Область техники, к которой относится изобретение
[0001] Изобретение относится к новому PSMA-связывающему антителу. Антитело к PSMA по изобретению не конкурирует перекрестно с известным из уровня техники PSMA-связывающим антителом J591, и характеризуется сниженной степенью индукции антигенного сдвига по сравнению с J591 и уникальной реактивностью в отношении клеток плоскоклеточной карциномы (SCC) разного происхождения. Кроме того, изобретение относится к молекуле биспецифического антитела PSMA×CD3. Изобретение также относится к способам получения молекулы антитела по изобретению, а также нуклеиновым кислотам, векторам и клеткам-хозяевам. Изобретение дополнительно относится к способам лечения или диагностирования заболевания с применением молекулы антитела к PMSA по изобретению.
Сведения о предшествующем уровне техники
[0002] В ходе научной работы, начавшейся в 1980-х годах, было установлено, что биспецифические антитела, направленные на опухоль-ассоциированный антиген (TAA) и комплекс T-клеточный рецептор (TCR)/CD3, способны активировать T-клетки, что приводит в результате к лизису экспрессирующих TAA опухолевых клеток активированными T-клетками (Staerz et al., Nature 1985, 314:628-631; Perez et al., Nature 1985, 316:354-356; Jung et al., Proc Natl Acad Sci USA 1986, 83:4479-4483). Поскольку CD3-антитела, связывающиеся с Fc-рецепторами (FcR) посредством их Fc-части, являются весьма эффективными в индуцировании активации T-клеток и высвобождении цитокинов, как нежелательных побочных эффектов, первостепенное значение имеет конструирование биспецифических TAA×CD3-антител, лишенных Fc или содержащих Fc с ослабленной функцией, с целью предотвращения связывания FcR и обеспечения ограниченной целевой клеткой, а не опосредованной FcR активации T-клеток (Jung et al., Immunol Today 1988; 9:257-260; Jung et al., Eur J Immunol 1991; 21:2431-2435).
[0003] Получение биспецифических антител, удовлетворяющих этому крайне важному требованию в контексте качества и количества в производстве, остается серьезной проблемой. Недавно было продемонстрировано, что рекомбинантное биспецифическое одноцепочечное (bssc) антитело с CD19×CD3-специфичностью, под названием блинатумомабом, являются весьма эффективными в лечении пациентов с ALL (Bargou et al.. Science 2008, 321:974-977), и оно прошло апробацию FDA, получив статус инновационного терапевтического средства. Следует отметить, что лекарственное средство применяют в виде непрерывной 24-часовой инфузии на протяжении нескольких недель ввиду его небольшого времени полужизни в сыворотке крови и довольно высокой токсичности: безопасно применяемые дозы составляют примерно 30 мкг на пациента в день, что в 10000 раз меньше доз, применяемых для лечения общепринятыми моноспецифическими противоопухолевыми антителами (Adams and Weiner. Nat Biotechnol 2005, 23:1147-57). Итоговые концентрации лекарственного средства в сыворотке крови составляют менее 1 нг/мл (Topp et al., J Clin Oncol 2011; 29:2493-2498). Такое существенное ограничение по дозе, также наблюдаемое в более ранних клинических испытаниях для разных биспецифических антител (Kroesen et al., Br J Cancer 1994; 70:652-661; Tibben et al., Int J Cancer 1996; 66:477-483), обусловлено нецелевой активацией T-клеток, приводящей в результате к системному высвобождению цитокинов. Очевидно, что такое явление препятствует оптимальной терапевтической активности биспецифических антител, стимулирующих комплекс TCR/CD3.
[0004] В целом, дозолимитирующая нецелевая активация T-клеток и возникающая в результате, связанная с токсичностью, проблема могут быть обусловлены проблемами P1 и P2, обсуждаемыми более детально далее; небольшое время полужизни в сыворотке крови обсуждается как проблема P3.
[0005] (P1): TAA, являющийся мишенью биспецифического антитела, не является абсолютно опухолеспецифическим, что приводит к опосредованной антителом активации T-клеток в результате связывания с нормальными экспрессирующими TAA клетками. Строго говоря это не относится к нецелевой активации, поскольку индуцируется экспрессирующими антиген целевыми клетками, хоть и «неправильными», то есть нормальными, а не злокачественными клетками. Блинатумомаб, упоминаемое выше биспецифическое антитело к CD19×CD3, определенно сталкивается с этой проблемой, поскольку его целевой антиген CD19 экспрессируется на поверхности нормальных B-лимфоцитов. Очевидно, специфичность антигена-мишени в отношении злокачественной ткани является решающей для предотвращения нецелевой активации T-клеток такого типа. В этом отношении PSMA является особенно подходящим антигеном, поскольку обширный иммуногистологический анализ позволил выявить, что экспрессия этого антигена в нормальной ткани ограничивается эпителием предстательной железы, молочной железой и проксимальными канальцами почки [Атлас белков человека, http://www.proteinatlas.org]. В злокачественной ткани указанный антиген в больших количествах экспрессируется на поверхности клеток карциномы предстательной железы и в ряде других солидных опухолей, таких как карцинома толстой кишки, молочной железы и поджелудочной железы и глиобластома (Chang et al., Cancer Res 1999, 59:3192; Ross et al., Cancer Met Rev 2005, 24:521). При таких последних опухолях, однако, экспрессия PSMA строго ограничивается сосудистой системой и «скрывает» собственно опухолевые клетки. Примечательно, что при карциноме предстательной железы, единственной известной на данный момент опухоли с экспрессией на поверхности опухолевых клеток, в большинстве случаев в клетках сосудистой системы экспрессия PSMA отсутствует (Chang et al., 1999), так что оптимальная ситуация, когда имеет место экспрессия в клетках сосудистой системы, а также на поверхности собственно опухолевых клеток, встречается редко (P1.1).
[0006] Помимо его специфичности, другое свойство антитела против клетки-мишени может иметь решающее значение для его терапевтической активности: антитело может быть причиной антигенного сдвига в результате «слущивания» или поглощения антигена целевой клеткой. Поглощение антигена является желательным в случае иммунотоксина, который представляет собой конструкцию, содержащую антитело и токсин, для которого обычно требуется поглощение клеткой для проявления его активности. Однако если антитела применяют для рекрутирования иммунных эффекторных клеток, антигенный сдвиг, независимо от механизма, может препятствовать активности антител. Действительно, было продемонстрировано, что терапевтические антитела к CD20 индуцируют антигенный сдвиг среди разных клеток лимфомы до различной степени, и что это явление, по меньшей мере частично, ответственно за непостоянные терапевтические эффекты этих антител (Glennie et al., Mol Immunol. 2007; 44:3823). В любом случае, в контексте активирующих T-клетки биспецифических антител, по-видимому, желательным является выбор антител против клеток-мишеней, которые индуцируют минимальный антигенный сдвиг (P1.2).
[0007] (P2): активация T-клеток не ограничивается, как это должно быть, целевой клеткой, то есть, даже моновалентный эффекторный сайт связывания CD3 в составе биспецифической конструкции антитела способен индуцировать некоторую активацию T-клеток в отсутствие целевых клеток, с которыми связывается антитело посредством его нацеливающего компонента. Это представляет собой нецелевую активацию в строгом смысле, поскольку для клеток, несущих целевой антиген, не требуется индукция данного явления. Авторы изобретения заметили, что степень выраженности этого явления существенно варьирует, если применяют разные антитела к CD3 в разных форматах, и если добавляют определенные стимулирующие «фоновые» клетки (SBC), такие как клетки лимфомы (SKW6.4) или эндотелиальные клетки (HUVEC), которые обеспечивают костимуляцию для активации T-клеток. Таким образом, для конструкции биспецифических антител следует выбирать CD3-связывающий компонент, индуцирующий минимальную «нецелевую» активацию T-клеток (P2.1).
[0008] В дополнение к активации T-клеток, индуцированной собственно стимуляцией мономерного CD3, в недавно опубликованной статье говориться об альтернативном механизме нецелевой активации, в который вовлечена нацеливающая часть биспецифического антитела; если такая часть состоит из одноцепочечного фрагмента, который индуцирует кластеризацию эффекторной части биспецифического антитела на поверхности T-клетки, может быть индуцирована фоновая передача сигнала, которая приводит в результате к истощению T-клеток (Long et al., Nat Med 2015; 6:581), что почти невозможно выявить с помощью традиционных быстрых in vitro анализов, но сильно влияет на эффективность in vivo. Эти наблюдения проводили с применением T-клеток, трансфицированных химерным антигенным рецептором (CAR-T-клетки). Химерные T-клеточные рецепторы содержат одноцепочечные антитела в качестве нацеливающих компонентов. Весьма вероятно, что результаты Long et al. (2015) также применимы по отношению к биспецифическим антителам с такой нацеливающей частью, поскольку эти реагенты после связывания с T-клеткой функционально эквивалентны в отношении Т-клетки, трансфицированной соответствующим CAR. В данной области хорошо известно, что большинство одноцепочечных антител имеют склонность к образованию мультимеров и агрегатов (Worn et al., J Mol Biol 2001, 305:989-1010), и, следовательно, не удивительно, что для всех, кроме одного из CAR, тестируемых Long et al. (2015), наблюдали явление кластеризации и фоновой передачи сигнала с участием CD3, хоть и в различной степени (Long et al., 2015). Для решения описанной в данном описании проблемы (P.2.2) требуется биспецифический формат, который обеспечивает предотвращение мультимеризации нацеливающей части и обусловленной ею кластеризации.
[0009] (P3): наиболее значительным недостатком биспецифических форматов является очень малое время полужизни в сыворотке крови (1-3 часа) ввиду сниженной молекулярной массы и отсутствия CH3-доменов. По этой причине являющееся прототипом антитело блинатумомаб применяют путем непрерывной 24-часовой i.v. инфузии в течение нескольких недель. Применение полных форматов на основе IgG с увеличенным временем полужизни в сыворотке крови, таких как IgGsc, представленное на фиг. 1B, оказалось неподходящим, поскольку возможное повышение нецелевой активации индуцировалось бивалентным C-концевым CD3-связывающим компонентом.
[0010] Исходя из вышесказанного, в данной области существует потребность в улучшенных молекулах антител, которые позволяют решить по меньшей мере одну из вышеописанных проблем.
Сущность изобретения
[0011] Изобретение относится к молекуле антитела или ее антигенсвязывающему фрагменту, способным к связыванию с простатическим специфическим мембранным антигеном (PSMA) человека, содержащим: (i) вариабельный домен тяжелой цепи, содержащий CDRH1-область, изложенную в SEQ ID NO: 03 (GFTFSDFYMY), CDRH2-область, изложенную в SEQ ID NO: 04 (TISDGGGYTSYPDSVKG), и CDRH3-область, изложенную в SEQ ID NO: 05 (GLWLRDALDY), или содержащий последовательности CDRH1, CDRH2 или CDRH3, характеризующиеся по меньшей мере 75% идентичностью последовательности или по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 03, SEQ ID NO: 04 или SEQ ID NO: 05; и (ii) вариабельный домен легкой цепи, содержащий CDRL1-область, изложенную в SEQ ID NO: 06 (SASSSISSNYLH), CDRL2-область, изложенную в SEQ ID NO: 07 (RTSNLAS), и CDRL3-область, изложенную в SEQ ID NO: 08 (QQGSYIPFT), или содержащий последовательности CDRL1, CDRL2 или CDRL3, характеризующиеся по меньшей мере 75% идентичностью последовательности или по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 06, SEQ ID NO: 07 или SEQ ID NO: 08.
[0012] Настоящее изобретение также относится к молекуле антитела или ее антигенсвязывающему фрагменту, способным к связыванию с PSMA человека, которые способны конкурировать за связывание с PSMA человека с молекулой антитела по изобретению или ее антигенсвязывающим фрагментом.
[0013] Настоящее изобретение дополнительно относится к молекуле биспецифического антитела, содержащей (i) вариабельную область, содержащую вариабельный домен тяжелой цепи и вариабельный домен легкой цепи молекулы PSMA-связывающего антитела по изобретению, где указанная вариабельная область содержит первый сайт связывания, способный к связыванию с простатическим специфическим мембранным антигеном (PSMA) человека; и (ii) вариабельную область тяжелой цепи и вариабельную область легкой цепи молекулы антитела, содержащие второй сайт связывания.
[0014] Настоящее изобретение дополнительно относится к фармацевтической композиции, содержащей молекулу антитела по изобретению или ее антигенсвязывающий фрагмент.
[0015] Настоящее изобретение дополнительно относится к молекуле антитела по изобретению или ее антигенсвязывающему фрагменту для применения в диагностике или лечении заболевания.
[0016] Настоящее изобретение дополнительно относится к in vitro способу диагностирования заболевания, предусматривающему приведение полученного от субъекта образца в контакт с молекулой антитела по изобретению или ее антигенсвязывающим фрагментом.
[0017] Настоящее изобретение дополнительно относится к молекуле нуклеиновой кислоты, кодирующей молекулу антитела по изобретению или ее антигенсвязывающий фрагмент, вектору, содержащему указанную молекулу нуклеиновой кислоты, и клетке-хозяину, содержащей указанную молекулу нуклеиновой кислоты или указанный вектор.
[0018] Настоящее изобретение дополнительно относится к способу получения молекулы антитела по изобретению или ее антигенсвязывающего фрагмента, предусматривающему обеспечение экспрессии нуклеиновой кислоты, кодирующей молекулу антитела, в условиях, обеспечивающих экспрессию нуклеиновой кислоты.
Подробное описание чертежей
[0019] Настоящее изобретение будет более понятно со ссылкой на подробное описание, при рассмотрении в сочетании с неограничивающими примерами и прилагаемыми чертежами, в которых:
[0020] На фиг. 1 изображены различные форматы молекул биспецифических антител, которые применяли в изобретении. Представлены биспецифические антитела к PSMA×CD3 в формате Fabsc (фиг. 1A) и формате IgGsc (фиг. 1B). У обоих форматов связывание CH2-домена с Fc-рецепторами предотвращается при определенных аминокислотных модификациях. Также представлен формат bssc (биспецифический одноцепочечный Fv) (фиг. 1C).
[0021] На фиг. 2 показаны результаты нецелевой активации T-клеток разными антителами к PSMA×CD3. PBMC инкубировали с указанными антителами в отсутствие и в присутствии клеток лимфомы SKW6.4. Через 3 дня с помощью проточной цитометрии анализировали экспрессию CD69 T-клеток.
[0022] На фиг. 3 показаны результаты активации T-клеток, которую оценивали по поглощению 3H-тимидина. На фиг. 3A показаны результаты по нецелевой активации T-клеток в отсутствие целевых клеток, а на фиг. 3B представлены, в сравнении, результаты по целевой активации T-клеток антителами к PSMA×CD3 в формате Fabsc и антителами к CD3, содержащими UCHT1 (NPCU) и OKT3 (NPCO) соответственно. На фиг. 3C продемонстрирован лизис экспрессирующих PSMA целевых клеток активированными T-клетками в анализе цитотоксичности Xelligence.
[0023] На фиг. 4 показаны результаты мультимеризации и агрегации разных форматов биспецифических антител. На фиг. 4A и фиг. 4B представлено сравнение антител со специфичностью к FLT3×CD3 (Fabsc- по сравнению с bscc-форматом), а на фиг. 4C-4F представлено сравнение таковых со специфичностью к PSMA×CD3 (Fabsc- по сравнению с IgGsc-форматом). Гель-фильтрацию осуществляли на колонках Superdex S200.
[0024] На фиг. 5 показаны результаты связывания известного из предшествующего уровня техники антитела к PSMA J591 и антитела по изобретению 10B3 с экспрессирующими PSMA клетками. Связывание (фиг. 5A), отсутствие конкурентного связывания (фиг. 5B) и антигенный сдвиг PSMA при связывании антитела (фиг. 5C) оценивали с помощью проточной цитометрии с применением трансфицированных PSMA клеток Sp2/0. На фиг. 5B продемонстрировано, что химерное (ch) J591, специфически выявляемое вторичным антителом козы к IgG человека, вытеснялось J591 мыши (mu), но не 10B3 мыши.
[0025] На фиг. 6 показаны полученные с помощью криостата срезы, окрашенные PSMA-связывающим известным из предшествующего уровня техники антителом J591 и антителом 10B3 по изобретению. На фиг. 6A и фиг. 6B, образец клеток карциномы предстательной железы окрашивали обоими антителами параллельно, и с применением полимерной системы от Zytomed, Берлин, Германия (POLHRP-100), а на фиг. 6C и фиг. 6D, образец плоскоклеточной карциномы окрашивали двумя антителами, снова параллельно, с применением полимерной системы от Zytomed. Стрелочками указаны строма опухоли (Tu) и кровеносные сосуды (Ve). Показаны репрезентативные результаты по 9 из 10 образцам рака предстательной железы и 7 из 10 образцам плоскоклеточной карциномы. В ряде различных нормальных тканей человека (полученных от BioCat, Гейдельберг, Германия, T6234701-2) картина окрашивания для двух антител была идентичной, за исключением незначительной реактивности 10B3 в отношении эпителиальных клеток кожи.
[0026] На фиг. 7 показаны результаты связывания гуманизированных антител 10B3 и антител 10B3 мыши. Молекулы биспецифических антител Fabsc со специфичностью к PSMA×CD3 (10B3×OKT3), содержащие либо гуманизированные вариабельные домены с привитыми CDR (h10B3), либо вариабельные домены антитела (m10B3) мыши, инкубировали с экспрессирующими PSMA клетками 22RV1 и анализировали с помощью проточной цитометрии.
[0027] На фиг. 8 показаны результаты связывания CD3-нацеленной части разных антител к PSMA×CD3. CD3-положительные клетки Jurkat инкубировали с указанными антителами и анализировали с помощью проточной цитометрии.
[0028] На фиг. 9 показаны результаты цитолитической активности разных антител к PSMA. Экспрессирующие PSMA клетки карциномы предстательной железы 22RV1 инкубировали с PBMC и указанными биспецифическими антителами к PSMA×CD3 при соотношении PBMC:мишень 5:1. Жизнеспособность адгезивных целевых клеток оценивали с применением системы Xelligence. Показаны репрезентативные результаты одного из четырех разных экспериментов с PBMC от разных здоровых добровольцев.
[0029] На фиг. 10 показана аминокислотная последовательность вариабельных областей тяжелой и легкой цепей мыши и гуманизированного антитела 10B3. На фиг. 10A показана аминокислотная последовательность вариабельной области тяжелой цепи антитела 10B3 мыши (SEQ ID NO: 01). Последовательности CDR подчеркнуты. На фиг. 10B показана аминокислотная последовательность вариабельной области легкой цепи антитела 10B3 мыши (SEQ ID NO: 02). Последовательности CDR подчеркнуты. На фиг. 10C показана аминокислотная последовательность вариабельной области тяжелой цепи гуманизированного антитела 10B3, в котором CDR-петли (CDRH1, CDRH2 и CHDR3) тяжелой цепи антитела 10B3 мыши привиты на вариабельный домен в пределах последовательности IGHV3-11*06 тяжелой цепи зародышевой линии (SEQ ID NO: 09). Последовательности CDR подчеркнуты. Кроме того, сериновый остаток, который присутствует в положении 49 последовательности IGHV3-11*06 тяжелой цепи зародышевой линии, подвергали обратной мутации в вариабельном домене SEQ ID NO:9 к аланину, который присутствует в антителе 10B3 мыши. Этот аланиновый остаток в положении 49 выделен жирным шрифтом и курсивом на фиг. 10C). На фиг. 10D показана аминокислотная последовательность вариабельной области легкой цепи гуманизированного антитела 10B3, в котором CDR-петли (CDRL1, CDRL2, CDRL3) легкой цепи антитела 10B3 привиты на вариабельный домен в пределах последовательности IGKV3-20*02 легкой κ-цепи человека (SEQ ID NO: 10). Последовательности CDR подчеркнуты. Кроме того, фенилаланин, присутствующий в положении 72 в последовательности вариабельного домена в пределах последовательности IGKV3-20*02 легкой цепи человека подвергали обратной мутации в вариабельном домене SEQ ID NO:10 к тирозиновому остатку, который присутствует в этом положении в последовательности антитела 10B3 мыши. Этот тирозиновый остаток в положении 72 выделен жирным шрифтом и курсивом на фиг. 10D.
[0030] На фиг. 11 показана аминокислотная последовательность тяжелой цепи молекулы биспецифического антитела формата IgGsc к PSMA (гуманизированное h10B3)×CD3 (гуманизированное hUCHT1) (SEQ ID NO: 11). Тяжелая цепь содержит гуманизированную вариабельную область тяжелой цепи (HC) 10B3, CH1-домен IgG1, шарнирую область IgG1, модифицированный CH2-домен IgG1, CH3-домен IgG1 и связывающийся с CD3 гуманизированный одноцепочечный Fv-фрагмент (UCHT1).
[0031] На фиг. 12 показана аминокислотная последовательность тяжелой цепи молекулы биспецифического антитела формата Fabsc к PSMA (гуманизированное h10B3)×CD3 (OKT3 мыши) (SEQ ID NO: 12). Тяжелая цепь содержит гуманизированную вариабельную область HC 10B3, CH1-домен IgG1, шарнирую область IgG1, модифицированный CH2-домен IgG1, начало CH3-домена IgG1 и связывающийся с CD3 одноцепочечный Fv-фрагмент мыши (OKT3).
[0032] На фиг. 13 показана аминокислотная последовательность легкой каппа-цепи (гуманизированного h10B3) антитела к PSMA (SEQ ID NO: 13). Эта легкая цепь дополняет конструкции на основе тяжелых цепей SEQ ID NO: 11 и SEQ ID NO: 12 с образованием молекулы IgGsc h10B3×UCHT1 и молекулы Fabsc h10B3×OKT3 соответственно (см. фиг. 1).
[0033] На фиг. 14 показана аминокислотная последовательность вариабельных доменов антитела J519, где на фиг. 14A показана аминокислотная последовательность вариабельного домена тяжелой цепи (SEQ ID NO: 15), а на фиг. 14B показана аминокислотная последовательность вариабельного домена легкой цепи (SEQ ID NO: 16) антитела J519.
[0034] На фиг. 15 показан терапевтический эффект биспецифического антитела по изобретению in vitro. Молекулу биспецифического антитела формата IgGsc к PSMA (гуманизированное h10B3)×CD3 (гуманизированное hUCHT1) по изобретению и контрольное биспецифическое антитело (NG2×CD3) инкубировали в присутствии PBMC с опухолевыми клетками или без них.
[0035] На фиг. 16 показана противоопухолевая активность in vivo молекулы биспецифического антитела формата IgGsc к PSMA (гуманизированное h10B3)×CD3 (гуманизированное hUCHT1) по изобретению в мышиной модели.
[0036] На фиг. 17 показаны результаты по активации T-клеток и ингибированию роста опухолевых клеток нацеленных не на PSMA биспецифических антител IgGsc с UCHT1 в качестве специфичного к CD3 антитела.
Подробное раскрытие изобретения
[0037] Изобретение относится к антителу, молекуле антитела или ее антигенсвязывающему фрагменту, которые способны к связыванию с простатическим специфическим мембранным антигеном (PSMA) человека. Антитело, молекула антитела или ее антигенсвязывающий фрагмент содержат (i) вариабельный домен тяжелой цепи, содержащий CDRH1-область, изложенную в SEQ ID NO: 3 (с аминокислотной последовательностью GFTFSDFYMY), CDRH2-область, изложенную в SEQ ID NO: 4 (с аминокислотной последовательностью TISDGGGYTSYPDSVKG), и CDRH3-область, изложенную в SEQ ID NO: 5 (с аминокислотной последовательностью GLWLRDALDY), или содержащий последовательности CDRH1, CDRH2 или CDRH3, характеризующиеся по меньшей мере 75% идентичностью последовательности или по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 3, SEQ ID NO:4 или SEQ ID NO: 5. Они дополнительно содержат (iii) вариабельный домен легкой цепи, содержащий CDRL1-область, изложенную в SEQ ID NO: 6 (с аминокислотной последовательностью SASSSISSNYLH), CDRL2-область, изложенную в SEQ ID NO: 7 (с аминокислотной последовательностью RTSNLAS), и CDRL3-область, изложенную в SEQ ID NO: 8 (с аминокислотной последовательностью QQGSYIPFT), или содержащий последовательности CDRL1, CDRL2 или CDRL3, характеризующиеся 75% идентичностью последовательности или 80% идентичностью последовательности с SEQ ID NO: 6, SEQ ID NO:7 или SEQ ID NO: 8. В изобретении предусмотрена молекула антитела, содержащая CDRH1-область, изложенную в SEQ ID NO: 3, CDRH2-область, изложенную в SEQ ID NO: 4, CDRH3-область, изложенную в SEQ ID NO: 5, CDRL1-область, изложенную в SEQ ID NO: 6, VLCDL2-область, изложенную в SEQ ID NO: 7, и VLCDL3-область, изложенную в SEQ ID NO: 8. В данном контексте следует отметить, что молекула антитела по изобретению или ее антигенсвязывающий фрагмент предпочтительно не конкурируют за связывание с антителом J591 (Liu et al., Cancer Res 1997; 57: 3629-34), которое является наиболее применимым в клинике антителом к PSMA человека (см. обзор Akhtar et al., “Prostate-Specific Membrane Antigen-Based Therapeutics”, Advances in Urology, том 2012 (2012), ID статьи 973820). Кроме того, молекула антитела по настоящему изобретению или ее антигенсвязывающий фрагмент могут характеризоваться сниженной степенью индукции антигенного сдвига при связывании с PSMA по сравнению с J591. Они также могут проявлять уникальную реактивность в отношении клеток плоскоклеточной карциномы разного происхождения.
[0038] Изобретение дополнительно относится к антителу, молекуле антитела или ее антигенсвязывающему фрагменту, содержащим вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с аминокислотной последовательностью, изложенной в SEQ ID NO: 01 или 09. Также изобретение отхватывает антитело, молекулу антитела или ее антигенсвязывающий фрагмент, содержащие вариабельную область легкой цепи, где вариабельная область легкой цепи содержит аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с аминокислотной последовательностью, изложенной в SEQ ID NO: 02 или SEQ ID NO: 10. Особенно предпочтительными являются антитело, молекула антитела или ее антигенсвязывающий фрагмент, содержащие вариабельный домен тяжелой цепи и вариабельный домен легкой цепи антитела к PSMA 10B3 мыши (m10B3), изложенные в SEQ ID NO: 01 и 02 соответственно. Также предпочтительными являются антитело, молекула антитела или ее антигенсвязывающий фрагмент, содержащие вариабельный домен тяжелой цепи и вариабельный домен легкой цепи гуманизированного антитела к PSMA 10B3 (h10B3), изложенные в SEQ ID NO: 09 и 10 соответственно.
[0039] PSMA является особенно перспективным антигеном для опосредованного антителом нацеливания, и было разработано несколько антител, направленных на внеклеточный участок этого белка. Наиболее новый препарат, J591 (Liu et al., Cancer Res 1997; 57: 3629-34), в настоящее время находится на стадии оценки в рамках клинических испытаний, являясь при этом либо меченным радиоактивным изотопом, либо соединенным с токсином DM1, производным майтансина, соединением, ингибирующим тубулин (Ross JS, et al., Cancer Met Rev 2005; 24:521; Akhtar et al., 2012, выше). Антитело к PSMA по настоящему изобретению характеризуется идентичным паттерном взаимодействия с нормальной тканью человека. Антитело к PSMA по настоящему изобретению, однако, отличается от антитела J591 в отношении его взаимодействия с клетками плоскоклеточной карциномы разного происхождения. Неожиданно было обнаружено, согласно данному описанию, что при таких опухолях, а также при злокачественной опухоли предстательной железы, как собственно опухолевые клетки, так и сосудистая система в пределах и вблизи опухоли окрашиваются антителом к PMSA, таким как антитело 10B3, которое содержит последовательности CDR вариабельных доменов тяжелой и легкой цепей, представленных в SEQ ID NO:3-SEQ ID NO: 8 (ср. фиг. 6). Формы плоскоклеточной карциномы составляют большую часть злокачественных опухолей, возникающих в области уха, носа и горла, пищеводе и шейке матки, а также 20-30% опухолей легкого, и экспрессия PSMA клетками таких опухолей ранее не была описана. Таким образом, подходящая реактивность антитела по изобретению в отношении таких злокачественных опухолей обеспечивает расширенные и улучшенные варианты диагностики и лечения.
