Настоящее изобретение относится к связывающей молекуле, включающей по меньшей мере три специфичности связывания, где (а) первая специфичность является специфичностью к поверхностному антигену вируса гепатита В (HVB), выбранному из малого поверхностного антигена HVB, среднего поверхностного антигена HVB и большого поверхностного антигена HVB; (b) (i) вторая и третья специфичность являются специфичностями к CD3 и CD28, соответственно; или (ii) вторая и третья специфичность выбраны из специфичностей к CD16, CD56, NKp30, NKp46, 4-1BB и NKG2D; и (с) каждая специфичность связывания обеспечена одним или несколькими участками связывания, где каждый участок связывания независимо обеспечен (i) набором из шести гипервариабельных областей (CDR), где каждый набор из шести CDR состоит из первого набора из трех CDR и второго набора из трех CDR, где каждый из указанного первого и указанного второго набора включен в иммуноглобулиновый домен; или (ii) набором из трех CDR, где каждый указанный набор из трех CDR включен в иммуноглобулиновый домен.
В данном описании цитирован ряд документов, включая патентные заявки и инструкции производителей. Раскрытие этих документов, не считающееся релевантным для патентоспособности настоящего изобретения, настоящим включено посредством ссылки во всей полноте. Более конкретно, все цитированные документ включены посредством ссылки в той же степени, как если бы каждый отдельный документ был специально и индивидуально указан для включения в качестве ссылки.
Примерно 350 миллионов человек хронически инфицированы вирусом гепатита В. HVB инфекция может приводить к циррозу печени и гепатоцеллюлярной карциноме (ГЦК), частота которой составляет примерно один миллион случаев в год (Ganem et al., «Hepatitis B virus infection - natural history and clinical consequences». N Engl J Med; 350:1118-29 (2004) («Инфекция вируса гепатита В - естественная история и клинические последствия»)). Инфекции HVB в настоящее время не удается контролировать примерно у 5% взрослых пациентов и примерно у 90% новорожденных. В этом случае HVB инфекция становится хронической. Вероятной причиной является недостаточный клеточный иммунный ответ. Имеющиеся в настоящее время противовирусные лекарства, применяемые для лечения HVB инфекции, ингибируют репликацию вирусов. Однако, ковалентно непрерывная кольцевая ДНК (кнкДНК) остается в ядре инфицированных гепатоцитов и может вызывать реактивацию HVB инфекции, как только пациент прекращает принимать медикаменты. Таким образом, необходимо устранить клетки, инфицированные HVB, несущие указанную кнкДНК, чтобы полностью вылечить инфекцию (Pроtzer et al., Nat Immunol Rev 12: 2013-213 (2012)).
Однако указанная цитотоксическая элиминация клеток, инфицированных HVB (посредством цитотоксических Т-лимфоцитов или естественных киллерных (NK) клеток), не происходит, или осуществляется в недостаточной степени.
Инфицированные клетки, несущие кнкДНК HVB, несут на своей поверхности вирусные поверхностные белки. Предполагается, что это происходит, хотя вирус высвобождается во внутриклеточные везикулы, потому что ряд поверхностных белков HVB остается интегрированным во внутриклеточную мембрану эндоплазматического ретикулума. В ходе процессов переноса везикул указанная внутриклеточная мембрана может сливаться с клеточной мембраной, вследствие чего на поверхности инфицированной клетки выявляются поверхностные белки HVB.
Bohne et al. («T cells redirected against hepatitis B virus surface proteins eliminate infected hepatocytes». Gastroenterology; 134:239-247 (2008) («Т-клетки, направленные против поверхностных белков вируса гепатита В, устраняют инфицированные гепатоциты»)) и Krebs et al. («T Cells Expressing a Chimeric Antigen Receptor That Binds Hepatitis B Virus Envelope proteins Control Virus Replication in Mice». Gastroenterology (2013) («Т-клетки, экспрессирующие химерный антигенный рецептор, связывающий белки оболочки вируса гепатита В, контролируют репликацию вируса у мышей»)) описывают химерные антигенные рецепторы, которые при ретровирусной доставке и экспрессии на поверхности Т-клетки позволяют первичным человеческим и мышиным Т-клеткам распознавать гепатоциты, несущие малый поверхностный антиген HVB и осуществлять лизис клеток, реплицирующих HVB.
EP 2 524 699 A1 описывает трифункциональные антитела. Эти антитела «имеют функциональную Fc часть» и «должны состоять из тяжелых цепей иммуноглобулинов различных подклассов». С другой стороны, Hornig und Färber-Schwarz описывают в главе 40 «Antibody Engineering» (ed. Patrick Channes, Humane Press, 2012) scFv конструкцию, которая не содержит Fc части.
Liao et al. (Oncology Reports 3, 637-644 (1996)) описывает биспецифические моноклональные антитела, переориентирующие эффекторные клетки для лизиса ксенотрансплантатов гепатомы человека у бестимусных мышей. Описанные биспецифические антитела генерируют путем слияния двух гибридом, с получением гибридомной клеточной линии, экспрессирующей комбинации тяжелой/легкой цепи двух разных антител. Это может приводить к конъюгации двух различных тяжелых цепей, но также к конъюгации идентичных тяжелых цепей, давая начало произвольной смеси моноспецифических исходных и биспецифических антител. Биспецифические антитела содержат тяжелую и легкую цепь, и димеризуются с образованием молекулы Ig.
С учетом предшествующего уровня техники технической проблемой является обеспечение альтернативных или улучшенных средств и способов лечения HVB инфекции, а также состояний, вызванных HVB инфекцией, таких как цирроз печени или гепатоцеллюлярная карцинома. Технической проблемой, которая должна быть решена, также является предоставление способа лечения HVB инфекции. При выражении в терминах клеточной биологии, технической проблемой является предоставление средств и способов для уничтожения клеток, несущих кнкДНК HVB. Эта техническая проблема решена посредством прилагаемой формулы изобретения.
Соответственно, настоящее изобретение относится в первом аспекте к связывающей молекуле, включающей по меньшей мере три специфичности связывания, где (а) первая специфичность является специфичностью к поверхностному антигену вируса гепатита В (HVB), выбранному из малого поверхностного антигена HVB, среднего поверхностного антигена HVB и большого поверхностного антигена HVB; (b) (i) вторая и третья специфичность являются специфичностями к CD3 и CD28, соответственно; или (ii) вторая и третья специфичность выбраны из специфичностей к CD16, CD56, NKp30, NKp46, 4-1BB и NKG2D; и (с) каждая специфичность связывания обеспечена одним или несколькими участками связывания, где каждый участок связывания независимо обеспечен (i) набором из шести гипервариабельных областей (CDR), где каждый набор из шести CDR состоит из первого набора из трех CDR и второго набора из трех CDR, где каждый из указанного первого и указанного второго набора включен в иммуноглобулиновый домен; или (ii) набором из трех CDR, где каждый набор из трех CDR включен в иммуноглобулиновый домен.
«Поверхностный антиген HVB» также обозначается аббревиатурой «HBs» или «HBsAg».
Связывающие способности связывающей молекулы в соответствии с настоящим изобретением функционально определяют с применением термина «специфичность связывания». Структурные элементы, обеспечивающие указанные специфичности связывания, обозначаются как «участки связывания». В связи каждой из указанных специфичностей предусматривается, что в связывающей молекуле в соответствии с настоящим изобретением присутствует один или несколько участков связывания, обеспечивающих данную специфичность. В той степени, в которой конкретная специфичность связывания реализуется в связывающей молекуле в соответствии с изобретением посредством более чем одного участка связывания, понятно, что предпочтение отдается участкам связывания, реализующим определенную специфичность связывания, идентичным как по структуре, так и по функции, где функцией является распознавание определенного эпитопа связываемого антигена. Это не исключает, однако, вариантов осуществления, где конкретная специфичность связывания реализуется множеством участков связывания, где участки связывания, при связывании с тем же антигеном, осуществляют это через различные эпитопы антигена. Это обычно также влечет за собой структурные различия между участками связывания, хотя в общих чертах они обладают способностью связываться с одним и тем же антигеном.
В зависимости от точного числа, примерные числа, являющиеся одним, двумя, тремя и четырьмя, предпочтительно двумя участками связывания, обеспечивающими конкретную специфичность общих свойств связывания, могут быть точно настроены или оптимизированы. Как правило, путем увеличения количества участков связывания, обеспечивающих определенную специфичность, можно улучшить кинетику связывания, а также термодинамику связывания.
Общее количество участков связывания также обозначается как «валентность» в данной области техники. Соответственно, хотя существует потребность в связывающей молекуле, которая является триспецифической, то есть является по меньшей мере трехвалентной, следует понимать, что связывающие молекулы в соответствии с настоящим изобретением могут также быть четырехвалентными, пятивалентными, шестивалентными, семивалентными или восьмивалентными. Тем не менее, предусмотрены более высокие валентности. Предполагается, что удаление клеток, несущих кнкДНК HVB, достигается также путем использования девятивалентных, десятивалентных связывающих молекул, или связывающих молекул с еще большим числом валентностей.
Для того чтобы связывающая молекула была, по меньшей мере, триспецифической, достаточно, чтобы каждая специфичность связывания была представлена точно одним участком связывания. Соответственно, триспецифическая связывающая молекула в соответствии с настоящим изобретением может представлять собой трехвалентную молекулу. Вообще говоря, предпочтение отдается тем триспецифическим молекулам, в которых каждая из одной, двух или трех специфичностей связывания реализуется с двумя участками связывания. Предпочтение также отдается тем триспецифическим молекулам, в которых каждая из одной, двух или трех специфичностей связывания реализуются с тремя участками связывания.
Также возможно, что три специфичности связывания реализуются различным количеством участков связывания каждая. Может быть предпочтительно, чтобы одна специфичность связывания была представлена большим числом участков связывания, чем две другие, или чтобы две специфичности связывания могли быть представлены большим числом участков связывания, чем третья.
Как видно из пункта (с) (i) первого аспекта, набор из шести CDR является предпочтительным осуществлением определенного участка связывания. Как хорошо известно в данной области техники, наборы из шести CDR содержат первый и второй набор, каждый из которых состоит из трех CDR. Эти два набора из трех CDR могут быть расположены в одной и той же полипептидной цепи или в различных полипептидных цепях, таким образом, давая начало различным молекулярным архитектурам. Также предполагается, что в данной молекуле связывания в соответствии с настоящим изобретением часть участков связывания такова, что все шесть CDR, составляющих данный участок связывания, расположены в одной полипептидной цепи, тогда как другие участки связывания таковы, что первый и второй набор из трех CDR, составляющих данный участок связывания, расположены на разных полипептидах. Различные полипептиды могут быть просто формально разными и в остальном идентичными, особенно в отношении аминокислотной последовательности. Альтернативно, различные полипептиды могут отличаться друг от друга по их аминокислотной последовательности.
Вообще говоря, и это будет очевидно из обсуждения предпочтительных вариантов осуществления ниже, связывающие молекулы в соответствии с настоящим изобретением могут иметь исключительно участки связывания, которые образованы одним полипептидом. Понятно, однако, что это не означает, что связывающая молекула в целом состоит из одной полипептидной цепи. С другой стороны, настоящее изобретение распространяется на связывающие молекулы, где все участки связывания образованы двумя полипептидными цепями. Точно так же последнее утверждение не является требованием, чтобы связывающая молекула в целом состояла только из двух полипептидных цепей. Вместо этого связывающие молекулы также могут состоять более чем из двух полипептидных цепей, таких как три или четыре, или более четырех полипептидных цепей. Наконец, как упомянуто выше, и как показано в приведенной ниже структуре (III), одна часть участков связывания, включающая связывающую молекулу по настоящему изобретению, может быть такой, что каждый из них образован двумя полипептидными цепями, тогда как остальная часть участков связывания такова, что они образованы одной полипептидной цепью.
В отношении пункта (b) (ii) необходимо понять, что вторая и третья специфичность отличаются друг от друга, тем самым обеспечивая, по меньшей мере, триспецифическую связывающую молекулу.
В той степени, в которой рассматривается пункт (c) (i), из требования шести CDR следует, что для образования единственного участка связывания требуется два домена иммуноглобулина. Это отличается от пункта (c) (ii), где один иммуноглобулиновый домен является достаточным для обеспечения участка связывания, причем участок связывания в этом случае состоит из трех CDR. Это также известно как однодоменный участок связывания.
Термин «гипервариабельная область», сокращенно называемая «CDR», имеет значение, установленное в данной области техники. Это короткие подпоследовательности, обычно в диапазоне от примерно 3 до примерно 25 аминокислот, которые придают антителу способность специфически распознавать эпитоп антигена. В общем случае вариабельный домен легкой цепи антитела обеспечивает три CDR, а вариабельный домен тяжелой цепи антитела обеспечивает три CDR. Если не указано иное, шесть CDR, определяющих участок связывания, обычно представлены в следующем порядке: CDR1 тяжелой цепи, CDR2 тяжелой цепи, CDR3 тяжелой цепи, CDR1 легкой цепи, CDR2 легкой цепи, CDR3 легкой цепи. Хотя CDR обычно являются частью доменов иммуноглобулина, в этом отношении нет требований в соответствии с настоящим изобретением. Достаточна аминокислотная последовательность, которая содержит указанные CDR, при условии, что указанная аминокислотная последовательность, когда она уложена в физиологических условиях, представляет указанные CDR в пространственной близости и поддерживает их способность к распознаванию когнатного антигена. Указанная пространственная близость и способность к связыванию антигена также могут быть названы «конфигурацией для связывания антигена». Термин «иммуноглобулиновый домен» известен в данной области техники, и относится к последовательности, как правило, от 70 до 100 аминокислот, предполагая трехмерную структуру двухслойного сэндвича от 7 и 9 антипараллельных β-цепей. В структурах, определяющих определенные варианты осуществления, а также на чертежах, эллипсы используются для отображения одного домена иммуноглобулина.
Набор из шести CDR обычно определяет участок связывания. В альтернативе, и как известно из верблюжьих антител и антител хрящевых рыб, набор из трех CDR (CDR1, CDR2, CDR3) может определять участок связывания. Как известно в данной области техники, и хотя это не требуется, предпочтение отдается таким наборам из трех CDR, где CDR3 состоит из более выступающей пептидной петли, чем во встречающихся в природе вариабельных иммуноглобулиновых доменах.
Термин «антиген» имеет свое значение, установленное в данной области техники. Он относится к молекуле, которая специфически распознается и связывается набором из шести CDR, которые обычно представлены доменами иммуноглобулина. Конкретная часть антигена, распознаваемая и связываемая указанными CDR, также известна как эпитоп.
Поверхностные белки М/С/Б HVB являются малым, средним и большим поверхностными белками во внешней оболочке HVB (Stibbe, W., and W. H. Gerlich. «Structural relationships between minor and major proteins of hepatitis B surface antigen». J. Virol. 1983 46:626-628 («Структурные взаимоотношения между второстепенными и основными белками поверхностного антигена гепатита В»)).
Три поверхностных антигена HVB транскрибируются и транслируются из одной открытой рамки считывания и отличаются друг от друга длиной N-концевой части. Соответственно, большой поверхностный антиген содержит часть, которая не присутствует ни в среднем, ни в малом поверхностном антигене, а средний поверхностный антиген содержит часть, которая, будучи в составе большого антигена, не содержится в малом антигене. Малый антиген состоит из последовательности, которая содержится в С-концевой части как среднего, так и большого антигена.
Большой поверхностный антиген HVB может быть встроен двумя способами в цитоплазматическую мембрану. Либо N-конец, либо С-конец может быть расположен на внеклеточной стороне. Обе конфигурации находятся в клетках, инфицированных HVB. Другими словами, N-конец большого поверхностного антигена HVB может быть расположен либо на цитозольной, либо на люминальной стороне мембраны ЭР, и следовательно, также присутствовать в этих конфигурациях во внешней оболочке частицы HVB и плазматической мембране инфицированного гепатоцита.
Неожиданно, объединение второй и третьей специфичностей в соответствии с (b) в триспецифические молекулы обеспечивает значительно более сильную активацию иммунных эффекторных клеток, чем комбинация двух биспецифических молекул. Обратимся к примерам.
В еще одном предпочтительном варианте осуществления участок связывания, обеспечивающий специфичность связывания для поверхностного антигена HVB, связывает эпитоп, который расположен (а) в указанном малом поверхностном антигене HVB; или (b) в части указанного большого поверхностного антигена HVB, которая не содержится в указанном малом поверхностном антигене HVB; или (с) в части указанного большого поверхностного антигена HVB, которая отличается по структуре от указанного малого поверхностного антигена HVB.
Пункт (а) относится к эпитопам, присутствующим в малом поверхностном антигене HVB. Благодаря описанной выше связи между малым, средним и большим поверхностным антигеном HVB, вся последовательность малого антигена содержится в среднем и большом антигене. В общем случае, но не обязательно, трехмерный эпитоп, представленный малым поверхностным антигеном, также будет представлен средним и/или большим поверхностным антигеном.
В соответствии с пунктом (b) предпочтительно, чтобы указанная часть указанного большого поверхностного антигена HVB также не содержалась в указанном среднем поверхностном антигене HVB. Что касается пункта (с), следует понимать, что «различная структура» включает эпитопы указанного большого поверхностного антигена HVB, которые включают или состоят из последовательностей, которые являются частью последовательности указанного малого поверхностного антигена HVB, где указанные эпитопы отсутствуют на указанном малом поверхностном антигене HVB. В соответствии с пунктом (с), также предпочтительно, чтобы указанный эпитоп находился в части указанного большого поверхностного антигена HVB, часть которого отличается по структуре также от среднего указанного поверхностного антигена HVB.
Указанный пункт (а), то есть указанный первый антиген, являющийся указанным малым поверхностным антигеном HVB, является особенно предпочтительным в сочетании со всеми аспектами и вариантами осуществления настоящего изобретения.
В соответствии с пунктами (b) и (c), полипептид или связывающая молекула, соответственно, будут специфически распознавать большой поверхностный антиген HVB.
Вторая и третья специфичности направлены на поверхностные антигены, представленные иммунными эффекторными клетками, предпочтительно специфически представленными NK-клетками и/или ЦТЛ. Иммунные эффекторные клетки - это клетки, которые должны быть переориентированы на клетки, инфицированные HVB, причем указанные клетки, инфицированные HVB, презентируют упомянутые поверхностные антигены HVB на своей поверхности.
Особо предпочтительно, чтобы вторая и третья специфичность были либо обе для ЦТЛ, либо обе для NK-клеток.
CD3 (кластер дифференцировки 3) Т-клеточный корецептор представляет собой белковый комплекс и состоит из четырех различных цепей. У млекопитающих комплекс содержит цепь CD3γ, цепь CD3δ и две цепи CD3ε. Специфичности связывания могут быть направлены на любую из этих цепей. Эти цепи связываются с молекулой, известной как Т-клеточный рецептор (TCR), и ζ-цепью, для генерации сигнала активации в Т-лимфоцитах. Молекулы TCR, ζ-цепи и CD3 вместе образуют комплекс TCR.
В предпочтительном варианте осуществления CD3 обозначает цепь CD3-эпсилон, которая является частью рецепторного комплекса CD3-Т-клеток (Borst, J. et al., «The delta- and epsilon-chains of the human T3/T-cell receptor complex are distinct polypeptides». Nature. 1984. 312: 455-458 («Дельта- и эпсилон-цепи человеческого рецепторного комплекса Т3-Т-клеток являются различными полипептидами»)). Предпочтительно, специфичность связывания для CD3 в соответствии с настоящим изобретением относится к указанной CD3 эпсилон цепи.
CD28 является главным Т-клеточным костимулирующим рецептором (Lesslauer, W. et al., «T90/44 (9.3 antigen). A cell surface molecule with a function in human T cell activation». Eur. J. Immunol. 1986. 16: 1289-1296 («Т90/44 (9.3 антиген). Клеточно-поверхностная молекула с функцией активации Т-клеток»)).
4-1BB (CD137) является костимулирующим рецептором активированных Т-клеток и NK клеток. (Kwon, B.S. et al., «cDNA sequences of two inducible T-cell genes». Proc. Natl. Acad. Sci. U. S. A. 1989. 86:.1963-1967 («Последовательности кДНК двух индуцибельных Т-клеточных генов).
CD16 (FcγRIIIa) является низкоаффинным Fc рецептором, экспрессируемым NK клетками, подгруппой активированных цитотоксических Т-клеток, а также клеточными типами миеломоноцитарной линии, связывающимися с Fc доменом молекул IgG. (Lanier, L.L. et al., «Functional Properties of a unique subset of cytotoxic CD3+ T lymphocytes that express Fc receptors for IgG (CD16/Leu-11 antigen)». J. Exp. Med. 1985. 162: 2089-2106 («Функциональные свойства уникальной подгруппы цитотоксических CD3+ T-лимфоцитов, которые экспрессируют Fc рецепторы для IgG (CD16/ Leu-11 антиген)»)).
CD56 (NCAM) является молекулой клеточной адгезии, экспрессируемой NK клетками. (Lanier, L.L. et al., «Identity of Leu-19 (CD56) Leukocyte differentiation antigen and neural cell adhesion molecule». J. Exp. Med. 1989. 169: 2233-2238 («Идентичность Leu-19 (CD56) антигена дифференцировки лейкоцитов и молекулы адгезии нервных клеток»)).
