Бактериальные инфекции остаются одной из наиболее распространенных причин смертельных заболевания человека. Появление антибиотикорезистентных бактериальных штаммов является одной из основных клинических проблем во всем мире [1, CDC, 2013; 2, O'Neill, 2014; 3, O'Neill, 2016]. Резкий рост и распространение резистентности к антибиотикам среди бактерий требует открытия и разработки новых противомикробных препаратов и расширения арсенала структурных мотивов, подходящих для создания на их основе новых антибактериальных соединений.
Заявленные в данном изобретении тиазол-оксазол модифицированные пептиды, выступают ингибиторами синтеза белка в бактериалоьной клетке, воздействуя на рибосомы. Они связываются в участке выходного туннеля рибосомы (nascent peptide exit tunnel, NPET), через который синтезированные de novo на рибосоме белки покидают ее.
1. Тиазол/оксазол-модифицированные микроцины (ТОММ) представляют собой подмножество более крупного класса натуральных продуктов - рибосомально синтезируемых и посттрансляционно модифицируемых пептидов (РиПП) [4, Paul G. Arnison et al., 2013]. ТОММ обладают множеством биологических активностей, среди них встречаются ингибиторы ДНК-гиразы, ингибиторы трансляции и гемолитические токсины. [4, Paul G. Arnison et al., 2013; 5, Molloy EM, 2011; 6, Lee SW et al. 2008; 7,  et al. 1991; 8, Bagley MC et al. 2005]. Антибиотики по степени воздействия на клетки бактерий делят на а) бактериостатические, которые тормозят рост и развитие клеток; б) бактерицидные, которые приводят к клеточной гибели.
et al. 1991; 8, Bagley MC et al. 2005]. Антибиотики по степени воздействия на клетки бактерий делят на а) бактериостатические, которые тормозят рост и развитие клеток; б) бактерицидные, которые приводят к клеточной гибели.
Из всех известных на сегодняшний день мишеней соединения по заявляемому изобретению ингибируют аппарат биосинтеза белка, осуществляемый на рибосомах. Бактериальная рибосома является одной из важнейших клеточных мишеней антибиотиков, используемых в клинической практике. Рибосомы представляют собой крупные, сложноорганизованные, состоящие из белков и рибосомных рибонуклеиновых кислот (рРНК) клеточные органеллы, осуществляющие синтез белка. Эти рибонуклеопротеины содержат примерно две трети клеточной РНК и одну треть всего белка клетки.
Антибиотики, ингибирующие рибосому, имеют различные сайты связывания и механизмы действия [9, Wilson DN, 2009]. К таким препаратам относятся классы макролидов и их производных; тетрациклинов, аминогликозидов; хлорамфеникол, клиндамицин, спектиномицин, стрептомицин и амикацин, а также некоторые антимикробные пептиды.
Большое количество доступных сайтов связывания и низкая частота мутационной резистентности делают бактериальную рибосому удобной мишенью для новых антибактериальных агентов.
Наиболее распространенными фармацевтическими препаратами, воздействующими на бактериальную рибосому, выступают макролидные антибиотики. Макролиды представляют собой широкий класс клинически значимых антибиотиков, основу химической структуры которых составляет макроциклическое лактонное кольцо с присоединенными одним или несколькими фрагментами сахара [10, Gaynor М, 2003]. В зависимости от числа атомов углерода в кольце макролиды подразделяются на 14-членные (эритромицин, рокситромицин, кларитромицин), 15-членные (азитромицин) и 16-членные (мидекамицин, спирамицин, джозамицин). Макролиды активны в отношении грамположительных кокков (стафилококки, стрептококки) и быцилл (роды Listeria monocytogenes, Bacillus anthracis, Corynebacterium); некоторых грамотрицательных кокков и коккобацилл (Neisseria gonorrhoeae, Moraxella catarrhalis, Haemophilus influenzae, Bordetella pertussis, Campylobacter, Helicobacter, и Legionella species), а также ряда внутриклеточных паразитов (Mycobacteria, Mycoplasma, Ureaplasma, Chlamydia, Spirochetes) [11, Comprehensive Information on Prescription Drugs, 2017]. Макролиды, как правило, оказывают бактериостатическое действие, они связываются в выходном туннеле рибосомы, чем препятствуют прохождению новосинтезированных полипептидов через туннель, приводя к остановке трансляции на ранних этапах белкового синтеза. [8, Bagley МС et al. 2005; 12, Gale EF at al. 1981; 13, Vazquez D, 1977]. Большинство макролидных антибиотиков связывается с основаниями 2058 и 2059 23S рРНК, 50S субъединицы рибосомы. Внедрение химических модификаций в структуру макролидов оказывает прямое влияние на механизм их связывания с рибосомой, что позволяет макролидам оставаться потенциально перспективными кандидатами на роль ингибирующих рибосому агентов.
Недостатки антибиотиков макролидного ряда обусловлены следующими факторами.
Ряд макролидных антибиотиков являются причиной возникновения резистентности к ним самим, поскольку выступают сильными индукторами экспрессии генов специальных метилаз, метилирующих аденин в положения 2058 23S рРНК [14, Leclercq R, 2002]. Таких металаз к настоящему моменту описано более 30 и их гены могут распространяться в популяции бактерий как на плазмидной ДНК, так и присутствовать в их геноме. Метилирование аденина в положения 2058 23S рРНК является наиболее распространенной причиной возникновения устойчивости к макролидным антибиотикам [15, 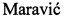 G, 2004; 16, Bailey М et al. 2008].
G, 2004; 16, Bailey М et al. 2008].
Антибиотики макролидного ряда длительное время находятся в клиническом применении - более 60-ти лет, что привело к снижению их антимикробного потенциала за счет возникновения среди чувствительных патогенов множества устойчивых штаммов.
Многолетние усилия в области химического синтеза для получения новых модифицированных макролидов с улучшенными свойствами позволили разработать ряд замечательных лекарств (азитромицин, кларитромицин, кетолиды и др.), успешно используемых в клинической практике, однако, со временем такой подход исчерпывает себя и становится все более трудоемок. Поэтому быстро появляющаяся среди патогенных микроорганизмов резистентность к применяемым в клинической практике препаратам, требует разработки новых эффективных антибиотиков.
Поскольку макролидные антибиотики имеют не пептидное происхождение и синтезируются в клетках бактерий в результате довольно сложного многостадийного процесса [17, Zhang Н et al. 2010], современные методы молекулярной генетики практически не применяются для создания новых вариантов макролидных антибиотиков.
К другим недостаткам макролидов можно отнести их необширный сайт связывания с рибосомой: они преимущественно взаимодействуют только с двумя основаниями 23S рРНК - 2058 и 2059, не связываясь в других сайтах рибосомы. Из-за своих некрупных размеров макролиды не полностью блокируют выходной туннель рибосомы, лишь частично перекрывая его, что в некоторых случаях позволят новосинтезированным пептидам преодолевать тоннель, фактически отменяя ингибирующее действие макролидов [18, Kannan K et al. 2012; 19, Kannan K et al. 2014; 20, Davis AR et al. 2014].
Широкий спектр антимикробного действия макролидных антибиотиков не позволяет использовать их качестве строго таргетированного лекарства, что для современной медицины является ощутимым недостатком.
Перспективным классом соединений, которые, однако, еще не имеют широкого фармацевтического применения, являются антибиотики пептидного происхождения.
Известные на сегодняшний день противомикробные пептиды, взаимодействующие с бактериальной рибосомой, относятся к классу пролин-богатых антимикробных пептидов (proline-rich AntiMicrobal Peptides, PrAMPs). Они обнаруживаются в гемолимфе нескольких видов насекомых, например онкоцин (патент US 8937155) и апиадецин (патент US 5300629), более устойчивая к протеазам форма - патент WO-2009/013262) [21, Mardirossian М et al. 2014; 22, Krizsan A et al. 2014], а также в нейтрофилах многих млекопитающих, например, бактеницин-7 (патент ЕР 0665239 А1, 1995) [23, Scocchi М et al. 2011].
