких выборок (групп) и вычисления общего уровня значимости различий. Статистический анализ выполнялся с помощью программного обеспечения Statistica 6.0. (StatSoft, USA).
Заключения и выводы ЗАО «НПО «Дом фармации» об оценке офтальмотоксичности и общего токсического действия препаратов белков на основе F(ab')2-фрагмента ранибизумаба и сравнительного препарата Lucentis® (Novartis) при однократном интравитреальном введении кроликам породы Советская шиншилла:
1. Тестируемые объекты T1, Т2, Т3, Т4, Т5, Т6, Т7, Т8, так же, как и референсный препарат Луцентис®, не повлияли на динамику массы тела животных.
2. Во всех группах (T1, Т2, Т3, Т4, Т5, Т6, Т7, Т8,) не было статистически значимых отклонений от контрольных групп по частоте возникновения изучаемых клинически значимых отклонений.
Результаты оценки параметров фармакокинетики и токсикокинетики препаратов белков на основе F(ab’)2-фрагмента ранибизумаба при однократном интравитреальном введении кроликам породы Советская шиншилла, полученные в ООО «МБЦ «Генериум»
С целью определения концентрации препаратов в биологических образцах кроликов был разработан специфичный и высоко чувствительный метод в варианте «сэндвич»-ИФА на основе поликлональных крысиных антител к ранибизумабу, полученных в ООО «МБЦ «Генериум».
Кратко, поликлональные крысиные антитела к ранибизумабу вносили в лунки ИФА планшетов по 100 μl в концентрации 1 μg/ml в карбонат-бикарбонатном буфере (pH 9.5) и иммобилизовали в течение ночи при +4С. Остаточные свободные центры связывания пластика блокировали раствором PBS-Tha («ПанЭко», Кат. № Э4100-110) в течение 1 часа при комнатной температуре. После этого в лунки планшетов вносили биологические образцы, разведенные раствором PBS-0,05%Tw20, в последовательных 2-кратных разведениях. Планшеты инкубировали в течение 1 часа, отмывали несвязавшиеся антитела и вносили раствор биотинилированных поликлональных кроличьих антител к ранибизумабу (ООО «МБЦ «Генериум»). После инкубации в течение 1 часа отмывали несвязавшиеся антитела и добавляли конъюгированный с пероксидазой хрена стрептавидин в разведении, рекомендованном производителем (Abcam, ab7403). Инкубировали 40 минут при комнатной температуре и вносили субстрат для пероксидазы хрена. В качестве субстрата для HRP использовали ТМБ (США, Партия: TMBPD10222-2-10273-А), реакцию останавливали раствором 2М H2SO4. Реакцию оценивали по оптической плотности, измеренной на микропланшетном спектрофотометре Bio-Rad xMark при длине волны 450 нм. Полученные данные обсчитывали с помощью программного обеспечения Bio-Rad Microplate Manager 6 для микропланшетного спектрофотометра.
Концентрация исследуемых препаратов в стекловидном теле после однократного ИВТ введения и предварительная оценка соотношения периодов полувыведения из стекловидного тела препаратов белков на основе F(ab’)2-фрагмента ранибизумаба и препарата Lucentis® (Novartis)
Дизайн проведенного сравнительного скринингового исследования включал всего три временных точки отбора биологических образцов, в соответствии с основными принципами этических норм обращения с лабораторными животными и минимизации количества экспериментальных животных, поскольку основной задачей исследования было сравнительное изучение параметров фармакокинетики при ИВТ введении большой панели изучаемых препаратов и препарата Lucentis®. Мы не стремились к точному определению параметров фармакокинетики, таких как скорость и период полувыведения препаратов из стекловидного тела, эта задача потребовала бы значительно больше лабораторных животных, а хотели лишь установить соотношение данных параметров у каждого из изучаемых препаратов и препарата Lucentis®. Образцы стекловидного тела забирали у экспериментальных животных после эвтаназии и энуклеации глаз на 1-й день (через 4 часа), на 14-й и 30-й день после ИВТ инъекций. Концентрацию препаратов белков на основе F(ab’)2-фрагмента ранибизумаба определяли описанным выше методом относительно соответствующей калибровочной кривой для каждого из изучаемых белков.
На рисунках 2-4 приведены графики зависимости медиан концентрации определяемых белков в стекловидном теле, водянистой влаге и плазме крови в молярном выражении, от времени забора биологических образцов после однократной ИВТ инъекции.