[0040] Антитело к PSMA J591 и антитело по изобретению отличаются в контексте еще одного важного аспекта: В целом, многие антитела индуцируют значительный антигенный сдвиг при связывании с целевой клеткой, что является желательным свойством, например, для конструкции иммунотоксинов, в случае которых требуется поглощение клетками для проявления биологической активности. Для этой цели применяют, например, эталонное антитело к PSMA J591 (Ross et al., 2005, выше). Однако если требуется привлечение иммунных эффекторных клеток, предпочтительной является стабильная экспрессия антигена, а не его быстрое поглощение клеткой. На фиг. 5 продемонстрировано, что связывание антитела по изобретению с трансфицированными PMSA клетками Sp2/0 сопоставимо с таковым у J591 (фиг. 5A), и что два типа антител не конкурируют перекрестно друг с другом, что свидетельствует о том, что они связываются с разными эпитопами молекулы PSMA (фиг. 5B). Наиболее важно, что антитело по изобретению индуцирует сниженную степень антигенного сдвига по сравнению с J591 (фиг. 5C). В этом контексте, однако, следует отметить, что эпитоп на PMSA, с которым связывается антитело 10B3, еще не известен. В этом отношении также следует отметить, что эпитоп, с которым антитело 10B3 связывается на поверхности клеток плоскоклеточной карциномы, не обязательно может быть таким же, как эпитоп на PMSA, в частности, поскольку об экспрессии PMSA на поверхности клеток плоскоклеточной карциномы еще не сообщалось. Таким образом, эпитоп или эпитопы, с которыми молекула антитела по изобретению связывается на поверхности клеток плоскоклеточной карциномы, могут быть связаны с эпитопом на PMSA только в отношении их аминокислотной последовательности или их конформации. Однако природа соответствующего эпитопа на PMSA или клетках плоскоклеточной карциномы не актуальна для изобретения до тех пор, пока молекула антитела по изобретению связывается с клетками, экспрессирующими PMSA, или с клетками плоскоклеточной карциномы, как описано в данном описании. В описании также отмечено, что связывание молекулы антитела по изобретению с клеткой не обязательно должно вызвать физиологический ответ. То есть достаточно, чтобы антитело по изобретению связывалось с данной клеткой (эпитоп присутствует на ее поверхности). Например, в случае конъюгирования с цитотоксическим средством, таким как токсин или радиоактивный лиганд, антитело выполняет функцию, в терапевтических целях, компонента для доставки или нацеливания, который доставляет цитотоксическое средство к клетке, в отношении которой цитотоксическое средство должно проявить свою цитотоксическую (обеспечивающую уничтожение клетки) активность. Аналогично, при применении в диагностических целях антитело по изобретению может быть конъюгировано с визуализируемым компонентом, который обеспечивает формирование выявляемого сигнала, который можно применять для выявления клетки, с которой связалось антитело.
[0041] Изобретение также относится к гуманизированной версии 10B3, которая была гуманизирована путем прививания CDR, что означает, что CDR-области антитела 10B3 мыши вставляют в каркасную область тяжелой цепи и легкой цепи антитела человека. В целом, в качестве каркаса для прививания CDR может служить любая вариабельная область легкой цепи и/или вариабельная область тяжелой цепи человека. Согласно одному иллюстративному примеру гуманизированного антитела по настоящему изобретению CDR-области легкой цепи антитела 10B3 (что означает CDR-петли SEQ ID NO: 6-SEQ ID NO: 8) можно вставлять в (вариабельный домен) последовательность IGKV3-20*02 легкой κ-цепи человека, внесенную в базу данных IMGT/LIGM под номером доступа L37729, см. также Ichiyoshi Y., Zhou M., Casali P. A human anti-insulin IgG autoantibody apparently arises through clonal selection from an insulin-specific 'germ-line' natural antibody template. Analysis by V gene segment reassortment and site-directed mutagenesis' J. Immunol. 154(1):226-238 (1995). Согласно другому иллюстративному примеру гуманизированного антитела по изобретению CDR-области тяжелой цепи антитела 10B3 (что означает CDR-петли SEQ ID NO: 3-SEQ ID NO: 5) можно включать в (вариабельные домены) последовательность IGHV3-11*06 тяжелой цепи, внесенную в базу данных IMGT/LIGM под номером доступа AF064919 (см. также Watson C.T., et al., Complete haplotype sequence of the human immunoglobulin heavy-chain variable, diversity, and joining genes and characterization of allelic and copy-number variation. Am. J. Hum. Genet. 92(4):530-546 (2013)). Согласно дополнительному иллюстративному варианту осуществления гуманизированного антитела, описанного в данном описании, CDR-петли тяжелой цепи антитела 10B3 прививают на вариабельный домен в пределах последовательности IGHV3-11*06 тяжелой цепи зародышевой линии, а CDR-петли легкой цепи антитела 10B3 прививают на вариабельный домен в пределах последовательности IGKV3-20*02 легкой κ-цепи человека. С целью сохранения свойств связывания исходного антитела 10B3 мыши, остатки каркасной области человека можно подвергать обратной мутации к аминокислотному остатку, который находится в конкретном положении в последовательности антитела 10B3 мыши. Согласно иллюстративному примеру такого гуманизированного антитела в вариабельном домене тяжелой цепи в пределах последовательности IGHV3-11*06 зародышевой линии человека серин в положении 49 подвергали обратной мутации к аланину, который находится в последовательности антитела 10B3 мыши (см. также фиг. 10C, на которой аланиновый остаток в положении 49 выделен жирным шрифтом и курсивом), тогда как в вариабельном домене в пределах последовательности IGKV3-20*02 легкой цепи фенилаланин в положении 72 в последовательности зародышевой линии человека подвергали обратной мутации к тирозиновому остатку, который находится в этом положении в последовательности антитела 10B3 мыши (см. также фиг. 10D, на которой тирозиновый остаток в положении 72 выделен жирным шрифтом и курсивом). Такое гуманизированное антитело, включенное в биспецифическое Fabsc-антитело, связывается с клетками экспрессирующей PSMA клеточной линии с такой же авидностью, что и исходное антитело мыши (ср. фиг. 7).
[0042] Термин «антитело», как правило, относится к белковой связывающей молекуле на основе иммуноглобулина. Типичные примеры такого антитела представляют собой производные или функциональные фрагменты иммуноглобулина, который сохраняет специфичность связывания. Методики получения антител и фрагментов антител являются хорошо известными в данной области. Термин «антитело» также включает иммуноглобулины (Ig) разных классов (т. е. IgA, IgG, IgM, IgD и IgE) и подклассов (таких как, IgG1, lgG2 и т. д.). Как также упоминалось выше, иллюстративные примеры производного или молекулы антитела включают Fab-фрагменты, F(ab’)2, Fv-фрагменты, одноцепочечные Fv-фрагменты (scFv), диатела или доменные антитела (Holt LJ et al., Trends Biotechnol. 21(11), 2003, 484-490). Определение термина «антитело», таким образом, также включает варианты осуществления, например, химерные, одноцепочечные и гуманизированные антитела.
[0043] «Молекула антитела» в контексте настоящего описания может нести один или несколько доменов, которые имеют последовательность, характеризующуюся по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 92%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98% или по меньшей мере приблизительно 99% идентичностью последовательности с соответствующим встречающимся в природе доменом иммуноглобулина M, иммуноглобулина G, иммуноглобулина A, иммуноглобулина D или иммуноглобулина E. В этом отношении следует отметить, что термин «приблизительно» или «примерно» в контексте настоящего описания означает отклонение в пределах 20%, например, отклонение в пределах 10% или в пределах 5% от данного значения или диапазона.
[0044] «Процент (%) идентичности последовательностей» в контексте настоящего изобретения означает процентную долю попарно идентичных остатков - согласно результатам выравнивания областей гомологии последовательности полипептида по настоящему изобретению с рассматриваемой последовательностью - по отношению к числу остатков в более длинной из этих двух последовательностей. Выравнивание с целью определения процента идентичности аминокислотных последовательностей можно осуществлять различными способами, известными в данной области, к примеру, с применением имеющегося в открытом доступе компьютерного программного обеспечения, такого как программное обеспечение BLAST, ALIGN или Megalign (DNASTAR). Специалисты в данной области смогут определить соответствующие параметры для проведения выравнивания, включая любые алгоритмы, необходимые для обеспечения максимального выравнивания по всей длине последовательностей, подлежащих сравнению. Это же справедливо и для нуклеотидных последовательностей, раскрытых в данном описании. В этом контексте идентичность последовательностей по меньшей мере 75% или по меньшей мере 80%, как описано в данном описании, представлена в отношении последовательности CDR антитела по настоящему изобретению. Упоминая сперва CDR H1, антитело по настоящему изобретению имеет последовательность CDRH1 GFTFSDFYMY (SEQ ID NO: 3) или аминокислотную последовательность, характеризующуюся 80% идентичностью последовательности с такой последовательностью. Поскольку эта последовательность CDRH1 имеет длину 10 аминокислот, 2 из этих 10 остатков могут быть замещены так, чтобы идентичность последовательности с SEQ ID NO:3 составляла 80%. Например, возможным является замещение треонинового остатка в положении 3 CDR H1 серином (выполнение консервативной замены) и замещение серинового остатка в положении 5 CDRH1 треониновым остатком (также введение консервативной замены). Таким образом, полученная в результате последовательность GFSFTDFYMY (SEQ ID NO: 14) CDRH1, которая несет такие две консервативные аминокислотные замены по сравнению с SEQ ID NO: 3, характеризуется 80% идентичностью последовательности с последовательностью SEQ ID NO: 3, тогда как последовательность CDR H1, в которой выполняют только одну из этих двух консервативных замен, характеризуется 90% идентичностью последовательности с последовательностью SEQ ID NO: 3. Аналогичным образом, в случае последовательности CDRH2, изложенной в SEQ ID NO: 04 (TISDGGGYTSYPDSVKG), содержащей 17 аминокислотных остатков, идентичность последовательности 80% допускает вплоть до 3 мутаций по сравнению с последовательностью SEQ ID NO: 04 (поскольку 20% теоретически соответствует 3,4 разным аминокислотам). Например, первый треониновый остаток SEQ ID NO: 4 может быть замещен серином. Аналогичным образом, CDRH3-область, изложенная в SEQ ID NO: 05 (GLWLRDALDY), имеет длину 10 аминокислотных остатков. Таким образом, последовательность CDRH3, которая характеризуется 80% или 90% идентичностью последовательности с SEQ ID NO: 05 (GLWLRDALDY), может предусматривать два аминокислотных замещения, например, консервативные замены, по сравнению с SEQ ID NO: 5. CDRL1-область, изложенная в SEQ ID NO: 06 (SASSSISSNYLH), содержит 12 аминокислотных остатков. Таким образом, последовательность CDRL1, которая несет одну или две аминокислотные замены по сравнению с аминокислотной последовательностью SEQ ID NO: 06, характеризуется более 80% идентичностью последовательности с последовательностью SEQ ID NO: 06. CDRL2-область, изложенная в SEQ ID NO: 07 (RTSNLAS), имеет длину 7 аминокислотных остатков. Таким образом, последовательность CDRL2, которая предусматривает одну аминокислотную замену по сравнению с последовательностью CDRL2 SEQ ID NO: 7, характеризуется 84% идентичностью последовательности с SEQ ID NO: 07. Наконец, CDRL3-область, изложенная в SEQ ID NO: 08 (QQGSYIPFT), имеет длину 9 аминокислотных остатков. Соответственно, последовательность CDRL3, которая содержит одну замененную аминокислоту по сравнению с последовательностью SEQ NO: 08, характеризуется 89% идентичностью последовательности с SEQ ID NO: 8, а последовательность CDL3, которая предусматривает две аминокислотные замены по сравнению с SEQ ID NO: 08, характеризуется 78 % идентичностью последовательности с последовательностью SEQ ID NO: 08. В настоящем описании отмечено, что из вышеприведенного объяснения и при рассмотрении описанных в данном описании последовательностей CDR-областей, специалисту в данной области будет понятно, что любые последовательности, которые характеризуются по меньшей мере 80% идентичностью последовательности с последовательностью любой из описанных в настоящем описании CDRH1, CDRH2, CDHL3, CDRL1, CDRL2 и CDRL3 (SEQ ID NO: 03-SEQ ID NO: 08), и которые способны связываться с PMSA, и также предпочтительно с клетками плоскоклеточной карциномы, как описано в настоящем описании, охвачены объемом настоящего изобретения. Хотя последовательность CDR, которая характеризуется по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85% или по меньшей мере 90% идентичностью последовательности с соответствующей последовательностью CDR из любой из SEQ ID NO: 03-SEQ ID NO: 08, предусматривает предпочтительно одну или несколько консервативных мутаций, также возможной является ситуация, когда отклонение от последовательности любой из шести «исходных» CDR-областей (SEQ ID NO: 3-SEQ ID NO: 8) антитела по настоящему изобретению, и, таким образом, идентичность последовательности 75% или больше, обусловлено наличием неконсервативных мутаций в CDR-областях, при условии, что антитело сохраняет способность связываться с PMSA, и также предпочтительно с клетками плоскоклеточной карциномы.
[0045] «Иммуноглобулин», при применении в настоящем описании, обычно представляет собой тетрамерный гликозилированный белок, состоящий из двух легких (L) цепей массой примерно 25 кДа каждая и двух тяжелых (H) цепей массой примерно 50 кДа каждая. В иммуноглобулинах могут встречаться два типа легких цепей, называемых лямбда и каппа. В зависимости от аминокислотной последовательности константного домена тяжелых цепей иммуноглобулины относят к пяти основным классам: A, D, E, G и M, и несколько из них могут быть дополнительно разделены на подклассы (изотипы), например, IgG1, lgG2, IgG3, IgG4, IgA1 и IgA2. IgM-иммуноглобулин состоит из 5 основных гетеротетрамерных единиц вместе с дополнительным полипептидом, называемым J-цепью, и содержит 10 антигенсвязывающих сайтов, тогда как IgA-иммуноглобулины содержат 2-5 из основных 4-цепочечных единиц, которые могут полимеризоваться с образованием поливалентных комплексов в сочетании с J-цепью. В случае IgG масса 4-цепочечной единицы, как правило, составляет приблизительно 150000 Дальтон.
[0046] В иммуноглобулинах класса IgG содержатся несколько иммуноглобулиновых доменов тяжелой цепи. Под «иммуноглобулиновым (Ig) доменом» в настоящем описании подразумевают область иммуноглобулина с отличающейся третичной структурой. В контексте IgG-антител каждый из изотипов IgG содержит три CH-области: «CH1» относится к положениям 118-220, «CH2» относится к положениям 237-340 и «CH3» относится к положениям 341-447 согласно EU-индексу по Kabat et al. Под «шарниром», или «шарнирной областью», или «шарнирной областью антитела», или «шарнирной областью иммуноглобулина» или «H» в настоящем описании подразумевают гибкий полипептид, содержащий аминокислоты между первым и вторым константными доменами антитела. В структурном отношении CH1-домен IgG заканчивается положением 220 согласно EU, и CH2-домен IgG начинается с остатка 237 согласно EU. Таким образом, для IgG шарнир в настоящем описании определен как охватывающий положения от 221 (D221 в IgG1) до 236 (G236 в IgG1), где нумерация указана в соответствии с EU-индексом по Kabat et al. Константная тяжелая цепь, как определено в настоящем описании, охватывает область от N-конца CH1-домена до C-конца CH3-домен, включая, таким образом, положения 118-447, где нумерация указана в соответствии с EU-индексом.
[0047] Термин «вариабельный» относится к участкам иммуноглобулиновых доменов, которые характеризуются вариабельностью в отношении их последовательности, и которые вовлечены в определение специфичности и аффинности связывания конкретного антитела (т. е. «вариабельный(-ые) домен(-ы)»). Вариабельность распределена неодинаково по вариабельным доменам антител; она сконцентрирована в субдоменах каждой из вариабельных областей тяжелой и легкой цепей. Такие субдомены называют «гипервариабельными областями», «HVR», или «HV», или «определяющими комплементарность областями» (CDR). Более консервативные (т.е. не являющиеся гипервариабельными) участки вариабельных доменов называют «каркасными» областями (FR). Каждый из вариабельных доменов встречающихся в природе тяжелых и легких цепей включает четыре FR-области, преимущественно принимающие конфигурацию β-листа, соединенные тремя гипервариабельными областями, которые образуют петли, соединяющие, и, в некоторых случаях, образующие часть структуры β-листа. Гипервариабельные области в каждой цепи удерживаются вместе в непосредственной близости посредством FR и, вместе с гипервариабельными областями из другой цепи, способствуют образованию антигенсвязывающего сайта (см. Kabat et al., см. ниже). Как правило, встречающиеся в природе иммуноглобулины включают шесть CDR (см. ниже); три в VH (H1, H2, H3) и три в VL (L1, L2, L3). Во встречающихся в природе иммуноглобулинах H3 и L3 проявляют наиболее широкое разнообразие из шести CDR, и в частности H3, как полагают, играет особую роль в придании иммуноглобулинам высокой специфичности. Константные домены непосредственно не вовлечены в связывание антигена, но они проявляют различные эффекторные функции, такие как, например, антителозависимая клеточноопосредованная цитотоксичность и активация системы комплемента.
[0048] Термины «VH» (также называемый VH) и «VL» (также называемый VL) применяют в настоящем описании для обозначения соответственно вариабельного домена тяжелой цепи и вариабельного домена легкой цепи иммуноглобулина. Вариабельная область легкой или тяжелой цепи иммуноглобулина состоит из «каркасной» области, прерываемой тремя гипервариабельными областями. Таким образом, термин «гипервариабельная область» относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Гипервариабельная область включает аминокислотные остатки «определяющей комплементарность области» или «CDR». В вариабельном участке иммуноглобулина находятся три CDR (или CDR-области) тяжелых цепей и три CDR легких цепей. Таким образом, «CDR» в контексте настоящего описания относятся ко всем трем CDR тяжелой цепи (CDRH1, CDRH2 и CDRH3), или ко всем трем CDR легкой цепи (CDRL1, CDRL2 и CDRL3), или как ко всем CDR тяжелой цепи, так и ко всем CDR легкой цепи, если уместно. Три CDR определяют свойства связывания вариабельной области легкой цепи, и три определяют свойства связывания вариабельной области тяжелой цепи. CDR определяют антигенную специфичность молекулы иммуноглобулина, и они разделены аминокислотными последовательностями, которые включают скелетные или каркасные области. Конкретные определительные границы и значения длины CDR основываются на различных типах классификации и системах нумерации. Структура и сворачивание белка у антитела могут предусматривать, что другие остатки считаются частью антигенсвязывающей области, и это будет понятно специалисту в данной области. CDR обеспечивают основную часть участвующих в контакте остатков для связывания иммуноглобулина с антигеном или эпитопом.
[0049] CDR3 обычно является наиболее значительным источником молекулярного разнообразия в пределах сайта связывания антитела. H3, например, может состоять только из двух аминокислотных остатков или более чем 26 аминокислот. Субъединичные структуры и пространственные конфигурации разных классов иммуноглобулинов хорошо известны в данной области. Для обзора структуры антител см. Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, eds. Harlow et al., 1988. Специалисту в данной области будет понятно, что каждая субъединичная структура, например, структура CH, VH, CL, VL, CDR, FR, включает активные фрагменты, например, часть субъединицы VH, VL или CDR связывается с антигеном, т. е. антигенсвязывающий фрагмент, или, например, участок субъединицы CH, который связывается, например, с Fc-рецептором и/или комплементом, и/или активирует их. CDR обычно относятся к CDR по Kabat, как описано в Sequences of Proteins of immunological Interest, Министерство здравоохранения и социальных служб США (1991), ред. Kabat et al. Другой стандарт определения характеристик антигенсвязывающего сайта относится к гипервариабельным петлям, как описано Chothia. См., например, Chothia, et al. (1992; J. MoI. Biol. 227:799-817; и Tomlinson et al. (1995) EMBO J. 14:4628-4638. Еще одним стандартом является определение на основе модели антитела, применяемое в программном обеспечении для моделирования антител AbM Oxford Molecular. В общих чертах см., например, Protein Sequence and Structure Analysis of Antibody Variable Domains, в: Antibody Engineering Lab Manual (ред.: Duebel, S. и Kontermann, R., Springer-Verlag, Гейдельберг). Варианты осуществления, описанные в отношении CDR по Kabat, в качестве альтернативы могут быть реализованы с применением аналогичным образом описанных принципов в отношении гипервариабельных петель по Chothia или петель, определенных на основе модели антитела.
[0050] Соответствующие тяжелая мю-цепь, тяжелая гамма-цепь, тяжелая альфа-цепь, тяжелая дельта-цепь, тяжелая эпсилон-цепь, легкая лямбда-цепь или легкая каппа-цепь иммуноглобулина может принадлежать любому виду, например, виду млекопитающего, включая виды грызунов, амфибии, например, подкласса Беспанцирные, который включает, например, лягушек, жаб, саламандр или тритонов, или виду беспозвоночного. Примеры млекопитающих включают без ограничения крысу, мышь, кролика, морскую свинку, белку, хомяка, ежа, утконоса, американскую пищуху, броненосца, собаку, лемура, козу, свинью, корову, опоссума, лошадь, летучую мышь, лесного сурка, орангутана, макака-резуса, шерстистую обезьяну, макака, шимпанзе, тамарина (saguinus oedipus), мармозетки или человека.
[0051] Как упоминалось в настоящем описании, иммуноглобулин обычно представляет собой гликопротеин, который содержит по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, соединенные дисульфидными связями, или его антигенсвязывающий участок. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (в настоящем описании сокращено как VH) и константную область тяжелой цепи. Согласно некоторым вариантам осуществления константная область тяжелой цепи содержит три домена, CH1, CH2 и CH3. Каждая легкая цепь содержит вариабельную область легкой цепи (в настоящем описании сокращено как VL) и константную область легкой цепи. Константная область легкой цепи содержит один домен, CL. VH- и VL-области дополнительно могут быть разделены на гипервариабельные области, называемые определяющими комплементарность областями (CDR), перемежающиеся с областями, которые являются более консервативными, называемыми каркасными областями (FR). CDR содержит большинство остатков, ответственных за специфические взаимодействия антитела с антигеном. Каждая VH и VL содержит три CDR и четыре FR, расположенные, от амино-конца к карбокси-концу, в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелых и легких цепей содержат связывающий домен, который взаимодействует с эпитопом антигена.
[0052] Остатки «каркасной области» или «FR» представляют собой остатки вариабельного домена, отличные от таковых гипервариабельной области. Последовательности каркасных областей разных легких или тяжелых цепей являются относительно консервативными среди видов. Таким образом, «каркасная область человека» представляет собой каркасную область, которая является практически идентичной (на приблизительно 85% или больше, обычно на 90-95% или больше) каркасной области встречающегося в природе иммуноглобулина человека. Каркасная область антитела, то есть каркасные области составных легких и тяжел цепей в совокупности, служит для определения положений и выравнивания CDR. CDR главным образом отвечают за связывание с эпитопом антигена.
[0053] Термины «Fab», «Fab-область», «Fab-участок» или «Fab-фрагмент» понимают как определяющие полипептид, который содержит иммуноглобулиновые домены VH, CH1, VL и CL. Fab может относиться к такой области в отдельности, или такой области в контексте молекулы антитела, а также полноразмерного иммуноглобулина или фрагмента иммуноглобулина. Обычно Fab-область содержит полную легкую цепь антитела. Fab-область может быть определена как «плечо» молекулы иммуноглобулина. Она содержит связывающий эпитоп участок такого Ig. Fab-область встречающегося в природе иммуноглобулина может быть получена в виде образующегося в результате протеолиза фрагмента посредством расщепления папаином. «F(ab’)2-участок» представляет собой образующийся в результате протеолиза фрагмент иммуноглобулина, расщепленного пепсином. «Fab’-участок» является продуктом, образующимся в результате восстановления дисульфидных связей F(ab’)2-части. В контексте настоящего описания термины «Fab», «Fab-область», «Fab-участок» или «Fab-фрагмент» могут дополнительно включать шарнирную область, которая определяет C-конец плеча антитела. Эта шарнирная область соответствует шарнирной области, обнаруживаемой со стороны C-конца CH1-домена в пределах полноразмерного иммуноглобулина, в которой плечи молекулы антитела определяют форму Y. Термин «шарнирная область» применяют в данной области ввиду того, что иммуноглобулин обладает некоторой гибкостью в этой области. Под «тяжелой цепью Fab» в контексте настоящего описания понимают тот участок или полипептид Fab-фрагмента, которые содержат VH и CH1, тогда как под «легкой цепью Fab» в контексте настоящего описания понимают тот участок или полипептид Fab-фрагмента, которые содержат VL и CL.
[0054] Термин «Fc-область» или «Fc-фрагмент» применяют в настоящем описании для определения C-концевой области тяжелой цепи иммуноглобулина, включая Fc-области с нативными последовательностями и вариантные Fc-областей. Fc-часть опосредует эффекторную функцию антител, например, активацию системы комплемента, и несущих Fc-рецепторы иммунных эффекторных клеток, таких как NK-клетки. В молекулах IgG человека Fc-область образуется при расщеплении папаином в сторону N-конца от Cys226. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьировать, Fc-область тяжелой цепи IgG человека обычно определяется отрезком от аминокислотного остатка в положении Cys226, или от Pro230, до ее карбокси-конца. C-концевой лизин (остаток 447 в соответствии с системой нумерации EU) Fc-области может быть удален, например, в ходе получения или очистки молекулы антитела, или при конструировании с применением рекомбинантной технологии нуклеиновой кислоты, кодирующей тяжелую цепь молекулы антитела. Fc-области с нативными последовательностями относятся к IgG1, IgG2 (IgG2A, IgG2B), IgG3 и IgG4 млекопитающих, например, человека или мыши. Fc-область содержит два или три константных домена, в зависимости от класса антитела. Согласно вариантам осуществления, в которых иммуноглобулином является IgG, Fc-область содержит CH2- и CH3-домен.
[0055] Термин «одноцепочечный вариабельный фрагмент» (scFv) применяют в настоящем описании для определения фрагмента антитела, в котором вариабельные области тяжелых (VH) и легких цепей (VL) иммуноглобулина слиты вместе, при этом они соединены коротким линкерным пептидом длиной от десяти до приблизительно 25 аминокислот. Линкер обычно обогащен глицином для обеспечения гибкости, а также серином или треонином для обеспечения растворимости, и может либо соединять N-конец VH с C-концом VL, либо соединять N-конец VL с C-концом VH. В scFv-фрагменте сохраняется специфический антигенсвязывающий сайт, но отсутствуют константные домены иммуноглобулинов.