NKG2D является рецептором активации, экспрессируемым NK клетками (Houchins, J. et al., «DNA sequence analysis of NKG2, a family of related cDNA clones encoding type II integral membrane proteins on human natural killer cells». 1991. J. Exp. Med. 173: 1017-1020 («Анализ последовательности ДНК NKG2, семейства родственных кДНК клонов, кодирующих интегральные мембранные белки II типа на естественных киллерных клетках человека»)).
NKp30 (NCR3) является рецептором активации, экспрессируемым NK клетками (Pende, D. et al., «Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells». 2000. J. Exp. Med. 192: 337-346 («Идентификация и молекулярная характеристика NKp30, нового триггерного рецептора, вовлеченного в естественную цитотоксичность, опосредованную естественными киллерными клетками человека»)).
NKp46 (NCR1) является рецептором активации, экспрессируемым NK клетками (Pessino et al., «Molecular cloning of NKp46: a novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicity». 1998. J. Exp. Med. 188: 953-960 («Молекулярное клонирование NKp46: новый член семейства иммуноглобулинов, вовлеченный в индукцию естественной цитотоксичности»)).
CD3, CD28 и 4-1BB присутствуют на поверхности ЦТЛ. Связывание полипептида из настоящего изобретения с любым из этих поверхностных антигенов приводит к стимуляции или костимуляции ЦТЛ.
CD16, CD56, NKG2D, NKp30, NKp46 и 4-1BB присутствуют на поверхности NK клеток. Связывание полипептида из настоящего изобретения с любым из этих поверхностных антигенов приводит к стимуляции или костимуляции NK клеток.
Что касается человеческих ЦТЛ, предпочтение отдается CD3 и CD28. Что касается человеческих NK клеток, предпочтение отдается CD16 и CD56.
Упомянутые поверхностные антигены обозначаются посредством наименований, установленных в данной области техники (см. также Kenneth Murphy, Janeway's Immunobiology, 7th edition, Garland Science; William E. Paul, Fundamental Immunology, 7th edition, Lippincott Williams & Wilkins).
Особенно предпочтительно, связывание в соответствии с изобретением, в частности между CDR и антигенами, является специфическим. Термины «специфически связывают» и «специфически связывающиеся» (имеющие то же значение, что и «специфически взаимодействующие»), используемые в соответствии с настоящим изобретением, означают, что эти связывающие части не реагируют, или практически не реагируют перекрестно с эпитопом или структурой, подобными таковым у целевого антигена. Перекрестная реактивность исследуемой группы молекул может быть проверена, например, путем оценки связывания указанной группы молекул в обычных условиях с представляющим интерес эпитопом, а также с рядом более или менее (структурно и/или функционально) тесно связанных эпитопов. Только те молекулы, которые связываются с представляющим интерес эпитопом в его соответствующем контексте (например, конкретным мотивом в структуре белка), но не связываются или по существу не связываются с каким-либо из других эпитопов, считаются специфическими для представляющего интерес эпитопа.
Хроническая инфекция HBV характеризуется иммунотолерантным статусом. Более конкретно, ЦТЛ и NK-клетки пациентов действуют так, что полное уничтожение инфицированных клеток или полный контроль репликацией HVB или полное уничтожение HVB не происходит. Связывающие молекулы согласно изобретению специфически распознают поверхностный антиген HVB, с одной стороны, и поверхностный антиген иммунной эффекторной клетки, с другой стороны. Связывающие молекулы по изобретению можно рассматривать как придающие искусственную специфичность иммунным эффекторным клеткам. На самом деле, ЦТЛ и NK-клетки переориентируются связующими молекулами по изобретению так, что они привлекаются к инфицированным HVB клеткам и убивают их.
С одной стороны, связывание связывающих молекул по изобретению с инфицированными HVB клетками и, с другой стороны, рекрутинг иммунных эффекторных клеток может происходить в любом порядке или одновременно.
В частности, предполагается системно применять связывающие молекулы по изобретению либо путем инъекции, либо в виде пероральной формы, и позволить им связываться с HVB-инфицированными или экспрессирующими HVB-антиген клетками-мишенями, и рекрутировать указанные иммунные эффекторные клетки к указанным клеткам-мишеням.
Говоря это, предполагается также привести связывающие молекулы из настоящего изобретения в контакт с иммунными эффекторными клетками (или популяцией мононуклеарных клеток периферической крови, содержащей указанные эффекторные клетки), так чтобы упомянутые эффекторные клетки загружались упомянутыми связывающими молекулами. Такие эффекторные клетки, которые были загружены in vitro или ex vivo (или популяцию МНКПК, содержащую такие загруженные эффекторные клетки), можно затем вводить пациенту, страдающему HVB инфекцией, или связанным с ней состоянием, как определено ниже. Такое введение может быть осуществлено внутрисосудистым путем, например, через печеночную артерию. Иммунная эффекторная клетка со связывающей молекулой в соответствии с настоящим изобретением, связанной с поверхностным антигеном указанной иммунной эффекторной клетки, также является аспектом настоящего изобретения. Этот аспект раскрыт ниже.
Этот лизис, в частности в сочетании с противовирусными иммунными медиаторами (например, цитокинами), секретируемыми иммунными клетками, обеспечивает искоренение HVB инфекции или постоянный контроль HVB инфекции, или устранение опухолевых клеток, экспрессирующих поверхностные антигены HVB.
Предпочтительные показатели эффективности связывающих молекул по изобретению включают Т-клеточно-опосредованную цитотоксичность в культуре клеток, рекрутинг Т-клеток в печень, воспалительный ответ печени, снижение титров вируса в сыворотке, снижение вирусных антигенов HBsAg и/или HBeAg в сыворотке, снижение репликации вируса в печени и уменьшение количества инфицированных клеток.
Каждое из этих свойств может быть измерено в соответствии с процедурами, установленными в данной области техники.
Для дальнейшего разъяснения, рекрутинг Т-клеток в печень можно количественно определить путем анализа уровней одной или нескольких мРНК маркерного гена Т-клеток, в частности, CD3, CD4, CD8, ИФНγ и IP-10 (CXCL10), с помощью RT-PCR в печени. В соответствии с настоящим изобретением, индукция этих маркерных генов значительно повышается по сравнению с отрицательным контролем, и может достигать уровней от 2-кратной до 1000-кратной индукции или выше, предпочтительно более чем 2-кратной, предпочтительно более чем 5-кратной, предпочтительно более чем 10-кратной, предпочтительно более чем 25-кратной, предпочтительно более чем 50-кратной, предпочтительно более чем 100-кратной, предпочтительно более чем 500-кратной, предпочтительно более чем 750-кратной, предпочтительно более чем 1000-кратной.
В соответствии с настоящим изобретением, Т-клеточно-опосредованная цитотоксичность в культуре клеток находится в пределах от 5 до 100%, предпочтительно более 5%, предпочтительно более 10%, предпочтительно более 20%, предпочтительно более 30%, предпочтительно более 40%, предпочтительно более 50%, предпочтительно более 75%, предпочтительно 100%.
T-клеточно-опосредованную цитотоксичность in vivo можно оценить, например, путем анализа трансаминаз печени (предпочтительно аланин-трансаминазы (АлАт) и/или аспартат-трансаминазы (АсАт), которые высвобождаются при лизисе гепатоцитов. Нормальные уровни АлАт у индивидуумов составляют ниже 40 международных единиц на литр (Ед./л) и могут достигать уровней от 50 Ед./л до 5000 Ед./л при эффективной цитотоксичности путем переориентации иммунных эффекторных клеток. Предпочтительно, уровни будут составлять более 50 Ед./л, предпочтительно более 100 Ед./л, предпочтительно более 200 Ед./л, предпочтительно более 300 Ед./л, предпочтительно более 400 Ед./л, предпочтительно более 500 Ед./л, предпочтительно более 750 Ед./л, предпочтительно более 1000 Ед./л.
В соответствии с настоящим изобретением, воспалительные реакции печени могут быть определены с помощью окрашивания ГЭ. ГЭ означает гематоксилин и эозин, и является стандартной методикой окрашивания для патологии и позволяет визуализировать клеточные структуры в оттенках синего, пурпурного, красно-розового и оранжевого. С помощью этого метода инфильтрирующие иммунные эффекторные клетки могут быть визуализированы и количественно оценены по сравнению с отрицательным контролем, и их количество будет увеличиваться в соответствии с настоящим изобретением в диапазоне от 5 до 5000 раз по сравнению с необработанным контролем, предпочтительно более чем в 5 раз, предпочтительно более чем в 10 раз, предпочтительно более чем в 20 раз, предпочтительно более чем в 50 раз, предпочтительно более чем в 100 раз, предпочтительно более чем в 250 раз, предпочтительно более чем в 500 раз, предпочтительно более чем в 1000 раз.
CD3-окрашивание биоптатов, взятых из печени, и/или окрашивание на триспецифические связывающие молекулы по изобретению, выявляет специфическую инфильтрацию Т-клеток и переориентированных Т-клеток, соответственно. С помощью этого способа переориентированные CD3+ Т-клетки могут быть визуализированы и количественно оценены по сравнению с отрицательным контролем, и их количество будет увеличиваться в соответствии с настоящим изобретением в диапазоне от 5 до 5000 раз по сравнению с необработанным контролем, предпочтительно более чем в 5 раз, предпочтительно более чем в 10 раз, предпочтительно более чем в 20 раз, предпочтительно более чем в 50 раз, предпочтительно более чем в 100 раз, предпочтительно более чем в 250 раз, предпочтительно более чем в 500 раз, предпочтительно более чем в 1000 раз.
Репликацию вируса можно количественно определить с помощью ПЦР и/или Саузерн-блоттинга образцов сыворотки или ткани печени. Применение связывающей молекулы по изобретению у зараженного HVB субъекта вызывает снижение вирусных титров в сыворотке. В соответствии с настоящим изобретением, предусматривается уменьшение более чем на 10%, предпочтительно более чем на 20%, предпочтительно более чем на 30%, предпочтительно более чем на 40%, предпочтительно более чем на 50%, предпочтительно более чем на 60%, предпочтительно более чем на 70%, предпочтительно более чем на 80%, предпочтительно более чем на 90%, предпочтительно ниже уровня обнаружения (50 копий ДНК HVB на мл) вирусных титров в сыворотке крови, достигаемое после применения эффективного количества связывающей молекулы. Например, уровни ДНК HVB в сыворотке достигают до 109 копий на мл у HVB-трансгенных мышей и могут быть уменьшены до 107 копий/мл при лечении переориентированными Т-клетками (Krebs et al. «T-Cells Expressing a Chimeric Antigen Receptor That Binds Hepatitis B Virus Envelope Proteins Control Virus Replication in Mice». Gastroenterology (2013) («Т-клетки, экспрессирующие химерный рецептор антигена, связывающий белки оболочки вируса гепатита В, контролируют репликацию вируса у мышей»)).
Количество оставшихся инфицированных клеток можно определить, окрашивая биоптаты печени на HBeAg, HBcAg или HBsAg. По сравнению с необработанными контрольными образцами, уменьшение HVB-реплицирующих клеток в соответствии с настоящим изобретением будет составлять от минимум 5% до максимум 100%, предпочтительно более 5%, предпочтительно более 10%, предпочтительно более 20%, предпочтительно более 30%, предпочтительно более 50%, предпочтительно более 70%, предпочтительно более 90%, предпочтительно 100%.
«Вирусный титр» (также известный как вирусная нагрузка) является термином, хорошо известным в данной области техники, указывающим количество вируса в данном биологическом образце. Количество вируса указывается различными способами, включая, количество вирусной нуклеиновой кислоты; присутствие вирусов (таких как полные вирионы, определяемые путем осаждения при градиентном центрифугировании) и субвирусных частиц (таких, как HBsAg или частицы поверхностного антигена вируса гепатита B, определенные с помощью иммунологических анализов, таких как ИФА), но не ограничиваясь ими. Как правило, для образцов жидкости, таких как кровь и моча, количество вируса определяют на единицу жидкости, например, на миллилитр. Для твердых образцов, таких как образцы тканей, количество вируса определяют на единицу массы, например, в граммах. Способы определения количества вируса известны в данной области техники. В настоящее время доступно рутинное обычное тестирование для вируса гепатита B, такое как измерение вирусной нагрузки, качественное и количественное определение HBsAg и HBeAg.
«Эффективное количество» относится к количеству агента, достаточному для достижения вышеописанной цели. В частности, количество связывающей молекулы в соответствии с изобретением является эффективным для снижения уровня вирусного титра у субъекта, инфицированного вирусом гепатита B, подлежащего лечению. Эффективное количество можно вводить посредством одного или нескольких применений. Эффективное количество может варьировать в зависимости от предполагаемого применения (in vitro или in vivo) или субъекта и патологического состояния, подлежащего лечению, например, массы тела и возраста субъекта, тяжести патологического состояния, способа введения и тому подобного, что может быть легко определено специалистом в данной области техники.
Есть, например, много разных способов на молекулярной основе для количественной оценки вирусной нагрузки с использованием технологии амплификации нуклеиновых кислот (NAT). Исходный материал для амплификации можно использовать для разделения этих молекулярных методов на группы. Метод полимеразной цепной реакции (ПЦР) для синтеза ДНК in vitro использует ДНК-матрицу, полимеразу, буферы, праймеры и нуклеотиды для умножения ВИЧ в образце крови. Затем химическая реакция маркирует вирус. Маркеры измеряются и используются для расчета количества вируса. ПЦР используют для количественной оценки интегрированной ДНК.
Полимеразная цепная реакция с обратной транскриптазой (RT-PCR) представляет собой вариацию ПЦР, которая может использоваться для количественной оценки вирусного транскрипта. РНК используют в качестве исходного материала для этого способа, и превращают в двухцепочечную ДНК с использованием фермента обратной транскриптазы (RT).
Терапевтической целью для хронического гепатита В является «функциональное» излечение HVB-инфекции, которое не достигается с доступными в настоящее время противовирусными агентами. Как отмечено выше, основными препятствиями для функционального излечения являются сложная устойчивость кнкДНК HVB и дисфункциональный иммунный ответ против HVB. Функциональное излечение HVB в контексте настоящего изобретения означает, что количество геномов HVB на мл составляет ниже 100000, предпочтительно ниже 10000, более предпочтительно ниже 1000, 100 или 10. Особо предпочтительным является полное уничтожение или уменьшение ниже предела обнаружения.
Учитывая, что связывающие молекулы в соответствии с настоящим изобретением обеспечивают специфические особенности для иммунных эффекторных клеток, естественно присущая специфичность иммунных эффекторных клеток или презентация антигенов для них становятся несущественными. Таким образом, большой пул потенциальных эффекторных клеток поддается переориентированию. Кроме того, связывающие молекулы по изобретению имеют биодоступность и период полужизни, которые по меньшей мере сопоставимы с показателями для моноклональных антител.
В предпочтительном варианте осуществления первого аспекта указанные по меньшей мере три специфичности связывания представляют собой (а) три специфичности; или (b) четыре специфичности, и при этом четвертая специфичность является специфичностью для клеток печени.
Предпочтение отдается связывающей молекуле, являющейся триспецифической связывающей молекулой. Другими словами, присутствуют только три специфичности связывания. Альтернатива присутствия четвертой специфичности может быть выгодной для нацеливания на определенные клетки, в частности на печеночные клетки. В этом отношении предпочтительным маркером на печеночных клетках является NTCP (полипептид, совместно транспортирующий таурохолат натрия). NTCP известен в данной области техники как рецептор, опосредующий проникновение HVB.
В еще одном предпочтительном варианте осуществления указанная молекула представляет собой (а) мультимер из полипептидов; или (б) полипептид. Понятно, что пункт (б) этого предпочтительного варианта осуществления определяет связывающую молекулу, которая состоит из одного полипептида.
Термин «полипептид» определяет молекулу, которая представляет собой поликонденсат аминокислот, которые образуют одну целую цепь с одним N-концом и одним С-концом. Составные аминокислоты включают 20 встречающихся в природе протеиногенных аминокислот. Предпочтительно указанный полипептид состоит исключительно из указанных естественных протеиногенных аминокислот. Говоря это, этот термин распространяется на молекулы, которые помимо указанных встречающихся в природе протеиногенных аминокислот содержат до 20%, 10%, 5%, 2% или 1% аминокислот, которые выбраны из не встречающихся в природе α-аминокислот, β-аминокислот, D-аминокислот, селеноцистеина, селенометионина, гидроксипролина, пирролизина и орнитина. Кроме того, понятно, что одна или более, такое количество, как 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот, могут быть фосфорилированы. Последнее относится, в частности, к серину, треонину и тирозину. Также могут присутствовать другие посттрансляционные модификации, известные в данной области, включая гликозилирование. Гликозилирование включает N-связанные гликозилирования, как правило, на аспарагине, и O-связанные гликозилирования, обычно в остатках серина или треонина. N- и/или С-конец могут быть защищены; защитные группы включают ацетил для N-конца и амин для С-конца. Тип связи между аминокислотами, содержащимися в указанном полипептиде, ограничен амидными связями (CONH). Термин «амидная связь» включает пептидные связи, которые связывают α-карбоксилат данной аминокислоты с α-аминогруппой последующей аминокислоты. «Амидная связь» также распространяется на изопептидные связи, которые представляют собой амидную связь, которая отсутствует в основной цепи полипептида. Например, вместо α-аминогруппы может быть задействована аминогруппа боковой цепи лизина. Аналогично, вместо α-карбоксильной группы может быть задействован карбоксилат боковой цепи глутамата или аспартата. Предусматривается наличие одной или нескольких, такое количество, как 2, 3, 4, 5, 6, 7, 8, 9 или 10 изопептидных связей. Однако предпочтение отдается полипептидам, в которых составляющие аминокислоты связаны друг с другом исключительно пептидными связями.
В общем, нет верхнего предела количества аминокислот в полипептиде. Как можно видеть из примерных полипептидных последовательностей, включенных в список последовательностей, полипептиды по настоящему изобретению обычно содержат несколько сотен аминокислот, предпочтительно от 250 до 1000, от 400 до 900, или от 700 до 800 аминокислот. Обычно различают пептиды, с одной стороны, и полипептиды, с другой стороны, где пептиды имеют 30 или менее аминокислот, а полипептиды имеют более 30 аминокислот.
Термин «мультимер» в соответствии с настоящим изобретением включает димеры, тримеры, тетрамеры, пентамеры, гексамеры, гептамеры и октамеры.
В самой широкой форме нет никаких ограничений в отношении местоположения по меньшей мере трех требуемых специфичностей на соответствующих составных полипептидах мультимера. Хотя можно предположить, что используют составные полипептиды, где каждый полипептид обеспечивает точно одну специфичность связывания, реализуемую одним, двумя, тремя, четырьмя или более участками связывания, предпочтение отдается мультимерам, причем каждый из составных полипептидов несет две или три специфичности, особо предпочтительными являются составные полипептиды с двумя специфичностями.
В той степени, в которой связующая молекула по настоящему изобретению, которая предпочтительно представляет собой триспецифическую связывающую молекулу, представляет собой мультимер полипептидов, предпочтительно димер или тетрамер, предпочтительно, чтобы (а) каждый полипептид представлял собой биспецифический полипептид; или (б) каждый полипептид был независимо выбран из моноспецифических, биспецифических и триспецифических полипептидов.
Само собой разумеется, что этот предпочтительный вариант осуществления охватывает только конструкции, которые также соответствуют терминам самого широкого определения первого аспекта настоящего изобретения. Например, димер из двух моноспецифических полипептидов не соответствует требованию «триспецифический» и поэтому не охватывается этим предпочтительным вариантом осуществления.
В соответствии с приведенным выше объяснением различия между специфичностью связывания, с одной стороны, и участком связывания, с другой стороны, понятно, что моноспецифический полипептид может быть одновалентным, двухвалентным, трехвалентным, четырехвалентным или может содержать даже более четырех валентностей. Аналогично, биспецифические полипептиды могут быть двухвалентными, трехвалентными, четырехвалентными или содержать более четырех валентностей. Наконец, триспецифические полипептиды могут быть трехвалентными, четырехвалентными или содержать более четырех валентностей. Как отмечено выше, участок связывания может быть обеспечен двумя иммуноглобулиновыми доменами, расположенными на одном и том же полипептиде, но также двумя иммуноглобулиновыми доменами, расположенными на разных полипептидах. Поэтому в тех случаях, когда связывающая молекула в соответствии с настоящим изобретением представляет собой мультимер из полипептидов, количество валентностей связывающей молекулы может превышать количество валентностей всех составляющих полипептидов, взятых вместе. Разница между общим числом валентностей в связывающей молекуле и суммарным числом валентностей, обеспечиваемых всеми составляющими полипептидами, представляет собой число тех участков связывания, которые образованы двумя иммуноглобулиновыми доменами, расположенными на разных полипептидных цепях.
Используя структуру (III), как описано ниже в качестве примера, общее число участков связывания в этой молекуле составляет пять, четыре из которых образованы парами иммуноглобулиновых доменов, где каждая пара расположена на одной полипептидной цепи, и один участок связывания образован парой иммуноглобулиновых доменов, расположенных на разных полипептидных цепях. Общее количество полипептидов в структуре (III) равно двум.