Рассчитывали один из основных критериев фармакокинетики, отражающий экспозицию каждого из исследуемых препаратов в стекловидном теле, - площадь под каждой кривой (AUCIVT, нМ/(мл*ч), (рис 33), в соответствии с линейным правилом трапеций с помощью приложения Excel PK Solver 2.0. Для расчета скорости элиминации (клиренса, Cl) использовали следующую формулу;
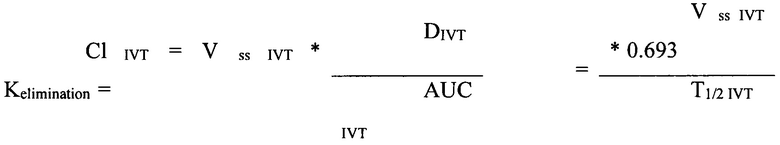
где Vss IVT - равновесный объем распределения в стекловидном теле, DIVT - ИВТ доза, Т1/2 IVT - период полувыведения из стекловидного тела.
Исходя из предположения, что равновесный объем распределения в стекловидном теле одинаков для всех исследованных молекул и для Lucentis®, получается следующее равенство:
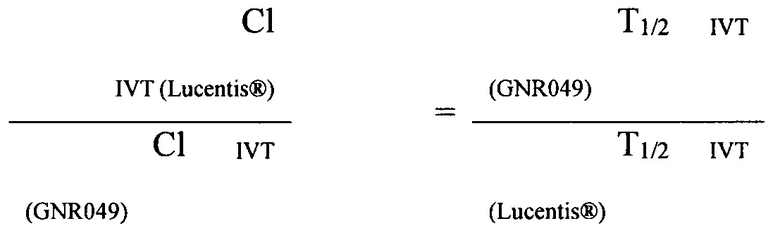
Данные расчетов приведены в таблице 9.
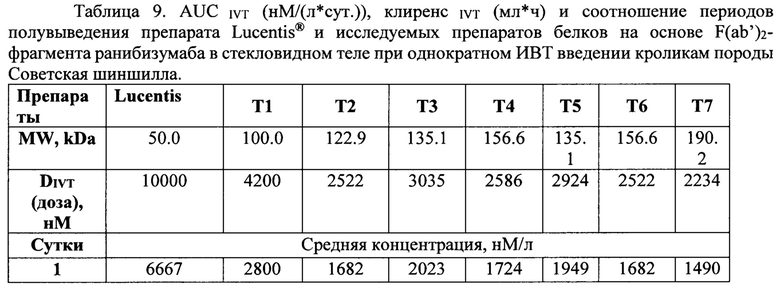
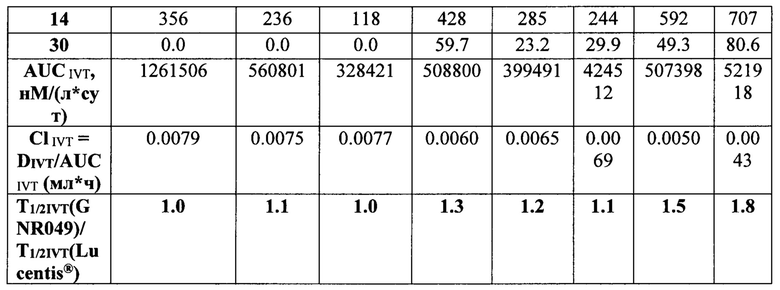
Как видно из данных таблицы 9, максимальное время экспозиции в стекловидном теле было определено для препарата Lucentis® при его большей, относительно других препаратов, молярной DIVT - 6667 нМ, молярные ИВТ дозы других препаратов были значительно (от 4.5 до 2.4 раза) меньше DIVT луцентиса и, в соответствии с этим, их показатели AUCIVT были меньше показателя AUCIVT для Lucentis® от 2.3 до 3 раз. При самой малой DIVT препарата Т7 (меньше в 4.5 раза дозы Lucentis®) его показатель AUCIVT был меньше соответствующего значения препарата Lucentis® лишь в 2.5 раза. Скорость элиминации исследуемых молекул была достоверно меньше для препаратов белков на основе F(ab’)2-фрагмента ранибизумаба, модифицированных НГП, чем для препарата Lucentis® и немодифицированных НГП вариантов белков. Самая низкая скорость элиминации была показана для препарата Т7. Соотношение периодов полувыведения исследуемых препаратов и препарата Lucentis® из стекловидного тела было больше 1 для всех модифицированных НГП вариантов, а для вариантов Т6 и Т7 данное соотношение составило 1.5 и 1.8, соответственно. Это означает, что период полувыведения последних больше T1/2IVT препарата Lucentis® как минимум в 1.5 и 1.8 раза, поскольку расчет AUCIVT, на основании которого рассчитывали скорость выведения, производили только до 30 дня, когда уже не определялась концентрация Lucentis®, но еще достоверно определялись и Т6, и Т7. Для точного определения периода полувыведения препаратов Т6 и Т7 из стекловидного тела необходимо провести дополнительное исследование с большим количеством временных точек отбора в интервале от 1 до 45-60 дня исследования.