[0056] Термин «эпитоп», также известный как «антигенная детерминанта», относится к участку антигена, с которым специфически связывается антитело или T-клеточный рецептор, с образованием тем самым комплекса. Таким образом, термин «эпитоп» включает любую детерминанту молекулы или белка, способную специфически связываться с иммуноглобулином или T-клеточным рецептором. Сайт(-ы) связывания (паратоп) описанной в настоящем описании молекулы антитела могут специфически связываться/взаимодействовать с конформационными или непрерывными эпитопами, и являются уникальными для целевой структуры. Антигенные детерминанты обычно состоят из химически активных групп на поверхности молекул, таких как аминокислоты и боковые цепи из сахаров, и обычно обладают специфическими характеристиками трехмерной структуры, а также специфическими характеристиками заряда. Согласно некоторым вариантам осуществления антигенные детерминанты включают химически активные группы на поверхности молекул, такие как аминокислоты, боковые цепи из сахаров, фосфорильные группы или сульфонильные группы, и, согласно определенным вариантам осуществления, могут обладать специфическими характеристиками трехмерной структуры и/или специфическими характеристиками заряда. Что касается полипептидных антигенов, конформационный или прерывающийся эпитоп характеризуется наличием двух или более отдельных аминокислотных остатков, разделенных в рамках первичной последовательности, но собирающихся в единую структуру на поверхности молекулы, когда полипептид сворачивается в нативный белок/антиген (SeIa, M., Science (1969) 166, 1365-1374; Laver, W.G., et al. Cell (1990) 61, 553-556). Два или более отдельных аминокислотных остатков, участвующих в образовании эпитопа, могут находиться на отдельных участках одной или нескольких полипептидных цепей. Такие остатки объединяются на поверхности молекулы, когда полипептидная(-ые) цепь(-и) сворачивается(-ются) в пространственную структуру с образованием эпитопа. В отличие от этого, непрерывный или линейный эпитоп состоит из двух или более отдельных аминокислотных остатков, которые находятся в одном линейном участке полипептидной цепи.
[0057] В этом контексте термин «специфический» или «специфическое связывание», также применяемый как «направленный на», в соответствии с настоящим изобретением означает, что антитело или фрагмент иммунного рецептора способны специфически взаимодействовать и/или связываться со специфическим антигеном или лигандом или набором специфических антигенов или лигандов, но практически не связываются с другими антигенами или лигандами. Примером такого связывания может быть специфичность по «принципу ключа и замка». Говорят, что антитела «связываются с одним и тем же эпитопом», если антитела перекрестно конкурируют так, что только одно антитело может связываться с эпитопом в данный момент времени, т. е. одно антитело препятствует связыванию или модулирующему эффекту другого.
[0058] Обычно связывание считается специфическим, если аффинность связывания превышает 10-6 M или 10-7 M. В частности, связывание считается специфическим, если аффинность связывания составляет от приблизительно 10-8 до 10-11 M (KD) или от приблизительно 10-9 до 10-11 M или даже больше. При необходимости степень неспецифического связывания сайта связывания может быть снижена без существенного влияния на специфическое связывание путем изменения условий связывания.
[0059] Термин «выделенная молекула антитела» в контексте настоящего описания относится к молекуле антитела, которая была идентифицирована и отделена от и/или извлечена из компонента ее естественного окружения. Контаминирующие компоненты ее естественного окружения представляют собой материал, который будет негативным образом влиять на диагностические или терапевтические применения антитела, и могут включать ферменты, гормоны и другие белковые или небелковые растворимые вещества. Согласно некоторым вариантам осуществления молекулу антитела очищают с достижением значения более чем 95% по весу антитела, как определено с помощью способа Лоури, например, более чем 99% по весу. Согласно некоторым вариантам осуществления антитело очищают до однородности состава, что определяют с помощью SDS-PAGE в восстанавливающих или невосстанавливающих условиях с применением Кумасси бриллиантового синего или, предпочтительно, серебряного красителя. Выделенная молекула антитела согласно некоторым вариантам осуществления может присутствовать в рекомбинантных клетках с одним или несколькими компонентами, отсутствующими в естественном окружении антитела. Обычно выделенное антитело получают в ходе по меньшей мере одной стадии очистки.
[0060] (Рекомбинантную) молекулу антитела по настоящему изобретению, которая связывается с PMSA и/или клетками плоскоклеточного рака, как описано в настоящем описании, можно применять в любом подходящем формате рекомбинантного антитела, например, в виде Fv-фрагмента, scFv, одновалентного антитела без шарнирной области, миниантитела, Fab-фрагмента, Fab’-фрагмента, F(ab’)2-фрагмента. Рекомбинантная молекула антитела по настоящему изобретению также может содержать константные домены (области), например, константную область IgG человека, CH1-домен (как у Fab-фрагментов) и/или всю Fc-область. В качестве альтернативы, молекула антитела по настоящему изобретению также может представлять собой полноразмерное (полное) антитело.
[0061] Существует целый ряд возможных механизмов, посредством которых антитела опосредуют эффекты в отношении клеток, включая антипролиферативное действие посредством блокирования необходимых сигнальных путей, вовлеченных в рост, внутриклеточную передачу сигнала, приводящую к апоптозу, усиленное подавление и/или обновление рецепторов, комплементзависимую цитотоксичность (CDC), антителозависимую клеточноопосредованную цитотоксичность (ADCC), антителозависимый клеточноопосредованный фагоцитоз (ADCP) и стимуляцию адаптивного иммунного ответа (Cragg et al, 1999, Curr Opin Immunol 11 541-547, Glennie et al., 2000, Immunol Today 21 403-410). Эффективность антитела может быть обусловлена сочетанием этих механизмов, и их относительная значимость в клинической терапии при онкологии, по-видимому, зависит от типа злокачественной опухоли.
[0062] Значимость FcγR-опосредованных эффекторных функций для активности некоторых антител была продемонстрирована на мышах (Clynes et al., 1998, Proc Natl Acad Sci USA 95:652-656, Clynes et al, 2000, Nat Med 6:443-446) и из наблюдаемой корреляции между клинической эффективностью у людей и их аллотипом высоко- (V158) или низкоаффинных (F158) полиморфных форм FcγRIIIa (Cartron et al., 2002, Blood 99 754-758, Weng & Levy, 2003, Journal of Clinical Oncology, 21 3940-3947). Вместе эти данные свидетельствуют о том, что антитело, оптимизированное для связывания с определенными FcγR, может лучше опосредовать эффекторные функции, и, тем самым, обеспечивать более эффективное разрушение целевых клеток у пациентов. Таким образом, перспективные способы усиления противоопухолевой активности антител действуют посредством усиления их способности опосредовать цитотоксические эффекторные функции, такие как ADCC, ADCP и CDC. Кроме того, антитела могут опосредовать противоопухолевый механизм за счет передачи сигнала, вовлеченной в ингибирование роста или апоптоз, которая может иметь место при связывании антитела с его мишенью на поверхности опухолевых клеток. Такая передача сигнала может быть усилена, если антитела представлены опухолевым клеткам в связанном с иммунными клетками через FcγR состоянии. Поэтому повышенная аффинность антител к FcγR может обеспечить усиленные антипролиферативные эффекты.
[0063] Определенный успех был достигнут при модификации антител, предусматривающей избирательно усиленную способность к связыванию с FcγR для обеспечения усиленной эффекторной функции. Конструирование антител для оптимизации эффекторной функции было выполнено с применением аминокислотных модификаций (см., например, публикацию заявки на патент США US 2004-0132101 или публикацию заявки на патент США 2006-0024298).
[0064] Таким образом, в настоящем изобретении также предусмотрено, что молекула антитела по настоящему изобретению или ее антигенсвязывающий фрагмент модифицированы так, что они характеризуются повышенной аффинностью к рецептору FcγRIIIa или усиленной эффекторной функцией в виде ADCC, по сравнению с исходным антителом. Одним из способов обеспечения усиленной ADCC является введение аминокислотных замен 239D и 332E в CH2-домен Fc-части молекулы антитела, например, в мышином или гуманизированном антителе 10B3. Обеспечивающая уничтожение клетки активность таких антител после этого может быть значительно повышена, или даже может быть выявлена и получена впервые. Согласно одному варианту осуществления аминокислотные замены представляют собой S239D и I332E.
[0065] Молекула антитела по настоящему изобретению способна к связыванию с PSMA человека. Термины «простатический специфический мембранный антиген» или «PSMA» в настоящем описании применяют взаимозаменяемо, и они включают варианты, изоформы и видовые гомологи PSMA человека. PSMA также называют глутаматкарбоксипептидазой II, NAALAD-азой I = N-ацетил-L-аспартил-L-глутаматпептидаза I, фолатгидролазой I (FOLH1). PSMA человека в UniProt имеет номер доступа Q04609 (версия 175 от 9 декабря 2015 года). Соответственно, антитела по настоящему изобретению могут, в определенных случаях, перекрестно реагировать с PSMA вида, отличного от человека, или с другими белками, которые в структурном отношении подобны PSMA человека (например, гомологи PSMA человека). Как упоминалось выше, предпочтительным вариантом осуществления настоящего изобретения является антитело 10B3 или его гуманизированная версия. Однако другие молекулы антител, которые связываются с тем же эпитопом, что и 10B3, также входят в объем настоящего изобретения.
[0066] Для определения эпитопа можно применять стандартные известные в данной области способы картирования эпитопов. Например, фрагменты (пептиды) PMSA (например, синтетические пептиды), которые связывают антитело, можно применять для определения того, связывает ли антитело-кандидат или его антигенсвязывающий фрагмент тот же эпитоп. Для линейных эпитопов синтезируют перекрывающиеся пептиды определенной длины (например, 8 или более аминокислот). Пептиды могут быть смещены на 1 аминокислоту так, что получают набор пептидов, перекрывающих каждый фрагмент из 8 аминокислот белковой последовательности PSMA. Меньшее количество пептидов может быть получено с применением больших смещений, например, в 2 или 3 аминокислоты. Кроме того, можно синтезировать более длинные пептиды (например, 9-, 10- или 11-меры). Связывание пептидов с антителами или антигенсвязывающими фрагментами можно определять с применением стандартных методик, включая метод поверхностного плазмонного резонанса (BIACORE) и ELISA-анализы. Для изучения конформационных эпитопов можно применять более длинные фрагменты PSMA. Другие способы, в которых для определения конформационных эпитопов используется масс-спектрометрия, были описаны и их также можно применять (см., например, Baerga-Ortiz et al., Protein Science 11:1300-1308, 2002, и цитируемые в ней ссылки). Еще одни способы определения эпитопов представлены в стандартных лабораторных справочных изданиях, например, глава 6.8 (“Phage Display Selection and Analysis of B-cell Epitopes”) и глава 9.8 (“Identification of Antigenic Determinants Using Synthetic Peptide Combinatorial Libraries”) Current Protocols in Immunology, Coligan et al., eds., John Wiley & Sons. Эпитопы можно подтверждать путем введения точечных мутаций или делеций в известный эпитоп, а затем тестировать в отношении связывания с одним или несколькими антителами или антигенсвязывающими фрагментами с целью определения того, какие мутации опосредуют снижение связывающей способности антител или антигенсвязывающих фрагментов.
[0067] Также в объем настоящего изобретения входят молекулы антител, которые конкурируют с молекулой антитела по настоящему изобретению, такой как 10B3, за связывание с PSMA, например, для конкурентного ингибирования связывания 10B3 с PSMA. Для определения конкурентного ингибирования можно использовать ряд анализов, известных специалисту в данной области. Например, для определения того, ингибирует ли конкурентным образом антитело или его антигенсвязывающий фрагмент связывание с PSMA другого антитела или его антигенсвязывающего фрагмента, можно применять анализы конкурентного связывания. Такие случаи включают способы с применением клеток, в которых используют цитометрию или твердофазный анализ связывания. Также можно применять другие анализы, с помощью которых оценивают способность антител или их антигенсвязывающих фрагментов перекрестно конкурировать за молекулы PSMA, которые не экспрессируются на поверхности клеток, в твердой фазе или в жидкой фазе. Анализ, с помощью которого можно тестировать перекрестную конкуренцию, например, приведен в примере 10.
[0068] Как упоминалось в описании, изобретение охватывает антитела, которые индуцируют сниженную степень антигенного сдвига при связывании с PSMA по сравнению с J591. Такую сниженную степень антигенного сдвига можно оценивать, например, с применением способа, в общем описанного в примере 10. Согласно предпочтительному варианту осуществления такую сниженную степень антигенного сдвига выявляют в трансфицированных PMSA клетках Sp2/0.
[0069] Молекула антитела в соответствии с настоящим изобретением может содержать две цепи, более короткую цепь, которая, согласно некоторым вариантам осуществления, может представлять собой легкую цепь, и основную цепь, которая, согласно некоторым вариантам осуществления, также может называться тяжелой цепью. Молекула антитела обычно представляет собой димер этих двух цепей.
[0070] Молекула антитела по настоящему изобретению предпочтительно может представлять собой молекулу биспецифического антитела. Молекула биспецифического антитела может содержать (i) вариабельную область, содержащую вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, как определено в любом из предыдущих пунктов, где указанная вариабельная область содержит первый сайт связывания, способный к связыванию с простатическим специфическим мембранным антигеном (PSMA) человека, и (ii) вариабельную область тяжелой цепи и вариабельную область легкой цепи молекулы антитела, содержащие второй сайт связывания. Следует понимать, что сайт связывания для PSMA предпочтительно является сайтом связывания PSMA-связывающего антитела по изобретению, описанному в описании.
[0071] «Биспецифическая» или «бифункциональная» молекула антитела представляет собой молекулу антитела, которая содержит два разных сайта связывания эпитопа/антигена, и, соответственно, характеризуется специфичностями связывания в отношении двух разных целевых эпитопов. Такие два эпитопа могут представлять собой эпитопы одного и того же антигена или разных антигенов. В отличие от этого, «бивалентное антитело» может содержать сайты связывания идентичной антигенной специфичности.
[0072] «Биспецифическое антитело» может представлять собой молекулу антитела, которая связывает один антиген или эпитоп с помощью одного из двух или более связывающих плеч, образуемых первой парой тяжелой и легкой цепей или основной и более короткой/меньшей цепями, и связывает другой антиген или эпитоп на втором плече, образуемом второй парой тяжелой и легкой цепей или основной и меньшей цепями. Такой вариант осуществления биспецифического антитела содержит два отдельных антигенсвязывающих плеча, причем оба имеют определенную специфичность и последовательности CDR. Обычно биспецифическое антитело является моновалентным в отношении каждого антигена, с которым оно связывается, то есть оно связывается с соответствующим антигеном или эпитопом с помощью только одного плеча. Однако биспецифические антитела также могут подвергаться димеризации или мультимеризации. Например, в димерном формате IgGsc, описанном в данном описании, антитело может содержать два сайта связывания для каждого антигена. Биспецифическое антитело может представлять собой гибридную молекулу антитела, которая может содержать первую связывающую область, образуемую первой вариабельной областью легкой цепи и первой вариабельной областью тяжелой цепи, и вторую связывающую область, образуемую второй вариабельной областью легкой цепи и второй вариабельной областью тяжелой цепи. В настоящем изобретении предусмотрено, что одна из таких связывающих областей может быть образована парой тяжелой/легкой цепи. В контексте настоящего изобретения молекула биспецифического антитела может содержать первый сайт связывания, образуемый вариабельными областями основной цепи и меньшей цепи, и второй, отличный сайт связывания, образуемый вариабельной областью scFv-фрагмента, который включен в основную цепь молекулы антитела.
[0073] Способы получения молекулы биспецифического антитела известны в данной области, например, химическая конъюгация двух разных моноклональных антител, или, например, также химическая конъюгация двух фрагментов антител, например, двух Fab-фрагментов. В качестве альтернативы, молекулы биспецифических антител получают с помощью квадромной технологии, то есть слияния гибридом, продуцирующих исходные антитела. Ввиду независимой рекомбинации H- и L-цепей, потенциально образуется набор из десяти разных структур антител, из которых только одна обладает требуемой специфичностью связывания.
[0074] Молекула биспецифического антитела по настоящему изобретению может функционировать в качестве моноклонального антитела (MAb) в отношении каждой мишени. Согласно некоторым вариантам осуществления антитело является химерным, гуманизированным или полностью человеческим. Молекула биспецифического антитела, например, может представлять собой биспецифический тандемный одноцепочечный Fv, биспецифический Fab2 или биспецифическое диатело.
[0075] В зависимости от доменов, включенных в молекулу антитела по настоящему изобретению, молекула биспецифического антитела по настоящему изобретению может содержать Fab-фрагмент, который, как правило, может включать шарнирную область, CH2-домен и одноцепочечный Fv-фрагмент. Такие молекулы биспецифических антител называют молекулами антител «Fabsc», и впервые они были описаны в международной заявке на патент WO 2013/092001. Более конкретно, в контексте описания молекула антитела формата «Fabsc» обычно относится к молекуле биспецифического антитела по настоящему изобретению с Fab-фрагментом, который, как правило, содержит шарнирную область, которая находится на C-конце Fab-фрагмента, соединенном с N-концом CH2-домена, C-конец которого, в свою очередь, соединен с N-концом scFv-фрагмента. Такое «Fabsc» не содержит или в основном не содержит CH3-домен. В этом контексте «не содержащий» или «в основном не содержащий» означает, что молекула антитела не содержит полноразмерный CH3-домен. Это предпочтительно означает, что молекула антитела содержит 10 или менее, предпочтительно 5 или менее, предпочтительно 3 или даже менее аминокислот CH3-домена. Иллюстративный пример молекулы антитела формата Fabsc показан на фиг. 1A, другой иллюстративный пример молекулы антитела формата Fabsc показан на фиг. 12. Согласно иллюстративным вариантам осуществления (в этом отношении ср. также фиг. 1A) молекула антитела Fabsc по настоящему изобретению может содержать CH2-домен, у которого отсутствует способность к димеризации посредством дисульфидных связей, образуемых цистеиновым остатком в положении 226 в последовательности шарнирной области и/или цистеиновым остатком в положении 229 в последовательности одного из шарнирных доменов, в соответствии с нумерацией по Kabat [EU-индекс]. Таким образом, согласно таким вариантам осуществления цистеиновые остатки в положении 226 в последовательности и/или в положении 229 в последовательности удаляют или замещают, например, сериновым остатком. Кроме того, или в качестве альтернативы, молекула антитела «Fabsc» по настоящему изобретению также может иметь CH2-домен «с ослабленной функцией связывания Fc» (который включает шарнирную область). Такое «ослабление функции связывания Fc» обеспечивается путем удаления и/или замены (мутирования) по меньшей мере одного из выбранных аминокислотных остатков в CH2-домене, которые способны опосредовать связывание с Fc-рецептором. Согласно иллюстративным вариантам осуществления по меньшей мере один аминокислотный остаток шарнирной области или CH2-домена, который способен опосредовать связывание с Fc-рецепторами, и который отсутствует или подвержен мутации, выбран из группы, состоящей из положений 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 265, 297, 327 и 330 в последовательности (нумерация положений в последовательности указана в соответствии с EU-индексом). Согласно иллюстративному примеру такая молекула антитела с ослабленной функцией связывания Fc может предусматривать меньшей мере одну мутацию, выбранную из группы, состоящей из делеции аминокислоты 228, делеции аминокислоты 229, делеции аминокислоты 230, делеции аминокислоты 231, делеции аминокислоты 232, делеции аминокислоты 233, замены Glu233→Pro, замены Leu234→Val, делеции аминокислоты 234, замены Leu235→Ala, делеции аминокислоты 235, делеции аминокислоты 236, делеции аминокислоты 237, делеции аминокислоты 238, замены Asp265→Gly, замены Asn297→Gln, замены Ala327→Gln и замены Ala330→Ser (нумерация положений в последовательности указана в соответствии с EU-индексом, также в этом отношении см., например, фиг. 1O и фиг. 1P международной заявки на патент WO 2013/092001). В случае биспецифических антител, которые активируют T-клетки, например, в отношении опухолевых клеток, ослабление функции связывания Fc может требоваться для предотвращения связывания антител с несущими Fc-рецептор клетками, которое может приводить к нежелательной нецелевой активации T-клеток.
[0076] Согласно публикации Coloma and Morrison (Nat Biotechnol 15:159-63, 1997), молекула биспецифического антитела по настоящему изобретению также может содержать CH3-домен, как правило, расположенный со стороны C-конца от CH2-домена. Такую молекулу в данном описании также называют молекулой антитела формата «IgGsc», и она означает молекулу биспецифического антитела по настоящему изобретению с Fab-фрагментом, который, как правило, содержит шарнирную область, которая находится на C-конце Fab-фрагмента, обычно соединенном с N-концом CH2-домена, C-конец которого, в свою очередь, обычно соединен с N-концом CH3-домена, C-конец которого, в свою очередь, обычно соединен с N-концом scFv-фрагмента. Иллюстративный пример молекулы антитела формата IgGsc показан на фиг. 1B, последовательности тяжелой и легкой цепей такой молекулы представляют собой молекулы, показанные на фиг. 11 и 13 соответственно.
[0077] Для обоих форматов антитела Fabsc и IgGsc общим является то, что N-концевая нацеливающая часть состоит из «физиологических» Fab- или Fab2-областей соответственно, что, тем самым, позволяет избежать применения одноцепочечных фрагментов в этой части молекулы. В этих форматах, подлежащих применению для ограниченной целевыми клетками активации T-клеток, можно использовать ослабление связывания Fc-рецептора (FcR) (по желанию или необходимости) для предотвращения опосредованной FcR активации. Это может обеспечиваться, например, путем введения определенных и хорошо известных мутаций в CH2-домен молекулы, как описано выше, в также в международной заявке на патент WO 2013/092001 и в Armour et al., Eur J Immunol 1999; 29:2613. Соответственно, также CH2-домен (включая шарнирную область) может содержать молекула антитела IgGsc по настоящему изобретению, в которой по меньшей мере один аминокислотный остаток шарнирной области или CH2-домена, который способен опосредовать связывание с Fc-рецепторами, отсутствует или подвержен мутации. Как объяснялось выше, этот остаток в CH2 и шарнирной области, соответственно, может быть выбран из группы, состоящей из положений 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 265, 297, 327 и 330 в последовательности (нумерация положений в последовательности указана в соответствии с EU-индексом). Однако благодаря присутствию в молекуле IgGsc CH3-домена две отдельные молекулы будут (спонтанно) гомодимеризоваться посредством CH3-домена с образованием тетравалентной молекулы (в этом отношении см. опять же фиг. 1B). Таким образом, удаление или мутация цистеиновых остатков в положении 226 в последовательности и/или в положении 229 в последовательности шарнирной области не является необходимым. Таким образом, такая молекула тетрамерного антитела IgGsc по настоящему изобретению может содержать цистеиновый остаток в положении 226 в последовательности и/или в положении 229 в последовательности одного из соответствующих шарнирных доменов, согласно нумерации по Kabat [EU-индекс].
[0078] Согласно вышеприведенному раскрытию молекул биспецифических антител, которые содержат набор CDR-областей (например, последовательности CDR SEQ ID NO: 3-SEQ ID NO:8 или последовательность с 80% идентичностью последовательности), которые опосредуют связывание PSMA и/или связывание с клетками плоскоклеточного рака, молекула антитела по настоящему изобретению может содержать второй сайт связывания, который специфически связывается с рецептором на поверхности иммунной клетки, такой как T-клетка или NK-клетки. Этот рецептор, присутствующий на поверхности иммунной клетки, может представлять собой рецептор, который способен активировать иммунную клетку или стимулировать иммунный ответ иммунной клетки. Индуцированный иммунный ответ предпочтительно может представлять собой цитотоксический иммунный ответ. Такой подходящий рецептор может, например, представлять собой CD3, антигенспецифический T-клеточный рецептор (TCR), CD28, CD16, NKG2D, Ox40, 4-1 BB, CD2, CD5, белок 1 запрограммированной гибели клеток (PD-1) и CD95. Особенно предпочтительной является молекула антитела, в которой второй сайт связывания связывается с CD3, TCR или CD16. Наиболее предпочтительной является молекула антитела, в которой второй сайт связывания специфически связывается с CD3. Предпочтительная молекула антитела содержит второй сайт связывания, который соответствует антигенсвязывающему сайту антитела к CD3 OKT3. Последовательности VH и VL антитела OKT3 представлены на фиг. 12. Аминокислотные последовательности вариабельного домена тяжелой цепи и вариабельного домена легкой цепи антитела OKT3 также описаны, например, в Arakawa et al., J. Biochem. 120, 657-662 (1996) и международной заявке на патент WO 2015/158868 (см. SEQ ID NO: 17 и 18 в перечне последовательностей WO 2015/158868). Другая предпочтительная молекула антитела содержит второй сайт связывания, который соответствует антигенсвязывающему сайту антитела к CD3 UCHT1. Последовательности VH и VL гуманизированного антитела UCHT1 представлены на фиг. 11 и, например, также описаны в международной заявке на патент WO 2013/092001. Другие примеры CD3-связывающих молекул антител, который можно применять в настоящем изобретении, включают молекулы антител, описанные в в заявке на европейский патент 2155783 B1 или заявке на европейский патент EP 2155788 B1, которые способны к связыванию с эпитопом CD3ε-цепи человека и Callithrix jacchus, Saguinus oedipus или Saimiri sciureus.
[0079] Соответственно, молекула биспецифического антитела по настоящему изобретению может представлять собой молекулу биспецифического антитела, такую как Fabsc- и IgGsc-молекула, которая содержит Fab-фрагмент и scFv-фрагмент, как описано в настоящем документе. В такой молекуле первый сайт связывания может связываться с PSMA и может содержаться в пределах Fab-фрагмента, как описано в настоящем документе, а второй сайт связывания (который может связываться с иммунным рецептором) может содержаться в пределах scFv-фрагмента. В качестве альтернативы, первый сайт связывания, который связывается с PMSA, содержится в пределах одноцепочечного Fv-фрагмента, а второй сайт связывания (который может связываться с иммунным рецептором) содержится в пределах Fab-фрагмента.
[0080] Согласно некоторым вариантам осуществления молекула биспецифического антитела по настоящему изобретению при связывании, например, связывании с CD3, сама по себе не активирует иммунную клетку, например, T-клетку. Вместо этого, только когда оба сайта связывания, например, PMSA-специфический сайт связывания и CD3-специфический сайт связывания, связываются с рецептором на поверхности T-клетки и с PMSA на поверхности целевой клетки, первый может перекрестно связывать активирующий рецептор, запуская эффекторные клетки для уничтожения конкретной целевой клетки. Для оценки и/или скрининга способности молекулы антитела активировать рецептор, с которым она связывается, можно проводить стандартные функциональные анализы для изучения способности лимфоцитов уничтожать целевые клетки в присутствии и отсутствие молекулы биспецифического антитела по настоящему изобретению.
[0081] Не вдаваясь в теорию, как правило, полагают, что биспецифические CD3-связывающие антитела по настоящему изобретению не активируют T-клетки в отсутствие целевых клеток, поскольку моновалентной стимуляции CD3 недостаточно для инициации эффективной активации T-клеток. Однако в некоторых случаях может иметь место некоторое отклонение от этого предполагаемого характера действия. Согласно настоящему документу, в случае применения UCHT1 в качестве антитела к CD3 в составе биспецифической конструкции Fabsc, отмечали некоторую активацию T-клеток в отсутствие экспрессирующих PSMA целевых клеток. Эти результаты были значительно более выраженными, когда эксперименты проводили в присутствии стимулирующих «фоновых» клеток, таких как клетки лимфомы SKW6.4. В отличие от этого, авторы настоящего изобретения также неожиданно обнаружили, согласно настоящему документу, что содержащие OKT3 антитела и - неожиданно - также содержащее UCHT1 антитело IgGsc, индуцируют заметно более слабую нецелевую активацию T-клеток. Неожиданно низкая степень нецелевой активации содержащим UCHT1 антителом IgGsc по меньшей мере частично объяснялась показателями авидности с применением экспрессирующих CD3 клеток Jurkat. В этих экспериментах было продемонстрировано, что UCHT1 утрачивает, а OKT3 приобретает некоторую степень авидности в случае экспрессии в составе формата IgGsc (а не Fabsc) (ср. фиг. 8). При тестировании в расширенном анализе цитотоксичности (Xelligence) с использованием экспрессирующих PSMA клеток 22RV1, авторы настоящего изобретения неожиданно наблюдали следующую градацию цитолитической активности: IgGsc(UCHT1) ~ Fabsc(UCHT1) > Fabsc(OKT3) > IgGsc(OKT3). Таким образом, в составе формата IgGsc предпочтительным может быть антитело к CD3 UCHT1 (подходящая нецелевая активация и цитолитическая активность), тогда как в составе формата Fabsc предпочтительным может быть OKT3 ввиду нежелательной нецелевой активации содержащим UCHT1 Fabsc (ср. фиг. 9).