В терминах механизмов мультимеризации или олигомеризации отмечается, что полипептиды могут быть связаны вместе ковалентно и/или нековалентно. Например, если связывающая молекула представляет собой димер двух полипептидов, указанные полипептиды могут быть исключительно связаны друг с другом одной или несколькими ковалентными связями, причем упомянутые ковалентные связи предпочтительно являются дисульфидными связями. Альтернативно, два полипептида могут быть связаны исключительно нековалентными взаимодействиями. В равной степени можно предположить, что два полипептида связаны друг с другом как ковалентными, так и нековалентными взаимодействиями. Аналогично, и учитывая мультимеры высшего порядка, понятно, что удерживать мультимер вместе могут только ковалентные взаимодействия, только нековалентные взаимодействия или комбинация ковалентных и нековалентных взаимодействий. Комбинации ковалентных и нековалентных взаимодействий могут быть такими, что для данного бинарного взаимодействия внутри мультимера существует только ковалентное или только нековалентное взаимодействие, или комбинация как ковалентного, так и нековалентного взаимодействий.
Термины «мультимеризация» и «олигомеризация» (применяемые эквивалентно) включают димеризацию.
Как будет дополнительно детально описано ниже, в некоторых случаях и для некоторых предпочтительных связывающих молекул по настоящему изобретению может быть предпочтительным ковалентное взаимодействие над нековалентным взаимодействием, или наоборот.
В той степени, в которой изобретение относится к мультимерам, составные полипептиды дополнительно содержат область димеризации или мультимеризации. Указанная область димеризации или мультимеризации может обеспечивать ковалентную и/или нековалентную димеризацию или мультимеризацию.
В особо предпочтительном варианте осуществления указанная область димеризации, которая соединяет два полипептида по изобретению, состоит из шарнирной области тяжелой цепи IgG или содержит цистеиновые остатки, ответственные за димеризацию тяжелых цепей антитела. Другие природные дисульфидные связи, которые могут быть использованы для димеризации и мультимеризации, образуются между соответствующими С-концами первого константного домена тяжелой цепи и константного домена легкой цепи.
Предпочтительно, указанная область димеризации состоит из подпоследовательности длиной в 32 аминокислоты, так называемой шарнирной области тяжелой цепи (EPKSSDKTHTCPPCPAPEFEGAPSVFLFPPKP, см. SEQ ID NO: 43-46) и содержит два остатка цистеина (подчеркнутые в вышеуказанной последовательности), ответственные за димеризацию тяжелых цепей. Предпочтительно, единственный остаток цистеина в шарнирной области тяжелой цепи IgG, который опосредует межмолекулярную дисульфидную связь между константными доменами тяжелой и легкой цепи IgG в натуральном антителе, мутирует в серин, чтобы предотвратить аберрантные дисульфидные мостики.
Домены димеризации, подходящие для нековалентной димеризации, известны в данной области техники, и включают лейциновые «застежки» (см., например, de Kruif and Logtenberg, «Leucine zipper dimerized bivalent and bispecific scFv antibodies from a semi-synthetic antibody phage display library». J. Biol. Chem 1996. 271: 7630-7634 («Лейциновая застежка димеризует двухвалентные и биспецифические scFv антитела библиотеки фагового дисплея полусинтетических антител»)). Еще одним вариантом запуска димеризации являются естественные взаимодействия между иммуноглобулиновыми доменами. Эти взаимодействия происходят, например, в молекуле IgG между вариабельными доменами тяжелой и легкой цепей, между первым константным доменом тяжелой цепи и константным доменом легкой цепи, соответствующими вторыми константными доменами тяжелых цепей и соответствующими третьими константными доменами тяжелых цепей.
В еще одном предпочтительном варианте осуществления составной полипептид дополнительно содержит спейсерную область, причем указанная спейсерная область предпочтительно содержит CH2 домен и CH3 домен. В случае составного полипептида, содержащего scFv-фрагменты, указанная спейсерная область может быть расположена между первым scFv-фрагментом и вторым scFv-фрагментом.
Предпочтительной является спейсерная область, содержащая или состоящая из CH2 домена и CH3 домена, в частности из IgG. Преимущества включают эффективную секрецию продуцируемыми клетками. Кроме того, способность такой спейсерной области связывать белок А обеспечивает средство для последующей очистки от реагентов.
С другой стороны, оба указанных СН2 и СН3 домена, и с другой стороны, указанная область димеризации, могут быть обеспечены соответствующей областью молекулы IgG, предпочтительно молекулы IgG1 или IgG2, еще более предпочтительной является молекула человеческого IgG1 или lgG2 (hIgG1, hlgG2). Предпочтительную подпоследовательность молекулы hIgG1, обеспечивающую CH2 домен и CH3 домен, можно видеть в SEQ ID NO: 43-46. Последовательности SEQ ID NO: 50 и 51 реализуют предпочтительный вариант нековалентной димеризации, более конкретно через домены лейциновой застежки. Предпочтительно - и это относится к указанным последовательностям - часть hIgG1, в частности, указанный CH2 домен, подвергнут мутации в нескольких положениях, чтобы уменьшить связывание с Fc-рецепторами (выделено жирным курсивом в последовательностях, приведенных ниже). В более общем случае, Fc область, в частности, CH2 домен и/или CH3 домен, может быть подвергнута мутации в одном или нескольких положениях, чтобы уменьшить связывание с Fc-рецепторами. Такая процедура известна в данной области техники и описана, например, в Armour et al., «Recombinant human IgG molecules lacking Fc gamma receptor I binding and monocyte triggering activities». Eur. J. Immunol. 1999. 29: 2613-2624 («Рекомбинантные человеческие IgG-молекулы, не связывающиеся с Fc-гамма-рецептором I и не способные к активации моноцитов») и Shields et al., «High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R». J. Biol. Chem. 2001. 276: 6591-6604 («Картирование высокого разрешения сайта связывания на человеческом IgG1 для Fc гамма RI, Fc гамма RII, Fc гамма RIII и FcRn, и разработка вариантов IgG1 с улучшенным связыванием с Fc гамма R»). Это является преимуществом, поскольку инициирование антителозависимой клеточно-опосредованной цитотоксичности (ADCC) не является предпочтительным в соответствии с настоящим изобретением.
Другими словами, фрагмент Fc антитела, предпочтительно модифицированный, как определено выше, может быть использован для реализации спейсерной области и области димеризации. Термин «фрагмент Fc» известен специалисту в данной области, и определяет фрагмент IgG, который получен расщеплением папаином и содержит домены CH2 и CH3.
Между указанным первым scFv-фрагментом и указанной спейсерной областью и/или между указанной спейсерной областью и упомянутым вторым scFv-фрагментом (а) может присутствовать линкерная последовательность (последовательности). Предпочтительные линкерные последовательности раскрыты выше. Как можно видеть из предпочтительных последовательностей, включенных в список последовательностей, в частности последовательностей SEQ ID NO: 43-46, такие линкерные последовательности могут состоять из глицинов, или глицинов и серинов.
Термины «CH2 домен» и «CH3 домен» имеют свое значение, установленное в данной области техники. Они относятся ко второму и третьему константному домену тяжелых цепей антител.
Обращаясь к предпочтительным молекулярным архитектурам, показанным на прилагаемых фигурах 1 и 2, димеризация и мультимеризация, соответственно, предпочтительно происходят следующим образом. Гомодимеризацию идентичных полипептидных цепей, таких как форматы B, D, G, H, L, M и O, предпочтительно осуществляют путем образования межмолекулярных дисульфидных связей шарнирного домена человеческого IgG1. То же самое относится к гетеродимеризации полипептидных цепей различных аминокислотных последовательностей, как в формате C. В форматах B и C, CH2 и CH3-домены вносят вклад с помощью нековалентных взаимодействий в стабильность димера. В форматах G, H, N и O природный межмолекулярный дисульфидный мостик между Cκ и доменом CH1 вносит свой вклад в ковалентный механизм димеризации, а также нековалентное взаимодействие между CH2 и CH3 доменами. Первый из этих двух механизмов способствует также димеризации и форматам D, L и M.
Для целей гетеродимеризации полипептидов с различными аминокислотными последовательностями, таких как в форматах A, E, F и K, предпочтение отдается вышеупомянутым доменам лейциновой застежки, также известным как домены димеризации амфипатической спирали. Примеры таких доменов димеризации описаны в deKruif and Logtenberg, J. Biol. Chem. 271, 7630-7634 (1996) и Arndt et al., J. Mol. Biol. 312, 221-228 (2001). В форматах A и E домены CH2 и CH3 способствуют димеризации. В формате F межмолекулярный дисульфидный мостик между Cκ и CH1-доменом, а также нековалентное взаимодействие между CH2 и CH3 доменами человеческого IgG1 способствуют димеризации.
В еще одном предпочтительном варианте осуществления один, несколько или все из указанных полипептидов состоят из одной или нескольких молекул scFv, или содержат их. Этот вариант осуществления распространяется на тандемные scFv-молекулы, также обозначенные (scFv)2, а также молекулы (scFv)3, такие как структура (VII).
Этот предпочтительный вариант осуществления определяет предпочтительные составные элементы полипептидов, входящих в связывающие молекулы из настоящего изобретения. Полипептиды могут либо состоять, либо содержать эти предпочтительные составляющие молекулы. Например, когда полипептид должен использоваться для обеспечения единственной специфичности связывания, это может быть достигнуто с использованием полипептида, состоящего из молекулы scFv. С другой стороны, когда необходимо использовать полипептид, обеспечивающий две специфичности связывания, это может быть осуществлено полипептидом, содержащим две молекулы scFv, или диателом.
Термин «scFv» хорошо известен в данной области техники. Аббревиатура означает «одноцепочечный вариабельный фрагмент» антитела и определяет полипептид, способный специфически распознавать и связывать эпитоп антигена. Как отмечено выше, три CDR представлены вариабельным доменом легкой цепи антитела (VL), и три CDR представлены вариабельным доменом тяжелой цепи (VH) антитела. Поскольку VL и VH не ковалентно связаны, они составляют Fv-фрагмент. В scFv, с другой стороны, два вариабельных домена связаны друг с другом пептидным линкером. Полученная гибридная конструкция представляет собой одну полипептидную цепь. Это обеспечивает легкую экспрессию молекулы scFv.
Термины «VH домен» и «VL домен» известны в данной области техники. Таким образом, они относятся к вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL) иммуноглобулинов, соответственно. Обычно VH и VL домены содержат три гипервариабельных области каждый (CDR) каждый.
Термины «диантитело», «триантитело» и «тетраантитело» известны в данной области и относятся к одноцепочечным конструкциям, которые содержат два, три или четыре фрагмента Fv. Вообще говоря, диантитела, триантитела и тетраантитела состоят из двух, трех или четырех полипептидных цепей. Они могут быть связаны друг с другом таким образом, что получается единственная полипептидная цепь. Обзор конструкций рекомбинантных антител представлен Little et al., Immunology Today, 21, 364-370 (2000).
Пептидный линкер предпочтительно используют для связывания любых вариабельных областей scFv или для связывания scFv с областью димеризации/мультимеризации и/или спейсерной областью, или с Fc, как определено ниже. Аналогичным образом, короткие пептидные линкеры могут быть использованы при конструировании диантител, триантител и тетраантител, как показано в вышеупомянутом обзоре.
Обычно пептидные линкеры имеют длину от 3 до 30 аминокислот, предпочтительно от 5 до 25 или 10 и 20 аминокислот. Предпочтение отдается тем линкерам, которые не влияют, или не оказывают существенного влияния на структуру и/или функцию доменов или полипептидов, которые их соединяют (соединение дает единственную непрерывную полипептидную цепь). Линкеры включают богатые глицином линкеры, такие как линкер (Гли4Сер)3 (SEQ ID NO: 47), который используется в предпочтительных полипептидах по изобретению для связывания VH/VL доменов ЦТЛ- или NK-специфических scFvs, и Yol-линкер (SEQ ID NO: 48, AKTTPKLEEGEFSEARV, как описано в Sellrie et al., Journal of Biochemistry and Molecular Biology, Vol. 40, No. 6, November 2007, pp. 875-880), который используют в предпочтительных полипептидах по изобретению для соединения VH/VL доменов scFvs, специфичных к поверхностным антигенам HVB. Кроме того, линкер (Гли4Сер)3 (SEQ ID NO: 49) может быть использован для соединения VH/VL доменов scFvs, специфичных к поверхностным антигенам HVB, или других строительных блоков связывающих молекулы по настоящему изобретению.
В еще одном предпочтительном варианте осуществления, относящемся к тем связывающим молекулам, которые являются мультимерами полипептидов, указанные мультимеры предпочтительно представляют собой димеры или тетрамеры, причем указанная связывающая молекула предпочтительно представляет собой триспецифическую связывающую молекулу, где один или несколько или все участки связывания обеспечены двумя указанными полипептидами вместе.
Структурный фон для этого предпочтительного варианта осуществления был описан выше. В частности, этот предпочтительный вариант осуществления предусматривает два домена иммуноглобулина, обеспечивающие два набора из трех CDR, составляющих участок связывания, расположенный на разных полипептидных цепях.
В особенно предпочтительном варианте осуществления указанная связывающая молекула содержит Fv-фрагмент; Fab фрагмент; диантитело; триантитело; тетраантитело; или молекулу иммуноглобулина G, при необходимости с модифицированной Fc частью.
Термины, используемые в связи с этим особо предпочтительным вариантом осуществления, имеют значение, установленное в данной области техники. В частности, термин «Fv фрагмент» обозначает нековалентный комплекс из двух доменов иммуноглобулина, причем указанные иммуноглобулиновые домены являются вариабельным доменом легкой цепи и вариабельным доменом тяжелой цепи.
Со структурной точки зрения, Fv фрагмент содержится в Fab-фрагменте, где Fab-фрагмент дополнительно содержит константный домен CL легкой цепи и первый постоянный домен тяжелой цепи CH1 антитела. В целом Fab-фрагмент содержит четыре домена иммуноглобулина, причем два из указанных четырех доменов иммуноглобулина, а именно VL и VH-домен, образуют вместе участок связывания. Fab-фрагменты могут быть получены из антитела путем расщепления ферментом папаином.
Когда связывающая молекула согласно изобретению содержит Fv фрагмент в определенном месте, понятно, что в этом конкретном месте он не имеет соседнего константного домена легкой цепи и не имеет соседнего константного домена 1 тяжелой цепи, отмечая, что в противном случае он будет Fab фрагментом в рассматриваемом месте. Аналогично, требование для Fab фрагмента исключает присутствие CH2 и CH3 домена в рассматриваемом месте. «Смежный» в этом контексте относится к близости, которая обнаруживается во встречающихся в природе антителах.
Модификации в Fc-части указанной молекулы иммуноглобулина G включают модификации, которые уменьшают или отменят связывание с Fc-рецепторами; см. ниже.
Предпочтительно, чтобы в соответствии с настоящим изобретением связывающие молекулы были полностью лишены Fc участков. Также предпочтительным является присутствие Fc частей, где указанные Fc участки модифицированы, как описано выше, или, кроме того, модифицированы так, что Fc часть не является N-гликозилированной. Наконец, также могут использоваться нативные Fc части.
Модификация, которая предусматривает уменьшение связывания Fc и/или уменьшение активации комплемента, представляет собой замену Fc, полученного из hIgG1, на Fc из hIgG2 или hIgG4.
Термин «антитело», используемый в настоящей заявке, имеет значение, установленное в данной области техники. Предпочтительно, он относится к моноклональному антителу. Моноклональные антитела могут быть получены, например, с помощью методик, описанных в Köhler and Milstein, Nature 256 (1975), 495 и Galfré, Метh. Enzymol. 73 (1981), 3, которые включают гибридизацию клеток миеломы мыши с клетками селезенки, полученными от иммунизированных млекопитающих, с модификациями, разработанными в данной области техники. Кроме того, антитела или их фрагменты, направленные на вышеупомянутые поверхностные белки HVB, могут быть получены с использованием способов, которые описаны, например, в Harlow and Lane «Antibodies, A Laboratory Manual», CSH Press, Cold Spring Harbor, 1988 («Антитела, лабораторное руководство»). Производство химерных антител описано, например, в WO89/09622. Другим источником антител, используемых в соответствии с настоящим изобретением, являются так называемые ксеногенные антитела. Общий принцип получения ксеногенных антител, таких как человеческие антитела у мышей, описан, например, в WO 91/10741, WO 94/02602, WO 96/34096 и WO 96/33735. Антитела, используемые в соответствии с изобретением, или их соответствующая иммуноглобулиновая цепь (цепи), могут быть дополнительно модифицированы с использованием общепринятых методов, известных в данной области техники, например, с использованием делеции (делеций), вставки (вставок), замены (замен), добавления (добавлений) и или перегруппировки (перегруппировок) аминокислот, и/или любой другой модификации (модификаций), известных в данной области, по отдельности или в комбинации. Способы введения таких модификаций в ДНК или полипептидную последовательность, лежащую в основе аминокислотной последовательности цепи иммуноглобулина, хорошо известны специалисту в данной области техники; см., например, Sambrook, «Molecular Cloning: A Laboratory Manual», Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989 («Молекулярное клонирование: лабораторное руководство»). Модификации полипептидов включают также посттрансляционные модификации, такие как гликозилирование.
В предпочтительном варианте осуществления, относящемся к тем связывающим молекулам настоящего изобретения, которые являются мультимерами полипептидов, и где каждый полипептид является биспецифическим полипептидом, первая специфичность каждого биспецифического полипептида представляет собой первую специфичность, определенную в соответствии с первым аспектом, а вторая специфичность каждого полипептида является специфичностью либо для CD3, либо для CD28.
Соответственно, необходимо понять, что предпочтение отдается составляющим биспецифическим полипептидам, которые разделяют способность к связыванию поверхностного антиген HVB, и которые отличаются друг от друга, в зависимости от их специфичности к поверхностному маркеру ЦТЛ.
Особенно предпочтительными связывающими молекулами в этом отношении являются димеры и тетрамеры полипептидов. В пределах этой группы димеров, которая состоит из димеров биспецифических двухвалентных полипептидов, предпочтение отдается нековалентным димерам.
Особенно предпочтительными димерами являются структуры (I), (II) и (III), а особенно предпочтительным тетрамером является структура (IV):
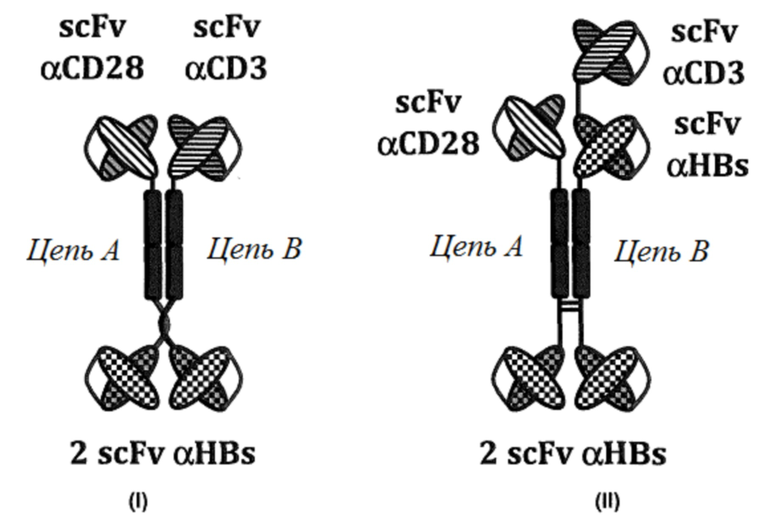
«Цепь А» и «Цепь В» обозначают полипептиды с различными аминокислотными последовательностями. «Цепь А» и «Цепь A'» обозначают различные полипептиды с идентичными аминокислотными последовательностями. Специфичности указаны вблизи каждого участка связывания. Как принято в данной области техники, термин «анти» сокращен как «α».
Структура (I) представляет собой нековалентный димер, состоящий из двух биспецифических двухвалентных полипептидов. Структура (I) показана на фигуре 1, «Формат A».
Структура (II) представляет собой ковалентный димер двух полипептидов, где полипептид, обозначенный как «цепь А», представляет собой биспецифический двухвалентный полипептид, а полипептид, обозначенный как «цепь В», представляет собой биспецифический трехвалентный полипептид. Структура (II) показана на фигуре 1, «Формат C».
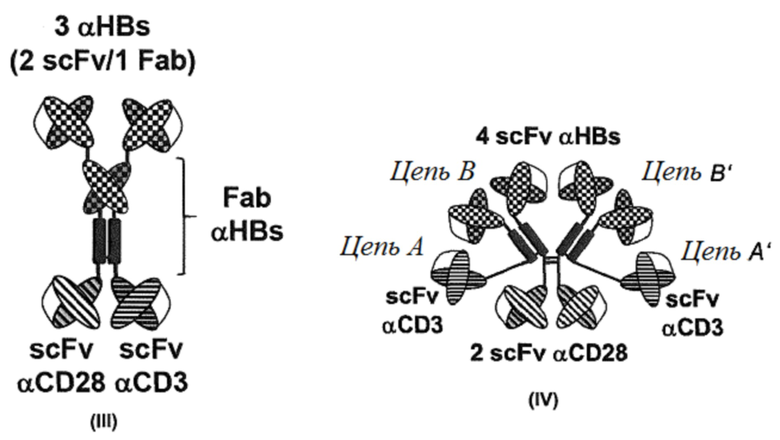
Мы отмечаем, что структура (III) имеет четыре участка связывания, каждый из которых обеспечен двумя иммуноглобулиновыми доменами, причем указанные два иммуноглобулиновых домена расположены в одной и той же полипептидной цепи, и пятый участок связывания, который образован двумя иммуноглобулиновыми доменами, расположенными на различных полипептидных цепях. Примеры включают данные для структуры (III) связывающей молекулы из настоящего изобретения.