PrAMPs активны в отношении грамотрицательных бактерий, включая широкий спектр ассоциированных с растениями, и некоторых патогенов человека. Апиадецина, онкоцин и бактеницин эффективны уже в микромолярных концентрациях, 1. 8 мкМ [22, Krizsan A et al. 2014], 3, 4 мкМ, [22] и 1 мкМ [24, Benincasa М et al. 2004] соответственно.
По своей структуре PrAMPs представляют собой альфа-спиральные пептиды, высоко обогащенные положительно заряженными аминокислотами. PrAMPs, обнаруженные в насекомых, как правило, имеют длину <21 аминокислот, в то время как PrAMPs из млекопитающих значительно крупнее, например, бычий бактинецин-7 (Вас7), который также встречается у овец и коз, состоит из 60 остатков [23]. Укороченные с С-конца производные апиадецина (Api88 и Api137), бактинецина-7 (1-16) и онкоцина (Onc72 и Onc112) ингибируют синтез белка путем связывания с 70S рибосомы с константами диссоциации в наномолярном диапазоне [22]. Сокристаллизация с 70S рибосомой бактинецина-7 и онкоцина показали схожий механизм их связывания. Оба пептида взаимодействуют с 50S субъединицей в трех функциональных сайтах: в выходном туннеле с обратной ориентацией, с Р- и с А-сайтом, где блокируют связывание аминоацил-тРНК [25, Seefeldt AC et al. 2016; 26, Roy RN et al. 2015; 27, Seefeldt AC et al. 2015]. Связывание пролин-богатых антибактериальных пептидов сразу в трех функциональных областях рибосомы значительно снижает вероятность появления резистентности, вызванной одиночными мутациями в рибосоме. Однако это не отменяет возникновения резистентности, обусловленной мутациями мембранного транспортера, через который антибактериальный пептид проникает в клетку.
Несмотря на перспективность обнаруженных антимикробных пептидов, ингибирующих бактериальную рибосому, их немногочисленность не позволяет рассматривать этот класс пептидов как опорную ступень в масштабном развитии категории фармацевтически значимых антибиотиков на их основе.
Так, все описанные на данный момент рибосомоориентированные антимикробные пептиды имеют схожий механизм ингибирования рибосомы и источник происхождения - организмы животных, а также происходят из одного класса - пролин-богатых антимикробных пептидов. Более того, среди них не обнаружено пептидов, эффективных в отношении грамотрицательных микроорганизмов.
Для расширения класса рибосомоориентированных антимикробных пептидов требуется открытие и разработка разнообразных пептидов из других структурных классов, имеющих иные механизмы ингибирования рибосомы и действующих на грамотрицательные микроорганизмы. Техническая результат заключается в расширении класса ингибиторов раннего этапа синтеза белка, за счет создания семейства тиазол-оксазол модифицированных пептидов, связывающих рибосому.
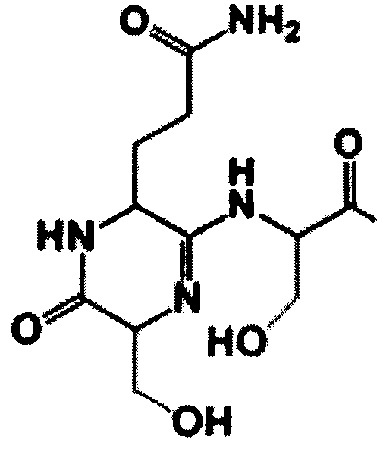
Технический результат достигается тем, что экспериментально разработаны тиазол-оксазол-модифицированные пептиды, общего вида NH2-Ser1-Gln2-Ser3-Pro4-Gly5-Asn6-Cys7-Ala8-Ser9-Cys10-Ser11-Asn12-Ser13-Pp-Ac формулы I
где NH2 - N-концевая амидная груцпа, Ас - С-концевая ацетильная
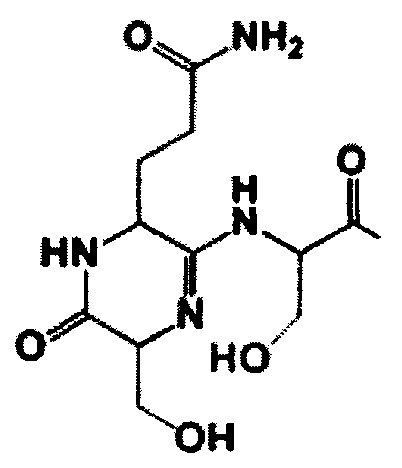
группа, Рр - полипептидная цепь, выбранная из ASANCTGGLG и KSANCTGGLG, Thz - аминокислота, образующая тиазол, Oxz - аминокислота, образующая оксазол, где пептид содержит а) три азольных цикла, образованных остатками Cys7, Cys10, Ser13, и б) модификацию первых двух аминокислот, приводящую к образованию на N-конце молекулы 6-ти-членного амидинового кольца общей формулы II
обладающие способностью ингибировать бактериальную рибосому Е. coli, K. pneumoniae и Yersinia pseudotuberculosis за счет связывания в ее выходном туннеле
В отличие от известных аналогов соединения по данному изобретению имеют нетипичную для пептидов химическую модификацию и способ блокирования выходного туннеля рибосомы, не показанный ранее для других известных антибиотиков. Некоторые представители соединений по изобретению имеют узкий спектр антибактериальной активности, и в отличие от аналогов, таргетирующих рибосому пептидов, воздействуют строго на грамположительные бактерии, что доказано экспериментально.
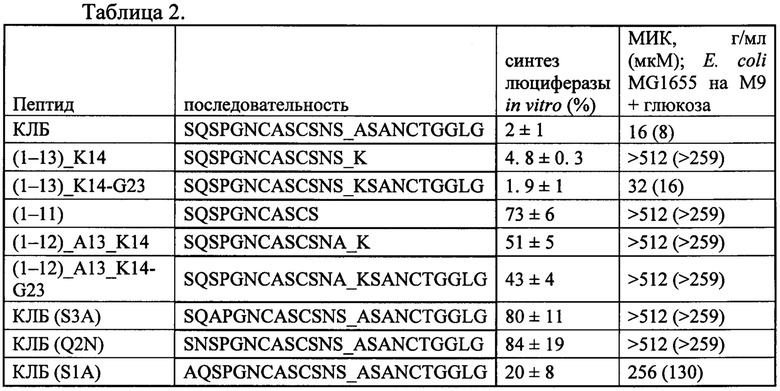
На фигуре 1 представлена химическая структура клебсазолицина и его аминокислотная последовательность. Аминокислоты, образующие тиазолы, обозначены как Thz, оксазол - Oxz.