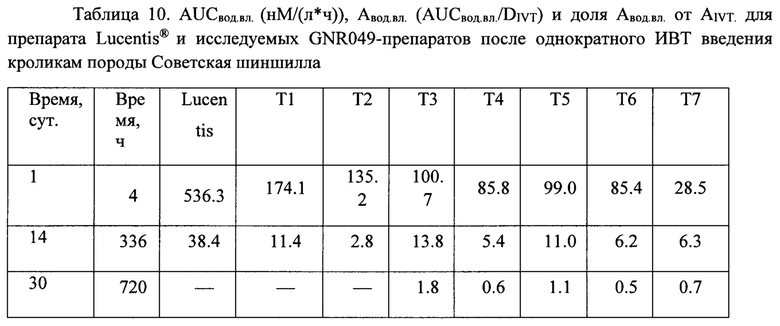
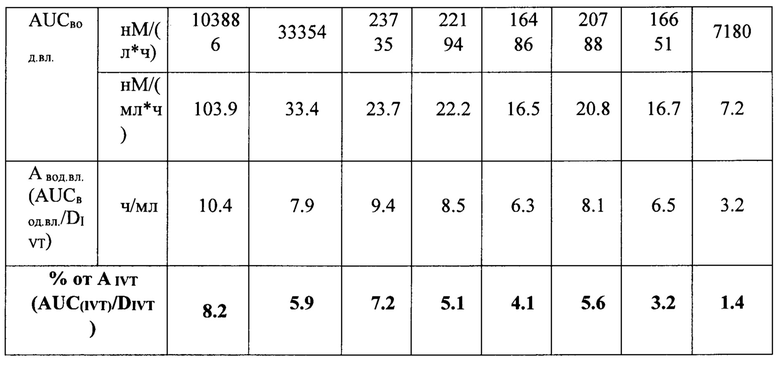
Данные, представленные в таблице 10, говорят о том, максимальные медианы концентрации препаратов в водянистой влаге были определены через 4 часа после ИВТ инъекций, максимальное значение медианы концентрации через 4 часа наблюдали для препарата луцентис, минимальное - для препарата Т7. На 14 сутки концентрация препаратов во всех группах снижалась, на 30-е сутки в образцах водянистой влаги определяли только модифицированные НГП варианты белков, что свидетельствует о том, что на 30-й день исследования данные препараты еще содержались в стекловидном теле, в отличие от луцентиса и немодифицированных НГП белков, поскольку в водянистую влагу белки поступают именно из стекловидного тела. Данный факт согласуется с данными определения концентрации исследуемых препаратов в стекловидном теле. Показатель AUCвод.вл. был максимальным для препарата луцентис в соответствии с его DIVT, поэтому для более корректного сравнения данного показателя между экспериментальными группами мы вычисляли нормированный относительно соответствующей DIVT показатель AUCвод.вл.-Авод.вл., который отражает эффективность поступления препарата из стекловидного тела в переднюю камеру глаза. Данный показатель (Авод.вл.) был максимальным для луцентиса и минимальным - для препарата Т7. Процентная доля показателя Авод.вл. от соответствующего нормированного показателя в стекловидном теле - AIVT была также максимальной для препарата луцентис и минимальной - для препарата Т7, данный показатель также отражает эффективность выведения препаратов из стекловидного тела через водянистую влагу передней камеры глаза.
В таблице 11 представлены основные фармакокинетические параметры исследуемых препаратов Т6 и Т7, для которых показано увеличенное время экспозиции в стекловидном теле, в сравнении с препаратом Lucentis®.