[0082] Независимо от конкретного применяемого антитела к CD3, кластеризация биспецифических антител на поверхности T-клетки, индуцируемая взаимодействием нацеливающих одноцепочечных фрагментов, может также приводить к нецелевой активации T-клеток в отсутствие целевой клетки. С целью предотвращения этого явления может потребоваться применение Fab- или Fab2-компонента, такого как компонент, содержащийся в формате Fabsc или IgGsc соответственно, вместо одноцепочечного антитела в качестве нацеливающей части в составе биспецифической конструкции. Степень мультимеризации и агрегации биспецифических антител, экспрессируемых в таких форматах, заметно снижена по сравнению с таковой, наблюдаемой в случае биспецифических одноцепочечных антител. На фигурах 4A и 4B показано, что агрегация биспецифического одноцепочечного формата со специфичностью FLT3×CD3 менее выражена по сравнению с таковой другого идентичного антитела в виде биспецифического одноцепочечного (bssc) или BiTE-формата, как определено с помощью эксклюзионной хроматографии. На фигурах 4C-F показано, что, аналогично, склонность четырех антител к PSMA×CD3 в формате Fabsc, а также IgGsc, образовывать мультимеры или агрегаты сильно снижена. Следует отметить, что авторы настоящего изобретения неожиданно обнаружили, что образование мультимеров является еще значительно менее выраженным у двух содержащих антитело UCHT1 конструкций, чем у содержащих антитело OKT3 конструкций. В любом случае полагают, что агрегация, если таковая имеет место, обусловлена эффекторной частью CD3, экспрессируемой в виде одной цепи. Как полагают, и как было обнаружено, согласно настоящему документу, что физиологические Fab- и Fab2-компоненты в N-концевой нацеливающей части не агрегируют, и, таким образом, полагают, что кластеризация одноцепочечных компонентов N-концевой целенаправленно воздействующей части, вероятно приводящая в результате к истощению T-клеток in vivo, не может происходить у соответствующей молекулы антитела по настоящему изобретению.
[0083] Кроме того, авторы настоящего изобретения, согласно настоящему документу, рассчитывают, что время полужизни молекулы антитела в сыворотке крови преимущественно определяется взаимодействием CH3-доменов с FcRn-рецептором. Поскольку у большинства биспецифических антител этот домен отсутствует, их время полужизни в сыворотке крови может быть коротким (в лучшем случае несколько часов). В отличие от этого, молекула полного IgG обычно характеризуется временем полужизни в сыворотке крови, составляющим несколько дней. Хотя биспецифические форматы на основе IgG являются доступными уже в течение нескольких лет, их редко применяли для конструирования CD3-стимулирующих антител, поскольку считалось, что бивалентная стимуляция CD3 может приводить к нецелевой активации T-клеток. Однако авторы настоящего изобретения неожиданно обнаружили, что обратное справедливо для двух разных форматов, содержащих антитело UCHT1: нецелевая активация T-клеток форматом Fabsc заметно превышает таковую, обусловленную антителом в составе формата IgGsc. Не вдаваясь в теорию, полагают, что это связано с тем, что авидность CD3-компонента в составе последнего формата уменьшена (ср. опять же фиг. 8). Это означает, в случае молекулы антитела, содержащей вариабельную область UCHT1, что IgGsc-формат неожиданно обеспечивает не только заметно уменьшенное время полужизни в сыворотке крови, но также сниженную степень нецелевой активации T-клеток. Следовательно, если требуется продленное время полужизни в сыворотке крови, например, во избежание необходимости длительной непрерывной инфузии, предпочтительным может быть биспецифическое антитело 10B3×UCHT1 в формате IgGsc, где Fab-компонент содержит антигенсвязывающий сайт антитела, происходящего из 10B3, и где scFv-компонент содержит антигенсвязывающий сайт антитела, происходящего из UCHT1. Если продленное время полужизни в сыворотке крови не требуется, предпочтительным может быть биспецифическое антитело 10B3×OKT3 в формате Fabsc, где Fab-компонент содержит антигенсвязывающий сайт антитела, происходящего из 10B3, и где scFv-компонент содержит антигенсвязывающий сайт антитела, происходящего из OKT3.
[0084] Таким образом, другими словами, настоящее изобретение согласно альтернативному аспекту дополнительно относится к молекуле тетравалентного и гомодимерного биспецифического антитела (биспецифическое антитело в описанном в настоящем документе формате IgGsc), содержащей в каждом мономере: (i) N-концевой Fab-фрагмент, содержащий вариабельную область, содержащую вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, где указанная вариабельная область содержит первый сайт связывания, способный к связыванию с антигеном; (ii) C-концевой scFv-фрагмент, содержащий вариабельную область тяжелой цепи и вариабельную область легкой цепи гуманизированного UCHT1, и где (i) и (ii) соединены с помощью CH2- и CH3-домена.
[0085] Согласно некоторым вариантам осуществления предпочтительной является молекула тетравалентного биспецифического антитела по настоящему изобретению, где по меньшей мере один аминокислотный остаток CH2-домена, который способен опосредовать связывание с Fc-рецепторами, отсутствует или подвержен мутации (см. также в другом месте настоящего документа).
[0086] Согласно некоторым вариантам осуществления предпочтительной является молекула тетравалентного биспецифического антитела по настоящему изобретению, где Fab-фрагмент не является Fab-фрагментом негуманизированного, химерного или гуманизированного антитела 10B3 или J591, где предпочтительно первый сайт связывания не способен к связыванию с PSMA. Таким образом, согласно некоторым предпочтительным вариантам осуществления первый сайт связывания, обеспечиваемый N-концевым Fab-фрагментом, является специфическим в отношении не PSMA, а отличного от PSMA антигена, предпочтительно опухоль-ассоциированного антигена за исключением PSMA. Термин «опухоль-ассоциированный антиген» относится к белкам, которые в нормальных условиях, т.е. у здорового субъекта, специфично экспрессируются в ограниченном числе органов и/или тканей или на определенных стадиях развития, например, опухоль-ассоциированный антиген в нормальных условиях может специфично экспрессироваться в ткани желудка, предпочтительно в слизистой оболочке желудка, в репродуктивных органах, например, в яичке, ткани трофобласта, например, в плаценте, или в клетках зародышевой линии, и которые экспрессируются или аномально экспрессируются в одной или нескольких тканях опухоли или злокачественной опухоли. В таком контексте «ограниченное число» предпочтительно означает не более 3, более предпочтительно не более 2 или 1. Опухоль-ассоциированные антигены в контексте настоящего изобретения включают, например, дифференцировочные антигены, предпочтительно дифференцировочные антигены, специфичные для типа клеток, т.е. белки, которые в нормальных условиях специфично экспрессируются в клетках определенного типа на определенной стадии дифференцировки, антигены злокачественной опухоли/яичка, т.е. белки, которые в нормальных условиях специфично экспрессируются в яичке и в некоторых случаях в плаценте, и специфичные для зародышевой линии антигены. В контексте настоящего изобретения опухоль-ассоциированный антиген предпочтительно не экспрессируется или экспрессируется только иногда в здоровых тканях, или является мутированным в опухолевых клетках. Предпочтительно опухоль-ассоциированный антиген или аномальная экспрессия опухоль-ассоциированного антигена определяют клетки злокачественной опухоли. В контексте настоящего изобретения опухоль-ассоциированный антиген, который экспрессируется клеткой злокачественной опухоли у субъекта, например, пациента, страдающего от заболевания, связанного со злокачественной опухолью, предпочтительно является собственным белком организма указанного субъекта. Согласно предпочтительным вариантам осуществления опухоль-ассоциированный антиген в контексте настоящего изобретения в нормальных условиях специфично экспрессируется в ткани или органе, которые не являются критически важными, т. е. тканях или органах, которые при повреждении с участием иммунной системы не приводят к смерти субъекта, или в органах или структурах организма, которые являются недоступными или труднодоступными для иммунной системы. Предпочтительно опухоль-ассоциированный антиген представляется в контексте молекул MHC клеткой злокачественной опухоли, в которой он экспрессируется.
[0087] Примерами TAA, которые могут быть пригодны в настоящем изобретении, и которые не являются PSMA - в соответствии с предпочтительными вариантами осуществления настоящего аспекта - являются p53, ART-4, BAGE, бета-катенин/m, Bcr-abL CAMEL, CAP-1, CASP-8, CDC27/m, CDK4/m, CEA, КЛАУДИН-12, c-MYC, CT, Cyp-B, DAM, ELF2M, ETV6-AML1, G250, GAGE, GnT-V, Gap 100, HAGE, HER-2/neu, HPV-E7, HPV-E6, HAST-2, hTERT (или hTRT), LAGE, LDLR/FUT, MAGE-A, предпочтительно MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11 или MAGE-A12, MAGE-B, MAGE-C, MART-1/Melan-A, MC1R, миозин/m, MUCl", MUM-1, -2, -3, NA88-A, NF1, NY-ESO-1, NY-BR-1, p190 минорный BCR-abL, Pml/RARa, PRAME, протеиназа 3, PSA, RAGE, RU1 или RU2, SAGE, SART-1 или SART-3, SCGB3A2, SCP1, SCP2, SCP3, SSX, СУРВИВИН, TEL/AML1, TPl/m, TRP-1, TRP-2, TRP-2/INT2, TPTE и WT.
[0088] В соответствии с данным аспектом настоящего изобретения гуманизированное UCHT1 предпочтительно представляет собой scFv-фрагмент, содержащий последовательность вариабельной области легкой цепи и последовательность вариабельной области тяжелой цепи, как показано в SEQ ID NO: 11, начинающейся с аминокислотной последовательности DIQMT… и оканчивающейся…VTVSS (как указано на фиг. 11). Предпочтительно последовательность вариабельной области легкой цепи и последовательность вариабельной области тяжелой цепи в scFv-фрагменте соединены посредством линкера, как представлено на фиг. 11 («линкерная область»).
[0089] В этом контексте следует отметить, что в объем настоящего изобретения входит тот факт, что молекула антитела может содержать один или несколько мутированных аминокислотных остатков. Термины «мутированный», «мутант» и «мутация» в отношении нуклеиновой кислоты или полипептида относятся к изменению, делеции или вставке одного(одной) или нескольких нуклеотидов или аминокислот соответственно по сравнению с встречающейся в «природе» нуклеиновой кислотой или полипептидом, т.е. эталонной последовательностью, которая может быть использована для определения дикого типа. Например, вариабельные домены молекулы антитела 10B3, полученной путем иммунизации и описанной в настоящем документе, могут быть использованы в качестве последовательности дикого типа.
[0090] В этом отношении следует понимать, что термин «положение», при применении в соответствии с настоящим изобретением, означает положение аминокислоты в пределах аминокислотной последовательности, представленной в настоящем документе. Это положение может быть указано относительно сходной нативной последовательности, например. последовательности домена или цепи встречающегося в природе IgG. Термин «соответствующий» в контексте настоящего документа также предусматривает, что положение не обязательно определяется номером предшествующих нуклеотидов/аминокислот, или определяется не только им. Таким образом, положение данной аминокислоты в соответствии с настоящим изобретением, которую можно заменять, может варьировать в связи с делецией или добавлением аминокислот в другом месте цепи антитела.
[0091] Таким образом, под «соответствующим положением» согласно настоящему изобретению следует понимать, что аминокислоты могут отличаться по указанному номеру, но при этом могут характеризоваться аналогичными соседними аминокислотами. Указанные аминокислоты, которые можно изменять, удалять или добавлять, также охвачены термином «соответствующее положение». С целью определения того, соответствует ли аминокислотный остаток в данной аминокислотной последовательности определенному положению в аминокислотной последовательности домена или цепи встречающегося в природе иммуноглобулина, специалист в данной области может применять средства и способы, хорошо известные в данной области, например, варианты выравнивания, либо вручную, либо с применением компьютерных программ, таких как BLAST2.0, что расшифровывается как «средство поиска основного локального выравнивания», или ClustalW или другая подходящая программа, которая подходит для выполнения выравниваний последовательностей.
[0092] Согласно некоторым вариантам осуществления замена (или замещение) представляет собой консервативную замену. Консервативные замены обычно представляют собой следующие замены, перечисленные в соответствии с аминокислотой, подлежащей мутации, каждая из которых следует за одной или несколькими замещениями, которые можно считать консервативными: Ala → Gly, Ser, Val; Arg → Lys; Asn → Gln, His; Asp → Glu; Cys → Ser; Gln → Asn; Glu → Asp; Gly → Ala; His → Arg, Asn, Gln; Ile → Leu, Val; Leu → Ile, Val; Lys → Arg, Gln, Glu; Met → Leu, Tyr, Ile; Phe → Met, Leu, Tyr; Ser → Thr; Thr → Ser; Trp → Tyr; Tyr → Trp, Phe; Val → Ile, Leu. Другие замены также возможны, и они могут быть определены экспериментально или в соответствии с другими известными консервативными или неконсервативными заменами. В качестве дополнительного ориентира представлены следующие восемь групп, каждая из которых содержит аминокислоты, которые обычно можно использовать для определения консервативных замен одной на другую:
[0093] Аланин (Ala), глицин (Gly);
[0094] Аспарагиновая кислота (Asp), глутаминовая кислота (Glu);
[0095] Аспарагин (Asn), глутамин (Gln);
[0096] Аргинин (Arg), лизин (Lys);
[0097] Изолейцин (Ile), лейцин (Leu), метионин (Met), валин (Val);
[0098] Фенилаланин (Phe), тирозин (Tyr), триптофан (Trp);
[0099] Серин (Ser), треонин (Thr); и
[00100] цистеин (Cys), метионин (Met)
[00101] Если такие замены приводят к изменению биологической активности, то можно вводить более значительные изменения, такие как следующие, или как дополнительно описано ниже в отношении классов аминокислот, и продукты подвергать скринингу в отношении требуемой характеристики. Примерами таких более значительных изменений являются: Ala → Leu, Ile; Arg → Gln; Asn → Asp, Lys, Arg, His; Asp → Asn; Cys → Ala; Gln → Glu; Glu → Gln; His → Lys; Ile → Met, Ala, Phe; Leu → Ala, Met, норлейцин; Lys → Asn; Met → Phe; Phe → Val, Ile, Ala; Trp → Phe; Tyr → Thr, Ser; Val → Met, Phe, Ala.
[00102] Согласно некоторым вариантам осуществления молекула антитела в соответствии с настоящим изобретением содержит один или несколько аминокислотных остатков, включая два, три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать, двенадцать, тринадцать, четырнадцать, пятнадцать, шестнадцать, семнадцать или восемнадцать аминокислотных остатков, которые подвергают мутации с целью предотвращения димеризации посредством цистеиновых остатков или с целью модуляции функции Fc (см. выше). Согласно некоторым из таких вариантов осуществления один или несколько аминокислотных остатков CH2-домена и/или шарнирной области, которые способны опосредовать связывание с Fc-рецепторами, подвержены мутации. Если присутствуют, один или несколько аминокислотных остатков, способных опосредовать связывание с Fc-рецепторами, могут представлять собой аминокислотные остатки, которые способны активировать антителозависимую клеточную цитотоксичность (ADCC) или комплементопосредованную цитотоксичность (CDC). Согласно некоторым вариантам осуществления соответствующий аминокислотный остаток, способный опосредовать связывание с Fc-рецепторами, заменяют другой аминокислотой, как правило, при сравнении последовательности с последовательностью соответствующего встречающегося в природе домена иммуноглобулина, такого как IgG. Согласно некоторым вариантам осуществления такой аминокислотный остаток, способный опосредовать связывание с Fc-рецепторами, удаляют, как правило, по сравнению с последовательностью соответствующего встречающегося в природе домена иммуноглобулина, такого как IgG.
[00103] Согласно некоторым вариантам осуществления один или несколько подверженных мутации, например, замененных или удаленных, аминокислотных остатков представляют собой аминокислоты, расположенные в одном из положений 226, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 265, 297, 327 и 330. Опять же, применяемая нумерация аминокислот соответствует положениям в последовательности в соответствии с нумерацией по Kabat [EU-индекс]. Соответствующая делеция аминокислоты может представлять собой, например, делецию аминокислоты 228, как правило, пролина в IgG, делецию аминокислоты 229, как правило, цистеина в IgG, делецию аминокислоты 230, как правило, пролина в IgG, делецию аминокислоты 231, как правило, аланина в IgG, делецию аминокислоты 232, как правило, пролина в IgG, делецию аминокислоты 233, как правило, глутаминовой кислоты в IgG, делецию аминокислоты 234, как правило, лейцина в IgG, делецию аминокислоты 235, как правило, лейцина в IgG, делецию аминокислоты 236, как правило, глицина в IgG, делецию аминокислоты 237, как правило, глицина в IgG, делецию аминокислоты 238, как правило, пролина в IgG и делецию аминокислоты 265, как правило, аспарагиновой кислоты в IgG. Соответствующая замена аминокислоты может представлять собой, например, замену аминокислоты 226, как правило, цистеина в IgG, замену аминокислоты 228, как правило, пролина в IgG, замену аминокислоты 229, как правило, цистеина в IgG, замену аминокислоты 230, как правило, пролина в IgG, замену аминокислоты 231, как правило, аланина в IgG, замену аминокислоты 232, как правило, пролина в IgG, замену аминокислоты 233, как правило, глутаминовой кислоты в IgG, замену аминокислоты 234, как правило, лейцина в IgG, замену аминокислоты 235, как правило, лейцина в IgG, замену аминокислоты 265, как правило, аспарагиновой кислоты в IgG, замену аминокислоты 297, как правило, аспарагина в IgG, замену аминокислоты 327, как правило, аланина в IgG и замену аминокислоты 330, как правило, аланина в IgG. Соответствующая замена может представлять собой одну из замены Cys226→Ser, замены Cys229→Ser, замены Glu233→Pro, замены Leu234→Val, замены Leu235→Ala, замены Asp265→Gly, замены Asn297→Gln, замены Ala327→Gln, замены Ala327→Gly и замены Ala330→Ser. Как можно сделать вывод из вышеописанного, согласно некоторым вариантам осуществления один или два из цистеиновых остатков в положениях 226 и 229 в шарнирной области заменены другой аминокислотой, к примеру, заменены сериновым остатком. С помощью этого можно предотвратить образование дисульфидной связи с другой основной цепью. Кроме того, и как также объясняется ниже, удаление и/или замена (мутирование) выбранных аминокислотных остатков в CH2-домене, который способен опосредовать связывание с Fc-рецепторами, может обуславливать тот факт, что молекула антитела по настоящему изобретению характеризуется менее выраженной активностью в отношении антителозависимой клеточноопосредованной цитотоксичности и связывания комплемента, или отсутствием такой активности.
[00104] Другой тип аминокислотной модификации антитела обеспечивает изменение исходного паттерна гликозилирования (если таковой имеется) молекулы антитела. Под изменением подразумевается удаление одного или нескольких углеводных компонентов, обнаруживаемых в антителе, и/или добавление одного или нескольких сайтов гликозилирования, которые отсутствуют в антителе. Гликозилирование антител обычно является либо N-сцепленным, либо O-сцепленным. N-сцепленное относится к присоединению углеводного компонента к боковой цепи аспарагинового остатка. Последовательности трипептида аспарагин-X-серин и аспарагин-X-треонин, где X представляет собой любую аминокислоту за исключением пролина, являются последовательностями распознавания для ферментативного присоединения углеводного компонента к боковой цепи аспарагина. Таким образом, присутствие любой из таких последовательностей трипептида в полипептиде создает потенциальный сайт гликозилирования. O-сцепленное гликозилирование относится к присоединению одного из N-ацетилгалактозамина, галактозы или ксилозы, являющихся сахарами, к гидроксиаминокислоте, чаще всего серину или треонину, хотя также можно применять 5-гидроксипролин или 5-гидроксилизин. Добавление в антитело сайтов гликозилирования успешно выполняют путем изменения аминокислотной последовательности так, чтобы она содержала одну или несколько из вышеописанных последовательностей трипептида (для сайтов N-сцепленного гликозилирования). Изменение также можно выполнять путем добавления к последовательности исходного антитела одного или нескольких сериновых или треониновых остатков, или путем замены ими (для сайтов O-сцепленного гликозилирования).
[00105] В контексте настоящего изобретения, согласно некоторым вариантам осуществления, участок основной цепи молекулы антитела по настоящему изобретению, который представляет собой Fc-область иммуноглобулина, обычно является инертным, или по меньшей мере в основном оказывает слабое влияние на связывание с Fc-рецепторами. Как уже говорилось, «ослабление функции Fc» обеспечивается путем удаления и/или замены (мутирования) по меньшей мере одного из выбранных аминокислотных остатков в CH2-домене, которые способны опосредовать связывание с Fc-рецептором. Такие молекулы также называют в настоящем документе молекулами антител с «ослабленной функцией связывания Fc» или молекулами антител «Fcko». В соответствии с настоящим изобретением участок цепи антитела, который может быть использован в качестве участка Fc-фрагмента, т. е. CH2-домена, и, где это применимо, CH3-домена, может, таким образом, определять «остов» без обеспечения конкретной биологической функции, такой как, например, эффекторная функция. Однако в настоящем изобретении было обнаружено, что такой остов может обеспечивать существенные преимущества с точки зрения очистки, эффективности продукции и/или стабильности молекул антител по настоящему изобретению по сравнению с известными молекулами антител.
[00106] Согласно некоторым вариантам осуществления степень распознавания и, соответственно, связывания этого соответствующего Fc участка с данным Fc-рецептором в приблизительно 2 раза, приблизительно 5 раз, приблизительно 8 раз, приблизительно 10 раз, приблизительно 12 раз, приблизительно 15 раз, приблизительно 20 раз меньше, чем таковая у Fc-области встречающегося в природе иммуноглобулина. Согласно некоторым вариантам осуществления этот соответствующий Fc участок полностью лишен его способности связываться с Fc-рецепторами. Связывание антитела с Fc-рецепторами, включая определение константы диссоциации, без труда может быть определено специалистом в данной области с применением стандартных методик, таких как метод поверхностного плазмонного резонанса, например, с применением измерения Biacore™. Аналогичным образом можно применять любой другой способ измерения параметров связывания биомолекул, к примеру, с использованием спектроскопических, фотохимических, фотометрических или радиологических методов. Примерами соответствующих способов выявления являются флуоресцентная корреляционная спектроскопия, фотохимическое сшивание и применение соответственно фотоактивных или радиоактивных меток Некоторые из этих способов могут включают дополнительные методики разделения, такие как электрофорез или HPLC.
[00107] С этой целью, при необходимости, можно осуществлять замену или делецию аминокислотных остатков, как объяснялось выше. Примеры подходящих мутаций могут быть заимствованы из Armour et al., (например, Eur. J. Immunol. [1999] 29, 2613-2624). Дополнительные примеры подходящих положений для осуществления мутаций в последовательности цепи антитела могут быть заимствованы из опубликованных данных о кристаллической структуре комплекса FcγRIII и Fc-фрагмента IgG1 человека (Sondermann et al., Nature [2000] 406, 267-273). В дополнение к измерению аффинности связывания, как описано выше, с целью оценки уровня «ослабления функции связывания Fc» или утраты аффинности связывания, также можно выполнять функциональную оценку (в отношении отсутствия) способности опосредовать связывание с Fc-рецептором. В случае молекул антител, которые связывают CD3 в качестве мишени, связывание можно оценивать, например, по митогенному потенциалу CD3-связывающих молекул антител по отношению к клеткам. Митогенный потенциал опосредован связыванием антител к CD3 с Fc-рецепторами на поверхности антиген-представляющих клеток, таких как моноциты. Если молекула антитела по настоящему изобретению, которая содержит один сайт связывания CD3, не оказывает митогенный эффект, тогда как исходное моноклональное антитело к CD3, которое содержит функциональную Fc-часть, индуцирует интенсивный митоз у T-клеток, очевидно что, как следует из отсутствия митоза, у молекулы антитела по настоящему изобретению отсутствует способность связывания Fc, и она может, таким образом, считаться «нокаутной по Fc» молекулой. Иллюстративные примеры способа оценки опосредованного антителами к CD3 митогенного потенциала были описаны Davis, Vida & Lipsky (J.Immunol (1986) 137, 3758) и Ceuppens, JL, & van Vaeck, F, (см. J.Immunol. (1987) 139, 4067 или Cell. Immunol. (1989) 118, 136). Дополнительно иллюстративные подходящие примеры анализа для оценки митогенного потенциала антитела были описаны Rosenthal-Allieri et al. (Rosenthal-Allieri MA, Ticcioni M, Deckert M, Breittmeyer JP, Rochet N, Rouleaux M, and Senik A, Bernerd A, Cell Immunol. 1995 163(1):88-95) и Grosse-Hovest et al. (Grosse-Hovest L, Hartlapp I, Marwan W, Brem G, Rammensee H-G, and Jung G, Eur J Immunol. [2003] May; 33(5):1334-1340). Кроме того, отсутствие связывания Fc можно оценивать по способности молекулы антитела по настоящему изобретению опосредовать одну или несколько из хорошо изученных эффекторных функций Fc-части.
[00108] Как отмечалось выше, замены или делеции цистеиновых остатков можно осуществлять с целью введения или удаления одной или нескольких дисульфидных связей, включая введение или удаление потенциальной или уже существующей дисульфидной связи. Следовательно, можно управлять связью между основной цепью и цепью с меньшим весом/меньшей длиной молекулы антитела в соответствии с настоящим изобретением, включая ее создание, усиление или устранение. Путем введения или удаления одного или нескольких цистеиновых остатков можно вводить или удалять дисульфидный мостик. В качестве иллюстративного примера, молекула антитела в соответствии с настоящим изобретением, как правило, содержит одну или несколько дисульфидных связей, которые соединяют две молекулы димерных антител. Одна такая дисульфидная связь обычно образуется между цистеином в основной цепи первой молекулы димерного антитела и цистеином в шарнирной области второй молекулы димерного антитела. В этом отношении, согласно некоторым вариантам осуществления антитело в соответствии с настоящим изобретением по сравнению с последовательностью иммуноглобулина IgG человека может предусматривать аминокислотную замену нативного цистеинового остатка в положениях 226 и/или 229, в соответствии с нумерацией по Kabat [EU-индекс], другим аминокислотным остатком.
[00109] Замены или делеции аминокислотных остатков, таких как аргининовые, аспарагиновые, сериновые, треониновые или тирозиновые остатки, также можно осуществлять для модификации паттерна гликозилирования антитела. В качестве иллюстративного примера, молекула IgG содержит один N-сцепленный углевод c двумя ветвями при Asn297 CH2-домена. В случае IgG, либо из сыворотки крови, либо продуцируемого ex vivo в гибридомах или сконструированных клетках, IgG являются гетерогенными в отношении Asn297-сцепленного углевода. В случае IgG человека, центральный олигосахарид обычно состоит из GlcNAc2Man3GlcNAc, с разным числом внешних остатков.
[00110] Как указано, помимо связывания антигенов/эпитопов, иммуноглобулин, как известно, выполняет дополнительные «эффекторные функции», виды биологической активности, приписываемые Fc-области (нативной последовательности Fc-области или аминокислотной последовательности вариантной Fc-области) иммуноглобулина, и варьирующие в зависимости от изотипа иммуноглобулина. Примеры эффекторных функций антитела включают: связывание Clq и комплементзависимую цитотоксичность (CDC); связывание Fc-рецептора; антителозависимую клеточноопосредованную цитотоксичность (ADCC); фагоцитоз; подавление рецепторов клеточной поверхности (например, B-клеточных рецепторов) и активацию B-клеток. Проявлением эффекторных функций антитела, как правило, является рекрутирование эффекторных клеток. Некоторые эффекторные функции иммуноглобулина опосредованы Fc-рецепторами (FcR), которые связывают Fc-область антитела. FcR определяются их специфичностью в отношении изотипов иммуноглобулина; Fc-рецепторы для IgG-антител называют FcγR, для IgE FcεR, для IgA FcαR и так далее. Любую из этих эффекторных функций (или утрату таких эффекторных функций), такую как CDC или ADCC, можно применять для оценки того, отсутствует ли у молекулы антитела по настоящему изобретению способность связываться с Fc.