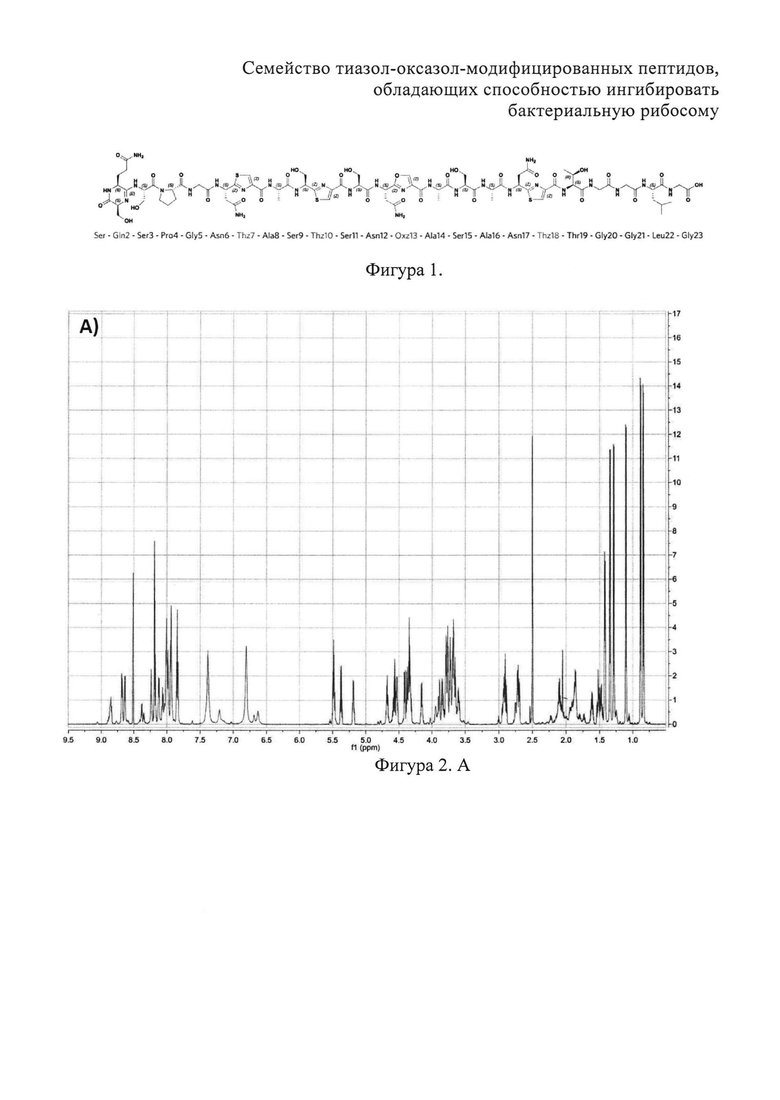
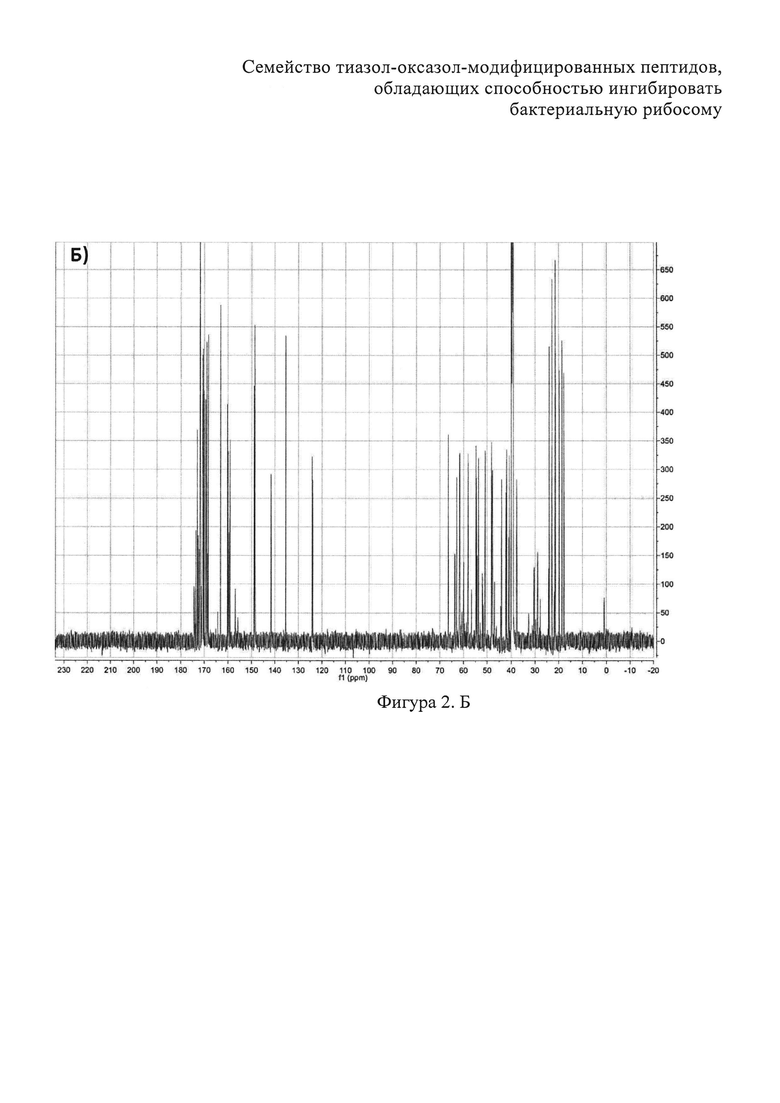
На фигуре 2 представлены двумерные ЯМР-спектры клебсазолицина:
A) 1Н спектр клебазолицина в растворителе DMSO-d6. Б) 13С спектр клебсазолицина в растворителе DMSO-d6.
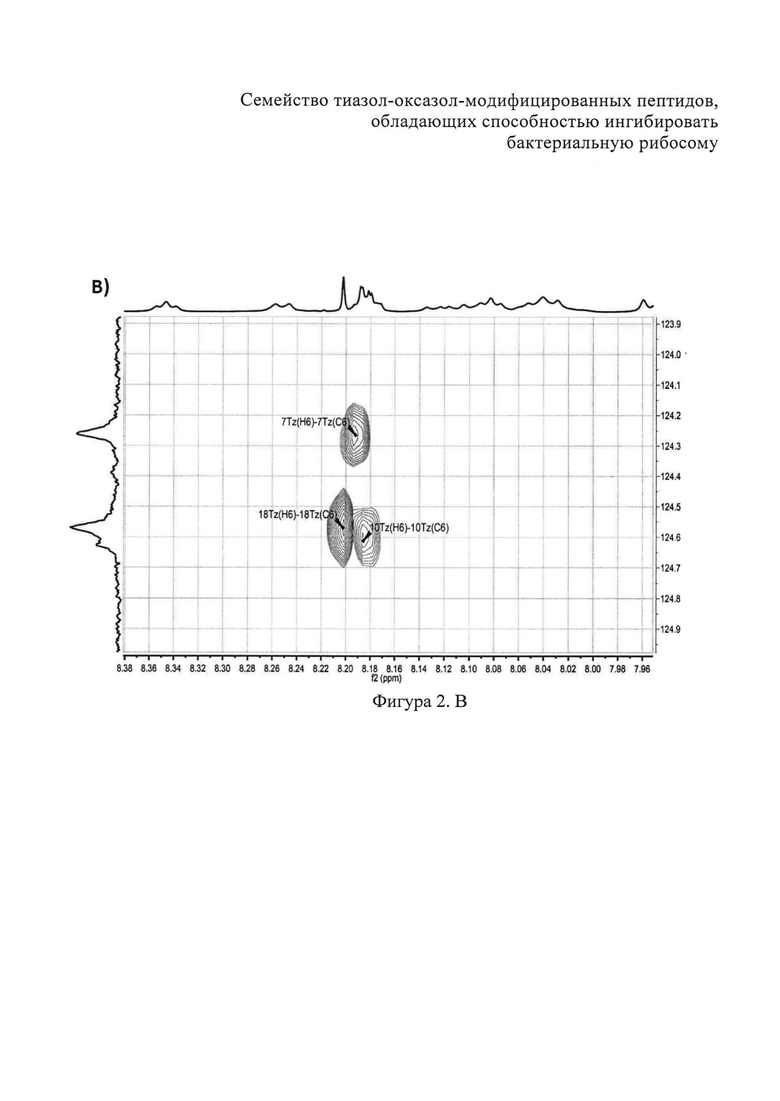