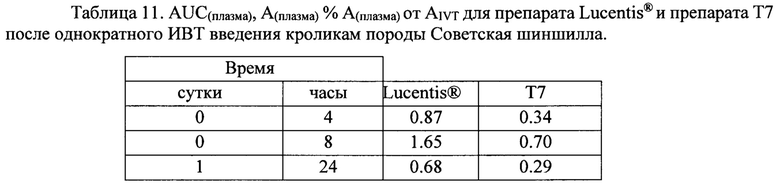
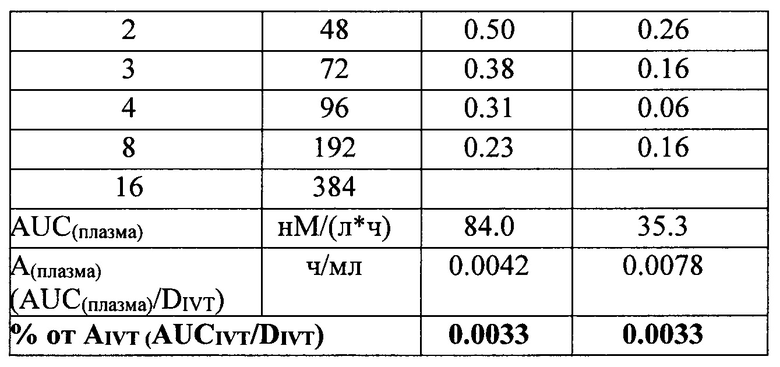
Вычисленное значение AUC(плазма) для препарата Lucentis® в 2.4 раза больше, чем соответствующее значение для препарата GNR049/T7, но ИВТ доза препарата Lucentis® в 4 раза превышает DIVT препарата GNR049/T7. Нормированные относительно DIVT значения AUC(плазма)-А(плазма) более информативно отражают как способность препаратов поступать в кровоток, так и время их экспозиции в кровеносном русле. Значение А(плазма) для препарата Т7 больше соответствующего значения для препарата Lucentis® лишь в 1.9 раза, что при доказанной безопасности применения препарата Lucentis® в клинической практике, скорее всего, не приведет к нежелательным явлениям, обусловленным системным влиянием препарата Т7, который представляет модифицированный вариант препарата Lucentis®.
Таким образом, новый белок Т7 экспрессируется в эукариотической системе с приемлемой для производства продуктивностью, обладает высокой, сопоставимой с ранибизумабом, аффинностью к VEGF-A человека и кролика, высокой эффективностью в ингибировании VEGF-А-индуцированной пролиферации клеток HUVEC, предварительно оцененный период полувыведения Т7 из стекловидного тела при ИВТ введении кроликам примерно в два раза превышает соответствующий показатель для ранибизумаба при сопоставимых показателях системного влияния этих препаратов.
Изобретение относится к области медицины, а именно к офтальмологии, и предназначено для лечения заболевания, сопровождающегося отеком макулы вследствие повышенной экспрессии VEGF-A. Полипептид содержит ранибизумаб с добавленным F(аb')-фрагментом и содержит на тяжелой цепи hinje-участок с цистеиновыми остатками для образования дисульфидных связей. Указанный полипептид вводят в стекловидное тело. Полипептид дополнительно может содержать полипептидные последовательности, выбранные из домена СН3 антитела, неструктурированных полипептидов, в том числе НАР-полипептидных последовательностей S(G4S)16 и S(G4S)20, или их комбинации. Использование изобретения позволяет повысить удобство анти-VEGF-A терапии при отеке макулы. 1 з.п. ф-лы, 4 ил., 11 табл., 14 пр.
1. Полипептид, содержащий ранибизумаб с добавленным F(аb')-фрагментом и содержащий на тяжелой цепи hinje-участок с цистеиновыми остатками для образования дисульфидных связей, для лечения и/или профилактики заболевания, в отношении которого указанное антитело имеет терапевтическую активность, при его введении в стекловидное тело.
2. Полипептид по п. 1, отличающийся тем, что дополнительно содержит полипептидные последовательности, выбранные из домена СН3 антитела, неструктурированных полипептидов, в том числе НАР-полипептидных последовательностей S(G4S)16 и S(G4S)20, или их комбинации.
| MAGDELAINE-BEUZELIN C et al | |||
| Therapeutic antibodies in ophthalmology | |||
| Old is new again | |||
| MAbs, 2010, 2(2), p.176-180 | |||
| СПОСОБЫ ИНГИБИРОВАНИЯ ГЛАЗНОГО АНГИОГЕНЕЗА | 2009 |
|
RU2530583C2 |
| СПОСОБ ХИРУРГИЧЕСКОГО ЛЕЧЕНИЯ СУБРЕТИНАЛЬНЫХ КРОВОИЗЛИЯНИЙ НА ФОНЕ ВОЗРАСТНОЙ МАКУЛОДИСТРОФИИ С СУБРЕТИНАЛЬНОЙ НЕОВАСКУЛЯРНОЙ МЕМБРАНОЙ | 2008 |
|
RU2366388C1 |
| JP 2012072152 A, 12.04.2012 | |||
| КОРОЛЬ А.Р | |||
| и др | |||
| Влияние интравитреального введения ранибизумаба, пегаптаниба натрия, бевацизумба на гемодинамику глаз кроликов | |||
| Таврический медико-биологический вестник, 2012, том 15, no.3, ч.3 (59), c.74-77. | |||