Структура (IV) показана на фигуре 1, «Формат D». Примеры включают данные для структуры (IV) связывающей молекулы по изобретению.
В еще одном предпочтительном варианте осуществления связывающей молекулы по настоящему изобретению упомянутая связывающая молекула предпочтительно является триспецифической связывающей молекулой, каждый полипептид представляет собой триспецифическую связывающую молекулу.
Другими словами, каждый составной полипептид уже содержит три специфичности в соответствии с первым аспектом настоящего изобретения. С использованием мультимера триспецифических полипептидов, предпочтительно димера, связывающие свойства могут быть доработаны или оптимизированы.
Особенно предпочтительным в этом отношении является димер из идентичных полипептидов, который наиболее предпочтительно имеет структуру (V):
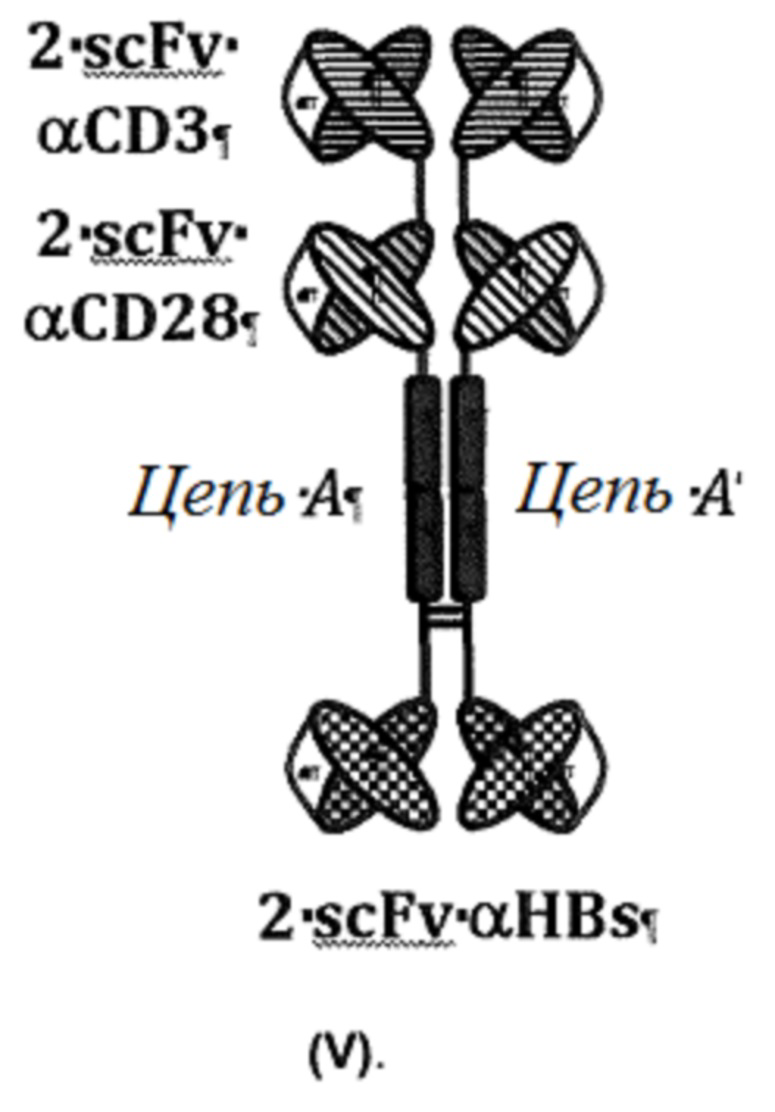
Структура (V) иллюстрирована на фигуре 1, «Формат В».
Настоящее изобретение также относится к связывающим молекулам, где участки связывания образуются исключительно с помощью иммуноглобулиновых доменов, расположенных на разных полипептидных цепях. Примером является связывающая молекула, имеющая структуру (VI):
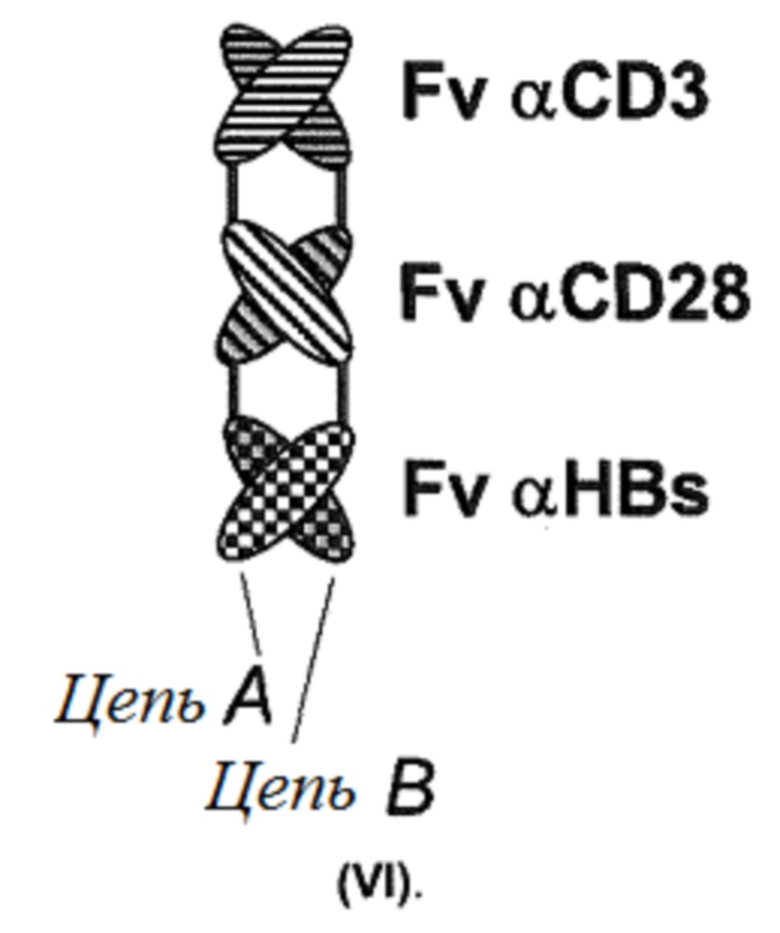
Структура (VI) представляет собой триантитело. Структура (VI) показана на фигуре 2, «Формат J».
Для обеспечения функциональности связывающей молекулы, имеющей структуру (VI), также может быть использована другая конфигурация. Соответственно, настоящее изобретение в еще одном предпочтительном варианте осуществления распространяется на связывающие молекулы, имеющие структуру (VII):
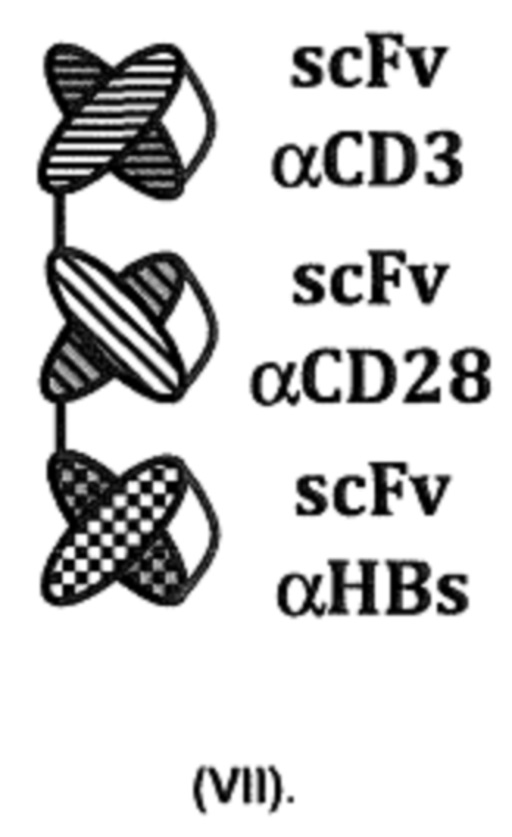
Структура (VII) состоит из одного полипептида. Он реализует описанную выше архитектуру (scFv)3. Структура (VII) показана на фигуре 2, «Формат I».
Предпочтительные варианты осуществления настоящего изобретения показаны на фигуре 1. Антигены указаны с целью иллюстрации. Понятно, что антигены не являются частью связывающих молекул в соответствии с настоящим изобретением. Дополнительные молекулярные архитектуры в соответствии с настоящим изобретением показаны на фигуре 2.
Вышеупомянутая первая специфичность предпочтительно обеспечивается набором из шести CDR, причем набор из шести CDR предпочтительно представляет собой последовательность SEQ ID NO: от 1 до 6, от 7 до 12 или от 13 до 18.
Указанная выше вторая специфичность связывающих молекул по изобретению, направленная на CD3, предпочтительно обеспечивается набором из шести CDR, имеющих последовательности SEQ ID NO: 19-24.
Указанная выше третья специфичность связывающих молекул по изобретению, направленная на CD28, предпочтительно обеспечивается набором из шести CDR, имеющих последовательности SEQ ID NO: 25-30.
В той степени, в которой вторая и третья специфические особенности выбраны из специфичностей CD16, CD56, NKp30, NKp46, 4-1BB и NKG2D, предпочтение отдается второй специфичности для CD16, а третьей - для CD56. Предпочтительными соответствующими наборами из шести CDR являются наборы из SEQ ID NO: 31-36 и 37-42, соответственно.
C8, 5F9, 5A19, OKT3, 9.3, A9 и NCAM29.2, используемые в перечне последовательностей, обозначают антитело, из которого происходят соответствующие CDR, и относятся к предпочтительному анти-HBs-антителу, ко второму другому анти-HBs-антителу, к антителу против большого поверхностного антигена HVB, к антителу против человеческого CD3, к антителу против человеческого CD28, к антителу против человеческого CD16 и к антителу против человеческого CD56, соответственно. «HBs» обозначает малый поверхностный антиген HVB.
В той степени, в которой используют биспецифические двухвалентные полипептиды, особенно предпочтительно, чтобы указанный полипептид содержал или состоял из аминокислотной последовательности любой из SEQ ID NO: 43-46 или аминокислотной последовательности, которая имеет по меньшей мере 80% идентичности с любой из SEQ ID NO: 43-46 при условии, что CDR указанной аминокислотной последовательности, имеющей идентичность по меньшей мере на 80%, идентичны тем, которые содержатся в любой из SEQ ID NO: 43-46, соответственно. В SEQ ID NO: 43 последние три остатка «GNS» являются необязательными.
Предпочтительные уровни идентичности последовательности включают по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% и по меньшей мере 99%. Средства и способы определения идентичности последовательностей хорошо известны в данной области техники. Предпочтительным алгоритмом определения идентичности парной последовательности является базовый инструмент поиска локального выравнивания (BLAST), как описано, например, в McGinnis and Madden (Nucleic Acid Research 32, W20-W25 (2004)).
Расположение указанных CDR в данной последовательности, в этом случае в последовательностях SEQ ID NO: 43-46, может быть определено с помощью способов, хорошо известных в данной области техники, включая системы Chothia, Kabat и LeFranc/IMGT, соответственно. При отсутствии каких-либо указаний на противоположное, понимается, что CDR в соответствии с вышеприведенным особо предпочтительным вариантом осуществления являются такими, как определено выше, а именно первый набор, имеющий последовательности SEQ ID NO: 1-6, 7-12 или 13-18, и второй набор, имеющий последовательности SEQ ID NO: 19-24, 25-30, 31-36 или 37-42. Как можно видеть из последовательностей, включенных в приложенный список последовательностей, эти специфические CDR-последовательности (подчеркнутые в последовательностях, воспроизведенных ниже) действительно включены в последовательности SEQ ID NO: 43-46.
Последовательности SEQ ID NO: 1-6 определяют CDR, а SEQ ID NO: 37-40 определяют биспецифические двухвалентные полипептиды, способные связывать специфический эпитоп в пределах малого поверхностного антигена HVB. Этот эпитоп расположен в а-детерминанте, которая подвергается воздействию поверхности инфицированных клеток и вирионов, соответственно. Термин «a-детерминанта» используется для обозначения области внутри малого поверхностного антигена HVB, где расположены основные эпитопы для индукции защитного гуморального иммунного ответа. Эти CDR, а также полипептиды SEQ ID NO: 43-46 имеют то преимущество, что их могут использовать для всех серотипов HVB.
Во втором аспекте настоящее изобретение относится к набору нуклеиновых кислот, кодирующих полипептиды, составляющие связывающую молекулу, причем связывающая молекула представляет собой мультимер полипептидов. Указанный второй аспект, в альтернативе, относится к нуклеиновой кислоте, кодирующей связывающую молекулу, которая представляет собой одиночный полипептид. Предпочтительные варианты осуществления полипептидов приводят к соответствующим предпочтительным вариантам осуществления указанной нуклеиновой кислоты.
Термин «нуклеиновая кислота» имеет свое установленное в данной области техники значение и особо не ограничен. Предпочтительными являются ДНК, такие как геномная ДНК или кДНК, а также РНК, такая как мРНК. Хотя это и не является предпочтительным, предполагается использование нуклеотидных производных, включающих 2'-дериватизированные нуклеотиды, такие как 2'-метилнуклеотиды; пептидных нуклеотидов, встречающихся в пептидных нуклеиновых кислотах, и тому подобных.
В следующем аспекте настоящее изобретение относится к фармацевтической композиции, содержащей или состоящей из одной или нескольких связывающих молекул по первому аспекту и/или одной или нескольких нуклеиновых кислот по второму аспекту.
Фармацевтическая композиция может дополнительно содержать фармацевтически пригодные носители, наполнители и/или разбавители. Примеры подходящих фармацевтических носителей, наполнителей и/или разбавителей хорошо известны в данной области техники, и включают фосфатные буферно-солевые растворы, воду, эмульсии, такие как эмульсии масло/вода, различные типы смачивающих агентов, стерильные растворы и т.д. Композиции, содержащие такие носители, могут быть изготовлены известными общепринятыми способами. Эти фармацевтические композиции можно применять у субъекта в подходящей дозе. Применение подходящих композиций можно осуществлять различными способами, например, путем внутривенного, подкожного или перорального введения, причем эти три варианта являются предпочтительными, а кроме того, посредством интраперитонеального, внутримышечного, местного, интрадермального, интраназального или интрабронхиального введения. Препараты для перорального введения включают таблетки и сиропы. Особенно предпочтительно, указанное введение осуществляют путем инъекции. Композиции также могут быть введены непосредственно в целевой участок, например, путем биолистической доставки к внешнему или внутреннему целевому участку. Режим дозировки будет определяться лечащим врачом и клиническими факторами. Как известно в медицине, дозировки для любого одного пациента зависят от многих факторов, включая рост и массу тела пациента, площадь поверхности тела, возраст, конкретное вводимое соединение, пол, время и путь введения, общее состояние здоровья и другие лекарства, применяемые одновременно. Белковое фармацевтически активное вещество может присутствовать в количествах от 1 нг до 10 мг / кг массы тела на дозу; однако предусматриваются дозы ниже или выше этого примерного диапазона, особенно учитывая вышеупомянутые факторы. Если режим представляет собой непрерывную инфузию, доза также должна находиться в диапазоне от 1 мкг до 10 мг на килограмм массы тела в минуту.
Особо предпочтительным является внутривенное применение.
В следующем аспекте настоящее изобретение относится к одной или нескольким связывающим молекулам, нуклеиновым кислотам и/или фармацевтическим композициям из изобретения для использования в способе лечения или профилактики HVB инфекции и/или состояния, вызванного указанной HVB инфекцией, где указанное состояние, вызванное HVB инфекцией, выбрано из цирроза печени, гепатоцеллюлярной карциномы и рака печени, причем указанный рак печени характеризуется экспрессией одного или нескольких поверхностных антигенов HVB. Предпочтительно, чтобы указанная гепатоцеллюлярная карцинома характеризовалась экспрессией одного или нескольких вышеуказанных поверхностных антигенов HVB.
В следующем аспекте настоящее изобретение обеспечивает способ лечения или профилактики HVB инфекции и/или состояния, вызванного указанной HVB инфекцией, причем указанное состояние, вызванное указанной HVB инфекцией, выбрано из цирроза печени и гепатоцеллюлярной карциномы, где указанный способ включает введение терапевтически эффективного количества или профилактического количества, соответственно, одной или нескольких связывающих молекул, нуклеиновых кислот и/или фармацевтических композиций по изобретению пациенту, нуждающемуся в этом.
В одном варианте осуществления указанная фармацевтическая композиция, указанная связывающая молекула (молекулы) и/или нуклеиновая кислота (кислоты) для использования в способе лечения и указанный способ лечения, соответственно, являются такими, что указанные связывающие молекулы и/или нуклеиновые кислоты являются единственными содержащимися или используемыми фармацевтически активными агентами.
Говоря это, также целенаправленно предполагается включать один или несколько дополнительных фармацевтически активных агентов в комбинированную терапию. Такие дополнительные фармацевтически активные агенты могут быть выбраны из интерферонов или других иммуномодуляторов (например, таких как интерферон альфа 2а или 2b, интерферон лямбда), непосредственно действующих противовирусных средств, таких как нуклеозидные (нуклеотидные) аналоги (например, такие как ламивудин (Epivir-HBV, Zeffix или Heptodin), адефовир дипивоксил (HepСерa, Preveon), энтекавир (Baraclude, Entaliv), телбивудин (Tyzeka, Sebivo), тенофовир (Viread)), ингибиторы входа (например, Myrcludex-B), других противовирусных препаратов или цитокинов, таких как интерлейкин-2.
Соответственно, предполагается, что в указанной фармацевтической композиции и в соответствии с указанной связывающей молекулой (молекулами) для использования в способе лечения и указанном способе лечения указанные связывающие молекулы и, кроме того, один или несколько из вышеуказанных определенных конкретных фармацевтически активных агентов являются единственными содержащимися и используемыми фармацевтически активными агентами.
Альтернативно, могут присутствовать и другие фармацевтически активные агенты. Дополнительные ингредиенты или фармацевтически активные агенты, которые повышают стабильность связывающей молекулы, такие как белки, например, молочные белки, питательные вещества или солевые растворы, могут быть добавлены для увеличения биодоступности и/или времени полужизни.
В следующем аспекте настоящее изобретение обеспечивает способ лизиса in vitro клеток, инфицированных HVB, включающий культивирование указанных клеток, инфицированных HVB, с (i) иммунными эффекторными клетками и (ii) одной или несколькими связывающими молекулами по изобретению.
В предпочтительном варианте осуществления способа in vitro указанные иммунные эффекторные клетки (i) включают мононуклеарные клетки периферической крови; или (ii) представляют собой или включают NK-клетки и/или ЦТЛ.
В следующем аспекте настоящее изобретение обеспечивает иммунную эффекторную клетку in vitro или ex vivo, которая (а) имеет связывающую молекулу из первого аспекта, связанную с поверхностным антигеном указанной иммунной эффекторной клетки; и/или (б) трансформированную, трансфицированную или трансдуцированную одной или несколькими нуклеиновыми кислотами в соответствии со вторым аспектом. Предпочтительные иммунные эффекторные клетки и предпочтительные поверхностные антигены, представленные иммунными эффекторными клетками, являются такими, как определено выше. Такая иммунная эффекторная клетка полезна для введения пациенту, страдающему от HVB инфекции, цирроза печени или гепатоцеллюлярной карциномы. Соответственно, также обеспечивается фармацевтическая композиция, содержащая или состоящая из иммунной эффекторной клетки, которая связана на её поверхностном антигене со связывающей молекулой по изобретению. Также обеспечивается иммунная эффекторная клетка, которая связана на ее поверхностном антигене со связывающей молекулой по изобретению для использования в способе лечения или профилактики HVB инфекции, цирроза печени или гепатоцеллюлярной карциномы.
Настоящее изобретение распространяется на некоторые варианты осуществления, которые являются менее предпочтительными и, соответственно, могут быть исключены из более широких вариантов осуществления при указании условий.
Менее предпочтительной в соответствии с настоящим изобретением является триспецифическая и четырехвалентная связывающая молекула, состоящая из первого и второго полипептидов, причем указанные первый и второй полипептиды являются биспецифическими и двухвалентными, где первая специфичность любого полипептида относится к поверхностному антигену HVB, выбранному из малого поверхностного антигена HVB, среднего поверхностного антигена HVB и большого поверхностного антигена HVB, а вторая специфичность является специфичностью для CD3 и CD28, соответственно.
В частности, менее предпочтительной является триспецифическая и четырехвалентная связывающая молекула, состоящая из первого и второго полипептидов, причем каждый из указанного первого и второго полипептидов является биспецифическим и двухвалентным, где существует по меньшей мере одна ковалентная связь между указанным первым и указанным вторым полипептидом, предпочтительно по меньшей мере один дисульфидный мостик между остатком Цис указанного первого полипептида и остатком Цис указанного второго полипептида, и где первая специфичность любого полипептида относится к поверхностному антигену HVB, выбранному из малого поверхностного антигена HVB, среднего поверхностного антигена HVB и большого поверхностного антигена HVB, а вторая специфичность является специфичностью для CD3 и CD28, соответственно.