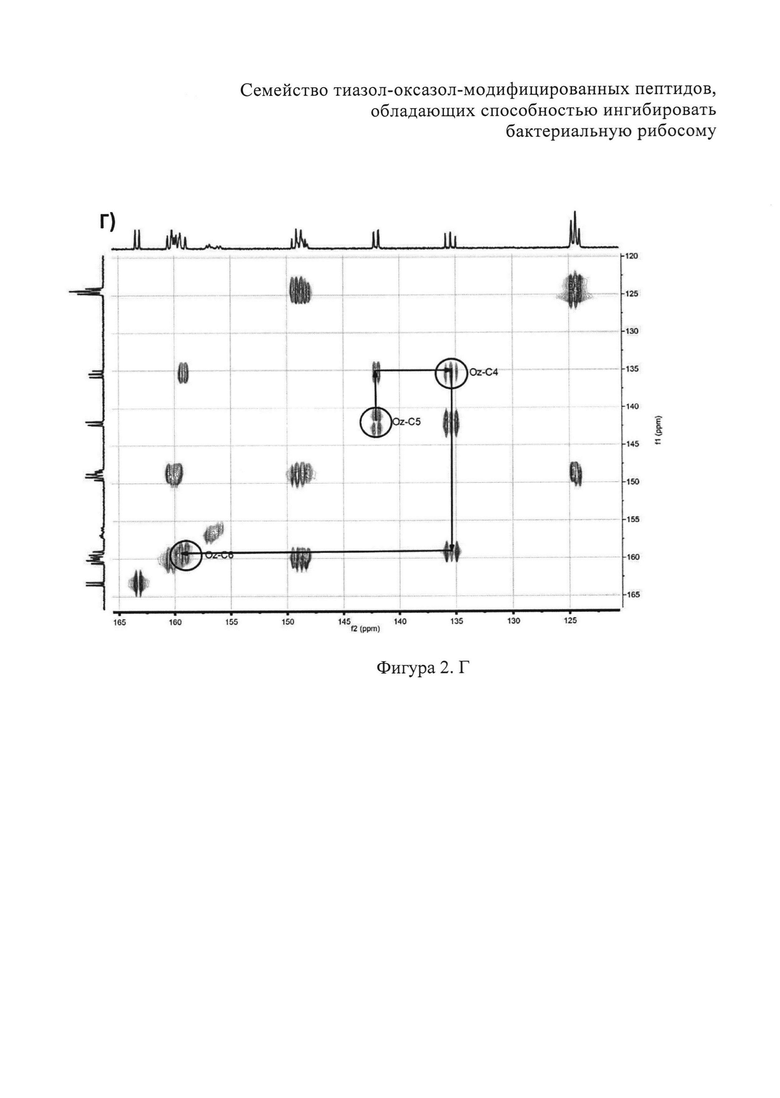
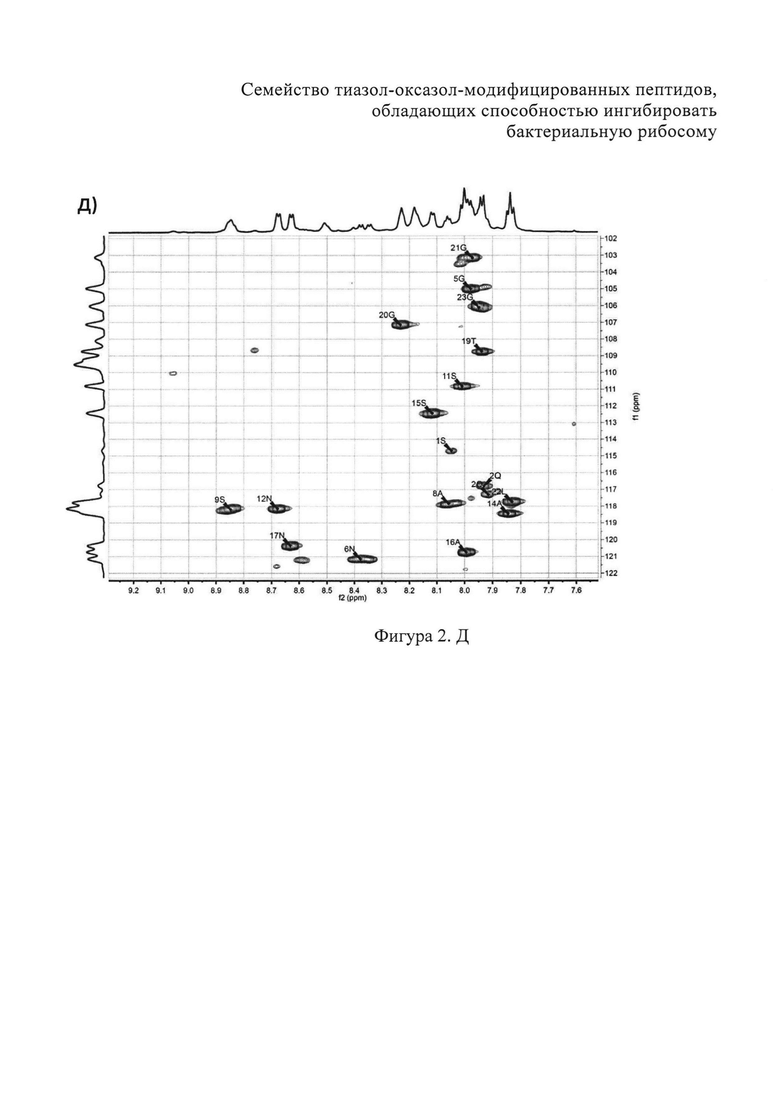
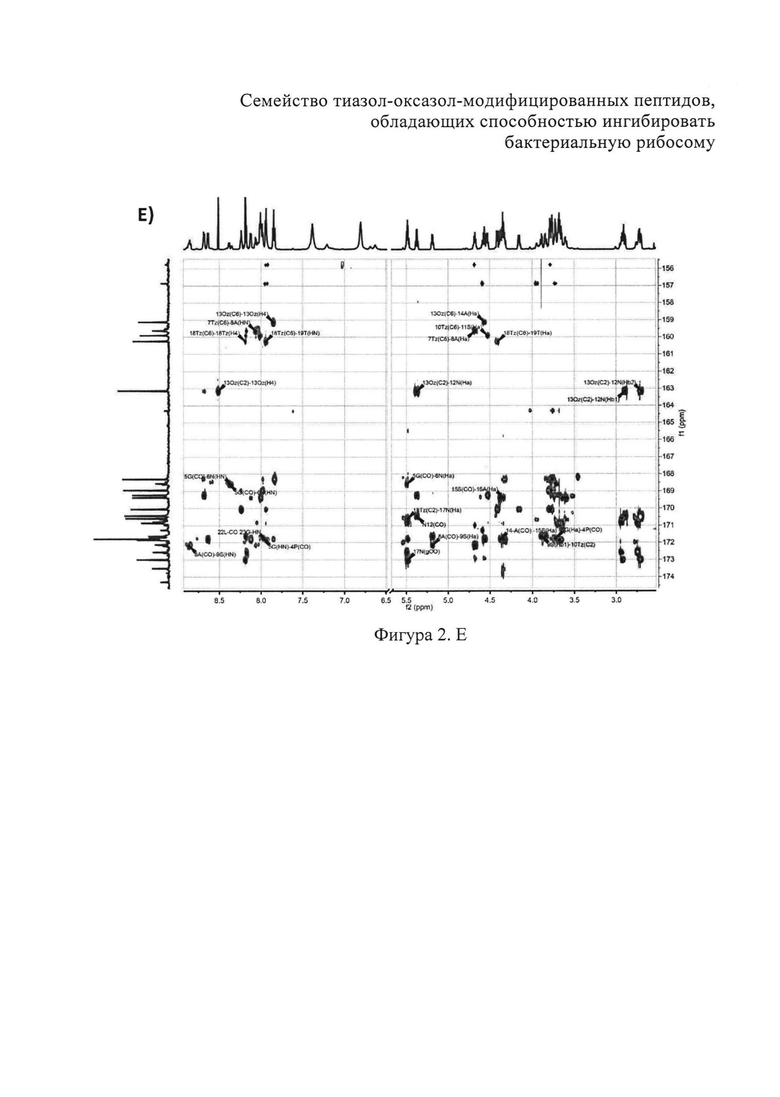
B) Фрагменты 13C-HSQC спектра с определением тиазолов. Г) Фрагменты 13C-COZY спектра с определением оксазол а. Д) Амидновый регион 1H-15N HSQC-спектра клебсазолицина. Е) Фрагменты 13С-НМВС-спектра.