[00111] В этом контексте следует отметить, что термином «Fc-рецептор» или «FcR» определяется рецептор, как правило, белок, который способен к связыванию с Fc-областью антитела. Fc-рецепторы находятся на поверхности определенных клеток иммунной система организма, например, естественных клеток киллеров, макрофагов, нейтрофилов и тучных клеток. In vivo Fc-рецепторы связываются с иммуноглобулинами, которые иммобилизованы на поверхности инфицированных клеток или представлены на инвазивных патогенах. Их активность стимулирует фагоцитарные или цитотоксические клетки в отношении разрушения микроорганизмов или инфицированных клеток посредством опосредованного антителом фагоцитоза или антителозависимой клеточноопосредованной цитотоксичности. Некоторые вирусы, такие как флавивирусы, используют Fc-рецепторы для содействия им в отношении инфицирования клеток с помощью механизма, известного как антителозависимое усиление инфекции. FcR рассмотрены в Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-92 (1991); Capel et al., Immunomethods 4: 25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126: 330-41 (1995).
[00112] «Комплементзависимая цитотоксичность» или «CDC» относится к лизису целевой клетки в присутствии системы комплемента. Активация системы комплемента по классическому пути инициируется связыванием первого компонента системы комплемента (Clq) с антителами (соответствующего подкласса), которые связаны с их когнатным антигеном. Для оценки активации системы комплемента можно осуществлять анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1997).
[00113] Термин «система комплемента» применяют в данной области для обозначения совокупности небольших белков - называемых факторами комплемента - обнаруживаемых в крови, как правило, циркулирующих в виде неактивных предшественников (пре-белков). Термин относится к способности такой неизменной и неадаптируемой системы «дополнять» способность антител и фагоцитарных клеток устранять из организма патогены, такие как бактерии, а также комплексы антиген-антитело. Примером факторов комплемента является комплекс C1, который включает C1q и две сериновые протеазы, C1r и C1s. Комплекс C1 является компонентом пути CDC. C1q представляет собой шестивалентную молекулу с молекулярной массой примерно 460000 и структурой, напоминающей букет тюльпанов, в которой шесть коллагеноподобных «стеблей» соединены с шестью глобулярными областями, представляющими собой «головку». Для активации каскада реакций системы комплемента C1q должен связаться по меньшей мере с двумя молекулами IgG1, IgG2 или IgG3.
[00114] «Антителозависимая клеточная цитотоксичность» или ADCC относится к форме цитотоксичности, при которой молекулы иммуноглобулинов, связанные на Fc-рецепторах (FcR), представляются определенным цитотоксическим клеткам - таким как естественные клетки-киллеры (NK), нейтрофилы и макрофаги - что позволяет этим цитотоксическим эффекторным клеткам специфически связываться с несущей антиген целевой клеткой и, впоследствии, уничтожать целевую клетку с помощью цитотоксинов. Антитела «вооружают» цитотоксические клетки и требуются для уничтожения целевой клетки посредством этого механизма. Основные клетки для опосредования ADCC, NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRlI и FcγRIII. Данные по экспрессии FcR на поверхности гемопоэтических клеток обобщены в таблице 3 на странице 464 Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-92 (1991). Для оценки ADCC-активности представляющей интерес молекулы можно осуществлять in vitro анализ ADCC, например, описанный в патентах США №№ 5500362 или 5821337. Пригодные эффекторные клетки для таких анализов включают без ограничения мононуклеарные клетки периферической крови (PBMC) и естественные клетки-киллеры (NK). Согласно некоторым вариантам осуществления ADCC-активность представляющей интерес молекулы можно оценивать in vivo, например, на животной модели, например, раскрытой в Clynes et al., PNAS USA 95: 652-656 (1998).
[00115] Молекулу антитела по настоящему изобретению можно получать с применением любой известной и хорошо отработанной системы экспрессии и технологии культивирования рекомбинантных клеток, например, путем обеспечения экспрессии в бактериальных хозяевах (прокариотические системы), или в эукариотических системах, таких как клетки дрожжей, грибов, насекомых или клетки млекопитающих. Например, в случае молекулы антитела по настоящему изобретению, применяемой в формате «IgGsc», молекула антитела может быть получена (как уже говорилось), как описано Coloma and Morrison (Nat Biotechnol 15:159-63, 1997), или как описано в разделе Примеры настоящей заявки. Аналогично, молекулу антитела по настоящему изобретению, используемую в формате «Fabsc», можно получать, как описано в международной заявке на патент WO 2013/092001 или как описано в данном документе в разделе Примеры. Молекулу антитела по настоящему изобретению также можно получать в трансгенных организмах, таких как коза, растение или трансгенная мышь XENOMOUSE, полученный с помощью методов генной инженерии штамм мыши, у которого имеются крупные фрагменты локусов генов иммуноглобулинов человека, и который является дефектным по продукции антител мыши. Антитело также можно получать путем химического синтеза.
[00116] Для получения рекомбинантной молекулы антитела по настоящему изобретению обычно выделяют кодирующий антитело полинуклеотид, и вставляют его в реплицируемый вектор, такой как плазмида, для дальнейшего клонирования (амплификации) или экспрессии. Иллюстративным примером подходящей системы экспрессии является система на основе подавления экспрессии глутаминсинтетазы (например, продаваемая Lonza Biologics), с использованием клетки-хозяина, представляющей собой, к примеру, CHO или NS0. Кодирующий антитело полинуклеотид без труда выделяют и подвергают секвенированию с применением традиционных процедур. Векторы, которые можно применять, включают плазмиду, вирус, фаг, транспозоны, мини-хромосомы, из которых плазмиды являются типичным вариантом осуществления. Как правило, такие векторы дополнительно включают сигнальную последовательность, точку начала репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательности терминации транскрипции, функционально связанные с кодирующим легкую и/или тяжелую цепь полинуклеотидом, с целью содействия экспрессии. Кодирующие легкие и тяжелые цепи полинуклеотиды можно вставлять в отдельные векторы и трансфицировать в одну и ту же клетку-хозяина или, при необходимости, последовательности как для тяжелой цепи, так и для легкой цепи можно вставлять в один и тот же вектор для трансфекции в клетку-хозяина. Обе цепи могут находиться, например, под контролем бицистронного оперона и экспрессироваться с образованием функциональной и надлежащим образом свернутой молекулы антитела, как описано в Skerra, A. (1994) Use of the tetracycline promoter for the tightly regulated production of a murine antibody fragment in Escherichia coli, Gene 151, 131-135, или Skerra, A. (1994) A general vector, pASK84, for cloning, bacterial production, and single-step purification of antibody Fab fragments, Gene 141, 79-8. Таким образом, в соответствии с одним аспектом настоящего изобретения предусмотрен способ конструирования вектора, кодирующего легкие и/или тяжелые цепи антитела или его антигенсвязывающего фрагмента по настоящему изобретению, причем способ предусматривает вставку в вектор полинуклеотида, кодирующего легкую цепь и/или тяжелую цепь молекулы антитела по настоящему изобретению.
[00117] В случае применения рекомбинантных методик молекула антитела может продуцироваться внутриклеточно, в периплазматическое пространство или секретироваться прямо в среду (ср. также Skerra 1994, выше). Если антитело продуцируется внутриклеточно, в качестве первой стадии, частицы в виде дебриса, относящиеся либо к клетке-хозяину, либо лизированным фрагментам, удаляют, например, путем центрифугирования или ультрафильтрации. В Carter et al., Bio/Technology 10: 163-167 (1992) описана процедура выделения антител, которые секретируются в периплазматическое пространство E coli. Антитело также может продуцироваться в любую окисляющую среду. Такая окисляющая среда может обеспечиваться периплазмой грамотрицательных бактерий, таких как E. coli, во внеклеточном пространстве грамположительных бактерий или в полости эндоплазматического ретикулума эукариотических клеток (включая клетки животных, такие как клетки насекомых или млекопитающих), и обычно способствует образованию участвующих в формировании пространственной структуры дисульфидных связей. Однако также возможным является продуцирование молекулы антитела по настоящему изобретению в цитозоле клетки-хозяина, такой как E. coli. В этом случае полипептид может быть либо непосредственно получен в растворимом и свернутом состоянии, либо выделен в форме телец включения, с последующей ренатурацией in vitro. Дополнительным вариантом является применение специфических штаммов-хозяев с окисляющей внутриклеточной средой, которая может, таким образом, обеспечивать образование дисульфидных связей в цитозоле (Venturi M, Seifert C, Hunte C. (2002) “High level production of functional antibody Fab fragments in an oxidizing bacterial cytoplasm.” J. Mol. Biol. 315, 1-8).
[00118] Продуцируемую клетками молекулу антитела можно очищать с применением любой традиционной технологии очистки, например, хроматографии на гидроксиапатите, гель-электрофореза, диализа и аффинной хроматографии, причем аффинная хроматография является одной из предпочтительных методик очистки. Молекулы антител можно очищать с помощью аффинной очистки с использованием белков/лигандов, которые специфически и обратимо связывают константные домены, такие как CH1- или CL-домены. Примерами таких белков являются иммуноглобулин-связывающие белки бактерий, такие как белок A, белок G, белок A/G или белок L, где связывание белка L ограничено молекулами антител, которые содержат легкие каппа-цепи. Альтернативным способом очистки антител с легкими цепями является применение связанных на гранулах антител к каппа-цепи (KappaSelect). Пригодность белка A в качестве аффинного лиганда зависит от вида и изотипа, к которому относится любой Fc-домен иммуноглобулина, который присутствует в антителе. Для очистки антител можно применять белок A (Lindmark et al., J. Immunol. Meth. 62: 1-13 (1983)). Белок G рекомендуется для всех изотипов мыши и для гамма 3 человека (Guss et al., EMBO J. 5: 15671575 (1986)). Выбор способа очистки, который применяют для конкретной молекулы антитела по настоящему изобретению, находится в пределах знаний специалиста в данной области.
[00119] Также можно обеспечивать одну из цепей молекулы антитела по настоящему изобретению одной или несколькими аффинными метками. Аффинные метки, такие как Strep-tag® или Strep-tag® II (Schmidt, T.G.M. et al. (1996) J. Mol. Biol. 255, 753-766), myc-метка, FLAG™-метка, His6-метка или HA-метка обеспечивают возможность несложного выявления, а также простой очистки рекомбинантной молекулы антитела.
[00120] Возвращаясь теперь к нуклеиновым кислотам по настоящему изобретению, молекула нуклеиновой кислоты, кодирующая одну или несколько цепей антитела в соответствии с настоящим изобретением, может представлять собой любую нуклеиновую кислоту в любой возможной конфигурации, например, однонитевую, двунитевую или их комбинацию. Нуклеиновые кислоты включают, к примеру, молекулы ДНК, молекулы РНК, аналоги ДНК или РНК, полученные с применением аналогов нуклеотидов или с применением химического синтеза нуклеиновых кислот, молекулы запертых нуклеиновых кислот (LNA) и молекулы пептидо-нуклеиновых кислот (PNA). ДНК или РНК могут быть геномного или синтетического происхождения, и могут быть одно- или двунитевыми. Такая нуклеиновая кислота может представлять собой, например, мРНК, кРНК, синтетическую РНК, геномную ДНК, кДНК, синтетическую ДНК, сополимер ДНК и РНК, олигонуклеотиды и т.д. Более того, соответствующая нуклеиновая кислота может содержать искусственные аналоги нуклеотидов и/или может быть соединена с аффинной меткой или маркером.
[00121] Согласно некоторым вариантам осуществления последовательность нуклеиновой кислоты, кодирующая цепь, например, основную цепь и/или меньшую цепь антитела в соответствии с настоящим изобретением, включена в вектор, такой как плазмида. Если в цепь антитела будет включена замена или делеция, по сравнению с встречающимися в природе доменом или областью антитела, кодирующая последовательность соответствующих нативных домена/области, например, включенных в последовательность иммуноглобулина, можно применять в качестве исходной точки для мутагенеза. Что касается мутагенеза по отношению к выбранным аминокислотным положениям, специалист в данной области имеет в своем распоряжении различные отработанные стандартные способы сайт-направленного мутагенеза. Широко применяемой методикой является введение мутаций с помощью ПЦР (полимеразной цепной реакции) с применением смесей синтетических олигонуклеотидов, которые несут набор вырожденных оснований в требуемых положениях в последовательности. Например, применение кодона NNK или NNS (где N = аденин, гуанин или цитозин или тимин; K = гуанин или тимин; S = аденин или цитозин) позволяет включать в ходе мутагенеза все 20 аминокислот и стоп-кодон «янтарь», тогда как кодон VVS ограничивает число аминокислот, которые можно включить, до 12, поскольку он исключает возможность включения аминокислот Cys, Ile, Leu, Met, Phe, Trp, Tyr, Val в выбранное положение в полипептидной последовательности; применение кодона NMS (где M = аденин или цитозин), например, ограничивает число аминокислот, которые можно включить, до 11 в выбранном положении в последовательности, поскольку он исключает возможность включения аминокислот Arg, Cys, Gly, Ile, Leu, Met, Phe, Trp, Val в выбранное положение в последовательности. В этом отношении следует отметить, что кодоны для других аминокислот (отличных от стандартных 20 встречающихся в природе аминокислот), таких как селеноцистеин или пирролизин, также можно включать в нуклеиновую кислоту, кодирующую молекулу антитела. Также можно, как описано Wang, L., et al. (2001) Science 292, 498-500 или Wang, L., and Schultz, P.G. (2002) Chem. Comm. 1, 1-11, применять «искусственные» кодоны, такие как UAG, которые обычно распознаются как стоп-кодоны, с целью вставки других нетипичных аминокислот, например, o-метил-L-тирозина или п-аминофенилаланина.
[00122] Применение элементарных звеньев в виде нуклеотидов со сниженной специфичностью спаривания оснований, как, например, инозина, 8-оксо-2’-дезоксигуанозина или 6-(2-дезокси-β-D-рибофуранозил)-3,4-дигидро-8H-пиримидо-1,2-оксазин-7-она (Zaccolo et al. (1996) J. Mol. Biol. 255, 589-603), является еще одним вариантом введения мутаций в выбранный сегмент последовательности. Дополнительным вариантом является так называемый тройной мутагенез. В этом способе применяют смеси нуклеотидных триплетов, каждый из которых кодирует одну аминокислоту для включения в кодирующую последовательность ( B, et al., 1994 Nucleic Acids Res 22, 5600-5607).
B, et al., 1994 Nucleic Acids Res 22, 5600-5607).
[00123] Молекула нуклеиновой кислоты, кодирующая цепь, такую как основная цепь и/или меньшая цепь, антитела в соответствии с настоящим изобретением, может быть экспрессирована с применением любой подходящей системы экспрессии, например, в подходящей клетке-хозяине или в бесклеточной системе. Полученную молекулу антитела можно подвергать обогащению с помощью отбора и/или выделения.
[00124] Настоящее изобретение также относится к фармацевтической композиции, которая включает молекулу антитела по настоящему изобретению и необязательно фармацевтически приемлемое вспомогательное вещество.
[00125] Молекулу антитела в соответствии с настоящим изобретением можно вводить посредством любого из парентерального или отличного от парентерального (энтерального) пути, который является терапевтически эффективным в случае белковых лекарственных средств. Способы парентерального применения предусматривают, например, методы внутрикожных, подкожных, внутримышечных, интратрахеальных, интраназальных, интравитреальных или внутривенных инъекции и инфузии, например, в форме инъекционных растворов, инфузионных растворов или настоев, а также аэрозольной инстилляции и ингаляции, например, в форме аэрозольных смесей, спреев или порошков. Обзор по легочному введению лекарственных средств, т. е. посредством либо ингаляции аэрозолей (которые также можно применять для интраназального введения), либо интратрахеальной инстилляции, дается J.S. Patton et al., (например, The lungs as a portal of entry for systemic drug delivery. Proc. Amer. Thoracic Soc. 2004, том 1, страницы 338-344). К способам доставки, отличной от парентеральной, относятся, к примеру, пероральный, например, в форме пилюль, таблеток, капсул, растворов или суспензий, или ректальный, например, в форме суппозиториев. Молекулы антител по настоящему изобретению можно вводить системно или местно в виде составов, содержащих, по необходимости, традиционные нетоксичные фармацевтически приемлемые вспомогательные вещества или носители, добавки и среды-носители.
[00126] Согласно одному варианту осуществления настоящего изобретения фармацевтический препарат вводят парентерально млекопитающему, и, в частности, людям. Соответствующие способы введения предусматривают без ограничения, например, методы внутрикожных, подкожных, внутримышечных, интратрахеальных или внутривенных инъекции и инфузии, например, в форме инъекционных растворов, инфузионных растворов или настоев, а также аэрозольной инстилляции и ингаляции, например, в форме аэрозольных смесей, спреев или порошков. Комбинация внутривенных и подкожных инфузии и/или инъекции может быть наиболее удобной в случае соединений с относительно коротким временем полужизни в сыворотке крови. Фармацевтическая композиция может представлять собой водный раствор, эмульсию типа «масло в воде» или эмульсию типа «вода в масле».
[00127] В этом отношении следует отметить, что для трансдермальной доставки описанной в настоящем документе молекулы антитела также можно применять такие методы трансдермальной доставки, как ионтофорез, сонофорез или доставка, опосредованная микроиглами, как описано в Meidan VM and Michniak BB 2004 Am. J. Ther. 11(4): 312-316. К способам доставки, отличной от парентеральной, относятся, к примеру, пероральный, например, в форме пилюль, таблеток, капсул, растворов или суспензий, или ректальное введение, например, в форме суппозиториев. Молекулы антител по настоящему изобретению можно вводить системно или местно в виде составов, содержащих ряд традиционных нетоксичных фармацевтически приемлемых вспомогательных веществ или носителей, добавок и сред-носителей.
[00128] Дозировка применяемой молекулы антитела для достижения требуемого профилактического эффекта или терапевтического ответа может варьироваться в широких пределах. Она будет зависеть, к примеру, от аффинности молекулы антитела к выбранной мишени, а также от времени полужизни комплекса молекулы антитела и лиганда in vivo. Кроме того, оптимальная дозировка будет зависеть от биораспределения молекулы антитела или конъюгата на ее основе, способа введения, тяжести подлежащих лечению заболевания/нарушения, а также медицинского состояния пациента. Например, в случае применения в виде мази для местного использования можно применять высокую концентрацию молекулы антитела. Однако при желании молекулу антитела можно также вводить в виде состава с замедленным высвобождением, например, в виде дисперсий липосом или полимерных микросфер на основе гидрогеля, таких как PolyActive™ или OctoDEX™ (ср. Bos et al., Business Briefing: Pharmatech 2003: 1-6). Другими доступными составами с замедленным высвобождением являются, например, полимеры на основе PLGA (PR pharmaceuticals), гидрогели на основе PLA-PEG (Medincell) и полимеры на основе PEA (Medivas).
[00129] Соответственно, молекулы антител по настоящему изобретению могут быть составлены в композиции с применением фармацевтически приемлемых ингредиентов, а также отработанных способов получения (Gennaro, A.L. and Gennaro, A.R. (2000) Remington: The Science and Practice of Pharmacy, 20-е издание, Lippincott Williams & Wilkins, Филадельфия, Пенсильвания). Для получения фармацевтических композиций можно применять фармацевтически инертные неорганические или органические вспомогательные вещества. Для получения, например, пилюль, порошков, желатиновых капсул или суппозиториев можно применять, например, лактозу, тальк, стеариновую кислоту и ее соли, жиры, воски, твердые или жидкие полиолы, натуральные и гидрогенизированные масла. Подходящие вспомогательные вещества для получения растворов, суспензий, эмульсий, аэрозольных смесей или порошков для восстановления растворов или аэрозольных смесей перед применением включают воду, спирты, глицерин, полиолы и их подходящие смеси, а также растительные масла.
[00130] Фармацевтическая композиция может также содержать добавки, такие как, например, наполнители, связующие вещества, смачивающими вещества, скользящие вещества, стабилизаторы, консерванты, эмульгаторы, а также растворители или солюбилизаторы или средства для обеспечения депо-эффекта. Последнее означает, что слитые белки могут быть включены в системы доставки с замедленными высвобождением или системы адресной доставки, такие как липосомы и микрокапсулы.
[00131] Составы можно стерилизовать с помощью ряда способов, включая фильтрование через удерживающий бактерии фильтр или путем включения стерилизующих средств в форме стерильных твердых композиций, которые можно растворять или диспергировать в стерильной воде или другой стерильной среде непосредственно перед применением.
[00132] Молекула антитела подходит и может применяться для лечения или предупреждения заболевания. Соответственно, согласно некоторым вариантам осуществления молекулу антитела в соответствии с настоящим изобретением можно применять в способе лечения и/или предупреждения медицинского состояния, такого как нарушение или заболевание. Аналогичным образом, молекулу антитела по настоящему изобретению можно применять в лечении заболевания. Подлежащее лечению или предупреждению заболевание может представлять собой пролиферативное заболевание. Такое пролиферативное заболевание может представлять собой предпочтительно опухоль или злокачественную опухоль. Ввиду способности молекулы антитела по настоящему изобретению связывать PMSA, такую молекулу антитела можно применять для лечения злокачественной опухоли, которая состоит из клеток, экспрессирующих PSMA, таких как клетки первичного и метастатического рака предстательной железы, и новообразованных сосудов при широком спектре злокачественных новообразований, таких как классическая (светлоклеточная) карцинома почки, переходно-клеточная карцинома мочевого пузыря, эмбриональная карцинома яичка, аденокарцинома толстой кишки, нейроэндокринная карцинома, мультиформная глиобластома, злокачественная меланома, карцинома протоков поджелудочной железы, немелкоклеточная карцинома легкого, саркома мягких тканей и карцинома молочной железы (в этом контексте см. Chang SS et al. Five different anti prostate specific membrane antigen (PSMA) antibodies confirm PSMA expression in tumor associated neovasculature. Cancer Res 1999; 59:3192). Таким образом, молекулу антитела по настоящему изобретению согласно одному аспекту можно применять в лечении солидной злокачественной опухоли, такой как рак предстательной железы, рак толстой кишки, рак молочной железы, рак поджелудочной железы или глиобластома. Однако в связи с неожиданным обнаружением того, что антитело по настоящему изобретению также связывается с клетками плоского эпителия, согласно другому аспекту молекулу антитела по настоящему изобретению предпочтительно можно также применять в лечении плоскоклеточной карциномы разного происхождения, включая без ограничения карциному кожи, головы и шеи, пищевода, легкого и шейки матки.
[00133] Субъектом, подлежащим лечению слитым белком, может быть человек или отличное от человека животное. Такое животное предпочтительно представляет собой млекопитающее, к примеру, человека, свинью, крупный рогатый скот, кролика, мышь, крысу, примата, козу, овцу, курицу или лошадь, наиболее предпочтительно человека.
[00134] Молекулу антитела по настоящему изобретению можно также применять в диагностировании заболевания, такого как описанное в настоящем документе заболевание. Для этой цели молекулу антитела можно метить подходящей выявляемой сигнальной меткой. Такая меченная молекула антитела может обеспечить возможность выявления или количественной оценки в образце или у субъекта уровня PSMA или злокачественной опухоли, такой как рак предстательной железы, рак толстой кишки, рак молочной железы, рак поджелудочной железы или глиобластома, плоскоклеточная карцинома, а также вышеперечисленные формы плоскоклеточной карциномы разного происхождения. Если предполагается in vivo применение, указанная выявляемая сигнальная метка предпочтительно является выявляемой in vivo.
[00135] Меченую молекулу антитела можно применять в рамках методики иммунной визуализации. Следовательно, может быть выбрана выявляемая сигнальная метка, к примеру, исходя из методики иммунной визуализации, используемой для диагностики, например, гамма-излучающий радионуклид (или гамма-излучатель) в случае методики визуализации с использованием гамма-камеры/SPECT, металл или излучатель позитронов в случае методик визуализации, представляющих собой МРТ или ПЭТ соответственно. В этом отношении, одна или несколько выявляемых сигнальных меток по настоящему раскрытию включают средства, визуализируемые с помощью гамма-камеры, ПЭТ-визуализируемые средства и МРТ-визуализируемые средства, такие как радионуклиды, флуорофоры, флуорогенные вещества, хромофоры, хромогены, фосфоресцирующие вещества, хемилюминесцентные вещества и биолюминесцентные вещества.
[00136] Подходящая выявляемая сигнальная метка может представлять собой радионуклид. Указанный радионуклид может быть выбран из группы, состоящей из 3H, 14C, 35S, 99Tc, 1231, 1251, 131I, mIn, 97Ru, 67Ga, 68Ga, 72As, 89Zr и 201T1.
[00137] Подходящая выявляемая сигнальная метка также может представлять собой флуорофор или флуорогенное вещество. Указанные флуорофор или флуорогенное вещество могут быть выбраны из группы, состоящей из флуоресцеина, родамина, дансила, фикоэритрина, фикоцианина, аллофикоцианина, o-фтальдегида, флуорескамина, производного флуоресцеина, красителей Oregon Green, Rhodamine Green, родол зеленый или техасский красный.
[00138] Меченная молекула антитела может быть связана с выявляемой сигнальной меткой либо прямо, либо опосредовано. Например, молекула антитела может быть связана с выявляемой сигнальной меткой либо прямо (например, через тирозиновые остатки молекулы антитела), либо опосредовано (например, через линкер, в качестве металлохелатора). Согласно некоторым другим вариантам осуществления молекула антитела может быть связана с молекулой, которая способна связываться (либо in vitro, либо in vivo) с выявляемой сигнальной меткой в определенный момент времени и в определенном месте применения.
[00139] Выявляемая сигнальная метка может быть связана с молекулой антитела через один или несколько остатков диэтилентриаминпентауксусной кислоты (DTPA), которые связаны с молекулой антитела.
[00140] Также в настоящем изобретении предусмотрен in vitro способ выявления или диагностирования заболевания, определенного в настоящем документе. Такой способ может предусматривать приведение полученного от субъекта образца в контакт с предпочтительно меченной молекулой антитела по настоящему изобретению. Образец может представлять собой образец крови, мочи или спинномозговой жидкости, но может предпочтительно представлять собой образец ткани или биоптат. Подлежащее выявлению или диагностированию заболевание предпочтительно представляет собой рак предстательной железы, рак толстой кишки, рак молочной железы, рак поджелудочной железы, глиобластому или плоскоклеточную карциному, наиболее предпочтительно плоскоклеточную карциному.
[00141] Настоящее изобретение дополнительно характеризуется следующими пунктами.
[00142] Пункт 1. Молекула антитела или ее антигенсвязывающий фрагмент, способные к связыванию с простатическим специфическим мембранным антигеном (PSMA) человека, содержащие: (i) вариабельный домен тяжелой цепи, содержащий CDRH1-область, изложенную в SEQ ID NO: 03 (GFTFSDFYMY), CDRH2-область, изложенную в SEQ ID NO: 04 (TISDGGGYTSYPDSVKG), и CDRH3-область, изложенную в SEQ ID NO: 05 (GLWLRDALDY), или содержащий последовательности CDRH1, CDRH2 или CDRH3, характеризующиеся по меньшей мере 75% идентичностью последовательности или по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 03, SEQ ID NO: 04 или SEQ ID NO: 05; и (ii) вариабельный домен легкой цепи, содержащий CDRL1-область, изложенную в SEQ ID NO: 06 (SASSSISSNYLH), CDRL2-область, изложенную в SEQ ID NO: 07 (RTSNLAS), и CDRL3-область, изложенную в SEQ ID NO: 08 (QQGSYIPFT), или содержащий последовательности CDRL1, CDRL2 или CDRL3, характеризующиеся по меньшей мере 75% идентичностью последовательности или по меньшей мере 80% идентичностью последовательности с SEQ ID NO: 06, SEQ ID NO: 07 или SEQ ID NO: 08.