В особенности, менее предпочтительной является триспецифическая и четырехвалентная связывающая молекула, состоящая из первого и второго полипептидов, где каждый из указанного первого и второго полипептидов является биспецифическим и двухвалентным, где существует две дисульфидные связи между указанным первым и указанным вторым полипептидом, и где первая специфичность любого полипептида относится к поверхностному антигену HVB, выбранному из малого поверхностного антигена HVB, среднего поверхностного антигена HVB, и большого поверхностного антигена HBV, а вторая специфичность является специфичностью для CD3 и CD28, соответственно.
В отношении каждого из вышеописанных менее предпочтительных вариантов осуществления особенно менее предпочтительно, чтобы участки связывания первого и второго полипептидов были обеспечены scFv-фрагментами. Кроме того, особенно менее предпочтительно по отношению к любому из предыдущих менее предпочтительных вариантов осуществления, чтобы в обоих из указанных первого и второго полипептидов имелся CH2 и CH3 домен между двумя scFv-фрагментами, обеспечивающими участки связывания.
Соответственно, особенно менее предпочтительной является триспецифическая и четырехвалентная связывающая молекула, состоящая из первого и второго полипептидов, причем каждый первый и второй полипептиды являются биспецифическими и двухвалентными, причем между указанным первым и вторым полипептидом существуют две дисульфидные связи, где первая специфичность каждого полипептида относится к поверхностному антигену HBV, выбранному из малого поверхностного антигена HVB, среднего поверхностного антигена HVB и большого поверхностного антигена HVB, а вторая специфичность является специфичностью для CD3 и CD28, соответственно, где участки связывания на первом и втором полипептидах представлены scFv-фрагментами, и где в обоих из указанных первого и второго полипептидов имеется CH2 и CH3 домен между двумя scFv фрагментами, обеспечивающими участки связывания.
Таким образом, для примера, настоящее изобретение также обеспечивает связывающую молекулу, содержащую по меньшей мере три специфичности связывания, где (а) первая специфичность является специфичностью для поверхностного антигена вируса гепатита В (HVB), выбранного из малого поверхностного антигена HVB, среднего поверхностного антигена HVB и большого поверхностного антигена HVB; (b) (i) вторая и третья специфичность являются специфичностями для CD3 и CD28, соответственно; или (ii) вторая и третья специфичность выбраны из специфичностей к CD16, CD56, NKp30, NKp46, 4-1BB и NKG2D; и (c) каждая специфичность связывания обеспечивается одним или несколькими участками связывания, причем каждый участок связывания независимо обеспечивается посредством (i) набора из шести гипервариабельных областей (CDR), где указанный набор из шести CDR состоит из первого набора из трех CDR и второго набора из трех CDR, причем указанный первый и указанный второй набор каждый входит в иммуноглобулиновый домен; или (ii) набора из трех CDR, где указанный набор из трех CDR содержится в иммуноглобулиновом домене; при условии, что триспецифические и четырехвалентные связывающие молекулы состоят из первого и второго полипептидов, где каждый из первого и второго полипептидов является биспецифическим и двухвалентным, где существует две дисульфидные связи между указанным первым и указанным вторым полипептидом, причем первая специфичность любого полипептида является специфичностью к поверхностному антигену HVB, выбранному из малого поверхностного антигена HVB, среднего поверхностного антигена HVB и большого поверхностного антигена HVB; а вторая специфичность является специфичностью для CD3 и CD28, соответственно; где участки связывания первого и второго полипептидов представлены scFv-фрагментами, и где в обоих из указанных первого и второго полипептидов имеется CH2 и CH3 домен между двумя scFv фрагментами, обеспечивающими участки связывания, исключаются.
Что касается вариантов осуществления, охарактеризованных в этом описании, в частности в формуле изобретения, предполагается, что каждый вариант осуществления, упомянутый в зависимом пункте формулы изобретения, объединяется с каждым вариантом осуществления каждого пункта формулы изобретения (независимого или зависимого), от которого зависит зависимый пункт. Например, в случае независимого пункта 1, указывающего 3 альтернативы A, B и C, зависимого пункта 2, излагающего 3 альтернативы D, E и F, и пункта 3, зависимого от пунктов 1 и 2 формулы и перечисляющего 3 альтернативы G, H и I, следует понимать, что в описании однозначно раскрыты варианты осуществления, соответствующие комбинациям A, D, G; A, D, H; A, D, I; A, E, G; A, E, H; A, E, I; A, F, G; A, F, H; A, F, I; B, D, G; B, D, H; B, D, I; B, E, G; B, E, H; B, E, I; B, F, G; B, F, H; B, F, I; C, D, G; C, D, H; C, D, I; C, E, G; C, E, H; C, E, I; C, F, G; C, F, H; C, F, I, если специально не указано иное.
Аналогичным образом, а также в тех случаях, когда независимые и/или зависимые пункты формулы изобретения не декларируют альтернативы, понятно, что если зависимые пункты формулы ссылаются на множество предшествующих пунктов формулы изобретения, любая комбинация предмета, охватываемого им, считается явно раскрытой. Например, в случае независимого пункта 1 формулы изобретения, зависимого пункта 2, ссылающегося на пункт 1 формулы изобретения, и зависимого пункта 3, ссылающегося на оба пункта 2 и 1, следует, что комбинация предмета обсуждения по пунктам 3 и 1 ясно и недвусмысленно раскрывается, как комбинация предмета по пунктам 3, 2 и 1 формулы изобретения. В случае присутствия дополнительного зависимого пункта 4, который относится к любому из пп.1-3, следует, что комбинация предмета обсуждения по пунктам 4 и 1, пунктам 4, 2 и 1, пунктам 4, 3 и 1, а также по пунктам 4, 3, 2 и 1 четко и недвусмысленно раскрыта.
Последовательности, раскрытые в настоящей заявке.
SEQ ID NO 1
C8 HC CDR1
Гли Фен Тре Фен Сер Гли Тир Ала
SEQ ID NO 2
C8 HC CDR2
Иле Сер Гли Сер Гли Гли Сер Тре
SEQ ID NO 3
C8 HC CDR3
Ала Лиз Про Про Гли Арг Глн Глу Тир Тир Гли Сер Сер Иле Тир Тир Фен Про Лей Гли Асн
SEQ ID NO 4
C8 LC CDR1
Асн Иле Гли Сер Лиз Сер
SEQ ID NO 5
C8 LC CDR2
Асп Асп Сер
SEQ ID NO 6
C8 LC CDR3
Глн Вал Трп Асп Сер Сер Сер Асп Лей Вал Вал
SEQ ID NO 7
5F9 HC CDR1
Гли Фен Тре Фен Асн Асн Тир Ала
SEQ ID NO 8
5F9 HC CDR2
Иле Асн Сер Асп Гли Арг Сер Тре
SEQ ID NO 9
5F9 HC CDR3
Ала Арг Тре Фен Тир Ала Асп Тир
SEQ ID NO 10
5F9 LC CDR1
Глн Асн Вал Асп Тре Тре
SEQ ID NO 11
5F9 LC CDR2
Трп Ала Сер
SEQ ID NO 12
5F9 LC CDR3
Глн Глн Тир Сер Иле Фен Про Тир Тре
SEQ ID NO 13
5A19 HC CDR1
Гли Фен Тре Фен Сер Сер Тир Ала
SEQ ID NO 14
5A19 HC CDR2
Вал Сер Сер Асп Гли Сер Тир Ала
SEQ ID NO 15
5A19 HC CDR3
Ала Сер Фен Асн Трп Асп Вал Ала Тир
SEQ ID NO 16
5A19 LC CDR1
Глн Сер Лей Лей Асн Тре Арг Тре Арг Лиз Сер Тир
SEQ ID NO 17
5A19 LC CDR2
Трп Ала Сер
SEQ ID NO 18
5A19 LC CDR3
Лиз Глн Сер Тир Сер Лей Тир Тре
SEQ ID NO 19
OKT3 HC CDR1
Гли Тир Тре Фен Тре Арг Тир Тре
SEQ ID NO 20
OKT3 HC CDR2
Иле Асн Про Сер Арг Гли Тир Тре
SEQ ID NO 21
OKT3 HC CDR3
Ала Арг Тир Тир Асп Асп ГисТир Цис Лей Асп Тир
SEQ ID NO 22
OKT3 LC CDR1
Сер Сер Вал Сер Тир
SEQ ID NO 23
OKT3 LC CDR2
Асп Тре Сер
SEQ ID NO 24
OKT3 LC CDR3
Глн Глн Трп Сер Сер Асн Про Фен Тре
SEQ ID NO 25
9.3 HC CDR1
Гли Фен Сер Лей Сер Асп Тир Гли
SEQ ID NO 26
9.3 HC CDR2
Иле Трп Ала Гли Гли Гли Тре
SEQ ID NO 27
9.3 HC CDR3
Ала Арг Асп Лиз Гли Тир Сер Тир Тир Тир Сер Мет Асп Тир
SEQ ID NO 28
9.3 LC CDR1
Глу Сер Вал Глу Тир Тир Вал Тре Сер Лей
SEQ ID NO 29
9.3 LC CDR2
Ала Ала Сер
SEQ ID NO 30
9.3 LC CDR3
Глн Глн Сер Арг Лиз Вал Про Тир Тре
SEQ ID NO 31
A9 HC CDR1
Гли Тир Тре Фен Тре Асн Тир Трп
SEQ ID NO 32
A9 HC CDR2
Иле Тир Про Гли Гли Гли Тир Тре
SEQ ID NO 33
A9 HC CDR3
Ала Арг Сер Ала Сер Трп Тир Фен Асп Вал
SEQ ID NO 34
A9 LC CDR1
Тре Гли Тре Вал Тре Тре Сер Асн Тир
SEQ ID NO 35
A9 LC CDR2
ГисТре Асн
SEQ ID NO 36
A9 LC CDR3
Ала Лей Трп Тир Асн Асн ГисТрп Вал
SEQ ID NO 37
NCAM29.2 HC CDR1
Гли Фен Тре Фен Сер Сер Фен Гли
SEQ ID NO 38
NCAM29.2 HC CDR2
Иле Сер Сер Гли Сер Тир Ала Иле
SEQ ID NO 39
NCAM29.2 HC CDR3
Вал Арг Гли Арг Арг Лей Гли Глу Гли Тир Ала Мет Асп Тир
SEQ ID NO 40
NCAM29.2 LC CDR1
Глн Сер Лей Лей Тир Сер Сер Асн Глн Лиз Асн Тир
SEQ ID NO 41
NCAM29.2 LC CDR2
Трп Ала Сер
SEQ ID NO 42
NCAM29.2 LC CDR3
Глн Глн Тир Сер Сер Трп Тре
SEQ ID NO 43
C8-hIgG1Fcmut-OKT3
MDFEVQIFSFLLISASVIMSRMAEVQLVESGGGLLQPGGSLRLSCAASGFTFSGYAMSWVRQAPGKGLEWVSSISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTALYYCAKPPGRQEYYGSSIYYFPLGNWGQGTLVTVSSASTKGPKLEEGEFSEARVQSALTQPASVSVAPGQTARITCGGNNIGSKSVHWYQQKPGQAPVLVVYDDSDRPSGIPERFSGSNSGNTATLTISRVEAGDEADYYCQVWDSSSDLVVFGGGTKLTVLGNSGGGGSGGGGSGGGGSASEPKSSDKTHTCPPCPAPPAAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSТИРVVSVLTVLHQDWLNGKEYKCAVSNKGLASSIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFЛИЗKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKDPGWSHPQFEKSRGGGGQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSGNSGGGGSGGGGSGGGGSASQIVLTQSPAIMSАСПGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEINGNS
SEQ ID NO 44
C8- hIgG1Fcmut-9.3
MDFEVQIFSFLLISASVIMSRMAEVQLVESGGGLLQPGGSLRLSCAASGFTFSGYAMSWVRQAPGKGLEWVSSISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTALYYCAKPPGRQEYYGSSIYYFPLGNWGQGTLVTVSSASTKGPKLEEGEFSEARVQSALTQPASVSVAPGQTARITCGGNNIGSKSVHWYQQKPGQAPVLVVYDDSDRPSGIPERFSGSNSGNTATLTISRVEAGDEADYYCQVWDSSSDLVVFGGGTKLTVLGNSGGGGSGGGGSGGGGSASEPKSSDKTHTCPPCPAPPAAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSТИРVVSVLTVLHQDWLNGKEYKCAVSNKGLASSIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFЛИЗKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKDPGWSHPQFEKSSGGGGQVQLQESGPGLVTPSQSLSITCTVSGFSLSDYGVHWVRQSPGQGLEWLGVIWAGGGTNYNSALMSRKSISKDNSKSQVFLKMNSLQADDTAVYYCARDKGYSYYYSMDYWGQGTTVTVSSRGGGSGGGGSGGGGSDIELTQSPASLAVSLGQRATISCRASESVEYYVTSLMQWYQQKPGQPPKLLIFAАСНVESGVPARFSGSGSGTNFSLNIHPVDEDDVAMYFCQQSRKVPYTFGGGTKLEIKR
SEQ ID NO 45
C8- hIgG1Fcmut-A9
MDFEVQIFSFLLISASVIMSRMAEVQLVESGGGLLQPGGSLRLSCAASGFTFSGYAMSWVRQAPGKGLEWVSSISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTALYYCAKPPGRQEYYGSSIYYFPLGNWGQGTLVTVSSASTKGPKLEEGEFSEARVQSALTQPASVSVAPGQTARITCGGNNIGSKSVHWYQQKPGQAPVLVVYDDSDRPSGIPERFSGSNSGNTATLTISRVEAGDEADYYCQVWDSSSDLVVFGGGTKLTVLGNSGGGGSGGGGSGGGGSASEPKSSDKTHTCPPCPAPPAAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSТИРVVSVLTVLHQDWLNGKEYKCAVSNKGLASSIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFЛИЗKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKDPGWSHPQFEKSRGGGGQVQLQQSGAELVRPGTSVKISCKASGYTFTNYWLGWVKQRPGHGLEWIGDIYPGGGYTNYNEKFKGKATVTADTSSRTAYVQVRSLTSEDSAVYFCARSASWYFDVWGAGTTVTVSSGNSGGGGSGGGGSGGGGSASQAVVTQESALTTSPGETVTLTCRSNTGTVTTSNYANWVQEKPDHLFTGLIGHTNNRAPGVPARFSGSLIGDKAALTITGAQTEDEAIYFCALWYNNHWVFGGGTKLTVL
SEQ ID NO 46
C8- hIgG1Fcmut-NCAM29.2
MDFEVQIFSFLLISASVIMSRMAEVQLVESGGGLLQPGGSLRLSCAASGFTFSGYAMSWVRQAPGKGLEWVSSISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTALYYCAKPPGRQEYYGSSIYYFPLGNWGQGTLVTVSSASTKGPKLEEGEFSEARVQSALTQPASVSVAPGQTARITCGGNNIGSKSVHWYQQKPGQAPVLVVYDDSDRPSGIPERFSGSNSGNTATLTISRVEAGDEADYYCQVWDSSSDLVVFGGGTKLTVLGNSGGGGSGGGGSGGGGSASEPKSSDKTHTCPPCPAPPAAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSТИРVVSVLTVLHQDWLNGKEYKCAVSNKGLASSIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFЛИЗKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKDPGWSHPQFEKSSGGGDVQLVESGGGLVQPGGSRKLSCAASGFTFSSFGMHWVRQAPEKGLEWVAYISSGSYAIYYADTVKGRFTISRDNPENTLFLQMTSLRSEDSAMYYCVRGRRLGEGYAMDYWGQGTSVTVSSGNSGGGGSGGGGSGGGGSASDIVMSQSPSSLAVSVGEKVTMSCKSSQSLЛИЗSNQKNYLAWYQQKPGQSPKLLIYWASTRKSGVPDRFTGSGSGTDFTLTISSVKAEDLAVYYCQQYSSWTFGGGTKLEIKR
SEQ ID NO 47
Гли Гли Гли Гли Сер Гли Гли Гли Гли Сер Гли Гли Гли Гли Сер
SEQ ID NO 48
Ала Лиз Тре Тре Про Лиз Лей Глу Глу Гли Глу Фен Сер Глу Ала Арг Вал
SEQ ID NO 49
Гли Гли Гли Гли Сер Гли Гли Гли Гли Сер Гли Гли Гли Гли Сер Гли Гли Гли Гли Сер
SEQ ID NO 50
C8-hIgG1Fcmut-OKT3 с Jun доменом димеризации с лейциновой застежкой (жирный шрифт, курсив, подчеркнуто)
MDFEVQIFSFLLISASVIMSRMAEVQLVESGGGLLQPGGSLRLSCAASGFTFSGYAMSWVRQAPGKGLEWVSSISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTALYYCAKPPGRQEYYGSSIYYFPLGNWGQGTLVTVSSASTKGPKLEEGEFSEARVQSALTQPASVSVAPGQTARITCGGNNIGSKSVHWYQQKPGQAPVLVVYDDSDRPSGIPERFSGSNSGNTATLTISRVEAGDEADYYCQVWDSSSDLVVFGGGTKLTVLGNSGGGGSGGGGSGGGGSASRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNYSSEPKSSDKTHTCPPCPAPPAAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSТИРVVSVLTVLHQDWLNGKEYKCAVSNKGLASSIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFЛИЗKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKDPGWSHPQFEKSRGGGGQVQLQQSGAELARPGASVKMSCKASGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSGNSGGGGSGGGGSGGGGSASQIVLTQSPAIMSАСПGEKVTMTCSASSSVSYMNWYQQKSGTSPKRWIYDTSKLASGVPAHFRGSGSGTSYSLTISGMEAEDAATYYCQQWSSNPFTFGSGTKLEINGNS
SEQ ID NO 51
C8- hIgG1Fcmut-9.3 с Fos доменом димеризации с лейциновой застежкой (жирный шрифт, курсив, подчеркнуто)
MDFEVQIFSFLLISASVIMSRMAEVQLVESGGGLLQPGGSLRLSCAASGFTFSGYAMSWVRQAPGKGLEWVSSISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTALYYCAKPPGRQEYYGSSIYYFPLGNWGQGTLVTVSSASTKGPKLEEGEFSEARVQSALTQPASVSVAPGQTARITCGGNNIGSKSVHWYQQKPGQAPVLVVYDDSDRPSGIPERFSGSNSGNTATLTISRVEAGDEADYYCQVWDSSSDLVVFGGGTKLTVLGNSGGGGSGGGGSGGGGSASLTDTLQAETDQLEDKKSALQTEIANLLKEKEKLEFILAAYSSEPKSSDKTHTCPPCPAPPAAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSТИРVVSVLTVLHQDWLNGKEYKCAVSNKGLASSIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFЛИЗKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKDPGWSHPQFEKSSGGGGQVQLQESGPGLVTPSQSLSITCTVSGFSLSDYGVHWVRQSPGQGLEWLGVIWAGGGTNYNSALMSRKSISKDNSKSQVFLKMNSLQADDTAVYYCARDKGYSYYYSMDYWGQGTTVTVSSRGGGSGGGGSGGGGSDIELTQSPASLAVSLGQRATISCRASESVEYYVTSLMQWYQQKPGQPPKLLIFAАСНVESGVPARFSGSGSGTNFSLNIHPVDEDDVAMYFCQQSRKVPYTFGGGTKLEIKR
На чертежах показано:
Фигуры 1 и 2 иллюстрируют предпочтительные триспецифические связывающие молекулы из настоящего изобретения.
Фигура 3. Т-клетки, активированные пятивалентными и восьмивалентными триспецифическими антителами. Определяли секрецию ИФНγ Т-клетками, культивированными в течение 48 часов на иммобилизованном HBsAg в присутствии би- и триспецифических антител. Совместное применение CD3- (bsAbαCD3) и CD28-активирующих биспецифических антител (bsAbαCD28) приводило к повышенной секреции ИФНγ в присутствии антигена, по сравнению с одиночной обработкой CD3-активирующими биспецифическими антителами. Применение пятивалентных (TriMAB) и восьмивалентных (ОКТА) конструкций из триспецифических антител индуцировало существенно более высокую секрецию интерферона, причем TriMAb была наиболее мощной конструкцией. Единичная обработка CD28-активирующими антителами, а также необработанные контрольные МНКПК не показали повышенных уровней ИФНγ.
Фигура 4. Т-клетки активировали пятивалентными и восьмивалентными триспецифическими антителами для их превращения в цитотоксические. (А) Определяли транслокацию LAMP-1, указывающую на цитотоксическую активацию Т-клеток, культивированных в течение 48 часов на иммобилизованном HBsAg в присутствии би- и триспецифических антител. Совместное применение CD3- (bsAbαCD3) и CD28-активирующих биспецифических антител (bsAbαCD28) приводило к усиленной транслокации маркера дегрануляции LAMP-1 в присутствии антигена, по сравнению с одиночной обработкой CD3-активирующими антителами. Применение пятивалентных (TriMAB) и восьмивалентных (OKTA) конструкций триспецифических антител индуцировало существенно повышенные уровни цитотоксической активации, где TriMAb была наиболее мощной конструкцией, индуцирующей дегрануляцию в 83% CD8+ T-клеток. Клетки, обработанные отдельно CD28-активирующими антителами, а также необработанные контрольные МНКПК не показали транслокации LAMP-1. (В) Экспрессия гранзима В, указывающая на цитотоксическую активацию. Конструкции триспецифических антител TriMAB и OKTA индуцировали существенно повышенные уровни цитотоксической активации, причем TriMAb была наиболее мощной конструкцией, индуцирующей экспрессию гранзима В в b 95% CD8+ T-клеток.