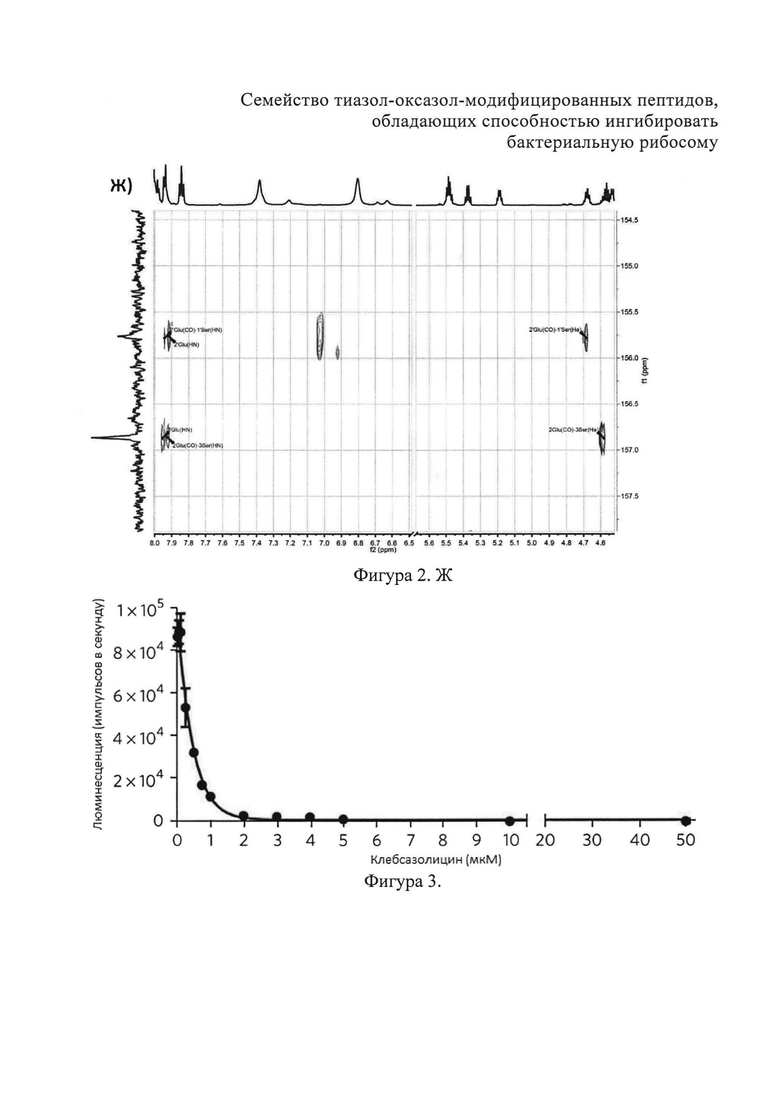
Ж) Фрагменты 13С-НМВС-спектра (Ser1, Glu2 и Ser3).
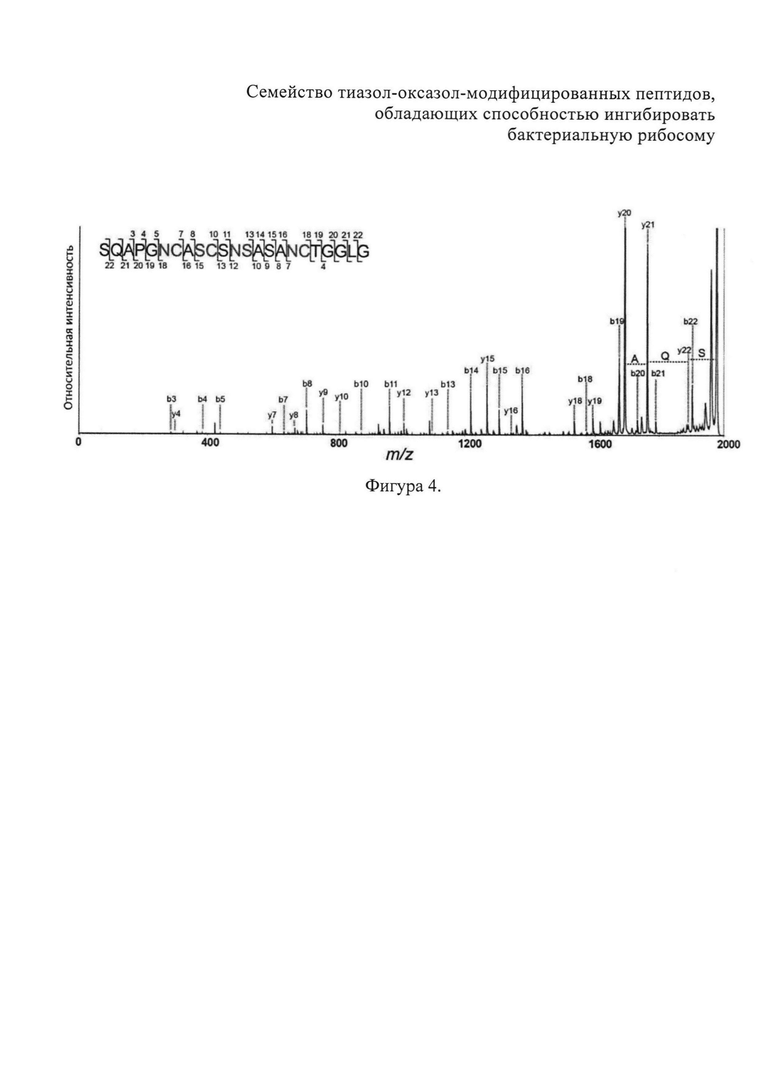
На фигуре 3 показан график зависимости интенсивности люминесценции люциферазы светлячков, синтезированной на рибосомах in vitro в бесклеточном экстракте Е. coli S30 от увеличения концентрации клебсазолицина. Клебсазолицин является ингибитором синтеза белка как in vitro, так и in vivo.
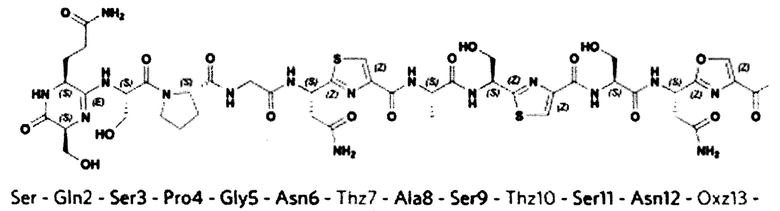
На фигуре 4 представлена МС-МС спектрограмма мутантной по третьей аминокислоте S3A формы клебсазолицина с наблюдаемыми b- и у-ионами, отмеченными в структуре и в спектрах
В Приложении 1 дана нуклеотидная последовательность полного генетического кластера биосинтеза клебсазолицина, 4616 п.н.; отдельно: нуклеотидные последовательности генов, кодирующих пептид-предшественник, белки процессинга и белок-транспортер, а также аминокислотные последовательности этих белков.
Описание изобретения
Изобретение относится к новым тиазол-оксазол модифицированным пептидам, имеющим, по меньшей мере, три азольных цикла, а также нетипичную модификацию аминокислот общей формулы I обладающих способностью ингибировать бактериальную рибосому за счет связывания в ее выходном туннеле, что приводит к остановке раннего этапа синтеза белка. В частности, к соединениям по изобретению принадлежит открытый авторами пептид, о котором сообщаем в рамках данного изобретения, названный клебсазолицином, который препятствует ранним стадиям элонгации при трансляции белка в опытах in vitro и ингибирует рост клеток ряда грамположительных штаммов.
Клебсазолицин - продукт микробного происхождения из бактерии Klebsiella pneumonia subsp. ozaenae АТСС 11296. За производство клебсазолицина этим микроорганизмом отвечает группа из 4-х генов, организованных в компактный кластер klpABCDE (Приложение 1). Кластер включает в себя ген пептида-предшественника klpA, гены белков процессинга klpB, klpC, klpD и ген белка-транспортера klpE, выкачивающий зрелый пептид из клеток наружу (Приложение 1).