[00143] Пункт 2. Молекула антитела или ее антигенсвязывающий фрагмент по пункту 1, где вариабельная область тяжелой цепи содержит аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с аминокислотной последовательностью, изложенной в SEQ ID NO: 01 или 09.
[00144] Пункт 3. Молекула антитела или ее антигенсвязывающий фрагмент по пункту 1 или 2, где вариабельная область легкой цепи содержит аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с аминокислотной последовательностью, изложенной в SEQ ID NO: 02 или 10.
[00145] Пункт 4. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, где антитело выбрано из группы, состоящей из scFv, одновалентного антитела без шарнирной области, миниантитела, Fab-фрагмента, Fab’-фрагмента, F(ab’)2-фрагмента и полного антитела.
[00146] Пункт 5. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, содержащие константный домен IgG человека.
[00147] Пункт 6. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, содержащие CH1-домен.
[00148] Пункт 7. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, содержащие Fc-область.
[00149] Пункт 8. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, где антитело выполняет эффекторную функцию в виде антителозависимой клеточноопосредованной цитотоксичности (ADCC).
[00150] Пункт 9. Молекула антитела или ее антигенсвязывающий фрагмент по пункту 8, характеризующиеся повышенной аффинностью к рецептору FcγRIIIa или усиленной эффекторной функцией в виде ADCC, по сравнению с исходным антителом.
[00151] Пункт 10. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, содержащие тяжелую цепь и легкую цепь и по меньшей мере одну аминокислотную замену в константной области, по сравнению с исходным антителом, где указанная по меньшей мере одна аминокислотная замена предусматривает аминокислотные замены S239D и I332E, где нумерация положений указана в соответствии с EU-индексом.
[00152] Пункт 11. Молекула антитела или ее антигенсвязывающий фрагмент, способные к связыванию с PSMA человека, которые способны конкурировать за связывание с PSMA человека с молекулой антитела или ее антигенсвязывающим фрагментом по пункту 1.
[00153] Пункт 12. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, где молекула антитела или ее антигенсвязывающий фрагмент не конкурируют за связывание с PSMA человека с J591.
[00154] Пункт 13. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, где молекула антитела или ее антигенсвязывающий фрагмент характеризуются сниженной степенью индукции антигенного сдвига при связывании с PSMA по сравнению с J591.
[00155] Пункт 14. Молекула антитела или ее антигенсвязывающий фрагмент по пункту 13, где антигенный сдвиг индуцирован в трансфицированных PMSA клетках Sp2/0.
[00156] Пункт 15. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, где молекула антитела или ее антигенсвязывающий фрагмент дополнительно связываются с клетками плоскоклеточной карциномы (SCC).
[00157] Пункт 16. Молекула биспецифического антитела, содержащая (i) вариабельную область, содержащую вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, как определено в любом из предыдущих пунктов, где указанная вариабельная область содержит первый сайт связывания, способный к связыванию с простатическим специфическим мембранным антигеном (PSMA) человека; и (ii) вариабельную область тяжелой цепи и вариабельную область легкой цепи молекулы антитела, содержащие второй сайт связывания.
[00158] Пункт 17. Молекула антитела по пункту 16, где первый сайт связывания и второй сайт связывания связываются с разными партнерами по связыванию.
[00159] Пункт 18. Молекула антитела по пункту 16 или 17, где первый или второй сайты связывания связываются с молекулой специфического рецептора T-клетки или естественной клетки-киллера (NK).
[00160] Пункт 19. Молекула антитела по пункту 16 или 18, где молекула специфического рецептора T-клетки или NK-клетки представляет собой одно из CD3, T-клеточного рецептора (TCR), CD28, CD16, NKG2D, Ox40, 4-1 BB, CD2, CD5, PD-1 и CD95.
[00161] Пункт 20. Молекула антитела по пункту 19, где TCR представляет собой TCR (альфа/бета) или TCR (гамма/дельта).
[00162] Пункт 21. Молекула антитела по пункту 19, где молекула специфического рецептора T-клетки или NK-клетки представляет собой CD3.
[00163] Пункт 22. Молекула антитела по пункту 21, где вариабельная область тяжелой цепи и вариабельная область легкой цепи молекулы антитела, содержащие второй сайт связывания, представляют собой вариабельную область тяжелой цепи и вариабельную область легкой цепи OKT3 или UCHT1.
[00164] Пункт 23. Молекула антитела по любому из пунктов 15-21, где (i) первый сайт связывания содержится в пределах Fab-фрагмента, а второй сайт связывания содержится в пределах scFv-фрагмента; или (ii) первый сайт связывания содержится в пределах одноцепочечного Fv-фрагмента, а второй сайт связывания содержится в пределах Fab-фрагмента.
[00165] Пункт 24. Молекула антитела по пункту 23, где Fab-фрагмент и одноцепочечный Fv-фрагмент соединены посредством CH2-домена и/или CH3-домена.
[00166] Пункт 25. Молекула антитела по пункту 24, где по меньшей мере один аминокислотный остаток CH2-домена, который способен опосредовать связывание с Fc-рецепторами, отсутствует или подвержен мутации.
[00167] Пункт 26. Молекула антитела по пункту 24 или 25, где один или несколько аминокислотных остатков в положениях 226, 228 и 229 в последовательности отсутствуют или подвержены мутации (нумерация положений в последовательности указана в соответствии с EU-индексом).
[00168] Пункт 27. Молекула антитела по любому из пунктов 24-26, где Fab-фрагмент соединен с CH2-доменом посредством CH1- и VH-доменов тяжелой цепи Fab-фрагмента или посредством CL- и VL-доменов легкой цепи Fab-фрагмента.
[00169] Пункт 28. Молекула антитела по пункту 27, где домены тяжелой цепи Fab-фрагмента или домены легкой цепи Fab-фрагмента расположены на N-конце полипептидной цепи молекулы антитела.
[00170] Пункт 29. Молекула антитела по любому из предыдущих пунктов, где Fab-фрагмент содержит шарнирную область.
[00171] Пункт 30. Молекула антитела по любому из пунктов 24-29, где по меньшей мере один аминокислотный остаток CH2-домена, который способен опосредовать связывание с Fc-рецепторами, отсутствующий или подверженный мутации, выбран из группы, состоящей из положений 230, 231, 232, 233, 234, 235, 236, 237, 238, 265, 297, 327 и 330 в последовательности (нумерация положений в последовательности указана в соответствии с EU-индексом).
[00172] Пункт 31. Молекула антитела по любому из пунктов 24-30, где цистеин в одном или обоих положениях 226 и 229 замещен другой аминокислотой.
[00173] Пункт 32. Молекула антитела по любому из пунктов 24-31, содержащая Fab-фрагмент, CH2-домен и scFv-фрагмент, где Fab-фрагмент содержит шарнирную область.
[00174] Пункт 33. Молекула антитела по пункту 32, где Fab-фрагмент представляет собой Fab-фрагмент гуманизированного антитела 10B3 и/или где scFv-фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи из антитела OKT3.
[00175] Пункт 34. Молекула антитела по пункту 33, где тяжелая цепь молекулы антитела содержит последовательность, как изложено в SEQ ID NO: 12, и/или где легкая цепь молекулы антитела содержит последовательность, изложенную в SEQ ID NO: 13.
[00176] Пункт 35. Молекула антитела по любому из пунктов 24-31, содержащая Fab-фрагмент, CH2-домен, CH3-домен и scFv-фрагмент, где Fab-фрагмент содержит шарнирную область.
[00177] Пункт 36. Молекула антитела по пункту 35, где Fab-фрагмент представляет собой Fab-фрагмент гуманизированного антитела 10B3 и/или где scFv-фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи из гуманизированного антитела UCHT1.
[00178] Пункт 37. Молекула антитела по пункту 36, где тяжелая цепь молекулы антитела содержит последовательность, как изложено в SEQ ID NO: 11, и/или где легкая цепь молекулы антитела содержит последовательность, изложенную в SEQ ID NO: 13.
[00179] Пункт 38. Молекула антитела по любому из пунктов 35-37, где молекула антитела представляет собой молекулу тетрамерного антитела.
[00180] Пункт 39. Молекула антитела по любому из пунктов 16-23, где молекула антитела представляет собой биспецифический тандемный одноцепочечный Fv, биспецифический Fab2 или биспецифическое диатело.
[00181] Пункт 40. Фармацевтическая композиция, содержащая молекулу антитела или ее антигенсвязывающий фрагмент, как определено в любом из предыдущих пунктов.
[00182] Пункт 41. Молекула антитела или ее антигенсвязывающий фрагмент, как определено в любом из пунктов 1-39, для применения в диагностике или лечении заболевания.
[00183] Пункт 42. Молекула антитела или ее антигенсвязывающий фрагмент для применения по пункту 41, где заболевание представляет собой пролиферативное заболевание.
[00184] Пункт 43. Молекула антитела или ее антигенсвязывающий фрагмент для применения по пункту 42, где пролиферативное заболевание представляет собой злокачественную опухоль.
[00185] Пункт 44. Молекула антитела или ее антигенсвязывающий фрагмент для применения по пункту 43, где злокачественная опухоль представляет собой рак предстательной железы, рак толстой и прямой кишки, рак желудка, карциному легкого, остеосаркому, рак молочной железы, рак поджелудочной железы или глиобластому.
[00186] Пункт 45. Молекула антитела или ее антигенсвязывающий фрагмент для применения по пункту 43, где злокачественная опухоль представляет собой плоскоклеточную карциному.
[00187] Пункт 46. In vitro способ диагностирования заболевания, предусматривающий приведение полученного от субъекта образца в контакт с молекулой антитела или ее антигенсвязывающим фрагментом, как определено в любом из пунктов 1-39.
[00188] Пункт 47. In vitro способ по пункту 46, в котором образец представляет собой образец ткани или биоптат.
[00189] Пункт 48. In vitro способ по пункту 46 или 47, в котором заболевание представляет собой злокачественную опухоль, предпочтительно рак предстательной железы, рак толстой и прямой кишки, рак желудка, карциному легкого, остеосаркому, рак молочной железы, рак поджелудочной железы, глиобластому или плоскоклеточную карциному.
[00190] Пункт 49. Молекула нуклеиновой кислоты, кодирующая молекулу антитела или ее антигенсвязывающий фрагмент, как определено в любом из пунктов 1-39.
[00191] Пункт 50. Вектор, содержащий молекулу нуклеиновой кислоты по пункту 49.
[00192] Пункт 51. Клетка-хозяин, содержащая молекулу нуклеиновой кислоты по пункту 49 или вектор по пункту 50.
[00193] Пункт 52. Способ получения молекулы антитела или ее антигенсвязывающего фрагмента по любому из пунктов 1-39, предусматривающий обеспечение экспрессии нуклеиновой кислоты, кодирующей молекулу антитела, в условиях, обеспечивающих экспрессию нуклеиновой кислоты.
[00194] Пункт 53. Способ по пункту 52, в котором экспрессию молекулы антитела или ее антигенсвязывающего фрагмента обеспечивают в клетке-хозяине или бесклеточной системе.
Краткое описание перечня последовательностей
[00195] SEQ ID NO: 01: вариабельный домен тяжелой цепи антитела 10B3 мыши (аминокислотная последовательность).
[00196] SEQ ID NO: 02: вариабельный домен легкой цепи антитела 10B3 мыши (аминокислотная последовательность).
[00197] SEQ ID NO: 03: CDRH1 антитела 10B3 мыши (аминокислотная последовательность).
[00198] SEQ ID NO: 04: CDRH2 антитела 10B3 мыши (аминокислотная последовательность).
[00199] SEQ ID NO: 05: CDRH3 антитела 10B3 мыши (аминокислотная последовательность).
[00200] SEQ ID NO: 06: CDRL1 антитела 10B3 мыши (аминокислотная последовательность).
[00201] SEQ ID NO: 07: CDRL2 антитела 10B3 мыши (аминокислотная последовательность).
[00202] SEQ ID NO: 08: CDRL3 антитела 10B3 мыши (аминокислотная последовательность).
[00203] SEQ ID NO: 09: вариабельный домен тяжелой цепи гуманизированного антитела 10B3 (аминокислотная последовательность).
[00204] SEQ ID NO: 10: вариабельный домен легкой цепи гуманизированного антитела 10B3 (аминокислотная последовательность).
[00205] SEQ ID NO: 11: тяжелая цепь молекулы биспецифического антитела формата IgGsc, содержащего гуманизированное h10B3 × гуманизированное hUCHT1
[00206] SEQ ID NO: 12: тяжелая цепь молекулы биспецифического антитела формата Fabsc, содержащего гуманизированное h10B3 × OKT3 мыши
[00207] SEQ ID NO: 13: легкая каппа-цепь гуманизированного антитела 10B3
[00208] SEQ ID NO: 14: иллюстративный пример мутированной CDRH1 антитела 10B3 мыши (аминокислотная последовательность), характеризующейся по меньшей мере 75% идентичностью последовательности с SEQ ID NO: 3.
[00209] SEQ ID NO: 15: аминокислотная последовательность вариабельного домена тяжелой цепи антитела J519.
[00210] SEQ ID NO: 16: аминокислотная последовательность вариабельного домена легкой цепи антитела J519.
Экспериментальные примеры
[00211] Настоящее изобретение дополнительно проиллюстрировано следующими неограничивающими примерами.
Пример 1. Получение, идентификация и продукция антитела 10B3
[00212] Антитело 10B3 получали после иммунизации самок мышей BALB/c облученными трансфицированными PMSA клетками Sp2/0-Ag14 (клетки Sp2/0-Ag14 были получены от ATCC, где они коммерчески доступны под названием ATCC® CRL-1581™). Через четыре дня после последней иммунизации клетки селезенки сливали с трансфицированными клетками Sp2/0 и культивировали в среде для селекции HAT. Супернатанты растущих клеток гибридомы подвергали скринингу в отношении продукции антител к PSMA с помощью проточной цитометрии с применением трансфицированных PSMA и нетрансфицированных клеток Sp2/0. После субклонирования путем предельного разведения получали стабильную моноклональную линию клеток гибридомы. Для продукции антител клетки гибридомы адаптировали к улучшенной среде DMEM (Gibco,Thermo Scientific, Уолтем, Массачусетс, USA02451), дополненной 1% FCS (Biochrom GmbH, Берлин, Германия) во избежание контаминации бычьих IgG при очистке. Антитело выделяли из супернатантов клеточных культур с помощью аффинной хроматографии с использованием белка А, и изотип очищенного антитело был идентифицирован как IgG2b/каппа (набор Rapid Mouse-Monoclonal Isotyping Kit, BioAssay Works, Ижамсвилл, Мэриленд, 21754). Показанные последовательности вариабельной области тяжелой цепи (показанной в SEQ ID NO: 1 и на фиг. 10A) и вариабельной области легкой цепи (показанной в SEQ ID NO: 2 и на фиг. 10B) были определены Alvedron GmbH, Фрайбург, Германия.
Пример 2. Получение гуманизированного антитела 10B3
[00213] Антитело 10B3 было гуманизировано путем прививания CDR-областей в последовательности зародышевой линии последовательности IGKV3-20*02 вариабельной области легкой κ-цепи человека (эта последовательность внесена в базу данных IMGT/LIGM под номером доступа L37729), см. также Ichiyoshi Y., Zhou M., Casali P. A human anti-insulin IgG autoantibody apparently arises through clonal selection from an insulin-specific 'germ-line' natural antibody template. Analysis by V gene segment reassortment and site-directed mutagenesis' J. Immunol. 154(1):226-238 (1995), и последовательность IGHV3-11*06 вариабельной области тяжелой цепи (эта последовательность внесена в базу данных IMGT/LIGM под номером доступа AF064919), см. также Watson C.T., et al., Complete haplotype sequence of the human immunoglobulin heavy-chain variable, diversity, and joining genes and characterization of allelic and copy-number variation. Am. J. Hum. Genet. 92(4):530-546 (2013). В данном случае следует отметить, что IMGT/LIGM-DB представляет собой всеобъемлющую базу данных IMGT нуклеотидных последовательностей иммуноглобулинов (IG) и T-клеточных рецепторов (TR) человека и других видов позвоночных; см. Nucleic Acids Res. 2006 Jan 1;34(Database issue):D781-4.
[00214] С целью сохранения свойств связывания исходного антитела мыши в каркасную область вариабельных доменов человека вводили следующие две обратные мутации. В вариабельном домене тяжелой цепи в пределах IGHV3-11*06 зародышевой линии человека серин в положении 49 подвергали обратной мутации к аланину, который присутствует в антителе 10B3 мыши (см. также фиг. 10C, на которой аланиновый остаток в положении 49 выделен жирным шрифтом и курсивом). В вариабельном домене легкой цепи в пределах последовательности IGKV3-20*02 фенилаланин в положении 72 в последовательности в пределах последовательности зародышевой линии человека подвергали обратной мутации к тирозиновому остатку, который присутствует в этом положении в последовательности антитела 10B3 мыши (см. также фиг. 10D, на которой тирозиновый остаток в положении 72 выделен жирным шрифтом и курсивом).
Пример 3. Получение рекомбинантных молекул антител и нецелевая активация T-клеток разными антителами к PSMA×CD3
[00215] Для конструирования рекомбинантных молекул биспецифических антител вариабельные домены антител к PSMA J591 и 10B3 сливали с константными областями и вариабельными областями антител к CD3 OKT3 или UCHT1 в следующем порядке. VL-CL для обоих форматов Fabsc и IgGsc. Тяжелые цепи были сконструированы следующим образом: VH-CH1-CH2mod-scFv(OKT3/UCHT1) для формата Fabsc, VH-CH1-CH2mod-CH3-scFv(OKT3/UCHT1) для формата IgGsc (ср. также фиг. 1A и 1B). В этих молекулах антител сайт связывания PSMA представлен как Fab-фрагмент, тогда как сайт связывания CD3 представлен как scFv-фрагмент (ср. опять же фиг. 1A и 1B). Для уменьшения степени FcR-связывания, сайтов гликозилирования и образования дисульфидных связей в шарнирную область и CH2-домен формата Fabsc вводили следующие модификации (EU-индекс): C226S; C229S; E233P; L234V; L235A; ΔG236; D265G; N297Q; A327Q; A330S (в этом отношении см. также международную заявку на патент WO 2013/092001). Модификации форматов IgGsc были идентичными, за исключением двух мутаций по цистеину в положениях 226 и 229 в последовательности шарнирной области, которые отсутствуют в формате IgGsc. Конструкции клонировали в вектор экспрессии, полученный на основе pcDNA3.1 (InVitrogen, Thermo Fisher), и трансфицировали в клетки Sp2/0 путем электропорации, как описано также в международной заявке на патент WO 2013/092001. Молекулы антител очищали из супернатантов трансфицированных клеток с помощью аффинной хроматографии с использованием смолы kappaSelect (Fabsc) или смолы со связанным белком A (IGGsc). Обе смолы для аффинной хроматографии были приобретены у GE Health Care, Фрайбург, Германия.
[00216] Для определения характеристик нецелевой активации T-клеток PBMC инкубировали с указанными молекулами антител J519 и 10B3 в отсутствие и в присутствии «фоновых» клеток лимфомы SKW6.4. Через 2 дня с помощью проточной цитометрии анализировали экспрессию CD69 CD4 T-клеток. Для этой цели PMBC инкубировали с детекторными антителами к CD4 (FITC-меченный клон HP2/6), CD8 (APC-меченный клон HIT8a) и CD69 (PE-меченный клон FN50) для идентификации T-клеток, экспрессирующих маркер активации CD69. Клетки анализировали с применением FACS Canto II от BD Biosciences.
[00217] Вывод: Как показано на фиг. 2 (на которой показаны результаты для молекул биспецифических антител, содержащих вариабельные домены антитела J519), молекула антитела Fabsc, содержащая scFv-фрагмент CD3-связывающего антитела UCHT1, индуцирует активацию T-клеток в отсутствие экспрессирующих PSMA клеток (нецелевая активация T-клеток), тогда как молекула антитела IgGsc, которая содержит такой же сайт связывания мишени, т.е. PSMA, и сайт связывания эффекторного антитела (т.е. scFv-фрагмент CD3-связывающего антитела UCHT1), не индуцирует такую активацию.
Пример 4. Целевая активация T-клеток антител к PSMA×CD3 в формате Fabsc
[00218] На фиг. 3, активацию T-клеток оценивали по поглощению 3H-тимидина. На фиг. 3A нецелевая активация T-клеток показана для сравнения. На фиг. 3C продемонстрирован лизис экспрессирующих PSMA целевых клеток активированными T-клетками в анализе цитотоксичности Xelligence. Для анализа поглощения 3H-тимидина PBMC (205/лунку) высевали в трех повторах в 96-луночные планшеты и инкубировали с молекулами биспецифических антител в различных концентрациях с (фиг. 3B) облученными (100 Гр) экспрессирующими PSMA клетками 22RV1 (105/лунку), или без них (фиг. 3A). Через 72 часа клетки экспонировали в течение еще 20 часов с 3H-тимидином (0,5 мкКи/лунку) и собирали на фильтр-подложках. Включенную радиоактивную метку определяли с помощью жидкостной сцинтилляционной спектрометрии на счетчике для микропланшетов 2450 Microplate counter (Perkin Elmer).
[00219] Для анализа Xelligence 50 мкл культуральной среды добавляли в 96-луночные E-планшеты (Roche) для определения фоновых значений. Затем при плотности 40000 клеток на лунку высевали целевые клетки (22RV1). В течение следующих 20-24 часов клеткам давали прикрепиться к лункам. Затем PBMC, выделенные путем центрифугирования в градиенте плотности, добавляли к целевым клеткам. Соотношение эффектора и мишени (E:T) составляло 5:1, а концентрация биспецифического антитела составляла 1 мкг/мл. Сопротивление отслеживали каждые 15 минут в течение нескольких дней в качестве показателя жизнеспособности адгезивных целевых клеток.
[00220] Вывод: Биспецифическое антитело к PSMA×CD3, содержащее OKT3 (NP-CO), является несколько менее эффективным в опосредовании «целевой» активации T-клеток и уничтожении целевых клеток по сравнению с реагентом, содержащим UCHT1 (NP-CU).
Пример 5. Мультимеризация и агрегация разных форматов биспецифических антител
[00221] На фиг. 4A и фиг. 4B представлено сравнение молекул антител со специфичностью к FLT3×CD3 (Fabsc- по сравнению с bscc-форматом), а на фиг. 4C-F представлено сравнение молекул антител со специфичностью к PSMA×CD3 (Fabsc- по сравнению с IgGsc-форматом). Гель-фильтрацию, которую применяли для анализа характера мультимеризации, осуществляли на колонках Superdex S200.
[00222] Выводы: (i) как показано на фиг. 4B, антитела в формате bssc (см. фиг. 1C в отношении биспецифического одноцепочечного формата, применяемого в эксперименте из фиг. 4B), характеризуются заметной склонностью к образованию мультимеров и агрегатов. Эта склонность является значительно менее низкой в случае Fabsc- и IGsc-форматов (фиг. 4A, фиг. 4C-F). (ii) Среднее количество мультимеров, образуемых конструкциями Fabsc/IgGsc, выше для антител, содержащих OKT3 (фиг. 4C, 4E), по сравнению с таковыми, содержащими UCHT1 (фиг. 4D, 4F). Хотя результаты, показанные на фигурах 4C-4F, были получены с молекулами Fabsc/IgGsc, которые содержат вариабельные домены антитела J591 в качестве сайта связывания PSMA, аналогичные характеристики наблюдали для соответствующих молекул Fabsc антитела 10B3 (данные не показаны).
Пример 6. Связывание антител к PSMA J591 и 10B3 с экспрессирующими PSMA клетками
[00223] Связывание (фиг. 5A), отсутствие конкурентного связывания (фиг. 5B) и антигенный сдвиг PSMA при связывании антитела (фиг. 5C) оценивали с помощью проточной цитометрии с применением трансфицированных PSMA клеток Sp2/0. Для этой цели разные антитела к PSMA инкубировали с этими клетками в 96-луночных планшетах в течение 30-45 мин. при 4°C. Затем клетки промывали и инкубировали с PE-конъюгированным F(ab)2-фрагментом козы, специфичным к IgG мыши (фиг. 5A, 5C) (Jackson ImmunoResearch), или PE-конъюгированным антителом козы, специфичным к FC-γ-фрагменту человека (Jackson ImmunoResearch) (B). Клетки анализировали на FACSCalibur (BD Biosciences). На фиг. 5B продемонстрировано, что химерное (ch) PSMA-связывающее антитело J591, специфически выявляемое вторичным антителом козы к антителу человека, вытеснялось антителом J591 мыши (mu), но не антителом 10B3 мыши, т. е. антителом по настоящему изобретению. Для определения антигенного сдвига (фиг. 5C) экспрессирующие PSMA клетки инкубировали с указанными антителами вначале эксперимента и повторно через 24 часа и перед FACS-анализом с насыщающим количеством (10 мкг/мл) соответствующего антитела. Экспрессия PSMA необработанными клетками выступала в качестве стандарта сравнения (100% экспрессия PSMA).
[00224] Выводы: (i) связывание обоих антител, J591 и 10B3, является сопоставимым, (ii) антитела не конкурируют перекрестно друг с другом, то есть они связываются с разными эпитопами, и (iii) антигенный сдвиг, индуцированный антителом 10B3, заметно менее выражен по сравнению с таковым, характерным для антитела J591.
Пример 7. Полученные с помощью криостата срезы, окрашенные антителами к PSMA J591 и 10B3
[00225] На фиг. 6A, B, образец клеток карциномы предстательной железы окрашивали обоими антителами, а на фиг. 6C и фиг. 6D, образец плоскоклеточной карциномы окрашивали обоими антителами. В обоих экспериментах окрашивание осуществляли параллельно и с применением полимерной системы от Zytomed, Берлин, Германия (POLHRP-100). Стрелочками указаны строма опухоли (Tu) и кровеносные сосуды (Ve). Показаны репрезентативные результаты по 9 из 10 образцам рака предстательной железы и 7 из 10 образцам плоскоклеточной карциномы.
[00226] В ряде различных нормальных тканей человека (полученных от BioCat, Гейдельберг, Германия, T6234701-2) картина окрашивания для двух антител была идентичной, за исключением незначительной реактивности антитела 10B3 в отношении эпителиальных клеток кожи.
[00227] Выводы: (i) окрашивание образцов карциномы предстательной железы и нормальной ткани является сопоставимым для обоих антител, (ii) в случае образцов плоскоклеточной карциномы антитело J5191 окрашивает только клетки сосудов, тогда как антитело 10B3 окрашивает клетки сосудов (более интенсивно, чем J591) и собственно опухолевые клетки.
Пример 8. Связывание гуманизированных антител 10B3 и антител 10B3 мыши
[00228] Биспецифические антитела Fabsc со специфичностью к PSMA×CD3 (10B3×OKT3), содержащие вариабельные домены либо гуманизированного, с привитыми CDR (h10B3), либо антитела (m10B3) мыши, инкубировали с экспрессирующими PSMA клетками 22RV1 и анализировали с помощью проточной цитометрии (фиг. 7). Для этой цели клетки инкубировали с указанными антителами в 96-луночных планшетах в течение 30-45 мин. при 4°C. После инкубации клетки промывали и инкубировали с вторичным PE-конъюгированным антителом козы, специфичным к FC-γ-фрагменту человека (Jackson ImmunoResearch). Клетки анализировали на FACSCalibur (BD Biosciences).
[00229] Вывод: Связывание мышиной и гуманизированной версий 10B3 (включенных в качестве Fab-компонента вариабельных доменом 10B3 в молекуле антитела Fabsc) с экспрессирующими PSMA клетками является идентичным.