Фигура 5. Интактные Т-клетки от здоровых доноров, активированные TriMAb. Оценивали секрецию ИФНγ в МНКПК здоровых доноров при совместном культивировании с HBsAg-позитивными (Huh7S) или негативными (Huh7) клетками-мишенями в присутствии TriMAb. Только при добавлении TriMAb клетки-мишени экспрессировали HBsAg, и определялась активация Т-клеток, выявляемая по секреции ИФНγ.
Фигура 6. Интактные Т-клетки здоровых доноров активировали для лизиса HBSAg-позитивных клеток-мишеней посредством TriMAb. Осуществляли совместное культивирование МНКПК от здоровых доноров с HBSAg-негативными (Huh7) и HBsAg-позитивными клетками-мишенями (Huh7S). После добавления TriMAb оценивали жизнеспособность клеток посредством анализа XCelligence в трех повторностях. В контроле МНКПК клетки совместно культивировали с Huh7S клетками, но не добавляли антител. Только когда добавляли TriMAb, и клетки-мишени экспрессировали HBsAg, наблюдался специфический лизис, начиная от 24 часов после добавления антитела.
Фигура 7. Нейтрализующие анти-HBs антитела конкурируют с активацией Т-клеток TriMAb. Совместное культивирование TriMAb-обработанных МНКПК с HBsAg-позитивными Huh7 (Huh7S) проводили в присутствии человеческой сыворотки, содержащей нейтрализующие αHBs антитела. 0%: совместная культура без сыворотки, содержащей антитела; 50% - половину ЭТС заменяли человеческой сывороткой, содержащей антитела, с получением концентрации αHBs 117,5 МЕ/л; 100%: ЭТС заменяли на сыворотку, содержащую антитела, с получением концентрации αHBs 235 МЕ/л. (А) Без добавления нейтрализующих антител Т-клетки были активированы и секретировали ИФНγ и другие цитокины; и (В) Т-клетки были активированы для лизиса клеток-мишеней в типичном временном профиле, приводящем к 50% потере жизнеспособности клеток в течение 72 часов. Нейтрализующие антитела конкурировали с активностью TriMAb, приводя к частичной или даже полной потере Т-клеточной активации и цитотоксичности.
Фигура 8. МНКПК пациентов с хроническим гепатитом В вызывают лизис HBsAg-позитивных клеток-мишеней при активации TriMAb. Осуществляли совместное культивирование МНКПК от пациентов с хроническим гепатитом В (ХГВ) с HBSAg-негативными (Huh7) и HBsAg-позитивными клетками-мишенями (Huh7S). После добавления TriMAb оценивали жизнеспособность клеток с применением анализа XCelligence в течение времени в трех повторностях. В контроле МНКПК клетки совместно культивировали с Huh7S, но не добавляли антител. Только когда добавляли TriMAb, и клетки-мишени экспрессировали HBsAg, наблюдался специфический лизис, начиная спустя 24-30 часов после добавления антитела.
Фигура 9. МНКПК от пациентов с хроническим гепатитом В активируются TriMAb. Оценивали секрецию ИФНγ МНКПК пациентов с хроническим гепатитом В (ХГВ) при совместном культивировании с HBsAg-позитивными (Huh7S) или негативными (Huh7) клетками-мишенями в присутствии TriMAb. Только когда добавляли TriMAb, и клетки-мишени экспрессировали HBsAg, определялась активация Т-клеток, выявляемая по секреции ИФНγ.
Примеры иллюстрируют изобретение.
Пример 1.
Материалы и методы
Клонирование и продукция триспецифических антител
Комплементарную ДНК, кодирующую вариабельную тяжелую и вариабельную легкую цепи анти-CD3 (OKT3) и анти-CD28 (9.3), получали путем ПЦР-амплификации полученных обратной транскрипцией мРНК из соответствующей гибридомы с применением набора из праймеров, охватывающих все субтипы VH и Vκ/Vλ. Продукты ПЦР лигировали в pCR2.1-TOPO (Invitrogen, Life Technologies) и секвенировали. Отбирали анти-HBsAg scFv C8 из библиотеки кДНК В-клеток с вакциной от гепатита В с применением технологии фагового дисплея, и осуществляли оптимизацию кодонов. С применением праймеров, содержащих подходящие участки рестрикции в 5' и 3' фланкирующих областях, собирали вариабельную тяжелую и вариабельную легкую цепи кДНК, кодирующую вышеупомянутые антитела, с глицин-сериновым линкером в scFvs. Для получения пятивалентной молекулы scFvs OKT3 и 9.3 гибридизировали с С-концами Fab фрагмента антитела 5F9, а два компонента scFv C8 добавляли к его N-концам. Для получения восьмивалентной молекулы, четыре scFv C8 домена и два домена OKT3 и 9.3, соответственно, соединяли с IgG-подобной молекулой.
Последовательность, кодирующую C8 scFv, клонировали на 5' конце упомянутого 5' глицин-серинового линкера. Полную последовательность scFv-линкер-hIgG1Fc-линкер-scFv субклонировали в вектор экспрессии млекопитающих pcDNA3.1(-) (Invitrogen). Плазмидную ДНК Maxi-prep использовали для трансфекции HEK293 клеток с применением реагента для трансфекции peqFECT (Peqlab). Отбирали стабильные трансфектанты с применением 0,8-1,0 мг/мл G418, и размножали. Собирали надосадочную жидкость от НЕК трансфектантов, и анализировали посредством ИФА концентрацию секретируемых триспецифических антител, а посредством вестерн-блоттинга - целостность секретируемых антител с применением козьих антител, специфичных к IgG-Fc человека, меченых пероксидазой.
Условия культивирования клеток и генерации Huh7S клеток
Клетки гепатомы HuH7 (Nakabayaski, et al. 1982. «Growth of human hepatoma cell lines with differentiated functions in chemically defined medium». Cancer Res. 42: 3858-3863 («Выращивание клеточных линий гепатомы человека с дифференцированными функциями в химически определенной среде»)) и HEK293 клетки поддерживали в среде Игла с модификацией Дульбекко (DMEM) с добавлением 10% эмбриональной телячьей сыворотки (ЭТС), пенициллина (100 Ед./мг), стрептомицина (100 мкг/мл), и L-глутамина (2 ммоль/л) (все от GIBCO, Life Technologies).
Для генерации Huh7S клеток, Huh-7 клетки трансфицировали плазмидой, кодирующий малый поверхностный антиген HVB генотипа D подтипа ayw1 и ген устойчивости к антибиотикам, с применением реагента для трансфекции FuGene (Promega). После селекции с антибиотиками выделяли отдельные клеточные клоны и выращивали в 24-луночных планшетах. Отбирали клоны с наивысшей секрецией HBsAg, и использовали для получения клеточной линии Huh7-S, используемой в качестве клеток-мишеней.
Выделение МНКПК
Мононуклеарные клетки периферической крови (МНКПК) выделяли с применением центрифугирования в градиенте плотности из гепаринизированной цельной крови с применением среды для выделения лимфоцитов LSM 1077 (РАА). 25 мл крови наслаивали примерно на 13 мл LSM 1077. После центрифугирования при 2000 об./мин в течение 20 минут (без перерыва) при комнатной температуре МНКПК собирали и культивировали в среде RPMI 1640 с добавлением 10% эмбриональной телячьей сыворотки (ЭТС), пенициллина (100 Ед./мг), стрептомицина (100 мкг/мл), и L-глутамина (2 ммоль/л) (все от GIBCO, Life Technologies). После этапа выстаивания в течение ночи при 37°С в CO2, МНКПК использовали в экспериментах с совместным культивированием.
Культивирование МНКПК на иммобилизованном HBsAg
100 мкл рекомбинантного HBsAg в ФБР (5 мкг/мл) переносили в 96-луночный планшет и выдерживали при 4°С в течение ночи. На следующий день раствор HBsAg отделяли декантацией, и планшет промывали фосфатно-солевым буфером. 1x105 МНКПК/лунку переносили в планшет, покрытый HBsAg, и смешивали с 100 мкл надосадочной жидкости, содержащей триспецифическое антитело. Клетки без иммобилизованного HBsAg служили в качестве отрицательного контроля. МНКПК культивировали на планшете со связанным HBsAg в течение 48 часов, затем собирали надосадочную жидкость с клеток и клетки для дальнейшего анализа. Определяли секрецию цитокинов после активации эффекторных Т-клеток в надосадочной жидкости посредством ИФА с применением набора для определения человеческого ИФН-γ IFN-γ ELISA MAX™ (BioLegend). Клетки окрашивали антителами против LAMP-1 и гранзима В, и анализировали посредством проточной цитометрии. Все эксперименты проводили в трех последовательностях.
Совместная культура HVB-позитивных клеток-мишеней и переориентированных клеток-эффекторов
Huh7 или Huh7S клетки-мишени культивировали в 96-луночном планшете при слиянии. Добавляли 1x105 МНКПК клеток в объеме 100 мкл среды на лунку. В лунку вносили 100 мкл надосадочных жидкостей НЕК, содержащих триспецифические антитела. Для определения синергетических эффектов активации CD3 и CD28, 50 мкл надосадочной жидкости с каждым биспецифическим антителом добавляли на лунку. Необработанные клетки-мишени, инкубированные с эффекторными клетками по отдельности (с добавлением или без добавления антител), служили в качестве контроля. Спустя 96 или 120 часов, надосадочную жидкость собирали, и анализировали секрецию ИФНγ посредством ИФА (см. выше).
Для определения цитотоксичности HBsAg-экспрессирующие Huh7S клетки культивировали в 96-луночном Е-планшете, совместимом с системой xCELLingence RATC (ACEA, Вioscience, Inc.), до достижения слияния. Свежевыделенные МНКПК и 100 мкл надосадочной жидкости, содержащей антитело, добавляли в трех повторностях на образец. Не трансформированные Huh7 клетки служили в качестве контроля специфичности. МНКПК, совместно культивируемые с Huh7S без антитела, служили в качестве отрицательного контроля. Жизнеспособность клеток-мишеней контролировали в течение 120 часов в режиме реального времени, с применением системы xCELLingence RATC.
Для определения способности триспецифических конструкций к активации ЦТЛ от пациентов с хроническим гепатитом В, мы выделяли МНКПК от хронического носителя и использовали в экспериментах.
Пример 2
Активация ЦТЛ пяти- и восьмивалентными конструкциями из триспецифических антител, направленных против HBsAg.
В первой серии экспериментов мы оценивали активность пяти- и восьмивалентных конструкций триспецифических антител, направленных против поверхностных антигенов ЦТЛ CD3 и CD28, и малого поверхностного антигена HVB. Мы наносили HBsAg на пластиковую поверхность чашек для культивирования клеток. В эти покрытые ячейки мы добавляли иммунные эффекторные клетки, а именно МНКПК, выделенные от здоровых доноров, и конструкции биспецифических, а также триспецифических антител. В качестве отрицательного контроля мы культивировали МНКПК на покрытых HBsAg лунках при отсутствии конструкций би- или триспецифических антител для оценки неспецифической активации иммунных клеток.
Эти эксперименты показали специфическую активацию цитотоксических Т-лимфоцитов (ЦТЛ) при совместном культивировании в присутствии CD3-активирующих биспецифических конструкций, как было определено по секреции провоспалительного цитокина интерферона-гамма (ИФНγ) до 5000 пг/мл (фигура 3). Этот эффект был дополнительно усилен при совместном применении CD3- и CD28-специфических конструкций, демонстрируя синергетический эффект, в то время как CD28-активирующая конструкция сама по себе была не способна к активации Т-клеток.
Неожиданно, при комбинации CD3- и CD28- стимулирующих и костимулирующих доменов в триспецифических молекулах была показана значительно более выраженная активация ЦТЛ, чем при комбинации двух биспецифических конструкций (фигура 3), как было определено по секреции ИФНγ. Это было подтверждено при анализе цитотоксического потенциала иммунных клеток проточной цитометрией после окрашивания на LAMP-1 транслокацию, указывающую на дегрануляцию цитотоксических везикул и экспрессию гранзима В в подавляющем большинстве клеток, активированных конструкциями триспецифических антител (фигура 4А и В).
Таким образом, пятивалентная конструкция обеспечивает наибольшую эффективность, и была способна к индукции цитотоксической функции у 80-95% иммунных клеток (фигура 4). Поскольку МНКПК содержат около 70% Т-клеток, но большинство из них являются интактными, активация ЦТЛ требует сильной активации TCR-комплекса.
Пример 3
Специфический лизис HBsAg-позитивных гепатоцитов при активации ЦТЛ конструкциями триспецифических антител
Во втором наборе экспериментов мы оценивали способность конструкций триспецифических антител, направленных против поверхностных антигенов ЦТЛ CD3 и CD28 и малого поверхностного антигена HVB, к специфическому лизису HBsAg-экспрессирующих клеток. Стабильно трансфицированные клеточные линии гепатомы Huh7, продуцирующие поверхностный антиген HVB (Huh7S клетки), применяли в качестве клеток-мишеней. После установки экспрессии HVB белка, эти клетки-мишени совместно культивировали с иммунными эффекторными клетками, в частности, МНКПК, выделенными от здоровых доноров, и пятивалентной конструкцией из триспецифических антител. В качестве отрицательного контроля мы анализировали совместные культуры с HVB-негативными клетками-мишенями, которые предварительно инкубировали с HBsAg-содержащими надосадочными жидкостями. Этот контроль применяли для исключения активации эффекторных клеток в результате неспецифического связывания частиц HVB на поверхности HVB-негативных клеток-мишеней. Далее, HVB-позитивные клетки-мишени совместно культивировали с иммунными эффекторными клетками при отсутствии конструкций триспецифических антител для оценки неспецифической фоновой цитотоксичности.
Эти эксперименты показали специфическую активацию ЦТЛ при совместном культивировании с HuH7S клетками, но не с Huh7 клетками в присутствии CD3- и CD-28-активирующей пятивалентной триспецифической конструкции. Активацию ЦТЛ определяли по секреции провоспалительного цитокина интерферона-гамма (ИФН-γ) до 2 нг/мл. При отсутствии триспецифических молекул не наблюдалось активации ЦТЛ (фигура 5).
Далее, триспецифическая конструкция опосредовала специфическую цитотоксическую элиминацию HBsAg-продуцирующих клеточных линий гепатомы HuH7 (фигура 6), как показано по >90% снижению жизнеспособности клеток-мишеней Huh7S, по сравнению с HuH7 или контролем, инкубированным без добавления конструкций антител.
Для дополнительного подтверждения HBsAg-специфичности ЦТЛ активации, проводили конкурентный анализ с человеческой сывороткой, содержащей нейтрализующие анти-HBs антитела (фигура 7). Эксперимент отражает только частичную конкуренцию ЦТЛ активации, а также лизиса клеток-мишеней, когда добавляли 117,5 МЕ/л анти-HBs, в то время как литический эффект был полностью отменен при концентрации 235 МЕ/л анти-HBs.
Пример 4
Переориентация и активация Т-клеток от пациентов с хроническим гепатитом В.
Для анализа способности триспецифических конструкций антител к активации функции ЦТЛ у больных хроническим гепатитом В, выделяли МНКПК от пациента, который был носителем гепатита В с высокой виремией. Стабильно трансфицированные клеточные линии гепатомы Huh7, продуцирующие поверхностный антиген HVB (Huh7S клетки), применяли в качестве клеток-мишеней. Эти клетки-мишени совместно культивировали с МНКПК, выделенными от пациента с гепатитом В, в присутствии или при отсутствии пятивалентных конструкций триспецифических антител. В качестве отрицательного контроля анализировали совместные культуры с HVB-негативными клетками-мишенями.
Эти эксперименты показали специфический лизис HuH7S клеток, но не Huh7 клеток ЦТЛ, активированными в МНКПК пациента с хроническим гепатотом В посредством CD3- и CD-28-активирующей пятивалентной триспецифической конструкции (фигура 8). При отсутствии триспецифической конструкции антител не отмечалось лизиса, что демонстрировало специфичность подхода и отсутствие спонтанной активности ЦТЛ в МНКПК пациента. Специфическая активация ЦТЛ была подтверждена секрецией провоспалительного цитокина интерферона-гамма (ИФН-γ) (фигура 9).
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Helmholtz Zentrum München - Deutsches Forschungszentrum für
Gesundheit und Umwelt (GmbH) et al.