Нами клебсазолицин был получен при помощи экспрессии биосинтетического кластера klpABCDE из Klebsiella pneumonia в гетероллогическом суррогатном хозяине Е. coli с последующей очисткой из культуральной среды путем твердофазной экстракции на картридже С18 и ВЭЖХ с обращенной фазой.
Клебсазолицин предсталяет собой полипептдную цепь из 23 аминокислотных остатков с формулой
SerGlnSerProGlyAsnCysAlaSerCysSerAsnSerAlaSerAlaAsnCysThrGly
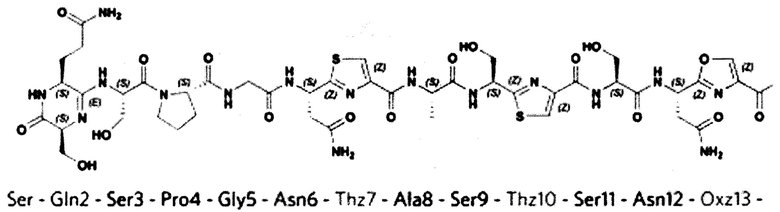
GlyLeuGly, содержащую ряд модификаций (фигура 1): а) остатки Cys7, Cys10, Cys18 и Ser13 образуют три тиазольных кольца и одно оксазольное соответственно; б) первые два N-концевых остатка (S1Q2) вовлечены в модификацию общей формулы I, приводящую к потере молекулы воды (-18 Да) и формированию 6-ти членного амидинового кольца. Такая модификация является уникальной среди группы линейных азол(ин)-содержащих пептидов (ЛАП).
Для установления химической структуры клебсазолицина, авторы использовали двумерный ядерный магнитный резонанс (ЯМР).
ЯМР-спектры регистрировали на растворенном в DMSO-D6 образце 5 мМ клебсазолицина, обогащенного изотопами 13С и 15N, находящегося в трубке Шигеми при 55°С. Все спектры ЯМР регистрировались на спектрометре Varian DirectDrive NMR System 700 МГц, снабженном 5-миллиметровым инверсным солеустойчивым датчиком тройного резонанса 1H/13C/15N. Данные ЯМР обрабатывали и анализировали с помощью Mestrelab Mnova версии 10.
Полное определение отдельных аминокислот, включая один пролин, четыре глицина, три аспарагина, три аланина, один лизин, пять серинов и один неканонический глутамин, а также три тиазола и один оксазол, получили из спектров 1Н, 13С, 1H-TOCSY, 1H-COZY, 13C-HSQC, 13С-НМВС, 13C-COZY, 13С-HSQCTOXY, 15N-HSQC, 15N-HSQCTOXY и HNCO (фигура 2).
Аланины, глицины, аспарагины, треонин и лейцин были определены из 1Н, 1H-TOCSY, 1H-COZY спектров 1H-сигнала (фигура 2. А) и из 13С-сигнала спектра 13C-HSQC (фигура 2. Б). Определение серинов, пролина и глутамина сделали на основе 13C-HSQCTOXY спектра. Боковые цепи аспарагинов и глутамин были определены из 13С-НМВС спектра. Тиазолы и оксазол были установлены из 13C-HSQC, 13C-HSQCTOXY и 13С-НМВС спектров. Корреляции С5-Н5 были установлены из 13C-HSQC спектра (фигура 2. В). Сигналы С4, С6 и Oxz13 были определены из 13C-COZY (фигура 2. Г). Сигналы карбонильных атомов углерода установили из 13С-НМВС, 13C-COZY и Н-С проекций HNCO спектра. NH-группы были определены из спектров 15N-HSQC (фигура 2. Д) и 15N-HSQCTOXY спектров. Последовательность связей между различными аминокислотами была установлена на основе анализа наблюдаемых корреляций НМВС 1Н-13С сигналов HSQC (фигура 2. Е). Связь Pro4-Gly5 была определена из НМВС-корреляций Pro4-СО с Gly5-HN и Pro4-СО с Gly5-Hα. Связи Gly5-Asn6, Ala8-Ser9, Ser11-Asn12 и Thr19-Gly20 были установлены аналогичным образом. Связи между Ser15-Ala16 и Ala16-Asn17 была установлена через НМВС-корреляции Ser15-CO с Ala16-Нα и Ala16-СО с Asn17-Hα соответственно. Связь между Gly20 и Gly21 была установлена через НМВС-корреляции Gly20-CO с Gly21-HN. Связи Gly21-Leu22 и Leu22-Gly23 были определены аналогичным образом. Связь Asn6-Thz7 очевидна из НМВС-корреляций Thz7-C2 и Thz7-C4 с Asn6-Hα и Asn6-Hβ соответственно. Аналогично обнаружено соединение Ser9-Thz10 и Asn17-Thz18. Связь Oxz13-Ala14 очевидна из НМВС-корреляций Oxz13-C6 с Ala14-Нα и Ala14-HN. Аналогичным образом были установлены связи Ala8-Thz7, Thz10-Ser11 и Thz18-Thr19. Связь Ala12-Oxz13 очевидна из НМВС-корреляций Oxz13-C2 с Ala12-Нα и Ala12-Нβ. Связывание Ser1'-Glu2 и Glu2-Ser3 очевидно из НМВС-корреляций Glu2-CO с Serl'-Нα, Ser3-Hα и Serl'-HN, Ser3-HN (фигура 2. Ж).
Наблюдаемые две группы сигналов от Ser1, Gln2, Ser3 и Asn6, где совокупность состояний, связанных как 5/3, указывают на наличие таутомерного равновесия между двумя протонированными формами амидного фрагмента. Эти данные доказывают, что N-концевые аминокислотные остатки Ser1 и Gln2 образуют шестичленный гетероцикл через амидиновую связь.
Антимикробная активность
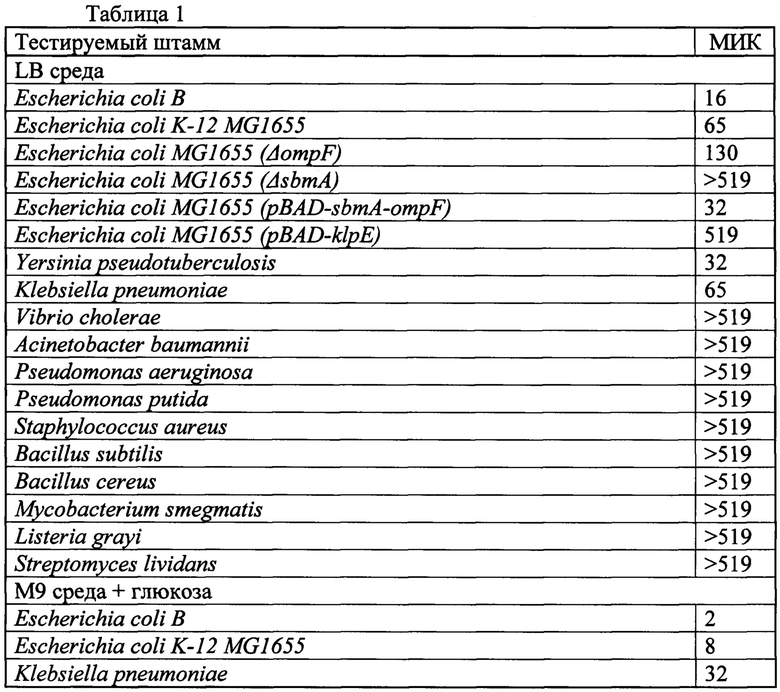
Антибактериальная активность клебсазолицина была оценена с использованием панели грамположительных и грамотрицательных микроорганизмов при помощи спот-теста на бактериальных газонах и методом микродилюции в жидкой среде на основе рекомендаций Института клинических и лабораторных стандартов (ИКЛС). КЛБ ингибирует рост ряда протестированных грамотрицательных штаммов - Е. coli, K. pneumoniae, и Yersinia pseudotuberculosis в микромолярных концентрациях 16-65 мкМ и не активен в отношении грамотрицательных микрорганизмов Acinetobacter baumannii, Pseudomonas aeruginosa, Pseudomonas putida и Vibrio cholerae, a также всех тестируемых грамположительных бактерий (таблица 1).