Пример 9. Связывание разных антител к PSMA×CD3 с CD3
Клетки Jurkat инкубировали с молекулами антител (Fabsc (UCHT1) = молекула Fabsc, содержащая CD3-связывающие вариабельные домены антитела UCHT1 и вариабельные домены антитела J591; IgGsc (UCHT1) = молекула IgGsc, содержащая CD3-связывающие вариабельные домены антитела UCHT1 и вариабельные домены антитела J591, Fabsc (OKT3) = молекула Fabsc, содержащая CD3-связывающие вариабельные домены антитела OKT3 и вариабельные домены антитела J591, IgGsc (OKT3) = молекула IgGsc, содержащая CD3-связывающие вариабельные домены антитела OKT3 и вариабельные домены антитела J591) в 96-луночных планшетах в течение 30-45 мин при 4°C. Затем клетки промывали и инкубировали с конъюгированным с биотином IgG козы, специфическим к F(ab')2-фрагменту человека (Jackson ImmunoResearch), и стрептавидином-PE (Life Technologies). Клетки анализировали на FACSCalibur (BD Biosciences).
[00230] Вывод: Как показано на фиг. 8, авидность к CD3 является наиболее высокой в случае Fabsc-антитела, содержащего UCHT1, а наиболее низкой - в случае такового, содержащего OKT3. В составе формата IgGsc конструкция UCHT1 утрачивает, а конструкция OKT3 приобретает некоторую степень авидности. Поскольку авидность молекул биспецифических антител к CD3 очевидно зависит исключительно от (сайта) связывания CD3, следует считать, что градация авидности молекул биспецифических антител, которые содержат сайт связывания антитела 10B3, будет такой же, как определено в данном документе с применением вариабельных доменов антитела J591 в качестве репрезентативного сайта связывания PSMA (целевой сайт связывания).
Пример 10. Цитолитическая активность, опосредуемая разными антителами к PSMA
[00231] Экспрессирующие PSMA клетки карциномы предстательной железы 22RV1 инкубировали с PBMC и указанными молекулами биспецифических антител к PSMA×CD3 (Fabsc (UCHT1) = молекула Fabsc, содержащая CD3-связывающие вариабельные домены антитела UCHT1 и вариабельные домены антитела J591; IgGsc (UCHT1) = молекула IgGsc, содержащая CD3-связывающие вариабельные домены антитела UCHT1 и вариабельные домены антитела J591, Fabsc (OKT3) = молекула Fabsc, содержащая CD3-связывающие вариабельные домены антитела OKT3 и вариабельные домены антитела J591, IgGsc (OKT3) = молекула IgGsc, содержащая CD3-связывающие вариабельные домены антитела OKT3 и вариабельные домены антитела J591) при соотношении PBMC:мишень 5:1. Жизнеспособность адгезивных целевых клеток оценивали с применением системы Xelligence, как описано в примере 4. Соотношение эффектора и мишени (E:T) составляло 5:1, а концентрация антител составляла 5 нМ.
[00232] Репрезентативные результаты одного из четырех разных экспериментов с PBMC от разных здоровых добровольцев показаны на фиг. 9.
[00233] Вывод: Градация литической активности: Fabsc (UCHT1) ≈ IgGsc (UCHT1) > Fabsc (OKT3) > IgGsc (OKT3). Поскольку литическая активность, опосредуемая молекулами биспецифических антител, зависит от (сайта) связывания CD3, следует считать, что градация литической активности, опосредуемой молекулами биспецифических антител, которые содержат сайт связывания антитела 10B3, будет такой же, как определено в данном документе с применением вариабельных доменов антитела J591 в качестве репрезентативного сайта связывания PSMA (целевой сайт связывания).
[00234] Пример 11. Терапевтический эффект IgGsc к PSMA×CD3 (CC-1) in vitro
[00235] Биспецифическое антитело IgGsc к PSMA×CD3 (h10B3×UCHT1) (CC-1) по настоящему изобретению и контрольное биспецифическое антитело (NG2×CD3) инкубировали с опухолевыми клетками (клетками 22Rv1, клетками карциномы предстательной железы человека; см. Sramkoski RM et al., In Vitro Cell Dev Biol Anim. 1999 Jul-Aug;35(7):403-9.), или без них. Активацию CD4 и CD8 T-клеток анализировали с помощью FACS с применением CD69 и CD25 в качестве маркеров клеточной поверхности через три дня после инкубации. Оба типа T-клеток подвергались активации (фиг. 15A), и наблюдали повышение уровней интерферона гамма (фиг. 15B) и пролиферации T-клеток (фиг. 15C). В анализах высвобождения хрома (спустя 20 ч, соотношение E:T 10:1) и FACS (через 72 ч, соотношение E:T 1:1) дополнительно была показана значительная степень лизиса опухолевых клеток при применении CC-1 (фиг. 15D и E). Обработка CC-1 по настоящему изобретению также обеспечивала ингибирование роста опухоли in vitro. При соотношении E:T 2:1 рост опухолевых клеток в присутствии CC-1 значительно уменьшался, согласно результатам анализа с использованием системы Xelligence (фиг. 15F, слева) и визуального изучения с применением микроскопа (фиг. 15F, справа).
[00236] Вывод: CC-1 индуцирует ограниченную опухолевыми клетками активацию T-клеток и продукцию цитокинов, что обеспечивает в результате пролиферацию T-клеток и противоопухолевую активность.
Пример 12. Терапевтический эффект IgGsc к PSMA×CD3 (CC-1) in vivo
[00237] Далее CC-1 (биспецифическое антитело IgGsc по настоящему изобретению) тестировали в отношении противоопухолевой активности in vivo в мышиной модели. Мышам NSG (NOD scid gamma, (NOD.Cg-Prkdcscid Il2rg™1Wjl/SzJ)) путем внутривенной инъекции вводили 1,5 × 106 клеток 22Rv1 (n=4/группу). Через три дня путем инъекции вводили 3 × 106 PBMC человека, и в день 3 и 5 обрабатывали 10 мкг CC-1 или PBS в качестве отрицательного контроля. Через 8 дней с применением FACS анализировали образование метастазов в легком. Существенное снижения числа образованных метастазов наблюдали в группе обработки CC-1 (фиг. 16A). Затем путем инъекции мышам NSG (n=3/группу) вводили 2 × 107 PBMC на мышь и обрабатывали «сверхтерапевтическими» дозами CC-1 по схеме 2 × 20 мкг с интервалом в 4 дня и контрольным антителом (UCHT1 в качестве положительного контроля, который неспецифически активирует T-клетки) или PBS в качестве отрицательного контроля. Высвобождение IFN-гамма и показатели веса тела анализировали в качестве маркера аутоиммунной активности (фиг. 16B). CC-1 по сравнению с UCHT1 не индуцировало продукцию IFN-гамма, и у обработанных CC-1 мышей снижения веса тела не наблюдали, в отличие от группы обработки UCHT1. В другом эксперименте мышам NOS/SCID путем инъекции вводили клетки 22Rv1 (PSMA-положительные опухолевые клетки) или клетки DU145 (PSMA-отрицательные клетки опухоли предстательной железы человека), и после развития опухолей спустя 35 дней, мышей обрабатывали меченным радиоизотопом антителом CC-1 (50 мкКи) и через 24 ч умерщвляли для измерения радиоактивности в разных органах. Существенное повышение радиоактивности можно было наблюдать только в опухолях обработанных 22Rv1 мышей (фиг. 16C). Для тестирования отличий во времени полужизни биспецифических антител формата BiTE или IgGsc по настоящему изобретению в сыворотке крови, мышам Balb/C путем инъекции вводили 50 мкг либо CC-1 (формат IgGsc), либо биспецифического антитела к PSMA×CD3 BiTE-формата. Концентрацию в сыворотке измеряли в динамике. Биспецифические антитела BiTE-формата не возможно было выявить спустя 2-4 ч после инъекции, тогда как CC-1 спустя 24 ч все еще можно было выявить в сыворотке крови (фиг. 16D). Таким образом, формат IgGsc по настоящему изобретению обеспечивает повышенную стабильность в сыворотке крови по сравнению с другими форматами биспецифических антител.
[00238] Вывод: CC-1 подавляет рост опухоли in vivo без индуцирования каких-либо неспецифических иммунных ответов. CC-1 специфически нацеливается в опухолевую ткань, в которой экспрессируется PSMA, и по сравнению с другими биспецифическими форматами характеризуется увеличенным временем полужизни в сыворотке крови.
Пример 13. Сравнение UCHT1 и OKT3
[00239] Чтобы дополнительно исследовать показанное в настоящем документе преимущество UCHT1 над OKT3 в качестве сайта связывания CD3 в биспецифическом антителе формата IgGsc (см. выше), Fab- и IgG-версии обоих моноспецифических антител тестировали в сравнении. OKT3 и UCHT1 очищали с помощью аффинной хроматографии с использованием белка А. Fab-фрагменты получали посредством расщепления пепсином с последующим восстановлением и модификацией дисульфидных связей шарнирной области, как было описано ранее (Jung et al., Target cell induced T cell activation with bi- and trispecific antibody fragments. Eur J Immunol 1991; 21,2431-2435). Fab-фрагменты очищали с помощью эксклюзионной хроматографии на колонке Superdex S200.
[00240] Затем экспрессирующие CD3 клетки Jurkat инкубировали с возрастающими концентрациями указанных антител, меченным биотином детекторным антителом (к Fab человека), а затем с PE-конъюгированным стрептавидином. Затем образцы анализировали с помощью проточной цитометрии (таблица 1). Указаны концентрации (нМ), при которых наблюдали полумаксимальное связывание.
Таблица 1. Авидность Fab- и IgG-версий антител к CD3
[00241] Вывод:
[00242] OKT3 утрачивает некоторую степень авидности при применении в качестве одновалентного Fab-фрагмента по сравнению с бивалентной интактной молекулой IgG. В отличие от этого, авидность UCHT1-фрагмента не меняется. Не вдаваясь в теорию, это указывает на одновалентное связывание интактного антитела UCHT1, авидность которого является сопоставимой с таковой бивалентно связывающегося антитела OKT3. Заметное отличие в связывании одновалентных Fab-фрагментов в пользу UCHT1 позволяет объяснить соответствующее отличие в связывании двух антител в составе формата Fabsc (фиг. 8, пример 9). В этом формате антитела присутствуют в качестве моновалентной одноцепочечной молекулы, присоединенной к C-концу антитела против клетки-мишени (фиг. 1). Одновалентное связывание UCHT1 также позволяет объяснить, почему это антитело утрачивает, тогда как OKT3 приобретает некоторую степень авидности, когда присутствует в составе IgGsc, а не в составе формата Fabsc (фиг. 1 и фиг. 8, пример 9).
[00243] Хотя в IgG-формате авидность содержащей UCHT1 молекулы (IgGsc-UCHT1) сопоставима, или даже несколько ниже таковой IgGsc-OKT3, активность IgGsc-UCHT1 в отношении опухолевых клеток является, что неожиданно, более высокой (фиг. 9, пример 10).
[00244] Вкратце, IgGsc-UCHT1 представляет собой формат с оптимизированными свойствами, объединяющими низкую степень нецелевой активации (фиг. 2, пример 3) с оптимальной литической активностью в отношении опухолевых клеток (фиг. 9, пример 10).
Пример 14. UCHT1 является эффективным в других биспецифических антителах формата IgGsc
[00245] Предпочтительное применение UCHT1 в качестве антитела к CD3 в формате антитела на основе IgGsc дополнительно тестировали для других антител, специфических не к PSMA. Облученные клетки меланомы M21, экспрессирующие ганглиозид GD2, а также O-ацетилированную форму этого антигена (oaGD2), инкубировали с мононуклеарными клетками периферической крови (PBMC), выделенными из периферической крови здоровых доноров-людей, и биспецифическими антителами IgGsc с указанными типами специфичности. Исходные моноспецифические антитела, применяемые в составе этих конструкций, представляли собой hu14.18 (к GD2), 8B6 (к oaGD2), J591 (к PSMA) и UCHT1 (к CD3), соответственно. Через 3 дня оценивали активацию T-клеток с применением анализа включения 3H-тимидина. Кроме того, клетки M21 инкубировали с PBMC и указанными биспецифическими антителами IgGsc (50 нМ). Затем с применением системы Xelligence отслеживали рост опухолевых клеток. Биспецифическое антитело IgGsc с другой целевой специфичностью (MOPC) применяли в качестве контроля.
[00246] Вывод:
[00247] В присутствии опухолевых клеток, экспрессирующих GD2 и oaGD2, и биспецифических антител, нацеленных на такие антигены, наблюдали эффективную активацию T-клеток в популяции PBMC. Контрольное антитело, нацеленное на PSMA, было неэффективным, поскольку M21 не экспрессируют PSMA (фиг. 17A).
[00248] Биспецифические антитела IgGsc, направленные на GD2 или oaGD2 в качестве целевой специфичности и на CD3 в качестве эффекторной специфичности, эффективно уничтожают опухолевые клетки, экспрессирующие эти антигены (фиг. 17B). Применяемые специфические антитела к GD2 и CD3 перечислены в подписи к фиг. 17A.
[00249] Изобретение, описанное в настоящем документе в иллюстративных целях, может быть подходящим образом реализовано на практике при отсутствии какого-либо элемента или элементов, ограничения или ограничений, конкретно не раскрытых в настоящем документе. Таким образом, например, термины «предусматривающий», «включающий», «содержащий» и т.д., следует толковать как с возможностью расширения и без ограничения. Кроме того, используемые в настоящем документе термины и выражения применялись как описательные, а не ограничивающие термины, и не подразумевается применение таких терминов и выражений для исключения любых эквивалентов показанных и описанных признаков или их частей, а признано, что возможными являются различные модификации в пределах объема заявленного изобретения. Таким образом, следует понимать, что хотя настоящее изобретение было конкретно раскрыто с помощью иллюстративных вариантов осуществления и необязательных признаков, специалисты в данной области могут использовать модификацию и вариацию осуществленных на практике изобретений, раскрытых в настоящем документе, и что такие модификации и вариации считаются включенными в объем настоящего изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДОЙЧЕС КРЕБСФОРШУНГСЦЕНТРУМ ШТИФТУНГ ДЕС ЭФФЕНТЛИХЕН