<120> Trispecific binding molecules for treating HBV infection and associated conditions
<130> Y1213 PCT
<150> EP15159274.8
<151> 2015-03-16
<160> 51
<170> BiSSAP 1.2
<210> 1
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv C8 heavy chain
<400> 1
Gly Phe Thr Phe Ser Gly Tyr Ala
1 5
<210> 2
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv C8 heavy chain
<400> 2
Ile Ser Gly Ser Gly Gly Ser Thr
1 5
<210> 3
<211> 21
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv C8 heavy chain
<400> 3
Ala Lys Pro Pro Gly Arg Gln Glu Tyr Tyr Gly Ser Ser Ile Tyr Tyr
1 5 10 15
Phe Pro Leu Gly Asn
20
<210> 4
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv C8 light chain
<400> 4
Asn Ile Gly Ser Lys Ser
1 5
<210> 5
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv C8 light chain
<400> 5
Asp Asp Ser
1
<210> 6
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv C8 light chain
<400> 6
Gln Val Trp Asp Ser Ser Ser Asp Leu Val Val
1 5 10
<210> 7
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv 5F9 heavy chain
<400> 7
Gly Phe Thr Phe Asn Asn Tyr Ala
1 5
<210> 8
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv 5F9 heavy chain
<400> 8
Ile Asn Ser Asp Gly Arg Ser Thr
1 5
<210> 9
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv 5F9 heavy chain
<400> 9
Ala Arg Thr Phe Tyr Ala Asp Tyr
1 5
<210> 10
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv 5F9 light chain
<400> 10
Gln Asn Val Asp Thr Thr
1 5
<210> 11
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv 5F9 light chain
<400> 11
Trp Ala Ser
1
<210> 12
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv 5F9 light chain
<400> 12
Gln Gln Tyr Ser Ile Phe Pro Tyr Thr
1 5
<210> 13
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv 5A19 heavy chain
<400> 13
Gly Phe Thr Phe Ser Ser Tyr Ala
1 5
<210> 14
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv 5A19 heavy chain
<400> 14
Val Ser Ser Asp Gly Ser Tyr Ala
1 5
<210> 15
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv 5A19 heavy chain
<400> 15
Ala Ser Phe Asn Trp Asp Val Ala Tyr
1 5
<210> 16
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv 5A19 light chain
<400> 16
Gln Ser Leu Leu Asn Thr Arg Thr Arg Lys Ser Tyr
1 5 10
<210> 17
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv 5A19 light chain
<400> 17
Trp Ala Ser
1
<210> 18
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv 5A19 light chain
<400> 18
Lys Gln Ser Tyr Ser Leu Tyr Thr
1 5
<210> 19
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv OKT3 heavy chain
<400> 19
Gly Tyr Thr Phe Thr Arg Tyr Thr
1 5
<210> 20
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv OKT3 heavy chain
<400> 20
Ile Asn Pro Ser Arg Gly Tyr Thr
1 5
<210> 21
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv OKT3 heavy chain
<400> 21
Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr
1 5 10
<210> 22
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv OKT3 light chain
<400> 22
Ser Ser Val Ser Tyr
1 5
<210> 23
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv OKT3 light chain
<400> 23
Asp Thr Ser
1
<210> 24
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv OKT3 light chain
<400> 24
Gln Gln Trp Ser Ser Asn Pro Phe Thr
1 5
<210> 25
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv 9.3 heavy chain
<400> 25
Gly Phe Ser Leu Ser Asp Tyr Gly
1 5
<210> 26
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv 9.3 heavy chain
<400> 26
Ile Trp Ala Gly Gly Gly Thr
1 5
<210> 27
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv 9.3 heavy chain
<400> 27
Ala Arg Asp Lys Gly Tyr Ser Tyr Tyr Tyr Ser Met Asp Tyr
1 5 10
<210> 28
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv 9.3 light chain
<400> 28
Glu Ser Val Glu Tyr Tyr Val Thr Ser Leu
1 5 10
<210> 29
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv 9.3 light chain
<400> 29
Ala Ala Ser
1
<210> 30
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv 9.3 light chain
<400> 30
Gln Gln Ser Arg Lys Val Pro Tyr Thr
1 5
<210> 31
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv A9 heavy chain
<400> 31
Gly Tyr Thr Phe Thr Asn Tyr Trp
1 5
<210> 32
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv A9 heavy chain
<400> 32
Ile Tyr Pro Gly Gly Gly Tyr Thr
1 5
<210> 33
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv A9 heavy chain
<400> 33
Ala Arg Ser Ala Ser Trp Tyr Phe Asp Val
1 5 10
<210> 34
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv A9 light chain
<400> 34
Thr Gly Thr Val Thr Thr Ser Asn Tyr
1 5
<210> 35
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv A9 light chain
<400> 35
His Thr Asn
1
<210> 36
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv A9 light chain
<400> 36
Ala Leu Trp Tyr Asn Asn His Trp Val
1 5
<210> 37
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv NCAM29.2 heavy chain
<400> 37
Gly Phe Thr Phe Ser Ser Phe Gly
1 5
<210> 38
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv NCAM29.2 heavy chain
<400> 38
Ile Ser Ser Gly Ser Tyr Ala Ile
1 5
<210> 39
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv NCAM29.2 heavy chain
<400> 39
Val Arg Gly Arg Arg Leu Gly Glu Gly Tyr Ala Met Asp Tyr
1 5 10
<210> 40
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR1 of scFv NCAM29.2 light chain
<400> 40
Gln Ser Leu Leu Tyr Ser Ser Asn Gln Lys Asn Tyr
1 5 10
<210> 41
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR2 of scFv NCAM29.2 light chain
<400> 41
Trp Ala Ser
1
<210> 42
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CDR3 of scFv NCAM29.2 light chain
<400> 42
Gln Gln Tyr Ser Ser Trp Thr
1 5
<210> 43
<211> 794
<212> PRT
<213> Artificial Sequence
<220>
<223> Anti-HBsAg scFv [C8]^Glycin-Serin-Linker^hIgG1-Fc[C245S,L259F,L26
0E,G262A,A355T]^StrepTag^Glycin-Linker^anti-CD3 scFv [OKT3]
<400> 43
Met Asp Phe Glu Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Met Ala Glu Val Gln Leu Val Glu Ser Gly Gly
20 25 30
Gly Leu Leu Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
35 40 45
Gly Phe Thr Phe Ser Gly Tyr Ala Met Ser Trp Val Arg Gln Ala Pro
50 55 60
Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser Gly Gly Ser
65 70 75 80
Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
85 90 95
Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
100 105 110
Asp Thr Ala Leu Tyr Tyr Cys Ala Lys Pro Pro Gly Arg Gln Glu Tyr
115 120 125
Tyr Gly Ser Ser Ile Tyr Tyr Phe Pro Leu Gly Asn Trp Gly Gln Gly
130 135 140
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Lys Leu Glu
145 150 155 160
Glu Gly Glu Phe Ser Glu Ala Arg Val Gln Ser Ala Leu Thr Gln Pro
165 170 175
Ala Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly
180 185 190
Gly Asn Asn Ile Gly Ser Lys Ser Val His Trp Tyr Gln Gln Lys Pro
195 200 205
Gly Gln Ala Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser
210 215 220
Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
225 230 235 240
Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys
245 250 255
Gln Val Trp Asp Ser Ser Ser Asp Leu Val Val Phe Gly Gly Gly Thr
260 265 270
Lys Leu Thr Val Leu Gly Asn Ser Gly Gly Gly Gly Ser Gly Gly Gly
275 280 285
Gly Ser Gly Gly Gly Gly Ser Ala Ser Glu Pro Lys Ser Ser Asp Lys
290 295 300
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Ala Pro
305 310 315 320
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
325 330 335
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
340 345 350
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
355 360 365
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
370 375 380
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
385 390 395 400
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Thr Pro Ile Glu Lys
405 410 415
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
420 425 430
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
435 440 445
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
450 455 460
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
465 470 475 480
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
485 490 495
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
500 505 510
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
515 520 525
Lys Asp Pro Gly Trp Ser His Pro Gln Phe Glu Lys Ser Arg Gly Gly
530 535 540
Gly Gly Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro
545 550 555 560
Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
565 570 575
Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu
580 585 590
Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln
595 600 605
Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr
610 615 620
Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr
625 630 635 640
Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly
645 650 655
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Asn Ser Gly Gly Gly Gly
660 665 670
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Ser Gln Ile Val
675 680 685
Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val
690 695 700
Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr
705 710 715 720
Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser
725 730 735
Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser Gly
740 745 750
Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala Ala
755 760 765
Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly Ser
770 775 780
Gly Thr Lys Leu Glu Ile Asn Gly Asn Ser
785 790
<210> 44
<211> 793
<212> PRT
<213> Artificial Sequence
<220>
<223> Anti-HBsAg scFv [C8]^Glycin-Serin-Linker^hIgG1-Fc[C245S,L259F,L26
0E,G262A,A355T]^StrepTag^Glycin-Linker^ anti-CD28 scFv [9.3]
<400> 44
Met Asp Phe Glu Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Met Ala Glu Val Gln Leu Val Glu Ser Gly Gly
20 25 30
Gly Leu Leu Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
35 40 45
Gly Phe Thr Phe Ser Gly Tyr Ala Met Ser Trp Val Arg Gln Ala Pro
50 55 60
Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser Gly Gly Ser
65 70 75 80
Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
85 90 95
Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
100 105 110
Asp Thr Ala Leu Tyr Tyr Cys Ala Lys Pro Pro Gly Arg Gln Glu Tyr
115 120 125
Tyr Gly Ser Ser Ile Tyr Tyr Phe Pro Leu Gly Asn Trp Gly Gln Gly
130 135 140
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Lys Leu Glu
145 150 155 160
Glu Gly Glu Phe Ser Glu Ala Arg Val Gln Ser Ala Leu Thr Gln Pro
165 170 175
Ala Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly
180 185 190
Gly Asn Asn Ile Gly Ser Lys Ser Val His Trp Tyr Gln Gln Lys Pro
195 200 205
Gly Gln Ala Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser
210 215 220
Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
225 230 235 240
Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys
245 250 255
Gln Val Trp Asp Ser Ser Ser Asp Leu Val Val Phe Gly Gly Gly Thr
260 265 270
Lys Leu Thr Val Leu Gly Asn Ser Gly Gly Gly Gly Ser Gly Gly Gly
275 280 285
Gly Ser Gly Gly Gly Gly Ser Ala Ser Glu Pro Lys Ser Ser Asp Lys
290 295 300
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Ala Pro
305 310 315 320
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
325 330 335
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
340 345 350
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
355 360 365
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
370 375 380
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
385 390 395 400
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Thr Pro Ile Glu Lys
405 410 415
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
420 425 430
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
435 440 445
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
450 455 460
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
465 470 475 480
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
485 490 495
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
500 505 510
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
515 520 525
Lys Asp Pro Gly Trp Ser His Pro Gln Phe Glu Lys Ser Ser Gly Gly
530 535 540
Gly Gly Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Thr Pro
545 550 555 560
Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Ser
565 570 575
Asp Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Gln Gly Leu Glu
580 585 590
Trp Leu Gly Val Ile Trp Ala Gly Gly Gly Thr Asn Tyr Asn Ser Ala
595 600 605
Leu Met Ser Arg Lys Ser Ile Ser Lys Asp Asn Ser Lys Ser Gln Val
610 615 620
Phe Leu Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Val Tyr Tyr
625 630 635 640
Cys Ala Arg Asp Lys Gly Tyr Ser Tyr Tyr Tyr Ser Met Asp Tyr Trp
645 650 655
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Arg Gly Gly Gly Ser Gly
660 665 670
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Glu Leu Thr Gln Ser
675 680 685
Pro Ala Ser Leu Ala Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys
690 695 700
Arg Ala Ser Glu Ser Val Glu Tyr Tyr Val Thr Ser Leu Met Gln Trp
705 710 715 720
Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Phe Ala Ala
725 730 735
Ser Asn Val Glu Ser Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser
740 745 750
Gly Thr Asn Phe Ser Leu Asn Ile His Pro Val Asp Glu Asp Asp Val
755 760 765
Ala Met Tyr Phe Cys Gln Gln Ser Arg Lys Val Pro Tyr Thr Phe Gly
770 775 780
Gly Gly Thr Lys Leu Glu Ile Lys Arg
785 790
<210> 45
<211> 792
<212> PRT
<213> Artificial Sequence
<220>
<223> Anti-HBsAg scFv [C8]^Glycin-Serin-Linker^hIgG1-Fc[C245S,L259F,L26
0E,G262A,A355T]^StrepTag^Glycin-Linker^ StrepTag^anti-CD16 scFv
[A9]
<400> 45
Met Asp Phe Glu Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Met Ala Glu Val Gln Leu Val Glu Ser Gly Gly
20 25 30
Gly Leu Leu Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
35 40 45
Gly Phe Thr Phe Ser Gly Tyr Ala Met Ser Trp Val Arg Gln Ala Pro
50 55 60
Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser Gly Gly Ser
65 70 75 80
Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
85 90 95
Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
100 105 110
Asp Thr Ala Leu Tyr Tyr Cys Ala Lys Pro Pro Gly Arg Gln Glu Tyr
115 120 125
Tyr Gly Ser Ser Ile Tyr Tyr Phe Pro Leu Gly Asn Trp Gly Gln Gly
130 135 140
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Lys Leu Glu
145 150 155 160
Glu Gly Glu Phe Ser Glu Ala Arg Val Gln Ser Ala Leu Thr Gln Pro
165 170 175
Ala Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly
180 185 190
Gly Asn Asn Ile Gly Ser Lys Ser Val His Trp Tyr Gln Gln Lys Pro
195 200 205
Gly Gln Ala Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser
210 215 220
Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
225 230 235 240
Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys
245 250 255
Gln Val Trp Asp Ser Ser Ser Asp Leu Val Val Phe Gly Gly Gly Thr
260 265 270
Lys Leu Thr Val Leu Gly Asn Ser Gly Gly Gly Gly Ser Gly Gly Gly
275 280 285
Gly Ser Gly Gly Gly Gly Ser Ala Ser Glu Pro Lys Ser Ser Asp Lys
290 295 300
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Ala Pro
305 310 315 320
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
325 330 335
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
340 345 350
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
355 360 365
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
370 375 380
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
385 390 395 400
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Thr Pro Ile Glu Lys
405 410 415
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
420 425 430
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
435 440 445
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
450 455 460
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
465 470 475 480
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
485 490 495
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
500 505 510
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
515 520 525
Lys Asp Pro Gly Trp Ser His Pro Gln Phe Glu Lys Ser Arg Gly Gly
530 535 540
Gly Gly Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro
545 550 555 560
Gly Thr Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
565 570 575
Asn Tyr Trp Leu Gly Trp Val Lys Gln Arg Pro Gly His Gly Leu Glu
580 585 590
Trp Ile Gly Asp Ile Tyr Pro Gly Gly Gly Tyr Thr Asn Tyr Asn Glu
595 600 605
Lys Phe Lys Gly Lys Ala Thr Val Thr Ala Asp Thr Ser Ser Arg Thr
610 615 620
Ala Tyr Val Gln Val Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr
625 630 635 640
Phe Cys Ala Arg Ser Ala Ser Trp Tyr Phe Asp Val Trp Gly Ala Gly
645 650 655
Thr Thr Val Thr Val Ser Ser Gly Asn Ser Gly Gly Gly Gly Ser Gly
660 665 670
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Ser Gln Ala Val Val Thr
675 680 685
Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu Thr Val Thr Leu Thr
690 695 700
Cys Arg Ser Asn Thr Gly Thr Val Thr Thr Ser Asn Tyr Ala Asn Trp
705 710 715 720
Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly Leu Ile Gly His Thr
725 730 735
Asn Asn Arg Ala Pro Gly Val Pro Ala Arg Phe Ser Gly Ser Leu Ile
740 745 750
Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala Gln Thr Glu Asp Glu
755 760 765
Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Asn Asn His Trp Val Phe Gly
770 775 780
Gly Gly Thr Lys Leu Thr Val Leu
785 790
<210> 46
<211> 799
<212> PRT
<213> Artificial Sequence
<220>
<223> Anti-HBsAg scFv [C8]^Glycin-Serin-Linker^hIgG1-Fc[C245S,L259F,L26
0E,G262A,A355T]^StrepTag^Glycin-Linker^ StrepTag^anti-CD56 scFv
[NCAM29.2]
<400> 46
Met Asp Phe Glu Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Met Ala Glu Val Gln Leu Val Glu Ser Gly Gly
20 25 30
Gly Leu Leu Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
35 40 45
Gly Phe Thr Phe Ser Gly Tyr Ala Met Ser Trp Val Arg Gln Ala Pro
50 55 60
Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser Gly Gly Ser
65 70 75 80
Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
85 90 95
Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
100 105 110
Asp Thr Ala Leu Tyr Tyr Cys Ala Lys Pro Pro Gly Arg Gln Glu Tyr
115 120 125
Tyr Gly Ser Ser Ile Tyr Tyr Phe Pro Leu Gly Asn Trp Gly Gln Gly
130 135 140
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Lys Leu Glu
145 150 155 160
Glu Gly Glu Phe Ser Glu Ala Arg Val Gln Ser Ala Leu Thr Gln Pro
165 170 175
Ala Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly
180 185 190
Gly Asn Asn Ile Gly Ser Lys Ser Val His Trp Tyr Gln Gln Lys Pro
195 200 205
Gly Gln Ala Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser
210 215 220
Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
225 230 235 240
Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys
245 250 255
Gln Val Trp Asp Ser Ser Ser Asp Leu Val Val Phe Gly Gly Gly Thr
260 265 270
Lys Leu Thr Val Leu Gly Asn Ser Gly Gly Gly Gly Ser Gly Gly Gly
275 280 285
Gly Ser Gly Gly Gly Gly Ser Ala Ser Glu Pro Lys Ser Ser Asp Lys
290 295 300
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Ala Pro
305 310 315 320
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
325 330 335
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
340 345 350
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
355 360 365
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
370 375 380
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
385 390 395 400
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Thr Pro Ile Glu Lys
405 410 415
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
420 425 430
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
435 440 445
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
450 455 460
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
465 470 475 480
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
485 490 495
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
500 505 510
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
515 520 525
Lys Asp Pro Gly Trp Ser His Pro Gln Phe Glu Lys Ser Ser Gly Gly
530 535 540
Gly Gly Asp Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro
545 550 555 560
Gly Gly Ser Arg Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
565 570 575
Ser Phe Gly Met His Trp Val Arg Gln Ala Pro Glu Lys Gly Leu Glu
580 585 590
Trp Val Ala Tyr Ile Ser Ser Gly Ser Tyr Ala Ile Tyr Tyr Ala Asp
595 600 605
Thr Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Pro Glu Asn Thr
610 615 620
Leu Phe Leu Gln Met Thr Ser Leu Arg Ser Glu Asp Ser Ala Met Tyr
625 630 635 640
Tyr Cys Val Arg Gly Arg Arg Leu Gly Glu Gly Tyr Ala Met Asp Tyr
645 650 655
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Gly Asn Ser Gly Gly
660 665 670
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Ser Asp
675 680 685
Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly Glu
690 695 700
Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser Ser
705 710 715 720
Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser
725 730 735
Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Lys Ser Gly Val Pro
740 745 750
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
755 760 765
Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln Tyr
770 775 780
Ser Ser Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
785 790 795
<210> 47
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 47
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 48
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 48
Ala Lys Thr Thr Pro Lys Leu Glu Glu Gly Glu Phe Ser Glu Ala Arg
1 5 10 15
Val
<210> 49
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 49
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 50
<211> 835
<212> PRT
<213> Artificial Sequence
<220>
<223> C8-hIgG1Fcmut-OKT3 with Jun leucine zipper dimerization domain
<400> 50
Met Asp Phe Glu Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Met Ala Glu Val Gln Leu Val Glu Ser Gly Gly
20 25 30
Gly Leu Leu Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
35 40 45
Gly Phe Thr Phe Ser Gly Tyr Ala Met Ser Trp Val Arg Gln Ala Pro
50 55 60
Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser Gly Gly Ser
65 70 75 80
Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
85 90 95
Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
100 105 110
Asp Thr Ala Leu Tyr Tyr Cys Ala Lys Pro Pro Gly Arg Gln Glu Tyr
115 120 125
Tyr Gly Ser Ser Ile Tyr Tyr Phe Pro Leu Gly Asn Trp Gly Gln Gly
130 135 140
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Lys Leu Glu
145 150 155 160
Glu Gly Glu Phe Ser Glu Ala Arg Val Gln Ser Ala Leu Thr Gln Pro
165 170 175
Ala Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly
180 185 190
Gly Asn Asn Ile Gly Ser Lys Ser Val His Trp Tyr Gln Gln Lys Pro
195 200 205
Gly Gln Ala Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser
210 215 220
Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
225 230 235 240
Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys
245 250 255
Gln Val Trp Asp Ser Ser Ser Asp Leu Val Val Phe Gly Gly Gly Thr
260 265 270
Lys Leu Thr Val Leu Gly Asn Ser Gly Gly Gly Gly Ser Gly Gly Gly
275 280 285
Gly Ser Gly Gly Gly Gly Ser Ala Ser Arg Ile Ala Arg Leu Glu Glu
290 295 300
Lys Val Lys Thr Leu Lys Ala Gln Asn Ser Glu Leu Ala Ser Thr Ala
305 310 315 320
Asn Met Leu Arg Glu Gln Val Ala Gln Leu Lys Gln Lys Val Met Asn
325 330 335
Tyr Ser Ser Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro
340 345 350
Cys Pro Ala Pro Pro Ala Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
355 360 365
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
370 375 380
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
385 390 395 400
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
405 410 415
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
420 425 430
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Ala Val Ser Asn
435 440 445
Lys Gly Leu Ala Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
450 455 460
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
465 470 475 480
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
485 490 495
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
500 505 510
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
515 520 525
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
530 535 540
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
545 550 555 560
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Asp Pro Gly Trp Ser His
565 570 575
Pro Gln Phe Glu Lys Ser Arg Gly Gly Gly Gly Gln Val Gln Leu Gln
580 585 590
Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser
595 600 605
Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val
610 615 620
Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro
625 630 635 640
Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr
645 650 655
Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser
660 665 670
Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp
675 680 685
Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val
690 695 700
Ser Ser Gly Asn Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
705 710 715 720
Gly Gly Gly Ser Ala Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile
725 730 735
Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser
740 745 750
Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser
755 760 765
Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
770 775 780
Ala His Phe Arg Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
785 790 795 800
Ser Gly Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp
805 810 815
Ser Ser Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn
820 825 830
Gly Asn Ser
835
<210> 51
<211> 834
<212> PRT
<213> Artificial Sequence
<220>
<223> C8- hIgG1Fcmut-9.3 with Fos leucine zipper dimerization domain
<400> 51
Met Asp Phe Glu Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Met Ala Glu Val Gln Leu Val Glu Ser Gly Gly
20 25 30
Gly Leu Leu Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
35 40 45
Gly Phe Thr Phe Ser Gly Tyr Ala Met Ser Trp Val Arg Gln Ala Pro
50 55 60
Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser Gly Gly Ser
65 70 75 80
Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
85 90 95
Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
100 105 110
Asp Thr Ala Leu Tyr Tyr Cys Ala Lys Pro Pro Gly Arg Gln Glu Tyr
115 120 125
Tyr Gly Ser Ser Ile Tyr Tyr Phe Pro Leu Gly Asn Trp Gly Gln Gly
130 135 140
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Lys Leu Glu
145 150 155 160
Glu Gly Glu Phe Ser Glu Ala Arg Val Gln Ser Ala Leu Thr Gln Pro
165 170 175
Ala Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly
180 185 190
Gly Asn Asn Ile Gly Ser Lys Ser Val His Trp Tyr Gln Gln Lys Pro
195 200 205
Gly Gln Ala Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser
210 215 220
Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
225 230 235 240
Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys
245 250 255
Gln Val Trp Asp Ser Ser Ser Asp Leu Val Val Phe Gly Gly Gly Thr
260 265 270
Lys Leu Thr Val Leu Gly Asn Ser Gly Gly Gly Gly Ser Gly Gly Gly
275 280 285
Gly Ser Gly Gly Gly Gly Ser Ala Ser Leu Thr Asp Thr Leu Gln Ala
290 295 300
Glu Thr Asp Gln Leu Glu Asp Lys Lys Ser Ala Leu Gln Thr Glu Ile
305 310 315 320
Ala Asn Leu Leu Lys Glu Lys Glu Lys Leu Glu Phe Ile Leu Ala Ala
325 330 335
Tyr Ser Ser Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro
340 345 350
Cys Pro Ala Pro Pro Ala Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
355 360 365
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
370 375 380
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
385 390 395 400
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
405 410 415
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
420 425 430
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Ala Val Ser Asn
435 440 445
Lys Gly Leu Ala Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
450 455 460
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
465 470 475 480
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
485 490 495
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
500 505 510
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
515 520 525
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
530 535 540
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
545 550 555 560
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Asp Pro Gly Trp Ser His
565 570 575
Pro Gln Phe Glu Lys Ser Ser Gly Gly Gly Gly Gln Val Gln Leu Gln
580 585 590
Glu Ser Gly Pro Gly Leu Val Thr Pro Ser Gln Ser Leu Ser Ile Thr
595 600 605
Cys Thr Val Ser Gly Phe Ser Leu Ser Asp Tyr Gly Val His Trp Val
610 615 620
Arg Gln Ser Pro Gly Gln Gly Leu Glu Trp Leu Gly Val Ile Trp Ala
625 630 635 640
Gly Gly Gly Thr Asn Tyr Asn Ser Ala Leu Met Ser Arg Lys Ser Ile
645 650 655
Ser Lys Asp Asn Ser Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu
660 665 670
Gln Ala Asp Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Lys Gly Tyr
675 680 685
Ser Tyr Tyr Tyr Ser Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr
690 695 700
Val Ser Ser Arg Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
705 710 715 720
Gly Ser Asp Ile Glu Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser
725 730 735
Leu Gly Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Glu
740 745 750
Tyr Tyr Val Thr Ser Leu Met Gln Trp Tyr Gln Gln Lys Pro Gly Gln
755 760 765
Pro Pro Lys Leu Leu Ile Phe Ala Ala Ser Asn Val Glu Ser Gly Val
770 775 780
Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asn Phe Ser Leu Asn
785 790 795 800
Ile His Pro Val Asp Glu Asp Asp Val Ala Met Tyr Phe Cys Gln Gln
805 810 815
Ser Arg Lys Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
820 825 830
Lys Arg
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННЫЕ АНТИ-FLT3 АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2830982C2 |
| НОВОЕ PSMA-СВЯЗЫВАЮЩЕЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЯ | 2017 |
|
RU2762704C2 |
| МУЛЬТИВАЛЕНТНЫЕ FV-АНТИТЕЛА | 2016 |
|
RU2785766C2 |
| ПОЛИПЕПТИД ДЛЯ ЛЕЧЕНИЯ ПАТОЛОГИЧЕСКИХ ТРОМБОВ | 2019 |
|
RU2778566C1 |
| МОДИФИЦИРОВАННАЯ J-ЦЕПЬ | 2015 |
|
RU2761118C2 |
| ДОСТАВКА ТЕРАПЕВТИЧЕСКИХ ПОЛИПЕПТИДОВ ПОСРЕДСТВОМ ПСЕВДОТИПИРОВАННЫХ ОНКОЛИТИЧЕСКИХ ВИРУСОВ | 2017 |
|
RU2758007C2 |
| УЛУЧШЕННЫЙ БЕЛОК REP ДЛЯ ПРИМЕНЕНИЯ В ДИАГНОСТИЧЕСКИХ АНАЛИЗАХ | 2018 |
|
RU2736275C1 |
| БИСПЕЦИФИЧЕСКИЕ ТРЕХВАЛЕНТНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С КЛАУДИНОМ 6 ИЛИ КЛАУДИНОМ 18.2, И CD3, ДЛЯ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ С ЭКСПРЕССИЕЙ КЛАУДИНА | 2017 |
|
RU2798988C2 |
| ИНДУЦИБЕЛЬНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И СПОСОБЫ ИСПОЛЬЗОВАНИЯ | 2017 |
|
RU2800034C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕСЯ С 5T4 И 4-1BB, И ОТНОСЯЩИЕСЯ К НИМ КОМПОЗИЦИИ И СПОСОБЫ | 2018 |
|
RU2804458C2 |
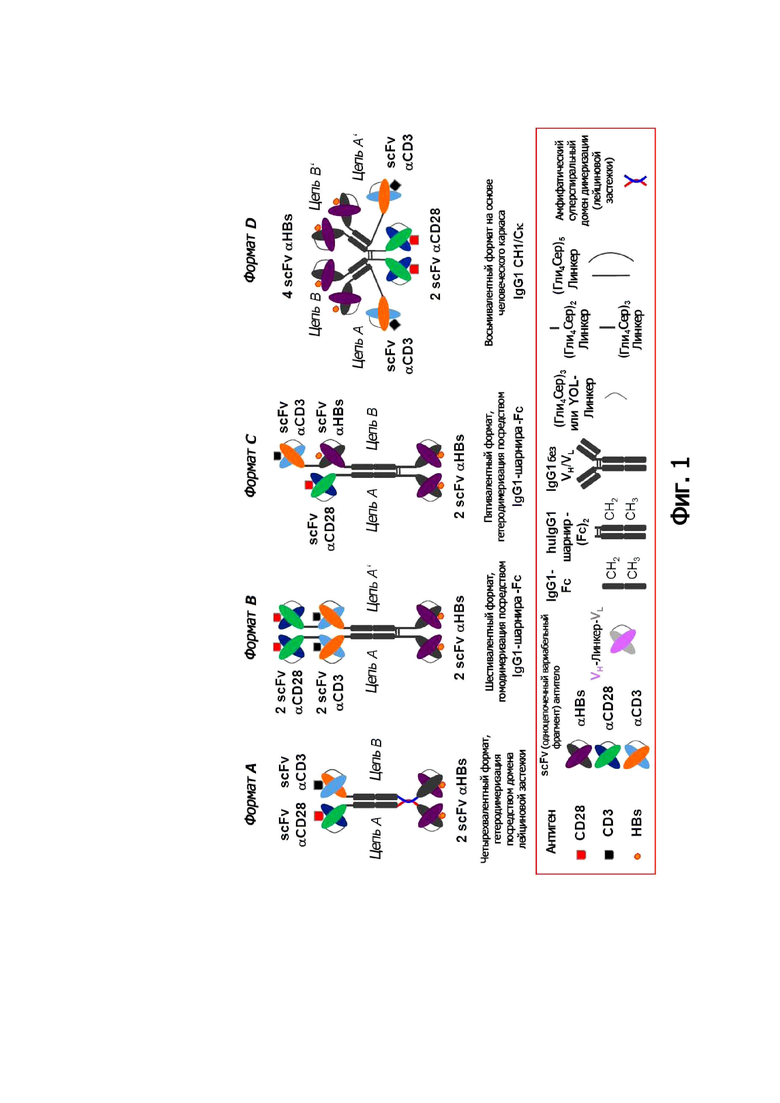
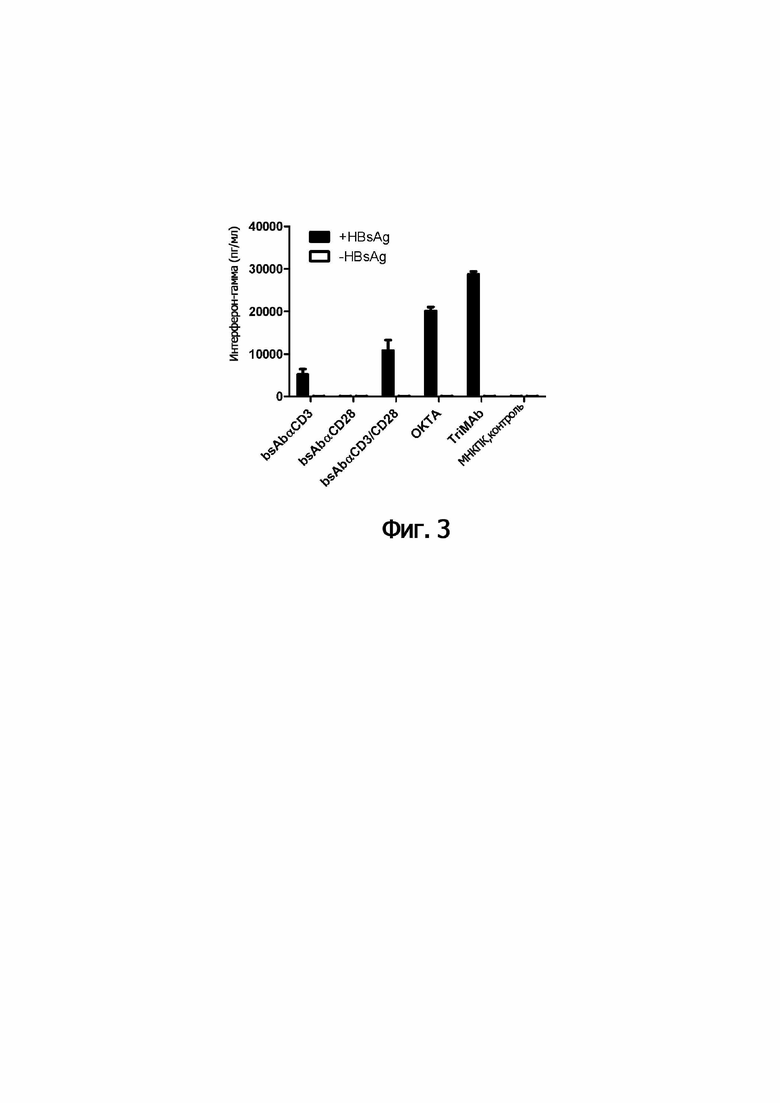
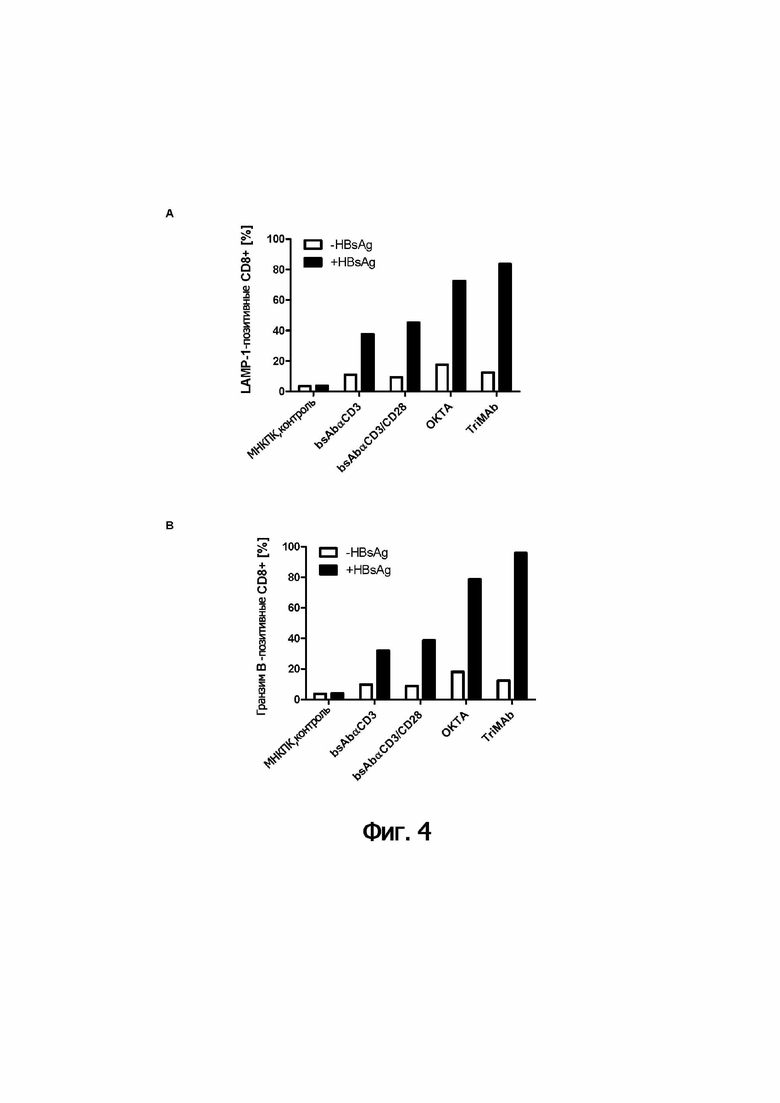
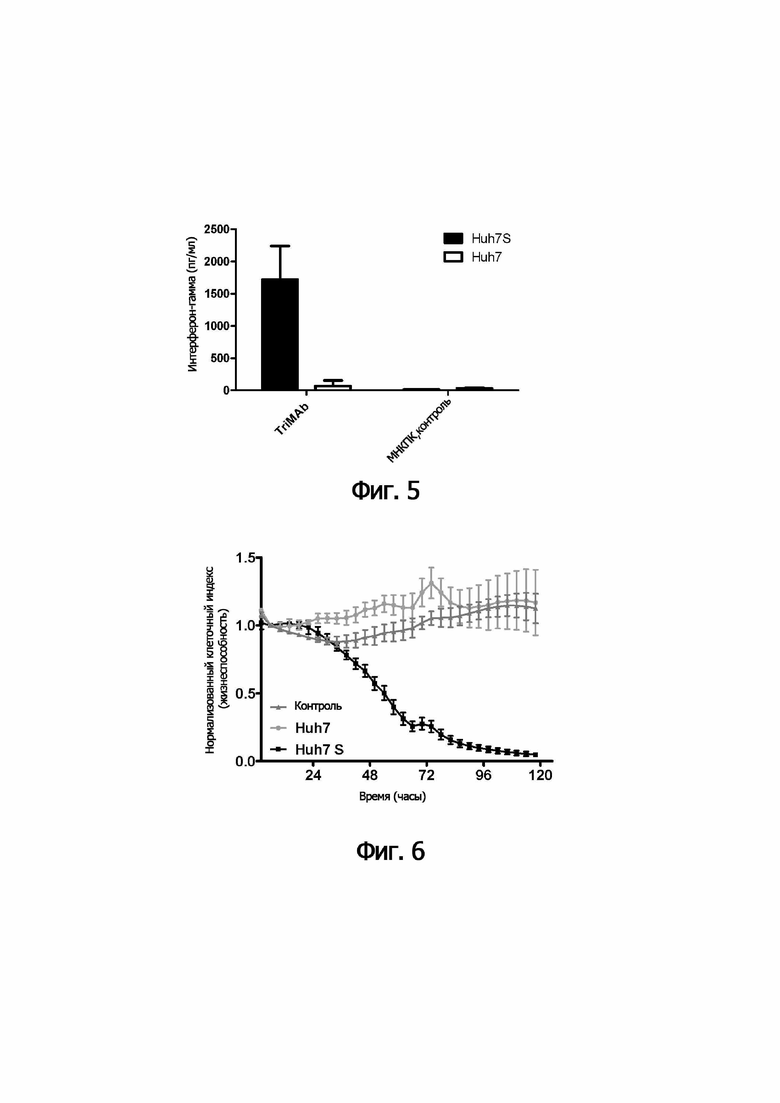
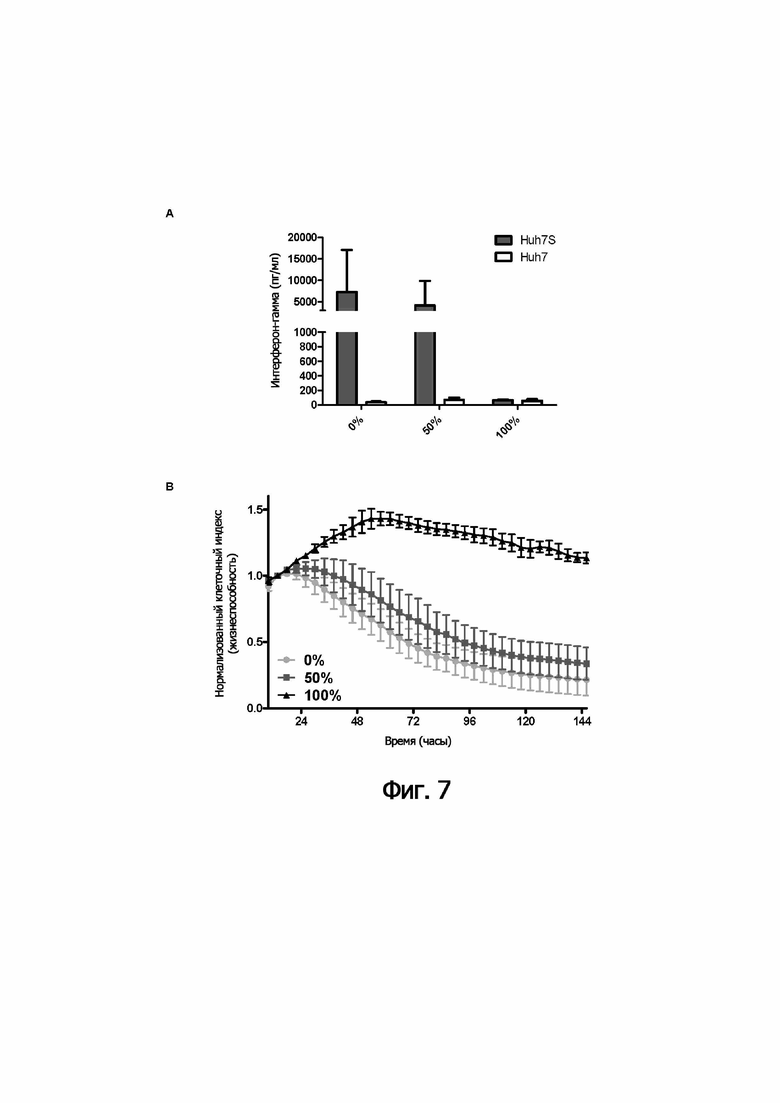
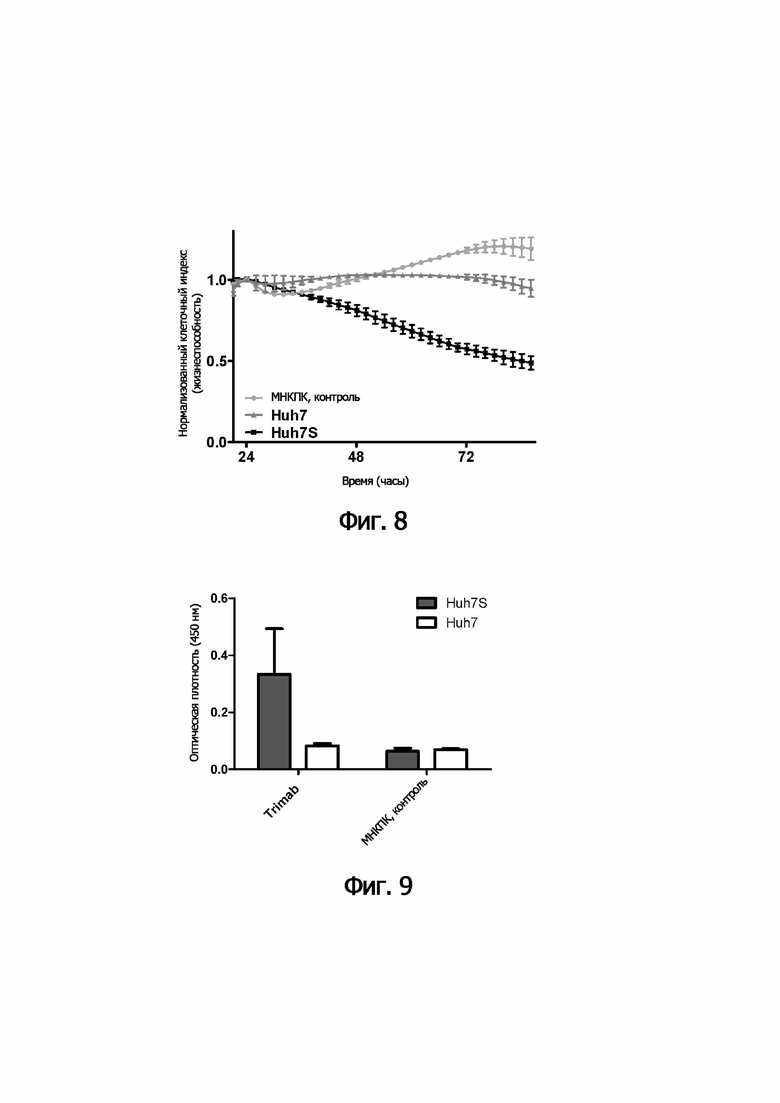
Изобретение относится к области биотехнологии, конкретно к получению молекулы специфично связывающейся с HBsAg, CD3 и CD28, и может быть использовано в медицине. Полученная связывающая молекула состоит из двух полипептидов с SEQ ID NO:43 и SEQ ID NO:44, соединённых между собой лейциновой молнией, и может быть использована для эффективного лечения или профилактики HVB инфекции или состояния, обусловленного указанной HVB инфекцией. 4 н.п. ф-лы, 9 ил., 4 пр.
1. Связывающая молекула для применения в способе лечения или профилактики HVB инфекции и/или состояния, обусловленного указанной HVB инфекцией, где указанное состояние, обусловленное указанной HVB инфекцией, выбрано из цирроза печени, гепатоцеллюлярной карциномы и рака печени, характеризующихся экспрессией одного или нескольких поверхностных антигенов HVB, где указанная связывающая молекула специфично связывается с HBsAg, CD3 и CD28 и состоит из двух полипептидов, соединённых между собой лейциновой молнией, где первый полипептид с аминокислотной последовательностью SEQ ID NO:43 имеет участки связывания с HBsAg и CD3 и второй полипептид c с аминокислотной последовательностью SEQ ID NO:44 имеет участки связывания с HBsAg и CD28.
2. Набор нуклеиновых кислот для получения молекулы по п.1, кодирующих полипептиды, составляющие связывающую молекулу по п.1.
3. Нуклеиновая кислота, кодирующая связывающую молекулу по п.1.
4. Фармацевтическая композиция для применения в способе лечения или профилактики HVB инфекции и/или состояния, обусловленного указанной HVB инфекцией, где указанное состояние, обусловленное указанной HVB инфекцией, выбрано из цирроза печени, гепатоцеллюлярной карциномы и рака печени, характеризующихся экспрессией одного или нескольких поверхностных антигенов HVB, включающая эффективное количество молекулы по п.1 и фармацевтически приемлемый носитель.
| СЫРЬЕВАЯ СМЕСЬ ДЛЯ ИЗГОТОВЛЕНИЯ БЕТОНА | 2013 |
|
RU2524699C1 |
| LIU J | |||
| et al., A new format of single chain tri-specific antibody with diminished molecular size efficiently induces ovarian tumor cell killing, Biotechnology letters, 2005, V | |||
| Прибор с двумя призмами | 1917 |
|
SU27A1 |
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| DEYEV S | |||
| M | |||
| et al., Multivalency: the hallmark of antibodies used for optimization of tumor targeting by design, Bioessays, | |||