Чтобы показать клеточную мишень нового антибиотика, в первую очередь, авторы прибегли к in vivo биосенсорной репортерной системе на основе Е. coli [28, Osterman IA et al. 2016], которая позволяет выявить ингибиторов синтеза белка и репликации ДНК. В такой системе соединения, ингибирующие трансляцию белка (например, эритромицин), индуцируют экспрессию красного флуоресцентного белка-репортера Катюшка 2S, в то время как соединения, вызывающие в клетках SOS-ответ, такие как ингибиторы ДНК-гиразы (например, левофлоксацин и микроцин В17), индуцируют экспрессию репортерного красного флуоресцентного белка (Red Fluorescent Protein, RFP). В дополнительных материалах на фотографии показано, что клебсазолицин сильно индуцирует экспрессию Катюшка 2S, но не вызывает экспрессию RFP, что указывает на то, что он ингибирует синтез белка, а не ДНК-репликацию.
Для подтверждения ингибирующего действия клебсазолицина в отношении ранних этапов трансляции, были осуществлены следующие биохимические, структурные и генетические эксперименты.
Авторы протестировали способность клебсазолицина останавливать синтез люциферазы светлячков in vitro в бесклеточном экстракте Е. coli S30. Бесклеточные системы синтез белка (БССБ) является удобным лабораторным инструментом для наблюдения за процессом транскрипции и трансляции in vitro, которые при БССБ обеспечиваются клеточными экстрактами, получаемыми путем физического разрушения клеток. Клебсазолицин тестировали в бесклеточном бактериальном экстракте Е. coli S30, содержащем мРНК, кодирующую в качестве репортерного белка люциферазу светлячков. На фигуре 3 приведен график зависимости интенсивности люминесценции от концентрации клебсазолицина в бесклеточных бактериальных экстрактах Е. coli S30 при совместной инкубации в течение 30 мин при 37С. Из этих данных наблюдалось сильное дозозависимое ингибирование трансляции клебсазолицином.
Сокристаллизация с 70S рибосомой.
Чтобы определить точный способы взаимодействия клебсазолицина с рибосомой, мы сокристаллизовали его с 70S рибосомой Thermits thermophilics и определили структуру комплекса методом рентгеновской кристаллографии с разрешающей способностью 2, 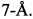 Полученные данные свидетельствовали в пользу единственного сайта связывания клебсазолицина - в выходном туннеле 50S большой рибосомной субъединицы.
Полученные данные свидетельствовали в пользу единственного сайта связывания клебсазолицина - в выходном туннеле 50S большой рибосомной субъединицы.
N-концевой амидиновый цикл в значительной степени обуславливает биологическую активности соединения, устанавливая множественные контакты в пептидил-трансферазном центре (дополнительный материал, рисунок 2. А-В): линкер между Ser1 и Gln2 образует две водородные связи с U2584 23S рРНК (дополнительный материал, рисунок 2. А, Б), гидроксильные группы боковых цепей остатков Ser1 и Ser9 устанавливают водородные связи с U2585 и U2506 соответственно, основными функциональными остатками ПТЦ. Кроме того, клебсазолицин напрямую взаимодействует с нуклеотидами А2058 и А2059 23S рРНК (дополнительный материал, рисунок 2. Б, В) и образует обширные Ван-дер-Ваальсовы контакты с U2609 23S рРНК (дополнительный материал, рисунок 2. А). Гетероциклы клебсазолицина устанавливают несколько стекинговых взаимодействий с нуклеотидами 23S рРНК (дополнительный материал, рисунок 2. А-В): плоскость амидинового кольца стекает по С2610 23S рРНК, первое и второе тиазольные кольца (Thz7 и Thz10) стекают над С2586-С1782 и А2062-А2503 парами оснований соответственно. Также после связывания с клебсазолицина значительно меняется конформация А2062 23S рРНК. Этот нуклеотид вращается более чем на 90°. В отличие от небольшого и компактного эритромицина, крупная молекула КЛБ сворачивается в глобулу в выходном туннеле пептида, чем полностью его закупоривает. При этом сайт связывания клебсазолицина не перекрывается с А- и Р-центрами рибосомы.
Другие представители семейства
Помимо клебсазолицина, ингибирующая активность была показана для еще двух соединений по изобретению общей формулы I, где Рр представлена: 1) Lys (пептид (1-13)_K14) и 2) LysSerAlaAsnCysThrGlyGlyLeuGly (пептид (1-13)_K14-G23). В опытах in vitro они оказались почти таким же активными, как и клебсазолицин (КЛБ), содержащий Рр в виде AlaSerAlaAsnCysThrGlyGlyLeuGly (таблица 2). Любопытно, что версия пептида по изобретению, содержащая усеченный фрагмент С-конца ((1-13)_K14) не способна при этом ингибировать рост чувствительных клеток. Это свидетельствует в пользу того, что С-концевые остатки необходимы для поглощения пептида чувствительными клетками, но не для взаимодействия их с рибосомой.
Однако усеченная до 11 аминокислот форма пептида (пептид (1-11)) практически полностью потеряла ингибирующую активность как в опытах in vitro, так и in vivo. Так же, как и формы пептида, лишенные третьего азольного кольца (пептиды (1-12)_А13_K14 и (1-12)_A13_K14-G23) (таблица 2).
Наиболее значительной особенностью, которая отличает пептид по изобретению от других тиазол-оксазол модифицированных пептидов, является наличие шестичленного амидинового цикла на N-конце молекулы, которое при связывании с рибосомой взаимодействует с нуклеотидами U2584, U2585 и C2610 23S pPHK.
Неспособность сформировать этот амидиновый цикл существенно снижает ингибирующую активность клебсазолицина. Так, его мутантная по третьей аминокислоте форма (КЛБ (S3A)), не несущая амидиновый цикл на N-конце (фигура 4), практически полностью теряет ингибирующую активность.
Мутации клебсазолицина по первой (КЛБ (S1A)) и второй (КЛБ (Q2N)) аминокислотам приводят к существенному снижению и полной потере активности клебсазолицина соответственно.
Таким образом, соединение, состоящее из 13-ти аминокислот по общей формуле I, включающее три азольных и один азолиновый цикла, является минимальным функциональным ядром, способным ингибировать рибосому.
Преимущества
1) Изобретение отличается от существующих аналогов, макролидных антибиотиков, например, азитромицина (патент US 8701612, 1987) и пролин-богатых антибактериальных пептидов, производных онкоцина (патент US 8937155) или бактеницин-7 (патент ЕР 0665239) тем, что ингибирует синтез белка, связываясь с бактериальной рибосомой в участке, который не идентичен сайтам связывания аналогов.
2) Соединения по данному изобретению имеют обширный сайт связывания с рибосомой, значительно превосходящий таковой в случае макролидных антибиотиков. Это снижает вероятность возникновения резистентости, вызванной одиночными мутациями в рибосоме.
3) Соединения по данному изобретению полностью закупоривают выходной тоннель рибосомы и не позволяют новосинтезированным пептидам его преодолевать, в отличие от некрупных макролидов, которые лишь частично перекрывают тоннель.
4) В отличие от рибосомоориентированных пролин-богатых антибактериальных пептидов, которые высоко обогащены положительно заряженными аминокислотами и взаимодействуют с отрицательно заряженной рРНК, соединения по изобретению взаимодействуют с рибосомой главным образом путем укладки гетероциклов над основаниями рРНК. Этот уникальный способ укладки в выходном туннеле пептида рибосомы может служить перспективным химическим каркасом для разработки новых антибиотиков, таргетирующих бактериальную рибосому, а именно - ее выходной туннель.