РЕХТС ЭБЕРХАРД КАРЛЬС УНИФЕРЗИТЕТ ТЮБИНГЕН
<120> НОВОЕ PSMA-СВЯЗЫВАЮЩЕЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЯ
<130> D31545WO
<150> EP16151281.9
<151> 2016 01 14
<160> 16
<170> PatentIn version 3.5
<210> 1
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> murine 10B3 antibody heavy chain variable region
<400> 1
Gln Val Gln Leu Gln Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Phe
20 25 30
Tyr Met Tyr Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Gly Gly Gly Tyr Thr Ser Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Ile Ala Lys Asn Asn Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Lys Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Trp Leu Arg Asp Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser
115
<210> 2
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> murine 10B3 antibody light chain variable region
<400> 2
Glu Ile Val Leu Thr Gln Ser Pro Ile Thr Met Ala Ala Phe Leu Gly
1 5 10 15
Glu Arg Ile Thr Ile Thr Cys Ser Ala Ser Ser Ser Ile Ser Ser Asn
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Phe Ser Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Thr Ser Asn Leu Ala Ser Gly Val Pro Ile Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Gly Thr Met Glu
65 70 75 80
Ala Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Gly Ser Tyr Ile Pro
85 90 95
Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 3
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR-H1 of 10B3 antibody
<400> 3
Gly Phe Thr Phe Ser Asp Phe Tyr Met Tyr
1 5 10
<210> 4
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR-H2 of 10B3 antibody
<400> 4
Thr Ile Ser Asp Gly Gly Gly Tyr Thr Ser Tyr Pro Asp Ser Val Lys
1 5 10 15
Gly
<210> 5
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR-H3 of 10B3 antibody
<400> 5
Gly Leu Trp Leu Arg Asp Ala Leu Asp Tyr
1 5 10
<210> 6
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR-L1 of 10B3 antibody
<400> 6
Ser Ala Ser Ser Ser Ile Ser Ser Asn Tyr Leu His
1 5 10
<210> 7
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR-L2 of 10B3 antibody
<400> 7
Arg Thr Ser Asn Leu Ala Ser
1 5
<210> 8
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR-L3 of 10B3 antibody
<400> 8
Gln Gln Gly Ser Tyr Ile Pro Phe Thr
1 5
<210> 9
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> humanized 10B3 antibody heavy chain variable region
<400> 9
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Phe
20 25 30
Tyr Met Tyr Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Gly Gly Gly Tyr Thr Ser Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Trp Leu Arg Asp Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 10
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> humanized 10B3 antibody light chain variable region
<400> 10
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Ile Ser Ser Asn
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Arg Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ser Tyr Ile Pro
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 11
<211> 714
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain of h10B3xhUCHT1 bispecific IgGsc format antibody
<400> 11
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val
20 25 30
Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr
35 40 45
Phe Ser Asp Phe Tyr Met Tyr Trp Ile Arg Gln Ala Pro Gly Lys Gly
50 55 60
Leu Glu Trp Val Ala Thr Ile Ser Asp Gly Gly Gly Tyr Thr Ser Tyr
65 70 75 80
Pro Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
85 90 95
Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Gly Leu Trp Leu Arg Asp Ala Leu Asp Tyr
115 120 125
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
210 215 220
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
225 230 235 240
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Pro Val
245 250 255
Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Gly Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gln Leu Pro Ser Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Lys Ser Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Ser
465 470 475 480
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
485 490 495
Gln Asp Ile Arg Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys
500 505 510
Ala Pro Lys Leu Leu Ile Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val
515 520 525
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr
530 535 540
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
545 550 555 560
Gly Asn Thr Leu Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
565 570 575
Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
595 600 605
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
610 615 620
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
625 630 635 640
Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn Gln Lys Phe
645 650 655
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn Thr Ala Tyr
660 665 670
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
675 680 685
Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val Trp
690 695 700
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
705 710
<210> 12
<211> 606
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain of h10B3xOKT3 bispecific Fabsc format antibody
<400> 12
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val
20 25 30
Lys Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr
35 40 45
Phe Ser Asp Phe Tyr Met Tyr Trp Ile Arg Gln Ala Pro Gly Lys Gly
50 55 60
Leu Glu Trp Val Ala Thr Ile Ser Asp Gly Gly Gly Tyr Thr Ser Tyr
65 70 75 80
Pro Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
85 90 95
Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Gly Leu Trp Leu Arg Asp Ala Leu Asp Tyr
115 120 125
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
210 215 220
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
225 230 235 240
Ser Cys Asp Lys Thr His Thr Ser Pro Pro Ser Pro Ala Pro Pro Val
245 250 255
Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Gly Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gln Leu Pro Ser Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Ser Gly Gln Val
355 360 365
Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val
370 375 380
Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met
385 390 395 400
His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr
405 410 415
Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp
420 425 430
Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln
435 440 445
Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg
450 455 460
Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr
465 470 475 480
Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
485 490 495
Gly Gly Gly Ser Asp Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
500 505 510
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser
515 520 525
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
530 535 540
Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His
545 550 555 560
Phe Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly
565 570 575
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
580 585 590
Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn
595 600 605
<210> 13
<211> 235
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain of humanized 10B3 antibody
<400> 13
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Ile Ser Ser Asn Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
50 55 60
Pro Arg Leu Leu Ile Tyr Arg Thr Ser Asn Leu Ala Ser Gly Ile Pro
65 70 75 80
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile
85 90 95
Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly
100 105 110
Ser Tyr Ile Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
115 120 125
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
130 135 140
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
145 150 155 160
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
165 170 175
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
180 185 190
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
195 200 205
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
210 215 220
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 14
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> mutated CDR-H1 of 10B3 antibody
<400> 14
Gly Phe Ser Phe Thr Asp Phe Tyr Met Tyr
1 5 10
<210> 15
<211> 115
<212> PRT
<213> Artificial Sequence
<220>
<223> variable heavy chain domain of J519 antibody
<400> 15
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Thr
1 5 10 15
Ser Val Arg Ile Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Thr Ile His Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Thr Tyr Asn Gln Lys Phe
50 55 60
Glu Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Gly Trp Asn Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
100 105 110
Val Ser Ser
115
<210> 16
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> variable light chain domain Jof J519 antibody
<400> 16
Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Ile Cys Lys Ala Ser Gln Asp Val Gly Thr Ala
20 25 30
Val Asp Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ala Ile Thr Asn Val Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Asn Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННЫЕ АНТИ-FLT3 АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2830982C2 |
| CD3-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2826453C2 |
| АКТИВИРУЕМЫЕ ПРОТЕАЗОЙ СВЯЗЫВАЮЩИЕ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИЕ МОЛЕКУЛЫ | 2017 |
|
RU2830288C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| АГЕНТЫ ДЛЯ ЛЕЧЕНИЯ ЭКСПРЕССИРУЮЩИХ КЛАУДИН РАКОВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2798990C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CD3 | 2017 |
|
RU2790288C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА ПРОТИВ CEACAM5 И CD3 | 2020 |
|
RU2827106C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CD3 | 2017 |
|
RU2807216C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
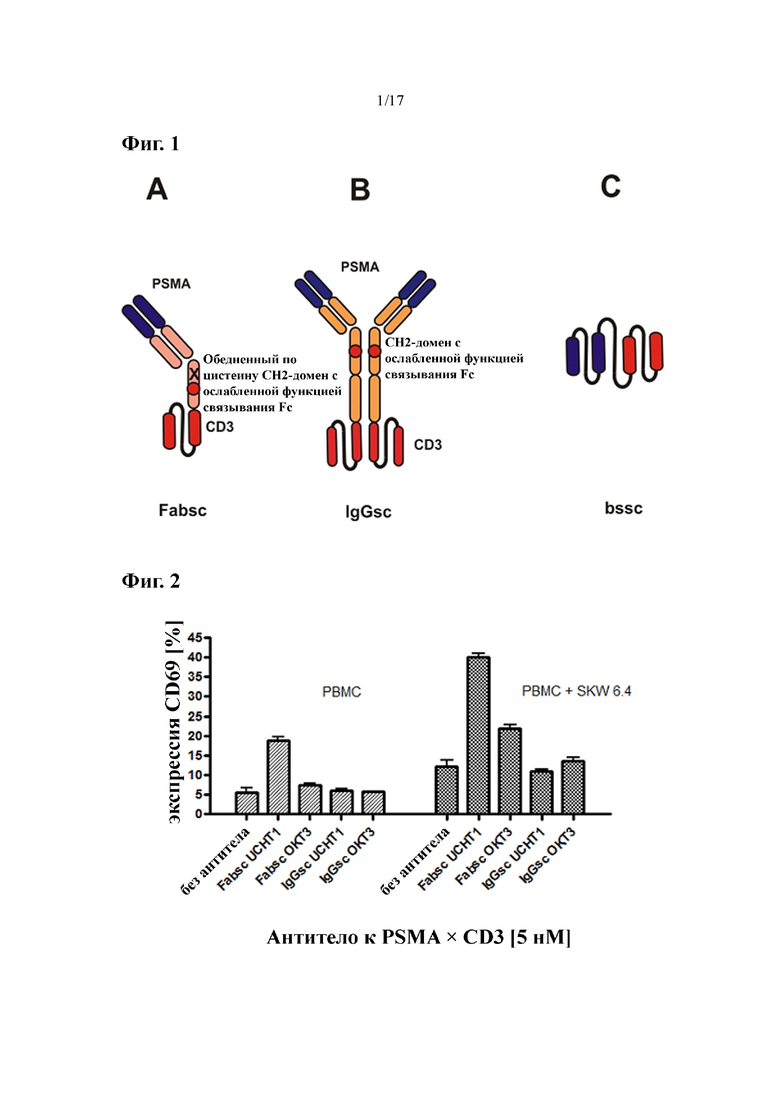
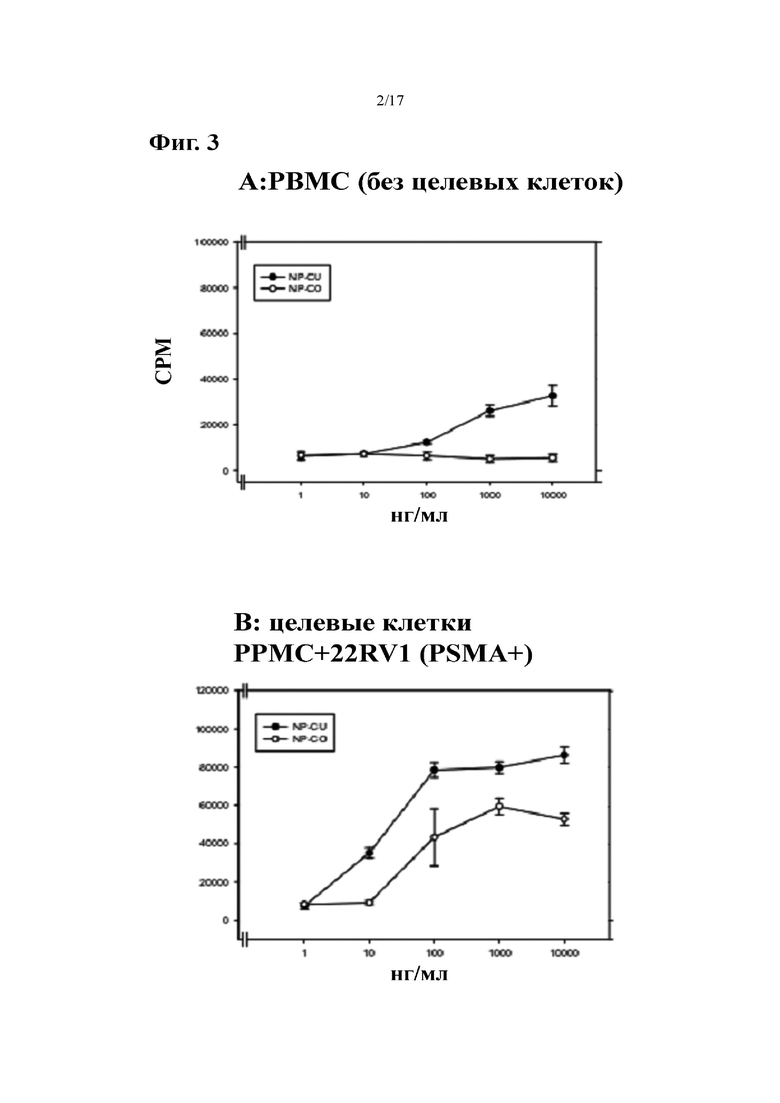
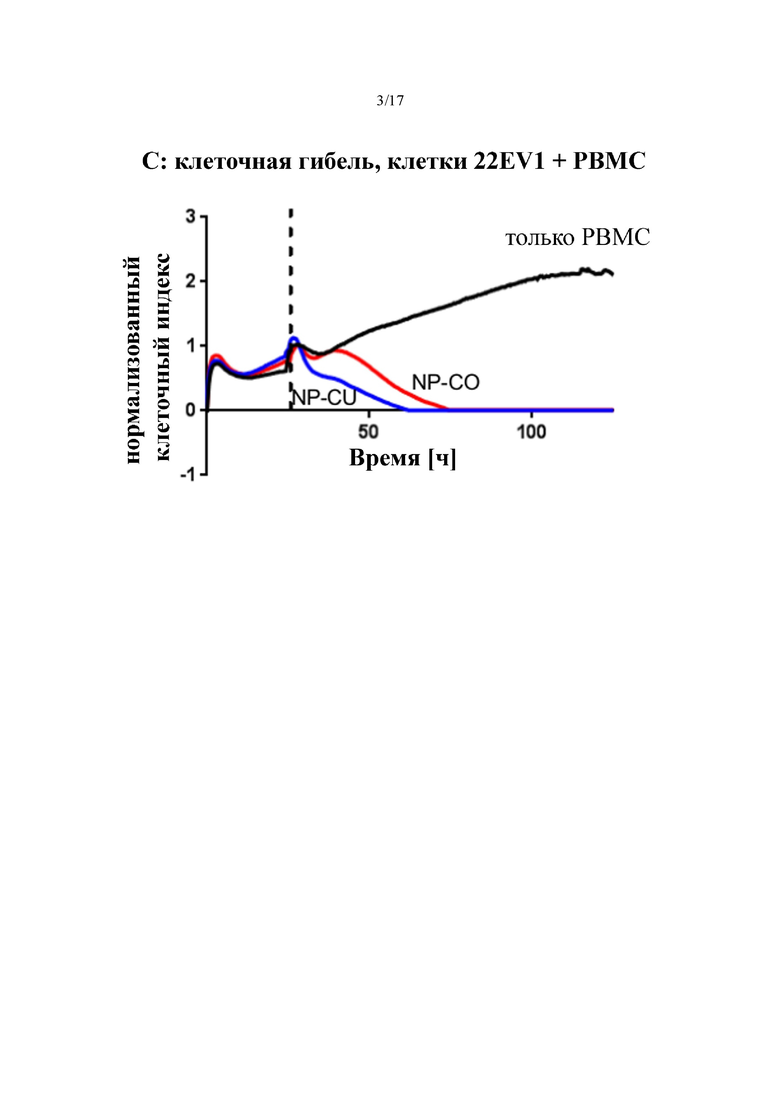
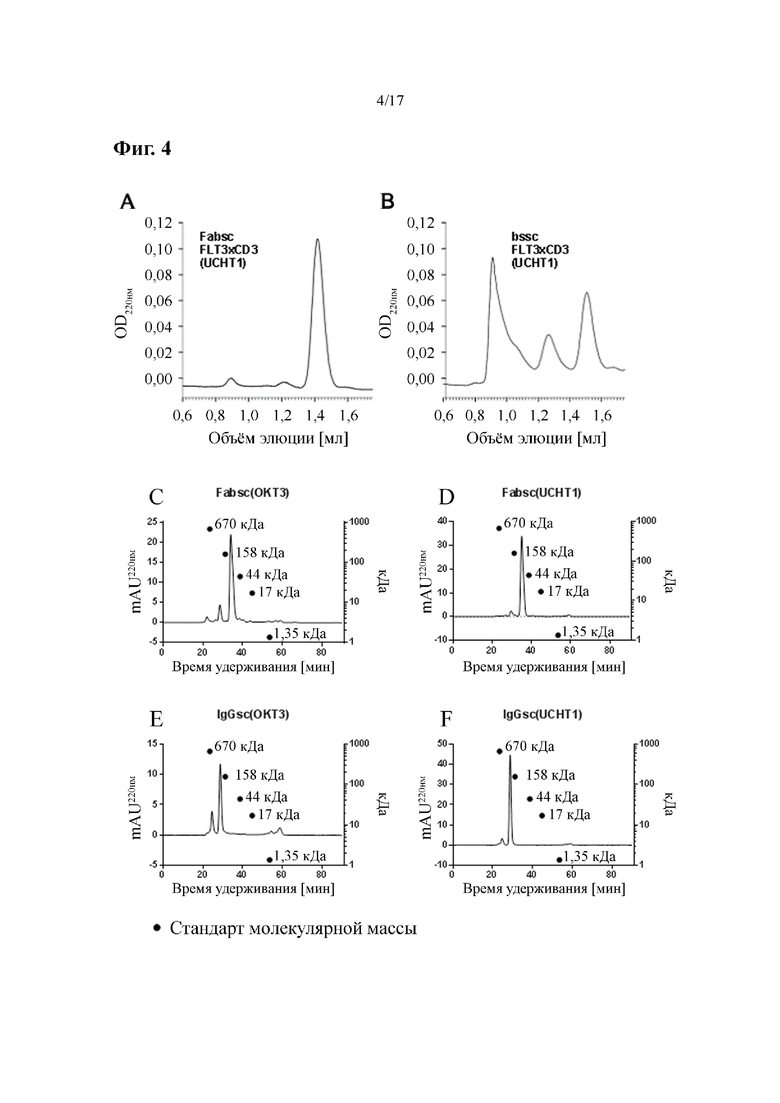
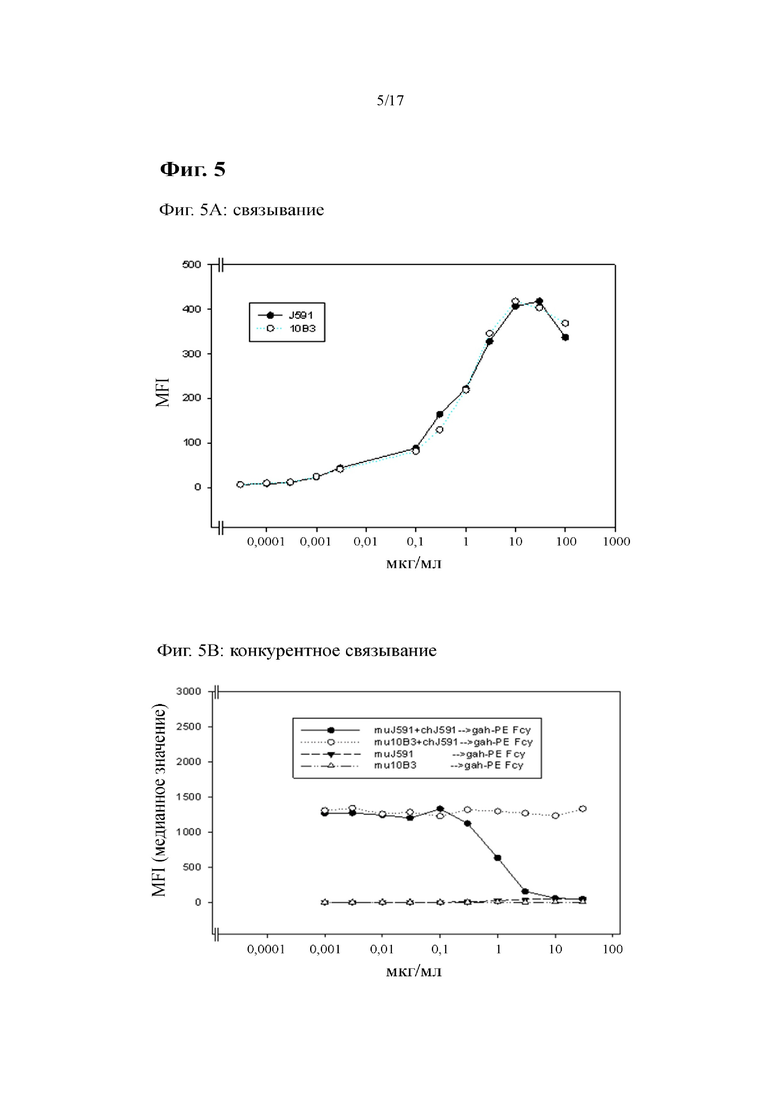
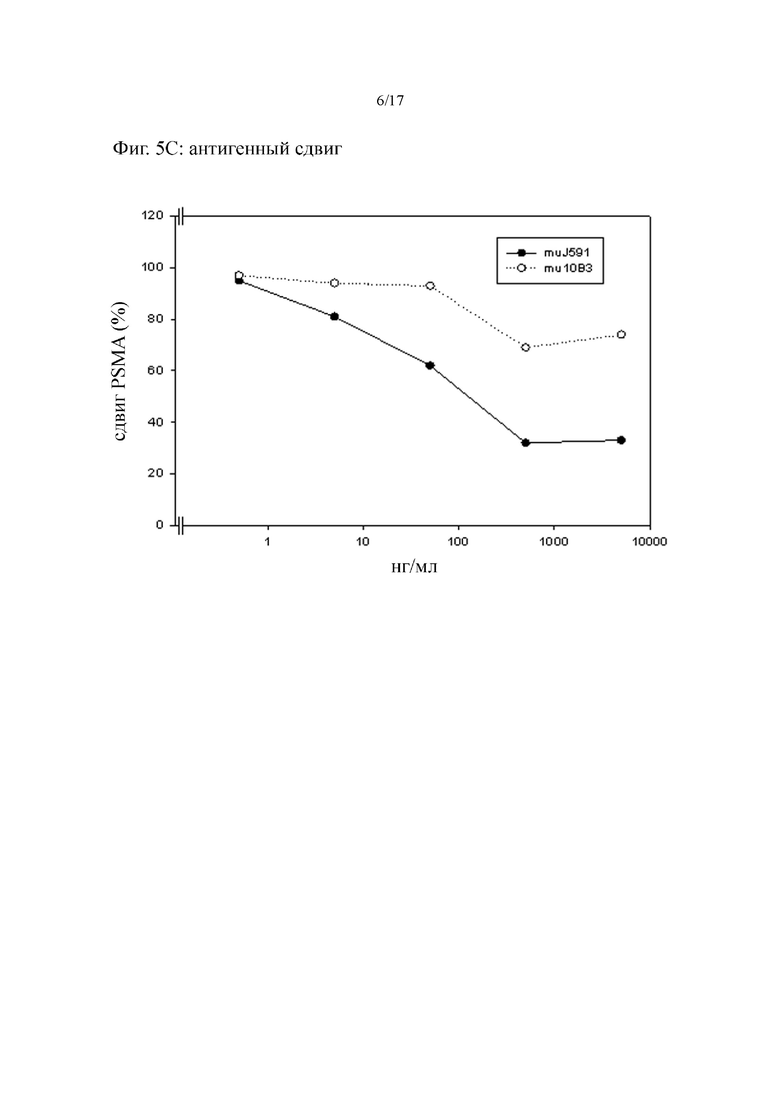
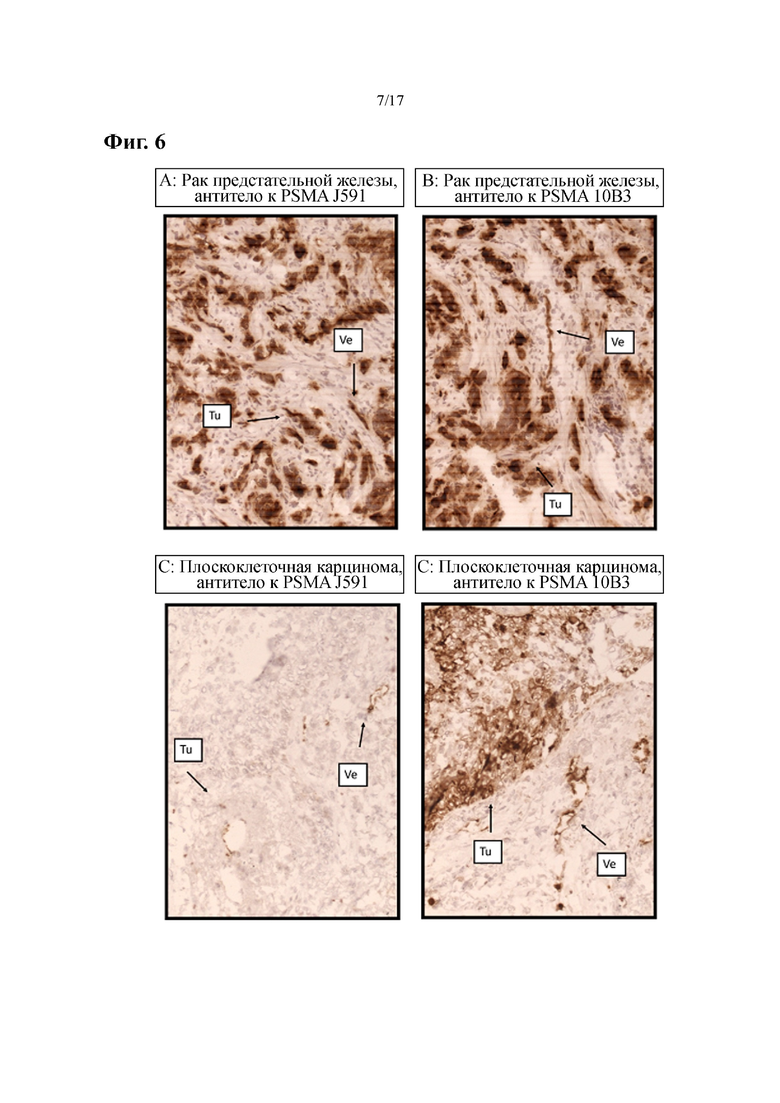
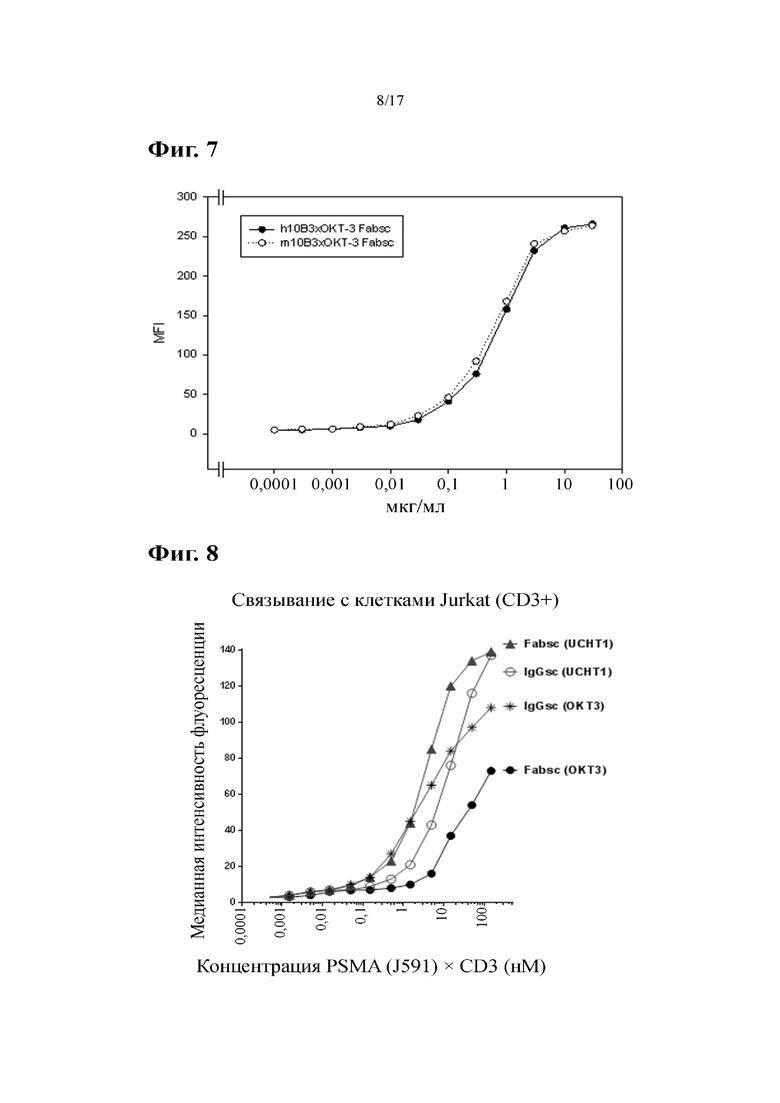
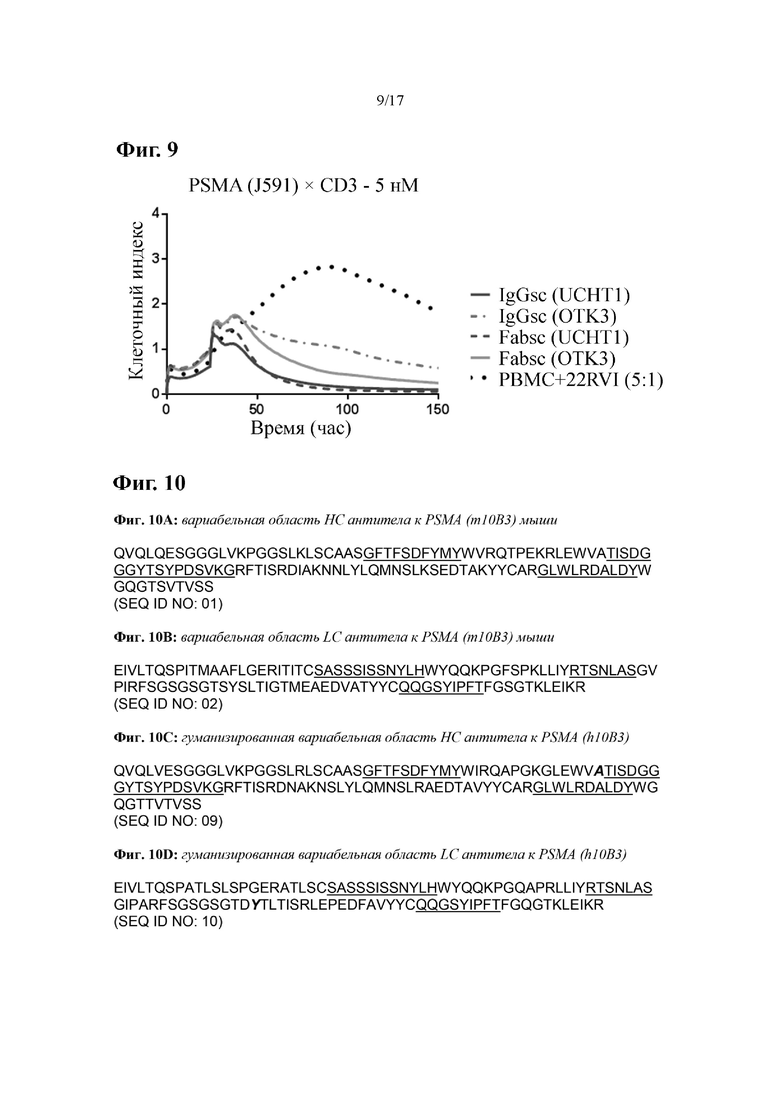




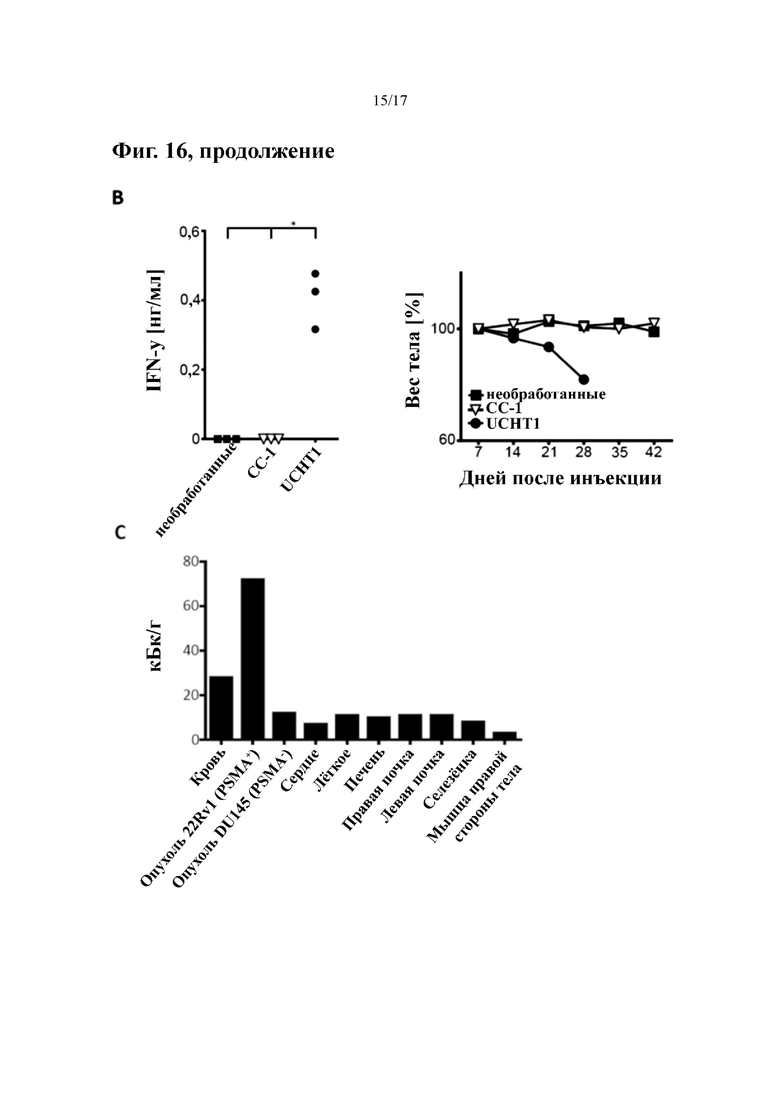
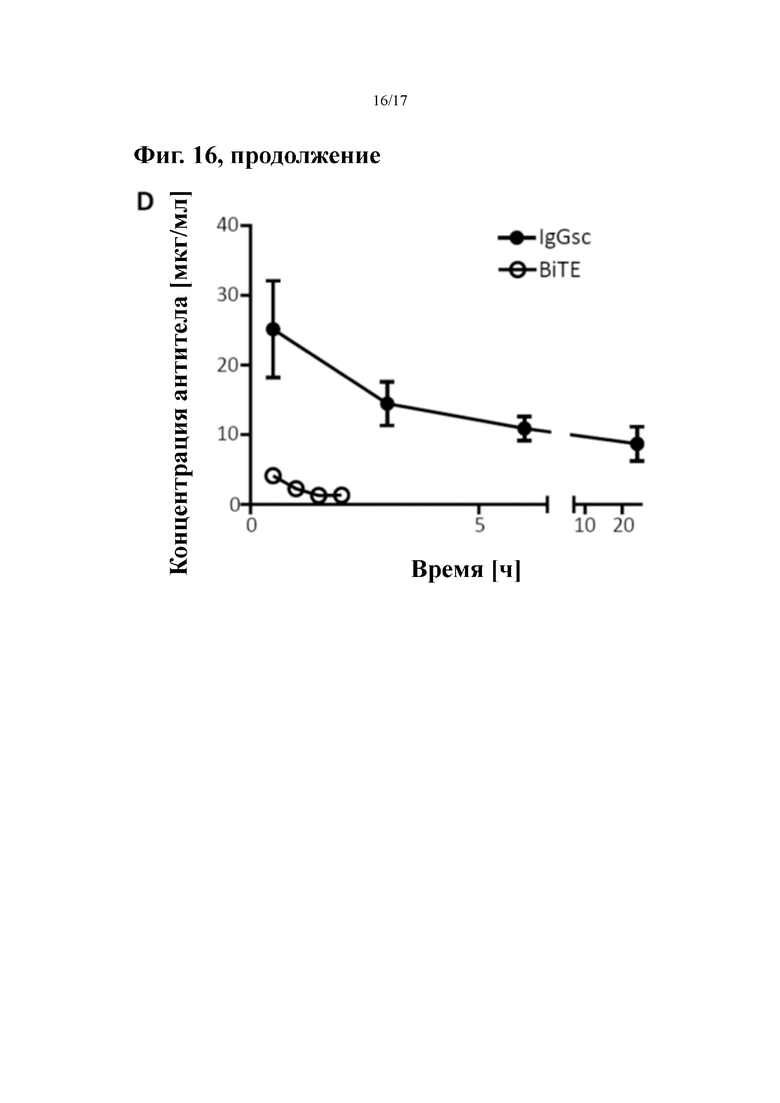
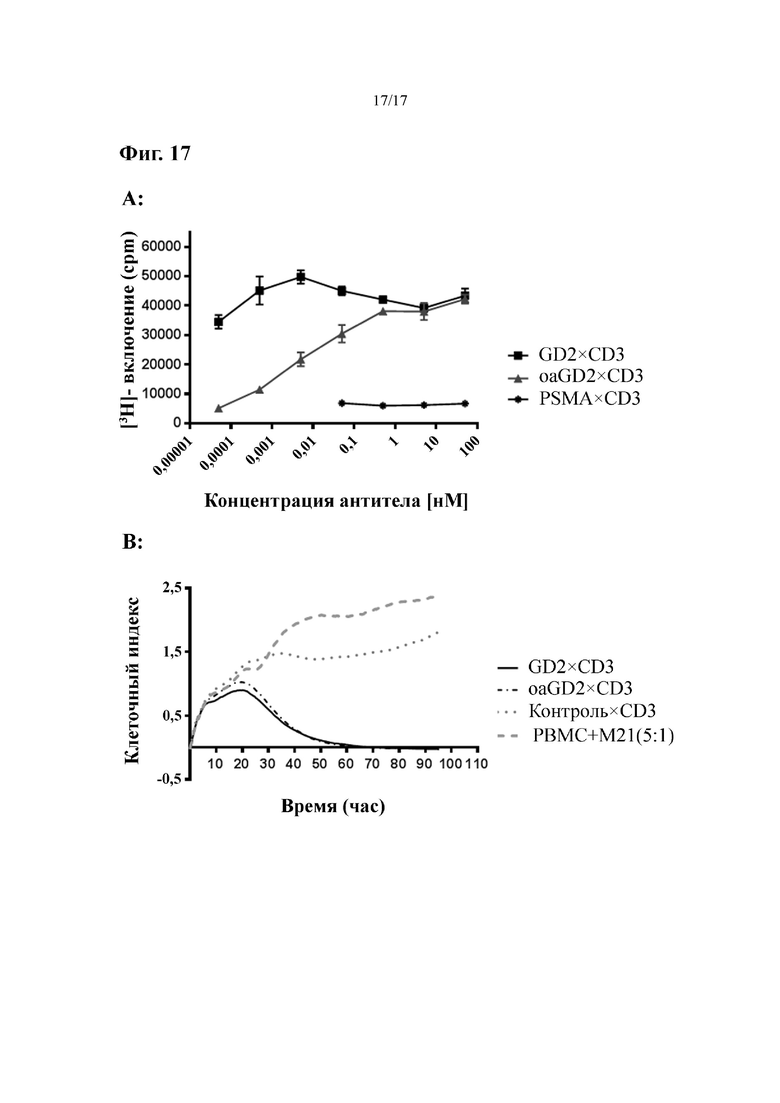
Изобретение относится к области биотехнологии. Предложено PSMA-связывающее антитело. Антитело характеризуется сниженной степенью индукции антигенного сдвига и уникальной реактивностью в отношении клеток плоскоклеточной карциномы (SCC) разного происхождения. 2 з.п. ф-лы, 17 ил., 1 табл., 14 пр.
1. Молекула антитела или ее антигенсвязывающий фрагмент, способные к связыванию с простатическим специфическим мембранным антигеном (PSMA) человека, содержащие:
(i) вариабельный домен тяжелой цепи, содержащий область CDRH1, представленную SEQ ID NO: 03 (GFTFSDFYMY), область CDRH2, представленную SEQ ID NO: 04 (TISDGGGYTSYPDSVKG), и область CDRH3, представленную SEQ ID NO: 05 (GLWLRDALDY); и
(ii) вариабельный домен легкой цепи, содержащий область CDRL1, представленную SEQ ID NO: 06 (SASSSISSNYLH), область CDRL2, представленную SEQ ID NO: 07 (RTSNLAS), и область CDRL3, представленную SEQ ID NO: 8 (QQGSYIPFT).
2. Молекула антитела или ее антигенсвязывающий фрагмент по п. 1, где вариабельная область тяжелой цепи содержит аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO: 01 или 09, или где вариабельная область легкой цепи содержит аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO: 02 или 10.
3. Молекула антитела или ее антигенсвязывающий фрагмент по любому из предыдущих пунктов, где молекула антитела или ее антигенсвязывающий фрагмент не конкурируют за связывание с PSMA человека с J591, или где молекула антитела или ее антигенсвязывающий фрагмент характеризуются сниженной степенью индукции антигенного сдвига при связывании с PSMA по сравнению с J591, или где молекула антитела или ее антигенсвязывающий фрагмент дополнительно связываются с клетками плоскоклеточной карциномы (SCC).
| WO 1998003873 A1, 29.01.1998 | |||
| WO 2015143033 A1, 24.09.2015 | |||
| ТЕХНЕЦИЙ- И РЕНИЙ-БИС(ГЕТЕРОАРИЛЬНЫЕ) КОМПЛЕКСЫ И МЕТОДЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ИНГИБИРОВАНИЯ PSMA | 2009 |
|
RU2532912C2 |
| FORTMULLER K | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| ROSS J.S | |||
| et al | |||
| Antibody-based therapeutics: Focus on prostate | |||