5) Описанные в изобретении пептиды, по сравнению с пролин-богатыми антибактериальными пептидами, имеют отличную химическую структуру, что существенно расширяет класс антимикробных пептидов, таргетирующих рибосому, а также расширяет класс модифицированных пептидов в целом.
6) Описанные в изобретении пептиды является не аппробированным в широком клиническом применении, и, как следствие, не имеющим распространенных устойчивых мутантов в популяции чувствительных штаммов.
7) Поскольку соединения по данному изобретения являются рибосомально синтезируемыми пептидами, в отношении их модифицирования применимы методы как химического синтеза, так и молекулярной генетики.
8) Представитель патентуемого семейства, клебсазолицин, имеет узкий антимикробный спектр, что является существенным преимуществом клебсазолицина перед аналогами, антибиотиками широкого спектра действия, поскольку позволяет использовать его как основу для строго таргетированных лекарств.
9) Кроме того, клебсазолицин в сравнении с онкоцином и бактинецином имеет филогенетически далекий источник происхождения - прокариотические бактерии, что расширяет группу источников антимикробных пептидов, таргетирующих рибосому.
Заключение
Пептиды описанного семейства могут стать источником как новых антибиотиков, сконструированных на их основе, так и представляют интересный структурный каркас для разработки новых химических соединений, способных укладываться в выходном туннеле бактериальной рибосомы. Так или иначе, структурные сравнения различных химически несвязанных соединений, которые ингибируют рибосому, связываясь с выходным туннелем, показывают, что во всех случаях задействованы очень похожие структурные элементы рибосомы. Эта информация может быть использована для разработки гибридных ингибиторов. Более того, уникальная сложенная консистенция клебсазолицина в туннеле выхода пептида обеспечивает новый структурный эшафот для дальнейших улучшений.
| название | год | авторы | номер документа |
|---|---|---|---|
| Генетическая конструкция на основе двух индуцибельных экспрессионных векторов для экспрессии тиазол/оксазол модифицированного пептида в клетках бактерий E. coli; способ получения рекомбинантного штамма бактерий E. coli и способ получения модифицированного пептида на его основе | 2017 |
|
RU2694044C2 |
| Способ выявления наличия мутаций, приводящих к резистентности у Mycoplasma genitalium к макролидным и фторхинолоновым антибиотикам | 2019 |
|
RU2725477C1 |
| ОЛИГОНУКЛЕОТИДНЫЙ БИОЧИП ДЛЯ ИДЕНТИФИКАЦИИ ГЕНЕТИЧЕСКИХ ДЕТЕРМИНАНТ РЕЗИСТЕНТНОСТИ Neisseria gonorrhoeae К АНТИМИКРОБНЫМ ПРЕПАРАТАМ, НАБОР ОЛИГОНУКЛЕОТИДОВ, ИСПОЛЬЗУЕМЫХ ДЛЯ ИММОБИЛИЗАЦИИ НА БИОЧИПЕ | 2016 |
|
RU2636457C2 |
| СПОСОБ СИНТЕЗИРОВАНИЯ ФЛУОРЕСЦЕНТНО-МЕЧЕНОГО БЕЛКА | 1995 |
|
RU2162104C2 |
| СПОСОБ ВЫЯВЛЕНИЯ МУТАЦИЙ, ПРИВОДЯЩИХ К РЕЗИСТЕНТНОСТИ У Mycoplasma genitalium И Mycoplasma pneumoniae К МАКРОЛИДНЫМ АНТИБИОТИКАМ | 2017 |
|
RU2646123C1 |
| ПРЕМЕРСАЦИДИН, ПРОМЕРСАЦИДИН, ДНК (ВАРИАНТЫ), ВЕКТОР (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ МЕРСАЦИДИНА | 1995 |
|
RU2198924C2 |
| Способ получения промотора для конструирования вектора и способ конструирования вектора для экспрессии белков в клетках | 1990 |
|
SU1836424A3 |
| Штамм Streptomyces rochei MP21 - продуцент антибиотика кирромицина | 2022 |
|
RU2798207C1 |
| Способ определения Helicobacter pylori | 2017 |
|
RU2744190C1 |
| СЕМЕЙСТВО ПЕПТИДОВ - ИНГИБИТОРОВ АКТИВНОСТИ БЕЛКА RecA, БЛОКИРУЮЩИХ SOS-ОТВЕТ У БАКТЕРИЙ | 2016 |
|
RU2621862C1 |
Изобретение относится к области биотехнологии и медицины, конкретно к новым тиазол-оксазол модифицированным пептидам, обладающим способностью ингибировать бактериальную рибосому Е. coli, K. pneumoniae и Yersinia pseudotuberculosis за счет связывания в её выходном туннеле. Модифицированный пептид имеет общий вид NH2-Ser1-Gln2-Ser3-Pro4-Gly5-Asn6-Cys7-Ala8-Ser9-Cys10-Ser11-Asn12-Ser13-Pp-Ac со структурой I, как представлено в формуле изобретения, где NH2 - N-концевая амидная группа, Ас - С-концевая ацетильная группа, Рр - полипептидная цепь, выбранная из ASANCTGGLG и KSANCTGGLG, и где пептид содержит а) три азольных цикла, образованных остатками Cys7, Cys10, Ser13, и б) модификацию первых двух аминокислот, приводящую к образованию на N-конце молекулы 6-членного амидинового кольца. Изобретение может быть использовано в качестве антибактериального средства в медицине и экспериментальной биологии. Предложенные пептиды имеют обширный сайт связывания с рибосомой, значительно превосходящий таковой в случае макролидных антибиотиков, полностью закупоривают выходной тоннель рибосомы и не позволяют новосинтезированным пептидам его преодолевать. 4 ил., 2 табл.
Тиазол-оксазол-модифицированный пептид общего вида NH2-Ser1-Gln2-Ser3-Pro4-Gly5-Asn6-Cys7-Ala8-Ser9-Cys10-Ser11-Asn12-Ser13-Pp-Ac формулы I
где NH2 - N-концевая амидная группа, Ас - С-концевая ацетильная группа, Рр - полипептидная цепь, выбранная из ASANCTGGLG и KSANCTGGLG, Thz - аминокислота, образующая тиазол, Oxz - аминокислота, образующая оксазол, где пептид содержит а) три азольных цикла, образованных остатками Cys7, Cys10, Ser13, и б) модификацию первых двух аминокислот, приводящую к образованию на N-конце молекулы 6-членного амидинового кольца общей формулы II
 ,
,
обладающий способностью ингибировать бактериальную рибосому Е. coli, K. pneumoniae и Yersinia pseudotuberculosis за счет связывания в ее выходном туннеле.
| MELBY J | |||
| O | |||
| et al | |||
| "Thiazole/oxazole-modified microcins: complex natural products from ribosomal templates" | |||
| Current Opinion in Chemical Biology, 2011, 15(3), 369-378 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| УТКИНА И | |||
| Л | |||
| "Характеристика нового антибактериального тиазол-оксазол-модифицированного пептида из K | |||
| pneumoniae" | |||
| Бакалаврская работа | |||
| Санкт-Петербургский политехнический университет Петра Великого, Институт физики, нанотехнологий и телекоммуникаций | |||
| Санкт-Петербург, 2016, 49 с | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| СЕМЕЙСТВО ПЕПТИДОВ - ИНГИБИТОРОВ АКТИВНОСТИ БЕЛКА RecA, БЛОКИРУЮЩИХ SOS-ОТВЕТ У БАКТЕРИЙ | 2016 |
|
RU2621862C1 |
| US 8937155 B2, 20.01.2015 | |||
| US 5300629 A1, 05.04.1994 | |||
| WO 2009013262 A1, 29.01.2009 | |||
| Захват для испытаний цилиндрических образцов на растяжение | 1977 |
|
SU665239A1 |

